Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Review
Chemistry of phospholipid oxidation☆
Ana Reis 1, Corinne M. Spickett ⁎,1
School of Life and Health Sciences, Aston University, UK
a b s t r a c ta r t i c l e i n f o
Article history:Received 15 October 2011Received in revised form 14 January 2012Accepted 3 February 2012Available online 9 February 2012
Keywords:Lipid peroxidationRadical oxidationMass spectrometryOxidative lipidomicsNMRInflammation
The oxidation of lipids has long been a topic of interest in biological and food sciences, and the fundamentalprinciples of non-enzymatic free radical attack on phospholipids are well established, although questionsabout detail of the mechanisms remain. The number of end products that are formed following the initiationof phospholipid peroxidation is large, and is continually growing as new structures of oxidized phospholipidsare elucidated. Common products are phospholipids with esterified isoprostane-like structures and chain-shortened products containing hydroxy, carbonyl or carboxylic acid groups; the carbonyl-containing com-pounds are reactive and readily form adducts with proteins and other biomolecules. Phospholipids can alsobe attacked by reactive nitrogen and chlorine species, further expanding the range of products to nitratedand chlorinated phospholipids. Key to understanding the mechanisms of oxidation is the development of ad-vanced and sensitive technologies that enable structural elucidation. Tandem mass spectrometry has provedinvaluable in this respect and is generally the method of choice for structural work. A number of studies haveinvestigated whether individual oxidized phospholipid products occur in vivo, and mass spectrometry tech-niques have been instrumental in detecting a variety of oxidation products in biological samples such as ath-erosclerotic plaque material, brain tissue, intestinal tissue and plasma, although relatively few have achievedan absolute quantitative analysis. The levels of oxidized phospholipids in vivo is a critical question, as there isnow substantial evidence that many of these compounds are bioactive and could contribute to pathology. Thechallenges for the future will be to adopt lipidomic approaches to map the profile of oxidized phospholipidformation in different biological conditions, and relate this to their effects in vivo. This article is part of a Spe-cial Issue entitled: Oxidized phospholipids—their properties and interactions with proteins.
© 2012 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23751.1. Historical perspective on the oxidation of phospholipids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23751.2. Types and sources of oxidants relevant to phospholipid oxidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2375
2. Lipid peroxidation and other types of phospholipid oxidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23762.1. Basic concepts in peroxidation of polyunsaturated fatty acyl chains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23762.2. The diversity of oxidized phospholipid products from peroxidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23762.3. Other pathways of phospholipid oxidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2377
3. Factors affecting oxidized phospholipid formation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23783.1. Influence of liposome size and composition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23793.2. Influence of the oxidant system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23793.3. Selectivity of modification dependent on fatty acyl chain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2379
4. Recent advances in understanding of phospholipid oxidation mechanisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23795. Analysis and quantification of oxidized phospholipids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2381
5.1. Mass spectrometry in the study of oxPL . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23815.2. 31P NMR in the study of oxPL . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2382
Biochimica et Biophysica Acta 1818 (2012) 2374–2387
☆ This article is part of a Special Issue entitled: Oxidized phospholipids—their properties and interactions with proteins.⁎ Corresponding author at: School of Life and Health Sciences, Aston University, Aston Triangle, Birmingham, B4 7ET, UK. Tel.: +44 121 2044085; fax: +44 121 2044175.
E-mail address: [email protected] (C.M. Spickett).1 Both authors contributed equally to the preparation of the manuscript.
0005-2736/$ – see front matter © 2012 Elsevier B.V. All rights reserved.doi:10.1016/j.bbamem.2012.02.002
Contents lists available at SciVerse ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r .com/ locate /bbamem

Author's personal copy
6. Detection and quantification of oxPL in biological systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23836.1. Detection of oxPL may be complicated by issues of stability and metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23836.2. Quantification of oxPL in cells and tissues . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2383
7. Conclusions and future perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2384Abbreviations and definitions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2384Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2384References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2384
1. Introduction
1.1. Historical perspective on the oxidation of phospholipids
In the 21st century the role of lipid peroxidation in disease is still asubject of considerable research, and within this area there is growinginterest in the roles of oxidized phospholipids, which have beenfound in pathological conditions, often at raised levels compared tonormal physiology [1,2]. Studies with isolated cells and tissues havedemonstrated both pro-inflammatory and anti-inflammatory effects,and have identified receptors and signalling pathways that contributeto them [3–6]. The importance of oxidized phospholipids in biochemis-try and pathophysiology is thus well accepted, although many ques-tions about mechanisms and role in specific diseases remainunanswered. The understanding that we have currently is built uponknowledge of the types of oxidized phospholipids that can be formed,which in turn depends on studies of the process of phospholipid oxida-tion and peroxidation, together with constantly improving methodolo-gies for product analysis.
Research on oxidation of phospholipids goes backmore than a centu-ry, with very early studies by Thunberg in 1910 observing “respiration”of phospholipid suspensions and noting that iron salts accelerated theuptake of oxygen in tissues [7]. The term “oxidized phospholipids” wasreported for the first time in 1939 by Frederik and Mary Bernheim, fol-lowing the studies on the action of vanadium salts as catalysts in the ox-idation of phospholipids from heart and brain tissues [8]. Mechanisticstudies of lipid peroxidation were ongoing from the 1940s; the free rad-ical nature of the process was identified and a number of low molecularweight fragmentation products, such asmalonaldehyde, pentanal and 4-hydroxynonenal were identified [9], as will be described later in this ar-ticle. However, until the late 1980s knowledge about the oxidized phos-pholipid products was somewhat limited by the available methods ofstructural analysis. The most structurally-informative method was gaschromatography coupled with mass spectrometry detection (GC–MS),but this required saponification and derivatisation protocols in order togenerate volatile analytes, with resulting loss of information about theoriginal esterified structure, as well as the possibility of artefact genera-tion through extensive sample manipulation. The introduction of fastatom bombardment (FAB) and subsequently electrospray ionisation(ESI) paved the way for analysis of less volatile biomolecules and re-duced thermal degradation, thus enabling structural determination ofmany oxidized phospholipids. At the end of 1980s, a phosphatidylcho-line derivative that was a substrate for the platelet-activating acetylhy-drolase enzyme found in human plasma was synthesized [10], andfound to be 1-palmitoyl-2-(5-oxo-pentanoyl)-3-glycero-PC (POVPC).Later this compound and other structurally similar ones were found inminimally oxidized low density lipoproteins (LDL) and atheroscleroticplaques [11–13], leading to an association of oxPL with the processof vascular dysfunction and atherosclerosis, thus prompting expan-sion of this new field of research. Full understanding of the role ofphospholipids and their oxidized counterparts is only recentlyemerging with the development of user-friendly and robust massspectrometers that enable the complete mapping of phospholipidspecies present in biological samples, and are attractive to a widerrange of researchers such as biologists, biochemists, pharmaceuti-cals, and chemists.
This article will describe the paradigm of non-enzymatic phospho-lipid oxidation, and identify some aspects of the mechanisms that re-main controversial or poorly understood. The wide range ofphospholipid oxidation products formed and their dependence onthe oxidizing system will be discussed, as well as the model systemscommonly used in such studies. The challenges of measuring andquantifying oxidized phospholipids in vivo will be discussed, as wellas the limitations of existing methodology. The aim of this article isto provide an introduction to other articles appearing in this issuethat will focus more deeply on individual aspects of the interactionsof oxidized phospholipids with proteins.
1.2. Types and sources of oxidants relevant to phospholipid oxidation
The oxidative modifications to phospholipids depend greatly onthe nature of the oxidant species, as this determines the mechanismof the oxidative reactions. Radical species such as hydroxyl radicals(•OH) and, to a lesser extent, superoxide radicals (O2
−•) tend to beregarded as more reactive and poorly selective, whereas many non-radical species (e.g. hydrogen peroxide and HOCl) show more selec-tive oxidation reactions with biomolecules and are able to diffusefrom their point of origin [14,15]. However, some non-radicaloxygen-containing species are highly oxidizing, such as ozone (O3),singlet oxygen (1O2) and peroxynitrite (ONOO−), while nitric oxide(•NO) can act both as a radical electron donor or oxidant [16–18].The oxidants in aerobic systems can come from endogenous process-es such as respiration, metabolism and phagocytosis [19], or from ex-ogenous sources by inhalation of air pollutants and exposure toradiation. In fact, many oxidants relevant to phospholipid oxidationmay derive from both exogenous and endogenous sources, such asnitric oxide (generated by nitric oxide synthases but also found in ex-haust fumes), transition metal ions, and ozone, which has been sug-gested to be produced by neutrophils under certain conditions [18].Superoxide radicals are also produced at low levels by NADPH oxi-dases (NOX) and dual oxidases (DUOX) as part of cell signalling pro-cesses [20].
Table 1 shows some of the reactions that form the radical speciescapable of causing oxidation of phospholipids. Although superoxideradicals are produced by a variety of endogenous reactions, they arenot particularly effective at causing hydrogen abstraction, and
Table 1Reactions for the formation of radical species from different oxidants present in aerobicsystems.
Oxidant systems Reactions
Fenton reaction (•OH) Fe (II)+H2O2→Fe (III)+HO•+HO−
Fe2++O2→ [Fe2+―O2↔Fe3+―O2•]→Fe3++O2•
Fe2++Fe2+―O2→Fe2+―O2―Fe2+→2 Fe2+―ORadiation H2O→H2O*→H++ •OHPollution NO2+hv→NO+OSOD (O2
−) O2−•+O2
−•+2H+→H2O2+O2
MPO H2O2+X−+H+→HOX+H2O−OCl+H2O2→
1O2+Cl−+H2ONO synthases •NO+O2
−•→ONOO−
ONOO−+H+→ONOOHONOOH→ •NO2+ •OH
2375A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
moreover, owing to the negative charge, are not very lipophilic. Otherspecies, notably hydroxyl radicals, are much more effective; theyreact rapidly with various biological molecules, and have correspond-ingly a low degree of selectivity in their reactions. Hydroxyl radicalsare formed mainly by one-electron redox reactions between H2O2
and pre-existing hydroperoxides with transition metal ions, althoughradiation induced homolysis of water is another mechanism. Reac-tions of ferrous (Fe2+) iron with hydrogen peroxide and oxygencan generate ferryl (FeO2+) and perferryl species (FeO2
+), which arestrong oxidants and have also been suggested to be capable of initiatingradical reactions [21]. Ozone, although not a radical, also leads to lipidperoxidation through an addition reaction to yield an unstable speciesthat subsequently breaks down via homolytic cleavage reactions. Hypo-halous acids (HOX, X−=Cl−, Br−, I−) and isothiocyanite (HOSCN) canbe produced by the enzyme myeloperoxidase, which is present at highlevels in neutrophils and at lesser levels in other polymorphonuclearleukocytes. Eosinophil peroxidase is a similar enzymebutwith differentspecificity [22]. The hypohalous acids are capable of 2-electron oxida-tion of phospholipids, resulting in halogenated species such as chloro-hydrins and α-chlorofatty aldehydes. Myeloperoxidase also catalysesreactions to form the radical species nitrogen dioxide (•NO2) from hy-drogen peroxide and nitrite [23]. Nitric oxide (•NO) is a relatively stableand diffusible radical, but reacts rapidly with superoxide to form perox-ynitrite (ONOO−). Peroxynitrite and its protonated form ONOOH arenot radical species, but can undergo a variety of one- and two-electron reactions in biological environments, resulting in both oxidiz-ing and nitrating reactions [17].
