Embed Size (px)
Citation preview

Biogenic Amines Modulate Synaptic Transmission between Identified Giant lnterneurons and Thoracic lnterneurons in the Escape System of the Cockroach
Janet L. Casagrand',**+ and Roy E. Ritzrnann'?*
Departments of 'Biology and *Neuroscience, Case Western Reserve University, Cleveland, Ohio 441 06
SUMMARY
In the escape system of the cockroach, Peripfunetu umericuna, a population of uniquely identifiable thoracic interneurons (type A or T1,s) receive information about wind via chemical synapses from a population of ventral giant interneurons ( vCIS). The T1,s are involved in the integration of sensory information necessary for orient- ing the animal during escape. It is likely that there are times in an animal's life when it is advantageous to mod- ify the effectiveness of synaptic transmission between the vCIs and the TI,s. Given the central position of the T1,s in the escape system, this would greatly alter associated motor outputs. We tested the ability of octopamine, sero- tonin, and dopamine to modulate synaptic transmission
INTRODUCTION
Animals adjust their behavioral responses in rela- tion to their current internal and external environ- mental conditions. The neural circuits responsible for these behaviors must be flexible enough to ac- commodate a variety of dynamic influences and yet generate meaningful behaviors. As we try to understand how these neural circuits function to generate complex behaviors, it will become impor- tant to take this flexibility into account.
Received February 4, 1992; accepted May I, 1992 Journal of Neurobiology, Vol. 23, No. 6, pp. 644-655 (1992) 0 1992 John Wiley & Sons, Inc. CCC 0022-3034/92/060644- 12$04.00
Some ofthese data were previously reported in abstract form
* To whom correspondence should be addressed. t Present address: Arizona Research Laboratories, Division
of Neurobiology, 6 I 1 Gould Simpson Building, University of Arizona, Tucson, AZ 85721.
644
(Casagrand and Ritzmann, I99 I ).
between vGIs and TI,s. Both octopamine and dopamine significantly increased the amplitude of vGI-evoked excit- atory postsynaptic potentials (EPSPs) in T1,s at lo-"- lo-* M, and M, respectively. On the other hand, serotonin significantly decreased the vCI-evoked EPSPs in TI,s a t 10-4-10-3 M. These results indicate that octo- pamine, serotonin, and dopamine are capable of modulat- ing the efficacy of transmission of important neural con- nections within this circuit. 0 1992 John Wiley & Sons, Inc. Keywords: biogenic amine modulation, ventral giant in- terneuron, thoracic interneuron, cockroach escape system.
We have been investigating the neural circuitry involved in evoking the escape response in the cockroach, Periplaneta amevicana, in order to gain a better understanding of how populations of neu- rons act to generate a defined behavior. A cockroach will normally orient away from the wind source in response to a wind puff. However, it has been shown that under certain conditions this response can be altered (Camhi and Volman, 1978; Camhi and Nolen, 1981: Camhi, 1988). This would imply that some form of modification occurs in the processing events that take place in the escape circuit.
Neuromodulatory substances, such as biogenic amines and neuropeptides, are reasonable candi- dates for this role. They have been shown to have profound effects on a number of neural circuits in invertebrates (Selverston, 1985; Getting, 1989: Harris-Warrick and Marder, 199 1 ). Moreover, there is a great deal of evidence for the involvement of biogenic amines as neuromodulators in insects
(OShea and Evans, 1979; Evans and OShea, 1977, 1978; Sombati and Hoyle, 1984a,b; Kinna- mon, Klaassen, Klammer, and Claassen, 1984; Bellah, Fitch, and Kammer, 1984; Claassen and Kammer, 1986; Ramirez and Orchard, 1990; Ra- mirez and Pearson, 199 1 a,b; Goldstein and Camhi, 199 1 ) . In particular, octopamine, seroto- nin, and dopamine, three of the most prevalent amines found in the cockroach (Dymond and Evans, 1979; Sloley and Owen, 1982; Martin, Bai- ley, and Downer, 1984; Shafi et al., 1989), have been shown to have important modulatory effects on a number of neural circuits in insects. These amines are found widely distributed in the nervous system of the cockroach, P. americana, along with uptake (Evans, 1978; Orchard, 1989) and inactiva- tion (Bodnaryk, Brunet, and Koepe, 1974; Richter and Rutschke, 1977) mechanisms, suggesting that these biogenic amines could play roles as neuro- modulators in the cockroach. Finally, the targets of these neuromodulators often appear to be the neural networks that effect escape behaviors (Twarog and Roeder, 1957; O'Shea and Murphey, 1978; Glanzman and Krasne, 1983; Sombati and Hoyle, 1984a,b; Kinnamon et al. 1984; Hashemza- deh-Gargari and Friesen, 1989).
In the escape system of the cockroach, Peripfan- eta americana, a population of approximately 100 uniquely identifiable thoracic interneurons (type A or T1,s) receive information about wind direc- tion via short latency, apparently cholinergic in- puts from a population of ventral giant interneu- rons (vGIs) (Ritzmann and Pollack, 1988; Casa- grand and Ritzmann, 1992). The T I A S integrate vGI activity with additional sensory information (Ritzmann, Pollack, Hudson, and Hyvonen, 199 1 ). They then activate leg motor neurons via direct and indirect pathways (Ritzmann and Pol- lack, 1990). Thus, changes in the gain of synaptic inputs to the TI,s or their responsiveness to those inputs would result in changes in neural processing within the circuit, and could thereby alter motor outputs. Such changes would have important con- sequences for the escape behavior. In this paper, we report on experiments that test whether biogenic amines have the ability to influence the vGI to TI, connections in the escape circuit of the cockroach.
MATERIALS AND METHODS
The techniques employed were the same as previously described (Casagrand and Ritzmann, 1992). Briefly, an
A
I VGI- 1
0 2 4 d 8 10 12 14 16 18 20
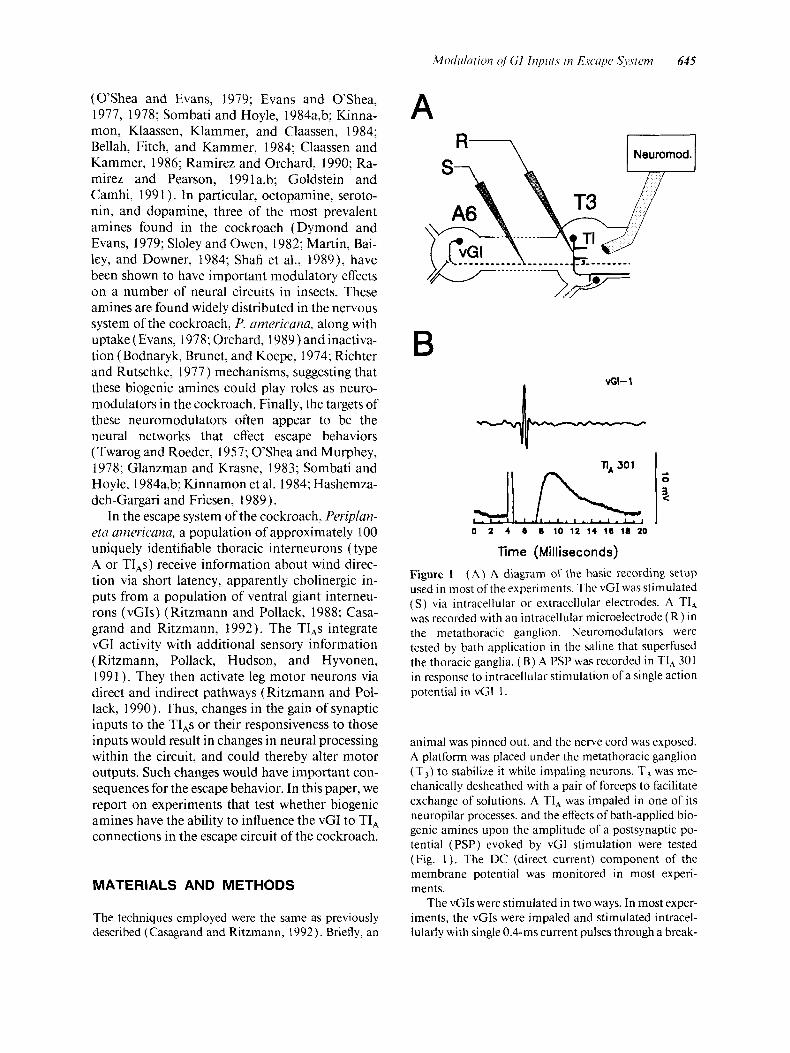
Time (Milliseconds) Figure I ( A ) A diagram of the basic recording setup used in most of the experiments. The vCI was stimulated ( S ) via intracellular or extracellular electrodes. A TI, was recorded with an intracellular microelectrode ( R ) in the metathoracic ganglion. Neuromodulators were tested by bath application in the saline that superfused the thoracic ganglia. ( B) A PSP was recorded in TI, 301 in response to intracellular stimulation of a single action potential in vGI I .
