Embed Size (px)
Citation preview

Neuroscience Vol. 54, No. I, pp. 37-47, 1993 Printed in Great Britain
0306-4522/93 s6.00 + 0.00 Pergamon Press Ltd
0 1993 IBRO
CO-ACTIVATION OF METABOTROPIC GLUTAMATE AND
N-METHYL-D-ASPARTATE RECEPTORS IS INVOLVED IN MECHANISMS OF LONG-TERM POTENTIATION
MAINTENANCE IN RAT HIPPOCAMPAL CA1 NEURONS
T. BEHNISCH and K. G. F&YMANN*
Department of Neurophysiology, Institute for Neurobiology, P.O.B. 1860, O-3010 Magdeburg, F.R.G.
Abstract-Slices of hippocampal area CA1 in the rat were employed to test the hypothesis that the activation of metabotropic glutamate receptors during tetanization is necessary for the late maintenance of long-term potentiation. If the metabotropic glutamate receptor antagonist L-2-amino-3-phosphono- propionate was present during tetanization, post-tetanic and early long-term potentiation of the population spike as well as field excitatory postsynaptic potential developed almost normally. However, 100 min after tetanization, long-term potentiation of the field excitatory postsynaptic potential decreased in an irreversible manner. The same concentration of D-2-amino-3-phosphonopropionate was ineffective. If L-2-amino-3- phosphonopropionate was applied 120 min after tetanization, it did not influence long-term potentiation. The presence of the metabotropic glutamate receptor agonist rrans-D,L-1-aminocyclopentane-1,3-dicarb- oxylic acid during tetanization weakly enhanced the slope of field excitatory postsynaptic potential long-term potentiation. The influence of L-2-amino-3-phosphonopropionate and D,L-1-amhiocyclopentane- 1,3-dicarboxylic acid on ionotropic glutamate receptors was studied using whole-cell voltage-clamp and pressure application techniques. No effect of L-2-amino-3-phosphonopropionate on either early or late components of excitatory postsynaptic currents could be detected at the concentration used to block long-term potentiation. It is therefore unlikely that the effect of L-2-amino-3-phosphonopropionate on long-term potentiation is due to an interaction with N-methyl-D-aspartate receptors or a-amino-3- hydroxy-S-methyl-4-isoxazole propionic acid receptors. However, bath-applied lS,3R-D,L-l-aminocyclo- pentane-l,3-dicarboxylic acid facilitated the N-methyl-D-aspartate-induced depolarization in response to N-methyl-o-aspartate pressure application in a reversible manner.
These data suggest that besides the involvement of ?v’-methyl-D-aspartate receptors the activation of a 2-amino-3-phosphonopropionate-sensitive metabotropic glutamate receptors during or immediately after tetanization is necessary for subsequent mechanisms responsible for the maintenance of long-term potentiation. A link between metabotropic glutamate receptors and protein kinase C activation during long-term potentiation is discussed considering the similar time course of long-term potentiation blockade after application of L-2-amino-3-phosphonopropionate and protein kinase C inhibitors.
Long-term potentiation (LTP) is a long-lasting increase in the efficacy of synaptic transmission following a brief period of high frequency stimulation of axons. This type of activity-dependent synaptic plasticity is widely used as a model for the investi- gation of elementary mechanisms of learning and memory. Ionotropic glutamate receptors play a sub-
*To whom correspondence should be addressed. Abbreviations : lS,3R-ACPD, lS,3R-D,L-l-aminocyclo-
pentane-1,3-dicarboxylic acid, ACSF, artificial cerebro- spinal fluid; AMPA, a-amino-3-hydroxy-S-methyl- 4-isoxazole propionic acid; AMPA-R, AMPA receptor; AP3, 2-amino-3-phosphonopropionate; AP4, 2-amino- 4-phosphonobutyrate; APS, 2-amino-5-phosphonopen- tanoic acid; EGTA, ethyleneglycoltetra-acetate; EPSCs, excitatory postsynaptic currents; fEPSP, field excitatory postsynaptic potential; HEPES, N-2_hydroxyethylpiper- azine-N’-2-ethanesulphonic acid; IP3, inositol 1,4,5-tris- phosphate; LTP, long-term potentiation; mGlu-R, alias Qp. metabotropic glutamate receptor; NMDA, N-methyl- o-aspartate; NMDA-R, NMDA receptor; PI, polyphos- phoinositide; PKC, protein kinase C; PLC, phospholipase C; PS, population spike; PTX, pertussis toxin; t-ACPD, lrU~-D,L-1-amino-1,3-cyclopentanedicarboxylic acid.
