Embed Size (px)
Citation preview

lable at ScienceDirect
Neuropharmacology 62 (2012) 1554e1563
Contents lists avai
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
Chronic phencyclidine (PCP)-induced modulation of muscarinic receptormRNAs in rat brain : Impact of antipsychotic drug treatment
Lucinda J. Steward a,*, Matthew D. Kennedy a, Brian J. Morris a,b, Judith A. Pratt a,c
a Psychiatric Research Institute of Neuroscience in Glasgow (PsYRING), University of Glasgow, Glasgow G12 8QQ, Scotland, UKb Institute of Neuroscience and Psychology, University of Glasgow, Glasgow G12 8QQ, Scotland, UKc Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, 161 Cathedral St, Glasgow G4ORE, Scotland, UK
a r t i c l e i n f o
Article history:Received 27 January 2011Received in revised form10 May 2011Accepted 17 May 2011
Keywords:M1 receptorM2 receptorM3 receptorSchizophreniaClozapine
* Corresponding author. Present address; F. Hoffmence Discovery, Grenzacherstrasse, Bldg. 070/326,Tel.: þ41 61 6889082; fax: þ41 61 6879566.
E-mail address: [email protected] (L.J. S
0028-3908/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.neuropharm.2011.05.016
a b s t r a c t
Many antipsychotics (APDs) have a high affinity for muscarinic receptors, which is thought to contributeto their therapeutic efficacy, or side effect profile. In order to define how muscarinic receptor geneexpression is affected by atypical or typical APDs, rats were treated with chronic (2.58 mg/kg) PCP(a psychotomimetic) or vehicle, plus clozapine (20 mg/kg/day) or haloperidol (1 mg/kg/day), and M1, M2and M3 receptor mRNA levels were determined in brain sections. Negligible changes in M2 or M3muscarinic mRNAwere detected in any region after clozapine or haloperidol. Chronic PCP administrationincreased M1 mRNA expression in the prefrontal cortex, which was not reversed by either chronicclozapine or haloperidol treatment. Chronic clozapine treatment in combination with PCP treatmentdecreased M1 receptor mRNA levels in the nucleus accumbens core, whereas chronic haloperidol incombination with PCP treatment increased M1 receptor mRNA levels in the ventromedial hypothalamusand medial amygdala. Thus M1 receptor gene expression is targeted by APDs, although the regionsaffected differ according to the APD treatment and whether PCP has been administered. The differentbrain circuitry modulated, may reflect the differing modes of action of typical and atypical APDs. Thesedata provide support for the dysregulation of M1 receptors in schizophrenia, and furthermore, modu-lation by antipsychotic agents in the treatment of schizophrenia.
This article is part of a Special Issue entitled ’Schizophrenia’.� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Schizophrenia is a complex heterogenous disease characterisedby a range of symptoms including positive symptoms such asdelusions and hallucinations, negative symptoms which describea poor ability to interact socially, avolition and anhedonia, andcognitive deficits. Current antipsychotic drug (APD) medication hasbeen categorised as being typical (1st generation) or atypical (2ndgeneration). Some atypical APDs such as clozapine and olanzapineand in fact some typical antipsychotics such as chlorpromazine andthioridazine, have high (nM) affinity for muscarinic receptors aswell as for dopaminergic and serotonergic receptors (Bymasteret al., 2003). As yet it is unclear whether their action at musca-rinic receptors plays a role in their APD therapeutic profile.
ann-La Roche Ltd, Neurosci-CH-4070 Basel, Switzerland.
teward).
All rights reserved.
The original dopamine hypothesis of schizophrenia proposedthat an enhanced mesolimbic dopamine system was key to thehallucinations and delusions in the disease. Current opinionsuggests that disease pathology may affect directly or indirectlymany neurotransmitter systems including the glutamatergic,serotonergic and cholinergic systems (Scarr and Dean, 2008; Hydeand Crook, 2001; Terry, 2008). The cholinergic system has beenassociated with both the treatment of schizophrenic symptoms andwith the amelioration of extrapyramidal-induced side effects (EPS)induced by some APDs. Moreover, it has been proposed that the useof anti-cholinergic medication may exacerbate positive symptomsduring an acute phase psychotic episode, but may be beneficial inreducing the negative symptoms of schizophrenia (Johnstone et al.,1983; Tandon et al., 1988). It is well documented that high doses ofmuscarinic antagonists such as scopolamine or atropine, or naturalplants such as belladonna (that contain scopolamine and atropine),can induce visual hallucinations characteristic of a psychoticepisode as well as cause cognitive impairment (reviewed in Perryand Perry, 1995; and Terry, 2008). Furthermore, the frequent useof drugs that have an antagonistic effect at muscarinic receptors in

L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e1563 1555
the treatment of Alzheimer’s disease and elderly patients, havebeen proposed to contribute to the genesis of psychotic symptomsin this patient population for which acetylcholine esterase inhibi-tors, as well as M1/M4 receptor agonists, have been shown toimprove this drug-induced psychosis as well enhance the cognitiveprofile (reviewed Cancelli et al., 2009; Conn et al., 2009).
In schizophrenia patients, it has been demonstrated that thereare decreases in muscarinic M1/M4 receptor levels in the caudateputamen, anterior cingulate cortex, sub temporal gyrus andhippocampus using the muscarinic (M1/M4) antagonist radio-ligand [3H]pirenzepine, suggesting that the muscarinic cholinergicsystem has been affected by the disease process, or the antipsy-chotic medication (Dean et al., 1996; Crook et al., 2000; Zavitsanouet al., 2004; Deng and Huang, 2005; Newell et al., 2007a). Deanet al. (2002), further demonstrated that not only was [3H]pir-enzepine binding decreased in dorsolateral prefrontal cortex (BA9)from patients with schizophrenia, but also, using a specific M1antibody as well as in-situ hybridisation, M1 receptor (M1R)protein and mRNA levels were decreased, whereas M4 receptor(M4R) protein was unchanged. More recently Salah-Uddin et al.(2009), in a schizophrenia patient cohort with reduced cortical[3H]pirenzapine binding, showed decreased muscarinic agonistpotency but increased efficacy in Gq/11 coupling in BA9 tissue using[35S]GTPgS binding, and proposed this was potentially an adaptiveresponse. Fewer studies have examined the M2 receptors (M2Rs)and M3 receptors (M3Rs) but Scarr et al. (2006), did not find anyalterations inM2RorM3R protein levels/function ormRNA levels incortical regions BA9 or BA40 of schizophrenic patients, althoughM2R mRNA was not detectable.
Animals models have provided further evidence of the potentialinteraction of cholinergic/dopaminergic systems. MuscarinicmRNAs are co-localised with dopaminergic neurones (Bernardet al., 1992; Weiner et al., 1990) and both muscarinic and dopami-nergic receptors modulate acetylcholine or dopamine release bothdirectly and indirectly (Allaoua et al., 1993; Bymaster et al., 1994;Friedman et al., 1990; Imperato et al., 1993; Moore et al., 1999;Yamada et al., 1990). The muscarinic compound ((5R,6R)-6-(3-propylthio-1,2,5-thiadiazol-4-yl)-1-azabicyclo[3,2,1]octane) PTAC,which is a partial agonist at M2/M4 receptors and an antagonist atM1, M3 andM5 receptors, was shown to be active in animal modelspredictive of antipsychotic behaviour, again consistent with a rolefor muscarinic receptors in the treatment of schizophrenia(Bymaster et al., 1999). Terry et al. (2006) investigated the effects ofchronic antipsychotic administration of haloperidol, chlorproma-zine, risperidone and olanzapine (most similar to clozapine)administered via drinking water over 90 or 180 days. They foundthat [3H]pirenzepine (M1/M4) binding levels were slightly higherin olanzapine treated animals (90 days) in the cortex lamina I andwere reduced in the CA1 of the hippocampus following chlor-promazine (180 days) treatment. In the same study, [3H]AF-DX 384(M1/M2) autoradiography analysis showed higher binding levels in10 brain regions including the cerebral cortex lamaniae I-V, the CA1and some thalamic nuclei (MD and AV) following 90 days olanza-pine treatment, whereas after 180 days these changes were nolonger evident. Zavitsanou et al. (2007) found that 36-day admin-istration of antipsychotics produced increases in [3H]pirenzepinebinding in rat dentate gyrus (DG) and CA1 hippocampal area forhaloperidol and the DG by clozapine, which was supported bya recent report of largely hippocampus-specific up-regulation ofM1 receptor mRNA after treatment with typical or atypical APDs(Han et al., 2008). Some studies have also investigated the effects ofpsychotomimetics upon muscarinic receptor binding. PCP treat-ment appears to have varied effects according to species. In rat aftera single acute injection of PCP (40 mg/kg) no effects on [3H]pir-enzapine (M1/M4) or [3H]AF-DX 384 (M1/M2) autoradiographical
receptor binding levels in rat brain were seen (Zavitsanou et al.,2008), whereas 1 and 14-day treatment of PCP (10 mg/kg) inmice had similar effects, elevating [3H]pirenzepine binding in thecingulate cortex, caudate putamen, nucleus accumbens andhippocampus (Newell et al., 2007b). Three week amphetaminetreatment in rats using [3H]pirenzapine binding and RT-PCR formRNA quantification in frontal cortex (FCx), hypothalamus (HYP)and striatum (Str) homogenates, showed a decrease in bindinglevels in the Ctx and Str which clozapine reversed while haloper-idol had no effect. No significant changes in M1R mRNA in thesebrain regions were seen in any treatment group (Malkoff et al.,2008).
Thus it remains ambiguous as to the role of the various musca-rinic receptors either in the aetiologyof schizophrenia, the treatmentof the disease or the APD side effects. In the present studywe soughtto determine the ability of both acute and chronic clozapine (atypicalAPD) and haloperidol (typical APD) treatment to modulate M1R,M2R andM3RmRNA. Inmany previous studies, intraperitoneal (i.p.)injections have been used to administer the antipsychotic dose. Asclozapinehas a relatively shorthalf life in the rat of approximately 2h(Baldessarini et al.,1993), intraperitoneal (i.p.) dosingwould result ina greater fluctuation in drug levels and receptor occupancy. Thisexperiment was designed to attempt to mimic (as near as possiblewithin the limitations of drug solubility and intolerable side effects(Kapur et al., 2003)) the human dosing regimen and consequentreceptor occupancy, by using osmoticmini-pumps to administer theantipsychotic drugs. This also facilitated a chronic treatment regimeof 21-days,whichwas chosen to approximate the time-lag period foronset of therapeutic effects within schizophrenic patients. Thistreatment regime also allowed us to determine whether a differentmode of drug administration, may influence the effect of clozapineupon the receptor mRNA and protein levels.
