Embed Size (px)
Citation preview

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

' :: Fe.de erratas-':c ., 'pp 18: 20 párrafo, g"linea:.. resultados en.. debe de,cit- ..r,'es,¡¿{üad,os son en.-) : : " , -"
'pp 25:3" " , Después de..P. nígra.. se añaci i ; 'á; . íRPP y RF$f res ect ivamente).
. : t{¡p a?: f ",
ú1,j*" frase:..acícuras...iiebe decir ..acíc¿¿res y hojas.' r . . : ' . . p 42: Zo "
, lot l inea: .- , t i3. .debe ciecir . .par trece.
pp 42: 3o ",...CoREX.. debe deci,r ...CORE'S.
pp 46: penúltima linea:..p.nígra.. debe ciecir ..p. ptrtasüet .pp 54: 20 párrafo: ..In-forrae Lucderme 1989..éebe Cecir . "-Mara 19g9.pp 82: últirno párrafo:....f ig.2-6.. ciebe ciecir .. Tab"La 2.ZOa.pp 82: úitimo párrafo: La frase "En términos.".4z" ha de suprirnirse puesto qu€vuej.ve a aDarecer en Ia pp 93.
pp 85: En ei apartado de increr¡rentos radiales, llrifiief párrafo :...COREX.. ira
cie toner.".COR.CS.n
pp 133: 1- párrafo, úiti:na iinea:...que supone ü.20--.debe decir ...que supone0.3 Kg/tndívíduo.año.
l:p 1E8: f pár:-afo" a.l final: --.iabLa 3.l8...debe decir' .. -t-eb!a. 3:Lfua-pp \92:20 párrafo, e.l finai: ...sisterna. se ha-de añariir .".\Tbbla 3.lgü.pp 215: 2c pánrafo. La última frase ccrresponde al pámafo anterior-pp 433: ulrimo párrafo. lo y zo linea: ...2.s r./ha.año (Rpp) y 2.04 en(RPM)..-debe ciecir:... 3.2 T/ha.año (Rpp) y 2.T T/hai¿o (npr).
pp '435: 30 pár'rafo, z"linea: ...7s en Rpp y 63 en Rply-.. debe decir: ...75Kg/ha.año en RPP y 63 Kg/t:ta.añ.o en RpiI. i:
pp 459: 3o referencia:..,.Tesi.s doctoral... debe
LLcenctatura.
Addendum bibliognafía.
Escr¡dero A,., Del ar,co J-M" y Sarrido M.v:' L99z- The efíiciency of flutrogeni'et¡^asnsiocation from ieaf .rnass in Quercus ilex ecosystens. Vegetatío 99-l.OO.pp 2?.5-2j,5.
eiecin ...Tesís de
Castillo Reqr.lena J. M.
i'egional ccn los tipos
Colección investigación
cie AlmerÍa.
1989a. El clima de AndalucÍa: ciasificación y análisis
de tierapo. Geografía -y
Ordenación del territoriof'!
n- 13. Instituto de Esi¡rdios Aknerienses. Diputación
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

$ffi
AtsREVIATURAS.
RFF. A¡'ea o .egión de la cuenca repobrada con p. ptnaster-.
RPN. Area o región de ia cuenca iepoblada con p. nígra.AD. ádenocarpus decoriícans
CL. Cistus lauri/ctius
PP. Pinus pin"aster
PN. Pinus ntgra
DtsH. Diámetrc rneCido a 130 crn.
DB. Diámet:-o basal
coRES. Muestra testigo de mad.era toraada con bari-ena tipo p:^essier.
PPN. Produ,cción primaria neta.
I-AI. Inciice de area foiia¡-
PF. Producción neta fcliar
PT. Frcducción neta total (:-eferioa a la parte aérea)tsT. Biomasa aérea total.
DF. Desfronde
DS. Deposicién seca
TN. Trascolación neta-
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Departamento de Ciencias Ambienta les y Recursos Natura lesFacul tad de CienciasUnivers idad de Al icante
Desfronde, descomposición y fi jación denitrógeno en una microcuenca con repoblación
joven de coníferas y matorral deCistus laurifolius y Adenocarpus decorticans
en la Sierra de los Filabres (Almería).
Memor ia p resentada por :
Mo José Moro Cuadr i l lero
para op tar a l g rado de Doctor en C ienc ias B io lóg icas
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Departamento de Cicnc ias Ambienta lesFacul tad dc Cicnc ia¡Univers idad de Al lcante
y Recursos Naturales
Desfronde, descomposición y fijación denitrógeno en una microcuenca con repoblación
joven de coníferas y matorral deCistus laurifolius y Adentocarpus decorticans
en la sierra de los Filabres (Almería).
Memoria presentada por:
M. José Moro Cuadri l leropara optar al grado de Doctor en ciencias Biológicas
Los directores de la tesis
Dr. Antonio Escarré EsteveCatedrát ico de EcologfaUnivers idad de Al icante
Dr. Juan Bel lot AbadProfesor T i tu lar de EcologfaUnivers idad de Al icante
A l i can te , Jun io de 1992 .
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

D. ANTONIO ESCARRE ESTEVE, CATEDRATICO DE ECOLOGIA DE LA
FACULTAD DE CIENCIAS DE LA UNIVERSIDAD DE ALICANTE
Y
D. JUAN BELLOT ABAD, PROFESOR TITULAR DE ECOLOGIA DE LA
FACULTAD DE CIENCIAS DE LA UNIVERSIDAD DE ALICANTE
CERTIFICAN, como Directores de la Tesis, que el presente trabajo se ha realizado en ellaboratorio del departamento de ciencias Ambientales y Recursos Naturales de launiversidad de Alicante y en la Estación Experimental de Zonas Aridas (c.S.l.C.) deAlmería, mientras que eltrabajo de campo se ha l levado a cabo en la sierra de los Fi labres.
Y para que conste a todos los efectos académicos, se firma el presente certif¡cado.
L ^r-/\-\-r< - '
D. ANTONIO ESCARRE ESTEVE D. JUAN BELLOT ABAD
Al icante, jun io de 1gg2
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

A mis padres
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

AGRADECIMIENTOS.
AL terminar un trabajo de varíos añ.os cuando Llega La hora de dar eL
"carpetazo", tlna empieza a recordar a toda La gente que de una u otra manera
colaboró, se interesó o símplemente te acompañó durante todo este ttempo.
EL estudio en cuencas en La províncía de Almería no se habría podído
Llevar a cabo de no ser por eL interés que D. Antonio Escarré, D, Juan Ramón
Verícad y D.Jose Angel Carreras pusíeron en eL empeñ,o.
Mí colaboración se debíó a una beca de FPI para eL período 1.986-89 que me
permítíó dedícarme por completo aL Proyecto.
EL apoyo inicíal del personaL del IARA (InstLtuto Andaluz de Reforma
Agrarid fue fundamental para La seLección deL área de estudio y La
construcción de La presa. En concreto he de agradecer eL ínterés
mostrado por D. Estanislao de Simón que síempre, y a pesar de todos sus
compromisos, hacía un hueco en su agenda para ínformarse de La marcha del
Proyecto. También Eduardo Seva (Uníversídad de ALícante) y Luis García-Rosell
(Uníversidad de Granada) particíparon en La primera fase del Proyecto.
En mis primeros muestreos de campo fue fundamental La ayuda de Antonio
que me acompañó durante Los tórrídos día del mes de agosto para colocar
trampas y medir pínos. Después fueron Los trabajadores "Caprícho" y compañía
así eomo .Mari
Carmen y Silvía García de La Universtdad de Alicante quienes
soportaron La ingrata Labor de Los muestreo de bíomasa y estructura. Mención
especial merece Jose ManueL de La guardería forestal que me acompaftó muchas
veces aL campo y cuando iba sola me hacía dos o tres visitas por sí acaso
tenía algún problema con Los jabalíes.
La Estación Experímental de Zonas Aridas me prestó su apoyo, tanto de
infraestructura como de personal. ALLí he pasado Largas temporadas y me he
sentído totalmente integrada. Concretamente he de agradecer a J. Ramón Verícad
y a Juan Puigedefábregas su preocupacLón por La marcha de Los trabajos, a
Sebastián por su apoyo técnico, a OLga, Mari Carmen, Paquita y Gaspar por eL
burocrático y eL de biblíoteca y a todos Los demás por su eompañía, ayuda y
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

buen humor.
AL personal de La casa forestal de Tíjola y más concretamente a La señora
María que me preparaba ricas y calLentes sopas de almendras en invíerno cuando
volvía helada de La Sierra (además me dio La receta).
A Cristina, Tere, Rafa, luús, Fernando, Ramoncin, BeLén y a todos
amígos de Almería gue compartieron mi tLempo de ocLo y con Los que pasé
buenos ratos.
A todos Los compañ,eros deL Departamento de Ecología y a Los que Lo fueron
durante eL período de realízación de La tesís por su apoyo y sugerencias. Ha
sído fundamentaL La ayuda de Silvia Garcta y Mari Carmen en La Labor de
preparación y anáLisís de Las muestras. La de Nico que síempre me ayudó con
Los anáLisis pendíentes que nunca había forma de terminar y La de StLvia Ivars
síempre dispuesta a todo. Jose, recíén incorporado también colaboró en La
preparacíón de aLgunas muestras. Chento me hizo Los díbujos de Las flujos.
Mucha gente del Departamento me ayudó con su.s sugerencias y consejos: Tachi,
Juan BeLLot, Antonio Pastor, Germán, Maria José LLedó, Antonío Estévez.
He de agradecer tambíén Los síempre sabíos consejos de CarLos Gracia,
Ramón VaLLejo y Vícente Hernándí2.
La parte de fijacíón de nítrógeno no habria sido posible sin La
colaboracíón de Paco Bermúdez de Castro que se prestó Lncondicionalmente a
coLaborar conmígo y que además,se hízo cargo de La Labor anáLítica.
A mís padres y a Los de Germán y en generaL a toda La familía que tenían
más ganas de que acabara que yo misma, por su comprensión y paciencia. También
a Toñi y a Carmen Las mujeres de mis compañeros de trabajo.
A Paco, ALfredo y Germán os he dejado para eL final, porque creo que no
os echaré nunca eL sufíciente número de flores para agradeceros toda vuestra
ayuda, apoyo y compañia. Paco ha sido mí compañ,ero de penas y glorias en eL
trabajo de Almería. Su colaboración en todos Los aspectos ha sido fundamental
para funcionar y seguír adelante. Alfredo, nuestro querído y entrañable
Los
tan
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Laborante y amígo siempre ha estado a nuestro Lad.o prestánd.onos una ayud.aeficientísima, sugeríéndonos ideas y poniénd.onos d.e buen humor. creo que Lostres hemos formado un buen equipo. Germán ha hecho d,e tod,o: ayud.arme en eLcampo durante algún muestreo, en La estadística y elaboracíón de d.atos, en Larealtzación del manuscríto, cuidar d.e La niñ,a míentras yo escríbía, aguantarmís malos humores, en fín, una joya. Por eso, junto a mLs padres, os d.ed.tco alos tres este trabajo aunque.soy con.scíente que no es tan divertid,o como unanovela de aventuras.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

INDICE.
SECCION PRIMERA. GENERALIDADES.
INTRODUCCION.
I. AREA DE ESTUDIO.
1.1. Localización.
L.2. Características geológicas y litológicas.
1.3. Climatología.
1.4. Cubierta vegetal.
SECCION SEGUNDA. FUNCIONALISMO DE LAS CUATRO ESPECIES DOMINANTES EN LACUENCA.
2. BIOMASA, MINERALOMASA Y PRODUCCION. 4I2.1. Introducción.
2.2. Materiales y métodos.
2.3. Resultados y discusión.
2.3.1. Biomasa.
2.3.2. Contenido en nutrientes.
2.3.3. Mineralomasa.
2.3.4. producción.
2.3.s. Nutrientes asociados a la producción primaria neta.
3. DESFRONDE.97
3.1. Introducción.
3.2. Material y métodos.
3.3. Resultados y discusión.
3'3'l ' Biomasa del desfronde en la cuanca Nacimiento. AspectosGenerales.
3.3.2. Desfronde en el pinar.
3.3.3. Desfronde en el matorral.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

3.3.4. Relación entre el desfronde total y desfrondefoliar con factores climáticos.
3.3.5. composición química de las hojas del desfronde-3.3.6. Retraslocación foliar.
3.3.7. Retorno de nutrientes al suelo vía desfronde
4. RETORNo DE NUTRIENTES zT7
4.1. Introducción.
4.2. Retorno de nutrientes por trascolación neta_4.2.L. Introducción.
4-2.2. principales caracterÍsticas de la trascoración en lacuenca.
4.2.3. Tasas de deposición seca e intercambio en er flujo detrascolación neta: Aplicación dei modelo de Lovett y Lindberg.4.2-4- contribución de la deposición seca y lixiviado en eraporte de nutrientes al suelo.
4.2.s. Importancia de la vía hidrológiea en er aporte denutrientes al suelo.
4.3. Retorno conjunto por desfnonde y trascolación neta.4.3.r. características dei retorno en ras distintasespecies.
4.3.2. Contribución de los aportes internos (desfronde vlixiviado).
4.3.3. Tasas de circulación de nutrientes.4.3.4. Origen de los iones retornados al suelo.
5. DESCOMPOSICION. 269
5.1. Introducción.
5.2. Materiales y métodos.
5.3. Resultados.
5.3.1. Pérdida de peso.
5'3'z' Dinámica de los nutrientes en el proceso dedescomposición.
I I
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

6. FIJACION DE NITROGENO EN A. d,ecorticans.
6.1. Introducción.
6.2. Material y métodos.
6.3. Resultados.
6.3.1. Nodulación.
6.3.2. Patrón estacional.
6.3.3. Patrón diario.
6.4. Discusión.
6.5. Estima de las entradas por fijación de nitrógeno
SECCION TERCERA. APROXIMACTON A LOS FLUJOS BIOGEOQUIMICOS.
7. FLUJOS INTERNOS Y BIOGEOQUIMICOS.
RESUMEN Y CONCLUSIONES
BIBLIOGRAFIA
APENDICES.
369
387
443
43r
III
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Sección primera.Generalidades.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Introducción.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Introducclón.
I.INTR.ODUCCION.
La utilización de pequeñas cuencas de sustrato impermeable surgió como
una aproximación experimental para el estudio de ecosistemas terestres desde
el punto de vista funcional e integrado. Los primeros precedentes de cuencas
aforadas aparecen en Estados Unidos en 1912 (Piñol l99O). Algo mas tarde, en1934' se puso en marcha la estación hidrológica de Coweeta. Durante estosprimeros ensayos la finalidad de las investigaciones eran basicamente elconocimiento del comportamiento hidrológico de las cuencas tomando a éstascomo meras cajas negras. Fue con la puesta en funcionamiento de la estaciónexperimental de Hubbard Brook cuando se avanzó un paso más en el estudio deecosistemas utilizando esta apnoximación experimental (Likens et aL. Lg17,Bormann y Likens L979). AsÍ, además del estudio de los balances de entrada ysalida de agua y nutrientes, se emprendió la cuantificación de loscompartimentos y flujos internos de la cuenca con vistas a conocer el papeldel componente biótico y edáfico en el funcionamiento global del sistema.Posteriormente, en esta misma estación se llevaron a cabo experimentos enforma de perturbaciones (tala, herbicidas) en las microcuencas con el fin deevaluar su efecto en la dinámica del agua y nutrientes.
El estudio de los ciclos de nutrientes en cuencas forestadas españolasrecogió el enfoque y la metodologÍa desarrollada en Hubbard Brook. Laprospección de cuencas adecuadas dio como resultado la elección de cuencas deencinar, como tipo de vegetación mas nepresentativo de la península ibérica(Bellot 19?8). Las dos areas escogidas fueron el macizo del Montseny y laSierra de Prades, ambas con microcuencas bien delimitadas de sustrato silíceoque reunian las condiciones requeridas de impermeabilidad.
En 1986, el ICONA como máximo responsable del Proyecto LUCDEME en España,consideró oportuno incluir dentro de su programa el estudio biogeoquímico depequeñas cuencas en el sureste español. Esta intención inicial se plasmó en unprimer proyecto de colaboración denominado " Funcionamiento biogeoquÍmico encuencas con vegetación natural, repoblaciones y cultivos: estima de pérdidaserosivas"en el que participaban el IARA (Instituto Andaluz de Reforma Agrariadependiente del ICONA), la Estación Experimental de Zonas Aridas de Almería(CSIC) y la Universida'd de Alicante y que tuvo como principales responsables a
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
D. Antonio Escarré, D.J.R. Vericad y D.Estanislao de Simón. Posteriormente en
el año 1988 el proyecto se continuó con el denominado "Respuestas de pequeñas
cuencas en ecosistemas mediterráneos " con financiación del programa LUCDEME y
de Comunidades Europeas. Actualmente algunas de la cuencas se encuentran
englobadas dentro del Programa ENCORÉ de la CEE (Hornung et al. 1990).
El estudio en la red de cuencas Almería tenía un doble enfoque. El
primero consistía en avanzar el conocimiento de la ecología básica en sistemas
de vegetación típicos del sureste español (matorral degradado, cultivos de
secano, encinares y repoblaciones). El segundo y más aplicado estaría centrado
en el papel de control que pueden ejercer las diferentes cubiertas vegetales
sobre los las pérdidas erosivas en forma disuelta o particulada que, dado el
régimen hidrológico irregular de estas zonas, pueden ser considerables. La
existencia dentro del mismo proyecto de microcuencas aforadas en zonas
mediterráneas de climatología menos extrema (Sierra de Prades) era importante
con vistas a la comparación de ambos tipos de sistemas. Un último objetivo
planteado en el proyecto, más a largo plazo y también con un claro interés
aplicado consistía en analizar los efectos de una. perturbación (aclareo,
fertilización..) sobre los flujos de agua y nutrientes en alguna de las
cuencas.
Con estos tres objetivos básicos, fueron seleccionadas dos cuencas con
repoblación de pinar, una de encinar y otra de matorral en la Sierra de los
Filabres (Almería). El aforo ¡se concluyó a finales de 1988. Hasta el presente
las únicas cuencas en las que se han realizado estudios son las denominadas
"Nacimiento" y "Barranco del agua" ambas de repoblación de Pinus pinaster y
Pinus nigra, pero es en la primera donde se ha concentrado la investigación.
Las repoblaciones con estas especies son representativas de amplias zonas no
sólo de la Sierra de los Filabres, sino de otros sistemas montañosos de la
provincia (Sierra Nevada, Sierra de las Estancias) y responden a la política
de repoblación del ICONA durante los años setenta.
Los trabajos en la cuenca Nacimiento comenzaron con un estudio preliminar
sobre la estructura, biomasa y mineralomasa de las especies dominantes
(finales de 1986 y 1987), instalación de cuñas y pluviómetros para la recogida
de agua de lluvia (1987), colocación de trampas de recogida de desfronde
2
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Introducción.
(final del 1986). Después siguieron la instalación de dispositivos para
recogida de agua de trascolación y escorrentía cortical y bolsas para la
descomposición (1988). Las salidas hidrológicas comenzaron a monitorizarse
también a finales de 1988 al igual que el estudio sobre fijación de nitrógeno
en una de las especies arbustivas dominantes.
Ciclos de nutrientes en sistemas forestales.
Los estudios sobre ciclos y flujos de nutrientes en ecosistemas
terrestres son numerosos, pero la mayor parte de ellos se refieren a sistemas
templado-fríos tanto de caducifolios como de coniferas. Existe una
bibliografía extensa en este campo y buena parte de esta información se
encuentra recopilada en trabajos de síntesis tales como los de Colé y Rapp
(1981), Rodin y Bazilevich (1967), Bray y Gorhman (1964) y Voght et al.
(1986). Sin embargo, los sistemas mediterráneos han sido muy poco estudiados y
dentro de ellos, menos aún los sistemas arbustivos. Uno de los primeros
estudios integrados sobre ciclos de nutrientes en sistemas mediterráneos fue
el realizado en el encinar de le Rouquet y la Madeleine (Lossaint y Rapp 1971)
así como el de la garriga de Quercus coccifera en St. Gély (Rapp y Lossaint
1981) ambas estaciones en el sureste de Francia. Este estudio fue el único
punto mediterráneo incluido en la recopilación de Colé y Rapp (1981) y en el
Programa IBP, lo que puede darnos una idea de la escasez de conocimientos que
en esos años se tenían al respecto en la zona mediterránea.
Poco a poco el número de trabajos sobre ciclos y flujos en el área
mediterránea se ha ido ampliando, apareciendo interesantes publicaciones de
recopilación (Di Castri et al. 1981, Specht 1988, Miller 1981, Kruger et al.
1983).
En España, la introducción de la técnica de microcuencas impulsó
considerablemente el estudio de este aspecto de la ecología de sistemas sobre
todo en la zona de Prades- Montseny. Existe ya un gran número de publicaciones
al respecto entre las que citaremos las de Ferrés 1984, Verdú 1984, Lledó
1990, Mayor 1991) interesados básicamente en los compartimento y flujos
internos de materia orgánica, y los de Roda (1983), Avila (1988), Bellot
(1989), Pinol (1990) o la propia Lledó (1990) interesados en los flujos
3
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
hidrológicos tanto de entrada y salida como de trasferencia interna. Otros
grupos de investigación que cuentan también con cierta tradición en el estudio
de ciclos de nutrientes, aunque no en microcueneas, son los que trabajan en
Salamanca y cuyas investigaciones se centran básicamente en el sistema dehesa
y en las relaciones del encinar con el pasto subyacente (Escudero eí al. 1985,
Escudero eí al. 1983, Del Arco eí a¿.1991, Escudero eí al. 1992) o bien, en
flujos de nutrientes en otros sistemas forestales como robledales, castañares
y pinares (Santa Regina eí al. 1989, Gallardo eí al. 1989).
Motivos que condujeron a la selección del área de estudio para la
realización del siguiente trabajo.
Uno de los puntos a cumplir dentro del proyecto consistía en el
conocimiento de los principales flujos nutrientes en la cuenca repoblada con
pinar y su relación con la vegetación natural. En este contexto la cuenca.
"Nacimiento" aforada en Almería fue seleccionada atendiendo básicamente a las
siguientes características:
1.- La cuenca "Nacimiento" es la más meridional de las cuenca aforadas en
España y en Europa.
2 . - El tipo de vegetación:
- La vegetación de la cuenca consiste en un pinar joven de
repoblación (13 años al comienzo del estudio, 1986). No se conocían
precedentes en España de estudios integrados sobre ciclos de nutrientes en
pinares tan jóvenes aunque en el presente ya estén en marcha estudios de este
tipo como por ejemplo el seguimiento de desfronde y descomposición en
repoblaciones de varias edades de P. halepensis en la provincia de Alicante
(Pastor y Martín 1989). En este sentido, los resultados de Almería podrían
resultar de interés para el conocimiento del comportamiento de un pinar en los
primeros estadios de desarrollo.
- El pinar no ha sufrido ningún tipo de tratamiento silvícola desde su
plantación lo que ha dado lugar a la aparición de un matorral espontáneo con
4
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Introducción.
importante grado de cobertura dominado por Cistus laurifolius y Adenocarpus
decorticans. Esta característica va a permitir abordar el estudio de especies
arbustivas típicamente mediterráneas y ampliar el conocimiento, muy escaso,
que existe sobre flujos de nutrientes en este tipo de arbustos en España
(Núñez 1989, Martín 1984, Merino et al. 1990).
- La coexistencia del pinar joven y del matorral espontáneo en la misma
cuenca ofrecía la posibilidad de abordar conjuntamente el comportamiento de
ambos sistemas respecto a determinados flujos de materia y nutrientes y poder
evaluar la importancia cuantitativa de uno u otro al conjunto del ecosistema.
- El sistema arbustivo de la cuenca se compone dé dos especies que, " a
priori" podían resultar diferentes en relación a su comportamiento
nutricional. Por un lado C. laurifolius es una cistácea de características
pioneras e invasoras englobadas dentro de lo que Herrera (1984) ha dado en
llamar síndrome de tipo I. La j a ra se implantó con mucha fuerza en la cuenca
t ras el aterrazamiento. Por su parte, A. decorticans es una leguminosa que
puede alcanzar porte considerable y es considerada por los fitosociólogos la
especie climácica, junto con la encina (Adenocarpus-quercetum-rotundifoliae)
del dominio meso-supramediterráneo silicícola de las cordilleras béticas (Losa
et al. 1986). Según hemos observado, también parece que esta especie tiene la
capacidad de implantarse con fuerza en sus zonas potenciales t ras una fuerte
perturbación y esto es lo que ha ocurrido en la zona de estudio. Parece pues,
que las poblaciones de A. decorticans pueden establecerse con facilidad en los
primeros estadios de la sucesión secundaría y mantenerse a lo largo de ésta en
sus aréas de distribución. La confirmación de su efectividad para la fijación
atmosférica de nitrógeno (Moro et al. 1989) permite suponer un comportamiento
diferente respecto a la economía del nitrógeno en relación con la otra especie
arbustiva.
Objetivos del presente t rabajo .
El siguiente trabajo se planteó como un primer objetivo general el
profundizar en el conocimiento de cuatro especies ampliamente distribuidas en
el área mediterránea del sureste español mediante el estudio de determinados
5
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
flujos de materia orgánica y nutrientes (desfronde, descomposición y fijación
de nitrógeno). Un segundo objetivo general consiste en esbozar el
comportamiento biogeoquímico de este sistema mixto a nivel de cuenca. Por
tanto el mayor interés se centra en las diferencias interespecíficas así como
en la interacción matorral-pinar, sin olvidar, por supuesto, la aproximación a
nivel de cuenca. Para alcanzar estos objetivos era necesario part i r de
información básica de la que no se disponía. Para ello se realizaron unos
muéstreos previos con vistas a conocer la estructura, biomasa, mineralomasa y
producción de estas cuatro especies, aspectos que se t ra tarán en el capítulo
2. También se recopilaron algunos datos climáticos de estaciones próximas,
información cartográfica y fotos aéreas.
A part ir de 1987-1988, los flujos hidrológicos en forma de deposición
global, trascolación, escorrentía cortical y avenamiento fueron estudiados por
Domingo (1991). Esta información resulta totalmente complementaria para
comprender el comportamiento global de un ecosistema joven como el que nos
ocupa. En algunos de los capítulos de esta memoria ha sido necesario utilizar
parte de estos resultados para intentar integrar el funcionamiento de los dos
vehículos principales de circulación de nutrientes en el ecosistema: la vía
hidrológica y la de la materia orgánica.
Los objetivos que específicamente se desarrollan en cada uno de los
capítulos son los siguientes:
Capítulo 1.
1.- Descripción de la estructura de las especies mayoritarias: Densidad,
cobertura, clases de tamaño y área basal. Variaciones espaciales de estos
parámetros en la cuenca.
Capitulo 2.
1.- Cuantificación de la biomasa y mineralomasa acumulada en las cuatro
especies dominantes
2 . - Composición elemental de las distintas fracciones de la biomasa en las
especies estudiadas. Comparación con bosques de coniferas y sistemas
6
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Introducción.
arbustivos.
3 . - Estima de la producción primaria neta y nutrientes asociados en las
especies mayoritarías.
Capítulo 3.
1.- Descripción de las pautas del desfronde en las cuatro especies
mayoritarías. Variación estacional e interanual. Influencia de factores
climáticos (temperatura, humedad, precipitación).
2 . - Análisis elemental del desfronde. Diferencias interespecíficas e
interfraccionales. Variabilidad estacional y su relación con la retraslocación
y lixiviación.
3 . - Estudio comparativo de las diferentes metodologías para el cálculo de la
retraslocación foliar.
4 . - Cuantificación del flujo de retorno de nutrientes vía desfronde al suelo.
Contribución de cada una de las cuatro especies.
5 . - El desfronde como indicador de la eficiencia en el uso de
nutrientes. Comparación entre especies y distintos tipos de
ecosistemas.
Capítulo 4.
1.- Compartimentalización de la vía de la trascolación neta en deposición seca
e intercambio utilizando un modelo sencillo (Lovett y Lindberg 1984)
2 . - Estima de la importancia del lixiviado en el flujo de
retorno. Comparación de la importancia de iones de origen
interno y externo que circulan por el sistema.
3 . - Participación de la vegetación en la restitución total de nutrientes al
suelo tanto por vía hidrológica como de detritus. Estima de las tasas de
circulación interna de cada nutriente.
4 . - Comparación en diferentes supuestos de estructura forestal del origen y
de las pautas de restitución de nutrientes al suelo.
Capítulo 5.
1.- Estudio de la dinámica de descomposición de la hojarasca utilizando el
7
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
método de incubación en bolsas (litterbags, Bocock y Gilbert 1957).
Comparación de la dinámica de pérdida de peso en las cuatro especies.
2 . - Aplicación de distintos modelos matemáticos para describir la dinámica de
descomposición de la hojarasca. Fiabilidad de estos modelos.
3 . - Estudio de la dinámica de liberación o acumulación de los distintos
nutrientes (N, P, K, Ca, Mg y Na) en la hojarasca de las cuatro especies.
Capítulo 6.
1.- Estimación de la fijación de nitrógeno por parte de los nodulos de A.
decorticans mediante la técnica del ARA (Hardy et al. 1973). Influencia de la
altitud en la variación estacional de este parámetro (capítulo 6).
2 . - Conocimiento de la variabilidad diaria en la fijación de nitrógeno:
Fijación y pautas diarias del ARA en dos momentos característicos del año, la
época del estrés hídrico estival, y la otoñal t ras las primeras lluvias
Capítulo 7.
1.- Ciclos y flujos de nutrientes en la cuenca según la estructura vegetal.
Comparación de las distintas especies.
8
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 1 Área de estudio.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