Thus the local concentration and nature of the oxidizing species,as well as the presence of antioxidants, is important. For instance,the low levels of [H2O2] in most cells (10−8–10−10 M) may limitthe potential for formation of hydroxyl radicals through the Fentonreaction, and hence their role in mediating lipid peroxidation reac-tions in phospholipids [21,24]. Instead, the higher ratio of [O2]/[H2O2] found in cells suggests that oxidant systems, such as iron com-plexes (ferryl/perferryl ions), may have more significant roles in theinitiation of radical oxidation of unsaturated fatty acid moietieswhen compared to hydroxyl radical [21]. On the other hand, in acidichypoxic environments the increased solubility of ferrous ions may ac-count for the phospholipid modification by the Fenton reaction withpre-existing hydroperoxides (LOOH), formed by enzymatic systemssuch as lipoxygenases (LOX) and cyclooxygenases (COX), althoughat physiological pH most iron ions are in complexed form [25,26].
2. Lipid peroxidation and other types of phospholipid oxidation
2.1. Basic concepts in peroxidation of polyunsaturated fatty acyl chains
Unsaturated fatty acids esterified to phospholipids are major tar-gets for modification under conditions of oxidative stress. In unsatu-rated acyl chains, the hydrogen atoms on methylene groupsimmediately adjacent to double bonds (allylic hydrogen atoms) ex-hibit low carbon–hydrogen (C―H) bond energies, and those locatedon methylenes between two double bonds (bis-allylic hydrogenatoms) have even lower C―H bond energies, which allows them tobe readily abstracted by reactive radical species, leading to the forma-tion of phospholipid radical species with the radical centred on the al-lylic the carbon atom (Fig. 1). Comparatively, saturated fatty acidchains such as palmitoyl and stearoyl are more resistant to oxidativemodification as they do not have allylic hydrogen atoms [25]. Oncephospholipid carbon centred radicals (L•) are formed, they readilyreact with molecular oxygen to form peroxyl radicals (LOO•). The ini-tial hydrogen abstraction followed by addition of O2 corresponds tothe initiation phase of lipid peroxidation. In polyunsaturated fattyacyl chains the addition of O2 molecules may occur several times ona single acyl chain because of the presence of other (bis)allylic
hydrogen atoms or following rearrangement of a peroxyl radical toa new carbon-centred radical.
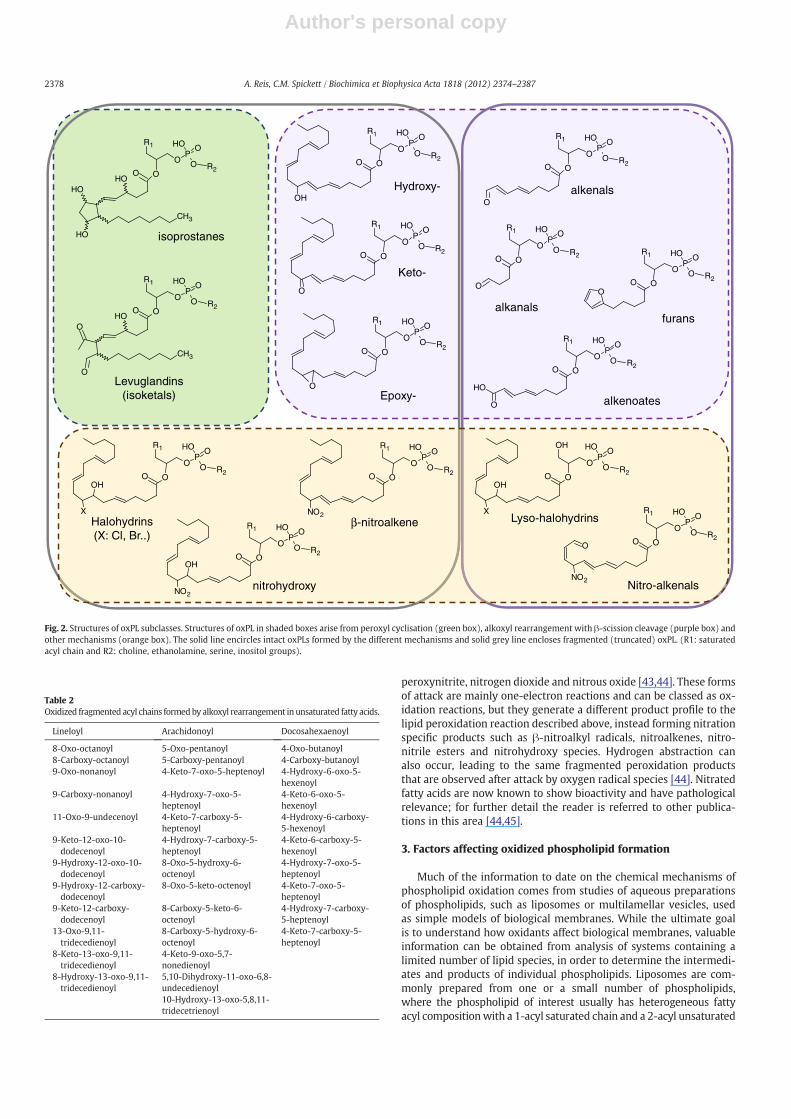
Subsequent steps in the intermediate stage of the process lead tothe formation of further carbon- or oxygen-centred radicals. These in-termediate peroxidation products, which include L• (alkyl), OL•(epoxy alkyl), HOL• (hydroxy-alkyl), LO• (alkoxyl) and OLOO•(epoxy-peroxyl) species, are responsible for perpetuating the radicaldamage, as they can stabilise themselves by abstracting hydrogenatoms from neighbouring molecules, thus leading to the formationof new carbon-centred radicals on adjacent fatty acyl chains. This rep-resents the propagation phase of lipid peroxidation, whereby a singleinitial free radical attack generates damage to multiple lipid mole-cules. Abstraction of a hydrogen by the peroxyl (or other) radicalgroup on the initially attacked lipid converts it to a non-radical spe-cies, such as a hydroperoxide. Fig. 2 shows the major subgroups ofoxPL that are generated by radical attack mechanisms.
The final step in lipid peroxidation is termination, which can beachieved in 2 ways. Under conditions where the concentration of ini-tiating radicals is sufficiently high to support radical–radical reactionsresult in the formation of non-radical phospholipids products [27]. Al-ternatively, termination may occur by the intervention of lipophilicchain-breaking antioxidants such as β-carotene or α-tocopherol,which have relatively stable conjugated radical forms [28].
The basic principles of the process of phospholipid peroxidation arequite well established, and apply to phospholipid peroxidation both invitro and in vivo. Nevertheless, questions over somemechanistic detailsremain, and while many fragmentation products of different nativephospholipids have been identified, it is quite possible that others areyet to be discovered. The next sections discuss in more depth some as-pects of the chemistry of phospholipid peroxidation. Much of thisknowledge has come from studies in carefully defined synthetic sys-tems such as liposomes, which limit the complexity of the productsand facilitate understanding of the mechanisms.
2.2. The diversity of oxidized phospholipid products from peroxidation
From the previous section it is clear that a diverse range of differentproducts of phospholipid peroxidation may arise from radical-basedperoxidation. For any individual phospholipid species, the profile ofoxPL generated is influenced by the oxidant species, the type of linkageof the FA to the glycerol backbone, and the fatty acyl chain present. Theposition of the double bonds in an unsaturated FAwill in turn influencethe site where the oxidant species attacks. However, while in acylchains with only one bis-allylic hydrogen atom (e.g. linoleoyl) identifi-cation of the products and therefore the mechanism is a fairly simpletask, it becomes far more complicated in acyl chains with more bis-allylic atoms (arachidonoyl, eicosapentanoyl and docosahexaenoyl),owing to the number of possible sites of attack and secondary radicalreactions.
The diverse oxPL species that are formed from the initial carbonand oxygen centred radical PL species can be divided into several sub-classes, depending on themechanism bywhich they become stabilisedand therefore the structures of these final products. Stabilisation of theperoxyl radical by inter- or intra-molecular hydrogen abstraction re-sults in formation of hydroperoxide derivatives, which may subse-quently rearrange or be reduced to hydroxy-, keto- or epoxy-containing species. Alternatively the peroxyl radical may carry out anintramolecular attack leading to endoperoxide formation and subse-quent cyclisation reactions to form compounds with substituted cyclo-pentane or cyclopentene rings (isoprostanes and neuroprostanes). Gaoand colleagues have shown the formation of prostaglandins D2 and E2via an isoprostane-like pathway that appeared to be cyclooxygenase-independent and probably involved an epimerisation mechanism [29].Isolevuglandins (also known as isoketals) are another subclass of com-pounds formed following cyclisation of peroxyl radicals by cleavage ofthe 5-membered ring [30]. All of these latter groups of compounds
2376 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
retain the same number of carbons as the original fatty acyl chain, al-though not in a linear sequence, and therefore class as non-fragmentedoxPL.
In addition to these full chain length oxPLs, rearrangement ofalkoxyl radicals with carbon–carbon cleavages in turn generates a va-riety of oxPL that contain a shortened fatty acyl chain. Depending onthe terminal group present, these can be grouped in various sub-classes of oxPL, namely alkanals (saturated aldehydes), alkenals (un-saturated aldehydes), hydroxy-alka(e)nals, alkanoates (saturatedcarboxylic acids), and hydroxy-alkanoates [31]. Extreme chain-shortened oxidized products can also be formed: for example, attackby ozone or radicals on plasmenyl phospholipids results in formationof 1-formyl-2-acyl-PL [32]. In addition, of course, different carbonchain length alkanes, alkenes, aldehydes, carboxylic acids and hy-droxides are released by this oxidation process, depending on thenative acyl chain and site of attack. The phospholipid fragmentationproducts (secondary peroxidation products) formed with various oxi-dant systems have been well characterized in linoleoyl [33,34], arachi-donoyl [11,33,35–38] and docosahexaenoyl-containing phospholipids[39]. Over time the relative abundance of fragmentation products
increases, usually accompanied by the decrease in longer carbon chain(intact) oxPL, and the structural diversity of the oxPL subclasses is grad-ually decreased. It is important to note that some individual productscan result from several different unsaturated acyl chains, while othersare unique to a particular native fatty acyl chain (Table 2), thus actingas a fingerprint for the type of oxidation.
2.3. Other pathways of phospholipid oxidation
Unsaturated phospholipids are also susceptible to 2 electron, elec-trophilic attack by certain non-radical oxidants, which leads to someunique products (Fig. 2). For example, it is well-established thatmyeloperoxidase-derived hypohalous acids oxidize fatty acyl chains atthe alkenyl double bonds, yielding halohydrins (α-halogenated β-hydroxylated compounds), and more recently HOCl has been shownto attack vinyl ether bonds in plasmalogens, resulting in degradationof the phospholipid to α-chlorofatty aldehydes and lysolipids, whichcan subsequently be modified further to chlorohydrins [40–42].
Nitrated lipids can be formed by multiple pathways following at-tack of reactive nitrogen species derived from nitric oxide, including
ROS
, H2O2)
Conjugated dienes
OLOO
LO
LOO
LH
Alkanes, Alkenes
Aldehydes
OL
GSH
O2
L
LOH
γ-hydroxy-α,β-aldehydes
LOOH
LH
L
Fe3+ Fe2+
O2
LOXCOX
GSSG
LH
LH
L
OLOOH
Dicarboxylic acids
PLGPx
LOH
'R R
O
MPO
HOX
(X:Cl,Br)
L(HOX)H
RNS
( NO, NO2)L(NO2)H
(HO
Fig. 1. Schematic diagramof radical peroxidation of fatty acid chain in phospholipids. The shaded lilac box depicts the initiation stage, involving attack by a free radical andhydrogen abstractionfrom the fatty acyl chain. The shaded grey box illustrates the various reactions that contribute to the propagation phase of radical reaction. The shaded green box corresponds to secondary re-actions (either fragmentation or enzymation reduction) that can contribute to the termination phase. The solid green line (bottom left) delineates the formations of fragmentation products byβ-scission, whereas the solid grey line indicates full length phospholipids.
