Embed Size (px)
Citation preview

ES41CH06-Shiganova ARI 1 October 2010 14:33
Biotic Homogenizationof Inland Seas of thePonto-CaspianTamara ShiganovaP.P. Shirshov Institute of Oceanology, Russian Academy of Sciences, 117997 Moscow, Russia;email: [email protected]
Annu. Rev. Ecol. Evol. Syst. 2010. 41:103–25
First published online as a Review in Advance onAugust 2, 2010
The Annual Review of Ecology, Evolution, andSystematics is online at ecolsys.annualreviews.org
This article’s doi:10.1146/annurev.ecolsys.110308.120148
Copyright c© 2010 by Annual Reviews.All rights reserved
1543-592X/10/1201-0103$20.00
Key Words
algae, ballast water, canal construction, disturbance, ecosystemfunctioning, fishes, invasions, invertebrates, non-native species, seas andoceans, shipping intensity
Abstract
Comparative analysis of patterns of species invasion, dispersal, and impacton ecosystems in the Ponto-Caspian seas has been ongoing since 1900. TheBlack Sea is an important international shipping destination. High shippingintensity has facilitated species invasions into the Black Sea. Many specieshave successfully established because of Black Sea disturbances. The BlackSea serves as a hub for species that then spread further to the Sea of Azov,the Caspian Sea, the Sea of Marmara, and in some cases the eastern Mediter-ranean Sea. The Black Sea has thus become the main recipient for non-nativespecies and acts as a donor to the seas of Eurasia. Native biodiversity has de-clined and invaders now dominate the Ponto-Caspian seas. This process hascaused biotic homogenization of the Ponto-Caspian, as the same invadersdetermine community structure and in some cases ecosystem functioning inthe different seas. Among these invaders, gelatinous species have become themain drivers of ecosystem functioning. The invasion rate of new species isaccelerating.
103
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Non-native species:organism introducedby anthropogenicmeans to a sea inwhich it did notpreviously occur andcould establish aself-sustainingpopulation
1. INTRODUCTION
Ecosystems of seas, particularly inland seas, are changing at an alarming rate. These changes haveaccelerated dramatically in the past few decades owing to new technology and increased shippingintensity, with expanded shipping routes globally and increased basin connectivity. These eventshave facilitated invasions of aquatic species from their native ranges to distant locations. Everyday, many non-native species are being transported around the world (Carlton et al. 1999, Rilov& Crooks 2009). The need to understand the causes and impacts of these alien invasions has neverbeen more urgent, and the development of effective responses to existing and threatened invasionsis a critical issue. Using the Ponto-Caspian basin as a case study, we try to analyze which marineecosystems have been vulnerable to invasions and which species are able to overcome ecologicalbarriers and establish in new areas. We attempt to track the process of non-native species invasions,the growth of invading populations, the details of their geographical range expansion, and theirroles and biotic interactions in recipient ecosystems.
The Black, Azov, and Caspian Seas (Ponto-Caspian) were united as a single basin several timesin the past, most recently in the Pliocene, when they were connected in the almost freshwaterPontian Lake–Sea. The marine biota was eliminated and a brackish water biota then formed.Its representatives still dwell in the Caspian Sea, in the Sea of Azov, and in desalinated regionsof the northwestern Black Sea; these are referred to as Ponto-Caspian species. All three seaswere reconnected again by the Volga–Don Canal in 1952. The Black Sea is also a part of theMediterranean basin and is connected via the Bosporus Strait with the Sea of Marmara and furtherby the Dardanelles Strait with the Mediterranean Sea. Owing to accelerating human activities suchas shipping, deliberate stocking, unintentional releases, and canal construction, many non-nativespecies have arrived and established in these seas. After construction of ballast water tanks in shipsthis process became global. In addition, since the 1980s, warming of the upper water layer ofthe Black Sea has led to increased populations of thermophilic species and northward expansionof their ranges. The Black Sea has thus become the main recipient for non-native marine andbrackish water species and acts as a donor to the other Ponto-Caspian seas.
An important detrimental role of the whole Ponto-Caspian basin is dispersal of its native brack-ish water Ponto-Caspian species outside their geographical range, because of canals connectingpreviously separated waters and accelerating shipping intensity (Ketelaars 2004, Mills et al. 1993).
2. COMMON ORIGIN OF THE PONTO-CASPIAN AND MAINFEATURES OF ENVIRONMENT AND BIOTA
The three seas are all temperate basins characterized by lower salinity than standard ocean values.All basins are isolated from the World Ocean, and all seas have low biodiversity but high pro-ductivity (Table 1). The physical evolution of the marine environments on the southern flanks ofEurope and Asia during the Tertiary and Quaternary eras affected their biotas in many ways. Theparamount variable causing biotic change was salinity, which fluctuated strongly. This turbulenthistory generated the heterogeneity of the regional fauna and flora.
The present biotas of the seas are largely relicts, the remains of an ancient Tethys fauna(including Sarmatian, Pontian, and Caspian species) after it was exposed to repeated alternation ofdesalination and salination phases. This biota is supplemented by comparatively recent invadersof freshwater origin and by immigrants (pseudorelics) from the Arctic. The latter worked theirway south via the Volga River and are therefore mainly restricted to the Caspian Sea. In addi-tion, Atlantic-Mediterranean elements advanced from the west through the Mediterranean withdecreasing forms from one sea to another in an eastward direction.
104 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.
ES41CH06-Shiganova ARI 1 October 2010 14:33
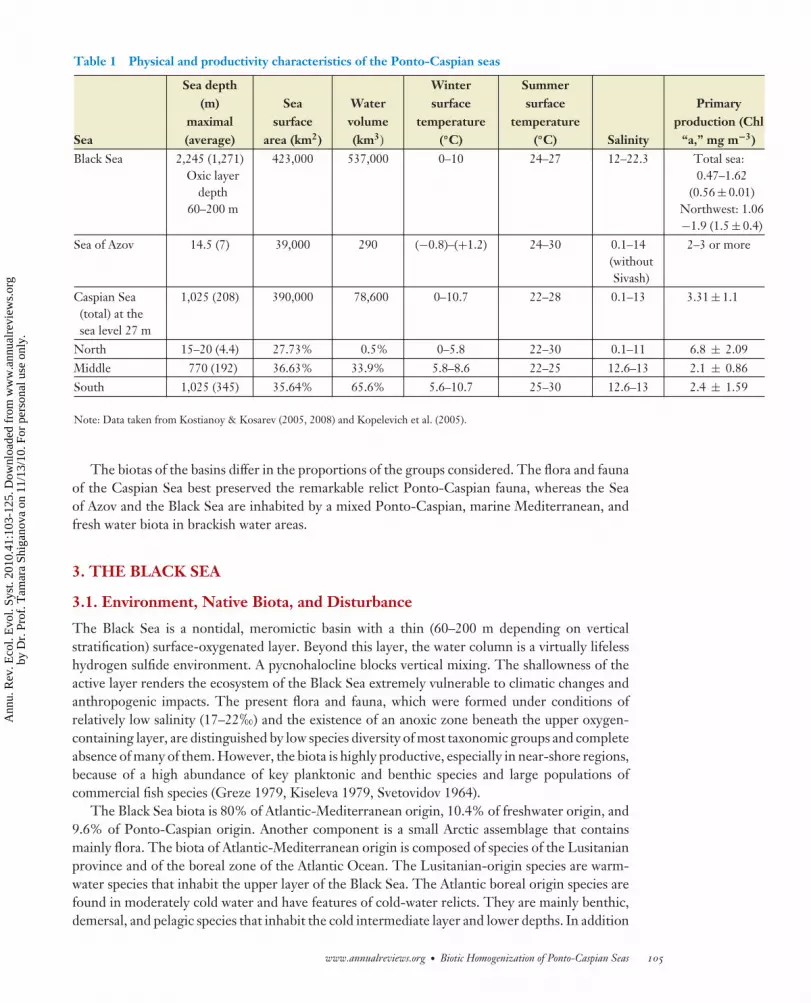
Table 1 Physical and productivity characteristics of the Ponto-Caspian seas
Sea
Sea depth(m)
maximal(average)
Seasurface
area (km2)
Watervolume(km3)
Wintersurface
temperature(◦C)
Summersurface
temperature(◦C) Salinity
Primaryproduction (Chl
“a,” mg m−3)Black Sea 2,245 (1,271)
Oxic layerdepth
60–200 m
423,000 537,000 0–10 24–27 12–22.3 Total sea:0.47–1.62
(0.56 ± 0.01)Northwest: 1.06−1.9 (1.5 ± 0.4)
Sea of Azov 14.5 (7) 39,000 290 (−0.8)–(+1.2) 24–30 0.1–14(withoutSivash)
2–3 or more
Caspian Sea(total) at thesea level 27 m
1,025 (208) 390,000 78,600 0–10.7 22–28 0.1–13 3.31 ± 1.1
North 15–20 (4.4) 27.73% 0.5% 0–5.8 22–30 0.1–11 6.8 ± 2.09Middle 770 (192) 36.63% 33.9% 5.8–8.6 22–25 12.6–13 2.1 ± 0.86South 1,025 (345) 35.64% 65.6% 5.6–10.7 25–30 12.6–13 2.4 ± 1.59
Note: Data taken from Kostianoy & Kosarev (2005, 2008) and Kopelevich et al. (2005).
The biotas of the basins differ in the proportions of the groups considered. The flora and faunaof the Caspian Sea best preserved the remarkable relict Ponto-Caspian fauna, whereas the Seaof Azov and the Black Sea are inhabited by a mixed Ponto-Caspian, marine Mediterranean, andfresh water biota in brackish water areas.
3. THE BLACK SEA
3.1. Environment, Native Biota, and Disturbance
The Black Sea is a nontidal, meromictic basin with a thin (60–200 m depending on verticalstratification) surface-oxygenated layer. Beyond this layer, the water column is a virtually lifelesshydrogen sulfide environment. A pycnohalocline blocks vertical mixing. The shallowness of theactive layer renders the ecosystem of the Black Sea extremely vulnerable to climatic changes andanthropogenic impacts. The present flora and fauna, which were formed under conditions ofrelatively low salinity (17–22�) and the existence of an anoxic zone beneath the upper oxygen-containing layer, are distinguished by low species diversity of most taxonomic groups and completeabsence of many of them. However, the biota is highly productive, especially in near-shore regions,because of a high abundance of key planktonic and benthic species and large populations ofcommercial fish species (Greze 1979, Kiseleva 1979, Svetovidov 1964).