In order to identify additional potential "therapeutic" effects ofthe drugs, following on from the results of the chronic antipsychotictreatment, we also examined the ability of these drugs to modulatemuscarinic M1 receptor mRNA expression in a schizophrenia-related animal model. Based upon accumulating evidence tosupport a glutamate hypofunction hypothesis of schizophrenia(Jentsch and Roth, 1999; Coyle, 2006), we employed repeatedtreatment with a low dose of the psychotomimetic non competitiveNMDA receptor antagonist phencyclidine (PCP), a schizophrenia-related animal model, that produces deficits in brain imaging,neurochemistry and behaviour (Cochran et al., 2003; Egerton et al.,2008; Pratt et al., 2008). We have examined the ability of sub-chronic PCP to modulate muscarinic mRNA expression, andwhether any PCP-induced charges of M1 mRNA can be modified bychronic APD treatment. Muscarinic mRNA levels were assessedusing oligonucleotide in-situ hybridisation (ISH).
2. Materials and methods
2.1. Acute and chronic treatment regime for administration of the antipsychoticsclozapine and haloperidol
Clozapine was dissolved in 3% acetic acid in saline and haloperidol in 3% aceticacid in water which were then sterile filtered (0.2 mm DLL syringe filter, GelmanSciences, Ann Arbor, MI, USA) for the M2R mRNA study and sterile filtered 3% aceticacid in water for the M1R and M3R studies. Osmotic mini-pumps (Alzet, Palo Alto,CA, USA; Model 2ML4) were aseptically filled (according to the Alzet protocol) 18 hbefore implantation, then placed in sterile tubes at room temperature and coveredto prevent light penetration. Male Hooded Long-Evans rats (Harlan Olac, 186e248 g)were housed in groups of two with food and water freely available. These studieswere performed in accordance with the Animals (Scientific Procedures) Act (1986)and Guide for the Care and Use of Laboratory Animals as adopted by the NationalInstitutes of Health. The animals were placed in a constant 12 h lightedark cycle.Osmotic mini-pumps were implanted on day �1 using halothane anaesthesia,allowing the pumps to equilibrate for one day within the animal.
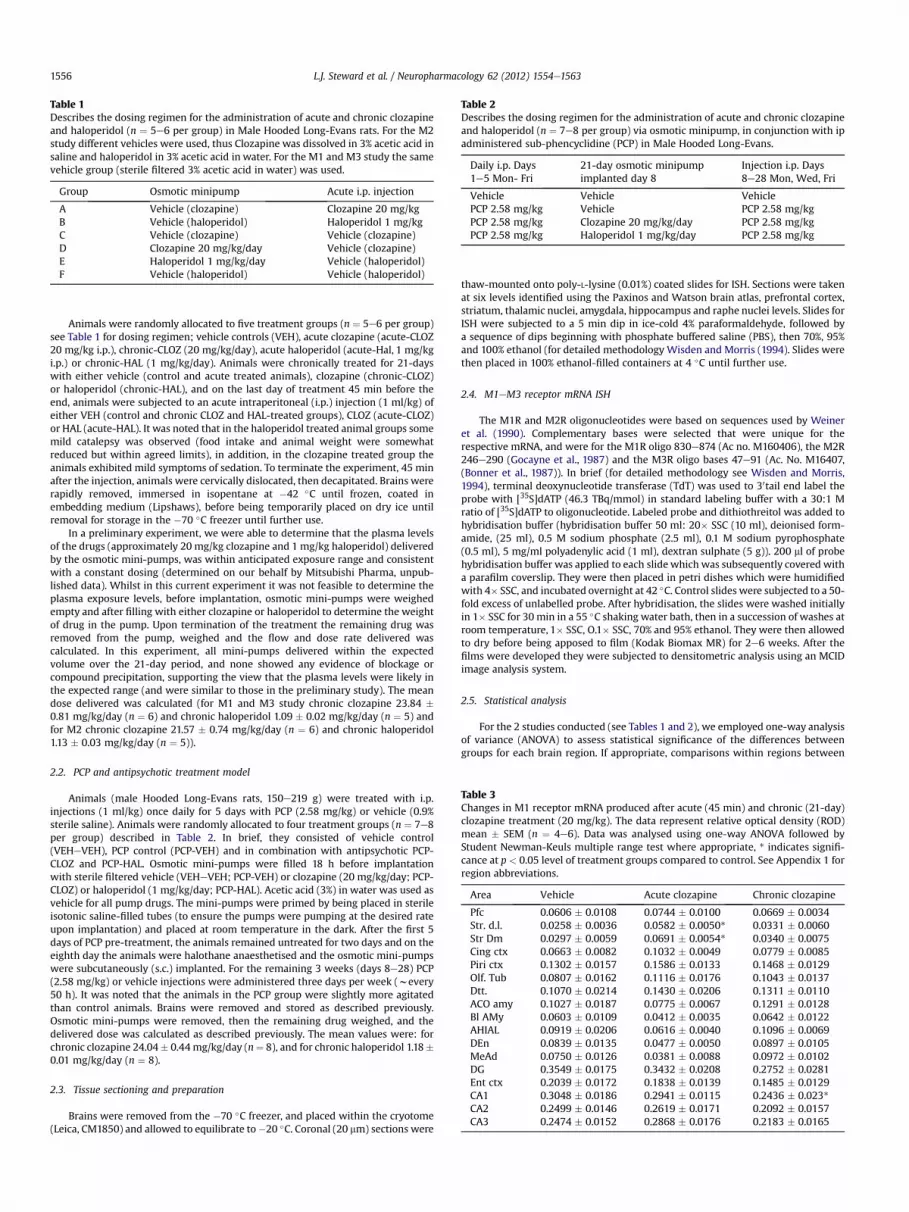
Table 2Describes the dosing regimen for the administration of acute and chronic clozapineand haloperidol (n ¼ 7e8 per group) via osmotic minipump, in conjunction with ipadministered sub-phencyclidine (PCP) in Male Hooded Long-Evans.
Daily i.p. Days1e5 Mon- Fri
21-day osmotic minipumpimplanted day 8
Injection i.p. Days8e28 Mon, Wed, Fri
Vehicle Vehicle VehiclePCP 2.58 mg/kg Vehicle PCP 2.58 mg/kgPCP 2.58 mg/kg Clozapine 20 mg/kg/day PCP 2.58 mg/kgPCP 2.58 mg/kg Haloperidol 1 mg/kg/day PCP 2.58 mg/kg
Table 3Changes in M1 receptor mRNA produced after acute (45 min) and chronic (21-day)clozapine treatment (20 mg/kg). The data represent relative optical density (ROD)mean � SEM (n ¼ 4e6). Data was analysed using one-way ANOVA followed byStudent Newman-Keuls multiple range test where appropriate, * indicates signifi-cance at p < 0.05 level of treatment groups compared to control. See Appendix 1 forregion abbreviations.
Area Vehicle Acute clozapine Chronic clozapine
Pfc 0.0606 � 0.0108 0.0744 � 0.0100 0.0669 � 0.0034Str. d.l. 0.0258 � 0.0036 0.0582 � 0.0050* 0.0331 � 0.0060Str Dm 0.0297 � 0.0059 0.0691 � 0.0054* 0.0340 � 0.0075Cing ctx 0.0663 � 0.0082 0.1032 � 0.0049 0.0779 � 0.0085Piri ctx 0.1302 � 0.0157 0.1586 � 0.0133 0.1468 � 0.0129Olf. Tub 0.0807 � 0.0162 0.1116 � 0.0176 0.1043 � 0.0137Dtt. 0.1070 � 0.0214 0.1430 � 0.0206 0.1311 � 0.0110ACO amy 0.1027 � 0.0187 0.0775 � 0.0067 0.1291 � 0.0128Bl AMy 0.0603 � 0.0109 0.0412 � 0.0035 0.0642 � 0.0122AHIAL 0.0919 � 0.0206 0.0616 � 0.0040 0.1096 � 0.0069DEn 0.0839 � 0.0135 0.0477 � 0.0050 0.0897 � 0.0105MeAd 0.0750 � 0.0126 0.0381 � 0.0088 0.0972 � 0.0102DG 0.3549 � 0.0175 0.3432 � 0.0208 0.2752 � 0.0281Ent ctx 0.2039 � 0.0172 0.1838 � 0.0139 0.1485 � 0.0129CA1 0.3048 � 0.0186 0.2941 � 0.0115 0.2436 � 0.023*CA2 0.2499 � 0.0146 0.2619 � 0.0171 0.2092 � 0.0157CA3 0.2474 � 0.0152 0.2868 � 0.0176 0.2183 � 0.0165
Table 1Describes the dosing regimen for the administration of acute and chronic clozapineand haloperidol (n ¼ 5e6 per group) in Male Hooded Long-Evans rats. For the M2study different vehicles were used, thus Clozapine was dissolved in 3% acetic acid insaline and haloperidol in 3% acetic acid in water. For the M1 and M3 study the samevehicle group (sterile filtered 3% acetic acid in water) was used.
