Embed Size (px)
Citation preview

Augmentation of Type-1 Polarizing Ability ofMonocyte-Derived Dendritic Cells from Chronically
Immunosuppressed Organ-Transplant Recipients
Camila Macedo,1,2 Iulia Popescu,1,2 Kareem Abu-Elmagd,1,2 Jorge Reyes,1,2 Ron Shapiro,1,2
Adriana Zeevi,1,2,3 Jan Mueller Berghaus,2,4 Lian Fu Wang,1,2 Lina Lu,1,2 Angus W. Thomson,1,2,5
Walter J. Storkus,2,4,5 John J. Fung,1,2 and Diana Metes1,2,5,6
Background. Chronic immunosuppressive (IS) therapy impairs normal T-cell immune surveillance and may predis-pose to opportunistic infections and malignancies that represent life-threatening complication of solid-organ trans-plantation (SOTx). Our study was designed to ascertain the impact of chronic in vivo administration of IS on the abilityof monocyte-derived dendritic cells (MoDC) to differentiate, mature, and function ex vivo. The potential of these cellsto be implemented for DC-based adoptive immunotherapy was also considered.Methods. MoDCs were propagated by conventional procedures, their phenotype was analyzed by flow cytometry, andtheir function was assessed by mixed leukocyte reaction, enzyme-linked immunoadsorbent assay, and ELISPOT assays.Nuclear translocation of nuclear factor (NF)-kB was analyzed by electrophoretic mobility shift assay.Results. Circulating DC1s in peripheral blood were reduced in SOTx patients. MoDCs generated from patients dis-played higher endocytic activity versus normal DCs, indicating their comparative immaturity. Patients’ DCs exposed topro-inflammatory cytokines (tumor necrosis factor-�, interleukin [IL]-1�, and IL-6) were less able to mature, tostimulate recall antigen (Ag)- or allo–Ag-induced proliferation responses, or to secrete IL-12p70. These deficiencieswere associated with a decrease in NF-kB translocation. In contrast, combination of pro-inflammatory cytokines andinterferon (IFN)-� (a Th1-polarizing factor) augmented patients’ DC1-type function and IL-12p70 production by wayof an NF– kB-independent mechanism.Conclusions. Chronic IS restrains DC differentiation, maturation, and function at a transcriptional level; however,type-1 polarizing potential of patients’ DC1 can be augmented ex vivo by a two-signal stimulation provided bypro-inflammatory cytokines and IFN-�. These results may have implications for DC-based immunotherapy of malig-nancies in the transplantation setting.
Keywords: Solid organ transplantation, Immunosuppressive drugs, Human, Dendritic cells, Cytokines.
(Transplantation 2005;79: 451–459)
Dendritic cells (DCs) are uniquely well-equipped pro-fessional antigen-presenting cells (APCs) that activate
“naı̈ve” T cells and boost “memory” T-cell responses (1).Mature DCs express high levels of surface major histocom-patibility complex (MHC) class I and II and costimulatoryand adhesion molecules required for effective T-cell acti-vation. Several human and mouse DC subsets have beencharacterized that display distinct phenotypes and func-tions (2). Depending on the type of antigen (Ag) pre-sented, the cytokine environment, and the level and natureof costimulatory molecule expression, DCs can induce im-munity or exhibit tolerogenic properties. For example,
fully mature DC1 produce interleukin (IL)-12p70, apivotal cytokine that selectively promotes Th1-type im-mune responses (3). However, in their immature state, thesame DCs may predominantly secrete IL-10, exerting reg-ulatory properties that promote peripheral T-cell toler-ance (4, 5). Thus, DCs are promising tools for immuno-therapy of malignancies, infectious and autoimmunediseases, and prevention of transplant rejection.
The standard immunosuppression (IS) drug regimensnecessary to prevent graft rejection include a combination ofcalcineurin inhibitors and steroids, with or without adjunctiveantiproliferative agents (6, 7). The long-term administration ofthese agents can lead to unwanted complications that includedrug toxicity, opportunistic infections, and malignancies. Post-transplant lymphoproliferative disorders (PTLD), such as Ep-stein-Barr virus (EBV)-associated lymphomas, have high ratesof morbidity and mortality (8). Although numerous therapeuticapproaches have been used to treat PTLD, approximately 20%of cases remain refractory to currently available treatments (9–11). Restoration of anti-EBV T-cell immunity, by infusion of exvivo expanded EBV-specific cytotoxic T lymphocytes (CTLs),has been shown to be a safe and effective approach to prevent ortreat PTLD in bone–marrow-transplant recipients (12, 13). Es-tablishment of DC-based adoptive immunotherapy protocols totreat refractory PTLD may provide a valuable alternative thera-peutic approach in the solid-organ transplantation (SOTx) set-ting (14, 15).
This work was supported by grants from the American Cancer Society andthe American Heart Association (DM).
1 The Thomas E. Starzl Transplantation Institute, University of Pittsburgh,Pittsburgh PA.
2 Department of Surgery, University of Pittsburgh, Pittsburgh PA.3 Department of Pathology, University of Pittsburgh, Pittsburgh PA.4 University of Pittsburgh Cancer Institute, University of Pittsburgh, Pitts-
burgh PA.5 Department of Immunology, University of Pittsburgh, Pittsburgh PA.6 Address Correspondence to: Dr. Diana Metes, Section of Cellular Trans-
plantation, Thomas E. Starzl Transplantation Institute, E1551 Biomed-ical Science Tower, 200 Lothrop Street, Pittsburgh, PA 15261. E-mail:[email protected].
Received 30 June 2004. Accepted 18 August 2004.Copyright © 2005 by Lippincott Williams & WilkinsISSN 0041-1337/05/7904-451DOI: 10.1097/01.TP.0000146589.49756.7F
Transplantation • Volume 79, Number 4, February 27, 2005 451

Despite considerable in vitro experiments and animaldata, no information is currently available on the phenotypeand function of monocyte-derived dendritic cells (MoDC)generated from IS SOTx patients. Here, we examine the im-pact of prednisone and tacrolimus (FK506) on ex vivo MoDCdifferentiation, maturation, and function. The ability of pro-inflammatory cytokines (tumor necrosis factor [TNF]-�, IL-1�, and IL-6) to reverse the potential “tolerogenic” state of ISpatients’ MoDCs and to induce molecular pathways that im-prove DC1 function has been advocated (16). In addition,interferon (IFN)-� has been shown to synergize with inflam-matory cytokines in boosting uncommitted immature DC(iDC) progenitors to become DC1s with strong Th-1 polar-izing capacity (high production of IL-12p70) (17). These cy-tokines are known to enhance the nuclear translocation ofnuclear factor (NF)-kB in a variety of cell types and to coregu-late and activate genes encoding for cell-surface receptors andsoluble mediators (18, 19). Such approaches could improvethe immunogenicity of Ags presented by DC1s when appliedin ex vivo generated vaccines (20, 21).
