Embed Size (px)
Citation preview

ACE2 deficiency is associated with impaired gestational weightgain and fetal growth restriction
Manish S Bharadwaj1, William B Strawn1, Leanne Groban2, Liliya M Yamaleyeva1, Mark CChappell1, Carina Horta1, Katie Atkins1, Luciana Firmes1, Susan B Gurley3, and K BridgetBrosnihan1
1The Hypertension and Vascular Research Center, Wake Forest School of Medicine, Winston-Salem, North Carolina, USA2Department of Anesthesiology, Wake Forest School of Medicine, Winston-Salem, NorthCarolina, USA3Department of Medicine, Duke University and Durham Veterans Affairs Medical Center, Durham,North Carolina, USA
AbstractAngiotensin converting enzyme 2 (ACE2) is a key enzyme of the renin-angiotensin system (RAS)that influences the relative expression of angiotensin (Ang) II and Ang-(1–7). Although ACE2expression increases in normal pregnancy, the impact of ACE2 deficiency in pregnancy has notbeen elucidated. We determined the influence of ACE2 deficiency on circulating and tissue RAScomponents, fetal and maternal growth characteristics, and maternal hemodynamics (mean bloodpressure (MBP) and cardiac output (CO)) at day 18 of gestation. Gestational body weight gain waslower in the ACE2 knock out (KO) vs C57BL/6 (WT) mice (30.3 ± 4.7 vs 38.2 ± 1.0 g, p<0.001).Fetal weight (0.94 ± 0.1 vs 1.24 ± 0.01 g, p<0.01) and length (19.6 ± 0.2 vs 22.2 ± 0.2 mm,p<0.001) were less in KO. MBP was significantly reduced in WT with pregnancy; it was elevated(p<0.05) in the KO virgin and pregnant mice, and this was associated with an increased CO inboth WT and KO pregnant mice (p<0.05). Plasma Ang-(1–7) was reduced in pregnant KO mice(p<0.05). Placenta Ang II levels were higher in KO mice (52.9 ± 6.0 vs 22.0 ± 3.3 fmol/mgprotein, p<0.001). Renal Ang II levels were greater in KO virgin mice (30.0 ± 1.7 vs 23.7 ± 1.1fmol/mg protein, p<0.001). There was no change in the Ang-(1–7) levels in the KO placenta andvirgin kidney. These results suggest that ACE2 deficiency and associated elevated placenta Ang IIlevels impact pregnancy by impairing gestational weight gain and restricting fetal growth.
KeywordsAng-(1–7); ACE2; renin-angiotensin system; pregnancy; fetus
IntroductionMaternal weight is an important determinant of optimal fetal development, pregnancyoutcome, and lifelong health as an adult.(1) Fetal growth restriction (FGR) remains a leadingcause of perinatal morbidity and mortality in humans.(2) Although there are many potential
Corresponding Author: K. Bridget Brosnihan, Ph.D., The Hypertension and Vascular Research Center, Wake Forest School ofMedicine, Medical Center Boulevard, Winston Salem, NC 27157-1032, Telephone: 336-716-2795, Fax: 336-716-2456,[email protected] of interest. None
NIH Public AccessAuthor ManuscriptHypertension. Author manuscript; available in PMC 2012 November 1.
Published in final edited form as:Hypertension. 2011 November ; 58(5): 852–858. doi:10.1161/HYPERTENSIONAHA.111.179358.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

causes of FGR, the most common identifiable cause is suboptimal uteroplacental perfusion.(3) Vasoactive mediators play an important role in the regulation of the vasculature of theuteroplacental bed. Among these are the peptide components of the renin-angiotensinsystem (RAS), primarily angiotensin II (Ang II) and Ang-(1–7). Angiotensin convertingenzyme 2 (ACE2) is a pleiotropic monocarboxypeptidase shown to efficiently metabolizeAng II to Ang-(1–7).(4) Ang II has well-characterized vasoconstrictive, proliferative, andangiogenic actions. In contrast, Ang-(1–7) has vasodilatory, antiproliferative, andantiangiogenic actions. In Ang II-induced cardiac hypertrophy and remodeling and pressure-overload heart failure, simultaneous administration of Ang-(1–7) counterbalances thedetrimental effects of Ang II.(5–8) ACE2 deficient mice have impaired cardiovascularfunction, enhanced oxidative stress, and inflammatory cytokine expression.(9) These studiesillustrate a critical role for the ACE2-dependent balance of Ang II and Ang-(1–7).