Group Osmotic minipump Acute i.p. injection
A Vehicle (clozapine) Clozapine 20 mg/kgB Vehicle (haloperidol) Haloperidol 1 mg/kgC Vehicle (clozapine) Vehicle (clozapine)D Clozapine 20 mg/kg/day Vehicle (clozapine)E Haloperidol 1 mg/kg/day Vehicle (haloperidol)F Vehicle (haloperidol) Vehicle (haloperidol)
L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e15631556
Animals were randomly allocated to five treatment groups (n ¼ 5e6 per group)see Table 1 for dosing regimen; vehicle controls (VEH), acute clozapine (acute-CLOZ20 mg/kg i.p.), chronic-CLOZ (20 mg/kg/day), acute haloperidol (acute-Hal, 1 mg/kgi.p.) or chronic-HAL (1 mg/kg/day). Animals were chronically treated for 21-dayswith either vehicle (control and acute treated animals), clozapine (chronic-CLOZ)or haloperidol (chronic-HAL), and on the last day of treatment 45 min before theend, animals were subjected to an acute intraperitoneal (i.p.) injection (1 ml/kg) ofeither VEH (control and chronic CLOZ and HAL-treated groups), CLOZ (acute-CLOZ)or HAL (acute-HAL). It was noted that in the haloperidol treated animal groups somemild catalepsy was observed (food intake and animal weight were somewhatreduced but within agreed limits), in addition, in the clozapine treated group theanimals exhibited mild symptoms of sedation. To terminate the experiment, 45 minafter the injection, animals were cervically dislocated, then decapitated. Brains wererapidly removed, immersed in isopentane at �42 �C until frozen, coated inembedding medium (Lipshaws), before being temporarily placed on dry ice untilremoval for storage in the �70 �C freezer until further use.
In a preliminary experiment, we were able to determine that the plasma levelsof the drugs (approximately 20 mg/kg clozapine and 1 mg/kg haloperidol) deliveredby the osmotic mini-pumps, was within anticipated exposure range and consistentwith a constant dosing (determined on our behalf by Mitsubishi Pharma, unpub-lished data). Whilst in this current experiment it was not feasible to determine theplasma exposure levels, before implantation, osmotic mini-pumps were weighedempty and after filling with either clozapine or haloperidol to determine the weightof drug in the pump. Upon termination of the treatment the remaining drug wasremoved from the pump, weighed and the flow and dose rate delivered wascalculated. In this experiment, all mini-pumps delivered within the expectedvolume over the 21-day period, and none showed any evidence of blockage orcompound precipitation, supporting the view that the plasma levels were likely inthe expected range (and were similar to those in the preliminary study). The meandose delivered was calculated (for M1 and M3 study chronic clozapine 23.84 �0.81 mg/kg/day (n ¼ 6) and chronic haloperidol 1.09 � 0.02 mg/kg/day (n ¼ 5) andfor M2 chronic clozapine 21.57 � 0.74 mg/kg/day (n ¼ 6) and chronic haloperidol1.13 � 0.03 mg/kg/day (n ¼ 5)).
2.2. PCP and antipsychotic treatment model
Animals (male Hooded Long-Evans rats, 150e219 g) were treated with i.p.injections (1 ml/kg) once daily for 5 days with PCP (2.58 mg/kg) or vehicle (0.9%sterile saline). Animals were randomly allocated to four treatment groups (n ¼ 7e8per group) described in Table 2. In brief, they consisted of vehicle control(VEHeVEH), PCP control (PCP-VEH) and in combination with antipsychotic PCP-CLOZ and PCP-HAL. Osmotic mini-pumps were filled 18 h before implantationwith sterile filtered vehicle (VEHeVEH; PCP-VEH) or clozapine (20 mg/kg/day; PCP-CLOZ) or haloperidol (1 mg/kg/day; PCP-HAL). Acetic acid (3%) in water was used asvehicle for all pump drugs. The mini-pumps were primed by being placed in sterileisotonic saline-filled tubes (to ensure the pumps were pumping at the desired rateupon implantation) and placed at room temperature in the dark. After the first 5days of PCP pre-treatment, the animals remained untreated for two days and on theeighth day the animals were halothane anaesthetised and the osmotic mini-pumpswere subcutaneously (s.c.) implanted. For the remaining 3 weeks (days 8e28) PCP(2.58 mg/kg) or vehicle injections were administered three days per week (wevery50 h). It was noted that the animals in the PCP group were slightly more agitatedthan control animals. Brains were removed and stored as described previously.Osmotic mini-pumps were removed, then the remaining drug weighed, and thedelivered dose was calculated as described previously. The mean values were: forchronic clozapine 24.04� 0.44mg/kg/day (n¼ 8), and for chronic haloperidol 1.18�0.01 mg/kg/day (n ¼ 8).
2.3. Tissue sectioning and preparation
Brains were removed from the �70 �C freezer, and placed within the cryotome(Leica, CM1850) and allowed to equilibrate to�20 �C. Coronal (20 mm) sections were
thaw-mounted onto poly-L-lysine (0.01%) coated slides for ISH. Sections were takenat six levels identified using the Paxinos and Watson brain atlas, prefrontal cortex,striatum, thalamic nuclei, amygdala, hippocampus and raphe nuclei levels. Slides forISH were subjected to a 5 min dip in ice-cold 4% paraformaldehyde, followed bya sequence of dips beginning with phosphate buffered saline (PBS), then 70%, 95%and 100% ethanol (for detailed methodology Wisden and Morris (1994). Slides werethen placed in 100% ethanol-filled containers at 4 �C until further use.
2.4. M1eM3 receptor mRNA ISH
The M1R and M2R oligonucleotides were based on sequences used by Weineret al. (1990). Complementary bases were selected that were unique for therespective mRNA, and were for the M1R oligo 830e874 (Ac no. M160406), the M2R246e290 (Gocayne et al., 1987) and the M3R oligo bases 47e91 (Ac. No. M16407,(Bonner et al., 1987)). In brief (for detailed methodology see Wisden and Morris,1994), terminal deoxynucleotide transferase (TdT) was used to 30tail end label theprobe with [35S]dATP (46.3 TBq/mmol) in standard labeling buffer with a 30:1 Mratio of [35S]dATP to oligonucleotide. Labeled probe and dithiothreitol was added tohybridisation buffer (hybridisation buffer 50 ml: 20� SSC (10 ml), deionised form-amide, (25 ml), 0.5 M sodium phosphate (2.5 ml), 0.1 M sodium pyrophosphate(0.5 ml), 5 mg/ml polyadenylic acid (1 ml), dextran sulphate (5 g)). 200 ml of probehybridisation buffer was applied to each slide which was subsequently covered witha parafilm coverslip. They were then placed in petri dishes which were humidifiedwith 4� SSC, and incubated overnight at 42 �C. Control slides were subjected to a 50-fold excess of unlabelled probe. After hybridisation, the slides were washed initiallyin 1� SSC for 30 min in a 55 �C shaking water bath, then in a succession of washes atroom temperature, 1� SSC, O.1� SSC, 70% and 95% ethanol. They were then allowedto dry before being apposed to film (Kodak Biomax MR) for 2e6 weeks. After thefilms were developed they were subjected to densitometric analysis using an MCIDimage analysis system.
2.5. Statistical analysis
For the 2 studies conducted (see Tables 1 and 2), we employed one-way analysisof variance (ANOVA) to assess statistical significance of the differences betweengroups for each brain region. If appropriate, comparisons within regions between

Table 4Changes in M1 receptor mRNA produced after acute (45 min) and chronic (21-day)haloperidol treatment (1 mg/kg). The data represent relative optical density (ROD)mean � SEM (n ¼ 4e6). Data was analysed using one-way ANOVA followed byStudent Newman-Keuls multiple range test where appropriate, * indicates signifi-cance at p < 0.05 level of treatment groups compared to control. See Appendix 1 forregion abbreviations.
Area Vehicle Acute haloperidol Chronic haloperidol
Pfc 0.0606 � 0.0108 0.1151 � 0.0209* 0.0768 � 0.0084Str. d.l. 0.0258 � 0.0036 0.0677 � 0.0082* þ 0.0467 � 0.0101Str Dm 0.0297 � 0.0059 0.0726 � 0.0114* 0.0543 � 0.0132Cing ctx 0.0663 � 0.0082 0.1205 � 0.0189* 0.0852 � 0.0110Piri ctx 0.1302 � 0.0157 0.2267 � 0.0160* þ 0.2207 � 0.0164* þ
Olf. Tub 0.0807 � 0.0162 0.1655 � 0.0219* 0.1722 � 0.0298*Dtt. 0.1070 � 0.0214 0.2020 � 0.0210* þ 0.1726 � 0.0118ACO amy 0.1027 � 0.0187 0.0925 � 0.0098 0.0947 � 0.0186Bl AMy 0.0603 � 0.0109 0.0576 � 0.0088 0.0519 � 0.0101AHIAL 0.0919 � 0.0206 0.0956 � 0.0217 0.0797 � 0.0065DEn 0.0839 � 0.0135 0.0711 � 0.0130 0.0655 � 0.0058MeAd 0.0750 � 0.0126 0.0575 � 0.0174 0.0595 � 0.0107DG 0.3549 � 0.0175 0.3647 � 0.0311 0.4211 � 0.0153Ent ctx 0.2039 � 0.0172 0.2283 � 0.0230 0.2368 � 0.0082CA1 0.3048 � 0.0186 0.3201 � 0.0082 0.3384 � 0.0104CA2 0.2499 � 0.0146 0.2801 � 0.0119 0.3118 � 0.0171*CA3 0.2474 � 0.0220 0.2785 � 0.0220 0.2837 � 0.0188
L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e1563 1557
individual treatment group means were assessed using the Student Newman-Keulsmultiple range test. To test the hypothesis that drug treatment changed mRNAexpression in a defined brain circuit, we employed two-way ANOVA using the’defined region’ (within a brain circuit) and treatment as the factor. If appropriatethis was followed by a Tukeys post-hoc test.
3. Results
3.1. Effects of acute and chronic antipsychotic treatmenton muscarinic mRNA M1R mRNA
The distribution of M1R mRNA was consistent with previousstudies (Table 3, Fig. 1; Bonner et al., 1987; Weiner et al., 1990). Thehighest levels of expression were observed at the hippocampallevel in the CA1, CA2 and CA3 fields, and the dentate gyrus (DG). Inthe cortical regions, M1R mRNA expression was high in the ento-rhinal and piriform cortices, whereas more moderate levels weredetected in the prefrontal (PFC) and cingulate cortices. Moderatelevels of M1RmRNAwere also observed in some amygdaloid nucleiand the olfactory tubercle with the lowest levels detected in thestriatum.
After chronic clozapine treatment there was an apparentgeneral decrease in M1R mRNA within the hippocampal regionsCA1, CA2, CA3 and dentate gyrus, although the CA1 was the onlyregion inwhich this attained statistical significance (Table 3; Fig. 2).Conversely chronic haloperidol caused a significant increase inM1RmRNA in the CA2 field of the hippocampus (Table 4; Fig. 2). Therewere no significant changes in the prefrontal cortex with theexception of acute haloperidol (Table 4; Fig. 2). Acute clozapine andhaloperidol produced increases in M1R mRNA in the dorsolateraland dorsomedial striatum (Tables 3 and 4; Fig. 2).
