Embed Size (px)
Citation preview

10.1128/MCB.20.22.8602-8612.2000.
2000, 20(22):8602. DOI:Mol. Cell. Biol. Juan Méndez and Bruce Stillman Prereplication Complexes in Late Mitosisduring the Cell Cycle: Assembly of Minichromosome Maintenance ProteinsRecognition Complex, Cdc6, and Chromatin Association of Human Origin
http://mcb.asm.org/content/20/22/8602Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/20/22/8602#ref-list-1at:
This article cites 68 articles, 42 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/00/$04.0010
Nov. 2000, p. 8602–8612 Vol. 20, No. 22
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Chromatin Association of Human Origin Recognition Complex,Cdc6, and Minichromosome Maintenance Proteins during
the Cell Cycle: Assembly of PrereplicationComplexes in Late MitosisJUAN MENDEZ AND BRUCE STILLMAN*
Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 11724
Received 7 July 2000/Returned for modification 2 August 2000/Accepted 15 August 2000
Evidence obtained from studies with yeast and Xenopus indicate that the initiation of DNA replication is amultistep process. The origin recognition complex (ORC), Cdc6p, and minichromosome maintenance (MCM)proteins are required for establishing prereplication complexes, upon which initiation is triggered by theactivation of cyclin-dependent kinases and the Dbf4p-dependent kinase Cdc7p. The identification of humanhomologues of these replication proteins allows investigation of S-phase regulation in mammalian cells. Usingcentrifugal elutriation of several human cell lines, we demonstrate that whereas human Orc2 (hOrc2p) andhMcm proteins are present throughout the cell cycle, hCdc6p levels vary, being very low in early G1 andaccumulating until cells enter mitosis. hCdc6p can be polyubiquitinated in vivo, and it is stabilized byproteasome inhibitors. Similar to the case for hOrc2p, a significant fraction of hCdc6p is present on chromatinthroughout the cell cycle, whereas hMcm proteins alternate between soluble and chromatin-bound forms.Loading of hMcm proteins onto chromatin occurs in late mitosis concomitant with the destruction of cyclin B,indicating that the mitotic kinase activity inhibits prereplication complex formation in human cells.
The molecular mechanism that restricts firing of origins ofreplication to once per cell cycle invokes the ordered bindingto and/or release of different replication proteins from specificDNA sequences (replicators) located in the vicinity of theactual origins of DNA replication. Following separation ofsister chromatids at mitosis and during the subsequent G1phase, prereplication complexes (pre-RCs) are formed at ori-gins of DNA replication. Initiation of DNA replication is trig-gered by the action of at least two sets of protein kinaseactivities, cyclin-dependent kinases (CDKs) and Dbf4p-Cdc7p.After initiation, the protein complex at each origin changes toa postreplication state (post-RC), thereby preventing furtherinitiation events for the rest of the cell cycle (reviewed inreferences 15 and 57).
The origin recognition complex (ORC), a six-subunit initia-tor protein (2), is present in both pre- and post-RCs (10), andone of its functions is to mark the position of replicationorigins in the genome. The pre-RC is established by the reg-ulated binding of additional factors, which include Cdc6p andthe minichromosome maintenance (MCM) proteins. YeastCDC6 displays a genetic interaction with the ORC and is acritical factor for establishing the competence of replicationorigins once per cell cycle (12, 33, 47, 48). Besides its functionin DNA replication, it may also be involved in a mitotic check-point control, because Cdc6-deprived yeast cells that do notreplicate DNA still undergo a reductional mitosis (4, 47, 63).
Cdc6p is a member of the large AAA1 superfamily ofATPases, which includes Orc1p, Orc4p, Orc5p, MCM, pro-teins and replication factor C (42). Based on sequence simi-larities between Cdc6p, replication factor C, and other AAA1
family members and on the characterization of a dominant-
negative CDC6 mutant, it has been proposed that yeast Cdc6pmight function as an ATP-dependent MCM protein loader(45, 63). Indeed, the association of MCM proteins with chro-matin is dependent on Cdc6p (1, 12, 34, 59). Biochemicalstudies with Xenopus provided additional support for the ideaof Xenopus Cdc6p (XCdc6p) being an essential factor for es-tablishing pre-RCs. In Xenopus egg cell extracts, XCdc6 couldbind to chromatin only in the presence of XOrc2, and it wasabsolutely required for the subsequent loading of XMcm3 (6).
Yeast Cdc6p is a highly unstable protein, and many factorsseem to be involved in its degradation, including the CDC4-CDC34-CDC53-Skp1 pathway (13, 14, 54). However, ectopicexpression of Cdc6p in G2 cells is not deleterious for the cell,and it has been shown that Cdc6p cannot induce MCM proteinbinding to chromatin at this point unless CDKs are inactivated(9, 59). Interestingly, a dominant gain-of-function allele ofCDC6 causes persistent MCM protein binding to chromatinand overreplication of the genome in a single cell cycle (34).
Cdc181 is the Schizosaccharomyces pombe homologue ofCDC6 and performs similar functions in regulating initiation ofDNA replication and possibly entry into mitosis (28, 40, 43).Gross overexpression of Cdc181 results in repeated rounds ofDNA replication in the absence of mitosis (23, 43). p65Cdc18 isalso a very labile protein targeted for destruction by CDKphosphorylation after cells enter S phase (24). At least in S.pombe and Xenopus, another protein, called Cdt1, participatesin the assembly of pre-RCs (37, 44).
MCM proteins are essential for DNA replication (reviewedin reference 27). A subcomplex of human MCM proteins mayfunction as a replicative helicase (22). This idea has beenrecently reinforced by the finding that yeast MCM proteins arerequired for replication fork progression (32) and by the char-acterization of a processive helicase activity in an archaealMCM protein (5, 29).
The regulation of initiation of replication in mammaliancells is much less understood, mainly for two reasons: discrete
* Corresponding author. Mailing address: Cold Spring Harbor Lab-oratory, P.O. Box 100, 1 Bungtown Rd., Cold Spring Harbor, NY11724. Phone: (516) 367 8384. Fax: (516) 367 8879. E-mail: [email protected].
8602
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

replicator sequences have not been defined, and the informa-tion regarding mammalian proteins involved in this process hasbeen limited. The recent identification of human homologuesof ORC subunits (18, 49, 50, 62), Cdc6 (65), and MCM pro-teins (reference 27 and references therein) suggests that theoverall mechanism for replication initiation is conserved be-tween mammals and yeast.
We have therefore investigated the dynamics of chromatinassociation of endogenous human Orc2, Cdc6, and Mcm pro-teins (referred to as hOrc2p, hCdc6p, and hMcm proteins)throughout the cell cycle. We used centrifugal elutriation as amethod to synchronize cells without interfering with their nor-mal metabolism, and we raised new monoclonal anti-hCdc6pantibodies and polyclonal anti-hMcm antibodies. Our datashow that a fraction of hCdc6p is targeted to chromatin duringthe entire cell cycle and help to define the point at which thehMcm proteins are loaded onto chromatin during late mitosis.We also report a previously unrevealed regulated destructionof hCdc6p and discuss the possible execution points for hCdc6pduring the cell cycle.
MATERIALS AND METHODS
Antibodies. To generate anti-hCdc6p monoclonal antibodies, full-length hCdc6pwas overexpressed in Escherichia coli and purified as a glutathione S-transferase(GST) fusion protein. After cleavage of the GST moiety, hCdc6p was furtherpurified in a Prep Cell gel (Bio-Rad) and used as an immunogen. Standardtechniques for immunization of mice, analysis of test bleeds, generation ofhybridoma cell lines, and production of ascitic fluid were used (19). A doublescreening of hybridomas was done by dot blotting, using either native or dena-tured hCdc6p as the antigen immobilized to nitrocellulose. Hybridoma cell lineshCdc6-26 and hCdc6-37 were positive in both tests. Several other positive celllines (hCdc6-34, hCdc6-39, and hCdc6-41) turned out to produce immunoglob-ulin Ms (IgMs) and were not used in this work (unpublished data). Anti-hOrc2pand anti-hCdc6p polyclonal antibodies have been described before (18, 65).Different anti-hMcm3p, anti-hMcm4p, and anti-hMcm5p polyclonal sera wereraised in rabbits against synthetic peptides corresponding to amino acids 674 to693 of hMcm3p, amino acids 21 to 40 of hMcm4p, and amino acids 19 to 38 ofhMcm5p, conjugated to keyhole limpet hemocyanin. Anti-pan-Mcm polyclonalserum was raised against a synthetic peptide that is highly conserved in all MCMproteins (amino acids 405 to 421 of hMcm3p). Antibodies against TFIIB were agift from N. Hernandez (Cold Spring Harbor Laboratory). Monoclonal antibody12CA5 (antihemagglutinin [anti-HA]) was obtained from C. Bautista (ColdSpring Harbor Laboratory). Anti-MEK2 (M24520) was obtained from Trans-duction Laboratories, anti-cyclin A and anti-p53 were obtained from Santa CruzBiotechnology, and Texas Red-conjugated anti-rabbit IgG was obtained fromJackson Immunoresearch Laboratories, Inc.
Cell manipulations and centrifugal elutriation. Cell lines HeLa, U2OS, 293,Manca (B-cell lymphoma), and Raji (Burkitt lymphoma) were obtained from theCold Spring Harbor Laboratory cell culture facility or the American Type Cul-ture Collection. Separation of logarithmically growing cells into distinct cell cyclephases was accomplished by centrifugal elutriation in a Beckman J2-21 M cen-trifuge and a JE-6B rotor with a large (40-ml) separation chamber. The rotor waskept at a speed of 1,500 rpm, the temperature was 20°C, and the medium flowwas controlled with a Cole-Parmer Masterflex pump. Consecutive fractions of250 ml were collected at increasing flow rates, ranging from 40 to 120 ml/min.Cells in all fractions were counted, and extracts were prepared after normaliza-tion by cell number. For cytofluorometric analyses, an aliquot of 106 cells wasfixed by rapid submersion in 1 ml of ice-cold 90% ethanol. After at least 1 h offixation at 4°C, cells were collected by centrifugation and incubated for 30 min at37°C in 0.5 ml of staining solution (25 mg of propidium iodide per ml and 10 mgof RNase per ml in phosphate-buffered saline [PBS]). Stained cells were ana-lyzed on a Becton-Dickinson FACScan.
