Embed Size (px)
Citation preview

Neuroscience Vol. 45, No. 2, pp. 461477, 1991 Printed in Great Britain
0306-4522/91 $3.00 f 0.00 Pergamon Press plc
0 1991 IBRO
A NOVEL EPITOPE EXPRESSED ON THE SURFACE OF DEVELOPING AND MATURE ASTROCYTES
D. M. D. LANDIS,* E. WELTER and G. SKOKDELB
Departments of Neurology and Neurosciences, School of Medicine, E-604, Case Western Reserve University, 2119 Abington Road, Cleveland, OH 44106, U.S.A.
Abstract-Plasmalemmal fractions from cultured astrocytes have been used as the immunogen in 8enerating a monoclonal antibody, termed 8Cl0, which binds to the surface of cultured astrocytes of the rat. 8ClO immunorcactivity is present on the membrane surface of cultured type 1 astrocytes, type 2 astrocytes, oligodendrocytes, meningeal cells, and 02A progenitor cells, and it persists after aldehyde fixation. The antibody also stains aldehyde-tixcd central nervous system, in a pattern which suggests that the plasma membranes of fine astrocytic processes in adult neuropil express the epitope. Astrocytic perikarya and processes in white matter are also stained, but there is no immunoreactivity present in neuronal processes or perikarya. Astrocytic processes in developing cerebellar cortex are stained at postnatal ages when some of these processes are guiding the migration of neuronal perikarya.
Astrocytes in the mature mammalian central nervous system are characterized by an enormous surface areaBs The initial indications of this very large surface area were provided by studies using thin-section electron microscopic methods, which revealed myriad fine astrocytic processes interwoven among neural elements. The protein composition of the astrocyte plasma membrane is not uniform across the surface of the cell. For example, freeze-fracture electron microscopic methods have identi~ed orthogonally packed arrays of intramembrane particles in astro- cyte plasma membranes, and have shown that these particle arrays are concentrated where astrocytic membranes are adjacent to vascular structures and where astrocytic processes form the surface of the brain.6-* Many astrocytic functions involve transport across the cell membrane, which further emphasizes the importance of this aspect of the cell’s architecture.
It has been relatively difficult to study processes which may alter the extent or specialization of astro- cytic membranes. Of the antibodies which have been found to be selective for astrocytes, most bind to intracellular epitopes, such as glial fibrillary acid protein (GFAP). The monoclonal antibody A2B5 does bind to a surface epitope, and has been extremely useful in the study of glial cell lineage and differentiation.‘2~‘3*‘6-18 The epitope, unfortunately, is not unique to astrocytes in tissue, and its expression in tissue is so weak as to limit the utility of the marker.
*To whom correspondence should be addressed. Abbreuiurions: EDTA, ethylen~iaminetetra acetate; GFAP,
glial fibrillary acid protein; HEPES, ~-2-hydroxye~yl- ~i~r~n~~-2~th~nesulphonic acid; PBS; phosphate- buffered saline; SDS-PAGE, sodium dodecyl sulphatc- polyacrylamide gel electrophoresis.
We have isolated a monoclonal antibody which binds to an epitope present on the surfaces of astro- cytes in developing and mature brain. Antibody binding can be demonstrated in vitro and in tissue sections, with and without aldehyde fixation. We describe here the binding characteristics of the anti- body in immunocytochemical preparations, and dis- cuss certain of the biochemical characteristics of the epitope.
EXPERIMENTAL PROCEDURES
Tissue culture Asfrocytes from neonatal ruts. The tissue culture tcch-
niques were based on the approach devised by McCarthy and de Vellis.” After the meninges, choroid plexus, cer- ebellum and brainstem had been removed, cerebral hemi- spheres from 24-48-h-old Sprague-Dawley rat pups were minced in L15 plating medium, exposed to 0.25% trypsin in the absence of divalent cations for 15 min at 37°C with agitation, and then to 0.1 mg/ml DNAse in the absence of divalent cations for 10min at 37°C. Dissociated cells were sedimented, and 5 x 106 viable cells (as judged by Trypan Blue exclusion) were inoculated into 75 cm2 Falcon flasks in 20 ml of L15-C0, growth medium supplemented with 20% fetal calf serum (GIBCO). The medium was changed every three days. Cells were confluent at seven to 10 days of culture. The llasks were sealed, placed on a rotary shaker and shaken for 12-18 h at 37°C to suspend oligodendrocytes and other cells growing over the confluent monolayer. The suspended cells were discarded (or used for oligodendrocyte culture; see below) and the flasks washed vigorously with fresh medium. The cells of the monolayer were incubated for 10min at 37°C in 5 mM EDTA in Hank’s balanced salt solution without divalent cations, and then collected after a 3-min exposure to trypsin-EDTA (GIBCO) in the absence of divalent cations. The cells were sedimented, rinsed, resuspended, and I x 10’ viable cells were inoculated into Falcon polystyrene tissue culture dishes (60 x 15 mm). The cultures were fed every three days with L15-CO~ growth medium supplemented with 20% fetal calf serum (GIBCO) and were usually confluent by t4 days.