1.1. LOCALIZACION.
Área de estudio.
La Cuenca Nacimiento se localiza en el sector central de la Sierra de los
Filabres (Figura 1.1), provincia de Almería, entre las coordenadas 3713' y
37°14' de latitud norte y entre los 2°19' y 2°20' de longitud oeste, con
orientación NE y una altitud de 1560 (cota mínima) y 1770 (cota máxima)
m.s.n.m. La tabla 1.1 contiene resumidas algunas características físicas de la
cuenca.
1.2. CARACTERÍSTICAS GEOLÓGICAS Y LITOLOGICAS.
La Sierra de los Filabres se sitúa hacia el centro de la provincia de
Almería, siendo la prolongación oriental del macizo de Sierra Nevada,
apareciendo en su conjunto como un anticlinal tan regular y monótono como el
de aquella, pero de altura más reducida.
Geológicamente la región pertenece a la Zona Bética, el más interno de
los tres grandes dominios paleogeográficos en que se dividen las cordilleras
Béticas. La zona Bética está constituida fundamentalmente por un apilamiento
de mantos de corrimiento, distinguiéndose en dicho conjunto cuatro unidades o
complejos superpuestos. Estos, de arriba a abajo son: Complejo maláguide,
Complejo alpujárride, Complejo Baballona-Cucharón y el Complejo
nevado-filábride.
La cuenca se sitúa dentro del Complejo nevado-filábride, dentro del cual
se distinguen varias secuencias litoestratigráficas que han sido
individualizadas como formaciones. La roca madre es silícea formada por
micaesquistos cuarcíticos de la Formación Tahal. Los esquistos y cuarcitas de
esta formación se componen principalmente de cuarzo, mica blanca y clorita con
algunos de los siguientes minerales: albita, granate, epidota s.l., anfíbol,
cloritoide, biotita, distena y plagioclasa calcica, y como accesorios:
minerales metálicos, turmalina, apatito, circón y rutilo; también se presentan
localmente carbonatos. Las rocas carbonatadas tienen una composición
mineralógica bastante simple, pues existen casi exclusivamente en gruesos
cristales de carbonato con algo de mica blanca, albita y cuarzo.
9
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
LOCALIZACION DEL ÁREA DE ESTUDIO
Lorca
Cabo de Gata
FIGURA 1 . 1
TABLA 1.1. CARACTERÍSTICAS FÍSICAS GENERALES DE LA CUENCA NACIMIENTO.
LOCALIZACION
SUPERFICIE
COTA MAX
COTA MIN
ORIENTACIÓN
PENDIENTE MEDIA
TEMPERATURA MEDIA
PRECIPITACIÓN MEDIA
VEGETACIÓN
37013'-37014* LAT;2°19 ,-2O20' LONG
34 HA
1779 MSNM
1560 MSNM
NE
o 15 (aterrazada)
12
480 mm
Plnus plnaster
Plnus nlgra
Adenocarpus decortlcan*
Clstus laurlfollus
10
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Desde el punto de vista geomorfológico la cuenca se haya inscrita en la
superficie de erosión más alta que arrasa toda la parte superior de la Sierra
de los Filabres. En consecuencia el relieve es suave y presenta un
recubrimiento de regolito de potencia considerable, de alrededor de un metro,
constituido por gravas finas y limos, en ocasiones denotando su origen
periglaciar.
El suelo es de tipo cambisol-regosol-eutrico, visible en superficie, y
corresponde a los depósitos que se originaron por la erosión de los esquistos
del Paleozoico. Son suelos en general muy sueltos, poco cohesivos, con escasos
elementos finos y fácilmente erosionables. No obstante, en los rellanos
topográficos de la parte alta de la cuenca los suelos parecen ser bastante
potentes, ya que además contienen en mayor proporción elementos finos.
1.3. CLIMATOLOGÍA.
Las estaciones de referencia que han servido para conocer la temperatura
y precipitación en la zona cercana y circundante a la Cuenca Nacimiento, en un
amplio rango de años, son las de Bacares (1200 m.s.n.m., 12.4°C, media de 16
años) y 396.8 mm de precipitación (26 años hasta 1978), Calar Alto (2168
m.s.n.m., 350 mm, media de los últimos 10 años) y Tahal (1010 m.s.n.m., 361.1
mm de precipitación, media de 30 años hasta 1988).
La bibliografía disponible sobre la climatología de la zona no es muy
extensa. Destacan los trabajos sobre el clima de lai provincia de Almería de J.
Capel Molina (1976, 1977, 1981, 1984, 1986), el estudio de los pisos
bioclimáticos y de los ombroclimas de Lázaro (1988), o en sentido más amplio
la monografía sobre el clima de Andalucía de J.M. Castillo Requena (1989a y
1989b). Por su parte la Universidad de Granada en colaboración con el ICONA y
el proyecto LUCDEME ha realizado durante los últimos años estudios sobre los
suelos de la provincia que además incluyen para cada punto de muestreo la
realización de un diagrama bioclimático (ICONA 1989). Si bien estos diagramas
están calculados en muchas ocasiones a partir de datos estimados con
regresiones múltiples, puede ser en cierta medida indicativos de las
relaciones temperatura-precipitación- evapotranspiración en zonas próximas a
la cuenca de estudio.
11
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
M.J. Moro Cuadrillero
5 índice de aridez de Thornthwaite (efectividad de las precipitaciones) Figura 1.2
Prectpitaadrí mm 250 r
150
Temperatura -i30«C
20*
10*
Utüúarjon
•
Régimen de humedad- Xtheo
. — — riaüuiujuán en mra
. . . . . . Temperatura
Figura 1.3
12
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
En cuanto a las precipitaciones, Capel (1986) indica para la Sierra de
los Filabres (con datos hasta 1985) un ritmo pluviométrico anual que sigue un
esquema uniforme con máximo en primavera, seguido de otoño e invierno, y
fuerte sequía estival, y con una irregularidad interanual característica de
todo el sudeste español. En 1987, 1988 y 1989 en los pluviómetros instalados
en la Cuenca Nacimiento las estaciones más lluviosas han sido otoño e invierno
seguidas de primavera y verano (Domingo 1991). Los años precedentes al
comienzo del estudio reflejan la irregularidad antes comentada. Así de 1981 a
1985 las precipitaciones fueron inferiores a los valores medios anuales,
especialmente 1981, 1983 y 1984. El observatorio de Calar Alto registra desde
1981 a 1985 una media de 230 mm, frente a los 403 mm obtenidos en 1986, donde
comienza un ciclo húmedo que aún se mantiene. En el área de estudio se
registraron 400 mm en 1987, 514 en 1988 y 1073 mm en 1989 (Domingo 1991).
Las variaciones pluviométricas en el transecto Almería-Cuenca Nacimiento
son notables. La precipitación media anual es de unos 450 mm en la parte alta
de la Sierra de los Filabres, de unos 225 mm en Tabernas (490 m.s.n.m.) y de
217.4 mm en la ciudad de Almería, e incluso niveles del orden de 160 mm en el
extremo sureste de la provincia (Cabo de Gata).
Las precipitaciones en forma de nieve son frecuentes en toda la S. de los
Filabres. Los meses de diciembre, enero y febrero son los que presentan el
mayor número de nevadas, aunque también se producen en marzo, abril y
noviembre, e incluso en mayo y octubre (Bacares y Calar Alto). Durante el
seguimiento de la precipitación en la cuenca "Nacimiento" realizado por
Domingo (1991) (Abril 1988- Septiembre 1989) se produjeron siete eventos en
forma de nieve (de un total de 33 ) fundamentalmente durante los meses de
noviembre de 1988 y marzo de 1989, con un volumen de 373 mm frente al total de
1587 mm recogidos en los dos años.
Los centros meteorológicos más próximos al aréa de estudio en los que se
disponen de series temporales respecto a la dirección y velocidad del viento
son los de Calar Alto y Almería capital. Según el registro de la primera
estación, los vientos predominantes en periodos lluviosos fueron de componente
N y NW en un 507. y de componente S y SE en un 307... El 207. restante
13
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
presentaron componentes complejos. En 1989 el 647. de los vientos tuvieron
componente N y el 367. de S. Las velocidades oscilaron entre 1.5 m/s y 12 m/s,
con una media de 6 m/s.
La provincia de Almería participa de la caracterología del clima
mediterráneo, muy rica en bioclimas, debido fundamentalmente a su situación
geográfica y a su accidentada orografía. Abarca desde 0 hasta 2609 m de
altitud, con 4 sierras de más de 2000 m, y se dan en ella precipitaciones
desde menos de 200 mm/afto hasta más de 800 mm/año, con fuerte variabilidad
interanual. En base a la clasificación propuesta por Rivas-Martínez (1987),
Lázaro (1988) identifica la zona de estudio con el piso Supramediterráneo.
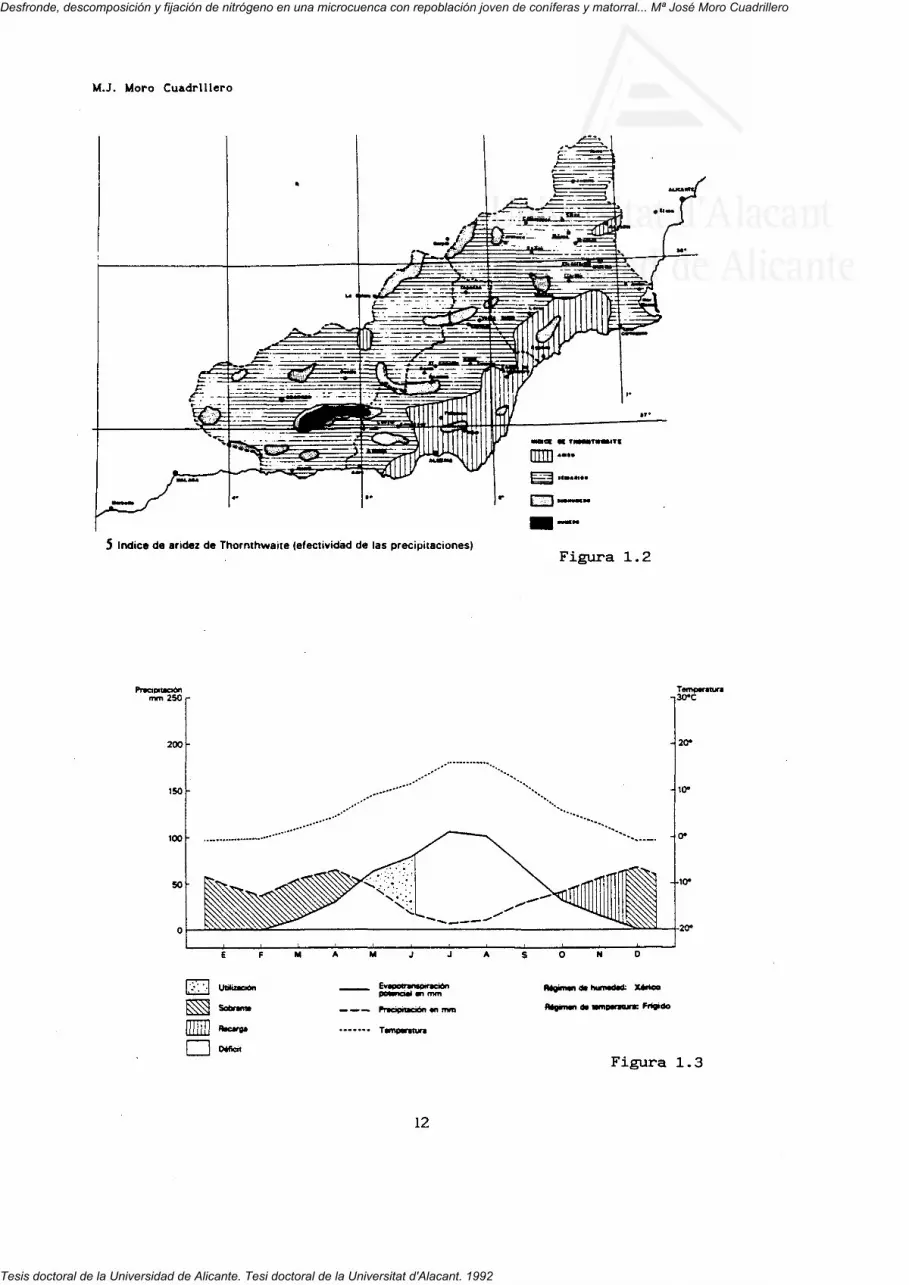
Capel Molina (1984) aplicando el índice de Tornwhaite a la provincia de
Almería sitúa la Sierra de los Filabres dentro del ombroclima subhúmedo
(figura 1.2). Hemos reproducido en la figura 1.3 el diagrama bioclimático
correspondiente al Collado del Conde (ICONA 1989), a pocos Km de la cuenca y
por otro lado similar en cuenta litología, orientación, altitud y tipo de
vegetación. La ficha climática indica una precipitación media de 480 mm, T*
media de 6.4 C, con una ETP de 510 mm y ETR de 261 mm. Existe exceso de agua
(219 mm) de Diciembre a Abril, detectándose déficit hídrico acusado en
Julio-Agosto y menos acentuado en Junio y Septiembre.
Fenómenos muy característicos de la zona son el registro de las llamadas
lluvias de barro. Según Domingo (1991), el 807. de las precipitaciones
registradas en el período 1988-1989 presentaban en mayor o menor medida un
polvo fino en Almería capital. En la cuenca de estudio tan sólo 5 eventos
lluviosos fueron clasificados como lluvias de barro durante ese período. Otra
característica del sudeste y área mediterránea española, en general, es la
presencia de tormentas y gotas frías. Durante el presente trabajo se registró
una gota fría el 7 de septiembre de 1989 durante la cual cayeron en hora y
media 115.9 mm.
1.4. ESTRUCTURA DE LA CUBIERTA VEGETAL.
Aún no siendo el objetivo principal de este trabajo el estudio de la
estructura de la vegetación, esta información es necesaria para poder
14
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Ares de estudio.
integrarla en el funcionamiento de los flujos internos considerados. Por
tanto, en este apartado se muestra, con más detalle de lo que es habitual,
información sobre la estructura del bosque.
La cuenca "Nacimiento" se encuentra repoblada con una plantación mixta de
Pinus pinaster CAiO (pino resinero) y Pinus nigra (AitXpino laricio)
instalados en 1973 excepto un pequeño rodal de P. nigra algo posterior. Las
dos especies no se solapan en la cuenca sino que cada una de ellas ocupa
aproximadamente un 507. ("figura 1.4). El año que se seleccionó la cuenca, 1986,
el pinar presentaba un aspecto semiarbustivo, con una gran porcentaje de pies
de DBH menor de 5 cm y encontrándose lejos de formar una cubierta continua.
Otra característica era la no realización de trabajos silvícolas (poda,
aclarado etc) en el pinar desde la plantación. Ello ha dado lugar a la
aparición de un estrato arbustivo espontaneo de baja diversidad pero
importante cobertura vegetal compuesto casi exclusivamente por Adenocarpus
decorticans CBoissJ, conocido con el nombre de gaznacho, y Cistus Laurifolius
L. Este matorral leñoso ocupa preferentemente los huecos dejados por el pinar
por lo que no constituye un sotobosque en sentido estricto. Esta última
característica ha de conllevar unas interelaciones ecológicas (que se irán
viendo a lo largo del trabajo) entre ambos grupos (pinar-Adenocarpus/Cistus)
diferentes a las esperables a partir de una dominancia clara del pinar.
Situaciones como la descrita si bien poco estudiadas, no son infrecuentes. En
zonas mediterráneas españolas, cuando las plantaciones de coniferas van
destinadas prioritariamente al bosque de protección y no tanto a un interés
maderero, es frecuente el que no se realicen desbroces de matorral hasta
edades avanzadas de la plantación. En estos casos es fácil encontrar
situaciones similares a la nuestra con pinares en estado semiarbustivo y un
matorral leñoso rellenado espacios libres y acumulando un buen porcentaje de
la biomasa total.
Además existe también en la cuenca un estrato subarbustivo dominado por
Artemisia glutinosa, Helichrysum serotinum y distintas especies de Thymus. El
estrato herbáceo tiene menor importancia exceptuando el sector más alto y
llano de la cuenca. Tanto el matorral subarbustivo como el estrato herbáceo no
15
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
han sido contemplados en este trabajo. El estudio solamente se refiere a las
cuatro especie mayoritarias.
UNIDADES DE MUESTREO.
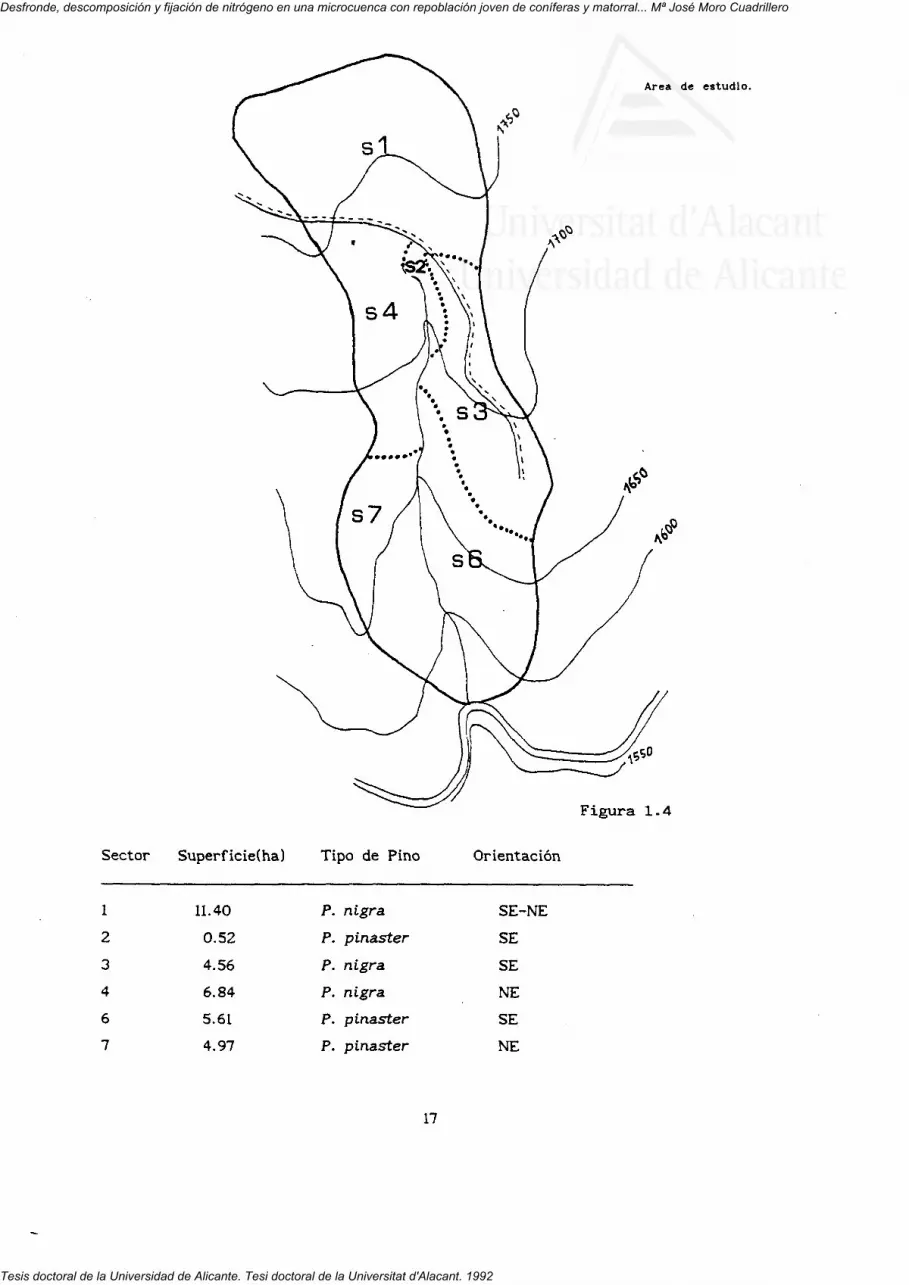
Para el estudio de la estructura vegetal, la cuenca de estudio fue
dividida en cinco transectos paralelos siguiendo las curvas de nivel situadas
a 1720, 1680, 1640, 1600 y 1560 m. Con este sistema se pretendía captar la
posible variación en la estructura vegetal siguiendo un gradiente de altitud.
Cada transecto tenía una anchura de cinco metros, lo que permitía abarcar la
totalidad estructural de la terraza (talud y rellano) y doscientos cincuenta m
de largo. Cada uno de ellos fue subdividido en diez parcelas de 40 m , cinco
en solana y cinco en umbría Esta parcelas se emplearon para el muestreo
estructural de las cuatro especies mayoritarias.
En cada parcela se contaron todos los individuos de cada especie. En el
pinar se midió en cada individuo el DBH (diámetro a 130 cm) y la altura. En A.
decorticans, se midió el diámetro basal tomado en la superficie del suelo ya
que a pocos centímetros de éste ya aparecen ramificaciones en los individuos.
En la jara , incluso el disco basal debía ser medido a unos pocos mm por debajo
del suelo ya que era allí donde se iniciaba la ramificación. Debido a la
irregularidad de los troncos, se tomaron dos y hasta t res medidas del diámetro
basal. Igualmente, se midió la altura de los individuos .Los pinos que no
alcanzaban los 150 cm de altura son considerados pertenecientes al estrato
arbustivo y su diámetro es medido a los 50 cm.
La fotografía aérea y las observaciones de campo sugerían una fuerte
heterogeneidad en la distribución de la vegetación, sobre todo en lo referente
a la densidad de las dos especies del estrato arbustivo e incluso en la
repoblación arbórea. Esta heterogeneidad no parecía seguir un gradiente
dependiente de la altitud. Por este motivo se establecieron una serie de
sectores (figura 1.4) delimitados en base a fotografía aérea y observación y
recorrido de la cuenca como una posible alternativa a utilizar como unidad de
muestreo en lugar de los transectos.
A partir de los datos obtenidos en cada parcela se calcularon los valores
16
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
ea de estudio.
F i g u r a 1.4
Sector Superficie(ha) Tipo de Pino Orientación
1
2
3
4
6
7
11.40
0.52
4.56
6.84
5.61
4.97
P. nigra
P. pinaster
P. nigra
P. nigra
P. pinaster
P. pinaster
SE-NE
SE
SE
NE
SE
NE
17
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
medios de las distintas variables. Estos resultados fueron utilizados para
calcular en cada transecto y sector variables como densidad, distribución de
clases diamétricas, altura y diámetro medio, área basal etc. El siguiente paso
consiste en utilizar estos resultados parciales para el cálculo de parámetros
estructurales de las cuatro especies a nivel de cuenca. Además, esta
información se utilizará en la selección de individuos a cortar para el
estudio posterior de biomasa y mineralomasa.
La validez tanto de los transectos como de los sectores como unidad de
muestreo representativa de la estructura vegetal se testó mediante ANOVA/test
de Fischer y Análisis factorial. Para los transectos, se incluyeron en ambos
análisis las variables densidad y diámetro medio de las cuatro especies en
cada una de las parcelas muestreadas no apreciándose diferencias
significativas para casi ninguna variable exceptuando la densidad de A.
decorticans y C. laurifolius en el transecto 1. Por su parte, al aplicar el
análisis factorial no se apreció nigún tipo de asociación de las parcelas de
cada transecto. Estos resultados en buena parte reflejo de la gran
variabilidad en las medias para los distintos parámetros en las parcelas
incluidas dentro de cada transecto. Por tanto, la validez de esta unidad como
representativa para su utilización en el estudio de la estructura de la cuenca
"Nacimiento " no parece demasiado adecuada.
En relación a los sectores, las variables incluidas en los análisis
estadísticos fueron igualmente el número de individuos y diámetro medio de
cada especie en cada parcelas a las que se añadió el número de individuos
pertenecientes a determinadas clases diamétricas. Se estableció una primera
separación de parcelas en dos grupos: las incluidas en los sectores 1, 3 y 4
cuya característica común es la presencia de P. nigra y las pertenecientes a
los sectores 2, 6 y 7 con presencia de P. pinaster. La no yuxtaposición de
ambas especies de pino es ya de entrada un factor claramente determinante de
la estructura del bosque.
El análisis de la varianza reveló diferencias significativas entre los
distintos sectores para las variables utilizadas (tabla 1.2a). El diámetro
medio de P. pinaster sirve claramente para diferenciar los distintos sectores
18
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Tabla 1.2. Arriba: Resultados de la aplicación del ANOVA a las parcelas de muestreo de estructura en
la cuenca "Nacimiento". Los resultados significativos indican diferencias entre los sectores. Las
variables consideradas son el número de individuos por parcela y el diámetro medio en cada una de
ellas. Para el matorral el análisis se ha efectuado incluyendo todas las parcelas (T) o sólo
aquellas con al menos un individuo de cada especie considerada. Abajo: resultados del test de
Neuman-Keuls. La misma tetra indica que no existen diferencias significativas para una variable dada
en los sectores considerados. *** p< 0.001; ** p<0.01; * p<0.05.
a) AHOVA VARIABLES
NUMERO PIES DIÁMETRO MEDIO
PARCELAS PP
PARCELAS PN
PARCELAS AO (>0)
PARCELAS AD (T)
PARCELAS CL (>0)
PARCELAS CL (T)
b) Resultados del test de Mn—m Keuls.
S1 S2 S3 S4 S6
VARIABLES
NPP DBHPP
NPN DBHPN
NAD (T)
NAD (>0)
DBAD
NCL (T)
NCL <>0)
DBCL
A A B B A AB A A A A
B B A A B C BA B B A
A A C C B BC CAEB
B B B
A A B C B C DB BC CB C
C B A A B C EAB B B A
***
#**
*
*
**
**
S7
B
C
A
A
B
C
FACB
C
C
C
19
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en los que esta especie está presente. Lo mismo ocurre para P. nigra. Para el
diámetro de ambas especies, los sectores son significativamente distintos (p<
0.001). Respecto a la densidad de P. pinaster, los sectores 2, 6 y 7 son
distintos (p< 0.001), no ocurriendo lo mismo para la densidad de P. nigra en
los sectores 1, 3 y 4 entre los que no se encontraron diferencias
significativas.
Los sectores son diferentes en relación tanto con la densidad como con el
diámetro respecto a las especies del estrato arbustivo. El nivel de
significación es siempre mayor en C. laurifolius.
Se aplicó un test de Newman- Keul para discriminar entre qué sectores
eran significativas (p<0.05) esta diferencias encontradas mediante el ANOVA
(tabla 1.2b) El mayor número de diferencias significativas entre sectores se
produce en relación al tamaño medio de A. decorticans.
Los resultados del análisis factorial realizado en parcelas incluidas en
la zona de P. pinaster se muestran en la tabla 1.3a. Los dos factores rotados
resultantes del Análisis explican en conjunto un 71% de la varianza. El factor
Fl agrupa significativamente al diámetro de ambas especies arbustivas y a su
densidad, ambas variables inversamente relacionadas. El F2 agrupa a la
variables diámetro y densidad de P. pinaster ambas, igualmente, inversamente
relacionadas. En la figura 1.5a se aprecian las tendencias antes comentadas.
Los sectores 7, 6 y 2 quedan claramente diferenciados respecto al Factor 1. El
gradiente en F2, relacionado con la densidad y diámetro medio del pino
resinero, está bastante menos marcado.
En la zona de la cuenca repoblada con P. nigra, los resultados del
análisis factorial aparecen en la tabla 1.4.a para las mismas variables,
densidad y diámetro medio de cada una de las especies presentes. La varianza
explicada por los dos factores resultantes es de un 77.19 %. También, en este
caso los sectores aparecen diferenciados. El sector 1 se caracteriza por una
clara asociación del pino laricio y gaznacho, (figura 1.6a) con altas
densidades de ambas especies y mayores tamaños en P. nigra. (parte positiva
del Fl). En esta zona parece que pueden darse condiciones favorables para el
20
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Tabla 1.3.Resultados del análisis factorial aplicado a las parcelas inclufdas en el área
repoblada con P. pinaster. £1 primer análisis incluye el número de individuos y el diámetro
medio de cada especie en cada parcela. El segundo análisis incluye el número de individuos de
cada parcela incluidos en unas determinadas clases diamétricas. Se representan solamente el
factor 1 y factor 2.
a) ANÁLISIS 1 b) ANÁLISIS 2
VARIABLE FACTOR 1 FACT0R2 VARIABLE FACT0R1 FACT0R2 FACT0R3
NPP
NAD
NCL
DBHPP
DBAD
DBCL
.001
.609
.605 -.390 -.814 -.885
-.920 -.218 -.564
.831
.141
.077
PP 0-4
PP 4-8
PP 8-+
A0 0-4
AO 4-8
AD 8-+
CL 0-3
CL 3-6
CL 6-+
.278
.663 -.564
.072
.089 -.737
.796
.858
.153
-.243 .294
-.207 .825 .889 .016 .384
-.224 -.296
-.798 -.385
.591 -.027 -.088
.123 -.179
.221
.787
2
1.5
1
0,5
0,5
- 1
1,5
=2
• ;
•
~
+
•+
+
+
+
+ ..
+
/"»
\ *
+
i
*
i
*
+ + •
»**\
* /
sector 2 + sector 6 * sector 7
Figura 1.5. Ordenación de las parcelas repobladas con P. pinaster en los ejes
definidos por los análisis factoriales.
21
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
crecimiento de este pino puesto que densidad y tamaño están correlacionadas
positivamente. El sector 4 puede considerarse de transición, con parcelas que
se extienden en buena parte del factor 1. El extremo negativo de este eje,
corresponde a parcelas con altas densidades de jara. En F2, el gradiente se
extiende desde mayores tamaños de C. laurifolius (parte positiva) hacia altas
densidades de esta especie y diámetros grandes de A. decorticans.i zona
negativa del eje dos).
Los análisis factoriales realizados anteriormente tienen el problema de
no incluir aquellas parcelas sin, al menos, algún individuo de las especies
estudiadas. Por ello se realizaron otros análisis del mismo tipo en el que se
utilizan como variables el número de individuos de cada especie que
pertenezcan a una determinada clase diamétrica. Para el cálculo de estos
intervalos de clase, se atendió al histograma de distribución de frecuencias
de diámetros de los individuos muestreados. Los resultados del análisis (tabla
1.3b y 1.4.b y figuras 1.5.b y 1.6.b) no añaden mucha más información a la
aportada anteriormente con las variables densidad y diámetro medio, sin
embargo incluyen una mayor número de parcelas.
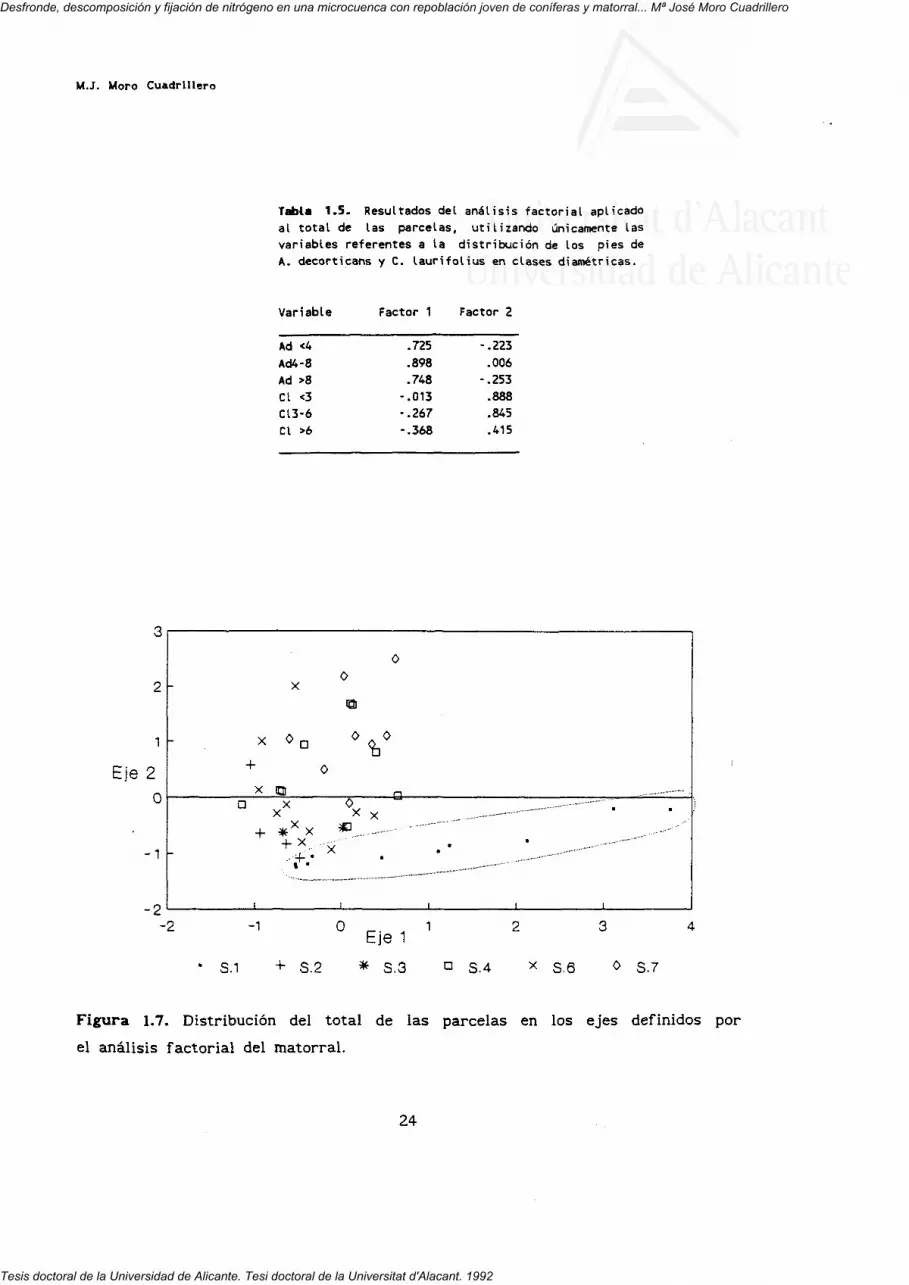
Un último análisis factorial se efectuó empleando únicamente las clases
de tamaño correspondientes a las dos especies arbustivas. Los resultados
aparecen en las tabla 1.5 y figura 1.7. El 64.7 7. de la varianza queda
explicada por dos factores. Fl separa a todos los gaznachos, aumentado su
número hacia la parte positiva del eje. F2 separa a todas las jaras con un
aumento en el número de estas hacia la parte positiva del eje. No existe buena
discriminación entre sectores, pues únicamente queda bien delimitado el sector
1.
Por tanto, a modo de resumen, en el estudio de la estructura de la cuenca
"Nacimiento", se ha llegado a las siguientes conclusiones:
-La separación de dos grandes áreas definidas por la especie de pino
presente constituye el principal criterio clasificatorio para el estudio de la
vegetación en la cuenca que utilizaremos de modo habitual en este trabajo.
-Los sectores delimitados por foto aérea y observación directa han
22
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Tabla 1.4. Resultados del análisis factorial aplicado a las parcelas incluidas en el área
repoblada con P. niara. El primer análisis incluye el número de individuos y el diámetro medio
de cada especie en cada parcela. El segundo análisis incluye el número de individuos de cada
parcela incluidos en unas determinadas clases diamétricas. Se representan solamente el factor
1 y factor 2.
a) ANÁLISIS 1
VARIABLE
NPN NAO NCL DBHPN
DBAO
DBCL
FACTOR 1
.793
.916
-.521
.798
-.350
-.290
FACT0R2
-.066
.101
-.636
.437
-.768
.918
b) ANÁLISIS 2
VARIABLE
PN 0-4
PN 4-8
PN 8-+
AD 0-4
AD 4-8
AD 8-+
CL 0-3
CL 3-6
CL 6-+
FACTOR1
.840
-.002
-.517
-.491
-.064
-.133
.847
.809
.765
FACT0R2
.133
-.055
.158
.470
.922
.893
-.053
-.239
-.295
FACT0R3
-.263
.861
.690
.619
.002
.087
-.090
-.028
-.202
F2
-2
Sector 1 + Sector 3 » Sector 4
/-.-" /
i •
1
. "\
i
A1* * > •
y v -
• • - - —
* *
»
* ¡
F1
• Sector 1 + Sector 3 * Sector 4
Figura 1.6. Distribución de las parcelas repobladas con P. nigra en los ejes
definidos por los análisis factoriales.
23
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
M.J. Moro Cuadrillero
Tabla 1.5. Resultados del análisis factorial aplicado
al total de las parcelas, utilizando únicamente las
variables referentes a la distribución de los pies de
A. decorticans y C. laurifolius en clases diamétricas.
Variable
Ad <4
Ad4-8
Ad >8
Cl <3
C13-6
Cl >6
Factor 1
.725
.898
.748
-.013
-.267
-.368
Factor 2
-.223
.006
-.253
.888
.845
.415
Eje 2
Figura 1.7. Distribución del total de las parcelas en los ejes definidos por
el análisis factorial del matorral.
24
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de e i tudlo.
resultado ser válidos y significativamente distintos para el estudio de la
estructura vegetal agrupando conjuntamente a pinar y matorral, no así los
transectos.
-Los sectores no han resultado ser válidos para el estrato arbustivo
considerado separadamente del arbóreo, ya que solo un sector quedaba
claramente diferenciado.
-Para el cálculo de variables estructurales a nivel de cuenca tales
como área basal, densidad, biomasa etc se tendrán en cuenta los valores
obtenidos para cada sector y la superficie ocupada por cada uno de ellos. Con
esta dos parámetros podrá obtenerse un valor medio poderado por superficie
para cada una de las variables en las dos grandes áreas repobladas con P.
pinaster o P. nigra.
PARÁMETROS ESTRUCTURALES.
Una vez definidos los sectores, se emplearon los resultados de las
parcelas incluidas en cada uno de ellos para estimar la densidad de cada
especie, distribución de clases diamétricas y área basal y la media ponderada
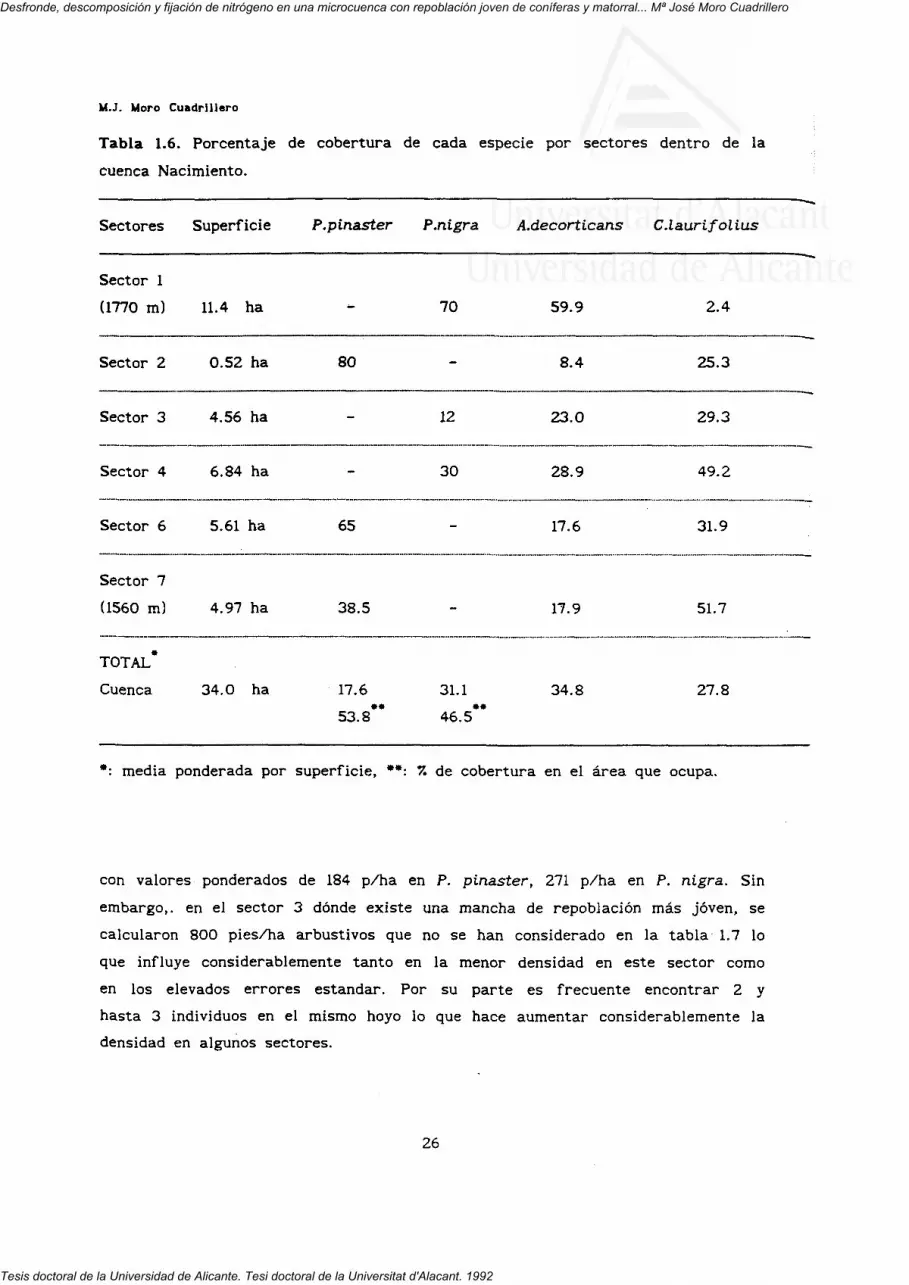
para el conjunto de la cuenca de cada parámetro. Las. estimas de cobertura de
suelo para cada especie se estimaron también en cada sector. La metodología
empleada para la estima de cobertura fue la de los trasectos lineares en las
dos especies de pino, realizándose 4 transectos perpendiculares de 50 m en
cada sector. Para el matorral, debido a la baja superposición de los
individuos, se midieron los diámetros básales de 40 individuos de cada especie
representativos de las diferentes clases diamétricas y sus correspondientes
coberturas calculando después una ecuación de regresión y aplicándola
posteriormente teniendo en cuenta las diferentes clases diamétricas de cada
sector (tabla 1.6).
Densidad.
Pinar. •
Se ha encontrado una densidad para la superficie repoblada de P. nigra
de 3671 P/ha mientras que para P. pinaster ésta resulta algo menor alcanzando
2238 pies/ha (tabla 1.7). Estos altos valores reflejan la ausencia de cortas y
aclareos en la repoblación. La contribución a la densidad total de los pinos
pertenecientes al estrato arbustivo (DBH< lcm o altura <130 cm) es muy baja
25
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
M.J. Moro Cuadrillero
Tabla 1.6. Porcentaje de cobertura de cada especie por sectores dentro de la
cuenca Nacimiento.
Sectores
Sector 1
(1770 m)
Sector 2
Sector 3
Sector 4
Sector 6
Sector 7
(1560 m)
Superficie
11.4 ha
0.52 ha
4.56 ha
6.84 ha
5.61 ha
4.97 ha
P.pinaster
-
80
-
-
65
38.5
P.nigra
70
-
12
30
-
-
A.decorticans
59.9
8.4
23.0
28.9
17.6
17.9
C.laurifolius
2.4
25.3
29.3
49.2
31.9
51.7
TOTAL*
Cuenca 34.0 ha 17.6 31.1 34.8 27.8
53.8 46.5
*: media ponderada por superficie, **: 7. de cobertura en el área que ocupa.
con valores ponderados de 184 p/ha en P. pinaster, 271 p/ha en P. nigra. Sin
embargo,, en el sector 3 dónde existe una mancha de repoblación más joven, se
calcularon 800 pies/ha arbustivos que no se han considerado en la tabla 1.7 lo
que influye considerablemente tanto en la menor densidad en este sector como
en los elevados errores estándar. Por su parte es frecuente encontrar 2 y
hasta 3 individuos en el mismo hoyo lo que hace aumentar considerablemente la
densidad en algunos sectores.
26
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Tabla 1.7. Densidad (pies/ha) por sectores y total ponderado por
superficie para el pinar en la cuenca "Nacimiento". Se indica
también el error estándard.
SECTOR P.pinaster P.nigra ES
1
2
3
4
6
7
3829
1514
2889
4800
-
1286
3365
-
-
528
491
250
571
222
595
RPP
RPN
2238
3671
RPP.Densidad media ponderada para la superficie ocupada por
el pino resinero.
RPN.Densidad media ponderada para la superficie ocupada por
el pino laricio.
La densidad del pinar no debiera estar sujeta a grandes variaciones en
los distintos sectores por t ra tarse de una repoblación de la misma edad, (si
exceptuamos una pequeña mancha del sector 3). Sin embargo esto no es lo que
ocurre en la realidad tal y como puede apreciarse en la tabla 1.7. En este
pinar no se han realizado nunca labores de aclareo, por tanto los valores de
densidad encontrados en el momento del muestreo deberían estar próximos a los
valores de stock iniciales. Las diferencias encontradas han de buscarse pues,
en la diferente mortalidad de plantones en cada sector en los primeros años
t ras la plantación ya que durante el período de estudio no se han observado
fenómenos de autotala entre individuos que comparten el mismo hoyo. Otra
explicación posible sería que realmente el stock inicial no coincida en las
distintas zonas.
27
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La distribución ponderada de clases diamétricas medidas a 130 cm se ha
representado en las figuras 1.8a para P. nigra y 1.8b para P. pinaster. En la
figura 1.9 se muestran los histogramas para cada sector. En esta primera etapa
del crecimiento del bosque, el pino laricio muestra una mayor regularidad en
la distribución de tamaños, con densidades altas pero repartidas en muy pocas
clases diamétricas. El 72% de los pies están comprendidos entre 2 y 6 cm de
DBH, alcanzando un valor máximo de 12 cm. Por el contrario, para el pino
resinero, se obtienen densidades más bajas en cada clase, pero abarcando un
mayor número de clases obteniéndose valores hasta de 20 cm.
pies/ha
« 2-4 4-8 8-8 8-10 10-12 12-14 14-1» 18-18 18-20
dbh(cm)
P.pinaster
800
400
300
100
0
pies/ha
A, decorticans
•-12
db(cm)
1900
1400
1200
1000
800
«00
400
200
pies/ha
« 2-4 4-« 8-8 8-10 10-12 12-14 14-1» 18-18 18-40
dbh(cm)
P. nigra
1800
1000
800
0
ples/Ha
WyM m
: « ;
ii i.::.:::*::;:;:;
1 0-5 3-» S-S B-12 12-18 18-18
db(cm)
C. laurifolius
Figura 1.8. Distribución ponderada de clases diamétricas en la cuenca
"Nacimiento" de las 4 especies estudiadas.
28
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Los sectores situados a mayor altitud son los que presentan una mayor
densidad en las especies de pino (sectores 1 y 2) con 4800 y 3828 pies/ha en
relación al resto de sectores. Una posible causa podría estar relacionada con
el fuerte grado de supervivencia que se ha observado para los dos e incluso 3
pies de pino por hoyo que fueron plantados en el año en que se realizó la
repoblación. Esta supervivencia pudiera estar causada por las condiciones
climáticas y edáficas más favorables que convergen en las zonas altas de la
cuenca sobre todo en el sector 1, como por ejemplo una mayor profundidad de
suelo (datos inéditos), menor pendiente (28 7. frente al 54-56 % de los demás
sectores ) o la incidencia de nieblas que se manifiestan de forma más
persistente que en las partes bajas etc. Además, si se observa las figura 1.9
en el que se representan los histogramas de clases diamétricas para cada
sector, puede comprobarse que los sectores 1 (para P. nigra) y 2 (para P.
pinaster) son rodales con una configuración más madura acercándose al aspecto
típico del bosque regular mientras que rodales como los de los sectores 4 y 7
reflejan un crecimiento claramente inferior. La coexistencia de densidades
superiores con mayor diámetro medio característicos de la zona alta de la
cuenca en relación a otros sectores parecen contradecir las explicaciones
clásicas (Yoda 1963, O'Neill & De Angelis 1981, Gracia en prensa) en las que
la tendencia apunta a una disminución de densidad con el aumento del diámetro
medio. En este caso en concreto, esas condiciones climáticas y edáficas más
favorables que hemos comentado podrían haber sido suficientes para soportar,
en estos primeros estadios iniciales del bosque un crecimiento superior de los
individuos presentes en las zonas más altas y llanas sin que los fenómenos de i
autotala hayan actuado aún de manera importante.
En ambas especies son los sectores de umbría los que presentan una
distribución más inmadura, con el mayor porcentaje de pies en las clases
diamétricas más bajas. Esta característica contrasta también con los
resultados normalmente encontrados en plantaciones de pino en zonas
mediterráneas para los que suelen encontrarse mayores crecimiento en umbrías.
Así, en poblaciones interiores de P. halepensis de la provincia de Alicante
existen diferencias significativas respecto al área basal en estaciones de
solana y umbría siendo este parámetro superior en umbrías (López y Moro n.p.).
El estrés hídrico al que se ven sometidos con mayor intensidad y duración las
29
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
des/ha i w o
1000
•00
•00
400
too
I 1 .. 1 _ 1_ .._
pies/ha
• - • »-• t - n <o-9 « - « >4-» » • • '
«h(cm)
• • — t
• - • • - « »-if *** dbh(cm)
plaa/ha
1400
i too
400
no
.piea/ha
« t-4 4-« »T4) » -» » - « « 1 4 M - »
dt¡h(cm)
tZItamr
BOO
•HO
neo
1M0
1000
•00 „
« t -4 4-« * 4 H O XMt
dbh(cm)
P.pinaster T o t a l pi es/ha
400
too
< t-4 4-« •"• »-» » - « *-M H - * «HO M-tO
toh(cm) D M O I
Wes/ha
1400
itoo
1000
*-4 4-« M » - « » - * «-M M - * 1
aerean)
1400
1M0
1000
•00
HO
0 « »-t 4-4 M »-«> «Mt
dbh(cm)
P. n/ffra T o t a l
Figura 1.9. Distribución de clases diamétricas en los diferentes sectores
definidos en la cuenca "Nacimiento" para las dos especies de pino.
3 0
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
zonas de solana suele barajarse como causa fundamental de los menores
crecimientos encontrados en estas orientaciones. En la cuenca de estudio, las
condiciones mediterráneas se encuentran muy suavizadas dados sus límites
altitudinales (1550-1740). Las nevadas son muy frecuentes desde Noviembre y se
prolongan incluso hasta finales de Marzo (Domingo 1991). Hemos observado un
considerable retraso en la fusión de la nieve en la umbría en relación a la
solana. Las heladas invernales son detectables sobre todo en umbría. En estas
condiciones y dada la precipitación media anual 450-500 mm en la zona parece
que sea el frío y no tanto el estréO estival el principal factor limitante del
crecimiento dado que el tipo de suelo y pendiente media es similar en ambas
exposiciones.
Matorral.
La densidad media para el conjunto de la cuenca alcanza para las dos
especies arbustivas mayoritarias 5912 pies/ha en los que 4080 son pies de C.
laurifolius mientras que 1832 correspoden a A. decorticans (tabla 1.8). La
distribución de esta clases diamétricas por sectores y total ponderada aparece
en las figuras 1.10 y 1.11 (sectores) y 1.8a y b (ponderadas).
Tabla 1.8. Densidad (p i e s /ha ) por s e c t o r e s y media ponderada
porsuperficie en A.decorticans y C. laurifolius.
SECTOR A.decorticans C. laurifolius.
1 3314 229
2 343 2800
3 1429 2429
4 1048 7206
6 800 4171
7 1175 10222
total p. 1832 4080
31
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La umbría de la cuenca (sectores 4 y 7) se caracteriza por una mayor
homogeneidad respecto al número de individuos de cada una de las especies,con
alrededor de 1000 pies /ha en el gaznacho y 7000-10000 pies/ha en la jara ,
mientras que en solana, la densidad varía considerablemente entre sectores. En
el sector 1, se advierte la clara dominancia del gaznacho (3314p/ha) frente a
los 228 p/ha de la jara. Estas zonas más altas, llanas y con mayor disposición
de agua (García Rosell com. personal) podrían favorecer la presencia de A.
decorticans. C. laurifolius, dado su carácter de planta invasora y resistente
a condiciones adversas (Herrera 1984, Núñez 1989) está mejor representada en
zonas de fuertes pendientes, con suelo poco profundo (taludes de terrazas por
ejemplo) así como en zonas más bajas e iluminadas (sectores 6, 7 y 4)
La distribución ponderada de clases diamétricas en la cuenca (figura
1.8.c para Adenocarpus y 1.8.d para C. laurifolius.) ha de estar relacionada
con la edad del matorral y por tanto ser indicativa de la evolución temporal
de ambas poblaciones. En ambas especies, la forma del histograma refleja unas
altas densidades en edades o clases de tamaño más bajas indicando una
situación de regeneración muy activa manifestada por el alto porcentaje de
individuos en las clases de tamaño más bajas, disminuyendo éste paulatinamente
con la edad. En Adenocarpus, esta tendencia se observa muy claramente en el
sector 1 que es el que mayor superficie ocupa en la cuenca (11 ha, figura
1.11), sin embargo el sector 4 posee una distribución normal de clases
diamétricas que confiere al rodal de A. decorticans de esta zona
características de mayor madurez (figura 1..11). En Cistus laurifolius, los
sectores de umbría (4 y 7) constituyen los representantes más claros de la
tendencia general observada en la distribución ponderada de tamaños (fig
1.12). El sector 7, al igual que ocurre con el gaznacho, es el área donde los
arbustos alcanzan menores tamaños, provocados probablemente por la competencia
intraespecífica dada su alta densidad (10000 p/ha). En los sectores de solana
(2 y 6), las poblaciones siguen evoluciones similares encontrándose una
distribución algo más madura. El mayor porcentaje de individuos grandes
(mayores de 6 cm de diámetro basal) lo tiene el sector 3. Esta población está
situada en un enclave donde la cobertura del pinar solamente alcanza un 10-15%
y el suelo es prácticamente inexistente (es un pedregal de esquistos). Con
estas condiciones se puede haber favorecido la colonización de la j a ra que
32
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
140
120
100
80
90
40
20
0
pies/ha
0-3 8-9 9-12
db
O SI
1400
1200
1000
800
900
400
200
0
pisa Aia
i i 9-S 9-12
db
O 32
1200
1000
800
900
400
200
0 9-9 9-12
db
• 33
3600
3000
2S00
2000
1500
1000
500
0
piea/ha
i 9-9 9-12 12-16
db
D s 4
2500
2000
1500
1000
500
pisa /Ha
8-9 9-12
db
C U 39
8
6
4
3
2
1
piea/ha (mil •»)
| 9-9 9-12
db
O 87
Figura 1.10. Distribución en clases diamétricas de ios pies de C.
laurifolius, en los diferentes sectores.
33
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
pies/Ha pisa/Ha 1ZOO
1000
800
800
400
200
0 0-3 3-8 e-9
D 9-12
B 12-18 16-18
100
80
80
40
20
0
• si
8-9 9-12
DB
D 3 2
pías/Ha pisa /Ha
200
0-3 3-8 8-9
D 9-12
B 12-1S 16-18
500
400
300
200
100
0 0-3 3-8 8-9
D 9-12
B 12-16 16-18
• S3 • S4
360
300
260
200
160
100
60
O
Piaa/Ha
8-9 9-12
DB
• se
800
600
400
300
200
100
O
pies/Ha
8-9 9-12
DB
• S7
Figura l.ll. Distribución en clases diamétricas
decorticans, en los diferentes sectores.
de los pies de
34
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
habría comenzado con un buen número de individuos que, posteriormente por
efecto de la competencia fueron autoeliminándose dando lugar a la situación
actual de bajas densidades y tamaños mayores con individuos que cubren buena
parte de la superficie del suelo. Una vez establecidos éstos, los huecos que
han dejado pueden ser colonizados por nuevos individuos. Esto se manifiesta,
también, por la abundancia relativa de pies con tamaños pequeños (entre 1-3 cm
de diámetro basal). Esta tendencia ha sido descrita para otros jarales de
Cistus ladanifer en Extremadura (Núñez, 1989).
ÁREA BASAL.
Pinar. 2
El área basimétrica del pinar medida a 130 cm es de 6.52 m /ha para P.
pinaster y 5.59 para P.nigra. Estos valores resultan inferiores a los dados
para plantaciones de zonas templadas con edades similares, P. taedea en USA,
P. nigra en Inglaterra, P. sylvestris en Suecia, según los datos recopilados
por Canell (1982). Así mismo, en relación con zonas mediterráneas españolas se
han encontrado valores medios de área basal superiores (16.5 en P. sylvestris,
13.8 en P. halepensis y 18.5 en P. nigra para una presencia mayor del 807. en
la comarca de Solsones (Gracia en prensa) aunque hay que tener en cuenta que
estos datos son medias de bosque o plantaciones de edades muy diferentes y con
DBH mínimo de 5 cm. Considerando sólo los mínimos que da este autor (2.9, 2.01
y 1.89), nuestros valores resultan ligeramente superiores a ellos. En bosques
de pino en la comarca del Pía de L'Estany, con una estructura ligeramente más
madura que la nuestra y con un 100% de presencia, se han encontrado valores de 2
área basal ligeramente superiores a la nuestra (de 11-23 m /ha en P. 2
halepensis y de 16 m /ha en P. nigra (Gracia en prensa).
La relación entre densidad, área basal, altura y diámetro medio en los
sectores y en la cuenca puede observarse en la tabla 1.9. El área basal total
de cada una de las especies es similar con 6.52 m /ha en P. pinaster y 5.59 2
m /ha en P. nigra quedando compensadas las diferencias de densidad y diámetro
medio que existen entre ambas poblaciones. La contribución al área basal de
cada una de las clases diamétricas puede apreciarse en la figura 1.12. En P.
35
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
800 r
600 -
400
200 "
p/Ha PP % AB PP 35
30
25
20
15
10
5
O
Densidad PP i '-'¿si % AB PP
p/Ha PN % AB PN - i50
Densidad PN ÜMi% AB pn
Figura 1.12. Contribución de cada clase diamétrica al área basal en las dos
especies de pino.
36
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
Tabla.1.9. Area basal (m/Ha), DBH medio (cm), altura (m) y densidad
(pies/ha) del pinar en cada sector, en la superficie repoblada con cada
especie, (TPP y TPN) y en el conjunto de la cuenca (T).
SECTOR DENSIDAD DBH ALTURA AB
* 1
+ 2
* 3
* 4
+ 6
+ 7
4800
3828
1285
3365
1514
2888
4.80
6.30
2.62
2 .43
8.30
2.49
5.60
3.65
2.74
2.94
2.02
1.96
3.40
2.08
2.79
2.36
9.72
13.67
0.79
1.86
10.04
1.79
6.52
5.59
TPP
TPN
2237
3671
3203 4.28 2.50 5.88
TPP= area repoblada con pino resinero,TPN= area repoblada con
pino laricio, * = pino laricio, + = pino resinero
pinaster, más del 70 7. del área basal es aportado por los pinos menores o
iguales a nueve cm de DBH, mientras que en P. nigra esta contribución aumenta
al 907.. En P. pinaster, las clases diamétricas bajas contribuyen en menor
medida al área basal que a la densidad. Sin embargo en P. nigra, las clases de
tamaño más frecuentes son las que contribuyen más al área basal.
Matorral.
El área basal del matorral mayoritario en el conjunto de la cuenca es de
12.8 m /ha, no comparable con la del pinar ya que los diámetros no se tomaron
a la misma altura. La distribución del área basimétrica por sectores y
especies aparece en la tabla 1.10 junto con los datos de densidad y diámetro
medio en cada uno de ellos.
37
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 1.10a. Densidad (pies/Ha), diámetro basal medio (cm) y área basal
(m /Ha) de A. decorticans en los sectores y en el conjunto de la cuenca.
SECTOR
1
2
3
4
6
7
TOTAL P.
DENSIDAD
3314
343
1428
1048
800
1175
1832
D.BASAL
5.01
6.72
4.49
7.53
6.04
4.27
5.53
A.BASAL
9.69
1.41
3.05
5.52
3.35
1.99
5.66
Tabla.1.10b. Densidad (pies/Ha), diámetro basal medio (cm) y área basal 2
(m /ha) de C.laurifolius en los sectores y en el conjunto de la cuenca.
SECTOR
1
2
3
4
6
7
TOTAL P
DENSIDAD
229
2800
2429
7206
4171
10222
4080
D.BASAL
5.65
4.98
6.34
3.99
4.36
3 .20
4.82
A.BASAL
0.60
7.10
9.03
13.55
8.11
10.58
7.14
Las variables densidad y área basal en los distintos sectorer presentan
una correlación significativa pero débil, (r= 0.888, p<0.01) en A. decorticans
y r= 0.774, p<0.05 en C. laurifolius), por lo que existe una sincronización
aunque no muy acusada entre el área basal del matorral y la densidad en las
diferentes zonas de la cuenca. No existe, sin embargo, correlación
significativa entre diámetro medio y área basal en los sectores para ninguna
38
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Área de estudio.
de las dos especies, ya que mientras los valores de diámetro presentan muy
poca variación (CV= 22.9 % en A. decorticans y CV= 24 7. en C. laurifolius), no
ocurre lo mismo con las áreas básales. Así, el factor densidad es el principal
causante de las variaciones que presenta el área basimétrica en la cuenca.
39
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
40
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Sección segunda. Funcionalismo de las especies estudiadas.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 2 Biomasa, mineralomasa
y producción.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mlneralomasa y producción.
2.1. INTRODUCCIÓN.
La cuantificación de la biomasa en bosque constituye una medida indirecta
de la organización de éste (Puigdefábregas 1981). La mayor parte de esta
biomasa consiste en un conjunto de árboles que forman la "canopy" o estrato
superior. Por ello los estudios sobre este tema se han centrado básicamente en
el estrato arbóreo dominante, mientras que el sotobosque y vegetación herbácea
subyacente han recibido menor consideración, si bien están cuantitativa y
cualitativamente relacionados e influenciados por por el estrato superior
(Satoo 1982). La biomasa en bosques es primariamente una función de la edad y
de la madurez del bosque ya que la materia seca se va acumulando año a año en
los troncos y estructuras leñosas (Puigdefábregas 1981, Satoo 1982). A su vez,
biomasa y producción primaria neta son parámetros claves para definir las
características de ecosistemas vegetales y el flujo de C y nutrientes
asociados, imponiendo límites superiores a la transpiración a través de la
fotosíntesis. En los estadios jóvenes existen aumentos correlativos de PPN e
índice foliar (Gholz 1982, Waring y Schlesinger 1985). En esta fase ambos
parámetros aumentan hasta llegar a una meseta que generalmente precede al
cierre de la bóveda. El tiempo en el que esta situación se alcanza es muy
variable dependiendo de las condiciones climáticas, especies, calidad del
sitio y factores ambientales. En zonas mediterráneas las principales
restricciones para la los límites máximos de LAI y de PPN están impuestas por
la reducida disponibilidad hídrica marcada fundamentalmente por la escasez de
precipitación y la irregular distribución de ésta. i
2.2. MATERIALES Y MÉTODOS.
Para el cálculo de la biomasa aérea de las cuatro especies, se
desmontaron 16 individuos de P. pinaster, 12 de P. nigra, 16 de A. decorticans
y 12 de C. laurifolius atendiendo a la distribución de clases diamétricas de
la cuenca. En los pinos se cortaba el tronco en secciones de distinto tamaño
que eran pesados en fresco tomándose después un disco de cada fracción para el
cálculo del peso seco y para los análisis químicos. Las ramas eran separadas
del árbol, se medía el diámetro basal y se pesaban individualmente. Una
muestra representativa de tres ramas de cada individuo se almacenaban para
41
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
llevar al laboratorio. En el matorral, se separaba, medía y pesaba el pequeño
tronco basal puesto que ambas especies poseen las primeras ramificaciones muy
ceca del suelo. Las ramas eran asimismo pesadas en fresco y medido su diámetro
basal. Una rama de cada individuo de A. decorticans y dos de C. laurifolias
eran seleccionadas para su fraccionamiento y análisis químico. En el
laboratorio, las ramas de los pinos eran separadas en hojas y madera de
diferente edad. Una alícuota de cada fracción se secaba en la estufa para el
cálculo del peso seco. En los matorrales se separaba la fracción foliar,
brotes, ramillas menores de dos cm y ramas. La determinación de nutrientes se
llevó a cabo sobre varias muestras compuestas de cada fracción y especie
mediante los métodos habituales para material vegetal (ver capítulo 3). Para
el cálculo de biomasa se ensayaron ecuaciones de regresión del tipo potencial
Ln(y)= Ln a + b (Ln DBH ó DB) para las distintas fracciones. La biomasa se
calculó aplicando estas ecuaciones para los individuos muestreados en cada
parcela, expresando después los resultados por ha en cada sector. Los
resultados de cada sector se poderaron por superficie para obtener la media
para cada especie en las dos grandes regiones de la cuenca. Los resultados de
biomasa se multiplicaron por la concentracción química media de cada fracción
obteniéndose así una estima de los nutrientes almacenados en la biomasa aérea
de las cuatro especies dominantes.
La Producción Primaria Neta (PPN), se estimó a par t i r de las producciones
parciales de las distintas fracciones (Satoo 1982, Whitakker y Woodwell 1971):
madera y corteza de tronco, ramas, tallos y acículas. i
La producción de madera de tronco se estimó a part ir de COREX tomados a
130 cm en 17 individuos de P. pinaster y 13 individuos de P. nigra
representativos de las diferentes clases diámetricas. En cada una de las
muestras, fueron medidos con lupa binocular y precisión de 0.1 mm. Los
incrementos radiales de los cinco últimos años, que se contrastaron con el
correspondiente DBH del árbol mediante análisis de regresión, estimándose
posteriormente el cambio en DBH experimentado para todos los pies de pino
medidos en cada sector. Aplicando a los nuevos DBH las alometrías calculadas
para madera de tronco frente a diámetro a 130 se calculó el cambio de biomasa
de esta fracción y subsequentemente la producción media anual/ha de madera del
42
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mlneralomasa y producción.
pinar en cada sector. Por último la producción se ponderó por la superficie de
cada sector para la obtención de la producción ponderada en la RPP y RPN.
La tasa de incremento de corteza fue asumida ser igual a la
correspondiente a la madera (Whittaker y Woodwell 1971, Mooney en Di Castri et
al. 1981) y fue aplicada a la biomasa total de corteza para cada especie.
La producción de ramas de dos y t res años se calculó a part i r de su
biomasa dividida por la edad correspondiente. Igualmente, para ramas mayores
de t res años se obtuvo el cociente biomasa/ edad (en este caso 10 años). El
valor medio de estas t res medidas puede considerarse una estima de la
producción media anual de madera y corteza de la fracción ramas.
Se asume que la producción de tallos y acículas es equivalente a la
biomasa de tallos y acículas del año calculada en Octubre 1986. El incremento
de la biomasa foliar en el pinar se calculó de la misma manera que la madera
de tronco, sólo que aplicando la regresión peso total de acículas - DBH a los
nuevos diámetros obtenidos a part ir del estudio de incrementos radiales. Una
forma más sencilla de calcular este incremento sería simplemente restando a la
procucción neta foliar el peso retornado por el desfronde foliar. Sin embargo,
como se verá en el capítulo 3, el desfronde fenológico de las acículas se
encuentra muy enmascarado debido a un almacén transitorio de acículas de
distintas edades que permanecen, ya caídas, sobre las ramas secas de la base
del árbol. Una cantidad aleatoria de este almacén cae cada año a consecuencia
de eventos meteorológicos produciendo picos muy importantes de los que es
difícil discriminar la fracción que corresponde al año actual o a los
anteriores.
En las especies arbustivas la formula empleada para el calculo de
producción fue:
PPN = IMP+ DF.
Donde IMP es el incremento anual de material perenne (leño y corteza) y
DF es el desfronde foliar y material reproductivo (flor, frutos y semillas).
Asumimos, por tanto que el matorral es suficientemente maduro (13-16 años
43
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en el periodo de muestreo) para considerar condiciones constantes de biomasa
foliar y por tanto se asume la igualdad entre producción foliar y
desfronde de esa fracción.
Las condiciones fluctuantes e irregulares del clima mediterráneo, se
reflejan claramente en el ritmo de crecimiento del leño de matorrales y
encinares de estas zonas (Ferrés 1984, Terradas et al. 1989) de tal manera que
los anillos de crecimiento se manifiestan con muy poca claridad y es muy
difícil establecer la correspondencia de cada uno de ellos con un periodo
determinado. En estos casos, se suele recurrir a las cronosecuencias (Sabaté
1986, Merino et al. 1990, Núñez 1989), ó a establecer cocientes biomasa/edad
(Lossaint y Rapp 1981, Rapp y Lossaint 1971, Terradas et al. 1989). En este
estudio hemos estimado la producción por este último método dividiendo la
biomasa de material leñoso/13 años de edad del matorral. La producción de
brotes se equipara al peso de esta fracción en la biomasa, mientras que el
incremento de hojas, y material reproductivo se estima a par t i r de su
desfronde medio en el periodo 86-89 (capt 3).
Los nutrientes asociados a la PPN se han calculado multiplicando la
producción anual de cada fracción por la concentración química en biomasa de
cada uno de los elementos analizados.
2.3. RESULTADOS Y DISCUSIÓN. i
2.3.1. BIOMASA.
1. Relaciones alométricas.
La relación entre la masa de las diferentes fraccciones en función del
diámetro de los individuos se ha analizado medíante funciones alométricas que
desde el punto de vista matemático son funciones potenciales de la forma:
Y= a.x o su equivalente mediante transformación logarítmica:
LN(y)= LN(a)+ b-LN(x)
ya que con ellas se obtienen las mejores estimas y son las más fáciles de
44
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mlneraloma»» y producción.
interpretar desde el punto de vista biológico. (Ferrés 1984). En las tablas
2.1 y 2.2 se especifican los parámetros de las ecuaciones generales ensayadas
en las cuatro especies estudiadas. Las alometrías son negativas en todos los
casos, entendiendo por ello que el coeficiente de alometría "b" está por
debajo de b =3 (isometría con el diámetro). Un coeficiente de b =2 significa
isometría con el area basal. La representación gráfica de las funciones
alométricas entre DBH (cm) y peso de las fracciones (g) se muestra en las
figuras 2.1, 2.2 y 2.3.
La fiabilidad de la regresión se testó, no sólo por la significación del
coeficiente de correlación (r), sino calculando el error relativo E de las
ecuaciones logarítmicas siguiendo el criterio de Whitaker and Woodwell (1968).
Valores altos y significativos de r pueden resultar engañosos a la hora de
considerar a una determinada ecuación apropiada para la predicción, ya que
estos coeficientes elevados se deben en gran parte a la amplitud del intervalo
donde se mueven las variables, y en realidad, la dispersión de los puntos a lo
largo de la linea de regresión puede ser elevada (Puigdefábregas 1981) El
error relativo (E) se define para las ecuaciones logarítmicas como el
antilogaritmo del error estandard de la estima. Este valor multiplicado o
dividido por la media de la variable dependiente nos informa acerca de la
dispersión de los puntos alrededor de la linea de regresión y puede
considerarse análogo a un coeficiente de variación (Whitaker y Woodwell 1968).
Algunos autores han apuntado que la ecuación LN(y) = LN(a) + b.LN(x) i
comporta un error sistemático para las estimas realizadas a partir de ellas
(Puigdefábregas 1981). En este tipo de ecuaciones, la linea de regresión se
ajusta más a las medias geométricas para los sucesivos intervalos de "x" que a
las aritméticas. Por tanto una estimación de "y" a part i r de un "x" dado
resultará menor que una media aritmética de un conjunto de observaciones
actuales de "y" para ese mismo "x". Esto puede llevar a una subestimación de
hasta el 20% (Lieth y Whitaker 1975).
Son varios los autores que han propuesto distintos tipos de correciones a
este tipo de ecuaciones (Flewelling y Pienaar, Madgwick en Satoo 1982).Pero,
en este trabajo no se ha considerado necesario su utilización porque el
45
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.1. Parámetros de la relación alométrica (lny= lna + b-lnx) entre el DBH y la biomasa de las diversas fracciones en ambas especies de pino. Todas las regresiones son significativas con p<0.001.
P. nigra, n = 12 (»D
Variable a b r E
Peso Total
Hojas
Ramas
Tronco
Madera Tronco
Corteza Tronco
Copa
Hojas 1 año
Brotes 1 raño
5.81
4.78
2.16
5.00
4.57
3.99
5.22
3.76
1.85
1.81
1.77
2.05
1.68
1.74
1.51
1.88
1.82
2.14
0.96
0.94
0.92
0.96
0.96
0.96
0.93
0.93
0.90
1.29
1.37
1.50
1.29
1.31
1.27
1.43
1.41
1.66
P. pinaster, n = 16 (»2)
Variable a
Peso Total
Hojas
Ramas
Tronco
Madera Tronco
Corteza Tronco
Copa
ii • i e r -Hojas 1 ano
Brotes leraño
4.92
4.19
3.27
3.85
3.37
2.96
4.50
2.63
0.57
2.17
1.93
2.28
2.27
2.35
2.05
2.09
2.12
2.38
0.97
0.92
0.94
0.97
0.97
0.97
0.94
0.89
0.90
1.32
1.54
1.56
1.31
1.32
1.31
1.47
1.73
1.77
*1. Intervalo de tamaños de DBH: [2.9-18.U (cm)
•2. Intervalo de tamaño* de DBH: [1.8-10] (cm)
intervalo de tamaños de los individuos de las especies estudiadas es
relativamente pequeño.
Los errores relativos encontrados en las dos especies de pino para las
ecuaciones que contemplan los pesos globales las fracciones de copa (hojas,
ramas) son más altas en P. nigra, lo que parece indicar que existe una mayor
variabilidad en las relaciones de dimensionales de ramas con respecto a su
46
•x&ft
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
Tabla 2.2. Parámetros de las ecuaciones alométricas (lny= lna + b-lnx) que relacionan el peso de diversas fracciones (g) con el diámetro basal (DB, cm) de las especies de matorral estudiadas. E: Error relativo.
A. decorticans, n= 16, intervalo DB: 0.9-16 cm
Fracción a b r E
Hojas
Ramas+Tronco
Tallos
Total
1.95
3.42
1.64
3.58
1.91
2.00
1.37
1.89
0.99
0.99
0.91
0.97
1.21
1.22
1.48
1.22
C. laurifolius, n= 13, intervalo DB: 0.85-13 cm
Fracción a b r
Hojas
Ramas
Tallos
Total
2.17
2.53
1.12
3.19
1.92
2.14
1.49
2.02
0.97
0.97
0.96
0.97
1.46
1.51
1.47
1.47
peso en esta especie. En el matorral, la variabilidad en la relación
tamaño-peso de ramas es más acusada en C. laurifolius tal y como lo reflejan
los errores relativos que oscilan entre 1.4-1.5 en contraposición del 1.2 de
A. decorticans. Para los pinos, las mejores estimas (menor E), se obtuvieron
para los troncos y el peso total mientras que las peores se obtuvieron para
las acículas y ramas separadas por edades sobre todo la de acículas de mas de
3 años de edad cuya variabilidad puede ser importante a causa de la mayor o
menor caida de esta fracción por hojarasca.
En la figura 2.1 y 2.2 se han representado las relaciones alométricas
entre el DBH y el peso (g) de las principales fracciones en las dos especies
de pino.
Los mayores coeficientes alométricos corresponden al peso aereo total y a
las fracciones leñosas, sobrepasando los de las ramas a los coeficientes del
47
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Peso fracción (Kg)
P. pin áster
8 10 12 DBH (cm)
tronco ramas acicuias
14 16
total
18
Figura 2.1. Relaciones alométricas entre DBH (cm) y las principales fracciones
de la biomasa en Pinus pinaster.
P. nigra
Peso fracción (kg)
HOJAS RAMAS TRONCO TOTAL
Figura 2.2. Relaciones alométricas entre DBH (cm) y las principales fracciones
de la biomasa en Pinus nigra.
48
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, minera lomasa y producción.
HOJAS + RAMAS * TOTAL
hojas * tallos x ramas A TOTAL
Figura 2.3. Relaciones alométricas entre diámetro basal (cm) y peso (Kg) de
las diferentes fracciones de la biomasa en A.decorticans (arriba) y
C.laurifolius (abajo).
49
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
tronco. Las hojas tienen coeficiente alométrico inferior a 2 pero no
significativamente distinto a este valor. La mayoría de los coeficientes en la
relación peso de acículas-DBH encontrados en pinares suelen oscilar
precisamente entre 1.5-2.5 (Whittaker y Woodwell 1968, Hendrickson et al.
1987, Leonardi et al. 1988, Lledó 1982, Gracia en prensa).
Los valores de peso seco de copa son siempre superiores a las de tronco
en P. nigra para el rango de tamaños manejado. El peso del tronco aumenta
proporcionalmente menos que el de copa (coeficiente alométrico del tronco
1.67, copa 1.88). Sorprende en esta especie este valor de "b" para el tronco
dado que en la bibliografía se barajan valores que por lo general son bastante
superiores a 2 (Puigdefábregas 1981, Hedrickson et al. 1987, Gracia en
prensa). Incluso en P.pinaster, estudiado en este trabajo el coeficiente de
alometría en troncos es de 2.27 valor que está totalmente de acuerdo con la
bibliografía. Hemos representado la relación entre peso de tronco y DBH en
ambas especie (figura 2.4). Las ecuaciones ajustadas reflejan claramente un
solapamiento de ambas curvas en la zona delimitada por diámetros pequeños cuyo
rango coincide prácticamente con el intervalo de tamaños de P. nigra (1.5-10
cm). Para este intervalo la configuración de la relación alométrica es
prácticamente idéntica en las dos especies y se manifestaría con valores de
"b" inferiores a 2. Sin embargo la relación DBH-peso cambia totalmente en el
pino resinero para tamaño superiores a lo 10 cm observándose un incremento
proporcionalmente mayor de Y frente a X que para individuos más pequeños. Al
no disponer de árboles mayores de 10 cm de DBH en P. nigra no podemos predecir
el cambio en las relaciones tamaño-peso, en esta especie aunque podemos
suponer que serán probablemente muy similares a P. pinaster y que provocarían
un aumento importante del coeficiente de alometría.
Otro aspecto a destacar en ambas especies es el elevado porcentaje de
hojas respecto al peso seco total, característica de árboles jóvenes (Satoo
1982, Puigdefábregas 1981). Un aumento absoluto del peso de hojas requiere un
aumento proporcional del soporte leñoso que las mantiene (Puigdefábregas
1981). Ello se ve claramente reflejado en las figuras 2.1 y 2.2. Las relación
alométricas del peso de hojas y peso de ramas respecto al tamaño son similares
50
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mineralomasa y producción.
Peso Tronco (kg) 35 i -
30 -
25 -
20 -
15 -
10 -
0 5 10 15 20 DBH
• P.plnaster D P.nlgra P.plnaster P.nlgra
Figura 2.4. Relación alométrica entre DBH (cm) y peso del tronco en ambas
especies de pino.
para cada una de las especies.
La relación madera de tronco/corteza de tronco es muy similar en ambas
especies, correspondiendo siempre un mayor incremento a la madera.
i
Las ecuaciones alométricas para las distintas fracciones de las especies
arbustivas de la tabla 2.2 se han representado en la figura 2.3. Interesa
señalar en principio la similitud de las pendientes de las curvas de cada
fracción entre las dos especies. Dado que el rango de tamaños manejado en los
dos arbustos es similar, podemos decir que las relaciones tamaño-peso siguen
un patrón parecido en las dos especies. Lógicamente es la fracción ramas la
que presenta un mayor incremento al aumentar el tamaño del individuo, seguido
de hojas y por último tallos. Las pendientes de las ecuaciones para la
fracción foliar no son significativamente diferentes de 2, al igual que
ocurría en el pinar. La relaciones entre el peso de las hojas/acículas y el
51
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
tamaño son pues, isométricas con el área basal.
La variada geometría y arquitectura de las especies arbustivas, permite
suponer una gran variabilidad en las relaciones y coeficientes alométricos de
estas especies pero son muy pocas las referencias que las describen. Por otro
lado, el diámetro de los arbustos es medido (dependiendo de su geometría y
tamaño) bien en la base del tronco (Gray 1983), a 130 (Puigdefábregas 1981) o
a 50 cm (Lledó 1990), lo que implica una mayor dificultad a la hora de
comparar los datos.
2. Biomasa total en las dos regiones.
Las dos especies de pino presentes fueron plantadas en el mismo año
(excepto un pequeño rodal) y por tanto las diferencias en sus biomasas se
deberían buscar en la densidad y en características específicas sin olvidar
por supuesto las condiciones microclimáticas que pueden afectar de diferente
manera a una u otra especie. Suponiendo una rápida regeneración del matorral
después del aterrazamiento la edad de éste puede oscilar entre 12 y 13 años.
Los resultados que se exponen a continuación se refieren a las cuatro especies
mayoritarias. No se incluye ni la biomasa de pies de pino menores de 1 cm de
DBH ó menores de 1 m. de altura ni la biomasa herbácea y subarbustiva.
La biomasa ponderada total para cada una de las dos grandes regiones
(tabla 2.3) es de 22.3 T/ha en la región repoblada con P.pinaster (denominada
desde ahota RPP) y de 25.1 en la repoblada con P.nigra (RPN). De estas
cantidades totales el pinar almacena el 747. en la primera y el 687. en la
segunda. La distribución de la biomasa por sectores refleja muy bien la
heterogeneidad de la cuenca, pasando de zonas donde el pinar es claramente
dominante (aunque nunca sobrepase el 75% del total) a otras en que estas dos
especies arbustivas acumulan mayor cantidad de biomasa que los pinos. Los
sectores de solana, así como el sector 1 caracterizado por una mayor
profundidad de suelo y menor pendiente, poseen biomasas alrededor de las 36
Tm/ha totales. Los sectores de umbría y el S3 ( que incluye a un pequeño rodal
de P. nigra de plantación posterior) acumulan biomasas considerablemente
menores (9.4 a 16.8 Tm/ha). En los tres sectores con mayor biomasa total
52
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mlneralomasa y producción.
1/5 0"\ 00
O. ce
n
cu cu es
t/5
\0
00 00
*-< n
en
oo
n
n
o ^
o
vo CS]
n oo
n
^3
00
n
1/5
vO
m en
O
OO
O
00
n n
n
u o a
ti
o
ni w e o m
fli i-H
A H
en
c* 00
vO
k,
d nj C - H
a. ,
ü .
a¡ U <W
•-H
c .
a.
(fl C <8 Ü
•-* k, O
u <u •a ,
• * ;
o 3
• H - H O
•-í
>-3
m —
6
j
o H
I/)
>J3
53
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
destaca ia fuerte contribución del pinar con alrededor del 75% del valor
total. En los sectores 3, 4 y 7 la contribución de las especies arbustivas
supera la biomasa del pinar alcanzando porcentajes del 60 y 75 7.. Existen
considerables diferencias en las proporciones relativas de Adenocarpus/Cistus
en algún sector. Así, en el SI la biomasa de Adenocarpus es predominante
mientras que la de la j a ra domina en todos los demás. Adenocarpus parece tener
preferencia por las zonas más llanas, húmedas y de mayor profundidad de suelo.
C. laurifolius ocupa las zonas aterrazadas de mayor pendiente características
del resto de los sectores.
Densidad y tamaño son los dos componentes de la variable biomasa con lo
que ésta seguirá básicamente las variaciones que sufran aquellos en los
distintos sectores. Diremos que las altas biomasas relativas de matorral en
algunos sectores (S4, S7) se relacionan con zonas en las que podría existir un
cierto grado de competencia intraespecífica en el pinar, de la que se han
descrito algunos indicios (informe LUCDEME 1989). Esta conllevaría menores
crecimientos en el pinar que influirían en el descenso relativo de la biomasa
de éste con respecto a la del matorral. A ello podría añadírsele el posible
efecto positivo que esta competencia pudiera tener sobre el crecimiento y
expansión de los arbustos.
La biomasa ponderada por superficie de las principales fracciones para las
especies consideradas puede observarse en la figura 2.5. También se adjunta en
la tabla 2.4 la biomasa de hojas y ramas de diferente edad para cada uno de
las especies de pino.
En C. laurifolius, el material foliar supone un 23-25 7. del total, valor
bastante concordante con el calculado para C. ladanifer en el Suroeste
español por Núñez (1989). Esta autora obtiene un porcentaje del 217. para una
población de 15 años mientras que la que nos ocupa tiene unos trece. Parece
que la tendencia apunta hacia una disminución del cociente conforme aumenta la
edad. En el estudio antes citado éste valor pasa de 14 a 0.21 en jarales desde
0 a 15 años. Igualmente Merino et ai.(1990) estudiando cronosecuencias en
sistemas arbustivos del coto de Doñana obtienen porcentajes de material foliar
que pasan del 447. a los 3 años al 137. a los 30 en el matorral de cistáceas y
54
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bloma»a, mineralomasa y producción.
Área PP Área PN
1.33
C, laurifoAua (T/ha) C. laurlfotus (T/ha)
A.decorticans (T/ha) A.decortícans (T/ha)
P.pinaster (T/ha) P.nigra /T/tia)
Figura 2.5. Distribución de la biomasa por especies y fracciones en las dos
grandes zonas de la cuenca.
55
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
del 337. al 137. en el brezal. La fracción hojas en A. decorticans supone un
alrededor de un 13 7. del peso total, porcentaje inferior al de la j a ra (figura
2.5). La tribu a la que pertenece el género Adenocarpus, (tribu Cityseae)
incluye géneros y especies áfilas (Polunin y Smythies 1977) ó con porcentajes
de hojas pequeños en relación al material leñoso. La relación material
foliar/resto es de 0.14.
La fracción tronco no aparece en la tabla como componente de la biomasa
aérea de la jara . Ello se debe a que las ramas de este arbusto se ramifican a
partir de la propia cepa a unos milímetros por debajo del suelo, no de un
tronco propiamente dicho. En A. decorticans, los troncos no suponen más que un
77. de la biomasa total ya que las ramas principales se originan a muy pocos
centímetros del suelo. En ambas especies son las ramas la fracción que acumula
el máximo porcentaje de biomasa, alrededor de un 707. muy similar entre
especies y regiones (figura 2.5).
En el pinar, la juventud de la plantación es el factor determinante por
el que las hojas constituyen un alto porcentaje de la biomasa total, (287. en
P.pinaster y 337. en P.nigra). A ello se le une la longevidad foliar observada
en las acículas ya que están caen de forma natural por desfronde a part i r del
cuarto año. Según Satoo (1982), la diferente longevidad de las acículas más
que la propia producción foliar, es el principal factor determinante de la
fuerte variabilidad observada en la biomasa de esta fracción observada en
coniferas japonesas.
En plantaciones, la biomasa foliar aumenta más o menos rápidamente hasta
que se cierra la bóveda (Marks y Bormann en Satoo 1982, Waring y Schlesinger
1985). En estos estadios la contribución relativa de hojas al total es
importante. Algo parecido ocurre con el ramaje cuyo incremento discurre
paralelo al de las hojas ya que constituye su armazón leñoso de sostén.
Conforme aumenta la madurez, las proporciones entre los distintos componentes
varían sustancialmente. Mientras que los pesos de las hojas se mantienen con
pocas variaciones, los troncos van acumulando proporcionalmente más biomasa
(Ovington en Satoo 1982) Este autor encuentra porcentajes relativos de tronco,
ramas y hojas respecto de la biomasa total muy similares a los nuestros en
56
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fracción
Acículas de 1 año
2 años
3 años
> 3 años
P. pinaster
1.51
0.70
1.10
1.27
Blomasa, mineralomasa y producción.
Tabla 2.4. Distribución de la biomasa en fracciones en P. pinaster y P. nigra
para una ha de superficie repoblada con cada una de ellas (T/ha).
P. nigra
1.70
1.40
1.18
0.87
Brotes 0.35 0.53
Ramas de 2 años 0.34 0.75
3 años 0.62 0.97
> 3 años 2.36 2.49
Madera tronco 5.38 4.55
Corteza tronco 1.79 1.74
Total 15.40 16.88
plantaciones de Pinus sylvestris de similar edad. Los valores absolutos de
biomasa foliar en las dos especies de pinos (4.75 y 5.7 T/ha) resultan
ciertamente elevados si los comparamos con pinares mediterráneos situadas en
comarcas mediterráneas más áridas o situadas a menor altitud (Gracia en
prensa). Conociendo la densidad foliar (mg/cm ) que en los rodales estudiados
es de 42.6 en el pino resinero y de 46.2 en el laricio puede estimarse de modo
grosero el índice foliar resultando éste de 1.1 para la primera especie y de
1.2 para la segunda. Estos resultados parecen indicar que en la cuenca de
estudio existen unas condiciones especialmente favorables que permiten el
mantenimiento de biomasas foliares relativamente altas. En la tabla 2.4 se
indica el peso de acículas de cada clase de edad. Las acículas de primer año,
pueden ser equiparables a la producción foliar del año 86 dado el momento del
muestreo de su biomasa (Octubre) siempre, que supongamos un aumento anual
despreciable en el peso del resto de las acículas. Los valores obtenidos son
57
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
igualmente elevados (1.5 T/ha en P. pinaster y 1.7 en P.nigra) y suponen el
31- 327. de toda la biomasa foliar mantenida en el árbol. En el pino laricio el
brote del año 86 fue especialmente intenso dado que se observaron dos pulsos
de crecimiento. Las acículas de los años 2 y 3 se mantienen en el árbol y son
posibles las pérdidas más o menos grandes por efectos atmosféricos (viento y
nieve son los principales en esta zona) ya que la acción de fitófagos no ha
sido observada. La biomasa encontrada para el segundo año es relativamente
baja respecto al tercero especialmente en P. pinaster indicando probablemente
una menor intensidad del brote del 1985. A partir del cuarto año la
variabilidad en la cantidad de acículas por árbol aumenta ya que éstas
empiezan a caer de forma natural por desfronde.
2.3.2. CONTENIDO EN NUTRIENTES.
Pinar.
En las tablas 2.5 y 2.6 aparecen las concentraciones en mg/g de las
fracciones acículas, ramas y tronco así como la concentraciones medias
ponderadas parciales y total para cada una de las dos especies. Las medias
ponderadas de hojas y ramas se han calculado ponderando por la biomasa de cada
clase de edad. La del tronco se calculó ponderando por la biomasa de madera y
corteza correspondiente. Lo mismo se hizo para hallar la media ponderada (2).
En lineas generales las concentraciones químicas elementales son similares
entre P.pinaster y P.nigra, con excepción del Mg y del K, sistemáticamente más
altos en el pino resinero, sea cual sea la fracción que se considere. Así
mismo, las concentraciones de Ca son superiores en hojas y en ramas de
P.pinaster.
Para las distintas fracciones es patente la tendencia, ampliamente
generalizada para muchas especies, consistente en la disminución de la
concentración de nutrientes al aumentar la edad de la fracciones hojas o
madera,(excepto para algunos elementos). El tronco es en general pobre en
nutrientes existiendo claramente una gran diferencia entre los niveles de los
elementos en madera y corteza. Esta última es especialmente rica en Ca,
elemento que es inmovilizado en esta estructura. Lo mismo puede observarse en
la madera y corteza de la fracción ramas.
58
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mineralomasa y producción.
Tabla 2 .5 . Concentraciones medias (mg/g) para cada elemento y f racc ión y medias ponderadas para ho ja s , ramas, t ronco y t o t a l en P.pinaster.
FRACCIÓN
ACICULAS 1 ANO
ACICULAS 2 "
ACICULAS 3 "
ACICULAS N "
MEDIA P.
BROTES AÑO
RAM 2 AÑO
RAM 3 AÑO
RAM N AÑOS
MEDIA P(l)
MADERA TRONCO
CORTEZA TRONCO
MEDIA P
MADERA RAMAS
CORTEZA RAMAS
MEDIA P(2)
Media P. TOTAL
N
9.22
8.53
8.05
6.65
8.12
7.50
4.80
3.65
2.92
3.67
1.18
2.44
1.50
2.00
2.85
2.26
4.01
P
1.05
0.83
0.75
0.76
0.86
1.10
0.58
0.39
0.30
0.42
0.19
0.13
0.10
0.22
0.27
0.23
0.43
K
4.53
3.19
2.04
2.35
3.13
3.74
2.52
2.01
2.00
2.22
1.45
1.80
1.54
2.50
2.80
2.58
2.18
Ca
2.27
5.98
7.02
5.30
4.83
3.88
4.82
5.27
5.02
4.11
1.50
3.85
2.10
1.10
8.65
3.33
3.30
Mg
1.67
2.04
2.02
1.60
1.79
1.37
0.96
0.93
0.90
0.96
0.50
0.65
0.53
0.60
0.95
0.73
1.01
Na
0.16
0.16
0.20
0.17
0.17
0.13
0.14
0.10
-
0.10
0.14
0.10
0.12
0.12
0.39
0.19
0.12
El orden de los distintos elementos en el conjunto de la biomasa es:
N> Ca> K> Mg> P> Na
Aciculas.
Las aciculas constituyen una fracción metabólicamente muy activa y de
ciclo rápido ; por ello es la más rica en todos los nutriente excepción hecha
del Ca, elemento que se encuentra en mayor concentración en tejidos
59
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.6. Concentraciones medias (mg/g) para cada elemento y fracción y medias ponderadas para hojas, ramas, tronco y tntal *»n P.rtiera. total en P.nigra
FRACCIÓN
ACICULAS 1 AÑO
ACICULAS 2 "
ACICULAS 3 "
ACICULAS N "
MEDIA P.
BROTES AÑO
RAM 2 AÑO
RAM 3 AÑO
RAM N AÑOS
MEDIA P(l)
MADERA TRONCO
CORTEZA TRONCO
MEDIA P
MADERA RAMAS
CORTEZA RAMAS
MEDIA P(2)
Media P. TOTAL
N
9.17
8.60
8.29
7.25
8.57
4.85
4.10
3.46
2.86
3.40
1.32
2.73
1.71
2.00
3.43
2.5
4.54
P
0.90
0.83
0.84
0.86
0.86
0.71
0.53
0.45
0.39
0.46
0.09
0.13
0.10
0.17
0.40
0.25
0.46
K
2.15
1.92
1.63
1.54
1.89
2.08
1.31
1.27
1.20
1.33
0.80
0.50
0.71
1.40
1.80
1.54
1.29
Ca
1.69
3.03
4.74
4.20
3.01
2.87
3.08
3.16
3.24
3.00
1.85
3.90
2.42
1.10
5.70
2.77
2.78
Mg
1.13
1.16
1.18
1.55
1.13
1.20
0.79
0.84
0.80
0.85
0.30
0.62
0.38
0.50
1.00
0.68
0.77
Na
0.17
0.09
0.10
0.09
0.12
0.09
0.10
0.09
0.09
0.09
0.08
0.06
0.07
0.09
0.10
0.09
0.09
lignificados. Los valores encontrados son muy similares entre una y otra
especie a excepción del K.(tablas 2.5 y 2.6). El orden de importancia de los
nutrientes en acículas de distintas edades es para ambas especies:
N > Ca > K > Mg > P > Na
La única excepción a este orden es el segundo lugar que ocupa el K en las
acículas del año por delante del Ca, ya que este último se encuentra em menor
60
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, tnineralomasa y producción.
cantidad en los tejidos jóvenes que se encuentran menos lignificados.
En las acículas de coniferas puede observarse una variación de
concentración, a veces marcada, entre las acículas de diferente edad (Rodin y
Bazilevich 1967). En este trabajo pueden advertirse también algunas de estas
tendencias. Para P.pLnaster, N, P y K presentan correlación negativa y
significativa (p< 0.05) con la edad de las acículas. Hay, en los t res casos,
una disminución de la concentración con la edad, si bien, la tendencia es
lineal para el N y potencial (tendiendo a una estabilización en las mayores
edades), para el P y el K. El Ca presenta correlación positiva y logarítmica
(potencial) con la edad, es decir, se produce un aumento en el contenido de Ca
tendiendo a alcanzar un valor constante o ligeramente inferior en las hojas
más viejas. La correlación no resultó significativa pero esto puede deberse a
los grados de libertad que se están manejando (2 grados).
En P.nigra, las tendencias del N y K son similares a las de P.pinaster,
existiendo una correlación significativa y negativa entre concentración-edad
(p<0.05) aunque las pendientes son algo menos marcadas. El Ca es el único
elemento que se concentra al aumentar la edad, no apreciándose disminución
alguna en la acículas de más edad como ocurría en la especie anterior. La
correlación encontrada sí resultó ser significativa (r=0.95, p<0.05). Rodin y
Bazilevich (1967) señalan disminuciones en la concentración de N, usualmente
en el P y particularmente en el K, en pinares y otras coniferas lo que
coincide con nuestros resultados. Para el Ca y Na describen un incremento al
aumentar la edad lo que solamente acontece de forma significativa para el Ca
en acículas de P. nigra.
Se han recopilado en la tabla 2.7 las concentraciones (7. peso seco) en
acículas de algunos pinares así como la concentración media de éstos. También
se ha incluido una media calculada por Ferrés (1984) a partir de 15 bosques
templados de conifera recopilados por Colé and Rapp (1981). Como puede
observarse las concentraciones de N y Ca para las especies estudiadas son en
ambos casos inferiores a las medias calculadas, aunque solamente el contenido
en Ca de P.nigra queda fuera de los límites de confianza de la media obtenida
para las especies de la tabla 2.7. Algunos autores han destacado que los
61
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla.2.7. Concentraciones ('/. m.s.) en acículas de distintas especies del género Pinas. L.C.: Límite de confianza 95%.
Especie
P.contorta P. resinosa P.nigra 3
P.sylvestris P.sylvestris P.pinnea P.halepensis P.halepensis P.pinaster P.radiata P.sylvestris P.uncinata P.nigra9
P.halepensis P.halepensis
MEDIA
L.C.
MEDIA(Ferrés)
P.pinaster P.nigra
Localidad
Wyoming(USA) USA Inglaterra Inglaterra URSS SE Francia Murcia SE Francia SE Francia N.Zelanda Cataluña Cataluña Cataluña Cataluña Grecia
[.7-1.
Almería Almería
N
.88
.68 1.15 -1.29 .82
1.00 -------
.94
.96
.11 [.07-
1.03
.81
.85
P
.08
.07
.07
.13
.16 -
.08 --
.11
.09
.07
.09
.07
.06
.09
.11] [.39-
.12
.08
.08
K
.43
.30
.38
.56
.45 -
.51
.53
.43
.73
.54
.47
.44
.27 —
.46
.53] [.
.58
.31
.19
CA
.46
.76 .74 .62 .34 -
.70
.41
.24
.48
.40
.34
.45
.93 —
.53
,41-.651
.61
.48
.30
MG
.11
.18
.12
.09
.10 -.20 .17 .12 .14 .11 .09 .10 .22 —
.13
[.11-.16]
.12
.18
.11
(1). Pearson et al. (1987). Wyoming (USA). (2). Rodin & Bazilevich (1967). Datos de Lutz & Chandler, (1947). (3). Rodin & Bazilevich (1967). Datos de Ovington,(1956). (4). Rodin & Bazilevich (1967). Datos de Remezov (1959). (5). Rapp et al. (1979). Provenza (Francia). (6). Lledó. (1982). Sierra Espuña (Murcia). (7). Passama et al (1970). S.E. Francia (8). Forrest & Ovington (1971). (9) Gracia (en prensa).Inventary forestal de Catalunya. Solsones. (10). Specht et al (1988). Datos de Margaris et al. (1984). Maquis griego. (11). Presente trabajo.
niveles de Ca en órganos vegetales están influenciados por la mayor o menor
abundancia de este elemento en el suelo (Passama 1970). La roca madre silícea
en la que crece el pinar de estudio podría ser una de las causas para los
bajos contenidos en Ca encontrado en las acículas de las dos especies. Los
contenidos de K son bajos en las dos especies estudiadas en relación a otros
62
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
pinares y quedan situado en ambos caso fuera del límite de confianza de la
media. Estos bajos niveles podrían indicar en principio, menores
requerimientos nutricionales respecto al K.
Tallos y ramas.
Los valores de concentración en tallos del año son bastante similares a
los encontrados en las acículas del año en ambas especies. Existe alguna
excepción clara como la del N que es siempre más alta en la fracción foliar.
El cambio de concentración al pasar de primero a segundo año está mucho más
acentuada en tallos que en acículas ya que los cambios estructurales que
experimentan los primeros son bastante más drásticos. Las concentraciones
medias ponderadas para el N, P, Mg en ramas de P.pinaster y para el N, P en
P.nigra son del orden de la mitad a las encontradas en acículas y más
similares respecto al Ca ,K, y Na.
El valor medio ponderado para madera y corteza de ramas que aparece en la
tabla 2.5 y 2.6 con un (2) ha sido calculada para ramas de 1 y más años, por
ello,al no estar incluidos los brotes que constituyen la fracción más rica en
nutrientes de las ramas, se obtienen valores medios algo más bajos que en en
la media ponderada (1) que sí considera los brotes. En P.nigra, el contenido
en N es superior al de Ca si se incluyen los tallos del año junto con el resto
de las ramas debido al la alta contribución del N y la baja del Ca en brotes.
Si se excluyen éstos, el Ca pasa a ser el elemento mayoritario en esta
fracción (media ponderada 2 en la tabla), puesto que se conjugan menores
niveles de N con mayores de Ca.
Ferrés (1984), a partir de datos de 17 bosques templados de coniferas
calcula unos valores medios que se presentan en la tabla 2.8 junto con los
nuestros.
El orden que ocupan los elementos es en el conjunto de ramas y brotes de
las dos especies estudiadas es:
P.nigra N > Ca > K > Mg > P > Na
P.pinaster Ca> N > K > M g > P > N a
63
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
En general este orden coincide con la bibliografía exceptuando el primer
lugar que ocupa el Ca en P.pinaster a causa de la relativamente alta
concentración de éste en ramas. El contenido en Ca de ramas en P.pinaster (4.1
mg/g) se encuentra prácticamente en el límite superior del intervalo de
confianza dado por Ferrés (1984) para esta fracción (4.2 mg/g). Niveles de Ca
para ramas de coniferas superiores a 5 mg/gr han sido encontrados en
bibliografía para un pinar de Pinus halepensis creciendo en suelo con roca
madre calcárea. Así, LLedó (1982) encuentra hasta 10 mg/g en ramas de esta
especie en Sierra Espuña. Al igual que para las acículas, en ramas, los
contenidos de K son más bajos que la media de las coniferas estudiadas por
Ferrés (1984) especialmente en P. nigra cuyo valor queda fuera del límite
inferior de confianza para la media (2 mg/g).
También en las ramas pueden apreciarse en algunos casos tendencias
químicas según la edad. Así, para P.pinaster, se aprecia una disminución
progresiva de concentración al aumentar la edad para los elementos N y P. Para
estos elementos las correlaciones concentración -edad son significativas
(p<0.001) ajustándose la curva a una ecuación potencial. Los niveles de Ca
aumentan rápidamente para luego estabilizarse posteriormente. La relación es
potencial pero no significativa. En P.nigra, el contenido en N de las ramas
disminuye de una manera casi constante con la edad; por tanto la relación
entre ambos parámetros es linear (r=-0.998, p<0.005) a diferencia de lo que
ocurre con P.pinaster. Para el P se repite el mismo patrón que en el pino
resinero siendo significativas las correlaciones a nivel (p< 0.001, r= -0.999)
El patrón que sigue el Ca es similar al de P.pinaster, pero se han obtenido
mejores correlaciones, (r=0.97,p<0.05).
Tronco.
Los resultados para madera y corteza y la media ponderada aparecen en
las tablas 2.5 y 2.6. La pauta general es la de una mayor riqueza en
nutrientes de la corteza exceptuando al Na y K en P.nigra junto con P y Na en
P. pinaster en que los valores son bastante similares en ambas fracciones.
Comparando nuestros valores ponderados con la media de Ferrés (1984) para
bosques templados de coniferas (tabla 2.9), destacan en lineas generales las
64
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomata, mlneralomasa y producción.
Tabla 2.8. Concentraciones (mg/g) para las ramas de
distintos bosques templados de coniferas (según Ferrés,
1984) y para las dos especies en estudio.
ESPECIE N P K Ca Mg Na
CONIFERAS 4.07 .53 2.77 3.53 0.61
P.pínaster 3.67 .42 2.22 4.11 0.96 0.10
P.nigra 3.40 .46 1.33 3.00 0.85 0.09
Tabla 2.9. Concentraciones medias (mg/g) para troncos de
bosques de coniferas (según Ferrés 1984) y de las dos
especies en estudio.
N P K Ca Mg
CONIFERAS 1.33 0.21 0.86 1.85 0.21
P.pínaster 1.50 0.10 1.54 2.10 0.53
P.nigra 1.70 0.10 0.71 2.42 0.38
concentraciones superiores en nuestras especies para casi todos los elementos
exceptuando el P que resulta inferior a la media de Ferrés tanto en P.
pinaster como en P. nigra.
En las tablas 2.10 y 2.11 se muestran algunos de los resultados obtenidos
para madera y corteza en troncos de diferentes especies de pino. El orden de
elementos coincide, en general, con lo apuntado por Rodin y Bazilevich (1967)
65
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.10. Contenido en nutrientes (% M.S.) en madera de diferentes pinos.
ESPECIE
P.sylvestris P.uncinata P.nigra1
P.nigra1 t
P.halepensis P.sylvestris P.halepensis White pine Red pine
MEDIA
4 P.pinaster P.nigra
Localidad
Cataluña Cataluña Cataluña Cataluña Cataluña Cataluña Murcia Canadá Canadá
Almería Almería
N
—
-----.160 .120 .056
.125
.120
.130
P
.015
.012
.013
.015
.014
.014
.014 -—
.014
.019
.009
K
.062
.030 -
.095
.077
.063
.099
.060
.025
.064
.145
.080
CA
.152
.196
.165
.166
.216
.129
.141
.072
.098
.148
.150
.185
MG
.024
.022
.017
.026
.026
.021
.036
.024 —
.024
.050
.030
(1). Gracia (en prensa). Inventari forestal de Catalunya. Solsones. (2). Lledó. (1982). Sierra de Espuña. (Murcia). (3). Hendrickson eí al. (1987). Great Lakes. Ontario. (Canadá) (4). Presente trabajo.
Tabla 2.11. Contenido en nutrientes en cortezas de diferentes pinos (% M.S)
ESPECIE
P.sylvestris P.uncinata P.nigra P.nigra1 t
P.halepensis P.sylvestris P.halepensis White pine Red pine
MEDIA.
4 P.pinaster P.nigra
LOCALIDAD
Cataluña Cataluña Cataluña Cataluña Cataluña Cataluña Murcia Canadá Canadá
Almería Almería
N
------.320 .630 .450
.460
.240
.270
P
.016 -.016 .017 .014 .014 .053 -—
.021
.013
.013
K
.039
.034
.035
.030
.060
.026
.390
.384
.238
.137
.180
.050
CA
.617
.821
.686
.816 2.360 .883 .666 .464 .547
.873
.385
.390
MG
.026
.018
.017
.021
.048
.011
.177
.149 —
.050
.065
.062
(1) Gracia (en prensa). Inventari Forestal de Catalunya. Solsones. (2) Lledó (1982). Sierra de Espuña (Murcia). (3) Hendrickson et al (1986). Great Lakes, Ontario (Canadá). (4) Presente trabajo.
66
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
para la madera, existiendo un predominio del Ca seguido por el N y el K. El
contenido en elementos minerales de la madera de tronco en las dos especies
estudiadas suma un 0.49 7. en P.pinaster y un 0.44 7. en P.nigra que les
aproxima al valor de 0.5% dados por estos autores para otros bosques de
coniferas. Respecto a la corteza, los valores de Ca y Mg expresados en la
tabla 2.11 son bastante variables. Las concentraciones de Ca encontrados en el
pino laricio de Lérida resultan inferiores a los encontrados para la misma
especie en Almería, sin embargo son superiores respecto al Mg. El contenido en
N de P.halepensis (Lledó 1982), P.nigra y P.pinaster (presente estudio), es
inferior a los de los pinos americanos que aparecen en la tabla.
Matorral .
En Europa, se disponen relativamente de pocos datos respecto a los
contenidos de nutrientes en matorrales o sistemas arbustivos. Una importante
recopilación aparece en Specht (1988) para sistemas arbustivos de Francia,
Australia, Sudafrica, Grecia y California, pero solamente incluyen contenidos
foliares. En España cabe destacar los trabajos de Martín (1984) para el
espinar costero alicantino, Clemente (1983) para arbustos y lianas de un
encinar en Sierra de Prades (Tarragona) y Picólo (com. personal) en arbustos
forrajeros. Son más abundantes los trabajos en la garriga francesa (Lossaint y
Rapp 1971, Rapp y Lossaint 1981, Mooney 1981) y sobre todo los realizados
para sistemas arbustivos de Chile y California de los que existen buenas
recopilaciones en Gray y Schlesinger (1981), Shaver (1981) y Gray (1983).
En las tablas 2.12 y 2.13 aparecen las concentraciones medias (mg/g)
para las distintas fracciones de la biomasa, así como la media ponderada para
cada especie arbustiva estudiada.
Tallos del año, y sobre todo hojas poseen las concentraciones más
altas en todos los elementos puesto que son metabólicamente más activas.
Además las hojas son lugares preferentes de traslocación de nutrientes, (Gray
y Schlesinger,1981). El orden que ocupan los elementos en las fracciones es:
67
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla.2.12. Contenido en nutrientes (mg/g) en las distintas fracciones
de A.decortica.ns junto con la media ponderada para la especie.
FRACCIÓN N P K Ca Mg Na
Hojas 29.00 1.19 6.40 4.43 2.10 0.16
Tallos 14.30 0.86 7.20 2.00 0.95 0.39
Ramillas 10.70 0.51 3.79 1.10 0.59 0.16
Madera 8.36 0.24 3.60 3.50 0.81 0.10
Total 10.10 0.36 3.90 2.70 0.89 0.11
Tabla 2.13. Contenido en nutrientes (mg/g) en las distintas fracciones
de C.laurifolius y media ponderada para la especie.
FRACCIÓN N P K Ca Mg Na
Hojas 13.71 1.50 4.89 10.50 2.17 0.12
Tallos 6.37 0.94 4.24 8.00 1.08 0.12 i
Ramas 4.90 0.50 1.24 8.90 0.53 0.09
Total 7.03 0.75 2.22 9.30 0.94 0.10
Cl: N > Ca > K > Mg > P > Na
Ad: N > K >Ca > Mg > P > Na
La principales diferencias entre las dos especies se refieren al N, K y
Ca. La riqueza de N y K es significativamente más elevado en A. decorticans
para cualquiera de las fracciones mientras que los contenidos de Ca en esta
68
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mlneralomasa y producción.
especie resultan del orden de 3-4 veces inferiores a los de C. laurifolius.
Por el contrario, en el gaznacho destacan los elevados valores de N en todas
las fracciones. Concretamente en hojas, el valor de 29 mg/g de N es
prácticamente el doble del de de Claurifolius y superior igualmente a los
encontrados en bibliografía para arbustos mediterráneos. La condición de
planta fijadora efectiva de A.decorticans parece una razón suficiente para
explicar estos valores elevados.
Hojas.
La riqueza en N de la fracción foliar de A. decorticans es, como hemos
comentado muy superior a la de Claurifolius y a la encontrada en
bibliografía para arbustos mediterráneos (tabla 2.14). La condición de planta
fijadora efectiva de A decorticans parece una razón suficiente para explicar
estos valores elevados. En la tabla 2.15 se han seleccionado datos de
distintos autores recopilados en Specht (1988) sobre composición química
foliar de otras leguminosas arbustivas procedentes de distintos ecosistemas
mediterráneos . Los altos contenidos en N (que oscilan entre los 14 y 37 mg/g)
parecen una característica común y definitoria de estas especies. Puede
apreciarse que la concentración de N en el gaznacho está próxima a la de
Cercis siliquastrum y Medicago arbórea. Los 29 mg/g de A.decorticans están
ligeramente por encima del límite de confianza superior de la media. Todo
parece apuntar a un consumo de lujo de N para las leguminosas mediterráneas y
en concreto A. decorticans.
A.decorticans se caracteriza también por unos bajos contenidos foliares
de Ca (4.43 mg/g) que, a pesar de un intervalo de confianza amplio para la
media en hojas de arbustos mediterráneos (tabla 2.14), queda por debajo del
límite inferior. La baja lignificación de las hojas de A.decorticans, en
contraste con la fuerte esclerofilia de la mayor parte de los arbustos
tipicamente mediterráneos, puede ser una de las causas para la escasa
contribución de este elemento. Según podía apreciarse en la tabla 2.15, otras
leguminosas propias del matorral del sureste francés presentaban contenidos de
Ca inferiores incluso a los del gaznacho. Por el contrario, los elementos P, K
y Mg en esta especie se encuentran dentro del rango encontrado para el resto
de los matorrales.
69
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.14. Composición química foliar (mg/g) para distintas especies/sistemas de matorral mediterráneo.
Especie/Sistema 1
Artemisia cal i fornica
Salvia leucophylla
Ceanothus megacarpus
Chaparral California
Matorral Chile
Garriga francesa
Phillyrea angustí folia
Arbutus unedo
Viburnun tinus
Kwongan Australia
Chaparral marítimo
Adenostoma fasciculatum
Arctostaphylos sp.
Vaccinium ovalis
Ceanothus sp
Matorral costero Chile
Matorral montano Chile
Maquis francés
Maquis francés
MEDIA L.C.
Adenocarpas decorticans Cistus laurifolius
Ftef.
(i)
(i)
(i)
(2)
(2)
(3)
(4)
(4)
(4)
(S)
(6)
(7)
(7)
(7)
(7)
(8)
(9)
(10)
(11)
[10.3-
(12)
(12)
N
18.8
18.5
16.9
11.0
12.0
13.5
10.8
11.0
9.1
6.7
8.4
10.8
7.2
6.5
15.0
13.4
14.6
15.3
11.0
12.1 -13.9] [.93-
29.6 13.7
P
2.25
2.00
0.82
2.50
1.70
0.80
0.80
1.11
0.97
0.30
0.70
1.20
0.80
0.50
1.00
1.40
1.8
1.5
1.1
1.2 -1.5] [6.3-9]
1.2 1.5
K
4.2
15.4
4.5
5.2
6.3
8.1
7.7
8.0
9.7
6.2
-
5.4
4.7
8.7
6.0
10.6
9.0
9.3
8.7
8.1 [9.2-12.5]
6.4 4.9
Ca
9.4
14.1
8.7
8.6
19.8
11.5
10.2
10.0
13.2
10.7
-
8.2
7.4
4.4
10.3
10.4
12.3
15.5
10.8
10.8 [1.9-3]
4.4 10.5
Mg
2.6
2.9
1.6
2.2
5.3
1.4
1.2
1.9
3.1
2.2
-
2.1
1.3
1.6
2.3
5.5
3.2
3.6
1.9
2.5
2.1 2.2
(1) Gray, J.T. (1983). Especies de chaparral y Coastal scrubs (2) Shaver, G.R.(1981) Chaparral californiano y matorral chileno (3) Rapp, M. & Lossaint P.(1981) .Garriga francesa (4) Clemente, A. (1983). Sotobosque de encinar (5) Brown, J.M. & Hopkins.A.J.M. (1983).Kwongan del Oeste Australia (6) Griffin, J.R. (1978). Chaparral marítimo,California (7) Specht, R.L.& Molí, E.J.(1983).Mediterranean shrublands.California (8) Rundel, P.W. (1981).Matorral costero chileno. (9) " " " .Matorral montano chileno. (10) Specht, R.L.(1988).Maquis francés,(Marsella) (11) Brun.B.,Conrad, L. & Gamisans J.(1975).Maquis francés,(Córcega) (12) Presente trabajo.Matorral del sureste español. Referencias (5) a la (11) recopiladas por Specht, R.L.(1988)
70
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomama, mineralomasa y producción.
Tabla 2.15. Composición química foliar para distintas especies de leguminosas en matorrales mediterráneos. Concentraciones en mg/g.
ESPECIE
Calicotome espinosa.
Ulex parviflorus.
Ceratonia siliqua.
Cercis siliquastrum. 2
Anthíllys hermanníae. Calicotome vi llosa.
Genista acanthoclada. 2
Medí cago arbórea. 2
Psoralea bituminosa. 3
Adenocarpus decorticans.
Media
L.C. [IÉ
N
16.5
14.1
15.2
23.1
20.5
18.9
17.1
34.2
36.9
29.6
21.8
¡.7-28]
P
1.0
0.6
0.7
1.8
1.3
0.9
1.7
1.3
1.8
1.2
1.2
[0.9-1.51
K
5.3
7.1
-
-
-
-
-
-
-
6.4
6.2
Ca
2.5
2.2
-
-
-
-
-
-
-
4.4
3.1
Mg
1.7
1.4
-
-
-
-
-
-
-
2.1
1.7
(l).Specht eí al. (1983). Maquis francés:(Herault y Provenza). (2).Margaris eí al. (1984). Maquis y frigana griega. (3).Presente estudio. Matorral mediterráneo del sureste español. (1) y (2).Datos recopilados en Specht (1988).
Los contenidos en nutrientes de C. laurifolius, se ajustan bien a los
rangos encontrados en otros matorrales para todos los elementos excepción
hecha del K que queda fuera del límite de confianza inferior de la media de
arbustos mediterráneos. Hemos recurrido a la tabla 2.16 en la que se recoge la
composición química foliar de algunas de las especies de Cistus presentes en
la zona mediterránea (Specht 1988) para estimar el rango de variación en la
concentración de K. Las variaciones de la cantidad de K en hojas no sólo puede
reflejar diferencias específicas sino que pueden ser indicativas del estado
nutricional de la planta y de la disponibilidad de este elemento en el suelo
(Epsten 1972). Puede comprobarse que es precisamente este elemento junto con
el Ca el que presenta una mayor variabilidad (cv= 647.). C. laurifolius posee
concentraciones de K encajadas dentro de los límites de confianza de la media
aunque más próxima al inferior asociada a C.ladanifer y Cmonspeliensis,
(Provenza, Francia). Estas especies podrían caracterizarse como frugales
71
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.16. Composición química foliar (mg/g) para distintas especies del
género Cistus en ecosistemas mediterráneos. L.C.: Límite de Confianza 95%.
ESPECIE
Calbidus
Cmonspel iensis
Csalvtfolius
C.ladanifer
Cmonspel iensis
Calbidus
Cmonspel iensis
Csalvifolius
Cincanus
Cmonspel iensis
Cparviflorus
Csalvifolius
Claurifolius
MEDIA
L.C.
(i)
(2)
(2)
(3)
(3)
(4)
(4)
(4)
(5)
(5)
(5)
(5)
(6)
N
13.0
13.0
14.5
12.1
14.5
13.6
14.9
14.6
18.8
14.1
15.1
12.4
13.7
14.2
[13.1-15.2]
P
1.3
-
2.3
1.1
1.3
1.6
2.1
1.9
1.2
1.3
0.8
1.1
1.5
1.4
ll.1-1.7]
K
7.3
-
20.6
5.5
5.5
7.5
5.5
7.4
-
-
-
-
4.9
8 .0
[3.7-12]
Ca
8.6
-
15.2
6.3
7.0
14.2
10.2
15.9
-
-
-
-
10.5
10.9
[7.8-15]
Mg
1.8
-
3.7
2.1
2.4
1.4
1.9
2.1
-
-
-
-
2.2
2.2
[1.6-2.7]
(l).Specht, R.L. (1988).Maquis francés (Marsella) (2).J3run, B., Conrad, L.& Gamisans, J.(1983).Maquis,(Córcega). (3) y (4). Specht, R.L. & Molí, E.J.( 1983).Matorral mediterráneo. Herault y Provenza (Francia) (S).Margaris, N.S.,Adamandiadou,t S.,Siafaca, L.& Diamantopoulus.J. (1984).Maquis griego. (6).Presente trabajo. Matorral sureste español. Del (1) al (5), recopilados en Specht, R. L.Ü988).
respecto al K y por tanto eficientes en el uso de éste. En este sentido, se
encontrarían más próximas a los encinares para los que se han dado valores
medios de K de 5.5 mg/gr (Ferrés 1984) y 7 mg/g (Clemente 1983). Ferrés hace
referencia precisamente a la frugalidad de los encinares respecto a este
elemento. En el otro extremo se encontraría Csalvifolius del maquis corso con
consumo de lujo y valores de 20 mg/gr. para el K foliar. La menor
72
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