To synchronize a cell culture in prometaphase, 50 ng of nocodazole per ml wasadded to 1 liter of HeLa cells growing in suspension medium (Joklik’s modifi-cation to minimum essential medium supplemented with 10% calf serum). After25 h, the culture was centrifuged at low speed (5 min at 3,000 rpm in a BeckmanCS-6R centrifuge), washed with PBS, and resuspended in prewarmed freshmedium without nocodazole. Every 20 min after the release, an aliquot was takenfrom the main culture, and the cells were collected, washed once in PBS, andused to prepare total cell extracts or subjected to biochemical fractionation asdescribed below. A total of 106 cells from each fraction were used for cytoflu-orometric analysis of DNA content.
Total cell extracts, nuclear extracts, and chromatin isolation. To prepare totalcell extracts, tissue-cultured cells were harvested by centrifugation, washed inPBS, and directly resuspended in Laemmli buffer, followed by sonication for 15 sin a Tekmar CV26 sonicator set at 25% amplitude. To prepare nuclear extracts,
the cells were washed once with PBS and lysed by Dounce homogenization inhypotonic buffer (20 mM Hepes-KOH [pH 8.0], 5 mM KCl, 1.5 mM MgCl2, 5mM Na butyrate, 0.1 mM dithiothreitol [DTT]). Nuclei were collected by cen-trifugation (10 min, 16,000 3 g, 4°C) and resuspended in nuclear extractionbuffer (15 mM Tris-HCl [pH 7.5], 1 mM EDTA, 0.4 M NaCl, 10% sucrose, 1 mMDTT). After 30 min on ice, insoluble proteins were removed from the nuclearextract by high-speed centrifugation (40 min, 100,000 3 g, 4°C).
To isolate chromatin, cells were resuspended (4 3 107 cells/ml) in buffer A (10mM HEPES, [pH 7.9], 10 mM KCl, 1.5 mM MgCl2, 0.34 M sucrose, 10%glycerol, 1 mM DTT, 5 mg of aprotinin per ml, 5 mg of leupeptin per ml, 0.5 mgof pepstatin A per ml 0.1 mM phenylmethylsulfonyl fluoride). Triton X-100(0.1%) was added, and the cells were incubated for 5 min on ice. Nuclei werecollected in pellet 1 (P1) by low-speed centrifugation (4 min, 1,300 3 g, 4°C). Thesupernatant (S1) was further clarified by high-speed centrifugation (15 min,20,000 3 g, 4°C) to remove cell debris and insoluble aggregates. Nuclei werewashed once in buffer A, and then lysed in buffer B (3 mM EDTA, 0.2 mMEGTA, 1 mM DTT, protease inhibitors as described above). Insoluble chromatinwas collected by centrifugation (4 min, 1,700 3 g, 4°C), washed once in buffer B,and centrifuged again under the same conditions. The final chromatin pellet (P3)was resuspended in Laemmli buffer and sonicated for 15 s in a Tekmar CV26sonicator using a microtip at 25% amplitude (see Fig. 3A). To release chromatin-bound proteins by nuclease treatment, cell nuclei (P1) were resuspended inbuffer A plus 1 mM CaCl2 and 0.2 U of micrococcal nuclease (Sigma). Afterincubation at 37°C for 1 min, the nuclease reaction was stopped by the additionof 1 mM EGTA. Nuclei were collected by low-speed centrifugation and lysedaccording to the chromatin isolation protocol described above.
Immunoblots, immunoprecipitation, and immunofluorescence. Standard pro-tocols for immunoblots, immunoprecipitation, and immunofluorescence wereused (19), with the following notations. Proteins were blotted to nitrocellulosemembranes for 1 h at 0.5 A using the buffer described by Bolt and Mahoney (3)(40 mM Tris, 20 mM sodium acetate, 2 mM EDTA, 20% [vol/vol] methanol,0.05% sodium dodecyl sulfate [SDS]). For immunofluorescence, HeLa cellsgrowing on coverslips were fixed with 3% paraformaldehyde in PBS (pH 7.4) for15 min at room temperature and permeabilized with 0.2% Triton X-100 in PBSfor 5 min at 4°C. Cells were incubated with PBS containing 1% normal goatserum as a blocking agent for 30 min and then with anti-hMcm4p antibody (1:200in PBS–1% normal goat serum) for 1 h. After being washed three times for 5 minwith PBS, the cells were incubated with Texas Red-conjugated anti-rabbit IgG(from goat) for 1 h. DNA was stained with 1 mg of Hoechst stain per ml for 30 s.
Proteasome inhibition and in vivo ubiquitination assays. HeLa cells (106)were treated for 12 h with protease inhibitor LLnL, LLM, MG132, or b-lactoneor proteasome inhibitor I (PSI) (all inhibitors were from Calbiochem) at aconcentration of 25 mM. Cells were then harvested and extracts were preparedexactly as described by Salghetti et al. (53). Ubiquitinated intermediates inhuman cells were detected using the His6-tagged-ubiquitin (His-Ubi) method ofTrier et al. (61) as used by Salghetti et al. (53). HeLa cells (106) were transfectedwith 2 mg of pCGN.CSH.FL42 (which encodes HA-tagged hCdc6p), pCGN.Myc,or pCGN.TFIIB, either in the absence or in the presence of plasmid pMT107,which encodes His-Ubi (a gift from D. Bohmann, EMBL, Heidelberg, Germa-ny). At 24 h posttransfection, 20 mM MG132 was added to the cells, whenindicated. Cells were harvested at 36 h posttransfection, and His-tagged (there-fore, ubiquitinated) proteins were purified on Ni-nitrilotriacetic acid–agaroseand subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE). HA-tagged proteins were detected by immunoblotting with anti-HA (12CA5) anti-body.
RESULTS
Generation and characterization of monoclonal anti-hCdc6pantibodies. To study the regulation of CDC6 in mammaliancells, monoclonal antibodies against full-length recombinanthCdc6p were raised (see Materials and Methods). Two mono-clonal antibodies (hCdc6-26 and hCdc6-37) were characterizedin detail and used throughout this study. In immunoblots, bothantibodies recognized a protein of approximately 62 kDa inprotein extracts from human cells (Fig. 1A). In some experi-ments, a second band of slightly lower molecular mass was alsodetected, most likely corresponding to a modified form ora degradation product of hCdc6p. The specificities of bothhCdc6-26 and hCdc6-37 in immunoblots were equal to or bet-ter than those of several anti-hCdc6p polyclonal sera tried (Fig.1A and data not shown). The new antibodies efficiently recog-nized recombinant hCdc6p expressed in E. coli or baculovirus-infected insect cells (not shown).
The antibodies also recognized hCdc6p in its native form, asshown by immunoprecipitation from human nuclear proteinextracts: both hCdc6-26 and hCdc6-37, but not a control anti-
VOL. 20, 2000 Cdc6 IN THE MAMMALIAN CELL CYCLE 8603
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
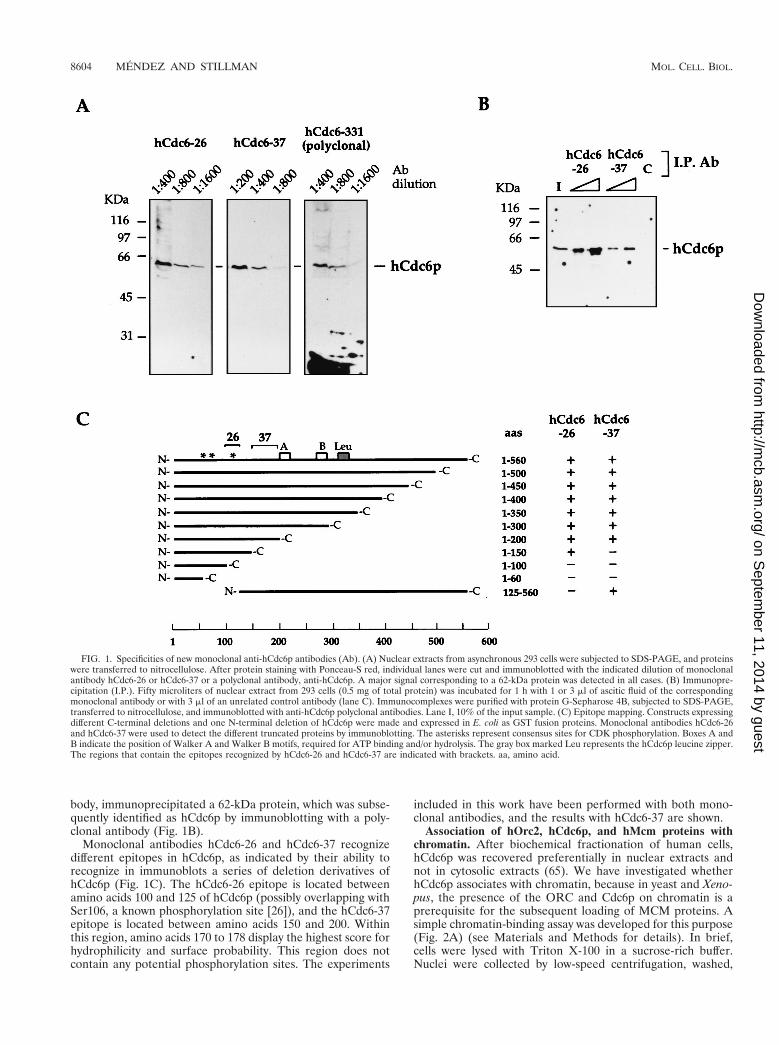
body, immunoprecipitated a 62-kDa protein, which was subse-quently identified as hCdc6p by immunoblotting with a poly-clonal antibody (Fig. 1B).