The circulating RAS is elevated in normal pregnancy, with hemodynamics characterized bya reduction in systemic vascular resistance and no increase or a reduction in systemic bloodpressure.(10;11) Pregnancy is associated with increased renal and uterine ACE2 mRNAexpression and activity as compared with virgin rats.(12;13) In the current study, wehypothesized that increases in ACE2 expression in the kidney and reproductive tissuesduring pregnancy have a protective role in renal, cardiovascular, and reproductive function.Therefore we determined the effect of pregnancy on, circulating and tissue RAScomponents, fetal and maternal growth characteristics, and maternal hemodynamics (bloodpressure, cardiac output) in both the ACE2 knockout (KO) and C57BL/6 (WT).
Materials/MethodsFor a description of Materials and Methods please see http://hyper.ahajournals.org
ResultsFigure S1 shows the ACE2 protein in placental tissue from ACE2 KO mice as well as WTfemales by Western blot. As expected, the glycosylated 125 kDa form of ACE2 wasdetected in the WT placenta, while it was not seen in the placenta from KO mice (FigureS1A). Upon preincubation of the ACE2 antibody with the antigenic peptide, the 125 kDaimmunoreactive band was absent in the WT, further confirming the specificity of ourantibody for mouse ACE2 (Figure S1B).
Maternal characteristicsFigure 1 shows the maternal characteristics of the WT and KO virgin and pregnant mice.There was no difference in body weight between virgin KO and WT females (16.1 ± 3.3 vs16.6 ± 0.4 g, respectively). Gestational body weight gain was significantly lower in the KOmice compared to WT mice (30.3 ± 4.7 vs 38.2 ± 1.0 g, respectively, p<0.001, KO vs WT).This difference (23 ± 1.3 vs 28 ± 0.5 g, p<0.05) persisted even when the total pup weight/pregnancy was subtracted. Mean blood pressure (MBP) was significantly higher in KOvirgin females as compared to WT virgins (86 ± 2 vs 76 ± 1 mmHg, p<0.01). MBP wasreduced in WT pregnant animals (76 ± 1 vs 69 ± 2 mmHg, p<0.05). In contrast to thepregnant WT, the higher pressure was maintained in the pregnant KO (84 ± 2.5 vs 86 ± 2mmHg, ns, pregnant vs virgin KO). There was a significant effect of pregnancy on cardiacoutput (20.8 ± 1.6 vs 25.3 ± 2.7 ml/min (WT, n = 10, 4) and 20.2 ± 1.3 vs 25.3 ± 2.8 ml/min(KO, n = 8, 9), virgin vs pregnant p<0.05).
Bharadwaj et al. Page 2
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fetal characteristicsFigure 2 provides the fetal characteristics for KO and WT mice. Average pup weight (0.94 ±0.1 vs 1.24 ± 0.01 g, p<0.01) and length (19.6 ± 0.2 vs 22.2 ± 0.2 mm, p<0.01) weresignificantly lower in KO as compared to WT mice. Total pup weight/pregnancy was alsosignificantly lower in KO mice (6.3 ± 0.9 vs 10.0 ± 0.4 g, p<0.01). There was a small butsignificant increase in fetal resorptions in the KO mice (approximately 1 fetus perpregnancy) compared to the WT mice (1 fetus per 5 pregnancies). There was a positivecorrelation of maternal weight to total pup weight/pregnancy (r = −0.88, p<0.001) (Figure1D).
RAS components in the plasma of KO and WT miceFigure 3A shows the circulating angiotensin peptide profile in the WT and KO virgin andpregnant mice. There was no change in the plasma levels of Ang I, Ang II and Ang-(1–7) invirgin KO and WT mice. Plasma Ang-(1–7) was significantly reduced in KO pregnantfemales, but plasma Ang I and Ang II were unchanged compared to pregnant WT. Figure3Bi shows the serum ACE activity using the synthetic substrate Hip-Gly-Gly. Pregnancywas associated with a significant reduction in serum ACE activity for WT and KO mice(185.17 ± 7.50 vs 143.50 ± 5.50 nmol/ml/min, p<0.01; 189.50 ± 2.10 vs 140.00 ± 17.16nmol/ml/min, p<0.05), respectively. In order to compare the relative amounts of ACE andACE2 in serum, we conducted additional measurements using 125I labeled Ang I and Ang II,respectively as substrates. There was a decrease in ACE activity with pregnancy in the WTmice (Figure 3Bii). Figure 3Biii shows that serum ACE2 activity is higher in the pregnant ascompared to virgin WT mice (13.2 ± 0.4 vs 8.9 ± 1.1 fmol/ml/min, p<0.001). In WT mice,ACE activity was more than 30-fold higher compared to ACE2 activity, indicating that ACEis considerably more abundant in the circulation than ACE2.