2377A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387
Author's personal copy
peroxynitrite, nitrogen dioxide and nitrous oxide [43,44]. These formsof attack are mainly one-electron reactions and can be classed as ox-idation reactions, but they generate a different product profile to thelipid peroxidation reaction described above, instead forming nitrationspecific products such as β-nitroalkyl radicals, nitroalkenes, nitro-nitrile esters and nitrohydroxy species. Hydrogen abstraction canalso occur, leading to the same fragmented peroxidation productsthat are observed after attack by oxygen radical species [44]. Nitratedfatty acids are now known to show bioactivity and have pathologicalrelevance; for further detail the reader is referred to other publica-tions in this area [44,45].
3. Factors affecting oxidized phospholipid formation
Much of the information to date on the chemical mechanisms ofphospholipid oxidation comes from studies of aqueous preparationsof phospholipids, such as liposomes or multilamellar vesicles, usedas simple models of biological membranes. While the ultimate goalis to understand how oxidants affect biological membranes, valuableinformation can be obtained from analysis of systems containing alimited number of lipid species, in order to determine the intermedi-ates and products of individual phospholipids. Liposomes are com-monly prepared from one or a small number of phospholipids,where the phospholipid of interest usually has heterogeneous fattyacyl compositionwith a 1-acyl saturated chain and a 2-acyl unsaturated
O R2
POOH
O
R1
OO
CH3
OH
OH
OH
isoprostanes
O R2
POOH
O
R1
OO
CH3
O
O
OH
Levuglandins (isoketals)
O R2
POOH
O
R1
OO
OH
O R2
POOH
O
R1
OO
O
O R2
POOH
O
R1
OO
O
Hydroxy-
Keto-
Epoxy-
O R2
POOH
O
R1
OO
Oalkenals
O R2
POOH
O
R1
OO
O
OH
alkenoates
O R2
POOH
O
R1
OO
O
alkanals
O R2
POOH
O
R1
OOO
furans
Halohydrins (X: Cl, Br..)
O R2
POOH
O
R1
OOOH
X
O R2
POOH
O
R1
OO
NO2β-nitroalkene
nitrohydroxy
O R2
POOH
O
R1
OOOH
NO2
O R2
POOH
O
R1
OO
NO2
O
Nitro-alkenals
O R2
POOH
O
OH
OOOH
XLyso-halohydrins
Fig. 2. Structures of oxPL subclasses. Structures of oxPL in shaded boxes arise from peroxyl cyclisation (green box), alkoxyl rearrangement with β-scission cleavage (purple box) andother mechanisms (orange box). The solid line encircles intact oxPLs formed by the different mechanisms and solid grey line encloses fragmented (truncated) oxPL. (R1: saturatedacyl chain and R2: choline, ethanolamine, serine, inositol groups).
Table 2Oxidized fragmented acyl chains formedby alkoxyl rearrangement in unsaturated fatty acids.
Lineloyl Arachidonoyl Docosahexaenoyl
8-Oxo-octanoyl 5-Oxo-pentanoyl 4-Oxo-butanoyl8-Carboxy-octanoyl 5-Carboxy-pentanoyl 4-Carboxy-butanoyl9-Oxo-nonanoyl 4-Keto-7-oxo-5-heptenoyl 4-Hydroxy-6-oxo-5-
hexenoyl9-Carboxy-nonanoyl 4-Hydroxy-7-oxo-5-
heptenoyl4-Keto-6-oxo-5-hexenoyl
11-Oxo-9-undecenoyl 4-Keto-7-carboxy-5-heptenoyl
4-Hydroxy-6-carboxy-5-hexenoyl
9-Keto-12-oxo-10-dodecenoyl
4-Hydroxy-7-carboxy-5-heptenoyl
4-Keto-6-carboxy-5-hexenoyl
9-Hydroxy-12-oxo-10-dodecenoyl
8-Oxo-5-hydroxy-6-octenoyl
4-Hydroxy-7-oxo-5-heptenoyl
9-Hydroxy-12-carboxy-dodecenoyl
8-Oxo-5-keto-octenoyl 4-Keto-7-oxo-5-heptenoyl
9-Keto-12-carboxy-dodecenoyl
8-Carboxy-5-keto-6-octenoyl
4-Hydroxy-7-carboxy-5-heptenoyl
13-Oxo-9,11-tridecedienoyl
8-Carboxy-5-hydroxy-6-octenoyl
4-Keto-7-carboxy-5-heptenoyl
8-Keto-13-oxo-9,11-tridecedienoyl
4-Keto-9-oxo-5,7-nonedienoyl
8-Hydroxy-13-oxo-9,11-tridecedienoyl
5,10-Dihydroxy-11-oxo-6,8-undecedienoyl10-Hydroxy-13-oxo-5,8,11-tridecetrienoyl
2378 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
chain. This simplifies the interpretation of the analysis. The majority ofstudies conducted on the formation, identification and quantificationof oxidized phospholipids have been focused on the choline class ofphospholipids, as they comprise nearly 50% of total PL in biologicalmembranes [46], and are thus important molecules. Being more abun-dant, they are alsomore readily available commercially in pure form. Fi-nally, they are the phospholipids most easily detected by positive ionelectrospray mass spectrometry, a frequently used analytical techniquefor studying their oxidation.
3.1. Influence of liposome size and composition
It is important to bear in mind that the type of liposome prepara-tion used may influence the oxidation process. Firstly, multilamellarvesicles (MLVs; formed during sonication) can be expected to havedifferent kinetics to unilamellar vesicles, or liposomes (formed by ex-trusion of MLVs through polycarbonate filters), in view of the differ-ent accessibility of the phospholipid layers to aqueous oxidants.Secondly, a small number of studies have reported differences in ox-idative susceptibility of liposomes of the same phospholipid composi-tion but different sizes, which can be generated using different poresizes and extrusion pressures. Small unilamellar vesicles (SUVs) pre-pared from phosphatidylcholines were found to undergo more oxida-tion than large unilamellar vesicles (LUVs), assessed by measurementof conjugated dienes, trienes, and thiobarbituric acid reactive sub-stances [47]. Similar results were obtained in another study that com-pared small sonicated liposomes and large non-sonicated liposomesprepared from retinal lipids and oxidized with Fe2+ and Fe3+ as radicalinitiators, and showed longer lag times in the larger liposomes [48]. Inboth cases, it was hypothesized that the greater oxidation of SUVs wasdue to higher surface curvature and more openly packed headgroupscompared to LUVs, which enabled greater penetration of water solubleradicals and consequently oxidation. The occurrence of asymmetric oxi-dation of phospholipids in the inner and outer leaflets of giant unilamel-lar vesicles has also been shown [49]. The situation is further complicatedin vivo by the fact that different fatty acid chains esterified to the glycerolcan also influence the acyl packing and bilayer curvature. These resultsmay explain the variability in phospholipid oxidation observed in vitrobetween different studies with apparently comparable conditions, as li-posome size is a parameter that is commonly overlooked. Thus it is im-portant to normalize liposome sizes for truly comparable results [50].
3.2. Influence of the oxidant system
There are many reports in the literature of phospholipid peroxida-tion, carried out on a wide variety of phospholipid starting materialand with an equally broad range of oxidizing systems. The most fre-quently studied methods of causing chemical radical reactions aremetal-catalysed reactions (generating •OH), autoxidation, radiation(generating •OH and 1O2) and ozone (O3); popular enzymatic radicalgenerating systems include lipoxygenases and myeloperoxidase. Thephospholipids that have been mostly studied in liposome models arethose containing an arachidonoyl chain, as these are readily oxidizable.Table 3 compares the oxidized products identified for arachidonoyl-based PC using different chemical and enzymatic oxidants in studiesof unilamellar model systems at 37 °C (liquid crystalline phase). Theconcentrations of oxidants and the lengths of the oxidation reactionare indicated, together with the analytical techniques employed, asthese have a direct influence on the oxidation products observed.
It can be seen that the oxidized products identified depend on theoxidant system, concentration and the length of the incubation used.As a generalization, modification by chemical oxidants leads to the for-mation of a wider variety of oxidized products compared to those de-scribed for enzymatic systems, which tend to yield specific products.However, some common features can also be found for the chemicalradical generating systems, and much of the reported variability relates
to modifications around C-5 position of arachidonoyl chain. Lysolipidspecies with either the saturated or unsaturated acyl chain remaininghave also been reported [40,51], both following free radical attack andoxidation with hypohalous acids (e.g. HOCl and HOBr) [52], althoughit is more common for the oxidized chain to be lost and the saturatedacyl chain to remain esterified to the headgroup. This involves an intra-molecular reaction initiated by reactive moieties in the oxidized chain,and a mechanism for several different full chain oxidation productshas recently been reported [53], demonstrating that the oxidizedchain is lost by spontaneous formation of a lactone. The oxidant alsomakes a difference to the products of plasmenyl phospholipids: radicaloxidations and attack by ozone result in the formation of 1-formyl-2-acyl-PL with cleavage of an n-1 alkenal moiety [32,54]. In contrast, at-tack at the vinyl ether by myeloperoxidase-generated oxidants leadsto formation of a lysolipid plus a 2-halofatty aldehyde [41]. These cleav-age reactions reflect the fact that the vinyl ether is more susceptible tochemical attack than the ester bond [55], and also that beyond thevinyl ether bond the chains are commonly saturated, while polysatu-rated chains are usually linked by ester bonds to the glycerol backbone,so the site of likely attack is different.
3.3. Selectivity of modification dependent on fatty acyl chain
As seen in the previous sections, oxidative damage to phospho-lipids results in a wide variety of different products, depending onthe initiating agent responsible and the phospholipid under attack.The free radical or oxidative modifications carried out directly by en-zymes as a general rule produce specific modifications, at least at theprimary product stage, with a moiety such as a hydroperoxide or hy-droxide introduced at a specific location in the fatty acyl chain. A goodexample of this is offered by cyclooxygenases (COX) and the lipoxy-genase (LOX) family: several lipoxygenases from different biologicalsources catalyse the formation of positional isomers of hydroperox-yeicosatetraenoic acid, e.g. 5-LOX and 12/15-LOX [56–58]. On theother hand, non-enzymatic oxidative damage by free radicals is usu-ally considered to lack the specificity of enzymes, as attack by freeradicals can theoretically take place at any of the carbon atoms con-taining bis-allylic hydrogen atoms to yield a larger variety of position-al isomers. Surprisingly, accumulated experimental observations bydifferent research groups have shown that arachidonoyl-containingphospholipids tend to form oxidation products with low carbonchain lengths with the predominance of modifications at either C-5or C-8 (Table 3). Modifications resulting from attack at the C-10 andC-13 bis-allylic hydrogen atoms are less common. Currently, it is notclear whether this is because hydrogen atoms closer to the polarhead are more likely to be abstracted, or that longer chain hydroper-oxide derivatives are less stable and fragment preferentially to theseproducts. Possible mechanisms and evidence for the former possibil-ity were discussed in Section 3.1 on the effect of liposome size; evi-dence for the second suggestion is discussed in Section 4. In view ofthe evidence that bilayer curvature affects the accessibility of thefatty acyl chains to oxidants, it would be interesting to know whetherthe location of the attack is also dependent on liposome size.
Many of the products of arachidonoyl-containing PC species withdifferent oxidants have also been observed for the arachidonoyl-containing phosphatidylethanolamines [38] and phosphatidylserines[59]. This suggests that differences in the hydration layer at thepolar group head between these species do not substantially influ-ence the accessibility of the fatty acyl chains to oxidants, and there-fore have little effect on the oxidized products formed.
4. Recent advances in understanding of phospholipid oxidationmechanisms
Despite the extensive knowledge already achieved on the reac-tions leading to formation of peroxyl, alkane, alkene and aldehyde
2379A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
derivatives, this is still an area of strong research activity with manyoutstanding aspects of the lipid peroxidation process remaining tounravelled. For instance, fatty acid chains in vivo exhibit a cis geome-try, though this geometry is lost after attack by radicals [60], but so farrelatively little attention has been devoted to the presence of transfatty acids in biological membranes and understanding of their bio-logical relevance remains scarce [61].
