Embed Size (px)
Citation preview

Adaptive and Pathological Inhibition of Neuroplasticity Associated WithCircadian Rhythms and Sleep
H. Craig Heller and Norman F. RubyStanford University
Asya RollsThe Technion, Haifa, Israel
Megha Makam and Damien ColasStanford University
The circadian system organizes sleep and wake through imposing a daily cycle of sleep propensity on theorganism. Sleep has been shown to play an important role in learning and memory. Apart from the dailycycle of sleep propensity, however, direct effects of the circadian system on learning and memory alsohave been well documented. Many mechanistic components of the memory consolidation processranging from the molecular to the systems level have been identified and studied. The question thatremains is how do these various processes and components work together to produce cycles of increasedand decreased learning abilities, and why should there be times of day when neural plasticity appears tobe restricted? Insights into this complex problem can be gained through investigations of the learningdisabilities caused by circadian disruption in Siberian hamsters and by aneuploidy in Down’s syndromemice. A simple working hypothesis that has been explored in this work is that the observed learningdisabilities are due to an altered excitation/inhibition balance in the CNS. Excessive inhibition is thesuspected cause of deficits in memory consolidation. In this article we present the evidence that excessiveinhibition in these cases of learning disability involves GABAergic neurotransmission, that treatmentwith GABA receptor inhibitors can reverse the learning disability, and that the efficacy of the treatmentis time sensitive coincident with the major daily sleep phase, and that it depends on sleep. The evidencewe present leads us to hypothesize that a function of the circadian system is to reduce neuroplasticityduring the daily sleep phase when processes of memory consolidation are taking place.
Keywords: Down’s syndrome, GABA, learning, memory, pentylenetetrazole, novel object recognition
Many hypotheses about possible functions of sleep and sleepstates have been advanced over the years (e.g., Benington &Heller, 1995; Crick & Mitchison, 1983; Diekelmann & Born,2010; Horne, 1988; Jouvet, 1998; Kreuger & Obal, 1993; McGinty& Szymusiak, 1990; Wehr, 1992; Tononi & Cirelli, 2003), but asubset of those hypotheses that focus on neural plasticity, and morespecifically on learning and memory has grown and gained supportin recent years (e.g., Frank, Issa, & Stryker, 2001; Maquet, 2001;Stickgold, 2005; Walker & Stickgold, 2004). Early evidencemostly came from experiments in which sleep deprivation wasshown to impair both declarative and procedural memories, andtherefore these results could have been due to indirect and non-
specific effects of sleep deprivation on the performance of subjectsduring retesting. However, continuing work pointed to specificfunctions of sleep in the processes of neural plasticity and ofmemory consolidation. Frank, Issa, and Stryker (2001) showedthat synaptic remodeling in the visual cortices of kittens subjectedto monocular deprivation depended on sleep. Experiments onhumans showed that when subjects were sleep deprived for a nightfollowing training in a perceptual skill task, but not tested untilafter two nights of recovery sleep, they showed no benefit from thetraining (Stickgold, James, & Hobson, 2000). In comparison, sub-jects allowed to sleep after task training showed large performanceimprovements the following day, and they continued to improvewith subsequent nights of sleep. The interpretation was that sleepwithin the same daily cycle following the training is essential foroptimal memory consolidation. Another study on human percep-tual skill training avoided sleep deprivation by examining theeffect of a daytime nap on acquisition of a perceptual skill andshowed performance benefits equivalent to those seen after a nightof sleep (Mednick, Nakayama, & Stickgold, 2003). These exper-iments and many others lead to the conclusion that sleep playsimportant roles in memory consolidation, and sleep deprivationinterferes with those processes.
A direct relationship between sleep and procedural memory waselegantly shown by Huber, Ghilardi, Massimini, and Tononi(2004) in experiments in which subjects were trained to use a
H. Craig Heller and Norman F. Ruby, Biology Department, StanfordUniversity; Asya Rolls, Rappaport School of Medicine, The Technion,Haifa, Israel; Megha Makam and Damien Colas, Biology Department,Stanford University.
The authors are deeply indebted to Bayarsaikhan Chuluun and GraceHagiwara for assistance with experiments. We wish to acknowledge thefollowing for financial support of our work: NIH Grant 1R01MH095837,Down Syndrome Research and Treatment Foundation, Research DownSyndrome, NARSAD (fellowship to AR), and NSF (fellowship to MM).
Correspondence concerning this article should be addressed to H. CraigHeller, Biology Department, Stanford University, 371 Serra Mall, Stan-ford, CA 94305-5020. E-mail: [email protected]
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
Behavioral Neuroscience © 2014 American Psychological Association2014, Vol. 128, No. 3, 273–282 0735-7044/14/$12.00 http://dx.doi.org/10.1037/a0036689
273

computer mouse to move the cursor to a target on the computerscreen. EEG recordings during subsequent sleep showed altera-tions in the quality of sleep specifically in the region of the motorcortex that was engaged by the training. The feature of sleep thatwas altered was the amount of slow wave activity (.5 Hz–4.5 Hz)in the EEG occurring during non-rapid eye movement (NREM)sleep. This band of EEG oscillations is called the delta band, andwhen quantified through Fourier analysis is referred to as deltapower. Furthermore, the performance improvement of the subjectsfollowing sleep was directly related to the measured increase indelta power. Thus, for a specific region of the brain, a clearrelationship was established between an electrophysiological fea-ture of sleep and improved performance in a motor-skill learningtask. The relationship between sleep and performance was alsodemonstrated in a study that manipulated EEG slow wave activityduring sleep using transcranial magnetic stimulation (TMS). In-ducing 0.75 Hz oscillations with TMS during early nocturnalNREM sleep enhanced the retention of hippocampal-dependentdeclarative memories in normal subjects (Marshall et al., 2006).
Electrophysiological studies of rodents are exploring the brainmechanisms involved in memory coding during wake and memoryconsolidation during sleep. Simultaneous recordings of many hip-pocampal neurons during maze running reveal activity patternsthat correlate with locations in the maze. These patterns are seen toreplay during quiet wakefulness and also during sleep (Lee &Wilson, 2002; Wilson & McNaughton, 1994), and during sleepthey seem to be relayed to the cortex in association with specificlocal field potentials (Carr et al., 2011; Ji and Wilson, 2007; Jones& Wilson, 2005; Ramadan et al., 2009; Siapas et al., 2005; Siapas& Wilson, 1998). The insights gained from these and many moreanimal studies along with fMRI studies in humans have beensynthesized into a hypothesis of how memory consolidation occursduring sleep (Born & Wilhelm, 2012). Thus, the concept that sleepplays an essential role in learning and memory is on very solidground. But, what role might be played by circadian rhythms?
The obvious connection between circadian rhythms and learn-ing/memory is the influence of the circadian system on sleep. Acircadian rhythm of alertness and sleep permissiveness interactingwith a wake dependent accumulation of sleep-need assures themaintenance of a daily sleep/activity pattern that corresponds tothe daily light/dark cycle in both humans and animals (Borbely &Achermann, 1992). Two potential consequences for learning andmemory experiments derive from the circadian control of sleep: (a)training during the circadian rest phase could result in sleepdeprivation that would impact learning and memory, or (b) lowerlevels of alertness during the rest phase could detract from theability to form memories when training occurs at this phase of thedaily cycle. Of course an additional possibility is that a factor witha circadian rhythm such as hormone release or gene expression ishaving an effect on learning and memory apart from sleep.
