Embed Size (px)
Citation preview

Antibody detection-based differential ELISA for
NDV-infected or vaccinated chickens versus
NDV HN-subunit vaccinated chickens
Andrea M. Makkaya, Peter J. Krellb, EÂ va Nagya,*
aDepartment of Pathobiology, Ontario Veterinary College, University of Guelph,
Guelph Ont., Canada N1G 2W1bDepartment of Microbiology, University of Guelph, Guelph Ont., Canada N1G 2W1
Received 27 May 1998; accepted 3 February 1999
Abstract
With the advent of subunit vaccines for microbial diseases it is becoming increasingly important
to be able to differentiate naturally infected animals from those vaccinated with the corresponding
subunit vaccine. For avian viruses such as Newcastle disease virus (NDV), a whole virus-based
ELISA cannot make such a differential diagnosis since in both cases the antisera would react with
the whole virus. The nucleocapsid protein (NP) gene of the NDV Hitchner B1 strain was cloned,
sequenced and expressed to develop a differential ELISA. The B1 NP had 95.7 and 96.1% amino
acid identities with the NP of the d26 and Ulster 2C strains, respectively. The B1 NP expressed in a
baculovirus expression vector (recNP) was the expected size and reacted with NDV-specific
antibodies (Ab) in Western blots and by radioimmunoprecipitation. The ELISA using recNP-coated
wells, tested on serum samples from flocks pretested with a commercial NDV kit gave results
corresponding to those of the kit. Furthermore, use of both the recNP-based ELISA and a whole
virus ELISA allowed the differentiation of birds vaccinated with a NDV haemagglutinin±
neuraminidase (HN) expressing fowlpox virus from birds infected with NDV. This provides the
basis for establishing an ELISA that discriminates between the antibody response to a recombinant
fowlpox vaccine (expressing NDV HN protein) and that to live and inactivated NDV. # 1999
Elsevier Science B.V. All rights reserved.
Keywords: Chicken±Viruses; Newcastle disease virus; Differential ELISA; Baculovirus expression; NDV
nucleocapsid protein
Veterinary Microbiology 66 (1999) 209±222
* Corresponding author. Tel.: +1-519-824-4120 x 4783; fax: +1-519-767-0809; e-mail:[email protected]
0378-1135/99/$ ± see front matter # 1999 Elsevier Science B.V. All rights reserved.
PII: S 0 3 7 8 - 1 1 3 5 ( 9 9 ) 0 0 0 1 6 - 4

1. Introduction
Newcastle disease virus (NDV) is a widespread and economically important poultry
pathogen. Although vaccines have long been available and administered to control
Newcastle disease, the virus remains an ongoing threat to commercial flocks. The forms
of the disease vary and are dependent upon several factors, but mainly on the strain of the
virus (Alexander, 1997).
NDV, a member of the family Paramyxoviridae (Rima et al., 1995), is an enveloped
virus with helical nucleocapsid symmetry and a single-stranded, negative sense RNA
genome, about 15 kb in length. In the 30±50 orientation, the genome encodes six major
proteins: nucleocapsid (NP), phosphoprotein (P), matrix (M), fusion (F), haemagglutinin±
neuraminidase (HN) and RNA-dependent RNA polymerase (L) (Chambers et al., 1986;
Wilde et al., 1986). The HN and F glycoproteins are important for virus infectivity and
virulence, and either of these two proteins can induce protective immunity (Meulemans
et al., 1986; Nagy et al., 1991).
NDV-specific antibodies (Ab) have traditionally been monitored by the haemagglu-
tination inhibition test (Alexander, 1997). ELISA procedures based on whole virus as
coating antigen have been described (Miers et al., 1983; Snyder et al., 1983; Wilson et al.,
1984; Rivetz et al., 1985), and, more recently, commercial kits (e.g. IDEXX Laboratories,
Westbrook, ME) are widely being used in diagnostic laboratories. Errington et al. (1995)
recently reported on an NDV NP-based ELISA using NP expressed in a baculovirus
system and compared their NP ELISA-based scores with HI and the commercial IDEXX
ELISA scores. They did not observe any false positive readings for samples scored
positive by HI. However, they did report that among sera expected to be negative were
some that scored positive by IDEXX ELISA and negative by their NP-based system.
Numerous, live and inactivated virus vaccines exist against NDV. There has recently
been growing interest in using subunit vaccines against poultry diseases including ND
(Meulemans et al., 1988; Nishino et al., 1991; Morgan et al., 1993; Nagy et al., 1993).
Consequently, there is a need to be able to differentiate between birds naturally infected
with NDV and vaccinated with such a recombinant subunit in the surveillance of NDV.
In this study, we sought to develop an ELISA test for Ab detection that would
discriminate between the antibody response to a subunit vaccine (HN expressed by a
recombinant fowlpox virus) and the response to live and inactivated NDV. We cloned the
NP gene of the Hitchner B1 NDV strain and expressed the NP in a baculovirus vector for
use as a coating antigen in the ELISA. Serum samples were analysed from chickens
vaccinated with a recombinant fowlpox virus expressing the NDV±HN and subsequently
infected with NDV.