stantial role in both induction and maintenance of LTP at glutamatergic synapses.“,49 The activation of N-methyl-D-aspartate receptors (NMDA-Rs) during tetanus is usually a critical prerequisite for induction, whereas a sensitivity increase in postsynaptic a-amino- 3-hydroxy-S-methyl-4-isoxazole propionic acid recep- tors (AMPA-Rs) could be at least a partial explanation of the expression of LTP. “J’ Furthermore, the prod- ucts of polyphosphoinositide (PI) hydrolysis follow- ing the activation of phospholipase C (PLC), i.e. inositol 1,4,5_trisphosphate (IP3) and diacylglycerol, as well as the subsequent intracellular Ca2+ release and protein kinase C (PKC) activation, seem to be involved in mechanisms regulating synaptic plasticity.6.22’0~35,43~61 Tetanic stimulation caused a high degree of PI turn- over6~6’ and an increase in IP335 in CA3 and dentate gyrus neurons, respectively. Several laboratories have shown that phorbol ester activation of PKC or intracellular injection of PKC itself into hippocampal neurons mimics LTP.22.24 Moreover, PKC inhibitors shorten the duration of LTP in both CA1 and dentate gyrus neurons.23,34.36,50
37

Since activation of Ni’vlDA-R usualIy results only in short-term enhancement,“’ other receptors may be involved in LTP. The receptor involved in PKC activation and subsequent stabilization of LTP has not yet been identi~ed. A likely candidate is the metabotropic glutamate receptor fmGlu-R, alias Qp). Previous pharmacological studies have shown that ibotenate, quisqualate, aspartate and glutamate. but not N-methyl-D-aspartate (NMDA) or kainate, increase the [ ‘Hjnositol monophosphate formation in hippocampal slice~.“‘,~’ From these and other studies, the existence of a new mGlu-R was postulated, which is specifically coupled to hydrolysis of phosphoinosi- tides and subsequent activation of PKC.?.4,:h.37,4’.J7.~~,~q,~~ However, direct evidence for a link of mGlu-R to activation of PKC exists only for cultured striatal neurons.3’ So far, ~-2-amino-3-phosphonopropionate (L-AP3) and fauns- or 1 S,3R-D,L- I -aminocyclo- pentane-1,3-dicarboxyli~ acid (t- and lS,3R-ACPD) are the commonly used antagonist and agonist at mGlu-R, respectively.‘5,47.55.57
Recent evidence from cDNA cloning studies suggests that a whole family of metabotropic glutamate receptors exists (mGlu-Rl~5), di~ering in signal transduction and agonist selectivity.” At least the mGlu-RI and mGlu-R5 subtypes are coupled to a pertussis toxin (PTX)-sensitive G protein, and stimulates hydrolysis of phosphatidylinositol-4,5-b& phosphate by activation of PLC.4.20.h’
At present there is only preliminary evidence for the involvement of the mGlu-R in PKC activation and subsequent stabilization of LTP. Our initial investigations have shown that the non-selective mGlu-R antagonist 2-amino-4-phosphonobutyrate (AP4), which antagonizes ibotenate-induced IP3 increase in hippocampal slices.“’ inhibits the mainten” ante of LTP in a similar manner as seen after inhi- bition of PKC.49.9’ This effect was not stereoselective
l=Ield EPSP
Stimulus Sl electrode
-----/ \
and was also seen ii‘ the drug was apphed aiicr tetanization. The more selective antagonist D,L.-AP.? also blocked LTP.*,‘” However, D,I,-AP~ 1s also it weak NMDA antagonist’” and both AP4 and AP3 were effective even if applied after tetanus,?“.J” an observation difficult to reconcile with the simple notion that mGlu-R are activated by [.-glutamate released during tetanus. Other, still contradictory evidence that mGlu-R may be involved in LTP comes from the effects of the selective agonist t-ACPD. In CA1 neurons, ACPD has been shown to aug~nent tetanus-induced LTP” or to convert short-term potentiation into LTP by a postsynaptic mechanism.’ In addition to these findings, others have even seen an induction of an LTP-like potentiation by simple bath application of ACPD without any tetanus.”
Therefore, in the present study we continued to examine the hypothesis that a metabotropi~ gluta- mate receptor is related to PKC-dependent late stages of LTP using the more selective stereoisomer I_-AP3 and the agonist ACPD. To investigate the possibility that t-AP3 and ACPD also affect ionotropic recep- tors, we studied the influence of both compounds on excitatory postsynapt~c currents (EPSCs) and NMDA pressure-induced currents in the same preparation.
EXPERIMENTAL PROCEDURES
H~ppo~mpal slices were prepared from seven- to eight- week-old Charles River Wistar rats (bred in the institute) as described previously.5o Animals were killed by a stun on the neck. After decapitation and dissection of the hippocampus, 400~pm-thick slices were cut in cold oxygenated physiologi- cal solution (ACSF, in mM: NaCl 124, KC1 4.9, <H;PO, ly2, MeSO, 1.3. CaCI, 2.0. NaHCO, 25.6. o-glucose IO) using a tissue chopper, and alfowed to recover for 1.5 h it 3.% Synaptic responses were elicited by a stainless steel stimulat- ing electrode in the striatum radiatum of the CA1 region {Fig. 1). Glass electrodes (filled with ACSF. R, = 1 4 MC?)
NMDA or ACPD intracellular
I.. . eiectrocie or PS electrode
; .- s!iri-iiJitIS s2
Fig. 1. Diagram of electrode placement for stimulation and intracellular recording in rat hippocampal slices. Either stimulation of Schaffer collateral-commissural pathway, NMDA or r-ACPD pressure
ejection to stratum radiatum were applied.

Long-term potentiation in rat hippocampal neurons 39
-
60: I I I :: : :: :
~l~fk$~~~“‘f’“““~r, -30 0 30 60 so 120 150 150 210 240
-30 0 30 60 9O 120 150 150 210 240
llme (mm) time (mln)
Fig. 2. Effects of 300 PM L-AP3 on the maintenance of the fEPSP slope (A) and PS (B). The tetanization (three 100 Hz trains, 0.5 s each, separated by 2 min intervals) was provided immediately after baseline recording (Omin). Data are plotted as an average of the percentage change from baseline responses. Standard error of the mean (k S.E.M., vertical bars) is shown only for selected intervals. Standard error of the mean (k S.E.M., vertical bars) is shown only for selected intervals. The symbol ( + ) indicates the beginning of the statistically significant difference between control and AP3 groups (Mann-Whitney, P < 0.05). In comparison to control LTP (n = 8, x ) perfusion with 300 PM L-AP3 for 30 min (horizontal bar) significantly reduces the potentiation of the fEPSP (n = 9, 4). In non-tetanized slices, 300 PM L-AP3
(n = 6, A) did not influence test responses. The bar indicates the duration of drug application.