tilomas», mlneralomama y producción.
variabilidad en las especies de Cistus se ha encontrado para el N (cv=ll 7.)
seguido de P y Mg (CV= 307.). Para el Ca, el alto coeficiente de variación
encontrado para estas especies puede ser en parte explicado por la diferente
naturaleza del sustrato en que viven. Sin embargo, en C. laurifolius la
acumulación de este elemento parece contradecir esta hipótesis dado que esta
especie crece en zona silíceas y por tanto pobres en Ca.
Tallos y ramas.
En las tablas 2.17 y 2.18 aparecen datos de distintos autores para
tallos y ramas de distintas especies y arbustos mediterráneos. Los intervalos
de confianza de la media calculada a partir de la bibliografía resultan
amplios debido al bajo número de datos. Además, en el caso de la madera, la
variabilidad depende también de la edad, y del tamaño de la madera que se haya
considerado para los análisis, cuestión que, a veces, no se especifica e
influye en que los datos manejados no sean del todo homogéneos.
Para A.decorticans son válidas en general, las mismas consideraciones
hechas para las hojas; el orden de elementos respecto a la fracción foliar no
se altera y los contenidos en N y Ca resultan alto y bajo respectivamente en
relación a la media de otros matorrales. Esta tendencia es más manifiesta en
tallos jóvenes que en ramas. En C. laurifolius lo más destacable son los bajos
contenidos de K en esta fracción.
Eficiencia en el uso de Nitrógeno y Potasio.
Epsten (1972), ha señalado que los contenidos de N y K en hojas pueden
ser indicativos del estado nutricional de éstas y por tanto, constituir una
medida indirecta del contenido de éstos en el suelo y de su disponibilidad.
Se ha realizado un ANOVA seguido de un test de Fischer para contrastar
los contenidos de estos dos elementos en hojas u acículas del año en las
especies estudiadas y detectar posibles diferencias significativas entre
ellas. Los resultados obtenidos indican diferencias significativas entre
especies para el N (F=158.38 p<0.001) pero no para el K (F=3.01, p=0.06). Al
73
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Moro Cuadrillero
Tabla 2.17. Concentraciones químicas (mg/g) en tallos de distintas
especies/sistemas de matorral mediterráneo.
ESPECIE
Phillyrea media.
Arbutus unedo.
Viburnum ti ñus. 2
Matorral california. Matorral Chile?
media
3
Adenocarpus decortícans. 3
Cistus laurifolius.
N
5.58
5.01
4.88
8.00
9.00
6.49
14.30
6.37
P
0.79
1.08
0.88
1.20
1.50
1.09
0.86
0.94
K
10.20
6.22
8.00
-
8.50
8.23
7.20
4.24
Ca
4.98
6.74
8.49
-
13.70
8.47
2.00
8.00
Mg
0.88
0.96
1.48
-
2.70
1.50
0.95
1.08
(D.Clemente A. (1983) (2).Shaver G.R.(1981) (3).Presente estudio.
Tabla 2.18..Composición química (mg/g) en ramas de distintos sistemas
de matorral mediterráneo.
SISTEMA
Chaparral.
Coastal sage. 2
Chaparral California. 2
Chaparral Chile. 3
Garriga. Media
4
Adenocarpus decortícans. 4
Cistus laurifolius.
N
5.40
3.70
7.00
10.00
5.4
6.30
8.60
4.90
P
0.40
0.42
1.00
1.60
0.30
0.74
0.20
0.50
K
2.30
3.60
7.80
7.00
2.70
4.68
3.60
1.20
Ca
4.80
2.40
6.50
14.80
22.20
10.14
3.20
8.90
Mg
0.53
0.58
1.40
2.60
0.30
1.08
0.70
0.50
(1) Gray J.T.(1983) (2) Shaver G.R. (1981). (3) Lossaint P. & Rapp, M.(1971) (4) Presente trabajo.
74
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mlneralomasa y producción.
aplicar posterioremente el test de Fisher no se encontraron diferencias
significativas entre las dos especies de pino respecto al contenido en N, pero
sí entre ellos y las especies arbustivas. Los valores medios son
significativamente más bajos (p<0.05) que en el matorral. (0.91 y 0.92 7. m.s).
Los pinos, por tanto, se comportan de manera más eficiente en el uso de este
nutriente. Como cabía esperar, el arbusto fijador A. decorticans con un
contenido media de 2.9 7. m.s., constituye un ejemplo de consumo de lujo para
este elemento. La media encontrada es significativamente más alta que la de
las otras t res especies. C.laurifolius ocupa una posición intermedia con un
contenido medio de 1.37 7. m.s. también significativamente diferente al resto
de las especies.
Respecto al K, sólo existen diferencias significativas entre P. nigra,
(la especie más frugal con un contenido medio de 0.279% m.s.) y A.decorticans
(la de mayores requerimientos : 0.64 7. d.w.). No existen diferencias
significativas entre los contenidos de K de P.pinaster, P.nigra y
C.laurifolius, por tanto, estas t res especies se comportan de una forma
similar respecto a este elemento.
Como resumen, puede hablarse en lineas generales de una especie con
consumo de lujo para el N (Adenocarpus decorticans) que probablemente mantenga
este status gracias a las entradas adicionales de N por fijación. Las dos
especies de pino son las más frugales respecto a ambos elementos,
característica coincidente con los datos de bibliografía para las coniferas en
general. Los bajos contenidos en nutrientes en acículas és una caractristica
que diferencia a las coniferas de otros sistemas (matorral, caducifolios etc )
(Rapp 1983, Vitusek 1988). C.laurifolius ocupa la posición intermedia. Al ser
un arbusto invasor y pionero sus requerimientos nutricionales no son altos,
siendo algo más exigente respecto al N que al K.
En sistemas mediterráneas, algunos autores han encontrado ciertos
patrones en las relaciones de contenidos foliares entre nutrientes
individuales tales como el N-P ó el N- Ca (Rundel 1988). A part ir de datos de
este autor y de las tablas 2.14, 2.15 y 2.16 se ha confeccionado la figura 2.6
relacionando los contenido de N y P. Puede observarse la estrecha agrupación
75
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
3
2,5
2
1,5
1
0,5
n
P (mg/g)
o
+ o + + «o +
+ 0e fe / x ° x C? *
£&&\ X X • 0 á X
o-xVfr * X '. . ° * I • : * * X a q-, ao
i i
X
X
1
¡o 20
N (mg/g)
Chap.mani. + Chaparral ° Chile * Francia ° Encinar A Jaral
30 40
a S.Africa
* Fijadores
Figura 2.6. Ordenación de algunos sistemas mediterráneos según la relación
entre la concentración foliar de nitrógeno y fósforo (mg/g).
3
2,5
2
1,5
1
0,5
n
P(mg/g)
* * *
-
•• J * x . f . + • XX X • • *
x 4-
1 1
+
+ +
1
4-
10
Motorr.esclerof. Pinos
20
N(mg/g)
+ Fijadores N * Encinar
30
* Jaras
40
Figura 2.7. Ordenación de sistemas arbustivos, pinares y encinares de la
región mediterránea según la relación entre la concentración foliar de N y P
(mg/g).
76
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
de los jarales con una mayor variabilidad respecto al P que respecto al N. Los
jarales quedan solapados con encinares, sistemas de garriga y encinar francés,
mientras que aquellos Cistus con un contenido superior de P están agrupados
con las especies de matorral chileno. El matorral sudafricano y el chaparral
marítimo de California se separan claramente dados su bajos contenidos en
ambos elementos. Las leguminosas fijadoras definen un gradiente amplio
respecto al nitrógeno, siendo sin embargo muy constantes con respecto al P.
Las fijadoras con menores contenidos de N y aquellos matorrales mediterráneos
con mayores niveles de N se asocian en una zona alrededor de lo 15 mg/g de N.
En la figura 2.7 se han situado también una serie de pinos mediterráneos,
incluidos los estudiados, situándose como era esperable en la parte inferior
del gradiente pero con valores bastante próximos a los encinares y solapados
con algunos matorrales escleróf ilos.
2.3.3. MINERALOMASA.
La mineralomasa aérea total en las especies estudiadas incluye 304 Kg/ha
en RPP y 331.5 kg/ha en RPN de los seis elementos analizados (N, P, Na, K, Ca,
Mg). El reparto por elementos se muestra en la tabla 2.19
Considerando cada estrato por separado, mientras que el matorral supone
un 28 y 317. de la biomasa y el pinar el 74 y 687. en RPP y RPN respectivamente,
los porcentajes relativos de mineralomasa suponen 40/507. en el matorral y
60/50 en el! pinar, lo que puede dar una idea de la importancia del estrato
arbustivo en el acumulo de nutrientes en la cuenca durante las primeras
etapas. Así, mientras que el matorral supone algo menos de un tercio de la
biomasa total, su mineralomasa supone aproximadamente entre el 40 y 507. del
total de nutrientes acumulados.
Gray y Schlesinger (1981) comparan la biomasa y mineralomasa media de un
conjunto de sistemas arbustivos mediterráneos con los correspondientes valores
medios de distintos bosques maduros encontrando que el matorral contiene un
847. menos de biomasa mientras que su mineralomasa es sólo menor de un 55 7. a
un 75 % a la obtenida para los bosques. En la cuenca de estudio estas
77
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.19. Mineralomasa to ta l (Kg/ha) y biomasa (T/ha) en cada una de las áreas de repoblación.
Área de
P.pinaster
P.pinaster
C.laurifolius
A.decorticans
Total
Área de
P.nigra
P.nigra
C.laurifolius
A.decorticans
Total
Biomasa
16.50
3.98
1.83
22.30
B i omasa
17.10
3.10
4.89
25.09
N
61
28.49
22.40
111.89
N
74.50
21.47
59.60
155.50
P
6.70
3.05
.75
10.50
P
7.60
2.29
1.98
11.87
K
32.80
8.99
7.49
49.28
K
21.40
6.68
20.03
48.11
Ca
66
37.15
5.45
108.60
Ca
49
28.62
14.65
92.27
Mg
16
3.85
1.75
21.59
Hg
13
2.86
4.67
20.53
Na
2.10
.41
.23
2.73
Na
1.20
.32
.60
2.11
Total
184.60
81.94
38.07
304.59
Total
166.70
62.24
101.53
330.39
relaciones resultan algo diferentes debido a que las diferencias en las
biomasas de cada estrato no son tan acusadas dada la juventud del estrato
arbóreo. Así, en RPP la biomasa del matorral es un 64% inferior a la del pinar
mientras que la mineralomasa acumulada sólo lo es en un 34%. Especialmente
interesante resulta la RPN en la que la biomasa arbustiva es inferior en un
55% mientras que la cantidad de nutrientes acumulados en este estrato es
prácticamente idéntica a la de P. nigra. La mayor presencia y abundancia de A.
decorticans es RPN es el motivo fundamental de este fuerte almacén arbustivo.
Solamente la leguminosa contiene el 62% de los nutrientes totales del estrato
arbustivo. La riqueza en elementos, básicamente N y K en la mayor parte de las
estructuras de esta especie (incluidas la fracción leñosa) junto a biomasas
más altas de este matorral presentes en RPN condicionan de forma decisiva la
mineralomasa aérea de esta región de la cuenca.
78
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Mineralomasa del pinar.
Biomasa, mineralomasa y producción.
La mineralomasa acumulada por el pinar para los pies con DBH > 1.5 cm es
de 184 Kg/ha en RPP y de 166.5 Kg/ha en RPN.(tabla 2.19). En la tabla 2.20 a y
b aparecen los resultados desglosados por fracciones en cada especie. Algo más
del 50 7. de la mineralomasa total se encuentra en el material fotosintético
(acículas). Esta alta contribución se debe no sólo a la mayor concentración
química de esta fracción sino a su importancia porcentual en el conjunto de la
biomasa.
El orden que ocupan los elementos en el conjunto de la mineralomasa es:
N > Ca > K > Mg > P > Na en P. nigra
Ca > N > K > Mg > P > Na en P. pinaster
La cantidad total de nitrógeno acumulada en el pino laricio es mayor que
la del res to de los elementos (74 Kg/ha) por encima incluso del Ca (49 Kg/ha)
(tabla 2.19 y 2.20b) que suele ser el elemento mayoritario en bosques o
plantaciones de edades y/o biomasa superiores a la nuestra, ya que en éstas
últimas la fracción más rica en Ca (madera) tiene una importancia cuantitativa
mayor en el conjunto de la biomasa. En el pino resinero la importancia de N y
Ca es similar. Sólo el nitrógeno acumulado en acículas supone un 20/28.5 7. del
total de la mineralomasa mientras que el Ca acumulado en ramas y tronco sólo
alcanza el 19/22 7.. El orden que ocupan el resto de los elementos coincide con
la mayoría de las serie bibliográficas. Las mayores cantidades de N, P
(alrededor del 60 7. ) y de Mg (alrededor del 507.) se acumulan en las acículas.
Por su par te , las fracciones leñosas acumulan el 63/66 7. del Ca, 50/58 7. del K
y 62/66 7. de Na. Se observa en todos los casos la escasa contribución que la
fracción tronco aporta al almacén de nutrientes. En el pinar de estudio, como
ya se ha comentado a lo largo de este capítulo, se combina la circustancia
part icular del escaso desarrollo y juventud del pinar, lo que implica baja
biomasa de troncos unido a las bajas concentraciones químicas en la mayaría de
elementos que aparecen en la fracción leñosa de las coniferas (Ferrés 1984)
Los resultados anteriores indican que respecto al acumulo total de
nutrientes en la biomasa aérea de los pinos no existen apenas diferencias
79
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.20a. Mineralomasa to ta l (Kg/ha) y biomasa (T/ha) por fracciones en la región de P.pinaster
--- T/ha Kg/ha
P.pinaster Biomasa N P K Ca Mg Na Total
37.13 3.95 13.85 24.18 8.70
13.14 1.50 7.95 26.51 3.43
10.70 1.25 11.02 14.96 3.85
Acfculas
Ramas
Tronco
4.75
4.18
7.20
79
38
93
' 88.60
52.91
42.71
Total
C.laurifolius
16.50
Biomasa
60.90
N
6.70
P
32.80
K
65.60
Ca
15.90
Mg
2.11
Na
184.22
Tota
Hojas
Tallos
Ramas
.99
.17
2.83
13.53 1.48 4.74 10.42 2.15
1.10 .16 .73 1.55 .19
13.86 1.41 3.52 25.17 1.51
.12
.02
.27
32.44
3.75
45.73
Total
A.decorticans
3.98
B i omasa
28.49
N
3.05
P
8.99
K
37.15
Ca
3.85
Mg
.41
Na
81.95
Tota
Hojas .24 7.20 .28 1.54 1.06
Tallos .06 .86 .05 .43 .12
Ramas, 1.52 14.30 .42 5.53 4.30
ramillas y tronco
.51
.05
1.19
.03
.02
.16
10.62
1.54
25.89
Total 1.83 22.40 .75 7.49 5.45 1.75 .23 38.05
entre ambas áreas. Solamente cabría considerar ciertas diferencias respecto a
algunos nutrientes en concreto. Así, la cantidad de Ca es un 257. más bajo en
P. nigra explicable más por las menores concentraciones de Ca en la mayoría de
sus fracciones en relación a P. pinaster que por diferencias en sus biomasas
respectivas. También existe diferencias, aunque menos acusadas para el N y K.
80
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
Tabla 2.20b. Mineralomasa total (Kg/ha) y biomas (T/ha) por fracciones en la región de P.nigra.
--- T/ha Kg/ha
P.nigra Biomasa H P K Ca Mg Na Total
Acículas 5.75 47.60 4.81 10.60 16.55 6.40 .40 86.36
Ramas 4.95 16.17 2.20 6.32 17.34 4.05 .31 46.39
Tronco 6.30 10.70 .63 4.51 15 2.44 .48 33.76
Total
C.laurifolius
17.10
Biomasa
74.50
N
7.65
P
21.40
K
48.90
Ca
12.80
Mg
1.19
Na
166.51
Total
Hojas .70 9.64 1.06 3.37 7.42 1.53 .09 23.11
Tallos .12 .75 .11 .50 1.06 .13 .02 2.54
Ramas 2.26 11.09 1.13 2.81 20.14 1.21 .21 36.58
Total
A.decorticans
3.10
B i omasa
21.47
N
2.29
P
6.69
K
28.62
Ca
2.87
Mg
.32
Na
62.26
Total
Hojas .62 18.70 .74 4 2.75 1.31 .10 27.59
Tallos .16 2.22 .13 1.12 .31 .14 .06 3.97
Ramas, 4.11 38.58 1.12 14.89 11.63 3.22 .43 69.87
ramilias y tronco
Total 4.89 59.56 1.98 20.03 14.65 4.67 .60 101.49
Las relaciones porcentuales de las distintas fracciones tanto en biomasa
como en mineralomasa se han representado en la figura 2.8. El cociente entre
la cantidad total de nutrientes y la biomasa es de (O.Oll en P. pinaster y
0.009 en P. nigra). En pinares mediterráneos más maduros que el nuestro, Lledó
(1982) obtuvo un valor de 0.0112 para P. halepensis similar al obtenido por
Rapp y Cabanettes (1980) en P. pinea (0.010). Lledó (1982) comenta que los
81
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
cocientes obtenidos en estos dos casos son bastante superiores al 0.0080
encontrado por Rodin y Bazilevich (1967) y al 0.0048 dado por Ovington (1959)
para bosques templados de coniferas y la explicación la basa en la hipótesis
de Osmond (1979) según la cual la mayor cantidad de nutrientes acumulada en
especies propias de zonas más áridas tendría la finalidad del aumento de la
presión osmótica en el interior de la planta para aumentar la captación de
agua. En nuestro caso, a pesar de que el valor es muy similar a la de los
pinares mediterráneos antes citados, los valores altos encontrados quedan
suficientemente explicados por los relativamente elevados porcentajes de
biomasa de las fracciones más ricas en nutrientes ya que las concentraciones
químicas de las dos especies de pino son muy similares a los valores medios
dados para bosques de coniferas.
Minelaromasa del matorral {A. decorticans,C. laurifolius).
La biomasa aérea de las dos especies arbustivas mayoritarias (7.26 T/ha)
en el conjunto de la cuenca contiene en total 145 Kg /ha de elementos
repartidos en 68.56 Kg /ha en C. Laurifolius y 80 Kg/ha en A. decorticans
No obstante, estas cantidades difieren sustancialmente al considerar las
dos grandes regiones, debido a la dominancia de una u otra especie en cada una
de ellas (tabla 2.19). En RPP, la contribución de A. decorticans tanto a la
biomasa arbustiva como a los nutrientes acumulados en ella es del 31%. Sin
embargo a pesar de su escasa contribución a la biomasa en esta región,
almacena similares cantidades de N Y K que la j a ra (figura 2.8) debido a sus
concentraciones significativamente más altas en hojas y ramas. En RPN la
predominancia del gaznacho es manifiesta para casi todos los elementos a
excepción del Ca y P elementos para los que la j a ra posee concentraciones más
elevadas sobre todo en lo que hace referencia a la fracción foliar (P) y ramas
(Ca). En términos relativos la fracción foliar del gaznacho contiene
aproximadamente el 27% de todos los nutrientes acumulados en esta especie,
mientras que el leño acumula alrededor del 68%. En la j a ra la importancia
porcentual de la fracción foliar es ligeramente superior: alrededor de un 38%,
el leño almacena el 55/58% y los tallos el 4%. el reparto de estas cantidades
por elementos difiere substancialmente para el N, P, K y Ca entre las dos
82
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mineralomasa y producción.
Área PP
AD 1,825
BIOMASA (T/ha)
PP 184
/ 6 0 %
A D X ^ - - L 38,5
:'27Í
81,95
MINERALOMASA (Kg/ha)
Área PN
AD 4,89
BIOMASA (T/ha) MINERALOMASA (Kg/ha)
Figura 2.8. Distribución de la biomasa y mineralomasa total en las especies
estudiadas en las dos zonas de la cuenca. Valores de biomasa en T/ha. Valores
de mineralomasa en Kg/ha.
83
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
especies. A. decorticans contiene aproximadamente el doble de N y K. que C.
laurifolius, explicable por las mayores concentraciones químicas que se dan
para estos elementos (sobre todo en lo que se refiere al N) en el gaznacho. La
jara, por el contrario, contiene 1.5 veces más P y 2.5 veces más Ca que
Adenocarpus, debido también fundamentalmente a las diferencias en
concentración química que se dan en estos elementos. En términos relativos, la
fracción leñosa del gaznacho (ramas, ramillas y tronco) contiene el 68 7. del
total de elementos frente al 27% de hojas. En la j a ra la importancia de las
hojas es algo mayor (38%) frente al 57.67. del leño. Los tallos del año en
ambas especies suponen porcentajes similares del total (4.2 % en jara , 3.9 %
en gaznacho).
Un aspecto remarcable en A. decorticans es el hecho de que las hojas que
sólo suponen un 13% de la biomasa aérea total contribuyan con porcentajes de
incluso más del doble para algunos nutrientes (31 % de N, 37 7. de P, y 28 % de
Mg). La cantidad de P contenido en los tallos supone un 6 % del P total,
cantidad importante teniendo en cuenta que esta fracción sólo supone el 3 % de
la biomasa. En C. laurifolius, los porcentajes de N, P, K y Mg en hojas son
alrededor del doble de los correspondientes a la biomasas de esta fracción El
porcentaje de K en tallos también duplica al correspondiente a la biomasa de
esta fracción respecto al total. La mayor importancia porcentual del Ca
(67/707.) se alcanza lógicamente en la fracción madera.
La relación contenido total en nutrientes/biomasa es de 0.020 en ambas
especies. A partir de datos de diferentes sistemas arbustivos con edades
comprendidas entre los 12 y 37 años recopilados por Gray y Schlensinger (1981)
se. ha calculado el cociente mineralomasa media/biomasa media hallando un valor
de 0.0175, que resulta ser muy similar al calculado en el presente trabajo. La
relativamente alta biomasa de hojas en ecosistemas arbustivos influencia el
almacén de nutrientes en este tipo de comunidades como resultado de las
mayores concentraciones químicas en las hojas (Gray y Schlesinger 1981) y por
tanto la relación masa nutrientes/biomasa se ve aumentada. En la citada
recopilación de Gray y Schlesinger se dan valores de porcentaje de hojas
respecto a biomasa total del 28 % para especies perennes del matorral chileno
de Fundo Sta Laura. Núñez (1989) encuentra un 30% en jarales de Extremadura.
84
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mlneralomasa y producción.
Ambos resultados resultan similares a los encontrados para la ja ra en el
presente estudio pero superiores a los de A. decorticans.
2.3.4. PRODUCCIÓN.
PINAR.
Incrementos radiales.-
La relación incremento de DBH (mm/año) frente a DBH (cm) se representa en
las figura 2.9 para las dos especies. El intervalo de diámetros manejado
oscila entre los 3.4-15.5 cm en P.pinaster y los 2-10.7 cm en P.nigra. Hay que
hacer notar que en aquellos COREX de P. pinaster con DBH inferior a los dos
cm, resultaba muy difícil distinguir los anillos, por tanto las muestras para
estas clases de tamaño no se han incluido en la regresión. La correlación
resulta positiva y altamente significativa para ambas especies (P. pinaster
r=0.81, p<0.001; P. nigra: r= 0.92, p<0.001). Si los datos son transformados a
logaritmos se obtienen una mejoría en el coeficiente de correlación aunque no
muy acusada. Las ecuaciones obtenidas han sido las siguientes:
P.pinaster; LN (mm/año) = .421 + 0.91 LN(DBH) r=0.86; p< 0.001
P. nigra; LN(mm/año)= -1.34 + 1.09 LN(dbh) r=0.96; p< 0.001
En ambas especies el coeficiente de alometría no es significativamente
distinto de 1, indicando una relación isométrica entre ambas variables o lo
que es lo mismo un incremento proporcional, de una con respecto a la otra para
el intervalo de tamaños manejados. La pequeña diferencia en los coeficientes
de correlación o regresión entre el ajuste linear y el potencial queda
suficientemente explicado por el valor de este coeficiente de alometría. La
relación positiva y significativa entre ambas variables (mayores crecimientos
al aumentar el tamaño del árbol), encontrada en este estudio parece apoyar la
idea de este tipo de comportamiento en poblaciones arbóreas con individuos de
tamaños pequeños (Gracia, com pers.). Al aumentar el tamaño medio del rodal,
la correlación desaparece y es muy común que en pinares más maduros que el
nuestro, no se encuentre ningún tipo de relación entre incremento radial y
DBH. Así, en un conjunto de rodales de P. halepensis y P. nigra en la comarca
85
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Incrementos Radiales
Inc. radial (mm/afio)
0 2 4 6 8 10 12 14 16
DBH (cm) P. plnaatar
INC. RADIAL (mm/arto).
O 2
P. nlgra
4 6 8
DBH (cm) 10 12
Figura 2.9. Relación entre DBH (cm) del árbol y el incremento radial (mm/año)
en los cores tomados en individuos de P.pinaster (gráfica superior) y P.nigra
(gráfica inferior).
86
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mlneralomasa y producción.
del Pía de l'Estany con DBH medios de 15 y 17 no se encontró relación
significativa entre el incremento radial de 5 años frente al DBH. (Gracia en
prensa). Cuando el pinar tiene un elevado grado de madurez, la relación pasa a
ser del tipo exponencial negativa (Gracia, com.pers.).
Otra característica a destacar a partir de los incrementos radiales,
expresados de forma acumulativa (incremento 5 años) o anual son sus altos
valores absolutos superiores a muchos pinares bien más maduros o bien de
similar edad o madurez pero que crecen en zonas con mayores condiciones de
aridez (Pastor, común, personal). Una vez más, se ha de tener en cuenta la
estructura y edad actual del bosque, que permite suponer que el rodal se
encuentra en una fase inicial de acumulación rápida de biomasa. Los rápidos
incrementos iniciales de biomasa en plantaciones jóvenes se ven frecuentemente
limitadas en zonas mediterráneas debido a el déficit hídrico que puede
prolongarse durante varios meses unido a la existencia de sustratos pobres y
muchas veces alterados. Las escasez de agua en la cuenca de estudio no parece
ser tan limitante dada la altitud, existencia de nieblas durante buena parte
del año y precipitaciones durante los últimos años superiores a los 500 mm.
Producción pr imar ia neta .
En la tabla 2.21 se expresa, para cada fracción del pinar, la producción
anual (T/ha.año) en las dos grandes regiones. La producción tanto total como
parcial resulta muy similar en ambas zonas. Existen pequeñas diferencias que
resultan lógicas teniendo en cuenta la distribución de tamaños en cada
especie. Por ejemplo la producción de madera en troncos es superior en P.
pinaster ya que el incremento de esta fracción es proporcional al tamaño del
árbol para las dos especies y en P.pinaster el tamaño medio del rodal es
superior al laricio. Las aproximadamente 3 T/ha. año en las dos especies se
encuentran dentro del rango de valores de 1.9-4.9 T/ha.año en P. halepensis o
2-5 T/ha.año en P. nigra. encontrados en determinadas parcelas de la comarca
del Pía de L'Estany (Gracia en prensaHtabla 2.22). Estos valores corresponden
a rodales con estructura ligeramente más madura que el pinar de estudio con
individuos comprendidos entre 5-20 ó 5-25 cm de DBH. Sin embargo, a pesar de
unas producciones netas totales del mismo orden entre el pinar estudiado y los
rodales del Pía de l'Estany, la compartimentalización de esta producción y sus
87
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.21. Producción neta total (T/ha.ano) y por especies en cada región.
Región de P.pinaster _^_^_
Especie Foliar Tallos Ramas Reproduc. Tronco(ma.) Tronco(cotz) Total
.82 .23 3.14
.55
1.02
.82 .23 4.41
P.pinaster
A.decorticans
C.laurifolius
Total
1.50
.25
.56
2.31
.30
.06
.17
.53
.25
.12
.22
.58
.04
.12
.07
.24
Región de P.nigra
Especie Foliar Tallos Ramas Reproduc. Tronco(ma.) Tronco(cotz) Total
.56 .18 3.30
1.40
.64
.56 .18 5.34
P.nigra
A.decorticans
C.laurifolius
Total
1.70
.62
.31
2.60
.50
.15
.12
.77
.31
.32
.17
.80
.08
.31
.04
.43
relaciones con la biomasa, indican un funcionalismo claramente distinto entre
las estaciones catalanas y la del presente estudio. Es conocido el aumento del
cociente biomasa/producción con la edad o madurez del bosque (Waring y
Schlesinger 1985). Este índice representa una tasa de acumulación de biomasa y
oscila entre 0.1 y 0.2 en los rodales catalanes en contraposición al pinar de
Almería en que los cocientes son de 2 a 4 veces inferiores reflejando de forma
ostensible la situación de inmadurez del sistema estudiado.
Sin duda, la característica diferencial más importante de los pinos
estudiados es la elevada producción de hojas (1.5 T/ha.año en P. pinaster y
1.7 en P.nigra). Aun suponiendo que ésta pueda estar ligeramente sobrestimada
debido a la fuerte intensidad del brote del año 1986 los valores siguen siendo
88
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Biomasa, mlnera lomasa y producción.
< o
-0
N I
00
N I
N I
u->
> í
v »
C0
"-
CO
o
NI - * *-
NI
O
00
o
o <\ j
5
o. o. -o
N I <- , -
< o
s CJ
oo
•o
NI
r- oo
oó 00 oo tn «- o
X E u 3: w
a
oo
-o' ° -o 00 «—
<0
u
s
o
-g
3
"8
O
5 u
I
N>
9 5
uu Q.
¡2 ¡C *~ <£ h- 3
c
a. c
Si 4>
C
41
O
<o
ÍM >o o
10
c o.
c a a.
c a.
89
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
muy elevados en relación con los otros pinares de la tabla 2.22. En los
primeros estadios de establecimiento de bosques o plantaciones, un
considerable porcentaje de producción va destinado al material fotosintético
(Waring y Schlesinger 1985) para permitir el crecimiento de las copas y, a su
vez compensar la pérdidas foliares por desfronde. En esta fase la producción
foliar aumenta hasta llegar a un máximo que suele coincidir con el cierre de
la bóveda. En el pinar de Almería nos encontramos probablemente en esta
situación, aunque tres años después (89/90) los contactos de las copas
observados podrían estar indicando ya un cierre de la bóveda por lo que los
valores obtenidos en el 86 estén posiblemente muy cerca de los máximos. La
fracción de producción foliar destinada al incremento de biomasa ha sido
calculada en 500 Kg/ha.año en P. pinaster y 770 Kg/ha.año en P. nigra. El
esfuerzo destinado a la producción foliar es muy elevado (48-52%) en relación
con los otros bosques de la tabla 2.22. LLedó (1982) también encontró
elevados porcentajes de producción foliar (34-54 %) en individuos aislados de
P. halepensis con tamaños comprendidos entre 1.1 y 15 cm DBH. Estos
porcentajes de producción foliar se encuentran más próximos a sistemas
arbustivos que a poblaciones arbóreas.Teniendo en cuenta la distribución de
clases diamétricas en la especies estudiadas con elevadas densidades de pies
menores de 5 cm. de DBH, se puede argumentar que la repoblación estudiada se
encuentra en una fase de transición arbustiva >arbóreo lo que implica
necesariamente características funcionales y dinámicas diferentes a las
esperables para un estrato arbóreo definido de las cuales el cociente PH/PT o
de forma más general la eficiencia en la producción (PT/BT) constituyen claros
ejemplos. No obstante en plantaciones de P. ponderosa que han alcanzado ya una
estabilización en la producción primaria neta (Gholz 1982) se ha encontrado
una relación PF/PT similar a nuestros resultados (507.). Satoo (1982) cita
ejemplos similares en plantaciones de P. densiflora de 15 años en Japón .
La productividad foliar expresada como cociente entre producción y
biomasa total de hojas es igualmente elevada en las especies estudiadas en
contraposición con los rodales de la tabla 2.22. Esto implica una tasa de
renovación foliar mucho más rápida en P. pinaster y P. nigra por lo menos en
estas primeras etapas de crecimiento.
90
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