Our results show that although SOTx patients’ DCs aretargeted by chronic IS therapy both in vivo and ex vivo, theseAPCs can be type-1 polarized in short-term cultures by two-signal cytokine stimulation, making DC-based adoptive im-munotherapy for PTLD a tenable therapeutic option.
METHODS
Human SubjectsTwelve healthy adult volunteers and 12 IS SOTx pa-
tients were recruited after informed consent under an insti-tutional review board-approved protocol. All patients werestable at the time of blood donation without evidence of re-jection or signs of EBV (re)activation. All patients were onchronic administration of prednisone and FK-506 for morethan 3 years. Human leukocyte antigen (HLA) typing forHLA-A, -B, and -DR loci using molecular-based techniqueswas performed in the Tissue Typing Laboratory, University ofPittsburgh Medical Center. PBMCs were isolated from 30 to40 mL of heparinized blood by Ficoll-Hypaque density gradi-ent centrifugation, as described previously (22).
Media, Cell Lines, and ReagentsFicoll-Paque was obtained from Amersham Bio-
sciences AB (Uppsala, Sweden). AIM-V medium, RPMI-1640 medium, and fetal calf serum (FCS) were purchasedfrom GIBCO (Grand Island, NJ). Normal human antibody(AB) serum (NHS) was purchased from Nabi-Boca Raton(Miami, FL). CD40L-transfected J558L plasmacytoma cellswere a gift from Dr. P. Lane, University of Birmingham, Bir-mingham, UK. bovine serum albumin (BSA) and phytohe-magglutinin (PHA) were obtained from Sigma (St. Louis,MO). Recombinant (r), human (h), granulocyte- macro-phage colony-stimulating factor (rhGM-CSF) was a gift ofAmgen (Seattle, WA), whereas rhIL-4 was kindly provided byfrom the Schering-Plough Corporation (Kenilworth, NJ).rhTNF-�, rhIL-6, and rhIL-1� were purchased from R&DSystems Inc. (Minneapolis, MN). RhIFN-� was purchasedfrom PeproTech (Rocky Hill, NJ). DQ-BSA was obtainedfrom Molecular Probes (Eugene, OR) and tetanus toxoid(TTox) from Wyeth-Ayerst (Narietta, PA).
DC Generation and MaturationMonocytes were isolated by 1-hour plastic adherence
(23) at 37°C, in 5% CO2, and cultured for 6 days in 10% FCSAIM-V media supplemented with 1,000 U/mL rhGM-CSFand 1,000 U/mL rhIL-4 (also referred to as DC media). Whenspecified, prednisone (10�6M) or FK-506 (10�6M) wereadded at the beginning of the cultures. iDC were incubatedwith rhTNF-� (10 ng/mL), rhIL-6 (0.9 �g/mL), and rhIL-1�(10 ng/mL), also referred to as “cyt” mix, in the presence orabsence of rhIFN-� (1,000 U/mL) for an additional 24 hoursat 37°C, 5% CO2, to induce DC maturation (mDC) (16, 17).The monocyte-depleted peripheral blood mononuclear cells(PBMC) were always frozen until required for use in func-tional assays.
EndocytosisiDC-mediated endocytosis was measured as cellular
uptake and activation of DQ (green)-BSA (24) as monitoredby flow cytometry. Briefly, 0.2x105 iDCs/sample were incu-bated in 100 �L DC media, containing any of six differentdilutions of DQ-BSA for 1 hour at 37°C, 5% CO2. After incu-bation, the cells were washed using flow wash (phosphate-buffered saline 1% FCS, 0.02% NaN3), and the relative fluo-rescence of DCs was assessed by flow cytometry. As negativecontrols, DCs stained with DQ-BSA were incubated at 4°C for1 hour before analysis to prevent uptake and processing of theprobe.
Flow Cytometric AnalysisThe frequency of DC1s in peripheral blood was deter-
mined by four-color flow cytometric analysis of PBMC (25).The cells were stained with monoclonal (m)Abs: anti-CD45,anti-Lin markers (a cocktail of mAbs: anti-CD3, CD14,CD16, CD19, CD20, and CD56), anti-HLA-DR, and anti-CD11c (Becton Dickinson [BD]-PharMingen, San Jose, CA).For DC phenotype, aliquots of iDCs and mDCs were stainedusing fluorescein isothiocyanate (FITC)- or phycoerythrin-conjugated mAbs anti-CD86, anti-CD80, anti-CD40 (Ancell,Bayport, MN), anti-CD83, and anti-HLA-ABC (BeckmanCoulter, Miami, FL), anti-CD11c, anti-CD1a, anti-CD54,and anti-HLA-DR (BD PharMingen). Data acquisition wasperformed on a Coulter EPICSElite flow cytometer (CoulterHialeah, FL), with results expressed as percentages or in arbi-trary mean fluorescence intensity units of positive cells.
Allo-Ag and Recall Ag-Induced Mixed LeukocyteReaction
For allospecific mixed leukocyte reaction (MLR),0.1�106 responder PBMCs in RPMI-1640 containing 5% v/vheat-inactivated NHS were incubated at a 50:1 ratio with dif-ferent types of irradiated (3,000 rads) allogeneic DCs (iDC,mDC, or mDC�IFN�) in 96-well, round-bottomed platesfor 5 days. The number of HLA mismatches for responderversus stimulator in control and patient alloMLRs was notsignificantly different. For recall Ag-specific responses,0.1�106 PBMCs from normal volunteers or patients werestimulated using different types of irradiated autologous DCs(iDC, mDC, mDC�IFN-�) loaded with 16 pfU/mL TTox in96-well plates for 5 days. Over the last 18 hours of culture, 1�Ci [3H]thymidine (NEN Life Science Products Inc., Boston
452 Transplantation • Volume 79, Number 4, February 27, 2005

MA) was added to each well. Cells were harvested and testedfor [3H]thymidine uptake using a LKB liquid �-scintillationcounter. Results are reported as the mean�SD of cpm ob-tained from triplicate determinations.