Monoclonal antibodies hCdc6-26 and hCdc6-37 recognizedifferent epitopes in hCdc6p, as indicated by their ability torecognize in immunoblots a series of deletion derivatives ofhCdc6p (Fig. 1C). The hCdc6-26 epitope is located betweenamino acids 100 and 125 of hCdc6p (possibly overlapping withSer106, a known phosphorylation site [26]), and the hCdc6-37epitope is located between amino acids 150 and 200. Withinthis region, amino acids 170 to 178 display the highest score forhydrophilicity and surface probability. This region does notcontain any potential phosphorylation sites. The experiments
included in this work have been performed with both mono-clonal antibodies, and the results with hCdc6-37 are shown.
Association of hOrc2, hCdc6p, and hMcm proteins withchromatin. After biochemical fractionation of human cells,hCdc6p was recovered preferentially in nuclear extracts andnot in cytosolic extracts (65). We have investigated whetherhCdc6p associates with chromatin, because in yeast and Xeno-pus, the presence of the ORC and Cdc6p on chromatin is aprerequisite for the subsequent loading of MCM proteins. Asimple chromatin-binding assay was developed for this purpose(Fig. 2A) (see Materials and Methods for details). In brief,cells were lysed with Triton X-100 in a sucrose-rich buffer.Nuclei were collected by low-speed centrifugation, washed,
FIG. 1. Specificities of new monoclonal anti-hCdc6p antibodies (Ab). (A) Nuclear extracts from asynchronous 293 cells were subjected to SDS-PAGE, and proteinswere transferred to nitrocellulose. After protein staining with Ponceau-S red, individual lanes were cut and immunoblotted with the indicated dilution of monoclonalantibody hCdc6-26 or hCdc6-37 or a polyclonal antibody, anti-hCdc6p. A major signal corresponding to a 62-kDa protein was detected in all cases. (B) Immunopre-cipitation (I.P.). Fifty microliters of nuclear extract from 293 cells (0.5 mg of total protein) was incubated for 1 h with 1 or 3 ml of ascitic fluid of the correspondingmonoclonal antibody or with 3 ml of an unrelated control antibody (lane C). Immunocomplexes were purified with protein G-Sepharose 4B, subjected to SDS-PAGE,transferred to nitrocellulose, and immunoblotted with anti-hCdc6p polyclonal antibodies. Lane I, 10% of the input sample. (C) Epitope mapping. Constructs expressingdifferent C-terminal deletions and one N-terminal deletion of hCdc6p were made and expressed in E. coli as GST fusion proteins. Monoclonal antibodies hCdc6-26and hCdc6-37 were used to detect the different truncated proteins by immunoblotting. The asterisks represent consensus sites for CDK phosphorylation. Boxes A andB indicate the position of Walker A and Walker B motifs, required for ATP binding and/or hydrolysis. The gray box marked Leu represents the hCdc6p leucine zipper.The regions that contain the epitopes recognized by hCdc6-26 and hCdc6-37 are indicated with brackets. aa, amino acid.
8604 MENDEZ AND STILLMAN MOL. CELL. BIOL.
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
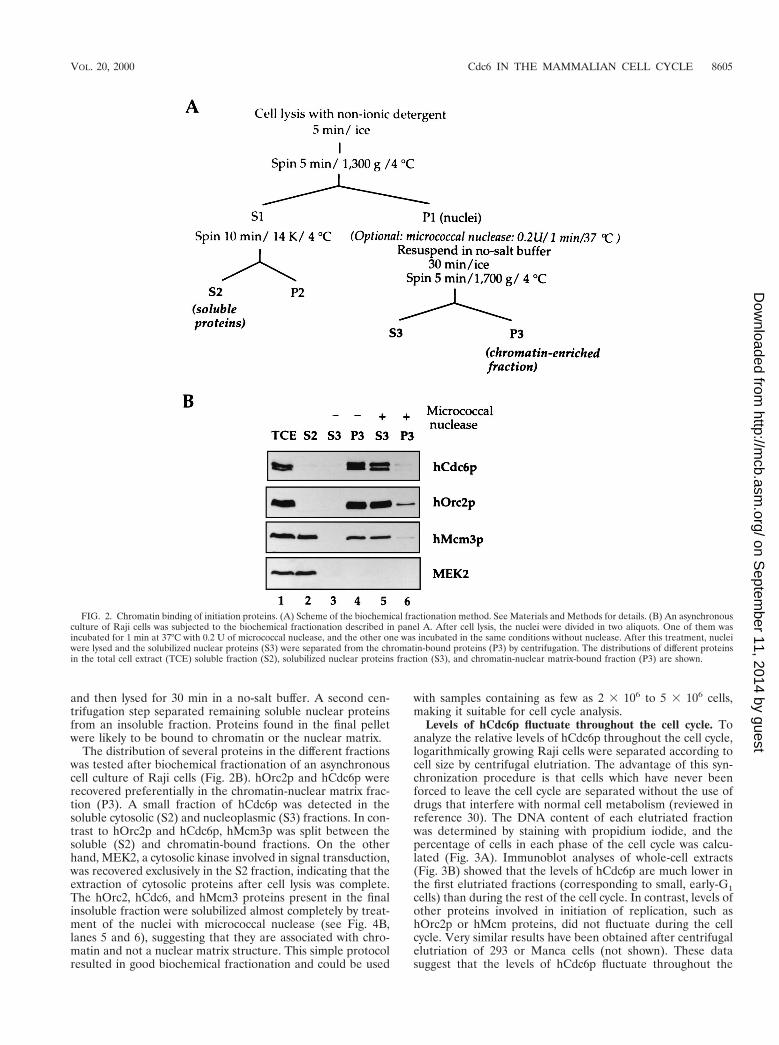
and then lysed for 30 min in a no-salt buffer. A second cen-trifugation step separated remaining soluble nuclear proteinsfrom an insoluble fraction. Proteins found in the final pelletwere likely to be bound to chromatin or the nuclear matrix.
The distribution of several proteins in the different fractionswas tested after biochemical fractionation of an asynchronouscell culture of Raji cells (Fig. 2B). hOrc2p and hCdc6p wererecovered preferentially in the chromatin-nuclear matrix frac-tion (P3). A small fraction of hCdc6p was detected in thesoluble cytosolic (S2) and nucleoplasmic (S3) fractions. In con-trast to hOrc2p and hCdc6p, hMcm3p was split between thesoluble (S2) and chromatin-bound fractions. On the otherhand, MEK2, a cytosolic kinase involved in signal transduction,was recovered exclusively in the S2 fraction, indicating that theextraction of cytosolic proteins after cell lysis was complete.The hOrc2, hCdc6, and hMcm3 proteins present in the finalinsoluble fraction were solubilized almost completely by treat-ment of the nuclei with micrococcal nuclease (see Fig. 4B,lanes 5 and 6), suggesting that they are associated with chro-matin and not a nuclear matrix structure. This simple protocolresulted in good biochemical fractionation and could be used
with samples containing as few as 2 3 106 to 5 3 106 cells,making it suitable for cell cycle analysis.
Levels of hCdc6p fluctuate throughout the cell cycle. Toanalyze the relative levels of hCdc6p throughout the cell cycle,logarithmically growing Raji cells were separated according tocell size by centrifugal elutriation. The advantage of this syn-chronization procedure is that cells which have never beenforced to leave the cell cycle are separated without the use ofdrugs that interfere with normal cell metabolism (reviewed inreference 30). The DNA content of each elutriated fractionwas determined by staining with propidium iodide, and thepercentage of cells in each phase of the cell cycle was calcu-lated (Fig. 3A). Immunoblot analyses of whole-cell extracts(Fig. 3B) showed that the levels of hCdc6p are much lower inthe first elutriated fractions (corresponding to small, early-G1cells) than during the rest of the cell cycle. In contrast, levels ofother proteins involved in initiation of replication, such ashOrc2p or hMcm proteins, did not fluctuate during the cellcycle. Very similar results have been obtained after centrifugalelutriation of 293 or Manca cells (not shown). These datasuggest that the levels of hCdc6p fluctuate throughout the
FIG. 2. Chromatin binding of initiation proteins. (A) Scheme of the biochemical fractionation method. See Materials and Methods for details. (B) An asynchronousculture of Raji cells was subjected to the biochemical fractionation described in panel A. After cell lysis, the nuclei were divided in two aliquots. One of them wasincubated for 1 min at 37°C with 0.2 U of micrococcal nuclease, and the other one was incubated in the same conditions without nuclease. After this treatment, nucleiwere lysed and the solubilized nuclear proteins (S3) were separated from the chromatin-bound proteins (P3) by centrifugation. The distributions of different proteinsin the total cell extract (TCE) soluble fraction (S2), solubilized nuclear proteins fraction (S3), and chromatin-nuclear matrix-bound fraction (P3) are shown.
VOL. 20, 2000 Cdc6 IN THE MAMMALIAN CELL CYCLE 8605
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
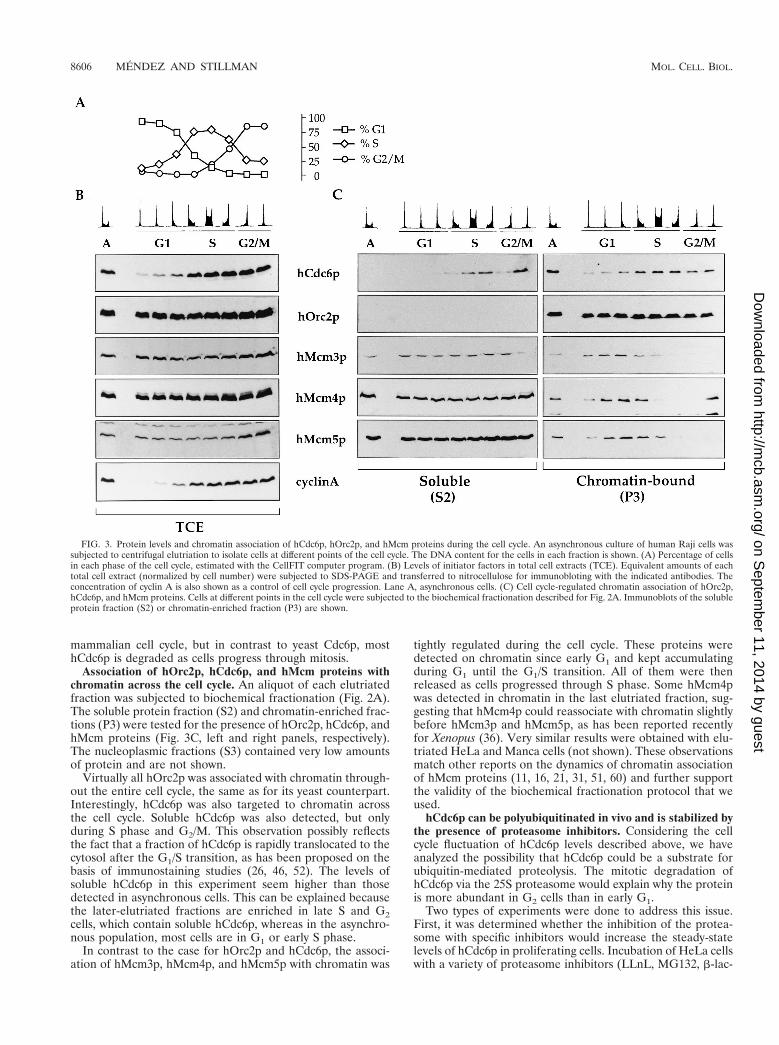
mammalian cell cycle, but in contrast to yeast Cdc6p, mosthCdc6p is degraded as cells progress through mitosis.