Another example relates to the mechanism underlying the forma-tion of HNE and its derivatives from ω-6 fatty acids, which has beenextensively studied. The most accepted mechanism involves the 4-hydroperoxy-nonenal intermediate arising from both linoleic acid(13-HPODE) and arachidonic acid (15-HPODE) by alkoxyl rearrange-ment with bond cleavage [62,63], although an alternative mechanisminvolving cross-chain peroxyl addition followed by decompositionhas been suggested for lineloyl chains from cardiolipin phospholipids[64]. However, the question about what factors govern the routes ofalkoxyl rearrangement and carbon–carbon chain cleavage to form un-saturated aldehydes (Fig. 3) still remains unsolved in the literature.Taking into consideration the literature on the formation of HNEthrough the decomposition of purified hydroperoxide derivatives[62,63], it seems that the most likely route for decomposition of inter-mediate oxidation products (alkoxyl species) is via the formation ofaldehydes of low carbon chain length [50].
Although phospholipids can be assembled with various polyunsatu-rated fatty acyl chains, some ofwhich contain several bis-allylic hydrogenatoms that are all theoretically equally likely to be abstracted duringradical initiation step, experimental data over the years has shown apredominance of secondary products with C9 carbon chains (oxo-nonanoyl) for lineloyl, C5 (oxo-valeroyl) for arachidonoyl, and C4
(oxo-butyroyl) for docosahexaenoyl chains. This is almost a finger-print of the unsaturated fatty acyl chains that have undergone
oxidation. Whether the consistence observed is the result of greaterstability of these saturated secondary products or preferential attackon the bis-allylic hydrogen atoms closer to the polar head is not yetfully understood. Arguments in favour of the first hypothesis havebeen proposed by Balamraju et al.; they suggested that the γ-hydroxy-α,β-unsaturated aldehydes formed during radical oxida-tion degrade via a Michael addition of a peracyl radical with forma-tion of a β-hydroxyperester, with concomitant loss of a 3 carbonchain resulting in the formation with accumulation of saturated al-dehydes (Fig. 4) [65]. Also, γ-hydroxy-alkenals can be involved inan alternative mechanism with cyclisation to a hemiacetal interme-diate followed by dehydration and conversion to furan [66].
This hypothesis is supported by the observation of longer carbonchain secondary products in the initial stages of peroxidation andtheir decrease over time and concomitant increase of shorter carbonchain length compounds. Further corroboration comes from spintrapping experiments conducted on the identification of phospholip-id and lipid radical species (intermediate peroxidation species),where linoleoyl- and arachidonoyl-containing phospholipid radicalspecies were located at C-11, C-13 and C-14 positions [67–69]. Theidentification of unstable and highly reactive radical species is possi-ble through the stabilisation of the unpaired electron by reactingwith a paramagnetic spin trap. The spin trap stabilises the radical(in the form of radical adducts) enabling their accumulation in solu-tion to detectable levels. Detection is usually conducted by electronparamagnetic resonance (EPR) or mass spectrometry (MS). Stabilisa-tion of radicals by the spin trap also enables the radical to be tagged,allowing information about the exact nature and location of the rad-ical within the unsaturated acyl chain to be obtained. Spin trappingexperiments conducted on oxidized linoleic acid by the Fenton reac-tion showed a higher relative abundance of smaller carbon chain
Table 3Dependence of phospholipid oxidation products on the oxidant system employed.
Oxidant system Oxidation products identified Detection Reference
Chemicaloxidants
Cu (II)/H2O2, 3 h Hydroxy (C-5); di-hydroxy (C-5, C-12) QQQ [54]Fe(II)/H2O2 POVPC, PGPC, 8-oxo-5-hydroxy-octenoylPC,
4-keto-nonadienoylPC, 13-oxo-10-hydroxy-tridecenoylPC,palmitoyl-(hydroxy-A)PC, palmitoyl-(hydroperoxide-A)-PC,PEIPC, palmitoyl-lysoPC, arachidonoyl-lysoPC
Reverse-phase LC-MS (Q-TOF) [37]
Autoxidation (48 h) POVPC, PGPC, PEIPC, ions up to addition of 9oxygen atoms are visible in the MS spectrum
QQQ [11]
Autoxidation (24 h) +3 oxygen atoms (m/z 830), + 4 oxygen atoms (m/z 846) Sector instrument (FAB-MS) [107]AAPH (2 mM), 3 h 5-Keto, 8,9-epoxy, 5,6-epoxy, 11,12-epoxy, 5,6-di-hydroxy QQQ [54]AAPH (10 mM) 5-HETE, 8-HETE, 9-HETE, 11-HETE, 5,6-ETE, 8,9-ETE, 5,
12-di-hydroxy, 1 formyl-APC, POVPC, lyso-PCQQQ [35]
AAPH 5-OH, 15-OH (where 5-OH>>15-OH) [108]AAPH (80 mM) 1-Acyl-2-(5-oxo-pentanoyl)-PC HPLC-fluorescence (DBD-H derivatives) [106]t-BHP (140 mM) Mono-OOH, di-OOH VG platform [109]HOCl/OCl (13 mM), 20 min–1 h Monochlorohydrins, bis-chlorohydrins,
stearoyl-lyso-PC, SOVPC, SGPCMALDI [42]
UVA, 80 Jcm−2 Mono-OOH (m/z 814), bis-OOH (m/z 846),tri-OOH (m/z 878), PEIPC (m/z 828),NO lyso-PC, NO fragmented-PC
QQQ [110]
UVB, 400 Jm2 1-Hexadecyl-2-(4-butanoyl)-PC QTrap [111]Enzymaticoxidants
MPO-H2O2―Cl Mono-chlorohydrins, di-chlorohydrins,tri-chlorohydrins, tetra-chlorohydrins,Lyso-PC (saturated), no fragmented PC
MALDI [112]
MPO Mono-chlorohydrins, di-chlorohydrins,tri-chlorohydrins, tetra-chlorohydrins,Lyso-PC (saturated), no fragmented PC
MALDI [52]
MPO-H2O2―NO2 8-Carboxy-5-OH-octenoylPC (HOdiA-PC),8-carboxy-5-keto-octenoylPC (KOdiA-PC),8-oxo-5-OH-octenoyl-PC (HOOA-PC),8-oxo-5-keto-octenoylPC (KOOA-PC)
QQQ [33]
LOX (reticulocyte, rabbit) 15(S)-OOH-arachidonoyl-PC HPLC-UV-MS [113]LOX (reticulocyte, rabbit) 15-OOH-arachidonoylPC GC–MS [56]LOX (reticulocyte, rabbit) 15(S)-OOH-arachidonoyl HPLC-UV [56]LOX (porcine) 12(S)-OOH-arachidonoyl-PC HPLC-UV-MS [113]LOX (reticulocyte, rabbit) 15-OOH-arachidonoylPC HPLC-CEC [57]
2380 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
alkoxyl (LO•) radicals over longer carbon chain alkoxyl radicals [70].Similarly, the identification of smaller alkyl (L•) DMPO radical adductsand the absence of longer alkyl DMPO adducts suggests that β-scission occurs with release of smaller carbon chain alkyl radicals[70]. The formation of smaller alkoxyl radicals at positions distantfrom the polar head with concomitant formation of longer carbonchain aldehydes and small alkyl radicals is consistent with random at-tack by free radicals at the various bis-allylic positions [50]. The longercarbon chain aldehydes formed may undergo further decompositionto more stable saturated aldehydes, in agreement with the findings de-scribed previously [65]. On the other hand, support for the concept thatattack closer to the headgroup is favoured comes from the observationthat smaller liposomes with higher curvature are more susceptible tooxidation [47,48], as described in Section 3.1.Whatever themechanismresponsible for this effect, this “specificity” is well-known, and almostresembles that found during enzymatic oxidative attack [56–58],which may also account for the fact that secondary oxPL such as alka-nals and alkanoates are predominantly detected in biological samples[12,13,38].
5. Analysis and quantification of oxidized phospholipids
The analysis of the complete family of phospholipid oxidation prod-ucts is challenging because of the diversity and extent of the products,as described above, and no singlemethod exists that enables quantifica-tion of all known oxPL products from the various subclasses. Somewellestablished methods are simple to perform, but may lack specificity orbe limited in the products observed [71]. Reliable and accuratemethodshave been developed for the quantification of primary PL products (hy-droperoxides) by chemiluminescence methods that allow the quantifi-cation of PLOOH up to pmol level [72–74] and for fragmented oxPL
(containing terminal carbonyl groups) by use of dinitrophenylhydra-zine (DNPH) post-column derivatisation followed by detection atλ=340–380 nm (UV range) enabling quantification up to μM [75].While the non-esterified breakdown products of oxidized phospho-lipids are relatively volatile and can be detected readily using GC–MS,this method cannot be applied directly to oxidized phospholipids [71].Instead, the most widely used method is electrospray ionisation MS,as this is not dependent on analyte volatility.
5.1. Mass spectrometry in the study of oxPL
Currently, mass spectrometry coupled with liquid-chromatographyis the best choice for the detection of a wide range of oxPL; it can offerqualitative and quantitative information about both primary andsecondary phospholipid oxidation products and lyso-PL withoutthe need for derivatisation procedures, while showing good linearityand reproducibility across a wide range of concentrations. The levelsof oxPL reported in literature in model membranes vary widely asthey depend on the extent of oxidation reaction at the time pointat which they were acquired. Moreover, there are a number of im-portant factors that affect the observation of analytes and complicateits application in biological samples.
One potentially problematic aspect of mass spectrometry as a de-tection technique for oxPL is that it relies largely on the ionisation ef-ficiency of molecules in the gas-phase; differences in this efficiencyespecially between PL classes can lead to ion suppression effects,resulting in spectrum with poor signals or even absence of PL classeswith lower ionisation efficiencies. Specifically, the ionisation processof phosphatidylcholines (PC) during mass spectrometry analysis is fa-cilitated by the fact that they are naturally positively charged mole-cules (trimethylamine group located at the polar head), and this
O
OH
O
OH
O
OH
CH
O
OH
O
a)
O
OH
O
CH2
O
OHO
O
OH
•
••
H abstraction
H abstraction
O2
H abstraction
Homolytic scission
b)
a) b)
+ +
O
OH
CH2
Fig. 3. Scheme of possible routes for homolytic β-scission of alkoxyl radical. Products formed via route (a) are hexanal and alkenyl radicals, while products formed via route (b) arepentyl radicals and 9,11-tridecedienoic acid.
2381A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
causes ion suppression of other phospholipid species [76]. Ion sup-pression can also result from even relatively low concentrations ofsalts in the samples. These factors are exacerbated when analysing bi-ological samples by shotgun lipidomics approaches, but can be over-come by interfacing with LC to remove salts and abundantunmodified species. Through this approach, identification of oxPCcan be achieved based on the m/z value, the retention time and thefragmentation pattern, narrowing the likelihood of misinterpretationof isobaric oxPC species present in complex mixtures. The carbonchain length, size of the analyte, and degree of unsaturation furtherinfluence the detection of oxPL [77]; the preferential ionisation oflower molecular weight oxPC results in mass discrimination effects,thus presenting a challenge in handling the diversity of oxPC prod-ucts. Moreover, differences in the chemical stability of relatively lessstable (e.g. carbonyl) and thermally labile chemical groups (e.g. hy-droperoxides) may lead to apparent differences in the relative abun-dances observed, which is crucial for accurate quantification. Finally,quantification depends on having internal standards, preferably iso-topically labelled versions of the analyte to give specific mass shifts,but the phospholipids available as internal standards are limitedand only cover a small part of the necessary mass range.