RAS components in the kidney of KO and WT miceFigure 4A shows the peptide profile in the KO and WT virgin and pregnant female kidneytissue. Ang II levels were greater in virgin KO vs WT mice (30.0 ± 1.7 vs 23.7 ± 1.1 fmol/mg protein, p<0.001), but not in the kidneys from pregnant KO vs WT mice. Ang I andAng-(1–7) levels were higher in WT pregnant animals compared to virgins. Figure 5Bshows ACE and NEP activity levels in the kidney. The KO animals had higher NEP activitycompared to WT mice. There was no difference in kidney tissue ACE activity in KO vs WTvirgin and pregnant mice.
RAS components in the placenta of KO and WT miceFigure 5A shows the peptide profile in the placenta of KO and WT mice. Ang II levels were2.5-fold greater in KO vs WT mice (52.9 ± 6.0 vs 22.0 ± 3.3 fmol/mg protein, p<0.01),however, Ang-(1–7) and Ang I levels were unchanged. Placental ACE activity was reducedin KO mice compared to WT (13.8 ± 1.7 vs 28.0 ± 2.6 nmol/hr/mg protein, p<0.005), andthere was no difference in the NEP activity (Figure 5B).
RAS peptide levels in the heart, uterus and fetal membranes of KO and WT miceSupplemental tables S1–S3 provide the Ang I, Ang II, and Ang-(1–7) levels in the heart,uterus, and fetal membranes from WT and KO virgin as well as pregnant mice. There wereno differences in the levels of peptides in the heart, uterus, or fetal membranes. Ang II andAng-(1–7) content in the fetal membranes was markedly higher than all other tissuesstudied.
Bharadwaj et al. Page 3
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

DiscussionThis is the first report to demonstrate the impact of ACE2 deficiency on maternal as well asfetal growth. ACE2, a carboxypeptidase, is a homologue of ACE that cleaves Ang II intoAng-(1–7) with high efficiency. ACE2 acts in a counter-regulatory manner to ACE to shiftthe balance between Ang II and Ang-(1–7). There is limited data on the specific functions ofACE2 in vivo, and no data on ACE2 function in pregnancy. Therefore, the present study wasundertaken to study the impact of ACE2 deficiency on maternal and fetal growth in miceduring pregnancy. We showed that gestational body weight gain and average and total pupweight were lower in the KO mice compared to the WT mice. We also evaluated the impactof the ACE2 deficiency on the major RAS peptides and maternal hemodynamics. Ang IIwas increased in the placenta of KO animals and in the kidney of virgin KO animals. Theincrease in Ang II in virgin kidney was absent in KO pregnant animals. Plasma Ang-(1–7)was decreased in the KO pregnant mice, a finding consistent with the circulating profile ofpreeclamptic women.(10)
Maternal gestational body weight gain was decreased in pregnant KO females compared topregnant WT females. Body weight increase during gestation is associated with increasedfood and fluid intake, expansion of plasma volume and enhanced cardiac output.(14;15)These changes are usually associated with a normal or decreased blood pressure, reflectingthe reduction in total peripheral resistance due to the prominent vasodilation.(15) Withpregnancy, WT mice showed a significant reduction in MAP as compared to virgins. Invirgin KO females MAP was higher than WT virgin females. The increase in MAP is inagreement with the original study by Gurley et al (16) where blood pressure was increasedin the male KO. These observations were made in KO mice on C57BL/6 background. In theKO mice on a 129/SvEv background, however, they did not see any difference in bloodpressure compared to WT, emphasizing that the different phenotypic outcomes for KO miceof different genetic backgrounds. In our study the increase in MAP in KO virgin mice mayreflect higher kidney Ang II content without a change in Ang-(1–7), indicating a shift in thebalance of local RAS tissues toward Ang II. An increase in renal Ang II could contribute toincreased vasoconstriction and fluid and water retention. In pregnant KO mice, MAPremained elevated compared to pregnant WT females. In these animals that exhibit noincrease in renal Ang II, the basis for the increase in blood pressure may reflect greaterplacental Ang II resulting in an increased placental bed resistance.
In spite of the increase in blood pressure, Gurley et al (16) did not report any difference incardiac dimensions in the KO mice on the two backgrounds, which is different fromCrackower’s study reporting that KO mice on a mixed background develop left ventriculardilation.(4) Pregnancy is well known to have substantial hemodynamic changes and COincreases by 35–40%.(15) As expected with normal pregnancy, CO increased in WTpregnant mice; in the pregnant KO mice CO increased to a similar level. Because CO wassimilarly increased in both WT and KO pregnant animals, our data would suggest that theimpaired gestational weight gain and fetal growth restriction in ACE2 KO pregnancy cannotbe attributed to inadequate maternal systemic cardiac output changes.