467
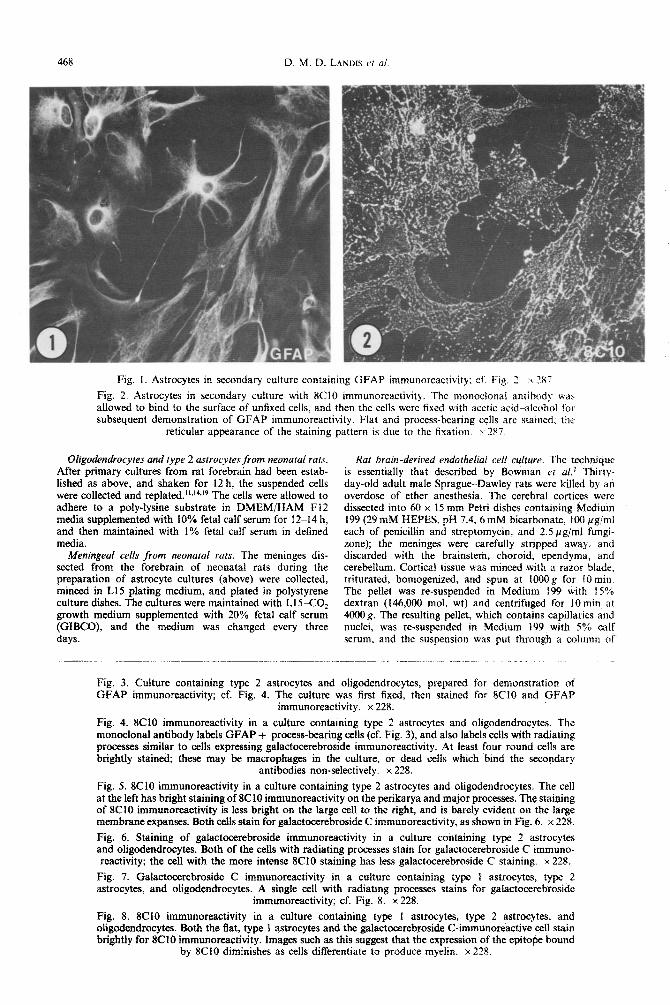
468 D. M. D. LANDIS C/ al.
Fig. I. Astrocytes in secondary culture containing GFAP immunoreactivity: cf: Fig. 7. \. 2X7
Fig. 2. Astrocytes in secondary culture with XC10 immunor~activity. The monoclonal antibody wah allowed to bind to the surface of unfixed cells, and then the ceils were fixed with acetic acid-alcohol foe subsequent demonstration of GFAP immunoreactivity. Flat and process-bearing cells are stained; the
reticular appearance of the staining pattern is due to the fixation. x 187.
Uligode~drocyte~ and type 2 ostrocyte~~rom neonatal rats. After primary cultures from rat forebrain had been estab- lished as above, and shaken for 12 h, the suspended cells were collected and replated.“,‘4,‘9 The cells were allowed to adhere to a poly-lysine substrate in DME~/HAM F12 media supplemented with 10% fetal calf serum for 12-14 h, and then maintained with 1% fetal calf serum in defined media.
Meningeal cells from neonatal rats. The meninges dis- sected from the forebrain of neonatal rats during the preparation of astrocyte cultures (above) were collected, minced in L15 plating medium, and plated in polystyrene culture dishes. The cultures were maintained with L15C0, growth medium supplemented with 20% fetal calf serum (GIBCO), and the medium was changed every three days.
Rat brain -derived endothetial celt c&we. The technique is essentially that described by Bowman c’t al.” Thirty- day-old adult male Sprague--Dawley rats were killed by an overdose of ether anesthesia. The cerebral cortices were dissected into 60 x 15 mm Petri dishes containing Medium 199 (29 mM HEPES, pH 7.4, 6 mM bicarbonate, LOO pgjml each of penicillin and streptomycin, and 2.5 pg/ml fungi- zone); the meninges were carefully stripped away, and discarded with the brainstem, choroid, ependyma, and cerebellum. Cortical tissue was minced with a razor blade, triturated, homogenized, and spun at 1OQOg for IO min. The pellet was re-suspended in Medium 199 with 15% dextran (146,000 mol. wt) and centrifuged for IOmin at 4OOOg. The resulting pellet, which contains capillaries and nuclei, was re-suspended in Medium 199 with 5% calf serum, and the suspension was put through a coiumn of
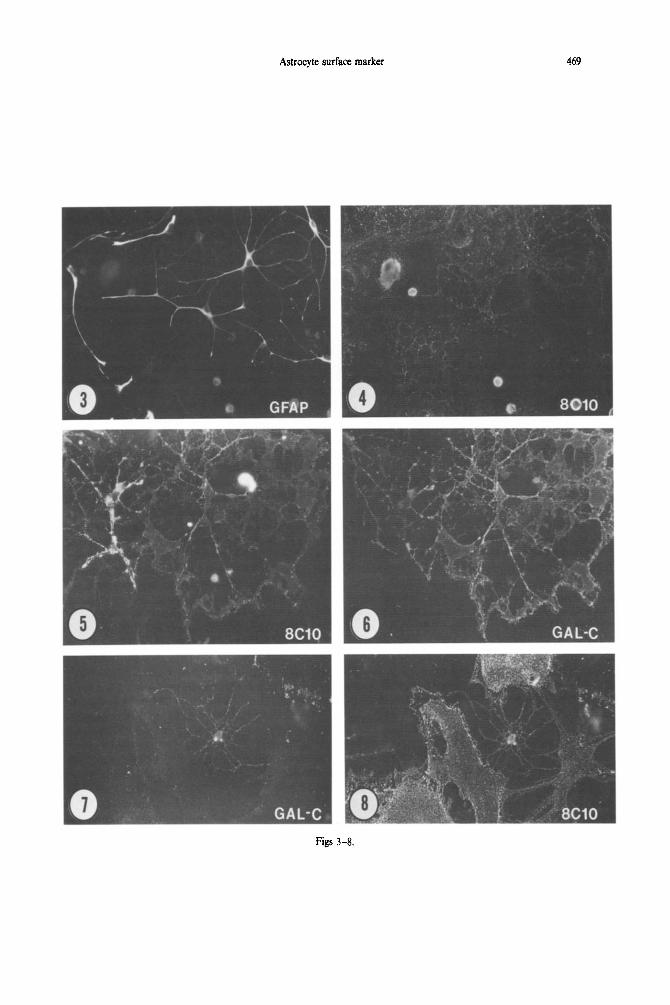
Fig. 3. Culture containing type 2 astrocytes and oligodendrocytes, prepared for demonstration of GFAP immunoreacti~ty; cf. Fig. 4. The culture was first fixed, then stained for 8CiO and GFAP
immunoreactivity. x 228.
Fig. 4. 8C10 immunoreactivity in a culture containing type 2 astrocytes and oligodendrocytes. The monoclonal antibody labels GFAP + process-bearing cells (cf. Fig. 3), and also labels cells with radiating processes similar to cells expressing galactocerebroside immunoreactivity. At least four round cells are brightly stained, these may be macrophages in the culture, or dead cells which bind the secondary
antibodies non-selectively. x 228.
Fig. 5. 8ClO immunoreactivity in a culture containing type 2 astrocytes and oligodendrocytes. The cell at the left has bright staining of 8ClO immunoreactivity on the perikarya and major processes. The staining of 8CIO immunoreactivity is less bright on the large cell to the right, and is barely evident on the large membrane expanses. Both cells stain for galactocerebroside C immunoreactj~ty, as shown in Fig. 6. x 228.
Fig. 6. Staining of gaiactocerebroside immunoreactivity in a culture containing type 2 astrocytes and oligodendrocytes. Both of the cells with radiating processes stain for galactocerebroside C immuno- reactivity; the cell with the more intense 8ClO staining has less galactocerebroside C staining. x 228.
