Embed Size (px)
Citation preview

Conservation and Management of a Threatened Madagascar Palm Species, Neodypsis decaryi, Jumelle JOELISOA RATSIRARSON,*~ JOHN A. SILANDER, JR.,* AND ALISON F. RICHARDt *Department of Ecology and Evolutionary Biology, 75 N. Eagleville Road, The University of Connecticut, Storrs, c r 06269, U.S.A. tYale School of Forestry and Environmental Studies, 205 Prospect Street, New Haven, CT 06520, U.S.A. ,Current Address: Yale School of Forestry and Environmental Studies, 205 Prospect Street, New Haven, CT 06520, U.S.A.
Abstract: The dynamics o f a remnant population o f the palm Neodypsis decaryi were characterized using a linear, stage-structured demographic model. This palm is a threatened keystone species restricted to a narrow ecological zone in southeastern Madagascar. The population showed high mortality rates in the early stages o f the life cycle, followed by a period o f lower adult mortality. Demographic results indicated that the popula- tion was either stable or increasing inside the r e s e r v e (A m close to 1.0). Sensitivity and elasticity analyses indi- cated that adult stage classes were the most sensitive to producing changes in population growth rates. Con- tinued biological monitoring is appropriate for the long-term conservation management o f this palm inside the reserve. Outside the protected area the population appears to be declining rapidly, mainly because o f an- thropogenic effects (especially f lre and grazing). I f the conservation of Neodypsis decaryi is to be successful in the long term, then conservation methods must be carried out in cooperation with local villagers, and the spe- cies should be managed as a renewable resource in situ and ex sit~ In situ conservation should include culti- vation o f this species throughout its natural range and protection o f the existing viable natural populations. Alternative conservation management practices, both ecologically and economically sustainable, may be use- fu l to alleviate the human pressures on this renewable resource. I f leaves of N. decaryi are to be harvested by local people, we recommend restricting annual harvesting to about 25% of leaves per tree per year. Based on sensitivity analysis, seed collection should be kept well below 95% o f the yearly crop i f harvesting is not to have a significant impact on population growth rates.
Conservaci6n y manejo de la especie de palmera de Madagascar en peligro Neodypsts decaryi, Jumelle
R e s u m e n : Se caracteriz6 la din~mica de las poblaciones remanentes de la palmera Neodypsis decaryi usando un modelo demogr~fico lineal estructurado por edades. Esta palmera es una especie clave en peligro restringida a una zona ecol6gtca angosta en el sudeste de Madagascar. La poblaci6n mostr6 altas tasas de mortalidad durante los estadfos tempranos del ciclo de vida seguidas de tasas mdm bajas para los estadios adultos. Los resultados demogrd~ficos indicaron que dentro de la reserva la poblaci6n permaneci6 estable o aument6 (A m cercano a 1.0). Los an~lisis de sensitividad y elasticidad indicaron que las classes correspon- dientes a los estadios adultos fueron las mdts sensibles siendo las principales responsables de cambios en los tasas de crecimiento poblacional. E1 monitoreo biol6gico continuado para los esfuerzos de manejo para la conservaci6n a largo plazo de esta palmera dentro de la reserva es apropiado. Fuera del d~rea protegida, la poblaci6n parece estar declinando rapidamente debido, principalmente, a los efectos antropog~nicos (espe- cialmente fuego y pastoreo). Para que la conservaci6n de Neodypsis decaryi sea exitosa la largo plazo, los m#todos de conservaci6n deben ser Ilevados a cabo en cooperaci6n con la poblaci6n local y la especie debe ser manejada como un recurso renovable tantoen en el lugar como fuera. La conservaci6n en el lugar debe incluir el cultivo de esta especie a lo largo de su dLrea natural de distribuci6n y la protecci6n de las poblacion- ens naturales viables existentes. Pr~cticas alternativas de manejo para la conservaci6n, sostenibles tanto econ6mica como ecol6gicamente, podrlan ser atiles para altviar las presiones humanas sobre este recurso
Paper submitted August 31, 1994; revised manuscript accepted February 17, 1995.
4O
Conservation Biology, Pages 40-52 Vo|ume 10, No. 1, February 1996

Ratsirarson et al. Conservation ofNeodypsis decaryi 41
renovable. Si las hojas de N. decaryi van a ser cosechadas por la poblaci6n local, se recomienda que la co- secha anual se restrinja a un 25% de hojas por drbol por ago. De acuerdo al andlisis de sensittvidad, la colec- ci6n de semtllas debe mantenerse bien por debajo de195% de la producci6n anual para que la cosecha no tenga un impacto significativo sobre las tasas de crecimiento poblacional.
Introduction
The conservation of biological diversity in the tropics is urgently needed because of high rates of habitat destruc- tion (Myers 1986, 1991; Murphy & Lugo 1986; Janzen 1988; Wilson 1988; World Resources Institute 1992). Many formerly c o m m o n species are declining and be- coming rare, threatened, or endangered. Novel conser- vation strategies and management practices for these species need to be developed, other than simply setting aside parks and protected areas (Frankel & Soul6 1981; Soul6 1986). We need to enhance our ability to predict and understand the population dynamics of rare, endan- gered, or keys tone species and to determine the best way to manage and preserve them inside (and outside) their natural habitats (Soul~ 1986; Terborgh 1986; Bawa & Ashton 1991; Du Puy et al. 1992). Management and conservation efforts focusing on keystone species are critical to maintaining the structure of natural biological systems (Terborgh 1986; Mills et al. 1993). The disap- pearance of these species may have a cascading effect, changing the competi t ive relationships and the relative abundance of other species in the communi ty (Howe & Westley 1988).
Demographic approaches have been useful in under- standIng the regulation of population fluctuations and are important in designing management strategies for rare or endangered species (Menges 1986; 1990; Soul6 1986). Demographic studies also provide insight into the response of population growth rates to changes in vital rates (survivorship, growth, and fecundity). This in- formation is useful In evaluating alternative management strategies for rare or endangered species in s i tu and ex
s i tu (Menges 1986). Projection matrices (Leslie 1945; Lefkovitch 1965) have been effectively used as tools for demographic analyses of organisms with complex life cycles (Caswell 1989). Several studies have successfully demonstrated the utility of projection matrices for exam- ining the effect of life-history parameters on population growth (Silvertown et al. 1993).
The triangle palm, Neodypsis decaryi Jumelle, is a threatened species restricted in southeastern Madagas- car to a narrow zone (about 20 km wide) be tween the humid rain forest of the southeast and the dry spiny for- est of the southwest (Ratsirarson & Silander 1995b). Neodypsis decaryi is important both economically and ecologically and not only provides a variety of resources for local people (food, fiber, building materials, Income
source) but also has the attributes of a keystone species because it provides food for animals in the ecological communi ty during critical periods of the year (Ratsirar- son 1993a; Mills et al. 1993; Ratsirarson & Silander 1995a). In many areas throughout the range of Neodyl~
sis decaryi, palm leaves are intensively harvested by lo- cal people for thatching, and fruits are heavily collected for food and revenue sources (Ratsirarson 1993a). The management and conservation of Neodypsis decaryi re- quire an understanding of the effects of leaf removal and fruit collection on individual plant performance and population dynamics.
The purpose of this study was to examine the popula- tion dynamics and demographic structure of the Neo-
• dypsis decaryi. The study also focused on the effects of varying levels of leaf and fruit harvesting on survival, growth, and reproduction. This information is useful in developing a management strategy for this species as a sustainable resource.
Like many palm species, Neodypsis decaryi offers an advantage in demographic studies because of its mono- podial growth form and the presence of leaf scars along the trunk (Pifiero et al. 1984; Ulh & Dransfield 1987). Each senescent leaf leaves a scar on the trunk and, if the rate of leaf product ion over t ime is determined, an indi- vidual's age can be estimated (Corner 1966). Demo- graphic data (growth, survival, fecundity) were col- lected for the population, and a matrix model was used to project the palm dynamics over time. Sensitivity anal- yses were used to determine the number of fruit that could be harvested sustainably from the population. A defoliation exper iment was performed to determine the effect of leaf harvest on palm populations.
Methods
Study Site
The field site was located in the third of the three non- contiguous parcels (25001 ' S 46o09 ' E) within the pro- tected reserve of Andohahela in southeastern Madagas- car. T41is parcel, about 500 ha, was established in 1939 to protec t the main viable populat ion of Neodypsis de-
caryi (Nicoll & Langrand 1989). Today it contains the only substantial population throughout the species range in which significant regeneration is occurring and a complete range of age and size classes can be found
Conservation Biology Volume 10, No. 1, February 1996
42 Conservation ofNe0dypsis decaryi Ratsirarson et al.
(Ratsirarson 1993b). Some successful recruitment also occurs in several small, isolated populations of Neodyp-
sis along ephemeral watercourses in the second parcel of the reserve. In extant populations elsewhere, seed- ling and juvenile stages are rare or absent.
The soils in the reserve are low in pH, shallow, and composed of lateritic clays with mixed hydroxides of aluminum and iron (Oxisols) (Jenkins 1987; Nicoll & Langrand 1989). The mean annual precipitation is 800 mm, with a marked dry season of up to 7 months (April- October) and a mean annual temperature of 22°C (Na- tional Weather Service, unpublished data). The emer- gent canopy is dominated by Neodyps is decaryi , and the subcanopy layer is composed mainly of shrubby mem- bers of families well adapted to the drought conditions (with the presence of latex, small leaves, thorns, over- developed root systems, etc.), including Apocynaceae, Didieraceae, Euphorbiaceae, Leguminosaceae, and Rubi- aceae (Nicoll & Langrand 1989; Eboroke 1994; Ratsirar- son, unpublished data).