MATORRAL.
Blomaaa, mlneralomasa y producción.
La PPN anual de cada especie y fracción arbustiva para las dos grandes
regiones se presenta en la tabla 2.21.
La producción total de las dos especies arbustivas es de 1.57 T/ha.año en
RPP y 2 T/ha.año en RPN. La dominancia de una u otra especie en las dos áreas
se manifiesta en sus porcentajes relativos de producción. En RPP la ja ra
produce el 64% del material arbustivo mientras que en RPN el gaznacho produce
el 707.. El material leñoso supone el 21-22% de la producción en el jara l y del
21-277. en A. decorticans. El esfuerzo reproductivo es considerable en ambas
especies sobre todo, en lo que se refiere al gaznacho en el que un 227. de la
producción total va destinado a la formación de flores, frutos y semillas. La
producción foliar está cercana al 50% en ambas especies y regiones.
Los sistemas arbustivos mediterráneos maduros de California suelen
producir entre 2.5-6.7 T/ha (Gray y Schlesinger 1981, Money (1981). Las
garrigas alrededor de 3 T/ha.año (Lossaint y Rapp 1971) y los encinares
5.4-9.3 T/ha.año (Lledó 1990). Por su parte Merino eí aZ.(1990) obtienen
producciones de 1.5 T/ha.año en jarales de Doñana y de 6.5 en brezales. En los
países catalanes se dan valores de 1-4 T/ha.año en sistemas arbustivos densos
y bien desarrollados (Terradas eí al. 1989), mientras que a part ir del trabajo
de Márquez eí al. (1989) puede calcularse una producción aérea elevada (6
T/ha.año) en jarales extremeños entre 10 y 20 años.
¡
La producción foliar en muchas formaciones arbustivas de la zona
mediterránea suele alcanzar el 507. de la producción total (Terradas et
al. 1989) porcentajes muy de acuerdo con los encontrados en el presente
trabajo. En el matorral fijador de nitrógeno Myrica gale el porcentaje es
también similar: (477.) (Schwintzer 1983) así como en jarales de Extremadura
(46%) (Márquez et al. 1989). La relación producción /biomasa es más elevada en
A. decorticans (28-307.) en relación a C. laurifolius (20-25%). Valores
próximos al 20% se han encontrado en chaparrales de California (Mooney 1981) y
ligeramente inferiores (14%) en la garriga francesa (Lossaint y Rapp 1971,
91
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.23. Nutrientes asociados a la PPM (Kg/ha.año) en el área de P. pinaster.
P.pinaster
N P K Ca Mg Na
Total
ACICULAS
13.95
1.58
6.81
3.43
2.52
.35
28.64
TALLOS
2.63
.96 1.31
1.36
.48
.05
6.79
RAMAS
.57
.06
.65
.83
.18
.05
2.33
TRONCO(M)
.96 ,-16
1.19
1.23
.41
.12
4.06
TRONCOCC)
.56
.03
.41
.88
.15
.02
2.05
REPRO
.30
.02
.04
.12
.05
.01
.53
TOTAL
18.97
2.80
10.40
7.85
3.79
.59
44.40
A. decorticariB
N P IC
Ca
Mg
Na
7.25
.30 1.60
1.10
.52
.04
.80
.05
.43
.12
.06
.02
1.11
.05
.43
.35
.08
.02
39 09 76 63 17 .02
11.55
.49 3.22
2.20
.83
.10
Total 10.81 1.48 2.03 4.06 18.39
C.laurifoliuB
N P K Ca Mg Na
7.50
.84 2.74
5.88
1.21
.07
1.40
.21
.95 1.79
.24
.02
1.06
.11
.27 1.93
.12
.02
.56
.06
.36
.12
.08
.01
10.52
1.22
4.32
9.72
1.65
.11
Total 18.24 4.62 3.51 1.20 27.56
TOTAL 57.69 12.89 7 .87 4.06 2.05 5.79 90.35
Mooney 1981). En Myrica gale, (Schwintzer 1983), la eficiencia es del 65%, del
orden del doble de la del gaznacho. En suelos pobres como las dunas del de
Doñana, Merino et al. (1990) obtienen eficiencias alrededor del 30 7. en jara l
y brezal y a part ir de Márquez et al. (1989) se obtiene un valor del 17% en
jarales ya maduros (20 años) con tasas de acumulación de biomasa cercanos a 6
frente a los 3-4 encontrados en el presente trabajo.
92
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mineralomasa y producción.
Tabla 2.24. Nutrientes asociados a la PPN (Kg/ha.año) en el área de P. m'gra (RPN).
P.nigra
ACIOJLAS TALLOS RAMAS TRONCO(M) TRONCO(C) REPRO TOTAL
.49
.02
.09
.70
.12
.01
.0580
.0030
.0078
.0180
.0130
.0020
20.06
2.03
5.71
6.91
3.03
.41
N 15.58 2.42 .77 .74 P 1.53 .35 .08 .05 K 3.65 1.04 .48 .45 Ca 2.87 1.43 .86 1.03 Mg 1.92 .60 . 21 . 1 7 Na . 28 .05 .03 .04
total 25.83 5.89 2.42 2.48 1.43 .10 38.15
A. decorticans
M
P
K
Ca
Mg
Na
total 26.73 3.80 5.51 0 0 10.11 46.15
17.90
.74 3.96
2.74
1.30
.09
2.10
.12 1.08
.30
.14
.06
3.01
.13 1.16
.95
.22
.04
C.laurifoliuB
N P K Ca Mg Na
4.24
.46 1.51
3.25
.67
.03
.74
.11
.50
.94
.13
.01
.85
.09
.22 1.55
.09
.02
5.940
.230
1.900
1.560
.430
.046
28.95
1.22
8.10
5.55
2.09
.24
.3030
.0350
.1970
.0650
.0460
.0037
6.13
.69 2.42
5.81
.94
.06
Total 10.16 2.43 2.81 0 0 .65 16.05
TOTAL 62.72 12.12 10.74 2.48 1.43 10.86 100.35
Si se compara la producción conjunta de las t res especies presentes en
cada una de las zonas de repoblación (RPP Y RPN) (Tabla 2.21), se pone
de manifiesto la importancia cuantitativa del matorral respecto a la PPN al
igual que ocurría para la biomasa. No obstante su efecto, es muy similar en
ambas áreas. En RPP el porcentaje invertido en PPN arbustiva es el 337. del
total y en RPN del 387.. En la primera zona, solamente la j a r a produce 1.02
T/ha.año, lo que supone un 21% de la producción total. Por el contrario en RPN
93
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
es el gaznacho el que produce el 267. de la producción aérea.
La contribución destacable de este estrato en los compartimentos y flujos
de materia orgánica en relación al pinar puede intensificarse si tenemos en
cuenta los nutrientes asociados a la PPN, dado la mayor riqueza elemental de
las especies arbustivas.
2.3.5. NUTRIENTES ASOCIADOS A LA PRODUCCIÓN PRIMARIA NETA.
A partir de los valores de producción y de las concentraciones químicas
en biomasa se han calculado los nutrientes incorporados anualmente por la
vegetación de cada región (Kg/ha.año) para la formación de las distintas
estructuras (tablas 2.23 y 2.24).
En ambas regiones la cantidad de nutrientes totales movilizados en la
producción son de un orden similar (90 Kg/ha.año en RPP y 100 Kg/ha.año en
RPN). Para todas las especies es el N el elemento mayoritario seguido del Ca y
K. La fracción hoja/acícula constituye la fracción con mayor demanda
nutricional. Los porcentajes de nutrientes destinados a la producción foliar
son del mismo orden en las cuatro especies oscilando entre el 63-67% en las
especies de pino, 58% en A. decorticans y 63-66 % en C. laurifolius. Interesa
resaltar la notable movilización de nutrientes para la formación de
estructuras reproductivas en el gaznacho (22%) en contraste con el jaral (4%).
En RPN, el pinar y el arbusto mayoritario (A. decorticans) utilizan para
la producción cantidades similares de nutrientes, 38.2 Kg/ha.año y 46
Kg/ha.año respectivamente (tabla 2.24). La dominancia en RPN del gaznacho,
especie productora de estructuras muy ricas en nutrientes y con un elevado
esfuerzo reproductivo es básicamente la responsable de las diferencias
existentes respecto a la contribución del matorral en ambas zonas (62% en RPN
y 50% en RPP). Si bien estas diferencias porcentuales no parecen demasiado
acusadas, vienen determinadas fundamentalmente por el gaznacho (tabla 2.23 y
2.24) puesto que la cantidad de nutrientes utilizados por el pinar o el jara l
en ambas regiones de la cuenca son mucho mas similares a las de A.
decorticans.
94
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Blomasa, mineralomasa y producción.
Tabla 2.25. Incremento neto anual de la mineralomasa en el material perenne (Kg/ha.año) calculado a p a r t i r de la d i ferencia entre nutr ientes asociados a la producción de estructuras perennes y sus pérdidas por desfronde.
(kg/ha año)
P.pinaster
N P K Ca
Mg Na
Total
A. decorticans
N P K Ca Mg Na
Total
C. laun
N P < Ca Mg Na
Total
folius
PRO
4.72
1.20
3.56
4.30
1.22
.24
15.23
PRO
1.91
.10
.86
.47
.14
.04
3.52
PRO
5.64
.79 3.32
7.69
.91
.11
18.45
RPf»
DF
.065
.005
.021
.212
.020
.002
.325
DF
.762
.030
.041
.063
.043
.012
.951
DF
.146
.009
.014
.337
.017
.003
.526
NETO
4.650
1.196
3.534
4.090
1.202
.233
14.905
NETO
1.148
.068
.819
.407
.096
.026
2.56
NETO
5.494
.781
3.306
7.353
.888
.103
17.925
(Kg/ha.año)
P.nigra
N P < Ca Mg Na
total
A. decorticans
N P K Ca Mg Na
total
C.laurifolius
N P K Ca Mg Na
Total
PRO
4.42
.50 2.06
4.02
1.10
.13
12.22
PRO
5.11
.25 2.24
1.25
.36
.10
9.31
PRO
1.59
.20
.72 2.49
.22
.03
5.24
RPM
DF
.058
.003
.007
.014
.009
.001
.093
DF
1.907
.074
.103
.157
.107
.031
2.380
DF
.079
.005
.008
.183
.009
.001
.285
NETO
4.36
.50 2.05
4.01
1.09
.13
12.13
NETO
3.20 .18
2.14
1.09
.25
.07
6.93
NETO
1.51 .19 .71
2.31
.21
.03
4.96
Respecto al incremento neto de la mineralomasa que podríamos denominar
también retención (Colé y Rapp 1981), puede ser equiparable a los nutrientes
incorporados anualmente para producir estructuras perennes que quedan
inmovilizados a largo plazo en la biomasa. Estrictamente, el cálculo de este
parámetro debería tener en cuenta la fracción perenne que retorna al suelo
anualmente por desfronde. No obstante, en este estudio estas cantidades son
prácticamente despreciables dado que la caída de ramas tanto finas como
gruesas es muy escasa (tabla 2.25). Únicamente en A. decorticans existe una
95
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 2.26. Aumento de la mineralomasa foliar en las dos especies de pino.
Valores en Kg/ha-año.
Especie Peso N P K Ca Mg Na
P.pinaster 500 4.6 0.52 2.26 1.15 0.80 0.08
P.nigra 700 6.4 0.63 1.50 1.20 0.77 0.11
ligera disyunción entre los nutrientes asociados a la producción perenne y el
incremento neto de mineralomasa puesto que la caída de ramas (básicamente
ramillas de muy pequeño calibre) tiene cierta importancia en el desfronde.
Los nutrientes retenidos son mayoritariamente N y Ca, con una importancia
incluso mayor en algunas especies del primer elemento. Normalmente en bosques
son mayores las cantidades de Ca retenidas respecto al N dadas sus superiores
concentraciones químicas en la fracción leñosa. En este estudio, la tasa de
producción de estructuras leñosas es elevada como puede deducirse de los
incrementos radiales, pero su valor absoluto es pequeño dado que también la
biomasa leñosa de partida es pequeña en relación a plantaciones o bosques más
maduros. Sin embargo la producción anual de tallos, fracción más rica en N que
en Ca es porcentualmente más importante en sistemas jóvenes como el estudiado
con respecto al leño total producido. Esta puede ser la principal
justificación al primer lugar ocupado por el N en el incremento neto de
mineralomasa en el pinar estudiado.
En el pinar ha de añadirse a estos incrementos perennes el de la
mineralomasa asociada al incremento de biomasa foliar dado que el pinar no ha
alcanzado aun el estado de equilibrio. A part ir del peso incrementado
anualmente por la fracción foliar multiplicado por la concentración de
nutrientes en acículas del año, se han obtenido los valores de la tabla
2.26.
96
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 3. Desfronde.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

I
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

3.1. INTRODUCCIÓN.
Desfronde
El desfronde está constituido por la materia orgánica (de origen vegetal
fundamentalmente) que cae al suelo de manera continua o estacional desde los
distintos estratos vegetales (Verdú 1984). Constituye el flujo principal de
tranferencia de energía y nutrientes al suelo sobre todo de N, P, Mg y Ca. Un
porcentaje importante de la biomasa acumulada y del carbono fijado anualmente
en la vegetación retorna al suelo por esta vía sobre todo en los sistemas
jóvenes. Bray y Gorham (1964) y Olson (1963) consideran al desfronde como un
indicador adecuado de la productividad de los ecosistemas.
La importancia de este flujo para la productividad y el ciclo de
nutrientes de un ecosistema ha sido reconocida por muchos autores y ha quedado
reflejada en una extensa literatura. Ejemplos de trabajos de síntesis con
particular referencia a los ecosistemas forestales son los de Bray y Gorham
(1964), Colé y Rapp (1981), Waring y Schlesinger (1985) y Rodin y Bazilevich
(1967).
Son muchos los factores que pueden influir sobre el desfronde. Bray y
Gorham (1964) los clasifican en cuatro grandes grupos: factores climáticos,
factores temporales, perturbaciones o tratamientos y tipo de bosque. En este
sentido, son muchos los trabajos que intentan relacionar la variabilidad del
desfronde (espacial o temporal) con diferentes parámetros. La mayoría de
autores consultados comparan el desfronde entre distintos tipos de
bosque/matorral ó distintas especies vegetales. Hay un menor número de
trabajos que aborden la relación con otro tipo de factores. Entre ellos
citaremos la edad del sistema: (Gholz y Fisher 1985, Bruijnzeel 1985, Márquez
et al. 1989, Sharma y Embash 1987); área basal: (Naik y Shrivastava 1985,
Turnbull y Madden 1983); fertilización (Baker et al. 1986); diversos factores
climáticos: (Turnbull y Madden 1983); precipitación: (Merino et al. 1990);
altitud: (Reiners y Lang 1987, Gosz et al. 1972).
Los ecosistemas mediterráneos están dominados por especies perennifolias.
En ellas el desfronde se caracteriza por ser un flujo continuo a lo largo del
año pero que presenta una estacionalidad muy marcada. Esta se manifiesta por
97
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
la existencia de unos pulsos de máxima caída que suelen producirse bien
inmediatamente antes de la sequía estival: encinares, garriga, algunos jarales
(Escudero et al. 1985, Garrido et al. 1989, Lossaint y Rapp 1971, Núñez 1989)
o bien durante ella (pinares fundamentalmente, Rapp 1967, Garrido et al. 1989,
Dannaoui 1981). En esta época del año, las precipitaciones son mínimas y por
tanto la materia orgánica y los nutrientes asociados que retornan al suelo son
muy poco suceptibles al lixiviado. A su vez, inmediatamente antes de la máxima
abcisión, suelen ser frecuentes en estas especies los mecanismos de
retraslocación de nutrientes (N y P principalmente). Todas estas
características conducen evidentemente a una minimización de la pérdida de
nutrientes, bien por reducción del lixiviado, bien por una reutilización
determinando así la composición final de la hojarasca que servirá de sustrato
a los organismos descomponedores. Es evidente pues, que la fenología de la
abcisión y los mecanismos asociados a ella tienen consecuencias importantes
para la dinámica de la materia orgánica y los nutrientes en la descomposición,
siguiente eslabón del ciclo (Garrido et al. 1989).
El desfronde es el flujo principal de circulación de algunos nutrientes
como N, Ca, P y Mg en la mayor parte de los ecosistemas (Gray y Schlesinger
1981). La magnitud del retorno es función del peso de desfronde y de su
composición elemental. La relación entre pérdida por desfronde y la cantidad
acumulada en la biomasa es para la mayoría de los nutrientes de un 10-157. en
ecosistemas en desarrollo tanto en matorrales mediterráneos (Gray y
Schlesinger 1981) como en bosques templados (Likens et al en Gray y
Schlesinger 1981). Sin embargo estas proporciones pueden variar
sustancialmente en sistemas muy jóvenes y en aquellos dominados por plantas
con una alta productividad.
En este capítulo abordaremos el retorno total de materia orgánica al
suelo por parte de las cuatro especies así como la contribución de cada una de
las fracciones. Se estudiará la variabilidad interanual y sobre todo
estacional de cada una de ellas. Dedicaremos especial atención al desfronde
foliar dada su mayoritaria contribución a la caída total. La relación de la
estacionalidad del desfronde con factores- climáticos se contempla en el punto
3.3.4. El siguiente apartado viene dedicado a la composición química de la
98
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