Experiments on humans and animals do not support the conceptthat memory impairment due to sleep deprivation caused by train-ing is a critical variable in studies of daily cycles of learning andmemory performance. When humans are placed on a forced de-synchrony protocol in which the circadian rhythm of body tem-perature does not remain in phase with sleep timing, the perfor-mance on memory and cognitive tasks follows the circadianrhythm of body temperature and not the sleep cycle (Johnson et al.,1992). When animals are trained and tested at different times
around the daily cycle of rest and activity, memory recall is seento have a locked phase relationship with time of training, meaningthat learning is just as effective when training occurs during thenormal sleep phase or the normal active phase (Holloway &Wansley, 1973a,b). And finally, studies of long-term potentiation(LTP) on hippocampal slices reflect daily cycles of magnitude inthe absence of sleep deprivation prior to animal sacrifice (Chaud-hury et al., 2005). It may be concluded, therefore, that the mam-malian brain undergoes a daily cycle of ability to acquire andconsolidate memories, or in other words, a daily cycle of neuralplasticity.
One view of the daily cycle of learning and memory propensityis that it is due to mechanisms promoting higher levels of alertnessduring the daily active phase. In this article we advance thealternative view that the daily cycles of low learning and memoryconsolidation ability are due to active suppression of neural plas-ticity during the daily rest phase. We propose that the mechanismof this suppression is GABAergic, and we show that abnormallyhigh levels of this GABAergic inhibition can be the cause of atleast one case of learning disability—Down’s syndrome. We re-view experiments on Siberian hamsters that indicate that the pos-sible origin of the GABAergic suppression of neural plasticity isthe circadian pacemaker, the suprachiasmatic nucleus. We con-clude by advancing the hypothesis that: (a) there is an adaptivefunction of the daily phases of suppression of neural plasticity, and(b) that function is to protect the integrity of memory traces duringthe processes of memory consolidation that occur during sleep. Werecognize that aspects of memory consolidation occur during wakeresulting in what has been referred to as intermediate-term mem-ory that lasts for minutes to hours. In this article, however, we arefocusing on the processes that result in durable long-term memorythat lasts from days to lifetimes.
Learning Ability in DS Model Mice Can be RescuedWith GABA Antagonists
Down’s syndrome (DS) in humans results from trisomy ofchromosome 21, which contains an estimated 250 genes makingDS a complex disease associated with many disorders. One featureof Down’s syndrome that seriously compromises quality of life forindividuals and families is intellectual disability. Because theincidence of DS is around 1 in 700 births, it is the leading geneticcause of learning disability. Many symptoms of DS such as hear-ing and vision disorders, thyroid and heart disorders, cancer, andgastrointestinal disorders can be medically treated resulting inincreased longevity, but no treatments exist for the intellectualdisability. Excellent mouse models of DS have been generatedthrough genetic engineering that has produced mice that are ane-uploid for genes syntenic to those on human chromosome 21. TheTs65Dn mouse is trisomic for about 150 syntenic genes and showsmany features of human DS including severe learning disabilitiesas revealed by traditional rodent learning tasks (Reeves et al.,1995).
In vitro studies of hippocampal neurons in brain slices indicatedthe possible involvement of inhibition in memory deficits inTs65Dn mice. Abnormalities in long-term potentiation (LTP) wereobserved in these preparations and were shown to involve exces-sive GABAergic inhibition (Costa & Grybko, 2005; Kle-schevnikov et al., 2004). Fabian Fernandez working with Craig
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
274 HELLER, RUBY, ROLLS, MAKAM, AND COLAS
Garner at Stanford University first tested the possibility that over-inhibition was responsible for learning disability in behavingTs65Dn mice. Three different GABAA receptor antagonists, pi-crotoxin (PTX), pentylenetetrazole (PTZ), and bilobilide, wereshown to normalize learning and memory in the Ts65Dn mice asassessed through the novel object recognition task for long-termmemory and the T maze spontaneous alternation task for workingmemory (Fernandez et al., 2007). In addition, this study alsoevaluated hippocampal LTP in Ts65Dn mice, and found it to begreatly reduced in comparison to 2N littermates. Treatment withPTZ rescued LTP in the Ts65Dn mice. A remarkable result of theFernandez et al. study was that the drug treatments were delivereddaily for 2 weeks, but the cognitive and LTP benefits were ob-served 1 week to 1 month later, long after the drugs had cleared thesystem. Thus, a short-term chronic drug treatment resulted inlong-term changes in neuroplasticity.
In recognition of the known involvements of circadian rhythmsand sleep in learning and memory, Ruby et al. (2010) investigatedwhether there were any circadian abnormalities in Ts65Dn mice.The findings were that the circadian rhythms of the Ts65Dn micewere perfectly intact, and in fact had higher periodogram scoresthan seen in the 2N littermates. The significance of these apparentnegative results were not appreciated at the time, but may reflectthe overinhibition mechanism.
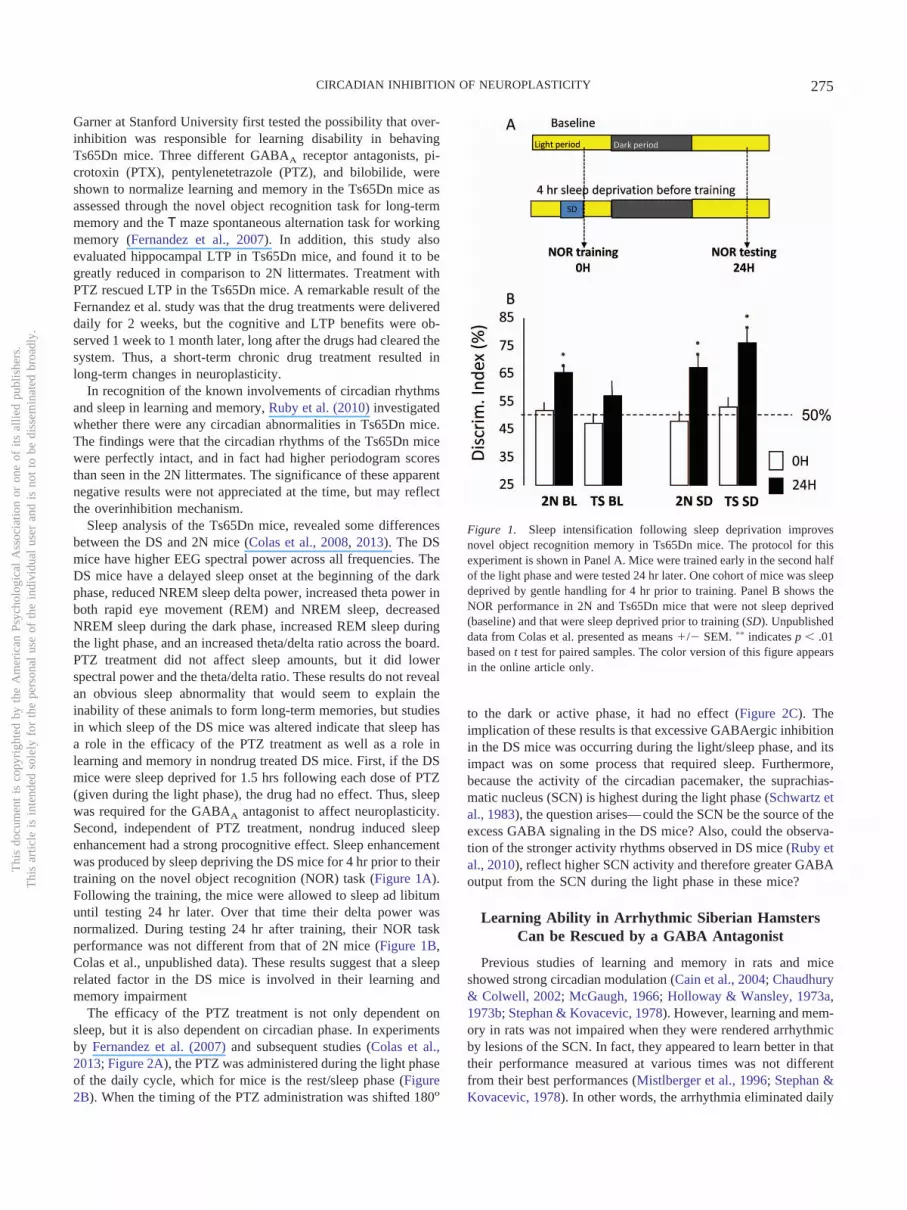