3.2. Muscarinic M2R mRNA
The level of M2R mRNA expression was much lower and lesswidespread than that seen for M1R mRNA necessitating a film
Fig. 1. A comparison between the distribution of M1, M2, and M3 muscarinic mRNA at the l(right column), using in-situ hybridisation.
exposure 2e3 times as long, consistent with previous studies(Vilaro et al., 1994). The highest levels of M2R mRNA wereexpressed in the piriform cortex and DG and the dorsal teniatectae (DTT; Fig. 1, Table 5). Moderate levels were observed inthe PFC, olfactory tubercle, hippocampus CA1, CA2 and CA3fields, the vertical diagonal band and the retrosplenial cortex(Table 5).
The M2R receptor mRNA was not significantly changed afteracute or chronic clozapine or haloperidol treatment in all theregions measured with the exception of the cingulate cortexwhere it was increased by acute haloperidol treatment (Tables 5and 6).
evel of the prefrontal cortex (left column), striatum (centre column) and hippocampus

0
50
100
150
*
Ac Cloz
Chr Cloz
Ac Hal
Chr Hal
Hippocampal CA1
% C
on
tro
l
0
50
100
150 *
Hippocampal CA2
% C
on
tro
l
0
100
200
300 **
Dorsolateral Striatum
% C
on
tro
l
Fig. 2. The effect of acute (ac., 45 min) and chronic (chr, 21-day) clozapine (20 mg/kg/day) and haloperidol (1 mg/kg/day) treatment upon changes in M1 mRNA within thedorsolateral striatum (StrDL) and the hippocampus (CA1 and CA2). Histogramsrepresent % control relative optical density (ROD) mean � SEM (n ¼ 4e6). Data wasanalysed using one-way ANOVA followed by Student Newman-Keuls multiple rangetest where appropriate, * indicates significance at p < 0.05 level of treatment groupscompared to control.
Table 6Expression of M2 receptor mRNA after acute (45 min) and chronic (21-day) halo-peridol treatment (1 mg/kg). The data represent relative optical density (ROD) mean� SEM (n ¼ 2e6). Data was analysed using one-way ANOVA followed by StudentNewman-Keuls multiple range test where appropriate, * indicates significance atp < 0.05 level of treatment groups compared to control. See Appendix 1 for regionabbreviations.
Area Vehicle Acute haloperidol Chronic haloperidol
Pfc 0.1062 � 0.0135 0.1379 � 0.0161 0.1246 � 0.0153Piri ctx 0.1999 � 0.0274 0.2639 � 0.0194 0.1508 � 0.0266Dtt 0.1376 � 0.0169 0.1950 � 0.0192 0.1305 � 0.0124Olf. Tub 0.0747 � 0.0228 0.1136 � 0.0306 0.0851 � 0.0221Cing ctx 0.0331 � 0.0164 0.0932 � 0.0181* 0.0412 � 0.0094VDb 0.1015 � 0.0185 0.1361 � 0.0190 0.1120 � 0.0153DG 0.1546 � 0.0348 0.1325 � 0.0368 0.2477 � 0.0398CA1 0.0815 � 0.0158 0.0817 � 0.0171 0.1242 � 0.0249CA2 0.0626 � 0.0204 0.0715 � 0.0328 0.0998 � 0.0086CA3 0.1065 � 0.0271 0.1136 � 0.0325 0.1810 � 0.0294RsGb 0.0940 � 0.0255 0.0864 � 0.0318 0.1237 � 0.0196
L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e15631558
3.3. Muscarinic M3R mRNA
Although expressed at a lower level than the M1R mRNA in thecurrent experiment relatively high levels of M3R mRNA weredetected compared to the M2R mRNA. Highest levels wereobserved in the CA1 and CA3 hippocampal fields, the piriformcortex, DTT, MHb and the claustrum (Fig. 1, Table 7). Moderatelevels were detected in the prefrontal and cingulate, retrosplenialand dorsal endopiriform (DEn) cortices and the CA2 region of thehippocampus. However, M3R receptor mRNA was not significantlychanged after acute or chronic clozapine or haloperidol treatments(Tables 7 and 8).
Table 5Changes in M2 receptor mRNA produced after acute (45 min) and chronic (21-day)clozapine treatment (20 mg/kg). The data represent relative optical density (ROD)mean � SEM (n ¼ 2e6). Data was analysed using one-way ANOVA followed byStudent Newman-Keuls multiple range test where. See Appendix 1 for regionabbreviation.
Area Vehicle Acute clozapine Chronic clozapine
Pfc 0.1131 � 0.0212 0.1326 � 0.0158 0.0634 � 0.0229Piri ctx 0.1901 � 0.0186 0.2471 � 0.0195 0.1561 � 0.0244Dtt 0.1600 � 0.0163 0.2120 � 0.0235 0.1255 � 0.0168Olf. Tub 0.0551 � 0.0133 0.1088 � 0.0153 0.0653 � 0.0254Cing ctx 0.0689 � 0.0150 0.0674 � 0.0207 0.0374 � 0.0200VDb 0.1182 � 0.0164 0.1487 � 0.0169 0.0901 � 0.0276DG 0.1362 � 0.0269 0.1806 � 0.0221 0.1506 � 0.0205CA1 0.0489 � 0.0116 0.0963 � 0.0162 0.0692 � 0.0149CA2 0.0149 � 0.0115 0.0495 � 0.0250 0.0431 � 0.0104CA3 0.0889 � 0.0211 0.1431 � 0.0204 0.1372 � 0.0179RsGb 0.0660 � 0.0158 0.0984 � 0.0149 0.0710 � 0.0147
3.4. Effects of chronic PCP treatment and antipsychoticon muscarinic M1R mRNA
Clozapine and haloperidol produced alterations in the pattern ofM1R mRNA expression in the PCP treated animals, which werequite different when compared to the study of APD treatmentalone. There were highly significant increases in M1R mRNA levelsin the retrosplenial cortex (RSGb), the medial amygdaloid nucleus(MeAd) and the ventromedial hypothalamus (VMH), in animalstreated with PCP-Hal combination as compared to all other treat-ment groups (Fig. 3, Table 9). There were no significant changes inany hippocampal regions in any of the treatment groups (Table 9).In the nucleus accumbens core, PCP-Clozapine significantlydecreased M1R mRNA compared to the PCP-Veh and PCP-Haltreatment groups, although it was not significantly different fromVeheVeh. The signal intensities in the PCP-Veh and PCP-Hal groupsalso appeared increased compared to VeheVeh, although this didnot reach statistical significance (Fig. 3, Table 9).
It was notable that PCP-Veh treatment tended to produce anincrease in M1R mRNA (of w23%) in individual PFC regionscompared to the VeheVeh treatment group, and therefore a two-way ANOVA (followed by a Tukey’s post-hoc) was performed oninteracting regions of the prefrontal cortex. The components of thePFC: the prelimbic cortex layers 1, 2, 3 and the ventral (Vo) andposterior orbital (Po) cortex, were analysed and a significant
Table 7Expression of M3 receptor mRNA after acute (45 min) and chronic (21-day) cloza-pine treatment (20 mg/kg). The data represent relative optical density (ROD) mean� SEM (n ¼ 4e6). Data was analysed using one-way ANOVA followed by StudentNewman-Keuls multiple range test where appropriate. See Appendix 1 for regionabbreviations.
Area Vehicle Acute clozapine Chronic clozapine
Pfc 0.1556 � 0.0124 0.1660 � 0.0162 0.1632 � 0.0146Cing ctx 0.1893 � 0.0197 0.1734 � 0.0138 0.1884 � 0.0129DEn 0.2245 � 0.0167 0.1897 � 0.0151 0.2028 � 0.0181Piri ctx 0.2965 � 0.0264 0.2427 � 0.0179 0.2230 � 0.0251Dtt. 0.2525 � 0.0286 0.2429 � 0.0222 0.1984 � 0.0199M1 ctx 0.1796 � 0.0144 0.1675 � 0.0077 0.1633 � 0.0190Ent ctx 0.1014 � 0.0281 0.1711 � 0.0174 0.1304 � 0.0263CA1 0.2628 � 0.0234 0.2536 � 0.0274 0.2700 � 0.0311CA2 0.1321 � 0.0261 0.1314 � 0.0140 0.1370 � 0.0346CA3 0.3125 � 0.0415 0.3580 � 0.0225 0.2963 � 0.0377MHb 0.2493 � 0.0469 0.1046 � 0.0160 0.1576 � 0.0155RSGb 0.1276 � 0.0192 0.0826 � 0.0128 0.1352 � 0.0156Claustrum 0.3138 � 0.0324 0.2994 � 0.0242 0.3033 � 0.0303

Table 8Expression of M3 receptor mRNA after acute (45 min) and chronic 21-day halo-peridol treatment (1 mg/kg). The data represent relative optical density (ROD) mean� SEM (n ¼ 4e6). Data was analysed using one-way ANOVA followed by StudentNewman-Keuls multiple range test where appropriate. See Appendix 1 for regionabbreviations.
Area Vehicle Acute haloperidol Chronic haloperidol
Pfc 0.1556 � 0.0124 0.1784 � 0.0190 0.1860 � 0.0192Cing ctx 0.1893 � 0.0197 0.1869 � 0.0022 0.1940 � 0.0126Den 0.2245 � 0.0167 0.2263 � 0.0339 0.2363 � 0.0150Piri ctx 0.2965 � 0.0264 0.2591 � 0.0321 0.3227 � 0.0223Dtt. 0.2525 � 0.0286 0.2549 � 0.0215 0.2748 � 0.0306M1 ctx 0.1796 � 0.0144 0.1776 � 0.0205 0.1946 � 0.0353Ent ctx 0.1014 � 0.0281 0.1396 � 0.0201 0.1184 � 0.0331CA1 0.2628 � 0.0234 0.2534 � 0.0186 0.2384 � 0.0208CA2 0.1321 � 0.0261 0.1189 � 0.0157 0.1049 � 0.0250CA3 0.3125 � 0.0415 0.3078 � 0.0260 0.2758 � 0.0322MHb 0.2493 � 0.0469 0.1891 � 0.0333 0.1698 � 0.0514RSGb 0.1276 � 0.0192 0.0802 � 0.0157 0.1575 � 0.0144Claustrum 0.3138 � 0.0324 0.2807 � 0.0287 0.3301 � 0.0240
L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e1563 1559
treatment effect upon M1R mRNA was detected (F(3,159) ¼ 5.99,P¼ 0.01). Further post-hoc testing revealed that PCP-Veh treatmentproduced a statistically significant increase in M1R mRNAcompared to the VeheVeh treatment group (P¼ 0.0003). To a lesserdegree the PCP-Cloz treatment group also produced significantincreases in M1R mRNA in this region as a whole (P ¼ 0.0460).