The Black Sea biota is 80% of Atlantic-Mediterranean origin, 10.4% of freshwater origin, and9.6% of Ponto-Caspian origin. Another component is a small Arctic assemblage that containsmainly flora. The biota of Atlantic-Mediterranean origin is composed of species of the Lusitanianprovince and of the boreal zone of the Atlantic Ocean. The Lusitanian-origin species are warm-water species that inhabit the upper layer of the Black Sea. The Atlantic boreal origin species arefound in moderately cold water and have features of cold-water relicts. They are mainly benthic,demersal, and pelagic species that inhabit the cold intermediate layer and lower depths. In addition
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 105
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
to salinity, impoverishment of the Black Sea biota is due to the absence of deep-water species atdepths below the oxic layer. The number of species in the Black Sea is presently relatively small:3,786. Of these, 1,619 are fungi, algae, and higher plants; 1,983 are invertebrates, 180 are fish,and 4 are mammals (Mordukhai-Boltovskoi 1969, Zaitsev & Mamaev 1997).
Since the 1970s, under the influence of climatic and anthropogenic factors, the biota hasgreatly changed. Among the most pronounced anthropogenic drivers are regulation of riverrunoff and an increased supply of dissolved phosphates and nitrates accompanied by reducedsilicate from great rivers. Changes include a switch in phytoplankton domination from diatomsto dinoflagellates, a significant increase in phytoplankton biomass, and massive development ofharmful algae, with subsequent eutrophication, corresponding to a two-fold increase in primaryproduction for the sea as a whole and a ten-fold increase in the northwestern part, and sub-sequent explosions of native gelatinous species such as the scyphomedusa Aurelia aurita (L.)and the dinoflagellate Noctiluca scintillans (Macartney) Kofoid et Swezy 1921 (Petranu 1997).In addition, fishing pressure resulted in decreasing stocks of the top predators: large pelagicfishes (migrants from the Aegean and Marmara seas) and dolphins. The Black Sea thus becamemesotrophic or eutrophic (in its northwestern parts), which facilitated invasion by non-nativespecies.
3.2. Vectors, Pathways, and Composition of Invaders
Among the Ponto-Caspian seas, only in the Black Sea has shipping accelerated, with routes todifferent regions of the World Ocean. Since the late twentieth century, enormous numbers ofmarine and brackish water species have arrived in its harbors in ballast water and on fouled hulls.Disturbance has facilitated their establishment. Biotic changes have also been caused by otherhuman activities: release by aquarists, deliberate stocking of commercial species and release ofspecies that accompany them, and penetration via canal systems connecting previously isolatedbasins. During recent decades, a new vector has accelerated expansion of warm-water species fromthe Sea of Marmara and Mediterranean Sea owing to rising upper-layer temperatures (Shiganova& Ozturk 2010).
Pathways of species penetration to the Black Sea based on an analysis of established non-native species and their donor regions are shown in Figures 1a,b and 2. The greatest numberof species (36%) arrived from the Mediterranean as a result of natural range expansion or inballast water (Figure 2). This proportion keeps increasing with rising upper-layer water temper-atures. Only a few of these species have become abundant and almost none have reached otherPonto-Caspian seas. Among these species are representatives of micro-, phyto- and zooplankton,demersal plankton, macrophytes, zoobenthos, and fishes. These species as a rule have subtropicalor sometimes even tropical origins; most still occur only in the southern parts of the Black Sea andnear the Bosporus Strait where salinity is higher. They are not considered to be established in theBlack Sea in this review (Table 2). Certain species have penetrated to the near-shore regions offBulgaria, Romania, and Ukraine (Odessa, Crimea) in the course of their migrations or have beencarried in currents or ballast water.
By contrast, some species of Adriatic origin (which compose 2% of the non-native speciesin the Black Sea) now have self-sustaining populations and are abundant. Chief among them is abivalve, Anadara inaequivalvis (Bruguiere 1789), which established in most coastal areas and spreadto the Sea of Azov. The successful establishment of propagules from the Adriatic Sea is explainedby the lower salinity of some Adriatic regions compared to other parts of the Mediterranean Seaand by the fact that Adriatic water temperatures are similar to those of the Black Sea. In addition,shipping traffic between ports of the Adriatic and Black Seas favors introductions.
106 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Lepomis gibbosusLepomis gibbosusUrnatella gracillisUrnatella gracillisLepomis gibbosusUrnatella gracillis
Rhithropanopeus harrisiiRhithropanopeus harrisiiGambusia holbrookiGambusia holbrooki
Rhithropanopeus harrisiiGambusia holbrooki
Eriocheirsinensis
Crassostreagigas
Ficopomatusenigmaticus
Anadarainaequivalvis
Potomopyrgusjenkinsi
a
b
Figure 1.Scheme of the donor areas of invasions into the Black Sea. (a) Primary pathway of selected species; (b) donorareas and scheme of pathways of invasion. Arrow colors correspond to geographic areas referenced inFigure 2.
Many established invaders from North American Atlantic regions were introduced mainly inthe 1980s–1990s (11% of all non-native species). Species of this group are mostly temperate,rather eurythermal and, importantly, euryhaline and widely distributed in coastal waters of theWorld Ocean. Invaders from this region have greatly affected the Black Sea. Among these arethe ctenophores Mnemiopsis leidyi (A. Agassiz 1865) and Beroe ovata (sensu Mayer 1912), and thecopepod Acartia tonsa (Dana 1849). The barnacles Balanus eburneus (Gould 1841) and B. improvisus(Darwin 1854) were introduced from the same area much earlier. This pathway also characterizesaccidentally introduced brackish water species represented by inhabitants of brackish bays andestuaries. The most successful of these are the hydromedusas Blackfordia virginica (Mayer 1910)and Bougainvillia (Perigonimus) megas (Kinne 1956).
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 107
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.
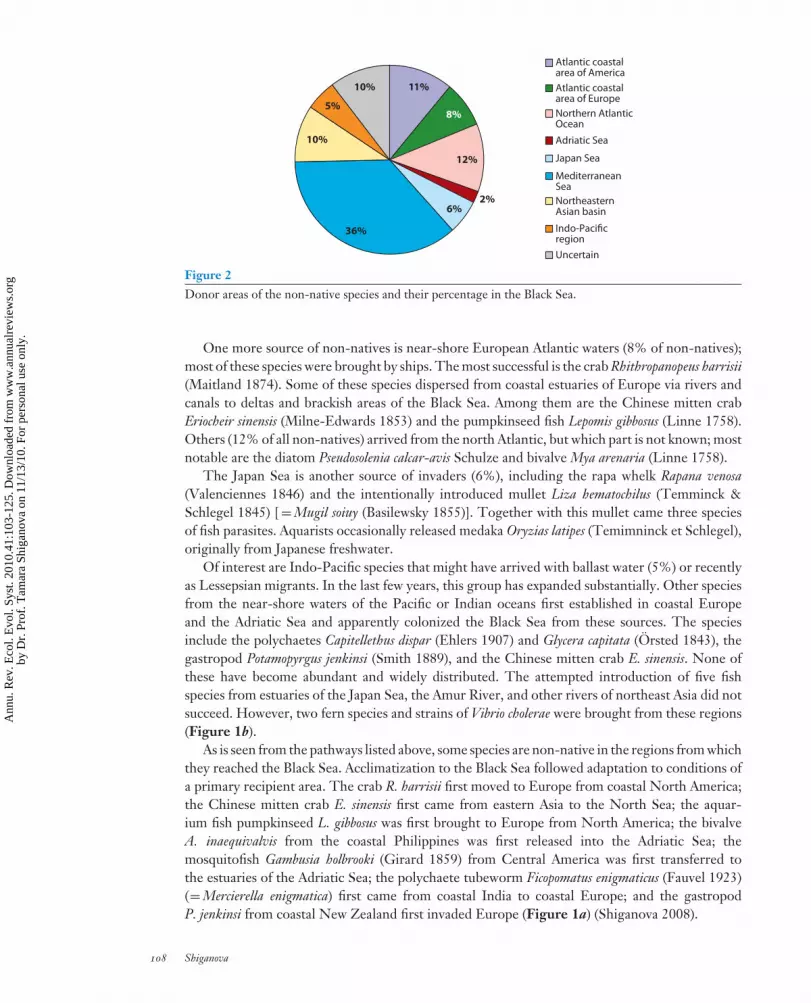
ES41CH06-Shiganova ARI 1 October 2010 14:33
11%
8%
12%
2%6%
36%
10%
5%
10%
Atlantic coastalarea of America
Atlantic coastalarea of Europe
Northern AtlanticOcean
Adriatic Sea
Japan Sea
MediterraneanSea
NortheasternAsian basin
Indo-Pacificregion
Uncertain
Figure 2Donor areas of the non-native species and their percentage in the Black Sea.
One more source of non-natives is near-shore European Atlantic waters (8% of non-natives);most of these species were brought by ships. The most successful is the crab Rhithropanopeus harrisii(Maitland 1874). Some of these species dispersed from coastal estuaries of Europe via rivers andcanals to deltas and brackish areas of the Black Sea. Among them are the Chinese mitten crabEriocheir sinensis (Milne-Edwards 1853) and the pumpkinseed fish Lepomis gibbosus (Linne 1758).Others (12% of all non-natives) arrived from the north Atlantic, but which part is not known; mostnotable are the diatom Pseudosolenia calcar-avis Schulze and bivalve Mya arenaria (Linne 1758).
The Japan Sea is another source of invaders (6%), including the rapa whelk Rapana venosa(Valenciennes 1846) and the intentionally introduced mullet Liza hematochilus (Temminck &Schlegel 1845) [ = Mugil soiuy (Basilewsky 1855)]. Together with this mullet came three speciesof fish parasites. Aquarists occasionally released medaka Oryzias latipes (Temimninck et Schlegel),originally from Japanese freshwater.