Generation of T-Cell ClonesT-cell lines specific for TTox were generated from se-
lected healthy control subjects expressing HLA-A2 and HLA-DR15 alleles by incubating NALs with TTox (16 pfU/mL)preloaded autoDC at a ratio 20:1, for 1 week, with subsequentrestimulation 1 week later, followed by direct cloning. CD4�
and/or CD8� sorted T cells were seeded by limiting dilutionfor cloning, together with feeder cells and IL–2-conditionedmedia (26). A total of seven TTox-specific Tc0 CD8� andfour Th0 CD4� T-cell clones were generated from one con-trol subject as well as four TTox-specific Th1 CD4� T-cellclones from another control subject. The clones were catego-rized using IFN-�/IL-4 intracellular cytokine staining andflow cytometry. Two Tc0 CD8�, two Th0 CD4�, and twoTh1 CD4� T-cell clones were further tested in ELISPOT as-say (22) against TTox loaded iDCs, mDC (cyt), and mDCs(cyt�IFN-�).
ELISPOT Assay for Single-Cell IFN-� ReleaseNinety–six-well nitrocellulose backed plates (Milli-
pore, Bedford, MA) were precoated with anti-IFN-� mAb1-D1K (10 �g/mL Mabtech, Sweden). TTox-specific CD4�or CD8� T-cell clones were added to triplicate wells at0.1�106 with either autologous or HLA-A2/HLA-DR15-matched TTox preloaded DCs at 0.02�106 cells/well for 20hours, at 37°C in 5% CO2. The wells were washed free of cellsand soluble factors, and a second, biotinylated anti-IFN-�mAb 7-B6 –1 (2 �g/mL, Mabtech) was added for an addi-tional 2 hours at 37°C. The reaction was developed as de-scribed previously (22). Spots were counted using an ELIS-POT plate reader (Karl Zeiss, Jenna, Germany).
Enzyme-Linked Immunoadsorbent Assay forCytokine Determination
iDCs (0.5x106/mL) were incubated overnight in 24-well plates either alone or with the cytokine cocktail (TNF-�,IL-6, and IL-1�) �IFN-�. MoDCs were subsequently incu-bated for an additional 24 hours in the presence or absence ofCD40L stimulation (irradiated J558L cells) at 1:2.5 ratio. Su-pernatants were collected, and IL-12p70 and IL-10 quantifiedby enzyme-linked immunoadsorbent assay. Primary and sec-ondary mAbs for IL-12p70 and IL-10 cytokine detection, aswell as standard recombinant cytokines, were purchasedfrom Endogen (Woburn, MA).
Nuclear Extracts and Electrophoretic MobilityShift Assay
Nuclear proteins from each category of DCs (iDC,mDC, mDC�IFN-�) were extracted, and electrophoreticmobility shift assay was performed using a commercial kit(Promega, Madison, WI) supplied with a NF-�B oligonucle-otide probe (sense sequence: 5'-AGTTGAGGGGACTTTC-CCAGGC-3'), which was end-labeled with [�-32P]adenosinetriphosphate (NEN) (27). Nuclear proteins (1.5 �g) wereloaded in each lane, and mobility shift was detected by run-
ning the mixture on a 4% polyacrylamide gel. Shifted bandswere visualized by autoradiography (27).
Statistical AnalysisA paired, two-tail Student’s t test was used for statistical
analysis of mean�SD values between groups. Results wereconsidered significant for P�0.05.
RESULTS
Frequency of CD11c�DC (DC1 Precursors) inPeripheral Blood of IS SOTx Patients is ReducedCompared with Healthy Controls
To ascertain the influence of chronic IS on MoDC dif-ferentiation in vivo, we analyzed the percentage of DC1 infreshly isolated PBMC from SOTx patients compared withhealthy volunteers (2, 25). The gating approach is illustratedin Figure 1A. The percentage of CD45�Lin�HLA-DR� cells,considered to represent DC precursors (25), was not signifi-cantly different when comparing patients with normal do-nors (Fig. 1B). The percentage of DC1 (CD45�Lin�HLA-DR�CD11c�) cells in patient peripheral blood was, however,decreased significantly (P�0.02) when compared with nor-mal controls (Fig. 1B). When the proportion of Lin�HLA-DR�CD11c� cells was further normalized to the percentageof Lin�HLA-DR� DC precursors, we noted a significant re-duction in the relative proportion of DC1 among Lin�HLA-DR� cells in SOTx patients (mean 30%�7) compared withnormal controls (mean 42%�16) (P�0.04) (Fig. 1B).
Superior Yields of iDCs can be Generated fromIS SOTx Patients
After 6 days of culture, the iDC yield from SOTx pa-tients ranged from 6% to 30% of PBMC (mean of15�8.7�106 iDC per 108 PBMC), which was significantlyhigher (P�0.004) than the yields obtained from normal do-nors, which ranged from 1.7% to 12.7% of PBMC (mean of6.6�3.6�106 iDCs per 108 PBMC). No increase in the abso-lute number or percent of peripheral blood monocytes orCD34� cells was observed between patient samples and nor-mal donor samples (data not shown).
DQ-BSA Uptake by In Vitro Generated DC from ISSOTx Patients is Enhanced Compared with Controls
DQ-BSA represents BSA that has been labeled at a highmolecular ratio with BODIPY-FL, rendering the dye stronglyself-quenched. Intracellular proteolysis of the conjugates byDCs can be easily monitored by flow cytometry because di-gestion results in dequenching and increased probe fluores-cence. It is well accepted that efficient Ag uptake is a featureassociated with iDCs (1). In our experiments, iDCs generatedfrom IS SOTx patients displayed a significantly higher capac-ity to endocytose and process DQ-BSA when compared withthose from normal donors iDCs, indicating that that patients’iDCs were functionally more immature (Fig. 2A).
In Vitro Generated DCs from IS SOTx Patients areLess Likely to Mature in Response to Cytokines
iDCs generated from IS SOTx patients expressed MHCclass I, CD86, CD40, and CD11c molecules at levels similar to
© 2005 Lippincott Williams & Wilkins 453Macedo et al.

normal control iDCs but expressed lower levels of MHC classII, CD80, and CD54 molecules (Table 1).