2. Materials and methods
2.1. Viruses and cells
NDV strains Hitchner B1 and LaSota were used (Nagy et al., 1990). Virus was propagated
in embryonated eggs and in chicken embryo lung (CELu) cells by standard procedures.
210 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222

Autographa californica nucleopolyhedrovirus (AcMNPV) and Spodoptera frugiperda
(Sf) 9, Sf21 and Trichoplusia ni High Five cells were from Invitrogen. Baculovirus and
insect cell techniques were performed as described by Summers and Smith (1987) and
O'Reilly et al. (1992).
Fig. 1. Construction of NP plasmid vector and baculovirus transfer vector.
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 211

2.2. Generation of recombinant baculovirus
The strategy for recombinant plasmid construction is outlined in Fig. 1. All molecular
biology techniques were based on Sambrook et al. (1989). Complementary DNA was
made (cDNA Synthesis System; Gibco-BRL, Burlington, ON) from genomic NDV RNA
using a primer (50-GCCTTCTGCCAAAATGTC-30; the initiation codon is in bold)
derived from the NP genes of d26 (Ishida et al., 1986) and Ulster 2C (GenBank accession
#Z300084). Blunt-ended cDNA was ligated into the SmaI site of pGEM-7Zf(�) to
generate pNP103, with a 2.5 kb insert containing the NDV NP gene. A PCR product
containing just the NP gene was generated using the same primer used for cDNA
synthesis, and primer B (50-TGTTGTTGGTCAGTACCC-30) with the complement of the
termination codon (in bold) and based on the NP gene of the Ulster 2C strain. The PCR
product was blunt-end ligated into the SmaI site of pGEM-7Zf(�) to generate
pNP103.22. The 1.5 kb EcoRI/Csp45I fragment from pNP103.22 was blunt-ended with
Klenow enzyme and NheI linkers (New England Biolabs, Beverly, MA) were added for
cloning into the NheI site of the baculovirus pETL vector (Richardson et al., 1992) to
generate pEN159 which contained the NP gene under the control of the polyhedrin (polh)
promoter. Recombinant baculovirus was generated as described by Summers and Smith
(1987) and purified by three rounds of plaque purification to generate one recombinant
referred to as rBNP.
2.3. Sequencing
The NP gene in pNP103 was sequenced at the Guelph Molecular Supercentre
(Laboratory Services Division, University of Guelph) and sequence data were analysed
with `Align' (version 1.02, #1989 Scientific and Education Software). Sequences were
deposited into GenBank, accession number AF060483.
2.4. Polyacrylamide gel electrophoresis and Western blot analysis
Sf9 cells infected with rBNP at a multiplicity of infection (m.o.i.) of 10 were analysed
by sodium dodecyl sulfate 10% polyacrylamide gel electrophoresis (SDS-PAGE;
Laemmli, 1970), and Western blotting. Infected cells, harvested at different times post-
infection (p.i.), were lysed in an electrophoretic sample buffer (ESB; 62.5 mM Tris [pH
6.8], 10% glycerol, 2% SDS, 5% b-mercaptoethanol, 0.002% bromophenol blue).
Western blotting onto nitrocellulose membrane with a 0.45 mm pore size (Schleicher and
Schuell, Keene, NH) was as described by Towbin et al. (1979). Blots were blocked in 5%
skim milk powder (SMP; Difco, Detroit, MI) in PBS for 3 h at room temperature. NDV-
specific rabbit polyclonal serum at a dilution of 1/2000 in 2% SMP in PBS±Tween
(0.05% Tween-20 in PBS) was added to the blots and incubated at room temperature for
2 h. Blots were then incubated with goat anti-rabbit alkaline phosphatase-conjugated Ab
(Bio-Rad, Mississauga, ON) at a dilution of 1/1000 in 2% SMP in PBS and developed in
a 1 : 1 : 200 (by volume) solution of BCIP (Bio-Rad; 15 mg/ml stock in N,N-
dimethylformamide), NBT (Bio-Rad; 30 mg/ml stock in 70% N,N-dimethylformamide)
in 0.1 M Tris (pH 9.5), 0.5 mM MgCl2.
212 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222

2.5. Radioimmunoprecipitation
Sf9 cells infected with rBNP were incubated at 48 h p.i. in methionine-free Grace's
medium with [35S] methionine at 100 mCi/ml (ICN; specific activity 1175 Ci/mmol) for
4 h at 288C. Cells were lysed in RIPA buffer (150 mM NaCl, 10 mM Tris [pH 7.2], 1%
sodium deoxycholate, 1% Triton X-100, 0.1% SDS). NDV-infected chicken CELu were
similarly labelled in methionine-free EMEM at 8 h p.i. for 2 h at 378C. Immunopreci-
pitation with NDV-specific rabbit serum and analysis was as described by Nagy et al.
(1990).