Of interest are Indo-Pacific species that might have arrived with ballast water (5%) or recentlyas Lessepsian migrants. In the last few years, this group has expanded substantially. Other speciesfrom the near-shore waters of the Pacific or Indian oceans first established in coastal Europeand the Adriatic Sea and apparently colonized the Black Sea from these sources. The speciesinclude the polychaetes Capitellethus dispar (Ehlers 1907) and Glycera capitata (Orsted 1843), thegastropod Potamopyrgus jenkinsi (Smith 1889), and the Chinese mitten crab E. sinensis. None ofthese have become abundant and widely distributed. The attempted introduction of five fishspecies from estuaries of the Japan Sea, the Amur River, and other rivers of northeast Asia did notsucceed. However, two fern species and strains of Vibrio cholerae were brought from these regions(Figure 1b).
As is seen from the pathways listed above, some species are non-native in the regions from whichthey reached the Black Sea. Acclimatization to the Black Sea followed adaptation to conditions ofa primary recipient area. The crab R. harrisii first moved to Europe from coastal North America;the Chinese mitten crab E. sinensis first came from eastern Asia to the North Sea; the aquar-ium fish pumpkinseed L. gibbosus was first brought to Europe from North America; the bivalveA. inaequivalvis from the coastal Philippines was first released into the Adriatic Sea; themosquitofish Gambusia holbrooki (Girard 1859) from Central America was first transferred tothe estuaries of the Adriatic Sea; the polychaete tubeworm Ficopomatus enigmaticus (Fauvel 1923)(= Mercierella enigmatica) first came from coastal India to coastal Europe; and the gastropodP. jenkinsi from coastal New Zealand first invaded Europe (Figure 1a) (Shiganova 2008).
108 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.
ES41CH06-Shiganova ARI 1 October 2010 14:33
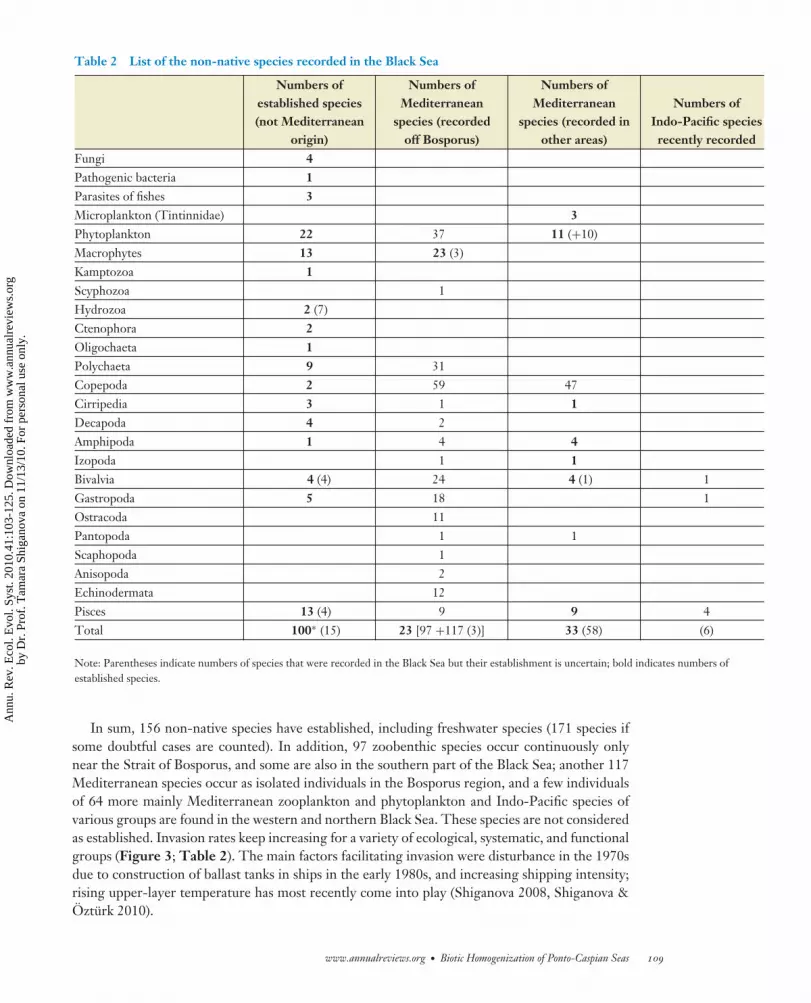
Table 2 List of the non-native species recorded in the Black Sea
Numbers ofestablished species(not Mediterranean
origin)
Numbers ofMediterranean
species (recordedoff Bosporus)
Numbers ofMediterranean
species (recorded inother areas)
Numbers ofIndo-Pacific speciesrecently recorded
Fungi 4Pathogenic bacteria 1Parasites of fishes 3Microplankton (Tintinnidae) 3Phytoplankton 22 37 11 (+10)Macrophytes 13 23 (3)Kamptozoa 1Scyphozoa 1Hydrozoa 2 (7)Ctenophora 2Oligochaeta 1Polychaeta 9 31Copepoda 2 59 47Cirripedia 3 1 1Decapoda 4 2Amphipoda 1 4 4Izopoda 1 1Bivalvia 4 (4) 24 4 (1) 1Gastropoda 5 18 1Ostracoda 11Pantopoda 1 1Scaphopoda 1Anisopoda 2Echinodermata 12Pisces 13 (4) 9 9 4Total 100∗ (15) 23 [97 +117 (3)] 33 (58) (6)
Note: Parentheses indicate numbers of species that were recorded in the Black Sea but their establishment is uncertain; bold indicates numbers ofestablished species.
In sum, 156 non-native species have established, including freshwater species (171 species ifsome doubtful cases are counted). In addition, 97 zoobenthic species occur continuously onlynear the Strait of Bosporus, and some are also in the southern part of the Black Sea; another 117Mediterranean species occur as isolated individuals in the Bosporus region, and a few individualsof 64 more mainly Mediterranean zooplankton and phytoplankton and Indo-Pacific species ofvarious groups are found in the western and northern Black Sea. These species are not consideredas established. Invasion rates keep increasing for a variety of ecological, systematic, and functionalgroups (Figure 3; Table 2). The main factors facilitating invasion were disturbance in the 1970sdue to construction of ballast tanks in ships in the early 1980s, and increasing shipping intensity;rising upper-layer temperature has most recently come into play (Shiganova 2008, Shiganova &Ozturk 2010).
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 109
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
0
5
10
15
20
25
30
< XX cen
Num
ber o
f spe
cies
Macrophytes Zoobenthos Zooplankton Phytoplankton
Microplankton Marine fungi Fish
Start of disturbance
Ballast tank construction
Temperature rising
2001–2006 1981–20001961–19801941–19601921–19401900–1920
Figure 3Chronology of species invasions into the Black Sea.
The proportions of the freshwater, brackish water, and euryhaline species, respectively, in thenon-native biota almost equal those within the native species (Figure 4). Many marine euryhalinespecies have established and are capable of occupying the entire sea. They compose 89% or 74%of the total non-native biota, depending on whether we include those species found only nearthe Bosporus. Brackish water species have established in the brackish northwestern area, andfreshwater species have established in the river mouths, bays, and estuaries.
Most planktonic euryhaline, eurythermal species have established and occur in the upper layer;several of them may temporarily penetrate below the thermocline. Other planktonic invaders arewarm-water species, which develop only during the warm seasons and inhabit the upper layer abovethe thermocline. Benthic non-native species are also represented by species with broad tolerancesfor various physical factors. The similarities in the proportions of native and non-native biotas ofthe various sorts might be due to the fact that, when there are stressful limiting environmentalfactors (low salinity, sharply continental climate, a virtually lifeless hydrogen sulfide layer below
80%
9.6%
10.4%a
Euryhaline Brackish water Fresh water
74%
22%
4%b
Figure 4(a) Ratio (%) of euryhaline (Atlantic-Mediterranean), brackish water (Ponto-Caspian), and freshwaterspecies in native biota; (b) ratio (%) of non-native euryhaline, brackish water, freshwater species of differentorigin in the Black Sea (without near Bosporus species).
110 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
60–200 m, etc.), only ecologically similar species can establish, but those are more competitivethan the natives.
3.3. Ecosystem Impacts
Most negative effects are caused by invaders that form massive populations. These include benthicspecies (e.g., the rapa whelk, which consumes oysters, mussels, and other bivalves), pelagic species(e.g., M. leidyi, which grazes on zooplankton, fish eggs, and larvae and affects all trophic levels), andfresh and brackish water species (e.g., the pumpkinseed fish L. gibbosus, which preys on zooplankton,fish eggs, larvae, fry, and small adult fish).
However, at the same time the rapa whelk has become a valuable commercial species. The mostpronounced event was the invasion by M. leidyi, which caused cascading effects at higher trophiclevels, from a decrease in zooplankton to collapsing planktivorous fish and drastic declines amonglarge pelagic fish and dolphins. Similar effects occurred at lower trophic levels, with a decreasein zooplankton leading to an increase in phytoplankton released from zooplankton grazing pres-sure, and increases in bacterioplankton, which led to increases in their predators, zooflagellates,and ciliates. Ten years later another warm-water ctenophore, B. ovata, an obligate predator onM. leidyi, was introduced via ballast water from North America (Shiganova et al. 2004a). Its in-vasion has been an example of invasion meltdown (Simberloff & Von Holle 1999). In this case,invasion of prey favored establishment of its predator. The B. ovata invasion reduced M. leidyipopulations and the ecosystem began to recover at all trophic levels.
The cholera bacterium V. cholerae may provoke epidemics from time to time in coastal popu-lated areas. The P. calcar-avis diatom is an additional food item for large zooplankton species, butwhen its populations explode, it supplants more valuable native phytoplankton species. The role ofthe fouling species B. improvisus is negative; nevertheless its larvae are consumed by small pelagicfishes. The introduction of other organisms may be regarded as positive from some perspectives.The bivalves A. inaequivalvis and M. arenaria, though replacing native species, constitute valuablefood for zoobenthic species and benthophagous fishes, while their larvae are consumed by smallpelagic fishes. The crab R. harrisii has become an additional food for benthophagous fishes. Themullet L. hematochilus competes with native mullets but is itself a valuable commercial species.Most established non-native species are temperate, euryhaline, and eurythermal. Species with allof these traits not only established but also became explosive and expanding, first reaching the Seaof Azov and the Caspian Sea; some then invaded the Sea of Marmara, the Aegean and AdriaticSeas, and other areas of the Mediterranean Sea. Some species, such as the rapa whelk, were widelydispersed via ballast water from the Black Sea to some areas of the Mediterranean and along thenorthern and southern Atlantic Ocean (Chandler et al. 2008). Thus, the Black Sea has become asecondary donor for the expansion of non-native species.