Association of hOrc2p, hCdc6p, and hMcm proteins withchromatin across the cell cycle. An aliquot of each elutriatedfraction was subjected to biochemical fractionation (Fig. 2A).The soluble protein fraction (S2) and chromatin-enriched frac-tions (P3) were tested for the presence of hOrc2p, hCdc6p, andhMcm proteins (Fig. 3C, left and right panels, respectively).The nucleoplasmic fractions (S3) contained very low amountsof protein and are not shown.
Virtually all hOrc2p was associated with chromatin through-out the entire cell cycle, the same as for its yeast counterpart.Interestingly, hCdc6p was also targeted to chromatin acrossthe cell cycle. Soluble hCdc6p was also detected, but onlyduring S phase and G2/M. This observation possibly reflectsthe fact that a fraction of hCdc6p is rapidly translocated to thecytosol after the G1/S transition, as has been proposed on thebasis of immunostaining studies (26, 46, 52). The levels ofsoluble hCdc6p in this experiment seem higher than thosedetected in asynchronous cells. This can be explained becausethe later-elutriated fractions are enriched in late S and G2cells, which contain soluble hCdc6p, whereas in the asynchro-nous population, most cells are in G1 or early S phase.
In contrast to the case for hOrc2p and hCdc6p, the associ-ation of hMcm3p, hMcm4p, and hMcm5p with chromatin was
tightly regulated during the cell cycle. These proteins weredetected on chromatin since early G1 and kept accumulatingduring G1 until the G1/S transition. All of them were thenreleased as cells progressed through S phase. Some hMcm4pwas detected in chromatin in the last elutriated fraction, sug-gesting that hMcm4p could reassociate with chromatin slightlybefore hMcm3p and hMcm5p, as has been reported recentlyfor Xenopus (36). Very similar results were obtained with elu-triated HeLa and Manca cells (not shown). These observationsmatch other reports on the dynamics of chromatin associationof hMcm proteins (11, 16, 21, 31, 51, 60) and further supportthe validity of the biochemical fractionation protocol that weused.
hCdc6p can be polyubiquitinated in vivo and is stabilized bythe presence of proteasome inhibitors. Considering the cellcycle fluctuation of hCdc6p levels described above, we haveanalyzed the possibility that hCdc6p could be a substrate forubiquitin-mediated proteolysis. The mitotic degradation ofhCdc6p via the 25S proteasome would explain why the proteinis more abundant in G2 cells than in early G1.
Two types of experiments were done to address this issue.First, it was determined whether the inhibition of the protea-some with specific inhibitors would increase the steady-statelevels of hCdc6p in proliferating cells. Incubation of HeLa cellswith a variety of proteasome inhibitors (LLnL, MG132, b-lac-
FIG. 3. Protein levels and chromatin association of hCdc6p, hOrc2p, and hMcm proteins during the cell cycle. An asynchronous culture of human Raji cells wassubjected to centrifugal elutriation to isolate cells at different points of the cell cycle. The DNA content for the cells in each fraction is shown. (A) Percentage of cellsin each phase of the cell cycle, estimated with the CellFIT computer program. (B) Levels of initiator factors in total cell extracts (TCE). Equivalent amounts of eachtotal cell extract (normalized by cell number) were subjected to SDS-PAGE and transferred to nitrocellulose for immunobloting with the indicated antibodies. Theconcentration of cyclin A is also shown as a control of cell cycle progression. Lane A, asynchronous cells. (C) Cell cycle-regulated chromatin association of hOrc2p,hCdc6p, and hMcm proteins. Cells at different points in the cell cycle were subjected to the biochemical fractionation described for Fig. 2A. Immunoblots of the solubleprotein fraction (S2) or chromatin-enriched fraction (P3) are shown.
8606 MENDEZ AND STILLMAN MOL. CELL. BIOL.
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
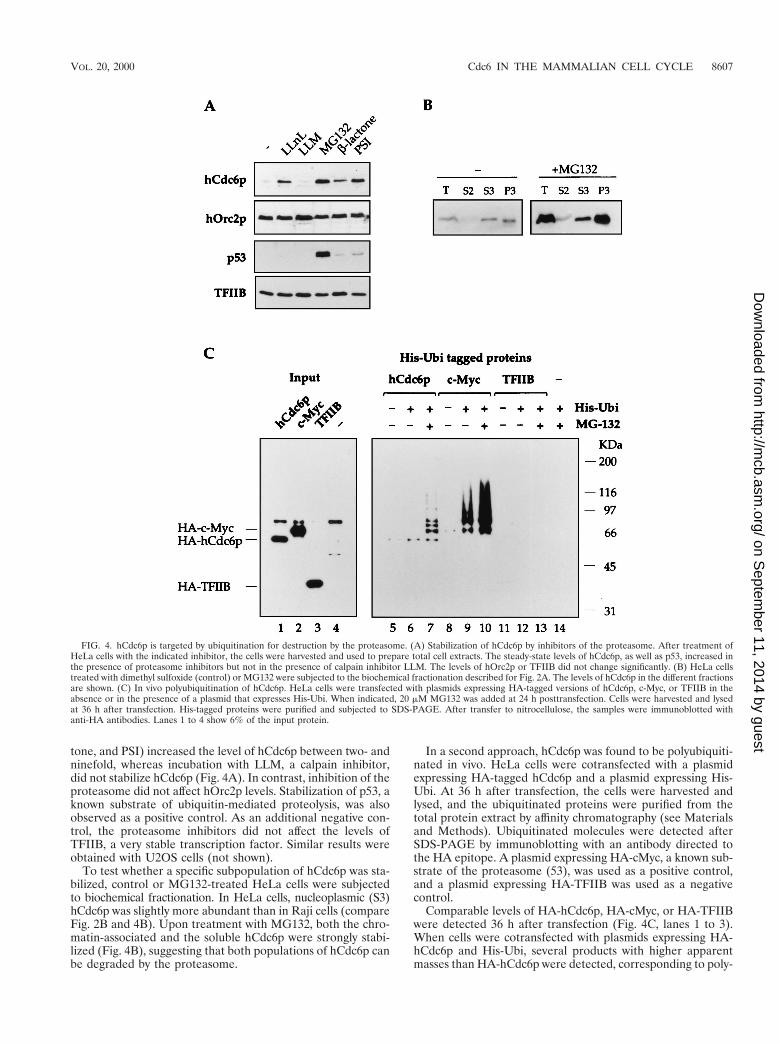
tone, and PSI) increased the level of hCdc6p between two- andninefold, whereas incubation with LLM, a calpain inhibitor,did not stabilize hCdc6p (Fig. 4A). In contrast, inhibition of theproteasome did not affect hOrc2p levels. Stabilization of p53, aknown substrate of ubiquitin-mediated proteolysis, was alsoobserved as a positive control. As an additional negative con-trol, the proteasome inhibitors did not affect the levels ofTFIIB, a very stable transcription factor. Similar results wereobtained with U2OS cells (not shown).
To test whether a specific subpopulation of hCdc6p was sta-bilized, control or MG132-treated HeLa cells were subjectedto biochemical fractionation. In HeLa cells, nucleoplasmic (S3)hCdc6p was slightly more abundant than in Raji cells (compareFig. 2B and 4B). Upon treatment with MG132, both the chro-matin-associated and the soluble hCdc6p were strongly stabi-lized (Fig. 4B), suggesting that both populations of hCdc6p canbe degraded by the proteasome.
In a second approach, hCdc6p was found to be polyubiquiti-nated in vivo. HeLa cells were cotransfected with a plasmidexpressing HA-tagged hCdc6p and a plasmid expressing His-Ubi. At 36 h after transfection, the cells were harvested andlysed, and the ubiquitinated proteins were purified from thetotal protein extract by affinity chromatography (see Materialsand Methods). Ubiquitinated molecules were detected afterSDS-PAGE by immunoblotting with an antibody directed tothe HA epitope. A plasmid expressing HA-cMyc, a known sub-strate of the proteasome (53), was used as a positive control,and a plasmid expressing HA-TFIIB was used as a negativecontrol.
Comparable levels of HA-hCdc6p, HA-cMyc, or HA-TFIIBwere detected 36 h after transfection (Fig. 4C, lanes 1 to 3).When cells were cotransfected with plasmids expressing HA-hCdc6p and His-Ubi, several products with higher apparentmasses than HA-hCdc6p were detected, corresponding to poly-
FIG. 4. hCdc6p is targeted by ubiquitination for destruction by the proteasome. (A) Stabilization of hCdc6p by inhibitors of the proteasome. After treatment ofHeLa cells with the indicated inhibitor, the cells were harvested and used to prepare total cell extracts. The steady-state levels of hCdc6p, as well as p53, increased inthe presence of proteasome inhibitors but not in the presence of calpain inhibitor LLM. The levels of hOrc2p or TFIIB did not change significantly. (B) HeLa cellstreated with dimethyl sulfoxide (control) or MG132 were subjected to the biochemical fractionation described for Fig. 2A. The levels of hCdc6p in the different fractionsare shown. (C) In vivo polyubiquitination of hCdc6p. HeLa cells were transfected with plasmids expressing HA-tagged versions of hCdc6p, c-Myc, or TFIIB in theabsence or in the presence of a plasmid that expresses His-Ubi. When indicated, 20 mM MG132 was added at 24 h posttransfection. Cells were harvested and lysedat 36 h after transfection. His-tagged proteins were purified and subjected to SDS-PAGE. After transfer to nitrocellulose, the samples were immunoblotted withanti-HA antibodies. Lanes 1 to 4 show 6% of the input protein.