We next evaluated whether iDCs generated from theblood of IS SOTx patients could be triggered to undergo mat-uration as compared with normal donor iDCs. As expected,the mixture of pro-inflammatory cytokines triggered normalDCs to enlarge and become more granular (data not shown)and to up-regulate expression of CD83, MHC class I and II,costimulatory, and adhesion molecules (Table 1). Combinedtreatment of normal iDCs with the pro-inflammatory cyto-kine mix and rhIFN-� further matured the DC phenotype. Bycontrast, treatment with rhIFN-� alone triggered MHC classI and II as well as CD54 up-regulation but had minimal effecton costimulatory molecule expression (CD40, CD80, andCD86) (Table 1). DC generated from SOTx patients exposedto the cytokine cocktail treatment up-regulated MHC class Iand II, as well as CD54 and CD83, but exhibited a diminishedcapacity to up-regulate the expression of the costimulatorymolecules (CD80, CD86, and CD40) as compared with nor-mal donor DCs (Table 1). The combined treatment with thecytokine mix and IFN-� further matured the patients DCs;however, the levels of CD80, CD86, and CD40 expressionremained suppressed relative to normal donor DCs (Table 1).
In Vitro Generated DCs from IS SOTx PatientsShow Impaired Capacity to Induce Proliferationto Recall- or Allo-Ags
Results in Figure 2B show that maturation with cytokinemix increased the ability of normal donor DCs to stimulate both“recall” and allo–T-cell proliferative responses, as expected. Ad-dition of rhIFN-� to the pro-inflammatory cytokine cocktailfurther increased the ability of normal donor DCs to promote
Ag- or allo-induced T-cell proliferation (Fig. 2B). By contrast,mDCs generated from SOTx patients were overall compara-tively poor in stimulating either TTox- or allo–Ag-induced T-cell proliferation (Fig. 2C). The cytokine mix�IFN-� only mod-estly increased the DCs’ ability to stimulate allospecific T-cellproliferation, whereas TTox-specific T-cell proliferation was notfurther augmented (Fig. 2C). In addition, when stimulated non-specifically with PHA, patients’ T cells proliferated as well asnormal control T cells (Fig. 2D).
IS SOTx Patients’ MoDCs Stimulated with Pro-Inflammatory Cytokines and IFN-� Exhibit aDC1-Functional Phenotype
Different groups of DCs (iDC, mDC, mDC�IFN-�)generated from HLA-A2�/DR15� IS SOTx patients and con-trol subjects were loaded with TTox and used in vitro as stim-ulators of HLA-A2�/DR15� ex vivo generated TTox-specificT-cell clones. Cloning from one normal control subject re-sulted in the isolation of only CD4�Th0 (mixed IFN-�/IL-5)and CD8�Tc0 (mixed IFN-�/IL-5) clones, suggesting thatnaı̈ve T cells were being primed in vitro, whereas TTox-spe-cific CD4�Th1 memory (IFN-� only) clones were generatedselectively from another subject. Figure 3 demonstrates thatnormal donor DCs matured with pro-inflammatory cyto-kines were efficient in boosting IFN-� responses from bothCD4� and CD8� T0 (Fig. 3A) and CD4� Th1 (Fig. 3B) clonesand that responses were enhanced further by exposure of theDCs to combined pro-inflammatory cytokines�IFN-� treat-ment. In contrast, IS SOTx patients’ DCs stimulated withpro-inflammatory cytokines exhibited a reduced T-cell stim-ulatory capacity that only marginally exceeded that of iDCs.However when pro-inflammatory cytokines�IFN-� were
FIGURE 1. The gating strategy and frequency of dendritic cell (DC)1s in peripheral blood of solid-organ transplant (SOTx)patients. (A) Peripheral blood mononuclear cells (PBMC) obtained from immunosuppressed IS SOTx patients or normal subjectswere stained with a lineage cocktail monoclonal antibodies (mAbs) in four-color flow cytometric analysis. The PBMC were gatedon CD45�Lin� cells and further evaluated for human leukocyte antigen (HLA)-DR�/CD11c� expression. One representativeexperiment performed out of several is shown for normal subjects (n�10) and patients (n�6). (B) Mean�SD of the percent ofHLA-DR� and CD11c� cells of CD45�Lin� cells, as well as the percent of CD11c� cells of CD45�Lin�DR� cells were calculatedfrom all experiments performed. *P�0.02. SS; FS; FITC, fluorescein isothiocyanate; ECD.AQ
454 Transplantation • Volume 79, Number 4, February 27, 2005

used to stimulate patients’ DCs, they significantly polarizedtype-1 responses in both Th0/Tc0 and Th1 clones (Fig. 3, Aand B). These results suggest a critical role of IFN-� as a sec-ond signal (in addition to pro-inflammatory cytokines) toaugment patients’ DC1 function.
Evaluation of Cytokine Production (IL-12p70 andIL-10) by DCs Generated from IS SOTx Patients
Normal control DCs stimulated with the cytokinemix�IFN-� produced elevated levels of IL-12p70 as com-
pared with iDCs (Fig. 4A). Maturation of normal iDCswith the cytokine mix�IFN-� and CD40 ligation had asynergistic effect on IL-12p70 secretion (Fig. 4B). In con-trast, all patients’ DC treatment conditions elicited lessIL-12p70 than normal donor DCs (Fig. 4A). However, theyapproached normal IL-12p70 production levels when pa-tients’ DCs were stimulated by cytokine mix�IFN-� andCD40 ligation (Fig. 4B).
IL-10 production by DCs generated from SOTx pa-tients was elevated compared with normal donor DCs (Fig.
FIGURE 2. The endocytic activity and stimulatory capacity of iDCs generated from IS SOTx patients. (A) iDCs from ISSOTx patients (n�6) or control subjects (n�8) were incubated with six different dilutions of DQ Green bovine serumalbumin (BSA) for 1 hour at 37°C. The antigen (Ag) uptake was evaluated by flow cytometry. Results shown as mean� Dvalues of percent of positive fluorescent iDCs. Next, iDCs generated from a representative normal control (B) and arepresentative SOTx patient (C) were matured for an additional 24 hours in the presence of the cytokine mix�interferon(IFN)-�, incubated with autologous or allogeneic PBMCs (1�105/well) and used in Ag-specific or in allogeneic mixedleukocyte reactions (MLR). There was an equal number of HLA mismatches between responder and stimulator in bothcontrol and patient alloMLRs. Results shown as means�SD of cpm triplicates obtained at 50:1 PBMC:DC ratio, and arerepresentative of several experiments (control n�4; patients n�4) performed. (D) Ability of PBMCs from normal controls orSOTx patients to proliferate in response to phytohemagglutinin (PHA (1 �g/mL). *P�0.01 to 0.05; ***P�0.001 to 0.005. Aone-tail Student’s t test was used for the statistical analysis in this set of experiments.