animal was pinned out, and the nerve cord was exposed. A platform was placed under the metathoracic ganglion (T,) to stabilize it while impaling neurons. T, was me- chanically desheathed with a pair of forceps to facilitate exchange of solutions. A TI, was impaled in one of its neuropilar processes, and the effects of bath-applied bio- genic amines upon the amplitude of a postsynaptic po- tential (PSP) evoked by vGI stimulation were tested (Fig. 1). The DC (direct current) component of the membrane potential was monitored in most experi- ments.
The vGIs were stimulated in two ways. In most exper- iments, the vGIs were impaled and stimulated intracel- lularly with single 0.4-ms current pulses through a break-

ttttttt1988). The breakaway box was necessary, as consider- able current is required to stimulate the large diameter vGIs. This method of stimulation resulted in large stimu- lus artifacts recorded in the Tl,s. However, these arti- facts did not interfere with data analysis. Activity from the stimulated vGI was recorded with bipolar silver hook electrodes placed between the second and third abdomi- nal ganglia. I n a few experiments, the vGIs were stimu- lated extracellularly through the hook electrodes on the abdominal nerve cord at an intensity just sufficient to evoke action potentials in large-diameter axons. The level was adjusted to evoke a single action potential. In these cases. an additional pair of hook electrodes be- tween T, and T, monitored extracellular activity from vGls. If during the experiment. another axon was re- cruited, then the data point was discarded. In general, it was not necessary to alter the stimulus strength during an experiment.
lntracellular and extracellular signals were recorded on VCR tape using a Medical Systems Corporation A / D VCR adapter. Data were analyzed on a Could digital oscilloscope. I Tests were performed to determine whether the changes in the EPSP amplitudes were statis- tically significant. In all but one case, the amplitudes of the EPSPs before exposure to biogenic amines were com- pared to the amplitudes at the height of the response. In one preparation [that shown in Fig. 6( B ) ] , it was neces- sary to use the amplitudes after wash rather than before exposure as the baseline level. In all preparations. the changes in the responses were significant at the p < 0.05 level or better.
Wafford and Sattelle ( 1986) saline for desheathed ganglia was used in all experiments to superfuse the nervous system. This will be referred to as normal .su- line. Thc following pharmacological agents were ob- tained from Sigma Chemical Co., (St. Louis, MO): ( DL) octopamine hydrochloride. j-hydroxytrypta- mine creatine sulfate (serotonin), 3-hydroxytyramine hydrochloride (dopamine), and D.L synephrine ( 1-[4- hydroxyphenyl]-2-methylaminoethanol). These were dissolved in saline at concentrations of M to lo-’ .M and bath-applied using a peristaltic pump (Rain in) to regulate flow. A flow rate of 2-4 ml /min was used throughout the experiments.
TI,s and vGls were stained with 4%) Lucifer Yellow (Molecular Probes) in 0.3 A4 LiAc and subsequently identified by their morphology when viewed in whole mount with a fluorescent microscope. When vGIs were stimulated intracellularly, the preparations were subse- quently embedded in paraplast and sectioned at 10 l m . The vGls were identified by the characteristic location of their axons in cross sections of the abdominal ganglia. The Tl,s werc named according to a numbering system (Westin, Ritzmann, and Goddard, 1988; Ritzmann and Pollack, 1990) similar to that used in locust (Robertson and Pearson, 1983). The general classes of neurons dis-
cussed in this paper belong to the 100, 300,500, and 700 series. Neurons in the 100 series are local interneurons. Those in the 300, 500, and 700 series all have axons contralateral to their somata. TI,s in the 700 series have axons that exit the ganglion both anteriorly and posteri- orly, those from the 500 series have axons that exit anteri- orly, and neurons in the 300 series have axons that exit the ganglion posteriorly.
RESULTS
The effects of perfusion of octopamine, serotonin, and dopamine upon the EPSPs evoked in TIAS by stimulation ofthe vGIs are summarized in Table 1. In all cases, changes in postsynaptic input imped- ance were tested by comparing rise time of EPSPs before and after agents were added. The only con- sistent significant change that was observed was an increase in rise time in the presence of synephrine, an analog of octopamine ( t test). We are, there- fore, confident that the changes noted below for the neuromodulators known to be present in cockroach are not the result of postsynaptic resis- tance changes. Even with synephrine, we feel that resistance changes cannot totally account for the increases that were observed.
Octopamine
Perfusion of octopamine at concentrations of lop4 to l o p 2 A4 resulted in significant potentiation of synaptic transmission between the vGIs and the TI,s of between 40% and 200% (Table 1; Fig. 2) . At a concentration of lop4 Moctopamine, we ob- served a 40% increase in the amplitude of the EPSP recorded in TI, 501 in response to extracellular stimulation of a vGI [Fig, 3 (A)] . The synaptic re- sponse began to decrement before the wash was begun, which may have been due to desensitization of the receptors, or to removal or uptake of the octopamine (Goosey and Candy, 1980; Evans, 1 9 7 8 ) . A slight hyperpolarization ( 5 mV) was noted in this preparation after 12 min of perfusion with octopamine. At this time, the amplitude of the PSP had already increased by 30%. After this, the DC component of the membrane potential re- mained at this level.
