Embed Size (px)
Citation preview

PALAIOS, 2012, v. 27, p. 78–89
Research Article
DOI: 10.2110/palo.2011.p11-058r
CAPITANIAN (MIDDLE PERMIAN) MASS EXTINCTION AND RECOVERY IN WESTERN TETHYS:A FOSSIL, FACIES, AND d13C STUDY FROM HUNGARY AND HYDRA ISLAND (GREECE)
PAUL B. WIGNALL,1* DAVID P. G. BOND,1 JANOS HAAS,2 WEI WANG,3 HAISHUI JIANG,4 XULONG LAI,4
DEMIR ALTINER,5 STEPHANIE VEDRINE,6 KINGA HIPS,2 NORBERT ZAJZON,7 YADONG SUN,4 andROBERT J. NEWTON 1
1School of Earth and Environment, University of Leeds, Leeds LS2 9JT, United Kingdom, [email protected], [email protected], [email protected];2Geological Research Group of the Hungarian Academy of Sciences, Budapest, Hungary, [email protected], [email protected]; 3Nanjing Institute of Geology and
Paleontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, People’s Republic of China, [email protected]; 4Key Laboratory of
Geobiology and Environmental Geology, China University of Geosciences, Wuhan 430074, People’s Republic of China, [email protected], [email protected];5Marine Micropalaeontology Research Unit, Department of Geological Engineering, Middle East Technical University, TR-06531 Ankara, Turkey, [email protected];
67 rue Albert 1er, 45000 Orleans, France, [email protected]; 7Department of Mineralogy and Petrology, University of Miskolc, Miskolc, Hungary,
ABSTRACT
The Capitanian (middle Permian) extinction and recovery event isexamined in carbonate platform settings from western Tethys (Hungaryand Hydra, Greece). The age model for these sections is poorly resolvedand we have constructed a d13C chemostratigraphic correlation scheme,supported by conodont and foraminifer data, which attempts correlationwith the well-dated events in China. This reveals the timing of events wassimilar in all Tethyan regions: extinction losses in the middle of theCapitanian produced late Capitanian assemblages in Hungary and Hydrawith a distinctive late Permian character (for example, they lack largefusulinaceans). There is no evidence for an extinction event at the end ofthe Guadalupian (Capitanian) suggesting that previous claims for an end-Guadalupian mass extinction are based on poorly dated records of a mid-Capitanian event. Base level was stable through much of the middle–latePermian transition with the exception of a major regression within theCapitanian Stage. The subsequent transgression established widespreadshallow-water carbonate deposition, such as the Episkopi Formation inHydra and the Nagyvisnyo Limestone Formation in Hungary.
INTRODUCTION
Until 20 years ago the Permian mass extinction was viewed as aninterval of protracted diversity decline throughout the final ,10 millionyears of the Permian (Newell, 1967; Teichert, 1990; Erwin, 1993) butmost subsequent studies favor rapid extinction in the latest Permian(Wignall and Hallam, 1992; Hallam and Wignall, 1997; Rampino andAdler, 1998). The discovery of a precursor extinction event at the end ofthe middle Permian (Guadalupian) suggests that claims for a prolongeddecline may be the result of conflating the extinction losses of twodiscrete events (Stanley and Yang, 1994; Jin et al., 1994). However,recent analysis of middle and late Permian marine invertebratecommunities led Clapham and Bottjer (2007) to conclude that diversitywas in long-term decline from the middle to the end of the Permian; afinding that reasserts the pre-1990s views of Newell and Teichert. LatePermian communities are also claimed to show a progressively moreMesozoic appearance as mollusk groups (bivalves and gastropodsespecially) became proportionally more important at the expense ofrhynchonelliform brachiopods (Clapham and Bottjer, 2007; Isozakiet al., 2007; Clapham et al., 2009). The status of a discrete Guadalupianmass extinction has thus been questioned (Fraiser et al., 2011).However, detailed studies of sections in South China have shown thatthere is a sharp extinction crisis for many groups within the middle ofthe Capitanian, the final stage of the Guadalupian (Wang andSugiyama, 2000; Wignall et al., 2009a, 2009b; Shen and Shi, 2009;
Bond et al., 2010a, 2010b). The loss of many fusulinacean foraminifersis constrained to the mid-Capitanian Jinogondolella altudaensisconodont Zone, an interval that coincides with the onset of eruptionof the Emeishan large igneous province in southwestern China (Wignallet al., 2009a). Similarly, many of the dominant large brachiopods of theearly Capitanian were extinct by the late Capitanian (Shen and Shi,2009). Brachiopod diversity recovered significantly in the Lopingian(e.g. Shen et al., 2010). The data compilation exercise of Clapham andPayne (2011) reveals that 41% of the 286 extant Capitanianbrachiopods failed to survive beyond the Stage. The extinction levelamong other groups is less well known but the generic extinctionintensity of rugose corals was twice that of the brachiopods (Wang andSugiyama, 2000).
Eustatic sea level probably reached its Phanerozoic low point aroundthe Guadalupian-Lopingian boundary (Hardenbol et al., 1998; Hallamand Wignall, 1999). It marks, for example, the point of peak regressionin the Paleozoic, carbonate-dominated successions of the Middle East(Weidlich and Bernecker, 2003; Al-Husseini and Matthews, 2010). Theapparent close temporal association between the extinction losses andthis major regression has led many to attribute the marine crisis to theloss of shallow shelf habitat area (Jin et al., 1994; Hallam and Wignall,1997, 1999; Lai et al., 2008; Bond and Wignall, 2009; Chen et al., 2009).However, recent conodont biostratigraphic dating in China has shownthe age of this major sequence boundary to be precisely constrained towithin the Jinogondolella xuanhanensis Zone of the middle Capitanian(Wignall et al., 2009b; Bond et al., 2010a; Sun et al., 2010; Fig. 1), this isone zone later than the mass extinction level in this region. Otherstudies have suggested cooling-driven extinction (Isozaki et al., 2007)or a crisis directly linked to the onset of Emeishan volcanism (Wignallet al., 2009a, 2009b).
This paper aims to examine the nature of faunal and sedimentarychanges in the Capitanian to Wuchiapingian interval in westernTethyan locations of Hungary and Greece (Fig. 2) and compare themwith changes in the well-known sections of South China. The nature ofthe post-extinction assemblages is also considered; are they typicalPermian types, or do they have high proportions of mollusks suggestinga transitional Mesozoic character (Clapham and Bottjer, 2007;Clapham et al., 2009)?