2.6. Antigen preparation and conditions for ELISA
The rBNP-infected Sf21 cell lysates were used as antigens for ELISA and the
appropriate conditions were determined by a series of checkerboard plates as described
by Carpenter (1992). Cells infected at an m.o.i. of 5 and harvested at 40 h p.i. were
washed twice, resuspended in PBS, and sonicated over a 1.5 min period on ice.
Concentrations of soluble protein were determined with a Bio-Rad protein assay kit.
ELISA plates (Becton Dickinson Laboratories, Lincoln Park, NJ) were coated at 2.5 mg
total protein/well in 0.5 M carbonate±bicarbonate buffer (pH 9.6), and blocked with 5%
BSA (Fraktion V, Boehringer Mannheim) for 1 h at 378C. Plates were incubated at 378Cfor 1.5 h with primary Ab diluted 1/500 in PBS±Tween. Plates were next incubated at
378C for 1.5 h with goat anti-chicken IgG alkaline phosphatase conjugate (Kirkegaard
and Perry, Gaithersburg, MD) diluted 1/1000 in PBS±Tween. Plates were developed with
Nitrophenyl±Phosphate (Sigma, p-NPP tablets; Sigma, Oakville, ON) and OD405 was
read in a Bio-Tek microplate autoreader.
Samples considered positive for NDV were from the Animal Health Laboratory
(University of Guelph) as a collection of serum samples from individual chickens from
flocks which tested positive for NDV Ab. For flock testing, random samples of five birds
per flock were tested with the commercial IDEXX kit. The individual samples were
divided into 48 pooled serum samples (five individuals per pooled sample) for testing
with our NP ELISA. Samples considered negative for NDV were from 58 birds reared in
isolation and confirmed to be negative by IDEXX ELISA.
Specificity and sensitivity of the ELISA were calculated according to Smith (1995).
Sensitivity was the number of true positive samples divided by the sum of the true
positive and false negative samples, and the specificity was the number of true negative
samples divided by the sum of the true negative and false positive samples. The true
positive and negative samples were those scored as positive or negative, respectively, by
both IDEXX and NP ELISA. False positives were those that scored negative by IDEXX
but positive by NP ELISA while false negatives were those that scored positive by
IDEXX but negative by NP ELISA. An OD405 cut-off value for the NP ELISA to
differentiate positive from negative samples was set at two standard deviations above the
average OD405 from IDEXX-negative samples. Values above this cutoff were scored as
positive.
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 213

2.7. Vaccination experiment
Three-week-old Leghorn chickens (Arkell Research Station, University of Guelph)
were separated into three groups of five birds each. Group 1 and Group 2 chickens were
vaccinated with a recombinant fowlpox virus expressing the HN of NDV with the
vaccinia virus promoter P7.5 (rFPV7.5) or P11 (rFPV11), respectively, at Weeks 0 and 2.
Group 3 chickens were mock-vaccinated. At Week 4 all birds were infected with 0.1 ml
of NDV (LaSota strain) via the intranasal route. Weekly blood samples were collected
beginning with a pre-immune bleed at Week 0, and ending with post-infection samples at
Week 5. Serum samples were tested using recNP ELISA and whole NDV ELISA. Plates
for whole NDV ELISA were coated with a hundred-fold dilution of NDV-infected
allantoic fluid (initial HA 1 : 256) at 100 ml/well.
3. Results
3.1. Sequence analysis of the NP gene
The open reading frame of the NP gene of the Hitchner B1 NDV strain was 1,467
nucleotides (nt) long and coded for a 489 aa protein with a predicted molecular mass (Mr)
of 53.03 kDa. The number of amino acids for the B1 strain NP gene was identical to that
from d26 and Ulster 2C. The nucleotide homology of the NP gene was 90.03 and 89.45%
with that of d26 (Ishida et al., 1986) and Ulster 2C (GenBank accession #Z30084),
respectively. Amino acid identities were 95.71 and 96.11%, respectively. The NP gene
sequences of the d26 and Ulster 2C strains showed nucleotide homology and amino acid
identity of 96.4 and 98.6%, respectively.
3.2. Expression of the recombinant NP protein in insect cells
Sf9 cells infected with rBNP were labelled with [35S] methionine and cell lysates were
immunoprecipitated (Fig. 2). A very prominent band was detected at 53 kDa (Lane 3) for
rBNP and the position of this band was identical to that of the NP band in NDV-infected
CELu cells (Lane 2). An NDV-specific band was also detected at 43 kDa for rBNP but a
corresponding band was not detected for NDV-infected CELu cells and may represent
degradation of the 53 kDa NP. Similarly, a 39 kDa protein band was detected in NDV-
infected CELu cells but not in the rBNP lanes. This represents the NDV matrix protein
that would co-precipitate with the anti-NDV Ab. No proteins of the uninfected CELu
(Lane 1) or Sf9 (Lane 5) cells reacted with the NDV-specific Ab. A single band at
approximately 30 kDa which corresponds to the position of the polyhedrin protein was
detected in the AcMNPV-infected Sf9 cell lysates treated with either anti-NDV or pre-
immune sera (Lanes 4 and 7, respectively). Polyhedrin is present as a protein complex
which does not solubilize in cell lysis buffer and pellets with immunoprecipitated proteins
(O'Reilly et al., 1992).