© 2005 Lippincott Williams & Wilkins 455Macedo et al.

4C). Addition of IFN-� to the cytokine cocktail used to ma-ture DCs down-modulated IL-10 production from normaldonor DCs but had little effect on IL-10 secreted by patients’DCs (Fig. 4C). Maturation and CD40L/CD40 ligationboosted IL-10 production by DCs generated from both SOTxpatients and normal donors at comparable levels (Fig. 4D).
MoDCs from IS SOTx Patients Exhibit ReducedNF-kB Nuclear Translocation in Response toCytokines Compared with Control DCs
Figure 5 shows that pro-inflammatory cytokines in-duce nuclear translocation of NF-kB in normal DCs, as ex-pected. In addition, IFN-� synergistically enhanced the abil-ity of pro-inflammatory cytokines to induce nucleartranslocation of NF-kB in normal DCs, even though IFN-�itself exhibited no capacity to do so. In contrast, patients’ DCsstimulated with pro-inflammatory cytokines displayed re-duced NF-kB DNA binding versus normal donor DCs. Inter-estingly, treatment of DCs with both the pro-inflammatorycytokine mix and IFN-� did not promote any further increasein NF-kB translocation (Fig. 5), suggesting that the IFN-�effects on patients’ DCs were NF-kB independent. Notably,nuclear extracts of untreated DCs showed little binding to aconsensus kB site on NF-kB (Fig. 5).
DISCUSSIONThis is the first study that supports the contention that
chronic in vivo administration of FK506 and prednisone inSOTx patients interferes with DC differentiation and func-tion, both in vivo and in short-term ex vivo cultures. Al-though pro-inflammatory cytokines only modestly maturedthe patients’ DCs phenotype in vitro and enhanced theirfunction (proliferation, IL-12p70 production, and type-1 po-larizing immunity) ex vivo, concomitant treatment with pro-inflammatory cytokines and IFN-� (as a second-signal stim-ulation) appeared critical for the development of Th1-typeresponses (Figs. 3 and 4).
Calcineurin inhibitors and corticosteroids in particularhave complex effects on MoDC, both in animal models and inhuman in vitro experiments. Dexamethasone and prednisone
FIGURE 3. Pro-inflammatory cytokines and IFN-� in-duce the generation of type-1 polarized DCs (DC1s) from ISSOTx patients. Tetanus toxoid (Ttox)-specific Tc0 CD8� orTh0 CD4� T-cell clones (A) or Th1 CD4� T-cell clones (B)were generated from two control subjects and used as re-sponder cells in IFN-� ELISPOT assays. Autologous iDCs orallogeneic iDCs generated from HLA-A2/HLA-DR15-matched SOTx patients (n�2) or normal donors (n�4) werecultured in the presence of DC media alone or stimulatedwith interleukin (IL)-1�, tumor necrosis factor (TNF)-�, andIL-6�IFN-�. After 24 hours, DCs were harvested, pulsedwith Ttox, and used as antigen-presenting cells (APC) inELISPOT assays. Results expressed as mean� SD of thenumber of IFN–�-producing cells/105 cells and are repre-sentative of two Tc0 CD8�, two Th0 CD4�, and two Th1CD4� T-cell clones. *P�0.01 to 0.05; **P�0.005 to 0.009.
TABLE 1. Phenotype of DCs differentiated from IS SOTx patients and normal donors
Marker
Study groups
Control Patients
iDC mDC mDC mDC iDC mDC mDCMedia IFN� Cyt Cyt�IFN� Media Cyt Cyt�IFN�
CD14 13�2 16�3 18�4 13�6 18�0 11�6 21�17MHC II 142�57 203�81 344�206 418�167 80�55 345�96 470�257MHC I 311�92 421�76 553�212 666�191 331�86 549�80 610�384CD80 23�4 20�4 20�5 31�9 16�1 16�3 18�6CD86 38�12 45�11 67�23 93�31 43�15 50�14 68�25CD40 84�25 98�12 127�40 180�36 87�30 110�56 151�75CD54 222�92 400�15 597�377 830�509 142�71 591�80 783�330CD83 18�4 31�4 56�7 56�12 16�3 52�18 107�76CD11c 627�206 595�33 539�229 597�238 630�172 740�274 866�461
DC were cultured with rhGM-CSF�rhIL-4 for 6 days. Maturation was further induced by culture with rhIL-1�, rhIL-6, and rhTNF-� (cyt)�rhIFN� asdescribed in the Materials and Methods. Results are expressed as mean MFI�SD. Data are of six separate experiments (controls n�7, patients n�4).
DC, dendritic cells; IS, immunosuppression; SOTx, solid-organ transplant; MHC, major histocompatibility complex; GM-CSF, granulocyte-macrophagecolony-stimulating factor; IL, interleukin; TNF, tumor necrosis factor; IFN, interferon.
456 Transplantation • Volume 79, Number 4, February 27, 2005
inhibit in vitro human monocyte differentiation into DCs byaltering the ability of DCs to up-regulate costimulatory mol-ecules and to acquire mature phenotypes. IL-12p70 secretion byDCs, and the capacity of these APCs to drive recall Ag- or allo–Ag-inducedT-cellproliferation, isalso inhibitedbycorticosteroids(28–30). In mice, dexamethasone impairs culture-induced as is T–cell-mediatedterminalmaturationofDCs(31). InvitroexposureofDCs to FK506 does not appear to affect MoDC differentiation ormaturation, but it has been reported to hamper the ability ofDCs to secrete TNF-� and to stimulate allogeneic T-cell prolif-eration (29, 32). These reports support the notion that cortico-steroids have a stronger effect on MoDC than FK506, whereasthe latter induces more subtle alterations in DCs. Because allSOTx patients analyzed in this study received prednisone andFK506 as maintenance therapy, it is difficult to dissect the impactof each individual drug on DC biology. However, on the basis ofthe available literature and our own results, we believe that pred-nisone most likely impacts DC functionality to a greater extent,whereas FK506 exerts only a mild influence on DCs.