VOL. 20, 2000 Cdc6 IN THE MAMMALIAN CELL CYCLE 8607
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
ubiquitinated forms of hCdc6p (lane 7). These products weredetected only in the presence of His-Ubi and the proteasomeinhibitor MG132 (compare lanes 5 to 7). The requirement forMG132 may reflect the extremely short half-life of hCdc6ponce it has been polyubiquitinated. Lanes 8 to 10 show thepolyubiquitination of c-Myc, which can be detected even in theabsence of MG132. On the other hand, TFIIB was not ubiqui-tinated under these conditions (lanes 11 to 13).
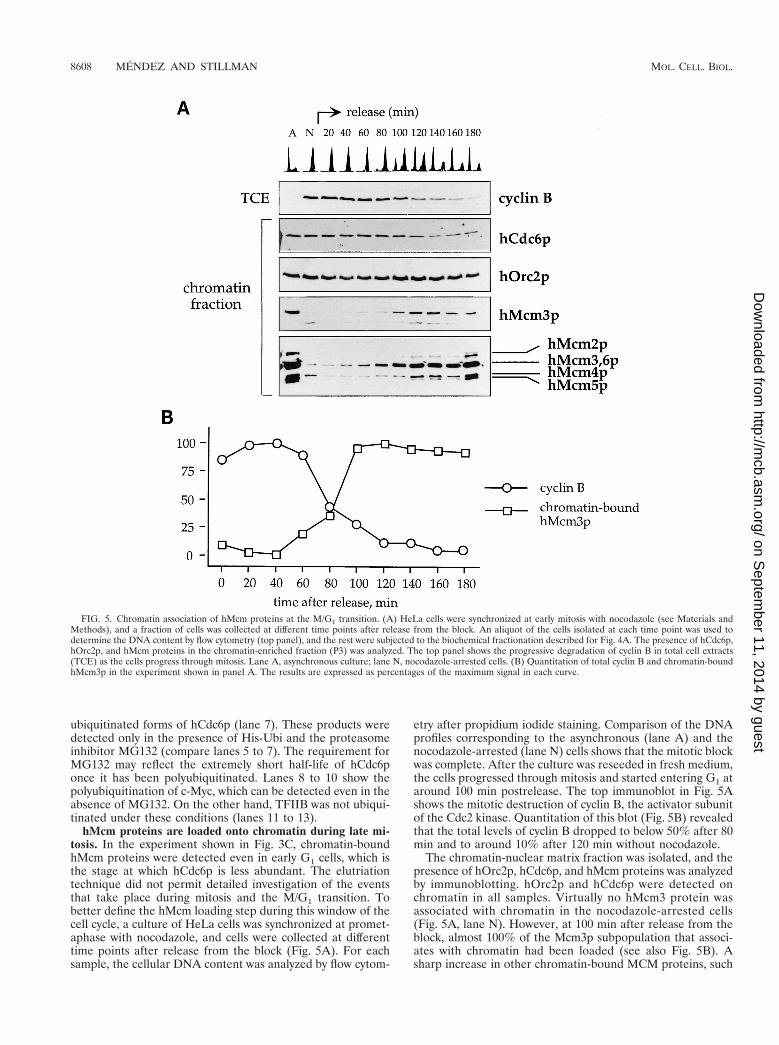
hMcm proteins are loaded onto chromatin during late mi-tosis. In the experiment shown in Fig. 3C, chromatin-boundhMcm proteins were detected even in early G1 cells, which isthe stage at which hCdc6p is less abundant. The elutriationtechnique did not permit detailed investigation of the eventsthat take place during mitosis and the M/G1 transition. Tobetter define the hMcm loading step during this window of thecell cycle, a culture of HeLa cells was synchronized at promet-aphase with nocodazole, and cells were collected at differenttime points after release from the block (Fig. 5A). For eachsample, the cellular DNA content was analyzed by flow cytom-
etry after propidium iodide staining. Comparison of the DNAprofiles corresponding to the asynchronous (lane A) and thenocodazole-arrested (lane N) cells shows that the mitotic blockwas complete. After the culture was reseeded in fresh medium,the cells progressed through mitosis and started entering G1 ataround 100 min postrelease. The top immunoblot in Fig. 5Ashows the mitotic destruction of cyclin B, the activator subunitof the Cdc2 kinase. Quantitation of this blot (Fig. 5B) revealedthat the total levels of cyclin B dropped to below 50% after 80min and to around 10% after 120 min without nocodazole.
The chromatin-nuclear matrix fraction was isolated, and thepresence of hOrc2p, hCdc6p, and hMcm proteins was analyzedby immunoblotting. hOrc2p and hCdc6p were detected onchromatin in all samples. Virtually no hMcm3 protein wasassociated with chromatin in the nocodazole-arrested cells(Fig. 5A, lane N). However, at 100 min after release from theblock, almost 100% of the Mcm3p subpopulation that associ-ates with chromatin had been loaded (see also Fig. 5B). Asharp increase in other chromatin-bound MCM proteins, such
FIG. 5. Chromatin association of hMcm proteins at the M/G1 transition. (A) HeLa cells were synchronized at early mitosis with nocodazole (see Materials andMethods), and a fraction of cells was collected at different time points after release from the block. An aliquot of the cells isolated at each time point was used todetermine the DNA content by flow cytometry (top panel), and the rest were subjected to the biochemical fractionation described for Fig. 4A. The presence of hCdc6p,hOrc2p, and hMcm proteins in the chromatin-enriched fraction (P3) was analyzed. The top panel shows the progressive degradation of cyclin B in total cell extracts(TCE) as the cells progress through mitosis. Lane A, asynchronous culture; lane N, nocodazole-arrested cells. (B) Quantitation of total cyclin B and chromatin-boundhMcm3p in the experiment shown in panel A. The results are expressed as percentages of the maximum signal in each curve.
8608 MENDEZ AND STILLMAN MOL. CELL. BIOL.
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
as hMcm2p and hMcm5p, was detected around the same timewith anti-pan-Mcm, an antibody that cross-reacts with differentmembers of the MCM protein family. This result has twoimplications: (i) despite the mitotic destruction of the bulk ofcellular hCdc6p, a fraction of it remained stable on chromatinduring mitosis, and (ii) even though hOrc2p and hCdc6p arepresent on chromatin during early mitosis, the bulk of hMcmproteins were not loaded until a later stage, concomitant withthe destruction of cyclin B.
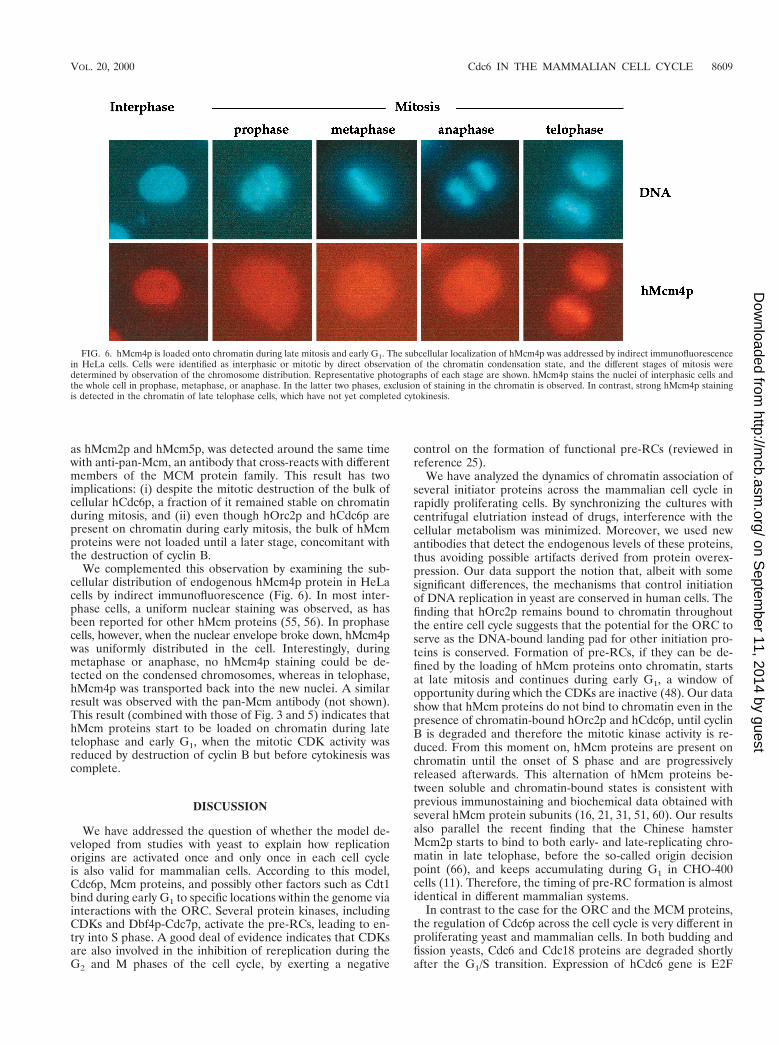
We complemented this observation by examining the sub-cellular distribution of endogenous hMcm4p protein in HeLacells by indirect immunofluorescence (Fig. 6). In most inter-phase cells, a uniform nuclear staining was observed, as hasbeen reported for other hMcm proteins (55, 56). In prophasecells, however, when the nuclear envelope broke down, hMcm4pwas uniformly distributed in the cell. Interestingly, duringmetaphase or anaphase, no hMcm4p staining could be de-tected on the condensed chromosomes, whereas in telophase,hMcm4p was transported back into the new nuclei. A similarresult was observed with the pan-Mcm antibody (not shown).This result (combined with those of Fig. 3 and 5) indicates thathMcm proteins start to be loaded on chromatin during latetelophase and early G1, when the mitotic CDK activity wasreduced by destruction of cyclin B but before cytokinesis wascomplete.