In a time course of expression of the recNP in Sf9 cells infected with rBNP at an m.o.i.
of 10, a Coomassie blue stained band corresponding in size to recNP (53 kDa) was seen
214 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222

by 36 h p.i. (arrow) and increased in intensity up to 48 h p.i., and remained constant
thereafter (Fig. 3, panel A). By Western blotting an NDV-specific protein band
corresponding to the unique 53 kDa band in the stained gel, was first detected at 24 h
p.i. (arrow) and increased in intensity over time (Fig. 3, panel B). Some bands of lower
molecular weight appeared at 36 h p.i. and increased in amount with time. The NP of
NDV grown in embryonated eggs, migrated at 55 kDa.
NP expression was compared in Sf9, Sf21 and High Five cell lines, at m.o.i. of 0, 1, 5
and 10 and harvested at 24, 36, 48 and at 60 h p.i. By Coomassie blue staining, recNP
expression in Sf9 and Sf21 cells was similar at all time points and m.o.i., but expression
in High Five cells was extremely low. Based on densitometric analysis of the gels, Sf9
and Sf21 cells infected with an m.o.i. of 5 and collected at 48 h p.i. contained
approximately 0.60 and 0.53%, respectively, of recNP relative to the total amount of
protein in the lane. RecNP expression was greater in Sf21 cells than in Sf9 cells by
Western blot analysis (data not shown). Since optimal production of recNP occurred in
Fig. 2. Radioimmunoprecipitation of rBNP-infected [35S] methionine-labelled Sf9 cell lysates. Lysates in Lanes
1 to 5 were precipitated with NDV-specific Ab, in Lanes 6 and 7 were precipitated with pre-immune serum.
Lane 1: uninfected CELu cells; Lane 2: NDV infected CELu cells; Lanes 3 and 6: rBNP infected cells; Lanes 4
and 7: AcMNPV-infected cells; and Lane 5: uninfected Sf9 cells. M: radiolabelled molecular weight marker.
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 215

Sf21 cells infected at an m.o.i. of 5 and harvested between 36 and 48 h p.i. these
conditions were used for production of recNP for subsequent ELISA studies.
3.3. Development of recombinant NP-based ELISA
Appropriate conditions of the recNP-based ELISA were determined through a series of
checkerboard ELISAs testing antigen concentrations, primary and secondary Ab
dilutions, and use of different blocking agents. The final conditions, as described in
Section 2, were then used to screen a number of known positive and negative samples.
The specificity and sensitivity of the ELISA were evaluated from a panel of 48 IDEXX-
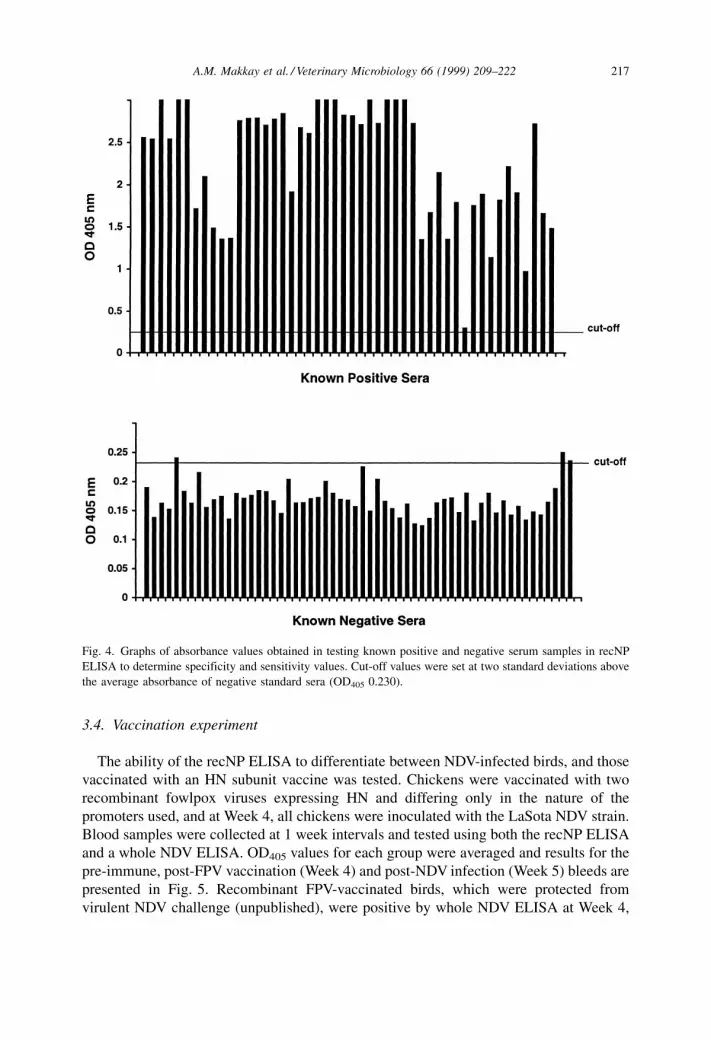
positive and 58 IDEXX-negative serum samples (Fig. 4). Of the samples tested, the
recNP ELISA gave a total of 48 true positive, 55 true negative, three false positive, and no
false negative readings (i.e. 48 of the 48 positive sera were scored as positive and 55 of
the 58 negative sera were scored as negative using the recNP ELISA). The OD405 values
for the three false positives were just above the cut-off OD405 value. The specificity and
sensitivity of the recNP ELISA were 94.8 and 100%, respectively. Raising the OD405 cut-
off value to three standard deviations above the average for the negative values (0.251)
would result in 48 true positive samples and 58 true negative samples for 100%
specificity and 100% sensitivity.