Detailed facies changes have been studied in four sections to evaluaterelative sea level and provide a context for fossil range charts: both areimportant factors in understanding this extinction event. Foraminifersand calcareous algae are the dominant bioclasts in Guadalupian Tethyancarbonates and their occurrence is strongly controlled by paleobathy-metry (e.g., Vachard et al., 2003; Insalaco et al., 2006). Consequently,rapid facies changes produce rapid compositional changes, and it istherefore important to constrain facies variation when evaluating rangecharts of these groups.* Corresponding author.
Copyright G 2012, SEPM (Society for Sedimentary Geology) 0883-1351/12/0027-0078/$3.00
Facies control of fossil distribution has also bedevilled biostrati-graphic study of many Permian limestone formations because dating isoften attempted using benthic groups such as foraminifers andostracodes. The result is conflicting age determinations. Carbon isotopechemostratigraphy offers an alternative approach to dating thanks tothe recent establishment of a Capitanian-Wuchiapingian d13C curvefrom carbonates in South China and the Panthalassa Ocean (Bondet al., 2010a). Thus, the d13C record of our study sections has beenanalyzed in combination with conodont sampling. Conodonts providethe most widely applicable and highest resolution zonal scheme for the
Permian and, although they are generally rare in the shallow-watercarbonate facies studied here, the occasional find has provided tiepoints for our chemostratigraphic correlation.
STUDY AREAS
Bukk Mountains, Hungary
The ,250-m-thick Nagyvisnyo Limestone Formation of the BukkMountains in northern Hungary contains abundant middle–late
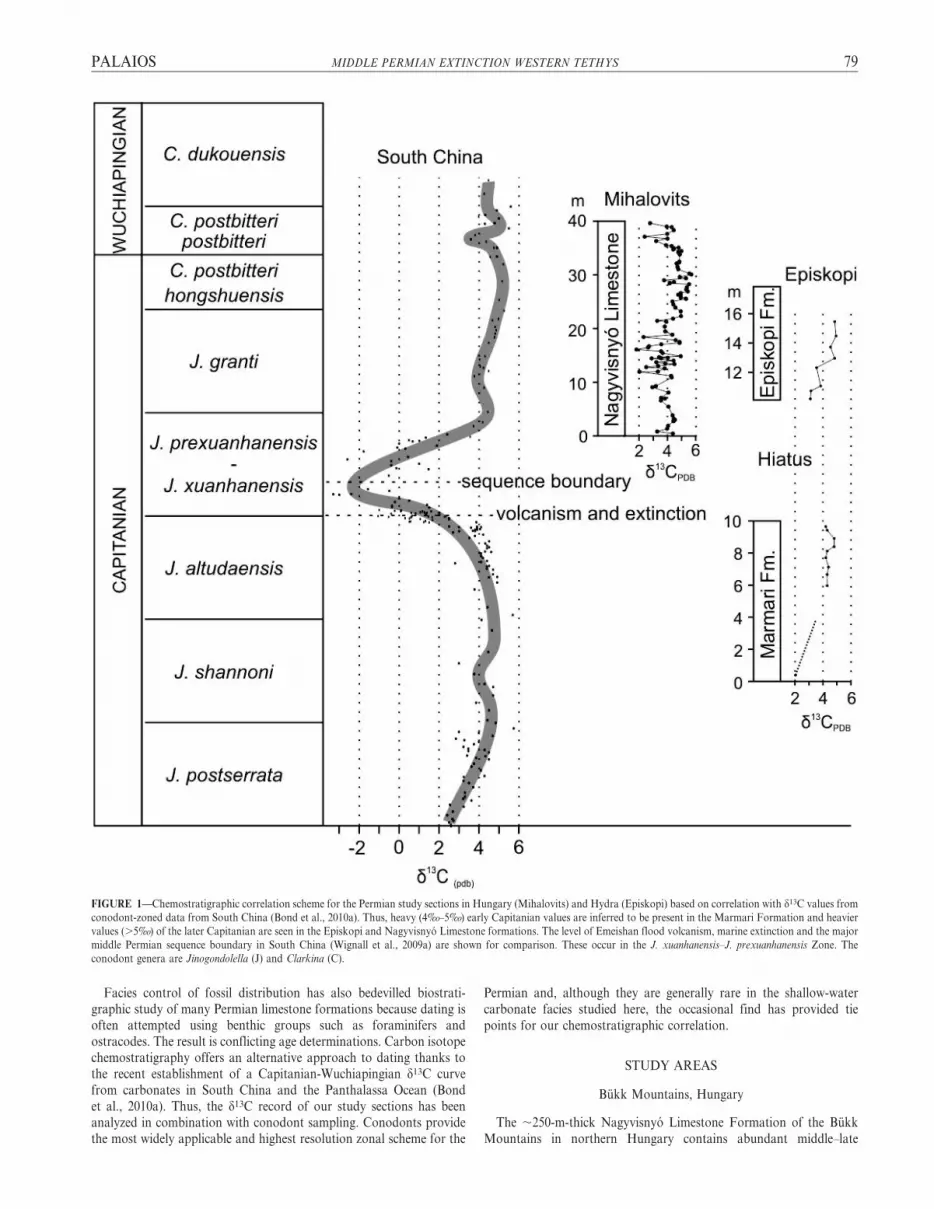
FIGURE 1—Chemostratigraphic correlation scheme for the Permian study sections in Hungary (Mihalovits) and Hydra (Episkopi) based on correlation with d13C values from
conodont-zoned data from South China (Bond et al., 2010a). Thus, heavy (4%o–5%o) early Capitanian values are inferred to be present in the Marmari Formation and heavier
values (.5%o) of the later Capitanian are seen in the Episkopi and Nagyvisnyo Limestone formations. The level of Emeishan flood volcanism, marine extinction and the major
middle Permian sequence boundary in South China (Wignall et al., 2009a) are shown for comparison. These occur in the J. xuanhanensis–J. prexuanhanensis Zone. The
conodont genera are Jinogondolella (J) and Clarkina (C).
