Embed Size (px)
Citation preview

Transcriptomic Analysis of the Gray short-tailed opossum
(Monodelphis domestica) B-cell Genes
Photo credit: J. Old
Andrea L. Schraven
A thesis presented as fulfilment of the requirements for the degree of Master of Research
Western Sydney University, Hawkesbury
School of Science and Health
Date: December 2019

2
Table of Contents
Table of Contents ......................................................................................................................... 2
Table of Figures ............................................................................................................................ 4
Table of Tables ........................................................................................................................... 10
Acknowledgements .................................................................................................................... 11
Statement of Authenticity .......................................................................................................... 12
Preface ......................................................................................................................................... 13
Publications and Prepared Manuscripts................................................................................... 14
Conference Presentations .......................................................................................................... 15
Abstract ........................................................................................................................................ 16
Introduction ............................................................................................................................ 18
1.1 Mammalian Evolution ......................................................................................................... 19
1.1.1 Marsupial Reproductive Strategies ........................................................................... 20
1.2 Marsupial Immunology ....................................................................................................... 22
1.3 Marsupial Genomics ............................................................................................................ 27
1.3.1 Transcriptomics ........................................................................................................... 29
1.4 Model Marsupial: The gray short-tailed opossum (Monodelphis domestica) ............. 32
1.5 Research Aim ....................................................................................................................... 34
1.6 Thesis Outline ...................................................................................................................... 35
1.7 Chapter 1 References........................................................................................................... 36
Immunogenetics of Marsupial B-cells .................................................................................. 44
2.1 Chapter Outline and Authorship ........................................................................................ 45
2.2 Introduction .......................................................................................................................... 46
2.3 Mammalian Evolution ......................................................................................................... 48
2.4 Marsupial Immunology ....................................................................................................... 49
2.5 B-cells .................................................................................................................................... 51
2.5.1 Immunoglobulins ......................................................................................................... 58
2.5.2 Cluster of Differentiation Markers ............................................................................ 66
2.5.3 Signal Transduction Molecules ................................................................................. 69
2.5.4 Transcriptional Regulators ........................................................................................ 71
2.6 Conclusions .......................................................................................................................... 72
2.7 Conflicts of Interest ............................................................................................................. 72
2.8 Chapter 2 References........................................................................................................... 73
Single-cell transcriptome analysis of the B-cell repertoire reveals the usage of
immunoglobulins in the gray short-tailed opossum (Monodelphis domestica) ................. 83

3
3.1 Chapter Outline and Authorship ........................................................................................ 84
3.2 Introduction .......................................................................................................................... 85
3.3 Materials and Methods ........................................................................................................ 87
3.3.1 Ethics Statement........................................................................................................... 87
3.3.2 Tissue Collection, Cell Sorting and RNA Extraction .............................................. 87
3.3.3 cDNA Synthesis, Sequencing and Alignment ........................................................... 87
3.3.4 Single-cell Identification and Statistical Analyses .................................................. 88
3.4 Results ................................................................................................................................... 89
3.4.1 Heavy and Light Chain Immunoglobulin Usage ..................................................... 89
3.4.2 Immunoglobulin Variable and Joining Domains .................................................... 90
3.4.3 VH and VL Gene Segment Pairing ............................................................................ 93
3.5 Discussion ............................................................................................................................. 95
3.6 Acknowledgements ............................................................................................................. 99
3.7 Conflicts of Interest ............................................................................................................. 99
3.8 Chapter 3 References......................................................................................................... 100
3.9 Chapter 3 Supplementary Figures ................................................................................... 103
Single-cell transcriptomic analysis of marsupial B-cells, the gray short-tailed opossum
(Monodelphis domestica) ...................................................................................................... 105
4.1 Chapter Outline and Authorship ...................................................................................... 106
4.2 Introduction ........................................................................................................................ 107
4.3 Materials and Methods ...................................................................................................... 109
4.3.1 Ethics Statement......................................................................................................... 109
4.3.4 Transcriptome Annotation and Gene Ontology .................................................... 110
4.3.5 Statistical Analyses .................................................................................................... 111
4.3.6 Phylogenetic Analyses and Motif Comparison ...................................................... 111
4.4 Results ................................................................................................................................. 112
4.4.1 Overview of the Transcriptome ............................................................................... 112
4.4.2 Highly Expressed Genes ........................................................................................... 114
4.5 Discussion ........................................................................................................................... 126
4.6 Acknowledgements ........................................................................................................... 130
4.7 Conflicts of Interest ........................................................................................................... 130
4.8 Chapter 4 References......................................................................................................... 131
Concluding Summary and Future Directions ................................................................... 135
5.1 Concluding Summary ........................................................................................................ 136
5.2 Future Directions ............................................................................................................... 140
5.3 Chapter 5 References......................................................................................................... 142

4
Table of Figures
Figure 2.1 Phylogenetic tree based on B-cell relationship in vertebrate groups and divergence
of marsupials referred to in this review, constructed using the NJ method.
Figure 2.2 Unrooted phylogenetic tree based on amino acid alignments of vertebrate IgD
sequences, constructed using the NJ method. Genbank accession numbers: I. punctatus IgD,
ADF56020; S. trutta IgD, AAV28076; E. coioides IgD, AEN71107; M. musculus IgD,
AAB59653; X. tropicalis IgD, ABC75541; P. sinensis IgD, ACU45375; O. anatinus IgD,
ACD31540; C. siamensis IgD, AFZ39208; E. macularius IgD, ABY67439; H. sapiens IgD,
AAA52771; P. troglodytes IgD, PNI88328; S. scrofa IgD, AAP23939; B. taurus IgD,
AAN03673; O. aries IgD, AAN03671.
Figure 2.3 Unrooted phylogenetic tree based on amino acid alignments of marsupial and
eutherian IgA and IgG subclasses sequences, constructed using the NJ method. Genbank
accession numbers: M. domestica IgG, AAC79675; T. vulpecula IgG, AAD55590; M.
domestica IgA, AAD21190; T. vulpecula IgA, AAD41690; N. eugenii IgA, ABV01995; M.
musculus IgA, AAB59662; H. sapiens IgA2, AAB59396; H. sapiens IgA1, AAC82528; H.
sapiens IgG1, BAC87418; H. sapiens IgG3, CAA27268; H. sapiens IgG2, AAB59393; H.
sapiens IgG4, CAC20457; M. musculus IgG1, CAD32497; M. musculus IgG3, ADK11691;
M. musculus IgG2b, ACX70084; M. musculus IgG2a, BAC44883; M. musculus IgG2c,
CAA34864.
Figure 2.4 Genomic organisation of the M. domestica IgH, Igκ, and Igλ loci. V, D (in the
IgH), and J gene segments are clustered, and C gene segments are shown (not to scale).
Direction of transcription is indicated by the Telomeric and Centromeric ends. Dotted lines
indicate where there are gaps in MonDom5 genome sequence. For the IgH locus, the C

5
domain is shown below the IgH main line, the presumptive IgM pseudogene is indicated with
a psi. H identifies the sequences with an encoding hinge region. Polyadenylation and
transmembrane protein domain are designated with Poly A and TM, respectively. Double
angled lines indicate where there are large regions of separation. The Igκ shows two large
clusters of Vκ gene segments with an 800 Kb region separating them. Circles on the Igκ
identify a recombining element, RE, and two kappa enhancers, Ke5’and KE3’P, respectively.
Pairing IgλJ and IgλC genes are designated according to their order.
Figure 2.5 Amino acid alignment of expressed (A) CD79α and (B) CD79α mammalian
sequences. Dots indicate identical bases to M. domestica CD79 transcripts. The Ig domain,
TM regions and CYT regions are indicated by arrows below the sequences, the ITAM motifs
are identified by a box around the amino acids. Genbank accession numbers: M. domestica
(CD79α, ACZ13436; CD79β, ACZ13438.1), N. eugenii (CD79α, ACZ13437; CD79β,
ACZ13439.1), (O. fraenata CD79α, AGO64767; CD79β, AII23405.1), H. sapiens (CD79α,
AAI13732; CD79β, AAH02975.2).
Figure 3.1 Fraction of cells expressing immunoglobulin heavy (IgM, IgG, IgA, IgE) and
light (Igκ and Igλ) chains in both spleen and blood single cells.
Figure 3.2 Fraction of cells expressing V and J domains of the Ig heavy (IgM, IgG, IgA, IgE)
chains in both spleen and blood single cells. NA indicates no information.
Figure 3.3 Fraction of cells expression variable and joining domains of the immunoglobulin
light (Igκ) chains in both spleen and blood single cells. NA indicates no information.
Figure 3.4 Fraction of cells expression variable and joining domains of the immunoglobulin
light (Igλ) chains in both spleen and blood single cells. NA indicates no information.
Figure 3.5 Heat map indicating the frequency of VH/Vκ gene segment pairs expressed in the
opossum repertoire.

6
Supplementary Figure 3.1 Previously annotated pseudogene VH1.10. Identification of ORF
on the reverse strand of frame 1, 3’ – 5’. Start codon, 2250; stop codon, 415; length, 612
amino acids. Stop codon is indicated by the asterisks.
Supplementary Figure 3.2 Previously annotated pseudogene Vκ1.17. Identification of ORF
on the reverse strand of frame 1, 3’ – 5’. Start codon, 273; stop codon, 37; length, 79 amino
acids. Stop codon is indicated by the asterisks.
Supplementary Figure 3.3 Previously annotated pseudogene Vκ7.11. Identification of ORF
on the reverse strand of frame 3, 3’ – 5’. Start codon, 990; Stop codon, 235; length, 252
amino acids. Stop codon is indicated by the asterisks.
Figure 3.6 Heat map indicating the frequency of VH/Vλ gene segment pairs expressed in the
opossum repertoire.
Figure 4.1 Overall distribution of GO categorical terms in the opossum cell transcriptome.
(A) Molecular function, (B) Biological process, and (C) Cellular components. Percentages
are relative of all genes which are categorised as a percentage of all annotated transcripts.
Figure 4.2 Overall distribution of GO categorical terms within the Immune System Process
Category. Percentages are relative of all genes which are categorised as a percentage of all
annotated transcripts.
Figure 4.3 (A) Multiple sequence alignment of SDC4 gene products and (B) a Phylogenetic
tree based on the SDC4 sequences, constructed using the NJ method. Protein sequences were
aligned using the ClustalW algorithm. Dots indicate identical matches to the opossum
sequences. Domains of the SDC4 protein are indicated by a line underneath the sequence.
Species designations are abbreviated to the first two letters of their scientific names. M.
domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus
(Oran), H. sapiens (Hosa), and M. musculus (Mumu). Genbank accession numbers are: Modo

7
SDC4, XP_007475896.1; Saha SDC4, XP_012395261.1; Phci SDC4, XP_020839750.1;
Vour SDC4, XP_027707442.1; Oran SDC4, XP_028927254.1; Hosa SDC4, NP_002990.2;
Mumu SDC4, NP_035651.1.
Figure 4.4 Two-dimensional non-transformed multidimensional scaling (MDS) plot visually
representing the single-cell transcriptomes of the top 10 highly expressed genes in the spleen
(grey upwards triangles) and blood (black downward triangles).
Figure 4.5 Two-dimensional non-transformed multidimensional scaling (MDS) plot visually
representing the single-cell transcriptomes of the top 10 highly expressed genes in the IgM
(black downward triangle), IgG (dark grey diamond), IgA (grey squares), and IgE (grey
upwards triangle).
Figure 4.6 (A) Multiple sequence alignment of HNRNPK gene products and (B) a
Phylogenetic tree based on the HNRNPK sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the HNRNPK protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues (2) Bullets mark nucleic acid binding regions, (3) Diamonds G-X-X-G motif.
Species designations are abbreviated to the first two letters of their scientific names. M.
domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), O. anatinus (Oran), H. sapiens
(Hosa) and M. musculus (Mumu). Genbank accession numbers are: Modo HNRNPK,
XP_016279557.1; Saha HNRNPK, XP_012398527.1; Phci HNRNPK, XP_020822678.1;
Oran HNRNPK, XP_016083874.1; Hosa HNRNPK, AAH14980.1; Mumu HNRNPK,
ACC69193.1.
Figure 4.7 (A) Multiple sequence alignment of ELMOD1 gene products and (B) a
Phylogenetic tree based on the ELMOD1 sequences, constructed using the NJ method.

8
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the ELMOD1 protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues. Species designations are abbreviated to the first two letters of their scientific names.
M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus
(Oran), and H. sapiens (Hosa). Genbank accession numbers are: Modo ELMOD1,
XP_007494949.1; Saha ELMOD1, XP_012400499.1; Phci ELMOD1, XP_020851842.1;
Vour ELMOD1, XP_027705369.1; Oran ELMOD1, XP_028904013.1; Hosa ELMOD1,
NP_061182.3.
Figure 4.8 (A) Multiple sequence alignment of GLG1 gene products and (B) a Phylogenetic
tree based on the GLG1 sequences, constructed using the NJ method. Protein sequences were
aligned using the ClustalW algorithm. Dots indicate identical matches to the opossum
sequences. Cysteine rich repeats are indicated by lines underneath the sequence. Symbols
above sequence are: (1) Inverted triangles mark cysteine residues. Species designations are
abbreviated to the first two letters of their scientific names. M. domestica (Modo), S. harrisii
(Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus (Oran), H. sapiens (Hosa), and M.
musculus (Mumu). Genbank accession numbers are: Modo GLG1, XP_007477569.1; Saha
GLG1, XP_012395893.2; Phci GLG1, XP_020819407.1; Vour GLG1, XP_027699023.1;
Oran GLG1, XP_028931633.1; Hosa GLG1, AAH60822.1; Mumu GLG1, EDL11491.1.
Figure 4.9 (A) Multiple sequence alignment of HLA-DRA gene products and (B) a
Phylogenetic tree based on the HLA-DRA sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the HLA-DRA protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine

9
residues, (2) Bullets mark heterodimer interface residues, (3) Diamonds mark MHC binding
domain interface residues. Species designations are abbreviated to the first two letters of their
scientific names. M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus
(Vour), and H. sapiens (Hosa). Genbank accession numbers are: Modo HLA-DRA,
XP_007483702.1; Saha HLA-DRA, XP_003770700.1; Phci HLA-DRA, XP_020829855.1;
Vour HLA-DRA, XP_027732036.1; Hosa HLA-DRA, ARB08447.1.
Figure 4.10 (A) Multiple sequence alignment of marsupial MHC DAB1 gene products and
(B) a Phylogenetic tree based on the MHC DAB sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the MHC DAB protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues. Species designations are abbreviated to the first two letters of their scientific names.
M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), N. eugenii (Noeu), and T.
vulpecula (Trvu). Genbank accession numbers are: Modo DAB1, NP_001028163.1; Saha
DAB, ABV58848.1; Phci DAB, ALX81653.1; Noeu DAB, AAX54675.1; Trvu DAB,
AAK84172.1.

10
Table of Tables
Table 1.1 Summary marsupial immune tissues, examined in the thymus, bone marrow,
lymph nodes, and spleen.
Table 2.1 List of B-cell genes that have either been expressed or predicted in marsupial
species, including their Genbank accession Identifications (ID).
Table 3.1 IgL isotypes Igκ:Igλ ratio expression in mammals.
Table 4.1 Top 100 most highly expressed genes in the opossum B-cell transcriptome. Total
percentages are calculated from spleen and blood counts.

11
Acknowledgements
Firstly, I’d like to thank Associate Professor Julie Old, for all the support and encouragement
throughout my Masters. Thank you for pushing me outside my comfort zone, especially when
it came to presenting at conferences. I’d like to thank my co-supervisors Dr. Hayley Stannard
and Dr. Oselyne Ong for reading and providing feedback on my endless drafts, and your
words of encouragement when tackling the journal revisions.
Thank you Professor Robert Miller for being a wonderful collaborator and sharing your data
with me. Thank you Kimberly Morrisey and Bethaney Fehrenkamp for your assistance in the
initial analysis, and Associate Professor Ricky Spencer for your help with the statistical
analyses. Special thanks to Danny Douek, Galina Petukhova and especially Victoria Hansen
for generating the data used in all analyses.
I’d like to give thanks to the friends I have made along the way, Blaire Vallin, Chandni
Sengupta, Rowan Thorley, Janice Vaz, Corrine Letendre and Kristen Petrov who have all
supported me during some of my highest and lowest points throughout my Masters. I am
forever grateful and wish you all well in your own degrees and future endeavours.
To my parents, Mark and Linda, whose work ethics have always been an inspiration to me.
Thank you for your support in everything I do and always showing an interest in my research.
Lastly to my sisters, Rebekah and Alicia, thank you for keeping my sanity in check.

12
Statement of Authenticity
This thesis is submitted to the Western Sydney University in fulfilment of the requirement for
the Degree of Master of Research.
The work presented in this thesis is, to the best of my knowledge, composed of my original
work, and contains no material previously published or written by another person, except
where due reference has been made in the text. I have clearly stated the contribution by others
to jointly authored works that I have included in my thesis.
........................................
Andrea L. Schraven

13
Preface
This thesis is presented as a series of manuscripts, which describes the gene expression of B-
cell in the gray short-tailed opossum (Monodelphis domestica). Each chapter presented is
self-contained, and includes a specific outline and authorship. Of the manuscripts presented
here, Chapter 2 has been published in a peer-review journal, Chapter 3 has been submitted to
a peer review journal and is currently under review, chapter 4 has been prepared for
submission to a peer-reviewed journal. An introductory and general conclusion has been
included in this thesis to assist with the continuity throughout. Format of manuscripts and
reference lists will vary according to the guidelines of journals they are submitted to. All
manuscripts are jointly authored, but in each case, I am first author.

14
Publications and Prepared Manuscripts
Schraven, A. L., Stannard, H. J., Ong, O. T. W., Old, J. M. 2020. Immunogenetics of
marsupial B-cells, Molecular Immunology. 117, 1-11.
https://doi.org/10.1016/j.molimm.2019.10.024
Schraven, A. L., Hansen, V. L., Morrissey, K. A., Stannard, H. J., Ong, O. T. W., Douek, D.
C., Miller, R. D., Old, J. M. Single-cell transcriptome analysis of the B-cell repertoire
exposes the usage of immunoglobulins in the gray short-tailed opossum (Monodelphis
domestica). Immunology and Cell Biology. Under Review.
Schraven, A. L., Hansen, V. L., Morrissey, K. A., Stannard, H. J., Ong, O. T. W., Douek, D.
C., Miller, R. D., Old, J. M. Single-cell transcriptomic analysis of marsupial B-cells, the gray
short-tailed opossum (Monodelphis domestica). Prepared for Immunology and Cell Biology.

15
Conference Presentations
Schraven, A. L., Miller, R. D., Stannard, H. J., Ong, O. T. W., Hansen, V. L., Morrissey, K.
A. Old, J. M. Bioinformatics analyses of B-cell repertoire expression in the gray short-tailed
opossum (Monodelphis domestica). June, 2019. Presented at the North American
Comparative Immunology Workshop, Waterloo, Canada.

16
Abstract
Marsupials and eutherians are mammals that differ in their physiological traits, predominately
their reproductive and developmental strategies; eutherians give birth to well-developed
young, while marsupials are born highly altricial after a much shorter gestation. These
developmental traits result in differences in the development of the immune system of
eutherian and marsupial species. B-cells are key to humoral immunity, are found in multiple
lymphoid organs, and have the unique ability to mediate the production of antigen-specific
antibodies in the presence of pathogens. Marsupial B-cell investigations have become
increasingly important in understanding an adaptive immune system that develops primarily
ex utero. In comparison to eutherians and monotremes, marsupial B-cells have four
Immunoglobulin (Ig) heavy (H) chain isotypes (IgA, IgG, IgM and IgE) and two light (L)
chain isotypes; lambda (Igλ) and kappa (Igκ). The gray short-tailed opossum (Monodelphis
domestica) is a well-established model marsupial species, with a well annotated genome. The
B-cell transcriptome of an individual opossum was investigated by Next Generation RNA-
Seq techniques at the single-cell level. A total of 273 single-cells and 575,721 contigs were
generated, annotation of the transcriptome identified 14,654 unique genes. The first study of
this thesis analysed the IgH and IgL usage in the opossum B-cell repertoire. Not surprisingly,
IgM had the highest expression in the repertoire, followed by IgA, IgG, and very few cells
expressing IgE. Despite Igκ being the most complex IgL isotype, the ratio of Igκ to Igλ was
35:65. IgL isotypes have been identified to have a greater contribution to antibody
diversification than IgH isotypes, due to the complexity and abundance of IgL variable (V)
gene segments. The second study of this thesis examined the whole opossum B-cell
transcriptome and analysed the most highly expressed genes. The most abundant gene
transcripts were Sydnecan-4, making up 0.66% of the entire transcriptome. IgM and IgG cells
produced significantly more transcripts of the golgi glycoprotein 1 and ELMO domain-

17
containing protein genes in comparison to IgA. Since IgE expressing cells were very low in
number, a definitive comparison could not be made between all IgH cells. Highly expressed
genes associated with the marsupial immune system included MHC class II DRα chain and
MHC class II DAβ chain. The diverse array of genes identified in the opossum single-cell
transcriptome reveals the importance of marsupial B-cells in producing endogenous antibody
responses, and has allowed for a comparative analysis with other mammalian lineages.

18
CHAPTER 1
Introduction

19
1.1 Mammalian Evolution
Marsupials, or metatherians, are one of three major mammalian lineages, along with
monotremes (Prototherians) and eutherian mammals (Selwood and Coulson, 2006).
Monotremes, the egg-laying mammals, diverged from viviparous mammals over 186 million
years ago (MYA) (Bininda-Emonds et al., 2007). There are two distinct living monotreme
clades comprising of five species (Rowe et al., 2008); the short-beaked echidna
(Tachyglossus aculeatus) (Griffiths, 2012), three long-beaked echidna species (Zaglossus
attenboroughi, Zaglossus bartoni, and Zaglossus bruijni) (Gambaryan and Kuznetsov, 2013),
and the platypus (Ornithorhynchus anatinus) (Johansson et al., 2002, Bininda-Emonds et al.,
2007). Eutherians and marsupials shared their last common ancestor more recently, between
130 to 180 MYA (Nilsson et al., 2004, Nilsson et al., 2010), resulting in more physiological
similarities than with monotremes, such as giving birth to live young (Tyndale-Biscoe and
Renfree, 1987).
One of the key differences between eutherians and marsupials is their prenatal and
postnatal investment in offspring. Marsupials have a relatively short gestational period in
comparison to eutherians, resulting in highly altrical young (Block, 1964, Ashman and
Papadimitriou, 1975, Tyndale-Biscoe and Renfree, 1987, Old and Deane, 2000); Block
(1964) described a newborn marsupial to be developmentally equivalent to a nine-week-old
human foetus. The major developmental stages which take place during the first few
postnatal months in marsupials have been the focus of many reproductive investigations of
marsupials, as these events occur outside a sterile uterine environment. (Block, 1964, Yadav
et al., 1972) In contrast, the major developmental milestones in eutherians only occur during
their prenatal period, and as a result low survival rates occur in underdeveloped individuals
(Hamilton et al., 2010). At birth marsupial neonates have well-developed forelimbs and
mouth parts, allowing them to climb towards the mother’s teat and successfully attach to the

20
teat (Edwards and Deakin, 2013). The development of the forelimbs and mouth parts allow
the neonate to embark on a lengthy and fixed lactation period , where important physical
(Cooper and Steppan, 2010) and immunological maturation occurs (Old and Deane, 2000). In
the past, the advantages of marsupial reproduction have been heavily debated (Kirsch, 1977a,
Kirsch, 1977b, Parker, 1977, Low, 1978, Cockburn, 1989). Specific benefits for short
gestation periods include reduced physiological energy cost on the mother, reduced
susceptibility to predator attack, and increased foraging capabilities (Kirsch, 1977a, Kirsch,
1977b). Since marsupials are ectothermic at birth and gradually become endothermic during
the lactation period (Ikonomopoulou and Rose, 2006), neonates have lower metabolic rates,
which increases postnatal parental care, postpartum (Hopson, 1973, Case, 1978).
1.1.1 Marsupial Reproductive Strategies
Protection from environmental risks is afforded to marsupials through the marsupium,
or pouch (Edwards and Deakin, 2013). Russell (1982) described six different types of
pouches that have varying structural distinction between species. Type one pouches only
have skin folds on the underbelly of the mother for the neonate to adhere to (or ‘pouchless’)
(Griffiths and Slater, 1988). Type two has a partial covering of the mammary area; type three,
the fold of skin is arranged in a circular pattern with a central opening to the teat. In type four
pouches, the mammary area is covered and the mammary glands are located in two pockets
on the underbelly of the mother. Type five and six pouches are completely enclosed and
deep, the difference being, type five is located on the underbelly (anterior), whereas type six
is located on the backside (posterior) of the mother (Russell, 1982). Different pouch types can
also influence the parental care given to neonates (Russell, 1982). Species with deep and
enclosed pouches will typically have 1-2 neonates per litter (Edwards and Deakin, 2013) in
which the neonate will remain in the pouch until a certain stage of development has occurred

21
and the mother can either leave the neonate in a nest, or the neonate has the ability to follow
the mother on foot (Russell, 1982). Deep and enclosed pouches are beneficial, as these
species have better control of environmental factors, such as humidity and warmth, which
allows the neonate to reach the developmental stage where their eyes are open, they have fur,
and can thermoregulate before leaving the pouch (Russell, 1982). On the other hand, the
‘pouchless’ marsupials like that of the gray short-tailed opossum (Monodelphis domestica)
(Fadem et al., 1982), have multiple offspring per litter and the mother leaves the young in the
nest at an early developmental stage where their eyes are still closed and they have no
apparent thermoregulation (Edwards and Deakin, 2013). Although ‘pouchless’ marsupials
leave their neonates in a nest at a substantially early developmental stage in comparison to
deep and enclosed pouched species, parental care of the neonate is still ongoing (Russell,
1982). When neonates of ‘pouchless’ species are left in the nest, the mother only leaves the
nest briefly for foraging purposes, otherwise the mother will remain in the nest keeping the
neonates close to her underbelly, allowing the neonates to voluntarily suckle from her
mammary glands (Aslin, 1974, Morton, 1978).
In addition to the pouch, marsupials also produce milk for the neonate as another form
of protection in the form of antibodies (or immunoglobulins) (Deane and Cooper, 1984). The
transfer of milk is multifaceted over four phases associated with different developmental
stages. Joss et al. (2009) described these phases in the tammar wallaby (Notamacropus
eugenii); phase 1 includes secretions of high levels of antibodies, lipids and proteins
(equivalent to colostrum), and ceases 48 h postpartum. Phase 2a occurs during the first 100
days postpartum, where the neonate adheres permanently to the teat and most immunological
development occurs (Nicholas, 1988, Joss et al., 2009). Phase 2b covers days 100-200 and
the neonate suckles less frequently, both phase 2a and 2b includes secretions of high levels of
carbohydrates and decreased levels of proteins and fats (Nicholas, 1988). At phase 3, the final

22
phase, milk secretions consist of high levels of proteins and fats, and low levels of
carbohydrates (Young et al., 1997). During this final phase, the neonate spends less time in
the pouch as it is less reliant on the milk (Edwards and Deakin, 2013).
1.2 Marsupial Immunology
It was once believed that the marsupial immune system was ‘primitive’ in comparison
to eutherians (Jurd, 1994), however copious studies have since described it to be just as
complex as the eutherian immune system (Bell, 1977, Harrison and Wedlock, 2000, Miller
and Belov, 2000, Belov et al., 2013, Old, 2016), despite the marsupial immune system
developing and maturing in a non-sterile environment (Old and Deane, 1998). The
development of marsupial immune tissues have been studied extensively over the past couple
of decades (Table 1.1) (Block, 1964, Ashman and Papadimitriou, 1975, Basden et al., 1996,
Basden et al., 1997, Old et al., 2003, Old et al., 2004b, Belov et al., 2013, Borthwick et al.,
2014, Borthwick and Old, 2016). Marsupials, like eutherians, have both primary and
secondary immune tissues, which have proven to be comparable in development, appearance,
and function (Old et al., 2003). Eutherian primary immune tissues are involved in the
production and maturation of lymphocytes (e.g. B- and T-cells), and include the thymus and
bone marrow (Block, 1964, Ashman and Papadimitriou, 1975, Old et al., 2004a). The
secondary tissues includes, but is not limited to, the spleen, lymph nodes, mucosal-associated
lymphoid tissues (MALT), gut-associated lymphoid tissues (GALT), bronchus-associated
lymphoid tissues (BALT), tonsils, and Peyer’s patches (Block, 1964, Ashman and
Papadimitriou, 1975, Basden et al., 1996, Cisternas and Armati, 1999). In contrast to
eutherians, marsupials are immunologically immature at birth (Block, 1964, Old et al., 2003,
Belov et al., 2013), and therefore the maturation of primary and secondary tissues occurs
during ‘pouch life’ (Old and Deane, 2000, Borthwick et al., 2014, Borthwick and Old, 2016).
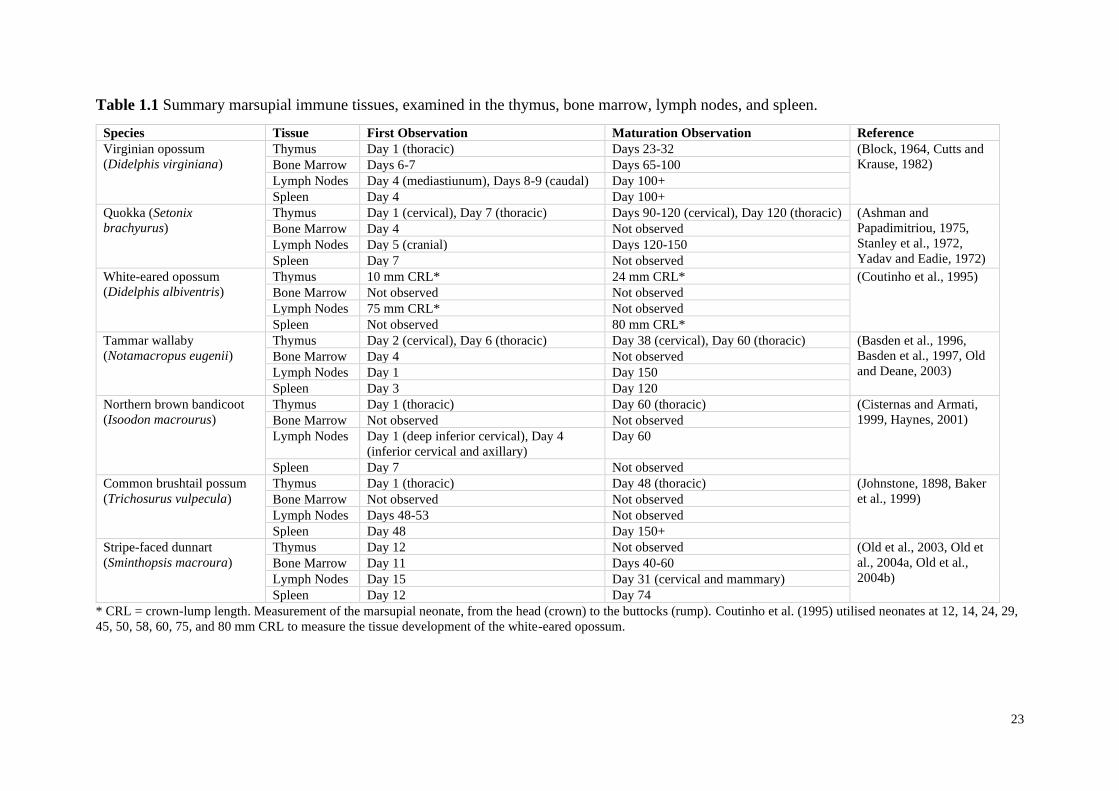
23
Table 1.1 Summary marsupial immune tissues, examined in the thymus, bone marrow, lymph nodes, and spleen.
Species Tissue First Observation Maturation Observation Reference
Virginian opossum
(Didelphis virginiana)
Thymus Day 1 (thoracic) Days 23-32 (Block, 1964, Cutts and
Krause, 1982) Bone Marrow Days 6-7 Days 65-100
Lymph Nodes Day 4 (mediastiunum), Days 8-9 (caudal) Day 100+
Spleen Day 4 Day 100+
Quokka (Setonix
brachyurus)
Thymus Day 1 (cervical), Day 7 (thoracic) Days 90-120 (cervical), Day 120 (thoracic) (Ashman and
Papadimitriou, 1975,
Stanley et al., 1972,
Yadav and Eadie, 1972)
Bone Marrow Day 4 Not observed
Lymph Nodes Day 5 (cranial) Days 120-150
Spleen Day 7 Not observed
White-eared opossum
(Didelphis albiventris)
Thymus 10 mm CRL* 24 mm CRL* (Coutinho et al., 1995)
Bone Marrow Not observed Not observed
Lymph Nodes 75 mm CRL* Not observed
Spleen Not observed 80 mm CRL*
Tammar wallaby
(Notamacropus eugenii)
Thymus Day 2 (cervical), Day 6 (thoracic) Day 38 (cervical), Day 60 (thoracic) (Basden et al., 1996,
Basden et al., 1997, Old
and Deane, 2003) Bone Marrow Day 4 Not observed
Lymph Nodes Day 1 Day 150
Spleen Day 3 Day 120
Northern brown bandicoot
(Isoodon macrourus)
Thymus Day 1 (thoracic) Day 60 (thoracic) (Cisternas and Armati,
1999, Haynes, 2001) Bone Marrow Not observed Not observed
Lymph Nodes Day 1 (deep inferior cervical), Day 4
(inferior cervical and axillary)
Day 60
Spleen Day 7 Not observed
Common brushtail possum
(Trichosurus vulpecula)
Thymus Day 1 (thoracic) Day 48 (thoracic) (Johnstone, 1898, Baker
et al., 1999) Bone Marrow Not observed Not observed
Lymph Nodes Days 48-53 Not observed
Spleen Day 48 Day 150+
Stripe-faced dunnart
(Sminthopsis macroura)
Thymus Day 12 Not observed (Old et al., 2003, Old et
al., 2004a, Old et al.,
2004b) Bone Marrow Day 11 Days 40-60
Lymph Nodes Day 15 Day 31 (cervical and mammary)
Spleen Day 12 Day 74
* CRL = crown-lump length. Measurement of the marsupial neonate, from the head (crown) to the buttocks (rump). Coutinho et al. (1995) utilised neonates at 12, 14, 24, 29,
45, 50, 58, 60, 75, and 80 mm CRL to measure the tissue development of the white-eared opossum.

24
The thymus is the first immunological tissue to mature in marsupials (Basden et al.,
1997), and due to its important role in cell-mediated immunology, via the production of T-
cells, it is the most widely investigated immune tissue in marsupial immunology (Johnstone,
1898, Yadav et al., 1972, Cisternas and Armati, 1999, Wong et al., 2011b). Anatomical
location and whether both the cervical and thoracic thymuses are present varies among the
marsupial lineage (Old and Deane, 2000). All marsupials (except wombats) have a thoracic
thymus, while the Diprotodontia order of marsupials also have a cervical thymus (Yadav et
al., 1972, Haynes, 2001). Yadav et al. (1972) reported the variance in presence of a cervical
thymus in the quokka (Setonix brachyurus) was due to the small size of the thoracic cavity,
whilst Ashman and Papadimitriou (1975) thought it may be due to the microbiological
enriched environment of the pouch. During the first few days after birth, the thymus begins to
mature rapidly and mature lymphocytes start to appear (Block, 1964, Rowlands et al., 1964,
Stanley et al., 1972, Johnstone, 1898, Baker et al., 1999). Detection of CD3+ T-cells were
detected at day 2 postpartum in both the white-eared opossum (Didelphis albiventris)
(Coutinho et al., 1994) and common brushtail possum (Trichosurus vulpecula) (Baker et al.,
1999), however full maturation of the thymus occurred in the possum on day 25 postpartum
(Baker et al., 1999). Whereas in the Virginian opossum (Didelphis virginiana), lymphocytes
appeared as early as day 1 postpartum, histological development of the Hassall’s corpuscles
began on day 5 and the opossum thymus continued to develop its medullary and cortical
areas until day 13-16, being fully matured by day 23 postpartum (Table 1.1) (Block, 1964).
Unlike eutherians, the bone marrow is not actively haematopoietic at birth, this role is
taken on by the liver due to the bones of the neonate being solely cartilaginous (Old and
Deane, 2000). During the first two weeks postpartum, haematopoiesis is increasingly
observed in the bone marrow of marsupial species and the liver’s involvement starts to
decline. In the Virginian opossum, primitive bone marrow appeared in the diaphysis of the

25
endochondral bones and primary bone marrow observed in the membranous bone on day 5
postpartum (Block, 1964). On days 13-16, the bone marrow continued to increase in the
endochondral bones and at days 23-32, they were filled with bone marrow (except in the
epiphysis). The opossum bone marrow appeared to be similar to that of adult mammals by
days 65-100 postpartum (Table 1.1) (Block, 1964). In other marsupial species, such as the
quokka (Setonix brachyurus) and tammar wallaby, primary bone marrow started to appear as
early as day 4 postpartum (Ashman and Papadimitriou, 1975, Basden et al., 1996), and by the
end of the second week postpartum, a diversity of cells including granulocytes, erythroblasts
and lymphocytes, had increased their numbers substantially (Ashman and Papadimitriou,
1975, Basden et al., 1996). By the end of the first month in the tammar wallaby, the bone
marrow had fully taken over the role of haematopoietic production (Block, 1964, Old et al.,
2004a).
Secondary tissues such as the spleen and lymph nodes start to develop and mature
much later than the thymus (Old and Deane, 2000). Characteristically, adult spleen and
lymph nodes have follicles and germinal centres throughout their respective tissues which are
the major sites for B-cell proliferation and continued development (Belov et al., 2013). White
and red pulp have been observed in adult marsupial splenic tissues, similar to eutherians,
however studies have reported very small differences in size, and number of sinuses and
follicles (Cutts and Krause, 1982, Baker et al., 1999, Old et al., 2004a). Spleen and lymph
node development has increasingly been studied in numerous species, including the Virginian
opossum (Block, 1964), quokka (Ashman and Papadimitriou, 1975), white-eared opossum
(Coutinho et al., 1995), tammar wallaby (Basden et al., 1996, Basden et al., 1997, Old and
Deane, 2003), northern brown bandicoot (Isoodon macrourus) (Cisternas and Armati, 1999),
common brushtail possum (Baker et al., 1999), and stripe-faced dunnart (Sminthopsis
macroura) (Old et al., 2004a, Old et al., 2004b). In marsupial neonates, the spleen is very

26
immature at birth. Composed of mesenchymal tissue, the spleen starts to develop as early
erythrocytes and megakaryocytes begin to populate the tissue, which is followed on by
granulopoietic cells (Cutts and Krause, 1982). The spleen begins to mature with the
appearance of red and white pulp, and reaches maturity when germinal centres can be found
(Old, 2016). In the Virginian opossum, large lymphocytes were apparent on day 2, and only
rare erythrocytes and megakaryocytes appeared on day 3 (Block, 1964). However,
megakaryocytes were more abundant in the tammar wallaby on day 3 (Basden et al., 1996).
Haematopoiesis, erythropoiesis, granulocytopoiesis, and size of the spleen continually
increased during the first two weeks postpartum, in the Virginian opossum, and between days
60 and 100, splenic nodules and germinal centres started to appear (Block, 1964, Cutts and
Krause, 1982). By day 60, in the tammar wallaby, the splenic tissue was much more distinct
with areas of red and white pulp, with the white pulp continuing to increase in abundance
until four months after birth (Basden et al., 1996). As the spleen continues to mature, the
germinal centre and follicles are utilised more for B-cell maturation, differentiation, and
proliferation (Old, 2016). In comparison to the spleen, rudimentary lymph node tissues are
not present until day 4 postpartum, of the Virginian opossum, quokka, and northern brown
bandicoot (Block, 1964, Ashman and Papadimitriou, 1975, Cisternas and Armati, 1999).
However, the developmental pattern of the lymph nodes is similar to that of the spleen
(Bryant and Shifrine, 1974), whereby during the first two weeks postpartum, various cell
types, lymphocytes, myelocytes and erythroblasts were visible in the Virginian opossum
(Block, 1964), and additional myeloid cells and megakaryocytes were seen in the quokka
(Ashman and Papadimitriou, 1975, Old, 2016).