were placed in the apical dendritic and cell body layers Neuroamin, U.K.) were dissolved as 1OmM in equimolar to record field excitatory postsynaptic potentials (fEPSPs) NaOH. and population spikes (PSs), respectively (Fig. 1). The slope-of the initial rising. phase was used as an estimation of the fEPSP. The PS amplitude was evaluated by measuring the voltage difference between the negative peak and the positive one preceding it. Tetanic stimulation used to induce near-maximal LTP under control conditions con- sisted of three 100-Hz trains, each 0.5 s long, separated by 2 min intervals, at twice test strength. Test stimuli (four pulses, 0.2 Hz), originally adjusted to elicit a population spike of 30% of its maximum amplitude, were applied every 10 min, starting 1 min after the third tetanus. All drugs were delivered to the slice for 30 min by the perfusion line (flow rate 1 ml/min), starting 20 min before the first tetanization or 120 min after it. A concentration of 100-300 PM AP3, which inhibits ibotenate- or quisqualate-stimulated [‘Hlinositol monophosphate formation in brain slices by 70-100%,58 was used. AP3 (lots 1 and 2) was added as 10mM water solution to the physiological buffer to yield the final bath concentration. ?-ACPD (Lot 8, Tocris Neuroamin, U.K.) and lS,3R-ACPD (Lot 7, Tocris
Excitatory postsynaptic currents measurements in the whole-cell configuration
The experiments for studying the effects of L-AP3 on EPSCs were performed in hippocampal slices from 18-20- day-old rats perfused continuously with ACSF (in mM: NaCl 125, NaCHO, 25, KC1 2.5, NaH,PG, 1.25, MgCl, 1, CaCl, 2, glucose 25) containing 20 PM picrotoxin. Whole- cell currents were recorded from intact CA1 pyramidal cells in 200-pm-thick slices using the EPC-7 amplifier (List). The patch pipettes (3-5 MR) were filled with a solution con- taining (in mM): CsCl 125, MgCl, 2, CaCl, 2, EGTA 10, HEPES 10, Na-ATP 2 (pH 7.3). The EPSCs were recorded from cells in normal bath solution and thereafter in solu- tions containing 100-300 /JM L-AP3 or 25 PM AP5 using holding potentials of - 70 mV to f40 mV. For data analysis we measured the amplitude of the early EPSC component (non-NMDA component) and, 15ms after peak current, a late EPSC component (more NMDA component). For
A 0’ .
10'
~ll?F
240’ 0
2/r z L 2 ma
control 100 PM L-AP3 300 FM L-APB
0'
control 300 MM L-APB
Fig. 3. Representative examples of averaged fEPSPs (A) and PS (B) from LTP control experiments and from experiments where L-AP3 was present during tetanization.

40 T‘. BEHNIKH and K. G. REVMANN
identifying the late EPSC as the NMDA component we used the NMDA inhibitor 2-amino-5phosphonopentanoic acid (AP5).
N-Methyl-waspartate- and lS,3R-D,L-l-aminoc_vclopentan~,- 1,3-dicarboxylic acid-induced currents in an intracellular recording mode
Intracellular signals were recorded and amplified usmg an npi SEC IL single-electrode current- and voltage-clamp amplifier (“pi, Tamm, F.R.G.). The intracellular electrodes (R, = 40~.50 MQ) were filled with 3 M potassium acetate (pH 7.3). A switching frequency of 18 kHz at a duty cycle of 25% was used. The amplifier headstage was continually monitored to ensure that any voltage developed across the microelectrode tip decayed completely before the next sampling period.@ Membrane currents were recorded at a holding potential of -70 mV. NMDA or ACPD were pressure-ejected into stratum radiatum (500 FM NMDA or 1 mM lS,3R-ACPD, 2-3 MO, I- 2 bar, 20-80ms). The position of the application pipette was justified to obtain a maximal response to NMDA or ACPD. In order to distinguish pure pressure artefacts in some of the NMDA ejection experiments, the NMDA antagonist AP5 (20pM) was added to the bath at the end of the experiment. Before this the effect of drop application of L-AP3 or lS,3R-ACPD into the bath on the micropressure-induced currents was tested.
RESULTS
Efects of 2-amino -3-phosphonopropionate on extra- cellular+ recorded long-term potentiation
Twenty minute perfusion of 300 PM L-AP3 did not influence baseline synaptic transmission (fEPSP slope) and firing (PS amplitude) before the tetanization. Furthermore. 300 PM L-AP3 (n = 6) had no con-
sistent effect on single pulse-evoked potentials in non-
tetanized control slices for at least 4 h. In LTP control experiments, tetanic stimulation produced a 20-60% enhancement of the fEPSP (Fig. 2A, B, Table 1). Following the 30min exposure to L-AP3 in another set of slices, tetanic stimulation produced a smaller potentiation of the fEPSP slope compared with LTP controls (Fig. 2A). Whereas the initial potentiation during the first 60 min was not significantly different. but the late potentiation was irreversibly reduced to the baseline level after 2-3 h (P > 0.05. Mann-- Whitney test).
In contrast to the effects of L-AP3 on fEPSP slope, we found only a weak reduction of the late LTP of PS 100 min after tetanization (Fig. 2B). Similar effects were seen with D,L-AP3. Table I summarizes the changes of fEPSP slope and PS amplitude at various isomers and concentrations (Fig. 3A, B). For ail different experiments a separate set of LTP controls was measured. D,L-AP~ and L-AP3 (100 PM) slightly reduced LTP after 100min but non-significantly (P > 0.005, Mann-Whitney test). Only 300 PM D,L-AP~ decreased fEPSP slope significantly up to the control value. If 300 p M L-AP3 (n = 6) was applied 120min after tetanization for 30min, no changes were observed in potentiation of EPSP slope and PS amplitude (Fig. 4A, B).
4

tude OF LTP fn ~7; Fig* SA, Bj. At the same time no effects of r-ACPD an fEPSP were seen in 8 non-tetzwked S2 input (Fig_ 6)s A cw&ined appka- tion of ISJR-ACPD and NMDA produced only short- and weak kg-lasting potentiations (n = 4; Fig. 73, To &xk the aWity of TS$R-BICPD to produce anathw well known e&x% of this metalro- tropic a&3&&, its inftuence on he afterhypqokriz- ation after a 100 ms depolarizing pulse was tested.

‘I’. BEHNISCH and K. G. REYMAN~
160
NMDA ,AP5
50 min
--If-+
90 110 130 150 170 190 Time(min)
Fig. 7. A combined bath application of IS,3R-ACPD and NMDA resulted in a sequence of depression and short-lasting facilitation (n = 4). APS was applied shortly (< 15 s) after NMDA to prevent excitotoxic
effects. The vertical bars in the graph characterize the standard error of the mean.