Despite the limitations described, the ability of mass spectrometryto detect a wide range of compounds with high sensitivity has madethis technique popular for use in both model and biological samples,and offers the benefit of obtaining information at a molecular level.The limit of quantification of oxPCs by mass spectrometry is similarto that reported for alternative chemiluminescence methods (pmol;μM range) [78], but can be lowered to femtomole levels through theuse of complimentary enrichment methodologies, with the additionaladvantage of obtaining structural information by tandem massspectrometry.
5.2. 31P NMR in the study of oxPL
In many ways, nuclear magnetic resonance spectroscopy (NMR) iscomplementary to mass spectrometry in terms of its advantages andlimitations. It is intrinsically a quantitative technique, whereby theintensity of the signal in the spectrum correlates linearly to theamount present in the sample, assuming appropriate pulse sequencesand settings for parameters such as relaxation time are employed. It isless affected by salt concentrations (so long as paramagnetic metalions are removed). Phospholipids can be conveniently detectedusing 1H-, 13C- or 31P-NMR, of which 1H is most sensitive and 13Cleast sensitive, in part owing to a natural abundance of only 1%. How-ever, 13C-NMR is usually carried out with C―H uncoupling to im-prove sensitivity and simplify the spectra, and under theseconditions the quantitative relationship is lost. 31P-NMR has an ad-vantage for analysis of phospholipids, as these are detected selective-ly over non-phosphorus containing lipids, and 31P has almost 100%natural abundance so strong signals are obtained. In phospholipidsthere is no significant splitting of the phosphorus signals due to cou-pling, and clearly resolved peaks can be observed; this is in contrastwith 1H NMR where overlapping signals and complex coupling pat-terns complicate the quantification of the signals even with modernhigh field strength instruments such as 800 MHz spectrometers[79]. Both 1H and 31P-NMR have been used in studies of phospho-lipids in vitro and analyses of biological lipid extracts, as they can dis-criminate between saturated, unsaturated and alkylacyl andalkenylacyl species for choline phospholipids and also between PLclasses [80–83]. However, to date very few studies have used theseapproaches to investigate phospholipid oxidation and for the analysisof oxidized phospholipids. The formation of lysophosphatidylcholinesfollowing attack by hypochlorous acid on PC multilamellar liposomes
γγ -hydroxy-alkenals γ -hydroxy-alkenals
Hydroperoxides Hydroperoxides
Alka(e)nodienoates Alka(e)nodienoates
Hock Cleavage
Alkoxyl rearrangement
Alkenals Alkenals
oxidationdecomposition
oxidationC3 decomposition
oxidation
Alkanals Alkanals
cyclisation
Alkanals Alkanals Alkanals Alkanals Furans Furans
Alkanoates Alkanoates
O
OC5H11
O OH
R
O
OOR
O
OOR
OH
O
OR O
O
OR O
O
OR O
O
OR O
OH
O
OR O
OH
O
OR O
dehydration
Fig. 4. Decomposition pathways of hydroperoxide OxPL primary products with formation of shortened OxPL products, and decomposition of (hydroxy)-alkenals by two alternativeroutes (cyclisation and C3 decomposition) with formation of stable oxidized phospholipids as proposed in the literature.
2382 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
[51] and in hepatocytes subjected to CCl4-induced stress [84] hasbeen observed using phosphorus NMR. The appearance of hydroper-oxides and conjugated double bonds in model liposomes subjectedto enzymatic peroxidation was detected by 1H-NMR, although theseproducts could not be detected in extracts of rat liver mitochondriaunder oxidative stress [85]. In addition to this, the oxidation product1-palmitoyl-2-(15-hydroperoxide-arachidonoyl)-PC was detectedafter incubation of PC with soybean lipoxygenase, using 31P-NMRwith an improved solvent system that prevented aggregation [80].The small number of studies using NMR probably reflects the limita-tions of the technique, which include lower sensitivity compared toMS, and for 1H-NMR of complex mixtures, multiple overlapping reso-nances from similar molecules that complicate identification of minorspecies such as oxPL. However, the manufacture of spectrometerswith higher field strengths and methodologies developed for metabo-lomics are steadily improving sensitivity and selectivity, and it is pos-sible that in the future that this technique will become more usefulfor profiling of oxPL in biological samples.
6. Detection and quantification of oxPL in biological systems
There is considerable interest in identifying and quantifying oxi-dized phospholipids in biological samples, as it is increasingly clearthat they are produced in a wide variety of inflammatory situations,and play a role in changes in cell behaviour during pathology[86,87]. The wide variety of oxPLs complicates the acquisition of com-prehensive knowledge about the formation of oxPL in biological sam-ples, and moreover several factors may complicate the detection andquantification of oxPL in biological samples, as described below. It isalso important to bear in mind that oxidized phospholipids may notbe homogenously distributed within cells or tissues with polyunsatu-rated PL in colocalised areas, but during most extraction protocols in-formation on subcellular location is lost. The levels of individualspecies of oxPL are often very low as a percentage of unmodified phos-pholipids, which owing to ion suppression increases the difficulty ofanalysing them by mass spectrometry, unless efficient separationmethods are used.
6.1. Detection of oxPL may be complicated by issues of stability andmetabolism
In attempting to quantify oxPL in biological samples and relatethese to the extent of oxidation damage and lipid peroxidation, it isimportant to bear in mind that some products of lipid peroxidationare inherently chemically unstable, particularly the radical speciesformed in the initial stages of the process, and to a lesser extent, thehydroperoxides that result from the propagation step. The latter canbe metabolized by enzymatic systems in vivo, such as phospholipid-dependent glutathione peroxidase (PhGPx), thioredoxin or other sele-noproteins; these reduce phospholipid hydroperoxides to hydroxy de-rivatives, thus lowering the concentration of primary oxPL productsand increasing the concentration of more stable end products [1]. Fur-ther downstream in the process, the oxPL containing carbonyl groupsformed by fragmentations may react with proteins in the local environ-ment, especially integral membrane proteins, to form adducts, thuslowering the concentration of free secondary oxPL products.Carbonyl-containing products react readily with amines, and can at-tack lysine residues to form Schiff's bases; α,β-unsaturated prod-ucts, such as phospholipids containing a hydroxyalkenal moiety,can also react with histidine, cysteine and lysine residues by Michaeladdition to form protein-LPO adducts, and owing to the aldehydegroup these have the potential to cross-link proteins [71]. In additionto this, the findings that oxPL activates phospholipase A2 were recentlyreported [88], and may also influence the concentration of secondaryoxPL to very low levels in vivo with concomitant increase of lysolipids
and free aldehydic products. The importance of these reactions will bediscussed in more detail in other articles in this issue.
6.2. Quantification of oxPL in cells and tissues
As described above, oxidized phospholipid products can bedetected mostly conveniently by electrospray mass spectrometrycoupled to reverse phase HPLC, and a substantial number of studieshave used this approach to observe the presence of oxidized PL incells and tissue from a variety of oxidative conditions, and demon-strate increased levels compared to controls. Many of the data pub-lished on the analysis of phospholipids in biological samples focuson particular PL classes, especially PC as it is one of the most abundantphospholipids and its high ionisation efficiency facilitates its detec-tion, as mentioned in Section 5.1. For example, hydroxylated PCswere detected at increased levels in plasma samples from breast can-cer patients, and suggested as potential biomarkers based on moresignificant changes than other PL classes [89]. SAPC-HETEs were ob-served in liver of CCl4-treated rats [90], while POVPC, PGPC, PEIPCand PC hydroxyalkenals were observed in extracts of human athero-sclerotic lesions [91,92], but the amounts were not reported. Nakanishiet al. reported increased levels of several oxidized PCs in ischaemicmouse myocardium compared to non-ischaemic tissue, with the datareported as counts/mg tissue [93]. Increases of approximately 10–15-fold have been reported for POVPC, PGPC, PEIPC and PECPC as well aslyso-lipids in cholesterol-fed zebrafish compared to the ones on thecontrol diet [94].
The quantification of oxPL in biological systems is challenging anddepending on the types of biological sample studied, the levels of ox-idized PL analysed are quoted in a variety of different ways, whichcomplicates the comparison of concentrations between studies. Instudies of human monocytes and platelets activated by a calcium ion-ophore or thrombin respectively, levels of HETE-PC were reported as18 ng/4×107 platelets and HETE-PE were 6 ng/4×107 platelets and150 ng/4×106 monocytes [95]. Another group has studied the forma-tion of hydroperoxides of several PL classes in the small intestine ofmice following total body γ-irradiation, and found levels of20–60 mmol/mol parent lipid in treated animals compared to4–7 mmol/mol in un-irradiated animals [59]; interestingly, the levelsof CL and PS increased approximately 10-fold, muchmore than the 3–4-fold observed with more abundant PLs PE and PC. This interestingheadgroup selectivity probably represents a biological process involv-ing a specific interaction of cytochrome c with the anionic phospho-lipids [96]. The same group also measured rat cortical neurones,sheep lung endothelial cells and mouse lung tissue under conditionsthat led to apoptosis, and levels of cardiolipin hydroperoxides varyingfrom 1 to 100 pmol/nmol CL and levels of PS hydroperoxides in therange 3–25 pmol/nmol PS were observed; thus between 0.1 and10% of the parent lipid was oxidized under these conditions [97].There have been few studies of atherosclerotic plaque material bothfrom humans patients and from high-fat diet fed animals, whichhave also reported high levels and significant increases in oxidizedPLs. Watson et al. measured the levels of PGPC, POVPC and PEIPC inthe aorta of rabbits on an atherogenic diet in the range of 40–65 ng/mg wet tissue, representing a 2–3-fold increase over control tissue[11]; while in another study even higher concentrations (50–100 μg/gwet tissue) weremeasured for the same oxidized PLs and their stearoylanalogues [98]. PC core aldehydes POVPC, SOVPC, PONPC, SONPC andHNE-PCwere detected inmaterial from human atherosclerotic plaques,although by far the most abundant component was POVPC at a concen-tration of 20–40 μg/g tissue depending on the plaque type; levels of theother aldehydes were in the range 1–8 μg/g tissue [13]. By contrast,analysis of human or animal plasma samples has indicated ratherlower levels of fragmented oxidized PCs, with concentrations rangingfrom approximately 0.5–6 μM in disease or inflammatory conditions,corresponding to 2 to 10-fold increases relative to control samples
2383A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
[99,100]. However, the ratio of full chain oxidized and fragmented PCsto total PC measured in another study was 0.1–2% [101], which is com-parable to the ratios found in some inflammatory tissue samples men-tioned above.
There are relatively few reports on quantitative analysis ofnitrated and chlorinated PLs in vivo. Several studies have reportedon the plasma levels of nitrated fatty acids in free and esterifiedforms, but there is some disagreement in the literature. An earlystudy reported 0.2 μM and 0.5 μM total esterified nitro-linoleate iso-mers in red blood cells and plasma from healthy humans respectively[102], and subsequently slightly lower esterified nitro-oleate concen-trations of 0.15 μM and 0.3 μMwere measured [103]. However, a laterstudy reported much lower (~1 nM) levels of the free nitrated fattyacids using GC–MSMS, although the esterified forms were not mea-sured [104]. However, it is thought that PL-esterified forms of nitratedfatty acids occur at comparable levels to the free nitrated fatty acids[105], and increased levels of both are expected under pathologicalconditions. The presence of C16:0 and C18:0 lysoPC chlorohydrinshas been detected in human atherosclerotic lesions, and showed 69-fold and 82-fold increases respectively compared to normal aorta, al-though the absolute levels of these compounds were not determined[40].
Thus in studies published so far there is considerable variabilityin the levels of oxPL reported in physiological conditions, and themajority of published reports have analysed carbonyl and carboxylfragmentation products of oxidized PC, although some studieshave also investigated full length oxidized products and other PLclasses. Possible explanations for the variability of biological con-centrations of oxidized phospholipids include the different method-ologies used for extraction of phospholipid from the tissues as wellas the different analytical approaches (direct infusion or MS inter-faced to LC) and the instrumentation employed by the researchers.More rigorous analytical studies are needed to begin to establish trust-worthy concentration ranges for oxidized phospholipids in differentphysiological and pathological conditions. Overall it is clear that thequantification of oxPL in biological systems is challenging, and theamount of oxidized PL in disease states and in healthy conditions re-mains subject of debate.