27
1.3 Marsupial Genomics
In the past studying the genome of marsupials has been difficult and traditional
strategies required an abundance of pre-existing resources (Abts et al., 2015, Sette et al.,
2005). The utilisation of conventional experimental methods that generate primers based on
eutherian sequences for reverse-transcriptase polymerase chain reactions (RT-PCR) have
limited the practicability of marsupial genomic studies (Harrison and Wedlock, 2000).
Advancements in the field have enabled researchers to utilise sequencing technology, from a
whole shotgun sequencing (WGS) approach (Mikkelsen et al., 2007) to now using next
generation sequencing (NGS) technology for Sanger sequencing (Murchison et al., 2012,
Deakin, 2012). The marsupial genomes are similar in size to eutherians (Deakin, 2012),
although they are packaged into larger and fewer chromosomes. The gray short-tailed
opossum was the first marsupial species to have its genome sequenced and is available
through the Broad Institute and NCBI Genbank (Mikkelsen et al., 2007). This resource has
provided invaluable data to analyse the evolution of developmental biology (Samollow,
2006), immunogenetics (Deakin et al., 2006, Mikkelsen et al., 2007), cancer development,
and disease susceptibility (Wong et al., 2006) of the different mammalian lineages. The
opossum genome was Sanger-sequenced to ~7x coverage or draft quality, producing a highly
usable assembly, reflected by the large contig N50 of 108kb and scaffold N50 of 60mb
(Papenfuss et al., 2010). However, the annotation of the opossum was largely based on
human transcripts and protein sequences (Hubbard et al., 2007), due to no large-scale
transcriptomic data for the species (Papenfuss et al., 2010). Despite sequencing of large
amounts of ‘good quality’ conserved genes in the opossum, the genes predominately involved
in immune responses have failed to be identified and/or annotated due to the high rate of
divergence (Wong et al., 2011a). Immune genes, for the most part, succumb to intense
selective pressure due to the requirements of forever evolving pathogens (Zelus et al., 2000).

28
As a result, the Ensembl Consortium has missed many immune genes and less than a third of
the opossum immune genes previously annotated (Wong et al., 2006, Belov et al., 2006) have
been predicted by Ensembl (Curwen et al., 2004). Meanwhile, WGS and Sanger sequencing
approaches were used to sequence the genome of the tammar wallaby (Renfree et al., 2011).
From this data for the tammar wallaby, ~2x sequence coverage was submitted to the NCBI
archives and the initial assembly was constructed from low coverage Sanger sequences.
Additionally, a 5.9x sequence coverage was generated at Baylor College of Medicine Human
Genome Sequencing Centre and used for performing contig correction, super-scaffolding,
and improvements on the initial assembly (Renfree et al., 2011).
Since the initial sequencing of the opossum and tammar wallaby genomes there have
been multiple genome sequencing projects. The Tasmanian devil (Sarcophilus harrisii), for
example, has provided much needed insights into Devil Facial Tumour Disease (DFTD), the
tissue of origin, and the evolution of the tumour (Pyecroft et al., 2007). To understand the
responses to DFTD, two male devil genomes were first sequenced, each inhabiting different
locations (Miller et al., 2011). Sequencing was undertaken by multiple NGS platforms and
the data from each devil combined to produce a genome assembly of a 147.5 kb N50 contig
with ~140,000 scaffolds (Miller et al., 2011). The combination of genome information was
done with the aim of detecting genetic variation between the two individuals (Deakin, 2012).
At the conclusion of the first sequencing project, a second genome project was initiated, in
which a 5-year-old female devil was sequenced and annotated. This project utilised the
Illumina platform to assemble a 1.8 Mb N50 contig with ~35,000 scaffolds (Murchison et al.,
2012). Along with flow cytometry, 99% of the scaffolds were assigned to chromosomes,
which was an immense improvement on the initial Tasmanian devil genome project
(Murchison et al., 2012). Furthermore, annotated genes were identified using the Ensembl
Genebuild Pipeline from 12 pooled devil tissues to construct the first marsupial

29
transcriptome. As a result of such efforts, a total of 18,775 protein coding genes were
identified, 1,213 of these genes did not have a human or opossum orthologue (Murchison et
al., 2012).
Recently, Johnson et al. (2018) assembled the koala (Phascolarctos cinereus) genome
by long-read Illumina sequencing. Its genome was sequenced and assembled using 57.3-fold
PacBio long-read coverage. The primary contigs consist of 3.19 Gb with an N50 of 11.6Mb
and the alternate contigs totalled 230 Mb with an N50 of 48.8Kb. Additionally, ~30x
sequence coverage of Illumina short reads were used to improve the initial assembly
(Johnson et al., 2018). The koala genome project resulted in the highest coverage of coding
regions in any marsupial genome project, scoring 95.1% of mammalian single-copy
orthologues, as well as identifying 6,124 protein-coding genes from 2,118 gene families. An
analysis of the gene families showed 1,089 of these gene families have more members in the
koala than in any species investigated to date (Johnson et al., 2018). The quality of the koala
genome has initiated research into the genetic diversity of different populations across
Australia, provided a resource into the disease pathogenesis and defence of the Chlamydia
pecorum infection, and the characterised specific genes related to taste and smell (Burgess,
2018).
1.3.1 Transcriptomics
The development of NGS technologies has increased the potential to expand our
knowledge of genomics and transcriptomics (Abts et al., 2015), where a transcriptome is a
complete set of RNA molecules (transcripts) found at a specific stage of development or
physiological condition in a cell (Wang et al., 2009a). The aim of transcriptomics is to
identify and register all the species of the RNA transcript (coding and non-coding inclusive),

30
determine the transcriptional structure of the gene, and quantify the different expression
levels of the transcripts at different developmental stages and/or under numerous
physiological conditions (Wang et al., 2009a). Previously, the characterisation of the
marsupial genome has largely been based on the use of gene prediction tools developed for
the human genome (Papenfuss et al., 2010). Analysis of the transcriptome allows any species
genome to be sequenced at a higher quality than any traditional laboratory technique
completed previously (Allendorf et al., 2010). A number of marsupial species have since
been used for gene expression analyses.
Several transcriptomic studies have been undertaken to show the gene expression
profiles in various tissues in the tammar wallaby. Wong et al. (2011b) analysed the
transcriptome of the cervical and thoracic thymuses in a 178-day old tammar wallaby
neonate, which was the first investigation to compare the gene expression of both thymuses
in any mammalian species. Their results saw that T-cell development and basic thymic
function of both tammar wallaby cervical and thoracic thymuses were highly similar to
eutherian thymuses and found specific genes involved in the differentiation and function of
T-cell in both thymuses (Wong et al., 2011b). A complete transcriptome was sequenced in
the koala (Hobbs et al., 2014), whereby a number of different tissues were sampled from both
a female (liver, brain, heart, kidney, lung, adrenal gland, spleen, uterus, and pancreas) and
male (liver, kidney, spleen, bone marrow, lymph node, salivary gland, and testes) individual.
This study used RNA-sequencing technology and de novo assembly to generate about 100 Gb
of sequences per individual and covering ~15,000 koala genes (Hobbs et al., 2014). Recently,
the long-nosed bandicoot (Perameles nasuta) also had multiple tissues (liver, heart, kidney,
and spleen) sequenced from one male individual (Morris et al., 2018). Immune genes and
immune gene families of both the passive and adaptive immune systems were characterised,
including immunoglobulins, T-cell receptors, major histocompatibility complex (MHC I and

31
MHC II classes) receptors, natural killer cells, cytokines, chemokines, and pattern recognition
receptors. This survey indicated the bandicoot has a diverse range of expressed immune
genes which are similar to those previously identified in the opossum, Tasmanian devil and
koala (Morris et al., 2018).
With the aim to create a better understanding of the evolutionary gestational and
lactational periods which are unique to marsupials, transcriptomic data has been generated
and analysed from the milk and mammary glands of the koala (Morris et al., 2016), milk of
the Tasmanian devil (Hewavisenti et al., 2016), uterus of the opossum (Hansen et al., 2016),
and the mammary glands and placenta of the tammar wallaby (Lefèvre et al., 2007, Guernsey
et al., 2017). Hewavisenti et al. (2016) sequenced, annotated and characterised the genes of
the passive immune system of the Tasmanian devil. They successfully identified 233,660
transcripts, of which more than 17,000 were identified as protein-coding genes and 846 were
immune genes. They collected milk samples at mid-lactation with the assumption that
marsupial neonates have encountered new pathogens as they begin to emerge from the pouch
(Adamski and Demmer, 2000, Daly et al., 2007). The immune compounds in the milk of the
devil found highly expressed caseins, β-lactoglobulin, and α-lactoglobulin (Hewavisenti et
al., 2016) during the mid-lactation period. β-lactoglobulin was also highly expressed during
mid-lactation of the tammar wallaby (Lefèvre et al., 2007) and during the early and late
lactation periods in the koala (Morris et al., 2016). In comparison to the tammar wallaby
(Lefèvre et al., 2007), of the devil had a higher expression of early lactation proteins (ELP)
and late lactation proteins (LLP) during mid-lactation. Hewavisenti et al. (2016) made the
assumption that the discrepancies between expression levels of these genes between the two
species are due to either recruitment of different proteins during mid-lactation or ELP and
LLP having additional and unknown functions. A global uterine transcriptomic analysis has
been performed on the gray short-tailed opossum using NGS on virgin, late stage pregnant

32
and non-pregnant individuals with the objective to establish what genes are associated with
pregnancy in marsupials (Hansen et al., 2016). Hansen et al. (2016) generated paired-end
reads using the Illumina HiSeq 2000 platform and then aligned them to the opossum genome
(Mikkelsen et al., 2007). The outcome of this investigation found over 900 opossum genes
had significantly higher expression in pregnant individuals compared to non-pregnant
individuals, and another 1400 opossum genes expressed more so in non-pregnant individuals.
1.4 Model Marsupial: The gray short-tailed opossum (Monodelphis domestica)
It has been well established that the gray short-tailed opossum is an important, fully
developed laboratory-bred marsupial utilised for many comparative biological investigations
(Lucero et al., 1998). The opossum has been bred in captive colonies in North America
(Kraus and Fadem, 1987, Miller et al., 1998, Wang et al., 2009b), Australia (Fadem et al.,
1982), and Europe (Moore, 1996, Zeller and Freyer, 2001) since the 1970s (Samollow, 2008),
providing researchers with the unique opportunity to investigate various evolutionary
processes that have given rise in modern mammalian lineages (Samollow, 2006).
The gray short-tailed opossum belongs to the family Didelphidae (Szalay, 2006)
which is the most abundant of all the marsupial families in the Didelphimorphia order, with
opossums being the most species-rich genus within the family (Gardner, 1993). The gray
short-tailed opossum is mostly covered in a light grey fur with slight variations (red and/or
white) between populations (O'Connell, 2007). Their tails are naked, semi-prehensile, and
usually make up about half of their body length. The adult opossum body length ranges
between 10-15cm, adult males weigh between 90-155g, whereas adult females weigh
between 80-100g, making them one of the smallest didelphids (Gardner, 1993, O'Connell,
2007). The sexual maturity of the opossum is reached at about 18-20 weeks and their mating

33
behaviour is strongly linked to the olfactory system, in which males will mark their
surroundings via their sternal gland secretions (Trupin and Fadem, 1982). The opossum has a
short 14 day pregnancy, and produces young that are developmentally equivalent to a 40-day-
old human embryo (Block, 1964, Wang et al., 2009b, Rousmaniere et al., 2010). Opossums
have approximately three litters a year, of up to thirteen (average of eight), hence has
established itself as a true “laboratory marsupial” (Samollow, 2008). Unlike laboratory
rodents and guinea pigs, opossums are housed separately unless paired up for breeding
purposes. At 6-months of age, individuals can be housed together to induce oestrus, which
occurs within 8 days (Stone et al., 1996). Since opossums have no enclosed pouch, the
neonates are exposed on the underbelly of the mother, allowing for easy observation and
manipulation during the early postnatal period (Rousmaniere et al., 2010). The neonates are
attached to the teat of the mother for about two weeks, weaned at two months and reach
sexual maturity between five and six months of age. Reproductive decline commences
between 18 and 30 months of age and age-related death naturally occurs between 36 and 42
months (Stone et al., 1996).
The opossum has been actively used in a variety of research programs over the past
30 years, covering topics from behaviour, reproduction, physiology, immunology, genetics
and several human-related health processes. Now with availability of the opossum genome
(Mikkelsen et al., 2007), research areas have successfully progressed into comparative
genomics, detection and mapping of specific genes and gene expression. This resource is not
just vital for investigating normal verses abnormal physiological states, but also practical uses
for animals that are susceptible to disease in the real world (Samollow, 2006, Samollow,
2008).

34
1.5 Research Aim
To investigate the expression of immune genes in marsupials, a transcriptomic
analysis is required. RNA-sequencing (RNA-Seq) will help us to identify and annotate key B-
cell transcriptomes, and ultimately address uncertainties about immune function in
marsupials and advance the examination of the immunological divergence between
mammalian groups (Wang et al., 2009a). RNA-Seq was developed from various novel high-
throughput DNA sequencing data methods (Wang et al., 2009a). The RNA-Seq method
utilises deep-sequencing technologies as an approach to transcriptomic profiling (Wang et al.,
2009a, Chu and Corey, 2012). The methodology allows for a population of RNA to be
converted to a library of cDNA fragments (or primers) with adaptors attached to one or both
ends. As most reads are between 30-400 base pairs (bp) long (Toung et al., 2011), single-end
or pair-end sequencing (Wang et al., 2009a) is necessary to obtain shortened reads
(Mortazavi et al., 2008). Reads can then be mapped to a reference genome or assembled de
novo (Cloonan et al., 2008), and the expression levels for a gene can subsequently be
determined by counting the number of reads that align to its exons (Wang et al., 2009a).
B-cells have a unique role in the humoral immune response of mammals. B-cells are
able to provide detection of almost any type of foreign molecule, or antigen, through their
differentiated cells to produce antibodies and elicit an immune response (Painter et al., 2011).
Therefore, the aims of this thesis are focused on the expression of B-cell genes in the gray
short-tailed opossum. The research detailed here for the first time in a marsupial analyses the
use of IgH and IgL chains in the B-cell repertoire of single cells isolated from spleen and
peripheral blood. Secondly, the single-cell transcriptome of these B-cells was further
analysed and the genes most highly expressed compared between the different IgH producers.
These findings will contribute to our understanding of comparative immunology and
genomics, and aid in future studies of the opossum and other marsupial species.

35
1.6 Thesis Outline
This thesis is presented in five chapters, inclusive of this introduction (Chapter 1).
The following three chapters are presented as manuscripts for peer review journals, in which
Chapter 2 is published, Chapter 3 is Under Review, and Chapter 4 is a prepared manuscript.
These chapters are formatted according to the journal specifications, and includes the
referencing style for those journals. The final chapter of this thesis (Chapter 5) discusses the
overall findings of the research and presents an overall conclusion and future direction for
this field of study. The following chapters of this thesis are:
Chapter 2: Immunogenetics of marsupial B-cells
Chapter 3: Single-cell transcriptome analysis of the B-cell repertoire reveals the usage of
immunoglobulins in the gray short-tailed opossum (Monodelphis domestica)
Chapter 4: Single-cell transcriptomic analysis of marsupial B-cells, the gray short-tailed
opossum (Monodelphis domestica)
Chapter 5: Concluding summary and future directions

36
1.7 Chapter 1 References
ABTS, K. C., IVY, J. A. & DEWOODY, J. A. 2015. Immunomics of the koala
(Phascolarctos cinereus). Immunogenetics, 67, 305-21.
ADAMSKI, F. M. & DEMMER, J. 2000. Immunological protection of the vulnerable
marsupial pouch young: two periods of immune transfer during lactation in
Trichosurus vulpecula (brushtail possum). Developmental and Comparative
Immunology, 24, 491-502.
ALLENDORF, F. W., HOHENLOHE, P. A. & LUIKART, G. 2010. Genomics and the
future of conservation genetics. Nature Reviews - Genetics, 11, 697-709.
ASHMAN, R. B. & PAPADIMITRIOU, J. M. 1975. Development of lymphoid tissue in a
marsupial, Setonix brachyurus (quokka). Cells Tissues Organs, 91, 594-611.
ASLIN, H. J. 1974. The behaiviour in Dasyuroides byrnei (marsupialia) in captivity.
Zeitschrift fur Tierpsychologie, 35, 187-208.
BAKER, M. L., GEMMELL, E. & GEMMELL, R. T. 1999. Ontogeny of the immune system
of the brushtail possum, Trichosurus vulpecula. The Anatomical Record, 256, 354-
365.
BASDEN, K., COOPER, D. & DEANE, E. 1996. Development of the blood-forming tissues
of the tammar wallaby Macropus eugenii. Reproduction, Fertility and Development,
8, 989-994.
BASDEN, K., COOPER, D. W. & DEANE, E. M. 1997. Development of the lymphoid
tissues of the tammar wallaby Macropus eugenii. Reproduction Fertility and
Development, 9, 243-254.
BELL, R. G. 1977. Marsupial immunoglobulins: the distribution and evolution of macropod
IgG2, IgG1, IgM and light chain antigenic markers within the sub-class Metatheria.
Immunology, 33, 917-924.
BELOV, K., DEAKIN, J. E., PAPENFUSS, A. T., BAKER, M. L., MELMAN, S. D.,
SIDDLE, H. V., GOUIN, N., GOODE, D. L., SARGEANT, T. J., ROBINSON, M.
D., WAKEFIELD, M. J., MAHONY, S., CROSS, J. G. R., BENOS, P. V.,
SAMOLLOW, P. B., SPEED, T. P., GRAVES, J. A. M. & MILLER, R. D. 2006.
Reconstructing an ancestral mammalian immune supercomplex from a marsupial
major histocompatibility complex. PLOS Biology, 4, e46.
BELOV, K., MILLER, R. D., OLD, J. M. & YOUNG, L. J. 2013. Marsupial immunology
bounding ahead. Australian Journal of Zoology, 61, 24-40.
BININDA-EMONDS, O. R. P., CARDILLO, M., JONES, K. E., MACPHEE, R. D. E.,
BECK, R. M. D., GRENYER, R., PRICE, S. A., VOS, R. A., GITTLEMAN, J. L. &
PURVIS, A. 2007. The delayed rise of present-day mammals. Nature, 446, 507.
BLOCK, M. D. 1964. The blood forming tissues and blood of the newborn opossum
(Didelphys virginiana). I. Normal development through about the one hundredth day
of life. Ergeb Anat Entwicklungsgesch, 37, 1-237.
BORTHWICK, C. R. & OLD, J. M. 2016. Histological development of the immune tissues
of a marsupial, the red‐tailed phascogale (Phascogale calura). Anatomical Record,
299, 207-219.
BORTHWICK, C. R., YOUNG, L. J. & OLD, J. M. 2014. The development of the immune
tissues in marsupial pouch young. Journal of Morphology, 275, 822-839.
BRYANT, B. & SHIFRINE, M. 1974. The immunohematopoietic and lymphatic systems of
Marmosa mitis: a developmental survey. Journal of the Reticuloendothelial Society,
16, 105.
BURGESS, D. J. 2018. Koala genome insights. Nature Reviews Genetics, 19, 533-533.

37
CASE, T. J. 1978. Endothermy and parental care in the terrestrial vertebrates. The American
Naturalist, 112, 861-874.
CHU, Y. & COREY, D. R. 2012. RNA sequencing: platform selection, experimental design,
and data interpretation. Nucleic Acid Therapeutics, 22, 271-4.
CISTERNAS, P. A. & ARMATI, P. J. 1999. Development of the thymus, spleen, lymph
nodes and liver in the marsupial, Isoodon macrourus (northern brown bandicoot,
Peramelidae). Anatomy and Embryology, 200, 433-443.
CLOONAN, N., FORREST, A. R., KOLLE, G., GARDINER, B. B., FAULKNER, G. J.,
BROWN, M. K., TAYLOR, D. F., STEPTOE, A. L., WANI, S., BETHEL, G.,
ROBERTSON, A. J., PERKINS, A. C., BRUCE, S. J., LEE, C. C., RANADE, S. S.,
PECKHAM, H. E., MANNING, J. M., MCKERNAN, K. J. & GRIMMOND, S. M.
2008. Stem cell transcriptome profiling via massive-scale mRNA sequencing. Nature
Methods, 5, 613-9.
COCKBURN, A. 1989. Adaptive patterns in marsupial reproduction. Elsevier Current
Trends.
COOPER, W. J. & STEPPAN, S. J. 2010. Developmental constraint on the evolution of
marsupial forelimb morphology. Australian Journal of Zoology, 58, 1-15.
COUTINHO, H. B., NOGUEIRA, J. C., KING, G., COUTINHO, V. B., ROBALINHO, T. I.,
AMORIM, A. M., CAVALCANTI, V. M., ROBINS, R. A. & SEWELL, H. F. 1994.
Immunocytochemical study of the ontogeny of peyer's patches in the brazilian
marsupial Didelphis albiventris. Journal of Anatomy, 185 ( pt 2), 347-354.
COUTINHO, H. B., SEWELL, H. F., TIGHE, P., KING, G., NOGUEIRA, J. C.,
ROBALINHO, T. I., COUTINHO, V. B. & CAVALCANTI, V. M. 1995.
Immunocytochemical study of the ontogeny of the marsupial Didelphis albiventris
immune system. Journal of Anatomy, 187 (Pt 1), 37.
CURWEN, V., EYRAS, E., ANDREWS, T., CLARKE, L., MONGIN, E., SEARLE, S. &
CLAMP, M. 2004. The ENSEMBL automatic gene annotation system. Genome
Research, 14, 942-50.
CUTTS, J. H. & KRAUSE, W. J. 1982. Postnatal development of the spleen in Didelphis
virginiana. Journal of Anatomy, 135, 601-613.
DALY, K. A., DIGBY, M., LEFÈVRE, C., MAILER, S., THOMSON, P., NICHOLAS, K. &
WILLIAMSON, P. 2007. Analysis of the expression of immunoglobulins throughout
lactation suggests two periods of immune transfer in the tammar wallaby (Macropus
eugenii). Veterinary Immunology and Immunopathology, 120, 187-200.
DEAKIN, J. E. 2012. Marsupial Genome Sequences: Providing Insight into Evolution and
Disease. Scientifica, 2012, 22.
DEAKIN, J. E., OLP, J. J., GRAVES, J. A. M. & MILLER, R. D. 2006. Physical mapping of
immunoglobulin loci IgH@, IgK@, and IgL@ in the opossum (Monodelphis
domestica). Cytogenetic and Genome Research, 114, 94.
DEANE, E. M. & COOPER, D. W. 1984. Immunology of pouch marsupials. I. levels of
immunoglobulin transferrin and albumin in the blood and milk of euros and wallaroos
(hill kangaroos: Macropus robustus. marsupialia). Developmental & Comparative
Immunology, 8, 863-876.
EDWARDS, M. J. & DEAKIN, J. E. 2013. The marsupial pouch: implications for
reproductive success and mammalian evolution. Australian Journal of Zoology, 61,
41-47.
FADEM, B. H., TRUPIN, G. L., MALINIAK, E., VANDEBERG, J. L. & HAYSSEN, V.
1982. Care and breeding of the gray, short-tailed opossum (Monodelphis domestica).
The International Journal of Laboratory Animal Science, 32, 405-409.

38
GAMBARYAN, P. & KUZNETSOV, A. 2013. An evolutionary perspective on the walking
gait of the long‐beaked echidna. Journal of Zoology, 290, 58-67.
GARDNER, A. L. 1993. Order Didelphimorphia. In: WILSON, D. E. & REEDER, D. M.
(eds.) Mammal species of the world: a taxonomic and geographic reference. 2nd ed.
Washington D.C: Smithsonian Institution Press.
GRIFFITHS, M. 2012. The biology of the monotremes, Elsevier.
GRIFFITHS, M. & SLATER, E. 1988. The significance of striated muscle in the mammary
glands of marsupials. Journal of Anatomy, 156, 141-156.
GUERNSEY, M. W., CHUONG, E. B., CORNELIS, G., RENFREE, M. B. & BAKER, J. C.
2017. Molecular conservation of marsupial and eutherian placentation and lactation.
eLife, 6, e27450.
HAMILTON, M. J., DAVIDSON, A. D., SIBLY, R. M. & BROWN, J. H. 2010. Universal
scaling of production rates across mammalian lineages. Proceedings of the Royal
Society B: Biological Sciences, 278, 560-566.
HANSEN, V. L., SCHILKEY, F. D. & MILLER, R. D. 2016. Transcriptomic changes
associated with pregnancy in a marsupial, the gray short-tailed opossum Monodelphis
domestica. PLOS ONE, 11, e0161608.
HARRISON, G. A. & WEDLOCK, D. N. 2000. Marsupial cytokines structure, function and
evolution. Developmental & Comparative Immunology, 24, 473-484.
HAYNES, J. I. 2001. The marsupial and monotreme thymus, revisited. Journal of Zoology,
253, 167-173.
HEWAVISENTI, R. V., MORRIS, K. M., O'MEALLY, D., CHENG, Y., PAPENFUSS, A.
T. & BELOV, K. 2016. The identification of immune genes in the milk transcriptome
of the Tasmanian devil (Sarcophilus harrisii). PeerJ, 4, e1569.
HOBBS, M., PAVASOVIC, A., KING, A. G., PRENTIS, P. J., ELDRIDGE, M. D. B.,
CHEN, Z., COLGAN, D. J., POLKINGHORNE, A., WILKINS, M. R.,
FLANAGAN, C., GILLETT, A., HANGER, J., JOHNSON, R. N. & TIMMS, P.
2014. A transcriptome resource for the koala (Phascolarctos cinereus): insights into
koala retrovirus transcription and sequence diversity. BMC Genomics, 15, 786-786.
HOPSON, J. A. 1973. Endothermy, small size, and the origin of mammalian reproduction.
The American Naturalist, 107, 446-452.
HUBBARD, T. J. P., AKEN, B. L., BEAL, K., BALLESTER, B., CACCAMO, M., CHEN,
Y., CLARKE, L., COATES, G., CUNNINGHAM, F., CUTTS, T., DOWN, T.,
DYER, S. C., FITZGERALD, S., FERNANDEZ-BANET, J., GRAF, S., HAIDER,
S., HAMMOND, M., HERRERO, J., HOLLAND, R., HOWE, K., JOHNSON, N.,
KAHARI, A., KEEFE, D., KOKOCINSKI, F., KULESHA, E., LAWSON, D.,
LONGDEN, I., MELSOPP, C., MEGY, K., MEIDL, P., OUVERDIN, B., PARKER,
A., PRLIC, A., RICE, S., RIOS, D., SCHUSTER, M., SEALY, I., SEVERIN, J.,
SLATER, G., SMEDLEY, D., SPUDICH, G., TREVANION, S., VILELLA, A.,
VOGEL, J., WHITE, S., WOOD, M., COX, T., CURWEN, V., DURBIN, R.,
FERNANDEZ-SUAREZ, X. M., FLICEK, P., KASPRZYK, A., PROCTOR, G.,
SEARLE, S., SMITH, J., URETA-VIDAL, A. & BIRNEY, E. 2007. Ensembl 2007.
Nucleic Acids Research, 35, D610-D617.
IKONOMOPOULOU, M. P. & ROSE, R. W. 2006. The development of endothermy during
pouch life in the eastern barred bandicoot (Perameles gunnii), a marsupial.
Physiological and Biochemical Zoology, 79, 468-473.
JOHANSSON, J., AVESKOGH, M., MUNDAY, B. & HELLMAN, L. 2002. Heavy chain V
region diversity in the duck-billed platypus (Ornithorhynchus anatinus): long and
highly variable complementarity-determining region 3 compensates for limited
germline diversity. The Journal of Immunology, 168, 5155-5162.

39
JOHNSON, R. N., O’MEALLY, D., CHEN, Z., ETHERINGTON, G. J., HO, S. Y. W.,
NASH, W. J., GRUEBER, C. E., CHENG, Y., WHITTINGTON, C. M.,
DENNISON, S., PEEL, E., HAERTY, W., O’NEILL, R. J., COLGAN, D.,
RUSSELL, T. L., ALQUEZAR-PLANAS, D. E., ATTENBROW, V., BRAGG, J. G.,
BRANDIES, P. A., CHONG, A. Y.-Y., DEAKIN, J. E., DI PALMA, F., DUDA, Z.,
ELDRIDGE, M. D. B., EWART, K. M., HOGG, C. J., FRANKHAM, G. J.,
GEORGES, A., GILLETT, A. K., GOVENDIR, M., GREENWOOD, A. D.,
HAYAKAWA, T., HELGEN, K. M., HOBBS, M., HOLLELEY, C. E., HEIDER, T.
N., JONES, E. A., KING, A., MADDEN, D., GRAVES, J. A. M., MORRIS, K. M.,
NEAVES, L. E., PATEL, H. R., POLKINGHORNE, A., RENFREE, M. B., ROBIN,
C., SALINAS, R., TSANGARAS, K., WATERS, P. D., WATERS, S. A., WRIGHT,
B., WILKINS, M. R., TIMMS, P. & BELOV, K. 2018. Adaptation and conservation
insights from the koala genome. Nature Genetics, 50, 1102-1111.
JOHNSTONE, J. 1898. The thymus in the marsupials. Journal of the Linnean Society of
London, Zoology, 26, 537-557.
JOSS, J. L., MOLLOY, M. P., HINDS, L. & DEANE, E. 2009. A longitudinal study of the
protein components of marsupial milk from birth to weaning in the tammar wallaby
(Macropus eugenii). Developmental & Comparative Immunology, 33, 152-161.
JURD, R. D. 1994. "Not proper mammals": immunity in monotremes and marsupials.
Comparative Immunology, Microbiology and Infectious Diseases, 17, 41-52.
KIRSCH, J. A. W. 1977a. Bioloigcal aspects of the marsupial-placental dichotomy: a reply to
lillegraven. Evolution, 31, 898-900.
KIRSCH, J. A. W. 1977b. The six-percent solution: second thoughts on the adaptedness of
the marsupialia: features of their physiology and diversity suggest that marsupials
represent an alternative but not inferior kind of mammal, valuable in understanding
the courst of mammalian evolution. American Scientist, 65, 276-288.
KRAUS, D. B. & FADEM, B. H. 1987. Reproduction, development and phsiology of the
gray short-tailed opossum (Monodelphis domestica). The International Journal of
Laboratory Animal Science, 37, 487-482.
LEFÈVRE, C. M., DIGBY, M. R., WHITLEY, J. C., STRAHM, Y. & NICHOLAS, K. R.
2007. Lactation transcriptomics in the Australian marsupial, Macropus eugenii:
transcript sequencing and quantification. BMC genomics, 8, 417-417.
LOW, B. S. 1978. Environmental uncertainty and the parental strategies of marsupials and
placentals. The American Naturalist, 112, 197-213.
LUCERO, J. E., ROSENBERG, G. H. & MILLER, R. D. 1998. Marsupial light chains:
complexity and conservation of λ in the opssum Monodelphis domestica. The Journal
of Immunology, 161, 6724-6732.
MIKKELSEN, T. S., WAKEFIELD, M., AKEN, B., AMEMIYA, C., CHANG, J., DUKE
BECKER, S., GARBER, M., GENTLES, A. J., GOODSTADT, L., HEGER, A.,
JURKA, J., KAMAL, M., MAUCELI, E., SEARLE, S., SHARPE, T., BAKER, M.
L., BATZER, M. A., BENOS, P., BELOV, K. & LINDBLAD-TOH, K. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in non-coding
sequences. Nature 447, 167-77.
MILLER, R. D. & BELOV, K. 2000. Immunoglobulin genetics of marsupials.
Developmental & Comparative Immunology, 24, 485-490.
MILLER, R. D., GRABE, H. & ROSENBERG, G. H. 1998. VH repertoire of a marsupial
(Monodelphis domestica). The Journal of Immunology, 160, 259-365.
MILLER, W., HAYES, V. M., RATAN, A., C., P. D., WITTEKINDT, N. E., MILLER, J.,
WALENZ, B., KNIGHT, J., QI, J., ZHAO, F., WANG, Q., C., B.-R. O., KATIYAR,
N., TOMSHO, L. P., MCCLELLAN KASSON, L., R., H., WOODBRIDGE, P.,

40
TINDALL, E. A., FROST BERTELSEN, M., DIXON, D., PYECROFT, S.,
HELGEN, K. M., LESK, A. M., PRINGLE, T. H., PATTERSON, N., ZHANG, Y.,
KREISS, A., WOODS, G. M., JONES, M. E. & SCHUSTER, S. C. 2011. Genetic
diversity and population structure of the endangered marsupial Sarcophilus harrisii
(Tasmanian devil). Proceedings of the National Academy of Sciences, 108, 12348-
12353.
MOORE, H. D. 1996. Gamete biology of the new world marsupial, the grey short-tailed
opossum, Monodelphis domestica. Reproduction Fertility and Development, 8, 605-
615.
MORRIS, K. M., O'MEALLY, D., ZAW, T., SONG, X., GILLETT, A., MOLLOY, M. P.,
POLKINGHORNE, A. & BELOV, K. 2016. Characterisation of the immune
compounds in koala milk using a combined transcriptomic and proteomic approach.
Scientific Reports, 6, 35011-35011.
MORRIS, K. M., WEAVER, H. J., O'MEALLY, D., DESCLOZEAUX, M., GILLETT, A. &
POLKINGHORNE, A. 2018. Transcriptome sequencing of the long-nosed bandicoot
(Perameles nasuta) reveals conservation and innovation of immune genes in the
marsupial order peramelemorphia. Immunogenetics, 70, 327-336.
MORTAZAVI, A., WILLIAMS, B. A., MCCUE, K., SCHAEFFER, L. & WOLD, B. 2008.
Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nature Methods,
5, 621-8.
MORTON, S. R. 1978. An ecological study of Smithopsis crassicaudata (marsupialia:
dasyuridae). II. behaviour and social organisation. Australian Wildlife Research, 5,
163-182.
MURCHISON, E., SCHULZ-TRIEGLAFF, O., NING, Z., ALEXANDROV, L., BAUER,
M., FU, B., HIMS, M., DING, Z., IVAKHNO, S., STEWART, C., NG, B., WONG,
W., AKEN, B., WHITE, S., ALSOP, A., BECQ, J., BIGNELL, G., CHEETHAM, R.
K., CHENG, W., CONNOR, T., COX, A., FENG, Z.-P., GU, Y., GROCOCK, R.,
HARRIS, S., KHREBTUKOVA, I., KINGSBURY, Z., KOWARSKY, M., KREISS,
A., LUO, S., MARSHALL, J., MCBRIDE, D., MURRAY, L., PEARSE, A.-M.,
RAINE, K., RASOLONJATOVO, I., SHAW, R., TEDDER, P., TREGIDGO, C.,
VILELLA, A., WEDGE, D., WOODS, G., GORMLEY, N., HUMPHRAY, S.,
SCHROTH, G., SMITH, G., HALL, K., SEARLE, S. J., CARTER, N., PAPENFUSS,
A., FUTREAL, P. A., CAMPBELL, P., YANG, F., BENTLEY, D., EVERS, D. &
STRATTON, M. 2012. Genome sequencing and analysis of the Tasmanian devil and
its transmissible cancer. Cell, 148, 780-791.
NICHOLAS, K. R. 1988. Asynchronous dual lactation in a marsupial, the tammar wallaby
(Macropus eugenii). Biochemical and Biophysical Research Communications, 154,
529-536.
NILSSON, M. A., ARNASON, U., SPENCER, P. B. & JANKE, A. 2004. Marsupial
relationships and a timeline for marsupial radiation in South Gondwana. Gene, 340,
189-196.
NILSSON, M. A., CHURAKOV, G., SOMMER, M., VAN TRAN, N., ZEMANN, A.,
BROSIUS, J. & SCHMITZ, J. 2010. Tracking marsupial evolution using archaic
genomic retroposon insertions. PLoS biology, 8, e1000436.
O'CONNELL, M. A. 2007. American Opossums. Oxford University Press.
OLD, J. M. 2016. Haematopoiesis in marsupials. Developmental & Comparative
Immunology, 58, 40-46.
OLD, J. M. & DEANE, E. 2003. The detection of mature T- and B-cells during development
of the lymphoid tissues of the tammar wallaby (Macropus eugenii). Journal of
Anatomy, 203, 123-131.

41
OLD, J. M. & DEANE, E. M. 1998. The effect of oestrus and the presence of pouch young
on aerobic bacteria isolated from the pouch of the tammar wallaby, Macropus eugenii.
Comparative Immunology, Microbiology and Infectious Diseases, 21, 237-245.
OLD, J. M. & DEANE, E. M. 2000. Development of the immune system and immunological
protection in marsupial pouch young. Developmental & Comparative Immunology,
24, 445-454.
OLD, J. M., SELWOOD, L. & DEANE, E. M. 2003. Development of lymphoid tissues of the
stripe-faced dunnart (Sminthopsis macroura). Cells Tissues Organs, 175, 192-201.
OLD, J. M., SELWOOD, L. & DEANE, E. M. 2004a. The appearance and distribution of
mature T and B cells in the developing immune tissues of the stripe-faced dunnart
(Sminthopsis macroura). Journal of Anatomy, 205, 25-33.
OLD, J. M., SELWOOD, L. & DEANE, E. M. 2004b. A developmental investigation of the
liver, bone marrow and spleen of the stripe-faced dunnart (Sminthopsis macroura).
Developmental & Comparative Immunology, 28, 347-355.
PAINTER, M. W., DAVIS, S., HARDY, R. R., MATHIS, D., BENOIST, C. &
IMMUNOLOGICAL GENOME PROJECT, C. 2011. Transcriptomes of the B and T
lineages compared by multiplatform microarray profiling. Journal of Immunology,
186, 3047-57.
PAPENFUSS, A., HSU, A. & WAKEFIELD, M. 2010. Marsupial sequencing projects and
bioinformatics challenges. Marsupial Genetics and Genomics, 121-132.
PARKER, P. 1977. An ecological comparison of marsupial and placental patterns of
reproduction. The Biology of Marsupials. Springer.
PYECROFT, S., PEARSE, A.-M., LOH, R., SWIFT, K., BELOV, K., FOX, N., NOONAN,
E., HAYES, D., HYATT, A., WANG, L., BOYLE, D., CHURCH, J., MIDDLETON,
D. & MOORE, R. 2007. Towards a case definition for devil facial tumour disease:
what is it? EcoHealth, 4, 346-351.
RENFREE, M. B., PAPENFUSS, A. T., DEAKIN, J. E., LINDSAY, J., HEIDER, T.,
BELOV, K., RENS, W., WATERS, P. D., PHARO, E. A., SHAW, G., WONG, E. S.
W., LEFÈVRE, C. M., NICHOLAS, K. R., KUROKI, Y., WAKEFIELD, M. J.,
ZENGER, K. R., WANG, C., FERGUSON-SMITH, M., NICHOLAS, F. W.,
HICKFORD, D., YU, H., SHORT, K. R., SIDDLE, H. V., FRANKENBERG, S. R.,
CHEW, K. Y., MENZIES, B. R., STRINGER, J. M., SUZUKI, S., HORE, T. A.,
DELBRIDGE, M. L., PATEL, H., MOHAMMADI, A., SCHNEIDER, N. Y., HU, Y.,
O'HARA, W., AL NADAF, S., WU, C., FENG, Z.-P., COCKS, B. G., WANG, J.,
FLICEK, P., SEARLE, S. M. J., FAIRLEY, S., BEAL, K., HERRERO, J., CARONE,
D. M., SUZUKI, Y., SUGANO, S., TOYODA, A., SAKAKI, Y., KONDO, S.,
NISHIDA, Y., TATSUMOTO, S., MANDIOU, I., HSU, A., MCCOLL, K. A.,
LANSDELL, B., WEINSTOCK, G., KUCZEK, E., MCGRATH, A., WILSON, P.,
MEN, A., HAZAR-RETHINAM, M., HALL, A., DAVIS, J., WOOD, D.,
WILLIAMS, S., SUNDARAVADANAM, Y., MUZNY, D. M., JHANGIANI, S. N.,
LEWIS, L. R., MORGAN, M. B., OKWUONU, G. O., RUIZ, S. J., SANTIBANEZ,
J., NAZARETH, L., CREE, A., FOWLER, G., KOVAR, C. L., DINH, H. H., JOSHI,
V., JING, C., LARA, F., THORNTON, R., CHEN, L., DENG, J., LIU, Y., SHEN, J.
Y., SONG, X.-Z., EDSON, J., TROON, C., THOMAS, D., STEPHENS, A., YAPA,
L., LEVCHENKO, T., GIBBS, R. A., COOPER, D. W., SPEED, T. P., FUJIYAMA,
A., M GRAVES, J. A., et al. 2011. Genome sequence of an Australian kangaroo,
Macropus eugenii, provides insight into the evolution of mammalian reproduction and
development. Genome Biology, 12, 414-414.