Demographic Studies
Three permanent plots (A, B, C), each of 0.9 ha, were chosen inside the protected reserve to span a range of slopes and aspects (hence light availabilities) and densi- ties of palms (Ratsirarson 1993b). All individual palms inside the plots were labeled in February 1990. To de- velop a full demographic characterization, measure- ments were taken on each individual. These included leaf number, leaf size (length) of the two oldest live
leaves (for seedlings only), height of the visible stem to the base of the petioles of the living leaves (for juveniles and adults), diameter at breast height (dbh) for adult in- dividuals, and total number of leaf scars on the trunk (for juveniles and adults) (Table 1). Age (A) of stemmed individuals was estimated from the total number of leaf scars (N) left on the trunk, the number of live leaves present (n), the leaf production rate (/) of an individual, and the time (a) required for a new seedling to produce a visible stem (a = 12 years on average based on palm plantation data (Ratsirarson 1993b), following Corner's (1966) formula: A = ((n + l ~ / l + a). Fruit production was determined by counting the number of mature fruits per adult.
To construct life tables, demographic parameters in- cluding survivorship, growth, and reproduction were es- timated from observations on marked individuals in the population. Information on survivorship came from cen- suses carried out every 6 months for seedlings and juve- niles and annually for mature individuals. The census pe- riod was between February 1990 and February 1993. Individual growth was determined by counting the num- ber of new leaves produced per year. The youngest leaf was tagged on each individual palm at the beginning of the study, and all new leaves produced were recorded at each census. Each oew leaf produced by individuals with a visible trunk represented a stem-height increment of 2 cm on average, and the total height growth of the trunk over time was estimated from the number of new leaves produced (Ratsirarson 1993b).
The use of stage-structured models to project popula-
Table 1. Measurement attributes for Neodypsis decaryt in Madagascar.
Development Stage*
Attribute Measurement Adults Juveniles Seedlings
Number of leaves Mean (-+ SD) 15.16 + 2.12 12.05 -+ 3.16 3.24 +- 1.87 Range 9-23 4-18 1-5 Sample size 141 58 812
Stem height (cm) Mean (_+ SD) 320.0 -+ 83.7 15028 + 70.46 -- Range 109-629 44-355 Sample size 220 58
Stem diameter (cm) Mean (+- SD) 27.1 --+ 4.46 19.48 + 5.25 - - Range 14-38.10 9.7-37.02 Sample size 249 60
Age (year) Mean (_+ SD) 63.1 -+ 23.1 25 -+ 8 -- Range 30-204 15-35 Sample size 220 57
New leaves/year Mean (+ SD) 3.89 - 0.51 3.00 + 1.05 1.02 -+ 0.98 Range 1-5 2-4 0-2 Sample size , 220 58 812
Number of fnaits Mean ( - SD) 262.58 + 278.95 -- - - per individual Range O- 1293
Sample size 249
*Adults, mature reproductive individuals; juveniles, immature (nonreproductive) individuals with a visible stem; seedlings, young plants without a visible stem.
Conservation Biology Volume IO, No. 1, February 1996

Ratsirarson et al. Conservation ofNe0dypsis decaryi 43
tion growth is often preferable to age-structured models for plants and other organisms for which w e lack reli- able information on the age of immature individuals (Lefkovitch 1965). In addition, the growth plasticity of plants as well as the indeterminate growth often make the age structure a poor predictor of future population states (Lefkovitch 1965; Vandermeer 1978; Kirkpatrick 1984). We used multiple regression to determine the pc> tential state variables contributing to the population growth (Caswell 1989). Height and age were measured or estimated for each adult individual, and height was found to be a bet ter predicitor of fectmdity than age (ANOVA: age R 2 = 0 .102,p = 0.975; height: R 2 = 0.204, p = 0.002). Height was then chosen as a state variable for projecting future population growth.
Eight different stage classes were defined within the populat ion depending on the size of the leaves for seed- lings and the height of the s tem for juveniles and adults. Vandermeer (1978) and Moloney (1986) proposed an al- gori thm to minimize the sample and the distribution er- ror for choosing category sizes. The intersection of these two errors is the predicted optimal category size for adults (see Ratsirarson 1993b). The resulting catego- ries were seedlings S1, leaf length < 15 cm; seedlings $2, leaf length 15-60 cm; seedlings $3, leaf length > 60 cm; juvenile J1, s tem visible and height < 80 cm; juve- nile J2, immature individual with stem height > 80 cm; adult A1, mature individual wi th stem height < 200 cm; adult A2, mature individual with s tem height 200-350 cm; and adult A3, mature individual with stem height > 350 cm.
Transition matrices for the average over the three-year study period were constructed for each of the three per- manent plots. The general matrix includes F~, the effec- tive reproductive output of an individual (the first row); Pt, the probability that an individual remains in the same stage class (the main diagonal); and G i, the probability of individuals moving from one stage class to the next (the subdiagonal) (Caswell 1989). All individuals in a given stage are assumed to be subjected to identical mortality, growth, and fecundity schedules. Fecundity (F/) is esti- mated by the number of new seedlings, rather than the number of seeds. Reproductive plants in year t p roduce seeds that will either germinate to produce seedlings in the next year (t + 1) or die (no seed dormancy from one year to the next). In addition, dispersal is limited in this palm (mean dispersal distance = 2.78 m); and most seed- lings are fotmd in close proximity to the maternal plant (Ratsirarson & Silander 1995b). Parameters Gt and Pi were estimated from the size-specific survival probabil- ity, ¢ri, and the growth probability, ~/t, and G t = ¢rt X T~, Pi =¢r t × (1 - Tl) (CasweU 1989). Growth probability can be estimated f rom the stage duration distribution. Little is known about the variability of survival and growth rates within a stage. Therefore, in all individuals within a stage i that survive and grow to stage (i + 1),
the stage duration T t was specified by a geometric distri- bution with a mean T t = 1/~tt; thus, %. = 1 / T t following Caswell (1989).
The linear, time-invariant model of population growth, X ( t + 1) = A × X( t ) , follows Caswell (1989). The dominant eigenvalue Kin, asymptotically equal to e r, and the right and left eigen-vectors, corresponding to the stable stage structure and the reproductive values for the stage classes respectively, were determined us- ing RAMAS/stage software (Ferson 1991). The assump- tions of linearity and time invariance exclude the effects of density and environmental fluctuations on population growth.
Leaf Removal and Fruit Harvesting
To understand the effect of leaf harvest on the growth, reproduction, and survival of adult palms, w e initiated a series of defoliation experiments. Three levels of defolia- tion were carried out: 25%, 50%, and 100%, plus the con- trol (0% defoliation). Although the number of live leaves did vary slightly among individuals in the population (Table 1), for the experimental plants the initial leaf number was not significantly different among treat- ments (Ratsirarson 1993b). The older living leaves were removed with a machete, which is the standard local practice of leaf removal. Entire leaves were cut off at the base of the petiole. Twenty-five individuals were chosen for each level of defoliation. Ten individuals in each level were defoliated in March 1991, and 15 additional individuals were defoliated in March 1992. The number of leaves, the diameter of the s tem at breast height, and the height of individuals prior to defoliation were re- corded. The growth (number of new leaves produced), the reproductive output (number of fruits produced), and the fate of each palm one year after defoliation were then determined. Duncan 's multiple comparison proce- dure with analysis of variance was used to obtain all pair- wise comparisons among treatments.
To provide information on h o w this potentially re- newable resource could best be managed, the same level of defoliation was repeated in March 1992 on the 10 individuals defoliated in March 1991. We recorded growth (number of new leaves produced) as well as re- productive output (number of fruits produced) of the palms one year after the second defoliation. Leaf size of the three oldest, well-expanded new leaves on each indi- vidual was noted by measuring the rachis length from the top of the petiole to the tip of the leaf blade.
To tmderstand the effect of seed harvest on popula- tion growth we carried out demographic analyses. We estimated fecundity (the first row of the transition ma- trix) as the number of mature fruits p roduced by repro- ductive individuals. We counted mature fruits over two seasons using specialized climbing equipment 0umar as- cenders). The growth of the population over t ime was
Conservation Biology Volume 10, No. 1, February 1996
44 Conservation ofNe0dypsis decaryi Ratsirarson et aL
simulated using RAMAS/stage program (Ferson 1991). The dominant eigenvalue, X m of the matrix was used to assess the dynamics of the population. The population is predicted to remain constant if Xm is not different from 1, to decline if )~m < 1, and grow if ~m > 1 (Caswell 1989). We performed a sensitivity analysis by reducing the fecundity coefficients in the matrix model by 10 to 99.9% and calculating the dominant latent root for the al- tered matrix.