Sleep analysis of the Ts65Dn mice, revealed some differencesbetween the DS and 2N mice (Colas et al., 2008, 2013). The DSmice have higher EEG spectral power across all frequencies. TheDS mice have a delayed sleep onset at the beginning of the darkphase, reduced NREM sleep delta power, increased theta power inboth rapid eye movement (REM) and NREM sleep, decreasedNREM sleep during the dark phase, increased REM sleep duringthe light phase, and an increased theta/delta ratio across the board.PTZ treatment did not affect sleep amounts, but it did lowerspectral power and the theta/delta ratio. These results do not revealan obvious sleep abnormality that would seem to explain theinability of these animals to form long-term memories, but studiesin which sleep of the DS mice was altered indicate that sleep hasa role in the efficacy of the PTZ treatment as well as a role inlearning and memory in nondrug treated DS mice. First, if the DSmice were sleep deprived for 1.5 hrs following each dose of PTZ(given during the light phase), the drug had no effect. Thus, sleepwas required for the GABAA antagonist to affect neuroplasticity.Second, independent of PTZ treatment, nondrug induced sleepenhancement had a strong procognitive effect. Sleep enhancementwas produced by sleep depriving the DS mice for 4 hr prior to theirtraining on the novel object recognition (NOR) task (Figure 1A).Following the training, the mice were allowed to sleep ad libitumuntil testing 24 hr later. Over that time their delta power wasnormalized. During testing 24 hr after training, their NOR taskperformance was not different from that of 2N mice (Figure 1B,Colas et al., unpublished data). These results suggest that a sleeprelated factor in the DS mice is involved in their learning andmemory impairment
The efficacy of the PTZ treatment is not only dependent onsleep, but it is also dependent on circadian phase. In experimentsby Fernandez et al. (2007) and subsequent studies (Colas et al.,2013; Figure 2A), the PTZ was administered during the light phaseof the daily cycle, which for mice is the rest/sleep phase (Figure2B). When the timing of the PTZ administration was shifted 180o
to the dark or active phase, it had no effect (Figure 2C). Theimplication of these results is that excessive GABAergic inhibitionin the DS mice was occurring during the light/sleep phase, and itsimpact was on some process that required sleep. Furthermore,because the activity of the circadian pacemaker, the suprachias-matic nucleus (SCN) is highest during the light phase (Schwartz etal., 1983), the question arises—could the SCN be the source of theexcess GABA signaling in the DS mice? Also, could the observa-tion of the stronger activity rhythms observed in DS mice (Ruby etal., 2010), reflect higher SCN activity and therefore greater GABAoutput from the SCN during the light phase in these mice?
Learning Ability in Arrhythmic Siberian HamstersCan be Rescued by a GABA Antagonist
Previous studies of learning and memory in rats and miceshowed strong circadian modulation (Cain et al., 2004; Chaudhury& Colwell, 2002; McGaugh, 1966; Holloway & Wansley, 1973a,1973b; Stephan & Kovacevic, 1978). However, learning and mem-ory in rats was not impaired when they were rendered arrhythmicby lesions of the SCN. In fact, they appeared to learn better in thattheir performance measured at various times was not differentfrom their best performances (Mistlberger et al., 1996; Stephan &Kovacevic, 1978). In other words, the arrhythmia eliminated daily
Figure 1. Sleep intensification following sleep deprivation improvesnovel object recognition memory in Ts65Dn mice. The protocol for thisexperiment is shown in Panel A. Mice were trained early in the second halfof the light phase and were tested 24 hr later. One cohort of mice was sleepdeprived by gentle handling for 4 hr prior to training. Panel B shows theNOR performance in 2N and Ts65Dn mice that were not sleep deprived(baseline) and that were sleep deprived prior to training (SD). Unpublisheddata from Colas et al. presented as means �/� SEM. �� indicates p � .01based on t test for paired samples. The color version of this figure appearsin the online article only.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
275CIRCADIAN INHIBITION OF NEUROPLASTICITY

nadirs in performance scores. New insights into the role of thecircadian system were made possible by the development of a newmodel of circadian arrhythmia in the Siberian hamster (Ruby et al.,1996). This model is unique because it does not involve aninvasive, genetic, or pharmacological manipulation; it only in-volves a single exposure to an extra 2 hr of light late at nightfollowed by a 3-hr delay in the onset of the dark phase on the nextday. This combination of phase-advancing and phase-delayinglight signals has been termed a circadian disruptive phase shift(DPS) by Prendergast, Cisse, Cable, and Zucker (2012). The
majority of Siberian hamsters respond to the DPS by becomingcircadian-arrhythmic for the rest of their natural lives even thoughthey continue to live under a 24-hr light–dark cycle. Due to thefact that the DPS treatment does not involve brain damage (lesion)or a genetic alteration (e.g., gene knock out) that could havedevelopmental or other effects, it was seen as an ideal model forasking questions about the functional significance of circadianrhythms including their role in learning and memory.
In contrast to the earlier results on SCN lesioned, arrhythmicrats, the hamsters made arrhythmic by the DPS were severelylearning disabled. Whereas they performed normally in the NORtest when the training to testing interval was 20 min or less, theywere unable to form long-term memories of objects if the trainingto testing interval was 60 min or 24 hr (Ruby et al., 2008). Spatialworking memory also was impaired as revealed by the spontane-ous alternation in the T maze task (Ruby et al., 2013; Figure 3,vehicle-treated animals). What could possibly account for theseopposite results on the cognitive abilities of arrhythmic rats andarrhythmic hamsters?