The ventromedial hypothalamus appears to be integral in thecircuit between amygdaloid nuclei and also functionally congruouscortical nuclei. In a two-way ANOVA analysis of this circuit (ante-rocortico (Aco), basolateral (BlAmy), central (CeA), medial (MeAd)amygdaloid nuclei, RSGb, VMh) the treatment groups were signif-icantly different (F(3,191) ¼ 18.15, P < 0.0001). Tukey’s post-hoctests revealed that PCP-Hal treatment caused a significant
0
100
200
300
400
PCP-VEH
PCP-CLOZ
PCP-HAL
*#†
*#†
*#†
% C
on
tro
l
0
50
100
150
PCP-VEH
PCP-CLOZ
PCP-HAL
N. Acc. Core
#†
% C
on
tro
l
Fig. 3. Determination of M1 receptor mRNA levels in animals treated with chronic(described Table 2) PCP (2.58 mg/kg) or chronic PCP in combination with chronic(21-day) clozapine (20 mg/kg) or haloperidol (1 mg/kg) measured using in-situhybridisation. Data represents relative optical density (ROD) mean � SEM (n ¼ 7e8)the medial amygdaloid nucleus (MeAd), ventromedial hypothalamus (VmH) and retro-splenial cortex (RSGb) (upper panel) and in the nucleus accumbens core (N.Acc.Core)(lower panel). Datawere analysed using one-wayANOVA followed by Student Newman-Keuls multiple range test where appropriate, *p < 0.05 compared to vehicleevehicle;#p < 0.05 compared to PCP-vehicle and yp < 0.05 PCP-clozapine vs PCP-haloperidol.
increase in M1R mRNA compared to the other treatment groups(P < 0.00001). The prefrontal cortex areas Vo, Po and the prelimbic(PL) cortex 1,2,3, are also involved in the regulation of VMH. Theirinclusion of the circuit also showed the treatment groups weredifferent (F (3,351) ¼ 12.94, P < 0.0001) and post-hoc tests showedthat PCP-Hal treatment caused an M1 mRNA increase compared toPCP-Cloz (P < 0.001), PCP-Veh (P < 0.0035) and VeheVeh(P < 0.00001). However, the PCP-Veh group also increased M1mRNA compared to the VeheVeh group (P < 0.0410) although thisis more likely to be attributed to the PFC effect.
4. Discussion
The effect of antipsychotic treatment alone on muscarinicreceptor M1, M2, M3 R mRNA’s was investigated, and also inconjunction with PCP treatment in the case of M1R mRNA (sinceM1 mRNA was the only mRNA in the current study to be signifi-cantly altered following APD or PCP treatment). The distribution ofM1R, M2R and M3R mRNAs was consistent with previous studies(Bonner et al., 1987; Weiner et al., 1990, 1990; Vilaro et al., 1994),the M2R and M3R mRNA being of much lower abundance than theM1R mRNA.
The effect of acute and chronic antipsychotic treatment wasinvestigated on both M2R and M3R mRNA. It appeared that ingeneral neither acute, or chronic clozapine or haloperidol treat-ment produced any significant changes in M2R or M3R mRNAs, theonly exception being a change induced by acute haloperidol in thecingulate cortex. This was somewhat surprising in the case of clo-zapine, given its relatively high (Ki 20e50 nM) affinity for thesereceptors. The lack of change in either M2R or M3R mRNA does notnecessarily indicate that clozapine has no therapeutic effect via theM2 or M3 receptors. However, it suggests a lack of adaptiveresponse in the expression of these receptors, either because theyare less important sites of action than the M1R, or else because thegenes are less subject to regulatory control. Previous studies havebeen confounded by the lack of available ligands selective for theindividual muscarinic receptors. Twoweeks of clozapine treatmentreportedly had no effect on striatal muscarinic receptors (Cawleyet al., 1993). Terry et al. (2006) did not see any effect in rats afterchronic haloperidol treatment (90 and 180 days) using [3H]AF-DX384 (a relatively selective M2/M4 ligand) binding. A decrease in[3H]AF-DX 384 binding in striatal tissue from schizophrenicpatients compared to controls, has been reported. However, thisappeared to reflect adjunctive treatment of the muscarinic antag-onist benzotropine co-administered with their antipsychoticmedication (Crook et al., 1999). Equally chronic haloperidol orclozapine treatment in rats either increased, decreased or did notaffect striatal [3H]AF-DX 384 binding, depending on the dose andtime of treatment (Crook et al., 1999). Consequently, it remains tobe determined if either theM2 or M3 receptors participate in eitherclozapine’s or haloperidol’s antipsychotic action, and selectiveligands are eagerly awaited.
Acute haloperidol treatment produced substantial increases inM1R mRNA in a number of telencephalic regions including theprefrontal cortex and striatum. It is unlikely at the concentrationused (1 mg/kg), that haloperidol would be acting directly at M1receptors (Schotte et al., 1993), due to its relatively low affinity fortheM1 receptor (Ki 1500 nM) (Bolden et al., 1991) and it is probablyproducing these changes indirectly via D2 receptor antagonism. Insupport of this, it has been shown that 80% of striatal cholinergicneurones express D2 mRNA (LeMoine et al., 1990) suggesting thatD2 receptors are involved in cholinergic modulation. Furthermore,the changes in M1R mRNA in the striatum of the acute clozapinetreated animals, appear to be similar to those in the acute halo-peridol group. The levels of clozapine in the acute treated animals
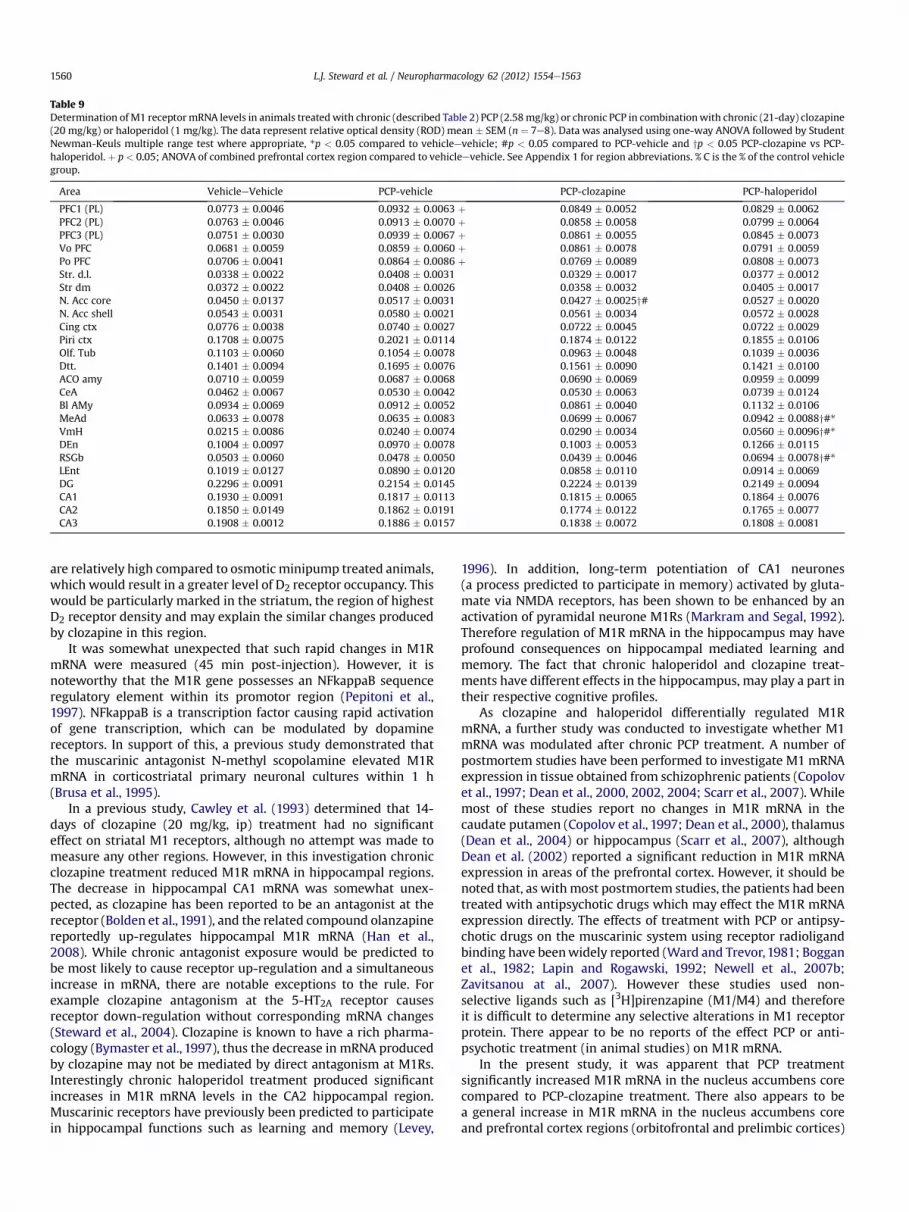
Table 9Determination ofM1 receptormRNA levels in animals treatedwith chronic (described Table 2) PCP (2.58mg/kg) or chronic PCP in combinationwith chronic (21-day) clozapine(20 mg/kg) or haloperidol (1 mg/kg). The data represent relative optical density (ROD) mean � SEM (n ¼ 7e8). Data was analysed using one-way ANOVA followed by StudentNewman-Keuls multiple range test where appropriate, *p < 0.05 compared to vehicleevehicle; #p < 0.05 compared to PCP-vehicle and yp < 0.05 PCP-clozapine vs PCP-haloperidol. þ p < 0.05; ANOVA of combined prefrontal cortex region compared to vehicleevehicle. See Appendix 1 for region abbreviations. % C is the % of the control vehiclegroup.