Results and Discussion
Demography
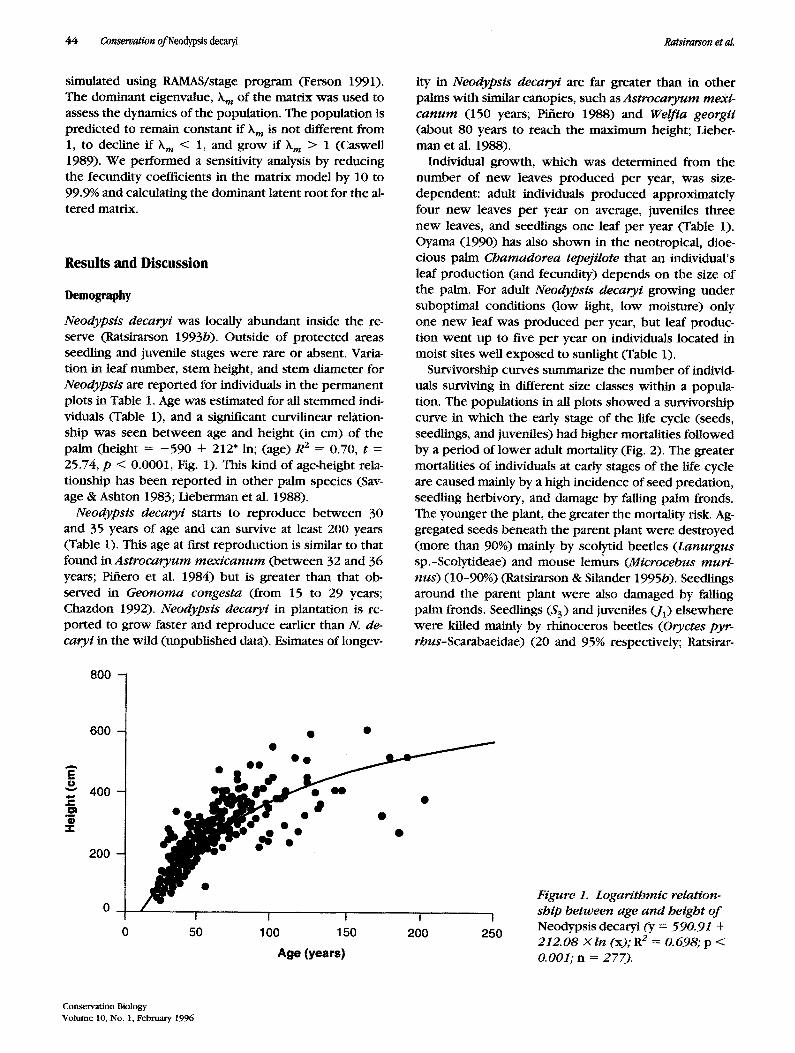
Neodypsis decaryi was locally abundant inside the re- serve (Ratsirarson 1993b). Outside of protected areas seedling and juvenile stages were rare or absent. Varia- tion in leaf number, stem height, and stem diameter for Neodypsis are reported for individuals in the permanent plots in Table 1. Age was estimated for all stemmed indi- viduals (Table 1), and a significant curvilinear relation- ship was seen between age and height (in cm) of the palm (height = - 5 9 0 + 212' in; (age) R 2 = 0.70, t = 25.74, p < 0.0001, Fig. 1). This kind of age-height rela- tionship has been reported in other palm species (Sav- age & Ashton 1983; Lieberman et al. 1988).
Neodypsis decaryi starts to reproduce between 30 and 35 years of age and can survive at least 200 years (Table 1). This age at first reproduction is similar to that found in Astrocaryum mexicanum (between 32 and 36 years; Pifiero et al. 1984) but is greater than that ob- served in Geonoma congesta (from 15 to 29 years; Chazdon 1992). Neodypsis decaryi in plantation is re- ported to grow faster and reproduce earlier than N. de- caryi in the wild (unpublished data). Esimates of longer-
ity in Neodypsis decaryi are far greater than in other palms with similar canopies, such as Astrocaryum mexi- canum (150 years; Pifiero 1988) and Welfia georgii (about 80 years to reach the maximum height; Lieber- man et al. 1988).
Individual growth, which was determined from the number of new leaves produced per year, was size- dependent: adult individuals produced approximately four new leaves per year on average, juveniles three new leaves, and seedlings one leaf per year (Table 1). Oyama (1990) has also shown in the neotropical, dioe- cious palm Chamadorea tepejilote that an individual's leaf production (and fecundity) depends on the size of the palm. For adult Neodypsis decaryi growing under suboptimal conditions (low light, low moisture) only one new leaf was produced per year, but leaf produc- tion went up to five per year on individuals located in moist sites well exposed to sunlight (Table 1).
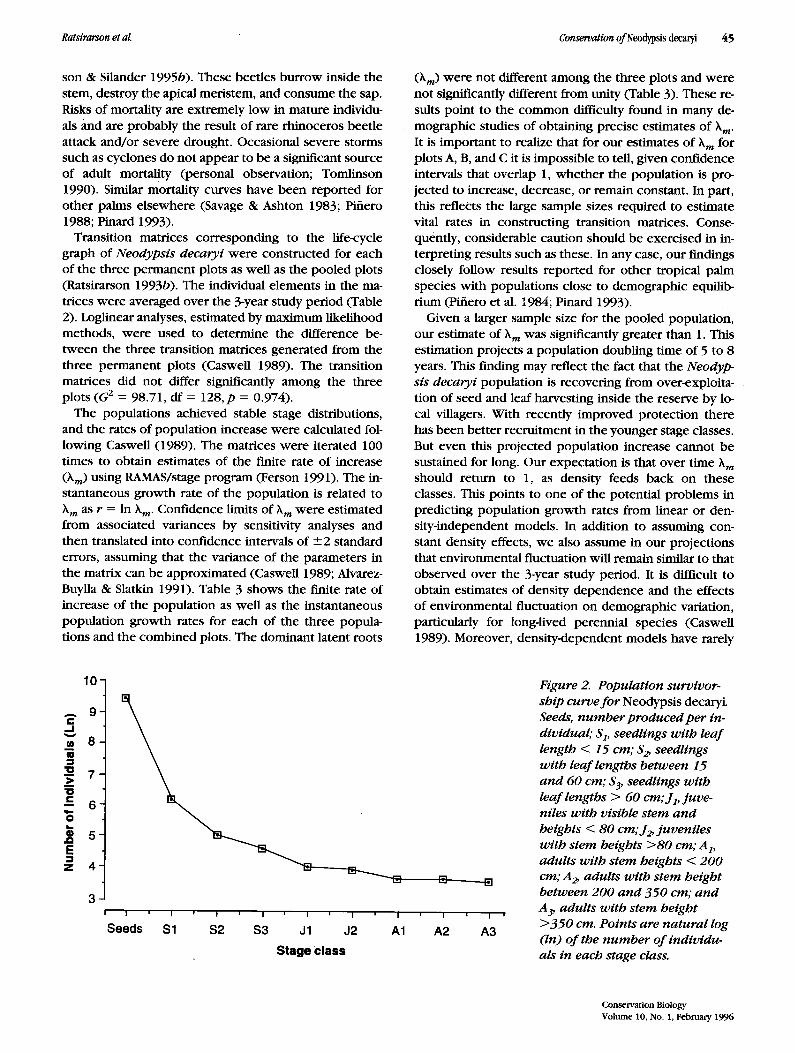
Survivorship curves summarize the number of individ- uals surviving in different size classes within a popula- tion. The populations in all plots showed a survivorship curve in which the early stage of the life cycle (seeds, seedlings, and juveniles) had higher mortalities followed by a period of lower adult mortality (Fig. 2). The greater mortalities of individuals at early stages of the life cycle are caused mainly by a high incidence of seed predation, seedling herbivory, and damage by falling palm fronds. The younger the plant, the greater the mortality risk. Ag- gregated seeds beneath the parent plant were destroyed (more than 90%) mainly by scolytid beetles (Lanurgus sp.-Scolytideae) and mouse lemurs (Microcebus muri- nus) (10-90%) (Ratsirarson & Silander 1995b). Seedlings around the parent plant were also damaged by falling palm fronds. Seedlings (S 3) and juveniles (J1) elsewhere were killed mainly by rhinoceros beetles (Oryctes pyr- rhus-Scarabaeidae) (20 and 95% respectively; Ratsirar-
A
_m .,r
800 -
600 -
400
200
0
O o o o
- •
o
e
N
I I I I I
50 1 O0 150 200 250
Age (years)
Figure 1. Logarithmic relation- ship between age and height of Neodypsis decaryi (y = 590.91 + 212.08 X ln (x); R 2 = 0.698; p < 0.001; n = 277).
Conservation Biology Volume 10, No. 1, February 1996
Ratsirarson et al. Conservation ofNeodypsis decaryi 45
son & Silander 1995b). These beetles bur row inside the stem, destroy the apical meristem, and consume the sap. Risks of mortality are extremely low in mature individu- als and are probably the result of rare rhinoceros beetle attack and/or severe drought. Occasional severe storms such as cyclones do not appear to be a significant source of adult mortality (personal observation; Tomlinson 1990). Similar mortality curves have been reported for other palms elsewhere (Savage & Ashton 1983; Pifiero 1988; Pinard 1993).