In accordance with the literature, ACPD diminished pound on the NMDA-R-induced current. The results the afterhyperpolarization (data not shown). are demonstrated in Fig. 8. In five cells the amplitude
Ejiicts of L-2-amino-3-phosphonopropionate on syn - was measured at different holding potentials between
apt&ally evoked N-methyl-D-aspartate current in the - 70 mV and i-40 mV. The Z-V plot for the peak
whose-dell conjuration amplitude (CNQX-~nsitive non-NMDA ~um~nent) is demonstrated in Fig. 8(a). L-AP3 (100-300 PM) and
The influence of 100-300 p M L-AP3 on EPSCs was AP5 (110 p M) did not influence this early AMPA-R- studied to exclude any possible effects of this com- mediated current (comparison to pre-drug values).
‘I 1 control ‘I I 10 ms
100 PM AP3
10 j&l AP5
Fig. 8. Effect of L-AP3 on early and late component of EPSCs. (A) Representative examples of averaged EPSCs before and after application of L-AP3 (100 and 300 FM) and APS (IOpM). The holding potential was changed from -7OmV up to +4OmV. Right: mean i-V curves for the early CNQX-sensitive component of EPSC (a; peak of responses) and the late, more APS-sensitive component of EPSC (b)
measured 15 ms after peak (n = 5).
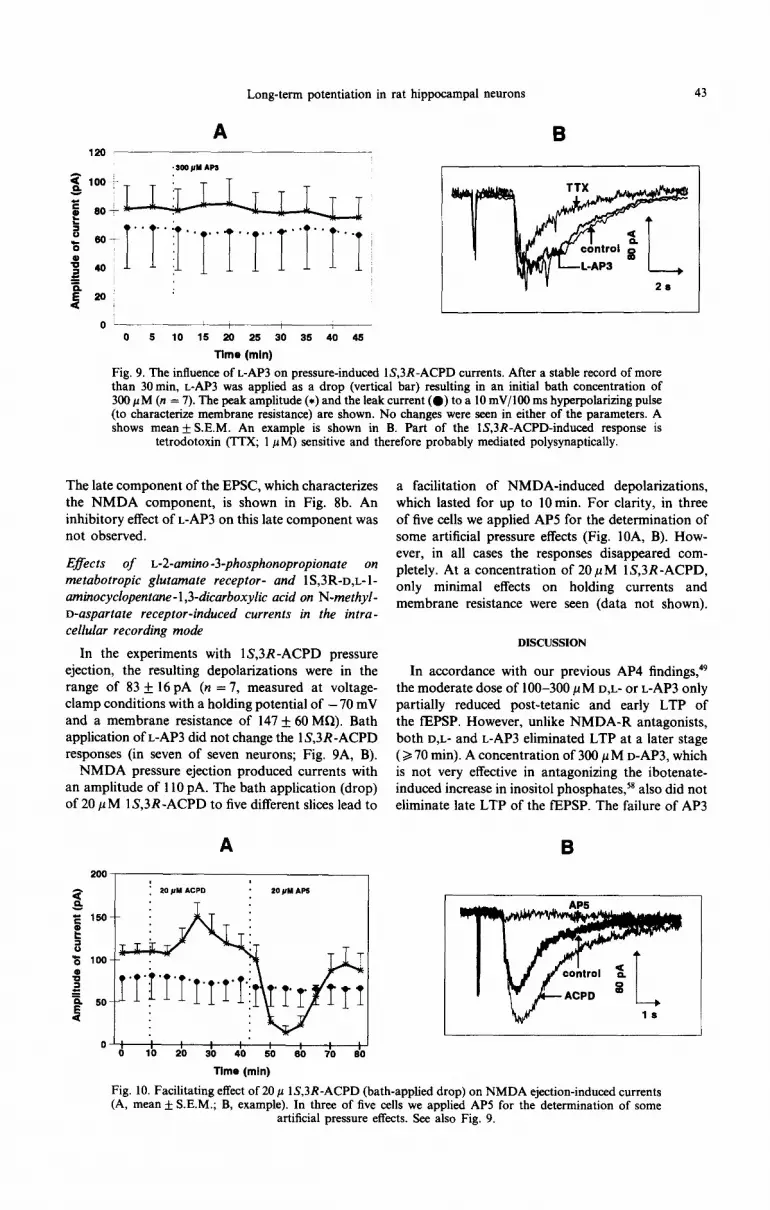
Long-term potentiation in rat hippocampal neurons 43
Fig. 9. The influence of L-AP3 on pressure-induced lS,3R-ACPD currents. After a stable record of more than 30min, L-AP3 was applied as a drop (vertical bar) resulting in an initial bath concentration of 300 PM (n = 7). The peak amplitude (*) and the leak current (0) to a 10 mV/lOO ms hyperpolarizing pulse (to characterize membrane resistance) are shown. No changes were seen in either of the parameters. A shows mean f S.E.M. An example is shown in B. Part of the lS,3R-ACPD-induced response is
tetrodotoxin (TTX; 1 PM) sensitive and therefore probably mediated polysynaptically.
The late component of the EPSC, which characterizes the NMDA component, is shown in Fig. 8b. An inhibitory effect of L-AP3 on this late component was not observed.
Effects of L-2-amino-3-phosphonopropionate on metabotropic glutamate receptor- and lS,3R-D,L-l- aminocyclopentane-1,3-dicarboxylic acid on N-methyl- D-aspartate receptor-induced currents in the intra- cellular recording mode
In the experiments with lS,3R-ACPD pressure ejection, the resulting depolarizations were in the range of 83 + 16 pA (n = 7, measured at voltage- clamp conditions with a holding potential of - 70 mV and a membrane resistance of 147 + 60 MR). Bath application of L-AP3 did not change the 1 S,3R -ACPD responses (in seven of seven neurons; Fig. 9A, B).
NMDA pressure ejection produced currents with an amplitude of 110 pA. The bath application (drop) of 20 p M 1 S,3R -ACPD to five different slices lead to
A
200,
0: : 0 10 20 30 40 so 50
I
70 50
Ttme (mln)
a facilitation of NMDA-induced depolarizations, which lasted for up to 10min. For clarity, in three of five cells we applied AP5 for the determination of some artificial pressure effects (Fig. lOA, B). How- ever, in all cases the responses disappeared com- pletely. At a concentration of 20 ,uM lS,3R-ACPD, only minimal effects on holding currents and membrane resistance were seen (data not shown).