We determined that the frequency of the DC1 subset ofDCs was significantly lower than in normal controls (Fig. 1).This might have occurred either because of the preferentialapoptosis of DC1s in patients or to a treatment-associatedinhibition in differentiation and maturation of DC1s by pred-nisone. This latter possibility is supported by an earlier reportby Moser et al. (31) who found that in mice, a single injectionof corticosteroids resulted in a loss of splenic DCs associatedwith a blockage in DC differentiation. In addition, we foundthe yields of MoDCs generated in vitro from peripheral bloodof IS SOTX patients to be significantly higher than the yieldsof MoDC obtained from normal controls, although both ex-hibited normal monocyte values in peripheral blood (datanot shown). Conceivably, greater numbers of DC precursorsmay circulate in the peripheral blood of IS SOTx patients.
Although patients’ DCs could differentiate ex vitro, adiminished expression of CD54, CD80, and MHC Class IImolecules on patients’ iDCs was observed after 6 days in vitroculture with rhIL-4 and rhGM-CSF. Upon exposure to pro-
inflammatory cytokines, patients’ iDCs were impaired intheir ability to up-regulate costimulatory molecules (CD86,CD80, and CD40) as compared with normal donor DCs (Ta-ble 1). This correlated with the inferior capacity of these DCsto promote recall Ag- and allo–Ag-induced T-cell prolifera-tion (Fig. 3) and IL-12p70 production (Fig. 4).
Given these findings, we sought to determine whethershort-term ex vivo DC stimulation with pro-inflammatorycytokines and IFN-� could induce their phenotypic and func-tional maturation that might allow for their integration inadoptive immunotherapy applicable for IS SOTx patients. Invitro generated MoDCs loaded with tumor- or viral-associ-ated Ags have been shown to be effective tools for vaccination
FIGURE 4. Cytokine produc-tion by DCs generated from ISSOTx patients. iDC generatedfrom IS SOTx patients or normalcontrols were exposed to pro-inflammatory cytokines�IFN-�for 24 hours. DCs (2�104/well)were then exposed to the CD40Ltransfected cell line J558 (5�104/well) for an additional 24 hours.The concentration of IL-12p70 (Aand B) and IL-10 (C and D) in thesupernatants was measured byenzyme-linked immunoadsor-bent assay. Results are shown aspg/mL and are the mean�SD ofseveral experiments performed.*P�0.01 to 0.05.
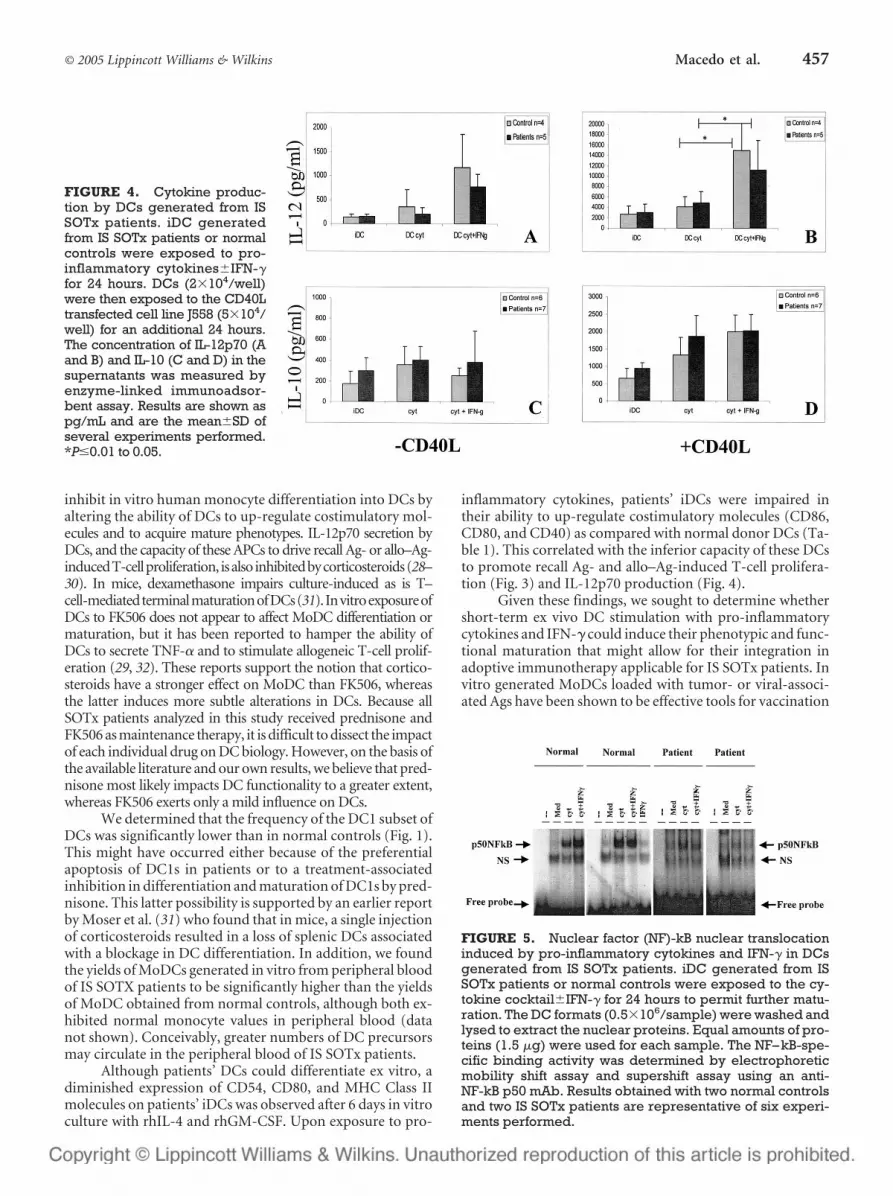
FIGURE 5. Nuclear factor (NF)-kB nuclear translocationinduced by pro-inflammatory cytokines and IFN-� in DCsgenerated from IS SOTx patients. iDC generated from ISSOTx patients or normal controls were exposed to the cy-tokine cocktail�IFN-� for 24 hours to permit further matu-ration. The DC formats (0.5�106/sample) were washed andlysed to extract the nuclear proteins. Equal amounts of pro-teins (1.5 �g) were used for each sample. The NF–kB-spe-cific binding activity was determined by electrophoreticmobility shift assay and supershift assay using an anti-NF-kB p50 mAb. Results obtained with two normal controlsand two IS SOTx patients are representative of six experi-ments performed.
© 2005 Lippincott Williams & Wilkins 457Macedo et al.