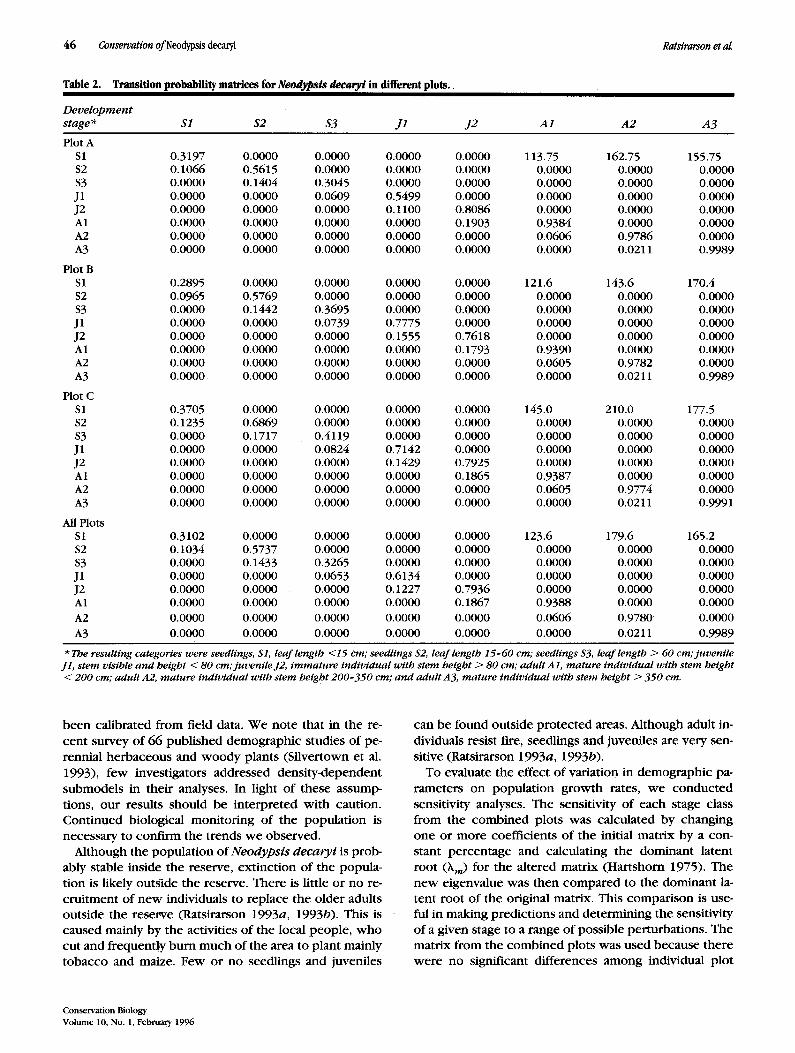
Transition matrices corresponding to the life-cycle graph of Neodypsis decaryi were constructed for each of the three permanent plots as well as the pooled plots (Ratsirarson 1993b). The individual elements in the ma- trices were averaged over the 3-year study period (Table 2). Loglinear analyses, estimated by maximum likelihood methods, were used to determine the difference be- tween the three transition matrices generated from the three permanent plots (Caswell 1989). The transition matrices did not differ significantly among the three plots (G 2 = 98.71, df = 128,p = 0.974).
The populations achieved stable stage distributions, and the rates of population increase were calculated fol- lowing Caswell (1989). The matrices were iterated 100 times to obtain estimates of the finite rate of increase (hm) using RAMAS/stage program (Ferson 1991). The in- stantaneous growth rate of the population is related to ~k m a s r = In hm' Confidence limits of ~k m w e r e estimated from associated variances by sensitivity analyses and then translated into confidence intervals of - 2 standard errors, assuming that the variance of the parameters in the matrix can be approximated (Caswell 1989; Alvarez- Buylla & Slatkin 1991). Table 3 shows the finite rate of increase of the population as well as the instantaneous populat ion growth rates for each of the three popula- tions and the combined plots. The dominant latent roots
(k m) were not different among the three plots and were not significantly different from unity (Table 3). These re- sults point to the c o m m o n difficulty found in many de- mographic studies of obtaining precise estimates of k m. It is important to realize that for our estimates of ~ m for plots A, B, and C it is impossible to tell, given confidence intervals that overlap 1, whether the population is pro- jected to increase, decrease, or remain constant. In part, this reflects the large sample sizes required to estimate vital rates in constructing transition matrices. Conse- quently, considerable caution should be exercised in in- terpreting results such as these. In any case, our findings closely follow results repor ted for other tropical paLm species with populations close to demographic equilib- rium (Pifiero et al. 1984; Pinard 1993).
Given a larger sample size for the pooled population, our estimate of ~k m w a s s i g n i f i c a n t l y greater than 1. This estimation projects a population doubling time of 5 to 8 years. This finding may reflect the fact that the Neodyp- sis decaryi population is recovering from over-exploita- tion of seed and leaf harvesting inside the reserve by lo- cal villagers. With recently improved protect ion there has been better recruitment in the younger stage classes. But even this projected population increase cannot be sustained for long. Our expectat ion is that over t ime hm should return to 1, as density feeds back on these classes. This points to one of the potential problems in predicting population growth rates from linear or den- sity-independent models. In addition to assuming con- stant density effects, we also assume in our projections that environmental fluctuation will remain similar to that observed over the 3-year study period. It is difficult to obtain estimates of density dependence and the effects of environmental fluctuation on demographic variation, particularly for long-lived perennial species (Caswell 1989). Moreover, density-dependent models have rarely
10-
w 8 -
"0 7 -
5-
:~ 4 -
3 I I ' I ' I ' I ' I ' I I ' I ' I
Seeds $1 S2 S3 J1 J2 A1 A2 A3
Stage class
Figure 2. Populat ion survivor- ship curve f o r Neodypsis decaryL Seeds, n u m b er produced per in- dividual; $1, seedlings with leaf length < 15 cm; S~ seedlings with leaf lengths between 15 a n d 60 cm; $3, seedlings with leaf lengths > 60 cm; Jv juve- niles wi th visible stem a nd heights < 80 cm; J~ juveni les wi th stem heights >80 cm; A v adults wi th stem heights < 200 cm; A~ adults wi th stem height between 200 a n d 350 cm; and A3, adults wi th stem height >350 cm. Points are natural log (In) o f the n u m b er o f individu- als in each stage class.
Conservation Biology Volume 10, No. 1, February 1996
46 Conservation ofNeodypsis decaryi Ratsirarson et al.
Table 2. Transition probability matrices for Neodypsis decaryi in different plots..
Development stage* $1 $2 $3 J1 J2 A1 A 2 A3
Plot A $1 0.3197 0.0000 0.0000 0.0000 0.0000 113.75 162.75 155.75 $2 0.1066 0.5615 0.0000 0:0000 0.0000 0.0000 0.0000 0.0000 $3 0.0000 O. 1404 0.3045 0.0000 0.0000 0.0000 0.0000 0.0000 J1 0.0000 0.0000 0.0609 0.5499 0.0000 0.0000 0.0000 0.0000 J2 0.0000 0.0000 0.0000 0.1100 0.8086 0.0000 0.0000 0.0000 A1 0.0000 0.0000 0.0000 0.0000 0.1903 0.9384 0.0000 0.0000 A2 0.0000 0.0000 0.0000 0.0000 0.0000 0.0606 0.9786 0.0000 A3 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0211 0.9989
Plot B S1 0.2895 0.0000 0.0000 0.0000 0.0000 121.6 143.6 170.4 $2 0.0965 0.5769 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 $3 0.0000 0.1442 0.3695 0.0000 0.0000 0.0000 0.0000 0.0000 J1 0.0000 0.0000 0.0739 0.7775 0.0000 0.0000 0.0000 0.0000 J2 0.0000 0.0000 0.0000 0.1555 0.7618 0.0000 0.0000 0.0000 A1 0.0000 0.0000 0.0000 0.0000 0.1793 0.9390 0.0000 0.0000 A2 0.0000 0.0000 0.0000 0.0000 0.0000 0.0605 0.9782 0.0000 A3 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0211 0.9989
Plot C S1 0.3705 0.0000 0.0000 0.0000 0.0000 145.0 210.0 177.5 $2 0.1235 0.6869 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 $3 0.0000 0.1717 0.4119 0.0000 0.0000 0.0000 0.0000 0.0000 J1 0.0000 0.0000 0.0824 0.7142 0.0000 0.0000 0.0000 0.0000 J2 0.0000 0.0000 0.0000 0.1429 0.7925 0.0000 0.0000 0.0000 A1 0.0000 0.0000 0.0000 0.0000 0.1865 0.9387 0.0000 0.0000 A2 0.0000 0.0000 0.0000 0.0000 0.0000 0.0605 0.9774 0.0000 A3 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0211 0.9991
MI Plots S1 0.3102 0.0000 0.0000 0.0000 0.0000 123.6 179.6 165.2 $2 0.1034 0.5737 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 $3 0.0000 0.1433 0.3265 0.0000 0.0000 0.0000 0.0000 0.0000 J1 0.0000 0.0000 0.0653 0.6134 0.0000 0.0000 0.0000 0.0000 J2 0.0000 0.0000 0.0000 0.1227 0.7936 0.0000 0.0000 0.0000 A1 0.0000 0.0000 0.0000 0.0000 0.1867 0.9388 0.0000 0.0000 A2 0.0000 0.0000 0.0000 0.0000 0.0000 0.0606 0.9780 0.0000 A3 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0211 0.9989
* The resulting categories were seedlings, $1, leaf length <15 cm; seedlings $2, leaf length 15-60 cm; seedlings $3, leaf length > 60 cm; juvenile J1, stem visible and height ~ 80 cm;juvenile J2, immature individual with stem height > 80 cm; adult A1, mature individual with stem height < 200 cm; adult A2, mature individual with stem height 200-350 cm; and adult A3, mature individual with stem height > 350 cm.
b e e n calibrated from field data. We note that in the re-
cen t survey of 66 pub l i shed demographic studies of pe- rennial he rbaceous and w o o d y plants (Silvertown et al.
1993), few investigators addressed dens i ty -dependent submodels in their analyses. In light of these assump- tions, our results should be in te rpre ted wi th caution. Con t inued biological moni to r ing of the popu la t ion is necessary to conf i rm the t rends we observed.
Although the popu la t ion of N e o d y p s i s d e c a r y i is prob- ably stable inside the reserve, ex t inc t ion of the popula- t ion is likely outside the reserve. There is little or no re- c ru i tmen t of n e w individuals to replace the older adults outside the reserve (Ratsirarson 1993a, 1993b). This is caused mainly by the activities of the local people , w h o cut and f r e q u e n t ~ b u r n m u c h of the area to plant mainly tobacco and maize. Few or no seedlings and juveniles
can be found outside p ro tec ted areas. Mthough adult in- dividuals resist fire, seedlings and juveniles are very sen-
sitive (Ratsirarson 1993a, 1993b). To evaluate the effect of variat ion in demographic pa-
rameters on popu la t ion growth rates, w e c onduc t ed sensitivity analyses. The sensitivity of each stage class f rom the c o m b i n e d plots was calculated by changing one or more coefficients of the initial matr ix by a con- stant percentage and calculating the dominan t latent root (K m) for the altered matr ix (Hartshorn 1975). The n e w eigenvalue was t hen compared to the dominan t la- tent root of the original matrix. This compar i son is use- ful in making predic t ions and de te rmin ing the sensitivity of a given stage to a range of possible per turbat ions . The matr ix from the c o m b i n e d plots was used because there were no significant differences among individual plot
Conservation Biology Volume 10, No. 1, February 1996
Ratsirarson et al. Conservation ofNe0dypsis decaryi 47
Table 3. Eigenvalues (Am) and instantaneous growth rates (r = In ~m) for populations ofNeodypsis decaryi.