42
ROUSMANIERE, H., SILVERMAN, R., WHITE, R., SASAKI, M., WILSON, S.,
MORRISON, J. & CRUZ, Y. 2010. Husbandry of Monodelphis domestica in the
study of mammalian embryogenesis. Lab Animal, 39, 219-226.
ROWE, T., RICH, T. H., VICKERS-RICH, P., SPRINGER, M. & WOODBURNE, M. O.
2008. The oldest platypus and its bearing on divergence timing of the platypus and
echidna clades. Proceedings of the National Academy of Sciences, 105, 1238.
ROWLANDS, D. T., LA VIA, M. F. & BLOCK, M. H. 1964. The blood forming tissues and
blood of the newborn opossum (Didelphys virginiana) II. ontogenesis of antibody
formation to flagella of salmonella typhi. The Journal of Immunology, 93, 157-164.
RUSSELL, E. M. 1982. Patterns of parental care and parental investment in marsupials.
Biological Reviews, 57, 423-486.
SAMOLLOW, P. B. 2006. Status and applications of genomic resources for the gray, short-
tailed opossum, Monodelphis domestica, an American marsupial model for
comparative biology. Australian Journal of Zoology, 54, 173-196.
SAMOLLOW, P. B. 2008. The opossum genome: insights and opportunities from an
alternative mammal. Genome Research, 18, 1199-1215.
SELWOOD, L. & COULSON, G. 2006. Marsupials as models for research. Australian
Journal of Zoology, 54, 137-138.
SETTE, A., FLERI, W., PETERS, B., SATHIAMURTHY, M., BUI, H.-H. & WILSON, S.
2005. A roadmap for the immunomics of category A–C pathogens. Immunity, 22,
155-161.
STANLEY, N. F., YADAV, M., WARING, H. & EADIE, M. 1972. The effect of
thymectomy on response to various antigens of a marsupial Setonix brachyurus
(quokka). Immunology and Cell Biology, 50, 689.
STONE, W., BRUUN, D., MANIS, G., HOLSTE, S., HOFFMAN, E., SPONG, K. &
WALUNAS, T. 1996. The immunobiology of the marsupial, Monodelphis domestica.
Modulators of Immune Responses, The Evolutionary Trail, Breckenridge Series, 2.
SZALAY, F. S. 2006. Evolutionary history of the marsupials and an analysis of osteological
characters, Cambridge University Press.
TOUNG, J. M., MORLEY, M., LI, M. & CHEUNG, V. G. 2011. RNA-sequence analysis of
human B-cells. Genome Research, 21, 991-8.
TRUPIN, G. L. & FADEM, B. H. 1982. Sexual behavior of the gray short-tailed opossum
(Monodelphis domestica). Journal of Mammalogy, 63, 409-414.
TYNDALE-BISCOE, H. & RENFREE, M. 1987. Reproductive physiology of marsupials,
Cambridge University Press.
WANG, Z., GERSTEIN, M. & SNYDER, M. 2009a. RNA-Seq: a revolutionary tool for
transcriptomics. Nature Reviews Genetics, 10, 57-63.
WANG, Z., HUBBARD, G. B., CLUBB, F. J. & VANDEBERG, J. L. 2009b. The laboratory
opossum (Monodelphis domestica) as a natural mammalian model for human cancer
research. International Journal of Clinical and Experimental Pathology, 2, 286-299.
WONG, E. S., PAPENFUSS, A. T. & BELOV, K. 2011a. Immunome database for
marsupials and monotremes. BMC Immunology, 12, 48.
WONG, E. S., PAPENFUSS, A. T., HEGER, A., HSU, A. L., PONTING, C. P., MILLER, R.
D., FENELON, J. C., RENFREE, M. B., GIBBS, R. A. & BELOV, K. 2011b.
Transcriptomic analysis supports similar functional roles for the two thymuses of the
tammar wallaby. BMC Genomics, 12, 420.
WONG, E. S., YOUNG, L. J., PAPENFUSS, A. T. & BELOV, K. 2006. In silico
identification of opossum cytokine genes suggests the complexity of the marsupial
immune system rivals that of eutherian mammals. Immunome Research, 2, 4.

43
YADAV, M. & EADIE, M. 1972. Passage of maternal immunoglobulins to the pouch young
of a marsupial Setonix brachyurus. Australian Journal of Zoology, 21, 171-81.
YADAV, M., STANLEY, N. & WARING, H. 1972. The thymus glands of a marsupial,
Setonix brachyurus (quokka), and their role in immune responses: effect of
thymectomy on somatic growth and blood leucocytes. Australian Journal of
Experimental Biology and Medical Science, 50, 357-364.
YOUNG, L., BASDEN, K., COOPER, D. & DEANE, E. 1997. Cellular components of the
milk of the tammar wallaby (Macropus eugenii). Australian Journal of Zoology, 45,
423-433.
ZELLER & FREYER 2001. Early ontogeny and placentation of the grey short-tailed
opossum, Monodelphis domestica (Didelphidae: Marsupialia): contribution to the
reconstruction of the marsupial morphotype. Journal of Zoological Systematics and
Evolutionary Research, 39, 137-158.
ZELUS, D., ROBINSON-RECHAVI, M., DELACRE, M., AURIAULT, C. & LAUDET, V.
2000. Fast evolution of interleukin-2 in mammals and positive selection in ruminants.
Journal of Molecular Evolution, 51, 234-244.

44
CHAPTER 2
Immunogenetics of Marsupial B-cells
Andrea L. Schraven1, Hayley J. Stannard2, Oselyne T.W. Ong3, Julie M. Old1
1School of Science and Health, Hawkesbury Campus, Western Sydney University, Locked
bag 1797, Penrith, NSW 2751, Austral00ia
2Charles Sturt University, School of Animal and Veterinary Sciences, Wagga Wagga, NSW
2678, Australia
3QIMR Berghofer Medical Research Institute, Brisbane, Queensland, Australia
Molecular Immunology; 2020, vol. 117, pp. 1-11

45
2.1 Chapter Outline and Authorship
Chapter 2 provides a detailed literature review on the knowledge of marsupial B-cell genetics
and biology. It discusses the similarities and differences between marsupial and eutherian B-
cells, in regard to genetic presence, homology, and developmental stages, as well as
highlights the areas requiring further research.
This chapter has been published in Molecular Immunology, and the manuscript is formatted
as per journal specifications. The manuscript is jointly authored; I am the primary author. I
collected and analysed the data and wrote the manuscript. Hayley Stannard, Oselyne Ong,
and Julie Old reviewed the manuscript and provided feedback.
Published version available as:
Schraven, A. L., Stannard, H. J., Ong, O. T. W., Old, J. M. 2020. Immunogenetics of
marsupial B-cells. Molecular Immunology. 117, 1-11.
https://doi.org/10.1016/j.molimm.2019.10.024

46
2.2 Introduction
The gray short-tailed opossum (Monodelphis domestica) was the first marsupial
species to have its genome sequenced and available through the Broad Institute and NCBI
Genbank (Mikkelsen et al., 2007). After the release of the gray short-tailed opossum genome,
the sequencing of other marsupials, in particular Australian marsupials, soon followed,
including the tammar wallaby (Notamacropus eugenii) (Renfree et al., 2009), Tasmanian
devil (Sarcophilus harrisii) (Murchison et al., 2012) and koala (Phascolartos cinereus)
(Hobbs et al., 2014). The opossum genome was Sanger-sequenced to ˜7x coverage or draft
quality, producing a highly usable assembly, reflected by the large contig N50 of 108 kb and
scaffold N50 of 60 mb (Papenfuss et al., 2010). However, at the time, the Ensembl
Consortium had no large-scale transcriptomic data available for the annotation of the species
(Papenfuss et al., 2010), and consequently, the annotation was based largely on human
(Homo sapiens) transcripts and protein sequences (Hubbard et al., 2007). Despite the
sequencing of large amounts of ‘good quality’ conserved genes, many genes predominately
involved in immune responses were not identified and/or annotated due to the high rate of
divergence (Wong et al., 2011a). Immune genes, for the most part, succumb to intense
selective pressure due to the requirements of forever evolving pathogens (Zelus et al., 2000),
and for this reason, the Ensembl Consortium began annotating the immune genes of the gray
short-tailed opossum. However, the majority of marsupial genes are yet to be identified
through expression and have therefore only been predicted by Ensembl (Curwen et al., 2004,
Wong et al., 2006, Belov et al., 2006). With recent bioinformatic advancements, in particular
genome annotation, researchers now have the ability to understand the immunocompetence of
different mammalian species through cell-mediated and humoral immune response
investigations.

47
To date, researchers have detected various genes and processes, including
immunoglobulins (Ig), T-cell receptors, and MHC genes, suggesting the marsupial immune
system is highly similar and just as complex as their eutherian counterparts using traditional
molecular techniques (Samollow, 2006). Responses to graft rejection, an example of a cell
mediated response have been investigated in the Virginian (Didelphis virginiana) and gray
short-tailed opossums, tammar wallaby and quokka (Setonix brachyurus) (Laplante et al.,
1969). In the Virginian opossum, pouch young accept skin grafts up to day 12 postpartum;
after this time, the rejection of tissue is apparent and has been suggested to occur as a result
of T-cell responses (Laplante et al., 1969). For humoral responses, Igs have been measured in
the serum of the Virginian opossum (Rowlands et al., 1964). Rowlands et al. (1964) found
that Igs could not be detected in the serum until after exposure to a bacterial antigen at day 8–
15, and Kalmutz (1962) only detected an Ig response at day 17 to viral antigens. Whilst
techniques used in immunochemistry have allowed investigators to detect tammar wallaby
CD79b+ lymphocytes at day 7 postpartum (Old and Deane, 2003).
In eutherians, B-cells are important for humoral immunity as their genes related to
immunity undergo vigorous developmental, differentiation, regulation, and functional
processes and checkpoints to enable them to mediate the production of antigen-specific
antibodies in the presence of foreign molecules. In contrast to eutherians, little is known
about marsupial B-cells due to the lack of investigation into the marsupial genome. For this
reason, the purpose of this paper is to review what is already known in regards to marsupial
B-cells, with a comparison to eutherian studies. Analyses of B-cell genes have the potential to
shine light on marsupial humoral immune responses as well as contribute to comparative
evolutionary studies of B-cells.

48
2.3 Mammalian Evolution
Mammals arose ˜200 million years ago (Archibald and Deutschman, 2001, Bininda-
Emonds et al., 2007) and are classified into three lineages; eutherians, marsupials
(metatherians), and monotremes (prototherians). Monotremes were the first to diverge ˜166
million years ago and exhibit both reptilian and mammalian features (Renfree et al., 2009).
Marsupials then separated from eutherians ˜160 million years ago (Kumar and Hedges,
1998). Both eutherians and marsupials give birth to live young, while monotremes are ‘egg
laying’ mammals (Warren et al., 2008). The major difference between eutherians and
marsupials is while eutherians are born fully developed after a long gestational period (e.g.
humans have nine month gestation period), marsupials have a shorter gestation period and are
born with no mature lymphoid tissues (Deane and Cooper, 1988, Block, 1964, Tyndale-
Biscoe and Renfree, 1987). The survival of the marsupial is heavily dependent on the young
having the ability to obtain passive acquired immunity from its mother. At birth marsupials
are considered to be at the same physical and physiological stage of development as a nine-
week-old human foetus (Block, 1960). Therefore, while marsupials continue developing
externally, eutherians are still undergoing prenatal development at a similar life stage (Wang
et al., 2012b). Marsupial pouch young can range from 3 mg (honey possum, Tarsipes
rostratus) to 828 mg (western grey kangaroo, Macropus fuliginosus) (Tyndale-Biscoe and
Renfree, 1987) at birth, with no developed hind legs (Graves and Westerman, 2002).
Newborn marsupials only have partially developed forelimbs to aid their upwards climb from
the vaginal opening to their mother’s teat, where they undergo a lengthy lactation period to
complete their physical and immunological development (Tyndale-Biscoe and Renfree,
1987).
For many decades our understanding of the marsupial immune system has lagged
behind that of the eutherian immune system (Morris et al., 2010) although there is evidence

49
that suggests the lymphoid tissues and cells examined in marsupials are highly similar to
eutherians (Ashman and Papadimitriou, 1975, Old and Deane, 2000, Wong et al., 2006,
Borthwick and Old, 2016). In the past, it was thought that marsupials (and monotremes) were
only evolutionary steps in the progression to eutherians and were therefore inferior (Jurd,
1994). However there is now evidence suggesting that marsupials and eutherians have similar
lymphoid tissues and cells (Block, 1964), and their immune system presumably functions in a
similar manner (Block, 1964, Deane and Cooper, 1984, Graves and Westerman, 2002, Old,
2016, Wong et al., 2006). In addition, the knowledge that marsupials give birth to
underdeveloped young could expand our knowledge on the evolution of developmental
immunity (Old and Deane, 2000), including defence mechanisms used via an adaptive
immune response (Edwards et al., 2012).
2.4 Marsupial Immunology
Marsupial haematopoiesis commences in the primitive yolk sac in utero, which
continues in the liver (postpartum) until the bone marrow and thymus have matured (Old,
2016). This process is already complete in newborn eutherians, where the haematopoietic
stem cells (HSCs) undergo development and maturation in the fetal liver and give rise to B-
cells prior to moving to the bone marrow in the last stages of foetal development (Doulatov et
al., 2012, Hadland and Yoshimoto, 2018). The liver has been identified as the major site for
haematopoietic activity for the first few weeks postpartum (Borthwick et al., 2014), and is
similar in the pattern of development as that in eutherians despite occurring prenatally in
eutherians. After this time, the liver ceases its involvement in haematopoiesis and the bone
marrow and thymus take over the role (Borthwick et al., 2014). Different studies have found
that both granulocyte and erythrocyte lineages were observed in the liver, day 1 postpartum
of the quokka, stripe-faced dunnart (Sminthopsis macroura) and tammar wallaby (Ashman

50
and Papadimitriou, 1975, Basden et al., 1996, Old et al., 2004). On day 3, the quokka’s
haematopoietic cells have started to aggregate and form islands throughout the liver (Old et
al., 2004), while on day 4, the northern brown bandicoot (Isoodon macrourus) liver has a
high abundance of erythrocytes and erythrocyte precursor islands (Cisternas and Armati,
1999). After the first week postpartum, haematopoiesis rapidly decreases in the liver of the
Virginian opossum (Didelphis virginiana) (Cutts et al., 1973) and northern brown bandicoot
(Cisternas and Armati, 1999). Over the next few weeks to months, haematopoiesis continues
to decrease, and the tammar wallaby and quokka are observed to have a few sparse
haematopoietic islands during development by the end of the third month (Ashman and
Papadimitriou, 1975, Basden et al., 1996).
Lymphoid tissues and cell types develop at different stages in marsupials from the
time of birth (Belov et al., 2013). The thymus is the first of the lymphoid tissues to develop
and mature in marsupials, and is also the case for humans where the thymus is responsible for
the maturation of T-cells (Wong et al., 2011b). The thymus has been identified in all
marsupials studied, but not all species investigated have both the thoracic and cervical
thymus, most only have the former (Old, 2016). For those marsupials that have both a
thoracic and cervical thymus, the cervical thymus develops earlier and is considerably larger
in size, however both perform the same function (Stanley et al., 1972, Wong et al., 2011b).
Block (1964) detailed the histological development of the thymus in the Virginian opossum,
observing on day 1 postpartum a single sheet of epithelial cells and between day 23 and 32
postpartum a fully functional thymus. The timing of bone marrow maturation varies across
different marsupial species (Old, 2016). The tammar wallaby has been observed to develop
early formation of erythrocytes and granulocytes on day 4, by day 8 megakaryocytes were
apparent, and by day 14, increased numbers and the first stages of diversification of
haematopoietic cells were observed (Old, 2016).

51
The spleen is the last of the lymphoid tissues to mature (Old, 2016), where
mesenchymal cells are the only representation of the spleen for the first couple of days
postpartum for the Virginian opossum (Cutts and Krause, 1982), tammar wallaby (Basden et
al., 1996), and quokka (Ashman and Papadimitriou, 1975). Haematopoietic activity slowly
increases over the next few weeks and by the end of the first month, erythroblastic
haematopoiesis is prominent in the quokka (Ashman and Papadimitriou, 1975). As for the
Virginian opossum, the appearance of white pulp increases exponentially in contrast to the
red pulp (Block, 1964). The spleen continues to undergo erythropoiesis through adulthood
and into old age of the Virginian opossum (Cutts and Krause, 1982, Old, 2016).
2.5 B-cells
B-cells are of particular importance in the immune system as they are found in
multiple lymphoid organs (Holmes et al., 2008), and are classified as an adaptive humoral
immune cell type (MacConmara and Lederer, 2005). They are able to detect foreign
molecules that invade the body through the production of antibodies released from their
differentiated cells (Painter et al., 2011). In mice (Mus musculus), B-cells are produced from
the pluripotent haematopoietic stem cells which are found in the liver during the late stages of
foetal development and continues development in the bone marrow after birth (Hardy and
Hayakawa, 2001). The appearance of marsupial B-cell development is similar to eutherians
except B-cells are still produced in the liver postpartum until the bone marrow fully matures
(Borthwick et al., 2014, Old, 2016).
Many genes are specifically expressed in B-cells, throughout multiple stages of B-cell
development, differentiation, and regulation. Signature genes associated with the B-cell
include Igs (heavy and light chains) (Palmer et al., 2006), cell surface markers (e.g. CD79α/β)
(Deneys et al., 2001), signal transduction molecules (e.g. BTK, Lyn, and Syk) (Kurosaki,
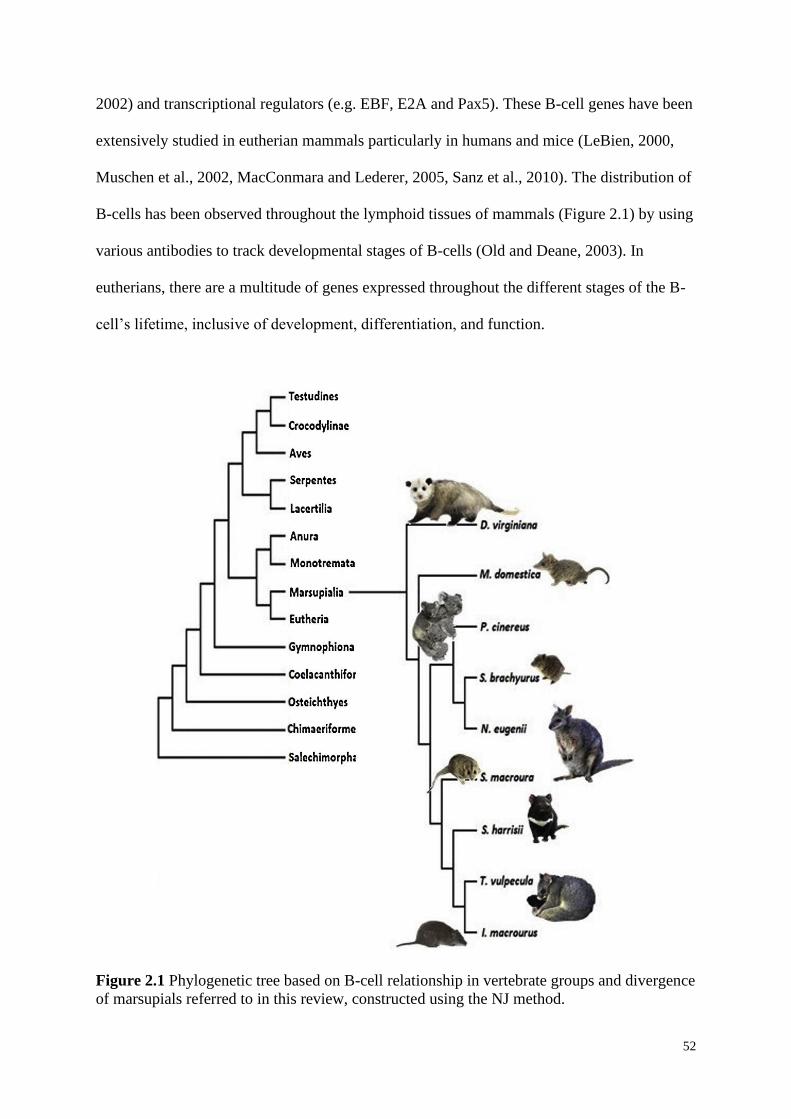
52
2002) and transcriptional regulators (e.g. EBF, E2A and Pax5). These B-cell genes have been
extensively studied in eutherian mammals particularly in humans and mice (LeBien, 2000,
Muschen et al., 2002, MacConmara and Lederer, 2005, Sanz et al., 2010). The distribution of
B-cells has been observed throughout the lymphoid tissues of mammals (Figure 2.1) by using
various antibodies to track developmental stages of B-cells (Old and Deane, 2003). In
eutherians, there are a multitude of genes expressed throughout the different stages of the B-
cell’s lifetime, inclusive of development, differentiation, and function.
Figure 2.1 Phylogenetic tree based on B-cell relationship in vertebrate groups and divergence
of marsupials referred to in this review, constructed using the NJ method.

53
In eutherians, developing B-cells are in constant contact with stromal cells that
facilitate their development. Stromal cells produce interleukin (IL)-7 to support the
proliferative expansion of progenitor (pro-) B-cells (Bagwell et al., 2015). Pro-B-cells are the
earliest of the B-cell developmental stages and express the IL-7 receptor, CD43, c-Kit, B220,
BP1, and terminal deoxynucleotide transferase (TdT). During this developmental stage, the Ig
heavy chain undergoes a genetic recombination called V(D)J (somatic) recombination where
variable (V), joining (J), and sometimes diversity (D) gene segments are rearranged to allow
for the Igs to adhere to the antigen, allowing for an adaptive immune response (Li et al.,
2004, Schatz and Swanson, 2011). Igs are glycoproteins constituting four polypeptide chains:
two identical light and heavy chains which are linked together by disulphide bonds and
noncovalent forces (Painter, 1998), and primarily involved in secondary immune responses.
Early pro-B-cells undergo D–J rearrangement while late pro-B-cell undergoes V-D–J
rearrangement. Eutherian B-cell development then transitions from a pro-B-cell to a
precursor (pre-) B-cell stage where chances of apoptosis are its highest. At this time, the Ig
heavy chain is expressed at the cell surface with the surrogate light chain, known as the pre-
B-cell receptor (pre-BCR), to determine if the Ig heavy chain has undergone a successful
recombination (Bagwell et al., 2015).
The maturation of eutherian B-cells is thought to be similar to marsupials and
involves several stages (Conley, 2003) defined by the rearrangement of different Igs,
expression of cluster of differentiation markers (CD), and location of the cells within the
bone marrow, spleen, and/or lymph nodes (Conley and Cooper, 1998). The few B-cell genes
that have been examined in marsupials, for example the gray short-tailed opossum,
demonstrate that a B-cell dependent immune response can only be initiated after the first
postnatal week, as all genes necessary to elicit an adaptive immune response have not yet

54
been expressed (Table 2.1) (Wang et al., 2012b). Marsupial B-cell dependent immune
responses are correlated with the appearance of cells expressing mature lymphocyte-specific
markers (Old and Deane, 2000, Duncan et al., 2010, Wang et al., 2012c). While developing
B-cells have been detected in late stage prenatal embryos, followed by B-cell maturation,
which occurs exclusively postnatal in the opossum (Wang et al., 2012c). Despite having the
knowledge of when specific B-cell stages are present, it remains unknown if these Ig
repertoire changes in marsupial B-cells are analogous to eutherians.
Table 2.1 List of B-cell genes that have either been expressed or predicted in marsupial
species, including their Genbank accession Identifications (ID). Gene Species Expressed/Predicted Genbank
accession ID
Expression Reference
CD10 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007502344.1
XM_020988925.1
XM_012546630.2
XM_027876124.1
CD19 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_016423695.1
XM_021006522.1
XM_023497850.1
XM_027862685.1
CD20 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001379088.3
XM_020976111.1
XM_003774020.3
XM_027858851.1
CD21 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007481319.2
XM_021003160.1
XM_023504041.1
XM_027871517.1
CD22 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001369696.3
XM_020977007.1
XM_012545693.2
XM_027855546.1
55
CD32 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007480653.2
XM_020988655.1
XM_012547618.2
XM_027873972.1
CD34 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007481323.2
XM_021007822.1
XM_003767657.3
XM_027871708.1
CD38 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007496701.2
XM_020992954.1
XM_012553075.2
XM_027836551.1
CD72 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007498802.1
XM_020977042.1
XM_023497642.1
XM_027839551.1
CD79a
(IgA)
M. domestica
P. cinereus
S. harrisii
V. ursinus
N. eugenii
O. fraenata
Expressed
Predicted
Predicted
Predicted
Expressed
Expressed
ACZ13436.1
XM_021004022.1
XM_023500926.1
XM_027858134.1
ACZ13437.1
AGO64767.1
(Duncan et al., 2010)
(Duncan et al., 2010)
(Suthers and Young, 2014)
CD79b M. domestica
P. cinereus
V. ursinus
N. eugenii
O. fraenata
Expressed
Predicted
Predicted
Expressed
Expressed
ACZ13438.1
XM_020997545.1
XM_027872889.1
ACZ13439.1
AII23405.1
(Duncan et al., 2010)
(Duncan et al., 2010)
(Suthers and Young, 2014)
IgG M. domestica
N. eugenii
T. vulpecula
Expressed
Expressed
Expressed
AF098643.1
EF599608.1
AF157619.1
(Wang et al., 2009)
(Daly et al., 2007)
(Belov et al., 1999a)
IgM M. domestica Expressed AF012109.2 (Miller et al., 1998)
IgE M. domestica
N. eugenii
T. vulpecula
Expressed
Expressed
Expressed
AF035194.1
EF599607.1
AF157617.1
(Aveskogh and Hellman, 1998)
(Daly et al., 2007)
(Belov et al., 1999b)
56
IGκ M. domestica
S. harrisii
N. eugenii
T. vulpecula
Expressed
Expressed
Expressed
Expressed
AAF25575.1
ALB75225.1
ABV02004.1
AAL17619.1
(Wang et al., 2012c)
(Ujvari and Belov, 2015)
(Daly et al., 2007)
(Belov et al., 2001)
IGλ M. domestica
P. cinereus
N. eugenii
T. vulpecula
Expressed
Predicted
Expressed
Expressed
AAC98628.1
XP_020834443.1
ABV02011.1
AAL37211.1
(Lucero et al., 1998)
(Daly et al., 2007)
(Belov et al., 2002b)
Pax5 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007497997.2
XM_021004585.1
XM_003761344.1
XM_027859120.1
POU2AF1 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007494855.2
XM_020989802.1
XM_023501417.1
XM_027847318.1
SPIB P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
XM_020971799.1
XM_023506369.1
XM_027849772.1
SPI1 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001370178.3
XM_003773728.3
XM_012552632.2
XM_027837740.1
Pu.1 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001370178.3
XM_021009704.1
XM_003773728.3
XM_027837740.1
E2A M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_016432215.1
XM_021009149.1
XM_012542196.2
XM_027853230.1
EBF1 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007473955.2
XM_020992364.1
XM_023505848.1
XM_027841851.1
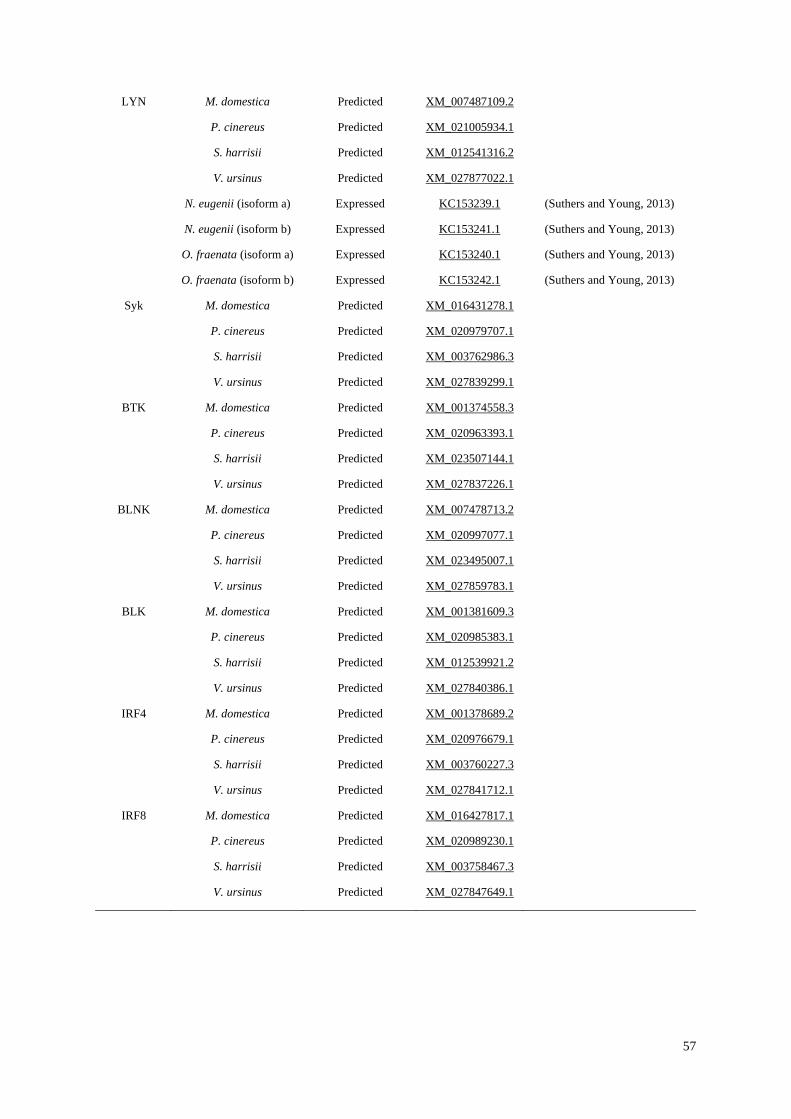
57
LYN M. domestica
P. cinereus
S. harrisii
V. ursinus
N. eugenii (isoform a)
N. eugenii (isoform b)
O. fraenata (isoform a)
O. fraenata (isoform b)
Predicted
Predicted
Predicted
Predicted
Expressed
Expressed
Expressed
Expressed
XM_007487109.2
XM_021005934.1
XM_012541316.2
XM_027877022.1
KC153239.1
KC153241.1
KC153240.1
KC153242.1
(Suthers and Young, 2013)
(Suthers and Young, 2013)
(Suthers and Young, 2013)
(Suthers and Young, 2013)
Syk M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_016431278.1
XM_020979707.1
XM_003762986.3
XM_027839299.1
BTK M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001374558.3
XM_020963393.1
XM_023507144.1
XM_027837226.1
BLNK M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_007478713.2
XM_020997077.1
XM_023495007.1
XM_027859783.1
BLK M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001381609.3
XM_020985383.1
XM_012539921.2
XM_027840386.1
IRF4 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_001378689.2
XM_020976679.1
XM_003760227.3
XM_027841712.1
IRF8 M. domestica
P. cinereus
S. harrisii
V. ursinus
Predicted
Predicted
Predicted
Predicted
XM_016427817.1
XM_020989230.1
XM_003758467.3
XM_027847649.1

58
2.5.1 Immunoglobulins
Early studies of the marsupial immune system included measurements of antibody
responses and comparing these levels of response to those of eutherians (Kalmutz, 1962).
Primary immune responses of marsupials are very similar to eutherians, however the
secondary immune responses are delayed and less effective than that of eutherian responses
(Kalmutz, 1962, Yadav and Eadie, 1972). In comparison to eutherians and monotremes,
marsupial B-cells have four Ig heavy (H) loci isotypes (IgA, IgG, IGM and IgE), defined by
their constant regions (Cα, Cγ, Cμ and Cε respectively) (Belov et al., 1999c). So far, multiple
Ig heavy chain genes, IgA, IgG, IgM and IgE inclusive, have been observed in a number of
marsupial species including the Virginian opossum (Rowlands and Dudley, 1968), quokka
(Bell et al., 1974b), common brushtail possum (Trichosurus vulpecula) (Ramadass and
Moriarty, 1982), koala (Aveskogh and Hellman, 1998) and the gray short-tailed opossum
(Aveskogh and Hellman, 1998). Interestingly, IgD described in eutherians and the platypus
(Ornithorhynchus anatinus) has not been identified in any marsupial species to date (Figure
2.2) (Aveskogh and Hellman, 1998, Aveskogh et al., 1999). Homologous-based searches in
the regions where IgD is expected to be located have been unsuccessful in the gray short-
tailed opossum (Wang et al., 2009), despite large duplications and insertions of a non-
functional copy of IgM being identified (Wang et al., 2009). The internal duplications of this
downregulated IgM may have contributed to the loss of the IgD gene in the gray short-tailed
opossum and possibly all modern marsupial species. IgD is still present in most eutherians,
monotremes and some non-mammalian species (Figure 2.2) suggesting this Ig isotype is
evolutionarily ancient (Ohta and Flajnik, 2006).

59
Figure 2.2 Unrooted phylogenetic tree based on amino acid alignments of vertebrate IgD
sequences, constructed using the NJ method. Genbank accession numbers: I. punctatus IgD,
ADF56020; S. trutta IgD, AAV28076; E. coioides IgD, AEN71107; M. musculus IgD,
AAB59653; X. tropicalis IgD, ABC75541; P. sinensis IgD, ACU45375; O. anatinus IgD,
ACD31540; C. siamensis IgD, AFZ39208; E. macularius IgD, ABY67439; H. sapiens IgD,
AAA52771; P. troglodytes IgD, PNI88328; S. scrofa IgD, AAP23939; B. taurus IgD,
AAN03673; O. aries IgD, AAN03671.
In all mammals, IgA is passively transferred from the mammary gland epithelium via
the colostrum to the young (during suckling), which provides protection in the mucus
membranes of both the respiratory and gastrointestinal systems of the newborn (Adamski and
Demmer, 1999). In eutherians, the mammary gland epithelium lymphocytes secrete IgA as a
dimer joined by the J chain (Koshland, 1985), in turn IgA is transported across the epithelial

60
cells in the milk by the polymeric Ig receptor (pIgR) and released by proteolytic cleavage of
the extracellular domain at the apical membrane (Mostov, 1994). sIgA was first identified in
marsupial milk and other secretory sites of the quokka (Bell et al., 1974b). Since then, IgA
has been isolated from the colostrum of the Virginian opossum (Hindes and Mizell, 1976)
and common brushtail possum (Ramadass and Moriarty, 1982). In the common brushtail
possum, Adamski and Demmer (1999) have reportedly cloned and characterised the J chain
and pIgR cDNA sequences and found these genes to be highly conserved when compared to
those of eutherians. The expression studies found both the J chain and pIgR had increased in
expression levels at two distinct time periods throughout the lactation period of the pouch
young and would concur with the time of exposure to a potential new pathogen (Adamski and
Demmer, 1999). The first IgA enhancement being at the beginning of lactation is consistent
with eutherians, however the second increase occurs at the end of the switch phase (80–120
days of lactation) and is unique to marsupials (Adamski and Demmer, 1999).
IgG makes up 80 % of serum antibody levels (Smith et al., 2019). Like IgA, IgG plays
a major role in respiratory protection, however IgG primarily acts to prevent local and
systemic infections in the lower rather than the upper respiratory tract (Stokes and
Rajasekaran, 2008). IgG has been isolated from the foetal serum of the tammar wallaby two
days prior to birth (Renfree, 1973), and again detected in the blood of pouch young, as well
as maternal colostrum and placental extracts (Deane et al., 1990), suggesting it can be
transferred transplacentally in this species. It is apparent that separate gene duplication events
have occurred in IgA and IgG isotypes in some mammalian species. In eutherians, humans
and mice have four functional IgG subclasses; humans have IgG1, IgG2, IgG3, and IgG4
(Vidarsson et al., 2014), whereas mice have IgG1, IgG2a/c, IgG2b, and IgG3 in which they
either present IgG2a or IgG2b depending on their strain (Zhang et al., 2012). Humans also
have two functional IgA subclasses, IgA1 and IgA2 with a higher preference for IgA1 in

61
lymphoid tissues (Kett et al., 1986). These gene duplications seem to have evolved quite
recently and independently within individual mammalian lineages as no IgA nor IgG
subclasses have been identified in any marsupial or monotreme (not shown) species, and not
all eutherians (Figure 2.3).
Figure 2.3 Unrooted phylogenetic tree based on amino acid alignments of marsupial and
eutherian IgA and IgG subclasses sequences, constructed using the NJ method. Genbank
accession numbers: M. domestica IgG, AAC79675; T. vulpecula IgG, AAD55590; M.
domestica IgA, AAD21190; T. vulpecula IgA, AAD41690; N. eugenii IgA, ABV01995; M.
musculus IgA, AAB59662; H. sapiens IgA2, AAB59396; H. sapiens IgA1, AAC82528; H.
sapiens IgG1, BAC87418; H. sapiens IgG3, CAA27268; H. sapiens IgG2, AAB59393; H.
sapiens IgG4, CAC20457; M. musculus IgG1, CAD32497; M. musculus IgG3, ADK11691;
M. musculus IgG2b, ACX70084; M. musculus IgG2a, BAC44883; M. musculus IgG2c,
CAA34864.