M, increases of 60%- 170% were observed in the EPSPs evoked in TIAs ( 30 1, 50 1, 70 1 ) in six different preparations (Fig. 2) . The time course of the effects of M octo-
At a concentration of
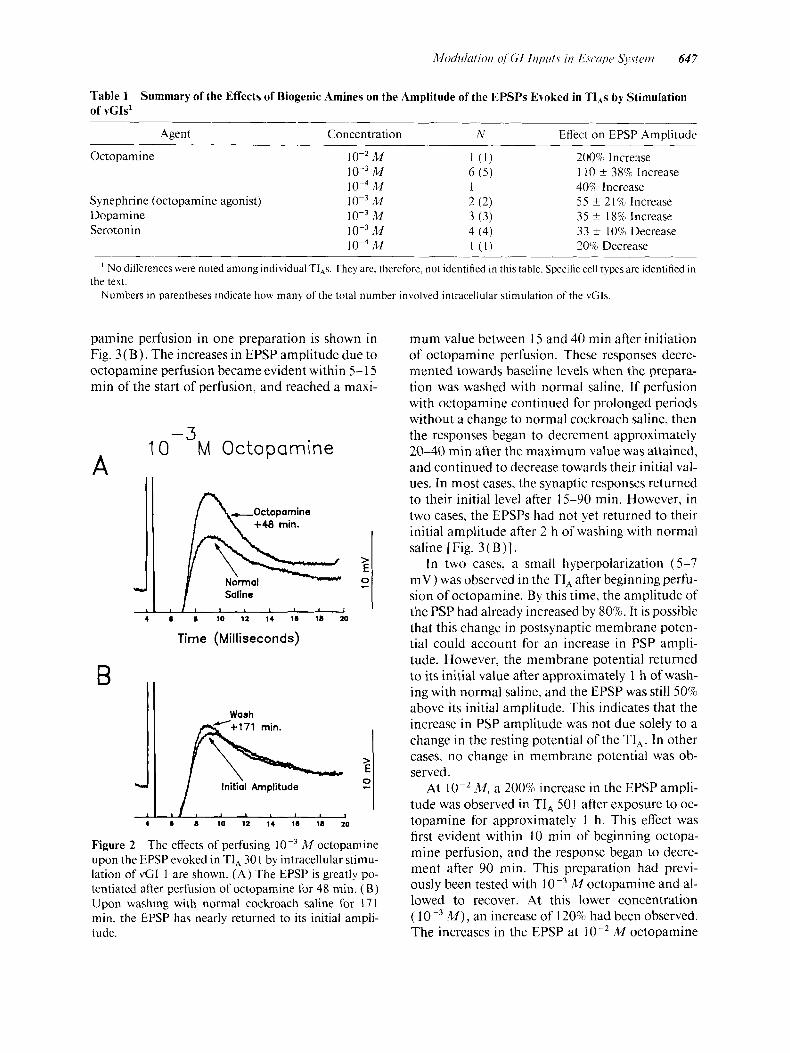
Table 1 of VCIS'
Summary of the Effects of Biogenic Amines on the Amplitude of the EPSPs Evoked in TI,s by Stimulation
Agent Concentration N EfTect on EPSP Amulitude
Octopamine
Synephrine (octopamine agonist) Dopamine Serotonin
M 10-3 M
M 10-3 M 10-3 M
M M
1 ( 1 ) 200% Increase 6 ( 5 ) 1 40%) Increase 2 ( 2 ) 3 ( 3 ) 4 (4) [ ( I ) 20% Decrease
110 f 38% Increase
5 5 f 21% Increase 35 f 18% Increase 33 f 10% Decrease
' No diferences were noted among individual TI,s. They are, therefore, not identified in this table. Specific cell types are identified in
Numbers in parentheses indicate how many of the total number involved intracellular stimulation of the vGIs. the text.
pamine perfusion in one preparation is shown in Fig. 3 (B) . The increases in EPSP amplitude due to octopamine perfusion became evident within 5- 15 min of the start of perfusion, and reached a maxi-
-3 10 M Octopamine
A
+48 rnin.
Normol Saline
8 I) 10 12 14 18 18 20
Time (Milliseconds)
> 0
E -
>
0
E Initial Amplitude -
4 8 8 10 12 14 18 18 20
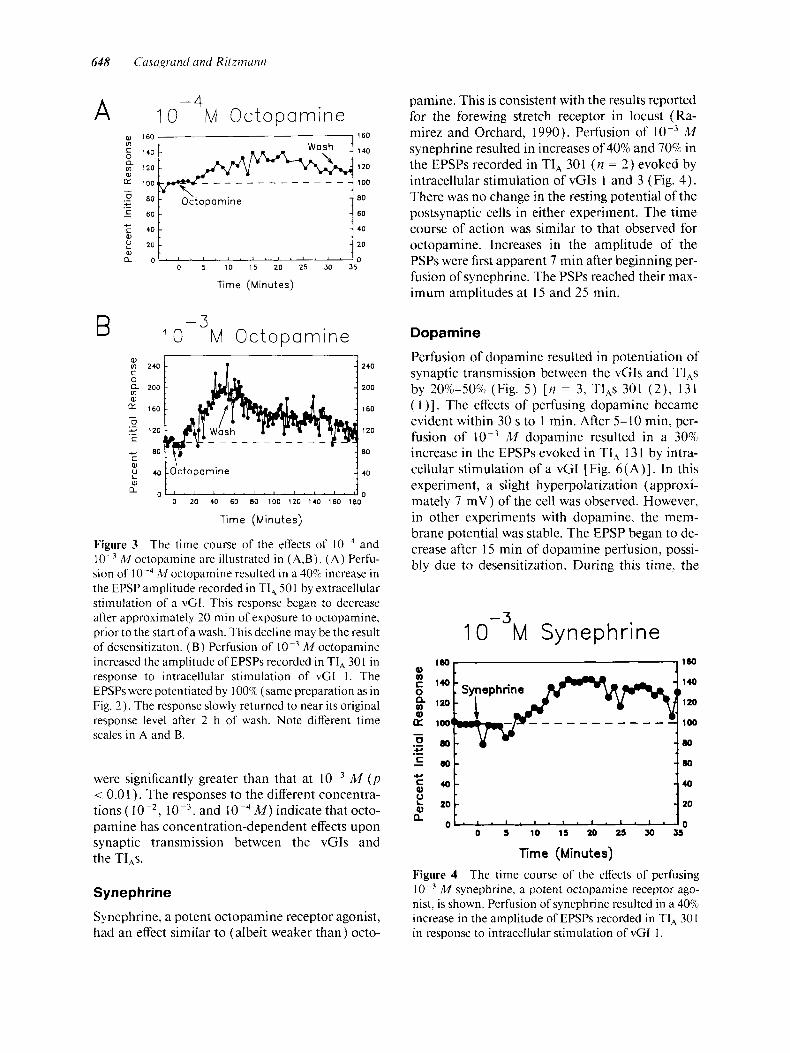
Figure 2 The effects of perfusing M octoparnine upon the EPSP evoked in TI, 30 1 by intracellular stimu- lation of vCI 1 are shown. ( A ) The EPSP is greatly po- tentiated after perfusion of octopamine for 48 min. (B) Upon washing with normal cockroach saline for 171 min, the EPSP has nearly returned to its initial ampli- tude.
mum value between I5 and 40 min after initiation of octopamine perfusion. These responses decre- mented towards baseline levels when the prepara- tion was washed with normal saline. If perfusion with octopamine continued for prolonged periods without a change to normal cockroach saline, then the responses began to decrement approximately 20-40 min after the maximum value was attained, and continued to decrease towards their initial val- ues. In most cases, the synaptic responses returned to their initial level after 15-90 min. However, in two cases, the EPSPs had not yet returned to their initial amplitude after 2 h of washing with normal saline [Fig. 3 ( B)] .
In two cases, a small hyperpolarization (5-7 mV) was observed in the TI, after beginning perfu- sion of octopamine. By this time, the amplitude of the PSP had already increased by 80%. It is possible that this change in postsynaptic membrane poten- tial could account for an increase in PSP ampli- tude. However, the membrane potential returned to its initial value after approximately 1 h of wash- ing with normal saline, and the EPSP was still 50% above its initial amplitude. This indicates that the increase in PSP amplitude was not due solely to a change in the resting potential of the TI,. In other cases, no change in membrane potential was ob- served.