DISCUSSION
We have addressed the question of whether the model de-veloped from studies with yeast to explain how replicationorigins are activated once and only once in each cell cycleis also valid for mammalian cells. According to this model,Cdc6p, Mcm proteins, and possibly other factors such as Cdt1bind during early G1 to specific locations within the genome viainteractions with the ORC. Several protein kinases, includingCDKs and Dbf4p-Cdc7p, activate the pre-RCs, leading to en-try into S phase. A good deal of evidence indicates that CDKsare also involved in the inhibition of rereplication during theG2 and M phases of the cell cycle, by exerting a negative
control on the formation of functional pre-RCs (reviewed inreference 25).
We have analyzed the dynamics of chromatin association ofseveral initiator proteins across the mammalian cell cycle inrapidly proliferating cells. By synchronizing the cultures withcentrifugal elutriation instead of drugs, interference with thecellular metabolism was minimized. Moreover, we used newantibodies that detect the endogenous levels of these proteins,thus avoiding possible artifacts derived from protein overex-pression. Our data support the notion that, albeit with somesignificant differences, the mechanisms that control initiationof DNA replication in yeast are conserved in human cells. Thefinding that hOrc2p remains bound to chromatin throughoutthe entire cell cycle suggests that the potential for the ORC toserve as the DNA-bound landing pad for other initiation pro-teins is conserved. Formation of pre-RCs, if they can be de-fined by the loading of hMcm proteins onto chromatin, startsat late mitosis and continues during early G1, a window ofopportunity during which the CDKs are inactive (48). Our datashow that hMcm proteins do not bind to chromatin even in thepresence of chromatin-bound hOrc2p and hCdc6p, until cyclinB is degraded and therefore the mitotic kinase activity is re-duced. From this moment on, hMcm proteins are present onchromatin until the onset of S phase and are progressivelyreleased afterwards. This alternation of hMcm proteins be-tween soluble and chromatin-bound states is consistent withprevious immunostaining and biochemical data obtained withseveral hMcm protein subunits (16, 21, 31, 51, 60). Our resultsalso parallel the recent finding that the Chinese hamsterMcm2p starts to bind to both early- and late-replicating chro-matin in late telophase, before the so-called origin decisionpoint (66), and keeps accumulating during G1 in CHO-400cells (11). Therefore, the timing of pre-RC formation is almostidentical in different mammalian systems.
In contrast to the case for the ORC and the MCM proteins,the regulation of Cdc6p across the cell cycle is very different inproliferating yeast and mammalian cells. In both budding andfission yeasts, Cdc6 and Cdc18 proteins are degraded shortlyafter the G1/S transition. Expression of hCdc6 gene is E2F
FIG. 6. hMcm4p is loaded onto chromatin during late mitosis and early G1. The subcellular localization of hMcm4p was addressed by indirect immunofluorescencein HeLa cells. Cells were identified as interphasic or mitotic by direct observation of the chromatin condensation state, and the different stages of mitosis weredetermined by observation of the chromosome distribution. Representative photographs of each stage are shown. hMcm4p stains the nuclei of interphasic cells andthe whole cell in prophase, metaphase, or anaphase. In the latter two phases, exclusion of staining in the chromatin is observed. In contrast, strong hMcm4p stainingis detected in the chromatin of late telophase cells, which have not yet completed cytokinesis.
VOL. 20, 2000 Cdc6 IN THE MAMMALIAN CELL CYCLE 8609
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
regulated (67), and the levels of hCdc6 mRNA peak at theG1/S transition (65). HCdc6p levels, however, are fairly con-stant in the cell cycle, with the exception of early G1 cells, inwhich the hCdc6p concentration is much lower. Therefore,most hCdc6p might be degraded during mitosis. We found thathCdc6p can indeed be polyubiquitinated in vivo and that inhi-bition of the proteasome results in increased steady-state levelsof hCdc6p. These data combined indicate that hCdc6p is abona fide substrate for ubiquitin-mediated proteolysis. Theregion of hCdc6p that targets it for degradation by the protea-some has been recently mapped to the N terminus of theprotein (B. O. Petersen, C. Wagener, M. Melixetian, F. Mari-nori, E. L. Denchi, C. Matteucci, and K. Helin, personal com-munication).
Our study also addresses the issue of hCdc6p subcellularlocalization. Some groups have concluded that the protein ismostly nuclear in G1 but is translocated to the cytosol after theG1/S transition (26, 46, 52). This conclusion is mostly sup-ported by experiments in which epitope-tagged versions ofhCdc6p were introduced in human cells and the overexpressedproteins were detected by indirect immunofluorescence. Thenuclear-cytoplasmic translocation seems to correlate with thephosphorylation of hCdc6p by cyclin A-Cdk2 (26, 46) anddepends on exportin-1 (26). On the other hand, other reportsindicate that endogenous hCdc6p is mostly nuclear (64, 65)and is actually associated with a nuclear structure (17).
Our present analysis with elutriated Raji cells revealed thata significant fraction of endogenous hCdc6p remained associ-
ated with chromatin from G1 until mitosis. It should be notedthat this is compatible with the previously published observa-tions, because the immunostaining technique cannot rule outthe possibility that some hCdc6p remains in the nuclei duringS and G2. Some soluble hCdc6p was detected after cells enterS phase, in agreement with the translocation model. Coverleyet al. (8) recently reported a series of experiments performedwith a cell-free system in which DNA replication was inducedin isolated 3T3 cell nuclei by incubation with cytosolic extractsfrom the same cells in the presence of defined amounts ofexogenous XCdc6p. The main conclusion from this study wasthat chromatin-bound Cdc6p persists in S and G2, while solu-ble Cdc6p is destroyed in a cyclin A-Cdk2 dependent process.This report is consistent with our own findings with endoge-nous hCdc6p and indicates that proteolysis of hCdc6p is likelyanother regulatory step in the mammalian cell cycle.
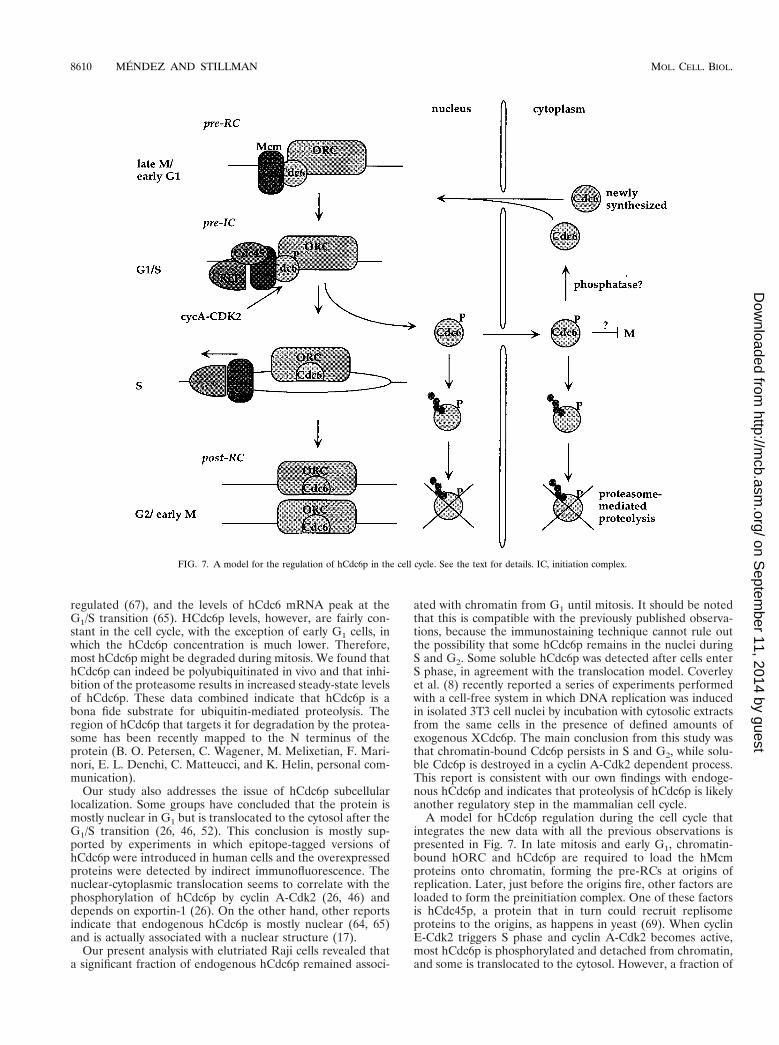
A model for hCdc6p regulation during the cell cycle thatintegrates the new data with all the previous observations ispresented in Fig. 7. In late mitosis and early G1, chromatin-bound hORC and hCdc6p are required to load the hMcmproteins onto chromatin, forming the pre-RCs at origins ofreplication. Later, just before the origins fire, other factors areloaded to form the preinitiation complex. One of these factorsis hCdc45p, a protein that in turn could recruit replisomeproteins to the origins, as happens in yeast (69). When cyclinE-Cdk2 triggers S phase and cyclin A-Cdk2 becomes active,most hCdc6p is phosphorylated and detached from chromatin,and some is translocated to the cytosol. However, a fraction of
FIG. 7. A model for the regulation of hCdc6p in the cell cycle. See the text for details. IC, initiation complex.
8610 MENDEZ AND STILLMAN MOL. CELL. BIOL.
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

hCdc6p remains associated with chromatin, perhaps because itis not accessible to the CDKs. A specific cellular mechanismmight contribute to keep a fraction of hypophosphorylatedhCdc6p, because hCdc6p has been found to associate with anovel regulatory subunit of protein phosphatase 2A (68). Nu-cleoplasmic and cytosolic hCdc6p are degraded by ubiquitin-mediated proteolysis.