Fig. 3. Time course analysis of recNP expression by SDS-PAGE and Western blotting. A: Coomassie blue
stained gel, and B: Western blot analysis with NDV-specific Ab. The numbers above the lanes indicate rBNP-
infected cells harvested at 12±72 h p.i. AcMNPV-infected (wt) and uninfected (un) cell lysates, and purified
NDV (NDV) were also run. M: molecular weight markers, in kDa. Arrows show recNP band.
216 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222
3.4. Vaccination experiment
The ability of the recNP ELISA to differentiate between NDV-infected birds, and those
vaccinated with an HN subunit vaccine was tested. Chickens were vaccinated with two
recombinant fowlpox viruses expressing HN and differing only in the nature of the
promoters used, and at Week 4, all chickens were inoculated with the LaSota NDV strain.
Blood samples were collected at 1 week intervals and tested using both the recNP ELISA
and a whole NDV ELISA. OD405 values for each group were averaged and results for the
pre-immune, post-FPV vaccination (Week 4) and post-NDV infection (Week 5) bleeds are
presented in Fig. 5. Recombinant FPV-vaccinated birds, which were protected from
virulent NDV challenge (unpublished), were positive by whole NDV ELISA at Week 4,
Fig. 4. Graphs of absorbance values obtained in testing known positive and negative serum samples in recNP
ELISA to determine specificity and sensitivity values. Cut-off values were set at two standard deviations above
the average absorbance of negative standard sera (OD405 0.230).
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 217

but were negative by recNP ELISA. These recombinant FPV-vaccinated birds would
produce antibodies only to HN of NDV and consequently react with only this NDV
polypeptide in the wells. The resulting OD405 values would consequently be lower than
that for NDV-inoculated birds whose antibodies would react against all NDV
polypeptides. Even though the OD405 values for the vaccinated birds were close to the
cut-off values they were clearly above those of the pre-immune sera. Mock-vaccinated
birds remained negative throughout Weeks 1±4 in both ELISAs. Serum samples taken at
Week 5, 1 week after live NDV inoculation, were positive in both tests for all birds.
4. Discussion
The NP of NDV was expressed in a baculovirus expression vector system and was used
as a coating antigen for a diagnostic and differential ELISA.
Fig. 5. Histograms of average OD405 values in ELISAs of serum samples from chickens vaccinated with
recombinant fowlpox virus expressing the haemagglutinin±neuraminidase of NDV and subsequently infected
with NDV. Panel A shows the results of the whole NDV ELISA, and panel B shows the results of the same serum
samples in the recNP ELISA. Average values per group are shown for Week 0 (pre-immune), Week 4 (post-
vaccination) and Week 5 (post-infection).
218 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222

The NP gene of the Hitchner B1 strain was sequenced and compared to those of strains
d26 and Ulster 2C (Ishida et al., 1986; GenBank accession #Z30084), the only complete
NP sequences of any NDV strains reported to date. The predicted molecular mass of the
B1 NP according to sequence information was 53,029 Da which is in excellent agreement
with the size of the virion NP and with the expressed NP determined by PAGE. The B1
NP amino acid sequence showed a high degree of identity to those of Ulster 2C and d26
(95.71 and 96.11%, respectively). Since there is a very high degree of amino acid
sequence conservation, the recNP ELISA based on the B1 strain should be effective
against all NDV strains. The recombinant NP expressed in insect cells was detected as a
single, Coomassie blue-stained band at approximately 53 kDa in size. The same protein
was shown to be NDV-specific by Western blotting and radioimmunoprecipitation with
rabbit polyclonal serum raised against purified NDV (Figs. 2 and 3, panel B). The
recombinant NP of 53 kDa migrated at a size comparable to that of NP (55 kDa) from
virus grown in embryonated eggs (Smith and Hightower, 1981; Ishida et al., 1986;
Samson, 1988). The slight discrepancy in size between the recNP (53 kDa) and the virion
NP (53 or 55 kDa) in Fig. 3 might reflect slightly different forms of the NP proteins from
NDV grown under different conditions as has previously been described (Smith and
Hightower, 1981). For radioimmunoprecipitation (Fig. 2), CELu cells infected with NDV
were used to compare with the rBNP-infected Sf9 cells. Both these samples showed NP
bands of the same mobility, estimated at 53 kDa. In addition to the major recNP band
several faster migrating bands were seen in Western blots and radioimmunoprecipitation
of rBNP-infected cell lysates. In time course experiments, these bands appeared only
after the major recNP band was seen, and they accumulated over time. These lower
molecular weight proteins may be due to proteolytic degradation of the recombinant NP,
since the NP of NDV is susceptible to such degradation (Mountcastle et al., 1970). This is
also common for other paramyxoviruses including simian virus 5 (Mountcastle et al.,
1974), measles virus (Rozenblatt et al., 1979) and canine distemper virus (Hall et al.,
1980). Lower molecular weight species were also obtained by Kamata et al. (1993) when
the rinderpest virus nucleocapsid protein was expressed using baculovirus. The 53 and
43 kDa bands present in the radioimmunoprecipitated rBNP-infected cell sample could
represent the undigested and digested forms of NP, respectively, as described by
Mountcastle et al. (1974).