Area VehicleeVehicle PCP-vehicle PCP-clozapine PCP-haloperidol
PFC1 (PL) 0.0773 � 0.0046 0.0932 � 0.0063 þ 0.0849 � 0.0052 0.0829 � 0.0062PFC2 (PL) 0.0763 � 0.0046 0.0913 � 0.0070 þ 0.0858 � 0.0058 0.0799 � 0.0064PFC3 (PL) 0.0751 � 0.0030 0.0939 � 0.0067 þ 0.0861 � 0.0055 0.0845 � 0.0073Vo PFC 0.0681 � 0.0059 0.0859 � 0.0060 þ 0.0861 � 0.0078 0.0791 � 0.0059Po PFC 0.0706 � 0.0041 0.0864 � 0.0086 þ 0.0769 � 0.0089 0.0808 � 0.0073Str. d.l. 0.0338 � 0.0022 0.0408 � 0.0031 0.0329 � 0.0017 0.0377 � 0.0012Str dm 0.0372 � 0.0022 0.0408 � 0.0026 0.0358 � 0.0032 0.0405 � 0.0017N. Acc core 0.0450 � 0.0137 0.0517 � 0.0031 0.0427 � 0.0025y# 0.0527 � 0.0020N. Acc shell 0.0543 � 0.0031 0.0580 � 0.0021 0.0561 � 0.0034 0.0572 � 0.0028Cing ctx 0.0776 � 0.0038 0.0740 � 0.0027 0.0722 � 0.0045 0.0722 � 0.0029Piri ctx 0.1708 � 0.0075 0.2021 � 0.0114 0.1874 � 0.0122 0.1855 � 0.0106Olf. Tub 0.1103 � 0.0060 0.1054 � 0.0078 0.0963 � 0.0048 0.1039 � 0.0036Dtt. 0.1401 � 0.0094 0.1695 � 0.0076 0.1561 � 0.0090 0.1421 � 0.0100ACO amy 0.0710 � 0.0059 0.0687 � 0.0068 0.0690 � 0.0069 0.0959 � 0.0099CeA 0.0462 � 0.0067 0.0530 � 0.0042 0.0530 � 0.0063 0.0739 � 0.0124Bl AMy 0.0934 � 0.0069 0.0912 � 0.0052 0.0861 � 0.0040 0.1132 � 0.0106MeAd 0.0633 � 0.0078 0.0635 � 0.0083 0.0699 � 0.0067 0.0942 � 0.0088y#*VmH 0.0215 � 0.0086 0.0240 � 0.0074 0.0290 � 0.0034 0.0560 � 0.0096y#*DEn 0.1004 � 0.0097 0.0970 � 0.0078 0.1003 � 0.0053 0.1266 � 0.0115RSGb 0.0503 � 0.0060 0.0478 � 0.0050 0.0439 � 0.0046 0.0694 � 0.0078y#*LEnt 0.1019 � 0.0127 0.0890 � 0.0120 0.0858 � 0.0110 0.0914 � 0.0069DG 0.2296 � 0.0091 0.2154 � 0.0145 0.2224 � 0.0139 0.2149 � 0.0094CA1 0.1930 � 0.0091 0.1817 � 0.0113 0.1815 � 0.0065 0.1864 � 0.0076CA2 0.1850 � 0.0149 0.1862 � 0.0191 0.1774 � 0.0122 0.1765 � 0.0077CA3 0.1908 � 0.0012 0.1886 � 0.0157 0.1838 � 0.0072 0.1808 � 0.0081
L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e15631560
are relatively high compared to osmotic minipump treated animals,which would result in a greater level of D2 receptor occupancy. Thiswould be particularly marked in the striatum, the region of highestD2 receptor density and may explain the similar changes producedby clozapine in this region.
It was somewhat unexpected that such rapid changes in M1RmRNA were measured (45 min post-injection). However, it isnoteworthy that the M1R gene possesses an NFkappaB sequenceregulatory element within its promotor region (Pepitoni et al.,1997). NFkappaB is a transcription factor causing rapid activationof gene transcription, which can be modulated by dopaminereceptors. In support of this, a previous study demonstrated thatthe muscarinic antagonist N-methyl scopolamine elevated M1RmRNA in corticostriatal primary neuronal cultures within 1 h(Brusa et al., 1995).
In a previous study, Cawley et al. (1993) determined that 14-days of clozapine (20 mg/kg, ip) treatment had no significanteffect on striatal M1 receptors, although no attempt was made tomeasure any other regions. However, in this investigation chronicclozapine treatment reduced M1R mRNA in hippocampal regions.The decrease in hippocampal CA1 mRNA was somewhat unex-pected, as clozapine has been reported to be an antagonist at thereceptor (Bolden et al., 1991), and the related compound olanzapinereportedly up-regulates hippocampal M1R mRNA (Han et al.,2008). While chronic antagonist exposure would be predicted tobe most likely to cause receptor up-regulation and a simultaneousincrease in mRNA, there are notable exceptions to the rule. Forexample clozapine antagonism at the 5-HT2A receptor causesreceptor down-regulation without corresponding mRNA changes(Steward et al., 2004). Clozapine is known to have a rich pharma-cology (Bymaster et al., 1997), thus the decrease in mRNA producedby clozapine may not be mediated by direct antagonism at M1Rs.Interestingly chronic haloperidol treatment produced significantincreases in M1R mRNA levels in the CA2 hippocampal region.Muscarinic receptors have previously been predicted to participatein hippocampal functions such as learning and memory (Levey,
1996). In addition, long-term potentiation of CA1 neurones(a process predicted to participate in memory) activated by gluta-mate via NMDA receptors, has been shown to be enhanced by anactivation of pyramidal neurone M1Rs (Markram and Segal, 1992).Therefore regulation of M1R mRNA in the hippocampus may haveprofound consequences on hippocampal mediated learning andmemory. The fact that chronic haloperidol and clozapine treat-ments have different effects in the hippocampus, may play a part intheir respective cognitive profiles.
As clozapine and haloperidol differentially regulated M1RmRNA, a further study was conducted to investigate whether M1mRNA was modulated after chronic PCP treatment. A number ofpostmortem studies have been performed to investigate M1 mRNAexpression in tissue obtained from schizophrenic patients (Copolovet al., 1997; Dean et al., 2000, 2002, 2004; Scarr et al., 2007). Whilemost of these studies report no changes in M1R mRNA in thecaudate putamen (Copolov et al., 1997; Dean et al., 2000), thalamus(Dean et al., 2004) or hippocampus (Scarr et al., 2007), althoughDean et al. (2002) reported a significant reduction in M1R mRNAexpression in areas of the prefrontal cortex. However, it should benoted that, as withmost postmortem studies, the patients had beentreated with antipsychotic drugs which may effect the M1R mRNAexpression directly. The effects of treatment with PCP or antipsy-chotic drugs on the muscarinic system using receptor radioligandbinding have beenwidely reported (Ward and Trevor, 1981; Bogganet al., 1982; Lapin and Rogawski, 1992; Newell et al., 2007b;Zavitsanou at al., 2007). However these studies used non-selective ligands such as [3H]pirenzapine (M1/M4) and thereforeit is difficult to determine any selective alterations in M1 receptorprotein. There appear to be no reports of the effect PCP or anti-psychotic treatment (in animal studies) on M1R mRNA.
In the present study, it was apparent that PCP treatmentsignificantly increased M1R mRNA in the nucleus accumbens corecompared to PCP-clozapine treatment. There also appears to bea general increase in M1R mRNA in the nucleus accumbens coreand prefrontal cortex regions (orbitofrontal and prelimbic cortices)

L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e1563 1561
after chronic PCP treatment. The nucleus accumbens core existswithin a circuit which projects to the ventral pallidum, which inturn projects to the mediodorsal thalamic nucleus (MD). The MDsends a glutamatergic projection to the prefrontal cortex which inturn sends a glutamatergic input to the nucleus accumbens core(Heimer et al., 1991; Zahm and Brog, 1992). This circuit isconsidered to be involved in spatial working memory (Kalivaset al., 1999; Kelley, 1999) which is known to be disrupted inschizophrenia and following PCP treatment (Jentsch and Roth,1999; Wolkin et al., 1992). The modulation of M1R mRNA in thiscircuitry may hence play some role in the actions of clozapine.While it is unclear whether this role might be detrimental or"therapeutic", we note that a clozapine metabolite, N-desmethyl-clozapine, has been proposed to exert beneficial effects oncognitive deficits via stimulation of M1Rs in prefrontal cortex (Suret al., 2003; Li et al., 2005).
PCP-Hal treatment produced large increases in M1R mRNAwithin the ventromedial hypothalamus (VMH) and regions con-nected to it, the medial amygdalal nucleus (MeAd) and the ret-rosplenial cortex (RSGb), compared to all the other treatmentgroups. It has also been demonstrated that the VMH has recip-rocal connections with the prefrontal cortices (Ongur et al., 1998).The VMH has been associated with the regulation of appetitiveand reproductive behaviours (Altman and Bayer, 1986). Both D1and D5 mRNAs have been located in the VMH (Zhou et al., 1999).Since the increases in M1R mRNA within the VMH were only seenafter PCP-Hal treatment and not chronic haloperidol treatmentalone, it is tempting to postulate that PCP caused an elevation indopamine levels within this region, which were subsequentlyblocked by haloperidol at D1 and D5 receptors (haloperidolhaving a higher affinity than clozapine for these receptors). Theincrease in M1R mRNA levels may represent an attempt to rectifythese abnormalities following treatment with haloperidol (whichhas low propensity to induce weight gain relative to atypicalAPDs).