De»fronde
hojarasca. Se establecen ciertas diferencias interespecíficas en la
composición elemental de las distintas fracciones. Posteriormente se t ratan
los aspectos de la variabilidad química temporal de la hojarasca, los
mecanismos que pueden estar implicados en ella (retraslocación, lixiviado) y
su relación con la fenología de la abcisión. Una última parte se dedica al
retorno de nutrientes por esta vía. Composición química y retorno de
nutrientes por desfronde enlazan directamente con los dos capítulo siguientes
en los que se abordará el estudio de las relaciones entre el retorno vía
hidrológica y vía detritus (capítulo 4)y la dinámica de la descomposición
(capítulo 5)
3.2. MATERIAL Y MÉTODOS.
3.2.1. Recogida de muestras.
El desfronde de las cuatro especies se recolectó mediante trampas de
recogida continua de diseño circular para los pinos y cuadrado para las 2
arbustivas. Las trampas redondas constan de un aro metálico que abarca 0.28 m
de superficie al que va cosida una red de nylon de 2 mm de luz. Estos
disposivos se colocan tan sólo a 25 cm del suelo con el fin de poder recoger
también hojarasca procedente de las ramas de la base del árbol. Por su parte 2
las trampas cuadradas tienen una superficie de 1 m con red de 2 mm de luz
disponiéndose a 10 cm del suelo ya que tanto la j a ra como el gaznacho tienen
ramas a muy poca altura. Se escogieron dos parcelas en la zona repoblada por
P. pinaster, coincidiendo con los puntos altitudinales extremos de la cuenca
plantados con esta especie (1540 y 1690 m.s.n.m.). En la parte alta de la
cuenca (1740 m.s.n.m.) se escogieron otras dos parcelas, una de ellas
representativa de la zona más densamente poblada de P. nigra y A. decorticans
y otra, muy próxima, dominada por el jaral de C. laurifolius. Se colocaron
diez dispositivos para cada una de las especies de pino y 5 para cada especie
arbustiva. La no existencia de una cubierta continua o cerrada por parte del
estrato arbóreo y la distribución horizontal del matorral rellenando los
huecos dejados por el pinar condicionó el tipo de muestreo. Así, los
99
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
dispositivos de recogida se colocaron bajo cubierta de individuos de cada
especie seleccionados al azar. Posteriormente los resultados de cada colector
se corregirán por el porcentaje de cobertura de cada especie en la cuenca.
Este tipo de muestreo estratificado es frecuentemente empleado en la recogida
de desfronde en aquellas poblaciones vegetales que presentan un patrón
espacial disperso con una baja cobertura de suelo: dehesas (Escudero et al.
1985), algunos matorrales mediterráneos (Merino et al. 1990), matorrales de
zonas desérticas (West 1979) o plantaciones jóvenes de coniferas (Cortina
1992).
La recogida de material se realizaba con una periodicidad mensual excepto
en algunas ocasiones, como por ejemplo durante algunas nevadas invernales en
las que no se pudo acceder a la zona de estudio. El desfronde de cada colector
era empaquetado y etiquetado de forma individual y transportado al
laboratorio. Las trampas se pusieron en el campo el 15 de Junio de 1986. El
período de seguimiento del presente estudio abarca el comprendido entre el 15
de Julio de 1986 y el 27 de Septiembre de 1989.
3.2.2. Preparación de las muestras.
Una vez en el laboratorio las muestras se dejan secar al aire o bien se
meten uno o dos días en la estufa para eliminar el exceso de humedad sobre
todo en las recogidas de otoño-invierno. Posteriormente se procede a la
separación en fracciones y especies de cada muestra individual. En los casos
en que el material recogido en determinadas trampas fuese muy abundante
solamente se separaba en fracciones una alícuota de éste. Posterioremente el
material se secaba en estufa a 70°C hasta peso constante.
3.2.3. Análisis químico.
3.2.3.1. Trituración.
En cada recogida mensual, se preparaba una muestra compuesta de cada
fracción y especie mezclando el contenido de las distintas trampas. Cada una
de ellas se tri turaba en un molinillo limpio. En algunos meses concretos en
100
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
los que las recogidas de algunas fracciones eran mínimas (meses de primavera
fundamentalmente), la muestra compuesta se preparaba agrupando los contenidos
de dos meses sucesivos. Una vez molidas las muestras se almacenaban hasta su
posterior análisis químico.
3.2.3.2. Técnicas analíticas.
Las muestras son mineralizadas por via húmeda o semihúmeda según los
casos.
Para el nitrógeno se realiza una digestión húmeda con ácido sulfúrico en
presencia de catalizador de selenio según el método Kjeldahl. La destilación y
determinación de N se realiza con un aparato Kjeldahl Autoanalizer. El peso de
la muestra utilizado fue de 0.2 g.
Para el P y cationes se realiza una digestión semihúmeda con alcohol
sulfúrico. Las muestras se introducen en el horno mufla durante una noche a
450 C. Posteriormente se procede a la disolución de las cenizas con ácido
nítrico y clorhídrico filtrando y enrasando con agua desionizada a 50 mi
obteniéndose de esta forma la solución madre. El peso de la muestra utilizado
fue de 0.5 g.
El P se determinó con el método del amarillo de vanadato-molibdato a
par t i r de las soluciones madre convenientemente diluidas en un
espectrofotómetro Shimadzu UV-160.
Los cationes se determinaron también a part ir de diluciones de las
soluciones madre en un espectrofotómetro de absorción atómica IL S-ll con
llama aire/acetileno tomando las siguientes precauciones:
- Para la determinación de Ca y Mg se añadió una disolución de óxido de
lantano al 0.17. tanto a patrones como a muestras para evitar interferencias.
- A los patrones de Na se les añadió un exceso de K para que la
proporción de ambos elementos sea similar al existente en las muestras.
101
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
- Para todos los cationes se aplica una corrección secuencia! por la
deriva instrumental de la linea base del aparato, utilizando las lecturas de
los patrones y blanco pasados cada diez-quince muestras. El tiempo de
integración de cada lectura fue de cinco segundos.
En el análisis de todos los elmentos se realizaron un buen número de
replicados a distintos niveles: muestra molida, solución madre y determinación
analítica.
La composición elemental de la hojarasca solamente se determinó durante
el período Julio 1986 - Junio 1988. Para calcular los aportes del año
siguiente se utilizó la concentración media mensual de los dos años
anteriores.
3.3. RESULTADOS Y DISCUSIÓN.
3.3.1. BIOMASA DEL DESFRONDE EN LA CUENCA "NACIMIENTO". ASPECTOS
GENERALES.
El desfronde medio anual es de 3.19 T/ha.año en RPP y de 2.67 en RPN
(tabla 3.1). La dominancia del pinar es patente en RPP zona en la que P.
pinaster aporta el 64% de la materia seca, mientras que en RPN son las
especies arbustivas quienes contribuyen en un mayor porcentaje al desfronde
total (567.).
Prácticamente la totalidad del desfronde recogido corresponde a la
denominada fracción fina (Verdú 1984). Dada la juventud de las especies
muestreadas, la fracción gruesa del desfronde, (ramas de diferente calibre)
carece prácticamente de importancia. Sin embargo, algunas tormentas de viento
y nieve produjeron caída o rotura de individuos enteros, sobre todo de
P.pinaster pero la contribución a la caída anual no ha sido cuantificada. La
fracción gruesa recogida en las trampas corresponde a ramas con diámetro
102
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.1.- Desfronde medio anual (g/m .año), desviaciones estándar y contribución de cada especie (%) respecto al desfronde total en las dos grande regiones de la cuenca "Nacimiento".
RPP
especie media ds 7. respecto al to ta l
P.pinaster 204 44.9 63.9
A.decorticans 46.4 18.8 14.5
C.laurifolius 68.7 8.6 21.5
TOTAL 319.1
RPN
especie media ds 7. respecto al total
P. nigra 114.3 18.1 43
A. decorticans 116.17 47.1 43.3
C.laurifolius 37.26 4.7 13.9
TOTAL 267.73
inferior a 1 cm que sólo supone el 2-5 7. del total anual medio (tabla 3.2). La
fracción fina del desfronde en este estudio ha sido separada en
hojas/acículas, flor, frutos, semillas y corteza, esta última sólo
diferenciada en el matorral dónde su contribución tiene cierta importancia. En
la tabla 3.2 se presentan estos datos para cada año de muestreo junto con los
valores anuales medios y correspondientes desviaciones standard.
Como era de esperar, la fracción hoja/acícula es la que contribuye de una
manera más destacable al desfronde total con valores medios de 2-2.5 T/ha.año
(76-81 %). Además, la caída de hojas/acículas se produce durante todo el año
con dos períodos de máxima producción, uno en el período estival o en otoño en
algún caso, y el segundo, más variable en intensidad en los meses de invierno
debido a factores predominantemente externos (ver más adelante).
103
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.2. Desfronde conjunto total en las dos regiones(g/m2).
En la parte inferior rporcentaje medio de cada fracción.
RPP HOJAS FLOR FRUTOS SEMILLAS RAMAS CORTEZA REPRO0UCCI0N TOTAL
jul86/jun87
jul87/jun88
jul88/jun89
media
ds
cv(%)
RPH
jul86/jun87
jul87/jun88
jul88/jun89
media
ds
cv(X)
Z •edio
RPP
RPM
228.80
307.99
242.90
259.90
42.24
16.25
HOJAS
215.24
166.64
230.20
204.03
33.23
16.29
%HOJA
81.36
76.20
9.70
20.88
18.46
16.35
5.88
35.98
FLOR
13.97
25.01
22.67
20.55
5.82
28.31
XFLOR
5.12
7.67
33.40
21.66
33.51
29.52
6.81
23.07
FRUTOS
28.51
4.04
35.69
22.75
16.59
72.94
7.FRUTOS
9.24
8.50
1.78
.23
1.38
1.13
.81
71.37
SEMILLAS
1.53
.14
2.77
1.48
1.32
88.96
%SEMILLAS
.35
.55
9.43
4.82
12.95
9.07
4.08
44.98
RAMAS
12.46
4.62
27.88
14.99
11.84
78.97
%RAMAS
2.84
5.60
.11
2.35
8.08
3.51
4.11
117.05
CORTEZA
.11
1.53
10.27
3.97
5.50
138.69
XCORTEZA
1.10
1.48
44.85
42.71
53.33
46.96
5.61
11.95
REPRODUCCIÓN
43.99
29.17
61.11
44.76
15.98
35.71
%REPRO0UCCI0N
14.70
16.72
283.04
357.90
317.34
319.43
37.47
11.73
TOTAL
271.80
201.96
329.53
267.76
63.88
23.86
La relación RESTO de FRACCIONES/ACICULAS-HOJAS es de 0.23-0.31 lo que en
principio puede indicarnos una etapa joven en el ecosistema estudiado ya que
según Kira y Shidei en Verdú (1984) el índice se aproxima a 1 en los bosques
cercanos al climax.
En los sistemas forestales en general se constata el predominio de la
fracción hojas en el desfronde, aunque su proporción relativa dependa del tipo
de bosque, estructura y edad de éste y otros factores externos. Read y
Mitchell (1983) citan valores del 697. en el scrub-heath australiano y del 78 7.
en el chaparral de California en estadios próximos a la madurez. Verdú (1984)
104
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
obtiene valores del 58 7. (encinar), 687. (abetal) y 75 7. (hayedo). Bray y
Gorham (1964) encuentran valores que oscilan entre el 60 y el 76 7. para
bosques perennes y caducos de distintas zonas de mundo. Para repoblaciones de
pino en zonas mediterráneas los porcentajes encontrados son igualmente altos.
79.7 7. en P.pinea y 64.7 % en P.brutia de Líbano (Dannaoui, 1981), 88/S para
P.nigra en Sicilia (Leonardi et ai. 1988), 65-72 7. en P.halepensis en Francia
(Rapp 1967)
La segunda fracción en importancia son los frutos/piñas. La caída media 2
anual fue de 22-29 g/m año de los cuales un aporte importante se debe a la
caída de pinas, que aunque escasa y bastante irregular en el tiempo contribuye
con un mayor peso relativo que los frutos del matorral. Las flores son 2
aportadas con una tasa media de 16-20 g/m año. Ramas y corteza aportan 9-14 y 2
3.5-3.9 g/m año respectivamente. La fracción semillas resulta poco 2
importante: 1.13-1.47 g/m año. Esta última fracción ha sido subestimada ya
que las semillas de la ja ra atraviesan la luz de las mallas de recogida y no
se contabilizaron.
Corteza y semillas presentan una fuerte variabilidad interanual seguidas
de ramas y frutos (Tabla 3.2). La caída de corteza y sobre todo de ramas
responde básicamente a factores externos y la variabilidad de éstos puede
fácilmente reflejarse en la caída de estas dos fracciones. En la corteza
intervienen además factores específicos (Bray y Gorham 1964). Por otro lado,
la dificultad de separación de la corteza en el laboratorio,(muy fragmentada y
de muy bajo peso en los matorrales) t rae consigo errores de calculo que pueden
aumentar esta variabilidad. El periodo 87/88 resultó ser un año de muy baja
producción de órganos reproductivos sobre en el matorral lo que provoca un
considerable aumento en la variabilidad para esta fracción. Debido a la
dominancia del desfronde arbustivo en RPN, los coeficientes de variación para
las fracciones reproductivas en esta región son mucho más acusados que en RPP.
En la tabla 3.3 se ha diferenciado el desfronde de cada especie en sus
diferentes fracciones. Los valores correspondientes al matorral están
ponderados para una Ha representativa de toda la cuenca. Los de P. pinaster y
P. nigra están referidos a una ha representativa de cada una de las zonas,
105
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.3. Desfronde de cada fracción para cada una de las especies estudiadas en los
diferentes periodos de muestreo. Se indica el valor medio y la desviación estandard.
Desfronde P.pinaster (g/a^.Csi^jerf icíe repoblada).
jul86/jun87
ju187/junas
jul88/jun89
•edia
S.D.
ACICULAS
140.74
228.69
158.26
175.90
46.55
FLOR
3.03
5.72
5.44
4.73
1.48
PINAS SEMILLAS
28.92
19.63
15.87
21.47
6.72
1.19
.18
.28
.55
.56
RAMAS
2.42
1.90
.83 1.71
.81
RAMAS CORTEZA REPRODUCCIÓN TOTAL
33.11 176.17
25.50 256.08
21.58 180.70
26.73 204.32
5.86 44.89
jul89/sep89 62.49 4.43 .28 4.77 67.28
Desfronde P.nigra (g/aO.Csiperf ic íe repoblada).
ACICULAS FLOR PINAS SEMILLAS RAMAS CORTEZA REPRODUCCIÓN TOTAL
jul86/jun87
jul87/jun88
jul88/junfi9
•edia
S.D.
95.92
90.53
126.58
104.34
19.45
1 .44 ,31
.79
.85
,57
22.19
2.91
0
8.37
12.06
.04 .13
.03 .40
0 1.71
.02 .75
.02 .84
23.66 119.72
3.25 94.13
.79 129.11
9.23 114.32
12.55 18.10
jul89/sep89 11.73 .15 .06 .15 11.94
Desfronde A.decorticana
HOJAS FLOR FRUTOS SEMILLAS RAMAS CORTEZA REPR00UCCI0N TOTAL
Jul86/jun87
jul87/jun88
jul88/jun89
avg std
73.42
33.96
59.21
55.53
19.99
9.14
16.90
15.21
13.75
4.08
4 .03
26.80
10.28
14.45
1.19
.09 2.23
1.17
1.07
8.75
2.70
20.11
10.52
8.84
.05
.26 6.04
2.12
3.40
14.33
17.01
44.23
25.19
16.54
96.56
53.96
129.59
93.37
37.92
jul89/sep89 37.04 6.68 .93
Desfronde C.laurifoliua (g/ar).
.10 .84 .64 7.70 46.21
HOJAS FLOR FRUTOS SEMILLAS RAMAS CORTEZA REPRODUCCIÓN TOTAL
jul86/jun87
jul87/jun88
jul88/jun89
•edia
S.D.
jul89/sep89
35.83
43.38
38.37
39.19
3.84
38.71
1.48
4.70
3.79
3.32
1.66
6.95
1.73
1.40
3 2.04
.84
.95
1.85
1.10
1.48
1.48
.38
.31
.06 1.54
3.53
1.71
1.74
.46
3.21
6.09
6.78
5.36
1.89
7.89
40.91
52.12
50.19
47.74
5.99
47.39
106
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
(mutuamente excluyentes) de la cuenca repobladas con cada una de ellas. P.
pinaster es la especie que aporta una mayor cantidad de desfronde mientras que
C. laurifolius es la de menor contribución. Aplicando un ANO VA se detectan,
diferencias significativas en el desfronde anual medio entre las distintas
especies (p<0.005, F= 13.6). Realizando posteriormente el test de
Newman-Keul's se constata que la cantidad media anual aportada por P. pinaster
es significativamente superior (p<0.05) a las correspondientes a P. nigra, A.
decorticans y C. laurifolius (Tabla 3.4). Por su parte, la contribución de
las especies arbustivas no difiere significativamente, mientras que la masa
media total aportada por P. nigra es superior de manera significativa a la de
la j a ra .
2
Tabla 3.4. Valores anuales medios (g/m ) de desfronde en las cuatro especies y
resultado de la aplicación a posteriori de un ANOVA del test de Newman-Keuls.
Para cada columna los valores seguidos de una misma letra no son
significativamente diferentes.
Especie Hojas/acíc. Flor Frutos Ramas Total
P. pinaster 175.8a 4 .73 a 21.47a 1.71a 204.3 a
P. nigra 104.8b 0.85 a 8.37a 0.75a 114.3b
A. decorticans 55.5C , b 13.75C 10.28a 10.52a 93.4b , C
C. laurifolius 39.2° 3.32a 2 .04 a 1.71a 47.7C
ANOVA P<0.005 P<0.001 n.s. n.s. p<0.005
Respecto al desfronde anual medio de la fracción hojas, las especies
estudiadas son significativamente distintas (p<0,005, F= 15.27) Las acículas
de ambas especies de pino son aportadas en cantidades significativamente más
altas que las hojas de C. laurifolius. Sin embargo, no existen diferencias
107
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
respecto al aporte medio anual de hojas entre las especies arbustivas. El
desfronde foliar en P. pinaster es, a su vez, significativamente superior al
de P. nigra pero no existen diferencias significativas entre esta especie con-
respecto a A. decorticans (tabla 3.4). La alta variabilidad anual de la
fracción frutos/piñas en las cuatro especies condiciona que no se detecten
diferencias significativas entre ellas (F= 1.77 ,n.s.). Lo mismo ocurre con
las ramillas (F= 3.2, n.s.). Respecto a las flores, los resultados del ANO VA
revelan diferencias significativas (F= 17.33, p<0.001) en las especies
estudiadas. El test de Newman-Keul muestra que estas diferencias se deben
solamente a la caída de flores en el gaznacho las cuales son aportadas al
suelo en cantidades significativamente más altas que las de j a ra y las flores
masculinas de los pinos.
Como hemos apuntado en varias ocasiones, resulta difícil a la hora de la
discusión encontrar datos en sistemas de "prebosque" en los que las que las
especies arbóreas resultan demasiado jóvenes para constituir un estrato
arbóreo en sentido estricto. Por ello quizá sea mejor discutir los datos
considerando el pinar y el matorral por separado. No obstante intentaremos
relacionar nuestro estudio con otro tipo de sistemas forestales y arbustivos
de distintas partes del mundo.
Así, en la tabla 3.5 aparecen los valores de desfronde anual para
distintos tipos de bosques en diferentes regiones forestales recopiladas por
Colé y Rapp (1981). La región mediterránea solo está representado por un punto
en el Sudeste francés que corresponde a bosque de Qu.ercus.ilex. El valor
encontrado es de 3.8 T/ha año que resulta bastante próximo a los 3.1
encontrados en este estudio. Hemos optado también por recopilar información
sobre desfronde en otros tipos de bosque (natural o artificial) y de matorral
pero centrándonos ya en la zona mediterránea (tabla 3.6). El valor medio de
3.6 T/ha año resultantes de promediar el desfronde total para estos tipos de
vegetación mediterránea es muy similar a nuestros resultados.
El clima es un factor determinante en la producción de hojarasca (Bray y
Gorham 1964). Estos autores dan unos valores medios de desfronde en distintas
zonas climáticas, valores que oscilan entre 1 T/ha año en la región
108
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.5. Desfronde anual medio en cinco regiones forestales (Según Colé y
Rapp, 1981).
DESFRONDE MEDIO ANUAL REGIÓN N* ESTACIONES (Kg/ha.año)
Boreal (Coniferas) 3 322
Boreal (Caducifolios) 1 2645
Templada (Coniferas) 13 4377
Templada (Caducifolios) 14 5399
Mediterránea 1 3842
TODAS 32 4373
Alpina-Ártica y los 10.9 T/ha año en zonas ecuatoriales. Los valores que estos
autores dan para la zona templada-cálida oscilan entre los 2.4 y 8.1 T/ha año.
El desfronde total en nuestro estudio estaría situado por tanto en el límite
inferior de este rango. Estos autores también señalan la importancia que la
edad del bosque (relacionada a su vez con la densidad y el área basal) tiene
en el retorno de materia por desfronde. A este respecto comentan que el
desfronde aumenta progresivamente hasta una cierta edad (que sitúan en unos 30
años) a partir de la cual no parece existir una variación significativa de
este flujo. Este tipo de comportamiento es muy similar al que sigue la
producción neta (Sharma y Pande 1989). Algunos otros autores han obtenido
resultados que apoyan esta hipótesis de Bray y Gorham (1964): (Sharma y
Ambasht 1989, Rodin y Bazilevich 1967, Colé y Rapp 1981, Gholz y Fisher 1985,
Nuñez 1989, Cortina 1992). Parece que la estabilización de este flujo está
relacionada con el cierre de la bóveda forestal. Sin embargo, esta
característica podría ser también el resultado de una compensación entre un
aumento de producción de desfronde por árbol y una disminución de la densidad
a medida que el bosque madura (Flower-Ellis y Olsson en Verdú 1984). Estos
últimos autores consideran que el desfronde expresado en cantidad de
materia/arbol o cantidad de materia/unidad de área basal podrían ser
parámetros mucho mas precisos cuando se t ra ta de comparar datos de desfronde
109
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro C u a d r i l l e r o
O 4) c
•tí u u, 4)
T3 4>
s W O >
01 3
X I u A
4)
W 4)
01 A a 4>
•U 01
«-4
w oí o
01 —4
T3
C 4>
o t e
A
(Ó J C N H
A 3 C A
V
C O
u
2 SO
A
j a A
Sí g i Sé
oo OS
1)
ü
08 = s = = s
T3 03 O)
06
00 CT>
*-< v *
u 4>
cf •^H
W H) , - s
—4 o s
x : oo O Os
C/5 «-4
*- ofl N
>•. 4)
2S o z
/—s
o o* os «-4
-' • - 1
tí
•w 4)
O = c - H
u V X
*~s
os 00 Os T - t
^ . — i
A
•«-» 4)
O •o A
> oí J
*~ 1/7 00 Os r>4 s - >
. —* A
+-> 4)
O U 4) •g 3 O oí w
*—* o O S
Os T H
>* >o T3 4)
» H
_J
^—1
<# 00 os 1-1
>- >3 T3 l -4)
>
*—«, .—•. * - « Q N
f - 00 ON CT* r-« T H "•w w
a . a - i A A
(X • i a
ofl 4)
* J O C TJ
- « U A A 01 --4 01 " H O «3
• J o
<*~\ « - I
00 Os r - l
^ • 3 O = A
2 A a
,—% p-vO O S * H V *
a Q. A ce
O S
00 *-> Os 00 «-• 00 w os
1-1
. > ^ ^ ~ i
A — i
- ia <tj ty ^-»
»3" «a A 00 4) C OS
f * i - i « ^ 6 0 - -O 4) l -
OC - 3 A T3 C
A U O •u 4) 4) ai > J
t>» (M <# O O
oo CM oo n os o o «* o o O OS Os 00 00
o
O n c i N n H N ^ H N N n
c u o
A
u
A «-4
O c A í- -u. u 3
C/3
A A - J ü i—i —t A U u <*«
•U -A 01 T3 3 3
< U)
A A « *
- t C H U A O u u-, * J - ^ 01 - H 3 A
< U
A U 3
• o A a 4) U
-4->
X u
A C A
¡C O a
A C A
á
A U 3
T3 A
a 4) U
j . ) X
u
A 1-, 4)
t/1 «fc. 3 - H
~< O >s Ü
O O O O O H j i n N i c
o o o o o o O * * ' - ' ' - < «S< 00
í 0 1 / 1 * 0 0 «* o «t f- n CM
sO
01 4)
_ -o A tí 2 ^ c o. A a o) A • «
^ H
A « W U l
A A u 5 ? - 1 A C O - o A C ai
•p A ca 01 u A Lu «i
u -3 U A 3 «
• J en en
o c A
X I V I
_J
O (3 A
J 2 V H
J
A U • ^ A O - 1 tí -4J A CQ u U . 0)
•o l_ 3 « •
en en
A >* tí A
* J 01 • A <
U U
A —l J en
. . 4)
I
o. o A
A 0) >•> O 1» • "
u u
01 0) 3 3
O O
01 - 1 3 A •-1 01 Ü Ü
—i 1H —i - ^ 01 01 01 A A •»-> ••* —4 —i
•"• « t u o ' H ' U C € U A C 0 C 3 4) 1» A - 4 A 4) 9
« (1 U , 3 Í H - ( S i« *», o s -< a o ^ - 4 ü - • 2 A o
- C 01 A —i —1 A A C 01 U O 01
• - « • O A 3 3 S A 3 » A O ~ 1 *-> S - M - M
A • U A —< 0 *-i >> o o o u j < j o a ; a . í :
A —H
~< 0
<*! - H
í 3 * J
0
•
X 4)
,
X 4)
~«4
a
X 4)
• i
,
A O
— i
a 4) U >•>
Q.
, O» O 1 O* O 1 o *
01 01 — 1 • - *
s¿ A A 4) 01 4) - « Q, 4) C * * 4) i C 3 —' —«
- i U A *» Q . X 1 < 01
oí oí oí oí oí
2 2 2 c -< •-4 * H * M " ^ í
o , cu a . o , •*:
A u t>0
c
al
A A A U U 1_ L. U U A A A
a a a u (0 (d (Q u
J3 3 u ü 01
A A 00 00
--£i 01 A A - 4
01
£ £ £ « * li 5, O U U U U U X k U
—t A A N U 4) A U
- I 00.
A 1_ U, O
•«->
A Z
A U U O
•4-)
A Z
u A C
»-4
O c Q
p - i
U, U U tí A A A 'O C C C 4)
v 4 » ) >H i-H O O ü jQ c e s o u u u a :
c c c c •o -o -o >o • H «-< »H -»H
o ü o o A A A A
•~\ ~-l r-t - H i—4 J3 X I X I J3 A Q 0 O 0 *>
a a. a a u 4) 4» 4) 4) X I ce ce os ce •<
u A c
- t
Cu
110
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
en relación a la edad. A pesar de ello, no suelen encontrarse trabajos en los
que el desfronde se exprese en estas unidades. En muchos casos ni siquiera se
podrían calcular puesto que no se indican ni la densidad ni el área basal de
las especies estudiadas.
En relación a lo dicho anteriormente, existe otra consideración indicada
por Bray y Gorham (1964) que consiste en que las diferencias de densidad entre
bosques con bóvedas forestales cerradas, no parecen ejercer mucha influencia
en el desfronde. Sin embargo, en bosques talados o aquellos que por su
juventud aún ni han cerrado sus bóvedas el factor densidad es digno de tener
en cuenta. Nuestro estudio constituye uno de estos últimos- casos, por ello
puede ser interesante, en la medida que los datos bibliográficos nos lo
permitan, comparar los valores de retorno expresados por individuo o unidad de
área basal. Hemos calculado para el conjunto de especies un valor de 1.8
Kg/arbol.año correspondiendo 0.91 a P.pinaster, 0.31 a P.nigra 0.51 a A.
decorticans y 0.12 para C.laurífolius. Lógicamente, en bosques más maduros o
por lo menos en los que la bóveda forestal se encuentra cerrada, los valores
resultan muy superiores a los nuestros dado el mayor tamaño de los individuos.
Verdú (1984) obtiene para tres bosques de encina, hayedo y abetal valores de
2.64, 2.95 y 6.05 Kg/árbol.año respectivamente. Un valor de 5.6 Kg/árbol.año
ha sido calculado para el pinar de Pínus halepensis estudiado por Rapp (1967)
con 60 años y 660 Pies/ha y 9.8 Kg/árbol.año para Pinus pínea estudiado por
Rapp eí al. (1979). En el bosque de Pinus sylvestris estudiado por Santa
Regina eí al. (1989a) puede calcularse un valor por árbol de 4.43
Kg/árbol.año. Por el contrario, en el trabajo de Leonardi eí al. (1988) sobre
bosques sicilianos de Pinus nigra. con 20-25 años de edad se obtiene una tasa
de 0.42 kg/árbol. año, valor muy similar al calculado para la misma especie en
este estudio. Tanto la densidad del pino laricio como el defronde de esta
especie calculado por unidad de superficie son muy diferentes en ambos
trabajos (6573 pies/ha y 2.8 T/ha. año en el trabajo de Leonardi eí al. (1988)
y de 3670 pies/ha y 1.14 T/ha.año en este estudio). Sin embargo los individuos
de cada uno de los pinares parecen comportarse de forma bastante parecida. La
hipótesis de Bray y Gorham (1964) puede ser apoyada con estos resultados: las
diferencias de densidad ejercen una influencia importante respecto al
desfronde en bosques, que como el nuestro y el de Sicilia son aún
111
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
relativamente jóvenes (14 y 20-25 años).
3.3.2 DESFRONDE EN EL PINAR.
3.3.2.1 Desfronde total . Contribución de las fracciones.
El retorno de materia seca procedente del pinar se ha estudiado en cada
una de las áreas repobladas con cada especie dentro de la misma cuenca. Para 2
P.pinaster, el desfronde medio anual supone 204 g/m .año (tabla 3.7). A pesar
de la variabilidad interanual (cv=22%), esta cantidad es significativamente
superior (ver apdo. anterior) a la aportada por P. nigra (114 g/m .año). Dado
que la edad, así como el porcentaje de cobertura y biomasa de cada una de.
estas especies son similares (Capt 2), es muy posible que el factor específico
explique en buena parte estas diferencias significativas. Expresando los
resultados en Kg/árbol.año se obtienen valores del orden de tres veces mayores
en P.pinaster, 0.9 Kg/árbol frente a los 0.3 Kg/árbol del pino laricio. La
diferencia de producción de desfronde entre ambas especies se debe básicamente
a la caída de acículas, sistemáticamente mayor en P.pinaster en todos los años 2
y con un valor medio anual de 175 g/m .año. Esta cantidad es del orden de un
41% mayor que en P.nigra para una cobertura de suelo como hemos dicho,
similar. Aun no teniendo en cuenta el desfronde del 87/88 en P.pinaster que
fue bastante superior al resto de períodos a causa de la fuerte caída invernal
de acículas verdes, esta especie sigue aportando un 28% más de materia seca
que P. nigra. Por otro lado la biomasa de hojas calculada para el año 1986
(4.75 T /ha en P. pinaster y 5.7 en P. nigra) presenta diferencias entre ambas
especies siendo superior en P. nigra especie que, por el contrario produce
menos desfronde. El factor accidental, que comentaremos con detalle en los
siguientes apartados ha de ser tomado también en cuenta a la hora de explicar
las diferencias entre ambas especies. La producción media de frutos resulta un
62% más alta en el pino resinero pero esta diferencia en el valor medio se
debe principalmente a la fuerte variabilidad anual en la caída de pinas en
P.nigra. La producción media de flores es también más elevada en P.pinaster 2
con un valor de 4.73 g/m año frente a los 0.85 del P.nigra. Los variabilidad
interanual de esta fracción es también mayor en P.nigra.
112
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.7. Desfronde de cada fracción para cada una de las especies de pino. Los valores
están referidos al área repoblada con cada especie. Se indica el valor medio y la S.D.
Desfronde P.pinaster (g/m ).(superficie repoblada).
jul86/jun87
jul37/jun88
jul88/jun89
Media
S.D.
jul89/sep89
ACICULAS
140.74
228.69
158.26
175.90
46.55
62.49
FLOR
3.03
5.72
5.44
4.73
1.48
4.43
PINAS
28.92
19.63
15.87
21.47
6.72
0
SEMILLAS
1.19
.18
.28
.55
.56
.28
RAMAS
2.42
1.90
.83 1.71
.81
0
REPRODUCCIÓN
33.11
25.50
21.58
26.73
5.86
4.77
TOTAL
176.17
256.08
180.70
204.32
44.89
67.28
Desfronde P.nigra (g/m ).(superficie repoblada).
jul86/jun87
jul87/jun88
jul88/jun89
Medía
S.D.
jul89/sep89
ACICULAS
95.92
90.53
126.58
104.34
19.45
11.73
FLOR
1f44
.31
.79
.85
.57
.15
PINAS
22.19
2.91
0 8.37
12.06
0
SEMILLAS
.04
.03 0
.02
.02
0
RAMAS
.13
.40 1.71
.75
.84
.06
REPRODUCCIÓN
23.66
3.25
.79 9.23
12.55
.15
TOTAL
119.72
94.13
129.11
114.32
18.10
11.94
La fracción acículas constituye la parte más importante del desfronde en
las dos especies, (833 en P.pinaster y 91 7. en P.nigra) seguido de las pinas
que constituyen el 7.3 % del total en P.nigra y 10.5 en P.pinaster. El resto
de fracciones tiene contribuciones poco importantes: la flor masculina 0.7 7.
en P.nígra y 2.3 en P.pinaster y las ramas con un 0.8 en P.pinaster y 0.6 7. en
P.nigra. La relación resto de fracciones vs. acículas es de 0.16 en P.pinaster
y 0.09 en P.nigra.
Respecto a la variación temporal del desfronde, prácticamente coincidente
con la registrada en las acículas, presenta un patrones bimodal en ambas
especies que, con ligeras variaciones, se mantienen en los distintos años de
113
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
estudio, (figuras 3.1 y 3.2). Se puede constatar la existencia de un primer
pulso relacionado con la abcisión fenológica distribuida en el tiempo de forma
diferente en las dos especies. Mientras que en P.pinaster ésta se produce
fundamentalmente durante los meses estivales (Agosto y Septiembre) en P.nigra
es claramente otoñal repartiéndose de una manera secuencial durante los meses
de Septiembre/Octubre y Noviembre. El segundo pulso se produce durante los
meses de invierno de tal forma que la recogidas durante este periodo son a
veces superiores a las registradas en verano (P. pinaster) u otoño (P. nigra)
(tabla 3.8). Sin embargo, hemos constatado que estos valores no corresponden
a acículas que se desprenden de los pinos durante esa época sino que se t ra ta
en su mayoría, de acículas, ya escindidas del árbol, que han permanecido
enganchadas en las ramas viejas de la base de los individuos desde el momento
de su abcisión natural. Factores externos que actúan con intensidad durante el
invierno (nevadas, fuertes vientos) provocan, entonces, la caída masiva de
parte de este almacén. Por tanto, la bimodalidad en las especies estudiadas
es, en realidad ficticia. Al agrupar las recogidas por estaciones, el verano e
invierno suponen contribuciones similares al total anual en P.pinaster (337. y
367. respectivamente. En P.nigra las estaciones predominantes son otoño e
invierno (437. y 39.8%) respectivamente (tabla 3.8). Por las razones comentadas
anteriormente la contribución invernal provoca una sobrestimación del
desfronde anual mientras que la caída fenológica puede estar subestimada ya
que un porcentaje de ésta tarda incluso años en llegar al suelo.
Se realizó un tes t - t de muestras emparejadas para comparar el desfronde
total entre las dos especies durante todo el periodo muestreado. Los
resultados señalan que las dos curvas resultan ser diferentes desde el punto
de vista estadístico (p<0.005, t=3.4) lo que indica que la curva que describe
P.pinaster a través del tiempo, se sitúa significativamente por encima de la
que sigue P.nigra. Por otro lado, existe una correlación positiva y
significativa pero muy débil (r=0.39, p<0.05) entre ambas curvas. Ello refleja
una débil sincronización de las oscilaciones que sigue el desfronde total
entre los dos pinos. Que esta sincronización sea poco marcada puede explicarse
observando de nuevo las figuras 3.1 y 3.2. Existe un desfase temporal respecto
a la abcisión fenológica entre ambas especies ya que la caída fenológica de P.
nigra está desplazada hacia el otoño en relación a la de P. pinaster. Por el
114
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
g/m2.día 2,5 r
tddftot
P. pin áster
Figura 3.1. Tasa de desfronde total de P. pinaster durante el período de
estudio.
g/m2.día
7 10 1 4 7 8 11 4 7 10 2 5
meses
— tddftot
P.nígra
Figura 3.2. Tasa de desfronde total de P. nlgra durante el período de estudio.
115
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3 .8 . Desfronde estacional (g/m ) para los d i s t i n tos períodos estudiados en ambas especies de pino.
a) P.pinaster b) P.nigra.
ACICULAS REPRODUCCIÓN RAMA TOTAL ACICULAS REPROOUCCION RAMA TOTAL
VERANO 86/87
VERANO 87/88
VERANO 88/89
VERANO 89/90
MEDIA VERANO
S.D. VERANO
OTOÑO 86/87
OTOÑO 87/88
OTOÑO 88/89
MEDIA OTOÑO
S.D. OTOÑO
INVIERNO 86/87
INVIERNO 87/88
INVIERNO 88/89
MEDIA INVIERNO
S.D. INVIERNO
PRIMAVERA 86/87
PRIMAVERA 87/88
PRIMAVERA 88/89
MEDIA PRIMAVERA
S.D. PRIMAVERA
49.10
89.72
40.57
62.49
60.47
21.49
21.61
58.85
63.40
47.95
22.93
57.84
72.42
52.43
60.90
10.34
12.20
7.70
1.86
7.26
4.13
2.48
16.05
3.58
4.77
6.72
6.29
.28
.21
.21
.23
.04
15.56
6.08
15.96
12.53
5.59
14.80
3.15
1.83
6.59
6.41
.89
.64
.12 0
.41
.42
.09 0
.21
.10
.11
1.19
.95
.49
.88
.36
.24
.31 0
.18
.64
52.43
106.44
44.27
67.28
67.61
27.59
21.98
59.06
63.83
48.29
22.91
74.53
79.42
68.90
74.29
5.26
27.24
11.16
3.70
14.03
5.66
11.85
12.62
13.19
11.73
12.35
.68
36.34
38.02
69.74
48.03
18.82
44.82
35.61
40.11
40.18
4.60
2.91
4.28
3.55
3.58
.51
5.51
2.98
.34
.15
2.25
2.53
2.52
.09
.12
.91 1.40
14.18
.03
.33
4.85
8.09
1.44
.15 0
.53
.78
.03 0 0
.06
.02
.03
0 .10 .06
.05
.05
.06
.21 1.65
.64
.88
.04
.09 0
.04
.05
17.39
15.61
13.53
11.94
14.62
3.24
38.86
38.21
69.91
49.00
20.27
59.06
35.85
42.08
45.66
13.57
4.40
4.51
3.55
4.15
1.35
116
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
contrario sí se aprecia una ligera sincronización en los meses de invierno
entre estas dos especies.
El desfronde en las dos especies de pino resulta menor que los resultados
obtenidos por otros autores en la zona mediterránea. Así, en el sur de Francia
se han obtenido valores de 4 T/ha año en una población de Pinus halepensis de
60 años (Rapp 1967) y 7 T/ha año para Pinus pinea de 35 años (Rapp et al.
1979). Dannaoui (1981), obtiene valores de 4.5 T/ha año en P.pinea y 3.4 en
P.brutia en el Líbano. Valores más similares encuentran Leonardi et al. (1988)
en Pinus laricio de Sicilia: 2.8 T/ha.año. En España, Santa Regina et al.
(1989a) obtienen valores muy elevados (7 T/ha) en Pinus sylvestris de la
Sierra de Béjar, (Salamanca) aunque en esta estación las condiciones climáticas
son de tipo mediterráneo húmedo. Alvera (1990) encuentra 4.4 T/ha.año en
poblaciones de Pinus sylvestris en el Pirineo. Verdú (1984) obtiene en el
abetar de la Castanya un valor de 3.43 T/ha. En otras especies perennifolias
mediterráneas como es el caso de la encina se han obtenido valores de 3.8 T/ha
(Lossaint y Rapp 1971), de 5.31 T/ha (Verdú, 1984) y 3.9 T/ha.año (Lledó
1990). Los datos para bosque templados de coniferas son igualmente bastante
superiores. Cromak y Monk (1975) obtienen para Pinus strobus en USA 3.25 T/ha
año, Weber (1987) obtiene 2.4 T/ha año en plantaciones de 21 años de Pinus
banksiana, valor muy cercano al obtenido en P. pinaster de este estudio. Para
otros bosques templados de coniferas de Norteamérica, la producción media
anual es de 5.43 T/ha año (Bray y Gorham 1964). En todos los casos se t r a t a de
bosques o plantaciones de mayor edad que los estudiados y con bóvedas cerradas
o semicerradas, lo que puede influir en su mayor producción.
La menor producción de desfronde en nuestro pinar en relación a la
bibliografía puede ser un efecto combinado de una menor productividad del
pinar estudiado en relación a pinares de otras zona y a su inmadurez. En
nuestro estudio la bóveda aun no está cerrada encontrándose el pinar en una
fase de activo crecimiento. Por lo tanto pueden ser esperables aumentos en el
desfronde de los próximos años hasta que la bóveda llegue a cerrarse y se
produzca una estabilización de este flujo (Bray y Gorham (1964)).
Evidentemente la edad a la que ésto se consigue depende de factores
específicos, ambientales y del tipo de bosque (natural, repoblación). Rodin y
117
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Bazilevich (1967) apuntan a un máximo desfronde a los 40-50 años en La zona
templada-fría europea a partir de los cuales el desfronde se mantiene o
disminuye. Colé y Rapp (1981) lo sitúan a los 30 años. Sin embargo, en otros
casos los valores máximos se consiguen a edades muy tempranas. Gholz y Fisher
(1985) lo encuentra en plantaciones de Pinus ellioti a los 10 años
coincidiendo con la máxima producción de hojas en la biomasa y Cortina (1992)
a los 7 en algunas plantaciones de P. radiata en Catalunya. La inmadurez del
pinar estudiado queda reflejado en el valor del cociente desfronde/biomasa:
6.57. en P. nigra y de 127. en P. pinaster. Rodin y Bazilevich (1967) consideran
como plantaciones maduras aquellas con índices comprendidos entre 1.5-27. y
jóvenes e inmaduras entre 9-127..
La variabilidad anual en el desfronde total de las especies de pino
(CV=167. en P. nigra y 227. en P. pinaster) es moderada si tenemos en cuenta que
son árboles perennifolios y que pueden sufrir caídas fortuitas en cualquier
momento del año. La importancia cuantitativa de aquellas fracciones cuya caída
no está regulada por el propio árbol sino que depende básicamente de factores
externos (madera, corteza) es muy escasa en este estudio. Quizá por ello, no
se hayan observado variaciones interanuales tan acusadas como las que
acontecen en coniferas más maduras (Rodin y Bazilevich 1967) en las que la
contribución de la fracción gruesa es mucho más importante. Sin embargo, en
P.pinaster la caída de acículas es más variable de lo esperable (267.) a causa
de una importante contribución de acículas verdes durante el segundo año.
3.3.2.2. Desfronde de las distintas fracciones.
ACÍCULAS.
En P.pinaster, la caída de esta fracción se produce a lo largo de todo el
periodo de estudio aunque en los meses de primavera el flujo diario resulte
ser muy poco importante (figura 3.3, tabla 3.9a). El valor medio para los t res
ciclos completos es de 1.7 T/ha año, oscilando entre 1.4 T/ha año y 2.2 T/ha
año y con un coeficiente de variación del 267. (tabla 3.7).
118
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.9 a y b. Tasas de desfronde (g/m'.dfa) para las distintas fracciones en P.pinaster y P. nigra.
a) P.pinaster
MES DÍA AñO ACICULAS FLOR PINA SEMILLAS RAMAS REPRO0UCCI0N TOTAL
7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 2 3 4 5 6 7 8 9 10 11 12 2 4 5 6 8 9 9
15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 18 20 22 25 20 22 23 28 31 29 7 11 27
86 86 86 86 86 86 87 87 87 87 87 87 87 87 87 87 87 87 88 88 88 88 88 88 88 88 88 88 88 89 89 89 89 89 89 89
.359
.857
.380
.248
.163
.305
1.474
.233
.176
.206
.085
.106
.249
1.569
1.085
.785
.488
.673
.963
.437
.184
.034
.032
.068
.288
.874
1.393
.230
.357
.376
.449
.026
.035
.067
1.452
.566
.018
.032
0 .002
0 .004
.003
0 0 0
.015
.025
.057
.015
.010
0 0
.005
0 0 0
.005
.095
.069
.013
.019
0 .006
.002
.001
.000
0 .062
.049
.068
.010
0 0 0 0 0 0
.496
0 0 0 0
.437
.451
0 0 0 0 0
.098
0 0 0 0 0 0 0 0 0 i
0 .252
0 0 0 0 0 0
.005
.009
.018
0 .001
.002
.002
0 0 0
.001
.001
.001
0 0 0 0
.003
0 0
.002
0 0 0
.004
.004
0 0 0 0 0
.001
0 .002
.006
0
.015
.009
.005
0 0
.003
.007
.018
.015
0 .001
.007
.021
0 0 0 0 0
.008
.016
.007
0 .003
0 .002
.002
.006
.001
0 .001
.007
0 0 0 0 0
.022
.040
.018
.002
.001
.006
.502
0 0 0
.016
.462
.510
.015
.010
0 0
.007
.098
0 .002
.005
.095
.069
.017
.023
0 .006
.002
.253
.000
.001
.062
.051
.075
.010
.396
.906
.401
.250
-•164
.314
1.982
.249
.191
.206
.102
.574
.780
1.585
1.095
.785
.488
.680
1.069
.453
.193
.039
.129
.136
.307
.899
1.399
.238
.359
.629
.457
.027
.097
.118
1.527
.577
119
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.9 (continuación)
b) P. nigra
MES DÍA AñO ACICULAS FLOR PINA SEMILLAS RAMAS REPRODUCCIÓN TOTAL
7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 2 3 4 5 6 7 8 9 10 11 12 3 4 5 ó 8 9 9
15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 18 20 22 25 20 22 7 28 31 29 7 11 27
86 86 86 86 86 86 87 87 87 87 87 87 87 87 87 87 87 87 88 88 88 88 88 88 88 88 88 88 88 89 89 89 89 89 89 89
.031
.118
.234
.306
.195
.703
.984
.348
.126
.058
.032
.005
.011
.102
.295
.410
.298
.549
.508
.141
.096
.020
.023
.010
.030
.359
.763
.740
.792
.535
.031
.034
.028
.015
.253
.142
.025
.012
.003
.004
0 .001
.001
0 0 0 0
.000
.001
.001
0 .001
0 .001
0 0 0 0
.005
.001
.001
.008
0 .005
0 .004
0 0 0 0
.004
0
.041
0 .099
.023
0 .054
.456
0 0
.046
0 0 0
.094
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0
.001
.000
0 0 0 0 0 0 0 0 0 0 0 0 0 0
.000
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
.001
0 0 0 0 0
.002
0 0 0
.001
0 0 0 0 0
.003
0 .003
0 .003
0 0 0 0 0
.002
0 0
.022
0 0 0 0
.002
0
.065
.012
.103
.028
0 .056
.457
0 0
.046
0 .000
.001
.095
0 .001
0 .001
.000
0 0 0
.005
.001
.001
.008
0 .005
0 .004
0 0 0 0
.004
0
.098
.130
.337
.335
.195
.759
1.443
.348
.126
.104
.033
.006
.012
.197
.295
.412
.301
.550
.512
.141
.099
.020
.027
.012
.032
.367
.765
.745
.792
.561
.031
.034
.028
.015
.259
.142
120
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Observando la curva de variación temporal (figura 3.3) en el desfronde de
acículas, puede apreciarse un comportamiento muy similar a la del desfronde
total, lo cual es esperable si tenemos en cuenta que esta fracción supone un
86 7. del desfronde total. Un primer pico de máxima caída aparece en los meses
secos (Agosto y Septiembre) en los dos primeros años y el cuarto, y algo más
retrasado en el tercer año: septiembre /octubre. Esta caída corresponde a la
abcisión fenológica. En el primer y cuarto año la produción estival de
hojarasca tiende a concentrarse en determinados meses (agosto) mientras que en
el segundo y tercer año se reparte más gradualmente entre Agosto, Septiembre y
Octubre.
Existe un segundo pico en los meses invernales. En esta época son muy
frecuentes en la zona las nevadas y los vientos fuertes que provocan incluso
caída de pinos enteros. Las ramas muertas o semimuertas de los individuos de
P.pinaster permanecen en el árbol ya que nunca se han realizado labores de
podado. Un buen número de acículas escindidas de ramas superiores se
encuentran enredadas en esta ramas y su caída en masa se ve favorecida por las
tormentas invernales que así mismo también provocan escisión de acículas
verdes. Por todo ello, las tasas de caída de acículas en las trampas durante
estos meses llegan a ser incluso más altas que las de los meses de verano
(tabla 3.9a). Por ejemplo, durante el primer año la máxima tasa se alcanzó en 2
Enero con 1.47 gr/m día mientras que la mayor tasa estival, (agosto) fue tan 2
sólo de 0.857 g/m día. En los restantes años fueron más elevadas las tasas de
desfronde estival, pero la contribución invernal siguió siendo importante.
Agrupando las recogida de acículas por estaciones, se encuentran los máximos 2 2
valores medios durante el verano (60.47 g/m ) e invierno (60.9 g/m )
quesuponen porcentajes similares respecto al total de acículas caídas (34.37. y
34.6% respectivamente).(tabla 3.8). Como ya se comentó en el apartado anterior
esta alta contribución invernal es ficticia. Se t ra ta de un error puesto que
no existe un desfronde real sino de un retraso en la llegada al suelo de
acículas escindidas en otros momentos del año. Solamente una pequeña parte de
las acículas recogidas en invierno (identificadas por su color más verde)
corresponde a material escindido en ese período pero este porcentaje no ha
sido calculado. Únicamente la caída invernal 87/88 en P. pinaster fue rica en
acículas verdes.
121
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
g/m2.día
10 i 4 10 2 5
meses
— tddfh
P. pin áster
Figura 3.3. Tasa de desfronde foliar (g/m día) de P. pinaster durante el
período de estudio.
g/m2.día
7 10 1 4 7 10 2 5 meses
8 11 4 7
tdfh
2 .,
P.nígra
Figura 3.4. Tasa de desfronde foliar (g/m6 día) de P. nigra durante el período
de estudio.
122
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Deafronde
En P. nigra, al igual que en P. pinaster, la caída de acículas se produce
durante todo el año obteniéndose las menores tasas en primavera y las mayores
en otoño e invierno (tabla 3.8 y 3.9b). La producción anual de esta fracción
alcanza las 1.04 T/ha año con valores anuales que oscilan entre los 0.9 y 1.26
T/ha.año (Tabla 3.7).
La abcisión fenológica se produce en otoño, alcanzándose las tasas
máximas en el mes de Octubre con valores que oscilan entre 0.3 y 0.7 g/m .día
(figura 3.4) si bien la caída resulta bastante escalonada comenzando a finales
de Agosto y prolongándose hasta Noviembre. El segundo momento de máxima
recogida coincide con los meses invernales y se debe a los mismos motivos que
ya hemos comentado en la especie anterior. Por la proximidad de ambos picos,
resulta difícil discernir dónde acaba la caída fisiológica propiamente dicha y
empieza la accidental sobre todo durante el tercer año en que los dos picos
llegan a unirse. Hemos considerado como la primera la registrada en Septiembre
Octubre y Noviembre y accidental (Diciembre, Enero y Febrero), aunque es muy
probable que pueda existir un solapamiento entre ambas. Durante los dos
primeros años de recogida, el pico invernal fue bastante superior al otoñal
(tabla 3.8).
Se ha realizado el tes t - t de muestras emparejadas para comparar las
curvas de caída de acículas entre ambas especies. En el caso de las acículas
de estas dos especies, la caída resulta superior en P. pinaster a nivel de
p<0.01 (t=2.83) por lo que se puede concluir que, por término medio la tasa de
caída diaria en P. pinaster es superior a la de P. nigra. Por otro lado, se ha
encontrado una correlación positiva y significativa muy débil (r=0.25,
p<0.005) entre ambas curvas. Esto indica que, al igual que para el desfronde
total, existe muy poca similitud en las oscilaciones que sigue la caída de las
hojas a lo largo del periodo de muestreo. Las causas son las mismas que la ya
comentadas en el desfronde total.
La fracción acículas es la predominante en el desfronde de las especies
arbóreas (Rodin y Bazilevich 1967). Bray y Gorham (1964) dan un valor medio de
60-76% de acículas respecto al desfronde total en coniferas. La proporción de
la fracción foliar varía, sin embargo, en función de la edad y desarrollo del
123
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
bosque. Al aumentar la edad, cobra una importancia progresiva la fracción
ramas en detrimento de las acículas (Rodin y Bazilevich 1967, Rapp 1967,
Briggs y Maher 1983). Los porcentajes encontrados en este trabajo son elevados
dada la juventud de la plantación (837. en P. pinaster y 91% en P. nigra)..
Entre los trabajos consultados en la bibliografía para plantaciones jóvenes
del género Pinus, Gholz y Fisher (1985) obtienen un 80% de acículas en P.
ellioti en Florida, Baker et al. (1986) un 907. en P. radiata de N.Zelanda y
Cortina (1992) un 70-83% en plantaciones de P. radiata en Catalunya, valores
todos ellos, muy similares a los nuestros. En pinares de la zona mediterránea,
ya con edades superiores a los nuestros, los valores son en general
inferiores. Leonardi et al. (1988) calculan un 88% en P. laricio, Dannaoui
(1981) un 74-857. en P. pinea y 63-65% en P. brutia de Líbano, Rapp (1967) un
65-72% en P. halepensis. Para otras especies perennifolias, Verdú (1984)
obtiene un 67% en un abetal del noreste de España. En encinares, se han
obtenido porcentajes de hoja del 58% (Ferrés et al. 1984), 52-64% (Lossaint y
Rapp 1971) y 59.1% (Lledó 1990). En Eucalyptus australianos los porcentajes de
hojas obtenidos en bosque maduros son muy bajos, alrededor del 30% (Briggs y
Maher 1983), pero ello parece también estar fuertemente ligado a
características genéticas del género Eucalyptus (Briggs y Maher 1983)
La producción media anual de acículas en los pinos estudiados, 1.7
T/ha.año en P. pinaster y 1.04 en P. nigra, resulta en general más baja que la
de otros pinares y coniferas mediterráneos, pero los trabajos consultados se
refieren a poblaciones más antiguas. En éstos, la caída de acículas oscila
entre las 2-3 T/ha.año (Leonardi et al. 1988; Dannaoui 1981; Rapp 1967; Verdú
1984).
Por su parte Rodin y Bazilevich (1967) dan un rango entre 1.4-6.2
T/ha.año en pinares rusos y de 1.9-3.8 en pinares americanos. Bray y Gorham
(1964) obtienen valores entre 2.2-5.1 T/ha.año para el desfronde acicular en
la zona templada cálida y 1.7-3.9 en la templada-fría. Una vez más nuestros
valores están en el límite inferior o incluso por debajo de estos rangos
debido a las razones ya comentadas en el desfronde total.
La estacionalidad en la caída de acículas ha sido constatada en este
trabajo, así como en pinares y coniferas en general de la zona mediterránea.
124
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