Whether there is a role for cytosolic hCdc6p remains un-known, but it could send a signal that prevents mitosis until itis degraded by the 26S proteasome. The possibility of Cdc6pcoordinating DNA replication and mitosis has been previouslysuggested for yeast (4, 28, 47), although the molecular mech-anism of this mitotic inhibition has not been elucidated.
In our model, the post-RC in human cells would contain thehORC as well as hCdc6p. But interestingly, this chromatin-bound pool of hCdc6p did not recruit hMcm proteins ontochromatin during G2 phase, or during mitosis until cyclin B wasdegraded. Therefore, the negative control exerted by activeCDKs on the formation of pre-RCs seems to be conserved.Consistent with this idea, inhibition of the CDKs permitted theMCM proteins to reload onto chromatin when mammalian G2nuclei were incubated in S-phase Xenopus extracts (7). It isnow clear that multiple mechanisms coexist to ensure the in-hibition of pre-RC formation after S phase. These additionalmechanisms likely include the regulated instability of the larg-est subunit of the hORC (41; H. Zou-Yang, H., J. Mendez, M.Hidaka, and B. Stillman, unpublished data) and geminin, aprotein first identified in Xenopus that is destroyed as cellsprogress through mitosis (38).
What is the execution point of human Cdc6p? Based on theevidence accumulated with yeast and Xenopus, it seems rea-sonable to assume that hCdc6p will be responsible for therecruitment of hMcm proteins onto ORC-bound chromatin atthe M/G1 transition. However, if the sole function of hCdc6pwas to load Mcm proteins onto pre-RCs during the M/G1transition, it would be difficult to explain why phosphorylationof hCdc6p by cyclin A-Cdk2 at the G1/S transition seems to beessential for entry into S phase (26). This conclusion is basedon the observation that overexpression of a nonphosphorylat-able form of hCdc6p (bearing the triple mutation S54A/S74A/S114A) inhibits cellular DNA replication. This result is differ-ent from that of Petersen et al. (46), where no significant defectin DNA replication was observed after transfection or micro-injection of the same hCdc6p mutant derivative. Interestingly,microinjection of mutant versions of hCdc6p which impairATP binding and/or hydrolysis also results in G1/S arrest (20),reinforcing the hypothesis that hCdc6p has a function at theG1/S-phase transition. One simple explanation that we favor isthat hCdc6p is required to load MCM proteins onto chromatinin late G1 phase, particularly in cells that are entering into theproliferation cycle from G0 phase or in cells that have anextended G1 phase. This situation would resemble that in S.cerevisiae cells undergoing a prolonged G1 phase (e.g., aftertreatment with a-factor), where there is a new peak of Cdc6pexpression just before the G1/S transition. Therefore, we pro-pose that hCdc6p is essential and rate limiting for initiation ofDNA replication, as has been observed (35, 58)
Is there another function for the chromatin-bound hCdc6pduring S phase and G2? Very recent data obtained with yeastindicate that Cdc6p cooperates with the ORC to determineorigin specificity, in an ATP-dependent manner (39). It istempting to speculate that the chromatin-bound form ofhCdc6p could cooperate with the hORC to bind to specificreplicator sequences. As the nature of such sequences inhigher eukaryotes remains unknown, this may be an excitingtopic for future research.
ACKNOWLEDGMENTS
We thank W. Tansey for his help with the proteasome inhibition andin vivo ubiquitination experiments, R. S. Williams for providing puri-fied recombinant hCdc6p, C. Bautista for her work at the Cold SpringHarbor Laboratory monoclonal antibody facility, K. Brown for herhelp with epitope mapping of the anti-hCdc6p antibodies, A. Koff forhis help with the elutriation protocol, N. Hernandez for anti-TFIIBantibodies, D. Bohmann for plasmid pMT107, A. Verreault for hissuggestions on the fractionation protocol, K. Cronin for excellent tech-nical assistance, and A. Losada and J. Chong for useful comments onthe manuscript.
This work was supported by the National Cancer Institute (grantCA13106). J.M. was the recipient of postdoctoral fellowships fromFundacion Ramon Areces (Spain) and the Human Frontier ScienceProgram Organization.
REFERENCES1. Aparicio, O. M., D. M. Weinstein, and S. P. Bell. 1997. Components and
dynamics of DNA replication complexes in S. cerevisiae: redistribution ofMCM proteins and Cdc45p during S phase. Cell 91:59–69.
2. Bell, S. P., and B. Stillman. 1992. ATP-dependent recognition of eukaryoticorigins of DNA replication by a multiprotein complex. Nature 357:128–134.
3. Bolt, M. W., and P. A. Mahoney. 1997. High-efficiency blotting of proteins ofdiverse sizes following sodium dodecyl sulfate-polyacrylamide gel electro-phoresis. Anal. Biochem. 247:185–192.
4. Bueno, A., and P. Russell. 1992. Dual functions of CDC6: a yeast proteinrequired for DNA replication also inhibits nuclear division. EMBO J. 11:2167–2176.
5. Chong, J. P., M. K. Hayashi, M. N. Simon, R. M. Xu, and B. Stillman. 2000.A double-hexamer archaeal minichromosome maintenance protein is anATP-dependent DNA helicase. Proc. Natl. Acad. Sci. USA 97:1530–1535.
6. Coleman, T. R., P. B. Carpenter, and W. G. Dunphy. 1996. The XenopusCdc6 protein is essential for the initiation of a single round of DNA repli-cation in cell-free extracts. Cell 87:53–63.
7. Coverley, D., H. R. Wilkinson, M. A. Madine, A. D. Mills, and R. A. Laskey.1998. Protein kinase inhibition in G2 causes mammalian Mcm proteins toreassociate with chromatin and restores ability to replicate. Exp. Cell Res.238:63–69.
8. Coverley, D., C. Pelizon, S. Trewick, and R. A. Laskey. 2000. Chromatin-bound Cdc6 persists in S and G2 phases in human cells, while soluble Cdc6is destroyed in a cyclin A-Cdk2 dependent process. J. Cell Sci. 113:1929–1938.
9. Dahmann, C., J. F. Diffley, and K. A. Nasmyth. 1995. S-phase-promotingcyclin-dependent kinases prevent re-replication by inhibiting the transitionof replication origins to a pre-replicative state. Curr. Biol. 5:1257–1269.
10. Diffley, J. F., J. H. Cocker, S. J. Dowell, and A. Rowley. 1994. Two steps inthe assembly of complexes at yeast replication origins in vivo. Cell 78:303–316.
11. Dimitrova, D. S., I. T. Todorov, T. Melendy, and D. M. Gilbert. 1999. Mcm2,but not RPA, is a component of the mammalian early G1-phase prereplica-tion complex. J. Cell Biol. 146:709–722.
12. Donovan, S., J. Harwood, L. S. Drury, and J. F. Diffley. 1997. Cdc6p-dependent loading of Mcm proteins onto pre-replicative chromatin in bud-ding yeast. Proc. Natl. Acad. Sci. USA 94:5611–5616.
13. Drury, L. S., G. Perkins, and J. F. Diffley. 1997. The Cdc4/34/53 pathwaytargets Cdc6p for proteolysis in budding yeast. EMBO J. 16:5966–5976.
14. Drury, L. S., G. Perkins, and J. F. Diffley. 2000. The cyclin-dependent kinaseCdc28p regulates distinct modes of Cdc6p proteolysis during the buddingyeast cell cycle. Curr. Biol. 10:231–240.
15. Dutta, A., and S. P. Bell. 1997. Initiation of DNA replication in eukaryoticcells. Annu. Rev. Cell Dev. Biol. 13:293–332.
16. Fujita, M., T. Kiyono, Y. Hayashi, and M. Ishibashi. 1996. hCDC47, ahuman member of the MCM family. Dissociation of the nucleus-bound formduring S phase. J. Biol. Chem. 271:4349–4354.
17. Fujita, M., C. Yamada, H. Goto, N. Yokoyama, K. Kuzushima, M. Inagaki,and T. Tsurumi. 1999. Cell cycle regulation of human CDC6 protein. Intra-cellular localization, interaction with the human MCM complex, and CDC2kinase-mediated hyperphosphorylation. J. Biol. Chem. 274:25927–25932.
18. Gavin, K. A., M. Hidaka, and B. Stillman. 1995. Conserved initiator proteinsin eukaryotes. Science 270:1667–1671.
19. Harlow, E., and D. Lane. 1998. Using antibodies: a laboratory manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
20. Herbig, U., C. A. Marlar, and E. Fanning. 1999. The Cdc6 nucleotide-binding site regulates its activity in DNA replication in human cells. Mol.Biol. Cell 10:2631–2645.
21. Holthoff, H. P., M. Baack, A. Richter, M. Ritzi, and R. Knippers. 1998.Human protein MCM6 on HeLa cell chromatin. J. Biol. Chem. 273:7320–7325.
22. Ishimi, Y. 1997. A DNA helicase activity is associated with an MCM4, -6, and-7 protein complex. J. Biol. Chem. 272:24508–24513.
VOL. 20, 2000 Cdc6 IN THE MAMMALIAN CELL CYCLE 8611
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

23. Jallepalli, P. V. and T. J. Kelly. 1996. Rum1 and Cdc18 link inhibition ofcyclin-dependent kinase to the initiation of DNA replication in Schizosac-charomyces pombe. Genes Dev. 10:541–552.
24. Jallepalli, P. V., G. W. Brown, M. Muzi-Falconi, D. Tien, and T. J. Kelly.1997. Regulation of the replication initiator protein p65cdc18 by CDK phos-phorylation. Genes Dev. 11:2767–2779.
25. Jallepalli, P. V., and T. J. Kelly. 1997. Cyclin-dependent kinase and initiationat eukaryotic origins: a replication switch? Curr. Opin. Cell Biol. 9:358–363.
26. Jiang, W., N. J. Wells, and T. Hunter. 1999. Multistep regulation of DNAreplication by Cdk phosphorylation of HsCdc6. Proc. Natl. Acad. Sci. USA96:6193–6198.
27. Kearsey, S. E., and K. Labib. 1998. MCM proteins: evolution, properties,and role in DNA replication. Biochim. Biophys. Acta 1398:113–136.