7. Conclusions and future perspectives
There is clearly undiminished interest in the field of phospholipidoxidation, both in terms of completing our understanding of themechanisms of formation of the wide gamut of products known andelucidating their biological effects. Advanced analytical techniques,especially electrospray mass spectrometry with fragmentationprotocols, have contributed enormously to the determination ofreaction pathways in model systems and detection of oxidizedphospholipids in biological samples. Nevertheless, our knowledgeabout the location and concentrations of these compounds inphysiological and pathological situations is still rather limited.
A fuller understanding of the mechanisms of phospholipid oxida-tion and the resulting products would allow profiling of physiologicaland pathological conditions. Apart from yielding information on typesof oxidants responsible, and therefore their roles in pathology, thiscould enable a “fingerprinting” approach to oxidized phospholipidsas biomarkers of inflammation and injury. Potentially, this opensthe door to a systems biology approach to oxidative lipidomics,which would allow theories of their formation and metabolism tobe tested. However, this will require improved methods of analysisand better (or more widely available) standards. The recent develop-ment of high resolution MS instruments with fast scan speeds andpolarity switching (i.e. Orbitrap instruments) that are suitable forthe screening of both positively and negatively charged compoundsin complex mixtures, will facilitate complete lipidome mapping ofbiological fluids and tissues. Combined with the development of
fluorescent probes for subcellular localization of oxidized phospho-lipids, this should provide novel insights into the biology of thesecompounds.
The interactions of oxidized phospholipids with proteins, eitherinvolving the formation of adducts by reactive carbonyl groups fromfragmented oxPL or noncovalent interactions with receptors areclearly important and need to be explored further. As the phospho-lipid types most susceptible to attack and the reactive productsformed from them are elucidated, this knowledge can be used toimplement targeted mass spectrometry-based identification of ad-ducts formed between oxidized phospholipids and proteins. Thesetopics are the subject of further, more detailed reviews in thisissue.
Abbreviations and definitions
AAPH 2,2′-azobis(2-amidinopropane) dihydrochlorideCer ceramideCL cardiolipinDNPH dinitrophenylhydrazineFA fatty acidLUV large unilamellar vesiclesL• alkyl lipid radicalOL• epoxy-alkyl lipid radicalLOO• peroxyl lipid radicalLO• alkoxyl lipid radicalOLOO• epoxy-peroxyl lipid radicalHOL• hydroxy-alkyl lipid radicalMALDI Matrix-assisted Laser Desorption IonisationMLV multi-lamellar vesiclesMPO myeloperoxidaseoxPL oxidized phospholipidPC phosphatidylcholines lipidsPE phosphatidylethanolamine lipidsPS phosphatidylserine lipidsPAPC 1-palmitoyl-2-arachidonoyl-phosphatidylcholinePDPC 1-palmitoyl-2-docohexaenoyl-phosphatidylcholinePGPC 1-palmitoyl-2-glutaroyl-phosphatidylcholinePL phospholipidPLPC 1-palmitoyl-2-lineloyl-phosphatidylcholinePLOOH phospholipid hydroperoxidesPONPC 1-palmitoyl-2-(oxo-nonanoyl)-phosphatidylcholinePOVPC 1-palmitoyl-2-(oxo-valeroyl)-phosphatidylcholinePhGPx phospholipid hydroperoxide glutathione peroxidaseROS reactive oxygen speciesRA relative abundanceSM sphingomyelinsSUV small unilamellar vesiclesTAG triacylglyceridesDMPO 5,5′-dimethyl-1-pyrrolidine-N-oxide
Acknowledgements
We gratefully acknowledge funding for a Marie-Curie Intra-European Fellowship (FP7-PEOPLE-2009-IEF Project ID 255076“ATHERO_MASS”). CMS would like to acknowledge the COST ActionB35 and 08-EuroMEMBRANES-FP-021 for intellectual support.
References
[1] E. Niki, Y. Yoshida, Y. Saito, N. Noguchi, Lipid peroxidation: mechanisms, inhibi-tion, and biological effects, Biochem. Biophys. Res. Commun. 338 (2005)668–676.
[2] A. Catala, Lipid peroxidation of membrane phospholipids in vertebrate retina,Front. Biosci. 3 (2011) 52–60.
2384 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
[3] P. Fu, K.G. Birukov, Oxidized phospholipids in control of inflammation and endo-thelial barrier, Transl. Res. 153 (2009) 166–176.
[4] I. Levitan, S. Volkov, P.V. Subbaiah, Oxidized LDL: diversity, patterns of recogni-tion and pathophysiology, Antioxid. Redox Signal. 13 (2010) 39–75.
[5] C.M. Spickett, N.M. Fauzi, Analysis of oxidized and chlorinated lipids by massspectrometry and relevance to signalling, Biochem. Soc. Trans. 39 (2011)1233–1239.
[6] O.V. Oskolkova, T. Afonyushkin, B. Preinerstorfer, W. Bicker, E. von Schlieffen, E.Hainzl, S. Demyanets, G. Schabbauer, W. Lindner, A.D. Tselepis, J. Wojta, B.R.Binder, V.N. Bochkov, Oxidized phospholipids are more potent antagonists of lipo-polysaccharide than inducers of inflammation, J. Immunol. 185 (2010) 7706–7712.
[7] K.A.C. Elliot, B. Libet, Oxidation of phospholipid catalysed by iron compoundswith ascorbic acid, J. Biol. Chem. 152 (1944) 617–626.
[8] F. Bernheim, M.L.C. Bernheim, The action of vanadium on the oxidation of phos-pholipids by certain tissues, J. Biol. Chem. 127 (1939) 353–360.
[9] G. Spiteller, The important role of lipid peroxidation processes in aging and agedependent diseases, Mol. Biotechnol. 37 (2007) 5–12.
[10] K.E. Stremler, D.M. Stafforini, S.M. Prescott, G.A. Zimmerman, T.M. McIntyre, Anoxidized derivative of phosphatidylcholine is a substrate for the platelet-activat-ing factor acetylhydrolase from human plasma, J. Biol. Chem. 264 (1989)5331–5334.
[11] A.D. Watson, N. Leitinger, M. Navab, K.F. Faull, S. Horkko, J.L. Witztum, W.Palinski, D. Schwenke, R.G. Salomon, W. Sha, G. Subbanagounder, A.M.Fogelmann, J.A. Berliner, Structural identification bymass spectrometry of oxidizedphospholipids in minimally oxidized low density lipoprotein that induce monocy-te/endothelial interactions and evidence for their presence in vivo, J. Biol. Chem.272 (1997) 13597–13607.
[12] H.F. Hoff, J. O'Neill, S. Wu, G. Hoppe, R. Salomon, Phospholipid hydroxyalkenals.Biological and chemical properties of specific oxidized lipids present in athero-sclerotic lesions, Arterioscler. Thromb. Vasc. Biol. 23 (2000) 275–282.
[13] A. Ravandi, S. Babaei, R. Leung, J.C. Monge, G. Hoppe, H. Hoff, H. Kamido, A.Kuksis, Phospholipids and oxophospholipids in atherosclerotic plaques at differ-ent stages of plaque development, Lipids 39 (2004) 97–109.
[14] O. Panasenko, The mechanism of the hypochlorite-induced lipid peroxidation,Biofactors 6 (1997) 181–190.
[15] D. Ford, Lipid oxidation by hypochlorous acid: chlorinated lipids in atherosclerosisand myocardial ischemia, Clin. Lipidol. 5 (2010) 835–852.
[16] P. Pacher, J.S. Beckman, L. Liaudet, Nitric oxide and peroxynitrite in health anddisease, Physiol. Rev. 87 (2007) 315–424.
[17] C. Szabo, H. Ischiropoulos, R. Radi, Peroxynitrite: biochemistry, pathophysiologyand development of therapeutics, Nat. Rev. Drug Discov. 6 (2007) 662–680.
[18] J. Ciencewicki, S. Trivedi, S.R. Kleeberger, Oxidants and the pathogenesis of lungdiseases, J. Allergy Clin. Immunol. 122 (2008) 456–468.
[19] W. Droge, Free radicals in the physiological control of cell function, Physiol. Rev.82 (2002) 47–95.
[20] J.D. Lambeth, Nox/Duox family of nicotinamide adenine dinucleotide (phosphate)oxidases, Curr. Opin. Hematol. 9 (2002) 11–17.
[21] S.Y. Qian, G.R. Buettner, Iron and dioxygen chemistry is an important route toinitiation of biological free radical oxidations: an electron paramagnetic resonancespin trapping study, Free Radic. Biol. Med. 26 (1999) 1447–1456.
[22] R. Senthilmohan, A.J. Kettle, Bromination and chlorination reactions ofmyeloperoxidase at physiological concentrations of bromide and chloride, Arch.Biochem. Biophys. 445 (2006) 235–244.
[23] J.P. Eiserich, M. Hristova, C.E. Cross, A.D. Jones, B.A. Freeman, B. Halliwell, A. vander Vliet, Formation of nitric oxide-derived inflammatory oxidants by myelo-peroxidase in neutrophils, Nature 391 (1998) 393–397.
[24] B. Halliwell, M.V. Clement, L.H. Long, Hydrogen peroxide in the human body,FEBS Lett. 486 (2000) 10–13.
[25] O. Sergent, I. Morel, J. Cillard, Involvement of metal ions in lipid peroxidation:biological implications, in: S.A.a.S. H (Ed.), Metal Ions in Biological Systems,vol. 36, Marcel Dekker, New York, 1999, pp. 251–262.
[26] M. Comporti, C. Signorini, G. Buonocore, L. Ciccoli, Iron release, oxidative stressand erythrocyte ageing, Free Radic. Biol. Med. 32 (2002) 568–572.
[27] W. Adam, D.V. Kazakov, V.P. Kazakov, Singlet-oxygen chemiluminescence inperoxide reactions, Chem. Rev. 105 (2005) 3371–3387.
[28] T. Grune, P. Schroder, H.K. Biesalski, Low molecular weight antioxidants, TheHandbook of Environmental Chemistry, vol. 2, Springer-Verlag, Heidelberg,2005, pp. 77–90.
[29] L. Gao, W.E. Zackert, J.J. Hasford, M.E. Danekis, G.L. Milne, C. Remmer, J. Reese, H.Yin, H.H. Tai, S.K. Dey, N.A. Porter, J.D. Morrow, Formation of prostaglandins E2and D2 via the isoprostane pathway, J. Biol. Chem. 278 (2003) 28479–28489.
[30] R.G. Salomon, Distinguishing levuglandins produced through the cyclooxygenaseand isoprostane pathways, Chem. Phys. Lipids 134 (2005) 1–20.
[31] M. Guichardant, S. Bacot, P. Moliere, M. Lagarde, Hydroxy-alkenals from theperoxidation of n-3 and n-6 fatty acids and urinary metabolites, Prostaglan-dins Leukot. Essent. Fatty Acids 75 (2006) 179–182.
[32] K.M. Wynalda, R.C. Murphy, Low concentration ozone reacts with plasmalogenglycerophosphoethanolamine lipids in lung surfactant, Chem. Res. Toxicol. 23(2010) 108–117.
[33] E.A. Podrez, E. Poliakov, Z. Shen, R. Zhang, Y. Deng, M. Sun, P.J. Finton, L. Shan, B.Gugiu, P.L. Fox, H.F. Hoff, R.G. Salomon, S.L. Hazen, Identification of a novel family ofoxidized phospholipids that serve as ligands for the macrophage scavenger receptorCD36, J. Biol. Chem. 277 (2002) 38503–38516.