62
IgM was the first of the Igs to appear in vertebrate evolution and is the first Ig to be
expressed during development of the immune system (Lawton et al., 1975). Membrane bound
IgM (mIgM) has a monomer structure which resides on the B-cell surface and acts upon the
presence of an antigen, whilst serum IgM (sIgM) is a pentamer and aids in the activation of
the complement system (Atwell and Marchalonis, 1977). In marsupials, studies of IgM have
mostly been limited to its initial characterisation including those in the Virginian opossum
(Rowlands and Dudley, 1968), quokka (Bell et al., 1974a), common brushtail possum
(Ramadass and Moriarty, 1982), koala (Wilkinson et al., 1991), and gray short-tailed
opossum (Shearer et al., 1995). Belov et al. (2002a) has also detected transcripts of IgM in
the spleen and liver as early as day 10 postpartum of the common brushtail possum which is
similarly expressed in humans at the same developmental stage (Cooper, 1987).
IgE is found in much lower concentrations in comparison to the other Ig classes,
however it plays a major role in immediate hypersensitivity reactions and defence against
parasitic infections (Ishizaka et al., 1966). Aveskogh and Hellman (1998) cloned IgE in the
gray short-tailed opossum and observed a single IgE isotype in the species, which has also
been identified in eutherians. During evolutionary divergence, where IgG was duplicated into
four isotypes, and two duplicates in IgA, IgE was not duplicated and still exists as only one
isotype in marsupials (Aveskogh and Hellman, 1998).
Light chain (Igκ and Igλ) genes have been extensively studied in eutherian mammals,
however, only a few key marsupials have characterised their respective Igκ (gray short tailed
opossum, AAF25575; tammar wallaby, ABV02004; common brushtail possum, AAL17619;
Tasmanian devil, ALB75226; human, ACJ1716; mouse, AAA65945) and Igλ (gray short-
tailed opossum, AAC98628; tammar wallaby, AAL37211; human, S25738; mouse
AAO53439) light chains (Lucero et al., 1998, Miller et al., 1999, Deakin et al., 2006). The L
loci in the opossum are highly diverse, with at least four Vκ families and three Vλ families

63
(Miller and Belov, 2000) that collectively contain more than 60 gene segments (Lucero et al.,
1998, Miller et al., 1999). Evidence has shown the IgH and IgL loci of marsupials appears to
be evolving and diversifying due to their complexity (Sitnikova and Su, 1998, Miller and
Belov, 2000), which may be due to the IgL loci compensating for the lack of gene segment
diversity in the IgH loci (Baker et al., 2005). In the gray short-tailed opossum and common
brushtail possum, the IgL loci appears to have a higher rate of complexity than the IgH loci
and therefore contributes more to overall antibody diversity (Figure 2.4) (Baker et al., 2005,
Wang et al., 2009). It is not yet known whether marsupials favour one light chain over the
other, and in eutherians, different species express Igκ and Igλ at different ratios, for example,
the Igκ:Igλ ratio in the mouse (Mus musculus) in 95:5, whereas in the horse (Equus caballus)
it is 10:90 (Belov et al., 2001). The gray short-tailed opossum does have two identified
conserved kappa enhancers (KE5′ and KE3′D) and one recombining element (RE), however
due to gaps the gray short-tailed opossum genome (MonDom5) specific kappa-deleting
elements (KDE) have not been detected (Das et al., 2009). The Igλ locus of the gray short-
tailed opossum has a confirmed eight J-C duplicated pairs which is also present in eutherians,
this proves to be conserved in all mammals, where in avians it only contain a single J-C
region in the Igλ locus (Lucero et al., 1998).
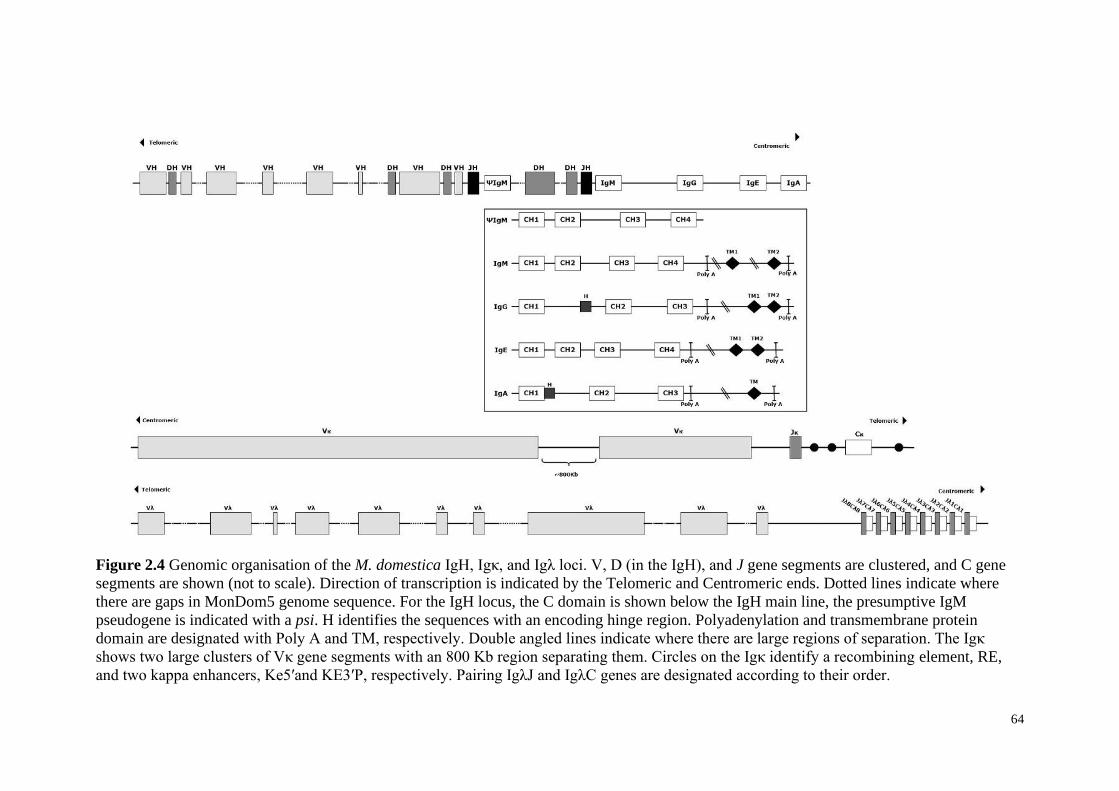
64
Figure 2.4 Genomic organisation of the M. domestica IgH, Igκ, and Igλ loci. V, D (in the IgH), and J gene segments are clustered, and C gene
segments are shown (not to scale). Direction of transcription is indicated by the Telomeric and Centromeric ends. Dotted lines indicate where
there are gaps in MonDom5 genome sequence. For the IgH locus, the C domain is shown below the IgH main line, the presumptive IgM
pseudogene is indicated with a psi. H identifies the sequences with an encoding hinge region. Polyadenylation and transmembrane protein
domain are designated with Poly A and TM, respectively. Double angled lines indicate where there are large regions of separation. The Igκ
shows two large clusters of Vκ gene segments with an 800 Kb region separating them. Circles on the Igκ identify a recombining element, RE,
and two kappa enhancers, Ke5′and KE3′P, respectively. Pairing IgλJ and IgλC genes are designated according to their order.

65
Of the Ig genes, IgA, IgG, IgE, Igκ and Igλ have all been found to be expressed in the gray
short-tailed opossum, tammar wallaby and common brushtail possum (Lucero et al., 1998,
Aveskogh and Hellman, 1998, Belov et al., 1999a, Belov et al., 1999b, Belov et al., 2001,
Duncan et al., 2010, Wang et al., 2009, Suthers and Young, 2014, Ujvari and Belov, 2015),
whereas IgM has only been expressed in the gray short-tailed opossum (Miller et al., 1998).
All three mammalian lineages therefore have fundamentally the same heavy (H)
(except IgD) and light (L) loci with respect to their Ig classes (Baker et al., 2005), and two
IgL isotypes kappa (Igκ) and lambda (Igλ) (Lucero et al., 1998, Miller et al., 1999). Antibody
diversity and specificity is due to the variation of amino acid residues at the N-terminal ends
of the H and L loci and are primarily produced by V(D)J recombination (Figure 2.4) (Baker
et al., 2005). Marsupials, unlike other mammals have contrasting diversity between the
germline H and L loci. Miller et al. (1998) reported that all but one variable (V) H loci gene
segment resided in the VH1 family, with the remaining (closely related) member residing in
the VH2 family observed in the gray short-tailed opossum. Wang et al. (2009) analysed the
gray short-tailed opossum genomic sequence and uncovered a third subgroup, IGHV3.1. This
previously unrecognizable partially germline-joined VH gene appears to be joined at the end
of the V and fused to the D segment in the IgH DNA germline of the opossum (Wang and
Miller, 2012). In a typical mammalian V(D)J recombination event, pro B-cells firstly
rearrange the DH to the JH, followed by the rearrangement of VH to the D-JH (Schatz and
Swanson, 2011). However, where a VH3.1 is apparent, there is a direct rearrangement of VH
to JH as the VH3.1 gene has previously joined the VH to the DH (Wang and Miller, 2012).
Uniquely, IGHV3.1 is significantly longer in length when compared to previously identified
mammalian IgH gene segments and shares <80 % of nucleotide identity with other gray
short-tailed opossum VH gene segments (Wang and Miller, 2012).

66
2.5.2 Cluster of Differentiation Markers
In eutherians, the first appearance and expression of CD markers can aid in detecting
and analysing the events involved in the development and differentiation of B-cells (Vale and
Schroeder, 2010). Various CD markers, such as CD10, CD19, CD34, CD79α and CD79β can
help to identify the stages of B-cell development, differentiation, and responses to antigens in
mammals. CD10, also known as neprilysin (Mishra et al., 2016) is a prime example of a
marker used to detect early stages of B-cell differentiation (Hystad et al., 2007). CD10 is
commonly expressed in early eutherian pro-B-cells (Torlakovic et al., 2005) with an
immature phenotype (Bachelard-Cascales et al., 2010), suggesting it plays a role in the
development and maturation of immature B-cells (Stamenkovic and Seed, 1988) before
ceasing its expression once B-cells mature (Ichii et al., 2010). The high expression levels of
CD10 in a common lymphoid progenitor (CLP) has been observed to aid in commitment
towards the development of early B-cells (Ichii et al., 2010). In eutherians, CD19 expression
is regulated by the transcription factor Pax5 (Wang et al., 2012a). Its initial expression begins
after the initiation of D–J gene rearrangement of the Ig heavy chain locus during the early
pro-B-cell stage (Nadler et al., 1983, Tedder, 2009) and then disappears during the terminal
plasma cell differentiation stages (Haas and Tedder, 2005, Tedder, 2009). CD34 plays a
major role in the developmental pathway from HSCs (Davi et al., 1997) through to the early
B-cell stages where it is highly expressed (LeBien and Tedder, 2008). CD34 and CD10 are
both expressed at the CLP stage through to the pro-B-cell stage, however CD19 expression is
initiated at the pro-B-cell stage (Sanz et al., 2010). From there B-cells develop into pre-B-cell
stages and the expression of CD34 ceases (Fluckiger et al., 1998). CD10, CD19, CD34 and
other CD marker genes have previously been described in many eutherians (Furness and
McNagny, 2006), and have now been identified in the gray short-tailed opossum, koala, and
Tasmanian devil (Wong et al., 2011a).

67
CD79α and CD79β are both essential for the differentiation and functionality of B-
cells in eutherians. These genes produce the BCR by forming a non-covalent heterodimer
with a sIg (Reth, 1992) and is required for BCR surface expression (Hashimoto et al., 1995).
The BCR is a complex hetero-oligomeric structure (Wienands, 2000), where ligand binding
surface Igs; Igα (CD79α) and Igβ (CD79β) are separated into distinct receptor subunits (Chu
and Arber, 2001). Both CD79 transcripts are expressed in all stages of a B-cell’s life;
immature and mature, as they are key to guiding the B-cell differentiation and antigen
response pathways (Mason et al., 1992, Chu and Arber, 2001). CD79α and CD79β have been
identified in various marsupials, and reported to have been transcribed and characterised in
the gray short-tailed opossum, tammar wallaby (Duncan et al., 2010) and bridled nail-tail
wallaby (Onychogalea fraenata) (Suthers and Young, 2014). Duncan et al. (2010) was able to
analyse the expression of CD79α and CD79β in pouch young tissues of the tammar wallaby.
At day 10 postpartum, both transcripts were detected in the bone marrow, cervical thymus
and lungs. CD79α was also detected at this time point in the spleen and blood, however
CD79β was not detected in these tissues until day 21 (Duncan et al., 2010), thus a tammar
wallaby would be unable to mount a specific immune response until at least day 21, when the
entire BCR is fully expressed. The genes encoding the CD79α (Figure 2.5A) and CD79β
(Figure 2.5B) protein sequences in these marsupials have shown a high similarity between
their highly conserved structural domains, inclusive of, an Ig domain, transmembrane (TM)
region, and a cytoplasmic tail (CYT). Their TM region in particular has been presented as
highly conserved within mammalian species, with 100 % identity in CD79α (Figure 2.5A)
and >80 % identity in CD79β (Figure 2.5B) between marsupials and the human protein
sequences. An immunoreceptor tyrosine activation motif (ITAM) was also identified on the
CYT in both CD79α and CD79β protein sequences (Figure 2.5). Reth (1989) first identified
ITAM in eutherians which has now also been characterised in different marsupials (Duncan

68
et al., 2010, Suthers and Young, 2014). CD79α and CD79β initiates an extracellular signal by
producing a specific antigen on the surface of the BCR through the cell membrane to the
cytoplasmic tail where it binds to the tyrosine residues within the ITAMs (Kim et al., 1993).
As a result, a rapid increase in protein tyrosine phosphorylation levels occurs (Gold et al.,
1991) and launches a cascade of intracellular signalling within the B-cells (Chu and Arber,
2001, Dal Porto et al., 2004, Radaev et al., 2010). The structural features of this ITAM are
highly conserved between marsupial species, even more so than with their eutherian
counterparts (Duncan et al., 2010). Transcribed CD79α of the gray short-tailed opossum and
tammar wallaby identified the presence of three conserved cysteine residues on the
extracellular domain (Duncan et al., 2010). The first two residues (C50 and C101) are able to
form an intrachain bond within the fold of the Ig protein, while the third residue (C113)
forms an intermolecular dimer between CD79α and CD79β on the BCR (Duncan et al.,
2010), and has also been observed in humans (Yu and Chang, 1992).

69
Figure 2.5 Amino acid alignment of expressed (A) CD79α and (B) CD79α mammalian
sequences. Dots indicate identical bases to M. domestica CD79 transcripts. The Ig domain,
TM regions and CYT regions are indicated by arrows below the sequences, the ITAM motifs
are identified by a box around the amino acids. Genbank accession numbers: M. domestica
(CD79α, ACZ13436; CD79β, ACZ13438.1), N. eugenii (CD79α, ACZ13437; CD79β,
ACZ13439.1), (O. fraenata CD79α, AGO64767; CD79β, AII23405.1), H. sapiens (CD79α,
AAI13732; CD79β, AAH02975.2).
2.5.3 Signal Transduction Molecules
Signal transduction is fundamental in the cellular processes used for communicating
molecular events from the cell surface and the nucleus, altering gene expression and inducing
a cellular response (Campbell, 1999). These pathways are activated by an array of external
stimuli and mediated by a series of physiological and pathological functions within the cell
(Niiro and Clark, 2002). In the instance where an Ig is expressed on the surface of a B-cell, it
results in the formation of a BCR. Upon this formation, the genes which are expressed within
the BCR bind to the BCR ligand, facilitating the signal to be sent from the cell surface to the
nucleus to elicit an immune response (Ha et al., 1992).

70
Key transducers involved in intracellular downstream BCR signalling are the
tyrosine-protein kinase (Lyn), spleen tyrosine kinase (Syk), and Bruton’s tyrosine kinase
(BTK) (Niiro and Clark, 2002). Lyn is a cytoplasmic non-receptor protein kinase that is
encoded by the Scr-family kinase Lyn gene and is associated with the cell surface receptor
proteins of the BCR. Syk is a protein kinase (Takata et al., 1994) with a structure that is
highly homologous to ZAP-70 which is expressed in T-cells and Natural Killer cells (Takata
et al., 1994) of the tammar wallaby (Flenady and Young, 2014). BTK is an intracellular
signalling molecule required by B-cells (Fluckiger et al., 1998) to link the signal responses
produced by the BCR through activation of various transcript factors (Afar et al., 1996). As
of yet, only the isoforms of Lyn tyrosine kinase have been characterised in marsupials
(Suthers and Young, 2013) due to its role in regulating the B-cell response and being the first
molecule to be activated in the downstream pathway of the BCR. Suthers and Young (2013)
detailed the expression of the Lyn isoforms, LynA and LynB in the spleen, lymph nodes,
leucocytes, whole and peripheral blood of marsupials and confirmed that the LynB arises
from the same splicing mechanism as humans. The coding domain of Lyn in both the tammar
wallaby and bridled nail-tail wallaby appears to have the same six structural regions (SH1, 2,
3, 4, a ‘unique’ domain, and COOH domains) as seen in other mammalian lineages (Suthers
and Young, 2013). The tammar wallaby Lyn gene has also been confirmed to have two
tyrosine residues (Y399 and Y510) corresponding to the human residues, Y397 and Y508
(Suthers and Young, 2013). These tyrosine residues in eutherians appear to phosphorylate,
hence regulates the activation and inhibition of intracellular kinase activity in the BCR (Dal
Porto et al., 2004). Therefore, the identification of Lyn isoforms in marsupials suggests that
Lyn is also utilised in the signalling mechanisms of marsupial B-cell responses.

71
2.5.4 Transcriptional Regulators
In eutherians, the fate of B-cells in early development and lineage commitment is
highly dependent on the activity of the paired box protein 5 (Pax5) as well as other
transcription factors including purine box factor 1 (PU.1), E box binding protein 2A (E2A)
and early B-cell factor-1 (EBF1) (Singh et al., 2005, Holmes et al., 2008). PU.1 primarily acts
in the promotion of lymphoid progenitor formation and the initiation of EBF1 expression
(Medina et al., 2004). Low concentrations of PU.1 are also required to secure the fate of B-
cells (Medina et al., 2004). Once these early lymphoid progenitors are formed, EBF1 and
E2A work together to initiate the expression of specific genes within the B-cell (O'Riordan
and Grosschedl, 1999). EBF1 is exclusively expressed by eutherian B-cell lineages, and has
an atypical helix-loop-helix zinc finger protein structure, while E2A has two basic helix-loop-
helix proteins; E12 and E47 (Hagman et al., 1993). E2A and EBF1 work together to initiate
the transcription of several B-cell genes (i.e. Vpreb1), which are encoded in the pre-B-cell
receptor and the V-D and J DNA rearrangements located in the IgH locus (Pongubala et al.,
2008).
The Pax5 gene belongs to the family of transcription factors which are important for
the control of tissue-specific transcription throughout many types of cellular differentiation
(Holmes et al., 2008). It is however the only one (of nine factors) that can be found
throughout the haematopoietic system and its expression is confined to that of B-cells in
eutherians (Fuxa and Busslinger, 2007). Pax5 expression is initiated in the pro- to pre- B-cell
stage and maintains a stable level of expression throughout the life of a B-cell until the
downregulation of plasma cell differentiation (Kallies et al., 2007). To date, Pax5 has only
been identified in a highly conserved transcriptional module that was generated to compare
human and tammar wallaby WNT4 promoter regions and shares 97 % homology between the
species (Yu et al., 2009). Other transcriptional regulators have been predicted in genome

72
databases such as ENSEMBL (Curwen et al., 2004) and Genbank (Benson et al., 2005). A
wider investigation into these key molecules in marsupials is required to confirm the
developmental and commitment process involved in marsupial B-cells.
2.6 Conclusions
Advances in genomics have and will continue to enable genome-wide investigations
of the marsupial immune system and will enable marsupials to be utilised in additional
studies in skin cancer, reproduction, evolutionary development (Selwood and Coulson, 2006),
new antimicrobial research (Wang et al., 2012c), and no doubt additional areas of research
yet to be defined. However, as many marsupial immune genes (equivalent to those identified
in eutherians) have yet to be expressed and/or characterised, our understanding is still lagging
that of the eutherian immune system. This review has highlighted the major similarities and
differences between known marsupial and eutherian B-cell genes, such as, only four of the
five Ig heavy isotypes being identified in marsupials. This has left scientists speculating that
the IgD isotype was lost in modern day marsupials during mammalian evolution.
Like eutherian B-cells, marsupial B-cells are vital in adaptive humoral immune
responses, as they are responsible for producing antibodies, via the expression of specific
genes, to be used to defend against the presence of extracellular pathogens and identify them
upon repeated exposure (Conley, 2003). Thereby, investigations into marsupial B-cell genes
continues to increase in importance, and will aid our understanding of the marsupial adaptive
immune system.
2.7 Conflicts of Interest
The authors declare no conflicts of interest.

73
2.8 Chapter 2 References
ADAMSKI, F. M. & DEMMER, J. 1999. Two stages of increased IgA transfer during
lactation in marsupial, Trichosurus vulpecula (brushtail possum). The Journal of
Immunology, 162, 6009-6015.
AFAR, D. E., PARK, H., HOWELL, B. W., RAWLINGS, D. J., COOPER, J. & WITTE, O.
N. 1996. Regulation of Btk by Src family tyrosine kinases. Molecular and Cellular
Biology, 16, 3465-3471.
ARCHIBALD, J. D. & DEUTSCHMAN, D. H. 2001. Quantitative Analysis of the Timing of
the Origin and Diversification of Extant Placental Orders. Journal of Mammalian
Evolution, 8, 107-124.
ASHMAN, R. B. & PAPADIMITRIOU, J. M. 1975. Development of lymphoid tissue in a
marsupial, Setonix brachyurus (quokka). Cells Tissues Organs, 91, 594-611.
ATWELL, J. L. & MARCHALONIS, J. J. 1977. Immunoglobulin gamma chains of a
monotreme mammal, the echnida (Tachyglossus aculeatus): amino acid composition
and partial amino acid sequence. Journal of Immunogenetics, 4, 73-80.
AVESKOGH, M. & HELLMAN, L. 1998. Evidence for an early appearance of modern post-
switch isotypes in mammalian evolution; cloning of IgE, IgG and IgA from the
marsupial Monodelphis domestica. European Journal of Immunology, 28, 2738-2750.
AVESKOGH, M., PILSTROM, L. & HELLMAN, L. 1999. Cloning and structural analysis
of IgM (μ chain) and the heavy chain V region repertoire in the marsupial
Monodelphis domestica. Developmental & Comparative Immunology, 23, 597-606.
BACHELARD-CASCALES, E., CHAPELLIER, M., DELAY, E., POCHON, G.,
VOELTZEL, T., PUISIEUX, A., CARON DE FROMENTEL, C. & MAGUER-
SATTA, V. 2010. The CD10 enzyme is a key player to identify and regulate human
mammary stem cells. Stem Cells, 28, 1081-8.
BAGWELL, C. B., HILL, B. L., WOOD, B. L., WALLACE, P. K., ALRAZZAK, M.,
KELLIHER, A. S. & PREFFER, F. I. 2015. Human B-cell and progenitor stages as
determined by probability state modeling of multidimensional cytometry data.
Cytometry Part B: Clinical Cytometry, 88, 214-26.
BAKER, M. L., BELOV, K. & MILLER, R. D. 2005. Unusually similar patterns of antibody
V segment diversity in distantly related marsupials. The Journal of Immunology, 174,
5665-5671.
BASDEN, K., COOPER, D. & DEANE, E. 1996. Development of the blood-forming tissues
of the tammar wallaby Macropus eugenii. Reproduction, Fertility and Development,
8, 989-994.
BELL, R. G., LYNCH, N. R. & TURNER, K. J. 1974a. Immunoglobulins of the marsupial
Setonix brachyurus (quokka). Immunology, 27, 1103-1115.
BELL, R. G., STEPHENS, C. J. & TURNER, K. J. 1974b. Marsupial immunoglobulins: an
immunoglobulin molecule resembling eutherian IgA in serum and secretions of
Setonix brachyurus (quokka). The Journal of Immunology, 113, 371-378.
BELOV, K., DEAKIN, J. E., PAPENFUSS, A. T., BAKER, M. L., MELMAN, S. D.,
SIDDLE, H. V., GOUIN, N., GOODE, D. L., SARGEANT, T. J., ROBINSON, M.
D., WAKEFIELD, M. J., MAHONY, S., CROSS, J. G. R., BENOS, P. V.,
SAMOLLOW, P. B., SPEED, T. P., GRAVES, J. A. M. & MILLER, R. D. 2006.
Reconstructing an ancestral mammalian immune supercomplex from a marsupial
major histocompatibility complex. PLOS Biology, 4, e46.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999a. Isolation and
sequence of a cDNA coding for the heavy chain constant region of IgG from the

74
Australian brushtail possum, Trichosurus vulpecula. Molecular Immunology, 36, 535-
541.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999b. Molecular
cloning of the brushtail possum (Trichosurus vulpecula) immunoglobulin E heavy
chain constant region. Molecular Immunology, 36, 1255-1261.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 2001. Characterisation
of the K light chain of the brushtail possum (Trichosurus vulpecula). Veterinary
Immunology and Immunopathology, 78, 317-324.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 2002a. Molecular
cloning of four lambda light chain cDNAa from the Australian brushtail possum
Trichosurus vulpecula. Journal of Immunogenetics, 29, 95-99.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 2002b. Molecular
cloning of four lambda light chain cDNAs from the Australian brushtail possum
(Trichosurus vulpecula). Euoropean Journal of Immunogenetics, 29, 95-99.
BELOV, K., HARRISON, G. A., ROSENBERG, G. H., MILLER, R. D. & COOPER, D. W.
1999c. Isolation and comparison of the IgM heavy chain constant regions from
Australian (Trichosurus vulpecula) and American (Monodelphis domestica)
marsupials. Developmental & Comparative Immunology, 23, 649-656.
BELOV, K., MILLER, R. D., OLD, J. M. & YOUNG, L. J. 2013. Marsupial immunology
bounding ahead. Australian Journal of Zoology, 61, 24-40.
BENSON, D. A., KARSCH-MIZRACHI, I., LIPMAN, D. J., OSTELL, J. & WHEELER, D.
L. 2005. GenBank. Nucleic Acids Research, 33, D34-D38.
BININDA-EMONDS, O. R. P., CARDILLO, M., JONES, K. E., MACPHEE, R. D. E.,
BECK, R. M. D., GRENYER, R., PRICE, S. A., VOS, R. A., GITTLEMAN, J. L. &
PURVIS, A. 2007. The delayed rise of present-day mammals. Nature, 446, 507.
BLOCK, M. 1960. Wound healing in the new-born opossum (Didelphis virginiana). Nature,
187, 340.
BLOCK, M. D. 1964. The blood forming tissues and blood of the newborn opossum
(Didelphis virginiana). I. normal development through about the one hundredth day
of life. Ergeb Anat Entwicklungsgesch, 37, 1-237.
BORTHWICK, C. R. & OLD, J. M. 2016. Histological development of the immune tissues
of a marsupial, the red‐tailed phascogale (Phascogale calura). Anatomical Record,
299, 207-219.
BORTHWICK, C. R., YOUNG, L. J. & OLD, J. M. 2014. The development of the immune
tissues in marsupial pouch young. Journal of Morphology, 275, 822-839.
CAMPBELL, K. S. 1999. Signal transduction from the B cell antigen-receptor. Current
Opinion in Immunology, 11, 256-264.
CHU, P. G. & ARBER, D. A. 2001. CD79: a review. Applied Immunochemistry and
Molecular Morphology, 9, 97-106.
CISTERNAS, P. A. & ARMATI, P. J. 1999. Development of the thymus, spleen, lymph
nodes and liver in the marsupial, Isoodon macrourus (northern brown bandicoot,
Peramelidae). Anatomy and Embryology, 200, 433-443.
CONLEY, M. E. 2003. Genes required for B cell development. Journal of Clinical
Investigation, 112, 1636-1638.
CONLEY, M. E. & COOPER, M. D. 1998. Genetic basis of abnormal B cell development.
Current Opinion in Immunology, 10, 399-406.
COOPER, M. D. 1987. Current concepts. B lymphocytes. normal development and function.
The New England Journal of Medicine, 317, 1452-1456.

75
CURWEN, V., EYRAS, E., ANDREWS, T., CLARKE, L., MONGIN, E., SEARLE, S. &
CLAMP, M. 2004. The ENSEMBL automatic gene annotation system. Genome
Research, 14, 942-50.
CUTTS, J. H. & KRAUSE, W. J. 1982. Postnatal development of the spleen in Didelphis
virginiana. Journal of Anatomy, 135, 601-613.
CUTTS, J. H., LEESON, C. R. & KRAUSE, W. J. 1973. The postnatal development of the
liver in a marsupial, Didelphis virginiana. I. light microscopy. Journal of Anatomy,
115, 327-346.
DAL PORTO, J., B GAULD, S., T MERRELL, K., MILLS, D., PUGH-BERNARD, A. &
CAMBIER, J. 2004. B cell antigen receptor signaling 101. Molecular Immunology,
41, 599-613.
DALY, K. A., DIGBY, M., LEFÈVRE, C., MAILER, S., THOMSON, P., NICHOLAS, K. &
WILLIAMSON, P. 2007. Analysis of the expression of immunoglobulins throughout
lactation suggests two periods of immune transfer in the tammar wallaby (Macropus
eugenii). Veterinary Immunology and Immunopathology, 120, 187-200.
DAS, S., NIKOLAIDIS, N. & NEI, M. 2009. Genomic organization and evolution of
immunoglobulin kappa gene enhancers and kappa deleting element in mammals.
Molecular Immunology, 46, 3171-3177.
DAVI, F., FAILI, A., GRITTI, C., BLANC, C., LAURENT, C., SUTTON, L., SCHMITT, C.
& MERLE-BÉRAL, H. 1997. Early onset of immunoglobulin heavy chain gene
rearrangements in normal human bone marrow CD34+ cells. Blood, 90, 4014-4021.
DEAKIN, J. E., OLP, J. J., GRAVES, J. A. M. & MILLER, R. D. 2006. Physical mapping of
immunoglobulin loci IgH@, IgK@, and IgL@ in the opossum (Monodelphis
domestica). Cytogenetic and Genome Research, 114, 94.
DEANE, E. & COOPER, D. 1988. Immunological development of pouch young marsupials.
The Developing Marsupial. Springer.
DEANE, E., COOPER, D. & RENFREE, M. 1990. Immunoglobulin G levels in fetal and
newborn tammar wallabies (Macropus eugenii). Reproduction, Fertility,
Development, 2, 369-375.
DEANE, E. M. & COOPER, D. W. 1984. Immunology of pouch marsupials. I. levels of
immunoglobulin transferrin and albumin in the blood and milk of euros and wallaroos
(hill kangaroos: Macropus robustus. marsupialia). Developmental & Comparative
Immunology, 8, 863-876.
DENEYS, V., MAZZON, A., MARQUES, J., BENOIT, H. & DE BRUYÈRE, M. 2001.
Reference values for peripheral blood B-lymphocyte subpopulations: a basis for
multiparametric immunophenotyping of abnormal lymphocytes. Journal of
Immunological Methods, 253, 23-36.
DOULATOV, S., NOTTA, F., LAURENTI, E. & DICK, JOHN E. 2012. Hematopoiesis: a
human perspective. Cell Stem Cell, 10, 120-136.
DUNCAN, L., WEBSTER, K., GUPTA, V., NAIR, S. & DEANE, E. 2010. Molecular
characterisation of the CD79a and CD79b subunits of the B cell receptor complex in
the gray short-tailed opossum (Monodelphis domestica) and tammar wallaby
(Macropus eugenii): delayed B cell immunocompetence in marsupial neonates.
Veterinary Immunology and Immunopathology, 136, 235-47.
EDWARDS, M., HINDS, L., DEANE, E. & DEAKIN, J. 2012. A review of complementary
mechanisms which protect the developing marsupial pouch young. Developmental &
Comparative Immunology, 37, 213-220.
FLENADY, S. & YOUNG, L. J. 2014. Molecular characterisation of the signaling molecules
TCRζ and ZAP-70 in the marsupial Macropus eugenii (tammar wallaby). Australian
Mammalogy, 36, 137-145.

76
FLUCKIGER, A. C., LI, Z., KATO, R. M., WAHL, M. I., OCHS, H. D., LONGNECKER,
R., KINET, J. P., WITTE, O. N., SCHARENBERG, A. M. & RAWLINGS, D. J.
1998. Btk/Tec kinases regulate sustained increases in intracellular Ca2+ following B-
cell receptor activation. The EMBO Journal, 17, 1973-1985.
FURNESS, S. & MCNAGNY, K. 2006. Beyond Mere Markers: Functions for CD34 Family
of Sialomucins in Hematopoiesis. Immunologic Research, 34, 13-32.
FUXA, M. & BUSSLINGER, M. 2007. Reporter gene insertions reveal a strictly B
lymphoid-specific expression pattern of pax5 in support of its B cell identity function.
The Journal of Immunology, 178, 8221.
GOLD, M., MATSUUCHI, L., KELLY, R. B. & DEFRANCO, A. 1991. Tyrosine
phosphorylation of components of the B cell antigen receptor following receptor
cross-linking. Proceedings of the National Academy of Sciences of the United States
of America, 88, 3436-40.
GRAVES, J. A. M. & WESTERMAN, M. 2002. Marsupial genetics and genomics. Trends in
Genetics, 18, 517-521.
HA, H. J., KUBAGAWA, H. & BURROWS, P. D. 1992. Molecular cloning and expression
pattern of a human genome homologous to the murine mb-1 gene. The Journal of
Immunology, 148, 1526-1531.
HAAS, K. M. & TEDDER, T. F. 2005. Role of the CD19 and CD21/35 receptor complex in
innate immunity, host defense and autoimmunity. Mechanisms of Lymphocyte
Activation and Immune Regulation X. Springer.
HADLAND, B. & YOSHIMOTO, M. 2018. Many layers of embryonic hematopoiesis: new
insights into B-cell ontogeny and the origin of hematopoietic stem cells. Experimental
Hematology, 60, 1-9.
HAGMAN, J., BELANGER, C., TRAVIS, A., TURCK, C. W. & GROSSCHEDL, R. 1993.
Cloning and functional characterisation of early B-cell factor, a regulator of
lymphocyte-specific gene expression. Genes and Development, 7, 760-773.
HARDY, R. R. & HAYAKAWA, K. 2001. B cell development pathway. Annual Review of
Immunology, 19, 595-621.
HASHIMOTO, S., CHIORAZZI, N. & GREGERSEN, P. K. 1995. Alternative splicing of
CD79a (Ig-αmb-1) and CD79b (Ig-βB29) RNA transcripts in human B cells.
Molecular Immunology, 32, 651-659.
HINDES, R. D. & MIZELL, M. 1976. The origin of immunoglobulins in opossum
“embryos”. Developmental Biology, 53, 49-61.
HOBBS, M., PAVASOVIC, A., KING, A. G., PRENTIS, P. J., ELDRIDGE, M. D. B.,
CHEN, Z., COLGAN, D. J., POLKINGHORNE, A., WILKINS, M. R.,
FLANAGAN, C., GILLETT, A., HANGER, J., JOHNSON, R. N. & TIMMS, P.
2014. A transcriptome resource for the koala (Phascolarctos cinereus): insights into
koala retrovirus transcription and sequence diversity. BMC Genomics, 15, 786-786.
HOLMES, M. L., PRIDANS, C. & NUTT, S. L. 2008. The regulation of the B-cell gene
expression programme by Pax5. Immunology and Cell Biology, 86, 47-53.
HUBBARD, T. J. P., AKEN, B. L., BEAL, K., BALLESTER, B., CACCAMO, M., CHEN,
Y., CLARKE, L., COATES, G., CUNNINGHAM, F., CUTTS, T., DOWN, T.,
DYER, S. C., FITZGERALD, S., FERNANDEZ-BANET, J., GRAF, S., HAIDER,
S., HAMMOND, M., HERRERO, J., HOLLAND, R., HOWE, K., JOHNSON, N.,
KAHARI, A., KEEFE, D., KOKOCINSKI, F., KULESHA, E., LAWSON, D.,
LONGDEN, I., MELSOPP, C., MEGY, K., MEIDL, P., OUVERDIN, B., PARKER,
A., PRLIC, A., RICE, S., RIOS, D., SCHUSTER, M., SEALY, I., SEVERIN, J.,
SLATER, G., SMEDLEY, D., SPUDICH, G., TREVANION, S., VILELLA, A.,
VOGEL, J., WHITE, S., WOOD, M., COX, T., CURWEN, V., DURBIN, R.,

77
FERNANDEZ-SUAREZ, X. M., FLICEK, P., KASPRZYK, A., PROCTOR, G.,
SEARLE, S., SMITH, J., URETA-VIDAL, A. & BIRNEY, E. 2007. Ensembl 2007.
Nucleic Acids Research, 35, D610-D617.
HYSTAD, M. E., MYKLEBUST, J. H., BØ, T. H., SIVERTSEN, E. A., RIAN, E.,
FORFANG, L., MUNTHE, E., ROSENWALD, A., CHIORAZZI, M. & JONASSEN,
I. 2007. Characterization of early stages of human B cell development by gene
expression profiling. The Journal of Immunology, 179, 3662-3671.
ICHII, M., ORITANI, K., YOKOTA, T., ZHANG, Q., GARRETT, K. P., KANAKURA, Y.
& KINCADE, P. W. 2010. The density of CD10 corresponds to commitment and
progression in the human B lymphoid lineage. PLoS One, 5, e12954.
ISHIZAKA, K., ISHIZAKA, T. & HORNBROOK, M. M. 1966. Physico-chemical properties
of human reaginic antibody. iv. presence of a unique immunoglobulin as a carrier of
reaginic activity. Journal of Immunology, 91, 75-85.
JURD, R. D. 1994. "Not proper mammals": immunity in monotremes and marsupials.
Comparative Immunology, Microbiology and Infectious Diseases, 17, 41-52.
KALLIES, A., HASBOLD, J., FAIRFAX, K., PRIDANS, C., EMSLIE, D., MCKENZIE, B.
S., LEW, A. M., CORCORAN, L. M., HODGKIN, P. D., TARLINTON, D. M. &
NUTT, S. L. 2007. Initiation of plasma-cell differentiation is independent of the
transcription factor blimp-1. Immunity, 26, 555-566.
KALMUTZ, S. E. 1962. Antibody production in the opossum embryo. Nature, 193, 851-853.
KETT, K., BRANDTZAEG, P., RADL, J. & HAAIJMAN, J. J. 1986. Different subclass
distribution of IgA-producing cells in human lymphoid organs and various secretory
tissues. The Journal of Immunology, 136, 3631-3635.
KIM, K. M., ALBER, G., WEISER, P. & RETH, M. 1993. Differential signalling throught
the Ig-a amd Ig-b components of the B-cell antigen receptor. European Journal of
Immunology, 23, 911-916.
KOSHLAND, M. E. 1985. The Coming of Age of the Immunoglobulin J Chain. Annual
Review of Immunology, 3, 425-453.
KUMAR, S. & HEDGES, S. B. 1998. A molecular timescale for vertebrate evolution.
Nature, 392, 917-920.
KUROSAKI, T. 2002. Regulation of B-cell signal transduction by adaptor proteins. Nature
Reviews Immunology, 2, 354.
LAPLANTE, S. E., BURRELL, L. R., WATNE, L. A., TAYLOR, L. D. & ZIMMERMANN,
L. B. 1969. Skin allograft studies in the pouch young of the opossum.
Transplantation, 7, 67-72.
LAWTON, A. R., KINCADE, P. W. & COOPER, M. D. 1975. Sequential expression of
germ line genes in development of immunoglobulin class diversity. Federation
Proceedings, 34, 33-39.
LEBIEN, T. W. 2000. Fates of human B-cell precursors. Blood, 96, 9-23.
LEBIEN, T. W. & TEDDER, T. F. 2008. B lymphocytes: how they develop and function.
Blood, 112, 1570-80.
LI, A., RUE, M., ZHOU, J., WANG, H., GOLDWASSER, M. A., NEUBERG, D.,
DALTON, V., ZUCKERMAN, D., LYONS, C., SILVERMAN, L. B., SALLAN, S.
E. & GRIBBEN, J. G. 2004. Utilization of Ig heavy chain variable, diversity, and
joining gene segments in children with B-lineage acute lymphoblastic leukemia:
implications for the mechanisms of VDJ recombination and for pathogenesis. Blood,
103, 4602.
LUCERO, J. E., ROSENBERG, G. H. & MILLER, R. D. 1998. Marsupial light chains:
complexity and conservation of λ in the opssum Monodelphis domestica. The Journal
of Immunology, 161, 6724-6732.