Initial ELISA experiments showed no cross-reactivity of chicken sera with uninfected
and AcMNPV-infected Sf21 cell lysates. ELISA plates were therefore coated with cell
lysates from rBNP-infected cells.
Baculovirus-expressed nucleocapsid proteins have been used as antigens in ELISAs for
measles virus (Hummel et al., 1992), rinderpest virus (Kamata et al., 1993), vesicular
stomatitis virus (Ahmad et al., 1993) and NDV (Errington et al., 1995). In comparison to
commercially available ELISAs, there was a better correlation of the recombinant NP
ELISAs with neutralising antibody levels than with commercial ELISAs (Hummel et al.,
1992; Errington et al., 1995).
The recNP-based ELISA was developed and optimised, and positive and negative sera
were tested. Since pooled samples are commonly used to determine overall flock
immunity, use of pooled samples for positive sera in determining the specificity and
sensitivity of this test was considered appropriate. Specificity and sensitivity of the test
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 219

were 94.8 and 100%, respectively, and no false negative readings were observed (Fig. 4).
Since IDEXX itself might not be 100% accurate this might lead to some lack of
correlation with the recNP ELISA. The few false positive samples were very close to the
cut-off values determined, and thus may be an artifact of a cutoff set too low. The cut-off
value used in the recNP ELISA was two standard deviations above the average OD405 of
the negative standards. If the cutoff was three standard deviations above the average
negative value, the three false positives would be true negatives. Since the sensitivity and
specificity of this recNP ELISA are high we feel that it is a reliable test for determining
the presence of NDV-specific antibodies.
Errington et al. (1995) earlier described a baculovirus-expressed NP-based ELISA for
NDV Ab. They did not report any false positive NP ELISA readings (for samples positive
by HI) although they reported some false negatives relative to some IDEXX-positive
samples which they felt may have been due to PMV-2 or PMV-3 interference. In our
recNP-based ELISA we did not observe any false negative samples, even using the
IDEXX ELISA kit which is more sensitive than HI. One of the differences in the
protocols of the two NP-based ELISAs was the cut-off values set; in our study we did not
use an S/P ratio as described by Errington et al. (1995). An S/P ratio is the ratio of the
unknown value to that of a weak positive control, after the subtraction of the negative
control value from both. It is possible that incorporating weak positive standards and an
S/P ratio cutoff into the recNP ELISA described herein could eliminate the false positives
and thus increase the specificity of our test to 100%. Thus our test is potentially more
reliable than that of Errington et al. (1995) for the detection of NDV Ab. Furthermore, we
showed that the recNP described can be used to differentiate between birds exposed to
NDV in either a whole NDV vaccine or by natural infection, from those vaccinated with a
subunit vaccine not containing the NP.
Presently, there is a great deal of interest in the development of recombinant and
protein subunit vaccines. For NDV, protective antibodies are induced against either
the HN or F proteins. The recNP ELISA described here could be used to dif-
ferentiate between birds naturally infected with NDV from those immunised with
HN or F protein subunit vaccines since antibody against the NP would be present
only in birds exposed to whole virus. To evaluate this, sera from chickens which
were vaccinated with recombinant fowlpox viruses expressing the HN only of NDV,
were tested. Chickens immunised with either recombinant FPV construct were
weakly positive using whole NDV ELISA after two vaccinations, whereas using recNP
ELISA they were negative (Fig. 5). Unvaccinated chickens were negative in both
ELISAs. All chickens, after receiving an intranasal inoculation of live NDV, became
strong positives according to both ELISAs. The OD405 values of the whole NDV ELISA
after vaccination with the FPV-HN were considerably lower than after exposure to
NDV, but were still positive for all birds within a group and significantly higher than
that in pre-immune sera. These indicate that a positive result in a whole NDV ELISA
can be due to antibodies to HN alone and that the recNP ELISA can be used to
differentiate between subunit-vaccinated birds and those exposed to whole virus. Most
importantly, a positive result with a recNP-based ELISA would indicate immunisation
with a whole NDV vaccine or/and infection with NDV and excludes birds immunised
with only a subunit vaccine.