4. THE SEA OF AZOV
4.1. Environment, Native Biota, and Disturbance
The Sea of Azov is a remnant of an ancient system of straits that formerly occupied the Caucasianforedeep. It is the smallest and shallowest among the southern Eurasian seas, with salinity 0.1–14�, and connects with the Black Sea by the narrow, shallow Kerch Strait. The history of its originand evolution is closely related to that of the Black Sea. The biota of the Sea of Azov was formedfrom representatives of freshwater, brackish water, and marine assemblages. The freshwater andbrackish water assemblages are mainly represented by Ponto-Caspian relics that populate rivers
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 111
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.
ES41CH06-Shiganova ARI 1 October 2010 14:33
and lagoons of the Sea of Azov and consist of 54 species in 32 genera. The bulk of the relics areconcentrated near river mouths, where the water is virtually fresh. The small marine assemblageis represented by Mediterranean flora and fauna. The biota includes 350 species of free-livinginvertebrates (without turbellaria and nematodes), 605 species of phytoplankton, 30 species ofmacrophytes, and 103 fish species. For well-studied groups, the Black Sea biota is 3.4 times as richas that of the Sea of Azov. Not only does the number of species decrease from the Black Sea to theSea of Azov, but the proportions of groups change as well. Many species with a Mediterraneanorigin that presently dwell in the north Atlantic disappear (Mordukhai-Boltovskoi 1960, 1969).
The regulation of the Don (1952) and Kuban (1973) Rivers and the withdrawal of river runoffto fill reservoirs reduced flooded and spawning areas. In the sea proper, one observed a growthin the vertical temperature and salinity gradients and an increase in the oxygen-deficient zonesnear the bottom. Increased salinity spurred dispersal of Black Sea species into the Sea of Azov;most of these species disappeared after salinity decreased to the original level, but a few persisted(Kosarev et al. 2008).
4.2. Vectors, Pathways, and Composition of Invaders
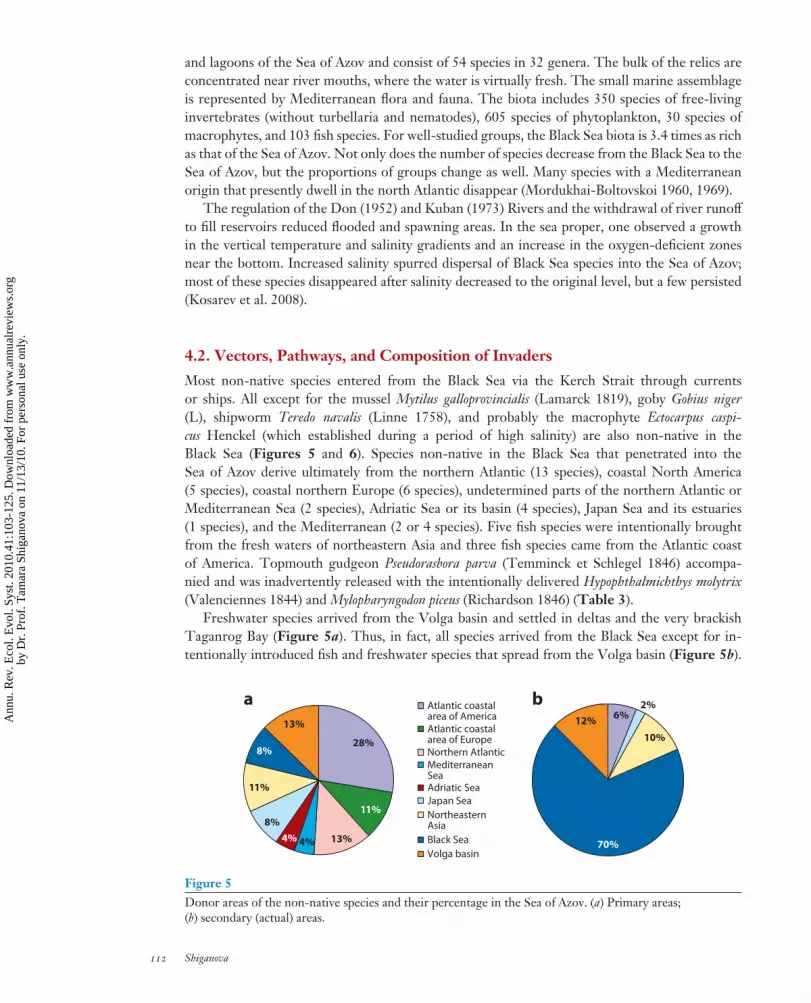
Most non-native species entered from the Black Sea via the Kerch Strait through currentsor ships. All except for the mussel Mytilus galloprovincialis (Lamarck 1819), goby Gobius niger(L), shipworm Teredo navalis (Linne 1758), and probably the macrophyte Ectocarpus caspi-cus Henckel (which established during a period of high salinity) are also non-native in theBlack Sea (Figures 5 and 6). Species non-native in the Black Sea that penetrated into theSea of Azov derive ultimately from the northern Atlantic (13 species), coastal North America(5 species), coastal northern Europe (6 species), undetermined parts of the northern Atlantic orMediterranean Sea (2 species), Adriatic Sea or its basin (4 species), Japan Sea and its estuaries(1 species), and the Mediterranean (2 or 4 species). Five fish species were intentionally broughtfrom the fresh waters of northeastern Asia and three fish species came from the Atlantic coastof America. Topmouth gudgeon Pseudorasbora parva (Temminck et Schlegel 1846) accompa-nied and was inadvertently released with the intentionally delivered Hypophthalmichthys molytrix(Valenciennes 1844) and Mylopharyngodon piceus (Richardson 1846) (Table 3).
Freshwater species arrived from the Volga basin and settled in deltas and the very brackishTaganrog Bay (Figure 5a). Thus, in fact, all species arrived from the Black Sea except for in-tentionally introduced fish and freshwater species that spread from the Volga basin (Figure 5b).
28%
11%
13%4%4%8%
11%
8%
13%
Atlantic coastalarea of America Atlantic coastalarea of Europe Northern AtlanticMediterraneanSeaAdriatic Sea
Japan Sea
NortheasternAsia
Black Sea
Volga basin70%
12%
10%
2%6%
a b
Figure 5Donor areas of the non-native species and their percentage in the Sea of Azov. (a) Primary areas;(b) secondary (actual) areas.
112 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Increase salinity
0
5
10
15
20
< XX cen
Num
ber o
f spe
cies
Macrophytes Zoobenthos Zooplankton
Phytoplankton Microplankton Fish
Ballast tank construction
2001–20081981–20001961–19801941–19601921–19401900–1920
Figure 6Chronology of species invasions into the Sea of Azov.
There are 1.6 times more euryhaline species in the non-native biota than in the native biota(Figure 7). The distribution of non-native species in terms of salinity tolerance is similar to thatof seasonal migrants from the Black Sea. Some of them (rapa whelk, blue crab Callinectes sapidusRathbun 1896) live only in the southern area and Kerch Strait, where salinity is higher. Others(the comb jellies M. leidyi and B. ovata) can survive only during warm seasons and are reintroducedevery spring-early summer (M. leidyi ) or later summer (B. ovata). The copepod A. tonsa and diatomP. calcar-avis can survive in the Sea of Azov but develop only in warm seasons.
The main events that favored non-native species were increased salinity, which allowed manyeuryhaline Black Sea species and Black Sea invaders to expand into the Sea of Azov (some ofwhich remained after salinity declined toward original levels), and ballast tank construction onships (Figures 6 and 7). It is important to note that the species that established are euryhaline,eurythermal, have wide oxygen-level tolerance, and thrive in shallow water. Some freshwaterspecies established in bays and deltas. There are 47 non-native species total. Establishment of 9intentionally introduced fish is uncertain.
Table 3 List of the non-native species recorded in the Sea of Azov
Numbers ofestablished species
Numbers ofestablished species
Pathogenic bacteria 1 Copepoda 1Microplankton 2 Cirripedia 1Phytoplankton 7 Decapoda 3Macrophytes 1 Amphipoda 5Kamptozoa 1 Bivalvia 4Hydrozoa 2 Gastropoda 3Ctenophora 2 Pisces 13 (9)Polychaeta 1 Total 47 (9)
Note: Parentheses indicate numbers of species that were recorded in the Caspian but their establishment is uncertain.
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 113
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
50%
17%
33%
b
33%
23%
44%
Euryhaline Brackish water Fresh water
a
Figure 7(a) Ratio (%) of euryhaline (Mediterranean), brackish water (Ponto-Caspian), and freshwater (includinganodromous and catadromous fish) species in native biota; and (b) ratio (%) of non-native euryhaline,brackish water, freshwater species of different origin in the Sea of Azov.
4.3. Ecosystem Impacts
Selected non-native species have negatively affected the ecosystem, whereas the others have en-riched its flora and fauna without substantial negative impacts. When analyzing the ecological roleof invaders in the Sea of Azov, one should first mention the enormous negative effect at all lev-els of its ecosystem, fish resources included, caused by the invasion of the predatory ctenophoreM. leidyi. In summer, after seasonal invasion by M. leidyi, only non-native A. tonsa from zoo-plankton and larvae of B. improvisus from meroplankon can resist the predator and survive in thesea or even in brackish Taganrog Bay (Povazhny 2009). Other invasive species that spread fromthe Black Sea have negative effects. The pathogenic bacterium V. cholerae is notorious, whereasthe common shipworm T. navalis and the acorn barnacle B. improvisus attack wooden structures.P. calcar-avis is not used by Azov zooplanktonic species owing to its large size, so it is harmful forthe Azov ecosystem during its bloom, replacing native species and sinking after elimination, thusprovoking hypoxia near the bottom (Studenikina et al. 1999).