Fig. 7. Galactocerebroside C imm~o~acti~ty in a culture containing type 1 astrocytes, type 2 astrocytes, and oligodendrocytes. A single cetl with radiating processes stains for galactocerebroside
immunoreactivity; cf. Fig. 8. x228.
Fig. 8. 8ClO immunoreactivity in a culture containing type 1 astrocytes, type 2 astrocytes, and oligodendrocytes. Both the flat, type 1 astrocytes and the galactocerebroside C-immunoreactive cell stain brightly for SC10 immuno~ctivity. Images such as this suggest that the expression of the epitope bound
by 8ClO diminishes as cells differentiate to produce myelin. x 228.
Astroqte surface marker
Figs 3-8.
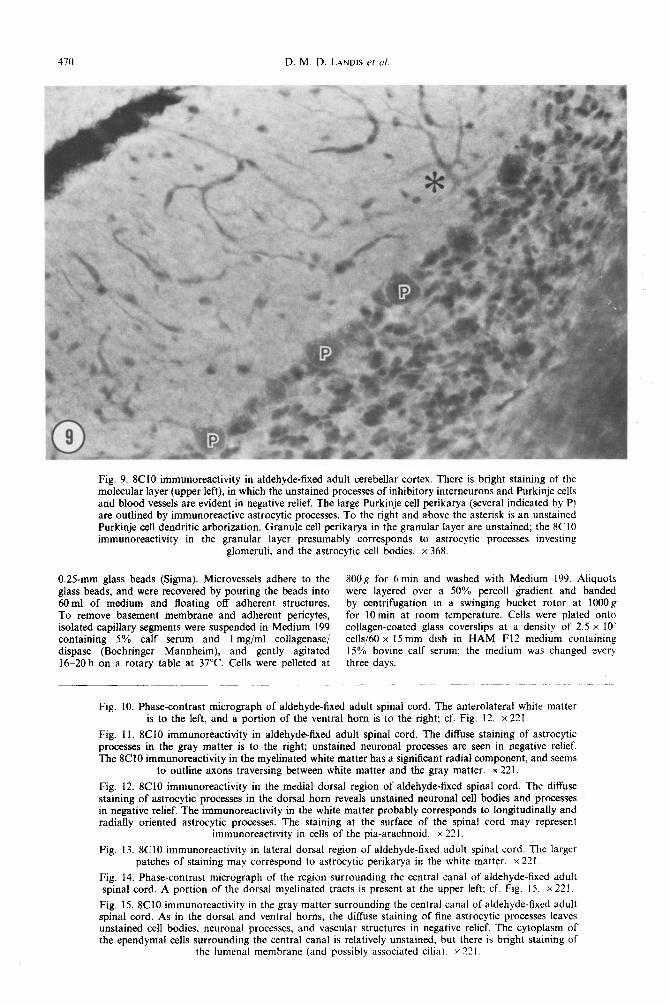
470 D. M. D. LANDIS rr u/
Fig. 9. 8C10 immunoreactivity in aldehyde-fixed adult cerebellar cortex. There is bright staining of the molecular layer (upper left), in which the unstained processes of inhibitory interneurons and Purkinje cells and blood vessels are evident in negative relief. The large Purkinje cell perikarya (several indicated by P) are outlined by immunoreactive astrocytic processes. To the right and above the asterisk is an unstained Purkinje cell dendritic arborization. Granule cell perikarya in the granular layer are unstained; the 8CI0 immunoreactivity in the granular layer presumably corresponds to astrocytic processes investing
glomeruli, and the astrocytic cell bodies. x 368.
0.25-mm glass beads (Sigma). Microvessels adhere to the 800g for 6 min and washed with Medium 199. Aliquots glass beads, and were recovered by pouring the beads into were layered over a 50% percoll gradient and banded 60ml of medium and floating off adherent structures. by centrifugation in a swinging bucket rotor at 1000~ To remove basement membrane and adherent pericytes, for 10min at room temperature. Cells were plated onto isolated capillary segments were suspended in Medium 199 collagen-coated glass coverslips at a density of 2.5 x 10‘ containing 5% calf serum and 1 mg/ml collagenase/ cells/60 x 15 mm dish in HAM F12 medium containing dispase (Boehringer Mannheim), and gently agitated 15% bovine calf serum; the medium was changed every 16-20 h on a rotary table at 37°C. Cells were pelleted at three days.
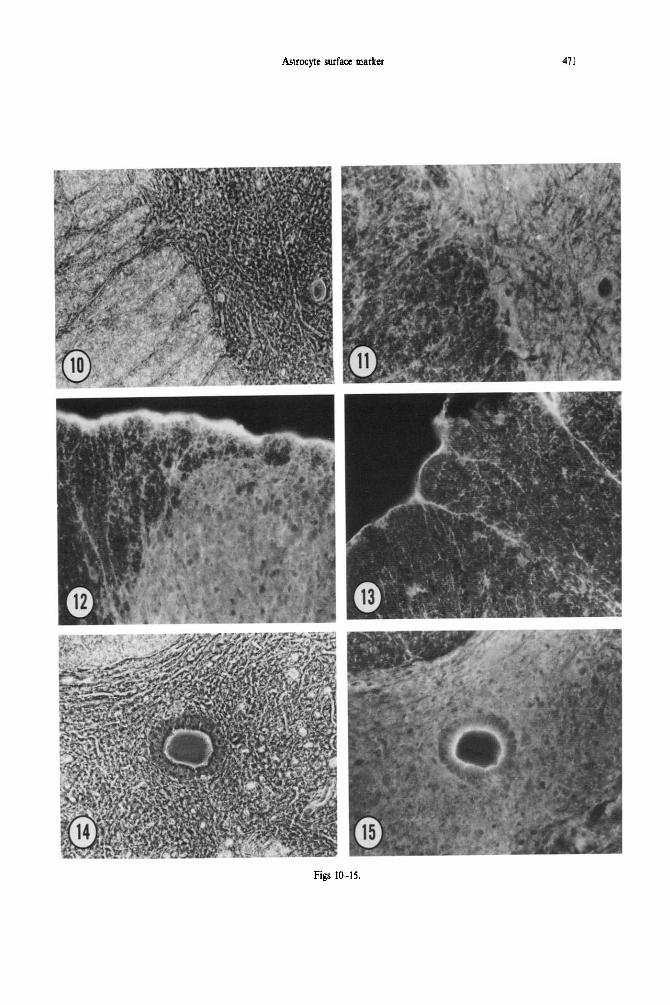
Fig. 10. Phase-contrast micrograph of aldehyde-fixed adult spinal cord. The anterolateral white matter is to the left, and a portion of the ventral horn is to the right; cf. Fig. 12. x 221.