At lop2 M, a 200% increase in the EPSP ampli- tude was observed in TI, 50 1 after exposure to oc- topamine for approximately 1 h. This effect was first evident within 10 min of beginning octopa- mine perfusion, and the response began to decre- ment after 90 min. This preparation had previ- ously been tested with 1 0-3 M octopamine and al- lowed to recover. At this lower concentration ( 1 0-3 M ) , an increase of 120% had been observed. The increases in the EPSP at M octopamine
648 CusagnLnd und Ritzinnnn
A
B
-4 10 M Octopamine
160
140
120
100
80
60
n o 20 40::. 20
0 5 10 15 20 25 30 35
Time (Minutes)
-3 10 M Octopamine
240 a Ul 240 t 0 a 200 200
0, Cf 160 160 - 0 Y 120 C
+- 80 80
120 .- .- -
c
0 20 40 60 80 100 120 140 160 180
Time (Minutes)
Figure 3 The time course of the erects of and M octopamine are illustrated in (A ,B) . ( A ) Perfu-
sion of I 0-4 M octopamine resulted in a 40% increase in the EPSP amplitude recorded in TI, 501 by extracellular stimulation of a vCI. This response began to decrease after approximately 20 min of exposure to octopamine, prior to the start of a wash. This decline may be the result of desensitizaton. ( B ) Perfusion of A4 octopamine increased the amplitude of EPSPs recorded in TI, 301 in response to intracellular stimulation of vGI 1. The EPSPs were potentiated by 100% (same preparation as in Fig. 2 ) . The response slowly returned to near its original response level after 2 h of wash. Note different time scales in A and B.
were significantly greater than that at M ( p < 0.0 1 ). The responses to the different concentra- tions ( lo-', and M ) indicate that octo- pamine has concentration-dependent effects upon synaptic transmission between the vGIs and the TI,s.
Synephrine
Synephrine, a potent octopamine receptor agonist, had an effect similar to (albeit weaker than) octo-
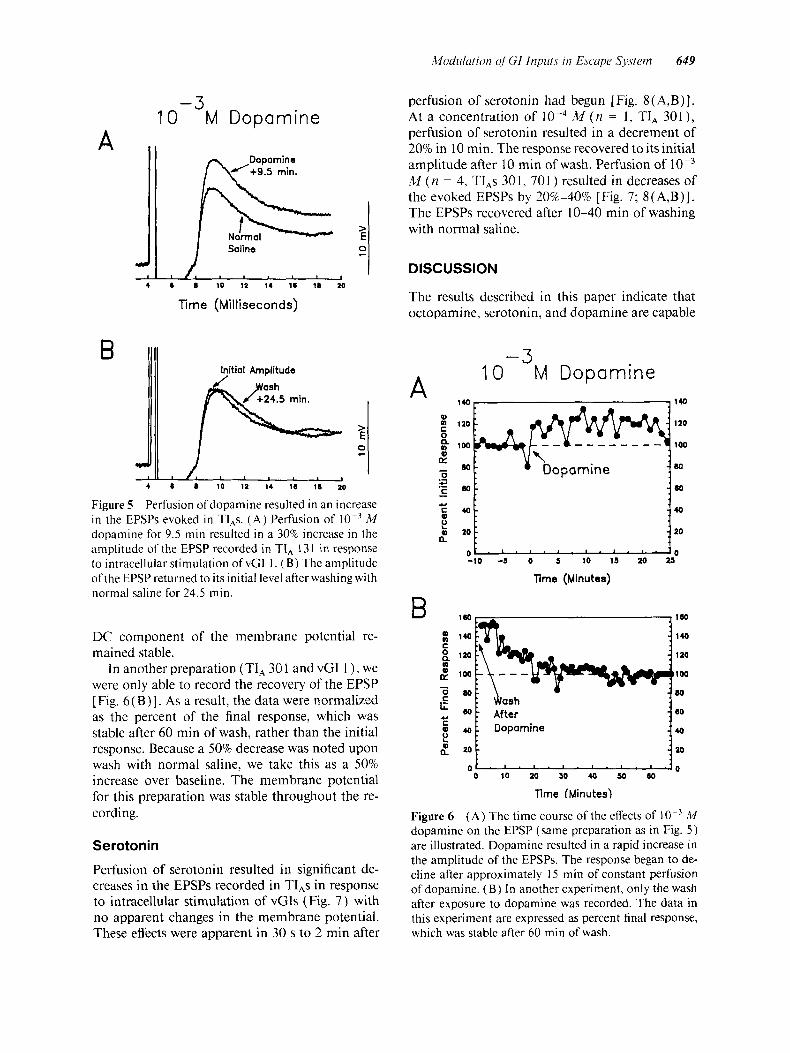
pamine. This is consistent with the results reported for the forewing stretch receptor in locust (Ra- mirez and Orchard, 1990). Perfusion of M synephrine resulted in increases of 40% and 70% in the EPSPs recorded in TI, 301 ( n = 2) evoked by intracellular stimulation of vGls I and 3 (Fig. 4). There was no change in the resting potential of the postsynaptic cells in either experiment. The time course of action was similar to that observed for octopamine. Increases in the amplitude of the PSPs were first apparent 7 min after beginning per- fusion of synephrine. The PSPs reached their max- imum amplitudes at 15 and 25 rnin.
Dopamine
Perfusion of dopamine resulted in potentiation of synaptic transmission between the vGIs and TI,s by 20%-50% (Fig. 5 ) [ n = 3, TI,s 301 (2) , 131 ( I ) ] . The effects of perfusing dopamine became evident within 30 s to 1 min. After 5-10 min, per- fusion of M dopamine resulted in a 30% increase in the EPSPs evoked in TI, 13 1 by intra- cellular stimulation of a vGI [Fig. 6 ( A)] . In this experiment, a slight hyperpolarization (approxi- mately 7 mV) of the cell was observed. However, in other experiments with dopamine, the mem- brane potential was stable. The EPSP began to de- crease after 15 min of dopamine perfusion, possi- bly due to desensitization. During this time, the
I O-3M Synephrine 1w I , 1 6 0
140
1 20
loo loo
Synephrine
W
W
40
20 20
0 0 0 5 10 15 20 25 30 35
Time (Minutes) Figure 4 The time course of the effects of perfusing
M synephrine, a potent octopamine receptor ago- nist, is shown. Perfusion of synephrine resulted in a 40% increase in the amplitude of EPSPs recorded in TI, 301 in response to intracellular stimulation of vGI 1.
A
-7 10 UM Dopamine
> E :
I 6 8 10 11 14 16 18 20
Time (Mi 1 ti se c o n d s)
B Initial Amplitude
+24.5 rnin.
> 0 E c
4 6 8 10 12 14 16 10 20
Figure 5 Perfusion of dopamine resulted in an increase in the EPSPs evoked in Tl,s. ( A ) Perfusion of M dopamine for 9.5 rnin resulted in a 30% increase in the amplitude of the EPSP recorded in TI, I3 1 in response to intracellular stimulation of vGI I . ( B ) The amplitude ofthe EPSP returned to its initial level after washing with normal saline for 24.5 min.
DC component of the membrane potential re- mained stable.
In another preparation (TI, 30 1 and vGI 1 ), we were only able to record the recovery of the EPSP [Fig. 6( B)]. As a result, the data were normalized as the percent of the final response, which was stable after 60 rnin of wash, rather than the initial response. Because a 50% decrease was noted upon wash with normal saline, we take this as a 50% increase over baseline. The membrane potential for this preparation was stable throughout the re- cording.
Serotonin
Perfusion of serotonin resulted in significant de- creases in the EPSPs recorded in TI,s in response to intracellular stimulation of VGIS (Fig. 7 ) with no apparent changes in the membrane potential. These effects were apparent in 30 s to 2 rnin after
perfusion of serotonin had begun [Fig. 8(A,B)]. At a concentration of M ( n = 1, TI, 301 ), perfusion of serotonin resulted in a decrement of 20% in 10 min. The response recovered to its initial amplitude after 10 rnin of wash. Perfusion of M ( n = 4, TI,s 30 1, 70 1 ) resulted in decreases of the evoked EPSPs by 20%-40% [Fig. 7; 8(A,B)] . The EPSPs recovered after 10-40 min of washing with normal saline.
DISCUSSION
The results described in this paper indicate that octopamine, serotonin, and dopamine are capable
-7 10 d M Dopamine A
I 8 i 140
120
100
W
Y
0 20 n 0
-10 -5 0 I 10 15 20 73
Time (Minutes)
B 1w
0 0 10 20 SO 40 50 60
Time IMinutes)
Figure 6 ( A ) The time course of the effects of M dopamine on the EPSP (same preparation as in Fig. 5 ) are illustrated. Dopamine resulted in a rapid increase i n the amplitude of the EPSPs. The response began to de- cline after approximately 15 rnin of constant perfusion of dopamine. ( B ) In another experiment, only the wash after exposure to dopamine was recorded. The data in this experiment are expressed as percent final response, which was stable after 60 rnin of wash.