78
MACCONMARA, M. & LEDERER, J. A. 2005. B cells. Critical Care Medicine, 33, S514-
S516.
MASON, D. Y., VAN NOESEL, C. J., CORDELL, J. L., COMANS-BITTER, W. M.,
MICKLEM, K., TSE, A. G., VAN LIER, R. A. & VAN DONGEN, J. J. 1992. The
B29 and mb-1 polypeptides are differentially expressed during human B cell
differentiation. European Journal of Immunology, 22, 2753 - 2756.
MEDINA, K. L., PONGUBALA, J. M. R., REDDY, K. L., LANCKI, D. W., DEKOTER, R.,
KIESLINGER, M., GROSSCHEDL, R. & SINGH, H. 2004. Assembling a gene
regulatory network for specification of the B cell fate. Developmental Cell, 7, 607-
617.
MIKKELSEN, T. S., WAKEFIELD, M., AKEN, B., AMEMIYA, C., CHANG, J., DUKE
BECKER, S., GARBER, M., GENTLES, A. J., GOODSTADT, L., HEGER, A.,
JURKA, J., KAMAL, M., MAUCELI, E., SEARLE, S., SHARPE, T., BAKER, M.
L., BATZER, M. A., BENOS, P., BELOV, K. & LINDBLAD-TOH, K. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in non-coding
sequences. Nature 447, 167-77.
MILLER, R. D. & BELOV, K. 2000. Immunoglobulin genetics of marsupials.
Developmental & Comparative Immunology, 24, 485-490.
MILLER, R. D., BERGEMANN, E. R. & ROSENBERG, G. H. 1999. Marsupial light
chains: IgK with four V families in the opossum Monodelpis domestica.
Immunogenetics, 50, 329-335.
MILLER, R. D., GRABE, H. & ROSENBERG, G. H. 1998. VH repertoire of a marsupial
(Monodelphis domestica). The Journal of Immunology, 160, 259-365.
MISHRA, D., SINGH, S. & NARAYAN, G. 2016. Role of B cell development marker CD10
in cancer progression and prognosis. Molecular Biology International, 2016,
4328697.
MORRIS, K., WONG, E. S. W. & BELOV, K. 2010. Use of genomic information to gain
insights into immune function in marsupials: a review of divergent immune Genes.
In: DEAKIN, J. E., WATERS, P. D. & MARSHALL GRAVE, J. (eds.) Marsupial
Genetics and Genomics. Springer, Dordrecht.
MOSTOV, K. E. 1994. Transepithelial transport of immunoglobulins. Annual Review of
Immunology, 12, 63.
MURCHISON, E., SCHULZ-TRIEGLAFF, O., NING, Z., ALEXANDROV, L., BAUER,
M., FU, B., HIMS, M., DING, Z., IVAKHNO, S., STEWART, C., NG, B., WONG,
W., AKEN, B., WHITE, S., ALSOP, A., BECQ, J., BIGNELL, G., CHEETHAM, R.
K., CHENG, W., CONNOR, T., COX, A., FENG, Z.-P., GU, Y., GROCOCK, R.,
HARRIS, S., KHREBTUKOVA, I., KINGSBURY, Z., KOWARSKY, M., KREISS,
A., LUO, S., MARSHALL, J., MCBRIDE, D., MURRAY, L., PEARSE, A.-M.,
RAINE, K., RASOLONJATOVO, I., SHAW, R., TEDDER, P., TREGIDGO, C.,
VILELLA, A., WEDGE, D., WOODS, G., GORMLEY, N., HUMPHRAY, S.,
SCHROTH, G., SMITH, G., HALL, K., SEARLE, S. J., CARTER, N., PAPENFUSS,
A., FUTREAL, P. A., CAMPBELL, P., YANG, F., BENTLEY, D., EVERS, D. &
STRATTON, M. 2012. Genome sequencing and analysis of the Tasmanian devil and
its transmissible cancer. Cell, 148, 780-791.
MUSCHEN, M., LEE, S., ZHOU, G., FELDHAHN, N., BARATH, V. S., CHEN, J.,
MOERS, C., KRONKE, M., ROWLEY, J. D. & WANG, S. M. 2002. Molecular
portraits of B cell lineage commitment. Proceedings of the National Academy of
Sciences of the United States of America, 99, 10014-10019.
NADLER, L., ANDERSON, K., MARTI, G., BATES, M., PARK, E., DALEY, J. &
SCHLOSSMAN, S. 1983. B4, a human B lymphocyte-associated antigen expressed

79
on normal, mitogen-activated, and malignant B lymphocytes. The Journal of
Immunology, 131, 244-250.
NIIRO, H. & CLARK, E. A. 2002. Regulation of B-cell fate by antigen-receptor signals.
Nature Review Immunology, 2, 945-56.
O'RIORDAN, M. & GROSSCHEDL, R. 1999. Coordinate regulation of B cell differentiation
by the transcription factors EBF and E2A. Immunity, 11, 21-31.
OHTA, Y. & FLAJNIK, M. 2006. IgD, like IgM, is a primordial immunoglobulin class
perpetuated in most jawed vertebrates. Proceedings of the National Academy of
Sciences of the United States of America, 103, 10723-10728.
OLD, J. M. 2016. Haematopoiesis in marsupials. Developmental & Comparative
Immunology, 58, 40-46.
OLD, J. M. & DEANE, E. 2003. The detection of mature T- and B-cells during development
of the lymphoid tissues of the tammar wallaby (Macropus eugenii). Journal of
Anatomy, 203, 123-131.
OLD, J. M. & DEANE, E. M. 2000. Development of the immune system and immunological
protection in marsupial pouch young. Developmental & Comparative Immunology,
24, 445-454.
OLD, J. M., SELWOOD, L. & DEANE, E. M. 2004. A developmental investigation of the
liver, bone marrow and spleen of the stripe-faced dunnart (Sminthopsis macroura).
Developmental & Comparative Immunology, 28, 347-355.
PAINTER, M. W., DAVIS, S., HARDY, R. R., MATHIS, D., BENOIST, C. &
IMMUNOLOGICAL GENOME PROJECT, C. 2011. Transcriptomes of the B and T
lineages compared by multiplatform microarray profiling. Journal of Immunology,
186, 3047-57.
PAINTER, R. H. 1998. IgG. In: DELVES, P. J. (ed.) Encyclopedia of Immunology (Second
Edition). Oxford: Elsevier.
PALMER, C., DIEHN, M., ALIZADEH, A. A. & BROWN, P. O. 2006. Cell-type specific
gene expression profiles of leukocytes in human peripheral blood. BMC Genomics, 7,
115.
PAPENFUSS, A., HSU, A. & WAKEFIELD, M. 2010. Marsupial sequencing projects and
bioinformatics challenges. Marsupial Genetics and Genomics, 121-132.
PONGUBALA, J. M., NORTHRUP, D. L., LANCKI, D. W., MEDINA, K. L., TREIBER,
T., BERTOLINO, E., THOMAS, M., GROSSCHEDL, R., ALLMAN, D. & SINGH,
H. 2008. Transcription factor EBF restricts alternative lineage options and promotes B
cell fate commitment independently of Pax5. Nature Immunology, 9, 203-15.
RADAEV, S., ZOU, Z., TOLAR, P., NGUYEN, K., NGUYEN, A., KRUEGER, P. D.,
STUTZMAN, N., PIERCE, S. & SUN, P. D. 2010. Structural and functional studies
of Ig alpha beta and its assembly with the B cell antigen receptor. Structure, 18, 934-
43.
RAMADASS, P. & MORIARTY, K. M. 1982. Immunoglobulins of the Australian brush-
tailed opossum, Trichosurus vulpecula. Developmental & Comparative Immunology,
6, 549-556.
RENFREE, M. B. 1973. The composition of the fetal fluids of the marsupial Macropus
eugenii. Developmental Biology, 33, 62-79.
RENFREE, M. B., HORE, T. A., SHAW, G., GRAVES, J. A. & PASK, A. J. 2009.
Evolution of genomic imprinting: insights from marsupials and monotremes. Annual
Reivew of Genomics and Human Genetics, 10, 241-62.
RETH, M. 1989. Antigen receptor tail clue. Nature, 338, 383-384.
RETH, M. 1992. Antigen Receptors on B Lymphocytes. Annual Review of Immunology, 10,
97-121.

80
ROWLANDS, D. T. & DUDLEY, M. A. 1968. The isolation of immunoglobulins of the
adult opossum (Didelphis domestica). Journal of Immunology, 100, 736-743.
ROWLANDS, D. T., LA VIA, M. F. & BLOCK, M. H. 1964. The blood forming tissues and
blood of the newborn opossum (Didelphys virginiana) II. ontogenesis of antibody
formation to flagella of salmonella typhi. The Journal of Immunology, 93, 157-164.
SAMOLLOW, P. B. 2006. Status and applications of genomic resources for the gray, short-
tailed opossum, Monodelphis domestica, an American marsupial model for
comparative biology. Australian Journal of Zoology, 54, 173-196.
SANZ, E., MUNOZ, A. N., MONSERRAT, J., VAN-DEN-RYM, A., ESCOLL, P., RANZ,
I., ALVAREZ-MON, M. & DE-LA-HERA, A. 2010. Ordering human CD34+CD10-
CD19+ pre/pro-B-cell and CD19- common lymphoid progenitor stages in two pro-B-
cell development pathways. Proceedings of the National Academy of Sciences of the
United States of America, 107, 5925-30.
SCHATZ, D. G. & SWANSON, P. C. 2011. V(D)J recombination: mechanisms of initiation.
Annual Review of Genetics, 45, 167-202.
SELWOOD, L. & COULSON, G. 2006. Marsupials as models for research. Australian
Journal of Zoology, 54, 137-138.
SHEARER, M. H., ROBINSON, E. S., VANDEBERG, J. L. & KENNEDY, R. C. 1995.
Humoral immune response in a marsupial Monodelphis domestica: anti-isotypic and
anti-idiotypic responses detected by species-specific monoclonal anti-
immunoglobulin reagents. Developmental & Comparative Immunology, 19, 237-256.
SINGH, H., MEDINA, K. L. & PONGUBALA, J. M. R. 2005. Contingent gene regulatory
networks and B cell fate specification. Proceedings of the National Academy of
Sciences of the United States of America, 102, 4949-4953.
SITNIKOVA, T. & SU, C. 1998. Coevolution of immuoglobulin heavy- and light-chain
variable-region gene families. Immunogenetics, 15, 617-625.
SMITH, K. G., KAMDAR, A. A. & STARK, J. M. 2019. Lung Defenses : Intrinsic, Innate,
and Adaptive. In: WILMOTT, R. W., DETERDING, R., LI, A., RATJEN, F., SLY,
P., ZAR, H. J. & BUSH, A. (eds.) Kendig’s Disorders of the Respiratory Tract in
Children (9th Ed).
STAMENKOVIC, I. & SEED, B. 1988. CD19, the earliest differentiation antigen of the B
cell lineage, bears three extracellular immunoglobulin-like domains and an epstein-
barr virus-related cytoplasmic tail. Journal of Experimental Medicine, 168, 1205-
1210.
STANLEY, N. F., YADAV, M., WARING, H. & EADIE, M. 1972. The effect of
thymectomy on response to various antigens of a marsupial Setonix brachyurus
(quokka). Immunology and Cell Biology, 50, 689.
STOKES, D. C. & RAJASEKARAN, S. 2008. Chapter 36 - Respiratory Infections in
Immunocompromised Hosts. In: TAUSSIG, L. M. & LANDAU, L. I. (eds.) Pediatric
Respiratory Medicine (Second Edition). Philadelphia: Mosby.
SUTHERS, A. N. & YOUNG, L. J. 2013. Molecular identification and expression of Lyn
tyrosine kinase isoforms in marsupials. Molecular Immunology, 55, 310-318.
SUTHERS, A. N. & YOUNG, L. J. 2014. Isoforms of the CD79 signal transduction
component of the macropod B-cell receptor. Developmental & Comparative
Immunology, 47, 185-90.
TAKATA, M., SABE, H., AKIKO, H., INAZU, T., HOMMA, Y., NUKADA, T.,
YAMAMURA, H. & KUROSAKI, T. 1994. Tyrosine kinases lyn and syk regulate B
cell receptor-couples Ca2+ mobilization through distinct pathways. The EMBO
Journal, 13, 1341-1349.

81
TEDDER, T. F. 2009. CD19: a promising B cell target for rheumatoid arthritis. Nature
Reviews Rheumatology, 5, 572.
TORLAKOVIC, E., TENSTAD, E., FUNDERUD, S. & RIAN, E. 2005. CD10+ stromal
cells form B-lymphocyte maturation niches in the human bone marrow. The Journal
of Pathology: A Journal of the Pathological Society of Great Britain and Ireland,
205, 311-7.
TYNDALE-BISCOE, H. & RENFREE, M. 1987. Reproductive physiology of marsupials,
Cambridge University Press.
UJVARI, B. & BELOV, K. 2015. Characterization of antibody V segment diversity in the
Tasmanian devil (Sarcophilus harrisii). Veterinary Immunology and
Immunopathology, 167, 156-165.
VALE, A. M. & SCHROEDER, H. W. 2010. Clinical consequences of defects in B-cell
development. Journal of Allergy and Clinical Immunology, 125, 778-787.
VIDARSSON, G., DEKKERS, G. & RISPENS, T. 2014. IgG subclasses and allotypes: from
structure to effector functions. Frontiers in Immunology, 5, 520-520.
WANG, K., WEI, G. & LIU, D. 2012a. CD19: a biomarker for B cell development,
lymphoma diagnosis and therapy. Experimental Hematology and Oncology, 1.
WANG, X. & MILLER, R. D. 2012. Recombination, transcription, and diversity of a
partially germline-joined VH in a mammal. Immunogenetics, 64, 713-7.
WANG, X., OLP, J. J. & MILLER, R. D. 2009. On the genomics of immunoglobulins in the
gray, short-tailed opossum Monodelphis domestica. Immunogenetics, 61, 581-96.
WANG, X., PARRA, Z. E. & MILLER, R. D. 2012b. A VpreB3 homologue in a marsupial,
the gray short-tailed opossum, Monodelphis domestica. Immunogenetics, 64, 647-52.
WANG, X., SHARP, A. R. & MILLER, R. D. 2012c. Early postnatal B cell ontogeny and
antibody repertoire maturation in the opossum, Monodelphis domestica. PLoS One, 7,
e45931.
WARREN, W. C., HILLIER, L. W., GRAVES, J. A. M., BIRNEY, E., PONTING, C. P.,
GRÜTZNER, F., BELOV, K., MILLER, W., CLARKE, L. & CHINWALLA, A. T.
2008. Genome analysis of the platypus reveals unique signatures of evolution. Nature,
453, 175.
WIENANDS, J. 2000. The B-Cell Antigen Receptor: Formation of Signaling Complexes and
the Function of Adaptor Proteins. In: JUSTEMENT, L. B. & SIMINOVITCH, K. A.
(eds.) Signal Transduction and the Coordination of B Lymphocyte Development and
Function I: Transduction of BCR Signals from the Cell Membrane to the Nucleus.
Berlin, Heidelberg: Springer Berlin Heidelberg.
WILKINSON, R., ALLANSON, M., KOLEGA, V., LAWRENCE, D. & NEVILLE, S. 1991.
Purification and initial characterisation of koala immunoglobulins. Veterinary
Immunology and Immunopathology, 29, 189-195.
WONG, E. S., PAPENFUSS, A. T. & BELOV, K. 2011a. Immunome database for
marsupials and monotremes. BMC Immunology, 12, 48.
WONG, E. S., PAPENFUSS, A. T., HEGER, A., HSU, A. L., PONTING, C. P., MILLER, R.
D., FENELON, J. C., RENFREE, M. B., GIBBS, R. A. & BELOV, K. 2011b.
Transcriptomic analysis supports similar functional roles for the two thymuses of the
tammar wallaby. BMC Genomics, 12, 420.
WONG, E. S., YOUNG, L. J., PAPENFUSS, A. T. & BELOV, K. 2006. In silico
identification of opossum cytokine genes suggests the complexity of the marsupial
immune system rivals that of eutherian mammals. Immunome Research, 2, 4.
YADAV, M. & EADIE, M. 1972. Passage of maternal immunoglobulins to the pouch young
of a marsupial Setonix brachyurus. Australian Journal of Zoology, 21, 171-81.

82
YU, H., PASK, A. J., SHAW, G. & RENFREE, M. B. 2009. Comparative analysis of the
mammalian WNT4 promoter. BMC Genomics, 10, 416-416.
YU, L. M. & CHANG, T. W. 1992. Human mb-1 gene: complete cDNA sequence and its
expression in B cells bearing membrane Ig of various isotypes. The Journal of
Immunology, 148, 633.
ZELUS, D., ROBINSON-RECHAVI, M., DELACRE, M., AURIAULT, C. & LAUDET, V.
2000. Fast evolution of interleukin-2 in mammals and positive selection in ruminants.
Journal of Molecular Evolution, 51, 234-244.
ZHANG, Z., GOLDSCHMIDT, T. & SALTER, H. 2012. Possible allelic structure of IgG2a
and IgG2c in mice. Molecular Immunology, 50, 169-171.

83
CHAPTER 3
Single-cell transcriptome analysis of the B-cell
repertoire reveals the usage of
immunoglobulins in the gray short-tailed
opossum (Monodelphis domestica)
A. L. Schraven1, V. L. Hansen2, K. A. Morrissey2, H. J. Stannard3, O. T.W. Ong4, D. C.
Douek5, R. D. Miller2, J. M. Old1
1School of Science and Health, Hawkesbury Campus, Western Sydney University, Locked
bag 1797, Penrith, NSW 2751, Australia
2Center for Evolutionary and Theoretical Immunology, Department of Biology, University of
New Mexico Albuquerque, New Mexico, USA
3Charles Sturt University, School of Animal and Veterinary Sciences, Wagga Wagga, NSW
2678, Australia
4QIMR Berghofer Medical Research Institute, Brisbane, Queensland, Australia
5Human Immunology Section, Vaccine Research Center, National Institute of Allergy and
Infectious Diseases, National Institutes of Health, Bethesda, Maryland, USA
Manuscript has been submitted to Immunology and Cell Biology and is currently under
review.

84
3.1 Chapter Outline and Authorship
Chapter 3 evaluates the usage of Immunoglobulin Heavy and Light chains in the gray short-
tailed opossum (Monodelphis domestica) B-cell repertoire. The B-cell transcriptome was
generated by single-cell RNA-Seq technology. Analysis of the opossum B-cell repertoire
identified the fraction of cells expressing each heavy chain, as well as the overall light chain
isotopic ratio.
This chapter is a prepared manuscript and is jointly authored; I am the primary author,
analysed the raw data, identified paired immunoglobulin chains and gene segments,
annotated pseudogenes, performed all statistical analysis, and prepared the manuscript.
Victoria Hansen performed Cell sorting, RNA extraction and cDNA synthesis, Daniel Douek
performed the sequencing, and Kimberly Morrissey assisted with the initial analysis. Robert
Miller, Hayley Stannard, Oselyne Ong and Julie Old provided advice on data analysis, and
have reviewed and provided feedback on the manuscript.
This chapter is a manuscript that has been submitted to the Journal of Immunology and Cell
Biology for publication and is currently under review, and is therefore formatted to the
journal specifications.

85
3.2 Introduction
Typically, Ig molecules comprise of two identical IgH and two identical IgL chains,
which are paired together on the surface of the B-cell. Both of these chains have a constant
(C) domain which determines the effector function of the Ig molecule, and a variable (V)
domain responsible for binding to an antigen (Alt et al., 1992). Ig isotypes are determined by
their CH domain, where four of the five mammalian IgH isotypes have been confirmed in the
gray short-tailed opossum (Monodelphis domestica). Previous investigations have isolated
IgM, IgG, IgA, and IgE heavy chain isotypes (Miller et al., 1998, Aveskogh and Hellman,
1998, Belov et al., 1998, Aveskogh et al., 1999, Belov et al., 1999a, Belov et al., 1999b,
Belov et al., 1999c), however, unlike eutherians and monotremes, IgD has not been found in
any marsupial species to date. The genomic region predicted to encode the IgD constant
domain in the opossum, appears to be replaced with insertions of large duplicated regions of
repetitive DNA (Wang et al., 2009). It is not known whether these insertions are typical for
all marsupials, or just the opossum. Although, it is clear the absence of IgD is due to a gene
loss in the opossum, the presence of IgD in both eutherian and monotreme species (and some
non-mammalian vertebrates) is consistent with IgD being ancient in vertebrates (Ohta and
Flajnik, 2006, Wang et al., 2009). Like all mammals, marsupials have two IgL isotypes;
kappa (Igκ) and lambda (Igλ) (Lucero et al., 1998). In the opossum Igκ, the V and joining (J)
gene segments are present in different clusters, upstream of a single Cκ domain (Miller et al.,
1999), whereas Igλ has multiple Jλ-Cλ pairs (Lucero et al., 1998, Wang et al., 2009).
Antibody diversity and specificity is determined by the V domain located on the N-
terminal ends of the IgH and IgL chains (Baker et al., 2005). The abundance and diversity of
the VH domain appears to be highly variational within the eutherian lineage, humans and
mice have a large set of VH gene segments, whereas other eutherians, such as rabbits and
cattle, have a limited VH diversity (Butler, 1997). This pattern of VH diversity seems to have a

86
coevolving relationship with the diversity of VL, where species with high VH diversity
(humans and mice) also have a high diversity of VL gene segments, and species with low VH
diversity (rabbits and cattle) have low VL diversification (Sitnikova and Su, 1998). In the
opossum however, this coevolution of VH and VL diversification doesn’t appear to apply, the
VH is both limited in diversity and abundance (Miller et al., 1998, Baker et al., 2005) while
VL has a set of highly diverse gene segments (Lucero et al., 1998, Miller et al., 1999, Wang et
al., 2009). This pattern appears true for other marsupial species as well (Baker et al., 2005). A
highly complex VL domain seems to compensate for limited VH diversity, and therefore
contribute more to overall antibody diversity in marsupials (Lucero et al., 1998, Miller et al.,
1999, Baker et al., 2005, Wang et al., 2009).
The availability of the fully mapped and annotated IgH and IgL chains of the gray
short-tailed opossum has facilitated the comparative analysis of mammalian B-cells (Deakin
et al., 2006, Wang et al., 2009). The opossum is born highly altrical and like all marsupials,
B-cell competence develops postpartum. IgH transcripts have been detected as early as the
first 24 h postpartum, though IgL transcripts have not been detected until day 7 after birth
(Wang et al., 2012). Therefore, the earliest the opossum neonate would be able to generate an
endogenous antibody response is at least one week postpartum (Wang et al., 2012), leaving
the neonate completely dependent on the antibodies transferred via the maternal milk for the
first week (Samples et al., 1986). During the second week after birth opossum antibody
diversification increases, seemingly due to the complexity of the IgL chain (Wang et al.,
2012). Here we identify and compare the usage of IgH and IgL isotypes in the B-cell
repertoire of the opossum. In doing so, we present the first study to investigate the usage of Ig
chains in any marsupial species and compare opossum IgL isotype ratios to other mammalian
species.

87
3.3 Materials and Methods
3.3.1 Ethics Statement
This study was approved under the protocol numbers 16-200407-MC and 15-200334-
B-MC from the University of New Mexico Institutional Animal Care and Use Committee and
protocol numbers BIO-17-969 from the Uniformed Services University of the Health
Sciences (USUHS) Animal Care and Use Committee. The opossums used in this study
originated from a captive-bred research colony housed at USUHS (Bethesda, MD, USA).
3.3.2 Tissue Collection, Cell Sorting and RNA Extraction
A 10-month old male gray short-tailed opossum was euthanized by carbon dioxide-
induced asphyxiation, whole spleen and blood samples extracted, and splenocytes (spleen)
and peripheral blood mononuclear cells (blood) isolated by density gradient centrifugation
with Ficoll-Paque Plus (GE Healthcare, Chicago, IL). The purified spleen and blood were
counted and then stained with LIVE/DEAD Aqua (1:800) (Invitrogen, Waltham, MA) to
differentiate viable cells, and then sorted into individual wells on 96-well plates. Single cells
were sorted into individual wells of 96-well plates containing 5 L 1% (v/v) 2-
mercaptoethanol (Sigma-Aldrich, St. Louis, MO) in TCL buffer (Qiagen, Hildon, Germany)
per well. All sample plates were stored at -80C until further use.
3.3.3 cDNA Synthesis, Sequencing and Alignment
The Sciclone G3 Automated Liquid Handler (PerkinElmer, Waltham, MA) was used
for most of the liquid handling. RNA was isolated using the Agencourt RNA Clean XP SPRI
beads (Beckman Coulter, Brea, CA). First strand Master Mix (Illumina, San Diego, CA) was
added to the RNA to conduct first strand cDNA synthesis. Second strand synthesis was

88
conducted using Second Strand Master Mix (Illumina) in a thermocycler to produce the
double-stranded cDNA. Agencourt AMPure XP Beads (Beckman Coulter) and Zephyr G3
NGS workstation (PerkinElmer) were used to isolate the double stranded DNA. The 2100
Bioanalyzer (Agilent, Santa Clara, CA) was used for quality assessment of the PCR products
and concentration were calculated using a Qubit 2.0 Fluorometer (Life Technologies, Grand
Island, NY, USA). Nextera indexed adapters were ligated to cDNA libraries according to
manufacturer’s instructions (Illumina). Nextera libraries were then quantified by qPCR using
KAPA SYBR FAST Master Mix (Kapa Biosystems, Wilmington, MA) according to
manufacturer’s instructions on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad,
Hercules, CA). Up to 96 Nextera libraries were pooled together in 10 nM equimolar
concentrations, were by Illumina HiSeq 3000/4000 instrument (Illumina) performed all
Illumina sequencing to create paired end reads which were demultiplexed to generate FASTQ
files. FASTQC was used to perform the quality assessment of the read data. Reads were
assembled de novo and aligned to the annotated opossum genome. The combined assembled
reads of all tissues resulted in 273 single-cells and 575,721 contigs, with a mean contig length
of 892 bp, and a transcript sum of 513.7 Mb.
3.3.4 Single-cell Identification and Statistical Analyses
Briefly, the identification of paired IgH and IgL chains in the single-cell
transcriptome was performed using BLASTx searched through the NCBI database with an E
value cut-off of 10-5. Open reading frames (ORF) were searched for in the ORFfinder (NCBI)
using the contig sequences which were previously annotated as pseudogenes in the opossum
genome to test whether these sequences were functional in the individual opossum used.
Prism 8.2 (Graphpad) software was used for all statistical analyses and graphing purposes. To
determine the Igλ:Igκ ratio for spleen and blood single-cells, Student’s (unpaired) t-tests was
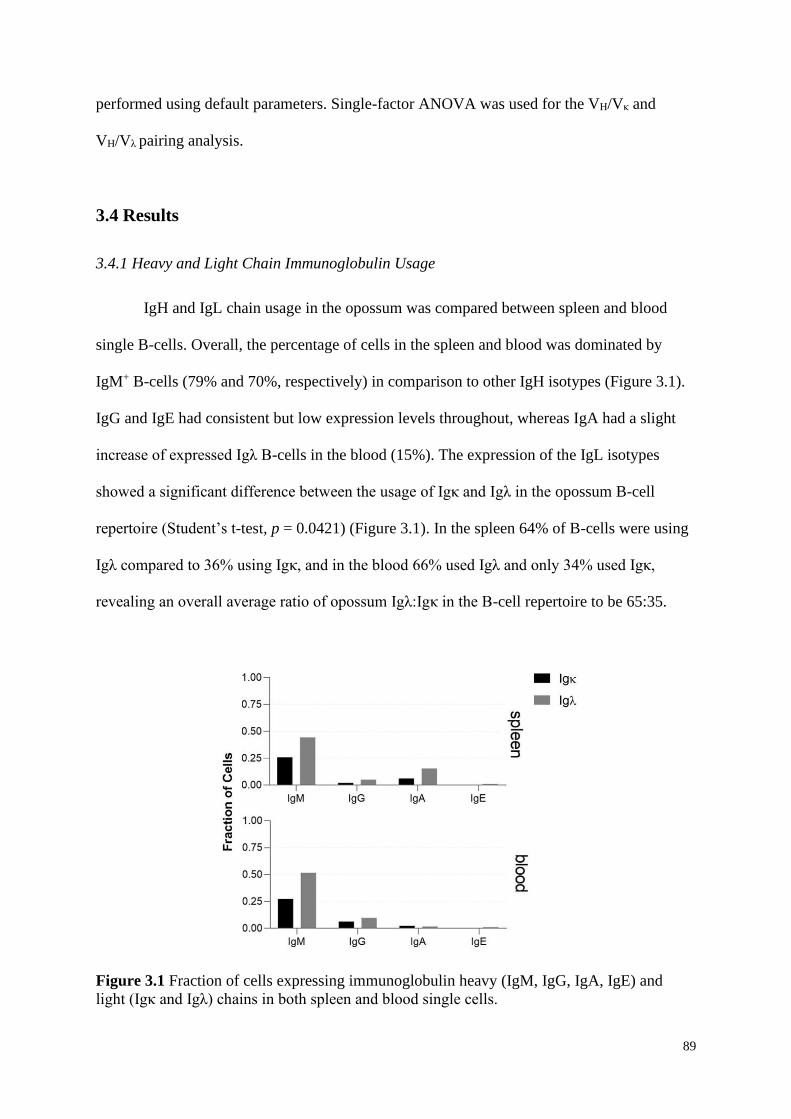
89
performed using default parameters. Single-factor ANOVA was used for the VH/Vκ and
VH/Vλ pairing analysis.
3.4 Results
3.4.1 Heavy and Light Chain Immunoglobulin Usage
IgH and IgL chain usage in the opossum was compared between spleen and blood
single B-cells. Overall, the percentage of cells in the spleen and blood was dominated by
IgM+ B-cells (79% and 70%, respectively) in comparison to other IgH isotypes (Figure 3.1).
IgG and IgE had consistent but low expression levels throughout, whereas IgA had a slight
increase of expressed Igλ B-cells in the blood (15%). The expression of the IgL isotypes
showed a significant difference between the usage of Igκ and Igλ in the opossum B-cell
repertoire (Student’s t-test, p = 0.0421) (Figure 3.1). In the spleen 64% of B-cells were using
Igλ compared to 36% using Igκ, and in the blood 66% used Igλ and only 34% used Igκ,
revealing an overall average ratio of opossum Igλ:Igκ in the B-cell repertoire to be 65:35.
Figure 3.1 Fraction of cells expressing immunoglobulin heavy (IgM, IgG, IgA, IgE) and
light (Igκ and Igλ) chains in both spleen and blood single cells.

90
3.4.2 Immunoglobulin Variable and Joining Domains
Annotation of the opossum IgH and IgL chains used in the single-cell repertoire
revealed limited functional usage of the IgH variable (VH) gene segment. A total of 27 VH
gene segments were used in the opossum B-cell repertoire, in which 6 VH gene segments
were originally annotated as being among the unaligned VH in the opossum genome assembly
(Figure 3.2, (Wang et al., 2009)). A VH gene segment, VH1.10, previously annotated as being
a pseudogene was found in the repertoire and appears to be functional in the individual
opossum used in this study, consistent with it having non-functional alleles rather than being
a pseudogene (Supplementary Figure 3.1). As IgM+ B-cells had the highest overall frequency
among the individual spleen and blood cells, it was expected that it would utilise the majority
of the variable gene segments. Interestingly, a high number of VH1.18 (11%) and VH1.Ue
(14%) gene segments were identified in the blood cells largely due to a high frequency of
IgA+ B-cells; 36% of VH1.18 and 100% of VH1.Ue were utilised by B-cells that had switched
to IgA.
The pattern of recombination between VH and JH was also analysed. There are 6 JH in
the opossum IgH locus, 4 appear functional and 2 pseudogenes (Wang et al., 2009). Only
two, JH1 and JH2, appeared be used in the single-cell repertoires. These are the two functional
JH in closest proximity to the functional C (Wang et al., 2009). JH2 was used in V(D)J
recombinations in 78% of splenic B-cells, while JH1 was used in 22% of splenic B-cells.
There was little difference between blood B-cells using JH1 (44%) and JH2 (56%) (Figure
3.2). The repertoire of both the IgM+ and IgG+ B-cells recombined JH2 more so than JH1
(57% and 89%, respectively) in all cells examined, however blood IgA+ B-cells utilised more
JH1 (70%) than JH2 (30%).
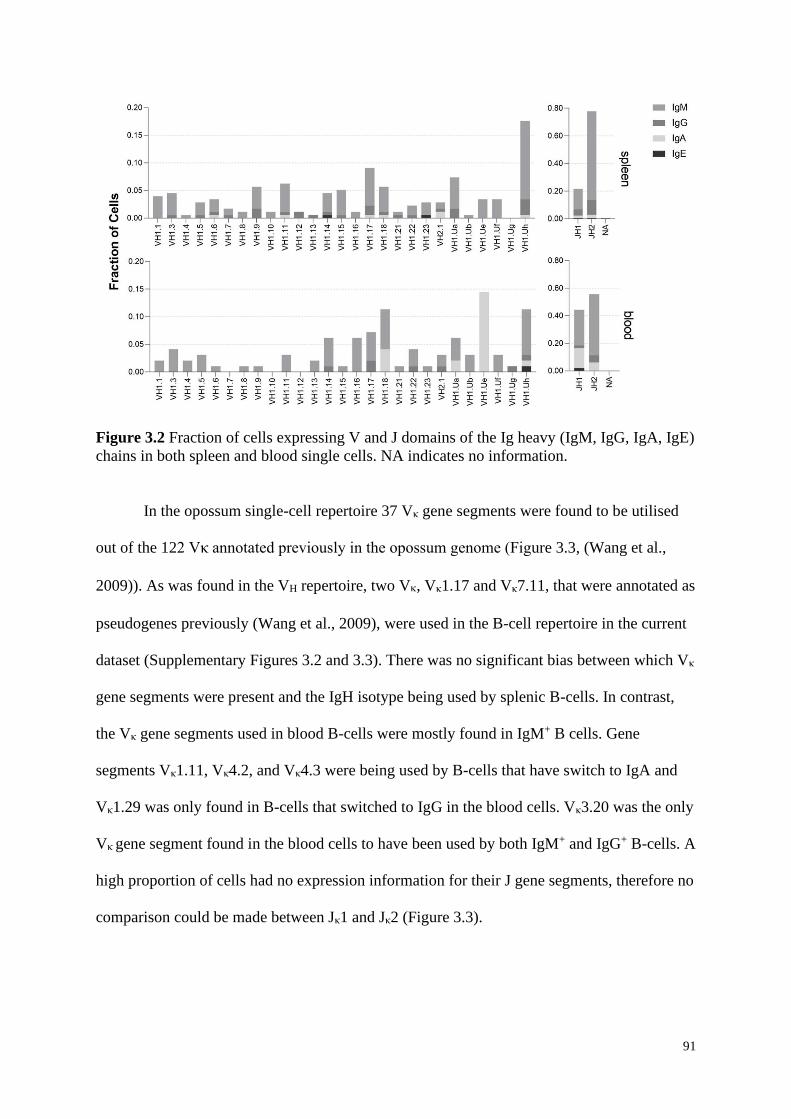
91
Figure 3.2 Fraction of cells expressing V and J domains of the Ig heavy (IgM, IgG, IgA, IgE)
chains in both spleen and blood single cells. NA indicates no information.
In the opossum single-cell repertoire 37 Vκ gene segments were found to be utilised
out of the 122 Vκ annotated previously in the opossum genome (Figure 3.3, (Wang et al.,
2009)). As was found in the VH repertoire, two V, Vκ1.17 and Vκ7.11, that were annotated as
pseudogenes previously (Wang et al., 2009), were used in the B-cell repertoire in the current
dataset (Supplementary Figures 3.2 and 3.3). There was no significant bias between which Vκ
gene segments were present and the IgH isotype being used by splenic B-cells. In contrast,
the Vκ gene segments used in blood B-cells were mostly found in IgM+ B cells. Gene
segments Vκ1.11, Vκ4.2, and Vκ4.3 were being used by B-cells that have switch to IgA and
Vκ1.29 was only found in B-cells that switched to IgG in the blood cells. Vκ3.20 was the only
Vκ gene segment found in the blood cells to have been used by both IgM+ and IgG+ B-cells. A
high proportion of cells had no expression information for their J gene segments, therefore no
comparison could be made between Jκ1 and Jκ2 (Figure 3.3).
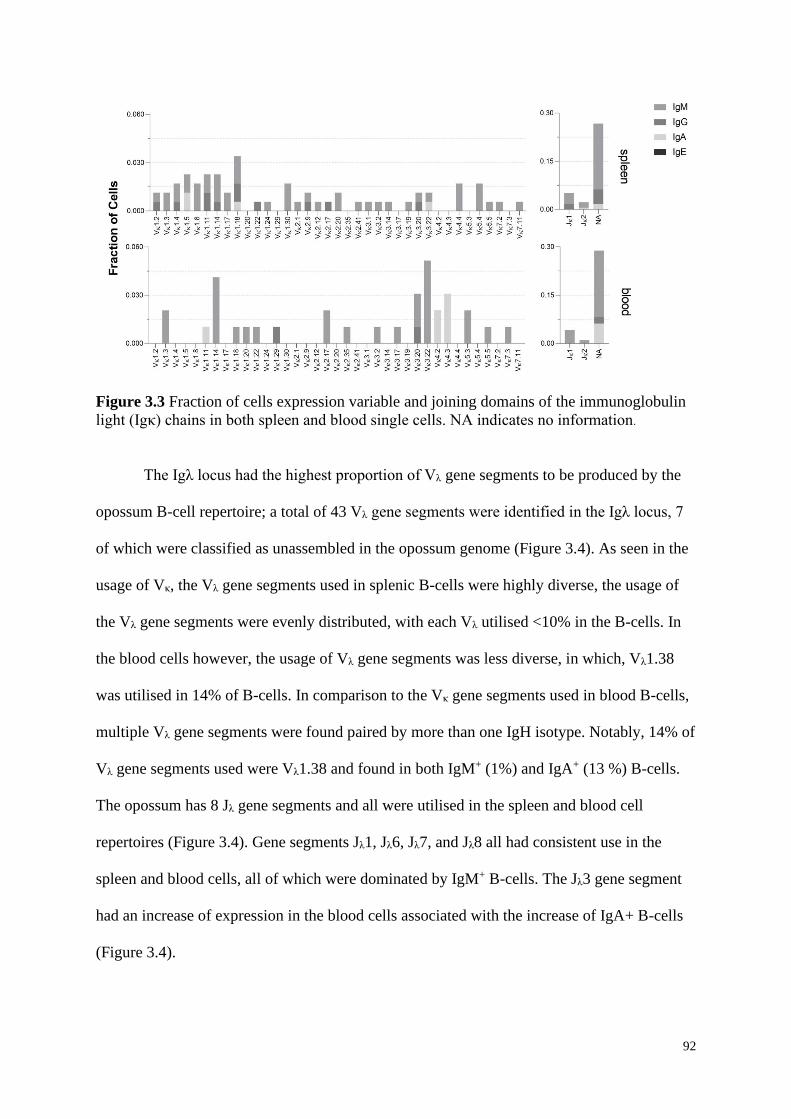
92
Figure 3.3 Fraction of cells expression variable and joining domains of the immunoglobulin
light (Igκ) chains in both spleen and blood single cells. NA indicates no information.
The Igλ locus had the highest proportion of Vλ gene segments to be produced by the
opossum B-cell repertoire; a total of 43 Vλ gene segments were identified in the Igλ locus, 7
of which were classified as unassembled in the opossum genome (Figure 3.4). As seen in the
usage of Vκ, the Vλ gene segments used in splenic B-cells were highly diverse, the usage of
the Vλ gene segments were evenly distributed, with each Vλ utilised <10% in the B-cells. In
the blood cells however, the usage of Vλ gene segments was less diverse, in which, Vλ1.38
was utilised in 14% of B-cells. In comparison to the Vκ gene segments used in blood B-cells,
multiple Vλ gene segments were found paired by more than one IgH isotype. Notably, 14% of
Vλ gene segments used were Vλ1.38 and found in both IgM+ (1%) and IgA+ (13 %) B-cells.
The opossum has 8 Jλ gene segments and all were utilised in the spleen and blood cell
repertoires (Figure 3.4). Gene segments Jλ1, Jλ6, Jλ7, and Jλ8 all had consistent use in the
spleen and blood cells, all of which were dominated by IgM+ B-cells. The Jλ3 gene segment
had an increase of expression in the blood cells associated with the increase of IgA+ B-cells
(Figure 3.4).
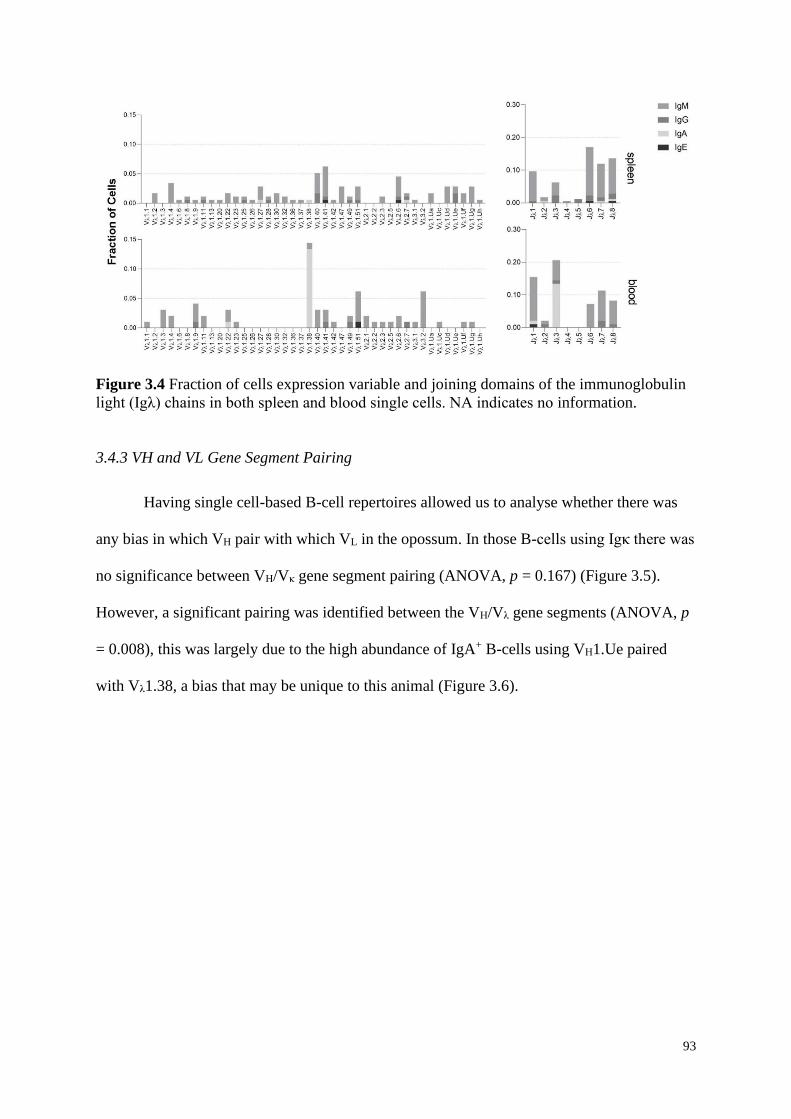
93
Figure 3.4 Fraction of cells expression variable and joining domains of the immunoglobulin
light (Igλ) chains in both spleen and blood single cells. NA indicates no information.
3.4.3 VH and VL Gene Segment Pairing
Having single cell-based B-cell repertoires allowed us to analyse whether there was
any bias in which VH pair with which VL in the opossum. In those B-cells using Igκ there was
no significance between VH/Vκ gene segment pairing (ANOVA, p = 0.167) (Figure 3.5).
However, a significant pairing was identified between the VH/Vλ gene segments (ANOVA, p
= 0.008), this was largely due to the high abundance of IgA+ B-cells using VH1.Ue paired
with Vλ1.38, a bias that may be unique to this animal (Figure 3.6).
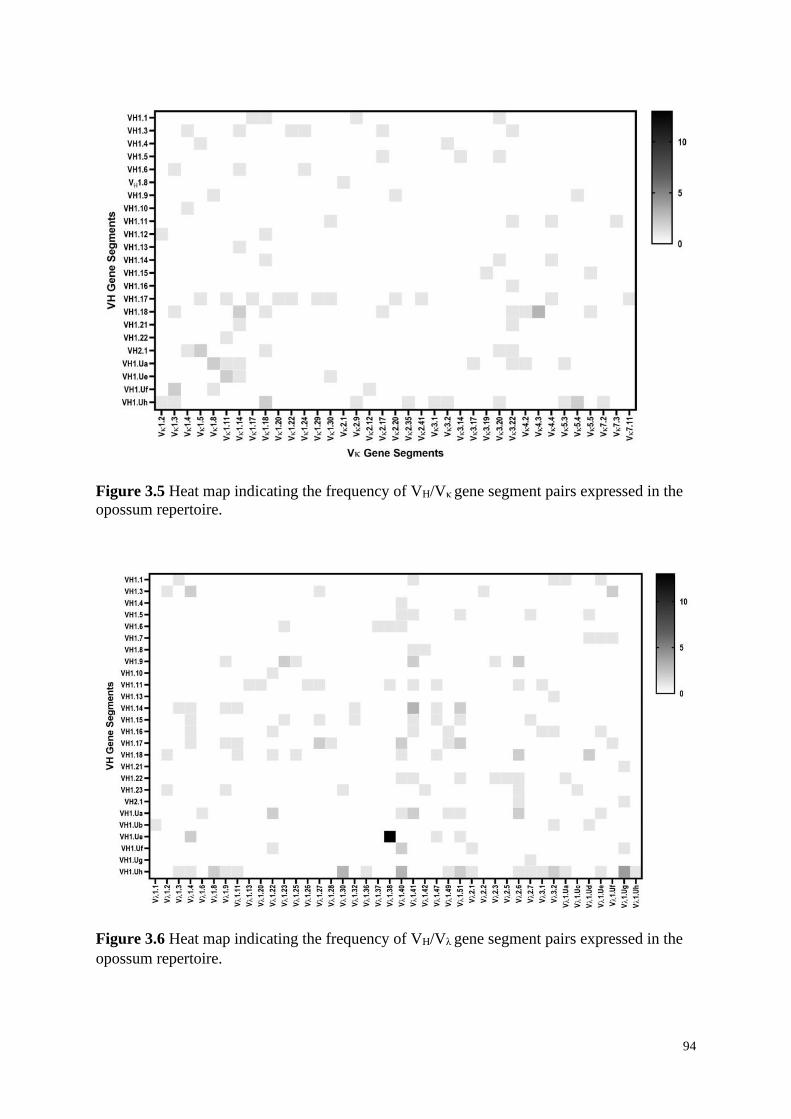
94
Figure 3.5 Heat map indicating the frequency of VH/Vκ gene segment pairs expressed in the
opossum repertoire.
Figure 3.6 Heat map indicating the frequency of VH/Vλ gene segment pairs expressed in the
opossum repertoire.