Fig. 11. 8C10 immunoreactivity in aldehyde-fixed adult spinal cord. The diffuse staining of astrocytic processes in the gray matter is to the right; unstained neuronal processes are seen in negative relief. The 8C10 immunoreactivity in the myelinated white matter has a significant radial component, and seems
to outline axons traversing between white matter and the gray matter. x 221.
Fig. 12. 8C10 immunoreactivity in the medial dorsal region of aldehyde-fixed spinal cord. The diffuse staining of astrocytic processes in the dorsal horn reveals unstained neuronal cell bodies and processes in negative relief. The immunoreactivity in the white matter probably corresponds to longitudinally and radially oriented astrocytic processes. The staining at the surface of the spinal cord may represent
immunoreactivity in cells of the pia-arachnoid. x 221.
Fig. 13. 8ClO immunoreactivity in lateral dorsal region of aldehyde-fixed adult spinal cord. The larger patches of staining may correspond to astrocytic perikarya in the white matter. x 221.
Fig. 14. Phase-contrast micrograph of the region surrounding the central canal of aldehyde-fixed adult spinal cord. A portion of the dorsal myelinated tracts is present at the upper left: cf. Fig. 15. x 221,
Fig. 15. 8C10 immunoreactivity in the gray matter surrounding the central canal of aldehyde-fixed adult spinal cord. As in the dorsal and ventral horns, the diffuse staining of fine astrocytic processes leaves unstained cell bodies, neuronal processes, and vascular structures in negative relief. The cytoplasm of the ependymal cells surrounding the central canal is relatively unstained, but there is bright staining of
the lumenal membrane (and possibly associated cilia). x 221.
Astrocyte surface marker 471
Figs IO-IS.

472 D. M. D. LANDIS P( ui
Preparation of immunogen from cultured astrocytes
Confluent secondary cultures of astrocytes were rinsed twice with chilled L 1 S-air medium, then with chilled 20 mM HEPES buffer (containing 1 mM EGTA, 1 mM MgCl,, 145 mM NaCl and 50pg/ml leupeptin, pH 7.35). Cells were scraped from the dish with a rubber policeman, and m~hanically lysed with 40 strokes of a pestle in a glass homogenizer. The lysate was sedimented for 10 mm at 4000 r.p.m., and the resulting supematant was then spun for - _ 60min at 30,000 r.p.m. The pellet was resuspended in HEPES buffer. lavered over a discontinuous sucrose nradi- ent (85, 50, and 15% sucrose), and centrifuged for 90 min at 25,000 r.p.m. Fractions were collected from the 15/50 interface, resuspended in HEPES buffer and spun for 60 min at 30,000 r.p.m. The supernatant was discarded, the pellet resuspended in HEPES and used as the immunogen.
Generation of m~noclonai antibodies
A Balb/c mouse was injected subcutaneously on one side of the abdomen with 1OOng of the astrocyte microsome preparation; the animal then received a second subcu- taneous injection of RIB1 adjuvant (RIB1 Immun~hem Res) into the other side of the abdomen. The mouse received two subsequent injections of the immunogen, each time with accompanying RIBI adjuvant. It was killed after an over- dose of ether anesthesia, the spleen was fused with NSl myeloma cells, and fused cells were plated in 96-well plates. Supematants were screened by ELISA against live astro- cytes (cultured with techniques as above, but the secondary cultures were placed into 96-well plates), and by ELISA against the immunogen bound to the plastic of 96-well plates (2.5 _ug of protein/well). The 8C10 clone has been sub-cloned by limiting dilution on several occasions.
Ascites containing the monoclonal antibody were ob- tained by injecting a pristane-pretreated balb/c mouse in- traperitoneally with hybridoma cells. Ascites were tapped at 24-48-h intervals after accumulation of a&tic fluid became evident, to avoid undue discomfort to the host in accordance with NIH guidelines.
Purl~cation of the monorlonai ~ti~ody
As&tic fluid containing SC10 activity was treated first with Lipoclean Clearing Agent (Calbiochem), diluted 1:2 with Tris buffer at pH 7.3, and then applied to a Protein A/G column (ImmunoPure, Pierce). The bound antibody was eluted with 0.1 M glycine at pH 2.5. The elution fractions were adjusted to neutral pH, and then concen- trated by ultrafiltration (Centriprep, Amicon). The immuno- globulin in such preparations was p~dominantly IgG,, as assessed by ELISA (ScreenType, Boehringer Mannheim). The isotype of the 8C10 monoclonal antibody has been confirmed using the supematant of hybridoma cells grown in defined media.
Immunoprecipitation
Microsomal fractions were prepared from cultured astro- cytes as described above. Several fractions from the sucrose gradient were found to contain material which bound 8C10,
as assessed by dot blotting. These fractions (15% s~~rost‘ 50% sucrose, 15/50 interface. and 50/85 interface) were pooled and treated with 0.5% (w/v) CHAPS. The detergent- extracted microsomal preparations continue to show binding to 8ClO as determined by dot blotting.
Purified 8C10 was bound to Protein A,‘G bead&, washed with phosphate-bused saline (PBS) and mixed with detergent extracts of microsomal fractions isolated from cultured astrocytes. After 16 h at 4°C the samples were centrifuged, the pellet washed in detergent-containing buffer, and the pellet then prepared for sodium dodecyl- sulphatepolyacrylamide gel electrophoresis (SDS-PAGE).
Purified SC10 (300/1g/ml) was added to the medium of cultured astrocvtes (50 ul of 8ClO to 1 .O ml of L15--CO? medium containing ‘20% fetal calf serum or to 1.0 mi L15C0, without serum supplementation; this concen- tration of 8ClO is equivalent to or exceeds that which gives maximal labeling in immunocyto~hemical preparations). After incubation for 2-4 h at 4°C the cells were rinsed with PBS, scraped from the dish, and homogenized m either 1% Triton X-100, 0.5-1% CHAPS, or in a Triton deoxy~holat~SDS mixture. The homogenate was centri- fuged at 35,000 r.p.m. for 50 min, and the supernatant was then mixed with Protein A/G beads in a 10: 1 or 25: 1 ratio. The mixture was incubated 16 h at 4°C’. and then prepared for SDS-PAGE.