The arrhythmic rats had no SCN and the arrhythmic hamstershad an intact SCN that was active, but not rhythmic. The activityof the SCNs of arrhythmic hamsters has been confirmed by the factthat the expression of clock genes continued in the SCNs of theseanimals, but that expression was arrhythmic (Grone et al., 2011).Furthermore, firing rates of SCN neurons are also arrhythmic inthese animals, and maintain an average daily firing rate that doesnot differ from normal rhythmic hamsters (Margraf et al., 1992).Thus, it is reasonable to expect that some output of the SCNcontinued in the arrhythmic hamsters, but that output did not cycle.Because, as mentioned above, the SCN is a GABAergic nucleus,could the constant output of GABA in the arrhythmic hamsterscreate the same problem of overinhibition as seen in the Ts65Dnmice?
Arrhythmic hamsters were treated with a 10-day regimen ofdaily IP injections of the GABAA receptor antagonist PTZ (1mg/kg). The day after the PTZ injections ended, these animalswere tested on the NOR and SA tasks, and they performed nor-mally with no apparent learning and memory disability (Ruby etal., 2013; Figure 3, PTZ-treated animals). As with the Ts65Dnmice, the performance of the PTZ treated hamsters on the memorytests was still normal when the animals were tested 1 month later.These results support, but do not prove, the hypothesis that in thearrhythmic hamsters, continuous GABAergic output from theirintact SCNs impairs learning and memory. The implication of thishypothesis is that in the intact animals, there is one particular phaseof the daily rhythm when the SCN is actively inhibiting neuralplasticity. What could be the adaptive significance of rhythmic,daily suppression of neural plasticity?
Long-Term Memory Consolidation Occurs During theSleep Phase of the Daily Rhythm
As discussed earlier, sleep is critical for memory consolidation.What aspect of sleep is critical for this function? We hypothesized thatcontinuity of NREM sleep was the critical factor, so we conducted anexperiment on mice that were prepared for nonstressful optogeneticdisruption of sleep continuity (Rolls et al., 2011). These mice receivedsurgically implanted cannulas into their lateral hypothalami as well asEEG/EMG recording electrodes. Lentivirus containing constructs of
Figure 2. PTZ improves novel object recognition memory in Ts65Dnmice only if the animals are dosed during the light phase of the daily cycle.(A) Treatment protocol was 2 weeks of daily IP injections of PTZ at 0.3mg/kg or of saline. At least 1 week was allowed between end of treatmentand testing. (B) NOR test results for 2- to 3-month-old mice treated withPTZ (0.3 mg/kg) or saline. Two objects were used, and discriminationindex is the time spent with the novel object divided by time spent withnovel and familiar objects. Gray bars are training and black bars are novelrecognition results 24 hr later. (C) NOR test results from mice receivingPTZ treatment (0.3 mg/kg) or saline during the dark phase. Data fromColas et al., 2013 presented as means �/� SEM. � p � .05. �� p � .01based on t test for paired samples.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
276 HELLER, RUBY, ROLLS, MAKAM, AND COLAS

the hypocretin promotor, light sensitive Channelrhodopsin-2 (ChR2)cation channel, and mCherry red fluorescent protein were injectedthrough the cannulas. Constructs injected into control mice did notinclude the ChR2 sequence. Prior to experiments, optical fibers wereinserted through the cannulas so they resided just above the lateralhypothalami. Experiments consisted of training mice in the NOR taskwith testing 24 hr later. However, for 4 hr immediately following theNOR training, the mice were either: undisturbed, totally sleep de-prived by gentle handling, or had their sleep fragmented by optoge-netic stimulation of the hypocretinergic (Hct) cells in the lateralhypothalamus by pulses of blue light at 30-, 60-, 120-, or 240-sintervals. Mice were allowed to sleep ad lib. for the remaining 20 hrsbefore testing.
Analysis of the EEG/EMG recordings revealed the effects ofthe optogenetic stimulation on sleep and on learning and mem-ory. Except for the sleep deprivation group, the groups did notdiffer in their total amounts of REM or NREM sleep. However,the distribution of NREM bouts by duration showed that theChR2 mice receiving optogenetic stimulation had more shortbouts and fewer long bouts of NREM sleep than did the control
or nonsleep deprived mice. Performance on the NOR test wasnormal in the control, nonsleep deprived mice. As expected, themice that were sleep deprived for 4 hr showed no discriminationbetween the two test objects. Performance differed in thegroups receiving optogenetic stimulation of the Hct neurons.Mice whose sleep was fragmented at 30-s or 60-s intervalsshowed no long-term memory of the NOR training. But, micewhose sleep was fragmented at 120-s or 240-s intervals showednormal long-term memory of the NOR training (see Figure 4).Our conclusion was that there was a minimal quantum ofcontinuous NREM sleep that was essential for consolidation ofthe NOR training memory into long-term NOR memory.
A circadian influence on this process of memory consolidationduring sleep was revealed by conducting the same experiment 12hr out of phase with the initial experiment. The NOR training inthe initial experiment was at the beginning of the light phase—thesleep phase for the mice. In this second experiment, the NORtraining was at the beginning of the dark phase—the active phasefor the mice. Following training, the different groups were treatedas in the previous experiment. NOR testing was conducted 24 hr
Figure 3. PTZ rescued memory in circadian-arrhythmic hamsters. The loss of circadian rhythms impairedspatial working memory as assessed by alternation behavior in a T maze (left panel). The ability to discriminatea familiar object from a novel one was impaired in a novel object recognition test (right panel). Test performanceof arrhythmic hamsters (ARR) after a 10-day injection regimen of PTZ was restored to levels observed in normalentrained animals (ENT). PTZ had no effect in ENT hamsters. �� p � .01 compared with random chanceperformance. Data from Ruby et al., 2013.
Figure 4. Effect of sleep fragmentation on novel object recognition memory. Mice were trained in the NORtask early in the light phase. Then for 4 hr they were left undisturbed, were totally sleep deprived, or weresubjected to different protocols of sleep fragmentation administered by brief trains of optogenetic stimulation ofhypocretinergic cells at 30-, 60-, 120-, or 240-s intervals. For the remaining 20 hr they were allowed to sleep adlibitum until testing 24 hr after training. Significant NOR memory was seen only in mice that slept ad libitumfor 24 hr or had their early sleep fragmented at 120- or 240-s intervals. Data from Rolls et al., 2011 are presentedas means �/� SEM. ��� p � .001.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
277CIRCADIAN INHIBITION OF NEUROPLASTICITY

later also during the dark phase. The results showed no deficits inNOR long-term memory due to any of the optogenetic stimulationregimes. What was the difference between this experiment and theinitial one? In this experiment, the mice had undisturbed sleepduring the normal daily sleep phase following the NOR training,even though that training was 12 hr earlier at the beginning of theactive phase. These results suggest that not just sleep, but sleepduring the proper daily or circadian phase is essential for memoryconsolidation. That daily phase for the mice is the light phase, andthis is the phase when the activity of the SCN is high. So, if theoutput of an active SCN is suppressing neural plasticity during thesleep phase of these mice, what could be its adaptive significance?Why should it be adaptive to suppress the ability to consolidatenew memories?
Memories are Vulnerable to Change During theProcess of Memory Consolidation During Sleep
The process of memory consolidation involves communicationbetween different brain regions during sleep, and that communi-cation involves several characteristic patterns of electrophysiolog-ical activity (Born & Wilhelm, 2012). These patterns have beendescribed most extensively in association with spatial learning inrats. Areas of the brain definitely involved in the formation ofspatial memories are the hippocampus and the cortex. The closeassociation of activities in these two brain areas is demonstrated bysimultaneous recordings of local field potentials (LFPs). Duringexploration of a new environment or a novel object, the EEG isdominated by theta oscillations (4 Hz–10 Hz). Recordings from thehippocampus and the prefrontal cortex show that the theta oscil-lations in these two areas are phase locked with the hippocampalactivity preceding the cortical activity by about 50 ms indicating adirectionality of information flow (Jones & Wilson, 2005; Siapaset al., 2005). What is that information and how is it coded?