95
3.5 Discussion
All three modern mammalian lineages, eutherians, marsupials, and monotremes, have
typical Ig molecules consisting of two identical heavy (H) and light (L) chains (Baker et al.,
2005). However, unlike eutherians and monotremes, marsupials only have four of the five
IgH isotypes (IgD has not been identified) (Miller and Belov, 2000), along with both IgL
isotypes; Igλ and Igκ (Lucero et al., 1998, Miller et al., 1999). The goal of this study was to
analyse the expression of IgH and IgL loci in the single-cell transcriptome of the opossum B-
cells and examine their usage of the variable and joining gene segments.
In contrast to other mammals, marsupial IgH constant (C) regions appear to only
contain a single functional copy of IgM, IgG, IgA and IgE subclasses (Aveskogh and
Hellman, 1998, Miller et al., 1998). Both the monotremes investigated, the platypus and
short-beaked echidna (Tachyglossus aculeatus), each have two IgA and IgG subtypes
(Vernersson et al., 2002, Belov and Hellman, 2003). There is a second IgM locus identified in
the gray short-tailed opossum, however, it appears to be a non-functional pseudogene (Wang
et al., 2009, Miller, 2010). Analyses found more IgM+ B-cells than other IgH isotypes in the
opossum repertoire. This is not surprising given the cells analysed came from a naïve animal
and IgM is the first Ig to be expressed during ontogeny of the immune system (Lawton et al.,
1975, Belov et al., 1999c),
The VH domain is encoded by an exon assembled from gene segments: the variable
(V), diversity (D), and joining (J) gene segments, through V(D)J recombination (Alt et al.,
1992, Schatz and Swanson, 2011). In contrast, IgL isotypes use only V and J gene segments
to encode the V domain. The limitation in the VH domain diversity appears to have occurred
during early marsupial evolution, with previous investigators reporting a limited number of
VH families in marsupials (Miller et al., 1998, Baker et al., 2005, Wang et al., 2009). Three
major groups of VH genes are found across vertebrates (Kabat et al., 1992). These groups are

96
further divided into families (or subgroups) based generally on nucleotide sequence identity
and/or cross-hybridization (Brodeur and Riblet, 1984). Most of the VH in the opossum belong
to a single family (VH1), and there are two other families (VH2 and VH3) that have a single
gene segment in each (Miller et al., 1998, Baker et al., 2005, Wang et al., 2009, Wang and
Miller, 2012). Our study found 21 of the annotated VH segments (VH2.1 included) used in the
B-cell repertoire, one of which, VH1.10, had been previously designated as a pseudogene
(Wang et al., 2009). Our results suggest that VH1.10 is actually functional in the individual
animal investigated due to the presence of an ORF identified in the transcript contigs,
indicating that VH1.10 can be a functional allele, rather than a pseudogene. Curiously, VH3.1
was not found in our dataset, in spite of it having previously shown to be used in the opossum
IgH repertoire (Wang and Miller, 2012). Only two of the six identified JH gene segments (JH1
and JH2) were found to be used in the repertoire analysed in this dataset. JH gene segments are
grouped into two sets of three, in which JH1 and JH2 are directly upstream of the functional
IgM constant region in the IgH genomic organisation (Wang et al., 2009). JH1 and JH2 are
also the only JH gene segments to have been previously found in IgH encoding cDNAs
isolated from adult opossum spleen (Aveskogh et al., 1999).
Our study is the first to determine the relative usage of Igλ and Igκ in any marsupial
species. The opossum has a Igλ:Igκ ratio of 65:35, whereas in eutherian mammals this ratio is
highly diverse among species (see table 3.1; reviewed in Nezlin (1998)).
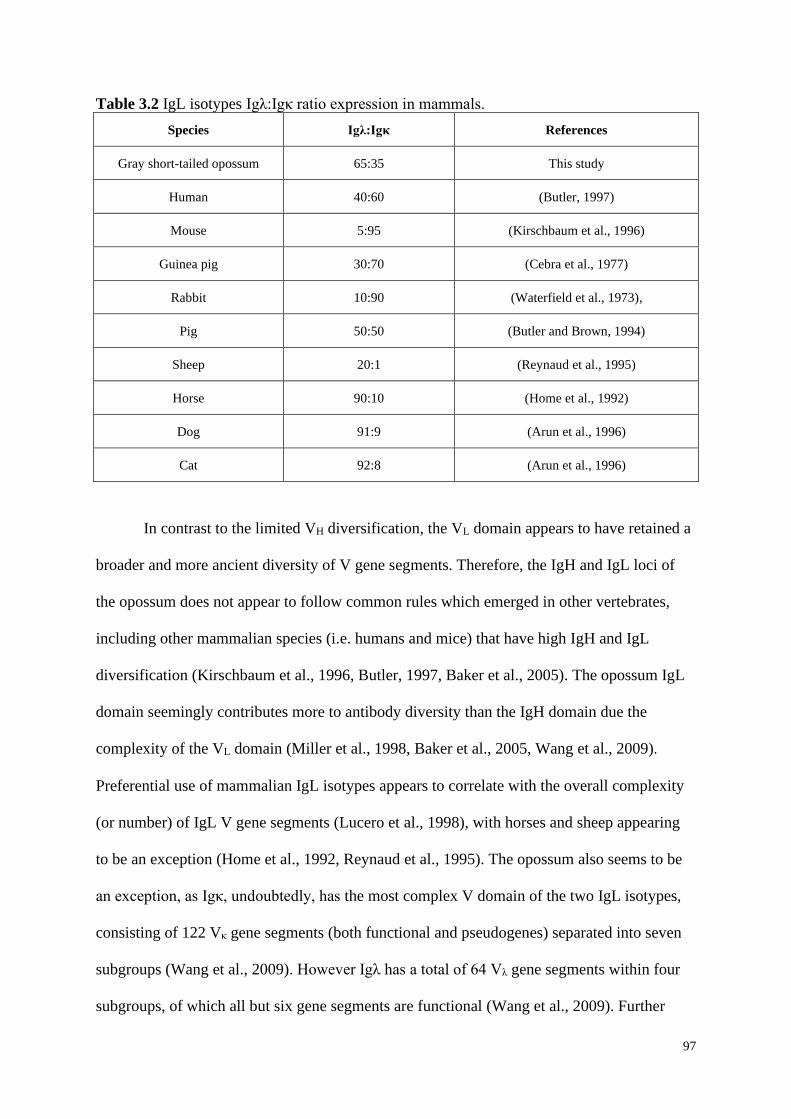
97
Table 3.2 IgL isotypes Igλ:Igκ ratio expression in mammals.
Species Igλ:Igκ References
Gray short-tailed opossum 65:35 This study
Human 40:60 (Butler, 1997)
Mouse 5:95 (Kirschbaum et al., 1996)
Guinea pig 30:70 (Cebra et al., 1977)
Rabbit 10:90 (Waterfield et al., 1973),
Pig 50:50 (Butler and Brown, 1994)
Sheep 20:1 (Reynaud et al., 1995)
Horse 90:10 (Home et al., 1992)
Dog 91:9 (Arun et al., 1996)
Cat 92:8 (Arun et al., 1996)
In contrast to the limited VH diversification, the VL domain appears to have retained a
broader and more ancient diversity of V gene segments. Therefore, the IgH and IgL loci of
the opossum does not appear to follow common rules which emerged in other vertebrates,
including other mammalian species (i.e. humans and mice) that have high IgH and IgL
diversification (Kirschbaum et al., 1996, Butler, 1997, Baker et al., 2005). The opossum IgL
domain seemingly contributes more to antibody diversity than the IgH domain due the
complexity of the VL domain (Miller et al., 1998, Baker et al., 2005, Wang et al., 2009).
Preferential use of mammalian IgL isotypes appears to correlate with the overall complexity
(or number) of IgL V gene segments (Lucero et al., 1998), with horses and sheep appearing
to be an exception (Home et al., 1992, Reynaud et al., 1995). The opossum also seems to be
an exception, as Igκ, undoubtedly, has the most complex V domain of the two IgL isotypes,
consisting of 122 Vκ gene segments (both functional and pseudogenes) separated into seven
subgroups (Wang et al., 2009). However Igλ has a total of 64 Vλ gene segments within four
subgroups, of which all but six gene segments are functional (Wang et al., 2009). Further

98
analyses of marsupial IgL isotype expression will be required to determine whether the entire
marsupial lineage follows the same expression pattern as the opossum, or like eutherians,
whereby a high dissimilarity between species occurs. Only 37 Vκ gene segments were found
to be used in the opossum Ig repertoire; 14 from Vκ1, seven from Vκ2 and Vκ3, four from Vκ4
and Vκ5, and three from Vκ7. Previously, Vκ1.17 and Vκ7.11 have both been annotated as
pseudogenes in the opossum genome (Mikkelsen et al., 2007, Wang et al., 2009). However,
our findings reveal that these gene segments are functional due to an ORF in the assembled
sequence contigs, suggesting that there are functional alleles in some individuals. No
transcripts were found to express any of the gene segments from the Vκ6 subgroup. In
comparison, 35 previously annotated Vλ gene segments were used in the opossum Ig
repertoire; 27 from Vλ1, 6 from Vλ2, and 2 from Vλ3. No Vλ4.1. The Vλ4 gene family is only
newly identified in the opossum and appears functional (Wang et al., 2009), and it still
remains unclear if Vλ4 contributes to antibody diversity. Analysis of the gene segment pairing
showed a significant difference between cells using VH/Vλ pairs, which was largely due the
high abundance of cells expressing VH1.Ue:Vλ1.38 in IgA+ B-cells. This may reflect selection
that has occurred on the B-cell pool in this opossum that expanded those B-cells using
VH1.Ue:Vλ1.38.
J and C domains vary between opossum IgL isotypes. The opossum Ig locus
contains only a single Cκ gene and two Jκ gene segments (Wang et al., 2009). Although the
dataset contained full length Ig transcripts, we could not distinguish which Jκ were being
used. The opossum has eight Jλ-Cλ pairs that have previously been identified (Lucero et al.,
1998, Wang et al., 2009), all of which were being used in the B-cell repertoire. The presence
of multiple Jλ-Cλ gene pairs in the opossum is a feature conserved across mammals
(Kirschbaum et al., 1996), whereas birds only contain one Jλ-Cλ pairing (McCormack et al.,
1989).

99
It should also be mentioned that a number of “unaligned” gene segments were utilized
in the opossum B-cells; six from VH belonging to the VH1 subgroup, and seven from Vλ1
subgroup. These sequences have previously been identified in the opossum genome, but it is
still unclear if any of these are alleles of VH annotated in the IgH locus or were unique gene
segments that are located in gaps in the assembly (Mikkelsen et al., 2007).
Our study has provided valuable insights into the Ig gene repertoire in opossum B-
cells. Until now, no studies have investigated the usage of IgL isotypes, Igλ and Igκ, in any
marsupial species at the single-cell level, and this information can now be utilised as a key
resource for future analysis of marsupial Ig usage. However, as it has already been seen in the
eutherian lineage, the Igλ:Igκ ratio is highly variable between species, therefore, we cannot
suppose the same variation occurs in all marsupials.
3.6 Acknowledgements
The authors would like to thank Galina Petukhova (USUHS) for generously making
available M. domestica for this project and assistance with animal husbandry and tissue
collection. This research was funded in part by a National Science Foundation award IOS-13531232.
Research reported in this publication was also supported in part by the National Institute of General
Medical Sciences of the National Institutes of Health under award P30 GM110907. The content is
solely the responsibility of the authors and does not necessarily represent the official views of the
National Institutes of Health.
3.7 Conflicts of Interest
The authors declare no conflicts of interest.

100
3.8 Chapter 3 References
ALT, F. W., OLTZ, E. M., YOUNG, F., GORMAN, J., TACCIOLI, G. & CHEN, J. 1992.
VDJ recombination. Immunology today, 13, 306-314.
ARUN, S., BREUER, W. & HERMANNS, W. 1996. Immunohistochemical examination of
light‐chain expression (Λ/k ratio) in canine, feline, equine, bovine and porcine plasma
cells. Journal of Veterinary Medicine Series A, 43, 573-576.
AVESKOGH, M. & HELLMAN, L. 1998. Evidence for an early appearance of modern post-
switch isotypes in mammalian evolution; cloning of IgE, IgG and IgA from the
marsupial Monodelphis domestica. European Journal of Immunology, 28, 2738-2750.
AVESKOGH, M., PILSTROM, L. & HELLMAN, L. 1999. Cloning and structural analysis
of IgM (μ chain) and the heavy chain V region repertoire in the marsupial
Monodelphis domestica. Developmental & Comparative Immunology, 23, 597-606.
BAKER, M. L., BELOV, K. & MILLER, R. D. 2005. Unusually similar patterns of antibody
V segment diversity in distantly related marsupials. The Journal of Immunology, 174,
5665-5671.
BELOV, K., HARRISON, G. A. & COOPER, D. W. 1998. Molecular cloning of the cDNA
encoding the constant region of the immunoglobulin A heavy chain (Cα) form a
marsupial: Trichosurus vulpecula (common brushtail possum). Immunology Letters,
60, 165-170.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999a. Isolation and
sequence of a cDNA coding for the heavy chain constant region of IgG from the
Australian brushtail possum, Trichosurus vulpecula. Molecular Immunology, 36, 535-
541.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999b. Molecular
cloning of the brushtail possum (Trichosurus vulpecula) immunoglobulin E heavy
chain constant region. Molecular Immunology, 36, 1255-1261.
BELOV, K., HARRISON, G. A., ROSENBERG, G. H., MILLER, R. D. & COOPER, D. W.
1999c. Isolation and comparison of the IgM heavy chain constant regions from
Australian (Trichosurus vulpecula) and American (Monodelphis domestica)
marsupials. Developmental & Comparative Immunology, 23, 649-656.
BELOV, K. & HELLMAN, L. 2003. Immunoglobulin genetics of Ornithorhynchus anatinus
(platypus) and Tachyglossus aculeatus (short-beaked echidna). Comparative
Biochemistry and Physiology, Part A, 136, 811-819.
BRODEUR, P. H. & RIBLET, R. 1984. The immunoglobulin heavy chain variable region
(Igh‐V) locus in the mouse. I. one hundred Igh‐V genes comprise seven families of
homologous genes. European Journal of Immunology, 14, 922-930.
BUTLER, J. E. 1997. Immunoglobulin gene organization and the mechanism of repertoire
development. Scandinavian Journal of Immunology, 59, 455-462.
BUTLER, J. E. & BROWN, W. R. 1994. The immunoglobulins and immunoglobulin genes
of swine. Veterinary Immunology and Immunopathology, 43, 5-12.
CEBRA, J., BRUNHOUSE, R., CORDLE, C., DAISS, J., FECHHEIMER, M., RICARDO,
M., THUNBERG, A. & WOLFE, P. 1977. Isotypes of guinea pig antibodies:
restricted expression and bases for interactions with other molecules. Progress in
Immunology, 111, 269.
DEAKIN, J. E., OLP, J. J., GRAVES, J. A. M. & MILLER, R. D. 2006. Physical mapping of
immunoglobulin loci IgH@, IgK@, and IgL@ in the opossum (Monodelphis
domestica). Cytogenetic and Genome Research, 114, 94.

101
HOME, W. A., FORD, J. E. & GIBSON, D. M. 1992. L chain isotype regulation in horse. i.
characterization of Ig lambda genes. Journal of Immunology 149, 3927-3936.
KABAT, E. A., TE WU, T., PERRY, H. M., FOELLER, C. & GOTTESMAN, K. S. 1992.
Sequences of proteins of immunological interest, DIANE publishing.
KIRSCHBAUM, T., JAENICHEN, R. & ZACHAU, H. G. 1996. The mouse
immunoglobulin ϰ locus contains about 140 variable gene segments. European
Journal of Immunology, 26, 1613-1620.
LAWTON, A. R., KINCADE, P. W. & COOPER, M. D. 1975. Sequential expression of
germ line genes in development of immunoglobulin class diversity. Federation
Proceedings, 34, 33-39.
LUCERO, J. E., ROSENBERG, G. H. & MILLER, R. D. 1998. Marsupial light chains:
complexity and conservation of λ in the opssum Monodelphis domestica. The Journal
of Immunology, 161, 6724-6732.
MCCORMACK, W., CARLSON, L., TJOELKER, L. & THOMPSON, C. 1989.
Evolutionary comparison of the avian Ig sub(L) locus: combinatorial diversity plays a
role in the generation of the antibody repertoire in some avian species. International
Immunology, 1, 332-341.
MIKKELSEN, T. S., WAKEFIELD, M., AKEN, B., AMEMIYA, C., CHANG, J., DUKE
BECKER, S., GARBER, M., GENTLES, A. J., GOODSTADT, L., HEGER, A.,
JURKA, J., KAMAL, M., MAUCELI, E., SEARLE, S., SHARPE, T., BAKER, M.
L., BATZER, M. A., BENOS, P., BELOV, K. & LINDBLAD-TOH, K. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in non-coding
sequences. Nature 447, 167-77.
MILLER, R. D. 2010. Those other mammals: the immunoglobulins and T cell receptors of
marsupials and monotremes. Seminars in Immunology, 22, 3-9.
MILLER, R. D. & BELOV, K. 2000. Immunoglobulin genetics of marsupials.
Developmental & Comparative Immunology, 24, 485-490.
MILLER, R. D., BERGEMANN, E. R. & ROSENBERG, G. H. 1999. Marsupial light
chains: IgK with four V families in the opossum Monodelpis domestica.
Immunogenetics, 50, 329-335.
MILLER, R. D., GRABE, H. & ROSENBERG, G. H. 1998. VH repertoire of a marsupial
(Monodelphis domestica). The Journal of Immunology, 160, 259-365.
NEZLIN, R. 1998. CHAPTER 2 - Animal and Human Immunoglobulins. In: NEZLIN, R.
(ed.) The Immunoglobulins. New York: Academic Press.
OHTA, Y. & FLAJNIK, M. 2006. IgD, like IgM, is a primordial immunoglobulin class
perpetuated in most jawed vertebrates. Proceedings of the National Academy of
Sciences of the United States of America, 103, 10723-10728.
REYNAUD, C.-A., GARCIA, C., HEIN, W. R. & WEILL, J.-C. 1995. Hypermutation
generating the sheep immunoglobulin repertoire is an antigen-independent process.
Cell, 80, 115-125.
SAMPLES, N. K., VANDEBERG, J. L. & STONE, W. H. 1986. Passively acquired
immunity in the newborn of a marsupial (Monodelphis domestica). American Journal
of Reproductive Immunology and Microbiology, 11, 94-97.
SCHATZ, D. G. & SWANSON, P. C. 2011. V(D)J recombination: mechanisms of initiation.
Annual Review of Genetics, 45, 167-202.
SITNIKOVA, T. & SU, C. 1998. Coevolution of immuoglobulin heavy- and light-chain
variable-region gene families. Immunogenetics, 15, 617-625.
VERNERSSON, M., AVESKOGH, M., MUNDAY, B. & HELLMAN, L. 2002. Evidence
for an early appearance of modern post-switch immunoglobulin isotypes in
mammalian evolution (ii); cloning of IgE, IgG1 and IgG2 from a monotreme, the

102
duck-billed platypus, Ornithorhynchus anatinus. European Journal of Immunology,
32, 2145-2155.
WANG, X. & MILLER, R. D. 2012. Recombination, transcription, and diversity of a
partially germline-joined VH in a mammal. Immunogenetics, 64, 713-7.
WANG, X., OLP, J. J. & MILLER, R. D. 2009. On the genomics of immunoglobulins in the
gray, short-tailed opossum Monodelphis domestica. Immunogenetics, 61, 581-96.
WANG, X., SHARP, A. R. & MILLER, R. D. 2012. Early postnatal B cell ontogeny and
antibody repertoire maturation in the opossum, Monodelphis domestica. PLoS One, 7,
e45931.
WATERFIELD, M. D., MORRIS, J. E., HOOD, L. E. & TODD, C. W. 1973. Rabbit
immunoglobulin light chains: correlation of variable region sequences with allotypic
markers. The Journal of Immunology, 110, 227.

103
3.9 Chapter 3 Supplementary Figures
Supplementary Figure 3.1 Previously annotated pseudogene VH1.10. Identification of ORF
on the reverse strand of frame 1, 3’ – 5’. Start codon, 2250; stop codon, 415; length, 612
amino acids. Stop codon is indicated by the asterisks.

104
Supplementary Figure 3.2 Previously annotated pseudogene Vκ1.17. Identification of ORF
on the reverse strand of frame 1, 3’ – 5’. Start codon, 273; stop codon, 37; length, 79 amino
acids. Stop codon is indicated by the asterisks.
Supplementary Figure 3.3 Previously annotated pseudogene Vκ7.11. Identification of ORF
on the reverse strand of frame 3, 3’ – 5’. Start codon, 990; Stop codon, 235; length, 252
amino acids. Stop codon is indicated by the asterisks.

105
CHAPTER 4
Single-cell transcriptomic analysis of marsupial
B-cells, the gray short-tailed opossum
(Monodelphis domestica)
A. L. Schraven1, V. L. Hansen2, H. J. Stannard3, O. T.W. Ong4, D. C. Douek5, R. D. Miller2,
J. M. Old1
1School of Science and Health, Hawkesbury Campus, Western Sydney University, Locked
bag 1797, Penrith, NSW 2751, Australia
2Center for Evolutionary and Theoretical Immunology, Department of Biology, University of
New Mexico Albuquerque, New Mexico, USA
3Charles Sturt University, School of Animal and Veterinary Sciences, Wagga Wagga, NSW
2678, Australia
4QIMR Berghofer Medical Research Institute, Brisbane, Queensland, Australia
5Human Immunology Section, Vaccine Research Center, National Institute of Allergy and
Infectious Diseases, National Institutes of Health, Bethesda, Maryland, USA
Manuscript has been prepared for submission to Immunology and Cell Biology

106
4.1 Chapter Outline and Authorship
Chapter 4 presents an investigation of the gray short-tailed opossum (Monodelphis
domestica) B-cell transcriptome at the single cell level. This chapter annotates all the genes
identified in the opossum and analyses the most highly expressed genes, inclusive of those
which are associated with marsupial immune system.
This chapter is a prepared manuscript and is jointly authored; I am the primary author,
analysed the raw data, identified and annotated the genes, performed structural motif and
phylogenetic analysis, and prepared the manuscript. Victoria Hansen performed Cell sorting,
RNA extraction and cDNA synthesis, and Daniel Douek performed the sequencing. Robert
Miller, Hayley Stannard, Oselyne Ong and Julie Old provided advice on data analysis, and
have reviewed and provided feedback on the manuscript.
This chapter is a manuscript that has been prepared for submission to Immunology and Cell
Biology, and is therefore formatted to the journal specifications.

107
4.2 Introduction
The gray short-tailed opossum (Monodelphis domestica) is arguably a leading
marsupial model for evolutionary research (Samollow, 2006), as it has been widely used as a
model for comparative and biomedical studies (Vandeberg and Robinson, 1997). Since the
sequencing of the opossum genome (Mikkelsen et al., 2007), many components of the
marsupial innate and adaptive immune system have been characterised (Wong et al., 2011,
Edwards et al., 2012, Morris et al., 2018), and the opossum Immunoglobulin (Ig), T cell
receptor (TCR), and major histocompatibility complex (MHC) genes have been annotated
(Wang et al., 2009). The resulting information suggests that the marsupial immune system is
highly similar, and as complex, to that of eutherian mammals.
Marsupials are a unique group of mammals and of great interest in immune system
development and functionality studies (Old and Deane, 2000). As with all marsupials, the
opossum is born at an early stage of immunological development after a considerably short
gestational period of only 13.5 days (Mate et al., 1994). Unlike eutherian mammals, physical
and physiological development is completed ex utero while attached to the mother’s teat
(Edwards and Deakin, 2013). During this time, the young will endure a lengthy lactation
period and undergo major developmental stages including the maturation of lymphoid tissues
and cells.
B-cells are of particular importance in humoral (adaptive) immune responses and are
found in multiple lymphoid organs (Holmes et al., 2008). B-cells are unique because of their
production of antibodies released from their differentiated cells and are able to detect foreign
molecules that invade the (Painter et al., 2011). In mice, B-cells are produced from the
pluripotent haematopoietic stem cells, which are found in the liver during the late stages of
foetal development and haematopoiesis continues in the bone marrow after birth (Hardy and
Hayakawa, 2001). Several stages are involved in the maturation of eutherian B-cells,

108
including the rearrangement of Igs, expression of CD79α+ and CD79β+ cells, and their
population in both primary and secondary immune tissues (Conley and Cooper, 1998). In the
opossum, detection of Ig rearrangement has only been expressed as early as day 7,
postpartum, suggesting marsupial neonates cannot produce an endogenous antibody response
until after the first postnatal week (Wang et al., 2012). The earliest time point detected for
CD79β+ cells was identified in the gut of the tammar wallaby (Notamacropus eugenii) at day
7 postpartum (Old and Deane, 2003). By day 10, B-cell receptor CD79α+ and CD79β+
transcripts were detected in cervical thymus and bone marrow, along with CD79α+ in the
spleen, gut and blood of the wallaby (Duncan et al., 2010). In the common brushtail possum
(Trichosurus vulpecula) IgM transcripts were detected as early as day 10 postpartum, and
only after two months, switched to IgG (Belov et al., 2002). Likewise, this switch to IgG was
detected in the opossum during the fifth postnatal week, when permanent detachment of the
mother’s teat has ceased (Wang et al., 2012).
There are many genes that are specifically expressed in B-cells, throughout
development, differentiation and regulation. Recent advances in genomics have enabled
researchers to make comparisons between marsupial and eutherian immune systems
(Samollow, 2008), and several investigations have found many overlapping similarities of the
structural and functional components of B-cells in groups of mammals (Stone et al., 1996,
Miller and Belov, 2000, Wang et al., 2012). Here, we identify and annotate the expressed
genes of the gray short-tailed opossum B-cell transcriptome at the single cell level.

109
4.3 Materials and Methods
4.3.1 Ethics Statement
This study was approved under the protocol numbers 16-200407-MC and 15-200334-B-
MC from the University of New Mexico Institutional Animal Care and Use Committee and
protocol numbers BIO-17-969 from the USUHS Animal Care and Use Committee. The
opossums used in this study originated from a captive-bred research colony housed at the
Uniformed Services University of the Health Sciences (USUHS; Bethesda, MD, USA).
4.3.2 Tissue Collection, Cell Sorting and RNA Extraction
A 10-month old male gray short-tailed opossum was euthanized by carbon dioxide-
induced asphyxiation, whole spleen and blood samples extracted, and splenocytes (spleen)
and peripheral blood mononuclear cells (blood) isolated by density gradient centrifugation
with Ficoll-Paque Plus (GE Healthcare, Chicago, IL). The purified spleen and blood were
counted and then stained with LIVE/DEAD Aqua (1:800) (Invitrogen, Waltham, MA) to
differentiate viable cells, and then sorted into individual wells on 96-well plates. Single cells
were sorted into individual wells of 96-well plates containing 5 L 1% (v/v) 2-
mercaptoethanol (Sigma-Aldrich, St. Louis, MO) in TCL buffer (Qiagen, Hildon, Germany)
per well. All sample plates were stored at -80C until further use.
4.3.3 cDNA Synthesis, Sequencing and Alignment
The Sciclone G3 Automated Liquid Handler (PerkinElmer, Waltham, MA) was used
for most of the liquid handling. RNA was isolated using the Agencourt RNA Clean XP SPRI
beads (Beckman Coulter, Brea, CA). First strand Master Mix (Illumina, San Diego, CA) was
added to the RNA to conduct first strand cDNA synthesis. Second strand synthesis was
conducted using Second Strand Master Mix (Illumina) in a thermocycler to produce the

110
double-stranded cDNA. Agencourt AMPure XP Beads (Beckman Coulter) and Zephyr G3
NGS workstation (PerkinElmer) were used to isolate the double stranded DNA. The 2100
Bioanalyzer (Agilent, Santa Clara, CA) was used for quality assessment of the PCR products
and concentration were calculated using a Qubit 2.0 Fluorometer (Life Technologies, Grand
Island, NY, USA). Nextera indexed adapters were ligated to cDNA libraries according to
manufacturer’s instructions (Illumina). Nextera libraries were then quantified by qPCR using
KAPA SYBR FAST Master Mix (Kapa Biosystems, Wilmington, MA) according to
manufacturer’s instructions on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad,
Hercules, CA). Up to 96 Nextera libraries were pooled together in 10 nM equimolar
concentrations, were by Illumina HiSeq 3000/4000 instrument (Illumina) performed all
Illumina sequencing to create paired end reads which were demultiplexed to generate FASTQ
files. FASTQC was used to perform the quality assessment of the read data. Reads were
assembled de novo and aligned to the annotated opossum genome. The combined assembled
reads of all tissues resulted in 273 single-cells and 575,721 contigs, with a mean contig length
of 892 bp, and a transcript sum of 513.7 Mb.
4.3.4 Transcriptome Annotation and Gene Ontology
Functional transcriptome annotation was performed using the Galaxy analysis
platform (Afgan et al., 2018) (https://usegalaxy.org/). Briefly, BLASTn searches were
performed using the Ensembl database as the target with an E value cut-off of 10-5. The
annotated genes matched a total of 14,654 unique Ensembl or Genbank genes. Gene ontology
(GO) analyses of all annotated genes was performed using online Panther tools (Mi et al.,
2019) (http://pantherdb.org/) to gain on overview of the assembled opossum transcriptome.
Genes were mapped to the opossum reference genome and classified under GO-Slim
Molecular Function, Biological Process, and Cellular Component terms. Any “unclassified”

111
terms were ignored in all analyses. Prism 8.2 (Graphpad) software was used to graph GO
annotation percentages.
4.3.5 Statistical Analyses
For the comparison of the most highly expressed genes in the in the single-cell
transcriptome of the opossum B-cells, PRIMER-e (v7) (Clarke et al., 2014) was used.
Analysis of Similarity (ANOSIM) was used to compare the genes between the spleen and
blood cells. To compare the genes expressed in the individual transcriptomes, ANOSIM and
Similarity Percentage Analysis (SIMPER) was used, and Ig heavy isotypes were grouped
together. Non-transformed multidimensional scaling plots were used to visualise the
multivariate differences in gene expression.
4.3.6 Phylogenetic Analyses and Motif Comparison
Geneious Prime 2019.2 (Kearse et al., 2012) (http://www.geneious.com) software was
used to analyse the evolutionary relationship between mammalian lineages of the top 10
highly expressed genes and the ribosomal protein family genes. Protein sequences from the
gray short-tailed opossum, Tasmanian devil (Sarcophilus harrisii), koala (Phascolarctos
cinereus), bare-nosed wombat (Vombatus ursinus), tammar wallaby (Notamacropus eugenii),
common brushtail possum (Trichosurus vulpecula), platypus (Ornithorhynchus anatinus),
human (Homo sapiens), and mouse (Mus musculus) were used for phylogenetic tree
constructions. Sequences were aligned in BioEdit (Hall, 1999)
(http://www.mbio.nscu.edu./BioEdit/bioedit.html) using the ClustalW algorithm (Thompson
et al., 1994), and phylogenetic trees were constructed based on the alignments in Geneious
Prime using Blossum62 (Henikoff and Henikoff, 1992) with a global alignment. The
unrooted morphological phylogenetic trees were constructed using the neighbour-joining tree