An 8ClO immunoaffinity column was prepared by bind- ing purified 8ClO to a hydrazide-activated gel support (Hydrazide AvidChrom, BioProbe). A detergent extract of homogenized cultured astrocytes was prepared in 0.1% SDS, and the SDS subsequently removed by precipitation with 0.1 M potassium phosphate. The supernatant was shown to contain the antigen by dot blotting. Approxi- mately 4 ml of antigen-containing solution (800 pg protein; ml) were passed over a l.O-ml immunoafBnity column. Material bound to the column was eluted in 0.1 M glycine at pH 2.5 and prepared for SDS-PAGE.
Protease sensitivity
Trypsin (GIBCO) was added to a homogenate of cultured astrocytes in a I.5 to 4.0-fold excess (w/w protein). After a 3-30-min incubation at 37°C soybean trypsin inhibitor (Sigma) was added (1 or 1.5 pg inhibitor/l ng trypsin), and the samples were kept for 30 min at 4°C prior to dot blotting on nitrocellulose paper.
Immunocytochemistry
Cells to be studied with immunocytochemical techniques were grown on acid-cleaned glass coverslips in Petri dishes.
For demonstration of 8ClO immunoreaetivity without fixation, cultures were rinsed in L15-C02 medium and incu- bated with 8ClO ascites (diluted I :1-i : 50 with L15-COZ medium) for 1 h at 4°C. The cultures were then rinsed, and exposed to goat anti-mouse IgG conjugated to rhodamine or fluorescein (Cappel) for 1 h at 4°C. After rinsing, the coverslips were mounted in 5% ~-pFopyl-fate (Sigma) in glycerol, and examined in a Zeiss 405 inverted microscape fitted with epifluorescence illumination.
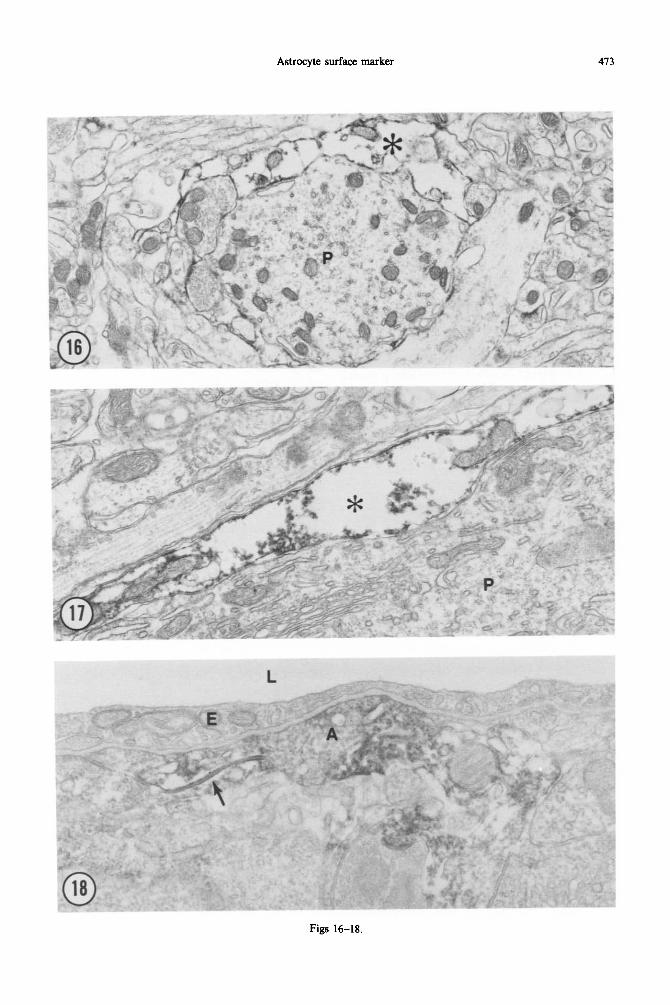
Fig. 16. SC10 immunoreactivity in adult cerebellar cortex prepared for electron microscopic study. The peroxidase reaction product is localized to the membrane and cytoplasm of an astrocytic process (asterisk) adjacent to the cross-sectioned Purkinje cell dendrite (p). There is no staining of neuronal processes. Most
of the fine astrocytic processes in this field are also unstained. x 14,352.
Fig. 17. 8ClO immunoreactivity in astrocytic processes (asterisk) investing the cell body of a Purkinje neuron (P). The peroxidase reaction product appears to adhere to the inner surface of the astrocytic membrane and to int~~ellular organelles, such as mitochondria. 8ClO jmmunoreactivity is especially
prominent in these peri-neuronal sheaths; cf., e.g. with Fig. 9. x 28,704.
Fig. 18. 8ClO immunoreactivity in a perivascular astrocytic process in adult cerebellar cortex. The lumen (L) of the capillary is above, lined by an endothelial cell (E). The immunoreactive astrocytic process (A) forms a gap junction with a second immunoreactive astrocytic process; the gap junction is lined by
peroxidase reaction product (arrow). x 36.800.
Astrocyte surface marker 413
Figs 16-18.

474 D. M. D. LANDIS et al
Cultures prepared for demonstration of 8C10 and GFAP immunoreactivity were first incubated with 8C10 and secondary antibody as above. Then the coverslips were washed in PBS with 2 mM CaCl,, and then fixed for 10 min in 5% acetic acid and 95% ethanol at - 20°C. Fixed cells were incubated with rabbit antisera to GFAP (Accurate Chemical & Scientific Corporation) at dilutions of 1: 50-1:200 for 1 h at room temperature, rinsed, and then incubated with goat anti-rabbit IgG conjugated to fluorescein or rhodamine (Cappel).
Cultures prepared for demonstration of 8ClO immuno- reactivity after fixation were rinsed, and then fixed in 4% parafonnaldehyde in 100 mM phosphate buffer at pH 7.35 for 20min. After rinsing in PBS with 1% bovine serum albumin, the cultures were exposed to 8ClO and secondary antibody as above, except at room temperature. Fixed cultures prepared for study of A2B5 immunoreactivity were treated in similar fashion. Cultures used for galacto- cerebroside immunoreactivity were first exposed to rabbit anti-galactocerebroside C (Chemicon), rinsed and incubated with goat anti-rabbit IgG conjugated with rhodamine or fluorescein, and then prepared for 8ClO immunoreactivity as above (in some instances, the cells were fixed with aldehydes before 8ClO staining).