De»fronde
La mayoría de los autores constatan un pico de caída fenológica en verano
causada en buena parte por la sequía estival (Dannaoui 1981, Rapp 1967, Verdú
1984, Cortina 1992). Rapp (1967) obtiene el máximo absoluto en Julio aunque la
caída de acículas es considerable en el periodo junio-septiembre. Dannaoui
(1981) obtiene caídas fenológicas entre Julio-Octubre en P. pinea y P. brutia
aunque aquí los máximos se producen más tarde (Septiembre-Octubre)
coincidentes con el periodo más seco en el Líbano. En el caso de P. pinaster
de este trabajo la caída de acículas se produce también en el periodo seco con
máximos en pleno verano en los dos primeros años y el cuarto) o a principios
de otoño (tercer año) (figura 3.3). Verdú (1984) también encuentra los máximos
en Agosto y Septiembre en el abetal de la Castanya y Cortina (1992) en Agosto
en plantaciones de P. radiata en Catalunya. En zonas mediterráneas de
Australia las hojas de Eucalyptus camaldulensis también caen en periodo seco
(Briggs y Maher 1983).
Sin embargo en otras especies arbóreas perennifolías mediterráneas, la
caída fenológica aparece a finales de primavera, inmediatamente antes de la
sequía estival,(Lossaint y Rapp 1971, Escudero et al. 1983). Lossaint y Rapp
(1971) y Rapp (1967) comentan que esta máxima abcisión foliar se debe
fundamentalmente a la retraslocación de nutrientes para la formación de nuevos
brotes foliares que en la encina se originan en Mayo. En P.halepensis (Rapp
1967) y en P. pinaster del presente trabajo, la caída no viene condicionada
por la aparición de brotes ya que en éstos se originan a principios de
primavera sino por la sequía estival. La caída estival de acículas pued^e
suponer un mecanismo de frenado a la evaporación durante el periodo seco.
Además, en esta época del año la lixiviación se reduce al mínimo permitiendo
así una mejor conservación de los nutrientes contenidos en la hojarasca (Núñez
1989).
La caída de acículas durante el periodo seco no aparece solamente en
coniferas sometidas a un fuerte estress hídrico estival como ocurre en zonas
mediterráneas. En otra zonas tropicales también se han encontrado máximas
abcisiones foliares para acículas de coniferas en los periodos menos húmedos
(Bruijnzell 1985, Sharma y Pande 1989).
125
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
El pico invernal que causa la bimodalidad en el patrón de caída de
acículas en las especies de pino estudiadas está sobrestimado por las razones
que ya han sido expuestas. No obstante existe caída de acículas verdes, aunque
sea en una menor proporción, debida a factores externos. Por tanto, puede
considerarse que tanto P. pinaster como P. nigra poseen realmente patrones
bimodales con un pulso de escisión fenológica y otro accidental. Este tipo de
abcisión no fenológica, coincidente o no con los meses invernales ha sido
también detectado en la encina (Escudero et al. 1983, Lossaint y Rapp 1971),
en P. halepensis (Rapp 1967) y P. laricio (Leonardi et al. 1988). Sin embargo,
en otros casos (Lossaint y Rapp 1971) el segundo pico acontece por factores
biológicos. Estos últimos autores constatan un pico a principios de otoño en
encina coincidente con una adicional producción de brotes foliares t ras la
sequía estival. En otras especies arbóreas de la zona mediterráneas son más
frecuentes, sin embargo, los patrones unimodales (Verdú 1984, Briggs y Maher
1983)
En zonas templadas, se han detectado patrones bimodales en el desfronde
foliar de alguna coniferas (Gosz et al. 1972, Bray y Gorham 1964) que suelen
coincidir con finales de primavera y principios de otoño. En el presente
estudio, la abcisión de las acículas del pino laricio se produce masivamente
bien entrado el otoño, caídas ya la primeras lluvias de otoño y transcurrido,
por tanto, el estrés hídrico estival. En poblaciones naturales sicilianas de
esta misma especie, Leonardi et al. (1988) observan también un máximo
desfronde foliar otoñal (periodo Septiembre-Noviembre) en el que un 60% de
todas las acículas caídas anualmente alcanza el suelo de bosque. Esta misma
pauta fue ya apuntada por Ranger (en Leonardi et al. (1988) para bosques de
pino laricio en Córcega. La fenología de esta especie no encaja con el patrón
temporal de otros pinos mediterráneos con abcisión estival sino que le acerca
al comportamiento exhibido por coniferas templadas con caída otoñal (Gosz et
al. 1972, Bray y Gorham 1964). Parece evidente que el factor específico o
genético puede jugar un importante papel. Leonardi et al. (1988) apuntan, sin
embargo, a las condiciones más favorables rspecto al estrés hídrico que
presenta el área de distribución de las masas de esta especie en Sicilia. En
esta isla los pinares del pino laricio se localizan en altitudes próximas a
los 1800 m.s.n.m.con lo que el clima es mediterráneo húmedo de montaña.
126
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
También en nuestro trabajo la parcela de pino laricio se sitúa a 1740 m s. n.
m. y quizá en esta zona no sea tan patente el estrés hídrico de verano. Sin
embargo P. pinaster ocupa zona no mucho más bajas (<1650m.s.n.m.) y presenta
una caída fenológica de hojas claramente estival. Por tanto nos inclinamos a
pensar en un predominio del factor genético probablemente regulado por el
fotoperíodo. P. nigra parece una especie capaz de explotar eficazmente zonas
montañosas presentando un comportamiento respecto al desfronde muy similar a
la de otros pinos de zonas templadas.
FRACCIONES REPRODUCTIVAS.
La caída media de flores en P. pinaster es de 4.73 g/m año y no presenta
en esta especie demasiada variación interanual (317.) pese a que tanto la
producción como caída de flores puede variar sustancialmente de un año a otro
debido a la influencia de factores externos. Las mayores tasas se producen
como norma general durante el mes de Junio (tabla 3.9a y figura 3.5), aunque
la caída puede comenzar en Mayo y prolongarse hasta Septiembre (años 87/88 y
88/89). El valor más bajo corresponde al primer año de muestreo (3.03 g/m )
(tabla 3.7). En P. nigra los periodos de caída de esta fracción coinciden
básicamente con las de P. pinaster pero las tasas de caída siempre resultan
inferiores (figura 3.6). La producción media fue en esta especie de 0.85 g/m
año con una producción máxima durante el primer año de 1.44 g/m año) (tabla
3.7). El tes t - t de muestras emparejadas muestra diferencias significativas en
la tasa de caída de flor entre las dos especies para el conjunto del periodo
estudiado (p<0.01, t=2.88).
2
La caída de pinas alcanza los 21.47 g/m año, producción algo inferior en
relación a bosques o plantaciones de coniferas más maduras (Verdú 1984, Rapp
1967) no presentando un patrón temporal claro (figura 3.7 y 3.8).
La fracción reproductiva representa un 87. y un 12.87. del desfronde total
en P. pinaster y P. nigra respectivamente. Estos valores se encuentran
próximos a los encontrados por otros autores en coniferas mediterráneas (Rapp
1967, Verdú 1984) y algo superiores a las encontradas por Leonardi et al.
(1988) en pino laricio (57.). La caída de pinas en este trabajo (10.5% en P.
127
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
g/m2.día
0,08
0,06 f-
0,04
0,02
7 10 1 4
tddfü
P-pin áster
Figura 3.5. Tasa de desfronde de flores (g/m'-día) de P. pinaster durante el
período de estudio.
g/m2.día 0,03
0,025 t-
1 [ t ^]~Nt^|Ni^TN| í i • i í i i
7 10 2 5
meses
— - tddífl
8 11 4 7
P.nlgra
Fisura 3.6. Tasa de desfronde de flores ( g / m2 . d í a ) de P. nigrz durante el
período de estudio.
128
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
0.6
0,5
0.4
0,3
0,2
0,1
n
g/m2.día
-
-
-
1 ' > ' -M- i i i i I i i | i Í ( i i ) i Í \ i i
7 10 2 5 8 11 ' 4 7
meses
— tddífr
7 10
P.pin áster
2 .-Figura 3.7. Tasa de caída de pinas (g/m -día) de P. pinaster durante el
período de estudio.
g/m2.día 0,5 r
A i f i \ | i i | i i | i i } i i | i i | i i ( i i
7 10 1 4 7 10 2 5 8 11 4 7
meses
tddífr
P.nlgra
Figura 3.8. Tasa de caída de pinas (g/m -día) de P. nigra durante el período
de estudio.
129
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
pinaster y 7.37. en P. nigra del desfronde total) también están dentro de
margen establecido por Bray y Gorham (1964) para la caída de esta fracción
(1-177,). La marcada variabilidad interanual respecto a la fracción
reproductiva patente en P. nigra ha sido también constatada por otros autores
en estudios con coniferas (Verdú 1984)
El patrón temporal en la caída de flores masculinas en los pinos
estudiados se caracteriza por ser bastante prolongada abarcando un periodo de
varios meses (Mayo-Septiembre). Esta pauta coincide básicamente con la
encontrada por Verdú (1984) en un abetal, aunque en este caso la caída está
ligeramente retrasada con respecto a las de los pinos estudiados
(Julio-Diciembre). Por su parte, Leonardi et al. (1988) encuentran los máximos
desfrondes de la fracción reproductiva en el periodo Junio-Septiembre. No se
aprecia ningún patrón estacional claro en la caída de pinas ni en P. pinaster
ni en P. nigra, sino más bien aportes aleatorios muy irregulares. Tampoco Rapp
(1967) observa ningún comportamiento estacional en la caída de pinas de P.
halepensis.
RAMAS.
La importancia cuantitativa de esta fracción es muy pequeña en este
estudio. Tan sólo se han detectado 1.75 g/m .año en P. pinaster y 0.75 2
g/m .año en P. nigra. La caída es muy irregular en el tiempo y la variabilidad
interanual es así mismo alta (tabla 3.7, figuras. 3.9 y 3.10).
Respecto a la fracción ramas todos los autores destacan su caída
irregular causada predominantemente por factores externos.
3.3.3. DESFRONDE ÉN EL MATORRAL.
3.3.3.1. Desfronde total. Contribución de las fracciones.
El retorno medio anual para el estrato arbustivo vía desfronde resultó 2 2
ser de 141 g/m .año de las que 93 g/m .año corresponden a A. decorticans y
47.7 a C. laurifolius (tabla 3.10). Teniendo en cuenta los similares
porcentajes de cobertura que ocupan en el área de estudio ambas especies
130
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
g/m2.día
7 10 1 4 10 2 5
meses
~— tddíram
8 11 4
P.plnaster
2 .. Figura 3.9. Tasa de caída de ramillas (g/m -día) de P. pinaster durante el
período de estudio.
g/m2.día 0,025
0,02
0,015
0,01 -
0,005
0 ^ i _ i i i / | \ i -fZbj. i>t t i i r i V I Y i \ i i | I / I \ . i. i [ i i f i \
7 10 1 4 7 10 2 5
meses
—*- tddíram
11 4 7
P.nígra
Figura 3.10. Tasa de caída de ramillas (g/m4 '-día) de P. nigra durante el
período de estudio.
2 . .
131
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.10. Oesfronde de cada fracción para cada una de las especies arbustivas en los diferentes periodos de muestreo. Se indica el valor medio y la desviación estandard.
Oesfronde A.decorticans (g/ar).
jul86/jun87
jul87/jun88
jul88/jun89
Media
S.D.
jul89/sep89
HOJAS
73.42
33.96
59.21
55.53
19.99
37.04
FLOR
9.14
16.90
15.21
13.75
4.08
6.68
FRUTOS
4 .03
26.80
10.28
14.45
.93
SEMILLAS
1.19
.09 2.23
1.17
1.07
.10
RAMAS
8.75
2.70
20.11
10.52
8.84
.84
CORTEZA
.05
.26 6.04
2.12
3.40
.64
REPRO0UCCI0N
14.33
17.01
44.23
25.19
16.54
7.70
TOTAL
96.56
53.96
129.59
93.37
37.92
46.21
Oesfronde C.laurifolius (a/«T).
Jul86/Jun87
Jul87/jun88
jul88/jun89
Media
S.D.
jul89/sep89
HOJAS
35.83
43.38
38.37
39.19
3.84
38.71
FLOR
1.48
4.70
3.79
3.32
1.66
6.95
FRUTOS
1.73
1.40
3 2.04
.84
.95
RAMAS
1.85
1.10
1.48
1.48
.38
.31
CORTEZA
.06 1.54
3.53
1.71
1.74
.46
REPRODUCCIÓN
3.21
6.09
6.78
5.36
1.89
7.89
TOTAL
40.91
52.12
50.19
47.74
5.99
47.39
(34.8% y 27.8% respectivamente), puede argumentarse que A. decorticans aporta
al suelo una masa bastante superior a la de C. laurifolius pero sin embargo,
estadísticamente no se han detectado diferencias significativas respecto al
desfronde medio anual entre ambas especies (ver apartado 3.3.1). En cambio, el
test t de muestras emparejadas sí refleja diferencias significativas entre las
tasas diarias de desfronde de ambas especies a lo largo de todo el periodo de
muestreo siendo más elevadas en A. decorticans (p<0.05, t= 2.46). De la tabla
3.10 también puede destacarse que mientras la j a ra manifiesta una caída más
regular en los distintos años (con un CV de 12%), A. decorticans presenta una
mayor variabilidad interanual (CV=40.6%) debida fundamentalmente a la
irregular caída de fracciones reproductivas en los t res periodos completos de
muestreo.
Como ya se comentó en el capítulo 2, las dos especies tienen biomasas
132
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
similares en la cuenca pero mientras que A. decorticans la acumula en pocos
individuos (1832 pies/ha), C. laurifolius tiene una estrategia más invasora
repartiendo la biomasa en un número mayor de individuos (4080 p/ha). Estas
diferencias estructurales se ponen de manifiesto cuando se expresa el
desfronde medio anual tomando como unidad el individuo. Así A. decorticans
retorna una media de 0.51 Kg/individuo.año. y C. laurifolius 0.11
Kg/individuo.año. El valor encontrado en A. decorticans resulta elevado si lo
comparamos con el de los pinos (0.9 y 0.3 Kg/árbol. año). Las diferencias
radican fundamentalmente en la importancia de la fracción reproductiva en el
gaznacho que supone 0.20
Las diferencias en la producción entre ambas especies se deben
básicamente a la fracción frutos (tabla 3.10), superior en A. decorticans
durante el periodo 86/87 y sobre todo en el 88/89 y a las flores cuya
producción es un 75% más alto en A. decorticans que en C. laurifolius (13.75 2 2
g/m .año frente a 3.32 g/m .año)(t=4.1, p<0.05). También la fracción ramillas
es superior en un 85.9% en A. decorticans. Por el contrario la caída media
anual de hojas es similar en ambas especies y no difiere de manera
significativa (t= 1.39, ns). La contribución de hojas en A. decorticans es de 2 2
55.53 g/m .año y 39.19 g/m año en C. laurifolius con una variabilidad anual
de CV= 35.99% en A. decorticans y CV= 9.7% en C. laurifolius (tabla 3.10).
La fracción hojas es la predominante en ambas especies suponiendo un
59.47% del desfronde total en A. decorticans y un 82% en C. laurifolius. La
fracción flor y frutos supone un 25% del total en el gaznacho y sólo un 11% en i
C. laurifolius. Cabe destacar la contribución de ramillas en A. decorticans :
un 11.26% del total, teniendo en cuenta que esta fracción sólo consiste en
tallos y ramillas muy finas. Esta fracción es menos importante en C.
laurifolius (un 3.1.% aunque su caída está más repartida en el tiempo. La
fracción corteza supone pequeños porcentajes del total en ambas especies, un
2.27% en A. decorticans y un 3.5% en C. laurifolius.
Respecto a la variación estacional aparece un patrón bimodal en las dos
especies aunque muy poco marcado en el caso de la jara . El pulso estival en
los dos arbustos corresponde a la caída fenológica mientras que el invernal,
133
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
cuando existe, responde a factores externos. En general, el desfronde de ambas
especies no parecen verse muy afectado por los factores ambientales invernales
contrariamente a lo que ocurría con el pinar. En al caso de la jara , el pico
invernal de caída no fenológica prácticamente no existe (figura 3.11) y en el
gaznacho su importancia varía según los años (figura 3.12) pero nunca llega a
superar a los pulsos estivales de caída fenológica. Varios factores puede
influir en la mayor o menor incidencia de estas condiciones ambientales sobre
las 4 especies estudiadas. Entre ellos, la propia estructura del bosque que
permite un mayor resguardo del viento a las especies arbustivas. Además en los
pinos concurren una serie de características no presentes en el matorral que
favorecen la caída de sus acículas. La presencia en los individuos de acículas
de edades superiores al año puede favorecer su abcisión prematura debido a la
incidencia de factores ambientales. Como ya se comentó anteriormente, también
existen considerables cantidades de acículas desprendidas del árbol que
permanenecen enganchadas sobre las ramas de la base de los pinos cuya caída se
ve favorecida por estos factores.
La caída fenológica comienza en A. decorticans en Mayo, Junio. Durante
estos meses es cuando se registra la máxima caída de flores y la mínima de
hojas, (tabla 3.11b). La caída se prolonga a lo largo de Julio, Agosto y
Septiembre (figura 3.12) durante los cuales caen fundamentalmente las hojas.
En la j a ra la abcisión fenológica se produce casi exclusivamente durante los
meses de verano. En esta época, las fracciones mayoritarias en el desfronde de
esta especie (hojas y flores) alcanzan su máxima producción (tabla 3.12a). Los
máximos absolutos están comprendidos entre 0.67-1.15 g/m .día y se registran
en el mes de Agosto excepto el año 87/88 en que se alcanzó en Julio. En A.
decorticans la máxima producción se reparte entre los meses de Julio y agosto
(primer y cuarto periodo) o se concentra más en uno de ellos (periodo segundo
y tercero (figura 3.12) (tabla 3.12). Las tasas máximas de producción están
comprendidas entre 0.62-1.3 g./m .día. Por tanto, la estación predominante
para el desfronde es el verano con valores medios anuales de 50.2g/m .año en 2
A. decorticans y 40.17g/m .año en C. Laurifolios.(tabla 3.12) que suponen un
54.4% y un 80% del total respectivamente.
134
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
g/mZdía
10 2 5
meses 8 11 4 7
C. laurltollus
tddítot
2 _,-Figura 3.11. Tasa de desfronde total (g/m -día) de C. laurifolius durante el
período de estudio.
g/m2.día
meses
tddítot
A. decorttcans
2 ,. Figura 3.12. Tasa de desfronde total (g/m -día) de A. decorticans durante el
período de estudio.
135
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.11 a y b. Tasa de desfronde (g/m .día) en C. laurifol ius y A. decorticans durante el periodo de estudio.
a) C. lauri fot iu».
MES OÍA AÑO HOJAS FLOR FRUTO CORTEZA RAMAS REPRODUCCIÓN TOTAL
7 8 9 10 11 12 1 2 3 4 5 6 7 3 9 10 11 12 2 3 4 5 6 7 8 9 10 11 12 3 4 5 6 8 9 9
15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 18 20 22 25 20 22 7 28 31 29 7 11 27
86 86 86 86 86 86 87 87 87 87 87 87 87 87 87 87 87 87 88 88 88 88 88 88 88 88 88 88 88 89 89 89 89 89 89 89
.175
.680
.180
.023
.013
.011
.061
.002
.002
.004
.005
.007
1.031
.168
.069
.029
.003
.009
.049
.008
.010
.002
.007
.139
.714
.171
.065
.017
.018
.008
.010
.002
.009
.542
.498
.009
.017
.003
.001
.011
.007
.005
0 0 0 0
.000
.005
.120
.033
0 0 0 0
.000
0 .000
.000
.000
.052
.016
.005
.011
.008
.004
.001
.002
0 .016
.118
.064
.006
.001
.002
.001
.026
.015
.009
.003
0 0 0
.000
0 0
.001
.011
.005
0 .007
.009
.003
0 0 0 0 0 0
.005
.003
.015
.010
.027
.004
0 .013
.012
.001
0 .001
.001
0 0 0 0 0 0 0 0 0
.001
.000
.004
.002
.003
.006
.011
.005
.004
.002
.002
.001
.001
.001
.002
.002
.002
.001
.033
.042
.003
.002
.005
.014
.015
.003
.001
.010
.005
.008
.006
.002
.001
.010
.001
0 0 0 0 0
.002
.005
.008
.006
.005
.000
.002
.001;
0 .006
.003
0 .003
.008
.004
.004
.004
.003
.004
.003
.018
.005
.002
.037
.022
.014
.003
0 0 0
.001
.005
.120
.034
.011
.005
0 .007
.009
.003
.000
.000
.000
.052
.016
.005
.015
.012
.019
.011
.028
.004
.016
.131
.077
.007
.208
.689
.184
.070
.040
.033
.070
.004
.003
.014
.006
.011
1.152
.203
.084
.036
.007
.027
.078
.021
.019
.005
.012
.193
.732
.183
.085
.031
.041
.028
.076
.053
.032
.678
.583
.033
136
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.11. (continuación).
b) A. decortfcare.
MES DÍA AñO HOJAS FLOR FRUTO SEMILLA RAMAS CORTEZA REPRODUCCIÓN TOTAL
7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 2 3 4 5 6 7 8 9 10 11 12 3 4 5 6 8 9 9
15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 18 20 22 25 20 22 7 28 31 29 7 11 27
86 86 86 86 86 86 87 87 87 87 87 87 87 87 87 87 87 87 88 88 88 88 88 88 88 88 88 88 88 89 89 89 89 89 89 89
.540
.561
.146
.104
.119
.348
.478
.027
.052
0 0
.734
.170
.023
.021
.023
.041
.011
.017
.068
.002
.001
.067
.978
.224
.097
.041
.019
.046
.048
.065
.150
.483
.507
.028
.170
.025
0 0 0 0 0
0 .019
.052
.035
0 .004
.005
.003
0 0 0 0
.003
.165
.372
.295
.082
.040
.013
.007
0 0 0 0
.029
.126
.026
.054
.003
.124
0 0 0 0
.002
0 0 0 0 0 0
.001
0 0 0 0 0 0 0 0
.002
.177
.318
.074
.165
.085
.012
.001
0 0
.014
.011
.002
0 .032
.003
.002
.001
0 0
0 0 0 0 0 0 0
.003
0 0 0 0 0 0 0 0
.042
.015
.002
.010
0 0 0 0 0 0
.001
.004
.015
.005
.004
.010
.015
.095
.133
.006
0 0 0 0
.001
.009
.009
.008
.014
.012
.005
.016
.002
.001
.025
.030
.003
.010
.003
0 .213
.019
.016
.009
.004
.019
.001
.002
0 0 0 0 0 0
0 0 0 0 0
.000
0 .001
0 .004
.000
.002
0 .001
.001
0 0
.001
.002
.002
.002
.070
.005
.008
.003
.003
.015
.002
.174
.181
.003
.002
.001
0 .002
-0
.019
.052
.035
0 .004
.005
.006
0 0 0 0
.003
.165
.372
.298
.302
.373
.089
.182
.085
.012
.001
0 .029
.139
.037
.060
.731
.747
.153
.116
.135
.443
.613
.033
.071
.052
.035
.734
.175
.038
.037
.031
.059
.023
.024
.087
.169
.375
.389
1.309
.601
.199
.228
.106
.341
.073
.089
.190
.629
.578
.091
137
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadr i l l ero
< o
«o w- rsi
O cO i n ^fl •# N N. >í St 2
£ 2 CO co N"l LA
¡X r . M CO
ru - * o o > T— NO
o r j m sO fM
•o r - w\ co n N
< í o ro
•O o « - CO fM O»
r\j «— •—
I * 3 3 c
»- -* N is."
3 s — 1*1 <~ o S 8 a
ie í » s « O >- l«1
wn Í M m o
r » o o o eo H 8 Pj
-O o' o u
8
5 § á á N8 « § fO M
«- oo eo sf «- v i •o — ro
O fM
N> eo
ES °
r«- -O f > r -fM *»• m K l ¡s § NÍ- O <\i
1*1 *- O R -* r - f » M fO N Í *— Pí 3 8 t:
K» KI •o r*-m <r r+* -<t *t t\i *t
N-~
fM fM ,-f -
* • -
fM O fM
N M
O fM
K fM
N . •O
IN. ir» ¡* í:
í 3
3 2 B JO o K fM
fM « - » •
3
8 ce a. tu ce
</> < O
fM 00 f > « -f > fM N . f*. s ¡8
ti "
fM « - «tf >» fM « - 8! 3 o N» in ta q -jr
>r 4 ca * fc *— (M ro
00
s o ac
< ce UJ
>
s CO
o at
< ce UJ
>
8 . 3
o
>
o o > s .
f > 00
o 3E
< ce LU
>
¡3 eo
§ 1 i 5 5 5
i 5 e o a i -_ o < ss
s a o
>» ^ x .
co co eo
o a o at 2 atoe ce ce UJ UJ UJ
• > > > a ! ae se
¡u ce
Ss —i ae
< 85
N 5 o CO 00 00 * s . ""s. •*%.
00 00 CO
< < < ac ac ac UJ LU UJ 2 * > >
í í í ce ce ce o . o . o .
s < I i S i . 0 . < 82
138
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Para establecer posibles diferencias en el patrón temporal del desfronde
total de cada especie, se realizaron análisis de correlación entre años. Los
resultados indican que la dinámica temporal es similar entre el primer y
tercer año en la ja ra (r=0.99, p<0.001) lo que significa una sincronía en las
oscilaciones de la caída total entre estos dos periodos. No se encontraron
relaciones significativas entre los demás periodos. En el gaznacho, tampoco se
obtuvo ninguna relación significativa entre años lo que indica variaciones de
un año a otro en la dinámica temporal del desfronde total en esta especie.
Los valores anuales medios en el desfronde de las especies arbustivas
(0.93 T/ha.año y 0.48 T/ha.año en A. decorticans y . C. laurifolius
respectivamente) son bajos en relación a la mayoría de los trabajos
consultados para sistemas arbustivos mediterráneos. Los valores más próximos
son los 0.71 T/ha-año que obtienen Merino et al. (1990) en un matorral de
Cistus libanotis y Rosmarinus officinalis en Doñana y los 0.87 T/ha.año para
chaparral de Adenostoma fasciculatum en California (Read y Mitchell, 1983).
Pero en la mayoría de los casos se obtienen producciones más altas. Read y
Mitchell (1983) observan un rango en matorral mediterráneo cercano a la
madurez entre 1- 3.4 T/ha-año. Núñez (1989) obtiene 2.8 T/ha-año en jarales
densos y maduros de Cistus ladantfer en Extremadura, Lossaint y Rapp (1971)
1.3-3.9 en coscoja con 100% de cobertura, Merino et al. (1990) 2 T/ha.año en
brezales de Doñana, Lavado et al. (1989) 1.2 y 2.3 T/ha.año para Myrtus
commnis y Pistacia lentiscus en Extremadura y Gray y Schlesinger (1981) 1.6 y
2.7 T/ha-año para chaparral californiano en distintos lugares.
Hay que tener en cuenta que los trabajos anteriormente citados se
refieren normalmente a sistemas maduros y sobre todo con coberturas
considerables. Sin embargo en nuestro trabajo, el porcentaje medio de suelo
que cubren los arbustos en el conjunto de la cuenca no es excesivamente
elevado (aproximadamente un 307.). Por tanto, aparte de otros muchos factores,
estas dos características han de tenerse en cuenta a la hora de discutir la
menor producción encontrada para A. decorticans y C. laurifolius. Si
calculamos el desfronde suponiendo que cada especie tuviera una cobertura del
100% los valores obtenidos serían entonces de 2.6 T/ha.año en A. decorticans y
de 1.72 T/ha.año en C. laurifolius que ya resultan muy similares a los
139
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
encontrados por otros autores.
El porcentaje de masa que supone el flujo de desfronde respecto a la
biomasa acumulada es del 237. en A. decorticans y de 14.2% en la jara. Este
cociente disminuye con la edad. Así, Gray y Schlesinger (1981) calculan en
otros matorrales mediterráneos más maduros un rango entre el 6% y el 157.. En
el trabajo de Márquez et al. (1989) se obtienen cocientes del 27.8% en jaral
de C. ladanifer de 10 años y del 9.2% a la edad de 20 años.
3.3.3.2. Desfronde en las distintas fracciones.
HOJAS.
Como se ha comentado anteriormente, la caída de hojas representa la
fracción predominante en el desfronde de A. decorticans con un valor medio de
55.5 g/m .año (59.5% del total) (tabla 3.10). Los valores extremos oscilan 2
entre 34-73 g/m .año. El pulso de caída más importante tiene lugar en los
meses secos (julio-agosto) (figura 3.14), bien concentrado en un. solo mes
(segundo y tercer periodo) o repartido entre ambos (primer y cuarto periodo).
Las tasas medias diarias para el estos dos meses son similares entre años
(tabla 3.11b) oscilando entre los 0.45 g/m .día del 87/88 y los 0.55 g/m2.día
del 86/87. La producción de hojas durante Julio 1987 es sin embargo
significativamente más baja que la de Julio de 1988 (p<0.05, t= 4.55) no
existiendo diferencias significativas, (t= 0.21, ns) en los aportes al suelo
durante julio-agosto de 1986 respecto al mismo periodo de 1989. Durante el
primer año, la fracción porcentual que Julio y Agosto suponen respecto a la
caída total de hojas es bastante inferior a los demás años (un 45.7%) debido a
un importante pulso de caída de hojas verdes en los meses de invierno. Hemos
realizado un análisis de la varianza para comparar las caídas invernales en
los diferentes periodos de estudio. Los resultados indican que sí existen
diferencias significativas entre estos pulsos (F= 7.28, p< 0.05). Aplicando
posteriormente un test de Fisher se constata que solamente la caída invernal
del primer período es significativamente superior (p<0.05) a la de años
subsiguientes. Su contribución al total anual es muy elevada (un 34%). Durante
este invierno se produjeron fuertes nevadas y ventiscas que sólo permitieron
el acceso a alguna de las trampas de A. decorticans situadas bajo individuos
140
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
g/m2.día
-i—l—L, •J^TN.
4 7 10 2 5
meses
— tddíh
C. laurUoltus
2 . . Figura 3.13. Tasa de desfronde foliar (g/m -día) de C. laurifolius durante el
período de estudio.
g/m2.día
- — tddíh
A. decortlcans.
Figura 3.14. Tasa de desfronde foliar (g/m2-día) de A. decorticans durante el
período de estudio.
141
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
que, además, habían sido seriamente dañados por la nieve con lo que es muy
probable que este pulso invernal haya sido sobreestimado.
Para el resto de los meses la caída de hojas es poco importante
predominando las hojas verdes sobre las secas. En Mayo y Junio no se detecta
caída de hojas (segundo y tercer año) o la cantidad es prácticamente
despreciable. Esta época coincide con el pulso fenológico de caída de flor.
2
La producción media de hojas en C. laurifolius fue de 39.19 g/m .año
presentando esta especie una fuerte regularidad interanual (CV= 2.55%) (tabla
3.10). La bimodalidad en el flujo de hojarasca es prácticamente inexistente en
la jara. Esta ausencia de un pulso invernal importante, mucho más aleatorio
que el fenológico estival, es probablemente la causa de esta regularidad en el
patrón de caída de hojas de jara. La dinámica temporal viene, pues,
determinada por un importante pico de máxima caída concentrada bien en julio o
en agosto (tabla 3.11a, figura 3.13). Aplicando el test t se constata que no
existen diferencias significativas entre las tasas de desfronde durante esos
pulsos de máxima caída. La cantidad de hojas recogidas en el periodo estival
varía poco entre años oscilando entre los 31.9 y 38.7 g/m .año, suponiendo
entre un 88.067. y 897. del total anual, (tabla 3.12)
Se ha encontrado una buena correlación (r=0.99, p<0.001)entre la caída de
hojas durante el primer y tercer año en C. laurifolius lo que indica una
sincronización en las oscilaciones del desfronde foliar durante estos años
(figura 3.13). Para el resto de ios periodos no se han encontrado relaciones
significativas. En A. decorticans, la correlaciones encontradas entre años no
son significativas o bien significativas pero muy bajas.
En los sistemas arbustivos es también la hoja la fracción mayoritaria del
desfronde. El valor obtenido para C. laurifolius (827.) se encuentra cerca del
límite superior del rango obtenido a part ir de los datos de varios autores:
60-857. (Gray y Schlesinger 1981, Gray y Schlesinger 1981, Read y Mitchell
1983, Núñez 1989, Rapp y Lossaint 1981, Schwintzer 1984).El relativamente bajo
porcentaje de hojas encontrado en A. decorticans (5975) está causado por un
considerable esfuerzo reproductivo en esta especie manifestado por la elevada
142
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
cantidad de flores en el desfronde (un porcentaje medio de 14.77.) sobre todo
en algunos años concretos. En otros matorrales fijadores de zonas templadas el
porcentaje de hojas se sitúa alrededor del 65% (Schwintzer 1984 en Myrica gale
y Wheeler et al. (1987) en Cytisus scoparius).
La masa aportada por las hojas en nuestras especies (0.55 T/ha.año y 0.39
T/ha.año) es inferior a los datos de la bibliografía por las mismas razones
que ya fueron comentadas en el desfronde total. La producción de hojas en
arbustos mediterráneos oscila entre 1-2 T/ha.año (Núñez 1989, Rapp y Lossaint
1981, Gray y Schlesinger 1981, Lavado et al. 1989). Existen algunas
excepciones pues por ejemplo, Márquez et al. (1989) obtienen valores muy altos
(3.2 T/ha.año) en una población de Cistus ladanifer de 10 años. En zonas
templadas se han obtenido de 1.1 a 1.4 T/ha.año en el matorral fijador Myrica
gale (Schwintzer 1984). Suponiendo una cobertura del 100% los valores de A.
decorticans y C. laurifolius serían respectivamente de 1.5 T/ha.año y de 1.4
T/ha.año, valores perfectamente encajados dentro del intervalo de los
matorrales mediterráneos.
Se han descrito fuertes pautas estacionales en el desfronde foliar para
sistemas arbustivos del matorral chileno, californiano y en la garriga
francesa (Gray y Schlesinger 1981). Estos pulsos se producen en primavera y
están relacionados con la producción de nuevas hojas en esta época del año
(Lossaint y Rapp 1971, Money et al. en Gray y Schlesinger (1981). En jarales
de Extremadura (Núñez 1989) el pico de máxima caída foliar se produce a final
de primavera pero en este caso no coincide con una nueva brotación ya que las
nuevas hojas de Cistus ladanifer se producen en otoño. En las especies
estudiadas de este trabajo la caída es claramente estival y parece más
relacionada con el estrés hídrico puesto que los nuevos brotes de estas
especies se generan básicamente a principios de primavera. En la jara, sin
embargo la abcisión foliar podría verse favorecida por una retraslocación de
nutrientes hacia las flores de nueva formación que aparecen en Junio.
Varios autores destacan la variabilidad interanual en la caída de hojas
causada fundamentalmente por fenómenos atmosféricos (Gray y Schlesinger 1981,
Lossaint y Rapp 1971, Rapp y Lossaint 1981) lo que también acontece en este
143
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

U.J. Moro Cuadrillero
estudio para A. decorticans. Por el contrario la j a ra presenta una gran
regularidad anual a este respecto siendo poco afectada por los factores
externos.
FRACCIÓN REPRODUCTIVA.
La producción media de flores es de 13.75 g/m .año en A. decorticans
(tabla 3.10) con una máxima producción durante el periodo 87/88 (16.9
g/m .año). La variabilidad anual no resulta muy elevada (CV=29.6%). Sin
embargo, esta baja variabilidad es un tanto engañosa ya que está calculada
para los periodos comprendidos entre junio y mayo del año siguiente periodo
que no se corresponden exactamente con el ciclo fenológico anual de la flor en
esta especie (desde Abril hasta Marzo del año siguiente). Observando la figura
3.16 entre los periodos Abril-Marzo de cada año sí que se detecta una cierta
irregularidad en la intensidad de los picos de máxima caída de flor así como
un cierto desfase temporal en ellos.
La caída de flor en A. decorticans comienza tempranamente: Abril y Mayo
en los años 87 y 88 respectivamente, retrasándose a veces hasta Junio (año
89). Los máximos absolutos se alcanzan al mes siguiente: mayo del 87, 0.05 2 2 2
g/m .día; junio del 88, 0.37 g/m .día y Julio año 89, 0.126 g/m .día. (tabla
3.11b).La caída de flores continúa con una menor intensidad durante uno o dos
meses más. Es destacable la importante producción de flores durante el pulso
Mayo-Septiembre de 1988 (figura 3.16) que supuso un peso de 28.9 g/m frente a
los correspondiente picos del año 87 (3.24 g/m ) y 89 (7.49 g/m2). El año 86
no refleja el ciclo completo de caída de flor ya que probablemente ésta
comenzó en mayo o junio y al iniciarse los muéstreos (julio 86) el desfronde
de esta fracción ya comenzaba a declinar. No obstante, en ese mes (Julio 1986)
se alcanzaban valores nada despreciables, (0.17 g/m .día) con lo que puede
intuirse que este ciclo fenológico fue bastante productivo en relación al
desfronde de las flores. El valor de máxima caída del año 88 (registrado en
Junio) es significativamente mayor que el máximo del año 87 (Mayo) (p<0.005,
t=22.9) y el de 1989 (Julio) (p<0.01, t=11.71).
La caída de flores en A. decorticans parece estar relacionada con el tipo
y la cantidad de lluvia registrada en los meses anteriores a la formación de
144
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
0,14
0,12
0,1
0,08
0,06
0,04
0,02
0
g/m2.d¡a
-
-
-
\vpn ' ' ! ' • 1 7 10 1 10 2 5
meses
— - tddffi
C. laurííalius
Figura 3.15. Tasa de caída de flores (g/m -día) de C. laurifolius durante el
período de estudio.
g/m2.día 0,4
0,3
0,2
0,1
I ' ' M n 7 10 1 10 2 5
meses
— tddífl
8 11
A. decartlcans.
Figura 3.16. Tasa de caída de flores (g/m2-día) de A. decorticans durante el
período de estudio.
145
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
la flores en esta especie. El periodo febrero -abril fue más seco en 1987 (39
mm) y más húmedo en el 86 y 88 (64 mm y 134 mm) lo que concuerda con el
desfronde más elevado durante el 86 y 88 y escaso en el 87. En el año 89, la
cantidad de lluvia durante ese periodo fue muy elevada (174 mm) y sin embargo
la producción de flores muy baja. La explicación puede buscarse en que más de
un 80% de esta precipitación cayó en forma de nieve cubriendo el suelo de la
cuenca hasta muy avanzado el mes de Marzo. Esta característica pudo haber
afectado negativamente la producción de flores en el gaznacho.
La producción media anual de flores de jara es de 3.32 g/m .año con un
coeficiente de variación interanual del 47% (tabla 3.10). La caída de esta
fracción se concentra básicamente en los meses de Julio y Agosto pero es
siempre en Julio cuando se alcanzan los máximos anuales (tabla 3.11). Fuera de
estos meses la recogida de flores fue escasa. La caída registrada en Julio de
1986 es significativamente menor que las correspondiente a los pulsos máximos
anuales registrados en Julio 1987 (p<0.005, t=5.96), Julio 88 (p<0.05, t=3.12)
y Julio 89 (p<0.05, t=3.76). La evolución temporal de la caída de esta
fracción es similar entre años como lo constatan las correlaciones
significativas entre todos los periodos. Sin embargo la correlación sólo puede
considerarse elevada entre el segundo y tercer periodo (r=0.95, p<0.001).
Parece existir un patrón cíclico en la producción de flores en esta
especie en el que se alternan un año con bajo desfronde de flores (86 y 88)
con otro de alta producción (87 y 89). (tabla 3.10). La menor eficacia
reproductiva de los períodos 86/87 y 88/89 se manifiesta también en el i
considerable porcentaje del total de flores producidas recogida durante el
invierno en forma de capullos abortados (figura 3.15). No se ha detectado
relación entre la cantidad de precipitación registrada durante los meses
previos a la floración (período Marzo-Junio) y el desfronde de esta fracción
en la jara . Sería necesario continuar el seguimiento del desfronde en esta
especie para establecer si esta conducta responde a un patrón fenológico o
bien a condiciones ambientales que han actuado de manera más marcada de un año
a otro.
La caída media de frutos en A. decorticans es de 10.28 g/m .año con una
146
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
fuerte variabilidad interanual (CV=1407.) (tabla 3.10). Las menores tasas
encontradas durante los años 87 y 89 se relacionan con la menor producción de
flores durante esos años. Los pulsos de caída coinciden con los meses de
verano: Julio y Agosto en el 86 y 89 y Agosto-Septiembre en el 88. Durante
1987 no se registró prácticamente caída de esta fracción (tabla 3.11b y figura
3.18). Durante el otoño-invierno 88-89, fundamentalmente debido a los vientos
y nevadas, se recogieron en las trampas cantidades adicionales de frutos que
permanecían unidos a la planta. En la jara, las cápsulas suelen quedar unidas
a la planta durante largos periodos incluso más de un año (Núñez 1989). A la
abcisión fenológica hay que añadir, pues, la causada por factores accidentales
que provocan la caída de cantidades adicionales de estas cápsulas viejas.
Quizá debido a ello, en este trabajo el patrón temporal del desfronde de
frutos de jara muestra un aspecto irregular (figura 3.17), con pulsos
considerablemente fuertes en invierno e incluso primavera debido a esa caída
tardía de cápsulas. La abcisión puramente fenológica podría considerarse la
producida en los meses finales de verano y principio de otoño (Agosto,
Septiembre, Octubre). Sin embargo, en esta época comienzan ya las primeras
lluvias fuertes con lo que es muy probable que se solape la caída fenológica
con la accidental. En conjunto el aporte medio anual de esta fracción en la 2
j a ra es de 2.04 g/m .año con una considerable variabilidad interanual (CV=
42%) (tabla 3.10).
Sólo se cuantificó el peso de las semillas en A. decorticans. La
producción fue mayor lógicamente durante los años en que se registró una mayor 2
caída de flores y frutos : periodo 86/87 y 88/89 con valores de 1.19 g/m y
2.23 g/m (Tabla 3.10). Los pulsos de caída se registran durante el mes de
Agosto (coincidente con la caída de frutos) y Septiembre (tabla 3.11b).
El esfuerzo reproductivo de A. decorticans es considerable. El gaznacho 2
produce una media de 13.7 g/m .año de flores (14.7% del desfronde total) con
un valor máximo durante el periodo 87/88 (16.9 g/m .año). En general, muchas
especies del grupo de las Cityseas producen gran cantidad de flores. La 2
producción media anual de 3.3 g/m .año aportadas por C. laurifolius es
significativamente inferior a la de A. decorticans y supone sólo un 6.9% del
desfronde total. Este valor resulta bajo si lo comparamos con otra cistácea:
147
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
0,03
0,025
g/m2.cíía
' t ' • I ' M 4 7 10 2 5
meses
tddffr
C. laurlíollus
Figura 3.17. Tasa de caída de frutos (g/m -día) de C. laurifolius durante el
período de estudio.
0,35
0,3
0,25
0,2
0,15
0,1
0,05
0
g/m2.día
-
-
-
-A i . 1 f ,1 1 I I I \ I , I f t 1 f t 1 [ i, í f 1 1 I t - l _ i ¿ 4 -
7 10 1 4 7 10 2 5 8 11 4 7
meses
—~ tddffr
A. decortícans.
Figura. 3.18. Tasa de caída de frutos (g/m -día) de A. decortícans durante el
período de estudio.
148
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Cistus ladanifer que en Extremadura alcanza producciones de 29 g/m2.año (107.
del desfronde total) (Núñez 1989). Pero para esta última especie la proporción
de suelo cubierta por la ja ra es alrededor de un 80% bastante superior a la
superficie cubierta por C. laurifolius. En otras arbustivas mediterráneas
Lavado et al. (1989) obtienen porcentajes de flores en el desfronde inferiores
a nuestros resultados (0.6% en Myrtus communis y 4.2% en Pistacia lentiscus).
Según Bray y Gorham (1964), existen grandes diferencias entre especies
respecto al aporte de frutos en el desfronde, lo que es debido básicamente a
la variabilidad interespecífica respecto al tamaño de los frutos y a su
producción. Hemos encontrado diferencias notables entre A. decorticans y C.
laurifolius en este aspecto. A. decorticans aporta una media de 10.7 g/m .año
(11% del desfronde total) y C. laurifolius 2.4 g/m .año (4% del desfronde
total), sin embargo dada la elevada variabilidad interanual presente en A.
decorticans las diferencias entre las dos especies no resultan significativas.
Lavado et al. (1989) obtienen valores de 4 g/m .año en Myrtus communis y 0.9 2
g/m .año en Pistacia lentiscus (3.3% y 0.4% del desfronde total
respectivamente). En C.ladanifer, Núñez(1989) obtiene 35 Kg/ha.año que sólo
suponen un 0.1 % de la hojarasca anual. Para la garriga francesa Rapp y
Lossaint (1981) obtienen una importancia de la fracción reproductiva en el
desfronde de un 20-27% (50-60 g/m2. año y Gray y Schlesinger (1981) en el
"coastal sage scrub" obtienen 10 g/m .año (5% del total).
Los pulsos de caída de la fracción reproductiva dependen evidentemente de
la fenología de cada especie y su duración es también variable. En A.
decorticans la flor cae tempranamente (normalmente entre abril y Junio)
similar a lo que ocurre en C. ladanifer (Abril-Mayo) (Núñez 1989) y en
Pistacia lentiscus (Abril-Junio) (Lavado et al. 1989).En C. laurifolius la
abcisión de flores se produce de una manera muy concentrada en Julio-Agosto de
forma similar al patrón seguido por Myrtus communis (Agosto-Septiembre)
(Lavado et al. 1989). La caída de frutos es normalmente más prolongada ya que
éstos pueden quedar unidos a la planta durante bastante tiempo tal y como
ocurre en C. laurifolius (presente trabajo) y C.ladanifer (Núñez 1989). El
periodo de máxima caída de esta fracción aparece entre Julio y Septiembre (A.
decorticans del prente trabajo), Octubre-Enero y Octubre-Noviembre en el mirto
149
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
y lentisco respectivamente (Lavado et al. 1989) y Septiembre-Diciembre en C.
ladanifer (Núñez 1989).
RAMILLAS Y CORTEZA.
Estas dos fracciones presentan en las dos especies patrones temporales
muy irregulares debida a su carácter de caída accidental (figuras 3.19 y
3.20). Además el bajo peso de la corteza y su fragmentación dificulta la
cuantificación correcta. En la j a r a la caída de ramillas se registra
prácticamente durante todo el año aunque es más continua durante el invierno
(figura 3.19). Las cantidades aportadas son muy pequeñas, (tabla 3.10) con un 2
valor medio es de 1.48 g/m .año. La fracción corteza es irrelevante en esta
especie durante el 86/87 mientras que en el 87/88 y 88/89 se registraron 1.54 2 2
g/m .año y 3.53 g/m .año respectivamente. Tanto para las ramillas como para la
corteza el periodo 88/89 fue el de máxima producción probablemente debido a la
mayor incidencia de los factores meteorológicos durante ese periodo. En A.
decorticans la producción media de ramillas fue bastante superior a la de la 2
j a ra con un valor medio de 10.52 g/m .año (tabla 3.10). El desfronde medio de 2
corteza alcanzó 2.12 g/m .año. También para ambas fracciones el año que aportó
cantidades más elevadas fue el 88/89 con una producción anual de 20.11 g/m 2
para las ramillas y 6.04 g/m para la corteza. Los principales pulsos de caída
se registraron en el periodo invernal (figura 3.20). 3.3.4. RELACIÓN DEL DESFRONDE TOTAL Y HOJARASCA CON FACTORES CLIMÁTICOS.
Para establecer la relación entre variables climáticas y la
estacionalidad del desfronde se ha realizado un análisis de correlación entre
las distintas variables ambientales (datos del Observatorio meteorológico de
Calr Alto exceptuando la precipitación 1987-1989 tomados de Domingo (1991))
con los datos de desfronde total y de hojas utilizando los datos tranformados
o no en forma logarítmica. El tipo de ecuación mejor ajustada, el índice de
correlación (r) y el nivel de significación se muestra en la tabla 3.13.
En términos generales, la variables de temperatura (bien expresada como
temperatura media máxima, media mínima o media nocturna) están positivamente
correlacionadas con el desfronde a excepción de P.nigra en que estas
150
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