The introduction of other organisms may be regarded as positive events. Benthic speciessuch as the bivalve A. inaequivalvis and M. arenaria spread widely over the regions with lowoxygen content unfavorable for other benthic representatives in the Sea of Azov; they providedvaluable food resources for benthophagous fishes, whereas their larvae are consumed by smallpelagic fishes. The crab R. harrisii also became an additional food object for benthophagousfishes (Frolenko 2006). The mullet L. hematochilus became a valuable commercial fish in theSea of Azov, where native species of mullet are not abundant and do not reproduce (Volovik& Chikharev 1998). The ctenophore B. ovata might be expected to be a useful invader as abiocontrol of M. leidyi. Unfortunately, because of its seasonal dynamics, it appears in the Seaof Azov too late, when M. leidyi has already reproduced, spread widely, and undermined thestocks of zooplankton. For that reason, no positive role of B. ovata in reducing the M. leidyipopulation in the Sea of Azov has been noted to date. Meanwhile, its development in the BlackSea influences the size of the M. leidyi population; therefore, after the B. ovata appearance,M. leidyi enters the Sea of Azov later and its abundance is significantly lower (Mirzoyan et al.2006).
114 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
5. THE CASPIAN SEA
5.1. Environment, Native Biota, and Disturbance
The Caspian Sea is the largest inland water body; its shelf zone (<100 m deep) occupies 62% ofits surface area. Physical geography and bottom topography divide the Caspian into Northern,Middle, and Southern regions. Sea-level oscillation is one of the main factors that determinesthe status of its ecosystems. During the twentieth century, environmental conditions deterioratedsignificantly, mainly owing to sea-level changes, river runoff, and pollution from multiple sourcesincluding petroleum hydrocarbons and phenols (Kosarev 2005).
Inhabitants belong to four groups. The most ancient and abundant are autochthonous(Ponto-Caspian) species (84%). Arctic species (3%) arrived during the last glaciations. Atlantic-Mediterranean species (1%) penetrated about 13,000 years ago. They have become full members ofCaspian communities, have evolved considerably, and have generated new species and subspecies.Freshwater species (13%) have entered on several occasions (Zenkevich 1963).
The present-day Caspian Sea is relatively species poor. Species richness is lower than that ofthe Black Sea by a factor of 2.5, although the biota contains 733 species and subspecies of plantsand 1,814 species and subspecies of animals, of which 1,069 are free-living invertebrates, 325 areparasites, and 415 are vertebrates (the latter are mainly represented by freshwater species; this listis still growing). The principal causes of the high degree of faunal endemism lie in the long-termisolation of the basin and its salinity regime. The low salinity (0.1–11% in the Northern Caspian,12.6–13% in the other parts) and its native biota restricted colonization by many marine speciesand, at the same time, constrained access by freshwater species. In spite of low biodiversity, theCaspian Sea has high productivity, particularly in the Northern Caspian, and rich fish stocks(Kasymov 1987).
5.2. Vectors, Pathways, and Composition of Invaders
The appearance of non-native species and changes in biodiversity may be divided into threephases (Figures 8 and 9). The first comprised deliberate large-scale introductions beginning inthe 1930s within the framework of the Soviet Union program for enriching commercial stocks andaimed at enlarging the resources of either commercial fishes themselves or their food organisms.However, among these introductions, only two finfish [the mullets Liza saliens (Popov 1930) andL. aurata (Popov 1930)] from the Black Sea and two benthic species [the polychaete Hediste (Nereis)diversicolor (O.F. Muller, 1776) and the bivalve Abra (Syndesmya) ovata (Philippi 1836)] from theSea of Azov achieved significance. Two Black Sea prawns, Palaemon adspersus (Rathke 1837) andP. elegans (Rathke 1837), accompanied the mullets, were inadvertently released with them, andbecame a valuable food source for benthic-feeding fishes. Eight rough fish and fish parasites werealso inadvertently introduced during these actions (Karpevich 1975) (Table 4).
The second introduction phase started when the Volga-Don Canal opened in 1952. First,most of these species were carried from the Black Sea by ships as fouling organisms. Amongthem zoobenthic animals and macrophytes dominate. Among macropytes found in late 1950 werethe green algae Acrochaete parasitica Oltm, Ectochaete leptochaete Huber (= Entocladia leptochaete),Enteromorpha flexuosa (Wulf ) (= E. tubulosa), E. maeotica Proshkina-Lavrenko, Monostroma latis-simum (Kuetzing) Witt; the brown algae Ectocarpus confervoides var. siliculosus (Dillwyn) Farlow,Entonema oligosporum Stromfelt (= Streblonema oligosporum); and the red algae Acrochaetium daviesii(Dillw) Nag, Ceramium diaphanum (Lightf ) Roth, Polysiphonia variegata (C. Ag.) (Zanardini)(= P. denudata) (Zevina 1994). The third phase began in the early 1980s, when mainly
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 115
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
13% 2%
5%
7%
67%
3% 3%aAtlantic coastalarea of AmericaAtlantic coastalarea of Europe
Northern Atlantic
NortheasternAsia
Black Sea
Sea of Azov
Uncertain
2%
7%
85%
3% 3%b
Figure 8Donor areas of the non-native species and their percentage in the Caspian Sea. (a) Primary areas;(b) secondary (actual) areas.
phyto- and zooplanktonic species began to arrive in ballast water after ballast tank constructionswere made on ships.
All established non-native species were brought in these different periods mainly from the BlackSea and partly from the Sea of Azov. A first group includes 23 widely distributed and often abundanteuryhaline Black Sea species. These are of Atlantic-Mediterranean origin but have lived for 1,500–2,000 years in the Black Sea and adapted to its low salinity. They are Cladocera (Pleopis polyphemoidesLeukart, Penilia avirostris Dana 1852), Podon intermedius (Lilljeborg), the amphipod Corophiumvolutator (Pallas 1766), the bivalve Mytilaster lineatus (Gmel.), phytoplankton, and macrophytes.Invasion by the Black Sea phyto- and zooplankton species is still going on. The copepod Oithonasilimis Claus 1866 has now been recorded for four years. Penilia avirostris, whose invasion wasdoubted, was found in 2009 in the Middle Caspian. Among new phytoplankton species Pseudo-nitzschia seriata (Cleve) has become abundant, whereas Pseudo-nitzschia pseudodelicatissima (Hasle),Chaetoceros peruvianus (Brightwell 1856), and the temporarily planktonic Tropidoneis lepidoptera(Greg. Cl.) have become widely distributed and are often recorded. In the Middle Caspian Seathe coccolithophore Braarudosphaera bigelowii (Gran & Braarud 1935) and globally significantcoccolithophore alga Emiliania huxleyi (Lohmann) Hay & Mohler have been observed. E. huxleyihas often bloomed in the Black Sea during the past several decades. In addition, two Black Seadinoflagellate species were recently found in the Caspian Sea: Gymnodinium sanguineum (Hirasaka)[(= G. splendens (Lebour)] and Protoperidinium crassipes (Kofoid) (Ballech 1974) (Shiganova et al.2005, Pautova et al. 2008).
0
5
10
15
20
< XX cen
Num
ber o
f spe
cies
Macrophytes Zoobenthos Zooplankton Phytoplankton Fish
Ballast tank construction
2001–2008 1981–20001961–19801952–19601921–19511900–1920
Volga-Don Canal construction
Deliberate stocking
Figure 9Chronology of species invasions into the Caspian Sea.
116 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Table 4 List of the non-native species recorded in the Caspian Sea
Numbers of established speciesParasites of fishes 9Phytoplankton 8 (3)Macrophytes 9Kamptozoa 1Scyphozoa (1)Hydrozoa 3Ctenophora 1Polychaeta 2Copepoda 2 (1)Cladocera 3Cirripedia 1 (1)Decapoda 4Amphipoda 3Bivalvia 4Gastropoda 2Bryozoa 3Pisces 14 (2)Total 60 (8) without parasites (15)
Notes: Parentheses indicate numbers of species that were recorded in the Caspian buttheir establishment is uncertain.
Another group consists of nine Black Sea brackish water species that were adapted to life inlow-salinity areas before invading similar areas of the Caspian: the hydromedusa Moerisia maeot-ica (Ostroumov); amphipods Gammarus aequicauda (Martyinov 1931) and Iphigenella shablensis(Carausu); molluscs Monodacna colorata (Eichwald 1829) (= Hypanis colorata), Dreissena bugensis(Andrusov 1897), Lithogliphus naticoides (Pfeiffer), and Tenellia adspersa (Nordmann 1845); kampto-zoan Barentsia benedeni (Foetinger 1887); and pearlwort Conopeum seurati (Canu) (= Membraniporacrustulenta) (Karpinsky et al. 2005). In addition to long-established Black Sea species, some BlackSea invaders have also been introduced. These include seven species that arrived first in the BlackSea from Atlantic inshore regions of North America, among them M. leidyi and A. tonsa. Alsoarriving via this route were the North American hydromedusae B. virginica and B. megas. Fromthe northern European Atlantic coast came the polychaete F. enigmaticus and two diatom species:P. calcar-avis and Cerataulina pelagica. The origins of other Caspian non-native species are un-certain, including the pearlwort Lophopodella carteri, three-spined stickleback Gasterosteus aculeatusaculeatus, and Chinese mitten crab E. sinensis.
About 60 species are established, although some fishes are known only from single individu-als; the persistence of eight more species is doubted, and four more species have only recentlybeen found. Among them the scyphomedusa A. aurita has been recorded only a few times in theMiddle and Southern Caspian. The fate of B. eburneus is not known. Recently, seven more phy-toplankton species and three zooplankton from the Black Sea were found in the Middle Caspian(Shiganova et al. 2005), but it is not known if they have established. Some introduced speciesalso failed to establish. Except for a few freshwater invertebrates and deliberately introducedfreshwater fishes, plus two species deliberately introduced from the Sea of Azov, all established in-vaders were introduced from the Black Sea (Figure 8a,b). Most established species are euryhaline
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 117
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Euryhaline Mediterranean
Brackish water
Euryhaline
Brackish waterFresh water Fresh water
Euryhaline Arctic
3% 2%
75%
20%
b4% 1%
35%
60%
a
65%
25%
10%
c
Figure 10Ratio (%) of euryhaline (Arctic and Mediterranean), brackish water, and freshwater native species in (a) theNorthern Caspian and (b) the Middle and Southern Caspian. (c) Ratio (%) of euryhaline, brackish water, andfreshwater species in non-native biotas of the Caspian Sea.