[34] C. Uhlson, K. Harrison, C.B. Allen, S. Ahmad, C.W. White, R.C. Murphy, Oxidizedphospholipids derived from ozone-treated lung surfactant extract reduce mac-rophage and epithelial cell viability, Chem. Res. Toxicol. 15 (2002) 896–906.
[35] N. Khaselev, RC Murphy, Structural characterization of oxidized phospholipidproducts derived from arachidonate-containing plasmenyl glycerophosphocholine,J. Lipid Res. 41 (2000) 564–572.
[36] M.S. Gopfert, F. Siedler, W. Siess, A. Sellmayer, Structural identification of oxi-dized acyl-phosphatidylcholines that induce platelet activation, J. Vasc. Res. 42(2005) 120–132.
[37] A. Reis, M.R. Domingues, F.M. Amado, A.J. Ferrer-Correia, P. Domingues, Separa-tion of peroxidation products of diacyl-phosphatidylcholines by reverse-phaseliquid chromatography-mass spectrometry, Biomed. Chromatogr. 19 (2005)129–137.
[38] B.G. Gugiu, C.A. Mesaros, M. Sun, X. Gu, J.W. Crabb, R.G. Salomon, Identificationof oxidatively truncated ethanolamine phospholipids in retina and their gener-ation from polyunsaturated phosphatidylethanolamines, Chem. Res. Toxicol. 19(2006) 262–271.
[39] K.A. Zemski-Berry, R.C. Murphy, Free radical oxidation of plasmalogen glycero-phosphocholine containing esterified docosahexaenoic acid: structure determi-nation by mass spectrometry, Antioxid. Redox Signal. 7 (2005) 157–169.
[40] M.C. Messner, C.J. Albert, F.-F. Hsu, D.A. Ford, Selective plasmenyl oxidation byhypochlorous acid: formation of lysophosphatidylcholine chlorohydrins, Chem.Phys. Lipids 144 (2006) 34–44.
[41] C.J. Albert, J.R. Crowley, F.F. Hsu, A.K. Thukkani, D.A. Ford, Reactive chlori-nating species produced by myeloperoxidase target the vinyl ether bond of plas-malogens: identification of 2-chlorohexadecanal, J. Biol. Chem. 276 (2001)23733–23741.
[42] J. Arnhold, A.N. Osipov, H. Spalteholz, O.M. Panasenko, J. Schiller, Effects of hypo-chlorous acid on unsaturated phosphatidylcholines, Free Radic. Biol. Med. 31(2001) 1111–1119.
[43] V.B. O'Donnell, J.P. Eiserich, P.H. Chumley, M.J. Jablonsky, N.R. Krishna, M. Kirk, S.Barnes, V.M. Darley-Usmar, B.A. Freeman, Nitration of unsaturated fatty acids bynitric oxide-derived reactive nitrogen species peroxynitrite, nitrous acid, nitrogendioxide, and nitronium ion, Chem. Res. Toxicol. 12 (1999) 83–92.
[44] B.A. Freeman, P.R. Baker, F.J. Schopfer, S.R. Woodcock, A. Napolitano, M. d'Ischia,Nitro-fatty acid formation and signaling, J. Biol. Chem. 283 (2008)15515–15519.
[45] B. Kalyanaraman, Nitrated lipids: a class of cell-signaling molecules, Proc. Natl.Acad. Sci. U. S. A. 101 (2004) 11527–11528.
[46] M.A. Yorek, Biological distribution, in: G. Cevc (Ed.), Phospholipids Handbook,Marcel Dekker, New York, 1993, pp. 745–775.
[47] Q.-T. Li, M.H. Yeo, B.K. Tan, Lipid peroxidation in small and large phospholipidunilamellar vesicles induced by water-soluble free radical sources, Biochem. Bio-phys. Res. Commun. 273 (2000) 72–76.
[48] N. Fagali, A. Catalá, Fe2+ and Fe3+ initiated peroxidation of sonicated andnon-sonicated liposomes made of retinal lipids in different aqueous media,Chem. Phys. Lipids 159 (2009) 88–94.
[49] J. Heuvingh, S. Bonneau, Asymmetric oxidation of giant vesicles triggerscurvature-associated shape transition and permeabilization, Biophys. J. 97(2009) 2904–2912.
[50] A. Reis-Pereira, Identification of free radicals and other oxidation products ofbiomolecules by mass spectrometry, Chemistry Department, vol. PhD, Universityof Aveiro, Aveiro, 2006.
[51] J. Arnhold, A.N. Osipov, H. Spalteholtz, O.M. Panasenko, J. Schiller, Formation oflysophospholipids from unsaturated phosphatidylcholines under the influenceof hypochlorous acid, Biochim. Biophys. Acta 1572 (2002) 91–100.
[52] O.M. Panasenko, T. Vakhrusheva, V. Tretyakov, H. Spalteholz, J. Arnhold, Influ-ence of chloride on modification of unsaturated phosphatidylcholines by themyeloperoxidase/hydrogen peroxide/bromide system, Chem. Phys. Lipids 149(2007) 40–51.
[53] J. Choi, W. Zhang, X. Gu, X. Chen, L. Hong, J.M. Laird, R.G. Salomon, Lysophospha-tidylcholine is generated by spontaneous deacylation of oxidized phospholipids,Chem. Res. Toxicol. 24 (2011) 111–118.
[54] N. Khaselev, RC Murphy, Peroxidation of arachidonate containing plasmenylglycerophosphocholine: facile oxidation of esterified arachidonate at carbon-5,Free Radic. Biol. Med. 29 (2000) 620–632.
[55] S. Mawatari, Y. Okuma, T. Fujino, Separation of intact plasmalogens and all otherphospholipids by a single run of high-performance liquid chromatography, Anal.Biochem. 370 (2007) 54–59.
[56] J.J. Murray, A.R. Brash, Rabbit reticulocyte lipoxygenase catalyzes specific 12(S)and 15(S) oxygenation of arachidonoyl-phosphatidylcholine, Arch. Biochem.Biophys. 265 (1988) 514–523.
[57] H. Arai, T. Suzuki, K. Takama, J. Terao, Reactivity of mammalian 15-lipoxygenasewith phospholipids in large unilamellar liposomes, Biochem. Biophys. Res. Com-mun. 228 (1996) 675–682.
[58] S.R. Clark, C.J. Guy, M.J. Scurr, P.R. Taylor, A.P. Kift-Morgan, V.J. Hammond, C.P.Thomas, B. Coles, G.W. Roberts, M. Eberl, S.A. Jones, N. Topley, S. Kotecha, V.B.O'Donnell, Esterified eicosanoids are acutely generated by 5-lipoxygenase in pri-mary human neutrophils and in human and murine infection, Blood 117 (2011)2033.
[59] Y.Y. Tyurina, V.A. Tyurin, M.W. Epperly, J.S. Greenberger, V.E. Kagan, Oxidativelipidomics of gamma-irradiation-induced intestinal injury, Free Radic. Biol.Med. 44 (2008) 299–314.
[60] C. Ferreri, C. Chatgilialoglu, Membrane lipidomics and the geometry of unsatu-rated fatty acids from biomimetic models to biological consequences, MethodsMol. Biol. 579 (2009) 391–411.
[61] M. Melchiorre, A. Torregianni, C. Chatgilialoglu, C. Ferreri, Lipid markers of “geo-metrical” radical stress: synthesis of monotrans cholesteryl ester isomers anddetection in human plasma, J. Am. Chem. Soc. 133 (2011) 15184–15190.
2385A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
[62] S.H. Lee, T. Oe, J.S. Arora, I.A. Blair, Analysis of Fe(II)-mediated decomposition ofa linoleic acid-derived lipid hydroperoxide by liquid chromatography/massspectrometry, J. Mass Spectrom. 40 (2005) 661–668.
[63] C. Schneider, N.A. Porter, A.R. Brash, Routes to 4-hydroxynonenal: fundamentalissues in the mechanisms of lipid peroxidation, J. Biol. Chem. 283 (2008)15539–15543.
[64] W. Liu, N.A. Porter, C. Schneider, A.R. Brash, H. Yin, Formation of4-hydroxynonenal from cardiolipin oxidation: intramolecular peroxyl radicaladdition and decomposition, Free Radic. Biol. Med. 50 (2011) 166–178.
[65] Y.N. Balamraju, M. Sun, R.G. Salomon, Gamma-hydroxyalkenals are oxidativelycleaved through Michael addition of acylperoxy radicals and fragmentation in-termediate beta-hydroxyperesters, J. Am. Chem. Soc. 126 (2004) 11522–11528.
[66] S. Gao, R. Zhang, M.E. Greenberg, M. Sun, X. Chen, B.S. Levinson, R.G. Salomon,S.L. Hazen, Phospholipid hydroxyalkenals, a subset of recently discovered en-dogenous CD36 ligands, spontaneously generate novel furan-containing phos-pholipids lacking CD36 binding activity in vivo, J. Biol. Chem. 281 (2006)31298–31308.
[67] S.Y. Qian, G.E. Yue, K.B. Tomer, R.P. Mason, Identification of all classes ofspin-trapped carbon centered radicals in soybean lipoxygenase-dependentlipid peroxidations of omega-6 polyunsaturated fatty acids via LC/ESR, LC-MSand tandem mass spectrometry, Free Radic. Biol. Med. 35 (2003) 33.
[68] A. Reis, P. Domingues, A.J. Ferrer-Correia, M.R. Domingues, Identification of freeradicals of glycerophosphatidylcholines containing omega-6 fatty acids usingspin-trapping coupled with tandem mass spectrometry, Free Radic. Res. 41(2007) 432–443.
[69] H. Iwahashi, High-performance liquid chromatography/electron spin resonance/massspectrometry analyses of lipid-derived radicals,MethodsMol. Biol. 477 (2008) 65–73.
[70] A. Reis, M.R. Domingues, F.M. Amado, A.J. Ferrer-Correia, P. Domingues, Identifi-cation of linoleic acid free radicals and other breakdown products usingspin-trapping with liquid chromatography-electrospray tandem massspectrometry, Biomed. Chromatogr. 20 (2006) 109–118.
[71] C.M. Spickett, I.Wiswedel,W. Siems, K. Zarkovic, N. Zarkovic, Advances inmethodsfor the determination of biologically relevant lipid peroxidation products, Free.Radic. Res. 44 (2010) 1172–1202.
[72] T. Miyazawa, T. Suzuki, K. Fujimoto, K. Yasuda, Chemiluminescent simultaneous de-termination of phosphatidylcholine hydroperoxide and phosphatidylethanolaminehydroperoxide in the liver and brain of the rat, J. Lipid Res. 33 (1992) 1051–1058.
[73] S.-P. Hui, H. Chiba, S. Jin, H. Nagasaka, T. Kurosawa, Analyses of phosphatidyl-choline hydroperoxides by LC/MS, J. Chromatogr. B 878 (2010) 1677–1682.
[74] T. Miyazawa, K. Yasuda, K. Fujimoto, T. Kaneda, Presence of phosphatidylcholinehydroperoxide in human plasma, J. Biochem. 103 (1988) 744–746.
[75] E. Koivusalmi, E. Haatainen, A. Root, Quantitative RP-HPLC determination ofsome aldehydes and hydroxyaldehydes as their 2,4-dinitrophenylhydrazone de-rivatives, Anal. Chem. 71 (1999) 86–91.
[76] X. Han, R. Gross, Electrospray ionisation mass spectroscopic analysis of humanerythrocyte plasma membrane phospholipids, Proc. Natl. Acad. Sci. U. S. A. 91(1994) 10635–10639.
[77] M. Koivusalo, P. Haimi, L. Heikinheimo, R. Kostiainen, P. Somerharju, Quantita-tive determination of phospholipid compositions by ESI-MS: effects of acylchain length, unsaturation, and lipid concentration on instrument response, J.Lipid Res. 42 (2001) 663–672.
[78] S.-P. Hui, H. Chiba, T. Kurosawa, Liquid chromatography-mass spectrometric de-termination of plasmalogen in human plasma, Anal. Bioanal. Chem. 400 (2011)1923–1931.