g/m2.día
1 ' ' 1 '
Desfronde
0,016
0,014
0,012
0,01
0,008
0,006
0,004
0,002
0 10 2 5
meses
tddfctz _ e - tddfram
C. laurlíollus
Figura 3.19. Tasa de caída de semillas y corteza (g/m -día) de C. laurifolius
durante el período de estudio.
g/m2,día -i 0,25 0,08
0,07
0,06
0,05
0,04
0,03
0,02
0,01
0
g/m2.día
-
-
r
WfT i ét¡$ trf^.
7 10 1 10 2 5 meses
8 11
tddfram tddíctz
A. decorticans.
Figura 3.20. Tasa de caída de semillas y corteza (g/m -día) de A. decorticans
durante el período de estudio.
151
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
i
3
« a <• « o -O *>
b -« 4 -* > O
ás
d&
o I I u
3
00
o i/)
o i a i .
a
3 o
I/)
i ° 5 •
51
3
i
S o
; a
a» o
*3
51
* s » • -i u
¡T -I u
2 —« C
—t o
1
o
o
M 1/1
3
•5E.
3
o I
o I I
f» l/>
i « o
o a 2
?
o i i u
~> m
3
1 V
1
«i as
! !
a a
íd
>i
152
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
relaciones son negativas. La temperatura constituye uno de los factores
predictores más importantes en relación con el desfronde anual en ecosistemas
de las distintas regiones climáticas (Bray y Gorham 1964). A nivel más local,
la relación entre esta variable y la estacionalidad del desfronde ha sido
también encontrada por Turnbull y Madden (1983) en bosques templados de Nueva
Zelanda. Tres de las especies estudiadas en este trabajo se caracterizan por
una abcisión estival. Read y Mitchell (1983) comentan que el desfronde
fenológico estival en muchas especies mediterráneas está fundamentalmente
causado por las altas temperaturas que favorecen la síntesis del ácido
abcísico. Resulta pues lógico que la máxima caída de hojas coincida con los
periodos del año con temperaturas más elevadas. Solamente en el matorral el
ciclo del desfronde oscila de una forma más o menos sincrónica con la
temperatura (relación positiva y significativa) (tabla 3.13 y figura 3.21). No
ocurre así en P.pinaster, a pesar de presentar también una abcisión estival.
En esta especie las recogidas invernales de acículas ya caídas altera y
enmascara de forma patente los resultados estacionales y ésta sea probablemnte
la causa de que no se haya llegado a ninguna relación significativa ni con la
temperatura ni con ninguna de las demás variable en esta especie.
Otras variables que han resultado estar correlacionadas con la
estacionalidad del desfronde en las especies arbustivas son el viento y/ó la
humedad ambiental. En el caso del viento resulta muy poco comprensible que
éste pudiera ejercer alguna influencia en los desfrondes estivales fenológicos
de A. decortícans y C. laurifolius aunque sí puede ser posible que participen
en las caídas accidentales del invierno o de otros momentos del año. En todo
caso parece que la correlación entre desfronde y viento para todo el conjunto
del periodo muestreado sea una pura coincidencia ya que el viento está
relacionado estrecha e inversamente con la temperatura (r= -0.77, p<0.001,
figura 3.22). Además, la correlación con el desfronde pasa a no ser
significativa cuando se calcula la correlación parcial con el viento
eliminando el efecto de la temperatura (A. decortícans total, r=-0.25, C.
laurifolius hojas y total, r=-0.48). Se puede concluir que a pesar de poder
ejercer un efecto local en las caídas de algunos meses no existe ningún tipo
de interacción directa del viento sobre el ciclo fenológico habitual de las
cuatro especies.
153
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
a, g/m2.día
b. TDDFHCL TMNOCT
7 10 1 7 10 2 8 11
TDDFHAD TMAX
Figura 3.21. A. Variación estacional de la caída de hojas en C. laurifolíus y
la temperatura nocturna durante el período de estudio. B. Variación estacional
de la caída de hojas en A. decorticans y la temperatura máxima durante el
período de estudio.
154
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
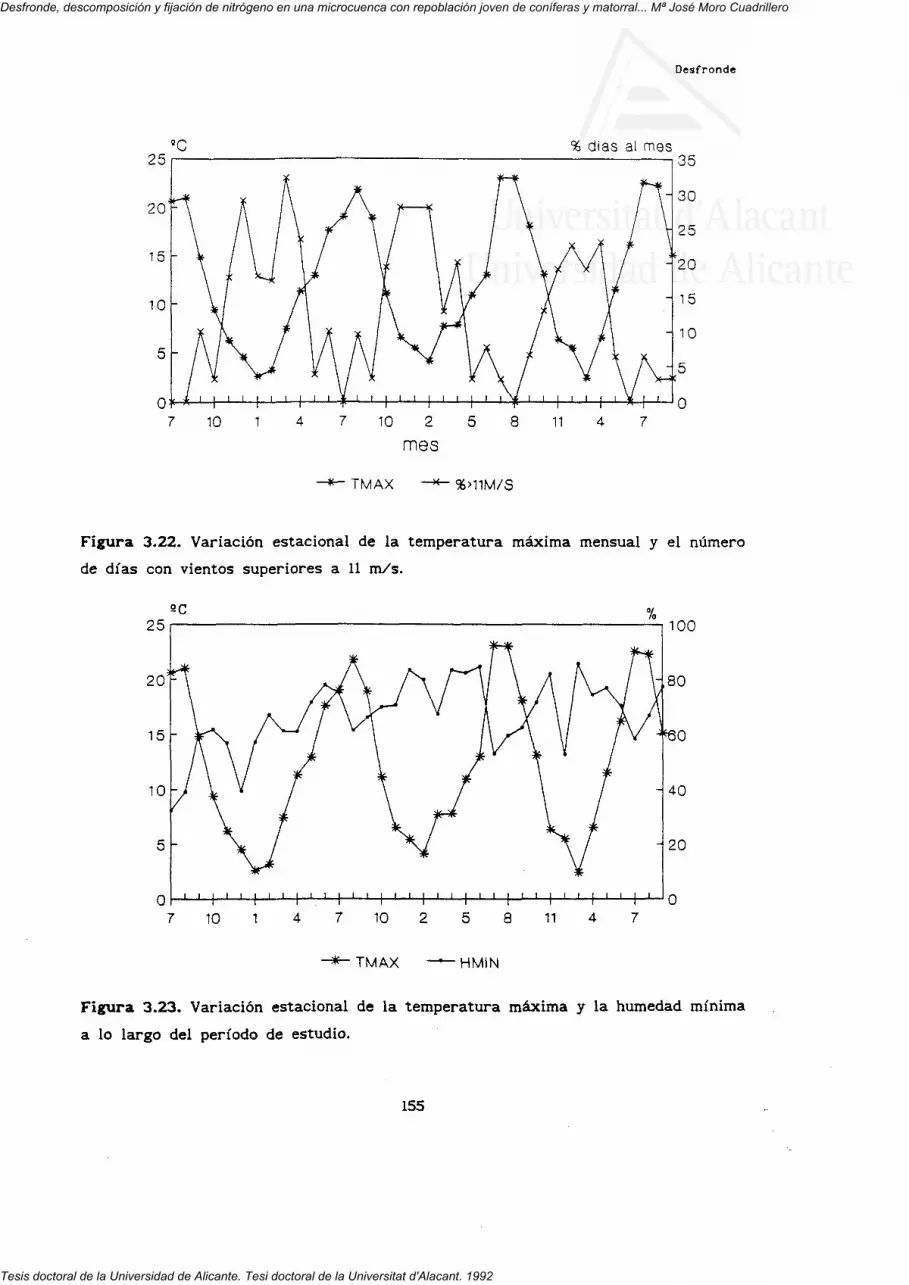
Desfronde
% dias al mes
7 10 1
TMAX *>11M/S
Figura 3.22. Variación estacional de la temperatura máxima mensual y el número
de días con vientos superiores a 11 m/s.
25 %
i i i ' J | L_J |—I—I | I I |_J l _ | I i | i ' | L-L.
100
7 10 1 4 7 10 2 5 8 11 4 7
TMAX HMIN
Figura 3.23. Variación estacional de la temperatura máxima y la humedad mínima
a lo largo del período de estudio.
155
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La humedad máxima y mínima junto con la temperatura son los dos factores
climáticos mejor correlacionados con el ritmo de desfronde en A. decorticans
(Humedad mínima-desfronde foliar, r= -0.57 p<0.001; Humedad máxima-defronde
total r= -0.47 p<0.01). Realizando la misma aproximación comentada en el
párrafo anterior, es decir eliminando el efecto de la temperatura, la
correlación desfronde-humedad sigue siendo significativa tanto para las hoja
(r= -0.39, p<0.05) como para el total (r=-0.42 p<0.05). En consecuencia la
humedad máxima con respecto al desfronde total y la humedad mínima respecto al
desfronde foliar explican un porcentaje de varianza no explicado por la
temperatura. Esto resulta lógico a la vista de las figura 3.23. Puede
apreciarse como ia humedad alcanza valores mínimos no solo en los meses
estivales coincidiendo con las temperaturas más altas, sino en determinados
meses invernales. De hecho, humedad y temperatura no están significativamente
correlacionadas (r= -0.30, ns). La figura 3.24 muestra que los descensos
invernales en el porcentaje de humedad coinciden con los meses invernales más
secos en los que no se registraron lluvias. El mínimo absoluto de humedad
invernal se registra en Diciembre del 86 (407.) con un porcentaje incluso más
bajo que el de los meses estivales (a excepción de Julio de 1986). Al mes
siguiente (Enero de 87) se observó un pico de caída de hojas en A. decorticans
(figura 3.25) que no se repite en años sucesivos. Es posible que los factores
antes mencionados generaran un cierto estrés hídrico invernal durante ese
primer año que pudiera haber provocado o influido en la importante caída de
hojarasca detectada entre Diciembre 86 y Enero del 87. En los inviernos
sucesivos el estrés no parece ser acusado y, según esta hipótesis no sería
suficiente para favorecer una posible caída de hojarasca.
Las correlación entre el desfronde mensual de P.nigra con respecto a la
temperatura media máxima es negativa y por tanto, opuesta con respecto a la
encontrada en las especies arbustivas (tabla 3.13). La caída fenológica en
esta especie como ya dijimos es claramente otoñal y va seguida de caídas
fortuitas en invierno de acículas ya desprendidas. La relación con la
temperatura parece por tanto bastante casual ya que en otoño y en invierno las
temperaturas son más bajas que en primavera-verano, cuando esta especie no
pierde apenas acículas (figura 3.26). A pesar de ello, sí que parece quedar
claro a la vista de estos resultados que el estrés hídrico relacionado con las
156
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
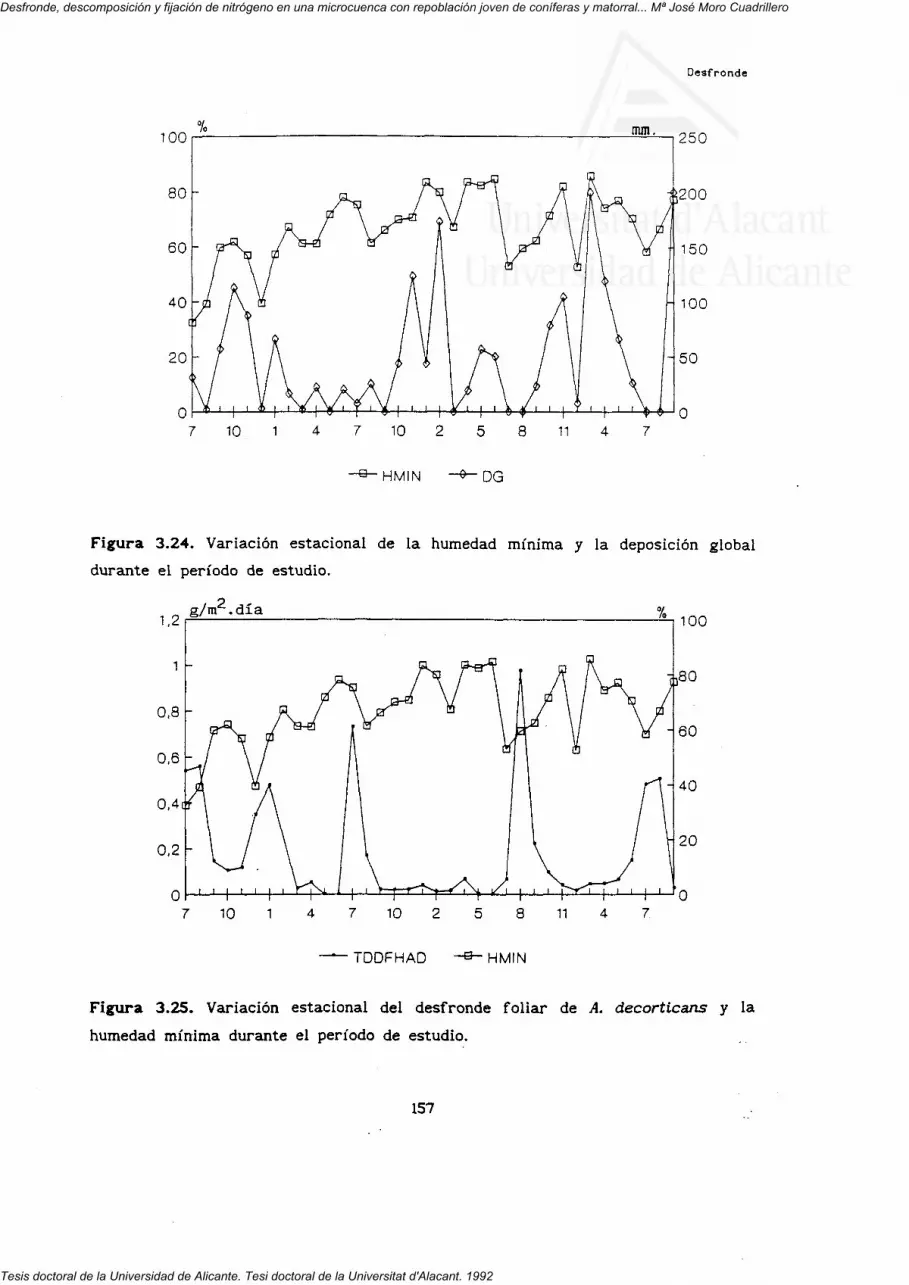
Desfronde
100 250
200
150
100
7 10 1 4 7 10 2 5 8 11 4 7
HMIN DG
Figura 3.24. Variación estacional de la humedad mínima y la deposición global
durante el período de estudio.
g/m . d í a 100
7 10 1 4 7 10 2 5 8 11 4 7
TDDFHAD HMIN
Figura 3.25. Variación estacional del desfronde foliar de A. decorticans y la
humedad mínima durante el período de estudio.
157
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
gr/m2.día °C 1,2
1
0,8
0,6
0,4
0,2
0 7 10 1 4 7 10 2 5 8 11 4 7
mes
TDDFHPN - * - T M A X
Figura 3.26. Variación estacional del desfronde foliar de P. nigra y la media
mensual de la temperatura máxima durante el período de estudio.
elevadas temperaturas estivales no es el causante de la abcisión fenológica en
esta especie. Al igual que ocurría con las especies de matorral, eliminando el
efecto de la temperatura desaparece la relación del desfronde mensual con el
viento aunque probablemente la mayor incidencia en invierno de vientos fuertes
sí pudiera favorecer el caída de las acículas desprendidas enganchadas en las
ramas.
No existe relación entre precipitación mensual y desfronde en ninguna de
las cuatro especies. Tampoco se ha encontrado relación entre la precipitación
total anual y el desfronde anual aunque hay que tener en cuenta el bajo número
de años de seguimiento en este trabajo (3 años). Algunos autores como Merino
et ai.(1990) sí han encontrado, una correlación positiva y significativa entre
estos parámetros en sistemas arbustivos de Doñana pero durante varios años con
precipitaciones anuales muy diferentes. Sería necesario disponer de series
158
¿ u
' ' i ' i i, i i i T ' - i . - i ' i i_ _i—i "f-r-i-j ,i i__i i i ["J'T-f i
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
temporales más largas para valorar la importancia que puede ejercer este
parámetro sobre la fenología del desfronde sobre todo en años con
precipitaciones escasas.
3.3.5. COMPOSICIÓN QUÍMICA DEL DESFRONDE FOLIAR.
En la tabla 3.14 se resumen los resultados de la composición química
elemental de la hojarasca. Los valores son medias ponderadas teniendo en
cuenta el peso de defronde cada mes. Los resultados están calculados para los
dos años en que se ha analizado la composición química. En el apéndice 3.5 se
muestra también la composición química ponderada para cada año por separado.
El orden que ocupan los elementos en cada fracción y especie según su
concentración química pueden visualizarse en la tabla 3.15.
En general N y Ca son los elementos con concentraciones más elevadas en
todas las fracciones exceptuando las flores del matorral y las pinas en P.
pinaster dónde el K sustituye al Ca. Los últimos lugares lo ocupan el P y el
Na. Exceptuando a las semillas cuyo contenido en todos los nutrientes es más
elevado que en cualquier otra fracción, existen considerables variaciones en
la concentración dependiendo la fracción que se considere (tabla 3.14). 4.
decorttcans, por su condición de fijadora, presenta los valores más elevados
de N en todas las fracciones incluso las ramillas-tallos (1.57%). En general,
todas las especies presentan las mayores concentraciones de N y P en flores.
El K es asimismo rico en las flores de las especies arbustivas (0.85% en A.
decorttcans y 0.71% en C. laurifolius) y en las pinas de P. pinaster (0.15%).
Las hojas son más ricas en Ca y Mg exceptuando el caso del Mg en acículas de
P. nigra. Características químicas similares se han descrito en otras
coniferas del área mediterránea (Verdú 1984, Sta Regina et al. 1989, Rapp et
al. 1979) y en sistemas de matorral mediterráneo (Núñez 1989).
159
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M,J. Moro Cuadrillero
Tabla 3.14. Concentraciones medias ponderadas (% de peso seco) para las dis
tintas fracciones de las cuatro especies (medias de dos años).
NITRÓGENO
P.pinaster
P.nigra
A.decorticans
C. laurifolius
FOSFORO
P.pinaster
P.niqra
A.decorticans
C.laurifolius
POTASIO
P.pinaster
P.niqra
A.decorticans
C.laurifolius
HOJA
.36
.44
2.12
.47
HOJA
.04
.04
.09
.10
HOJA
.09
.08
.59
.21
FLOR
.55
.71
2.48
.30
FLOR
.05
.03
.12
.12
FLOR
.07
.09
.85
.71 '
FRUTO
.67
-
.72
.58
FRUTO
.04
-
.01
.04
FRUTO
.15
-
.50
.07
SEMILLAS
2.87
2.87
5.46
-
SEMILLAS
.35
.34
.35
-
SEMILLAS
.45
.45
1.10
-
RAMAS
.38
.78
1.57
.68
RAMAS
.03
.05
.06
.05
RAMAS
.15
.10
.08
.07
160
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.14. (continuación)
CALCIO
P.Dinaster
P.niqra
A.decorticans
C.laurifolius
MAGNESIO
P.Dinaster
P.niqra
A.decorticans
C.laurifolius
SODIO
P.Dinaster
P.niqra
A.decorticans
C. laurifolius
HOJA
1.13
.85
1.00
1.68
HOJA
.20
.16
.22
.27
HOJA
.02
.01
.01
.01
FLOR
.19
.24
.76
.16
FLOR
.09
.19
.17
.15
FLOR
.01
.03
.02
.01
FRUTO
.08
-
.22
.23
FRUTO
.10
-
.08
.05
FRUTO
.02
-
.02
.01
SEMILLAS
.65
-
-
-
SEMILLAS
.22
.22
.13
-
SEMILLAS
.02
.02
.01
-
RAMAS
1.25
.19
.11
1.57
RAMAS
.12
.11
.09
.08
RAMAS
.01
.01
.05
.01
161
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.15. Orden que ocupan los elementos según su concentración química ponderada en las diferentes fracciones del desfronde.
HOJAS P. pinaster Ca > N > Mg > K > P > Na
P. nigra Ca > N > Mg > K > P > Na
A. decorticans N > Ca > K > Mg > P > Na
C. laurifolius Ca > N > Mg - K > P > Na
FLOR P. pinaster N > Ca > Mg > K > P> Na
P. nigra N > Ca > Mg > K > P > Na
A. decorticans N > » K > Ca > Mg > P > Na
C. laurifolius N > K > Ca > Mg > P > Na
FRUTO P. pinaster N > K > Mg - Ca > P > Na
A. decorticans N > K > Ca > Mg > Na > P
C. laurifolius N > Ca > K > P > Mg > Na
RAMAS P. pinaster Ca > N > K > Mg > P > Na
P. nigra N > Ca > Mg > K > P > Na
A. decorticans N » > Ca > Mg - K > P > Na
C. laurifolius Ca > N > Mg > K > P > Na
3.3.5.1. Variabilidad temporal en la composición química del desfronde
foliar. Mecanismos asociados.
Debido a que la fracción hojas constituye el grueso del desfronde total,
hemos analizado con más detalle la composición química de esta fracción y su
variabilidad temporal así como algunos de los posibles mecanismos asociados a
ésta (traslocación y lixiviado).
162
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
1.- Comparación estadíst ica de la composición química de la hojarasca en t re
las dis t intas especies.
Para poder realizar comparaciones estadísticas y estimar la variabilidad
en la concentraciones de cada elemento en las distintas especies hemos
utilizado las medias aritméticas (tabla 3.16) y no las ponderadas (tabla 3.14)
debido a la dificultades que conlleva un análisis estadístico con las valores
ponderados. Comparando estas dos tablas se observa que las diferencias entre
ambos métodos son inferiores al 2.07» exceptuando el N en la ja ra y el Na en P.
nigra. Por tanto la utilización de las medias aritméticas puede considerarse
aceptable exceptuando los casos anteriormente citados.
N y Ca son los elementos con concentraciones más altas en todas las
especies. En general, las concentraciones de la mayoría de los elementos son
superiores en las especies arbustivas. Aplicando un ANOVA se constata que para
todos los nutrientes existen diferencias significativas respecto a las
concentraciones medias entre las especies estudiadas (p<0.001 en todos los
casos). Los resultados de la aplicación posterior del test de Newman- Keul's
se muestra en la tabla 3.16. Los valores de cada columna seguidos de la misma
letra no son significativamente distintos (p<0.05). A. decorticans posee
concentraciones significativamente superiores de N y K (2.49% y 0.47%) a las
demás especies, la j a ra de Ca (1.34%) y P. pinaster de Na (0.024%). Por su
parte P. nigra presenta contenidos significativamente más bajos de Mg (0.16%)
que el resto de especies. Las dos especies de pino no presentan diferencias
respecto a la concentración de P (0.039% y 0.042%) y K (0.10% y 0.095%)
resultando éstas significativamente menores a las de matorral. Además, las
especies arbustivas tienen contenidos similares de Mg (0.21% y 0.22%) y P
(0.107% y 0.089%). Estos resultados son ligeramente diferentes a los
encontrados, aplicando el mismo test, para la hojarasca de estas cuatro
especies en la época de máxima abcisión fenológica utilizada para el
seguimiento de la descomposición.(capítulo 5). Los cambios más acusados entre
hojarasca senescente y la del conjunto del año se producen en el N, P y Ca.
El elemento más variable en todas las especies exceptuando P. nigra es el
P (CV entre 40-46%) seguido del Na (30-63%). Para este último los errores
163
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
analíticos causados por sus bajas concentraciones pueden tener cierta
importancia. La jara es la especie que presenta mayor variabilidad mensual en
la química de la hojarasca sobre todo en lo que se refiere a los elementos más
móviles y limitantes :N (457.), P (407.) y K (507.). En las demás especies el
coeficiente de variación oscila entre el 21-25% para el N y 36-397. para el K.
La especie que presenta menores fluctuaciones en su composición química es P.
nigra.
Otros autores han destacado la variabilidad intermensual en las
concentraciones químicas de la hojarasca (Rapp 1967, Sharma y Pande 1989,
Escudero et al. 1983, Verdú 1984, Gosz et al. 1972, Dannaoui 1981). Esta
variación a veces va acompañada de un cierto patrón temporal sobre todo en el
caso del N (Sharma y Pande 1989, Escudero et al. 1983, Gosz et al. 1972, Gray
1983, Verdú 1984) y P (Rapp 1967, Sharma y Pande 1989, Escudero et al. 1983,
Gray 1983, Verdú 1984) y menos frecuentemente en el caso del Ca (Escudero et
al. 1983 en Quercus pyrenaica) y K (Rapp 1967, Sharma y Pande 1989).
En el siguiente apartado se abordará la variación temporal de cada
elemento en los dos primeros años de muestreo en orden a establecer posibles
pautas temporales
2 . - Pautas temporales en la concentración química de la hojarasca.
Nitrógeno.
En los pinos la concentración de N es elevada durante el invierno y la
primavera (figura 3.27a y b). En verano del segundo año, coincidiendo con la
máxima caída de hojas, éstas descienden notablemente en P. pinaster. Durante
el primer año también existe un descenso hacia finales de verano. En P. nigra
el descenso de los niveles de N coincide con los meses de caída fenológica de
hojas (Septiembre-Octubre del primer año y Octubre del segundo). En A.
decorticans, el comportamiento descendente de la concentración de este
elemento es acusado en la época estival coincidiendo con la masiva caída de
hojarasca mientras que el resto del año los niveles de N son más elevados
(figura 3.27d). Un patrón parecido se observa en C. laurifolius (figura
164
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
C W 4) (d
>í
u r-OJ
^ c-oj
60
< t—1 OJ
O O OJ
<3
p~ 00
o' 2
I u n ID oí
OJ
o -
00
C\ co o
co <¿ oí ~-
«* n «•H
CO
oi 01
m
OJ -
co
01
U 00 n
^ co r> >o
u o OJ
OJ
o
o ~ *—i
O
a
co
OJ OJ
o ~
o o
o co n
o O* 00
o 3
0> «* OJ
01
VO 01
u 1—1
a. c/i u
I X
•o
> u
m
o ^ o ^
00 oi O -
O in 0^ o
n 1/1
o! (0
O —
CQ
CQ
00 00
o
OÍ
ui •»
I X > u
u
o
o 2
> u
n *# o o
, OJ
có 01
vO •sf
* O
co »-l 01
I X
a cu
> u
c cu
165
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
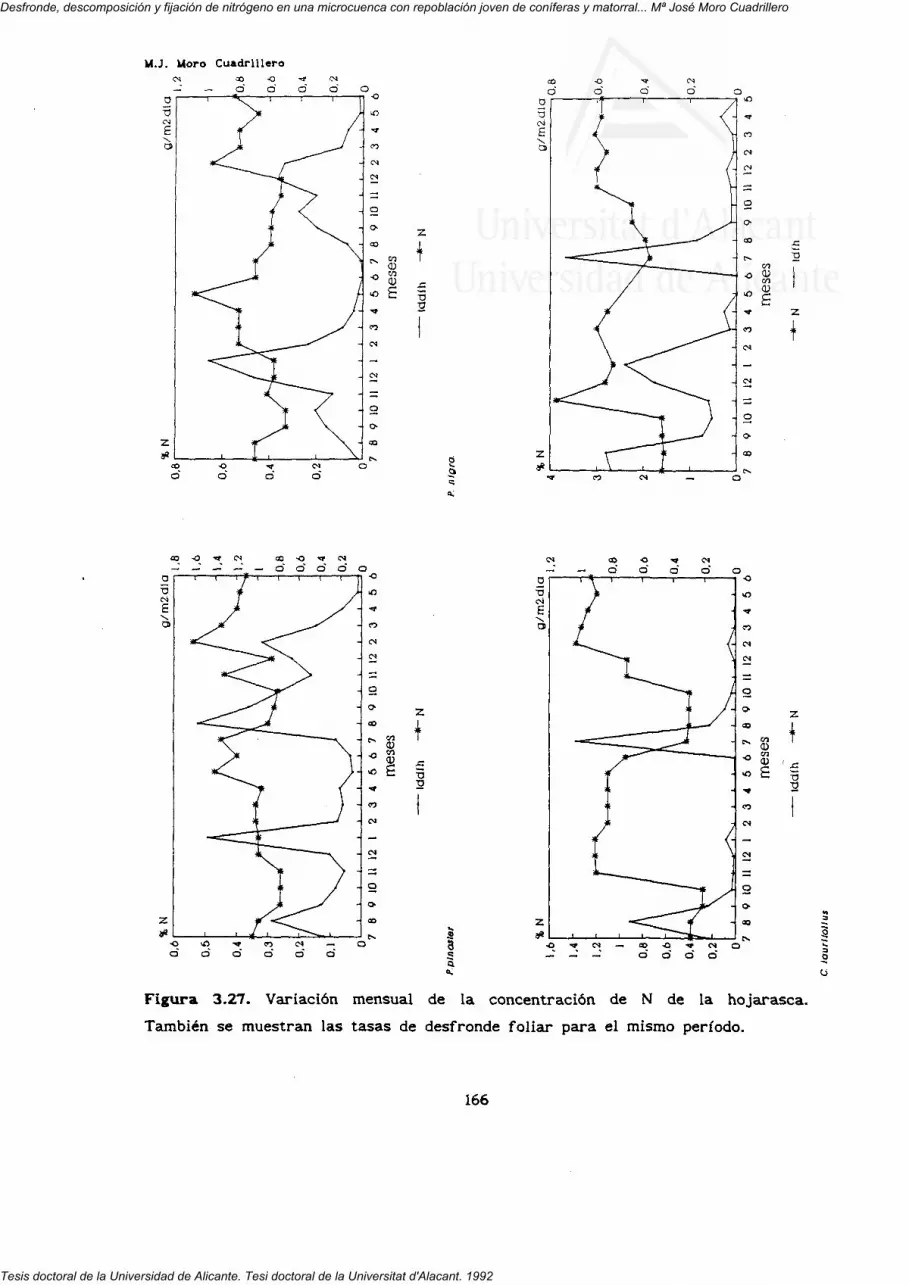
M.J. Moro Cuadrillero
o o
-O
£
Z
t
13
1
-o <D
s
o
•* <r>
Oi
<n 50
s TI
1
a; o
Figura 3.27. Variación mensual de la concentración de N de la hojarasca.
También se muestran las tasas de desfronde foliar para el mismo período.
166
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
3.27c). En el matorral, estos cambios estacionales resultan mucho más acusados
que en las especies de pino.
Fósforo.
La concentración de P se mantiene baja en el periodo otoñal de caída de
acículas de P. nigra, ascendiendo en los meses siguientes (figura 3.28b). En
Agosto de 1988 la concentración vuelve a descender manteniéndose en esos
niveles durante la caída otoñal de hojas para volver a aumentar
posteriormente. En la jara , por el contrario no se han observado disminuciones
de contenido en P durante el mes de máxima abcisión sino que lo hacen un mes
más tarde cuando ya la tasa de caída de hojas senescentes es mucho más pequeña
(figura 3.28c). Los niveles de P aumentan en invierno manteniéndose así hasta
finales del verano siguiente en que se repite el patrón del año anterior. Para
A. decorticans los cambios estacionales respecto al P resultan muy acusados
detectándose un patrón en el que predominan concentraciones bajas en la época
de abcisión estival y altas el resto del año.(figura 3.28d) En P. pinaster los
niveles de P tienden a ser más bajos en invierno pero no parece existir un
patrón temporal definido, (figura 3.28a).
Potasio.
Este elemento presenta un patrón muy irregular en todas las especies. En
P. pinaster y P. nigra las concentraciones de este elemento son más bajas en
invierno manteniéndose más altas durante el resto del año (figura 3.29a y b).
Los cambios son más acusados en P. nigra. En esta especie el descenso en la
concentración comienza ya a finales de verano. En C. laurifolius las
concentraciones tienden a ser más bajas en verano-otoño(primer año) y otoño
(segundo año) (figura 3.29c) aumentando de forma considerable en invierno y
primavera. Las oscilaciones que sufre este elemento en A. decorticans parecen
menos acusadas que en la j a r a pero parece existir la tendencia a que los
valores más bajos se produzcan en otoño durante los dos años de estudio
(figura 3.29d).
Calcio.
El Ca parece comportarse de forma opuesta al N y P por lo menos en las
especies arbustivas (figura 3.30c y d). En el matorral predominan
167
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
o o-
•o
iO
•*T
co es
0)
fci
a.
t .c •o n
I
S 5 I
O
Ü
Figura 3.28. Variación mensual de la concentración de P de la hojarasca.
También se muestran las tasas de desfronde foliar para el mismo período.
168
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
0) 0)
s 1
I
s
o
o
•O
U5
0 1
a
«
t .C
T3 •o
. g o-
1
3
Figura 3.29. Variación mensual de la concentración de K de la hojarasca.
También se muestran las tasas de desfrondé foliar para el mismo período.
169
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
concentraciones altas en la época de abcisión fenológica contrariamente a lo
que ocurría con N y P. Estos niveles se mantienen uno o dos meses después para
descender posteriormente. Los pinos no presentan pulsos estacionales marcados
sino que, por el contrario, los niveles de Ca son más o menos constantes a lo
largo de todo el año sufriendo solamente pequeñas oscilaciones (figura 3.30a y
b). De hecho los coeficientes de variación para el Ca y el Mg en los pinos son
más bajos que para cualquier otro elemento en las cuatro especies estudiadas
(tabla 3.16)
Magnesio.
Los cambios en la concentración de Mg son muy poco importantes en el
pinar lo que implica que no pueden definirse patrones estacionales (figura
3.32a y b). Únicamente puede destacarse el aumento durante el periodo de caída
de hojarasca estival en P. pinaster durante el segundo año y la subida en
primavera para este mismo año en P. nigra. Tampoco existe ninguna pauta clara
en A. decorticans (figura 3.31d) mientras que en C. laurifolius se aprecian
mayores niveles de Mg durante la caída estival en los dos periodos (figura
3.31c). El resto del año la concentración de Mg se mantiene mucho más baja con
pequeños pulsos que como veremos se relacionan con la cantidad de hojas
caídas.
Sodio.
En P. nigra las concentraciones más elevadas de este elemento se
registran durante los picos de máxima caída, tanto otoñales como invernales
(figura 3.32b) mientras que las más altas aparecen en primavera cuando la
caída de acículas es mínima. En las otras especies el Na presenta un un
comportamiento muy irregular y aleatorio (figura 3.32a, c y d).
3 . - Relación composición química-peso de hojarasca.
La concentración de algunos nutrientes en el desfronde foliar puede
relacionarse de manera significativa con la magnitud de ésta (tabla 3.17).
Para el conjunto del periodo C. laurifolius presenta una relación negativa y
altamente significativa (p<0.001) respecto al N y positiva respecto al Mg y Ca
170
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
a o
I
i
•o
-r s
I 0
u
o
1
1
o
1
o
Figura 3.30. Variación mensual de la concentración de Ca de la hojarasca.
También se muestran las tasas de desfrondé foliar para el mismo período.
171
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
0)
£
a 2
1
o
0)
S o 2
a 2
I
£1 o.
O 2
I
o u
Figura 3.31. Variación mensual de la concentración de Mg de la hojarasca.
También se muestran las tasas de desfronde foliar para el mismo período.
172
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
o z
3 a z
I <o <N O O
« O o
o —• o o
_ o o o o o o
o o-
•o
f>
•«1»
eo
CM
CQ
S
0
z + TÍ T3
1 1
- 2
§
en
£
0< iO — lO
« o — o o °. o" R o o o o o
o z
s
o
Figura 3.32. Variación mensual de la concentración de Na de la hojarasca.
También se muestran las tasas de desfronde foliar para el mismo período.
173
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
(p<0.001). Para A. decorticans sólo se han encontrado relaciones negativas y
significativas para el N (p<0.05) y para P. nigra respecto al P (p<0.05).
Estas relaciones sugieren la existencia de mecanismos de retraslocación para
el P y N en estas especies.
Considerando cada año de muestreo por separado, el número de
correlaciones significativas aumenta aunque en muchos casos con un nivel de
significación más bajo debido al menor número de datos (tabla 3.17). La
relación concentración de N y peso de hojarasca es significativa y negativa
para los dos años en C. laurifolíus y solamente para el segundo año en A.
decorticans. La baja correlación que existía entre nivel de N-caída de hojas
en A. decorticans para el conjunto del periodo (p<0.05) probablemente se deba
a la no significación de esta relación durante el primer año. Para esta
especie existe también relación entre el contenido en P y peso de hojarasca
durante el segundo año (p<0.05). Para aquellos elementos cuya concentración
tiende aumentar con el desarrollo y senescencia de la hoja como el Ca y Mg
(Waring y Schlesinger 1985), la j a ra presenta relaciones positivas y altamente
significativas (p<0.001) durante el primer año para ambos nutrientes y para el
Mg el segundo año (p<0.01). También A. decorticans presenta esta pauta con
respecto al Ca en el segundo año (p<0.05) y P. pinaster para el Mg el primer
año (p<0.05).
Otros autores han encontrado también relaciones negativas entre la tasa
de desprendimiento foliar y las concentraciones de N y P sobre todo en
aquellos estudios en que la caída accidental tiene una escasa importancia
cuantitativa. Gray (1983) calcula relaciones inversas entre concentración de N
y peso de hojarasca en especies de Ceanothus del chaparral californiano y
Sharma y Pande (1989) para el N y P en acículas de Pinus rosburghii en la
India. Estos resultados son indicativos, según estos autores, de una efectiva
traslocación. Sharma y Pande (1989) han encontrado también este tipo de
relaciones negativas respecto al K y Mg lo cual no se ha detectado en el
presente trabajo.
174
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
O
z
u
2 £
— o
• 10
i*
£3
CL
00
en
en
en z
a ai
w
en
?Í3
a ai
en
en
en
en
M
a ai
en
en
en
L 3 N 6
ii
en
H e ai
c i ai
en
en
en
en
en
c ai
5 3 o +
I II
">< 5
2-5 a
2 s o S
6 ii
en
en
X
p O
?r H U II
"x
• * ~ 5
? 5
i-3
oo °
u «o o
en
en z
d i II
u
X
3 «i o A a 1
»
m~5
c— •
V
en
en
en
- 5
o r* ' • o ? 7 ii
en
en z
en
en
en
CM
3 s o o I
o «
3
•<
en
en
•a
en
en
X
- 3 i.<-
a i
vO t - » o i
I II II
00
3 SO d
vO I
U II
•<
175
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
3.3.5.2.- Discusión. Mecanismos asociados a la variabilidad temporal .
A la vista de los resultados se observa que, en general, las
fluctuaciones mensuales de N, P y K son superiores a las de Ca y Mg en todas
las especies. Esta variaciones pueden estar asociadas con mecanismos de
traslocación antes de la caída de la hoja que, ademas, parecen estar muy
desarrolladas en especies perennes mediterráneas (Read y Mitchell 1983). Las
relaciones significativas entre el N y P con respecto a la caída de hojarasca
encontradas en algunas especies parecen confirmarlo. Para estos elementos
parecen descartarse los procesos de lixiviado vía trascolación ya que en las
especies estudiadas N y P son elementos minoritarios de este flujo e incluso
en muchas ocasiones son absorbidos a nivel de copas (Domingo 1991). Además,
excepto en P. nigra, la abcisión fenológica se produce en época estival
coincidente con los meses más secos del año. Las variaciones en la
concentración de K pueden estar relacionadas no sólo con la retraslocación
antes de la caída de hojas senescentes sino con el lixiviado, (antes o después
de la caída), sobre todo en los meses de otoño invierno (Gosz et al. 1972,
Sharma y Pande 1989).
Otra fuente de variabilidad adicional (Escudero et al. 1983, Sharma y
Pande 1989) en el pinar se refiere a la coexistencia en los árboles de
acículas de varias edades y por tanto, en distinto grado de desarrollo. Aunque
normalmente la hojarasca se compone mayoritariamente de hojas con un elevado
grado de desarrollo, factores predominantemente externos o bien biológicos
(plagas, fitofagia) pueden determinar la caída prematura y aleatoria de hojas
de edades inferiores. Evidentemente esto puede influenciar la composición
química final de la hojarasca pudiendo provocar fluctuaciones más evidentes
que en otras especies con hojas de ciclo más rápido. En nuestro estudio,
también va a resultar de importancia para la variabilidad química la caída de
acículas, ya escindidas del árbol incluso algún año atrás , que han permanecido
enganchadas a las ramas secas de la base del árbol.
176
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Mecanismos asociados a la variabilidad temporal en cada elemento.
NITRÓGENO.
La diminución en la concentración de N en A. decorticans durante el
periodo de máxima caída fenológica parece un buen indicador de la existencia
de mecanismos de retraslocación en A. decorticans. El nivel de N en hojas
durante el mes de Agosto oscila entre el 1.5-1.77. mientras que las
concentraciones máximas encontradas en otros momentos del año y asociadas a
caídas de hojas verdes están alrededor del 37.. Estos resultados se ven
confirmados por la existencia de una correlación significativa y negativa
entre la concentración de N y la tasa mensual de caída de hojarasca durante el
segundo año. Durante el primer año, la cocentración de N también disminuye en
verano pero no existe correlación significativa entre el N y peso de
hojarasca. La explicación más evidente es la existencia de un pulso importante
de caída de hojas verdes con niveles de N elevados en el invierno 86-87. Gosz
et a¿.(1972) han puntualizado la importancia que estas caídas accidentales
tienen respecto al retorno de nutrientes al suelo sobre todo en lo que se
refiere a N, P y K. Esta abcisión prematura retorna una buena cantidad de
elementos móviles antes que actúen los procesos naturales de retraslocación.
La existencia de retraslocación durante la abcisión fenológica parece
tener menos importancia en P. pinaster y P. nigra. La disminución en la
concentración de nitrógeno en acículas escindidas de P. nigra se manifiesta
durante el otoño, época en que tiene lugar la abcisión fenológica, pero este
descenso es muy poco acusado (figura 3.27b y apéndice 3.5). En P. pinaster
sólo existe una disminución clara durante el verano del segundo año. La no
existencia de correlaciones significativas se debe en gran medida a las mismas
razones comentadas para A. decorticans pero en el pinar tienen un mayor
sentido ya que en estas especies los pulsos accidentales de invierno son
intensos (incluso mayores que los de caída natural) y se repiten en los dos
años de muestreo. Durante el primer invierno, la concentración de N en
acículas sólo es ligeramente más alta que la de las caídas durante el
desfronde fenológico. Por el contrario durante el segundo invierno las
diferencias son mucho más acusadas. Esta característica podría indicar que el
pulso invernal del primer año correspondería básicamente a parte del almacén
177
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
acumulado en ramas secas con menor contribución de material verde. Durante el
segundo periodo de muestreo el material verde parece el dominante en ambas
especies. A pesar de ello, para la mayor parte del material foliar recogido en
las trampas se observan concentraciones de N inferiores a acículas verdes de
t res años (0.6% de N)lo que indicaría que son predominantemente acículas bien
senescentes, bien próximas a la senescencia las que caen al suelo en las que
ya han actuado previamente los mecanismos de reabsorción.
La caída de hojarasca en C. lawifolius está, a la vista de los resultados,
controlada más eficazmente por la propia planta a la manera descrita por
Escudero et al. (1983). En esta especie la incidencia de la abcisión prematura
y accidental es poco frecuente y poco importante mientras que en verano la
máxima caída fenológica coincide con una clara reabsorción de N (figura
3.27c). Se podría concluir que la j a ra posee un mecanismo eficaz de control
del N y que ésto puede reflejarse en la relación significativa para los dos
años entre concentración de N y magnitud de desfronde foliar (tabla 3.17).
Además la concentración media en hojas vivas de C. laurifolíus es del 1.37% lo
que implica una disminución del 69% del N en la época de máxima abcisión
foliar.
Otros autores han destacado también la existencia de una patente
disminución en la concentración de N en el momento de caída de hojas
senescentes. En matorrales, Núñez (1989) encuentra esta pauta en Cistus
ladanifer, Gray (1983) en Ceanothus y Schwintzer (1984) en la fijadora Myrica
gale aunque en este último caso la disminución de concentración no es
excesivamente elevado. En especies arbóreas perennes Escudero et al. (1983)
observan estos patrones en encina, Verdú (1984) en abeto.
FOSFORO.
El P junto con el N constituye uno de los elementos muchas veces
limitantes en ecosistemas mediterráneos por lo que son esperables mecanismos
de traslocación desarrollados para reutilizarlos de forma más eficaz (Read y
Mitchell 1983). En este sentido, la disminución de concentración en la época
de la abcisión puede actuar como indicador de este proceso (Escudero et al.
1983).
178
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Deifronde
En el presente estudio el descenso en el nivel de P foliar sólo se
manifiesta de una forma clara para los meses de caída estival en hojas del
gaznacho. La concentración en este período es de un 0.04% mientras que la
concentración en la biomasa de hojas es de un 0.17. lo que implicaría un
reabsorción del 60% del P en esta época del año. La existencia de una relación
significativa inversa entre contenido en P y la magnitud del desfronde foliar
(tabla 3.17) también sugiere la existencia de reabsorción durante el segundo
año. El primer año, por la razones ya comentadas para el N no se encontraron
relaciones significativas.
En especies arbustivas mediterráneas Núñez (1989) en Cistus ladanifer y
Gray (1983) en Ceanothus observan también disminuciones claras en la época de
máxima abcisión. Este patrón se ha observado también en la hojarasca de
diferentes especies arbóreas perennes de la zona mediterránea (Escudero et al.
1983, Verdú 1984)
La reabsorción de P parece evidente en otoño de 1987 en acículas de P.
nigra. Durante el primer año las concentraciones más bajas también se
registran en otoño pero éstas no difieren demasiado con los valores
encontrados el resto del año. (Fig 3.28b). No obstante existe una relación
significativa, aunque débil entre P y peso de hojarasca para el conjunto de
periodo muestreado (p<0.05) (tabla 3.17). Al igual que ocurría con el N,
durante casi todo el año los niveles de P son inferiores a los de acículas
vivas de más de t res años (alrededor del 0.08%) lo que indica que son acículas
con un elevado grado de desarrollo y senescencia las que caen buena parte del
año en las que ya se han producido mecanismos de reabsorción.
No parecen existir mecanismos internos de reabsorción en hojas de j a ra
antes de la abcisión. La figura 3.28c y apéndice 3.5 muestran elevadas
concentraciones de P (0.098%-0.ll2%) en el periodo de máxima caída muy
similares a los registrados en primavera e invierno cuando caen
predominantemente hojas verdes. De hecho la concentración media en la biomasa
foliar de esta especie (cap.2) es del 0.15% no muy diferente a los niveles
encontrados en la hojarasca estival. En el capitulo 5 se constata que también
179
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
al verano siguiente (Agosto de 1988) el nivel de fósforo en hojas escindidas
presentaba concentraciones elevadas (0.142) muy similares a las encontradas en
hojas vivas. También se comenta en ese capítulo la influencia que tiene para
la dinámica de descomposición la no existencia de reabsorción de P previa a la
caída natural en esta especie. Por su parte, se han detectado descensos en los
niveles de P durante la estación otoñal en ambos periodos de muestreo (figura
3.28c). Se ha constatado que las lluvias otoñales lixivian porcentajes
considerables de P en hojas de esta especie en la misma cuenca (Domingo 1991).
Este autor calcula para el fosfato unos coeficientes de enriquecimiento medios
en el agua que transcola por la j a r a de 5.43 lo que indica una clara cesión de
este elmento por parte de C. laurifolius. Es por tanto muy probable que el
empobrecimiento en P de la hojarasca caída en el periodo otoñal esté causada
por un lixiviado previo de éste elemento por el agua de lluvia de forma
similar a lo encontrado en Quercus pyrenaica por Escudero et a¿.(1983).
En P. pinaster tampoco se observan disminuciones en la concentración de P
en la época estival. El primer verano los niveles son similares a los
encontrados en acículas vivas de 3 años (0.07%) mientras que el segundo verano
las concentraciones sí son inferiores (0.0487.) lo que sí que indica un clara
reabsorción. Existe una fuerte disminución en los niveles de P en
otoño-inverno. Este descenso no puede explicarse por un lavado de acículas
antes de caer ya que el fosfato contenido en el agua de lluvia es retenido
cuando atraviesa las copas de esta especie (Domingo 1991, Domingo et al.
1991). En este caso la disminución de P podría achacarse a que el grueso de
hojarasca en esta época está compuesto de las acículas viejas escindidas que
permanecen enganchadas en las ramas bajas del árbol y que posiblemente sean
pobres en P no sólo por su grado de senescencia sino por el lavado al que
pueden haber estado sometidas. En este sentido la concentración de Ca para el
otoño-invierno es ligeramente más alta que en otra época del año, lo que
podría confirmar la existencia de este pulso importante de acículas viejas ya
escindidas. En esta especie la caída accidental alcanza una mayor magnitud
provocando escisión prematura de acículas de diferentes edades. Esta
característica afecta, probablemente, de forma más determinante la
variabilidad química de este elemento que los propios mecanismos de lixiviado
y traslocación. De hecho, esta especie presenta para el P un coeficiente de
180
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

variación muy elevado (44%).
Detfronde
POTASIO.
El K constituye un elemento extremadamente móvil cuyo contenido en hojas
varía sustancialmente dependiendo del estado fisiológico de éstas (Waring y
Schlesinger 1985). Debido a ello, puede ser fácilmente translocado desde las
hojas viejas antes de que caigan (Gosz et al. 1972). Además, su lixiviado por
el agua de lluvia (antes o después de la abcisión) constituye también una
importante fuente de variabilidad temporal en su concentración en la
hojarasca. Las cuatro especies estudiadas presentan fuertes enriquecimientos
de este elemento en el agua de trascolación (Domingo 1991, Domingo et al.
1991). Además la hojarasca pierde rápida y fácilmente K por lavado
(capitulo.5). Por tanto, a la hora de considerar la variabilidad mensual de
este elemento en hojarasca habrán de tenerse en cuenta los dos procesos
(reabsorción, lixiviado). Es evidente que ambos pueden interactuar aunque,
dependiendo del momento del año o de unas condiciones concretas, uno de ellos
domine sobre el otro.
En el matorral (figura 3.29c y d) se observa un descenso en el nivel de K
durante el otoño. En esta época del año están terminando de caer las hojas
senescentes y a su vez, han comenzado las primeras lluvias. Presumiblemente, a
las concentraciones más bajas de K propias de hojas maduras se añada el efecto
de lavado debido a la precipitación con lo que el resultado final es una
patente disminución de este nutriente en estos meses. En invierno se produce
fundamentalmente caída de hojas verdes y la concentración aumenta. En esta
época parece predominar el efecto de la superior concentración de K en estas
hojas sobre el lavado que ret i rar ía parte del K. Durante la época estival no
se observan disminuciones patentes en la concentración de K en C. laurifolius.
Sin embargo los valores alcanzados durante el mes de máxima caída (0.14% y
0.277.) están bastante por debajo del contenido en K en hojas vivas (0.49%)
(capítulo 2). Por tanto, dado que en esta época el lixiviado se reduce al
mínimo, puede concluirse que sí debe existir un cierto grado de movilización
de K a otros órganos de esta especie. En Julio y Agosto del 1987 los niveles
de K en A. decorticans (0.53% y 0.16%) (apéndice 3.5) también se encuentran
por debajo de los encontrados en la biomasa (0.64%). No ocurre lo mismo para
181
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
los mismos meses de 1986 (0.617.). Este elemento parece por tanto reabsorbido
únicamente durante el segundo periodo. En resumen, podemos decir que si bien
parecen existir en algunos casos mecanismos de reutilización de K durante la
época estival, el efecto del lixiviado es también importante retirando K de la
hojarasca otoñal.
En el pinar (figura 3.29a y b), las menores concentraciones se registran
en invierno. Como ya se ha comentado son acículas viejas las que caen
mayoritariamente en los pulsos invernales. Las bajas concentraciones pueden
estar causadas tanto por la menor riqueza en este elemento en las hojas como
por el lixiviado a part ir de lluvias invernales. Las concentraciones
encontradas en los meses de abcisión fenológica en P. pinaster (0.11-0.12%,
apéndice 3.5) resultan ligeramente más bajas que las encontradas para acículas
vivas con un mayor grado de desarrollo (0.20%). Existe, por tanto una
reabsorción pero poco importante. Lo mismo ocurre en P. nigra durante
Septiembre-Octubre de 1987 (figura 3.29b) en que los niveles de K son de
0.098% frente a un 0.15 de hojas de t res años. Sin embargo el aumento que
experimenta la hojarasca de esta especie durante Septiembre- Octubre 1986 es
un tanto inexplicable y quizá pueda deberse a un error analítico.
CALCIO.
En el pinar las variaciones estacionales en la concentración de este
elemento son poco importantes con valores extremos de 0.82-1.3% en P. pinaster
y 0.66-1.07% en P. nigra. Igualmente los coeficientes de variación son bajos
(9% y 12% respectivamente). No pueden establecerse, por tanto patrones
temporales en ninguna de las dos especies. Otros autores han destacado también
la baja variabilidad temporal en la concentración de Ca en arbóreas perennes.
Estos autores lo explican bien debido al carácter estructural y baja movilidad
del Ca (Sharma y Pande 1989 en Pinus rosburghii) o a que las hojas que caen
durante todo el año se encuentran en avanzado grado de desarrollo y por tanto
con altos contenidos de Ca (Escudero eí al. 1983 en encina). A pesar de ello,
a veces puede apreciarse un ligero aumento de concentración en aquellos meses
en que la senescencia de las hojas es más acentuado (Escudero eí al. 1983,
Verdú 1984). En este estudio, incluso las concentraciones de Ca más bajas
encontradas en ambas especies (0.82 en P. pinaster y 0.66% en P. nigra,
182
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
apéndice 3.5) son ligeramente superiores a las encontradas en acículas vivas
de tres y más años (0.77. en P. pinaster y 0.477. en P. nigra). Esto puede
indicarnos que la hojarasca de ambas especies está predominantemente
constituida por acículas, bien senescente, bien vivas pero con un elevado
estado de desarrollo.
En el matorral sin embargo, la concentración de Ca está sometida a una
mayor variabilidad temporal (cv= 26.27. en A. decorticans y 22.87. en C.
lawifolius, tabla 3.16) que además, presenta unas pautas bastante claras en
los dos períodos de estudio sobre todo en el caso de la jara . Tanto en C.
lawifolius como en A. decorticans las concentraciones son más elevadas en el
periodo estival de caída fenológica (figura 3.30c y d) manteniéndose en otoño
mientras continua la caída de hojas viejas. El resto del año los niveles
disminuyen claramente indicando que los pulsos de caída corresponden a hojas
verdes con menor contenido en Ca. Las concentraciones máximas encontradas en
verano son igualmente superiores a las de hojas vivas en estas especies.
Además estos niveles máximos coinciden con los pulsos de caída más fuertes del
año, de ahí que se hayan encontrado relaciones significativas y positivas
entre peso de hojarasca y contenido en Ca (tabla 3.17).
MAGNESIO.
Al igual que ocurre en el caso del Ca, no se manifiesta ningún patrón
estacional en la concentración de Mg. Existe bastante estabilidad en los
niveles de Mg a lo largo del año (CV entre el 10-117.). Los valores máximos
obtenidos (0.217. en P. pinaster y 0.187. en P. nigra) no difieren respecto al
contenido en acículas de t res años (0.187. y 0.117. respectivamente). En A.
decorticans la variabilidad en los niveles de Mg es algo mayor pero esto no
implica un patrón temporal en esta especie (figura 3.31d). La concentración de
Mg en el periodo estival de 1986 en A. decorticans (0.267.) es ligeramente
superior al contenido en hojas vivas (0.2%) lo que parece indicar una cierta
acumulación de este nutriente en hojas senescentes. En la j a ra (figura
3.31c), la acumulación de Mg en la época estival de máximo desfronde foliar es
patente sobre todo en el segundo año (0.29%). Las concentraciones disminuyen a
continuación coincidiendo con caídas de hojas más verdes. Esta relación se ve
reflejada en la significación de la ecuación entre la concentración de Mg y
183
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
peso de hojarasca (tabla 3.17). Los niveles de Mg encontrados en la hojarasca
estival (0.30%) son superiores a los de las hojas vivas (0.22 7.). El resto del
año las concentraciones están alrededor del 0.2% lo que confirma un desfronde
mayoritario de material verde fuera de la época estival.
El Mg al igual que el Ca tiende a acumularse en tejidos senescentes (Gosz
et al. 1972, Waring y Schlesinger 1985). Escudero et al. (1983) encuentran
correlaciones positivas entre el comportamiento del Mg y del Ca en hojarasca
de encina en el sentido de que estos elementos tiende a poseer una mayor
concentración en las hojas más senescentes. Este parece ser el caso de la ja ra
en el presente estudio. Sin embargo Sharma y Pande (1989) encuentran
disminuciones en la concentración de Mg durante la máxima caída de hojas
senescentes en y una relación negativa Mg-peso de hojarasca.
3.3.6.RETRASLOCACION FOLIAR.
3.3.6.1. Introducción.
En términos generales el concepto de retraslocación está relacionado con
la reutilización o la reabsorción "de nutrientes por parte de las especies
vegetales antes de que éstos lleguen a perderse (Chapín 1980). A nivel
individual, la retraslocación permite un mayor control interno de los
nutrientes mejorando la eficiencia en el uso, de éstos. Además esta reabsorción
disminuye la posibilidad de que los elementos sean inmovilizados en la
hojarasca (Mayor 1990) afectando, por tanto, a la velocidad de descomposición.
A nivel de ecosistema o de ciclo biogeoquímico este proceso puede reducir la
posibilidad de que los nutrientes sean exportados de forma disuelta en el agua
de avenamiento
La retraslocación puede ser considerada a un nivel global bien sea para
un ecosistema, una determinada especie o una planta individual. Puede
calcularse como la diferencia entre requerimientos y absorción (uptake) (Colé
y Rapp 1981) o bien entre requerimientos y retorno (Gray 1983). Según autores
esta retraslocación total puede denominarse también reabsorción (resorption) o
184
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

redistribución (redistribution).
Desfronde
La retraslocación total viene en buena parte condicionada por mecanismos
locales que actúan de manera más intensa en determinados tejidos vegetales y
en momentos concretos del ciclo vegetal. Así, es muy frecuente que exista una
fuerte retirada de algunos nutrientes por parte de las hojas senescentes antes
de caer. Estos nutrientes serán destinados a la formación inmediata de nuevas
estructuras o bien serán almacenados en órganos perennes para un uso
posterior. Este proceso es el denominado retraslocación o reabsorción foliar.
La prueba más evidente de su existencia es la dismininución de la
concentración química de las hojas durante el desfronde y la manera más
sencilla de determinarlo consiste en calcular la diferencia entre la
concentración de un elemento dado en hoja vivas y en hojas senescentes. Este
cálculo, si bien muy útil desde el punto de vista cualitativo, no debe
emplearse de forma estricta para cuantificar la traslocación debido
básicamente a dos motivos expuestos por Gray (1983). El primero consiste en
que la hojarasca puede haber estado sometida a lixiviado o contaminación y por
tanto no es un reflejo exacto del estado nutricional de las hojas. El segundo
motivo es la reducción de peso seco que sufren las hojas durante la abcisión
lo que puede hacer sobrestimar la reabsorción cunado se calcula a part i r de
diferencia en concentraciones químicas (en 7. de peso seco). En ecosistemas
mediterráneas, la abcisión suele coincidir con el periodo estival en que las
precipitaciones y por tanto el lixiviado se reducen al mínimo. Además la
vegetación dominante consiste en especies de hoja esclerófila dónde la
disminución de peso durante la abcisión es pequeña. Por ello este método de
cálculo puede ser empleado con mayor fiabilidad que en otras especies de hojas
más mesófilas o bien aquellas cuyo desfronde se produce en épocas más
favorables para el lixiviado.
Otra aproximación para estimar la retraslocación o reabsorción foliar es
la que utilizan Del Arco et al. (1991) para el caso concreto del nitrógeno. Se
utiliza la diferencia entre la cantidad máxima de N acumulada anualmente en
las hojas y el retorno anual de N por hojarasca expresando los valores en
forma de porcentaje. Según estos autores el valor obtenido constituye una
medida de la retraslocación real. Además, introducen la denominada
185
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
retraslocación potencial en la que el N retornado por hojarasca estaría
calculado suponiendo que toda la hojarasca caída anualmente tuviera la misma
concentración que la de la época de máxima abcisión fenológica cuando es de
esperar que los mecanismos de reabsorción funcionen con un mayor rendimiento.
La diferencia entre la retraslocación foliar potencial y real reflejaría la
disminución en la eficiencia de la retraslocación motivada por escisiones
accidentales de hojas verdes debido a factores externos (meteorológicos,
animales etc) (Del Arco et al. 1991).
Hemos intentado utilizar al máximo la información disponible y calcular
la retraslocación mediante distintos métodos. En este apartado sólo se
comentarán los resultados obtenidos para la reabsorción foliar. En el capítulo
7 se estimará la retraslocación total incluida dentro del ciclo interno global
para cada especie. Nos hemos centrado exclusivamente en los dos elementos que
suelen ser limitantes en ecosistemas mediterráneos: N y P.
3.3.6.2. Resultados y discusión.
1.- Calculo de la reabsorción foliar mediante diferencias de concentración.
Aunque este método puede incluir pérdidas de nutrientes debidas al
lixiviado, hemos de suponer que dada la época de máxima abcisión en las
especies estudiadas el error cometido debe ser mínimo al menos en P. pinaster,
C. laurifoiius y A. decorticans. En P.nigra, sin embargo podemos considerar
que las eficacias de retraslocación que se obtengan pueden haber sido
sobrestimadas ya que en otoño (época de máxima abcisión foliar en esta
especie) el lixiviado a part ir de las lluvias otoñales puede tener importancia
sobre todo respecto al P.
Un problema adicional que surge a la hora de aplicar el modelo a las
especies de pino consiste en definir si la retraslocación foliar se establece
como la diferencia de concentración desde que la hoja es joven (acículas del
primer año) hasta que cae cuatro o más años después o bien, si consideramos
como reabsorción solamente la que ocurre, inmediatamente antes de la abcisión,
es decir, la diferencia de concentración entre acícula presenescente y la
186
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.18a. Concentración química de N y P (% del peso seco) en hojas de diferentes edades en las distintas especies.
Hl H2 H3 H>3 Hps Hs Hps Hs * i i * 2 2
P.pinaster:
N 0.92 0.85 0.80 0.66 0.35 0.33 0.45 0.30
P 0.10 0.083 0.075 0.075 0.062 0.057 - 0.048
P. nigra:
N 0.91 0.86 0.82 0.72 0.46 0.33 0.46 0.39
P 0.09 0.083 0.084 0.08 0.055 0.033 0.051 0.034
C. laurifolius:
N 1.37 - - 0.38 0.95 0.40
P 0.15 - - 0.098 0.13 0.11
A. decorticans: N 3.0 - 1.55 - 1.87 P 0.12 - - 0.04 - 0.047
Tabla 3.18b. Mineralomasas de N y P (descontado la retención en las especies de pino) en hojas/acículas de 1 año para las cuatro especies (g/m ). Especie N P
P.pinaster 0.92 0.10
P. nigra 0.90 0.09
A. decorticans 1.50 0.06
C. laurifolius 0.54 0.06
Hl, H2, H3 y H>3= Concntración en hojas de uno, dos, tres y mas de tres años.
Hpsl y Hps2= Concentración en hojas presenescentes durante el primer y
sejundo año de muestreo
Hsl y Hs2= Concentración en hojas senescentes en la época de máxima abcisión
durante el primer y secundo año de muestreo.
senescente. En este sentido, hemos contemplado ambas posibilidades calculando
la retraslocación parcial hacia hojas vivas de distintas edades (1, 2, 3, más
de 3 y presenescentes) desde las acículas senescentes que caen en el período
de máximo desfronde. Los valores de concentración en hojas vivas de diferentes
187
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
edades se han tomado de los resultados existentes para estas fracciones en
biomasa. La concentración de acículas presenescentes se ha considerado la
obtenida a part i r de la hojarasca caída el mes anterior a la máxima abcisión
(tabla 3.18).
En la tabla 3.19 se muestran los valores de eficiencia de la reabsorción
en las acículas de pino de diferentes edades en relación a las senescentes. De
los resultados obtenidos para el N se desprende que la retraslocación de este
elemento en hojas de pino es un proceso gradual que se va sucediendo conforme
la hoja va madurando y envejeciendo. La eficacia de este proceso va
disminuyendo conforme aumenta la edad de las acículas puesto que parte del N
ha sido ya retirado en estadios anteriores. Así mientras la eficacia global
media (acículas primer año- acículas senescentes) alcanza valores elevados
(657. en P. pinaster y 60% en P.nigra) la reabsorción puntual inmediata a la
abcisión tiene mucho menos importancia obteniéndose porcentajes mucho más
bajos y muy similares en ambas especies (19.3 y 21.7% de rendimiento en P.
pinaster y P.nigra respectivamente). Sin embargo a pesar de esta menor
eficiencia en los momentos finales se retrasloca entre el 29.4% y 36% de todo
el N reabsorbido a lo largo del ciclo foliar.
Los resultados obtenidos para el P son algo más dispares entre las dos
especies. La eficiencia global es superior en P.nigra con un 62.5% de
reabsorción frente al 50% de P. pinaster. En la primera especie los momentos
anteriores a la escisión son muy importantes para la reabsorción de este
elemento obteniéndose rendimientos del 45% (tabla 3.19). Por tanto, un 72% del
P total retraslocado se movilizaría en ese período. En P. pinaster la
importancia de los momentos finales es inferior suponiendo sólo un 16% del P
total retraslocado a lo largo de la vida de la hoja. Como se comentó
anteriormente la eficiencia en P.nigra puede haber estado sobrestimada por el
efecto superpuesto de lixiviado durante la caída otoñal de hojas que habría
hecho disminuir la concentración química de P por debajo del valor esperable
si hubiesen actuado únicamente los mecanismos de reabsorción.
Escarré et al. (1987) utilizan una aproximación similar siguiendo un
gradiente de edad en hojas y tallos de encina en una misma rama comprobando la
188
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.19. Porcentajes de retraslocación calculados a part ir de las diferencias de concentración entre acículas vivas de distinta edad y senescentes.
P.pinaster:
Al-As
A2-As
A3-As
A>3-As
Aps-As
P. nigra:
Al-As
A2-As
A3-As
A>3-As
Aps-As
N
64
61
58.7
50
5.7
N
63.7
61.6
59.7
54.1
28.2
AÑOl
AÑ01
P
45.7
31.3
24
24
8
P
63
60
60
58.7
40
N
67.4
64.7
62.5
54.5
33
N
57.5
54.6
52.9
46.2
15.2
AÑ02 P
54.2
42
36
36
-
AÑ02 P
62
59
59.5
57.5
50
N
65.5
62.8
60.6
52.2
19.3
N
60 .6
58.1
56.3
50.1
21.7
MEDIA P
50
36.6
30
30
8
MEDIA P
62.5
59.5
59.7
58
45
Al, A2, A3 y A>3: Acículas de uno, dos, tres y más de tres años.
Aps y As: Acículas presenescentes y senescentes respectivamente.
reabsorción secuencial de N, P y K a distintos tiempos de brotada (abril a
agosto). Un 16.7 7. de N se retrasloca en los últimos momentos del ciclo foliar
es decir a part ir de las hojas senescentes, porcentajes muy parecidos a los
obtenidos en nuestro pinar (19-21%). La eficiencia en la retraslocación global
durante todo el periodo de brotación es en la encina del 487. frente al 60-65 7.
obtenido en las especies de pino estudiadas. Respecto al P un 17% se
retrasloca a part ir de hojas senescentes en la encina muy similar al 167.
obtenido en P. pinaster de este trabajo.
La eficiencia de retraslocación de N en las especies arbustivas calculada
por diferencia de concentración entre hoja viva y hoja senescente es bastante
distinta en ambas especies (tabla 3.20). Mientras que la j a ra reabsorbe un
189
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.20. Eficacia de la retraslocación (7.) en las especies arbustivas A. decorticans y C. laurifolius. calculada a part ir de la diferencia de concentración entre hojas vivas y senescentes.
AÑ01 AÑ02 MEDIA N P N P N P
C.laurifolius:
Hv-Hs 72 34 70.8 26.6 71.4 30
Hps-Hs - - 58 15 58 15
A. decorticans: Hv-Hs 48 66 37.6 60.5 43 63.2 Hv = concentración en hoja viva; hs; concentración en hoja senescente; hps:
concentración en hoja presenescente
71.4% de N, A. decorticans tan sólo lo hace con un 43%. Muy probablemente la
condición de fijadora en A. decorticans favorezca el status nutricional de
esta especie con respecto al nitrógeno y por tanto no se hagan tan necesarios
los mecanismos de reabsorción tendentes a economizar al máximo el uso de este
nutriente. Para el P es C. laurifolius la que posee un baja eficiencia de
reabsorción (un 30%)muy inferior a la de A. decorticans (63.2%). Las
concentraciones de P no disminuyen apenas cuando las hojas envejecen y este
patrón se repite sistemáticamente en los t res períodos de estudio (ver este
mismo capítulo y el capítulo 5).
En la j a ra se ha calculado también la eficacia de la retraslocación
inmediatamente antes de la abcisión fenológica. Hemos utilizado la
concentración de la hojarasca del mes de junio (un mes antes de la máxima
caída) como medida más aproximada del nivel de N y P en hojas presenescente.
En la tabla 3.20 puede verse como la eficacia en este momento concreto es del
58% para el N y del 15% para el P. Esto indica que de todo el N y P
retraslocado a lo largo de la vida de la hoja un 81% y un 50% respectivamente
se produce en los momentos anteriores a la caída. No hemos podido realizar el
mismo cálculo en A.- decorticans porque no disponemos de datos de concentración
foliar en hojas presenescentes ya que durante los meses anteriores a la máximo
abcisión (Mayo y Junio) no se registra caída de hojas. En otras épocas del año
fuera de la abcisión estival las hojas que caen poseen contenidos similares a
190
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
las de hojas vivas con Jo que podemos concluir que es material verde en el que
no han existido procesos de reabsorción. La retraslocación foliar en A.
decorticans viene pues determinado casi exclusivamente por la ret irada de los
nutrientes en un corto periodo de tiempo inmediatamente antes de la abcisión.
En la j a r a es también, como hemos visto, la reabsorción anterior a la máxima
caída la que controla fundamentalmente el contenido final de P y sobre todo de
N en la hojarasca.
En resumen, independientemente de la mayor o menor eficiencia global de
la retraslocación foliar en las cuatro especies, puede deducirse de estos
resultados que las especies perennes con una longevidad foliar mayor (pinar)
retrascolan de un forma muy gradual los nutrientes llegando a ser la
reabsorción muy pequeña en los momentos finales del ciclo fenológico foliar.
Sin embargo las otras especies, que a pesar de ser perennes tienen una
longevidad foliar mucho menor (unos 16 meses) presentan un patrón de
reabsorción muy diferente ya que es casi exclusivamente en los momentos que
preceden a la abcisión fenológica cuando se detecta una retirada patente de N
y P.
2 . - Cálculo en la reabsorción foliar mediante diferencias entre
mieneralomasas.
El segundo método consiste en calcular la retraslocación mediante la
fórmula (Del Arco et al. 1991): ,
[Máximo pool de N ó P foliar anual - Retorno anual por hojarasca de N ó P] /
Máximo pool de N ó P foliar
2
Las unidades empleadas son g/m -año y el resultado se expresa como
porcentaje. Esta fórmula representaría la retraslocación real. Si el segundo
término de la ecuación se sustituye por el retorno de N ó P calculado a part i r
de la concentración durante el máximo desfronde fenológico y del peso anual de
desfronde foliar se obtiene la retraslocación potencial. Los autores
anteriores suponen que la biomasa foliar es constante.
191
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