and adoptive immunotherapy of several types of cancer, in-cluding lymphoma (33, 34). The development of DC-basedvaccines to generate ex vivo EBV-specific CTLs for adoptiveimmunotherapy of refractory PTLD in SOTx recipients rep-resents an important clinical goal in the transplant setting.However, no study has yet determined whether MoDCs gen-erated from IS SOTx patients are suitable for suchapplication.
It is well accepted that for effective antiviral/antitumorimmunity, induction of Th1-type immune responses(IFN-�) is required and that this is a tightly regulated process(35, 36). IL-12p70 produced by monocyte/macrophages andDCs plays a pivotal role in the generation of protective Th1-type immunity (37). IFN-� is produced mainly by T and nat-ural killer cells and is important in boosting IL-12p70 pro-duction by DCs (38, 39). However, IFN-� by itself is notsufficient for the development of Th1-type immune re-sponses (35, 36). Our results suggest that IFN-� alone playsonly a minimal role in supporting normal donor or IS SOTxpatient DC maturation and in the ability of these APCs todrive recall Ag- or allo–Ag-induced T-cell proliferation andDC-associated cytokine production (Table 1, and data notshown). However, when provided as an additional stimulusto the pro-inflammatory cytokine cocktail, IFN-� facilitatesnormal DC1 to further mature, boost recall Ag- and allo-AgMLR, and to promote type-1 polarized T-cell responses invitro (Figs. 3 and 4). Although the coordinate exposure topro-inflammatory cytokines and IFN-� does not further trig-ger the IS SOTx patients’ DC stimulatory activity in MLR, itsignificantly augments DC1-type function and IL-12p70 pro-duction. Experiments are currently on going to test whetheraddition of exogenous T-cell growth factors to the patientsDC1s in MLR may rescue Ag-driven T-cell proliferation.
To further elucidate the molecular mechanisms in-volved in the phenotypic and functional changes observed inIS SOTx patients’ DCs, we analyzed the status of the NF-kBtranscription factor in these cells. Corticosteroids are de-scribed to potently inhibit NF-kB by triggering increased syn-thesis and cytoplasmic accumulation of IkB, which precludesNF-kB translocation to nucleus (40). Recently, Biswas et al.(41) showed that FK506 could also interfere with normal cy-toplasmic IkB inactivation triggered by calcineurin activity.Our data show that in normal donor DCs, pro-inflammatorycytokines enhance NF-kB nuclear translocation, which is fur-ther augmented in the presence of IFN-�. Interestingly,Cheshire et al. (19) have also reported that IFN-� and TNF-�synergize in the activation of NF-kB in preneuronal cells,which is consistent with our findings in normal donors DCs(20). However, IS SOTx patients’ DCs subjected to these sametreatments failed to significantly increase NF-kB transloca-tion to the nucleus (Fig. 5). We therefore speculate that othermolecular pathways (i.e., STAT1) might be dominantly in-volved in triggering the type-1 functional effects observed inpatient-derived DCs (42).
In summary, our results demonstrate that chronic IS inSOTx patients has a significant inhibitory impact on MoDCnumbers, phenotype, and function. The data also show thatIFN-�, in addition to pro-inflammatory cytokines, is a criticalsecond signal that can modulate and repolarize patientMoDCs, promoting the ex vivo development of DC1. Ourresults have important implications for future DC-based
adoptive immunotherapy against EBV infections and tumorsin SOTx patients. The usage of cyt�IFN-� mDCs to boost exvivo EBV-memory T cells might result in potent type-1 CTLs.
ACKNOWLEDGMENTSThe authors thank Sheila Fedorek, Clyde Harris, and
Lynn Ostrowski, the research coordinators, for their excellentsupport in collecting patients’ blood samples; Robbie Millardand Anna Kalinska-Wankowicz for their advice throughout;and Allison Logar for her excellent flow cytometric dataacquisition.
REFERENCES1. Banchereau J, Steinman RM. Dendritic cells and the control of immu-
nity. Nature 2001; 392: 245.2. Liu YJ. Dendritic cell subsets and lineages, and their functions in innate
and adaptive immunity. Cell 2001; 106: 259.3. Trinchieri G. Interleukin-12: a cytokine produced by antigen present-
ing cells with immunoregulatory functions in the generation ofT-helper cells type 1 and cytotoxic lymphocytes. Blood 1994; 84: 4008.
4. Steinman RM, Hawiger D, Liu K, et al. Dendritic cell function in vivoduring the steady state: a role in peripheral tolerance. Ann N Y Acad Sci2003; 987: 15.
5. Mocellin S, Panelli MC, Wang E, et al. The dual role of IL-10. TrendsImmunol 2003; 24: 36.
6. Jain A, Kashyap R, Dodson F, et al. A prospective randomized trial oftacrolimus and prednisone versus tacrolimus, prednisone and myco-phenolate mofetil in primary adult liver transplantation: a single centerreport. Transplantation 2001; 72: 1091.
7. Koenen HJ, Michielsen EC, Verstappen J, et al. Superior T-cell suppres-sion by rapamycin and FK506 over rapamycin and cyclosporine A be-cause of abrogated cytotoxic T-lymphocyte induction, impaired mem-ory responses, and persistent apoptosis. Transplantation 2003; 75:1581.
8. Nalesnik MA. Clinicopathologic characteristics of post-transplant lym-phoproliferative disorders. Recent Results Cancer Res 2002; 159: 9.
9. McDiarmid SV, Jordan S, Kim GS, et al. Prevention and preemptivetherapy of postransplant lymphoproliferative disease in pediatric liverrecipients. Transplantation 1998; 66: 1604.
10. Serinet MO, Jacquemin E, Habes D, et al. Anti-CD20 monoclonal an-tibody (Rituximab) treatment for Epstein-Barr virus-associated, B-celllymphoproliferative disease in pediatric liver transplant recipients.J Pediatr Gastroenterol Nutr 2002; 34: 389.
11. Green M, Reyes J, Webber S, et al. The role of antiviral and immuno-globulin therapy in the prevention of Epstein-Barr virus infection andpost-transplant lymphoproliferative disease following solid organtransplantation. Transpl Infect Dis 2001; 3: 97.
12. Aguilar LK, Rooney CM, Heslop HE. Lymphoproliferative disordersinvolving Epstein-Barr virus after hemopoietic stem cell transplanta-tion. Curr Opin Oncol 1999; 11: 96.