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 79

Permian taxa in dark, pyritic limestones (Fulop, 1994; Berczi-Makket al., 1995; Haas et al., 2004). In the lower member of the formation(50–100 m) dolomite and limestone beds alternate, showing an upward-decreasing trend of dolomitization. Hermatypic corals (Waagenophyl-lum) and calcareous sponges were found in the topmost bed of thelower member. The middle member (Mihalovits Member; 60–80 mthick) is characterized by dark-gray to black limestone, punctuated bythin shale layers. It is rich in calcareous algae, foraminifera, andostracodes; bivalves, gastropods, brachiopods, and nautiloids alsooccur. The upper member of the formation (60–80m) is typified by shaleand clayey nodular limestone interbeds. These beds are rich incalcareous algae, foraminifera, ostracodes, and in some beds, brachio-pods (Leptodus) and mollusks; trilobites also occur. The formation wasformed in a euphotic, shallow marine environment that was intermit-tently restricted. The faunal assemblage suggests usually normal marinebut periodically high salinity conditions. The rich biota and the low-energy depositional environment led to reducing diagenetic conditionsbelow the water/sediment interface and accumulation of organic-richand pyritic deposits. The precise age of this unit has proven difficultto constrain. Berczi-Makk et al. (1995) suggest a Capitanian toWuchiapingian age based on foraminifer species, whereas Kozur(1985) suggests a latest Permian (Changhsingian) age for the upperpart of the formation based on ostracodes. In contrast, Thery et al.(2007) have argued that the Nagyvisnyo Limestone belongs entirely tothe Changhsingian Stage based on the occurrence of Paradagmaritaand Multidiscus.
We have examined a 40-m-thick section in the Nagyvisnyo Limestonein an abandoned quarry at Nagyvisnyo (Mihalovits Quarry) (Fig. 3).This is one of the lowest sections available in the formation in the BukkMountains and it potentially records the Capitanian/Wuchipingiantransition. A total of 70 samples was collected for thin-section analysisof microfacies and microfossil identification, 90 samples were collected
for C and O isotope analysis, and 20 large samples were collected forconodont extraction.
Hydra Island (Greece)
Nearly 500 m of Permian strata, mostly limestone, are present on theGreek island of Hydra (Idhra) with the Guadalupian-Lopingianinterval reported from the Marmari and Episkopi formations(Romermann, 1968, 1969; Grant et al., 1991). Thanks to silicificationthe fauna of the Episkopi Formation is well preserved and thebrachiopods and ostracodes in particular are well studied (Grant et al.,1991; Grant, 1994; Crasquin-Soleau and Baud, 1998). The strataaround the boundary between the Marmari and Episkopi formationshave been logged in detail at Cap Bisti, Episkopi, and Lehusis on thesouthwest coast of the island (Fig. 4). A total of 71 thin sections wasanalyzed for their microfacies and microfossil content, and the d13C andd18O values of 18 samples straddling the formational boundary atEpiskopi were obtained.
Both the Marmari and Episkopi formations thicken toward thesouthwest of Hydra. Thus, the Marmari Formation is 117 m thick atLehusis, 145 m thick at Episkopi and 178 m thick 0.7 km to thesouthwest of this last location (Grant et al., 1991). The same thicknesstrend is seen in the Episkopi Formation: it is 200 m thick at its typelocality but thins to ,10 m over a distance of 10 km to the northwest(Grant et al., 1991). These thickness variations suggest half-graben stylestructural control on deposition. However, for much of the depositionalhistory sedimentation remained close to base level.
The Marmari Formation consists of two, extensively dolomitized,peritidal cycles (Baud et al., 1990) with an assemblage of brachiopods,calcareous algae and ostracodes that is of low diversity compared tothat found in the overlying Episkopi Formation (Grant et al., 1991;Crasquin-Soleau and Baud, 1998; Vachard et al., 2008). There are onlya few biostratigraphically useful taxa, but the presence of Neoschwa-
gerina margaritae in the middle and upper part of the formationsuggests a Wordian-Capitanian age (Baud et al., 1990; Grant et al.,1991), and also a pre-Capitanian extinction assemblage.
The contact between the Marmari and Episkopi formations iserosional, and in southwestern Hydra the basal bed of the EpiskopiFormation is a calcirudite composed of reworked clasts of the MarmariFormation (Grant et al., 1991; Jenny et al., 2004). The duration of thehiatus at this contact is difficult to precisely constrain, but the Yabeina
globosa fusulinid Zone is absent, implying much of the CapitanianStage is missing (Grant et al., 1991; Jenny et al., 2004). The presence ofParaglobivalvulina mira and Codonofusiella aff. schubertelloides in thebasal 90 m of the Episkopi Formation is used by Baud et al. (1990,
FIGURE 3—Location of the middle-upper Permian Mihalovits Quarry section in
northern Hungary.
FIGURE 4—Location of middle-upper Permian study sections in Hydra, Greece.
FIGURE 2—Global paleogeography of the middle Permian showing location of the
two study areas in western Tethys.
80 WIGNALL ET AL. PALAIOS

p. 195) and Jenny et al. (2004) to suggest ‘‘a late? Midian’’(5Capitanian) age. However, Codonofusiella populations are typicallyrecorded from the Wuchiapingian, and conodonts diagnostic of thisStage were obtained in a sample 80 m from the top of the formation(Nestell and Wardlaw, 1987). The Marmari/ Episkopi erosional contacttherefore marks either an intra-Capitanian hiatus or a hiatus straddlingthe Capitanian-Wuchiapingian boundary. The level of the Capita-nian mass extinction, identified in South China at the end of the J.altudaensis Zone (Wignall et al., 2009a), could be recorded in the upperMarmari Formation or it may be missing at the unconformable contact.
AGE OF STUDY UNITS
Biostratigraphic Evidence
Attempts to extract conodonts from Nagyvisnyo Limestone sampleswere generally unsuccessful; only a few fragments were found, with theexception of a well-preserved specimen of Jinogondolella granti from theupper part of the section (Fig. 5). This indicates the presence of the lateCapitanian J. granti Zone at this level. The occurrence of Rectostipulinaquadrata around this level further supports this age assignment becausethis foraminifer appears and is abundant in post-extinction assemblagesof the J. granti Zone in South China (Wignall et al., 2009b; Bond et al.,2010a). R. quadrata was also found to be common in the basal EpiskopiFormation in all three study sections in Hydra.
Carbon Isotope Analysis and Results
Well-screened, whole-rock carbonate (mostly micrite) samples fromEpiskopi and Mihalovics were measured for C and O isotopes at thestable isotope laboratory of the Nanjing Institute of Geology andPalaeontology (MAT 253). CO2 was generated by the addition of an-hydrous phosphoric acid to around 5 mg of powdered whole rock in avacuum. Values have been corrected with standard methods and arereported relative to the Vienna Pee Dee belemnite (VPBD) standard.The analytical precision for this analysis, based on replicate analyses ofan in-house strontium carbonate standard, is ,0.1%o.