28. Kelly, T. J., G. S. Martin, S. L. Forsburg, R. J. Stephen, A. Russo, and P.Nurse. 1993. The fission yeast cdc181 gene product couples S phase toSTART and mitosis. Cell 74:371–382.
29. Kelman, Z., J. K. Lee, and J. Hurwitz. 1999. The single minichromosomemaintenance protein of Methanobacterium thermoautotrophicum DeltaHcontains DNA helicase activity. Proc. Natl. Acad. Sci. USA 96:14783–14788.
30. Krek, W., and J. A. DeCaprio. 1995. Cell synchronization. Methods Enzymol.254:114–124.
31. Krude, T., C. Musahl, R. A. Laskey, and R. Knippers. 1996. Human repli-cation proteins hCdc21, hCdc46 and P1Mcm3 bind chromatin uniformlybefore S-phase and are displaced locally during DNA replication. J. Cell Sci.109:309–318.
32. Labib, K., J. A. Tercero, and J. F. Diffley. 2000. Uninterrupted MCM2–7function required for DNA replication fork progression. Science 288:1643–1647.
33. Liang, C., M. Weinreich, and B. Stillman. 1995. ORC and Cdc6p interactand determine the frequency of initiation of DNA replication in the genome.Cell 81:667–676.
34. Liang, C., and B. Stillman. 1997. Persistent initiation of DNA replicationand chromatin-bound MCM proteins during the cell cycle in cdc6 mutants.Genes Dev. 11:3375–3386.
35. Madine, M. A., M. Swietlik, C. Pelizon, P. Romanowski, A. D. Mills, andR. A. Laskey. 2000. The roles of the MCM, ORC, and cdc6 proteins indetermining the replication competence of chromatin in quiescent cells. J.Struct. Biol. 129:198–210.
36. Maiorano, D., J. M. Lemaıtre, and M. Mechali. 2000. Stepwise regulatedchromatin assembly of MCM2–7 proteins. J. Biol. Chem. 275:8426–8431.
37. Maiorano, D., J. Moreau, and M. Mechali. 2000. XCDT1 is required for theassembly of pre-replicative complexes in Xenopus laevis. Nature 404:622–625.
38. McGarry, T. J., and M. W. Kirschner. 1998. Geminin, an inhibitor of DNAreplication, is degraded during mitosis. Cell 93:1043–1053.
39. Mizushima, T., N. Takahashi, and B. Stillman. 2000. Cdc6p modulates thestructure and DNA binding activity of the origin recognition complex invitro. Genes Dev. 14:1631–1641.
40. Muzi-Falconi, M., G. W. Brown, and T. J. Kelly. 1996. cdc181 regulatesinitiation of DNA replication in Schizosaccharomyces pombe. Proc. Natl.Acad. Sci. USA 93:1566–1570.
41. Natale, D. A., C.-J. Li, W.-H. Sun, and M. L. DePamphilis. 2000. Selectiveinstability of Orc1 protein accounts for the absence of functional originrecognition complexes during the M-G1 transition in mammals. EMBO J.19:2728–2738.
42. Neuwald, A. F., L. Aravind, J. L. Spouge, and E. V. Koonin. 1999. AAA1: aclass of chaperone-like ATPases associated with the assembly, operation,and disassembly of protein complexes. Genome Res. 9:27–43.
43. Nishitani, H., and P. Nurse. 1995. p65cdc18 plays a major role controlling theinitiation of DNA replication in fission yeast. Cell 83:397–405.
44. Nishitani, H., Z. Lygerou, T. Nishimoto, and P. Nurse. 2000. The Cdt1protein is required to license DNA for replication in fission yeast. Nature404:625–628.
45. Perkins, G., and J. F. Diffley. 1998. Nucleotide-dependent prereplicativecomplex assembly by Cdc6p, a homolog of eukaryotic and prokaryotic clamp-loaders. Mol. Cell 2:23–32.
46. Petersen, B. O., J. Lukas, C. S. Sorensen, J. Bartek, and K. Helin. 1999.Phosphorylation of mammalian CDC6 by cyclin A/CDK2 regulates its sub-cellular localization. EMBO J. 18:396–410.
47. Piatti, S., C. Lengauer, and K. Nasmyth. 1995. Cdc6 is an unstable proteinwhose de novo synthesis in G1 is important for the onset of S phase and forpreventing a ‘reductional’ anaphase in the budding yeast Saccharomyces
cerevisiae. EMBO J. 14:3788–3799.48. Piatti, S., T. Bohm, J. H. Cocker, J. F. Diffley, and K. Nasmyth. 1996.
Activation of S-phase-promoting CDKs in late G1 defines a “point of noreturn” after which Cdc6 synthesis cannot promote DNA replication in yeast.Genes Dev. 10:1516–1531.
49. Quintana, D. G., Z. Hou, K. C. Thome, M. Hendricks, P. Saha, and A. Dutta.1997. Identification of HsORC4, a member of the human origin of replica-tion recognition complex. J. Biol. Chem. 272:28247–28251.
50. Quintana, D. G., K. C. Thome, Z. H. Hou, A. H. Ligon, C. C. Morton, and A.Dutta. 1998. ORC5L, a new member of the human origin recognition com-plex, is deleted in uterine leiomyomas and malignant myeloid diseases.J. Biol. Chem. 273:27137–27145.
51. Ritzi, M., M. Baack, C. Musahl, P. Romanowski, R. A. Laskey, and R.Knippers. 1998. Human minichromosome maintenance proteins and humanorigin recognition complex 2 protein on chromatin. J. Biol. Chem. 273:24543–24549.
52. Saha, P., J. Chen, K. C. Thome, S. J. Lawlis, Z. H. Hou, M. Hendricks, J. D.Parvin, and A. Dutta. 1998. Human CDC6/Cdc18 associates with Orc1 andcyclin-cdk and is selectively eliminated from the nucleus at the onset of Sphase. Mol. Cell. Biol. 18:2758–2767.
53. Salghetti, S. E., S. Y. Kim, and W. P. Tansey. 1999. Destruction of Myc byubiquitin-mediated proteolysis: cancer-associated and transforming muta-tions stabilize Myc. EMBO J. 18:717–726.
54. Sanchez, M., A. Calzada, and A. Bueno. 1999. The Cdc6 protein is ubiqui-tinated in vivo for proteolysis in Saccharomyces cerevisiae. J. Biol. Chem.274:9092–9097.
55. Schulte, D., R. Burkhart, C. Musahl, B. Hu, C. Schlatterer, H. Hameister,and R. Knippers. 1995. Expression, phosphorylation and nuclear localizationof the human P1 protein, a homologue of the yeast Mcm 3 replicationprotein. J. Cell Sci. 108:1381–1389.
56. Schulte, D., A. Richter, R. Burkhart, C. Musahl, and R. Knippers. 1996.Properties of the human nuclear protein p85Mcm. Expression, nuclear local-ization and interaction with other Mcm proteins. Eur. J. Biochem. 235:144–151.
57. Stillman, B. 1996. Cell cycle control of DNA replication. Science 274:1659–1664.
58. Stoeber, K., A. D. Mills, Y. Kubota, T. Krude, P. Romanowski, K. Marhei-neke, R. A. Laskey, and G. H. Williams. 1998. Cdc6 protein causes prema-ture entry into S phase in a mammalian cell-free system. EMBO J. 17:7219–7229.
59. Tanaka, T., D. Knapp, and K. Nasmyth. 1997. Loading of an Mcm proteinonto DNA replication origins is regulated by Cdc6p and CDKs. Cell 90:649–660.
60. Todorov, I. T., A. Attaran, and S. E. Kearsey. 1995. BM28, a human memberof the MCM2–3-5 family, is displaced from chromatin during DNA replica-tion. J. Cell Biol. 129:1433–1445.
61. Trier, M., L. M. Staszewski, and D. Bohmann. 1994. Ubiquitin-dependentc-Jun degradation in vivo is mediated by the delta domain. Cell 78:787–798.
62. Tugal, T., X. H. Zou-Yang, K. Gavin, D. Pappin, B. Canas, R. Kobayashi, T.Hunt, and B. Stillman. 1998. The Orc4p and Orc5p subunits of the Xenopusand human origin recognition complex are related to Orc1p and Cdc6p.J. Biol. Chem. 273:32421–32429.
63. Weinreich, M., C. Liang, and B. Stillman. 1999. The Cdc6p nucleotide-binding motif is required for loading Mcm proteins onto chromatin. Proc.Natl. Acad. Sci. USA 96:441–446.
64. Williams, G. H., P. Romanowski, L. Morris, M. Madine, A. D. Mills, K.Stoeber, J. Marr, R. A. Laskey, and N. Coleman. 1998. Improved cervicalsmear assessment using antibodies against proteins that regulate DNA rep-lication. Proc. Natl. Acad. Sci. USA 95:14932–14937.
65. Williams, R. S., R. V. Shohet, and B. Stillman. 1997. A human proteinrelated to yeast Cdc6p. Proc. Natl. Acad. Sci. USA 94:142–147.
66. Wu, J. R., and D. M. Gilbert. 1996. A distinct G1 step required to specify theChinese hamster DHFR replication origin. Science 271:1270–1272.
67. Yan, Z., J. DeGregori, R. Shohet, G. Leone, B. Stillman, J. R. Nevins, andR. S. Williams. 1998. Cdc6 is regulated by E2F and is essential for DNAreplication in mammalian cells. Proc. Natl. Acad. Sci. USA 95:3603–3608.
68. Yan, Z., S. A. Fedorov, M. C. Mumby, and R. S. Williams. 2000. PR48, anovel regulatory subunit of protein phosphatase 2A, interacts with Cdc6 andmodulates DNA replication in human cells. Mol. Cell. Biol. 20:1021–1029.
69. Zou, L., and B. Stillman. 2000. Assembly of a complex containing Cdc45p,RPA, and Mcm2p at replication origins controlled by S-phase cyclin-depen-dent kinases and Cdc7p-Dbf4p kinase. Mol. Cell. Biol. 20:3086–3096.
8612 MENDEZ AND STILLMAN MOL. CELL. BIOL.
on Septem
ber 11, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from





