Sprague-Dawley rats to be studied with light microscopic immunocytochemical techniques were anesthetized with intraperitoneal pentobarbital (50 mg/kg), and killed by perfusion with 4% paraformaldehyde in 1OOmM phos- phate buffer at pH 7.35. The perfusion was maintained for 5-10 min, and the animal was left undisturbed for 1 h prior to removal of the brain. Tissue was soaked in 30% sucrose in PBS for 12-18 h at 4°C frozen, and sectioned to lo-12 pm thickness with a Harris-Bright cryostat.
For light microscopic study, sections were incubated in the monoclonal antibody at various dilutions for 2-24 h, rinsed, and incubated in goat anti-mouse IgG conjugated to rhodamine.
Sprague-Dawley rats to be studied with electron micro- scopic immunocytochemical techniques were anesthetized with intraneritoneal nentobarbital(50 mR/kR). and then killed by perfusion with 4b/, paraformaldehyie,?l:5% glutaralde- hyde in 100 mM phosphate buffer at pH 7.35 for 10 min at 37°C. The brain was removed, postfixed by immersion in the same fixative for 2-4 h at room temperature, and rinsed in 1OOmM phosphate buffer. Sections 30-50pm thick were cut on a Vibratome, and incubated with 8ClO for 12-14 h at 4°C. The tissue was rinsed, and immunoreactivity local- ized with sequential exposure to biotin-conjugated anti- mouse IgG and avidin-biotin horseradish peroxidase (Vector Labs, Burlingame, CA). Peroxidase activity was demonstrated with 0.05% diaminobenzidine in 0.01% H,O,. The sections were briefly postfixed in 1% osmium tetroxide, and embedded in Epon. Some thin sections were stained with lead citrate and uranyl acetate.
RESULTS
Glial cells cultured from neonatal rats
The monoclonal antibody 8ClO bound to the surfaces of unfixed or fixed astrocytes in secondary cultures (Figs 1,2). All cells which contained GFAP immunoreactivity were stained by 8ClO. The inten- sity of staining was fairly uniform across the surface of each cell, but seemed to be accentuated at the edge, where one cell contacted or overlapped the membrane of another cell. Cell shape was typically hetero- geneous in these cultures, especially when they had become confluent. However, process-bearing and flat cells seemed to be stained with similar intensity.
Immunoreactivity was present at 24 h of culture, and persisted through three weeks in secondary culture.
In order to examine the distribution of 8ClO immunoreactivity in type 2 astrocytes and in myelin- forming cells, we prepared cultures enriched in these cell types by plating cells which had been suspended by shaking primary cultures, and growing the cells in defined media. Most of the GFAP +cells in such cultures had long unbranched processes. and resembled the type 2 cells described by Raff and colleagues in cultures derived from neonatal rat optic nerve.‘2~‘3~‘6-18 These cells also were stained with 8ClO (Figs 3, 4). Oligodendroglial cells in such cultures were recognized by their content of galactocere- broside immunoreactivity. We stained cells for both 8ClO and galactocerebroside immunoreactivity, and found that oligodendroglial cells also were stained with 8ClO (Figs 5-8). The 8CIO immunoreactivity was more intense in galactocerebroside-immuno- reactive cells that were relatively undifferentiated. When the oligodendrocytes acquired extensive sheets of galactocerebroside-immunoreactive membrane, the 8ClO staining became progressively fainter and more difficult to demonstrate.
These cultures also contained cells with A2B5 immunoreactivity, similar to the 02A progenitor cells originally described in optic nerve. We did not attempt to double label cells with both A2B5 and 8C10 monoclonal antibodies. In sister cultures, how- ever, we observed that 8ClO stained cells with the same shape and GFAP immunoreactivity as those stained by A2BS.
We prepared co-cultures of astrocytes derived from neonatal rat forebrain and endothelial cells derived from mature rat forebrain. The astrocytes in such co-cultures could be identified by their content of GFAP immunoreactivity. When the co-cultures were stained for both 8ClO and GFAP immunoreactivity. the 8ClO immunoreactivity was demonstrated only on cells which also labeled with GFAP (data not illustrated).
Cultures of meninges from neonatal rats contained both GFAP-immunoreactive cells, which resembled the astrocytes in our usual secondary cultures, and also flat cells which did not stain for the presence of GFAP. Both cell classes had 8ClO immunoreactivity when prepared with and without parafotmaldehyde fixation (data not illustrated).
Immunoreactivity in mature rat central nervous system
Sections of cerebral cortex, hippocampus, cerebellar cortex, and spinal cord all contained 8C10 immuno- reactivity. In cerebellar cortex (Fig. 9) and spinal cord (Figs lo-15), the 8ClO immunoreactivity was much more intense in gray matter than in white matter. The staining pattern was consistent with the interpret- ation that astrocytic membranes were being stained through the thickness of the section. Neurons and vascular elements stood out in negative relief.

Astrocyte surface marker 415
Since fixation, section thickness and other staining parameters are rarely constant, it is difficult to rigor- ously compare staining intensity in different brain regions. However, we noted consistently that hippo- campal immunoreactivity was less than that of the cerebellar cortex. We undertook to correlate this difference in staining intensity with the structure of the same regions as visualized in thin-sectioned elec- tron microscopic preparations. Astrocytic processes invest virtually all synaptic junctions in cerebellar cortex, but they are much less frequent in the stratum radiatum in area CA1 of the rat hippocampal cortex. The difference in staining pattern is thus correlated with the extent of astrocytic processes evident in thin sections.
We used pre-embedding staining followed by avidin-biotin horseradish peroxidase labeling to examine the distribution of 8ClO immunoreactivity in cerebellar cortex at the light and electron microscopic levels. In these preparations, the pattern of immuno- reactivity at the light microscopic level was some- what different from that seen in immunofluorescence preparations. The glutaraldehyde-fixed, peroxidase- treated tissue seemed to show immunoreactivity in proximal processes of astrocytes, similar to the distri- bution of GFAP immunoreactivity, in addition to the more diffuse stain that we had presumed to be due to binding of the 8ClO to cell surfaces.
At the electron microscopic level, 8ClO immuno- reactivity could only be demonstrated in the most superficial l-3 pm of the 50-pm-thick Vibratome sections. Only astrocytic processes were stained, though not all astrocytic processes were stained (Figs 16-18). The peroxidase reaction product some-
times appeared to be associated with the inner surface of the plasma membrane, and to he adherent to intracellular organelles and bundles of intermediate filaments.