(Figure 10); many are widely distributed in coastal waters and therefore have wide ecological toler-ances. Fewer established invaders are from brackish water, and the only freshwater species are de-liberately introduced fish. Among introduced species, there are many more euryhaline species thanin the native biota in both the brackish Northern Caspian and the Middle and Southern Caspian(Figure 10). Marine euryhaline non-native species have settled in the Middle and SouthernCaspian, often replacing native species. Brackish and freshwater species have settled in the North-ern Caspian, although the most euryhaline of them may penetrate the Middle and SouthernCaspian.
5.3. Ecosystem Impacts
The Caspian Sea ecosystem was the most vulnerable to invaders because of its long isolationand high level of endemism. Most Atlantic invaders had major impacts; for instance, M. leidyiaffected all trophic levels and finally ecosystem functioning. Though few in number, these speciesoccupy dominant community positions. They include the diatom P. calcar-avis, the cladoceranP. polyphemoides, and the copepod A. tonsa. The biomass of M. lineatus, A. ovata, H.(N). diversicolor,and B. improvisus makes up more than 60% of the total biomass of the benthos and fouled substrates.Fouling communities consist almost wholly of non-native species. Native species dominate onlyamong the fishes.
Some introductions may be construed as positive, but many are harmful, especially that ofM. leidyi. Hediste diversicolor and A. ovata became the favorite food of stellate and Russian sturgeons.The crab R. harrisii, competing with sturgeons for food, is itself one of their food items. Themussel M. lineatus, which forms the bulk of the benthic biomass, replaced native species and isscarcely used by benthophagous fish and sturgeons. But during the past few years M. lineatus hasincreasingly been found in stomach contents of benthophagous sturgeons owing to the absence ofother available food (Molodtzova et al. 2004). The mullet fishery achieved only limited importance.The diatom P. calcar-avis, having increased phytoplankton biomass, was of limited nutritive valuefor zooplankters and pelagic phytophagous fish.
After the invasion of M. leidyi, the functioning of the Caspian ecosystem changed as in theBlack and Azov Seas previously (Shiganova et al. 2004b). In addition, after invasion by M. leidyi,
118 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
only non-native zooplankton, meroplankton, and zoobenthic species substantially survived thispredator. Among them are Acartia tonsa, which at times composes up to 90% of zooplanktonannually by August, and larvae of B. improvisus. They also often become the main food of all threespecies of kilkas. Stocks of the anchovy kilka Clupeonella engrauliformis (Borodin 1904) and big-eyeC. grimmi Kessler 1877 have greatly decreased. Only Clupeonella cultriventris (Nordmann 1840)has survived because of its wide ecological tolerances and capacity to migrate to feed in brackishand fresh waters (Zarbalieva et al. 2006). It has also spread widely into brackish and fresh watersin the Volga River basin.
Among the factors that permitted the outbreak of M. leidyi was the absence of predators.There is much optimism, therefore, about the intentional or accidental arrival in the Caspian Seaof B. ovata, which preys exclusively on zooplanktivorous ctenophores and could be a successfulbiocontrol of the M. leidyi population as shown by the example of the Black Sea ecosystem. Beroeovata would be able to live in salinities of the Middle and Southern Caspian and perhaps controlMnemiopsis abundance if it were introduced (Kideys et al. 2004).
6. DISCUSSION AND CONCLUSIONS
We may conclude that in the latter half of the twentieth century, increased shipping and construc-tion of canals caused the Black Sea to become a recipient and donor area for marine and brackishwater species. It serves as a hub for species that then spread further to the Sea of Azov and theCaspian Sea, and also to the Sea of Marmara and in some cases to the eastern Mediterranean Sea.All three Ponto-Caspian seas have low diversity but high productivity and are inland semiclosed orclosed seas with limited water exchange with the ocean (the Black Sea) or with no such exchange(the Caspian Sea). All these features have made them vulnerable to invasions.
The location of the Black Sea is crucial. Situated between the species-rich Mediterranean Seaand the species-poor Azov and Caspian seas, the Black Sea is also intermediate between these seasin abiotic conditions: It has much lower salinity than the Mediterranean but higher salinity thanthe Sea of Azov and the Caspian Sea (Table 1). Anthropogenic disturbance of the Black Sea hasalso increased vulnerability; eutrophication and overfishing of top fish predators and dolphins areespecially important.
The introduced species in all three seas are mainly from temperate regions, widely distributedin the coastal areas of the World Ocean. They have high environmental tolerance, have abun-dant genetic variation, and are generally dominant in their native habitats. They often becamedominant first in the Black Sea, then in many instances in the other seas. Native biodiversitydeclined and invaders now dominate. The invaders that affected all Ponto-Caspian ecosystems tothe greatest extent are mainly euryhaline, in most cases from the northern Atlantic coastal regions.The proportion of euryhaline species has increased compared to that in the original native biota,with the increase least in the Black Sea and greatest in the Caspian Sea. In the Black Sea, theproportion of euryhaline species is almost the same in the native and non-native biotas. In the Seaof Azov, which has more limited environmental conditions, the proportion of euryhaline invadersis 1.5 times higher than it is in the native biota; and in the closed Caspian Sea, which has a veryspecific salt content and a high level of biotic endemism, it was mostly euryhaline species thatwere able to establish. The proportion of euryhaline species among invaders there is 13 timeshigher than the native biota (Figures 4, 7, and 10). Some Atlantic invaders were able to establishbecause their dispersal was in gradual steps. First they were introduced into the Black Sea, wheresalinity is lower than or equal to that in their native habitats. Then they spread to the Sea of Azovand ultimately to the Caspian, where salinity is lower than in the Black Sea. Therefore, salinitydecrease was gradual for such invaders and they were able to establish more easily.
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 119
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
156
214
15 6447
9
60
1 4 7
647
98
Established
Off-Bosporus orsingle findings
Uncertain
Recently found
Mediterranean Sea
Black Sea
Sea ofAzov
Caspian Sea
Figure 11Total numbers of the non-native species in the seas of Eurasia.
In addition, most of these successful invaders have physiological features that aid theirestablishment such as a wide food spectrum, high fecundity, hermaphroditism (barnaclesB. eburneus, B. improvisus; shipworm T. navalis; comb-jellies M. leidyi and B. ovata), and in some casesthe ability to self-fertilize (B. improvisus, T. navalis, M. leidyi, B. ovata). Many non-native speciesalso have abilities to survive in unfavorable conditions, for example, by decreasing metabolism,reducing movements, and stopping feeding (M. leidyi, B. ovata); by releasing eggs directly intothe water or producing resting eggs (copepod A. tonsa); by producing cysts (some algae); by rapidgrowth to maturity and reproducing quickly after hatching (within two weeks: M. leidyi, B. ovata;within one month: B. eburneus, B. improvisus).
The most impressive example is the ctenophore M. leidyi, which is highly preadapted to rapidcolonization of new areas. It is a simultaneous hermaphrodite, is able to self-fertilize, and has ahigh reproductive rate. High percentages of eggs and larvae survive during development. It isable to regenerate from fragments larger than one quarter of an individual. It is also a generalistcarnivorous feeder and can live without food for a long time by simply shrinking during unfavorableconditions. M. leidyi has high genetic diversity (Ghabooli et al. 2010) and occurs over a broad rangeof salinity and temperature conditions (Purcell et al. 2001).
The numbers of non-native species are proportional to the numbers of native species in the seasof Eurasia (Figures 11 and 12); however, the proportion of non-native species is not high in theseseas: in the Mediterranean it is 5.4% [if we do not include some doubtful cases (Zenetos et al. 2005)];in the Black Sea, 4.1%; in the Sea of Azov, 4.2%; and in the Caspian Sea, 2.6%. Both numbersof native species and numbers of non-native species decline with salinity decrease in the Eurasianseas, from the species-rich Mediterranean Sea, to the less-rich Black Sea, to the impoverishedAzov and Caspian seas. High species diversity in the Mediterranean Sea might be explained byits high salinity, which allows native oceanic species to exist and non-native oceanic species toestablish. In addition, diversity of environmental conditions and high, increasing temperaturefacilitate establishment of subtropical and tropical species. From the Mediterranean to the BlackSea salinity drops by 21�, the numbers of native species decrease by a factor of 3.5, and numbersof non-native species decrease by a factor of 3.5 as well.
Among the Ponto-Caspian seas, the highest native and non-native species diversities are in theBlack Sea, the next highest are in the Caspian Sea, and the lowest ones are in the Sea of Azov
120 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
0
100
200
300
400
500
600
700
800
0
2,000
4,000
6,000
8,000
10,000
12,000
14,000
MediterraneanN
umber of non-native species
Num
ber o
f nat
ive
spec
ies
Caspian SeaSea of AzovBlack Sea
Figure 12Ratio of native and non-native species in the inland southern seas of Eurasia.
(Figure 12). From the Black Sea to the Sea of Azov, salinity drops by only 7�, but total numberof native species decreases by a factor of 3.4 and the number of non-native species decreases by afactor of 3.3. The total number of native species falls by a factor of 2.5 between the Black Sea andthe Caspian Sea, while the number of non-native species falls by a factor of 2.6. The decrease inthe total numbers of both native and non-native species in the Azov and Caspian Seas might beexplained by the fact that their salinities drop below the level of that in mesohaline waters (about15�), which is a key biotic threshold (Mordukhai-Boltovskoi 1969).
It is evident that the widely cited theory that a species-rich marine basin will be less invasiblethan a species-poor one with many empty niches is not confirmed in the case of the inland southernEurasian seas. On the contrary, in these marine basins the number of non-native species establishedis proportional to the number of native species (Figures 11 and 12).
The most widely distributed, ecologically tolerant species, which established in the Black Seaand developed large populations there, are also abundant and widely distributed in other Ponto-Caspian seas. Along with changing conditions, the construction of the Volga–Don canal facilitatedthe colonization and naturalization of both non-native species established in the Black Sea andnative Black Sea species in the other Ponto-Caspian seas, and their numbers are high for allthree seas and rather high for the Sea of Marmara (Figure 13). These processes led to biotichomogenization of all the Ponto-Caspian Seas, as the same species became dominant numericallyin communities and often in ecosystem functioning.