DISCUSSION
In accordance with our previous AP4 findings,49 the moderate dose of 100-300 PM D,L- or L-AP3 only partially reduced post-tetanic and early LTP of the fEPSP. However, unlike NMDA-R antagonists, both D,L- and L-AP3 eliminated LTP at a later stage ( > 70 min). A concentration of 300 p M D-AP3, which is not very effective in antagonizing the ibotenate- induced increase in inositol phosphates,58 also did not eliminate late LTP of the fEPSP. The failure of AP3
B
Fig. 10. Facilitating effect of 20 p lS,3R-ACPD (bath-applied drop) on NMDA ejection-induced currents (A, mean + S.E.M.; B, example). In three of five cells we applied AP5 for the determination of some
artificial pressure effects. See also Fig. 9.

44 I BEHNISCH and K. G. REYMAM
to block early LTP is in accordance with findings 01’ Stanton et al.,‘” but in contrast to the observations of Izumi et ~1.‘~ In the latter study, which was performed in four-week-old rats, D,L-AP3 eliminated LTP immediately after tetanization.
The late NMDA component of the EPSC in CA1 pyramidal cells was not affected by L-AP3. Consider- ing this finding, as well as the failure of moderate doses of AP3 to influence NMDA binding,‘8 the partial reduction of the early LTP (< 1 h) by L-AP3 cannot be explained by side effects at the NMDA receptor.
The incomplete and non-significant blockage of spike potentiation after L- and D,L-AP3 differs from those recorded in a recent report by Izumi et ul. with D,L-AP~ and our previous data with AP4.‘8.4y This
might be due to a second mechanism for spike potentiation (so-called E/S potentiation), which is pronounced after repeated tetanization and is also insensitive to NMDA antagonists,” and under certain conditions to protein synthesis inhibitors too.44 In contrast to Izumi et ~l..‘~ we used triple tetanization.
There is convincing evidence for a contribution of a transient activation of PKC to the maintenance phase of LTP.23~““~So The late effects of both D,L- and L-AP3 in the present experiments exhibit the temporal par- ameters observed for the late, PKC-dependent phases of LTP.34 Therefore, the present data are consistent with the view that, in the CA1 region, I_-AP3 prevents PKC activation during LTP by interacting with an mGlu-R. Moreover, it seems unlikely that the diacyl- glycerol necessary to activate PKC originates from sources other than the PLC-coupled mGlu-R. mGlu-R must not necessarily be involved in LTP mechanisms
at other synapses, since LTP in the fimbria-CA3 pathway cannot be blocked by AP3.”
Previous investigations have shown that AP4 and AP3 block the late stages of LTP when they are applied several minutes after tetanization.‘x.s” Accord- ing to the present data, application of L-AP3 120 min after tetanization had no effect on LTP maintenance. Therefore, a time window of several minutes may exist for activation of metabotropic receptors, similar to activation of PKC.J” The effectiveness of the antag- onists, even if applied shortly after the tetanus, is an observation difficult to reconcile with the simple notion that mGlu-Rs are activated by L-glutamate released during tetanus. If the receptor interaction is not unusually long, one has to take into account that AP3 perhaps prevents glutamate-stimulated phosphoinositide hydrolysis at a level beyond the receptor. Independent of a possible involvement of a metabotropic receptor in the induction of those processes enabling LTP maintenance (late phases). the role of mGlu-Rs might not be restricted to the induction process. In a previous study we observed an increase in effectiveness of the mGlu-R in stimulating phosphoinositide hydrolysis several hours after tetanization.’
Studies with the agonist ACPD have also recently supported the involvement of an mGlu-R in LTP.
However, it is still a matter of debate whether ACPD alone can induce long-lasting potentiation. In some studies, t-ACPD only enhances the magnitude and prolongs duration of tetanus-induced LTP,‘,“ whereas in others t - or 1 S,3R-ACPD enhance excitatory trans- mission in the short term” or even permanently.“,“” Our results with I-ACPD rather support the former observations. Another study in which a concomitant activation of NMDA-R and mGlu-R activation. together with low frequency presynaptic stimulation. resulted in LTF’ is in better accordance with findings that NMDA application provokes short-term poten- tiation” and that both NMDA-R antagonists and AP3 prevent late maintenance of LTP in the CA 1 region’“,”
(present results). Although lS,3R-ACPD was effective in our hands in reducing the slow after hyperpolariz- ation it did not result in a convincing long-lasting potentiation, even if applied together with NMDA. Taking into account that dopamine antagonists also block late LTP,14 we postulate that the activation of both NMDA-R and mGlu-R is a necessary but not sufficient condition and that non-glutamate receptors must also be activated during tetanization to establish a real long-lasting potentiation.
The facilitating effect of ACPD on LTP is likely to be due, in part, to the ability of ACPD to potentiate responses mediated by NMDA receptors” through a mechanism which may involve PKC?” or another second messenger. ” The facilitation of NMDA- induced currents by lS,3R-ACPD in the present experiments extends these observations. It seems unlikely, however, that AP3 prevents late LTP simply by a mechanism or reduction of mGlu-R-mediated NMDA-R up-regulation, since the NMDA-R- dependent early LTP was not significantly reduced in the present experiments. Moreover, the facilitation of LTP by ACPD was shown not to be sensitive against PKC inhibitors. 4’ At present it is not known
whether the ACPD-induced enhancement of LTP can be blocked by AP3 and whether it is mediated via inositol polyphosphates. Taken together, both the failure of PKC inhibitors to block ACPD-induced facilitation and the early onset of the effect (already early LTP is enhanced), one can speculate that this
agonist additionally activates another subtype of mGlu-R not coupled to PLC.