Previous studies using the antagonist [D-Ala7]-Ang-(1–7) [A779] to block the endogenousactions of Ang-(1–7) in pregnant rats revealed reduced water and food intake, as well as alower amount of urine.(14) It is possible that in pregnancy, ACE2 deficiency and theassociated reduced plasma Ang-(1–7) adversely affected the thirst and hunger centers of thebrain resulting in decreased water and food intake, thereby leading to reduced body weight.Because the reduced water and food intake accompanied the antidiuresis, the preponderanceof effects in KO pregnant animals appears to be central actions of the reduced Ang-(1–7)controlling food and water intake. The central effects would be balanced by the reduced
Bharadwaj et al. Page 4
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

diuresis that is also observed in pregnant animals, which may contribute to the increase incardiac output. The basis for the maternal weight reduction together with the increased MAPand CO in the KO requires further exploration.
ACE2 deficiency resulted in significant inhibition of fetal growth in addition to impairedgestational weight gain. Fetal weight and length were lower in the KO mice compared toWT mice, and there was approximately 1 fetal resorption/pregnancy. These findings point toan important survival role of ACE2 in normal pregnancy. The mechanism for the resorptionsis unknown. Crackower et al. (4) reported in the first study describing the KO mice on amixed genetic background that mice appeared healthy, without any gross detectablealterations in all organs analyzed, and at the expected Mendelian frequency. However, theassessment of fetal characteristics was not the primary goal of their study, and onlyqualitative observations were presented. Maternal body size has an influence on fetalgrowth;(17–19) smaller mothers produce smaller offspring than larger mothers, and this maybe one of the causes of poor fetal growth in the KO females. Our data confirms thisobservation by the strong positive correlation between maternal growth and fetal growth. Inaddition, the uterine environment is a key determinant of fetal growth.(20) The increasedAng II in the placenta of the pregnant KO mice could lead to increased placental ischemia.Increased Ang II was observed in the chorionic villi of the placenta of preeclamptic women(21) and in both the maternal and fetal components of the placenta of the preeclampsia rattransgenic model arising from the cross of the female human angiotensinogen with the malehuman renin.(22) In both preeclamptic women and the human transgenic rat model ofpreeclampsia, fetal growth restriction accompanied the activated placental RAS. Finally,because of the observation of pre-pregnancy elevated blood pressure and elevated kidneyAng II levels in the virgin KO animals, the question of systemic maternal factorscontributing to later detrimental outcomes during pregnancy cannot be eliminated.
No differences in plasma Ang II were detected among KO and WT virgin and pregnantmice. Gurley et al (16) also reported no differences between WT and KO mice in circulatingAng II levels. These results differ from the report by Crackower et al (4) who showed thatplasma Ang II was increased in the KO model. The disparity in these findings may beattributed to the differing backgrounds, the gender and age of the animals, and theaccompanying cardiovascular disease in the ACE2 KO mice from Crackower’s study at thetime of measurement. On the other hand, plasma Ang-(1–7) in our study was significantlydecreased in KO pregnant mice. Previously we reported that preeclamptic women at lategestation showed lower plasma Ang-(1–7).(10) In the present study, the profile incirculating peptides was accompanied by a lower serum ACE activity in the pregnant ascompared to the virgin females in both WT and KO animals. The decrease of ACE inpregnancy observed in this study is consistent with previous studies conducted duringpregnancy (10) and with estrogen treatment in rats (23) and mice.(24) Another aspect ofthese studies is that we used the endogenous substrates Ang I and Ang II assayed underidentical conditions to provide an accurate assessment of the relative activities of ACE andACE2 in the serum. We demonstrated that serum ACE activity was 30-fold higher comparedto serum ACE2 activity. This differential expression of the two enzymes in the circulationwith ACE2 being present at substantially lower levels may explain why there was no changein serum peptides with ACE2 deletion. Serum ACE2 activity was increased in pregnantcompared to virgin WT mice. This pattern of opposite changes in ACE2 and ACE activitiesis consistent with the counter-regulatory mechanism of their actions on peptide formation.
In the kidney, Ang II levels were increased in virgin KO mice compared to virgin WT mice.Gurley et al (16) showed a 6-fold increase in renal Ang II in KO males following Ang IIinfusion, but did not report baseline Ang II values. Crackower et al (4) reported increasedAng II content in kidney of KO mice. Thus, the increase in Ang II in the kidney is consistent
Bharadwaj et al. Page 5
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

with the lack of ACE2 enzyme in the KO virgin mice. In the present study, this increase wasabsent in pregnant mice.
To further investigate the basis for the angiotensin peptide profile in pregnant KO mice, wedetermined NEP and ACE activity levels in the kidney. NEP activity was higher in virginand pregnant KO mice compared to WT, suggesting a negative feedback regulation betweenACE2 and NEP in the WT. NEP converts either Ang I or Ang-(1–9) to Ang-(1–7), but alsocontributes to the degradation of Ang II.(25) In ACE2 KO mice, a decrease in Ang-(1–7)was expected in the presence of elevated Ang II, because of the reduced degradation of AngII; however, the presence of elevated NEP may have contributed to the maintenance of Ang-(1–7) in the kidney.