95% con f idence Plots )~m limits for )k m r
A 1.067 0.081 0.0648 B 1.105 0.137 0.0998 C 1.161 0.220 0.1492 All plots 1.157 0.066 0.1458
matrices. Simulated changes in the popu la t ion parame-
ters had little or no effect on ~k m excep t among adult stage classes (Table 4). Surprisingly, a r educ t ion of seed- lings or juveni le survival up to 90% did no t significantly
affect )k m (Table 4). Adult classes were the mos t sensit ive stages. A 50% reduc t ion of adult survival caused greater changes in the d o m i n a n t la tent root (from ~k m = 1.157542 to 0.994199) than did equivalent reduc t ions in the survival of o ther stages (Table 4). A reduc t ion in all parameters by 50% had a great impact on popu la t ion
g rowth rates (from ~m = 1.157542 to 0.784606). A change of adult survival appears to have a greater influ-
ence on )k m than the same pe rcen t change in seedling or juveni le survival.
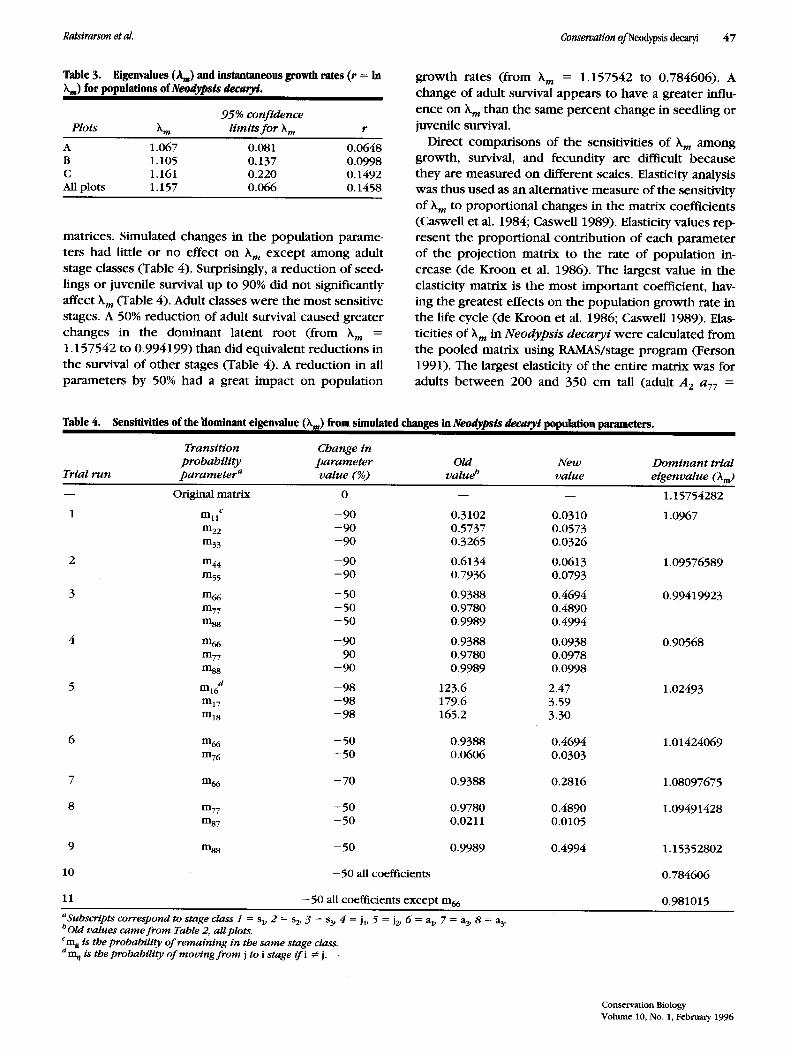
Direct compar i sons of the sensitivities of ~m among growth, survival, and fecundi ty are difficult because they are measured on different scales. Elasticity analysis was thus used as an alternative measure of the sensitivity of ~k m to p ropor t iona l changes in the matr ix coefficients (CasweU et al. 1984; Caswell 1989). Elasticity values rep- resent the p ropor t iona l con t r ibu t ion of each paramete r of the pro jec t ion matr ix to the rate of popu la t ion in- crease (de Kroon et al. 1986). The largest value in the
elasticity matr ix is the most impor tan t coefficient, hav- ing the greatest effects on the popu la t ion g rowth rate in the life cycle (de Kroon et al. 1986; Caswell 1989). Elas-
ticities of )~m in N e o d y p s i s d e c a r y i were calculated f rom the p o o l e d matr ix us ing RAMAS/stage program (Ferson 1991). The largest elasticity of the ent i re matr ix was for
adults b e t w e e n 200 and 350 cm tall (adult A 2 a77 =
Table 4. Sensitivities of the ~ominant eigenvalue (~m) from simulated changes in Neodypsis decaryi population parameters.
Trans i t ion Change in p r o b a b i l i t y p a r a m e t e r Old N e w D o m i n a n t tr ia l
Trial r u n p a r a m e t e r a va lue (%) va lue b va lue e igenva lue (A m)
- - Original matrix 0 - - - - 1.15754282
1 mll c - 9 0 0.3102 0.0310 1.0967 m22 - 9 0 0.5737 0.0573 m33 - 9 0 0.3265 0.0326
2 m44 - 9 0 0.6134 0.0613 1.09576589 m55 - 9 0 0.7936 0.0793
3 m ~ - 5 0 0.9388 0.4694 0.99419923 m77 - 5 0 0.9780 0.4890 mss - 5 0 0.9989 0.4994
4 m66 - 9 0 0.9388 0.0938 0.90568 m77 - 9 0 0.9780 0.0978 m ~ "90 0.9989 0.0998
d 5 ml 6 --98 123.6 2.47 1.02493 m17 --98 179.6 3.59 m18 --98 165.2 3.30
6 m66 - 5 0 0.9388 0.4694 1.01424069 m76 - 5 0 0.0606 0.0303
7 m66 - 7 0 0.9388 0.2816 1.08097675
8 m77 - 5 0 0.9780 0.4890 1.09491428 m87 - 5 0 0.0211 0.0105
9 mss -- 50 0.9989 0.4994 1.15352802
10 --50 all coefficients 0.784606
11 --50 all coefficients except m66 0.981015
aSubscripts correspond to stage class 1 = sl, 2 = s~ 3 = sv 4 = Jl, 5 = j~ 6 = al, 7 = a~ 8 = a 3. ~ Old values came f rom Table 2, all plots. Cmu is the probability o f remaining in the same stage class. d mij iS the probability o f moving f rom j to i stage i f i # j.
Conservation Biology Volume 10, No. 1, February 1996
48 Conservation ofNe0dypsis decaryi Ratsirarson et al.
0.20
0.10
0.00 , ~ r ~ ' ~ ' ' ' 0 $1 S2 S3 J1 J2 A1 A2 A3
Size class
Figure 3. Elasticities o f A m rela- tive to changes in survival, growth, a n d f ecund i ty in Neodypsis decaryt Points repre- sent the proport ional contribu- tion o f each p a r a m e t e r o f the matr i x in all plots to the rate o f populat ion increase (A). Size cat- egories o f the p a l m are the same as in Fig. 2. Survival (Pi) is shown as squares wi th a dot, growth (Gi) as a f i l led diamond, a n d f ecund i t y (F i) as open squares.
0.214). The elasticities for Pt were larger than those for G t or F t in all stage classes (Fig. 3). Thus the probabilities of surviving and remaining in the same size class are the most sensitive coefficients in the life cycle. Changes in fecundity have only a small or no effect on h m. But we note that recruitment is essential to population persis- tence. Obviously, outside the reserve where recruitment is close to nothing, populations are doomed. Changes in the probability of survival with growth G t are more im- portant, although changes in the probability of survival to remain in the same stage P~ contribute the most to hm. This result appears to be common in woody plants in general (Silvertown et al. 1993), as well as specifically in palm species (Pifiero et al. 1984; Enright & Watson 1992; Pinard 1993).