Immunoreactivity in developing central nervous system tissue
We used secondary cultures of astrocytes derived from neonatal rat brain to obtain the immunogen that generated the 8ClO monoclonal antibody, and we wanted to determine whether developing astro- cytes in vivo also bound the antibody. In cerebellar cortex, the astrocytic processes of the molecular layer in the first two postnatal weeks contained 8ClO immunoreactivity (Figs 19,20). There was no staining of neurons and no staining of the neuronal precursors in the external granular layer of the cerebellar cortex. The pattern suggests that radial glial processes guid- ing the migration of neuronal perikarya have 8ClO immunoreactivity.
Immunoreactivity in non-neural tissue
We studied several non-neural tissues at the light microscopic level, with and without prior para- formaldehyde fixation. We failed to recognize any immunoreactivity in renal cortex, gastric wall, the wall of the small intestine, and in the trachea. Thus far, we have not demonstrated any 8ClO immuno- reactivity outside the central nervous system in mature animals.
Biochemical characteristics of the epitope
The immunocytochemical studies indicate that the epitope is present on the surface of astrocytic
Fig. 19. 8ClO immunoreactivity in aldehyde-fixed developing cerebellar cortex, postnatal day 7. Stained astrocytic processes extend from perikarya in the nascent Purkinje cell layer (P) to the molecular layer (ML) and into the external granular layer (EGL). There is some staining in the granule cell layer (G), which could be associated with astrocytic processes investing glomerular synaptic complexes; the pattern is also consistent with staining of granule cell neuron surfaces. Neurons and proliferating cells in the external granular layer are unstained, but the pia arachnoid between the apposed folia does appear to
be stained. x 199.
Fig. 20. GFAP immunoreactivity in aldehyde-fixed cerebellar cortex at postnatal day 7, for comparison with Fig. 19 (these are adjacent, non-overlapping fields; the several layers of the cortex are in horizontal
register with Fig. 19). x 199.

476 D. M. D. LANDIS cl u/
membranes, and that binding to 8C10 is not prevented by brief exposure to paraformaldehyde.
The epitope which binds to 8ClO appears to be labile when treated by trypsin. Immunoreactivity, as assessed by dot blotting, declines with progressively longer intervals of incubation, and with increasing concentrations of trypsin.
We prepared Western blots by solubilizing the microsomal fraction in boiling SDS and then running it on a polyacrylamide gel. The electrophoresed pro- teins were transferred to a nitrocellulose substrate, but no immunoreactivity could be demonstrated.
We attempted to immunoprecipitate a polypeptide binding to 8Cl0, using three slightly different pro- cedures. We prepared detergent extracts of the micro- somal fractions prepared from cultured astrocytes, and found that immunoreactivity (as assessed by dot blotting) persisted in Triton X-100, CHAPS, and SDS if used at low concentrations. These detergent extracts were then mixed with 8ClO bound to a substrate by protein A/G or by a hydrazide link. In a third set of experiments, the purified 8ClO was first added to cultured astrocytes, and then the astrocytes were homogenized, and the homogenate incubated with Protein A/G beads to immobilize the 8C10 and material bound to it. We eluted material bound to the 8ClO and prepared it for SDS-PAGE and subsequent Western blotting. In other experiments, the complex of 8C10, substrate, and bound material was prepared for SDS-PAGE. None of these procedures revealed a band in the gels or the blots which seemed to be concentrated from the starting material by the 8C10. We presume that either the epitope is labile under the preparative conditions used, or it is not associated with a polypeptide.
DISCUSSION
The epitope recognized by the 8ClO monoclonal antibody is present on the surface of all cells in the rat central nervous system which manifest GFAP immunoreactivity. It is a useful marker for astrocytes, and because the epitope is located on the surface of the cell, it may be employed in experiments which preclude the use of intracellular label.
The monoclonal antibody binds to type 1 and type 2 astrocytes in culture, and also labels cultured cells which express galactocerebroside C immuno- reactivity. Less direct evidence indicates that cells similar to the 02A progenitor cell described in the optic nerve12313,16-18 also express the epitope labeled by 8ClO. Thus, both of the lineages which give rise to astrocytes in the rat optic nerve have in common the epitope labeled by 8ClO. It seems likely that expression of the 8ClO epitope. is transient in the differentiation of myelin-forming cells, since there is no 8ClO binding to myelin sheaths in the white matter of mature brain. The staining of radial glial processes in the molecular layer of developing cerebellar cortex suggests that astrocytes involved
in guiding neuronal migration may have XC i (1 immunoreactivity.
There is no staining of neuronal perikarya or large processes in the immunocytochemical preparations studied to date. This suggests, but does not prove that the epitope is restricted to glial and meningeal cells. In preliminary studies, we have stained co- cultures of neurons and glia from developing spinal cord and have not demonstrated 8ClO immuno- reactivity in cells with neuronal shape (Miller and Landis, unpublished observations).
The epitope may be present in the cytoplasm as well as the surface of the plasma membrane of astrocytes. Immunocytochemical studies of aldehydc- fixed brain at the electron microscopic level reveal peroxidase reaction product adjacent to the inner surface of the cell plasma membrane and adjacent to organelles and bundles of intermediate filaments. It remains to be determined whether the reaction product reflects the precise location of the epitope. or whether localization has been confused by migration of the reaction product.
Preliminary biochemical investigations have not yet succeeded in characterizing the epitope bound by the antibody. Immunoreactivity evident in dot blots seems to decline with exposure to detergents. Immunoreactivity in dot blots also declines with exposure to trypsin. In preliminary experiments, there is evidence that staining of astrocytes freshly dissoci- ated from optic nerve is eliminated by exposure to papain (Barres, personal communication). Both of these lines of evidence would suggest that the epitope has a polypeptide component. We have not succeeded in visualizing immunoreactivity in Western blots. Our attempts to immunoprecipitate a polypeptide from detergent extracts of cultured astrocytes have not yet succeeded.
The immunocytochemical results, however, can exclude several components of the astrocyte surface membrane. The pattern of immunoreactivity does not resemble the distribution of gap junctions or of assemblies, the specialization of intramembrane par- ticle distribution which we have shown to be present only in astrocytes in the central nervous system.r Other cell types outside the central nervous system which have assemblies have not yet been found to have 8ClO immunoreactivity, such as gastric parietal cells’ or the light cells of the distal collecting duct of the kidney.4 The pattern of immunoreactivity does not correspond to the distribution of caveolae in cultured astrocytes, as found in freeze-fractured preparations (Landis, unpublished observations).