112
build method, and the sequences were evolved according to the Jukes-Cantor model using
1000 bootstrap replicates (Jukes and Cantor, 1969). Additionally, sequence motif analyses
were conducted to compare the 10 most highly expressed genes in the opossum B-cell
transcriptome with other mammals. Protein sequences were entered into MOTIF (Pagni et al.,
2007) (http://www.genome.ad.jp/) using the PROSITE pattern notation. The functionally
important residues and sites were labelled and amino acids with >75% identity were
highlighted.
4.4 Results
4.4.1 Overview of the Transcriptome
The B-cell transcriptome was assembled and annotated using the opossum spleen and
blood samples. Of the 14,654 unique gene hits, 13,376 genes were assigned GO-slim
annotation classifications (Figure 4.1). In the molecular function category, the most common
GO terms included ‘binding’ (37.5%) and ‘catalytic activity’ (35.3%). The most common
cellular component categories were ‘cell’ (42.0% of genes), ‘organelle’ (30.9%), and ‘protein
coding complex’ (13.5%). In the biological process category, the highest abundance of genes
came under the ‘cellular process’ category (32.1%), followed by ‘metabolic process’ (32.1%)
and ‘biological regulation’ (15.1%).
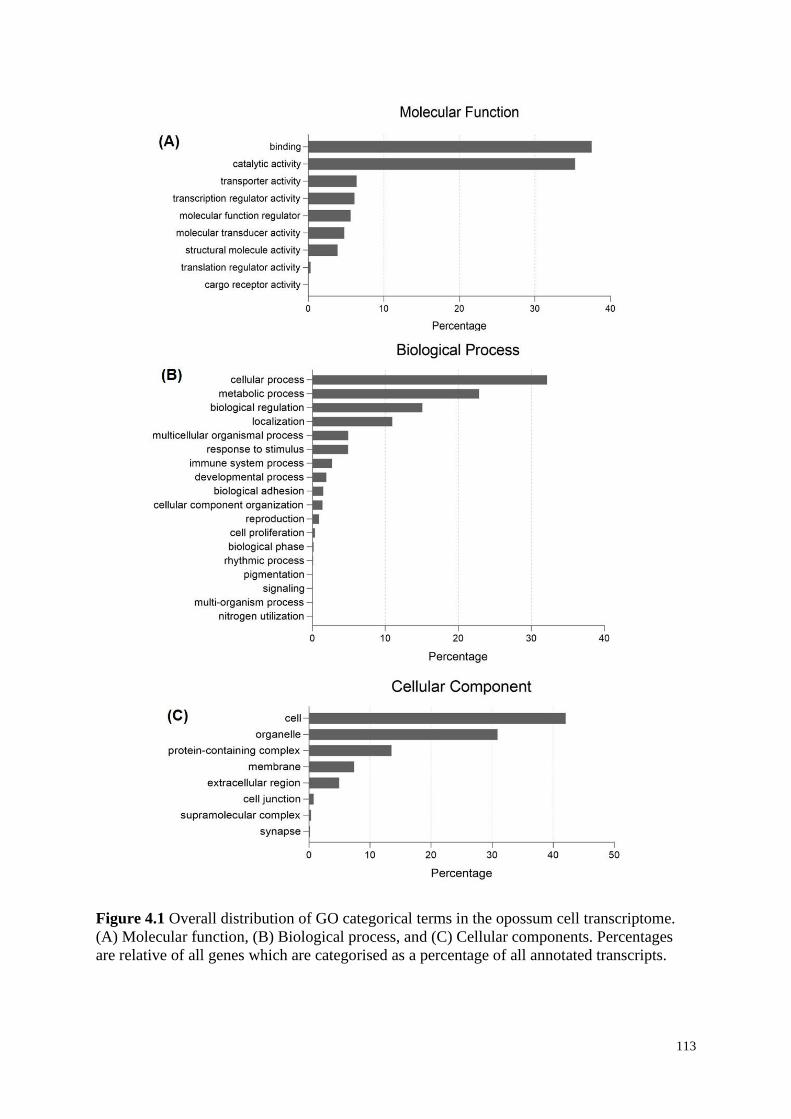
113
Figure 4.1 Overall distribution of GO categorical terms in the opossum cell transcriptome.
(A) Molecular function, (B) Biological process, and (C) Cellular components. Percentages
are relative of all genes which are categorised as a percentage of all annotated transcripts.
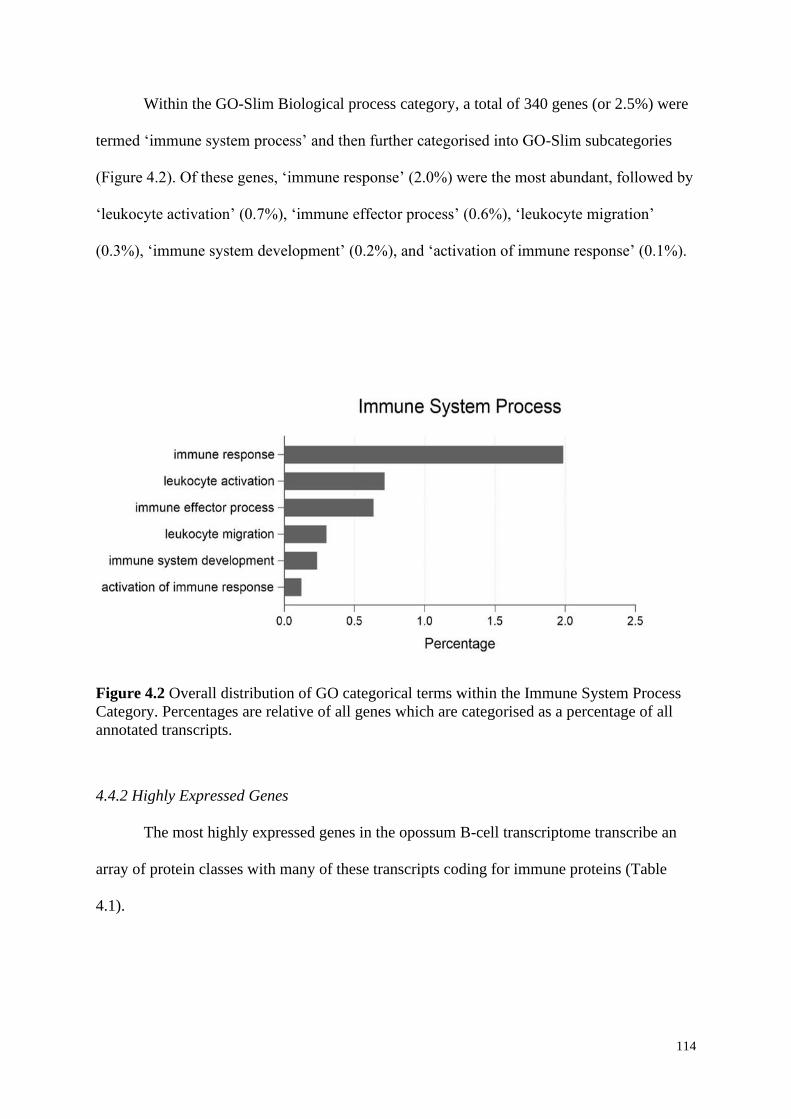
114
Within the GO-Slim Biological process category, a total of 340 genes (or 2.5%) were
termed ‘immune system process’ and then further categorised into GO-Slim subcategories
(Figure 4.2). Of these genes, ‘immune response’ (2.0%) were the most abundant, followed by
‘leukocyte activation’ (0.7%), ‘immune effector process’ (0.6%), ‘leukocyte migration’
(0.3%), ‘immune system development’ (0.2%), and ‘activation of immune response’ (0.1%).
Figure 4.2 Overall distribution of GO categorical terms within the Immune System Process
Category. Percentages are relative of all genes which are categorised as a percentage of all
annotated transcripts.
4.4.2 Highly Expressed Genes
The most highly expressed genes in the opossum B-cell transcriptome transcribe an
array of protein classes with many of these transcripts coding for immune proteins (Table
4.1).
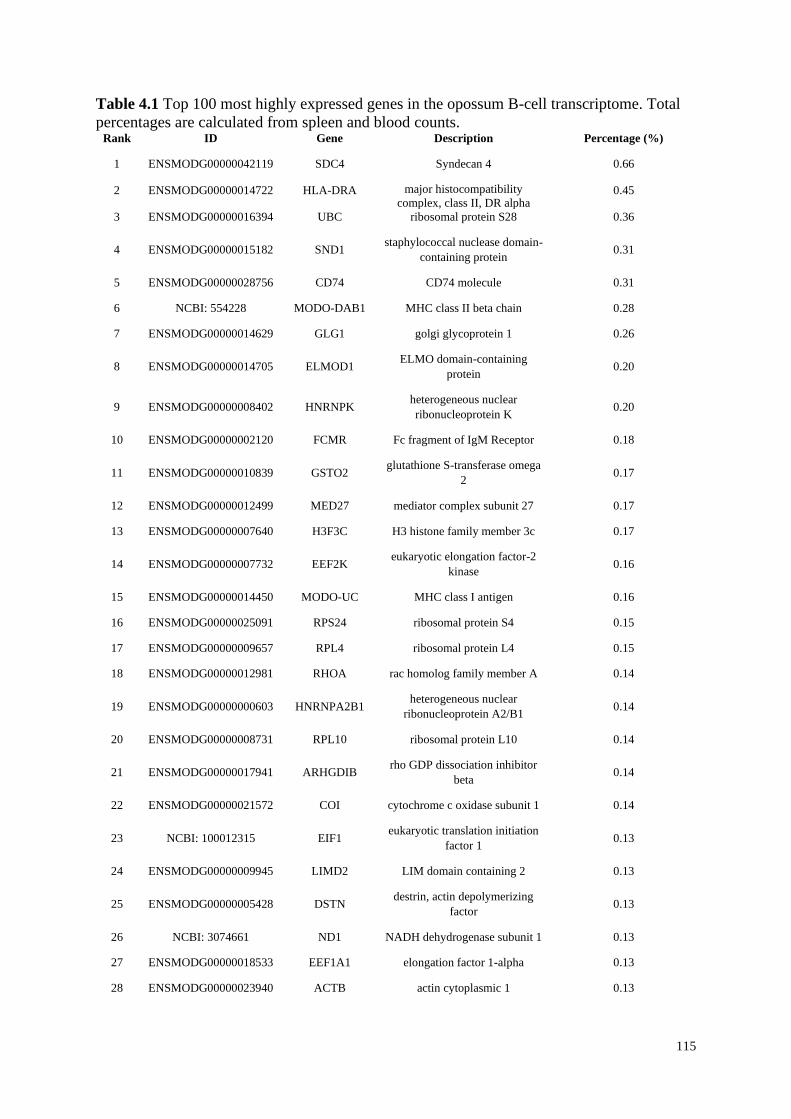
115
Table 4.1 Top 100 most highly expressed genes in the opossum B-cell transcriptome. Total
percentages are calculated from spleen and blood counts. Rank ID Gene Description Percentage (%)
1 ENSMODG00000042119 SDC4 Syndecan 4 0.66
2 ENSMODG00000014722 HLA-DRA major histocompatibility
complex, class II, DR alpha 0.45
3 ENSMODG00000016394 UBC ribosomal protein S28 0.36
4 ENSMODG00000015182 SND1 staphylococcal nuclease domain-
containing protein 0.31
5 ENSMODG00000028756 CD74 CD74 molecule 0.31
6 NCBI: 554228 MODO-DAB1 MHC class II beta chain 0.28
7 ENSMODG00000014629 GLG1 golgi glycoprotein 1 0.26
8 ENSMODG00000014705 ELMOD1 ELMO domain-containing
protein 0.20
9 ENSMODG00000008402 HNRNPK heterogeneous nuclear
ribonucleoprotein K 0.20
10 ENSMODG00000002120 FCMR Fc fragment of IgM Receptor 0.18
11 ENSMODG00000010839 GSTO2 glutathione S-transferase omega
2 0.17
12 ENSMODG00000012499 MED27 mediator complex subunit 27 0.17
13 ENSMODG00000007640 H3F3C H3 histone family member 3c 0.17
14 ENSMODG00000007732 EEF2K eukaryotic elongation factor-2
kinase 0.16
15 ENSMODG00000014450 MODO-UC MHC class I antigen 0.16
16 ENSMODG00000025091 RPS24 ribosomal protein S4 0.15
17 ENSMODG00000009657 RPL4 ribosomal protein L4 0.15
18 ENSMODG00000012981 RHOA rac homolog family member A 0.14
19 ENSMODG00000000603 HNRNPA2B1 heterogeneous nuclear
ribonucleoprotein A2/B1 0.14
20 ENSMODG00000008731 RPL10 ribosomal protein L10 0.14
21 ENSMODG00000017941 ARHGDIB rho GDP dissociation inhibitor
beta 0.14
22 ENSMODG00000021572 COI cytochrome c oxidase subunit 1 0.14
23 NCBI: 100012315 EIF1 eukaryotic translation initiation
factor 1 0.13
24 ENSMODG00000009945 LIMD2 LIM domain containing 2 0.13
25 ENSMODG00000005428 DSTN destrin, actin depolymerizing
factor 0.13
26 NCBI: 3074661 ND1 NADH dehydrogenase subunit 1 0.13
27 ENSMODG00000018533 EEF1A1 elongation factor 1-alpha 0.13
28 ENSMODG00000023940 ACTB actin cytoplasmic 1 0.13
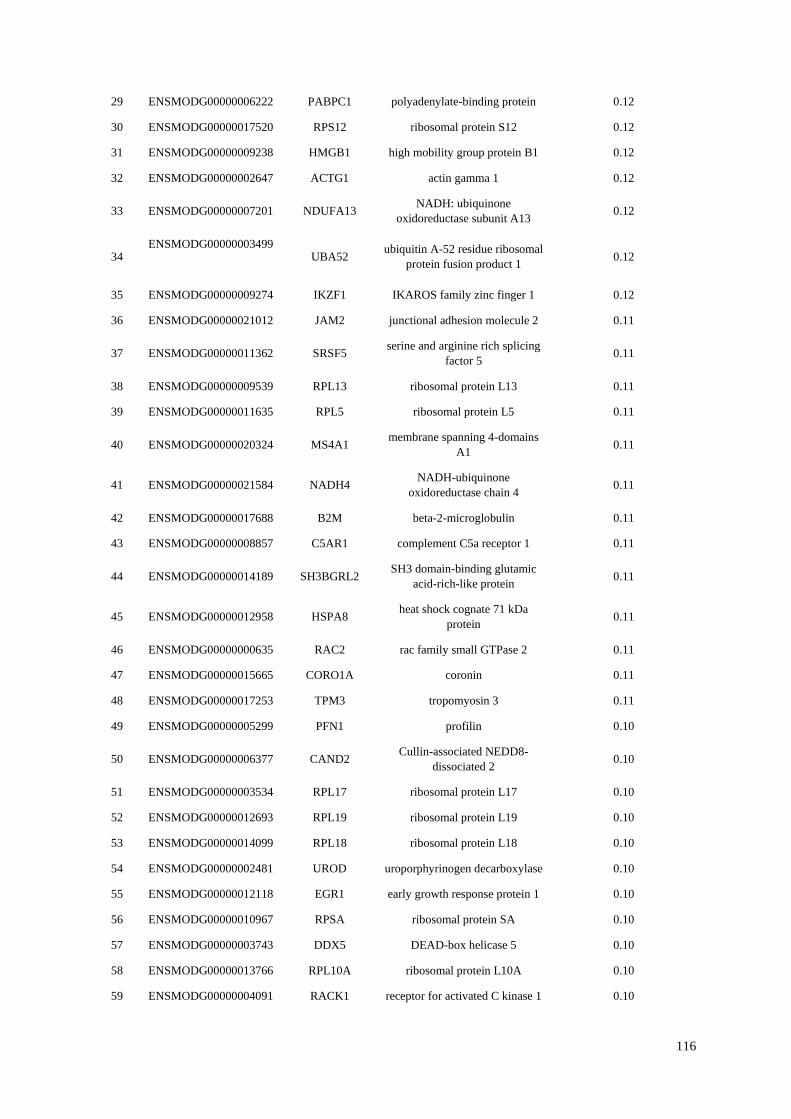
116
29 ENSMODG00000006222 PABPC1 polyadenylate-binding protein 0.12
30 ENSMODG00000017520 RPS12 ribosomal protein S12 0.12
31 ENSMODG00000009238 HMGB1 high mobility group protein B1 0.12
32 ENSMODG00000002647 ACTG1 actin gamma 1 0.12
33 ENSMODG00000007201 NDUFA13 NADH: ubiquinone
oxidoreductase subunit A13 0.12
34 ENSMODG00000003499
UBA52
ubiquitin A-52 residue ribosomal
protein fusion product 1 0.12
35 ENSMODG00000009274 IKZF1 IKAROS family zinc finger 1 0.12
36 ENSMODG00000021012 JAM2 junctional adhesion molecule 2 0.11
37 ENSMODG00000011362 SRSF5 serine and arginine rich splicing
factor 5 0.11
38 ENSMODG00000009539 RPL13 ribosomal protein L13 0.11
39 ENSMODG00000011635 RPL5 ribosomal protein L5 0.11
40 ENSMODG00000020324 MS4A1 membrane spanning 4-domains
A1 0.11
41 ENSMODG00000021584 NADH4 NADH-ubiquinone
oxidoreductase chain 4 0.11
42 ENSMODG00000017688 B2M beta-2-microglobulin 0.11
43 ENSMODG00000008857 C5AR1 complement C5a receptor 1 0.11
44 ENSMODG00000014189 SH3BGRL2 SH3 domain-binding glutamic
acid-rich-like protein 0.11
45 ENSMODG00000012958 HSPA8 heat shock cognate 71 kDa
protein 0.11
46 ENSMODG00000000635 RAC2 rac family small GTPase 2 0.11
47 ENSMODG00000015665 CORO1A coronin 0.11
48 ENSMODG00000017253 TPM3 tropomyosin 3 0.11
49 ENSMODG00000005299 PFN1 profilin 0.10
50 ENSMODG00000006377 CAND2 Cullin-associated NEDD8-
dissociated 2 0.10
51 ENSMODG00000003534 RPL17 ribosomal protein L17 0.10
52 ENSMODG00000012693 RPL19 ribosomal protein L19 0.10
53 ENSMODG00000014099 RPL18 ribosomal protein L18 0.10
54 ENSMODG00000002481 UROD uroporphyrinogen decarboxylase 0.10
55 ENSMODG00000012118 EGR1 early growth response protein 1 0.10
56 ENSMODG00000010967 RPSA ribosomal protein SA 0.10
57 ENSMODG00000003743 DDX5 DEAD-box helicase 5 0.10
58 ENSMODG00000013766 RPL10A ribosomal protein L10A 0.10
59 ENSMODG00000004091 RACK1 receptor for activated C kinase 1 0.10
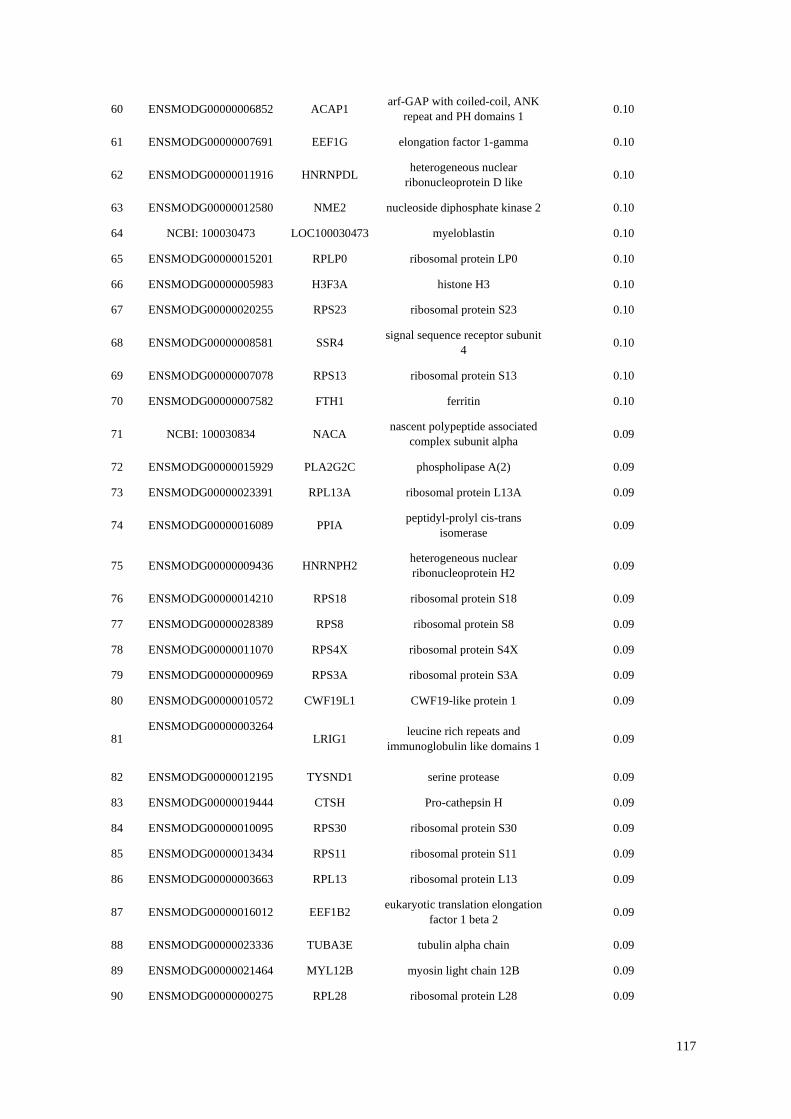
117
60 ENSMODG00000006852 ACAP1 arf-GAP with coiled-coil, ANK
repeat and PH domains 1 0.10
61 ENSMODG00000007691 EEF1G elongation factor 1-gamma 0.10
62 ENSMODG00000011916 HNRNPDL heterogeneous nuclear
ribonucleoprotein D like 0.10
63 ENSMODG00000012580 NME2 nucleoside diphosphate kinase 2 0.10
64 NCBI: 100030473 LOC100030473 myeloblastin 0.10
65 ENSMODG00000015201 RPLP0 ribosomal protein LP0 0.10
66 ENSMODG00000005983 H3F3A histone H3 0.10
67 ENSMODG00000020255 RPS23 ribosomal protein S23 0.10
68 ENSMODG00000008581 SSR4 signal sequence receptor subunit
4 0.10
69 ENSMODG00000007078 RPS13 ribosomal protein S13 0.10
70 ENSMODG00000007582 FTH1 ferritin 0.10
71 NCBI: 100030834 NACA nascent polypeptide associated
complex subunit alpha 0.09
72 ENSMODG00000015929 PLA2G2C phospholipase A(2) 0.09
73 ENSMODG00000023391 RPL13A ribosomal protein L13A 0.09
74 ENSMODG00000016089 PPIA peptidyl-prolyl cis-trans
isomerase 0.09
75 ENSMODG00000009436 HNRNPH2 heterogeneous nuclear
ribonucleoprotein H2 0.09
76 ENSMODG00000014210 RPS18 ribosomal protein S18 0.09
77 ENSMODG00000028389 RPS8 ribosomal protein S8 0.09
78 ENSMODG00000011070 RPS4X ribosomal protein S4X 0.09
79 ENSMODG00000000969 RPS3A ribosomal protein S3A 0.09
80 ENSMODG00000010572 CWF19L1 CWF19-like protein 1 0.09
81 ENSMODG00000003264
LRIG1
leucine rich repeats and
immunoglobulin like domains 1 0.09
82 ENSMODG00000012195 TYSND1 serine protease 0.09
83 ENSMODG00000019444 CTSH Pro-cathepsin H 0.09
84 ENSMODG00000010095 RPS30 ribosomal protein S30 0.09
85 ENSMODG00000013434 RPS11 ribosomal protein S11 0.09
86 ENSMODG00000003663 RPL13 ribosomal protein L13 0.09
87 ENSMODG00000016012 EEF1B2 eukaryotic translation elongation
factor 1 beta 2 0.09
88 ENSMODG00000023336 TUBA3E tubulin alpha chain 0.09
89 ENSMODG00000021464 MYL12B myosin light chain 12B 0.09
90 ENSMODG00000000275 RPL28 ribosomal protein L28 0.09

118
91 ENSMODG00000001819 RPS27A ribosomal protein S27A 0.09
92 ENSMODG00000003890 RPS28 ribosomal protein S28 0.08
93 ENSMODG00000019167 RPL18A ribosomal protein L18 0.08
94 ENSMODG00000028168 COX5A cytochrome c oxidase subunit 5A 0.08
95 ENSMODG00000003974 RPS9 ribosomal protein S9 0.08
96 ENSMODG00000021591 CYTB cytochrome b 0.08
97 ENSMODG00000025578 RPL36A ribosomal protein L36A 0.08
98 ENSMODG00000028326 CCDC184 coiled-coil domain containing
184 0.08
99 ENSMODG00000004571 RPL12 ribosomal protein L12 0.08
100 ENSMODG00000006682 RPL38 ribosomal protein L38 0.08
Syndecan 4 (SDC4) had the highest expression in the entire B-cell transcriptome,
accounting for 0.66% of the total gene expression in the opossum B-cell transcriptome (Table
4.1). Alignment comparison between the opossum SDC4 protein sequence and other
mammalian species showed a >90% identity between opossum and other marsupial SDC4
protein sequences, and >75% identity between the opossum and investigated eutherian
protein sequences (Figure 4.3).

119
Figure 4.3 (A) Multiple sequence alignment of SDC4 gene products and (B) a Phylogenetic
tree based on the SDC4 sequences, constructed using the NJ method. Protein sequences were
aligned using the ClustalW algorithm. Dots indicate identical matches to the opossum
sequences. Domains of the SDC4 protein are indicated by a line underneath the sequence.
Species designations are abbreviated to the first two letters of their scientific names. M.
domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus
(Oran), H. sapiens (Hosa), and M. musculus (Mumu). Genbank accession numbers are: Modo
SDC4, XP_007475896.1; Saha SDC4, XP_012395261.1; Phci SDC4, XP_020839750.1;
Vour SDC4, XP_027707442.1; Oran SDC4, XP_028927254.1; Hosa SDC4, NP_002990.2;
Mumu SDC4, NP_035651.1.
The spleen and blood single cells showed significant differences between their
expression of highly expressed genes identified in the opossum B-cell transcriptome
(ANOSIM Global R = 0.035, P = 0.046) (Figure 4.4). Average dissimilarity revealed that this
significant difference between the spleen and blood was largely due to the proportion of the
HNRNPK gene (heterogeneous nuclear ribonucleoprotein K) in the spleen (55%) compared
to the blood (36%). There was no significant difference between the different Ig expressing
cells (ANOSIM Global R = 0.063, P = 0.066), as the abundance of IgE cells were too small
to compare (Figure 4.4 and Figure 4.5). ANOSIM pairwise tests showed significant
differences between the expression of specific genes produced by IgM (R = 0.168, P = 0.002)
and IgG (R = 0.138, P = 0.001) cells in comparison to IgA cells (Figure 4.5). SIMPER
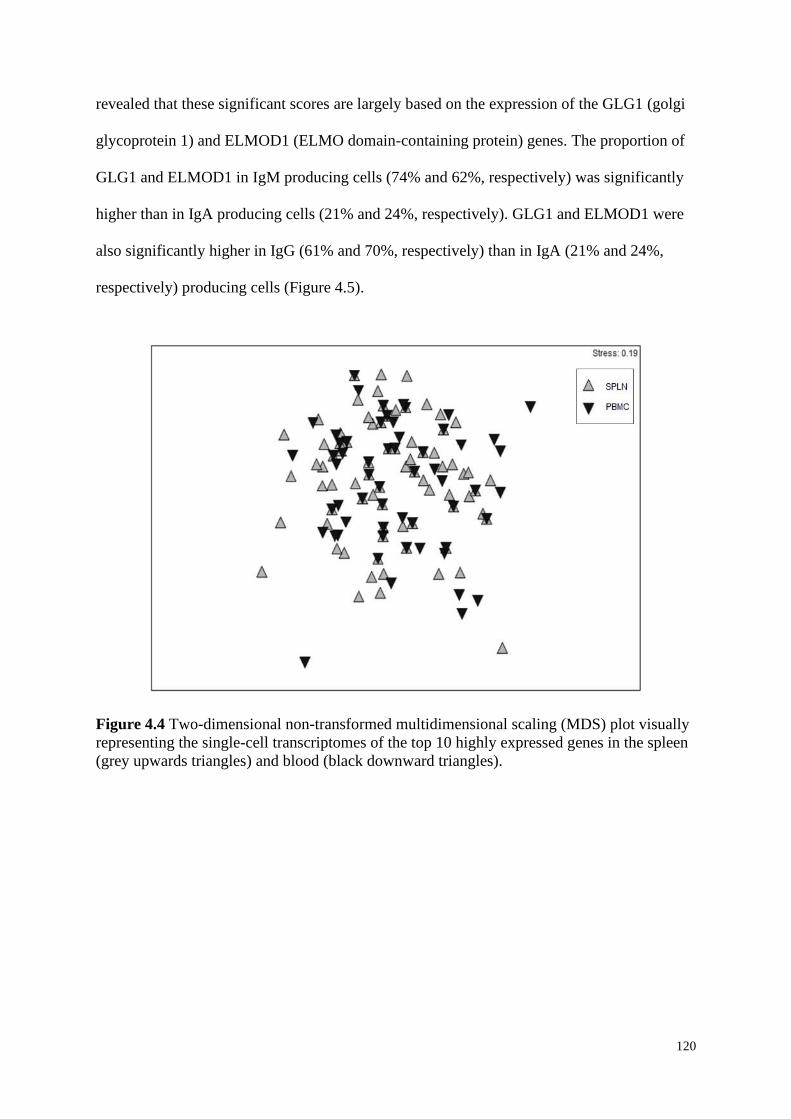
120
revealed that these significant scores are largely based on the expression of the GLG1 (golgi
glycoprotein 1) and ELMOD1 (ELMO domain-containing protein) genes. The proportion of
GLG1 and ELMOD1 in IgM producing cells (74% and 62%, respectively) was significantly
higher than in IgA producing cells (21% and 24%, respectively). GLG1 and ELMOD1 were
also significantly higher in IgG (61% and 70%, respectively) than in IgA (21% and 24%,
respectively) producing cells (Figure 4.5).
Figure 4.4 Two-dimensional non-transformed multidimensional scaling (MDS) plot visually
representing the single-cell transcriptomes of the top 10 highly expressed genes in the spleen
(grey upwards triangles) and blood (black downward triangles).
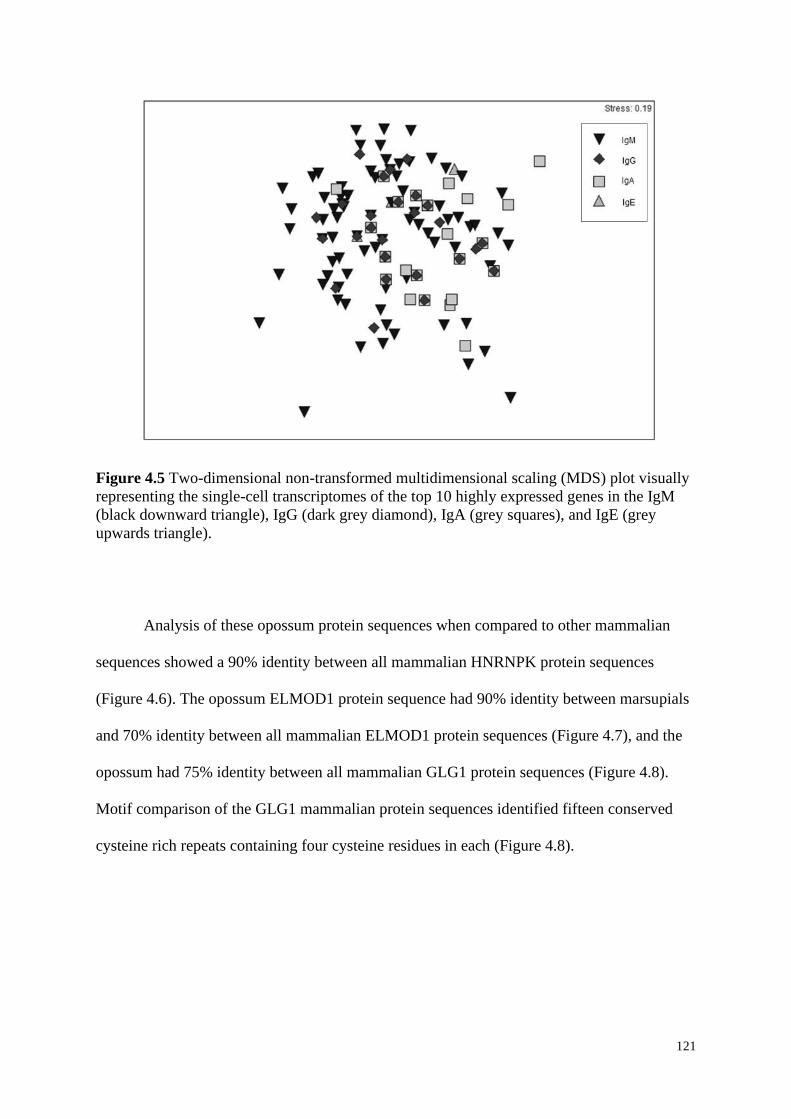
121
Figure 4.5 Two-dimensional non-transformed multidimensional scaling (MDS) plot visually
representing the single-cell transcriptomes of the top 10 highly expressed genes in the IgM
(black downward triangle), IgG (dark grey diamond), IgA (grey squares), and IgE (grey
upwards triangle).
Analysis of these opossum protein sequences when compared to other mammalian
sequences showed a 90% identity between all mammalian HNRNPK protein sequences
(Figure 4.6). The opossum ELMOD1 protein sequence had 90% identity between marsupials
and 70% identity between all mammalian ELMOD1 protein sequences (Figure 4.7), and the
opossum had 75% identity between all mammalian GLG1 protein sequences (Figure 4.8).
Motif comparison of the GLG1 mammalian protein sequences identified fifteen conserved
cysteine rich repeats containing four cysteine residues in each (Figure 4.8).
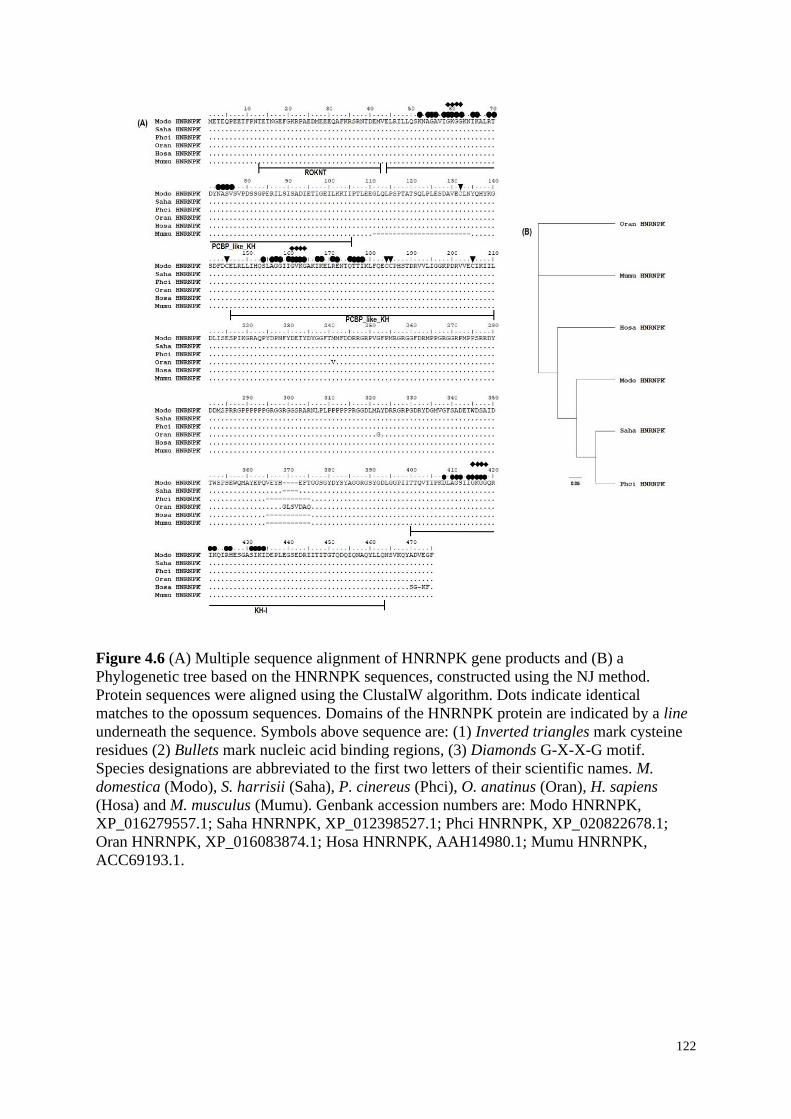
122
Figure 4.6 (A) Multiple sequence alignment of HNRNPK gene products and (B) a
Phylogenetic tree based on the HNRNPK sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the HNRNPK protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues (2) Bullets mark nucleic acid binding regions, (3) Diamonds G-X-X-G motif.
Species designations are abbreviated to the first two letters of their scientific names. M.
domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), O. anatinus (Oran), H. sapiens
(Hosa) and M. musculus (Mumu). Genbank accession numbers are: Modo HNRNPK,
XP_016279557.1; Saha HNRNPK, XP_012398527.1; Phci HNRNPK, XP_020822678.1;
Oran HNRNPK, XP_016083874.1; Hosa HNRNPK, AAH14980.1; Mumu HNRNPK,
ACC69193.1.

123
Figure 4.7 (A) Multiple sequence alignment of ELMOD1 gene products and (B) a
Phylogenetic tree based on the ELMOD1 sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the ELMOD1 protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues. Species designations are abbreviated to the first two letters of their scientific names.
M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus
(Oran), and H. sapiens (Hosa). Genbank accession numbers are: Modo ELMOD1,
XP_007494949.1; Saha ELMOD1, XP_012400499.1; Phci ELMOD1, XP_020851842.1;
Vour ELMOD1, XP_027705369.1; Oran ELMOD1, XP_028904013.1; Hosa ELMOD1,
NP_061182.3.

124
Figure 4.8 (A) Multiple sequence alignment of GLG1 gene products and (B) a Phylogenetic
tree based on the GLG1 sequences, constructed using the NJ method. Protein sequences were
aligned using the ClustalW algorithm. Dots indicate identical matches to the opossum
sequences. Cysteine rich repeats are indicated by lines underneath the sequence. Symbols
above sequence are: (1) Inverted triangles mark cysteine residues. Species designations are
abbreviated to the first two letters of their scientific names. M. domestica (Modo), S. harrisii
(Saha), P. cinereus (Phci), V. ursinus (Vour), O. anatinus (Oran), H. sapiens (Hosa), and M.
musculus (Mumu). Genbank accession numbers are: Modo GLG1, XP_007477569.1; Saha
GLG1, XP_012395893.2; Phci GLG1, XP_020819407.1; Vour GLG1, XP_027699023.1;
Oran GLG1, XP_028931633.1; Hosa GLG1, AAH60822.1; Mumu GLG1, EDL11491.1.
Immune genes that were highly expressed genes included the MHC class II DRα
chain (HLA-DRA) and MHC class II β chain (MODO-DAB1) genes (both ranked in the top
10 most highly expressed genes), and along with SDC4, make up >1.3% of the total
expressed genes in the opossum B-cell transcriptome (Table 4.1). Comparison of these
protein sequences with other mammals showed HLA-DRA to have a >90% identity between
marsupial and eutherian mammals while MODO-DAB1 had a >70% identity with other

125
marsupial MHC DAB protein sequences (Figure 4.9 and Figure 4.10). Motif analysis
identified an Ig C1-set domain and an α domain on both HLA-DRA and MODO-DAB1
protein sequences. Additionally, HLA-DRA protein sequences included seven heterodimer
interface residues and ten MHC binding domain interface residues (Figure 4.9).
Figure 4.9 (A) Multiple sequence alignment of HLA-DRA gene products and (B) a
Phylogenetic tree based on the HLA-DRA sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the HLA-DRA protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues, (2) Bullets mark heterodimer interface residues, (3) Diamonds mark MHC binding
domain interface residues. Species designations are abbreviated to the first two letters of their
scientific names. M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), V. ursinus
(Vour), and H. sapiens (Hosa). Genbank accession numbers are: Modo HLA-DRA,
XP_007483702.1; Saha HLA-DRA, XP_003770700.1; Phci HLA-DRA, XP_020829855.1;
Vour HLA-DRA, XP_027732036.1; Hosa HLA-DRA, ARB08447.1.