220 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222

The feasibility of using baculovirus-expressed nucleocapsid protein as an antigen for
an ELISA to differentiate between vaccinated and infected animals was also explored by
Ahmad et al. (1993) for vesicular stomatitis virus, and Kamata et al. (1993) for rinderpest
virus. As in our study, both groups showed that a baculovirus-expressed recombinant NP-
based ELISA was capable of differentiating subunit-vaccinated animals from those
infected with whole virus.
Acknowledgements
This work was funded by the Natural Sciences and Engineering Research Council
(NSERC) of Canada. We thank Paul Huber for his photographic and technical expertise
and Sean Walter for his technical assistance. We are indebted to Dr. D. Key (Animal
Health Laboratory, University of Guelph) for providing the pre-tested chicken serum
samples.
References
Ahmad, S., Bassiri, M., Banerjee, A.K., Yilma, T., 1993. Immunological characterization of the VSV
nucleocapsid (N) protein expressed by recombinant baculovirus in Spodoptera exigua larva: Use in
differential diagnosis between vaccinated and infected animals. Virology 192, 207±216.
Alexander, D.J., 1997. Newcastle disease and other avian Paramyxoviridae infections. In: Calnek, B.W., Barnes,
H.J., Beard, C.W., McDougald, L.R., Saif Y.M. (Eds.), Diseases of Poultry, 10th edn., Iowa State University
Press, Ames, IA, pp. 541±569.
Carpenter, A.B., 1992. Enzyme-linked immunoassays. In: Rose, N.E., de Macario, E.C., Fahey, J.L., Friedman,
H., Penn, G.M. (Eds.), Manual of Clinical Laboratory Immunology, 4th edn., American Society for
Microbiology. pp. 2±9.
Chambers, P., Millar, N.S., Bingham, R.W., Emmerson, P.T., 1986. Molecular cloning of complementary DNA
to Newcastle disease virus, and the nucleotide sequence analysis of the junction between the genes
encoding the haemagglutinin±neuraminidase and the large protein. J. Gen. Virol. 67, 475±486.
Errington, W., Steward, M., Emmerson, P.T., 1995. A diagnostic immunoassay for Newcastle disease virus based
on the nucleocapsid protein expressed by a recombinant baculovirus. J. Virol. Meth. 55, 357±365.
Hall, W.W., Lamb, R.A., Choppin, P.W., 1980. The polypeptides of canine distemper virus: Synthesis in infected
cells and relatedness to the polypeptides of other morbilliviruses. Virology 100, 433±449.
Hummel, K.B., Erman, D.D., Heath, J., Bellini, W.J., 1992. Baculovirus expression of the nucleocapsid protein
gene of measles virus and utility of the recombinant protein in diagnostic enzyme immunoassays. J. Clin.
Microbiol. 30, 2874±2880.
Ishida, N., Taira, H., Omata, T., Mizumoto, K., Hattori, S., Iwasaki, K., Kawakita, M., 1986. Sequence of 2,617
nucleotides from the 30 end of Newcastle disease virus genome RNA and the predicted amino acid sequence
of viral NP protein. Nucl. Acids. Res. 14, 6551±6564.
Kamata, H., Ohkubo, S., Sugiyama, M., Matsuura, Y., Kamata, Y., Tsukiyama-Kohara, K., Imaoka, K., Kai, C.,
Yoshikawa, Y., Yamanouchi, K., 1993. Expression in baculovirus vector system of the nucleocapsid protein
gene of rinderpest virus. J. Virol. Meth. 43, 159±166.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
Nature 227, 680±685.
Meulemans, G., Gonze, M., Carlier, M.C., Petit, P., Burny, A., Long, L., 1986. Protective effects of HN and F
glycoprotein-specific monoclonal antibodies on experimental Newcastle disease. Avian Path. 15, 761±768.
Meulemans, G., Letellier, C., Gonze, M., Carlier, M.C., Burny, A., 1988. Newcastle disease virus glycoprotein
expressed from a recombinant vaccinia virus vector protects chickens against live-virus challenge. Avian
Path. 17, 821±827.
A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222 221

Miers, L.A., Bankowski, R.A., Zee, Y.C., 1983. Optimizing the enzyme-linked immunosorbent assay for
evaluating the immunity of chickens to Newcastle disease. Avian Dis. 27, 1112±1125.
Morgan, R.W., Gelb Jr., J., Pope, C.R., Sondermeijer, P.J.A., 1993. Efficacy in chickens of a herpesvirus of
turkeys recombinant vaccine containing the fusion gene of Newcastle disease virus: Onset of protection and
effect of maternal antibodies. Avian Dis. 37, 1032±1040.