The d13C results from Mihalovits quarry show stable values around+4%o throughout much of the section, with values falling to +2%o–+3%o
between 11 and 17 m and above 35 m in the section (Figs. 1, 5). Thed18O values are also stable, and there is no correspondence between theminor fluctuations of the two isotopic curves, suggesting little dia-genetic influence.
The d13C results from the Marmari Formation of the Episkopi sectionshow an initial value of 2%o rising to stable values of 4.5%o in the topmost4 m (Figs. 1, 6). Values fall sharply to 3%o across the unconformablecontact at the base of the Episkopi Formation and then rise rapidly to5%o in the overlying 5 m of section. The O isotope values also show asharp fall across the formation boundary, but the second orderfluctuations do not correspond to variations in d13C values. The Cisotope record at Episkopi is therefore considered to be a primary one.
Chemostratigraphic Correlation
Using the occurrence of J. granti (and R. quadrata) as a tie point, it ispossible to align the Mihalovits d13C curve with the late Capitanianrecord from South China (Fig. 1). In this tentative correlation, the minornegative excursion at the top of the Hungarian section equates with anexcursion of similar size and magnitude seen in the base of theWuchiapingian in China (Wang et al., 2004). No conodonts have beenobtained from the Hydra sections, but the presence of abundant R.quadrata at the base of the Episkopi Formation suggests a J. granti Zoneage for this level; a correlation that accords with the similar d13C record.
By Phanerozoic standards, the Capitanian Stage is marked by heavyd13C values, and Isozaki et al. (2007) has called the +4%o plateau theKamura event. The +4%o values in the Marmari Formation are taken to
indicate a Capitanian age (Fig. 1), in accord with the age determinedusing foraminifer taxa (Baud et al., 1990; Grant et al., 1991). The rapidrise of d13C values in the basal Episkopi Formation can be matchedwith the rise seen around the J. xuanhanensis–J. granti zonal boundaryin the Bond et al. (2010a) curve. However, the major intra-Capitaniannegative excursion of this curve is missing in Hydra, presumably be-cause of a hiatus at the formation contact (Fig. 1).
The chemostratigraphic correlation proposed here allows the baseEpiskopi unconformity to be correlated with the major J. xuanhanensissequence boundary in China (Wignall et al., 2009b; Bond et al., 2010a;Fig. 1). The subsequent transgression saw the re-establishment ofthe platform carbonates of the Episkopi Formation. The onset ofNagyvisnyo Limestone Formation deposition in northern Hungary isprobably a similar response to base-level rise in the late Capitanian.
HUNGARY: FACIES AND FOSSILS
The Nagyvisnyo Limestone Formation in the Mihalovits Quarryconsists of tabular bedded wackestone and packstones ranging from 0.2to 3.0 m thick, separated by thin (1–10 cm thick) beds of calcareousshales (Fig. 5). Most limestone beds are massive but some show gradingwith bioclasts concentrated at the base and more micritic tops bur-rowed by Psilonichnus. Four facies types are recognized: (A) mollusk-algal-echinoderm packstone (Fig. 7A), (B) algal wacke-packstone(Fig. 7B), (C) echinoderm wackestone/packstone (Fig. 7C), and (D)ostracode wackestone (Fig. 7D). Facies A–C are very similar, beingdistinguished by their varying proportions of the same bioclasts (seebelow). Facies D is distinctly different because it contains fewer bioclasttypes, predominantly ostracode valves, and has patches of anhydritecrystals replaced by calcite (Fig. 7D).
With the exception of facies D, the limestone beds contain a diverseassemblage consisting of foraminifers, calcareous algae, Tubiphytesobscurus, echinoderm material (especially crinoids), brachiopods, ostra-codes, microgastropods, and bivalves. The foraminifers include rarefusulinaceans, of which only Codonofusiella is identifiable, while smallervarieties, mostly lagenides and miliolinids, are much more abundant andinclude Agathammina, Diplosphaerina, Globivalvulina, Hemigordius,Nodosinelloides, and Pachyphloia (Fig. 5). These are all genera knownto have survived the Capitanian mass extinction (Bond and Wignall,2009). Calcareous algae are the most abundant bioclasts in theMihalovits Quarry section and assemblages are dominated by Permo-calculus spp., Mizzia spp., and Gymnocodium bellerophontis that rangethroughout the section (Figs. 5, 7B). Such dominance by calcareousalgae is typical of Permian carbonates, however gastropods and bivalvesare dominant in beds of facies A. Mollusk groups are more typical ofMesozoic assemblages and in this regard the Nagyvisnyo Limestone hassome beds with the mixed Permian-Mesozoic faunal character suggestedfor late Permian carbonates by Clapham and Bottjer (2007).
In summary, the Mihalovits Quarry record consists of moderatelydiverse fossil assemblages that lack Capitanian mass extinction victimssuch as schwagerinids and neoschwagerinid fusulinaceans. This suggeststhat the Capitanian crisis predates the J. granti Zone in Hungary.Bioclastic packstone deposition was punctuated by intervals ofevaporation and anhydrite growth supporting previous restricted,lagoonal interpretations (Berczi-Makk et al., 1995) as do the absenceof some stenohaline groups such as ammonoids and bryozoans and therare occurrence of corals (Fulop, 1994). However, echinoderms,conventionally regarded as a stenohaline group, are present throughout.
HYDRA: FACIES AND FOSSILS
Facies Trends
The uppermost Marmari Formation consists of pale gray–whitebeds that contrast with the darker gray limestones of the Episkopi
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 81

Formation. There is modest lateral facies variation between the threesections and all show low diversity compared with the overlying EpiskopiFormation (Figs. 6, 8–9). Algal-ostracode wackestones and packstonesdominate at Lehusis and Cap Bisti. These facies are also present at
Episkopi but beds of pelmicrite are more common. In the topmost bedsat Cap Bisti some bioclasts are replaced by chert-calcite intergrowths.