The rate of introduction has accelerated in the Ponto-Caspian seas, particularly after ballasttank construction for ships and as upper sea layer temperature increased (Figure 14). In theBlack Sea, the rate of non-native species establishment increased by a factor of 2.5 after 1960, adate that coincides with the beginning of substantial anthropogenic disturbance there. In the Seaof Azov and the Caspian Sea, the rates accelerated by factors of 1.75 and 1.55, respectively, atthat time (Figure 14). During the next twenty years the rate of non-native species establishmentincreased by a factor of 2.4 in the Black Sea, 2.1 in the Sea of Azov, and 1.4 in the Caspian Sea.From 2001 to 2008 the rate of non-native species establishment increased in the Black Sea by afactor of 1.33, in the Sea of Azov by a factor of 1.16, and in the Caspian Sea by a factor of 1.15(Figure 14). The numbers of recorded species recently are much greater, particularly in the BlackSea (Table 2). As a consequence, the native biodiversity became poorer in all seas, and invadersin all ecological groups (except for fish) now play a dominant role in ecosystem functioning. Firstnon-native species affected the native communities simply by occupying certain ecological niches,after which some of them affected other trophic levels, and finally some caused drastic changes
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 121
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
regions70
30
83
13 13
204
76
42
94
Penetrated fromthe Black SeaProbably fromthe Black SeaFrom the Seaof AzovFrom other
Mediterranean Sea
Sea ofMarmara
Sea ofAzov
Caspian Sea
Figure 13Proportion of species (%) penetrated from the Black Sea, probably from the Black Sea, from the Sea of Azov,and from other regions.
in ecosystem functioning and a major rearrangement of the original energy fluxes. In most casesnon-native species suppress native species directly by preying on them or indirectly by competingwith them for food or habitat.
The main driver in all these ecosystems has been the most aggressive gelatinous invader,M. leidyi. We found cascading effects of M. leidyi in all Ponto-Caspian seas, including bottom-up control from zooplankton, to planktivorous fish, to top predators (dolphins, seals, predatoryfish), and top-down control from zooplankton to phytoplankton and from bacteria to ciliates andzooflagellates. Mnemiopsis leidyi has also greatly affected the ecosystem of the Sea of Marmarasince 1992 (Shiganova 1993) and has spread to the Aegean Sea, but no remarkable effect has beenrecorded in this oligotrophic sea (Shiganova et al. 2001). After B. ovata appeared in the BlackSea, the ecosystem began to recover. Beroe ovata also expanded in the Sea of Azov and the Seaof Marmara (Isinibilir et al. 2004, Mirzoyan et al. 2006), so these two ctenophores now largelydetermine ecosystem status in these seas. In 2009, M. leidyi continued its expansion into theMediterranean Sea. Outbreaks were observed in various areas of the Mediterranean Sea during
0
20
40
60
80
100
120
140
160
<1900
1900–1921
1921–1940
1941–1960
1961–1980
1981–2000
2001–2008
<1900
1900–1921
1921–1940
1941–1960
1961–1980
1981–2000
2001–2008
<1900
1900–1921
1921–1940
1941–1960
1961–1980
1981–2000
2001–2008
Num
ber o
f spe
cies
Black Sea Sea of Azov Caspian Sea
Figure 14Cumulative graphs of non-native species invasion rate for 20-year intervals (colored in shades of blue) in the seas of the Ponto-Caspian in1990–2008.
122 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
total warm seasons (Boero et al. 2009, Fuentes et al. 2009, Galil et al. 2009). Continued expansionof M. leidyi in different areas of the Mediterranean Sea is a great concern. The outbreak of the non-native gelatinous ctenophores M. leidyi and B. ovata in the Eurasian seas has significantly advancedour understanding of the complex nature of the role of invasive species in marine ecosystems.It offers an example of how lower gelatinous invaders can affect an entire system: One of themcompletely suppressed several productive ecosystems, while the invasion of the other led to therecovery of the Black Sea ecosystem (Shiganova et al. 2004a). These events featured a combinationof two large ecological problems of the seas and coastal oceans: species invasions and gelatinousplankton outbreaks.
The trend is for an acceleration of invasions by new gelatinous species. The Mediterraneangelatinous species are increasing their populations with rising temperature and have begun todisperse with the currents out of their native ranges. Recently, several Mediterranean gelatinousspecies arrived for the first time in the Sea of Marmara. Among them are the jellyfishes Chrysaorahysoscella (Linne 1766), Cassiopea andromeda Forskal 1775 and the trachimedusa Liriope tetraphyllaChamisso and Eysenhardt. In 2009, C. hysoscella was recorded for the first time in the IstanbulStrait and the Turkish part of the Black Sea (Shiganova & Ozturk 2010). This is a temperateplanktivorous species so we cannot exclude the possibility that it will establish in the Black Sea infavorable conditions with respect to its crustacean zooplankton prey if it can tolerate the salinity.In 2007, the ctenophore Bolinopsis vitrea (L. Agassiz 1860) was also recorded in the Turkish partof the Black Sea (Ozturk & Shiganova 2010).
Thus, there is a shift from native biodiversity to biotic homogenization in all of the Ponto-Caspian seas and, to an extent, in the Sea of Marmara, with domination by the same introducedspecies and by Black Sea invertebrates and algal species. Gelatinous invaders have become the maindrivers of ecosystem function. The invasion rate of non-native species and the rate of appearanceof new gelatinous invaders are accelerating in all inland southern seas of Eurasia.
DISCLOSURE STATEMENT
The author is not aware of any affiliations, memberships, funding, or financial holdings that mightbe perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
I greatly appreciate the invitation of Prof. Daniel Simberloff to write this review and his valuableediting of my manuscript. Research described herein was performed in the framework of projectEC “SESAME.”
LITERATURE CITED
Boero F, Putti M, Trainito E, Prontera E, Piraino S, Shiganova T. 2009. First records of Mnemiopsis leidyi(Ctenophora) from the Ligurian, Thyrrhenian and Ionian Seas (Western Mediterranean) and first recordof Phyllorhiza punctata (Cnidaria) from the Western Mediterranean. Aquat. Invasions 4(4):675–80
Carlton JT, Geller JB, Reaka-Kudla ML. 1999. Historical extinctions in the sea. Annu. Rev. Ecol. Syst. 30:515–38
Chandler EA, McDowell JR, Graves JE. 2008. Genetically monomorphic invasive populations of the rapawhelk (Rapana venosa). Mol. Ecol. 17:4079–91
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 123
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Dumont HJ, Shiganova TA, Niermann U, eds. 2004. The Ctenophore Mnemiopsis leidyi in the Black, Caspianand Mediterranean Seas and Other Aquatic Invasions, Vol. 35. Dordrecht/Boston/London: Kluwer Acad.NATO ASI Ser., IV Earth Environ. Sci. 314 pp.
Frolenko LN. 2006. Benthic invertebrate communities of the Sea of Azov. In The Main Problems of the Fisheriesand Protection of Water Bodies with Fisheries in the Azov and Black Sea Basin, ed. NV Voinova, pp. 88–96.Rostov-on-Don: AzNIIRKH. 596 pp. (In Russian)
Fuentes VL, Atienza D, Gili J-M, Purcell JE. 2009. First record of Mnemiopsis leidyi A. Agassiz 1865 off theNW Mediterranean coast of Spain. Aquat. Invasions 4(2):315–18
Galil B, Kress N, Shiganova T. 2009. First record of Mnemiopsis leidyi A. Agassiz, 1865 (Ctenophora; Lobata;Mnemiidae) off the Mediterranean coast of Istrael. Aquat. Invasions 4(2):356–62
Ghabooli S, Shiganova TA, Zhan A, Cristescu M, Eghtesadi-Araghi P, MacIsaac HJ. 2010. Multiple intro-ductions and invasion pathways for the invasive ctenophore Mnemiopsis leidyi in Eurasia. Biol. Invasions.In press
Greze VN. 1979. Zooplankton. In Biological Productivity of the Black Sea, ed. VN Greze, pp. 143–69. Kiev:Nauk. Dumka. 392 pp. (In Russian)
Isinibilir M, Tarkan AN, Kideys AE. 2004. Decreased levels of the invasive ctenophore Mnemiopsis in theMarmara Sea in 2001. See Dumont et al. 2004, pp. 155–66
Karpevich AF. 1975. Theory and Practice of Aquatic Animal Acclimatizations. Moscow: Pizhevaya Promyshlen-nostj. 432 pp. (In Russian)
Karpinsky MG, Shiganova TA, Katunin DN. 2005. Introduced species. See Kostianoy & Kosarev 2005,pp. 175–90
Kasymov AG. 1987. Biota of the Caspian Sea. Baku: Elm. 156 pp. (In Russian)Ketelaars HAM. 2004. Range extensions of Ponto-Caspian aquatic invertebrates in continental Europe. See
Dumont et al. 2004, pp. 209–36Kideys AE, Finenko GA, Anninsky BE, Shiganova TA. 2004. Physiological characteristics of the Beroe ovata
in the Caspian water. J. Mar. Ecol. Prog. Ser. 266:111–21Kiseleva MI. 1979. Zoobenthos. In Biological Productivity of the Black Sea, ed. VN Greze, pp. 208–39. Kiev:
Nauk. Dumka. 392 pp. (In Russian)Kopelevich OY, Burenkov VI, Sheberstov SV, Lukianova EA, Prokhorenko OV. 2005. Bio-optical character-
istics of the seas of Russia from data of the SeaWifs satellite ocean scanner. Moscow: P.P. Shirshov Inst.Oceanol. Russ. Acad. Sci. CD.
Kosarev AN. 2005. Physico-geographical conditions of the Caspian Sea. See Kostianoy & Kosarev 2005,pp. 5–31
Kosarev AN, Kostianoy AG, Shiganova TA. 2008. The Sea of Azov. See Kostianoy & Kosarev 2008, pp. 63–89Kostianoy AG, Kosarev AN, eds. 2005. The Caspian Sea Environment. Handb. Environ. Chem., Vol. 5, Part P.
Berlin/Heidelberg: Springer-Verlag. 271 pp.Kostianoy AG, Kosarev AN, eds. 2008. The Black Sea Environment. Handb. Environ. Chem., Vol. 5, Part Q.