In summary chronic clozapine and haloperidol treatmentaltered expression of M1R mRNA in the hippocampus withoutaffectingM2 andM3 receptor expression. This supports the conceptthat M1R gene expression shows more plasticity in relation to APDtreatment than other muscarinic receptors. These changes in M1receptor expressionwere not evident when these drugs were givenin conjunction with chronic PCP treatment. Interestingly PCPtreatment alone produced an overall increase in M1 receptorexpression in prefrontal cortex-accumbens circuitry which was notrestored by either clozapine or haloperidol. These data are consis-tent with the view that neither APD shows dramatic protectionagainst cognitive deficits clinically. Thus while the participation ofM2 and M3 receptors in APD action remains unclear, our datasupport the concept that actions involving the M1R may contributeeither directly or indirectly in the antipsychotic or side effects ofboth clozapine and haloperidol.
Acknowledgements
This study was supported by Mitsubishi Tanabe Pharma Corp.(formerly Mitsubishi Pharma Corp.). We are grateful to Dr H.Yasu-matsu, Dr. S.Hammond, Prof. T.W.Stone, and Prof. C.M.R.Turner, ofthe YRING Research Management team, for their many usefuldiscussions and comments during the course of this work.
References
Allaoua, H., Chaudieu, I., Alonso, R., Quirion, R., Boksa, P., 1993. Muscarinic poten-tiation of excitatory amino acid-evoked dopamine release in Mesencephalic
cells - specificity for the Nmda response and role of intracellular messengers.Synapse 15, 39e47.
Altman, J., Bayer, S.A., 1986. The development of the rat hypothalamus. Advances inAnatomy Embryology and Cell Biology 100, 1e173.
Baldessarini, R.J., Centorrino, F., Flood, J.G., Volpicelli, S.A., Hustonlyons, D.,Cohen, B.M., 1993. Tissue concentrations of clozapine and its metabolites in therat. Neuropsychopharmacology 9, 117e124.
Bernard, V., Normand, E., Bloch, B., 1992. Phenotypical characterization of the ratstriatal neurons expressing muscarinic receptor genes. Journal of Neuroscience12, 3591e3600.
Boggan, W.O., Evans, M.G., Wallis, C.J., 1982. Effect of phencyclidine on [H-3] Qnbbinding. Life Sciences 30, 1193e1200.
Bolden, C., Cusack, B., Richelson, E., 1991. Clozapine is a potent and selectivemuscarinic antagonist at the 5 cloned human muscarinic acetylcholine-receptors expressed in Cho-K1 Cells. European Journal of Pharmacology 192,205e206.
Bonner, T.I., Buckley, N.J., Young, A.C., Brann, M.R., 1987. Identification of a Family ofmuscarinic acetylcholine-receptor genes. Science 237, 527e532.
Brusa, R., Gamalero, S.R., Genazzani, E., Eva, C., 1995. Primary neuronal culturesmuscarinic M1 and M3 receptor messenger-Rna levels are regulated byagonists, partial agonists and antagonists. European Journal of Pharmacology-Molecular Pharmacology Section 289, 9e16.
Bymaster, F.P., Rasmussen, K., Calligaro, D.O., Nelson, D.L., DeLapp, N.W.,Wong, D.T., Moore, N.A., 1997. In vitro and in vivo biochemistry of olanza-pine: a novel, atypical antipsychotic drug. Journal of Clinical Psychiatry 58(Suppl. 10), 28e36.
Bymaster, F.P., Wong, D.T., Mitch, C.H., Ward, J.S., Calligaro, D.O., Schoepp, D.D.,Shannon, H.E., Sheardown, M.J., Olesen, P.H., Suzdak, P.D., Swedberg, M.D.B.,Sauerberg, P., 1994. Neurochemical effects of the M(1) muscarinic agonistxanomeline (Ly246708/Nnc11-0232). Journal of Pharmacology and Experi-mental Therapeutics 269, 282e289.
Bymaster, F.P., Shannon, H.E., Rasmussen, K., DeLapp, N.W., Ward, J.S.,Calligaro, D.O., Mitch, C.H., Whitesitt, C., Ludvigsen, T.S., Sheardown, M.,Swedberg, M., Rasmussen, T., Olesen, P.H., Jeppesen, L., Sauerberg, P., Fink-Jensen, A., 1999. Potential role of muscarinic receptors in schizophrenia. LifeSciences 64, 527e534.
Bymaster, F.P., Felder, C.C., Tzavara, E., Nomikas, G.G., Calligaro, D.O., McKinzie, D.L.,2003. Muscarinic mechanisms of antipsychotic atypicality. Progress in Neuro-Psychopharmacology and Biological Psychiatry 27, 1125e1143.
Cancelli, I., Beltrame, M., D’Anna, L., Gigli, G.L., Valente, M., 2009 Sep. Drugs withanticholinergic properties: a potential risk factor for psychosis onset in Alz-heimer’s disease? Expert Opinion on Drug Safety 8 (5), 549e557.
Cawley, T.A., Shickley, T.J., Ruggieri, M.R., Luthin, G.R., 1993. Effect of chronicneuroleptic treatment on central and peripheral muscarinic receptors. Journalof Pharmacology and Experimental Therapeutics 267, 134e139.
Cochran, S.M., Steward, L.J., Kennedy, M., McKerchar, C.E., Pratt, J.A., Morris, B.J.,2003. Induction of metabolic hypofunction and neurochemical deficits afterchronic intermittent exposure to phencyclidine: differential modulation byantipsychotic drugs. Neuropsychopharmacology 28, 265e275.
Conn, P.J., Jones, C.K., Lindsley, C.W., 2009 Mar. Subtype-selective allosteric modu-lators of muscarinic receptors for the treatment of CNS disorders. Trends inPharmacological Sciences 30 (3), 148e155.
Copolov, D., Crook, J., Opeskin, K., Hill, C., Keks, N., Dean, B., 1997. Muscarinic M1receptors in the caudate putamen of patients with schizophrenia: decreasedreceptor density is not accompanied by decreased receptor mRNA. Schizo-phrenia Research 24, 32.
Coyle, J.T., 2006. Glutamate and schizophrenia: beyond the dopamine hypothesis.Cellular and Molecular Neurobiology Vol. 26, 4e6.
Crook, J.M., Dean, B., Pavey, G., Copolov, D., 1999. The binding of [H-3]AF-DX 384 isreduced in the CaudateePutamen of subjects with schizophrenia. Life Sciences64, 1761e1771.
Crook, J.M., Tomaskovic-Crook, E., Copolov, D.L., Dean, B., 2000. Decreased musca-rinic receptor binding in subjects with schizophrenia: a study of the humanhippocampal formation. Biological Psychiatry 48, 381e388.
Dean, B., Crook, J.M., Opeskin, K., Hill, C., Keks, N., Copolov, D.L., 1996. The density ofmuscarinic M1 receptors is decreased in the caudate-putamen of subjects withschizophrenia. Molecular Psychiatry 1, 54e58.
Dean, B., Crook, J.M., Pavey, G., Opeskin, K., Copolov, D.L., 2000. Muscarinic(1) and(2) receptor mRNA in the human caudate-putamen: no change in M1 mRNA inschizophrenia. Molecular Psychiatry 5, 203e207.
Dean, B., McLeod, M., Keriakous, D., McKenzie, J., Scarr, E., 2002. Decreasedmuscarinic(1) receptors in the dorsolateral prefrontal cortex of subjects withschizophrenia. Molecular Psychiatry 7, 1083e1091.
Dean, B., Gray, L., Keriakous, D., Scarr, E., 2004. A comparison of M1 and M4muscarinic receptors in the thalamus from control subjects and subjects withschizophrenia. Thalamus & Related Systems 2, 287e295.
Deng, C., Huang, X.F., 2005. Decreased density of muscarinic receptors in thesuperior temporal gyrus in schizophrenia. Journal of Neuroscience Research 81,883e890.
Egerton, A., Reid, L., McGregor, S., Cochran, S.M., Morris, B.J., Pratt, J.A., 2008. Sub-chronic and chronic PCP treatment produces temporally distinct deficits inattentional set shifting and prepulse inhibition in rats. Psychopharmacology198, 37e49.
Friedman, E., Wang, H.Y., Butkerait, P., 1990. Decreased striatal release of acetyl-choline following withdrawal from long-term treatment with haloperidol -

L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e15631562
modulation by cholinergic, Dopamine-D1 and Dopamine-D2 mechanisms.Neuropharmacology 29, 537e544.
Gocayne, J., Robinson, D.A., Fitzgerald, M.G., Chung, F.Z., Kerlavage, A.R., Lentes, K.U.,Lai, J., Wang, C.D., Fraser, C.M., Venter, J.C., 1987. Primary structure of rat cardiacbeta-adrenergic and muscarinic cholinergic receptors obtained by automatedDna-sequence analysis - further evidence for a multigene family. Proceedings ofthe National Academy of Sciences of the United States of America 84,8296e8300.
Han, M., Newell, K., Zavitsanou, K., Deng, C., Huang, X.-F., 2008. Effects of antipsy-chotic medication on muscarinic M1 receptor mRNA expression in the rat brain.Journal of Neuroscience Research 86, 457e464.
Heimer, L., Zahm, D.S., Churchill, L., Kalivas, P.W., Wohltmann, C., 1991. Specificity inthe projection patterns of accumbal core and shell in the rat. Neuroscience 41,89e125.
Hyde, T.M., Crook, J.M., 2001. Cholinergic systems and schizophrenia: primarypathology or epiphenomena? Journal of Chemical Neuroanatomy 22,53e63.
Imperato, A., Obinu, M.C., Casu, M.A., Mascia, M.S., Dazzi, L., Gessa, G.L., 1993.Evidence that neuroleptics increase striatal acetylcholine- release throughstimulation of dopamine-D(1) receptors. Journal of Pharmacology and Experi-mental Therapeutics 266, 557e562.
Jentsch, J.D., Roth, R.H., 1999. The neuropsychopharmacology of phencyclidine:from NMDA receptor hypofunction to the dopamine hypothesis of schizo-phrenia. Neuropsychopharmacology 20, 201e225.
Johnstone, E.C., Crow, T.J., Ferrier, I.N., Frith, C.D., Owens, D.G., Bourne, R.C.,Gamble, S.J., 1983. Adverse effects of anticholinergic medication on positiveschizophrenic symptoms. Psychological Medicine Aug 13 (3), 513e527.
Kalivas, P.W., Churchill, L., Romanides, A., 1999. Involvement of the pallid-alethalamocortical circuit in adaptive behavior. Annals of the New YorkAcademy of Sciences 877, 64e70.