The nature of the transition between the Marmari and Episkopiformations varies substantially between sections. At Cap Bisti the
FIGURE 5—Section in the lowermost Nagyvisnyo Limestone Formation at Mihalovics quarry showing an alternating succession of the four facies: a 5 mollusk-algal-
echinoderm packstone, b 5 algal wacke-packstone, c 5 echinoderm packstone, d 5 ostracode wackestone; and C and O isotope data and range charts for foraminifers and
calcareous algae. Photograph shows Jinogondolella granti obtained from sample 68, scale bar is 100 mm.
82 WIGNALL ET AL. PALAIOS

uppermost bed of algal-ostracode wackestone in the MarmariFormation is deeply karstified with a network of vertical and horizontalfissures several decimeters in width penetrating up to 3 m downward(Fig. 8). This is infilled and overlain by a 2-m-thick bed of clast-supported breccia consisting of reworked clasts of the underlying beds.This bed is in turn partly truncated by erosion at the base of a furtherbed of clast-supported breccia with clasts approaching one meter indiameter. Reworked Marmari clasts continue to occur up to 4.5 mabove the base of the overlying Episkopi Formation where they arepresent as rounded, floating pebbles in a dark matrix of echinodermpackstones and pelsparites. Clast orientation varies from random, highangle orientations to bedding parallel pebble trains that do not showimbrication. Further east, at Episkopi, evidence for erosion of thetopmost Marmari Formation is less spectacular but reworked clasts arepresent in the uppermost part of the formation although coarse,angular pebbles are not present (Fig. 7E). Instead, rounded pebblesoccur either in sharp-based beds of conglomerate or as floating clastsin a bioclastic packstone or grainstone matrix. At the easternmost lo-cation (Lehusis), the formation contact is seen as an irregular surface,
with ,10 cm of erosional relief between the white Marmari Formationand the dark-gray Episkopi Formation. A few, small rounded Marmaripebbles are present immediately above this surface, but the conglom-erates and breccias of the more western localities are not seen at Lehusis(Fig. 9).
The onset of deposition of the Episkopi Formation saw the establish-ment of a uniform facies style in southwestern Hydra: foraminifer-algalpackstones dominate the basal beds at all three locations, althoughmollusk clasts are not uncommon (Fig. 7F). Higher levels, between 70and 100 m above the basal contact, were sampled at Episkopi. Thisrevealed that this facies persists, albeit with a larger component of largebioclasts (colonial Rugosa, calcareous sponges, and brachiopods).
Fossil Ranges
The uppermost Marmari beds sampled here contain a low diversityfauna of ostracodes, calcareous algae and foraminifers (Fig. 10). Theforaminiferal records are restricted to sparse occurrences of smaller
FIGURE 6—Episkopi section showing d13C and d18O fluctuations in the uppermost Marmari and lower Episkopi formations and foraminifer and calcareous algal range
charts. No extinction horizon is identifiable and so this section is considered to postdate the middle Capitanian crisis.
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 83

forms (mostly lagenides), including Dagmarita, Earlandia, Eotuberitina,Glomomidiella, Hemigordius, Nodosinelloides, and Pachyphloia. Anindeterminate staffellid fusulinacean (probably Staffella) was recoveredfrom sample EK6 at Episkopi, where it ranges up into the overlyingEpiskopi Formation (Fig. 10L); a unit with an abundant and diverseforaminifer assemblage including those genera listed above, pluscommon Agathammina pusilla, Climacammina, Diplosphaerina in-equalis, Geinitzina, Globivalvulina, Palaeotextularia, Pseudomidiella,Rectostipulina quadrata (and lesser R. pentamerata), Retroseptellina,as well as numerous less common genera (Fig. 10). Fusulinaceanforaminifera are limited to the smaller genera Codonofusiella andReichelina, plus Palaeofusulina and Staffella (?). Codonofusiella andReichelina are biostratigraphically useful because, although they arepresent before the Capitanian extinction in South China, they come todominate the fusulinacean components of post-extinction faunas in thatarea. Their presence in the Episkopi Formation, together with anabsence of fusulinacean extinction casualties, indicates that these strataare of post-extinction age, in keeping with its sequence- and chemo-stratigraphic age assignment. The extinction aftermath attributes of theHydra assemblages are further supported by the common Earlandia
and Diplosphaerina; a typical post-extinction disaster assemblage (e.g.,Groves and Altiner, 2005; Bond et al., 2010a, 2010b).
DISCUSSION
Relative Sea-Level Change
The overall uniformity of facies types in the Mihalovits sectionsuggests that there were no major changes of palaeobathymetry in thislate Capitanian record. The Hungarian record is here correlated withthe lower Episkopi Formation of Hydra (Fig. 1) that similarly showspersistent, uniform deposition in a shallow, platform setting. Incontrast, the sedimentological evidence from the Marmari-Episkopiboundary clearly records a major regression. At Cap Bisti, the mostwesterly location, sea level fell sufficiently for emergence and karsti-fication of the upper Marmari Formation to occur, but at the moreeasterly locations, the upper surface is only eroded to a depth of a fewcentimeters, which may have occurred in a shallow marine setting. Thecoarse breccia beds at Cap Bisti are probably a subaerial scree slopefacies, perhaps part of a fan conglomerate, but this facies is absent in
FIGURE 7—Photomicrographs of typical facies types at Mihalovits Quarry (A–D) and Hydra (E–F). Facies A) Mollusk-algal-echinoderm packstone, showing Permocalculus
cf. tenellus grains (5A), echinoderm grain (5E) and mollusk (5M), likely a gastropod. B) Algal wacke-packstone dominated by Gymnocodium bellerophontis, a complete thallus
is labelled A. C) Echinoderm wackestone. D) Ostracode wackestone, although the patch illustrated consists of anhydrite crystals now pseudomorphed by calcite and showing
relict zonation. E) The upper two-thirds of the field of view is a clast of calcareous algal packstone in a micrite matrix, topmost Marmari Formation, Episkopi. F) Bioclastic
packstone/grainstone showing mollusk clasts (thick-shelled bivalves (5B) and microgastropods (5M)), calcareous algae (5A) and peloids (5Pe), lower Episkopi Formation,
Episkopi. Width of field of view is 6 mm for all images.
84 WIGNALL ET AL. PALAIOS

the east. The scree slope facies is overlain by shallow marine limestoneswith rounded pebbles of Marmari Formation at Cap Bisti. This faciescharacterizes the uppermost Marmari Formation to the east. Therounding of pebbles suggests beach reworking prior to transport into ashallow marine setting.
A Capitanian Mass Extinction in Western Tethys?