Placental Ang II was higher in ACE2 deficient mice compared to WT mice. This findingsuggests that Ang II is a primary substrate for ACE2 in the placenta, since its degradation isprevented in the absence of ACE2. In contrast to the expected decrease in Ang-(1–7) in theplacenta of the KO, Ang-(1–7) levels were not changed as compared to the WT mice. Thisfinding suggests that redundant enzymes may contribute to the formation of Ang-(1–7) inthe placenta. Our study demonstrated no difference in the placental NEP but a decrease inACE activity in the KO. The absence of ACE2 with a reduction in ACE in the presence ofelevated Ang II suggests that ACE2 is the predominate enzyme that is responsible for thebreakdown of Ang II with a lesser contribution of ACE to the generation of Ang II. BecauseACE can degrade Ang-(1–7) and convert it to Ang-(1–5), a decrease in ACE activity in theplacenta is consistent with a decrease in degradation of Ang-(1–7) by ACE,(26) which couldcontribute to the maintenance of Ang-(1–7) in the placenta. In contrast to the kidney andplacenta, the lack of change of angiotensin peptides in the uterus, heart and fetal membranesindicates that there is tissue specific regulation of peptide expression and that differenttissues show a distinct profile of peptide changes.
This study surprisingly showed very little change in the Ang-(1–7) levels in tissue. The RASsystem is complicated and part of the complexity is the redundancy of enzymes to act onmultiple substrates. Thus, the endopeptidases neprilysin, prolyl oligopeptidase, and thimetoligopeptidase, as well as prolyl carboxypeptidase, all can generate Ang-(1–7). In this modelany of these enzymes can contribute to maintain Ang-(1–7) in the face of increased Ang II.Since ACE2 is an Ang-(1–7) synthesizing enzyme, a reduction in Ang-(1–7) levels in thetissue was anticipated in the KO animals, especially in the placenta and virgin kidney whereAng II was increased. In the placenta, the reduction in ACE activity is consistent with itscontribution to the maintenance of Ang-(1–7), but the lack of change in NEP point to a non-NEP enzyme that may be compensating for the reduction in ACE2. In the kidney, theincrease in NEP could contribute to the maintenance of Ang-(1–7).
Finally, ACE2 acts on substrates unrelated to the RAS. Apelin is a multifunctional peptidewhich is catabolized by ACE2 and has major actions related to cardiovascular function,body fluid homeostasis, and energy metabolism. Van Mieghem et al (27) showed that apelinlevels drop by 50% in the last week of gestation and proposed that the drop in apelin wasdue to the increased expression of ACE2 in the placenta which they propose is the site ofincreased clearance of the peptide.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe thank the Hypertension Core Laboratory for their measurement of angiotensin peptides.
Bharadwaj et al. Page 6
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Sources of funding. This work was supported in part by grants from the National Institutes of Health, NHLBI-P01HL51952 (KBB), K08-AG026764(LG), and R01-AG033727(LG). The authors gratefully acknowledge grantsupport in part provided by Unifi, In. Greensboro, NC and Farley-Hudson Foundation, Jacksonville, NC.
Reference List1. Kramer MS. Determinants of low birth weight: methodological assessment and meta-analysis. Bull
World Health Organ. 1987; 65:663–737. [PubMed: 3322602]2. McCormick MC. The contribution of low birth weight to infant mortality and childhood morbidity.
N Engl J Med. 1985; 312:82–90. [PubMed: 3880598]3. Murphy VE, Smith R, Giles WB, Clifton VL. Endocrine regulation of human fetal growth: the role
of the mother, placenta, and fetus. Endocr Rev. 2006; 27:141–169. [PubMed: 16434511]4. Crackower MA, Sarao R, Oudit GY, Yagil C, Kozieradzki I, Scanga SE, Oliveira-dos-Santo AJ, da
Costa J, Zhang L, Pei Y, Scholey J, Bray MR, Ferrario CM, Backx PH, Manoukian AS, ChappellMC, Yagil Y, Penninger JM. Angiotensin-converting enzyme 2 is an essential regulator of heartfunction. Nature. 2002; 417:822–828. [PubMed: 12075344]
5. Huentelman MJ, Grobe J, Vazquez J, Stewart JM, Mecca AP, Katovich MJ, Ferrario CM, RaizadaMK. Protection from angiotensin II-induced cardiac hypertrophy and fibrosis by systemic lentiviraldelivery of ACE2 in rats. Exp Physiol. 2005; 90:783–790. [PubMed: 16049057]
6. Ferrario CM, Brosnihan KB, Diz DI, Jaiswal N, Khosla MC, Milstea OA, Tallant EA. Angiotensin-(1–7): a new hormone of the angiotensin system. Hypertension. 1991; 18(Suppl III):III-126–III-133.[PubMed: 1937675]
7. Machado RD, Santos RA, Andrade SP. Mechanisms of angiotensin-(1–7)-induced inhibition ofangiogenesis. American Journal of Physiology - Regulatory Integrative & Comparative Physiology.2001; 280:R994–R1000.