650 Cusugrund and Ritzmunn
of modulating synaptic transmission at the vGI to TI, connections in the escape circuit of the cockroach. This provides an example of neuromod- ulation between two populations of identified in- terneurons in an insect central nervous system (CNS). Our results do not address the mechanism or the immediate locus of neuromodulatory influ- ence. Certainly, it is possible that the substances that were tested acted directly on other neurons that only secondarily alter the PSP amplitude re- corded in the T1,s. Because the changes that we recorded occurred gradually rather than as a step function, it is less likely that several spiking inter- neurons intervene between the site of neuromodu- latory action and the vGI to TI, synapses. How- ever, a definitive indication of the specific locus of action must await further experiments that were beyond the scope of this study. Nonetheless, it is clear from our results that release of neuromodula- tory substances into the neuropile of the thoracic
A
B
-L 4
-3 10 M Serotonin
I, 8 8 10 12 14 l a 18 20
Time (Milliseconds)
+ 23 rnin.
-L 4
Figure 7 The effects of perfusion of M serotonin on the EPSP evoked in T1,70 1 by intracellular stimula- tion of vGI 1 are shown. (A) After 10 min of perfusing, the EPSP is decreased by 40%. ( B ) Upon washing with normal saline for 23 min, the evoked EPSP has nearly returned to its initial amplitude.
A
B
-3 10 M Serotonin
- 20
d 0-0 0 5 1 0 1 5 2 0 2 5 3 0 S 5 4 0
Time (Minutes)
-7 1 0 "M Serotonin
Figure 8 The time course of the erects of perfusion of 1 0-3 M serotonin and wash out are illustrated. Perfusion of serotonin rapidly results in a decrement in the ampli- tude of EPSPs evoked in TI, 70 1 by intracellular stimula- tion ofvGI 1 ( A ) and in TI, 30 I by intracellular stimula- tion of vG1 2 ( B ) . When the preparation was perfused with normal saline. the response of the evoked EPSPs recovered to their initial response level.
ganglia has profound effects upon the escape system.
Our results are consistent with a large body of evidence in the neurobiological literature. Modula- tory effects of octopamine, serotonin, dopamine, and noradrenaline have all been described in in- sects (Twarog and Roeder, 1957; Hodgson and Wright, 1963; Evans and O'Shea, 1977; O'Shea and Murphey, 1978; Kinnamon et al., 1984; Dy- mond, 1978; Bellah et a]., 1984; Claassen and Kammer, 1986; Ramirez and Orchard, 1990; Goldstein and Camhi, 199 1 : Ramirez and Pear- son, 1991a,b), where they act to change the basic response properties of neurons and to alter motor patterns. Insect nervous tissue can also synthesize octopamine (Hoyle and Barker, 1975; Maxwell, Tait, and Hildebrand, 1978), serotonin (Colhoun,

1963), and dopamine (Colhoun, 1967), and the enzymes necessary for octopamine synthesis are known to be present in P. americana (Whitehead, 1969; Lake, Mills, and Brunet, 1970), along with adenylate cyclases activated specifically by octopa- mine, serotonin, and dopamine (Nathanson and Greengard, 1973, 1974; Robertson and Steele, 1973; Harmer and Horn, 1977). The amines octo- pamine, serotonin, and dopamine are widely dis- tributed in the nervous system of the cockroach, P. atnerrcana, and uptake (Evans, 1978; Orchard, 1989) and inactivation (Bodnaryk et al., 1974; Richter and Rutschke, 1977) mechanisms are also present, suggesting that these biogenic amines could play roles as neuromodulators in the cockroach.
Modulation of Behavior
It has been proposed that biogenic amines function to regulate behavioral responsiveness (i.e., sensiti- zation, arousal) in insects (Evans, 1980). For ex- ample, in many animals, octopamine appears to cause general excitation or increased responsive- ness (Kinnamon et al., 1984; Flamm and Harris- Warrick, 1986; Sombati and Hoyle, 1984a; O’Shea and Evans, 1979), and may serve in a general arousal phenomenon (Evans, 1980; Orchard, Loughton, and Webb, 1981; Glanzman and Krasne, 1983; Davenport and Evans, 1984) similar to the vertebrate sympathetic nervous system (Hoyle, 1974; Battelle and Kravitz, 1978; Evans and O’Shea, 1978; Candy, 1978; Hoyle and Dagan, 1978; O’Shea and Evans, 1979), whereas serotonin often has the opposite effect (Glanzman and Krasne, 1983; Harris-Warrick and Kravitz, 1984). Thus, it is possible that octopamine, dopamine, and serotonin act in the cockroach to modulate the threshold of escape in response to changes in the animal’s environment or physiological state.
Changes in the behavioral threshold for cockroach escape have been described under a vari- ety of behavioral conditions (Camhi and Volman, 1978; Camhi and Nolen, 1981; Camhi, 1988). For example, when a cockroach is walking, the thresh- old for wind-evoked escape decreases (Camhi and Nolen, 198 1 ). Under these conditions, there is no apparent change in the response ofthe GIs to wind, suggesting that the site of modulation is the meta- thoracic ganglion (Camhi, 1988). In a number of recent experiments, J. T. Watson and R. E. Ritz- mann ( unpublished observations) have found that threshold for escape varies markedly with different
physiological states. For example, cockroaches rarely escape in response to collisions with other cockroaches. This phenomenon may be related to the release of an aggregation pheromone ( Burk and Bell, 1973) detected by the antennae. If the anten- nae are covered or removed, then the animal escapes as normal (J . T. Watson and R. E. Ritz- mann, unpublished observations).
Although the mechanisms responsible for modu- lating the threshold of the escape response in the examples cited above are not known, it is possible that they could be mediated by the selective release of neuromodulatory substances. In the crayfish, for example, octopamine and serotonin differentially affect the threshold of a lateral giant escape reac- tion (Glanzman and Krasne, 1983). Octopamine enhances the lateral giant responsiveness, whereas serotonin depresses it. The results described in this paper indicate that the biogenic amines; octopa- mine, serotonin, and dopamine. similarly have the ability to differentially modulate synaptic trans- mission between the vGIs and TI,s in the escape circuit of P. arnericma. This modulation could have important consequences for the escape behav- ior if it translates into changes in the outputs of the TI,s, by bringing them closer or further away from threshold for action potentials. Indeed, since the TI, population has been estimated at between 76 and 100 interneurons (Ritzmann and Pollack. I990 ), the effects that we have reported would be multiplied many times throughout the system making even small synaptic changes very signifi- cant.
Goldstein and Camhi ( I99 I ) reported that topi- cal application of lop3 A4 serotonin and dopamine to the metathoracic ganglion resulted in changes (decrease, increase, respectively) in the extracellu- larly monitored activity of leg motor neurons evoked by either direct electrical stimulation of the abdominal nerve cord (including the GIs), or by wind stimulation. They also observed that seroto- nin increased the latency to the beginning of a mo- tor response, whereas dopamine decreased the la- tency. Since the TI,s make both direct and indirect connections to leg motor neurons (Ritzmann and Pollack, 19901, these effects are consistent with our observations. Moreover, they further suggest that the changes we noted do, in fact, have a func- tional consequence, and could serve to modify the escape behavior.