The fossils of the Nagyvisnyo Limestone and Episkopi formationshave distinctive post-Capitanian extinction attributes, while extinctionvictims (large fusulinaceans) are only known from low in the Marmari
FIGURE 8—Sedimentary log of the Cap Bisti section, westernmost Hydra Island showing a deeply karstified surface in the uppermost Maramari Formation and microfossil range chart.
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 85

Formation (Baud et al., 1990; Grant et al., 1991; Jenny et al., 2004). Themass extinction level therefore occurs, either within the upper MarmariFormation, or at the boundary with the Episkopi Formation. UpperMarmari assemblages are impoverished and of similar composition topost-extinction assemblages in South China (Lai et al., 2008; Wignall
et al., 2009b; Bond et al., 2010a). This could reflect an unfavorableenvironment because the facies at this level record peritidal/supratidalconditions (Grant et al., 1991; Jenny et al., 2004). The carbon isotopevalues from the upper Marmari Formation suggest an early Capitanianage (J. altudaensis Zone or older), which places this strata below the
FIGURE 9—Sedimentary log and microfossil range chart from the Lehusis section, south Hydra Island.
86 WIGNALL ET AL. PALAIOS

extinction level seen in China (Fig. 1). Therefore, it is likely that theCapitanian mass extinction occurred during the hiatus recorded by thesequence boundary between the Marmari and Episkopi formations.The clear signal of a major intra-Capitanian regression, in a stageotherwise characterized by little eustatic change, occurs in both Chinaand Hydra. In Hydra the regression and extinction may coincide,suggesting a possible causal connection (e.g., Jin et al., 1994; Hallamand Wignall, 1999; Lai et al., 2008). However, in China, the regressionslightly post-dates the extinction event, and a more circumspect
conclusion to be drawn from the western Tethyan record would bethat the extinction occurs during the hiatus caused by base level fall.
The Nature of Late Permian Assemblages
The contention that post-Capitanian extinction assemblages weremore mollusk-rich than earlier Permian times (Clapham and Bottjer,2007) receives some support from this study, particularly in themollusk-dominated facies A of the Nagyvisnyo Limestone. However,
FIGURE 10—Foraminifera from Episkopi, Hydra. A) Dagmarita chanakchiensis (sample EK14); B) Diplosphaerina sp. (EK19); C) Diplosphaerina reitlingerae (EK14); D)
Geinitzina araxensis (EK14); E) several Hemigordiopsis sp. (EK14); F) Hemigordius irregulariformis (EK13); G) Pachyphloia ovata (EK14); H) Pseudomidiella sp. (EK16); I)
Rectostipulina pentamerata (EK15); J) Rectostipulina quadrata (EK15); K) Reichelina sp. (EK19); L) Staffellid fusulinacean (indet, EK14). All scale bars 5 100 mm except E, H,
and L 5 300 mm.
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 87

the occasional presence of evaporite pseudomorphs in this sectionsuggests salinity restriction occurred at least periodically: conditions thatwill have favored euryhaline mollusks. In other regards the microfossilcontent of the normal marine assemblages is dominated by typicalPermian bioclasts such as calcareous algae and foraminifers. Theabundant and diverse Episkopi ostracode fauna is similarly typical ofshallow-water assemblages of late Permian age (Crasquin-Soleau andBaud, 1998). Thus, the ecological transition proposed to have occurredbetween the middle and late Permian, as a possible consequence of theCapitanian mass extinction, is only manifest in the reported increase inthe dominance of mollusks in units such as the Episkopi Formation(Clapham and Bottjer, 2007). In all other regards, the palaeoecologicalattributes of the assemblages are clearly Permian. It has also been arguedthat the Episkopi brachiopods have peculiar attributes more typical ofolder Paleozoic assemblages (Grant et al., 1991, p. 491) but more recentappraisal shows that they are typical Wuchiapingian taxa with a strongsimilarity to eastern Tethyan assemblages of South China and with alesser number of Perigondwanan taxa (Shen and Clapham, 2009).
CONCLUSIONS
Western Tethyan Capitanian-Wuchiapingian sections of Hungaryand Hydra (Greece) reveal that a typical post-Capitanian massextinction fauna is developed from the late Capitanian and persistswithout faunal break into the Wuchiapingian. This is comparable to thepaleontological record seen in South China (where conodonts define arefined zonation scheme) that shows the extinction event happenedaround the top of the J. altudaensis Zone. Sporadic conodonts, fora-minifers, and correlation of d13C records reveal that the timing of thecrisis is likely to be similar in western Tethys, although the precise levelis difficult to pin down in the extensively dolomitized middle MarmariFormation of Hydra. In Hungary, the extinction occurs below theNagyvisnyo Limestone Formation studied here.
A major regression within the Capitanian, seen in China, is alsorecorded by the major base-level fall recorded at the boundary betweenthe Marmari and Episkopi formations in Hydra. This is the only majorbase-level fluctuation in the middle–late Permian in Hydra and itswidespread development (i.e., occurrence in Hungary, Greece, andChina) clearly argues for a eustatic origin. Subsequent base level risesaw the establishment of platform carbonate deposition with a diversefauna of typical Permian aspect seen in both Hydra and Hungary.
ACKNOWLEDGMENTS
We thank Helen Cope for field assistance, Csaba Pero for infor-mation that facilitated fieldwork in Hungary, and Daniel Vachard forhelp with identifying calcareous algae. Matthew Clapham and JohnGroves are thanked for their reviews. This research was supported bythe following grants: Natural Environment Research Council grantNE/D011558/1 to Paul Wignall; National Natural Science Foundationof China (NSFC) and BK2010022 (Jiangsu Province) grants to WangWei; Natural Science Foundation of China (grant 40872002) andChinese State Administration of Foreign Experts Affairs (GrantB08030) to Xulong Lai.
REFERENCES
AL-HUSSEINI, M.I., and MATTHEWS, R.K., 2010, Calibrating Mid-Permian to Early
Triassic Khuff sequence with orbital clocks: Geoarabia, v. 15, p. 171–206.
BAUD, A., JENNY, C., PAPANIKOLAOU, D., SIDERIS, C., and STAMPFLI, G., 1990, New
observations on Permian stratigraphy in Greece and geodynamic interpretation:
Bulletin of the Geological Society of Greece, v. 25, p. 187–206.