8. Brosnihan KB, Li P, Ferrario CM. Angiotensin-(1–7) dilates canine coronary arteries through kininsand nitric oxide. Hypertension. 1996; 27:523–528. [PubMed: 8613197]
9. Oudit GY, Crackower MA, Backx PH, Penninger JM. The role of ACE2 in cardiovascularphysiology. Trends Cardiovasc Med. 2003; 13:93–101. [PubMed: 12691672]
10. Merrill DC, Karoly M, Chen K, Ferrario CM, Brosnihan KB. Angiotensin-(1–7) in normal andpreeclamptic pregnancy. Endocrine. 2002; 18:239–245. [PubMed: 12450315]
11. August P, Lenz T, Ales KL, Druzin ML, Edersheim TG, Hutson JM, Muller FB, Laragh JH, SealeyJE. Longitudinal study of the renin-angiotensin-aldosterone system in hypertensive pregnantwomen: Deviations related to the development of superimposed preeclampsia. Am J ObstetrGynecol. 1990; 163:1612–1621.
12. Neves LAA, Stovall K, Joyner J, Valdes G, Gallagher PE, Ferrario CM, Merrill DC, BrosnihanKB. ACE2 and Ang-(1–7) in the uterus during early and late gestation. Am J Physiol Regul IntegrComp Physiol. 2007; 294:R151–R161. [PubMed: 17977916]
13. Joyner J, Neves LA, Granger JP, Alexander BT, Merrill DC, Chappell MC, Ferrario CM, DavisWP, Brosnihan KB. Temporal-spatial expression of angiotensin-(1–7) and angiotensin convertingenzyme 2 in the kidney of normal and hypertensive pregnant rats. Am J Physiol Regul IntegrComp Physiol. 2007; 239:R169–R177. [PubMed: 17428896]
14. Joyner J, Neves LA, Stovall K, Ferrario CM, Brosnihan KB. Angiotensin-(1–7) serves as anaquaretic by increasing water intake and diuresis in association with downregulation ofaquaporin-1 during pregnancy in rats. Am J Physiol Regul Integr Comp Physiol. 2008;294:R1073–R1080. [PubMed: 18184763]
15. Dowell RT, Kauer CD. Maternal hemodynamics and uteroplacental blood flow throughoutgestation in conscious rats. Methods Find Exp Clin Pharmacol. 1997; 19:613–625. [PubMed:9500125]
16. Gurley SB, Allred A, Le TH, Griffiths R, Mao L, Philip N, Haystead TA, Donoghue M, BreitbartRE, Acton SL, Rockman HA, Coffman TM. Altered blood pressure responses and normal cardiacphenotype in ACE2-null mice. J Clin Invest. 2006; 116:2218–2225. [PubMed: 16878172]
17. Robinson J, Chidzanja S, Kind K, Lok F, Owens P, Owens J. Placental control of fetal growth.Reprod Fertil Dev. 1995; 7:333–344. [PubMed: 8606942]
Bharadwaj et al. Page 7
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

18. Thompson JMD, Wall C, Becroft DMO, Robinson E, Wild CJ, Mitchell EA. Maternal dietarypatterns in pregnancy and the association with small-for-gestational-age infants. British Journal ofNutrition. 2010; 103:1665–1673. [PubMed: 20211035]
19. Cogswell ME, Yip R. The influence of fetal and maternal factors on the distribution of birthweight.Semin Perinatol. 1995; 19:222–240. [PubMed: 7570074]
20. Brooks AA, Johnson MR, Steer PJ, Pawson ME, Abdalla HI. Birth weight: nature or nurture?Early Hum Dev. 1995; 42:29–35. [PubMed: 7671843]
21. Anton L, Merrill DC, Neves LAA, Stovall K, Gallagher PE, Diz DI, Moorefield C, Gruver C,Ferrario CM, Brosnihan KB. Activation of local chorionic villi angiotensin II levels but notangiotensin (1–7) in preeclampsia. Hypertension. 2008; 51:1066–1072. [PubMed: 18259034]
22. Brosnihan KB, Hering L, Dechend R, Chappell MC, Herse F. Increased angiotensin II in themesometrial triangle of a transgenic rat model of preeclampsia. Hypertension. 2010; 55:562–566.[PubMed: 20038747]
23. Gallagher PE, Li P, Lenhart JR, Chappell MC, Brosnihan KB. Estrogen regulation of angiotensin-converting enzyme mRNA. Hypertension. 1999; 33:323–328. [PubMed: 9931124]
24. Brosnihan KB, Hodgin JB, Smithies O, Maeda N, Gallagher P. Tissue-specific regulation of ACE/ACE2 and AT1/AT2 receptor gene expression by oestrogen in apolipoprotein E/oestrogenreceptor-alpha knock-out mice. Exp Physiol. 2008; 93:658–664. [PubMed: 18192335]
25. Allred AJ, Diz DI, Ferrario CM, Chappell MC. Pathways for angiotensin-(1–7) metabolism inpulmonary and renal tissues. Am J Physiol. 2000; 279:F841–F850.