It is interesting that both octopamine and dopa- mine had similar effects upon the vGI to TI, con- nections; an increase in synaptic efficacy. How-

652 Casagrund and Ritzinunn
ever, it is possible that they are acting in different capacities upon the entire circuit. For example, one neuromodulator might result in general acti- vation of the circuit, setting the whole circuit very close to threshold at particular times, as has been suggested for octopamine in other systems (O’Shea and Evans, 1979; Sombati and Hoyle, I984a; Kin- namon et al., 1984; Flamm and Harris-Warrick, 1986). On the other hand, another neuromodula- tor might selectively modify the responsiveness of the TI,s to individual sensory modalities, making the circuit more or less, sensitive to specific inputs, including, but not restricted to, the vGIs. The TI,s receive a wealth of sensory input in addition to that from the vGIs (Ritzmann et al., 199 I ), and each of these connections could be differentially affected by the various neuromodulators. For example, in the crustacean stomatogastric system, neuromodu- lators can selectively modify the properties of some, or all, neurons in the circuit. In the presence of a single neuromodulator, the activity of some neurons can increase, whereas in others it de- creases or is simply unaffected (Marder and Eisen, 1984; Flamm and Harris-Warrick, 1986; Hooper and Marder, 1987; Katz and Harris-Warrick, 1990). Thus, although two of the biogenic amines that we tested had the same effect upon the vGI to TI, connections, the functional consequences of their actions may indeed differ.
Possible Sources of Biogenic Amines
The present studies suggest that local release of oc- topamine, dopamine, and serotonin is capable of modulating transmission between the vGIs and TIAS. Although we have not yet identified neurons that may, under the appropriate environmental stimuli, release neuromodulators, there are a num- ber of candidates. In the locust, at least some of the dorsal unpaired median (DUM) neurons in the metathoracic ganglion synthesize octopamine (Evans and O’Shea, 1977, 1978), and they are ca- pable of modulating neural transmission ( O’Shea and Evans, 1979; Sombati and Hoyle, 1984a) and specific behaviors (Evans and O’Shea, 1977, 1978; O’Shea and Evans, 1979; Sombati and Hoyle, 1984b) via release of octopamine (Morton and Evans, 1984). The cockroach also possesses DUM neurons, and some in the metathoracic ganglion have been shown to synthesize octopamine (Dy- mond and Evans, 1979). Neurosecretory endings, similar to those described for the octopaminergic DUM neurons in locust (Hoyle, 1975; Hoyle, Col-
quhoun, and Williams, 1980) are found to be asso- ciated with the GI terminals in the metathoracic ganglion of the cockroach (Castel, Spira, Parnas, andYarom, 1976), which mayrepresenttheneuro- pi1 processes of the DUM cells (Dymond and Evans, 1979). In addition, several serotonin-im- munopositive somata have been stained in the metathoracic ganglion of the cockroach, P. ameri- cana (Bishop and O’Shea, 1983) and locust (Tag- hert and Goodman, 1984), and there is a dense serotonergic innervation within the cockroach metathoracic ganglion (Bishop and O’Shea, 1983). Putative dopaminergic neuronal somata have also been described in the metathoracic gan- glion of cockroach nymphs (Fleming and Pitman, 1983), and catecholaminergic processes are found in the neuropil of the metathoracic ganglion (Goldstein and Camhi, 199 1 ). More work needs to be done in this area to identify neurons that could serve to modulate the processing that occurs at the vGI to TI, connections, and to determine under what conditions these neurons are activated to re- lease their neuromodulatory substances.
The concentrations of octopamine, dopamine, and serotonin that were used in our experiments to modulate the vGI to TI, connections were lop4 to lo-’ M. Lower concentrations were not tried, but it is likely that the actual threshold concentration of these biogenic amines is lower. The threshold concentration for octopamine, for example, may be lop5 M or less, since a 40% increase was ob- served at lop4 M . These concentrations are of the same order of magnitude as were found to be effec- tive in other studies in which biogenic amines were bath-applied to the nervous systems of insects (Twarog and Roeder, 1957; O’Shea and Evans, 1979; Dymond, 1978; Kinnamon et al., 1984; Bel- lah et al., 1984; Goldstein and Camhi, 199 l ).
Insect nervous systems have been found to be far more responsive to direct injection of octopa- mine into the neuropil than to bath-applied octo- pamine (Sombati and Hoyle, 1984a; Kinnamon et al., 1984; Stevenson, 1989). This may be due to diffusion barriers or mechanisms of neuroniodula- tor uptake and inactivation. Effective uptake and inactivation mechanisms in the ganglia ( Bodnaryk et al., 1974; Richter and Rutschke, 1977; Evans, 1978; Orchard, 1989) may serve to keep biogenic amines in the hemolymph from reaching central synaptic regions. In insects, as well as other inverte- brates, biogenic amines are released not only near synaptic sites to locally modulate transmission at conventional synapses (Orchard, I982), but also

Modulation of GI Inpiits in E.scupc Sjistem 653
into the hemolymph as a neurohormone (David and Lafon-Cazal, 1979; Goosey and Candy, 1980, 1982; Orchard et al., 1981; Lang, Orchard, and Barnett, 1989). Sombati and Hoyle ( 1984a) have suggested that the highly compartmentalized orga- nization of insect ganglia (Watson and Burrows, 1982) serves to prevent actions of octopamine far from its intended targets.
We chose not to use injection methods, because the injection process tends to cause a slight move- ment that could jar the microelectrode used to re- cord synaptic potentials. This could alter the size of the PSPs. Such an artifact would have been a severe problem in assessing the effects of neuromodula- tory substances at these synapses.
CONCLUSIONS
The ability of neuromodulators to transiently change the effectiveness of synaptic transmission and responsiveness of neurons, and thus alter be- haviors, is exciting. It increases the flexibility of circuits, allowing animals to modify their behavior in response to particular internal or external cues, without increasing either the number of neurons in a circuit. or the number of circuits. Such modifica- tions in the gain of inputs to elements of the cockroach escape circuit could have a profound impact on the information processing that occurs at the vC1 to TI, connections, and, therefore, on the escape behavior. We know that behavioral changes i n the threshold of escape in the cockroach do occur (Camhi and Volman, 1978; Camhi and Nolen, 1981; Camhi, 1988; Ritzmann et al., 199 1 ), and thus, it would appear that the escape circuit can be modified under appropriate condi- tions. These studies suggest that neuromodulation could underlie such changes, and further studies o n the nature and locations in which it is utilized should provide useful insights into the mechanism whereby dedicated circuits can be modified to ad- just their outputs to suit a variety of situations.
The authors would like to thank Drs. Edmund Arbas, Richard Levine, David Morton, and Jane Witten for crit- ically reviewing the manuscript. This work was sup- ported by NIH grant NS17411 to R.E.R.
REFERENCES
BATTELLE, B. A. and KRAVITZ, E. A. ( 1978). Targets of octopamine action in the lobster: cyclic nucleotide
changes and physiological effects in haemolymph, heart, and exoskeletal muscle. J . Pharmacol. Exp. Ther. 205:438-448.
BELLAH, K. L., FITCH, G. K., and KAMMER, A. E. ( 1984). A central action ofoctopamine on ventilation frequency in Corjidalzu cornutiis. J. Exp. Zool.
BISHOP, C. A. and OSHEA, M. (1983). Serotonin reac- tive neurons in the central nervous system of an insect ( Prripluneta umericana). J. Neurobiol. 14: 15 1 - 169.
BODNARYK, R. D., BRUNET, P. C. J., and KOEPE, J. K. (1974). On the metabolism of N-acetyldopamine in Peripluneta americanu. .J. Insect Physiol. 20:9 I 1-923.
BURK, T. and BELL, W. J. (1973). Cockroach aggrega- tion pheromones: inhibition of locomotion (Orthop- tera: Blattidae). J. Kansas En/. Soc. 46:310.
CAMHI, J. M. ( 1988). Sites of multiple controls ofexcit- ability in the escape circuit of the cockroach Periplun- eta americanu. SOC. Neiirosci. Abstr. 14:3 10.