Recordings of single units show that the initial experience inrunning a maze is coded in the sequences of firing of hippocampalneurons (Wilson & McNaughton, 1993, 1994). These sequencesare coordinated with the theta oscillation (Siapas et al., 2005).During subsequent NREM sleep these sequences are replayed on afaster (about 20x) time scale (Lee & Wilson, 2002). These com-pressed sequences coincide with fast (about 200 Hz) LFP oscilla-tions called sharp-wave ripples. The sharp-wave ripples with theirassociated compressed sequences of single cell firing can also beobserved when the animal is resting. And, they can reflect bothforward and reverse replay of the original experience (Carr et al.,2011). Selective suppression of these sharp-wave ripples duringsleep following a spatial learning task impairs the subsequentmemory of the animal for that task (Girardeau et al., 2009). Theseresults support the concept that the information encoded duringtraining is contained in the neural activity associated with thesharp-wave ripples.
During NREM sleep, rapid sequences of single cell firing in theprefrontal cortex correlate closely with the fast sequences recordedin the hippocampus (Ji and Wilson, 2007; Peyrache, Khamassi,Benchenane, Wiener, & Battaglia, 2009). In the cortex, thesesequences are also expressed in association with sharp-wave rip-ples, and these sharp-wave ripples occur in phase with corticalspindle oscillations that originate in the thalamus (Siapas & Wil-son, 1998). Replays of these unique sequences observed in the
cortex occur in frames that are set off by the slow rhythm (1 Hz–4Hz) that characterizes NREM sleep (Molle, Bergmann, Marshall,& Born, 2011; Molle et al., 2006). Extended replays may involvemultiple bursts of sharp-wave ripples that extend over multipleslow wave frames (Davidson, Kloosterman, & Wilson, 2009).These results and others show that reactivations of memoriesduring sleep occur in a coordinated fashion in a brain networkincluding the cortex, the thalamus, and the hippocampus (Peyracheet al., 2011). The ensemble of these events are believed to beresponsible for the reactivation-related improvement of memoryconsolidation.
The interpretation of these and other elegant electrophysiolog-ical studies of spatial memory processing is that the experience iscoded in the hippocampus and the cortex, but consolidation ofthose relatively ephemeral memory traces into long-term storageinvolves communication between the hippocampus and the cortexfacilitated by the various brain electrical rhythms that have beenobserved: sharp-wave ripples, theta waves, spindle waves, andslow waves. One model of this memory consolidation systemcharacterizes the hippocampus as a rapid but short-term learningsystem and the cortex as a slow but durable learning system, andthat the process of memory consolidation involves an interactionbetween these two systems with the rapid learning system rein-forcing (or tutoring) the slow learning system (Born & Wilhelm,2012). These repeated and highly coordinated conversations be-tween cell assemblies in the hippocampus and in the cortex occurduring sleep. Our purpose in briefly reviewing this information onthe electrophysiology of memory consolidation is to suggest that itis important to stabilize memory traces while these processes aretaking place. High levels of neuroplasticity that characterize thewaking brain could jeopardize the fidelity of the memory beingconsolidated.
Are memory traces vulnerable to alteration when they are beingconsolidated during NREM sleep? Several recent studies in bothhumans and mice have shown that memories can be modifiedduring sleep. Two studies on humans showed that consolidation ofa specific memory can be strengthened during sleep by introducinga sensory stimulus during the learning experience and then replay-ing that stimulus during subsequent sleep. In one study subjectswere trained to locate card pairs on a computer screen grid. Theywere then tested following sleep by presenting one of each pair andasking the subject to indicate where the matching card shouldappear (Rasch et al., 2007). In some training sessions the subjectswere subjected to the scent of roses. The rose scent was thenreintroduced to the subjects during sleep. The results were thatlearning was significantly enhanced when the scent present duringtraining was reintroduced during NREM sleep. The scent aloneduring sleep had no effect if it had not been present duringlearning. The vehicle had no effect. And, the scent had no effect ifit were introduced during REM sleep. This study clearly showedthat memories could be modified (in this case strengthened) duringsleep.
A similar study using auditory stimuli showed that the sensorystimulation during sleep was specific to individual memories andnot just to the overall process of memory consolidation. Rudoy,Voss, Westerberg, and Paller (2009) trained subjects to associate50 images with their placement on a computer screen grid. Eachimage was paired with an appropriate sound—cat, meow; bell,ding; and so forth. The subjects were exposed to half of these
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
278 HELLER, RUBY, ROLLS, MAKAM, AND COLAS

sounds during subsequent NREM sleep. Positional recall duringsubsequent testing was significantly greater for the images thatwere recued by their sounds during sleep.
A study on mice demonstrated that the emotional valence of amemory could be strengthened or weakened during sleep (Rolls etal., 2013). Mice were exposed to foot shock that was paired withan odor as a conditioned stimulus (CS). When that odor wasreintroduced to the mice during sleep immediately following thefoot-shock conditioning or even during the sleep phase of thefollowing day, it did not disturb their sleep, but when the animalswere reexposed to the CS during subsequent wakefulness, theyresponded much more vigorously as measured by behavioralfreezing (Figure 5a). The fear memory was clearly enhanced. Incontrast, if the mice received bilateral microinjections of ani-somycin (a protein synthesis inhibitor) into the amygdala justprior to sleep, and the CS was introduced during sleep, thestrength of the fear memory during subsequent wakefulness wasreduced as evidenced by decreased freezing upon reexposure tothe CS (Figure 5b). These results show that the emotionalvalence of memories can be altered during sleep. The hypoth-esis is that the reintroduction of the CS reactivates the memorythat then must be reconsolidated, and through this process, theemotional valence of the memory can be either enhanced orreduced.
Additional experiments (Makam et al., unpublished data) dem-onstrated a fundamental difference of reactivating the memoryduring wake or during sleep. When the foot-shocked mice wererepeatedly reexposed to the CS during wake in a neutral environ-ment (not the environment in which the shocks had been experi-
enced), there was gradual extinction of the freezing response aswould be expected. That extinction was in contrast to the enhance-ment of the freezing response when the mice were reexposed to theCS during sleep (Figure 5a). The interpretation of these results wasthat experiencing the CS during wake resulted in the coding of anew memory that associated the CS with a safe environment. Thisis in contrast to reexperiencing the CS during sleep which did notresult in coding a new association, and therefore the emotionalvalence of the original memory was enhanced through the repeatedreactivation/reconsolidation process. When the anisomycin waspresent, it interfered with that reconsolidation process and the fearmemory was diminished. In another set of trials, the anisomycintreated mice were only reexposed to the CS when they werespontaneously awake. This treatment had no effect on the subse-quent fear response indicating that it interfered with the coding ofa new memory associating the CS with a safe environment. Over-all, these experiments on reactivating fear memories during wakeand sleep support the idea that the memories are coded duringwake, and long-term consolidation occurs during sleep, and ifreactivation and reconsolidation occurs during sleep, the memorytrace can be altered.