26. Chappell MC, Allred AJ, Ferrario CM. Pathways of angiotensin-(1–7) metabolism in the kidney.Nephrol Dial Transplant. 2001; 16(Suppl 1):22–26. [PubMed: 11369816]
27. Van Mieghem T, Van Bree R, Van Herck E, Pijnenborg R, Deprest J, Verhaeghe J. Maternalapelin physiology during rat pregnancy: the role of the placenta. Placenta. 2010; 31:725–730.[PubMed: 20580085]
Bharadwaj et al. Page 8
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Perspectives
In the present study we demonstrated that ACE2 deficiency is associated with impairedgestational weight gain and restricted fetal growth. ACE2 deficiency affected theangiotensin profiles in the kidney of virgin animals and the placenta with an increase inAng II. Only in the circulation was the shift in the Ang II/Ang-(1–7) balanceaccompanied with a decrease in Ang-(1–7). Our findings have uncovered a unique,protective role for ACE2 in normal pregnancy. Correcting the ACE2 deficiency duringabnormal pregnancy may provide the potential to prevent impaired gestational weightgain and fetal growth restriction. The study has also raised questions about thecomplexity and redundancy of enzymes of the RAS, which resulted in the unexpectedmaintenance of tissue Ang-(1–7) levels.
Bharadwaj et al. Page 9
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1.Maternal characteristics of virgin and pregnant C57BL/6 and ACE2KO females. Panel A.Pregnant ACE2KO body weight was significantly reduced compared to C57BL/6 pregnantfemales(n = 7). Panel B. ACE2KO maternal weight - total pup weight/pregnancy wassignificantly reduced compared to C57BL/6 females (n = 7). Panel C. Mean blood pressurewas significantly higher for virgin and pregnant ACE2KO (n = 5, 4) females compared tothe respective C57BL/6 females (n = 7, 5). Values are mean ± SEM. * p<0.05 vs virgin; #p<0.01; ## p<0.001 vs C57BL/6.
Bharadwaj et al. Page 10
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
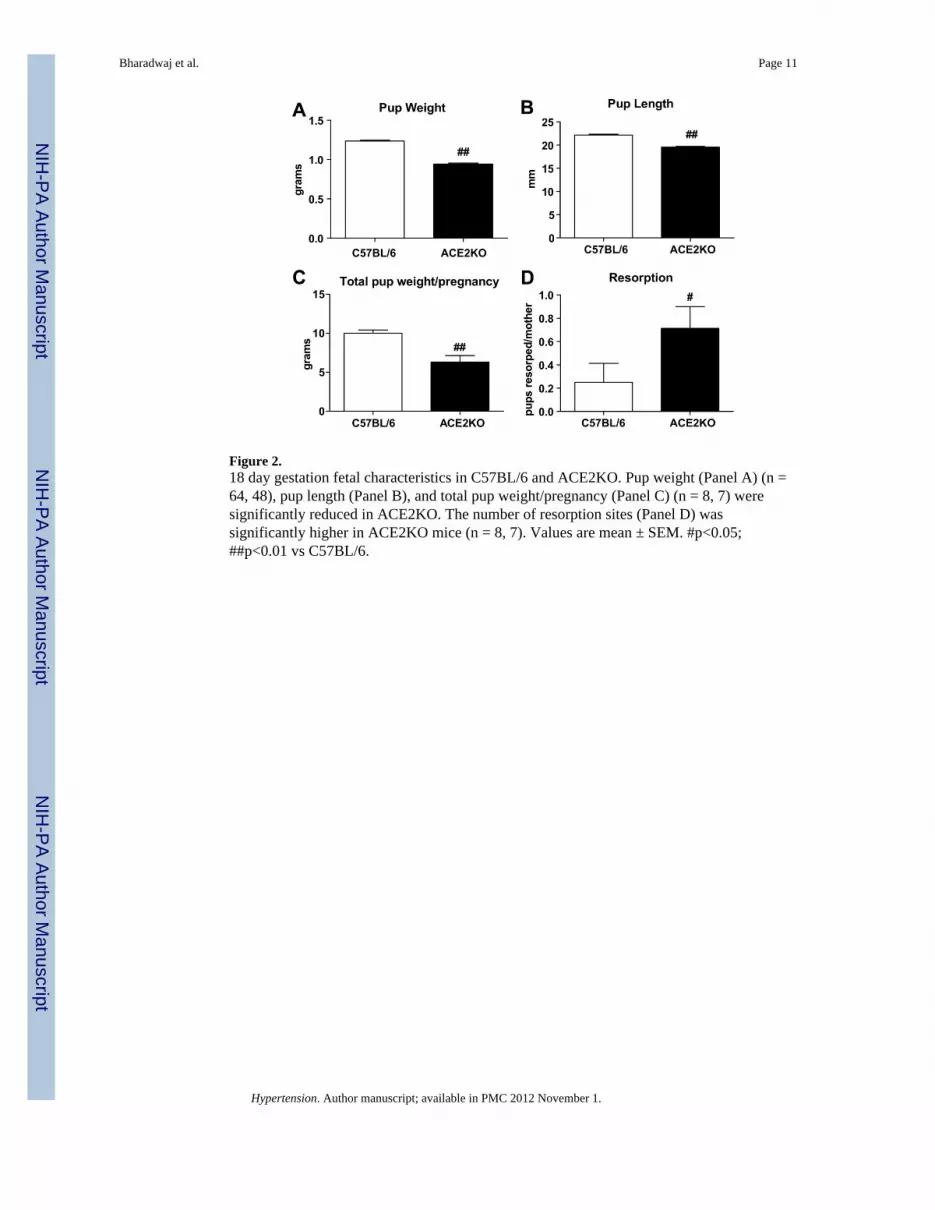
Figure 2.18 day gestation fetal characteristics in C57BL/6 and ACE2KO. Pup weight (Panel A) (n =64, 48), pup length (Panel B), and total pup weight/pregnancy (Panel C) (n = 8, 7) weresignificantly reduced in ACE2KO. The number of resorption sites (Panel D) wassignificantly higher in ACE2KO mice (n = 8, 7). Values are mean ± SEM. #p<0.05;##p<0.01 vs C57BL/6.