CAMHI, J. M. and VOLMAN, S. (1978). Modulation of the threshold in the escape behavior of the cockroach Peripluneta arnericana. SOC. Nezirosci. Ahstr. 4: 189.
CAMHI, J. M. and NOLEN, T. G. (1981). Properties of the escape system of cockroaches during walking. J . Corny. Phjisiol. 142:339-346.
CANDY, D. J. (1978). The regulation of locust flight muscle metabolism by octopamine and other com- pounds. Insect Biochrm. 8: 171- 18 1.
CASAGRAND, J. L. and RITZMANN, R. E. ( 1991 ). Bio- genic amines modulate synaptic transmission between ventral giant interneurons and thoracic interneurons in the escape system of the cockroach. Soc. Nezirosci. Ahstr. 17:276.
CASAGRAND, J. L. and RITZMANN, R. E. (1992). Evi- dence that synaptic transmission between giant inter- neurons and identified thoracic interneurons in the cockroach is cholinergic. J. Nezirohiol. 23:627-643.
CASTEL, M., SPIRA, M. E., PARNAS, I., and YAROM, Y. ( 1976). Ultrastructure of region of a low safety factor in inhomogeneous giant axon of the cockroach. J. Neurophysiol. 39:900-908.
CLAASSEN, D. E. and KAMMER, A. E. ( 1986). Effects of octopamine, dopamine and serotonin on production of flight motor output by thoracic ganglia of Mandzica .sexta. J . Neiirohid. 11: I 14.
COLHOUN, E. H. ( 1963). Synthesis of 5-hydroxytrypt- amine in the american cockroach. Experentiu 19:9- 10.
COLHOUN, E. H. ( 1967). Pharmacological tantalizers. In: Insects and Phj’siology. J. W. Beament and J. E. Treherne, Eds. Oliver and Boyd, Edinburg, pp. 201- 213.
DAVENPORT, A. P. and EVANS, P. D. (1984). Stress-in- duced changes in the octopamine levels of insect hae- molymph. Insect Biochem. 14: 135-143.
DAVID, J. C. and LAFON-CAZAL, M. ( 1979). Octopa- mine distribution in the Loczista migrutorin nervous
23 1 :2 8 9-292.

654 Cawgrand and R i t m u n n
and non-nervous systems. Cotnp. Biochctn. Phj:c.io/. 64C: 16 1-1 64.
DYMOND, G. R. ( 1978). Biogenic amines in the central nervous system of the cockroach. Periplunetu utncri- curia. Ph.D. Thesis, University of Cambridge, En- gland.
DYMOND, G. R. and EVANS, P. D. (1979). Biogenic amines in the nervous system of the cockroach, Pui- plunctu ameuicuna: association of octopamine with mushroom bodies and dorsal unpaired median ( D U M ) neurones. In.srct Biochcvn. 9:535-545.
EVANS, P. D. (1978). Octopamine: a high affinity up- take mechanism in the nervous system of the cockroach. .I. Nezirochern. 30: 101 5-1022.
EVANS, P. D. ( 1980). Biogenic amines in the insect ner- vous system. .4dr. Insect Ph~:siol. 15:3 17-473.
EVANS, P. D. and O'SHEA, M. ( 1977). The identification of an octopamine neurone which modulates neuro- muscular transmission in the locust. hrutiire 270:257- 259.
EVANS. P. D. and OSHEA, M. ( 1978). The identification of an octopaminergic neurone and the modulation of a myogenic rhythm in the locust. J . Ekp. Biol.
FLAMM, R. E. and HARRIS-WARRICK, R. M. ( 1986). Aminergic modulation in lobster stomatogastric gan- glion. 11. Target neurons of dopamine, octopamine, and serotonin within the pyloric circuit. J. NmropIzj>.s- iol. 555366-88 1.
FLEMING. J. R. and PITMAN, R . M. ( 1983). Histochemi- cal localization of biogenic amines in cockroach neu- rons. J. P h j W l . (1,ond.l 33439XP.
GETTING, P. A. ( 1989). Emerging principles governing the operation of neural networks. .&?nil. Rrv. Ncirro-
GLANZMAN, D. L. and KRASNE, F. B. ( 1983). Serotonin and octopamine have opposite modulatory effects on the crayfish's lateral giant escape reaction. J. Ncmro.sci. 3:2263-2269.
GOLDSTEIN. R. S. and CAMHI. J. M. ( 199 1 ). Different erects of the biogenic amines dopamine, serotonin and octopamine on the thoracic and abdominal por- tions of the escape circuit in the cockroach. J. Comp.
GOOSEY. M. W. and CANDY, D. J. ( 1980). The D-OCtO- pamine content of the haemolymph of the locust Schistocenu utnc~ricuna grquriu. and its elevation during flight. Insect Biochcin. 10:393-397.
GOOSEY, M. W. and CANDY, D. J. (1982). The release and removal of octopamine by tissues of the locust Schi.stocm~u arnrricwna grc2guriu. I n s ~ ~ c r BiochCtn.
HARMER. A. J. and HORN, A. S. ( 1977). Octopamine- sensitive adcnylate cyclase in cockroach brain: effects of agonists, antagonists and guanylyl nucleotides. Mo- l e ~ . Phurmucol. 13:s 12-520.
HARRIS-WARRICK. R. M. and KRAVITZ, E. A. (1984).
73~235-260.
S C ~ . 12: 185-204.
Phj'SiOl. [ i l l 168:103-1 12.
l2:68 1-685.
Cellular mechanisms for modulation of posture by oc- topamine and serotonin in thc lobster. J . Ncwosci.
HARRIS-WARRICK. R. M. and MARDER. E. ( 1991). Modulation of neural networks for behavior. Arinii .
Rev. Niwrosci. 14:39-57. HASHEMZADEH-GARGARI. H. and FRIESEN. W. 0.
( 1989). Modulation of swimming activity in the me- dicinal leech by serotonin and octopamine. Cornp. Biochcm. Pli~~siol. 94(7:295-302.
HODCSON, E. S. and WRIGHT, A. M. ( 1963). Action of epinephrine and related compounds upon the insect nervous system. Gen. C'omp. Endocrinol. 3:5 19-525.
HOOPER. S. L. and MARDER. E. ( 1987). Modulation of the lobster pyloric rhythm by the peptide proctolin. .I. (V~iirosci. 73097-2 1 12.
HOYLE. G. ( 1975). Evidence that insect dorsal unpaired median ( D U M ) neurons arc octopaminergic. J. E . Y ~ . Zool. 193:425-43 1.
HOYLE. G . ( 1974). A function for neurons ( D U M ) neu- rosecretory on skeletal muscles of insects. ./. E.\p. Zool. 189:40 1-406.
HOYLE, G. and BARKER, D. L. ( 1975). Synthesis of oc- topamine by insect dorsal median unpaired neurons. J . E,Y~. Zool. 193:433-439.
HOYLE, G. and DAGAN. D. ( 1978). Physiological char- acteristics and reflex activation of DUM (octopamin- ergic) neurons of the locust metathoracic ganglion. ./. Neiruohiol. 959-79.
HOYLE, G., COLQUHOUN, W.. and WILLIAMS. M. (1980). Fine structure of an octopaminergic neuron and its terminals. ./. Ncirrohiol. 11: 103- 126.
KATZ, P. and HARRIS-WARRICK, R. M. ( 1990). Neuro- modulation of the crab pyloric central pattern genera- tor by serotonergic/cholinergic proprioceptive affer- ents. J. Nmrosci. 10: 1495- 15 1 2.
KINNAMON. S. C., KLAASSEN, L. W., KAMMER, A. E., and CLAASSEN, D. ( 1984). Octopamine and chlordi- meform enhance sensory responsiveness and produc- tion of the flight motor pattern in developing and adult moths. .J. Ncirrohiol. 15983-293.