Defoliation
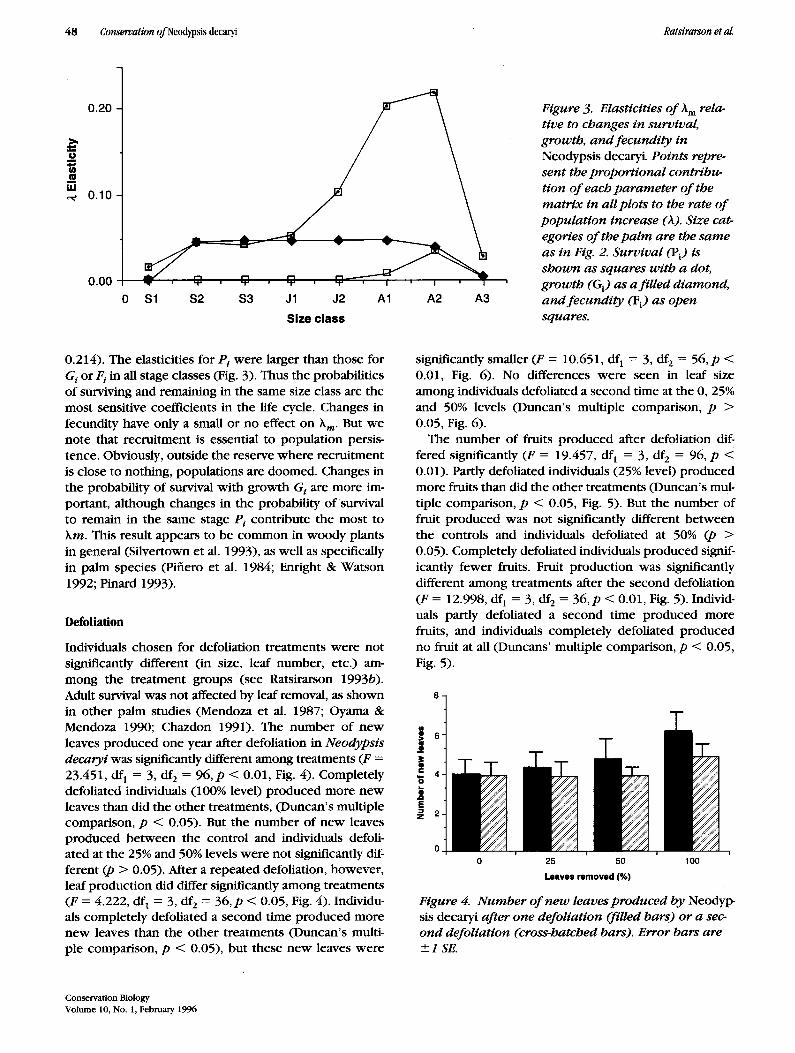
Individuals chosen for defoliation treatments were not significantly different (in size, leaf number, etc.) am- mong the treatment groups (see Ratsirarson 1993b). Adult survival was not affected by leaf removal, as shown in other palm studies (Mendoza et al. 1987; Oyama & Mendoza 1990; Chazdon 1991). The number of new leaves produced one year after defoliation in Neodypsis decaryi was significantly different among treatments (F = 23.451, df 1 = 3, df2 = 96 ,p < 0.01, Fig. 4). Completely defoliated individuals (100% level) produced more new leaves than did the other treatments, (Duncan's multiple comparison, p < 0.05). But the number of new leaves produced between the control and individuals defoli- ated at the 25% and 50% levels were not significantly dif- ferent (p > 0.05). After a repeated defoliation, however, leaf production did differ significantly among treatments (F = 4.222, df 1 = 3, df 2 = 36,p < 0.05, Fig. 4). Individu- als completely defoliated a second time produced more new leaves than the other treatments (Duncan's multi- ple comparison, p < 0.05), but these new leaves were
significantly smaller (F = 10.651, d f 1 = 3, df 2 = 56 ,p < 0.01, Fig. 6). No differences were seen in leaf size among individuals defoliated a second time at the O, 25% and 50% levels (Duncan's multiple comparison, p > 0.05, Fig. 6).
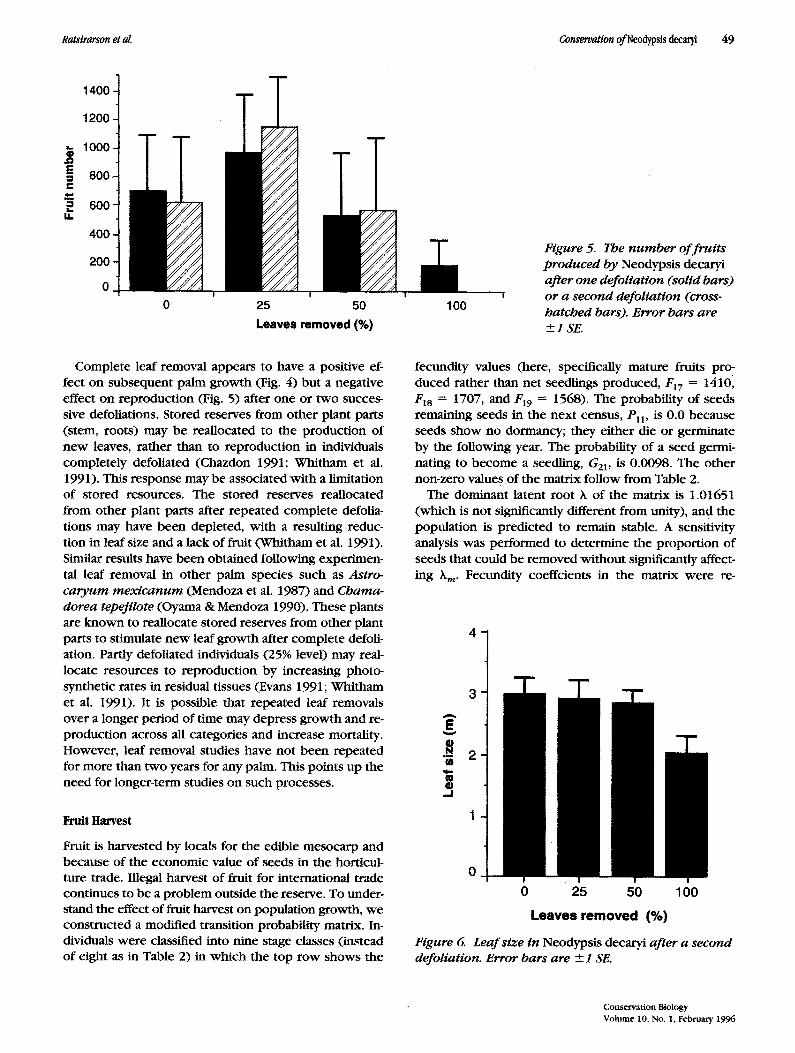
The number of fruits produced after defoliation dif- fered significantly (F = 19.457, df I = 3, df2 = 96, p < 0.01). Partly defoliated individuals (25% level) produced more fruits than did the other treatments (Duncan's mul- tiple comparison, p < 0.05, Fig. 5). But the number of fruit produced was not significantly different between the controls and individuals defoliated at 50% (p > 0.05). Completely defoliated individuals produced signif- icantly fewer fruits. Fruit production was significantly different among treatments after the second defoliation (F = 12.998, d f 1 • 3, df 2 = 36,p < 0.01, Fig. 5). Individ- uals partly defoliated a second time produced more fruits, and individuals completely defoliated produced no fruit at all (Duncans' multiple comparison, p < 0.05, Fig. 5).
g 6 _o | -~ 4 o
z
0 0 25 50 100
Leaves removed (%)
Figure 4. N u m b e r o f n e w leaves produced by Neodyp- sis decaryi after one defoliation (filled bars) or a sec- ond defoliation (cross-hatched bars). Error bars are +_I SE.
Conservation Biology Volume 10, No. 1, February 1996
gatsirarson et al. Conservation ofNe0dypsis decaryi 49
h ,
E I=
2
1400
1200
1000
800
600
400
200
0
25 50
Leaves removed (%)
100
Figure 5. The n u m b er o f f rui ts produced by Neodypsis decaryi after one defoliation (solid bars) or a second defoliation (cross- hatched bars). Error bars are +_1 SE.
Complete leaf removal appears to have a positive ef- fect on subsequent palm growth (Fig. 4) but a negative effect on reproduction (Fig. 5) after one or two succes- sive defoliations. Stored reserves from other plant parts (stem, roots) may be reallocated to the product ion of new leaves, rather than to reproduct ion in individuals completely defoliated (Chazdon 1991; Whitham et al. 1991). This response may be associated with a limitation of stored resources. The stored reserves reallocated f rom other plant parts after repeated complete defolia- tions may have been depleted, with a resulting reduc- tion in leaf size and a lack of fruit (Whitham et al. 1991). Similar results have been obtained following experimen- tal leaf removal in other palm species such as Astro- caryum mex icanum (Mendoza et al. 1987) and Chama- dorea tepejilote (Oyama & Mendoza 1990). These plants are known to reallocate stored reserves f rom other plant parts to stimulate new leaf growth after complete defoli- ation. Partly defoliated individuals (25% level) may real- locate resources to reproduct ion by increasing photo- synthetic rates in residual tissues (Evans 1991; Whitham et al. 1991). It is possible that repeated leaf removals over a longer period of t ime may depress growth and re- product ion across all categories and increase mortality. However, leaf removal studies have not been repeated for more than two years for any palm. This points up the need for longer-term studies on such processes.
Fruit Harvest
Fruit is harvested by locals for the edible mesocarp and because of the economic value of seeds in the horticul- ture trade. Illegal harvest of fruit for international trade continues to be a problem outside the reserve. To under- stand the effect of fruit harvest on population growth, we constructed a modified transition probability matrix. In- dividuals were classified into nine stage classes (instead of eight as in Table 2) in which the top row shows the
fecundity values (here, specifically mature fruits pro- duced rather than net seedlings produced, F17 = 1410, F18 = 1707, and F19 = 1568). The probability of seeds remaining seeds in the next census, P]I, is 0.0 because seeds show no dormancy; they either die or germinate by the following year. The probabili ty of a seed germi- nating to become a seedling, Gzl, is 0.0098. The other non-zero values of the matrix follow from Table 2.