Bharadwaj et al. Page 11
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3.Panel A. Angiotensin peptide levels in the plasma of virgin and pregnant C57BL/6 andACE2KO mice (n = 9, 6, 6, 6). No significant differences were seen among the groups forAng I or Ang II. Ang-(1–7) was reduced in pregnant ACE2KO vs virgin ACE2KO. Panel B.Serum ACE and ACE2 activity. Panel Bi. Serum ACE activity measured using an ALPCOdiagnostic Kit (n = 10, 4, 6, 3). ACE activity was significantly reduced in pregnant C57BL/6and ACE2KO. Panel Bii. Serum ACE activity using 125I labeled Ang I (n = 4). ACE activitywas significantly reduced in pregnant ACE2KO females compared to virgin ACE2KOfemales. Panel Biii. Serum ACE2 activity using 125I labeled Ang II (n = 4). ACE2 activitywas significantly increased in pregnant C57BL/6 mice. Values are mean ± SEM. *p<0.05;**p<0.001 vs virgin of the same background.
Bharadwaj et al. Page 12
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4.Panel A. Angiotensin peptide levels in the kidney of virgin and pregnant C57BL/6 andACE2KO mice (n = 20, 12, 8, 7). Ang II levels were significantly increased in virginACE2KO females. Ang I and Ang-(1–7) were significantly increased in pregnant C57BL/6mice. Panel B. ACE and NEP activity in the kidney (n = 5, 5, 4, 5). NEP was significantlyincreased in virgin and pregnant ACE2KO mice. Values are mean ± SEM. **p<0.01;***p<0.001 vs virgin of the same background; #p<0.05; ##p<0.01, ###p<0.001 vs C57BL/6.
Bharadwaj et al. Page 13
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5.Panel A. Angiotensin peptide levels in the placenta of pregnant C57BL/6 and ACE2KOmice. Ang II levels were increased in ACE2KO females compared to C57BL/6 (n = 6, 7).Panel B. ACE and NEP activity in the placenta (n = 6, 4). ACE was decreased in ACE2KO,whereas NEP was unchanged as compared to C57BL/6. Values are mean ± SEM. ##p<0.01vs C57BL/6.
Bharadwaj et al. Page 14
Hypertension. Author manuscript; available in PMC 2012 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript





