LAKE, C. R., MILLS. R. R., and BRUNET. P. C. J. ( 1970). beta-Hydroxylation of tyramine by cockroach haemo- lymph. Biochitn. Bioplz~*.s. Acts 2 15:226-228.
LANG, A. B., ORCHARD. I., and BARNETT, F. M. ( 1989). Changes in haemolymph serotonin levels associated with feeding in the blood-sucking bug. Rlzodniiis pro- l i s i i s . J . Insect Plrj:ciol. 35393-399.
MARDER, E. and EISEN, J. S. (1984). Electrically cou- pled pacemaker neurons respond differently to same physiological inputs and neurotransmitters. J. Neirro- physiol. 5 I : 1 362- 1 374.
MARTIN, R. J., BAILEY, B. A,, and DOWNER, R. G. H. ( 1984). In: Neirrohiologj~ (?/' Truce ..lminr.s. A. A. Boulton, G. B. Baker, W. G . Dewhurst, and M. Sandler, Eds. Humana Press, Clifton, NJ. pp. 9 1.
MAXWELL, G. D., TAIT, J . F.. and HILDEBRAND, J. G.
4: 1976- 1993.

( 1978). Regional synthesis of neurotransmitter candi- dates in the CNS of the moth Mundiicu .wm. Comp. Biochcm. P/ij..~iol. 6 1C: 109- I 19.
MORTON. D. B. and EVANS. P. D. ( 1984). Octopamine release from an identified neurone in the locust. .I. Ekp. Biol. 113269-287.
NATHANSON, J. A. and GREENGARD, P. ( 1973). Octo- pamine sensitive adenylate cyclase: evidence for a bio- logical role of octopamine in nervous tissue. Science 180:308-310.
NATHANSON, J. A. and GREENGARD, P. ( 1974). Seroto- nin-sensitive adenylate cyclases in neural tissue and its similarity to the serotonin receptor: a possible site of action of lysergic acid diethylamide. Proc. Nutl. Acud.
ORCHARD, I. ( 1982). Octopamine in insects: neurotrans- mitter, neurohormone. and neuromodulator. Cun. J . Zool. 60559-669.
ORCHARD, I. ( 1989). Serotonergic neurohaemal tissue in Rhodniirs pro/ixir.\: synthesis, release and uptake of serotonin. .I. In.rcct Plrj~.siol. 35:943-947.
ORCHARD, 1.. LOUGHTON. B. G., and WEBB, R. A. ( I98 1 ). Octopamine and short-term hyperlipaemia in the locust. Gcn. C'omp. Endocrinol. 45: 175- 1 80.
O'SHEA. M. and EVANS, P. D. (1979). Potentiation of neuromuscular transmission by an octopaminergic neurone in the locust. J . Exp. Biol. 79: 169- 190.
O'SHEA. M. and MURPHEY, R. K. ( 1978). Octopamine modulates sensitivity of identified insect interneurons. Soi~. h c i r rosci. .,I h s. 4 : 2 0 3 .
RAMIREZ. J. -M. and ORCHARD. I. ( 1990). Octopamin- ergic modulation of the forewing stretch receptor in the locust Locirstn migrutoriu. .I. E .Y~. Biol. 149:255- 279.
RAMIREZ. J . -M. and PEARSON, K. G. ( 1991a). Octopa- mine induces bursting and plateau potentials in insect neurones. Bruin Res. 549:332-337.
RAMIREZ. J . -M. and PEARSON, K. G. ( 1991b). Octopa- minergic modulation of interneurons in the flight sys- tem of the locust. .I. A'c~iiro~~li!sio/. 66: 1522- 1537.
RICHTER. D. and RUTSCHKE. E. ( 1977). Localization of monoamine oxidase ( M A O ) in the brain of Periplun- oiu umrric,unu ( L.). k t u . His todim. 60:304-3 1 1.
RITZMANN, R. E. and POLLACK, A. J. (1988). Wind activated thoracic interneurons of the cockroach: 11. Patterns of connection from ventral giant interneu- rons. J . Neirrohiol. 19589-6 1 1.
RITZMANN. R. E. and POLLACK, A. J. ( 1990). Parallel motor pathways from thoracic interneurons of the ventral giant interneuron system of the cockroach. Pmiifuncta uincricunu. J . Neiirohiol. 21: 1219-1235.
RITZMANN, R. E.. POLLACK, A. J., HUDSON, S., and HY- VONEN, A. ( 199 1 ). Thoracic interneurons in the escape system of the cockroach, Periplunetu ameri- c'unu. are multi-modal. Bruin Rcs. 563: 175-1 83.
.%.i. US.,1 71:797-780.
ROBERTSON, H. A. and STEELE, H. E. ( 1973). Octopa- mine in the insect central nervous system: distribu- tion, biosynthesis and possible physiological role. .I. Phj..siol. 237:34-35 P.
ROBERTSON, R. M. and PEARSON, K. G . (1983). Inter- neurons in the flight system of the locust: distribution. connections, and resetting properties. J. Comp. A1euro/. 21533-50.
SELVERSTON, A. 1. (Ed.). ( 1985). Model Neiirul N c - works und Behavior. Plenum, New York.
SHAFI, N., MIDGLEY, J. M., WATSON, D. G., SMAIL. G. A., STRANG, R.. and MACFARLANE, R. G. (1989). Analysis of biogenic amines in the brain of the ameri- can cockroach ( Periplunctu umericuna) by gas chro- matography-negative ion chemical ionisation mass spectroscopy. .I. Chrom7atogr. 490:9- 19.
SLOLEY. B. D. and OWEN, M. D. ( 1982). The effects of reserpine on amine concentrations in the nervous sys- tem of the cockroach ( PLv?plunrtu umcricnnu). Insect Biochem. 12:469-476.
SOMBATI, S. and HOYLE, G. (1984a). Central nervous sensitization and dishabituation of reflex action in an insect by the neuromodulator octopamine. J. Neziro- hiol. 15455-480.
SOMBATI, S . and HOYLE, G. ( 1984b). Generation of spe- cific behaviors in a locust by local release into neuropil of the natural neuromodulator octopamine. J . Ncicro- hiol. 1 5 4 8 1-506.
STEVENSON. P. ( 1989). Central effects of octopamine in the locust. In: Neiirul Mecliuni.~rn.c,fi)r Rohuvior: Pro- twrfings of ilrr 2nd Intcmui ionul Cbngress ~ ? ~ ~ ~ ~ ~ ~ r o ~ i t ~ i - o/ogj3. J . Erber, H. -J. Pfluger, and D. Todt. Eds. Theime, New York, pp. 23 I .
TAGHERT, P. H. and GOODMAN. C. S. ( 1984). Cell deter- mination and differentiation of identified serotonin- immunoreactive neurons in the grasshopper embryo. .J. Nezrrosci. 4:989- 1000.
TWAROG, B. M. and ROEDER, K. D. ( 1957). Pharmaco- logical observations on the desheathed last abdominal ganglion of the cockroach. Ann. Ent. Soc. Amer.
WAFFORD, K. A. and SATTELLE, D. B. ( 1986). Effects of amino acid neurotransmitter candidates on an identi- fied insect motoneurone. Ncirrosci. Left. 6: 135-140.
WATSON. A. H. D. and BURROWS, M. ( 1982). The ultra- structure of identified locust motor neurones and their synaptic relationships. .J. C'omp. Nezrrol. 205383- 397.
WESTIN, J., RITZMANN, R. E.. and GODDARD, D. J. ( 1988). Wind-activated thoracic interneurons of the cockroach: I. Responses to controlled wind stimula- tion. J. Neiirohiol. 19:573-588.
WHITEHEAD. D. L. ( 1969). New evidence for the control mechanisms of schlerotization in insects. Nutirrr 22472 1-123.
50:23 1-236.





