A dramatic demonstration of the vulnerability of memories wasa recent demonstration of the creation of a false memory (Ramirezet al., 2013). In this study, the optogenetic agent ChR2 wasintroduced into active hippocampal cells of mice while the animalswere in a particular environment (neutral context). They were thenfoot shocked in a different environment, which became a fearcontext. But, if the neurons that had incorporated ChR2 in theneutral context were activated by blue light during the foot-shock
Figure 5. The effects of introducing the Conditioned Stimulus from a foot-shock conditioning paradigm duringsubsequent sleep. (A) Mice received foot shocks paired with an odor (the CS) at the beginning of the light phase.During the sleep phase beginning 24 hr later the mice were reexposed to the CS or to a control odor duringepisodes of NREM sleep. They were then tested 48 hr after conditioning in a neutral context by reintroductionof the CS or the control odor and measurements of time spent freezing were recorded. The reexposure to the CSduring sleep intensified the subsequent fear response. (B) The experiment was repeated with a higher stimulusintensity, but the mice received bilateral injections of anisomycin or vehicle into the amygdala prior to the sleepphase. Responses to the CS in a neutral context were tested 48 hr following the conditioning. Animals thatreceived the anisomycin and were reexposed to the CS during sleep showed a reduction in their subsequent fearresponse to the CS. Data from Rolls et al., 2013.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
279CIRCADIAN INHIBITION OF NEUROPLASTICITY

experience, the mice showed increased fear (freezing) when reex-posed to the neutral context. The interpretation was that the opto-genetic stimulation reactivated the memory of the neutral context,and because that reactivation and subsequent reconsolidation waspaired with the foot shock, the memory took on a new emotionalvalence. Although this experiment did not involve sleep recording,it is reasonable to expect that the new fear memory was consoli-dated during subsequent sleep.
Conclusion: A Function of the SCN May be toStabilize Memory Traces as They are BeingConsolidated During the Daily Sleep Phase
The evidence reviewed in this article leads us to propose that thedaily phases of low learning and memory performance are notsimply due to an ebb of brain alerting mechanisms, but rather theycould be due to episodes of active suppression of neural plasticitythrough GABAergic inhibitory mechanisms. Overactivity of thesemechanisms can lead to pathological learning disabilities as occurin Down’s syndrome and perhaps also in Alzheimer’s disease.Pharmacological suppression of GABAergic inhibition restoreslearning and memory in mouse models of Down’s syndrome andAD (Braudeau et al., 2011; Colas et al., 2013; Fernandez et al.,2007; Rueda et al., 2008; Yoshike et al., 2008). But, this GABAe-rgic suppression only works when administered during the phaseof the circadian cycle when the SCN is most active, pointing to theSCN as a critical player in the daily suppression of neural plastic-ity. Studies in which the SCN of rats were eliminated by electro-lytic lesions produced no impairments of learning and memory, butrather resulted in performance at maximal levels at all times of day(Stephan & Kovacevic, 1978). Experiments on Siberian hamstersproduced a very interesting contrast to the SCN lesion experi-ments. The fact that Siberian hamsters can be rendered arrhythmicby a DPS made them an attractive model for studying the effect ofcircadian arrhythmia on learning and memory. When these animalsare rendered arrhythmic without destroying their SCN, they showsevere learning and memory disability (Ruby et al., 2008). How-ever, that disability can be reversed with pharmacological treat-ments that reduce GABAergic inhibition (Ruby et al., 2013).Taken together, these various results indicate an active role of theSCN in suppressing neural plasticity. What adaptive functioncould such suppression serve?
We have summarized some of the extensive evidence showingthat memory consolidation occurs during sleep. We also show thatspecific memories can be reactivated during sleep and modified byvarious treatments (Rasch et al., 2007; Rolls et al., 2013; Rudoy,Voss, Westerberg, & Paller, 2009). In conclusion, we propose thata function of the circadian system, and specifically the SCN inmammals, is to suppress neural plasticity during the sleep phase ofthe daily cycle when memory traces coded during the precedingactive phase are being reactivated and consolidated into long-termstorage. We suggest that the function of this suppression of neuralplasticity is to protect the fidelity of the memory traces that arebeing consolidated.
Our proposal that the circadian system stabilizes memory tracesduring the process of consolidation is not intended to suggest thatall forms of neural plasticity are suppressed during sleep. Early inthis article we referred to the elegant studies of Frank et al., 2001demonstrating the sleep dependency of the physical processes of
neuronal remodeling underlying plasticity of the visual system ofmonocularly deprived kittens. At another level, many studies inhumans have shown that information acquired during wake isprocessed and integrated during sleep resulting in extraction of gistand insight (e.g., Diekelmann, Born, & Wagner, 2010; Ellenbogenet al., 2007; Wagner et al., 2004) and reviewed in detail by Bornand Wilhelm (2012). Neural plasticity takes on many forms andplays numerous roles in the nervous system. At present we canonly suggest that the circadian suppression of neural plasticitydiscussed in this article applies specifically to the exchanges ofinformation between the hippocampus and the cortex that areessential for memory consolidation during sleep.
References
Benington, J., & Heller, H. C. (1995). Restoration of brain energy metab-olism as the function of sleep. Progress in Neurobiology, 45, 347–360.doi:10.1016/0301-0082(94)00057-O
Borbely, A., & Achermann, P. (1992). Concepts and models of sleepregulation: An overview. Journal of Sleep Research, 1, 63–79. doi:10.1111/j.1365-2869.1992.tb00013.x
Born, J., & Wilhelm, I. (2012). System consolidation of memory duringsleep. Psychological Research, 76, 192–203. doi:10.1007/s00426-011-0335-6
Braudeau, J., Delatour, B., Duchon, A., Pereira, P. L., Dauphinot, L., deChaumont, F., . . . Potier, M. C. (2011). Specific targeting of theGABA-A receptor �5 subtype by a selective inverse agonist restorescognitive deficits in Down syndrome mice. Journal of Psychopharma-cology, 25, 1030–1042. doi:10.1177/0269881111405366
Cain, S. W., Chou, T., & Ralph, M. R. (2004). Circadian modulation ofperformance on an aversion-based place learning task in hamsters.Behavioural Brain Research, 150, 201–205. doi:10.1016/j.bbr.2003.07.001
Carr, M. F., Jadhav, S., & Frank, L. M. (2011). Hippocampal replay in theawake state: A potential substrate for memory consolidation and re-trieval. Nature Neuroscience, 14, 147–162. doi:10.1038/nn.2732
Chaudhury, D., & Colwell, C. S. (2002). Circadian modulation of learningand memory in fear-conditioned mice. Behavioural Brain Research,133, 95–108. doi:10.1016/S0166-4328(01)00471-5
Chaudhury, D., Wang, L. M., & Colwell, C. S. (2005). Circadian regulationof hippocampal long-term potentiation. Journal of Biological Rhythms,20, 225–236. doi:10.1177/0748730405276352
Colas, D., Chuluun, B., Warrier, D., Blank, M., Wetmore, D. Z., Buck-master, P., . . . Heller, H. C. (2013). Short-term treatment with theGABAA antagonist pentylenetetrazole produces a sustained procognitivebenefit in a mouse model of Down’s syndrome. British Journal ofPharmacology, 169, 963–973. doi:10.1111/bph.12169
Colas, D., Valletta, J. S., Takimoto-Kimura, R., Nishino, S., Fujiki, N.,Mobley, W. C., & Mignot, E. (2008). Sleep and EEG features in geneticmodels of Down syndrome. Neurobiology of Disease, 30, 1–7. doi:10.1016/j.nbd.2007.07.014
Costa, A. C. S., & Grybko, M. J. (2005). Deficits in hippocampal CA1 LTPinduced by TBS but not HFS in the Ts65Dn mouse: A model of Downsyndrome. Neuroscience Letters, 382, 317–322. doi:10.1016/j.neulet.2005.03.031
Crick, F., & Mitchison, G. (1983). The function of dream sleep. Nature,304, 111–114. doi:10.1038/304111a0
Davidson, T. J., Kloosterman, F., & Wilson, M. A. (2009). Hippocampalreplay of extended experience. Neuron, 63, 497–507. doi:10.1016/j.neuron.2009.07.027
Diekelmann, S., & Born, J. (2010). The memory function of sleep. NatureReviews Neuroscience, 11, 113–126.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
280 HELLER, RUBY, ROLLS, MAKAM, AND COLAS

Diekelmann, S., Born, J., & Wagner, U. (2010). Sleep enhances falsememories depending on general memory performance. Behavioral BrainResearch, 208, 425–429. doi:10.1016/j.bbr.2009.12.021
Ellenbogen, J. M., Hu, P. T., Payne, J. D., Titone, D., & Walker, M. P.(2007). Human relational memory requires time and sleep. Proceedingsof the National Academy of Sciences of the United States of America,104, 7723–7728.