Kapur, S., VanderSpek, S.C., Brownlee, B.A., Nobrega, J.N., 2003. Antipsychotic dosingin preclinical models is often unrepresentative of the clinical condition: a sug-gested solution based on in vivo occupancy. Journal of Pharmacology andExperimental Therapeutics 305 (2), 625e631.
Kelley, A.E., 1999. Functional specificity of ventral striatal compartments in appe-titive behaviors. Annals of the New York Academy of Sciences 877, 71e90.
Lapin, I.P., Rogawski, M.A., 1992. Muscarinic antagonists attenuate dizocilpine-induced hypermotility in mice. Life Sciences 50, L59ePL64.
LeMoine, C., Tison, F., Bloch, B., 1990. D2-Dopamine receptor gene-expressionby cholinergic neurons in the rat striatum. Neuroscience Letters 117,248e252.
Levey, A.I., 1996. Muscarinic acetylcholine receptor expression in memory circuits:implications for treatment of Alzheimer disease. Proceedings of the NationalAcademy of Sciences of the United States of America 93, 13541e13546.
Li, Z., Huang, M., Ichikawa, J., Dai, J., Meltzer, H.Y., 2005. N-desmethylclozapine,a major metabolite of clozapine, increases cortical acetylcholine and dopaminerelease in vivo via stimulation of M1 muscarinic receptors. Neuro-psychopharmacology 30, 1986e1995.
Malkoff, A., Weizman, A., Gozes, I., Rehavi, M., 2008. Decreased M1 muscarinicreceptor density in rat amphetaminemodel of schizophrenia is normalizedby clozapine, but not haloperidol. Journal of Neural Transmission 115,1563e1571.
Markram, H., Segal, M., 1992. The inositol 1,4,5-trisphosphate pathway mediatescholinergic potentiation of rat hippocampal neuronal responses to Nmda.Journal of Physiology-London 447, 513e533.
Moore, H., Fadel, J., Sarter, M., Bruno, J.P., 1999. Role of accumbens and corticaldopamine receptors in the regulation of cortical acetylcholine release. Neuro-science 88, 811e822.
Newell, K.A., Zavitsanou, K., Jew, S.K., Huang, X.F., 2007a. Alterations of muscarinicand GABA receptor binding in the posterior cingulate cortex in schizophrenia.Progress in Neuro-Psychopharmacology and Biological Psychiatry 31 (1),225e233.
Newell, K.A., Zavitsanou, K., Huang, X., 2007b. Opposing short- and long-termeffects on muscarinic M1/4 receptor binding following chronic phencyclidinetreatment. Journal of Neuroscience Research 85, 1358e1363.
Ongur, D., An, X., Price, J.L., 1998. Prefrontal cortical projections to the hypothalamusin macaque monkeys. Journal of Comparative Neurology 401, 480e505.
Pepitoni, S., Wood, I.C., Buckley, N.J., 1997. Structure of the M1 muscarinic acetyl-choline receptor gene and its promoter. Journal of Biological Chemistry 272,17112e17117.
Perry, E.K., Perry, R.H., 1995 Aug. Acetylcholine and hallucinations: disease-relatedcompared to drug-induced alterations in human consciousness. Brain andCognition 28 (3), 240e258.
Pratt, J.A., Winchester, C., Egerton, A., Cochran, S.M., Morris, B.J., 2008 Mar.Modelling prefrontal cortex deficits in schizophrenia: implications for treat-ment. British Journal of Pharmacology 153 (Suppl. 1), S465eS470.
Salah-Uddin, H., Scarr, E., Pavey, G., Kriss Harris, K., Hagan, J., Dean, B., Challiss, R.A.J.,Watson, J.M., 2009. AlteredM1muscarinic acetylcholine receptor (CHRM1)-Gaq/11 coupling in a schizophrenia endophenotype. Neuropsychopharmacology 34,2156e2166.
Scarr, E., Dean, B., 2008. Role of the cholinergic system in the pathology andtreatment of schizophrenia. Expert Review of Neurotherapeutics 9, 73e86.
Scarr, E., Keriakous, D., Crossland, N., Dean, B., 2006. No change in corticalmuscarinic M2, M3 receptors or [35S]GTPgS binding in schizophrenia. LifeSciences 78, 1231e1237.
Scarr, E., Sundram, S., Keriakous, K., Dean, B., 2007. Altered hippocampal muscarinicM4, but not M1, receptor expression from subjects with schizophrenia. Bio-logical Psychiatry 61, 1161e1170.
Schotte, A., Janssen, P.F.M., Megens, A., Leysen, J.E., 1993. Occupancy of centralneurotransmitter receptors by risperidone, clozapine and haloperidol,measured ex-vivo by quantitative autoradiography. Brain Research 631,191e202.
Steward, L.J., Kennedy, M.D., Morris, B.J., Pratt, J.A., 2004. Phencyclidine and theatypical antipsychotic clozapine downregulate prefrontal cortex 5-HT2Areceptors. Neuropharmacology 47, 527e537.
Sur, C., Mallorga, P.J., Wittmann, M., Jacobson, M.A., Pascarella, D., Williams, J.B.,Brandish, P.E., Pettibone, D.J., Scolnick, E.M., Conn, P.J., 2003. N-desmethylclo-zapine, an allosteric agonist at muscarinic 1 receptor, potentiates N-methyl-d-aspartate receptor activity. Proceedings of the National Academy of Sciences100, 13674e13679.
Tandon, R., Greden, J.F., Silk, K.R., 1988. Treatment of negative schizophrenicsymptoms with trihexyphenidyl. Journal of Clinical Psychopharmacology 8,212e215.
Terry Jr, A.V., Gearhart, D.A., Mahadik, S.P., Warsi, S., Waller, J.L., 2006. Chronictreatment with first or second generation antipsychotics in rodents: effects onhigh affinity nicotinic and muscarinic acetylcholine receptors in the brain.Neuroscience 140 (4), 1277e1287.
Terry, A., 2008. Role of the central cholinergic system in the therapeutics ofschizophrenia. Current Neuropharmacology 6, 286e292.
Vilaro, M.T., Palacios, J.M., Mengod, G., 1994. Multiplicity of muscarinic autoreceptorsubtypes - comparison of the distribution of cholinergic cells and cells con-taining messenger-Rna for 5 Subtypes of muscarinic receptors in the rat-brain.Molecular Brain Research 21, 30e46.
Ward, D., Trevor, A., 1981. Phencyclidine-Induced alteration in rat muscariniccholinergic receptor regulation. European Journal of Pharmacology 74,189e193.
Weiner, D.M., Levey, A.I., Brann, M.R., 1990. Expression of muscarinic acetylcholineand dopamine receptor messenger-Rnas in rat basal ganglia. Proceedings of theNational Academy of Sciences of the United States of America 87, 7050e7054.
Wisden, W.W., Morris, B.J., 1994. In situ hybridization with synthetic oligonucleotideprobes. In: W., W., Morris, B.J. (Eds.), In-situ Hybridization Protocols for theBrain. Academic Press, pp. 9e34.
Wolkin, A., Sanfilipo, M., Wolf, A.P., Angrist, B., Brodie, J.D., Rotrosen, J., 1992.Negative symptoms and hypofrontality in chronic-schizophrenia. Archives ofGeneral Psychiatry 49, 959e965.
Yamada, S., Nishi, S., Kojima, H., Yokoo, H., Tsutsumi, T., Inanaga, K., 1990.Involvement of the cholinergic system in haloperidol-induced release ofdopamine from slices of striatum in the rat. Neuropharmacology 29, 55e59.
Zahm, D.S., Brog, J.S., 1992. On the significance of subterritories in the accumbenspart of the rat ventral striatum. Neuroscience 50, 751e767.
Zavitsanou, K., Nguyen, V., Kelly Newell, K., Patrice Ballantyne, P., Huang, X., 2008.Rapid cortico-limbic alterations in AMPA receptor densities after administrationof PCP: implications for schizophrenia. Journal of Chemical Neuroanatomy 36,71e76.
Zavitsanou, K., Katsifis, A., Filomena, M., Huang, X., 2004. Investigation of M1/M4Muscarinic receptors in the anterior cingulated cortex in schizophrenia, bipolardisorder and major depressive disorder. Neuropsychopharmacology 29,619e625.
Zavitsanou, K., Nguyen, V., Han, M., Huang, X., 2007. Effects of typical and atypicalantipsychotic drugs on rat brain muscarinic receptors. Neurochemical Research32, 525e532.
Zhou, D., Apostolakis, E.M., O’Malley, B.W., 1999. Distribution of D5 dopaminereceptor mRNA in rat ventromedial hypothalamic nucleus. Archives ofBiochemistry and Biophysics.
Appendix 1 Brain regions identified using a rat brain atlas(Paxinos and Watson (19.86.)
ACO amy: anterior cortical amygdaloid nucleus,AHIAL: amygdalohippocampal area anterolateral partBl Amy: basolateral amygdaloid nucleusCA1: hippocampal regionCA2: hippocampal regionCA3: hippocampal regionCeA: Central amygadala nucleusCing. Ctx: cingulate cortexCL: centrolateral thalamic nucleusCM: centro medial thalamic nucleusDen: dorsal endopiriform cortexDG: dentate gyrusDtt: dorsal tenia tectaeEnt ctx: entorhinal cortexLent: lateral entorhinal cortexM1 ctx: M1 motor cortexMeAd: medial amygdaloid nucleusMHb: medial habenullaN.Acc core: nucleus accumbens coreN.Acc shell: nucleus accumbens shellOlf. Tub: olfactory tubercle

L.J. Steward et al. / Neuropharmacology 62 (2012) 1554e1563 1563
Pfc: prefrontal cortexPfc 1: prefrontal cortex prelimbic layer IPfc 2: prefrontal cortex prelimbic layer IIeIIIPfc 3: prefrontal cortex prelimbic layer IVeVIPiri. Ctx: piriform cortexPo: posterior orbital cortex
RsGb: retrosplenial cortexStr. d.l: dorsal lateral striatumStr dm: dorsomedial striatumVDb: nucleus of the diagonal band of BrocaVmH: ventromedial hypothalamusVo: ventral orbital cortex





