Our immunocytochemical studies of cultured astrocytes suggest that cells expressing A2B5 also express 8ClO immunoreactivity, but it is clear that 8ClO immunoreactivity is present in cells lacking A2B5 immunoreactivity and vice versa.3.‘8.M The dis- tribution of 8ClO immunoreactivity is different from that described with antisera to the ganglioside GD3,” and is also different from the distribution of the NG:!

Astrocyte surface marker 411
antigen of chondroitin sulfate proteoglycan.9~‘0,22 Cells with galactocerebroside C immunoreactivity also express 8ClO immunoreactivity, but the presence of 8ClO immunoreactivity on the surface of type 1 astrocytes in culture indicates that the epitope recog- nixed by the 8ClO monoclonal antibody is different from the epitopes bound by 01 and 04.*l
CONCLUSION
Despite the present uncertainty about the nature of the epitope, we anticipate that the 8ClO monoclonal antibody will be a useful reagent in studies of astro- cyte differentiation and function. Thus far, it has been difficult to estimate the surface area of astrocytic membranes in tissue. 8ClO immunoreactivity, how- ever, appears to be distributed over astrocytic sur-
faces in vitro and in tissue, and thus may serve to identify conditions which alter the surface area of this cell class. The formation of glial scars, for example, is likely to involve an increase in the amount of astrocytic membrane in a given volume of brain.
Because the epitope is located on the surface of astrocytes, it may be possible to select positively (by FACS) or negatively (by complement-mediated lysis) for astrocytes in vitro. These approaches may permit one to examine neuronal differentiation and function in vitro in the presence or in the absence of astrocytes.
Acknowledgements-H. R. Payne and L. A. Weinstein provided excellent technical assistance. We wish to thank Drs Barbara Barres and Robert H. Miller for allowing us to report preliminary tindings of work carried out in collaboration with them. This research was supported by NS 22614.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
REFERENCES
Bordi C. and Perrelet A. (1978) Orthogonal arrays of particles in plasma membrane of the gastric parietal cell. Anat. Rec. 192, 297-304. Bowman P. D., Betz A. L., Ar D. et al. (1981) Primary culture of capillary endothelium from rat brain. In Vitro 17, 353-362. Eisenbarth G. S., Walsh F. S. and Nirenberg M. (1979) Monoclonal antibody to a plasma membrane antigen of neurons. Proc. natn. Acad. Sci. U.S.A. 76, 4913-4917. Humbert F.. Pricam C.. Perrelet A. and Orci L. (1975) Snecific nlasma membrane differentiations in the cells of the kidney collecting tubules. J. ultrastruct. Res. 52, 13-26. a a Landis D. M. D. (1986) Membrane structure in astrocytes. In CeN Biology and Pathology of Astrocytes (eds Fedoroff S. and Vernadakis A.), pp. 61-76. Academic Press, Orlando. Landis D. M. D. and Reese T. S. (1974) Arrays of particles in freeze-fractured astrocytic membranes. J. Ceil Biof. 60, 316-320. Landis D. M. D. and Reese T. S. (1981) Membrane structure in mammalian astrocytes: a review of freeze-fracture studies in adult, developing, reactive and cultured astrocytes. J. exp. Biof. 95, 35-48. Landis D. M. D. and Reese T. S. (1982) Regional organization of astrocytic membranes in cerebellar cortex. Neuroscience 7, 937-950. Levine J. M. and Card J. P. (1987) Light and electron microscopic localization of a cell surface antigen (NG2) in the rat cerebellum: association with smooth protoplasmic astrocytes. J. Neurosci. 7, 271 l-2720. Levine J. M. and Stallcup W. B. (1987) Plasticity of developing cerebellar cells in vitro studied with antibodies against the NG2 antigen. J. Neurosci. 7, 2721-2731. McCarthy K. D. and de Vellis J. (1980) Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. CeN Biof. 85, 890-902. Miller R. H., David S., Pate1 R., Abney E. R. and RatT M. C. (1985) A quantitative immunohistochemical study of macroglial cell development in the rat optic nerve: in uiuo evidence for two distinct astrocyte lineages. Devf Biof. 111, 35-42. Miller R. H. and RatTM. C. (1984) Fibrous and protoplasmic astrocytes are biochemically and developmentally distinct. J. Neurosci. 4, 585-592. Morrison R. S. and de Vellis J. (1981) Growth of purified astrocytes in a chemically defined medium. Proc. natn. Acad. Sci. U.S.A. 70, 7205-7209. Norton W. T. and Farooo M. (1989) Astrocvtes cultured from mature brain derive from elial orecursor cells. J. Neurosci. 9, 769-775. _ _
_ .
Raff M. C., Abney E. R., Cohen J., Lindsay R. and Noble M. (1983) Two types of astrocytes in cultures of developing white matter: differences in mornholoav. surface aanaliosides. and arowth characteristics. J. Neurosci. 3. 1289-1300. RatT M. C., Abney E. R. and-Milled-R. H. (1984)-Two ghal ceil lineages diverge prenatally in rat ‘optic nerve. Devf Biof. 106, 53-60. Raff M. C., Miller R. H. and Noble M. (1983) A glial progenitor cell that develops in vitro into an astrocyte or an oligodendrocyte depending on culture medium. Nature 303, 390-396. Saneto R. P. and de Vellis J. (1985) Characterization of cultured rat oligodendrocytes proliferating in a serum-free, chemically defined medium. Proc. mm. Acad. Sci. U.S.A. 8X 35093513.
20. SchnitzerJ. and Schachner M. (1982) Cell type specificity of a neural cell surface antigen recognized by the monoclonal antibody A2B5. Ceff Tim. Res. 224, 635-636.
21. Sommer I. and Schachner M. (1981) Monoclonal antibodies (01 to 04) to oligodendrocyte cell surfaces: an immunocytochemical study in the central nervous system. Deuf Biof. 83, 311-327.
22. Stallcup W. B. and Beasley L. (1987) Bipotential glial precursor cells of the optic nerve express the NG2 proteoglycan. J. Neurosci. 7, 2737-2744.
(Accepted 20 May 1991)




