BERCZI-MAKK, A., CSONTOS, L., and PELIKAN, P., 1995, Data on the (Upper Permian)
Foraminifer fauna of the Nagyvisnyo Limestone Formation from borehole
Malyninka-8 (Northern Hungary): Acta Geologica Hungarica, v. 38, p. 185–250.
BOND, D.P.G., and WIGNALL, P.B., 2009, Latitudinal selectivity of foraminifer
extinctions during the late Guadalupian crisis: Paleobiology, v. 35, p. 465–483.
BOND, D.P.G., WIGNALL, P.B., WANG, W., IZON, G., JIANG, H.-S., LAI, X.-L., SUN,
Y.-D., NEWTON, R.J., SHAO, L.-Y., VEDRINE, S., and COPE, H., 2010a, The mid-
Capitanian (Middle Permian) mass extinction and carbon isotope record of
South China: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 292,
p. 282–294.
BOND, D.P.J., HILTON, J., WIGNALL, P.B., ALI, J.R., STEVENS, L.G., SUN, Y.-D., and
LAI, X.-L., 2010b, The Middle Permian (Capitanian) mass extinction on land and
in the oceans: Earth-Science Reviews, v. 109, p. 100–116.
CHEN, Z.Q., GEORGE, A.D., and YANG, W.-R., 2009, Effects of Middle-Late Permian
sea-level changes and mass extinction on the formation of the Tieqiao skeletal
mound in the Laibin area, South China: Australian Journal of Earth Sciences,
v. 56, p. 745–763.
CLAPHAM, M.E., and BOTTJER, D.J., 2007, Prolonged Permian-Triassic ecological
crisis recorded by molluscan dominance in Late Permian offshore assemblages:
Proceedings of the National Academy of Sciences, v. 104, p. 12971–12975.
CLAPHAM, M.E., and PAYNE, J.L., 2011, Acidification, anoxia, and extinction:
A multiplelogistic regression analysis of extinction selectivity during the Middle
and Late Permian: Geology, v. 39, p. 1059–1062.
CLAPHAM, M.E., SHEN, S.-Z., and BOTTJER, D.J., 2009, The double mass extinction
revisited: Reassessing the severity, selectivity, and causes of the end-Guadalupian
biotic crisis (Late Permian): Paleobiology, v. 35, p. 32–50.
CRASQUIN-SOLEAU, S., and BAUD, A., 1998, New Permian ostracods from Greece
(Hydra Island): Journal of Micropalaeontology, v. 17, p. 131–152.
ERWIN, D.H., 1993, The Great Paleozoic Crisis: Life and Death in the Permian:
Columbia University Press, New York, 327 p.
FRAISER, M.L., CLAPHAM, M.E., and BOTTJER, D.J., 2011, Mass extinctions and
changing taphonomic processes, in Allison, P.A., and Bottjer, D.J., eds.,
Taphonomy: Process and Bias Through Time: Springer, Dordrecht, Heidelberg,
London, New York, p. 569–590.
FULOP, L.L., 1994, Geology of Hungary, Paleozoic II: Akademia Kiado, Budapest,
p. 222–229. [in Hungarian]
GRANT, R.E., 1994, Upper Permian brachiopods of the superfamily Orthotetoidea
from Hydra Island, Greece: Journal of Paleontology, v. 69, p. 655–670.
GRANT, R.E., NESTELL, K., BAUD, A., and JENNY, C., 1991, Permian stratigraphy of
Hydra Island, Greece: PALAIOS, v. 6, p. 479–497.
GROVES, J.R., and ALTINER, D., 2005 Survival recovery of calcareous foraminifera
pursuant to the end-Permian mass extinction: Comptes Rendus Palevolution, v. 4,
p. 419–432.
HAAS, J., HIPS, K., PELIKAN, P., ZAJZON, N., GOTZ, A.E., and TARDI-FILACZ, E., 2004,
Facies analysis of marine Permian/Triassic boundary sections in Hungary: Acta
Geologica Hungarica, v. 47, p. 297–340.
HALLAM, A., and WIGNALL, P.B., 1997, Mass Extinctions and their Aftermath: Oxford
University Press.
HALLAM, A., and WIGNALL, P.B., 1999, Mass Extinctions and Sea-Level Changes:
Earth-Science Reviews, v. 48, p. 217–250.
HARDENBOL, J., THIERRY, J., FARLEY, M.B., JACQUIN, T., DE GRACIANSKY, P.C., and
VAIL, P.R., 1998, Mesozoic and Cenozoic sequence chronostratigraphic framework
of European basins, in de Graciansky, P.C., Jacquin, T., and Vail, P.R., eds.,
Mesozoic and Cenozoic Sequence Stratigraphy of European Basins: SEPM
(Society for Sedimentary Geology) Special Publication 60, p. 3–13.
INSALACO, E., VIRGONE, A., COURME, B., GAILLET, J., KAMALI, M., MOALLEMI, A.,
LOTFPOUR, M., and MONIBI, S., 2006, Upper Dalan Member and Kangan
Formation between the Zagros Mountains and offshore Fars, Iran: Depositional
system, biostratigraphy and stratigraphic architecture: Geoarabia, v. 11, p. 75–176.
ISOZAKI, Y., KAWAHATA, H., and MINOSHIMA, K., 2007, The Capitanian (Permian)
Kamura cooling event: The beginning of the Paleozoic-Mesozoic transition:
Palaeoworld, v. 16, p. 16–30.
JENNY, C., IZART, A., BAUD, A., and JENNY, J., 2004, Le Permien l’ıle d’Hyrda (Grece),
micropaleontologie, sedimentologie et paleoenvironments: Revue de Paleobiologie,
Geneve, v. 23 (1), p. 275–312.
JIN, Y.-G., ZHANG, J., and SHANG, Q., 1994, Two phases of the end-Permian mass
extinction: Canadian Society of Petroleum Geologists, Memoir, v. 17, p. 813–822.
KOZUR, H., 1985, Biostratigraphic of the Upper Paleozoic conodonts, ostracods and
holothurian sclerites of the Bukk Mts. Part II, Upper Paleozoic ostracods: Acta
Geologica Hungarica, v. 28, p. 225–256.
LAI, X.-L., WANG, W., WIGNALL, P.B, BOND, D.P.G., JIANG, H.-S., ALI, J.R., JOHN,
E.H., and SUN, Y.-D., 2008, Palaeoenvironmental change during the end-
Guadalupian (Permian) mass extinction in Sichuan, China: Palaeogeography,
Palaeoclimatology, Palaeoecology, v. 269, p. 78–93.