Berlin/Heidelberg: Springer-Verlag. 457 pp.Mills EL, Leach JH, Carlton JT, Secor CL. 1993. Exotic species in the Great Lakes: History of biotic crises
and anthropogenic introductions. J. Great Lakes Res. 19(1):1–54Mirzoyan ZA, Martynuk ML, Vyazun EV. 2006. Development of ctenophores Beroe ovata and Mnemiopsis
leidyi in the Sea of Azov in the present-day-period. In The Main Problems of the Fisheries and Protection ofWater Bodies with Fisheries in the Azov and Black Sea Basin, ed. NV Voinova, pp. 136–48. Rostov-on-Don:AzNIIRKH. 596 pp. (In Russian)
Molodtzova AI, Polyaninova AA, Kashentseva AI, Kamelov AK. 2004. The state of feeding of sturgeons in theCaspian Sea. In Fisheries Research in the Caspian Sea, ed. MI Karpuyk, pp. 215–25. Astrakhan: CaspNIRKH.570 pp. (In Russian)
Mordukhai-Boltovskoi FD. 1960. Caspian Fauna in the Azov and Black Seas Basins. Moscow-Leningrad: Acad.Nauk USSR. 290 pp. (In Russian)
Mordukhai-Boltovskoi FD. 1969. Amphipoda Latreille of the Black Sea. In Guide to Fauna in the Black and AzovSeas, ed. VF Vodyanitsky, pp. 440–524. Kiev: Nauk. Dumka. 151 pp. (In Russian)
Ozturk B, Shiganova T. 2010. First finding of Bolinopsis vitrea: (Ctenophore, Lobata) in the Turkish Black Seacoasts. J. Biol. Invasions. In press
124 Shiganova
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41CH06-Shiganova ARI 1 October 2010 14:33
Pautova LA, Silkin VA, Vostokov SV. 2008. The phytoplankton of the present-day central Caspian Sea. InSaline Lakes Around the World: Unique Systems with Unique Values, ed. A Oren, DL Naftz, WA Wurtsbaugh,p. 184. Nat. Resourc. Res. Libr., Utah State Univ. Coll. Nat. Resourc. 172 (Abstr.)
Petranu A. 1997. Black Sea Biological Diversity—Romania. Black Sea Environ. Ser., Vol. 4. New York: UN.315 pp.
Povazhny VV. 2009. Zooplankton communities functioning in Taganrog Bay of the Sea of Azov. PhD thesis.Murmansk. 129 pp. (In Russian)
Purcell JE, Shiganova TA, Decker MB, Houde ED. 2001. The ctenophore Mnemiopsis leidyi in native and exotichabitats: U. S. estuaries versus the Black Sea basin. In Jellyfish Blooms: Ecological and Social Importance, ed.JE Purcell, WM Graham, HJ Dumont, pp. 145–76. Dordrecht/Boston/London: Kluwer Acad. 333 pp.
Rilov G, Crooks JA. 2009. Marine bioinvasions: conservation hazards and vehicles for ecological under-standing. In Biological Invasions in Marine Ecosystems, Ecol. Stud., ed. G Rilov, JA Crooks, 204:3–12.Berlin/Heidelberg: Springer. 641 pp.
Shiganova T. 2008. Introduced species. See Kostianoy & Kosarev 2008, pp. 375–406Shiganova T, Ozturk B. 2010. Trend on increasing Mediterranean species arrival into the Black Sea. In Towards
a “Mediterraneazation of the Black Sea? Physical and Biological Changes, pp. 75–91. CIESM WorkshopMonogr. 39, Trabzon, 3–6 June, 2009
Shiganova TA. 1993. Ctenophore Mnemiopsis leidyi and ichthyoplankton in the Sea of Marmara in October of1992. Oceanology 33(6):900–3
Shiganova TA, Dumont HJD, Mikaelyan AS, Glazov DM, Bulgakova YV, et al. 2004a. Interaction betweenthe invading ctenophores Mnemiopsis leidyi (A. Agassiz) and Beroe ovata Mayer 1912, and their influenceon the Pelagic ecosystem of the Northeastern Black Sea. See Dumont et al. 2004, pp. 33–70
Shiganova TA, Dumont HJD, Sokolsky AF, Kamakin AM, Tinenkova D, Kurasheva EK. 2004b. Populationdynamics of Mnemiopsis leidyi in the Caspian Sea, and effects on the Caspian ecosystem. See Dumont et al.2004, pp. 71–111
Shiganova TA, Mirzoyan ZA, Studenikina EA, Volovik SP, Siokou-Frangou I, et al. 2001. Population devel-opment of the invader ctenophore Mnemiopsis leidyi in the Black Sea and other seas of the Mediterraneanbasin. J. Mar. Biol. 139:431–45
Shiganova TA, Musaeva EI, Pautova LA, Bulgakova YV. 2005. The problem of invaders in the Caspian Sea inthe context of the findings of new zoo- and phytoplankton species from the Black Sea. Biol. Bull. 1:78–87
Simberloff D, Von Holle B. 1999. Positive interactions of nonindigenous species: invasion meltdown? Biol.Invasions 1:21–32
Studenikina EI, Aldakimova AYA, Gubina GS. 1999. Phytoplankton of the Sea of Azov in Conditions of Anthro-pogenic Impact. Rostov-on-Don: AzNIIRKH. 175 pp. (In Russian)
Svetovidov AN. 1964. Handbook of the Fauna of the USSR, Fishes of the Black Sea. Moscow: Nauka. 550 pp.Volovik SP, Chikharev AS. 1998. Anthropogenic alterations of ichthyofauna of the Azov basin. In The Main
Problems of Fisheries and Fishery Basin Conservation in Azov-Black Sea Basin, ed. EV Makarov, SP Volovik,YE Tyutina, pp. 7–22. Rostov-on-Don: AzNIIRKH. 736 pp. (In Russian)
Zaitsev YUP, Mamaev V. 1997. Marine Biological Diversity in the Black Sea. A Study of Change and Decline. BlackSea Environ. Ser. 3. New York: UN. 208 pp.
Zarbalieva TS, Gadjiev RV, Akhundov MM, Gasanov NG, Muradova IT. 2006. Characteristic of feedingof kilka in the western coastal areas of the Middle and Southern Caspian under new environmentalconditions. In Fisheries Research in the Caspian, ed. MI Karpyuk, AY Mazhnik, DN Katunin, pp. 140–56.Astrakhan: CaspNIRKH. 435 pp. (In Russian)
Zenetos A, Cinar ME, Pancucci-Papadopoulou MF, Hermelin JR, Furnari J, et al. 2005. Annotated list ofmarine alien species in the Mediterranean with records of the worst invasive species. Medit. Mar. Sci.6(2):1–56
Zenkevich LA. 1963. Biology of the Seas of USSR. Moscow: Acad. Nauk. USSR. 739 pp. (In Russian)Zevina GB. 1994. Biology of the Marine foulings. Moscow: Moscow State Univ. 135 pp. (In Russian)
www.annualreviews.org • Biotic Homogenization of Ponto-Caspian Seas 125
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41-FM ARI 1 October 2010 15:39
Annual Review ofEcology, Evolution,and Systematics
Volume 41, 2010Contents
What Animal Breeding Has Taught Us about EvolutionWilliam G. Hill and Mark Kirkpatrick � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
From Graphs to Spatial GraphsM.R.T. Dale and M.-J. Fortin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �21
Putting Eggs in One Basket: Ecological and Evolutionary Hypothesesfor Variation in Oviposition-Site ChoiceJeanine M. Refsnider and Fredric J. Janzen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �39
Ecosystem Consequences of Biological InvasionsJoan G. Ehrenfeld � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �59
The Genetic Basis of Sexually Selected VariationStephen F. Chenoweth and Katrina McGuigan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �81
Biotic Homogenization of Inland Seas of the Ponto-CaspianTamara Shiganova � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 103
The Effect of Ocean Acidification on Calcifying Organisms in MarineEcosystems: An Organism-To-Ecosystem PerspectiveGretchen Hofmann, James P. Barry, Peter J. Edmunds, Ruth D. Gates,
David A. Hutchins, Terrie Klinger, and Mary A. Sewell � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 127
Citizen Science as an Ecological Research Tool: Challengesand BenefitsJanis L. Dickinson, Benjamin Zuckerberg, and David N. Bonter � � � � � � � � � � � � � � � � � � � � � � � 149
Constant Final YieldJacob Weiner and Robert P. Freckleton � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 173
The Ecological and Evolutionary Consequences of Clonalityfor Plant MatingMario Vallejo-Marın, Marcel E. Dorken, and Spencer C.H. Barrett � � � � � � � � � � � � � � � � � � � 193
Divergence with Gene Flow: Models and DataCatarina Pinho and Jody Hey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 215
Changing Geographic Distributions of Human PathogensKatherine F. Smith and Francois Guegan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 231
v
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.

ES41-FM ARI 1 October 2010 15:39
Phylogenetic Insights on Adaptive RadiationRichard E. Glor � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 251
Nectar Robbing: Ecological and Evolutionary PerspectivesRebecca E. Irwin, Judith L. Bronstein, Jessamyn S. Manson, and Leif Richardson � � � � � 271
Germination, Postgermination Adaptation, and SpeciesEcological RangesKathleen Donohue, Rafael Rubio de Casas, Liana Burghardt, Katherine Kovach,
and Charles G. Willis � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 293
Biodiversity and Climate Change: Integrating Evolutionaryand Ecological Responses of Species and CommunitiesSebastien Lavergne, Nicolas Mouquet, Wilfried Thuiller, and Ophelie Ronce � � � � � � � � � � � 321
The Ecological Impact of BiofuelsJoseph E. Fargione, Richard J. Plevin, and Jason D. Hill � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 351
Approximate Bayesian Computation in Evolution and EcologyMark A. Beaumont � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 379
Indexes
Cumulative Index of Contributing Authors, Volumes 37–41 � � � � � � � � � � � � � � � � � � � � � � � � � � � 407
Cumulative Index of Chapter Titles, Volumes 37–41 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 410
Errata
An online log of corrections to Annual Review of Ecology, Evolution, and Systematicsarticles may be found at http://ecolsys.annualreviews.org/errata.shtml
vi Contents
Ann
u. R
ev. E
col.
Evo
l. Sy
st. 2
010.
41:1
03-1
25. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by D
r. P
rof.
Tam
ara
Shig
anov
a on
11/
13/1
0. F
or p
erso
nal u
se o
nly.





