As already mentioned. activation of an mGlu-R stimulates a dual pathway, leading to both PKC trans- location and IP3 production.‘,4’.47,55 Indeed, inhibitors of G proteins, of intraneuronal Ca’ + release” and of calmodulin block LTP of CA1 neurons faster than PKC inhibitors.‘h.43.50
t-ACPD has been reported to inhibit the normal synaptic transmission in hippocampal CA 1 neurons of immature rats, probably through a presynaptic mGlu-R controlling glutamate release.7.‘5.45 However, in the CA1 region of adult rat, we and others” have not observed a clear reduction of control EPSPs. However, the mGlu-R antagonized by AP3, which is necessary for the late LTP as shown by the present

Long-term potentiation in rat hippocampal neurons 45
data, seems to be localized postsynaptically. At least a PKC inhibitor prevents both LTP maintenance and increase in postsynaptic AMPA-R sensitivity.s1 Other, presumably postsynaptic, effects of ACPD were also described in CA1 pyramidal cells, for example depolarization, which is generated by an inhibition of K+ conductance and a depression of calcium- activated K+ currents (ix, ca~AHP~).12~21~62 Both ACPD- induced depolarization and block of slow after- hyperpolarization could not be prevented by AP3. It is possible, however, to prevent the inhibition of slow afterhy~~ola~~tion with the protein kinase inhibitor H-7.3 It seems likely, therefore, that perhaps three different mGlu-Rs are present in the CAl: one presynaptic, one postsynaptic with fast ionotropic consequences and one AP3-sensitive involved through the PLC cascade in slow regulation of AMPA-R transmission. It is as yet unknown to what extent and under which conditions an mGlu-R is involved in normal synaptic transmission in hippocampal CA1 neurons (for CA3 see Ref. 10).
The mGlu-R subtypes involved in LTP could be mGlu-RI or mGlu-R5, since to date they are the only members of the mGlu-R family coupled to the phosphoinositide second messenger system6s and since LTP in the CA1 can also be blocked by PTXib (however, see Ref. 27). However, mGlu-Rl expression in CA1 is rather weak,39 although a new study demonstrates the occurrence of mGlu-Rlol in post- synaptic CA1 dendritic shafts and spines.38 More-
over, AP3 and AP4, effective antagonists reported for LTP28*49 (present data) and glutamate-stimulated PI hydrolysis in brain slices,“,58 showed no clear effect in mGlu-RI transfected Chinese hamster ovary (CHO) cells.4 On the other site mGlu-R2-4 are not linked to the IP3/Ca2+ signal transduction, at least in the Xenopm oocyte expression system.65 Therefore, it seems possible that another G protein-coupled, AP3- sensitive subtype of mGluR could be involved in the initialization of mechanisms enabling the late main- tenance of LTP. It also remains to be found whether the metabotropic receptors involved in hippocampal LTP are the same or not as those involved in mechanisms of cerebellar long-term depression,33 visual cortical plasticity I3 and hypoxic-excititoxic damage.32~47~”
CONCLUSION
The stereoselective effect of L-AP3 in CA 1 neurons suggests the involvement of an AP3-sensitive mGlu-R in mechanisms of LTP maintenance, in addition to the obligatory NMDA-R activation. It remains to be determined, however, whether C kinase activation during LTP is actually blocked in the presence of AP3, and to which subtype of the mGlu-R family it is linked.
Acknowledgements-We would like to thank Prof. W. Singer for his support. This study was supported by a research grant from the Deutsche Forschungsgemeinsehaft, Re 847/1-i.
REFERENCES
1. Akers R. F., Lovinger D. M., Colley P. A., Linden D. J. and Routtenberg A. (1986) Translocation of protein kinase C activity may mediate hippocampal long-term potentiation. Science 231, 587-589.
2. Aniksztejn L., Otani S. and Ben-Ari Y. (1992) Quisqualate metabotropic receptors modulate NMDA currents and facihtate induction of long-term ~tentiation through protein kinase C. Eur. J. ~~~o~cj. 4, 500-505.
3. Anwyl R. (1992) Metabotropic glutamate receptors: el~trophysiologi~l properties and role in plasticity. Rev. Neurosci. 3. 217-237.
4. Aramori I. and Nakanishi S. (1992) Signal transduction and pharmacological characteristics of a metabotropic glutamate receptor, mGluR1, in transfected CHO cells. Neuron 8. 757-765.
5. kronica E., Frey U., WagnerM., Schrijder H., Krug M., Ruthrich H., Catania M. V., Nicoletti F. and Reymaun K. G. (1992) Enhanced sensitivity of “metabotropic” glutamate receptors after induction of long-term potentiation in rat hippocampus. J. Neurochem. 57, 376-383.
6. Bar P. R., Wiegant F., Lopes da Silva F. H. and Gispen W. H. (1984) Tetanic stimulation affects the meta~lism of phosphoinositides in hippocampal slices. Brain Res. 321, 381-385.
7. Baskys A. and Malenka R. C. (1991) Agonist at metabotropic glutamate receptors presynaptically inhibit EPSCs in neonatal rat hippocampus. J. Physiol. 444, 687-701.
8. Behnisch T., Fjodorow K. and Reymann K. G. (1991) L-2-Amino-3-phosphonopropionate blocks late synaptic long-term potentiation. NeuroReport 2, 386-388.
9. Bortolotto Z. A. and Collingridge G. L. (1992) Activation of glutamate metabotropic receptors induces long-term potentiation. Ear. J. Pharmac. 214, 297-298.
10. Charpak S., Gahwiler B. H., Do K. Q. and KnBpfel T. (1990) Potassium conductances in hippocampal neurons blocked by excitatory amino-acid transmitters. Nature 347, 765-767.
11. Collingridge G. L., Herron C. E. and Lester R. A. J. (1988) Synaptic activation of N-methyl-o-aspartate receptors in the Schaffer collateral-commisural pathway of rat hippocampus. J. Physiof., Land. 399, 288-300.
12. Desai M. A. and Conn J. (1991) Excitatory effects of ACPD recentor activation in the hinnocamous are mediated bv direct effects on pyramidal cells and blockade of synaptic inhibition. J. Neurophysiol. &,‘40-52:
13. Dudek S. M. and Bear M. F. (1991) A biochemical correlate of the critical oeriod for svnaotic modification in the visual cortex. Science 244, 673-675.~ ’
‘ - 1
14. Frey U., Matthies H., Reymann K. G. and Matthies H.-J. (1991) The effect of dopaminergic D, receptor blockade during tetanization on the expression of long-term potentiation in the rat CA1 region in vitro. Neurosci. Lett. 129, 111-114.