NESTELL, M.K., and WARDLAW, B.R., 1987, Upper Permian conodonts from Hydra,
Greece: Journal of Palaeontology, v. 61, p. 758–772.
NEWELL, N.D., 1967, Revolutions in the history of life: Geological Society of America
Special Paper 89, p. 63–91.
88 WIGNALL ET AL. PALAIOS

RAMPINO, M.R., and ADLER, A.C., 1998, Evidence for abrupt latest Permian mass
extinction of foraminifera: Results of tests for the Signor-Lipps effect: Geology, v.
26, p. 415–418.
ROMERMANN, H., 1968, Geologie von Hydra (Griechenland): Geologica et Palaeon-
tologica, v. 2, p. 163–171.
ROMERMANN, H., 1969, Geologie der Insel Hydra (Griechenland): Inaugural-
Dissertation, Phillips Universitat, Marburg, 62 p.
SHEN, S.-Z., and CLAPHAM, M.E., 2009, Wuchiapingian (Lopingian, Late Permian)
brachiopods from the Episkopi Formation of Hydra Island, Greece: Palaeontol-
ogy, v. 52, p. 713–743.
SHEN, S.-Z., and SHI, G.R., 2009, Latest Guadalupian brachiopods for the
Guadalupian/ Lopingian boundary GSSP section at Penglaitan in Laibin,
Guangxi, South China and implications for the timing of the pre-Lopingian crisis:
Palaeoworld, v. 18, p. 152–161.
SHEN, S.-Z., CAO, C.-Q., ZHANG, Y.-C., LI, W.-Z., SHI, G.R., WANG, Y., WU, Y.-S.,
UENO, K., HENDERSON, C.M., WANG, X.-D., ZHANG, H., WANG, X.-J., and CHEN,
J., 2010, End-Permian mass extinction and palaeoenvironmental changes in
Neotethys: Evidence from an oceanic carbonate section in southwestern Tibet:
Global and Planetary Change, v. 73, p. 3–14.
STANLEY, S.M., and YANG, X., 1994, A double mass extinction at the end of the
Paleozoic era: Science, v. 266, p. 1340–1344.
SUN, Y.-D., LAI, X-L., WIGNALL, P.B., WIDDOWSON, M., ALI, J.R., JIANG, H.-S.,
WANG, W., YAN, C.-B., BOND, D.P.G. and VEDRINE, S. 2010. Dating the onset and
nature of the Middle Permian Emeishan large igneous province eruptions in SW
China using conodont biostratigraphy and its bearing on mantle plume uplift
models. Lithos, v. 119, p. 20–33.
TEICHERT, C., 1990, The Permian-Triassic boundary revisited, in Kauffman, E.G., and
Walliser, O.H., eds., Extinction Events in Earth History: Springer-Verlag, Berlin,
p. 199–238.
THERY, J.M., VACHARD, D., and DRANSERT, E., 2007, Late Permian limestones and the
Permian-Triassic boundary: New biostratigraphic, palaeobiogeographical and
geochemical data in Caucasus and eastern Europe, in Alvaro, J.J., Aretz, M.,
Boulvain, F., Munnecke, A., Vachard, D., and Vennin, E., eds., Palaeozoic Reefs
and Bioaccumulations: Climatic and Evolutionary Control: Geological Society of
London, Special Publication 275, p. 255–274.
VACHARD, D., ZAMBETAKIS-LEKKAS, A., SKOURTSOS, E., MARTINI, R., and ZANINETTI,
L., 2003, Foraminifera, algae and carbonate microproblematica from the Late
Wuchiapingian/Dzhulfian (Late Permian) of Peloponnesus (Greece): Rivista
Italiana di Paleontologia e Stratigrafia, v. 109, p. 339–358.
VACHARD, D., RETTORI, R., ANGIOLINI, L., and CHECCONI, A., 2008, Glomomidiella
gen. n. (Foraminifera, Miliolata, Neodiscidae): A new genus from the late
Guadalupian-Lopinginian of Hydra Island (Greece): Rivista Italiana di
Paleontologia e Stratigrafia, v. 114, p. 349–361.
WANG, W., CAO, C.-Q., and WANG, Y., 2004, The carbon isotope excursion on GSSP
candidate section of Lopingian-Guadalupian boundary: Earth and Planetary
Science Letters, v. 220, p. 57–67.
WANG, X.-D., and SUGIYAMA, T., 2000, Diversity and extinction patterns of Permian
coral faunas of China: Lethaia, v. 33, p. 285–294.
WEIDLICH, O., and BERNECKER, M., 2003, Supersequence and composite sequence
carbonate platform growth: Permian and Triassic outcrop data of the Arabian
platform and Neo-Tethys: Sedimentary Geology, v. 158, p. 87–116.
WIGNALL, P.B., and HALLAM, A., 1992, Anoxia as a cause of the Permian/Triassic
extinction: Facies evidence from northern Italy and the western United States:
Palaeogeography, Palaeoclimatology, Palaeoecology, v. 93, p. 21–46.
WIGNALL, P.B., SUN, Y.-D., BOND, D.P.G., IZON, G., NEWTON, R.J., VEDRINE, S.,
WIDDOWSON, M., ALI, J.R., LAI, X.-L., JIANG, H.-S., COPE, H., and BOTTRELL, S.H.,
2009a, Volcanism, mass extinction, and carbon isotope fluctuations in the Middle
Permian of China: Science, v. 324, p. 1179–1182.
WIGNALL, P.B., VEDRINE, S., BOND, D.P.G., WANG, W., LAI, X.-L., ALI, J.R., and JIANG,
H.-S., 2009b, Facies analysis and sea-level change at the Guadalupian-Lopingian
Global Stratotype (Laibin, South China), and its bearing on the end-Guadalupian
mass extinction: Journal of the Geological Society, v. 166, p. 655–666.
ACCEPTED NOVEMBER 29, 2011
PALAIOS MIDDLE PERMIAN EXTINCTION WESTERN TETHYS 89

![Feasibility of using hyperpolarized [1-13C]lactate as a substrate for in vivo metabolic 13C MRSI studies](https://img.pdfslide.net/doc/110x75/6351313d137c5d316803b615/feasibility-of-using-hyperpolarized-1-13clactate-as-a-substrate-for-in-vivo-metabolic.jpg)



