[79] O. Soininen, K. Oorni, H. Maaheimo, R. Laatikainen, P.T. Kovanen, K. Kaski, M.Ala-Korpela, 1H NMR at 800MHz facilitates detailed phospholipid follow-up duringatherogenic modifications in low density lipoproteins, Biochem. Biophys. Res. Com-mun. 360 (2007) 290–294.
[80] J.M. Pearce, R.A. Komoroski, Analysis of phospholipid molecular species in brainby 31P NMR spectroscopy, Magn. Reson. Med. 44 (2000) 215–220.
[81] J. Schiller, O. Zschornig, M. Petkovic, M. Muller, J. Arnhold, K. Arhnold, Lipid anal-ysis of human HDL and LDL by MALDI-TOF mass spectrometry and 31P NMR, J.Lipid Res. 42 (2001) 1501–1508.
[82] M.J. Albers, M.D. Krieger, I. Gonzalez-Gomez, F.H. Gilles, J.G. McComb, M.D.Nelson, S. Bluml, Proton-decoupled 31P MRS in untreated pediatric brain tu-mours, Magn. Reson. Med. 53 (2005) 22–29.
[83] L. Yao, S. Jung, 31P NMR phospholipid profiling of soybean emulsion recoveredfrom aqueous extraction, J. Agric. Food Chem. 58 (2010) 4866–4872.
[84] A. Bauer, A. Schumann,M. Gilbert, C.Wilhelm, J.G. Hengstler, J. Schiller, B. Fuchs, Eval-uation of carbon tetrachloride-induced stress on rat hepatocytes by 31P NMR andMALDI-TOF mass spectrometry: lyso-phosphatidylcholine generation from unsatu-rated phosphatidylcholines, Chem. Phys. Lipids 159 (2009) 21–29.
[85] P. Pollesello, O. Eriksson, B.J. Kvam, S. Paoletti, N.E.L. Saris, 1H-NMR studies oflipid extracts of rat liver mitochondria, Biochem. Biophys. Res. Commun. 179(1991) 904–911.
[86] V. Bochkov, O.V. Oskolkova, K.G. Birukov, A.L. Levonen, C.J. Binder, J. Stockl, Gen-eration and biological activities of oxidized phospholipids, Antioxid. Redox Sig-nal. 12 (2010) 1009–1059.
[87] F.H. Greig, S. Kennedy, C.M. Spickett, Physiological effects of oxidizedphospholipidsand their cellular signaling mechanisms in inflammation, Free Radic. Biol. Med. 52(2012) 266–280.
[88] C. Code, A.K. Mahalka, K. Bry, P.K.J. Kinnunen, Activation of phospholipase A2 by1-palmitoyl-2-(9-oxo-nonanoyl)-sn-glycero-3-phosphocholine in vitro, Bio-chim. Biophys. Acta 1798 (2010) 1593–1600.
[89] L.A. Hammad, G. Wu, M.M. Saleh, I. Klouckova, L.E. Dobrolecki, R.J. Hickey, L.Schnaper,M.V. Novotny, Y.Mechref, Elevated levels of hydroxylated phosphocholine
lipids in the blood serum of breast cancer patients, Rapid Commun. Mass Spectrom.23 (2009) 863–876.
[90] H.Y. Yin, B.E. Cox, W. Liu, N.A. Porter, J.D. Morrow, G.L. Milne, Identification of in-tact oxidation products of glycerophospholipids in vitro and in vivo using nega-tive ion electrospray iontrap mass spectrometry, J. Mass Spectrom. 44 (2009)672–680.
[91] G. Subbanagounder, A.D. Watson, J.A. Berliner, Bioactive products of phospholipidoxidation: isolation, identification, measurement and activities, Free Radic. Biol.Med. 28 (2000) 1751–1761.
[92] H.F. Hoff, J. O'Neil, Z. Wu, G. Hoppe, R.L. Salomon, Phospholipid hydroxyalkenals:biological and chemical properties of specific oxidized lipids present in atheroscleroticlesions, Arterioscler. Thromb. Vasc. Biol. 23 (2003) 275–282.
[93] H. Nakanishi, Y. Iida, T. Shimizu, R. Taguchi, Analysis of oxidized phosphatidylcholinesasmarkers for oxidative stress, usingmultiple reactionmonitoring with theoreticallyexpanded data sets with reversed-phase liquid chromatography/tandemmass spec-trometry, J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 877 (2009) 1366–1374.
[94] L. Fang, R. Harkewicz, K. Hartvigsen, P. Wiesner, S.H. Choi, F. Almazan, J. Pattison, E.Deer, T. Sayaphupha, E.A. Dennis, J.L.Witztum, S. Tsimikas, Y.I.Miller, Oxidized choles-teryl esters and phospholipids in zebrafish larvae fed a high cholesterol diet: macro-phage binding and activation, J. Biol. Chem. 285 (2010) 32343–32351.
[95] A.H. Morgan, V.J. Hammond, L. Morgan, C.P. Thomas, K.A. Tallman, Y.R.Garcia-Diaz, C. McGuigan, M. Serpi, N.A. Porter, R.C. Murphy, V.B. O'Donnell,Quantitative assays for esterified oxylipins generated by immune cells, Nat. Pro-toc. 5 (2010) 1919–1931.
[96] H. Bayir, B. Fadeel, M.J. Palladino, E. Witasp, I.V. Kurnikov, Y.Y. Tyurina, V.A.Tyurin, A.A. Amoscato, J. Jiang, P.M. Kchanek, S.T. DeKosky, J.S. Greenberger,A.A. Shvedova, V.E. Kagan, Apoptotic interactions of cytochrome c: redox flirtingwith anionic phospholipids within and outside of mitochondria, Biochim. Biophys.Acta 1757 (2006) 648–659.
[97] V.A. Tyurin, Y.Y. Tyurina, M.Y. Jung, M.A. Tungekar, K.J. Wasserloos, H. Bayir, J.S.Greenberger, P.M. Kochanek, A.A. Shvedova, B. Pitt, V.E. Kagan,Mass-spectrometricanalysis of hydroperoxy- and hydroxy-derivatives of cardiolipin and phosphatidyl-serine in cells and tissues induced by pro-apoptotic and pro-inflammatory stimuli,J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 877 (2009) 2863–2872.
[98] G. Subbanagounder, N. Leitinger, D.C. Schwenke, J.W. Wong, H. Lee, C. Rizza, A.D.Watson, K.F. Faull, A.M. Fogelman, J.A. Berliner, Determinants of bioactivity ofoxidized phospholipids. Specific oxidized fatty acyl groups at the sn-2 position,Arterioscler. Thromb. Vasc. Biol. 20 (2000) 2248–2254.
[99] E.A. Podrez, T.V. Byzova, M. Febbraio, R.G. Salomon, Y. Ma, M. Valiyaveettil, E.Poliakov, M. Sun, P.J. Finton, B.R. Curtis, J. Chen, R. Zhang, R.L. Silverstein, S.L.Hazen, Platelet CD36 links hyperlipidemia, oxidant stress and a prothromboticphenotype, Nat. Med. 13 (2007) 1086–1095.
[100] B. Frey, R. Haupt, S. Alms, G. Holzmann, T. Konig, H. Kern, W. Kox, B. Rustow, M.Schlame, Increase in fragmented phosphatidylcholine in blood plasma by oxidativestress, J. Lipid Res. 41 (2000) 1145–1153.
[101] B. Davis, G. Koster, L.J. Douet, M. Scigelova, G. Woffendin, J.M. Ward, A. Smith, J.Humphries, K.G. Burnand, C.H. Macphee, A.D. Postle, Electrospray ionizationmass spectrometry identifies substrates and products of lipoprotein-associatedphospholipase A2 in oxidized human low density lipoprotein, J. Biol. Chem.283 (2008) 6428–6437.
[102] P.R.S. Baker, F.J. Schopfer, S. Sweeney, B.A. Freeman, Red cell membrane andplasma linoleic acid nitration products: synthesis, clinical identification, andquantitation, Proc. Natl. Acad. Sci. U. S. A. 101 (2004) 11577–11582.
[103] P.R.S. Baker, Y.M. Lin, F.J. Schopfer, S.R. Woodcock, A.L. Groeger, C. Batthyany, S.Sweeney, M.H. Long, K.E. Iles, L.M.S. Baker, B.P. Branchaud, Y.Q.E. Chen, B.A.Freeman, Fatty acid transduction of nitric oxide signaling — multiple nitratedunsaturated fatty acid derivatives exist in human blood and urine and serve asendogenous peroxisome proliferator-activated receptor ligands, J. Biol. Chem.280 (2005) 42464–42475.
[104] D. Tsikas, A.A. Zoerner, A. Mitschke, F.M. Gutzki, Nitro-fatty acids occur inhuman plasma in the picomolar range: a targeted nitro-lipidomics GC-MS/MSstudy, Lipids 44 (2009) 855–865.
[105] D. Tsikas, A.A. Zoerner, J. Jordan, Oxidized and nitrated oleic acid in biologicalsystems: analysis byGC-MS/MSand LC-MS/MS, and biological significance, Biochim.Biophys. Acta Mol. Cell Biol. Lipids 1811 (2011) 694–705.
[106] Y. Kawai, A. Ogamo, Y. Nakagawa, Formation of the aldehydic choline glycero-phospholipids in human red blood cell membrane peroxidized with an azo ini-tiator, J. Biochem. 126 (1999) 115–120.
[107] A.D. Watson, J.A. Berliner, S.Y. Hama, B.N. La Du, K.F. Faull, A.M. Fogelman, M.Navab, Protective effect of high density lipoprotein associated paraoxonase, In-hibition of the biological activity of minimally oxidized low density lipoprotein.J. Clin. Invest. 96 (1995) 2882–2891.
[108] X.H. Wang, H. Ushio, T. Ohshima, Distributions of hydroperoxide positionalisomers generated by oxidation of 1-palmitoyl-2-arachidonoyl-sn-glycero-3--phosphocholine in liposomes and in methanol solution, Lipids 38 (2003) 65–72.
[109] A. Jerlich, A.R. Pitt, R.J. Schaur, C.M. Spickett, Pathways of phospholipid oxida-tion by HOCl in human LDL detected by LC-MS, Free Radic. Biol. Med. 28(2000) 673–682.
[110] F. Gruber, O. Oskolkova, A. Leitner, M. Mildner, V. Mlitz, B. Lengauer, A. Kadl, P.Mrass, G. Kronke, B.R. Binder, V.N. Bochkov, N. Leitinger, E. Tschachler, Photoox-idation generates biologically active phospholipids that induce heme oxyge-nase-1 in skin cells, J. Biol. Chem. 282 (2007) 16934–16941.
[111] G.K. Marathe, C. Johnson, S.D. Billings, M.D. Southall, Y. Pei, D. Spandau, R.C.Murphy, G.A. Zimmerman, T.M. McIntyre, J.B. Travers, Ultraviolet B radiationgenerates platelet-activating factor-like phospholipids underlying cutaneousdamage, J. Biol. Chem. 280 (2005) 35448–35457.
2386 A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387

Author's personal copy
[112] O.M. Panasenko, H. Spalteholz, J. Schiller, J. Arnhold, Myeloperoxidase-inducedformation of chlorohydrins and lysophospholipids from unsaturated phosphati-dylcholines, Free Radic. Biol. Med. 34 (2003) 553–562.
[113] L.S. Huang, M.R. Kim, D.E. Sok, Regulation of lipoxygenase activity by polyunsat-urated lysophosphatidylcholines or their oxygenation derivatives, J. Agric. FoodChem. 56 (2008) 7808–7814.
2387A. Reis, C.M. Spickett / Biochimica et Biophysica Acta 1818 (2012) 2374–2387





