15. Garaschuk 0. V., Kovalchuk Y. N. and Krishtal 0. A. (1992) Trans-ACPD selectively inhibits excitability of hippocampal CA1 neurones. Eur. J. Pharmac. 212, 305-306.

I HEHNSC.H ;trd K. G. RLYMANN 46
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
Goh J. W. and Pennefather P. S. (1990) Pcrtussis toxin prevents induction of hippocampal long-term potrntiation rn the stratum radiatum and stratum oriens inputs to CA1 neurons. Brtrin lies. 511, 3455348. Harvey J. and Colhngridge G. L. (1992) A Cal’ dependent process mediates 1~,3R-ACPD-induced potentl~~ti~~l~ ,>t‘ NMDA responses in rat hippocampus. .S’UC,. ~Veurosci. AFstr. 18, 1218. Harvey J. and Collingridge G. L. (1992) Thapsigargin blocks the induction of long-term potentiation in rat hippocampai slices. Neurosci. Left. 139, 197 200. Harvey J., Frenguelli B. G., Sunter D. C.. Watkins J. C. and Colhngridge G. L. (1991) The actions of IS,ZR-ACPD. a glutamate metabotropic receptor agonist. in area CAI of rat hippocampus. Br. J. Pharmuc. 104, 75P. Houamed K. M., Kuijper J. L.. Gilbert T. L., Haldeman A., O’Hara P. J., Mulvihill E. R., Almers W. and Hagen F. S. (1991) Cloning, expression, and gene structure of a G protein-coupled glutamate receptor from rat brain. Scir~lce 252, 13 1% I32 1. Hu G.-Y and Storm J. F. (1991) Excitatory amino acids acting on metabotropic glutamate receptors broaden the action potential in hippocampal neurons. Brain Res. 568, 339- 344. Hu G.-Y., Hvalby O., Walaas S. I., Albert K. A., Skjeflo P., Andersen P. and Greengard P. (1987) Protein kmase C injection into hippocampal pyramidal cells elicits features of long term potentiation. Nature 328, 426.-429. Huang Y.-Y., Colley P. A. and Routtenberg A. (1992) Postsynaptic then presynaptic protein kinase C activity may be necessary for long-term potentiation. Neuroscience 49, 819.,827. Hvalby 0.. Reymann K. and Andersen P. ( t 988) Intracellular analysis of potentiation of CA 1 hippocampal synaptic transmission by phorbol ester application. Expl Brain Rer. 71, 588-596. Irving A. J., Schotield J. G. Watkins J. C., Sunter D. C. and Coliingridge G. L. (1990) IS.3R-ACPD stimulates and L-AP3 blocks Cal’ mobilization in rat cerebellar neurons. Eur. J. Pharmac. 186, 3633365. lshida M., Akagi H., Shimamoto K.. Ohfune Y. and Shinozaki H. (1990) A potent metabotropic glutamate receptor agonist: electrophysiological actions of a conformationally restricted glutamate analogue in the rat spinal cord and Xenopus oocyte. Bruin Rex 537, 3 1 I 3 14. Ito 1.. Okada D. and Sugiyama H. (1988) Pertussis toxin suppresses long-term potentiation of hippocampai mossy fiber synapses. Neurosci. Lett. 90, 181~ 185. Izumi Y.. Clifford D. B. and Zorumski C. F. (1991) ~-Amino-3-phosphonopropionate blocks the induction and maintenance of long-term potentiation in rat hippocampal slices. Neurosci. Left. 122, 187 191. Katsuki H., Saito H. and Satoh M. (1992) The involvement of muscarinic, a-adrenergic and metabotropic glutamate receptors in long-term potentiation in the fimbriaaCA3 pathway of the hippocampus. Neurosci. Left. 142, 249-525. Kauers J. A.. Malenka R. C. and Nicoll R. A. (1988) NMDA application potentiates synaptic transmission in the hippocampus. Nature 334, 250-252. Kelso S. R., Nelson T. E. and Leonard J. P. (1992) Protein kinase C-mediated enhancement of NMDA currents by metabotropic glutamate receptors in Xenopus oocytes. J. Physiol. 449, 705-718. Koh J.-Y., Palmer E. and Cotman C. W. (1991) Activation of the metabotropic glutamate receptor attenuates N-methyl-u-aspartate neurotoxicity in cortical cultures. Proc. natn. Acad. Sci. U.S.A. 88, 9431 -9435. Linden D. J., Dickinson M. H.. Smeyne M. and Conner J. A. (1991) A depression of AMPA currents in cultured cerebellar Purkinje neurons. ,‘Vrurotr 7. 81 89. Lovinger D. M., Wong K. L.. Murakami K. and Routtenberg A. (1987) Protein kinase C inhibitors eliminate hippocampal long-term potentiation. Bruin Rex. 436, 177- 183. Lynch M. A., Clements M. P., Errington M. L. and Bliss T. V. P. (1988) Increased hydrolysis of phosphatidyiinosito~- 4,5-bisphosphate in long-term potentiation. ~~ur~sci. Lett. 84, 291-296. Malinow R., Madison D. V. and Tsien R. W. (1988) Persistent protein kinase activity underlying long-term potentiation. Nature 335, 820-824. Manzoni 0. J. J.. Finiels-Marlier F.. Sassetti I., Blockaert J., le Peuch Ch. and Sladeczek F. A. J. (1990) The glutamate receptor of the Qp-type activates protein kinase C and is regulated by protein kinase C. Neurosci. Lett. 109, 146-l 5 1. Martin L. J., Blackstone C. D.. Huganir R. L. and Price D. L. (1992) Cellular localization of a metabotropic glutamate receptor in rat brain. Neuron 9, 2599270.