Mountcastle, W.E., Compans, R.E., Caliguiri, L.A., Choppin, P.W., 1970. Nucleocapsid protein subunits of
simian virus 5, Newcastle disease virus, and Sendai virus. J. Virol. 6, 677±684.
Mountcastle, W.E., Compans, R.E., Lackland, H., Choppin, P.W., 1974. Proteolytic cleavage subunits of the
nucleocapsid of paramyxovirus SV5. J. Virol. 14, 1253±1261.
Nagy, EÂ ., Derbyshire, J.B., Dobos, P., Krell, P.J., 1990. Cloning and expression of NDV hemagglutinin±
neuraminidase cDNA in a baculovirus expression vector system. Virology 176, 426±438.
Nagy, EÂ ., Krell, P.J., Dulac, G.C., Derbyshire, J.B., 1991. Vaccination against Newcastle disease with a
recombinant baculovirus hemagglutinin±neuraminidase subunit vaccine. Avian Dis. 35, 585±590.
Nagy, EÂ ., Krell, P.J., Heckert, R.A., Derbyshire, J.B., 1993. Vaccination of chickens with a recombinant fowlpox
virus containing the hemagglutinin±neuraminidase gene of Newcastle disease virus under the control of the
fowlpox virus thymidine kinase promoter. Can. J. Vet. Res. 57, 306±308.
Nishino, Y., Niikura, M., Suwa, T., Onuma, M., Gotoh, B., Nagai, Y., Mikami, T., 1991. Analysis of the
protective effect of the haemagglutinin±neuraminidase protein in Newcastle disease virus infection. J. Gen.
Virol. 72, 1187±1190.
O'Reilly, D.R., Miller, L.K., Luckow, V.A., 1992. Baculovirus Expression Vectors: A Laboratory Manual, W.H.
Freeman, New York.
Richardson, C.D., Banville, M., LalumieÁre, M., Vialard, J., Meighen, E.A., 1992. Bacterial luciferase produced
with rapid-screening baculovirus vectors is a sensitive reporter for infection of insect cells and larvae.
Intervirology 34, 213±227.
Rima, B., Alexander, D.J., Billeter, M.A., Collins, P.L., Kinsbury, D.W., Lipkind, M.A., Nagai, Y., OÈ rvell, C.,
Pringle, C.R., ter Meulen, V., 1995. Paramyxoviridae. In: Murphy, F.A., Fauquet, C.M., Bishop, D.H.L.,
Ghabrial, S.A., Jarvis, A.W., Martelli, G.P., Mayo, M.A., Summers, M.D. (Eds.), Virus Taxonomy:
Classification and Nomenclature of Viruses, 6th Report of the International Committee on Taxonomy of
Viruses, Springer, New York, pp. 268±274.
Rivetz, B., Weissman, Y., Ritterband, M., Fish, F., Herzberg, M., 1985. Evaluation of a novel rapid kit for the
visual detection of Newcastle disease virus antibodies. Avian Dis. 29, 929±942.
Rozenblatt, S., Gorecki, M., Shure, M., Prives, C.L., 1979. Characterization of measles virus-specific proteins
synthesized in vivo and in vitro from acutely and persistently infected cells. J. Virol. 29, 1099±1106.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A Laboratory Manual, Cold Spring Harbour
Laboratory, Cold Spring Harbour, New York.
Samson, A.C.R., 1988. Virus structure. In: Alexander, D.J. (Ed.), Paramyxoviridae, Kluwer Academic
Publishers, Hingham, MA, pp. 23±44.
Smith, R.D., 1995. Veterinary Epidemiology: A Problem Oriented Approach, 2nd edn., CRC Press, Boca Raton,
FL, pp. 31±52.
Smith, G.W., Hightower, L.E., 1981. Identification of the P protein and other disulfide-linked and
phosphorylated proteins of Newcastle disease virus. J. Virol. 37, 256±267.
Snyder, D.B., Marquardt, W.W., Mallinson, E.T., Russek, E., 1983. Rapid serological profiling by enzyme-
linked immunosorbent assay. I. Measurement of antibody activity titer against Newcastle disease virus in a
single serum dilution. Avian Dis. 27, 161±170.
Summers, M.D., Smith, G.E., 1987. A Manual of Methods for Baculovirus Vectors and Insect Cell Culture
Procedures, Texas Agricultural Experiment Station Bulletin No. 1555, Texas A&M University, College
Station, TX.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. U.S.A. 76, 4350±4354.
Wilde, A., McQuain, C., Morrison, T., 1986. Identification of the sequence content of four polycistronic
transcripts synthesized in Newcastle disease virus infected cell. Virus Res. 5, 77±95.
Wilson, R.A., Perrotta Jr., C., Frey, B., Eckroade, R.J., 1984. An enzyme-linked immunosorbent assay that
measures protective antibody levels to Newcastle disease virus in chickens. Avian Dis. 28, 1079±1085.
222 A.M. Makkay et al. / Veterinary Microbiology 66 (1999) 209±222





