Fernandez, F., Morishita, W., Zuniga, E., Nguyen, J., Blank, M., Malenka,R. C., & Garner, C. C. (2007). Pharmacotherapy for cognitive impair-ment in a mouse model of Down syndrome. Nature Neuroscience, 10,411–413.
Frank, M. G., Issa, N. P., & Stryker, M. P. (2001). Sleep enhancesplasticity in the developing visual cortex. Neuron, 30, 275–287. doi:10.1016/S0896-6273(01)00279-3
Girardeau, G., Benchenane, K., Wiener, S. I., Buzsaki, G., & Zugaro, M. B.(2009). Selective suppression of hippocampal ripples impairs spatialmemory. Nature Neuroscience, 12, 1222–1223. doi:10.1038/nn.2384
Grone, B. P., Chang, D., Bourgin, P., Cao, V., Fernald, R. D., Heller, H. C.,& Ruby, N. F. (2011). Acute light exposure suppresses circadianrhythms in clock gene expression. Journal of Biological Rhythms, 26,78–81. doi:10.1177/0748730410388404
Holloway, F. A., & Wansley, R. A. (1973a). Multiple retention deficits atperiodic intervals after active and passive avoidance learning. Behav-ioral Biology, 9, 1–14. doi:10.1016/S0091-6773(73)80164-6
Holloway, F. A., & Wansley, R. A. (1973b). Multiple retention deficits atperiodic intervals after passive-avoidance learning. Science, 180, 208–210. doi:10.1126/science.180.4082.208
Horne, J. (1988). Why we sleep: The functions of sleep in humans and othermammals. New York, NY: Oxford University Press.
Huber, R., Ghilardi, M. F., Massimini, M., & Tononi, G. (2004). Localsleep and learning. Nature, 430, 78–81. doi:10.1038/nature02663
Ji, D., & Wilson, M. A. (2007). Coordinated memory replay in the visualcortex and hippocampus during sleep. Nature Neuroscience, 10, 100–107. doi:10.1038/nn1825
Johnson, M. P., Duffy, J. F., Dijk, D. J., Ronda, J. M., Dyal, D. M., &Czeisler, C. A. (1992). Short-term memory, alertness and performance:A reappraisal of their relationship to body temperature. Journal of SleepResearch, 1, 24–29. doi:10.1111/j.1365-2869.1992.tb00004.x
Jones, M. W., & Wilson, M. A. (2005). Theta rhythms coordinatehippocampal-prefrontal interactions in a spatial memory task. PLoSBiology, 3, e402. doi:10.1371/journal.pbio.0030402
Jouvet, M. (1998). Pardoxical sleep as a programming system. JournalSleep Research, 7, 1–5. doi:10.1046/j.1365-2869.7.s1.1.x
Kleschevnikov, A. M., Belichenko, P. V., Villar, A. J., Epstein, C. J.,Malenka, R. C., & Mobley, W. C. (2004). Hippocampal long-termpotentiation suppressed by increased inhibition in the Ts65Dn mouse, agenetic model of Down syndrome. The Journal of Neuroscience, 24,8153–8160. doi:10.1523/JNEUROSCI.1766-04.2004
Kreuger, J., & Obal, F. (1993). A neuronal group theory of sleep function.Journal of Sleep Research, 2, 63–69. doi:10.1111/j.1365-2869.1993.tb00064.x
Lee, A. K., & Wilson, M. A. (2002). Memory of sequential experience inthe hippocampus during slow wave sleep. Neuron, 36, 1183–1194.doi:10.1016/S0896-6273(02)01096-6
Maquet, P. (2001). The role of sleep in learning and memory. Science, 294,1048–1052. doi:10.1126/science.1062856
Margraf, R. R., Puchalski, W., & Lynch, G. R. (1992). Absence of a dailyneuronal rhythm in the suprachiasmatic nuclei of acircadian Djungarianhamsters. Neuroscience Letters, 142, 175–178. doi:10.1016/0304-3940(92)90367-G
Marshall, L., Helgadottir, H., Molle, M., & Born, J. (2006). Boosting slowoscillations during sleep potentiates memory. Nature, 444, 610–613.doi:10.1038/nature05278
McGaugh, J. L. (1966). Time-dependent processes in memory storage.Science, 153, 1351–1358.
McGinty, D., & Szymusiak, R. (1990). Keeping cool: A hypothesis aboutthe mechanisms and functions of slow-wave sleep. Trends in Neurosci-ence, 13, 480–487. doi:10.1016/0166-2236(90)90081-K
Mednick, S., Nakayama, K., & Stickgold, R. (2003). Sleep-dependentlearning: A nap is as good as a night. Nature Neuroscience, 6, 697–698.doi:10.1038/nn1078
Mistlberger, R. E., de Groot, M. H. M., Bossert, J. M., & Marchant, E. G.(1996). Discrimination of circadian phase in intact and suprachiasmaticnuclei-ablated rats. Brain Research, 739, 12–18. doi:10.1016/S0006-8993(96)00466-0
Molle, M., Bergmann, T. O., Marshall, L., & Born, J. (2011). Fast and slowspindles during the sleep slow oscillation: Disparate coalescence andengagement in memory processing. Sleep, 34, 1411–1421.