39, Masu M., Tanabe Y., Tsuchida K., Shigemoto R. and Nakanishi S. (1991) Sequence and expression of a metabotropic
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
glutamate receptor. Nature 349, 760.-765. Matthies H., Behnisch T., Kase H., Matthies H. J. and Reymann K. G. (1991) Differential effects of protein kinase inhibitors on pre-established long-term potentiation in rat hippocampal neurons in vitro. Neurusci. Letf. 121, 259-262. McGuinnes N., Anwyl R. and Rowan M. (1991) T-ACPD enhances long-term potentiation in the hippocampus. Eur. J. Phar. 197, 231 -232. Nicoletti F., Iadarola M. J., Wroblewski J. T. and Costa E. (1986) Excitatory amino acid recognition sites coupled with inositol phospholipid metabolism: developmental changes and interaction with I-adrenoceptors. Prctc. nuln. Acaci. Sri. U.S.A. 83, 1931-1935. Obenaus A., Mody 1. and Bairnbridge K. G. (1989) Dantrolene-Na (Dantrium) blocks induction of long-term potentiation in hippocampal slices. N&msci. Letr. 98, 172.. 178. Otani S.. Marshall C. J.. Tate W. P.. Goddard G. V. and Abraham W. C. 11989) Maintenance of long-term uotentiation . I I . in rat dentate gyrus requires protein synthesis but not messenger RNA synthesis immediately post tetanization. Neuroscience 28, 5 199526. Pace& J. G. and Kelso S. R. (1993) T-ACPD reduces muitipfe components of synaptic transmission in the rat hippocampus. Neurmci. Left. (in press). Palmer E., Monaghan D. T. and Cotman C. W. (1988) Glutamate receptors and phosphoinositide metabolism: stimulation via quisqualate receptors is inhibited by N-methyl-D-aspartate receptor activation. M&c. Brain Res. 4, 161-165. Palmer E., Monaghan D. T. and Cotman C. W. (1989) Trans-ACPD, a selective agonist of the phosphoinositide- coupled excitatory amino acid receptor. Eur. J. Pharmac. 166, 5855587. Radpour S. and Thomson A. M. (1992) Synaptic enhancement induced by NMDA and Q, receptors and presynaptic activity. Neurosci. Lelt. 138, 119 ~122. Reymann K. G. and Matthies H. (1989) Z-Amino-4-phosphonobutyrate selectively eliminates iate phases of long-term potentiation in rat hippocampus. Neurosci. Lett. 98, 1666171.

Long-term potentiation in rat hippocampal neurons 41
50. Reymann K. G., Broedemann R., Kase H. and Matthies H. (1988) Inhibitors of calmodulin and protein kinase C block different phases of hippocampal long-term potentiation. Brain Res. 461, 388-392.
51. Reymann K. G., Davies S. N., Matthies H., Kase H. and Collingridge G. L. (1990) Activation of a K-252b-sensitive protein kinase is necessary for a post-synaptic phase of long-term potentiation in area CA1 of rat hippocampus. Eur. J. Neurosci. 2, 481-486.
52. Reymann K. G., Frey U., Jork R. and Matthies H.-J. (1988) Polymyxin B, an inhibitor of protein kinase C., prevents the maintenance of synaptic long-term potentiation in hippocampal CA, neurons. Brain Res. 440, 305-314.
53. Reymann K. G., Matthies H., Schulzeck K. and Matthies H. (1989) N-Methyl-u-aspartate receptor activation is required for the induction of both early and late phases of long-term potentiation in rat hippocampal slices. Neurosci. Lerr. 96, 96-101.
54. Sacaan A. I. and Schoepp D. D. (1992) Activation of hippocampal metabotropic excitatory amino acid receptors leads to seizures and neuronal damage. Neurosci. Left. 139, 7782.
55. Schoepp D., Bockaert J. and Sladeczek F. (1990) Pharmacological and functional characteristics of metabotropic excitatory amino acid receptors. Trends pharmac. Sci. 11, 508-515.
56. SchoeDD D. D. and Johnson B. G. (1991) Selective inhibition of excitatorv amino acid-stimulated nhonhoinositide hydrolysis in the rat hippocampus by activation of protein kinase C. Biociem. Pharmac. 37, 4299-4305:
57. Schoepp D. D., Johnson B. G., True R. A. and Monn J. A. (1991) Comparison of (lS,3R)-1-aminocyclopentane-1,3- dicarboxylic acid (lS,3R-ACPD)- and lR,3S-ACPD-stimulated brain phosphoinositide hydrolysis. Eur. J. Pharmac. 207, 351-353.
58. Schoepp D. D., Johnson B. G., Smith E. D. R. and McQuaid L. A. (1990) Stereoselectivity and mode of inhibition of phosphoinositide-coupled excitatory amino acid receptors by 2-amino-3-phosphonopropionic acid. Molec. Pharmac. 38, 222-228.
59. Sladeczek F., Reacasens M. and Bockaert J. (1988) A new mechanism for glutamate receptor action: phosphoinositide hydrolysis. Trends Neurosci. 11, 545-549.
60. Stanton P. K., Chattarji S. and Sejnowski T. J. (1991) 2-Amino-3-phosphonopropionic acid, an inhibitor of glutamate-stimulated phosphoinositide turnover, blocks induction of homosynaptic long-term depression, but not potentiation, in rat hippocampus. Neurosci. Left. 127, 61-66.
61. Stelzer A., Feasey K. J., Moneta M. E., Sincini E., Bruggencate G. and Noble E. P. (1989) Inositol l-phosphate formation in long-term potentiation and kindling. Bruin Res. 490, 41-47.
62. Stratton K. R., Worley P. F. and Baraban J. M. (1990) Pharmacological characterization of phosphoinositide-linked glutamate receptor excitation of hippocampal neurons. Eur. J. Pharmac. 186, 357-361.
63. Sugiyama H., Ito I. and Hirono C. (1987) A new type of glutamate receptor linked to inositol phospholipid metabolism. Nature 325, 531-533.
64. Sutor B. and Hablitz J. J. (1989) EPSPs in rat neocortical neurons in vitro. I. Electrophysiological evidence for two distinct EPSPs. J. Neurophysiol. 61, 607-620.
65. Tanabe Y., Masu M., Ishii T., Shigemoto R. and Nakanishi S. (1992) A family of metabotropic glutamate receptors. Neuron 8, 159-179.
66. Zheng F. and Gallagher J. P. (1992) Metabotropic glutamate receptors are required for the induction of long-term potentiation. Neuron 9, 163-172.
(Accepted I December 1992)





