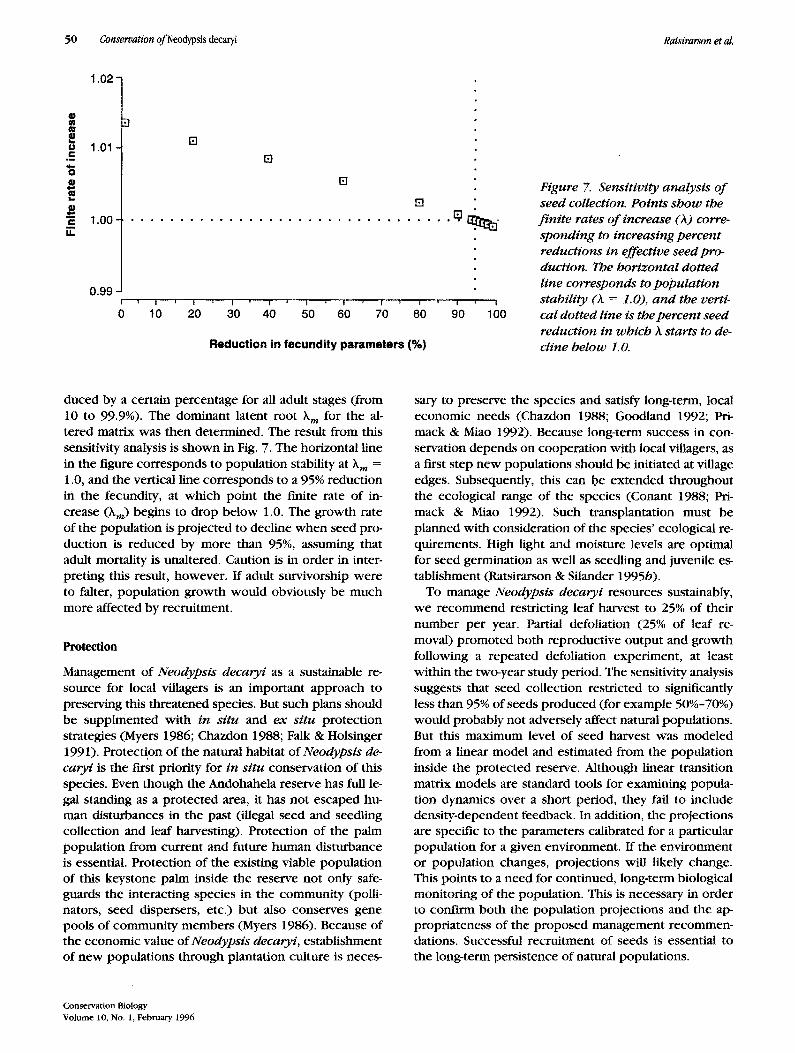
The dominant latent root h of the matrix is 1.01651 (which is not significantly different from unity), and the population is predicted to remain stable. A sensitivity analysis was per formed to determine the propor t ion of seeds that could be removed wi thout significantly affect- ing k m. Fecundity coeffcients in the matrix were re-
4
3
A
E
w ~ m
0 0 25 50 1 O0
Leaves removed (%)
Figure 6. Leaf size in Neodypsis decaryi after a second defoliation. Error bars are +_ 1 SE.
Conservation Biology Volume 10, No. 1, February 1996
50 Conservation ofNe0dypsis decaryi Ratsirarson et al.
Q W m
0 C
"6 .= 0. e~
t L
1.02"
1.01
1.00
0.99
0
[]
[]
[]
[] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . E] [ ~ .
' ' ' ' ' i o ¢ d o ' ' ' 10 20 30 40 50 0 90 100
Reduction in fecundity parameters (%)
Figure 7. Sensitivity analysis o f seed collection. Points show the f in i te rates o f increase (A) corre- sponding to increasing percen t reductions in effective seed pro- duction. The horizontal dotted line corresponds to popula t ion stability (A = 1.0), a n d the verti- cal dotted line is the percent seed reduction in which A starts to de- cline below 1. O.
duced by a certain percentage for all adult stages (from 10 to 99.9%). The dominant latent r o o t ~ m for the al- tered matrix was then determined. The result f rom this sensitivity analysis is shown in Fig. 7. The horizontal line in the figure corresponds to population stability a t )k m =
1.0, and the vertical line corresponds to a 95% reduction in the fecundity, at which point the finite rate of in- crease (km) begins to drop below 1.0. The growth rate of the population is projected to decline when seed pro- duction is reduced by more than 95%, assuming that adult mortality is unaltered. Caution is in order in inter- preting this result, however. If adult survivorship were to falter, population growth would obviously be much more affected by recruitment.
Protection
Management of Neodypsis decaryi as a sustainable re- source for local villagers is an important approach to preserving this threatened species. But such plans should be supplmented with in situ and ex situ protect ion strategies (Myers 1986; Chazdon 1988; Falk & Holsinger 1991). Protection of the natural habitat of Neodypsis de- caryi is the first priority for in situ conservation of this species. Even though the Andohahela reserve has full le- gal standing as a protected area, it has not escaped hu- man disturbances in the past (illegal seed and seedling collection and leaf harvesting). Protection of the palm population from current and future human disturbance is essential. Protection of the existing viable population of this keystone palm inside the reserve not only safe- guards the interacting species in the communi ty (polli- nators, seed dispersers, etc.) but also conserves gene pools of communi ty members (Myers 1986). Because of the economic value ofNeodyps is decaryi, establishment of new populations through plantation culture is neces-
sal T to preserve the species and satisfy long-term, local economic needs (Chazdon 1988; Goodland 1992; Pri- mack & Miao 1992). Because long-term success in con- servation depends on cooperat ion with local villagers, as a first step new populations should be initiated at village edges. Subsequently, this can be extended throughout the ecological range of the species (Conant 1988; Pri- mack & Miao 1992). Such transplantation must be planned with consideration of the species' ecological re- quirements. High light and moisture levels are optimal for seed germination as well as seedling and juvenile es- tablishment ~a ts i ra rson & Silander 1995b).
To manage Neodypsis decaryi resources sustainably, we recommend restricting leaf harvest to 25% of their number per year. Partial defoliation (25% of leaf re- moval) p romoted both reproductive output and growth following a repeated defoliation experiment, at least within the two-year study period. The sensitivity analysis suggests that seed collection restricted to significantly less than 95% of seeds produced (for example 50%-70%) would probably not adversely affect natural populations. But this maximum level of seed harvest was modeled from a linear model and estimated from the population inside the protected reserve. Although linear transition matrix models are standard tools for examining popula- tion dynamics over a short period, they fail to include density-dependent feedback. In addition, the projections are specific to the parameters calibrated for a particular population for a given environment. If the environment or population changes, projections will likely change. This points to a need for continued, long-term biological monitoring of the population. This is necessary in order to confirm both the population projections and the ap- propriateness of the proposed management recommen- dations. Successful recrui tment of seeds is essential to the long-term persistence of natural populations.
Conservation Biology Volume 10, No. 1, February 1996

Ratsirarson et al. Conservation ofNeodypsis decaryi 51
S u s t a i n a b l e l e a f a n d s e e d h a r v e s t c a n n o t b e e s t a b -
l i s h e d w i t h o u t t h e a c t i v e p a r t i c i p a t i o n o f l oca l c o m m u -
n i t i e s . Loca l n o n g o v e r n m e n t a l o r g a n i z a t i o n s o r sma l l lo-
ca l c o o p e r a t i v e s h a v e a c r u c i a l r o l e to p l a y i n p r o m o t i n g
g r a s s r o o t s p a r t i c i p a t i o n in c o n s e r v a t i o n e f f o r t s a n d i n es-
t a b l i s h i n g a s u c c e s s f u l c o n s e r v a t i o n p r o g r a m ( G 6 m e z -
P o m p a & B u r l e y 1991) . S u c h a p r o g r a m h a s b e e n i m p l e -
m e n t e d r e c e n t l y i n t h e r e g i o n , i n c o l l a b o r a t i o n w i t h t h e
e x i s t i n g I n t e g r a t e d C o n s e r v a t i o n a n d D e v e l o p m e n t Pro-
j e c t o f A n d o h a h e l a . C o o p e r a t i o n w i t h t h i s g r o u p is n e c -
e s s a r y to i m p l e m e n t t h e c o n s e r v a t i o n m a n a g e m e n t p ro -
g r a m .
Acknowledgments
W e a c k n o w l e d g e t h e D i r e c t i o n d e s E a u x e t For~ts -Min-
i s t~ re d e s E a u x e t F o r ~ t s in M a d a g a s c a r f o r g r a n t i n g pe r -
m i s s i o n t o w o r k i n t h e p r o t e c t e d r e s e r v e o f A n d o h a h e l a .
Spec i a l t h a n k s a re d u e t o S. O ' C o n n o r , A Ra je r i son , A.
R a m a n a n j a t o v o , a n d J . V. G. R a s o l o n i r i n a f o r t h e i r inva lu-
a b l e h e l p a n d a s s i s t a n c e in M a d a g a s c a r . W e t h a n k R. L.
C h a z d o n , K. H o l s i n g e r , S. Zack , a n d R. P r i m a c k f o r t h e i r
u s e f u l s u g g e s t i o n s a n d c o m m e n t s o n e a r l i e r v e r s i o n s o f
t h e m a n u s c r i p t . W e a c k n o w l e d g e e n c o u r a g e m e n t a n d
h e l p in m a n y d i f f e r e n t w a y s f r o m R. D e w a r a n d M.
S c h w a r t z . T h i s p r o j e c t w a s f i n a n c i a l l y s u p p o r t e d b y t h e
B i o d i v e r s i t y S u p p o r t P r o g r a m ( W o r l d Wi ld l i f e F u n d
G r a n t # 7 5 1 7 ) . A d d i t i o n a l s u p p o r t w a s p r o v i d e d b y T h e
W o r l d Wi ld l i f e F u n d U.S. a n d T h e U n i v e r s i t y o f C o n n e c t -
i c u t R e s e a r c h F o u n d a t i o n .
Literature Cited
Alvarez-BuyUa, E. R., and M. Slatkin. 1991. Finding confidence limits on population growth rates. Trends in Ecology and Evolution. 6:221- 224.