Molle, M., Yeshenkoo, O., Marshall, L., Sara, S. J., & Born, J. (2006).Hippocampal sharp wave-ripples linked to slow oscillations in rat slow-wave sleep. Journal of Neurophysiology, 96, 62–70. doi:10.1152/jn.00014.2006
Peyrache, A., Battaglia, F. P., & Destexhe, A. (2011). Inhibition recruit-ment in prefrontal cortex during sleep spindles and gating of hippocam-pal inputs. Proceedings of the National Academy of Sciences of theUnited States of America, 108, 17207–17212. doi:10.1073/pnas.1103612108
Peyrache, A., Khamassi, M., Benchenane, K., Wiener, S. I., & Battaglia,F. P. (2009). Replay of rule-learning related neural patterns in theprefrontal cortex during sleep. Nature Neuroscience, 12, 919–926. doi:10.1038/nn.2337
Prendergast, B. J., Cisse, Y. M., Cable, E. J., & Zucker, I. (2012).Dissociation of ultradian and circadian phenotypes in female and maleSiberian hamsters. Journal of Biological Rhythms, 27, 287–298. doi:10.1177/0748730412448618
Ramadan, W., Eschenko, O., & Sara, S. J. (2009). Hippocampal sharpwave/ripples during sleep for consolidation of associative memory.PLoS One, 4, e6697. doi:10.1371/journal.pone.0006697
Ramirez, S., Liu, X., Lin, R., Suh, J., Pignatelli, M., Redondo, R., . . .Tonegawa, S. (2013). Creating a false memory in the hippocampus.Science, 341, 387–391. doi:10.1126/science.1239073
Rasch, B., Buchel, C., Gais, S., & Born, J. (2007). Odor cues duringslow-wave sleep prompt declarative memory consolidation. Science,315, 1426–1429. doi:10.1126/science.1138581
Reeves, R. H., Irving, N. G., Moran, T. H., Wohn, A., Kitt, C., Sisodia,S. S., . . . Davisson, M. T. (1995). A mouse model for Down syndromeexhibits learning and behaviour deficits. Nature Genetics, 11, 177–184.doi:10.1038/ng1095-177
Rolls, A., Colas, D., Adamantidis, A., Carter, M., Lanre-Amos, T., Heller,H. C., & de Lecea, L. (2011). Optogenetic disruption of sleep continuityimpairs memory consolidation. PNAS, 108, 13305–13310. doi:10.1073/pnas.1015633108
Rolls, A., Makam, M., Kroeger, D., Colas, D., de Lecea, L., & Heller, H. C.(2013). Sleep to forget: Interference of fear memories during sleep.Molecular Psychiatry, 18, 1166–1170. doi:10.1038/mp.2013.121
Ruby, N. F. A., Saran, T., Kang, P., Franken, P., & Heller, H. C. (1996).Siberian hamsters free run or become arrhythmic after a phase delay ofthe photocycle. American Journal of Physiology, 271, R881–R890.
Ruby, N. F., Hwang, C. E., Wessells, C., Fernandez, F., Zhang, P.,Sapolsky, R., & Heller, H. C. (2008). Hippocampal-dependent learningrequires a functional circadian system. Proceedings of the NationalAcademy of Sciences of the United States of America, 105, 15593–15598. doi:10.1073/pnas.0808259105
Ruby, N. F., Fernandez, F., Garrett, A., Klima, J., Zhang, P., Garrett, A.,. . . Heller, H. C. (2013). Spatial memory and object recognition areimpaired by circadian arrhythmia and restored by the GABAA antagonist
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
281CIRCADIAN INHIBITION OF NEUROPLASTICITY

pentylenetetrazole. PLoS ONE, 8, e72433. doi:10.1371/journal.pone.0072433
Ruby, N. F., Fernandez, F., Zhang, P., Klima, J., Heller, H. C., & Garner,C. C. (2010). Circadian locomotor rhythms are normal in Ts65Dn“Down syndrome” mice and unaffected by pentylenetetrazole. Journalof Biological Rhythms, 25, 63–66. doi:10.1177/0748730409356202
Rudoy, J. D., Voss, J. L., Westerberg, C. E., & Paller, K. A. (2009).Strengthening individual memories by reactivating them during sleep.Science, 326, 1079. doi:10.1126/science.1179013
Rueda, N., Flórez, J., & Martínez-Cué, C. (2008). Chronic pentylenetetra-zole but not donepezil treatment rescues spatial cognition in Ts65Dnmice, a model for Down syndrome. Neuroscience Letters, 433, 22–27.doi:10.1016/j.neulet.2007.12.039
Schwartz, W. J., Reppert, S. M., Eagan, S. M., & Moore-Ede, M. C.(1983). In vivo metabolic activity of the suprachiasmatic nuclei: Acomparative study. Brain Research, 274, 184–187. doi:10.1016/0006-8993(83)90538-3
Siapas, A. G., Lubenov, E., & Wilson, M. A. (2005). Prefrontal phaselocking to hippocampal theta oscillations. Neuron, 46, 141–151. doi:10.1016/j.neuron.2005.02.028
Siapas, A. G., & Wilson, M. A. (1998). Coordinated interactions betweenhippocampal ripples and cortical spindles during slow-wave sleep. Neu-ron, 21, 1123–1128. doi:10.1016/S0896-6273(00)80629-7
Stephan, F. K., & Kovacevic, N. S. (1978). Multiple retention deficit inpassive avoidance in rats is eliminated by suprachiasmatic lesions.Behavioral Biology, 22, 456–462. doi:10.1016/S0091-6773(78)92565-8
Stickgold, R. (2005). Sleep-dependent memory consolidation. Nature, 437,1272–1278. doi:10.1038/nature04286
Stickgold, R., James, L. T., & Hobson, J. A. (2000). Visual discriminationlearning requires sleep after training. Nature Neuroscience, 3, 1237–1238. doi:10.1038/81756
Tononi, G., & Cirelli, C. (2003). Sleep and synaptic homeostasis: Ahypothesis. Brain Research Bulletin, 62, 143–150. doi:10.1016/j.brainresbull.2003.09.004
Wagner, U., Gais, S., Haider, H., Verleger, R., & Born, J. (2004). Sleepinspires insight. Nature, 427, 352–355. doi:10.1038/nature02223
Walker, M. P., & Stickgold, R. (2004). Sleep-dependent learning andmemory consolidation. Neuron, 44, 121–133. doi:10.1016/j.neuron.2004.08.031
Wehr, T. (1992). A brain-warming function for REM sleep. Neuroscience Biobe-havioral Reviews, 16, 379–397. doi:10.1016/S0149-7634(05)80208-8
Wilson, M. A., & McNaughton, B. L. (1993). Dynamics of the hippocam-pal ensemble code for space. Science, 261, 1055–1058. doi:10.1126/science.8351520
Wilson, M. A., & McNaughton, B. (1994). Reactivation of hippocampalensemble memories during sleep. Science, 265, 676–679. doi:10.1126/science.8036517
Yoshike, Y., Kimura, T., Yamashita, S., Furudate, H., Mizoroki, T., Mu-rayama, M., & Takashima, A. (2008). GABAA receptor-mediated ac-celeration of aging-associated memory decline in APP/PS1 mice and itspharmacological treatment by picrotoxin. PLoS ONE, 3, e3029. doi:10.1371/journal.pone.0003029
Received January 24, 2014Revision received March 17, 2014
Accepted March 25, 2014 �
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
282 HELLER, RUBY, ROLLS, MAKAM, AND COLAS





