126
Figure 4.10 (A) Multiple sequence alignment of marsupial MHC DAB1 gene products and
(B) a Phylogenetic tree based on the MHC DAB sequences, constructed using the NJ method.
Protein sequences were aligned using the ClustalW algorithm. Dots indicate identical
matches to the opossum sequences. Domains of the MHC DAB protein are indicated by a line
underneath the sequence. Symbols above sequence are: (1) Inverted triangles mark cysteine
residues. Species designations are abbreviated to the first two letters of their scientific names.
M. domestica (Modo), S. harrisii (Saha), P. cinereus (Phci), N. eugenii (Noeu), and T.
vulpecula (Trvu). Genbank accession numbers are: Modo DAB1, NP_001028163.1; Saha
DAB, ABV58848.1; Phci DAB, ALX81653.1; Noeu DAB, AAX54675.1; Trvu DAB,
AAK84172.1.
4.5 Discussion
The release of the opossum genome (Mikkelsen et al., 2007) and several companion
articles have both reported structural details of the genome and in depth comparisons between
eutherian and marsupial immunological components. Along with this report, the opossum has
become the first marsupial to begin analysing the gene expression of B-cells at the single-cell
level. Marsupials are born immunologically incompetent, unable to mount an endogenous
antibody response until after the first week postpartum (Wang et al., 2009). Initial studies of
primary humoral immune response in the opossum utilised sheep red blood cells and
haemolytic titration to conclude that humoral immune responses are similar to that of

127
eutherians (Croix et al., 1989). Our investigation identified the most highly expressed genes
in the opossum’s single-cell transcriptome and analysed these genes to show any preferential
production from IgM, IgG, IgA and IgE expressing cells, however since the number of cells
expressing IgE were far too small, a definitive comparison with IgE could not be made.
SDC4 had the highest accumulative expression in the single-cell transcriptome,
making up 0.66% of the opossum B-cells. Analyses of the SDC4 gene found no preferential
variation between the IgH expressing cells. SDC4 is a transmembrane heparin sulphate
proteoglycan located on the cell surface (Lee et al., 1998), encoding proteins that play major
roles in cell adhesion, signalling and defence (Tkachenko et al., 2005, Elfenbein and Simons,
2013). Although the specific immunological role of SDC4 remains unclear (Endo et al.,
2015), it has previously been expressed in the uterine transcriptome of a pregnant opossum
(Hansen et al., 2016), as well as in the splenic B-cells of mice (Yamashita et al., 1999).
Furthermore a 4.1 m domain (putative band 4.1 homologues’ binding motif) was identified in
both eutherian and marsupial SDC4 protein sequences and has been acknowledged for its
importance in transmembrane signalling and cell-matrix adhesion (Lee et al., 1998, Wang et
al., 2013). Other genes which had a high accumulative expression within the opossum B-cell
transcriptome including HNRNPK, ELMOD1, and GLG1. HNRNPK had a significant
abundance of transcripts expressed by spleen cells, though no difference was seen in between
the single IgH expressing cells. ELMOD1 and GLG1 on the other hand, had significantly
higher expression of these transcripts in both IgM and IgG in comparison to IgA. Due to low
expression of IgE in the transcriptome, it is still unclear whether these genes are definitively
produced by IgE expressing cells. HNRNPK was first identified to facilitate the processing
and/or transport of pre-mRNA between the nucleus and cytoplasm of cells (Matunis et al.,
1992). Though analysis of human HNRNPK distribution has shown greater function in signal
transduction and gene expression involved in the B-cell receptor signalling pathway

128
(Bomsztyk et al., 1997, Jeon et al., 2005). The KH-1 motif identified on the investigated
mammalian HNRNPK sequences suggests this gene is highly conserved between mammals
and present the similar function in the B-cell receptor (Adinolfi et al., 1999). The ELMOD1
gene encodes for a protein with GTPase-activating properties (Ivanova et al., 2014),
regulating actin-mediated processes such as chemotaxis and engulfment in many cell types
(Brzostowski et al., 2009). Previous investigations have expressed the ELMOD1 gene in the
uterine transcriptome of the opossum (Hansen et al., 2016), showing a decrease of expression
in non-pregnant opossum individuals. ELMOD1 has been identified in number of eukaryotes,
suggesting it is highly conserved and has a distinct set of cellular functions which are
homologous in eukaryotes (East et al., 2012). Lastly, GLG1 gene has not been fully
characterised in mammals (Ahn et al., 2005). Previous investigations of GLG1 have shown
significant localisations with basic fibroblast growth factors binding proteins, along with
number of cysteine rich repeats identified on the GLG1 mammalian alignments suggests
GLG1 may assist with processing growth factors in numerous cells (Zuber et al., 1997, Köhl
et al., 2000).
Specific immune genes that were highly expressed in the in transcriptome included
HLA-DRA (ranked 2nd) and MODO-DAB1 (ranked 6th). Both HLA-DRA and MODO-DAB1
belong to the MHC Class II family, which have extensive involvement in B-cell
immunological responses (Cheng et al., 2009). Neither of these genes showed any
preferential variation amongst the different IgH isotype expressing cells. Previously, both
HLA-DRA and MODO-DAB1 genes have been identified and characterised in marsupials.
The MHC DAB1 gene has been characterised in several marsupial species; two DAB alleles
have been isolated in the red-necked wallaby (Macropus rufogriseus) (one was identified as a
pseudogene) (Schneider et al., 1991), six DAB1 alleles have been isolated from the
Tasmanian devil (Siddle et al., 2007), five DAB1 alleles in the common brushtail possum

129
(Lam et al., 2001), and only a single DAB1 transcript isolated in the spleen from a male
opossum (Stone et al., 1999). The marsupial MHC DAB gene family, along with DBB (Belov
et al., 2004) and DCB (Belov et al., 2006), are not present in any eutherian species (Belov et
al., 2004). MHC DAB and DBB gene families are considered to have the highest expression
in marsupials (Jobbins et al., 2012), with DAB the most polymorphic (Holland et al., 2008),
yet this investigation could only identify the expression for DAB1 in the B-cell
transcriptome. HLA-DRA has been investigated in both the red-necked wallaby (Slade and
Mayer, 1995), and koala (Abts et al., 2015). Slade and Mayer (1995) showed that the
marsupial DRA sequence forms a distinct clade with DRA sequences observed in eutherians
and northern blot analysis revealed very high expression of DRA in the spleen of the red-
necked wallaby. Unlike DAB1, marsupial DRA is present in eutherians, suggesting some
MHC class II genes of mammals have evolved from different ancestral genes (Schneider et
al., 1991).
The investigation of these highly expressed genes expressed in the opossum B-cell
transcriptome has shown marsupial B-cells are comparable to eutherians. B-cells are vital for
the humoral immune system response which is fundamental in the investigatory studies of
mammalian immunity. This report provides a key resource for future transcriptomic studies
of the opossum B-cells and assist with comparisons between the immunological divergence
of marsupials and eutherians, and will contribute to the understanding of defence mechanisms
used by marsupials, particularly the adaptive immune response.

130
4.6 Acknowledgements
The authors would like to thank Ricky Spencer (WSU) for his help with the MDS
plots, and Galina Petukhova making available M. domestica and her assistance with animal
husbandry and tissue collection. This research was funded in part by a National Science
Foundation award IOS-13531232. Research reported in this publication was also supported in
part by the National Institute of General Medical Sciences of the National Institutes of Health
under award P30 GM110907. The content is solely the responsibility of the authors and does
not necessarily represent the official views of the National Institutes of Health.
4.7 Conflicts of Interest
The authors declare no conflicts of interest.

131
4.8 Chapter 4 References
ABTS, K. C., IVY, J. A. & DEWOODY, J. A. 2015. Immunomics of the koala
(Phascolarctos cinereus). Immunogenetics, 67, 305-21.
ADINOLFI, S., BAGNI, C., CASTIGLIONE MORELLI, M. A., FRATERNALI, F.,
MUSCO, G. & PASTORE, A. 1999. Novel RNA-binding motif: the KH module.
Peptide Science, 51, 153-164.
AFGAN, E., BAKER, D., BATUT, B., VAN DEN BEEK, M., BOUVIER, D., ČECH, M.,
CHILTON, J., CLEMENTS, D., CORAOR, N., GRÜNING, B. A., GUERLER, A.,
HILLMAN-JACKSON, J., HILTEMANN, S., JALILI, V., RASCHE, H.,
SORANZO, N., GOECKS, J., TAYLOR, J., NEKRUTENKO, A. &
BLANKENBERG, D. 2018. The galaxy platform for accessible, reproducible and
collaborative biomedical analyses: 2018 update. Nucleic Acids Research, 46, W537-
W544.
AHN, J., FEBBRAIO, M. & SILVERSTEIN, R. L. 2005. A novel isoform of human golgi
complex-localized glycoprotein-1 (also known as E-selectin ligand-1, MG-160 and
cysteine-rich fibroblast growth factor receptor) targets differential subcellular
localization. Journal of Cell Science, 118, 1725.
BELOV, K., DEAKIN, J. E., PAPENFUSS, A. T., BAKER, M. L., MELMAN, S. D.,
SIDDLE, H. V., GOUIN, N., GOODE, D. L., SARGEANT, T. J., ROBINSON, M.
D., WAKEFIELD, M. J., MAHONY, S., CROSS, J. G. R., BENOS, P. V.,
SAMOLLOW, P. B., SPEED, T. P., GRAVES, J. A. M. & MILLER, R. D. 2006.
Reconstructing an ancestral mammalian immune supercomplex from a marsupial
major histocompatibility complex. PLOS Biology, 4, e46.
BELOV, K., LAM, M. & COLGAN, D. 2004. Marsupial MHC class II beta genes are not
orthologous to the eutherian beta gene families. The Journal of Heredity, 95, 338-45.
BELOV, K., NGUYEN, M.-A. T., ZENGER, K. R. & COOPER, D. W. 2002. Ontogeny of
immunoglobulin expression in the brushtail possum (Trichosurus vulpecula).
Developmental & Comparative Immunology, 26, 599-602.
BOMSZTYK, K., VAN SEUNINGEN, I., SUZUKI, H., DENISENKO, O. & OSTROWSKI,
J. 1997. Diverse molecular interactions of the hnRNP K protein. FEBS Letters, 403,
113-115.
BRZOSTOWSKI, J. A., FEY, P., YAN, J., ISIK, N. & JIN, T. 2009. The elmo family forms
an ancient group of actin-regulating proteins. Communicative & Integrative Biology,
2, 337-340.
CHENG, Y., SIDDLE, H. V., BECK, S., ELDRIDGE, M. D. B. & BELOV, K. 2009. High
levels of genetic variation at MHC class II DBB loci in the tammar wallaby
(Macropus eugenii). Immunogenetics, 61, 111-118.
CLARKE, K. R., GORLEY, R., SOMERFIELD, P. & WARWICK, R. 2014. Change in
marine communities: an approach to statistical analysis and interpretation, Primer-E:
Plymouth, UK.
CONLEY, M. E. & COOPER, M. D. 1998. Genetic basis of abnormal B cell development.
Current Opinion in Immunology, 10, 399-406.
CROIX, D., SAMPLES, N., VANDEBERG, J. & STONE, W. 1989. Immune response of a
marsupial (Monodelphis domestica) to sheep red blood cells. Developmental &
Comparative Immunology, 13, 73-78.
DUNCAN, L., WEBSTER, K., GUPTA, V., NAIR, S. & DEANE, E. 2010. Molecular
characterisation of the CD79a and CD79b subunits of the B cell receptor complex in
the gray short-tailed opossum (Monodelphis domestica) and tammar wallaby

132
(Macropus eugenii): delayed B cell immunocompetence in marsupial neonates.
Veterinary Immunology and Immunopathology, 136, 235-47.
EAST, M. P., BOWZARD, J. B., DACKS, J. B. & KAHN, R. A. 2012. ELMO domains,
evolutionary and functional characterization of a novel GTPase-activating protein
(GAP) domain for Arf protein family GTPases. Journal of Biological Chemistry, 287,
39538-39553.
EDWARDS, M., HINDS, L., DEANE, E. & DEAKIN, J. 2012. A review of complementary
mechanisms which protect the developing marsupial pouch young. Developmental &
Comparative Immunology, 37, 213-220.
EDWARDS, M. J. & DEAKIN, J. E. 2013. The marsupial pouch: implications for
reproductive success and mammalian evolution. Australian Journal of Zoology, 61,
41-47.
ELFENBEIN, A. & SIMONS, M. 2013. Syndecan-4 signaling at a glance. Journal of Cell
Science, 126, 3799-3804.
ENDO, T., ITO, K., MORIMOTO, J., KANAYAMA, M., OTA, D., IKESUE, M., KON, S.,
TAKAHASHI, D., ONODERA, T., IWASAKI, N. & UEDE, T. 2015. Syndecan 4
regulation of the development of autoimmune arthritis in mice by modulating B cell
migration and germinal center formation. Arthritis & Rheumatology, 67, 2512-2522.
HALL, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and
analysis program for windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95-98.
HANSEN, V. L., SCHILKEY, F. D. & MILLER, R. D. 2016. Transcriptomic changes
associated with pregnancy in a marsupial, the gray short-tailed opossum Monodelphis
domestica. PLOS ONE, 11, e0161608.
HARDY, R. R. & HAYAKAWA, K. 2001. B cell development pathway. Annual Review of
Immunology, 19, 595-621.
HENIKOFF, S. & HENIKOFF, J. G. 1992. Amino acid substitution matrices from protein
blocks. Proceedings of the National Academy of Sciences of the United States of
America, 89, 10915.
HOLLAND, O. J., COWAN, P. E., GLEESON, D. M. & CHAMLEY, L. W. 2008. Novel
alleles in classical major histocompatibility complex class II loci of the brushtail
possum (Trichosurus vulpecula). Immunogenetics, 60, 449-460.
HOLMES, M. L., PRIDANS, C. & NUTT, S. L. 2008. The regulation of the B-cell gene
expression programme by Pax5. Immunology and Cell Biology, 86, 47-53.
IVANOVA, A. A., EAST, M. P., SLEE, L. Y. & KAHN, R. A. 2014. Characterization of
recombinant ELMOD (cell engulfment and motility domain) proteins as GTPase-
activating proteins (GAPs) for ARF family GTPases. Journal of Biological
Chemistry, 289, 11111-11121.
JEON, H.-K., AHN, J.-H., CHOE, J., PARK, J. H. & LEE, T. H. 2005. Anti-IgM induces up-
regulation and tyrosine-phosphorylation of heterogeneous nuclear ribonucleoprotein
K proteins (hnRNP K) in a Ramos B cell line. Immunology Letters, 98, 303-310.
JOBBINS, S. E., SANDERSON, C. E., GRIFFITH, J. E., KROCKENBERGER, M. B.,
BELOV, K. & HIGGINS, D. P. 2012. Diversity of MHC class II DAB1 in the koala
(Phascolarctos cinereus). Australian Journal of Zoology, 60, 1-9.
JUKES, T. H. & CANTOR, C. R. 1969. CHAPTER 24 - Evolution of Protein Molecules. In:
MUNRO, H. N. (ed.) Mammalian Protein Metabolism. Academic Press.
KEARSE, M., MOIR, R., WILSON, A., STONES-HAVAS, S., CHEUNG, M.,
STURROCK, S., BUXTON, S., COOPER, A., MARKOWITZ, S., DURAN, C.,
THIERER, T., ASHTON, B., MEINTJES, P. & DRUMMOND, A. 2012. Geneious
basic: An integrated and extendable desktop software platform for the organization
and analysis of sequence data. Bioinformatics, 28, 1647-1649.

133
KÖHL, R., ANTOINE, M., OLWIN, B. B., DICKSON, C. & KIEFER, P. 2000. Cysteine-
rich fibroblast growth factor receptor alters secretion and intracellular routing of
fibroblast growth factor 3. Journal of Biological Chemistry, 275, 15741-15748.
LAM, M. K. P., BELOV, K., HARRISON, G. A. & COOPER, D. W. 2001. Cloning of the
MHC class II DRB cDNA from the brushtail possum (Trichosurus vulpecula).
Immunology Letters, 76, 31-36.
LEE, D., OH, E., WOODS, A., COUCHMAN, J. R. & LEE, W. 1998. Solution structure of
syndecan-4 cytoplasmic domain and its interation with phosphatidylinositol 4,5-
bisphosphate. The Journal Of Biological Chemistry, 273, 13022-13029.
MATE, K. E., ROBINSON, E. S., PEDERSEN, R. A. & VANDEBERG, J. L. 1994.
Timetable of in vivo embryonic development in the grey short‐tailed opossum
(Monodelphis domestica). Molecular Reproduction and Development, 39, 365-374.
MATUNIS, M. J., MICHAEL, W. M. & DREYFUSS, G. 1992. Characterization and primary
structure of the poly(C)-binding heterogeneous nuclear ribonucleoprotein complex K
protein. Molecular and Cellular Biology, 12, 164.
MI, H., MURUGANUJAN, A., HUANG, X., EBERT, D., MILLS, C., GUO, X. &
THOMAS, P. D. 2019. Protocol Update for large-scale genome and gene function
analysis with the PANTHER classification system (v.14.0). Nature Protocols, 14,
703-721.
MIKKELSEN, T. S., WAKEFIELD, M., AKEN, B., AMEMIYA, C., CHANG, J., DUKE
BECKER, S., GARBER, M., GENTLES, A. J., GOODSTADT, L., HEGER, A.,
JURKA, J., KAMAL, M., MAUCELI, E., SEARLE, S., SHARPE, T., BAKER, M.
L., BATZER, M. A., BENOS, P., BELOV, K. & LINDBLAD-TOH, K. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in non-coding
sequences. Nature 447, 167-77.
MILLER, R. D. & BELOV, K. 2000. Immunoglobulin genetics of marsupials.
Developmental & Comparative Immunology, 24, 485-490.
MORRIS, K. M., WEAVER, H. J., O'MEALLY, D., DESCLOZEAUX, M., GILLETT, A. &
POLKINGHORNE, A. 2018. Transcriptome sequencing of the long-nosed bandicoot
(Perameles nasuta) reveals conservation and innovation of immune genes in the
marsupial order peramelemorphia. Immunogenetics, 70, 327-336.
OLD, J. M. & DEANE, E. 2003. The detection of mature T- and B-cells during development
of the lymphoid tissues of the tammar wallaby (Macropus eugenii). Journal of
Anatomy, 203, 123-131.
OLD, J. M. & DEANE, E. M. 2000. Development of the immune system and immunological
protection in marsupial pouch young. Developmental & Comparative Immunology,
24, 445-454.
PAGNI, M., IOANNIDIS, V., CERUTTI, L., ZAHN-ZABAL, M., JONGENEEL, C. V.,
HAU, J., MARTIN, O., KUZNETSOV, D. & FALQUET, L. 2007. MyHits:
improvements to an interactive resource for analyzing protein sequences. Nucleic
acids research, 35, W433-W437.
PAINTER, M. W., DAVIS, S., HARDY, R. R., MATHIS, D., BENOIST, C. &
IMMUNOLOGICAL GENOME PROJECT, C. 2011. Transcriptomes of the B and T
lineages compared by multiplatform microarray profiling. Journal of Immunology,
186, 3047-57.
SAMOLLOW, P. B. 2006. Status and applications of genomic resources for the gray, short-
tailed opossum, Monodelphis domestica, an American marsupial model for
comparative biology. Australian Journal of Zoology, 54, 173-196.
SAMOLLOW, P. B. 2008. The opossum genome: insights and opportunities from an
alternative mammal. Genome Research, 18, 1199-1215.

134
SCHNEIDER, S., VINCEK, V., TICHY, H., FIGUEROA, F. & KLEIN, J. 1991. MHC class
II genes of a marsupial, the red-necked wallaby (Macropus rufogriseus):
identification of new gene families. Molecular Biology and Evolution, 8, 753-766.
SIDDLE, H. V., SANDERSON, C. & BELOV, K. 2007. Characterization of major
histocompatibility complex class I and class II genes from the Tasmanian devil
(Sarcophilus harrisii). Immunogenetics, 59, 753-760.
SLADE, R. W. & MAYER, W. E. 1995. The expressed class II alpha-chain genes of the
marsupial major histocompatibility complex belong to eutherian mammal gene
families. Molecular Biology and Evolution, 12, 441-450.
STONE, W., BRUUN, D., MANIS, G., HOLSTE, S., HOFFMAN, E., SPONG, K. &
WALUNAS, T. 1996. The immunobiology of the marsupial, Monodelphis domestica.
Modulators of Immune Responses, The Evolutionary Trail, Breckenridge Series, 2.
STONE, W. H., BRUUN, D. A., FUQUA, C., GLASS, L. C., REEVES, A., HOLSTE, S. &
FIGUEROA, F. 1999. Identification and sequence analysis of an MHC class II B gene
in a marsupial (Monodelphis domestica). Immunogenetics, 49, 461-463.
THOMPSON, J. D., HIGGINS, D. G. & GIBSON, T. J. 1994. CLUSTAL W: improving the
sensitivity of progressive multiple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22,
4673-4680.
TKACHENKO, E., RHODES, J. M. & SIMONS, M. 2005. Syndecans. Circulation
Research, 96, 488-500.
VANDEBERG, J. L. & ROBINSON, E. S. 1997. The laboratory opossum (Monodelphis
domestica) in laboratory research. The Institute for Laboratory Animal Research
Journal, 38, 4-12.
WANG, W., TAO, C., ZHOU, P., ZHOU, X., ZHANG, Q. & LIU, B. 2013. Molecular
characterization, expression profiles of the porcine SDC2 and HSPG2 genes and their
association with hematologic parameters. Molecular Biology Reports, 40, 2549-2556.
WANG, X., OLP, J. J. & MILLER, R. D. 2009. On the genomics of immunoglobulins in the
gray, short-tailed opossum Monodelphis domestica. Immunogenetics, 61, 581-96.
WANG, X., SHARP, A. R. & MILLER, R. D. 2012. Early postnatal B cell ontogeny and
antibody repertoire maturation in the opossum, Monodelphis domestica. PLoS One, 7,
e45931.
WONG, E. S., PAPENFUSS, A. T. & BELOV, K. 2011. Immunome database for marsupials
and monotremes. BMC Immunology, 12, 48.
YAMASHITA, Y., ORITANI, K., MIYOSHI, E. K., WALL, R., BERNFIELD, M. &
KINCADE, P. W. 1999. Syndecan-4 Is expressed by B lineage lymphocytes and can
transmit a signal for formation of dendritic processes. The Journal of Immunology,
162, 5940.
ZUBER, M. E., ZHOU, Z., BURRUS, L. W. & OLWIN, B. B. 1997. Cysteine‐rich FGF
receptor regulates intracellular FGF‐1 and FGF‐2 levels. Journal of cellular
physiology, 170, 217-227.

135
CHAPTER 5
Concluding Summary and Future Directions

136
5.1 Concluding Summary
Marsupials are immunologically comparable to eutherians (Old et al., 2003), and like
eutherians, marsupial B-cells are necessary for an adaptive humoral response to occur
(Kalmutz, 1962). Early investigations into marsupial immunology included measuring
antibody responses (Kalmutz, 1962). These early antibody response studies showed
consistent differences between eutherians and marsupials in terms of their intensity of
primary and secondary antibody responses. Sustained primary antibody responses have been
investigated in the quokka (Yadav, 1971), opossum (Croix et al., 1989), and common
brushtail possum (Deakin et al., 2005), in which elevated IgG responses lasted 26, 37, and 15
weeks respectively. In marsupials secondary antibody responses were reportedly delayed and
considerably lower in magnitude compared to eutherian responses, as poor isotype switching
of IgM in secondary responses were observed (Yadav et al., 1972, Croix et al., 1989, Shearer
et al., 1995). Later studies utilised a variety of traditional immunisation procedures typically
used for eutherians to report more comparable responses between the two mammalian
lineages (Kitchener et al., 2002, Deakin et al., 2005). Prior to the completion and availability
of the opossum genome, comparative studies of eutherian and marsupial immune genes relied
heavily on the use of heterologous probes to isolate cDNA sequences from cDNA libraries.
Harrison and Wedlock (2000) designed consensus primers from eutherian gene sequences
with similar structure and function to isolate and clone a number of cytokines. However,
techniques such as these were limited to isolating only highly conserved genes (Wakefield
and Graves, 2005). The value of the opossum genome has been paramount for the
comparative analysis of distinctive traits of each mammalian lineage, as well as paving the
way for the identification of highly divergent genes (Mikkelsen et al., 2007).
The research presented in this thesis investigated the gene expression of gray short-
tailed opossum B-cells. My research has successfully analysed the usage of IgH and IgL

137
chain in the opossum B-cell repertoire of single cells, the first report for any marsupial
species. Furthermore, my research has investigated the single-cell transcriptome of opossum
B-cells and made a comparative analysis of the genes that were highly expressed.
The first objective of the thesis was to discuss the knowledge of marsupial B-cell
genetics in terms of genetic presence, homology and developmental stages in comparison to
eutherians. The literature review (Chapter 2) explored these key similarities and differences
between the two mammalian lineages and also identified the knowledge gaps and areas of
research that still require attention. This chapter gives a comprehensive review of signature
genes associated with B-cell development, differentiation and regulation. The maturation of
B-cells seems to be similar in both mammalian lineages, defined by several stages involving
the production of cells within the lymphoid tissues, the rearrangement of Igs and expression
of CD markers (Conley, 2003). Marsupial B-cells appear to develop differently compared to
eutherians, as the majority of development is delayed until after birth, and the production of
B-cells still occurs in the liver as the bone marrow is immature (Borthwick et al., 2014, Old,
2016). My review reports on the abundance and diversity of IgH and IgL isotypes
investigated in various marsupial species; four of the five Igs identified in eutherians and
monotremes have been reported in marsupials (Aveskogh and Hellman, 1998, Belov et al.,
1998, Aveskogh et al., 1999, Belov et al., 1999a, Belov et al., 1999b, Belov et al., 1999c).
IgD has not been identified in any investigated marsupial species to date, leaving researchers
to speculate its loss in all marsupials. The review also discusses the roles of CD markers,
signal transduction markers, and transcriptional regulators in a few marsupial species. A
number of CD markers have recently been identified in marsupials, CD79α and CD79β have
both been transcribed and characterised in the tammar wallaby (Duncan et al., 2010) and
bridled nail-tail wallaby (Suthers and Young, 2014). Duncan et al. (2010) investigated the
expression of CD79α and CD79β in developing pouch young, reporting the detection of

138
CD79α on day 10 postpartum, and CD79β on day 21 postpartum, concluding that the tammar
wallaby would only be able to elicit a humoral antibody response after both CD markers are
expressed. To date only the isoforms of the signal transduction molecule Lyn have been
characterised in marsupials (Suthers and Young, 2013). LynA and LynB in the tammar
wallaby and bridled nail-tail wallaby show the genomic organisation of these isoforms to be
highly conserved, suggesting their signalling mechanism is alike in all mammals (Suthers and
Young, 2013). Many transcriptional regulators have only been predicted in marsupial
genomic regions. My review highlights the importance of key regulators in terms of their role
in developmental and commitment processes in B-cells, as many of these key molecules are
yet to be identified in marsupials.
The usage of IgH and IgL isotypes has never been investigated in any marsupial
species before, which has left a major knowledge gap in the comparison of mammalian
immunological evolution. Therefore, the objective of the first study in the thesis (Chapter 3)
was to analyse the expression of IgH and IgL isotypes in the opossum B-cell repertoire. This
study determined IgM had the overall highest number of expressed cells, not surprisingly as
IgM is the first isotype to be formed during an immune response (Aveskogh et al., 1999). IgA
had an increased abundance of expressed blood cells, with a significantly high number of IgA
cells pairing with Igλ. IgG and IgE had low numbers of expressed cells in both the spleen and
blood. A major discovery in this study was the Igκ:Igλ ratio of expressed cells in the opossum
B-cell repertoire, a question which has been left unanswered for marsupials since the 1990s.
The analysis of the opossum single-cells shows a highly distinct Igκ:Igλ ratio of 35:65.
Previous investigations have shown this ratio to be highly diverse within the eutherian
lineage, in human, mice and rabbits expression of the IgL chain is clearly dominated by the
Igκ isotype (Butler, 1997, Kirschbaum et al., 1996, Waterfield et al., 1973). On the other end
of the scale, eutherian species dominated by Igλ usage include horses, dogs and cats (Home

139
et al., 1992, Arun et al., 1996), whilst pigs have a 50:50 ratio when it comes to IgL isotopic
expression (Butler and Brown, 1994). The V domain of the IgH and IgL chains contributes to
antibody diversity and establishes its specificity (Baker et al., 2005). My study identified a
limited set of VH gene segments in the expressed cells of the opossum in contrast to a much
higher variability of cells expressing Vκ and Vλ gene segments. Coevolution of opossum VH
and VL diversity does not appear to follow the same pattern of V diversification as seen in
other mammals investigated. The eutherian lineage has shown that species with highly
diverse VH domains also have a highly diverse VL domain, while eutherian species with low
VH diversity have low VL diversity (Butler, 1997, Sitnikova and Su, 1998).
A further investigation was undertaken to gain insights into the whole opossum B-cell
transcriptome (Chapter 4). By analysing the single-cells of paired Igs 14,654 unique genes
were identified, of which 13,376 were successfully annotated. Accumulation of all cells
found SDC4 was the most highly expressed gene throughout the entire transcriptome, with no
preferential expression between the IgH cells. SDC4 has previously been expressed in the
uterine transcriptome of a pregnant opossum (Hansen et al., 2016). ANOSIM pairwise and
SIMPER analyses found highly expressed ELMOD1 and GLG1 genes were significantly
expressed in IgM and IgG cells when compared to IgA cells. As cells expressing IgE were far
too low, no definitive comparison could be made between all marsupial IgH isotypes. MOTIF
comparison and phylogenetic analysis of ELMOD1 and GLG1 genes suggest their protein
sequences are highly conserved throughout mammalian evolution. Of all the genes annotated,
only 2.5% (340 genes) were termed immunological. Of these immune genes, the most
abundant were HLA-DRA and MODO-DAB1, both affiliated with the MHC Class II family.
These immune genes have previously been isolated and characterised in a number of
marsupial species; red-necked wallaby (Schneider et al., 1991), Tasmanian devil (Siddle et
al., 2007), common brushtail possum (Lam et al., 2001) and opossum (Stone et al., 1999).

140
Unlike the HLA-DRA gene, MODO-DAB1 has only been identified in marsupials,
suggesting that the MHC class II gene family has undergone a further evolutionary event
after the divergence of the mammalian lineages has occurred (Schneider et al., 1991).
5.2 Future Directions
The results presented in my thesis confirm the vastness of the opossum B-cell
transcriptome, and in some ways has raised more questions about the humoral immunity of
marsupials. Prior to this work, studies for IgH and IgL usage in the B-cell repertoire has been
limited to eutherian species. The opossum is now the first marsupial to have its ratio of
Igκ:Igλ usage identified in the B-cell repertoire, paving the way for future analysis. In
eutherians the IgL isotopic ratio is highly variable, therefore future transcriptomic analyses
are necessary to determine whether the marsupial lineage follows a similar pattern of IgL
isotopic ratio variability.
The results of this thesis have shown that marsupial VH and VL domain usage may be
coevolving in marsupial species. Due to the lack of VH gene segment diversity, IgL isotopic
V domains compensate by utilising a highly diverse pool of gene segments in the opossum B-
cell repertoire. The common brushtail possum is the only other marsupial species to be
investigated for VH and VL usage (Baker et al., 2005), and along with the opossum, this
pattern of VH and VL is contradictory to eutherians. Investigations with humans and mice
have demonstrated that IgH chains have a higher contribution to antibody-antigen
interactions than IgL chains (Wilson and Stanfield, 1994), leaving some researchers
questioning the importance of IgL chains (Pilström, 2002). Hence, further investigations are
needed on marsupial IgL isotype contribution to antibody diversity. Immune genes are known
to rapidly evolve within the genome and are therefore of great interest in comparative studies
of mammalian evolution. My thesis has provided an in depth analysis of the opossum B-cell

141
transcriptome, which can now be utilised as a reference point for future comparative studies
within and between the mammalian lineages.

142
5.3 Chapter 5 References
ARUN, S., BREUER, W. & HERMANNS, W. 1996. Immunohistochemical examination of
light‐chain expression (Λ/k ratio) in canine, feline, equine, bovine and porcine plasma
cells. Journal of Veterinary Medicine Series A, 43, 573-576.
AVESKOGH, M. & HELLMAN, L. 1998. Evidence for an early appearance of modern post-
switch isotypes in mammalian evolution; cloning of IgE, IgG and IgA from the
marsupial Monodelphis domestica. European Journal of Immunology, 28, 2738-2750.
AVESKOGH, M., PILSTROM, L. & HELLMAN, L. 1999. Cloning and structural analysis
of IgM (μ chain) and the heavy chain V region repertoire in the marsupial
Monodelphis domestica. Developmental & Comparative Immunology, 23, 597-606.
BAKER, M. L., BELOV, K. & MILLER, R. D. 2005. Unusually similar patterns of antibody
V segment diversity in distantly related marsupials. The Journal of Immunology, 174,
5665-5671.
BELOV, K., HARRISON, G. A. & COOPER, D. W. 1998. Molecular cloning of the cDNA
encoding the constant region of the immunoglobulin A heavy chain (Cα) form a
marsupial: Trichosurus vulpecula (common brushtail possum). Immunology Letters,
60, 165-170.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999a. Isolation and
sequence of a cDNA coding for the heavy chain constant region of IgG from the
Australian brushtail possum, Trichosurus vulpecula. Molecular Immunology, 36, 535-
541.
BELOV, K., HARRISON, G. A., MILLER, R. D. & COOPER, D. W. 1999b. Molecular
cloning of the brushtail possum (Trichosurus vulpecula) immunoglobulin E heavy
chain constant region. Molecular Immunology, 36, 1255-1261.
BELOV, K., HARRISON, G. A., ROSENBERG, G. H., MILLER, R. D. & COOPER, D. W.
1999c. Isolation and comparison of the IgM heavy chain constant regions from
Australian (Trichosurus vulpecula) and American (Monodelphis domestica)
marsupials. Developmental & Comparative Immunology, 23, 649-656.
BORTHWICK, C. R., YOUNG, L. J. & OLD, J. M. 2014. The development of the immune
tissues in marsupial pouch young. Journal of Morphology, 275, 822-839.
BUTLER, J. E. 1997. Immunoglobulin gene organization and the mechanism of repertoire
development. Scandinavian Journal of Immunology, 59, 455-462.
BUTLER, J. E. & BROWN, W. R. 1994. The immunoglobulins and immunoglobulin genes
of swine. Veterinary Immunology and Immunopathology, 43, 5-12.
CONLEY, M. E. 2003. Genes required for B cell development. Journal of Clinical
Investigation, 112, 1636-1638.
CROIX, D., SAMPLES, N., VANDEBERG, J. & STONE, W. 1989. Immune response of a
marsupial (Monodelphis domestica) to sheep red blood cells. Developmental &
Comparative Immunology, 13, 73-78.
DEAKIN, J. E., BELOV, K., CURACH, N. C., GREEN, P. & COOPER, D. W. 2005. High
levels of variability in immune response using antigens from two reproductive
proteins in brushtail possums. Wildlife Research, 32, 1-6.
DUNCAN, L., WEBSTER, K., GUPTA, V., NAIR, S. & DEANE, E. 2010. Molecular
characterisation of the CD79a and CD79b subunits of the B cell receptor complex in
the gray short-tailed opossum (Monodelphis domestica) and tammar wallaby
(Macropus eugenii): delayed B cell immunocompetence in marsupial neonates.
Veterinary Immunology and Immunopathology, 136, 235-47.

143
HANSEN, V. L., SCHILKEY, F. D. & MILLER, R. D. 2016. Transcriptomic changes
associated with pregnancy in a marsupial, the gray short-tailed opossum Monodelphis
domestica. PLOS ONE, 11, e0161608.
HARRISON, G. A. & WEDLOCK, D. N. 2000. Marsupial cytokines structure, function and
evolution. Developmental & Comparative Immunology, 24, 473-484.
HOME, W. A., FORD, J. E. & GIBSON, D. M. 1992. L chain isotype regulation in horse. I.
characterization of Ig lambda genes. Journal of Immunology 149, 3927-3936.
KALMUTZ, S. E. 1962. Antibody production in the opossum embryo. Nature, 193, 851-853.
KIRSCHBAUM, T., JAENICHEN, R. & ZACHAU, H. G. 1996. The mouse
immunoglobulin ϰ locus contains about 140 variable gene segments. European
Journal of Immunology, 26, 1613-1620.
KITCHENER, A., EDDS, L., MOLINIA, F. & KAY, D. 2002. Porcine zonae pellucidae
immunisation of tammar wallabies (Macropus eugenii): fertility and immune
responses. Reproduction, Fertility and Development, 14, 215-223.
LAM, M. K. P., BELOV, K., HARRISON, G. A. & COOPER, D. W. 2001. Cloning of the
MHC class II DRB cDNA from the brushtail possum (Trichosurus vulpecula).
Immunology Letters, 76, 31-36.
MIKKELSEN, T. S., WAKEFIELD, M., AKEN, B., AMEMIYA, C., CHANG, J., DUKE
BECKER, S., GARBER, M., GENTLES, A. J., GOODSTADT, L., HEGER, A.,
JURKA, J., KAMAL, M., MAUCELI, E., SEARLE, S., SHARPE, T., BAKER, M.
L., BATZER, M. A., BENOS, P., BELOV, K. & LINDBLAD-TOH, K. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in non-coding
sequences. Nature 447, 167-77.
OLD, J. M. 2016. Haematopoiesis in marsupials. Developmental & Comparative
Immunology, 58, 40-46.
OLD, J. M., SELWOOD, L. & DEANE, E. M. 2003. Development of lymphoid tissues of the
stripe-faced dunnart (Sminthopsis macroura). Cells Tissues Organs, 175, 192-201.
PILSTRÖM, L. 2002. The mysterious immunoglobulin light chain. Developmental &
Comparative Immunology, 26, 207-215.
SCHNEIDER, S., VINCEK, V., TICHY, H., FIGUEROA, F. & KLEIN, J. 1991. MHC class
II genes of a marsupial, the red-necked wallaby (Macropus rufogriseus):
identification of new gene families. Molecular Biology and Evolution, 8, 753-766.
SHEARER, M. H., ROBINSON, E. S., VANDEBERG, J. L. & KENNEDY, R. C. 1995.
Humoral immune response in a marsupial Monodelphis domestica: anti-isotypic and
anti-idiotypic responses detected by species-specific monoclonal anti-
immunoglobulin reagents. Developmental & Comparative Immunology, 19, 237-256.
SIDDLE, H. V., SANDERSON, C. & BELOV, K. 2007. Characterization of major
histocompatibility complex class I and class II genes from the Tasmanian devil
(Sarcophilus harrisii). Immunogenetics, 59, 753-760.
SITNIKOVA, T. & SU, C. 1998. Coevolution of immuoglobulin heavy- and light-chain
variable-region gene families. Immunogenetics, 15, 617-625.
STONE, W. H., BRUUN, D. A., FUQUA, C., GLASS, L. C., REEVES, A., HOLSTE, S. &
FIGUEROA, F. 1999. Identification and sequence analysis of an MHC class II B gene
in a marsupial (Monodelphis domestica). Immunogenetics, 49, 461-463.
SUTHERS, A. N. & YOUNG, L. J. 2013. Molecular identification and expression of Lyn
tyrosine kinase isoforms in marsupials. Molecular Immunology, 55, 310-318.
SUTHERS, A. N. & YOUNG, L. J. 2014. Isoforms of the CD79 signal transduction
component of the macropod B-cell receptor. Developmental & Comparative
Immunology, 47, 185-90.

144
WAKEFIELD, M. J. & GRAVES, J. A. 2005. Marsupials and monotremes sort genome
treasures from junk. Genome Biology, 6, 218.
WATERFIELD, M. D., MORRIS, J. E., HOOD, L. E. & TODD, C. W. 1973. Rabbit
immunoglobulin light chains: correlation of variable region sequences with allotypic
markers. The Journal of Immunology, 110, 227.
WILSON, I. A. & STANFIELD, R. L. 1994. Antibody-antigen interactions: new structures
and new conformational changes. Current Opinion in Structural Biology, 4, 857-867.
YADAV, M. 1971. The transmissions of antibodies across the gut of pouch-young
marsupials. Immunology, 21, 839.
YADAV, M., STANLEY, N. & WARING, H. 1972. The thymus glands of a marsupial,
Setonix brachyurus (quokka), and their role in immune responses: effect of
thymectomy on somatic growth and blood leucocytes. Australian Journal of
Experimental Biology and Medical Science, 50, 357-364.




