Bawa, K. S., and P. S. Ashton. 1991. Conservation of rare trees in tropi- cal rain forests: a genetic perspective. Pages 62-71 in D. A. Falk and K. E. Holsinger, editors. Genetics and conservation of rare plants. Oxford University Press, Oxford, England.
Caswell, H. 1989. Matrix population models. Sinauer Associates, Sun- derland, Massachusetts.
Caswell, H., R. J. Naiman, and R. Morin. 1984. Evaluating the conse- quences of reproduction in complex salmonid life cycles. Aquacul- ture 43:123-134.
Chazdon, R. L. 1988. Conservation-conscious collecting: concerns and guidelines. Principles 32(1):13-17.
Chazdon, R. L. 1991. Effects of leaf and ramet removal on growth and reproduction of Geonoma congesta, a clonal understory palm. Journal of Ecology 79:1137-1146.
Chazdon, R. L. 1992. Patterns of growth and reproduction of Geonoma congesta, a clustered understory palm. Biotropica 24(1):43-51.
Conant, S. 1988. Saving endangered species by translocation. Bio- science 38:254-257.
Corner, E. J. H. 1966. The natural history of palms. Weinfeld and Nicholson, London.
de Kroon, H., A. Plaisier, J. van Groenendael, and H. Caswen. 1986.
Elasticity: the relative contribution of demographic parameters to population growth rate. Ecology 67:1427-1431.
Du Puy, B., D. Du Puy, and V. Randrianasolo. 1992. Some aspects of the palms of Madagascar, and their cultivation at the parc de Tsim- bazza, Antananarivo. Principes 36(2):84-93.
Eboroke, S. 1994. Approche phytosociologique de la Parcelle 3 de la RNI d'Andohahela. Travaux de recherches. Departement Sciences Naturelles, E.E.S.S. Universit6 de Toliary, Madagascar.
Enright, N. J., and A. D. Watson. 1992. Population dynamics of the ni- kau palm, Rhopalostylis sapida (Wendl. et Drude), in a temperate rainforest remnant near Auckland, New Zealand. New Zealand Journal of Botany 30:29-43.
Evans, A. S. 1991. Whole plant responses ofBrassica campestris (Crn- ciferae) to altered source-sink relations. American Journal of Bot- any 78:394-400.
Falk, D. A., and K. E. Holsinger, Editors. 1991. Genetics and conserva- tion of rare plants. Oxford University Press, Oxford, England.
Ferson, S. 1991. RAMAS/Stage generalized stage-based modeling for population dynamics. Exeter Software, Setauket, New York.
Frankel, O. H., and M. E. Soul6. 1981. Conservation and evolution. Cambridge University Press, Cambridge, England.
G6mez-Pompa, A., and F. W. Burley. 1991. The management of natural tropical forests. Pages 3-18 in A. G6mez-Pompa, T. C. Whitmore, and M. Hadley, editors. Rain forest regeneration and management. Man and the biosphere series. UNESCO, Paris.
Goodland, R. J. A. 1992. Neotropical moist forests: priorities for the next two decades. Pages 416-433 in K. H. Redford and C. Padoch, editors. Conservation of neotropical forests. Columbia University Press, New York.
Hartshorn, G. S. 1975. A matrix model of tree population dynamics. Pages 41-51 in F. B. Golley and E. Medina, editors. Tropical ecolog- ical systems. Springer-Verlag, New York.
Howe, H. F., and L. C. Westley. 1988. Ecological relationships of plants and animals. Oxford Univergity Press, Oxford, England.
Janzen, D. H. 1988. Tropical dry forests: the most endangered major tropical ecosystem. Pages 130-137 in E. O. Wilson, editor. Biodi- versity. National Academy Press, Washington, D.C.
Jenkins, M. D. 1987. Madagascar an environment profile. International Union for the Conservation of Nature and Natural Resources, United Nations Environmental Program, World Wildlife Fund, Gland, Switzerland.
Kirkpatrick, M. 1984. Demographic models based on size, not age, for organisms with indeterminate growth. Ecology 65:1874-1884.
Lefkovitch, L. P. 1965. The study of the population growth in organ- isms grouped by stages. Biometrics 21:1-18.
Leslie, P. H. 1945. On the use of matrices in certain population mathe- matics. Biometrlka 33:183 -212.
Lieberman, M., D. Lieberman, and J. Vandermeer. 1988. Age size rela- tionships and growth behavior of Welfia georgii. Biotropica 20(4): 270-273.
Mendoza, A. D Pifiero, and J. Saruldaan. 1987. Effects of experimental defoliation on growth, reproduction and survival of Astrocaryum mexicamum. Journal of Ecology 75:545-554.
Menges, E. S. 1986. Predicting the future of rare plant populations: de- mographic monitoring and modeling. Natural Areas Journal 6:13- 25.
Menges, E. S. 1990. Population viability analysis for an endangered plant. Conservation Biology 4:52-62.
Mills, L. S., M. E. Soul6, and D. F. Doak. 1993. The keystone-species concept in ecology and conservation. Bioscience 43(4):219-224.
Moloney, K. A. 1986. A generalized algorithm for determining category size. Oecologia 69:176-180.
Murphy, P. G., and A. E. Lugo. 1986. Ecology of tropical dry forest. An- num Review of Ecology and Systematics 17:67-88.
Myers, N. 1986. Tropical deforestation and a mega-extinction spasm. Pages 394-409 in M. E. Soul6, editor. Conservation biology: The science of scarcity and diversity. Sinauer Associates, Sunderland, Massachusetts.
Conservation Biology Volume 10, No. 1, February 1996

52 Conservation ofNeodypsis decaryi Ratsirarson et aL.
Myers, N. 1991. Tropical deforestation: the latest situation. Bioscience 41.-282.
NicoU, M. E., and O. Langrand. 1989. Madagascar: revue de la conserva- tion et des aires prot6g6es. World Wildlife Fund, Gland, Switzerland.
Oyama, K. 1990. Variation in growth and reproduction in the neotropi- cal dioecious palm Chamadorea tepejilote. Journal of Ecology 78: 648-663.
Oyama, K., and A. Mendoza. 1990. Effects of defoliation on growth re- production and survival of a neotropical dioecious palm: Chama- dorea tepejilote. Biotropica 22(2):119-123.
Pinard, M. 1993. Impacts of stem harvesting on populations of Iriartea deltoidea (Palmae) in an extractive reserve in Acre, Brazil. Biotro- pica 25:2-14.
Pifiero, D. 1988. Demographic studies in Astrocaryum mexicanum and their use in understanding community dynamics. Principes 30(3):108-116.
Pifiero, D., M. Martinez-Ramos, and J. Sanfldmn. 1984. A population model of Astrocaryum mextcanum and a sensitivity analysis of its finite rate of increase. Journal of Ecology 72:977-991.
Primack, R. B., and S. L. Miao. 1992. Dispersal can limit local plant dis- tribution. Conservation Biology 6:513-519.
Ratsirarson, J. 1993a. Importance 6conomique et menace d 'un palmier en danger de Madagascar: Neodypsis decaryi. Akon'ny Ala 2:24-25.
Ratsirarson, J. 1993b. Population biology and conservation of an en- dangered Madagascar triangle palm: Neodypsis decaryi. Ph.D. the- sis. The University of Connecticut, Storrs.
Ratsirarson, J., and J. A. Silander, Jr. In press. Reproductive biology of a threatened Madagascar triangle palm: Neodypsis decaryi, Jumelle. Biotropica.
Ratsirarson, J., and J. A. Silander, Jr. In press. Factors affecting the dis- tribution of a threatened Madagascar palm species Neodypsis de- caryi Jumelle. Journal of Tropical Ecology.
Savage, A. J. P., and P. S. Ashton. 1983. The population structure of the double coconut and other Seychelles palms. Biotropica 15(1):15- 25.
Silvertown, J., M. Franco, I. Pisanty, and A. Mendoza. 1993. Compara- tive plant demography. Relative importance of life-cycle compo- nents to the finite rate of increase in woody and herbaceous peren- nials. Journal of Ecology 81:465-476.
Soul6, M. E., editor. 1986. Conservation biology: The science of scar- city and diversity. Sinauer Associates, Sunderland, Massachusetts.
Terborgh, J. 1986. Keystone plant resources in the tropical forest. Pages 330-344 in M. E. Soul6, editor. Conservation biology: The science of scarcity and diversity. Sinauer Associates, Sunderland, Massachusetts.
Tomlinson, P. B. 1990. The structural biology of palms. Clarendon Press, Oxford.
Uhl, N., and J. Dransfield. 1987. Genera Palmarum a classification of palms based on the work of Harold E. Moore Jr. Allen Press, Lawrence, Kansas.
Vandermeer, J. 1978. Choosing category size in a stage projection ma- trix. Oecologla 69:176-180.
Wittham, T. G.,J. Maschinski, K. C. Larson, and K. N. Page. 1991. Plant responses to herbivory: the continuum from negative to positive and underlying physiological mechanisms. Pages 227-256 in D. W. Price, T. M. Lewinsohn, G. W. Fernandes, and W. W. Benson, edi- tors. Plant-animal interactions. John Wiley & sons, New York.
Wilson, E. O. editor, 1988. Biodiversity. National Academy Press, Washington, D. C.
World Resources Institute, International Union for the Conservation of Nature and Natural Resources, United National Environmental Pro- gram. 1992. Global biodiversity strategy: Guidelines for action to save, study and use Earth's biotic wealth, sustainably, and equita- bly. World Resources Institute, Washington, D.C.
Conservation Biology Volume 10, No. 1, February 1996





