13. Rooney CM, Aguilar LK, Huls MH, et al. Adoptive immunotherapy ofEBV-associated malignancies with EBV-specific cytotoxic T-cell lines.Curr Top Microbiol Immunol 2001; 258: 221.
14. Metes D, Storkus WJ, Zeevi A, et al. Use of autologous dendritic cellsloaded with apoptotic LCL for ex vivo generation of specific CTL fromthe PBMC of EBV(-) individuals. Transplant Proc 2001; 33: 441.
15. Savoldo B, Cubbage ML, Durett AG, et al. Generation of EBV-specificCD4� cytotoxic T cells from virus naive individuals. J Immunol 2002;168: 909.
16. Jonuleit H, Knop J, Enk AH. Cytokines and their effects on maturation,differentiation and migration of dendritic cells. Arch Dermatol Res1996; 289: 1.
17. Vieira PL, de Jong EC, Wierenga EA, et al. Development of Th1-induc-ing capacity in myeloid dendritic cells requires environmental instruc-tion. J Immunol 2000; 164: 4507.
18. Denk A, Wirth T, Baumann B. NF-kappaB transcription factors: criti-cal regulators of hematopoiesis and neuronal survival. Cytokine GrowthFactor Rev 2000; 11: 303.
19. Cheshire JL, Williams BR, Baldwin AS JR. Involvement of double-stranded RNA-activated protein kinase in the synergistic activation of
458 Transplantation • Volume 79, Number 4, February 27, 2005

nuclear factor-kappaB by tumor necrosis factor-alpha and gamma-interferon in preneuronal cells. J Biol Chem 1999; 274: 4801.
20. Hoffmann TK, Meidenbauer N, Muller-Berghaus J, et al. Proinflam-matory cytokines and CD40 ligand enhance cross-presentation andcross-priming capability of human dendritic cells internalizing apopto-tic cancer cells. J Immunother 2001; 24: 162.
21. Fruh K, Yang Y. Antigen presentation by MHC class I and its regulationby interferon gamma. Curr Opin Immunol 1999; 11: 76.
22. Popescu I, Macedo C, Zeevi A, et al. Ex vivo priming of naı̈ve T cells intoEBV-specific Th1/Tc1 effector cells by mature autologous DC loadedwith apoptotic/necrotic LCL. Am J Transplant 2003; 3: 1369.
23. Romani, N, Reider D, Heuer M, et al. Generation of mature DC fromhuman blood: an improved method with special regard to clinical ap-plicability. J Immunol Methods 1996; 196: 137.
24. Bjorck P, Lie WR, Woulfe SL, et al. Progenipoietin-generated dendriticcells exhibit anti-tumor efficacy in a therapeutic murine tumor model.Int J Cancer 2002; 100(5): 586.
25. Hoffmann TK, Muller-Berghaus J, Ferris RL, et al. Alterations in thefrequency of dendritic cell subsets in the peripheral circulation of pa-tients with squamous cell carcinomas of the head and neck. Clin CancerRes 2002; 8: 1787.
26. Wankowicz-Kalinska A, Wijngaadr RM, Tigges BJ. Immunopolariza-tion of CD4� and CD8� T cells to type-1-like is associated with mela-nocyte loss in human vitiligo. Lab Invest 2003; 83: 683.
27. Andrews NC, Faller DV. A rapid micropreparation technique for ex-traction of DNA-binding proteins from limiting numbers of mamma-lian cells. Nucleic Acids Res 1991; 19: 2499.
28. Piemonti L, Monti P, Allavena P, et al. Glucocorticoids affect humandendritic cell differentiation and maturation. J Immunol 1999; 162:6473.
29. Woltman AM, de Fijter JW, Kamerling SW, et al. The effect of cal-cineurin inhibitors and corticosteroids on the differentiation of humandendritic cells. Eur J Immunol 2000; 30: 1807.
30. de Jong EC, Vieira PL, Kalinski P, et al. Corticosteroids inhibit theproduction of inflammatory mediators in immature monocyte-de-
rived DC and induce the development of tolerogenic DC3. J Leukoc Biol1999; 66: 201.
31. Moser M, De Smet T, Sornasse T, et al. Glucocorticoids down regulatedendritic cell function in vivo and in vitro. Eur J Immunol 1995; 25:2818.
32. Szabo G, Gavala C, and Mandrekar P. Tacrolimus and cyclosporine Ainhibit allostimulatory capacity and cytokine production of humanmyeloid dendritic cells. J Invest Med 2001; 49: 442.
33. Banchereau J, Paczesny S, Blanco P, et al. Dendritic cells: controllers ofthe immune system and a new promise for immunotherapy. Ann N YAcad Sci 2003; 987: 180.
34. Buchler T, Michalek J, Kovarova L, et al. Dendritic cell-based immu-notherapy for the treatment of hematological malignancies. Hematol-ogy 2003; 8: 97.
35. Abbas AK, Murphy KM, Sher A. Functional diversity of helper T lym-phocytes. Nature 1996; 383: 787.
36. O’Garra A. Cytokines induce the development of functionally hetero-geneous T helper cell subsets. Immunity 1998; 8: 275.
37. Scott P, Trinchieri G. IL-12 as an adjuvant for cell-mediated immunity.Semin Immunol 1997; 9: 285.
38. Gerosa F, Baldani-Guerra B, Nisii C, et al. Reciprocal activating inter-action between natural killer cells and dendritic cells. J Exp Med 2002;195: 327.
39. Snijders A, Kalinski P, Hilkens CM, et al. High-level IL-12 productionby human dendritic cells requires two signals. Int Immunol 1998; 10:1593.
40. Koski KG, Lyakh LA, Cohen PA, et al. CD14� monocytes as dendriticcell precursors: diverse maturation-inducing pathways lead to com-mon activation of NFkB/rRelB. Crit Rev Immunol 2001; 21: 179.
41. Biswas G, Anandatheerthavarada HK, Zaidi M, et al. Mitochondria tonucleus stress signaling: a distinctive mechanism of NFkappaB/Rel ac-tivation through calcineurin-mediated inactivation of IkappaBbeta.J Cell Biol 2003; 161: 507.
42. Ramana CV, Gil MP, Schreiber RD, et al. STAT-1-dependent and -in-dependent pathways in IFN-gamma-dependent signaling. Trends Im-munol 2002; 23: 96.
© 2005 Lippincott Williams & Wilkins 459Macedo et al.





