•1
M.J. Moro Cuadrillero
Tabla 3.21. Eficacia media de traslocación foliar (7.) estimada para el N y P en las cuatro especies estudiadas.
ESPECIE PP
MÉTODO N P
PN
N P
AD
N P
CL
N P
1 65.5 50 60 62 43 63 71 30
2A(Pot.) 41.6 20 59 59 40 64 72 33
2B(reai) 44 39 49 54 25 35 67 43
método 1 = Diferencia hoja* de 1 año con respecto a las senescentes (en %).
método 2a= Mineralomasa hojas del año (descontando la retención)- mineralomasa de hojas en desfronde si todas ellas tuvieran la concentración del mes de máxima abcislón (en X). método 2b= Mineralomasa hojas del año(descontando retención) ' - mineralomasa retornada por desfronde.
En este trabajo hemos considerado como máximo pool de N y P foliar anual
en las especies arbustivas a las cantidades de N y P asociadas con la
producción foliar calculadas en el capítulo 2 (ponderando por superficie los
valores en RPP y RPN para obtener un valor a nivel de cuenca).
Para las especies de pino, a los nutrientes asociados a la producción
de acículas (tablas 2.23 y 2.24) se ha descontado la fracción correspondiente
a la retención (tabla 2.26) para obtener una medida de la cantidad de N y P
utilizado para el mantenimiento del sistema.
Los resultados obtenidos aparecen en la tabla 3.21 (2a para la
retraslocación potencial y 2b para la real) a los que se han añadido los del
método anterior (1) con el fin de facilitar la comparación entre ambos. Los
porcentajes obtenidos según el método 1 y 2a (tabla 3.21) deberían ser
similares mientras que los del método 2b incluirían el efecto de las caídas
accidentales o traumáticas no afectadas por la reabsorción y por tanto es
perfectamente lógico que puedan obtenerse resultados algo diferentes.
Las mayores eficiencias reales (método 2b) respecto al N se registran en
192
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Deifronde
P.nigra y jara. (49 7. y 677.) y la más baja en A. decorticans (257.). La
influencia de la caída accidental en las que se pierden una gran cantidad de
hojas verdes es determinante para este bajo rendimiento encontrado en A.
decorticans no sólo para el N sino para el P. Respecto al P vuelve a ser
P.nigra la especie con mayor eficacia (547.) seguido por P. pinaster y C.
laurifolius con rendimientos medios de alrededor del 407. y por último A.
decorticans con un 35%.
La retraslocación potencial ha de ser, por definición mayor, en mayor o
menor medida, que la traslocación real. Sin embargo existen algunas
excepciones que se deben evidentemente a un error del método ya que el término
mineralomasa anual retornada puede verse afectada por otros factores que no
tengan que ver con la reabsorción (lixiviado etc). Así, la explicación más
plausible para la obtención de un coeficiente más alto de reabsorción real de
P en la j a ra es la retirada de P mediante lixiviado debido al agua de lluvia.
La disminución de la concentración de este elemento en otoño es muy patente
(figura 3.28c) llegando a estar bastante por debajo de la obtenida en el
momento de máxima abcisión (Julio o Agosto). Como consecuencia de ello la
eficiencia real está siendo sobrestimada porque está incluyendo una retirada
de P que no es debida a la reabsorción. Para P. pinaster la explicación es
similar aunque con algunos matices. Las acículas que se recogen en invierno
realmente ya se han desprendido meses antes (probablemente ese mismo verano).
Desde entonces hasta que caen en las trampas deben haberse producido con toda
seguridad no sólo lixiviado de P sino pérdida de peso en las acículas. Ambos
factores se conjugan para introducir un error al calcular la eficiencia real
por este método.
Comparando ahora el método 1 y 2a, las eficiencias calculadas son muy
similares tanto para el P como para el N en A. decorticans, C. laurifolius y
P. nigra. aunque existe una tendencia a que las eficiencias sean ligeramente
superiores por el método 1.. Sin embargo, existen diferencias destacables en
los resultados de ambos métodos para P. pinaster. Evidentemente, las
diferencias se deben en gran parte a la introducción del factor "peso de
desfronde foliar" en el cálculo del método 2a. ya se ha comentado en el
capítulo 3 que este parámetro puede estar sobrestimado por dos motivos. En
193
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
primer lugar la caída media de acículas en el pino resinero tiene una elevada
variabilidad (CV=26%) ya que durante el segundo año se registró una fuerte
caída de material verde en invierno. En segundo lugar, durante los t res años,
la llegada al suelo de acículas escindidas en años anteriores, supone una
entrada adicional que sobrestima el desfronde real del año considerado. La
retraslocación así calculada será entonces menor que la obtenida a par t i r de
las diferencias de concentración obteniéndose menores eficacias por el método
2a y 2b frente al método 1.
En la tabla 3.22 se han recogido datos sobre retraslocación existentes en
la bibliografía calculados por distintos métodos. La eficacia de
retraslocación real de N en P. pinaster es muy similar y la potencial
ligeramente inferior a la obtenida por Del Arco et al. (1991) para distintas
especies de pino. Los datos de reabsorción real y potencial de N en P. nigra
son del mismo orden a los obtenidos por Del Arco et ai.(1990). Los valores de
retraslocación de N a part ir del método 1 en los dos pinos estudiados son
ligeramente superiores a las obtenidas por el mismo método por Alvera (1990)
en P. sylvestris e inferiores respecto al P. Así, obtiene eficiencias del 587.
en el N frente a un 65 y 60% en los pinos estudiados y un 69% para el P frente
al 50 y 627. en P. pinaster y P. nigra. Sin embargo Alvera (1990) encuentra
embargo eficiencias muy elevadas en P.sylvestris del Pirineo tanto para el N
(75-85%) como para el P (83-89) a par t i r de diferencia de mineralomasas,
valores que siempre resultan muy superiores no sólo a los pinos de Almería
sino también a los estudiados por Del Arco et al (1990). La retraslocación en
encina (Mayor 1990, Del Arco et al. 1991, Escarré et al. 1987 ) es inferior a
la encontrada para los pinos estudiados oscilando entre 31-48% para el N y
31-43% para el P.
Entre las especies arbustivas A. decorticans presenta una baja eficiencia
potencial (40%) cercana la coscoja (37%) respecto al N. Además, la reabsorción
calculada por el método 1 en A. decorticans resulta muy similar al brezales
estudiados por Berendse et al. (1987) y Merino (1986). La j a r a posee
porcentajes de traslocación de N (71%) a part i r de diferencias de
concentración muy similares a otras cistáceas estudiadas por Merino (1986) en
Doñana (61-78%). La reabsorción inmediata a la abcisión es del 15% para el P
194
1
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
e -o
e o - I L . 01 O « <M u a •u -o 0 u. u
s
¡i £32
¡3
00 0>
o *
u u >> u tu 0) X u
n —
O 00
01
a.
3*
0>
1 3 X 3 3 3
á
i *•• -• vO * •* PJ «-• PJ n n «i i/i w * i/i
oo f- ui p-pj o
g — o M o PJ *o n n í ) 1 '
o "I V . V. 01 01 - g <«, t . I * oí 3
3 y «i * * i> <* - « * . o v »i a . v o o o * s w c "*« U y —* C -* C ~* oí oí oí a . J S o . § • 3 3 3 U u oí oí oí ui
« « S i S S II a a —• •-* —i -* —* c5 O a. a. a. a. —
00 os 0>
«-« •*** — • *
(4
4 ->
0
. * 3 U 19
j a
2 — 4
—-t i — *
i-i
« 4
^ c*-00 Cf» í - t
• W
—( a d i - * J
V
u 4)
+J -t-> O
CU
^ -—» f~ co OS
*** s _ *
F - *
td
t a •*-> 4)
V 01
1 41 u ¿
** J - *
>o 00 O
*-• *« o a
- i
1) X
o i o o o t - e - ^ a c M » * i / i p s i c i v o p - m p - i / i
O - O O O O O v O P - P J
2 id
- i 01
• - *
X
0
1 £
A i <•->
0 0
O
u
I « O J <«
- . n) » o - • y
i» . - « y —« •«* «i y y -a
-i * - . U «I B ••« —» 3
3 y i . << -o
o» -< 0) 3 3 C t» v. y -» 3
oí <« a x
C —* C - * - i u o «t 1- O U 1 -
y -» i-y >> v
5 ¿ á ^ 8 5 *
I» O - i
3 3 3 3 3
c* -« PJ oo N p-^ p» p- p~ v$ co
c >
á
35 =¡3 O - t
<••« y ••^ •••» • <H
- . v , - . o
-C 01
01
<« 0) .c - . y
a 8-3 Í « i
§5 -. -. a « - i —«
—< —• 55
H •-4 w 3 0) a
—1 —* 0) 3
« 01
5 3 •9 S 1 i
O y 01
<« y •«*
U -4 UJ
o C*N 3 3 3 3
n 00
N
0 '
00
o vO
P-
vO
t ~
U1 1/1 P- 00
n — CM p-oo \n n m ift « M
«i o 0 > a
—i —* l*
S fiSfS. - 1 - I 1 » I
0)
01
s oV
&o,Si " 3 o , X W 3 41 V X s o s
195
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en la j a ra algo inferior al encontrado en otros matorrales estudiados por
Killingbeck (1988) en el que se han encontrado reabsorciones de 287. en el
género Vaccinium y del 33% en Guylussacia.
3.3.7. RETORNO DE NUTRIENTES AL SUELO VIA DESFRONDE.
El desfronde es el flujo principal de circulación de algunos nutrientes
como el N, Ca, P y Mg en la mayor parte de los ecosistemas (Gray y Schlesinger
1981). La magnitud de retorno es función del peso de desfronde y de su
concentración elemental. La relación entre la pérdida por desfronde y la
cantidad acumulada en la biomasa es para la mayoría de los nutrientes de un
10-15% en ecosistemas en desarrollo tanto en matorrales mediterráneos (Gray y
Schlesinger 1981), como en bosques templados (Likens et al. en Gray y
Schlesinger 1981). Sin embargo estas proporciones pueden variar
sustancialmente en sistemas muy jóvenes y en aquellos dominados por plantas
con una alta productividad.
Aportes totales en la cuenca.
El desfronde total en las dos regiones supone un retorno medio anual de
75 Kg/ha.año de elementos (N, P, K, Ca, Mg y Na) en RPP y de 62 Kg/ha.año en
RPN (tabla 3.23). El orden de importancia de los nutrientes sigue la siguiente
sucesión:
Ca - N > K > Mg > P > Na
Este orden coincide con el encontrado para la mayor parte de autores
consultados (Gholz et al. 1985, Rapp y Lossaint 1981, Lossaint y Rapp 1971,
Rapp 1967, Verdú 1984, Escudero et al. 1985, Gray 1983) en distintos sistemas
arbóreos y arbustivos.
El cociente nutrientes retornados/nutrientes en biomasa es del 19-24% lo
que indica una etapa aún muy joven del ecosistema. El índice se ha calculado a
part i r de la mineralomasa calculada en el bosque en Octubre de 1986 y el
196
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfrondo
Tabla 3.23. Aportes tota les anuales (Kg/ha) vía desfronde en una ha. media para cada región. Se indica también el valor del peso del desfronde (g/m2).
RPP
jul86/.
jul87/.
jul88/.
aedia
std cv(%)
RPM
¡un87
¡un88
¡un89
M
28.72
29.15
29.49
29.12
.39 1.33
II
P
1.92
2.09
1.64
1.88
.23 12.14
P
K
a.32
8.25
6.52
7.69
1.02
13.26
K
Ca
26.02
37.22
27.04
30.09
6.19
20.58
Ca
Hg
5.49
6.51
5.40
5.80
.62 10.72
ig
Ha
.53
.42
.46
.47
.06 12.18
Ha
TOTAL
70.99
83.64
70.55
75.06
7.43
9.90
TOTAL
DESFROMDE
283.
357.
317.
.04
.90
.34
319.43
37, 11. .47 .73
DESFMXOE
j'ul86/jun87
jul87/jun88
jul88/jun89
•edía
std cv(%)
27.50
16.37
30.58
24.82
7.47
30.11
1.84
1.17
1.70
1.57
.35 22.34
7.96
4.63
6.76
6.45
1.69
26.13
24.91
20.90
28.04
24.62
3.58
14.53
5.25
3.66
5.59
4.83
1.03
21.36
.51
.23
.48
.41
.15 36.96
67.97
46.97
73.14
62.69
13.86
22.11
271 201 329
267 64.09
24.01
retorno por desfronde medio durante el período (86-89).
La variabilidad interanual en RPP es poco importante, no sólo para el
peso (CV=11%) sino para el aporte de nutrientes (tabla 3.23). Excepto el Ca,
cuyo coeficiente de variación interanual es del 207., los demás nutrientes
presentan valores inferiores (entre el 1.5 y 13%). En RPN la variabilidad
interanual es más acusada tanto para el peso como para los aportes
elementales. Los coeficientes de variación oscilan entre el 14% del Ca y el
377. del Na.
La distribución en fracciones de los aportes (tabla 3.24) muestra un
predomimio claro de la hojas/acículas que contienen entre el 79 y 83 7. de los
197
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
nutrientes aportados por este flujo (62.89 en RPP y 49.49 en RPN). Los órganos
reproductivos (flor, fruto, semillas y pinas) retornan al suelo porcentajes
(14-16%) y cantidades similares (alrededor de lOKg/ha.año) en ambas regiones.
Las ramas, dadas sus menores concentraciones químicas y su escasa importancia
cuantitativa, tan sólo suponen 2.3-4.47..
El orden de importancia de los nutrientes en las diferentes fracciones
queda, en algunos casos alterado respecto al patrón general:
HOJAS: Ca > N > K - Mg > P > Na
REPRODUCCIÓN: N> K > Ca > Mg > P >Na
RAMAS: N >Ca >Mg - K > P > Na
ó
N: REPRODUCCIÓN >RAMAS >HOJAS
P: HOJAS > REPRODUCCIÓN > RAMAS
K: REPRODUCCIÓN > HOJAS > RAMAS
Ca: HOJAS > RAMAS > REPRODUCCIÓN
Mg: HOJAS > RAMAS > REPRODUCCIÓN
El K y el N ocupan el primer lugar en los órganos de ciclo más rápido
(fracción reproductiva). El Mg, componente de la clorofila ocupa el primer
lugar en hojas. La fracción ramas es relativamente rica en N y pobre en Ca ya
que está compuesta básicamente por ramillas y tallos. Esto puede explicar el
orden de estos dos elementos en esta fracción en relación con las hojas.
Aportes por especies.
En la tabla 3.25 están indicados los aportes de cada nutriente para las
cuatro especies durante los t res años así como los valores medios. Se realizó
un análisis de la varianza seguido del test de Newman- Keul's para establecer
qué especies aportaban cantidades significativamente diferentes de los
198
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.24. Retorno de las principales fracciones por desfronde en la cuenca "Nacimiento".
RPP N P K Ca Mg Na
HOJA/ACICULA 21.7 1.53 5.50 28.6 5.27 0.29
REPRODUCCIÓN 6.2 0.31 2.13 1.55 0.56 0.07
RAMILLAS 1.2 0.05 0.07 0.31 0.08 0.02
TOTAL
RPN
29.1
N
1.89
P
7.70
K
30.47
Ca
5.90
Mg
0.39
Na
HOJA/ACICULA 17.1 1.20 4.32 22.5 4.14 0.23
REPRODUCCIÓN 5.9 0.30 2.03 1.48 0.53 0.07
RAMILLAS 1.9 0.08 0.11 0.51 0.13 0.03
TOTAL 24.9 1.58 6.46 24.49 4.80 0.33
distintos nutrientes. Los resultados del ANOVA indican diferencias
significativas para todos los nutrientes (p< 0.005) a excepción del P. Con el
test de Newman Keul's (tabla 3.26) puede establecerse que A. decorticans
presenta una tasa de circulación vía desfronde significativamente más alta que
las demás especies respecto al K y N mientras que P. pinaster aporta
cantidades significativamente superiores para los demás cationes, Ca, Na y Mg.
La circulación de P es superior en P. pinaster y A. decorticans con respecto a
P. nigra y C. laurifolius. Por tanto, el desfronde del gaznacho es la
principal vía de retorno de elementos móviles mientras que el de P. pinaster
moviliza preferencialmente nutrientes de carácter estructural y menos móviles.
Hay que destacar que mientras que el peso de desfronde en A. decorticans es
inferior desde el punto de vista estadístico al de P. pinaster (apartado
3.3.1) aporta al suelo cantidades significativamente superiores de N y K y
similares de P indicando un ciclo rápido en esta especie arbustiva.
La ja ra y el pino laricio tienen una contribución mucho más pequeña en el
aporte de nutrientes al suelo forestal. A pesar de que el desfronde de P.
199
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

U.J. Moro Cuadrillero
Tabla 3.25. Nineraloaasa total para cada eleaento en el desfronde de las cuatro especies.
Mineralomasa
jul86/jun87 '
jul87/jun88
jul88/jun89
jul89/sep89
•edia
std
Mineralomasa
jul86/jun87
jul87/j"un88
jul88/jun89
jut89/sep89
•edia
std
Mineralomasa
jul86/jun87
jul87/jun88
j'ul88/jun89
jul89/sep89
•edia
std
total
M
6890,
10715.
6585.
2095
(g/ha)
.39
.73
.67
.28
8063.93
2301,
total
M
.58
(g/ha)
«717.52
4421.
5564,
441,
4901
593
total
H
20632
11430
22105,
.88
.52
.35
.31
.07
P.pi
P
786.
901.
588.
1599.
inaster (supf.repoblada)
.54 ,30 ,28 .39
758.70
158.36
P.ni
P
468.
374.
igra
,78 .19
517.42
39. .21
453.47
72. .84
A.decorticans.
.40
.25
.07 8802.49
18055.91
5785 .04
P
923.
425.
781.
286.
710.
256.
.01
.92
.58
.24
.17
.12
K
1789.
2414.
.57
.39 2137.42
717.60
2113.79
313.
(supf.[
K
1329.
732.
1036.
135.
.08
Ca
16301.
26464.
17982.
7014.
.91
.42
.85
.06
20249.73
5447.31
-epoblada)
.54
.18
.77
.65
1032.83
298.
(g/ha).
t
5902.
3117,
4400.
.70
Ca
8235.
7852.
10690.
851.
.69
.86
.48
.99
8926.34
1539.
. (cuenca)
.84
.95
.00 1736.40
4473.60
1393. .90
Ca
8284.
4628,
8054.
.73
.09
.61
.24 4608.37
o)rao* B
2047.37
Ng
3296.
4599.
,79 ,81
3492.00
1406. .24
3796.20
702.
Hg
1800.
1436.
2039.
195.
,76
.80
.17
.61
.55
1758.86
303.
Ng
2093,
862,
1972.
934,
.90
.13
.70
.44
.81
1642.76
678.24
*a
491.
430.
402.
150.
441.
45.
Ma
136.
67.
,17 ,91 ,70 ,19
.59
.19
.74
.46 207.59
11. .39
137.26
70.
Ma
197.
63 159.
58,
.06
.15
.56
.77
.00
140.16
68 .92
TOTAL
29556.
45526.
31188.
12982.
,39 ,49 ,91 .76
35423.93
8787. ,07
TOTAL
16688.92
14884.
20056.
1675.
.81
.40
.13
17210.04
2624. .88
TOTAL
38032.
20529.
37473.
.59
.02
.09 16426.32
32011
9948,
.57
.11
DESFRONDE
176.
256.
180.
67.
,17 ,08 ,70 .28
204.32
44. .89
DESFROIDE
119.
94. 129.
11.
.72
.13
.11
.94
114.32
18. .10
DESFRONDE
96. 53, 129.
46.
.56
.96
.59
.21
93.37
37.92
Mineralomasa total C.laurifolius (g/ha) (cuenca)
jul86/jun87
jul87/jun88
jul88/jun89
jul89/sep89
•edia
std
a
1916.93
2714.29
2275.61
2997.11
2302.28
399.35
P
372.91
503.77
356.44
346.66
411.04
80.73
t
716.58
1435.33
892.70
957.25
1014.87
374.63
Ca
6180.04
7759.63
6631.35
6652.34
6857.01
813.61
Na
959.55
1274.90
1051.25
1095.37
1095.23
162.21
Ma
67.50
45.37
43.79
47.79
52.22
13.26
TOTAL
10213.54
13733.31
11251.14
12096.52
11732.66
1808.62
DESFRONDE
40.91
52.12
50.19
47.39
47.74
5.99
200
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.26. Resultados del test " a posteriori" de Newman-Keul aplicado al retorno de nutrientes vía desfronde en las cuatro especies estudiadas (Kg/ha.año). Para cada columna, los valores seguidos de una misma letra no son significativamente distintos (p<0.05)
ESPECIE N P K Ca Mg Na
a a a a a a P.pinaster 8.06 0.76 2.11 20.25 3.79 0.44
a a b b b b P.nigra 4.90 0.45 1.03 8.92 1.76 0.13
b a c b b,c b AAecorticans 18.00 0.71 4.47 6.99 1.64 0.14
c b b b c,d b C.laurifolius 2.30 0.41 1.01 6.86 1.09 0.052
nigra es significativamente superior al de la jara , aporta cantidades muy
similares de Ca y P y sólo ligeramente superiores de N, Mg y Na (Tabla 3.24 y
tabla 3.25). P. nigra parece pues, ser una especie altamente conservativa en
el uso de nutrientes.
La dinámica estacional en los aportes elementales para cada especie está
correlacionado con el peso del desfronde en todos los casos. Las oscilaciones
en las concentraciones químicas mensuales no son, por tanto suficientes para
modificar sustancialmente la dinámica mensual de los aportes respecto a la del
peso. De hecho la variabilidad interanual en los aportes de nutrientes en cada
especie (Tabla 3.24) es muy similar a la del peso seco. A este respecto, los
nutrientes que suelen presentar una mayor variabilidad que la del peso seco
son el Na en P. nigra, A. decorttcans y C. laurifolius y el K en P. nigra y C.
laurifolius. La especie con una mayor variación interanual respecto a la
cantidad caída y, por tanto respecto a los nutrientes aportados, es A.
decorticans.
En la tabla 3.27 pueden observarse los aportes totales de cada nutriente
diferenciando el estrato arbóreo del arbustivo. El valor correspondiente a las
dos especies de pino se ha ponderado para calcular los aportes en una hectárea
201
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.27. Aportes de nutrientes vía desfronde por parte del pinar y matorral en una ha
media de la cuenca "Nacimiento".
mineralomasa total (g/ha) MATORRAL.
jul86/jun87
jul87/jun88
jul88/]un89
jul89/sep89
•edia
std
M
22549.33
14144.54
24380.68
11799.60
20358.18
5458.53
P
1295.92
929.69
1138.02
632.90
1121.21
183.69
K
6619.42
4553.28
5292.69
2693.65
5488.46
1046.89
Ca
14464.13
12388.24
14685.59
11260.71
13845.99
1267.29
Ng
3052.68
2137.60
3023.69
2030.18
2737.99
520.16
Na
264.65
108.93
203.56
105.79
192.38
78.46
TOTAL
48246.13
34262.33
48724.23
28522.84
43744.23
8215.04
D€SFROMDE
g/m2
137.47
106.08
179.78
93.60
141.11
36.98
mineralomasa total (g/ha). PINAR
M P K Ca Ng Ha TOTAL DESFMNDE
jul86/jun87
jul87/jun88
jut88/jun89
Jul89/sep89
•edia
std
5430.75
6483.01
5901.44
982.61
5938.40
527.10
572.97
546.81
540.87
549.59
553.55
17.08
1480.71
1282.84
1397.35
326.09
1386.97
99.34
10878.57
13945.11
13081.49
2868.15
12635.06
1581.26
2291.10
2471.80
2515.77
591.69
2426.22
119.07
252.75
186.39
271.52
56.80
236.89
44.73
20906.76
24915.98
23708.45
5374.93
23177.06
2056.75
138.25
147.16
146.05
30.05
143.82
4.86
ideal de toda la cuenca. Destaca la contribución del matorral, que con un
desfrondei similar al pinar aporta al suelo un 477. más de nutrientes. La
presencia del estrato arbustivo en repoblaciones de coniferas parece, pues,
interesante por lo menos desde el punto de vista de una devolución rápida e
importante de nutrientes al suelo.
Comparando los aportes totales de cada estrato con los encontrados en la
bibliografía (tabla 3.28), puede señalarse que nuestros resultados son
inferiores a los que presentan muchos autores. Las razones argumentadas en
este mismo capítulo en relación con el peso de desfronde son igualmente
válidas en el caso de los aportes. La juventud del pinar estudiado y la baja
cobertura del matorral (alrededor del 30% en las dos especies para el conjunto
202
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.28. Aportes totales vía desfronde en distintos bosques y sistemas arbustivos (Kg/ha.año).
Especie dominante
Quercus coccifera Cistus ladanifer Ceanhotus megacarpus Artemisia, Salvia matorral mediterráneo
(vario»)
A. decorticans C.laurifolius
Quercus ilex Quercus ilex Quercus ilex Pinus radiata Pinus elliottii Pinus echinata Pinus banksiana Pinus pinaster Pinus nigra Pinus sylvestris Pinus sylvestris Pinus brutia Pinus pinnea Pinus halepensis
bosques templados (vario*)
bosque mixto
pinares (varios)
LOCALIDAD
Sur Francia Extremadura California California varias
Almería Almería
La Castanya Sur Francia Sierra Prades Nueva Zelanda
Tenesse
Almería Almería Pirineos Sierra Béjar Líbano Líbano Sur Francia
USA
Hubbard Brook
varias
Aportes (Kg/ha.año)
65-84 25.5 184 67 121
32 12
115 121 94 34 61 106 28.3 35 17 107 102 63 89 71-104
106
135
. 60-90
AUTOR
Rapp & Lossaint (1981) Núñez (1989) Gray (1983) tf ll
Gray & Schlesinger (1981) PT PT
Verdú (1984) Lossaint & Rapp (1971) LLedó (1990) Baker et al. (1986) Gholz & Fisher (1985) Johnson & Todd (1989) Weber (1987) PT PT Alvera 1990 Sta Regina et ai(1989) Dannaoui (1981) ff ti
Rapp (1967)
Gray & Schlesinger (1981)
Gosz et al. (1972)
Rodin & Bazilevich (1967)
de la cuenca) resultan determinantes en este sentido. Para la plantaciones de
pinar con edades similares a la nuestra que aparecen en la tabla 3.28 se
encuentran aportes de nutrientes más cercanos a los de P. pinaster y P. nigra.
Así, Baker et al. (1986) obtienen 34 Kg/ha.año en Pinus- radiata de 14 años con
una densidad de 2000 pies/ha y Weber (1987) 28.23 Kg/ha.año en plantaciones de
Pinus banksiana de 20 años. Sin embargo los valores obtenidos por Gholz y
Fischer (1985) para Pinus elliottii de 14 años (61.23 Kg/ha.año) son muy
203
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
superiores a los encontrados en el pinar de estudio (35 Kg/ha.año en P.
pinaster y 17 Kg/ha.año en P. nigra).
La mayoría de los sistemas arbustivos que aparecen en la tabla 3.28
corresponden a ecosistemas maduros, de mayor edad y con un porcentaje de
recubrimiento superior al estudiado. En ellos, los retornos de nutrientes son
superiores a los encontrados en las especies arbustivas estudiadas en este
trabajo. Considerando en A. decortlcans y C. laurifolias una cobertura del
1002 obtendríamos unos aportes de unos 60 y 100 Kg/ha.año en la j a ra y
gaznacho respectivamente, valores mucho más cercanos a los descritos en la
bibliografía. De la tabla 3.28 también puede deducirse que los sistemas
arbustivos maduros, con un desfronde muy inferior a bosques viejos de
coniferas y de otro tipo (Gray y Schlesinger 1981), aportan, sin embargo
cantidades muy similares de nutrientes. Ya hemos visto, que esta estrategia
también puede aplicarse en nuestro trabajo para C. lauri folius y A.
decorticans respecto a los pinos a pesar de que éstos aún no han alcanzado la
fase madura.
Los aportes de cada fracción en cada una de las especies se muestran en
la tabla 3.29. Para facilitar las comparaciones se ha añadido la tabla 3.30 en
la que aparece la ordenación de cada nutriente en cada fracción y especie.
El Ca ocupa el primer lugar en la fracción acículas de los pinos,
aportándose con una tasa de 19.7 y 8.8 Kg/ha.año en P. pinaster y P. nigra
respectivamente. Esto resulta lógico dado que las acículas que caen en las
trampas presentan altos contenidos de Ca debido a su elevado grado de
desarrollo. El Ca de las acículas supone el 61% y el 54% de los nutrientes
totales aportados por esta fracción y el 93 y 98% de todo el Ca retornado
anualmente en P. pinaster y P. nigra respectivamente. La hojarasca de C.
laurifolius se caracteriza también por sus elevadas concentraciones de Ca,
significativamente más altas que en las otras especies, lo que resulta
determinante para los aportes de este elemento al suelo. Los 6.54 Kg/ha.año de
Ca aportado por la hojarasca de la j a r a suponen un 62% de los nutrientes
aportados por esta fracción y un 95 % de todo el Ca aportado anualmente por C.
laurifolius.
204
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Tabla 3.29 a. Retorno de nutriente» via desfronde para las distintas fracciones de P.pinaster <Sup. r.
HOJA (Kg/ha)
Jul86/junio87 jul87/ji#i88 jul88/jun89 iul89/seo89
inedia std
FU* (g/ha)
Jut86/junio87 jul87/ j in88 jul88/jun89 ¡ul89/seD89 media std
TOTO (g/ha)
Jut86/junio87 jul87/jun88 ju(88/j"un89 iut89/seo89 inedia std
SSIILL(g/ha>
Jul86/junio87 jul87/jun88 jul88/j'un89 iul89/seo89 media std
UNAS (g/ha)
Jul86/junio87 jul87/jun88 jul88/jun89 iul89/seo89
media std
KPIOD (Kg/ha)
Jul86/junio87 jul87/ji»i88 jul88/jun89 ¡ul89/seo89
media std
TOTAL (Kg/ha)
Jul86/junio87 jul87/jun88 jul88/jun89 ¡ul89/seo89 media std
•
4.54 8.72 5.14 1.69
6.13 2.26
•
233.98 249.30 418.78 329.36 300.69 102.56
•
1695.32 1625.80 918.63
0 1413.25 429.76
II
328.93 47.90 78.96 78.96
151.93 154.07
•
92.50 71.44 30.37
0 64.77 31.60
•
2.26 1.92 1.42
.41 1.87
.42
•
6.89 10.72 6.59 2.10 8.06 2.30
P
.62
.79
.49 1.58
.63
.15
P
16.57 24.30 27.15 12.29 22.67 5.48
P
105.62 73.34 60.29
0 79.75 23.33
P
39.53 5.78 9.45 9.45
18.25 18.52
P
6.60 4.89 2.06
0 4.52 2.29
P
.16
.10
.10
.02
.12
.04
P
.79
.90
.59 1.60
.76
.16
K
1.43 1.82 1.48
.67 1.58
.21
K
31.76 28.22 55.74 38.93 38.57 14.97
t
240.71 530.05 583.86
0 451.54 184.56
K
51.60 7.52
12.38 12.38 23.83 24.17
r
35.06 27.60
1.49 0
21.38 17.631
K
.32
.57
.65
.05
.51
.17
r
1.79 2.41 2.14
.72 2.11
.31
Ca
15.60 25.99 17.58 6.81
19.72 5.52
Ca
96.45 65.69
186.17 204.02 116.10 62.60
Ca
232.21 154.78 115.82
0 167.60 59.25
Ca
74.47 10.85
1.79 1.79
29.04
39.61
Ca
295.43 238.11 102.76
0 212.10 98.93
Ca
.40
.23
.30
.21
.31
.09
Ca
16.30 26.46 17.98 7.01
20.25. 5.45
N>
2.93 4.32 3.24 1.35
3.50 .73
na
37.85 43.04 66.49 46.24
49.13 15.26
Hfl
277.30 205.28 165.00
0 215.86
56.89
"a
24.67 3.61 5.92 5.92
11.40 11.55
NB
28.22 23.17
9.82 0
20.40 9.50
Ng
.34
.25
.24
.05
.28
.06
NB
3.30 4.60 3.49 1.41 3.80
.70
•a
.43
.38
.36
.14
.39
.04
•a
6.57 5.20
12.11 10.64 7.96 3.66
•a
50.35 38.64
31.73 0
40.24 9.41
•a
2.11 .34 .51 .51
.98
.98
•a
1.86 2.23
.64 0
1.58 .83
te
.06
.04
.04
.01
.05
.01
te
.49
.43
.40
.15
.44
.05
to ta l
25.55 42.04 28.29 12.23 31.96
8.84
t o t a l
423.18 415.75 766.44 641.49 535.12 200.36
tota l
2601.51 2627.89 1875.34
0 2368.25 427.07
to ta l
521.31 76.00
109.01 109.01 235.44
248.12
total
459.68 367.45 147.15
0 324.76 160.58
tota l
3.55 3.12 2.75
.75 3.14
.40
tota l
29.56 45.53 31.19 12.98 35.42
8.79
DESFMHK (O/H2)
140.74 228.69 158.26 62.49
175.90 46.55
OESnUNDE (g/B 2 )
3.03 5.72 5.44 4.43 4.73 1.48
DESfMaW (a/ar2)
28.92 19.63 15.87
0 21.47 6.72
DESFIOaDE ( a / * 2 )
1.19 .18 .28 .28 .55 .56
KSFKMDE ( a / " 2 )
2.42 1.90
.83 0
1.71 .81
2 DEsnoBE (g /ar )
33.11 25.50 21.58 4.77 26.73 5.86
DESnOBE ( a / » 2 )
176.17 256.08 180.70 67.28
204.32 44.89
205
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 3.29 b. Retorno de nutrientes vi» desfronde prt las distintas fracciones de P.nigra (Sup. repobiada).
(Kg/ha) Ca Mg Na TOTAL DESFRON0E(g/me)
Jul86/junio87
julS7/jun88
jul88/jun89
¡utB9/sco89
3.89
4.36
5.38 .43
.38
.37
.51
.04
.42
.08
.86
.72 1.01
.13
.87
.14
8.01 7.84
10.64 .85
8.83 1.57
1.63 1.42 2.01
.19 1.69
.30
10 07 20 ,01 .12 07
14.87 14.77 19.76 1.65
16.47 2.85
95.92 90.53 126.58 11.73 104.34 19.45 std
4.54
.76
FU» (g/ha) Ca Mg Na TOTAL DESFRONDE(g/m2)
Jul86/ junio87 ju l87/ junS8 julS8/junB9 iul89/seo89
97.88 26.08 50.39 9.80
4.92 1.08 3.05
.64
13.53 2.69 7.20 1.34
35.08 5.98
15.15 2.15
16.57 15.71 7.09 1.49
3.98 .73
1.88 .30
171.95 52.27 84.76 15.72
1.44 .31 .79 .15
std
58.11
36.52
3.02
1.92
7.81
5.45
18.74
14.87
13.12
5.24
2.19
1.65
102.99
61.89 .57
«UTO (g/ha) Ca Mg Na TOTAL DESFROMOEÍg/m')
Jul86/junio87
jul87/jun88
jut88/jun89
¡ul89/sco89
705.51 0 0 0
235.17 407.33
81.35 0 0 0
27.12 46.97
453.95 0 0 0
151.32 262.09
191.13 0 0 0
63.71 110.35
147.95 0 0 0
49.32 85.42
30.75 0 0 0
10.25 17.76
1610.65 0 0 0
536.88 929.91
22.19 2.91 0 0
8.37 12.06 std
SEMUXg/ha) Ca Mg Na TOTAL DESFRON0E(g/m')
Jul86/junio87
jutS7/jun8S
jul88/jun89
iul89/sep69
10.68 8.00
0 0
6.23 5.56
1.29 .97
0 0
.75
.67
1.66 1.26
0 0
.98
.87
.30
.59
0 0
.07
.04
0 0
14.51 10.87
0 0
.04
.03 0 0
std .47
.42
.04
.04
8.46
7.55
.02
.02
(g/ha) Ca Mg Na TOTAL DESFRONOEÍg/nT)
Jul86/junio87
jul87/jun88
jul88/junS9
iul89/set>89
JulS6/junio87
jul87/jun88
jutB8/jun89
iul89/seo89
11.18 31.02
132.52 4.61
.64 1.80 7.68
.27
1.43 3.95
16.91 .59
2.76 7.66
32.79 1.14
1.63 4.56
19.42 .68
.12
.33 1.38
.05
17.76 49.32
210.70 7.33
.13
.40 1.71 JJ6
•adía s td
KFtODU (Kg/ha)
58.24 65.09
N
3.37 3.78
P
7.43 8.31
K
14.40 16.11
Ca
8.54 9.54
Mg
.61
.68
Na
92.59 .75 103.50 .84
TOTAL DESFRONOE(g/m2)
.814
.034
.050
.010
.300
.446
.088
.002
.003
.001
.031
.049
.469
.004
.007 ,001
.160
.268
.226
.006
.015
.002
.082
.125
.165
.016
.007
.001
.063
.089
035 ,001 002 ,000
.012 ,019
1.80 .06 .08 .02
.65
.99
23.66 3.25 .79 .15
9.23 12.55
•odia
std
TOTAL (Kg/ha) Ca Mg Na TOTAL DESFROWEíg/i»2)
JulB6/junio87
jul87/jun88
jul88/jun89
¡ut89/seo89
4.72
4.42
5.56
.44
,47 .37 .52 .04
1.33 .73
1.04 .14
8.24 7.85
10.69
.85
1.80 1.44 2.04
.20
14 07 21 01 .14 07
16.69 14.88 20.06
1.68 17.21 2.62
119.72 94.13 129.11 11.94
114.32 18.10 std
4.90
.59
.45
.07
1.03
.30
8.93
1.54
1.76
.30
206
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Tabla 3.29.C. Retorno de nutrientes
HOJA(Kg/ha) H P
Jul86/jun87 1.57 .34 jul87/jun88 2.19 .43 jul88/jun89 1.68 .30 ¡ul89/seo89 1^47 .27 •edia 1.81 .36 ds .33 .07
FLOR íg/ha) N P
Jul86/jun87 jul87/jun88 jul88/jun89 ¡ul89/seD89 •edia ds
116.11 379.57 377.42 1433.80 291.03 151.49
14.30 62.52 39.88 69.88 38.90 24.12
FRUTO (g/ha) N P
Jul86/jun87 jul87/jun88 jul88/jun89 iul89/seo89 •edia ds
110.21 70.51 114.59 71.80 98.44 24.28
6.83 4.97 8.66 5.09 6.82 1.85
RAMAS (g/ha) H P
Jul86/jun87 jul87/jun88 jul88/jun89 ¡ul89/seis89 •edia ds
122.92 77.05 104.30 20.49 101.42 23.07
8.27 4.98 6.52 1.40 6.59 1.65
REPHD.(9/ha) N P
Jul86/jun87 226.33 21.13 jul87/jun88 450.08 67.49 j"ul88/jun89 492.01 48.54 iul89/sep89 1505.60 74.97 •edia 389.47 45.72 ds 142.83 23.31
TOTAL (ka/ha) N P
Jul86/jun87 1.92 .37 jul87/jun88 2.71 .50 jul88/jun89 2.28 .36 iul89/sep89 LOO J l •edia 2.30 .41 ds .40 .08
via desfronde en las diferentes f
K Ca Mg
.59 1.08 .58 .46 .75 .29
5.79 7.52 6.31 6.49 6.54 .89
.92 1.19 .97 .98
1.02 .14
K Ca Mg
99.91 338.76 286.63 484.65 241.77 125.59
38.40 58.32 45.88 81.79 47.53 10.06
18.62 73.24 57.16 112.41 49.67 28.07
K Ca Mg
15.86 4.74 12.25 6.97 10.95 5.67
59.61 12.55 36.69 29.63 36.28 23.53
7.80 6.27 13.90 4.25 9.32 4.04
K Ca Mg
11.79 7.87 9.45 2.11 9.70 1.97
292.33 168.23 242.35 48.30 234.30 62.44
14.83 8.48 12.03 2.51 11.78 3.18
JC Ca Mg
115.77 343.49 298.88 491.62 252.71 120.68
98.03 70.86 82.57 111.42 83.82 13.63
26.43 79.51 71.06 116.66
59 28.52
K Ca Mg
.72 1.44 .89 .96
1.01 .37
6.18 7.76 6.63 6.65 6.86 .81
.96 1.27 1.05 1.10 1.10 .16
Desfrondo
facciones de Cistus laurifolius.
Na total DESFRONOEíg/m )
.062
.039
.036
.034
.046
.014
9.27 12.45 9.88 9.71 10.53 1.69
35.83 43.38 38.37 38.71 39.19 3.84
Na total DESFR0N0E(g/m )
1.26 4.54 4.16 12.73 3.32 1.80
288.60 916.95 811.14 2195.26 672.23 336.42
1.48 4.70 3.79 6.95 3.32 1.66
Na total 0£SFRONDE(g/ra')
1.61 .87 1.78 .87 1.42 .48
201.92 99.91 187.86 118.61 163.23 55.29
1.73 1.40 3 .95
2.04 .84
Na total DESFROND£(g/nT)
2.36 1.26 1.87 .40
1.83 .55
452.50 267.87 376.51 75.21
365.63 92.79
1.85 1.10 1.48 .31
1.48 .38
Na total 0ESFR0ND£(g/nT)
2.85 5.41 5.94 13.60 4.73 1.65
490.54 1016.84 999.00 2313.87 835.46 298.84
3.21 6.09 6.78 7.89 5.36 1.89
Na total DESFRONDE(g/m')
.07
.05
.04
.05
.05
.01
10.21 13.73 11.25 12.10 11.73 1.81
40.91 52.12 50.19 47.39 47.74 5.99
207
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
la 3.29.d. Retorno de nutrientes vía desfronde para las distintas
decorticans. rab A. decorticans
fracciones de
•OÍA (Kg/ha)
Jul86/jun87 jgl87/jun88 jul88/jun89 iul89/seo89 •odia std
nm (kg/tia)
Jul86/jun87 jul87/jun88 julS8/jun89 iul89/seo89
std
mnO (a /ha)
Jul86/jun87 jut87/jun88 jut88/jun89 iul89/seo89
•adía std
SEMILLA (g/ha
JulS6/jun87 jul87/jun88 jul88/jun89 ¡ul89/seo89
aadia std
IAMAS (o/ha)
Jul86/jun87 jul87/jun88 jul88/jun89 iut89/seo89 •adía std
BEPtO (fcs/ha)
Jul86/jun87 jul87/jun88 jul88/jun89 iul89/seo89
•adía std
TOTAL (Kg/ha)
lili
1I
II
sadia std
N
15.81 7.02
12.31 6.85
11.71 4.43
N
2.51 3.94 3.85 1.70 3.43
.80
N
288.40 2
1928.88 66.24
739.76 1039.72
N
651.72 49.40
1217.58 54.60
639.57 584.18
N
1372.45 425.91
2799.97 131.88
1532.78 1195.12
M
3.45 3.99 7.00 1.82
4.81 1.91
M
20.63 11.43 22.11 8.80
18.06 5.79
P
.72
.21
.45
.22
.46
.25
P
.11
.20
.13
.06
.14
.05
P
2.57 .02
17.15 .59
6.58 9.24
P
42.14 3.19
78.72 3.53
41.35 37.77
P
52.45 16.17
109.98 5.04
59.53 47.30
P
.15
.20
.22
.06
.19
.04
P
.92
.43
.78
.29
.71
.26
K
4.83 1.53 2.21 1.25 2.86 1.74
K
.68 1.54
.47
.42
.90
.57
K
198.67 1.38
1328.78 45.63
509.61 716.25
K
131.29 9.95
245.30 11
128.85 117.69
K
61.19 35.75
151.30 5.88
82.75 60.71
K
1.01 1.55 2.04
.48 1.54
.51
K
5.90 3.12 4.40 1.74 4.47 1.39
Ca
7.41 3.31 6.08 4.08 5.60 2.09
Ca
.69 1.29 1.11
.49 1.03
.31
Ca
88.13 .61
589.38 20.24
226.04 317.69
Ca
0 0
22.30 1
7.43 12.87
Ca
97.03 29.52
253.11 9.32
126.55 114.68
Ca
.77 1.29 1.72
.52 1.26
.47
Ca
8.28 4.63 8.05 4.61 6.99 2.05
M3
1.79 .58
1.25 .78
1.21 .61
Ng
.18
.26
.32
.14
.25
.07
Mg
32.04 .22
214.32 7.36
82.19 115.53
Mg
14.91 1.13
27.88 1.25
14.64 13.37
Mg
74.31 23.50
160.78 7.14
86.20 69.41
Mg
.23
.26
.56
.15
.35
.18
Mg
2.09 .86
1.97 .93
1.64 .68
Na
.127
.028
.077
.045
.077
.049
Na
.010
.029
.021
.011
.020
.010
Na
6.41 .04
42.86 1.47
16.44 23.11
Na
1.14 .08
2.12 .10
1.11 1.02
Na
52.45 6.33
15.98 .64
24.92 24.33
Na
.018
.029
.066
.012
.038
.025
Na
.20
.06
.16
.06
.14
.07
TOTAL
30.68 12.68 22.37 13.22 21.91
9.01
TOTAL
4.18 7.25 5.90 2.83 5.78 1.53
TOTAL
6 1 6 . 2 2
4.27 4121.37
141.53 1580.62 2221.53
TOTAL
841.20 63.75
1593.89 71.48
832.95 765.10
TOTAL
1709.88 537.18
3491.12 , 159.90 1912.73 1487.38
TOTAL
5.64 7.31
11.61 3.04 8.19 3.08
TOTAL
3 8 . 0 3
2 0 . 5 3
37.47 16.43 32.01
9.95
DESFRONDE(g/m2
73.42 33.96 59.21 37.04 55.53 19.99
DESFRÓN0E(g/m2
9.14 16.90 15.21 6.68 13.75 4.08
DESFRONDE(J/m2
4 .03
26.80 .93
10.28 14.45
0ESFRON0E(g/m2
1.19 .09
2.23 .10
1.17 1.07
DESFRONDE(J/m2
8.75 2.70 20.11
.84 10.52 8.84
DESFRONDE(g/m2
14.33 17.01 44.23 7.70 25.19 16.54
0ESFRON0E(g/n2
96.56 53.96
129.59 46.21 93.37 37.92
208
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
Tabla 3.30. Ordenación de los distintos nutrientes según su aporte medio anual vía desfronde para cada fracción y especie estudiada.
P.pinaster.
ACICULAS C a > N > M g > K > P > N a REPR. N > K > C a > M g > P > N a RAMAS Ca > N > K-Mg > P > Na
P.nigra. ACICULAS Ca > N > Mg > K > P > Na REPR. N > K > Ca > Mg > P > Na RAMAS N > Ca > Mg-K > P > Na
A. decorticans.
C.laurifolius
HOJAS N » Ca > K > Mg > P > Na REPR. N > K > C a > M g > P > N a RAMAS N > Ca > Mg-K > P > Na
HOJAS CA»N > Mg > K > P > Na REPR. N > K > C a > M g > P > N a RAMAS Ca > N > Mg > K > P > Na
El aporte más alto de N vía desfronde más elevada corresponde a A.
decorticans sea cual sea la fracción considerada. Es evidente la importancia
que a este respecto puede tener su condición de fijadora de nitrógeno. El
nitrógeno foliar del desfronde supone un 537. del total de nutrientes aportados
por las hojas y un 65 7. del N anualmente aportado por esta especie. Las
flores, pese a caer en un periodo de tiempo muy restringido aportan 3.94
Kg/ha.año valor que incluso supera a los aportes totales de N en la j a ra (2.3
Kg/ha.año) y es muy similar a los de P. nigra (4.9 Kg/ha.año). La flor de A.
decorticans supone además una proporción nada despreciable a los aportes
totales de este elemento en el gaznacho (19%). Las ramillas y tallos de A.
decorticans contribuyen con 1 Kg/ha-año. Los aportes de K tienen una
contribución relativa destacable en los órganos reproductivos más que en otras
fracciones debido a sus concentraciones elevadas en este tipo de tejidos
aportándose en cantidades que oscilan entre 0.16 Kg/ha. año y los 1.26
209
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Kg/ha.año en A. decorticans.
En la tabla 3.31 se han recogido los aportes medios de nutrientes en
algunos sistemas forestales y arbustivos. En las repoblaciones jóvenes de
pinos se ha mantenido la individualidad de los datos debido al bajo número de
estaciones y la heterogeneidad de los valores entre ellas. Los pinares maduros
corresponden básicamente a pinares mediterráneos.
Los aportes de P.pinaster son muy similares a los encontrados por Weber
(1987) y Baker et al. (1986) para plantaciones de Pinas banksiana de 22 años
y de Pinus radiata de 14. Esta similitud puede deberse en buena parte al
desfronde anual muy similar entre las t res repoblaciones. Las mayores
diferencias se refieren al P cuyos aportes por parte de P.pinaster son
ligeramente inferiores mientras que para el Ca resultan superiores. Las
cantidades de nutrientes aportadas por P. nigra resultan inferiores a las
repoblaciones anteriormente citadas principalmente debido al menor peso de
desfronde en P. nigra. No obstante, el Ca, K y Mg aportado por el laricio es
similar al de la repoblación de Pinus banksiana estudiado por Weber (1987).
Suponiendo una cobertura de suelo del 1007. en los arbustos estudiados, el
desfronde de A. decorticans aportaría al suelo cantidades ligeramente
superiores de N que los matorrales mediterráneos maduros y bastante más
superiores a los de otros sistemas como pinares o encinares. Este elevado
retorno de N resulta lógico teniendo en cuenta la condición de fijadora de
esta especie y sus elevadas concentraciones tanto en órganos vivos como en
hojarasca. Las cantidades aportadas de P y K son similares a los valores
medios para estos nutrientes encontrados en los sistemas arbustivos y sobre
todo a los encinares. La mayor diferencia de A. decorticans respecto a
matorrales y encinares estriba en el Ca, muy inferior en A. decorticans. Tanto
las bajas concentraciones de este elemento en hojarasca como la escasa
contribución al desfronde de la fracción gruesa pueden influir en este
aspecto.
La contribución de C. laurifolius aún suponiendo una cobertura del 100%
sigue siendo bastante inferior a A. decorticans tanto en peso como en
210
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
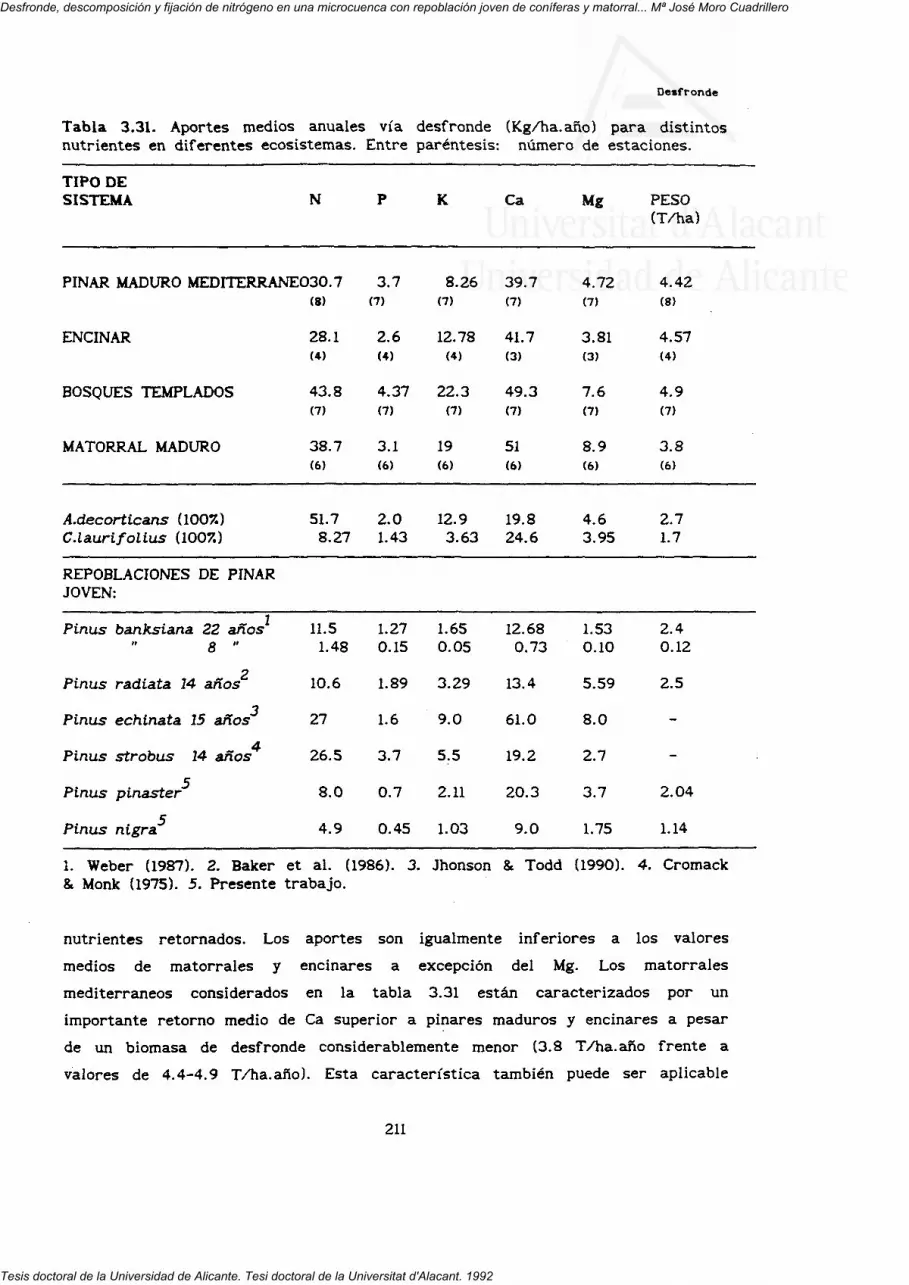
Detfronde
Tabla 3.31. Aportes medios anuales vía desfronde (Kg/ha.año) para distintos nutrientes en diferentes ecosistemas. Entre paréntesis: número de estaciones.
TIPO DE SISTEMA N
PINAR MADURO MEDITERRANE030.7
ENCINAR
BOSQUES TEMPLADOS
MATORRAL MADURO
A.decorticans (100%) C.laurifollus (1007.)
REPOBLACIONES DE PINAR JOVEN:
Pinus banksiana 22 años 8 "
2 Pinus radiata 14 años
3 Pinus echínata 15 años
4 Pinus strobus 14 años
Pinus pinaster
Pinus nigra
(8)
28.1 (4)
43.8 (7)
38.7 (6)
51.7 8.27
11.5 1.48
10.6
27
26.5
8.0
4.9
P
3.7 (7)
2.6 (4)
4.37 (7)
3.1 (6)
2.0 1.43
1.27 0.15
1.89
1.6
3.7
0.7
0.45
K
8.26 (7)
12.78 (4)
22.3 (7)
19 (6)
12.9 3.63
1.65 0.05
3.29
9.0
5.5
2.11
1.03
Ca
39.7 (7)
41.7 (3)
49.3 (7)
51 (6)
19.8 24.6
12.68 0.73
13.4
61.0
19.2
20.3
9.0
Mg
4.72 (7)
3.81 (3)
7.6 (7)
8.9 (6)
4.6 3.95
1.53 0.10
5.59
8.0
2.7
3.7
1.75
PESO (T/ha)
4.42 (8)
4.57 (4)
4.9 (7)
3.8 (6)
2.7 1.7
2.4 0.12
2.5
-
-
2.04
1.14
1. Weber (1987). 2. Baker et al. (1986). 3. Jhonson & Todd (1990). 4. Cromack &. Monk (1975). 5. Presente trabajo.
nutrientes retornados. Los aportes son igualmente inferiores a los valores
medios de matorrales y encinares a excepción del Mg. Los matorrales
mediterráneos considerados en la tabla 3.31 están caracterizados por un
importante retorno medio de Ca superior a pinares maduros y encinares a pesar
de un biomasa de desfronde considerablemente menor (3.8 T/ha. año frente a
valores de 4.4-4.9 T/ha.año). Esta característica también puede ser aplicable
211
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en la ja ra . Suponiendo un desfronde similar respecto a los demás sistemas, los
aportes de Ca en el matorral mediterráneo incluyendo a C. laurifolius
llegarían a ser del orden de un 25-357. superiores a la media de pinares,
encinares y caducifolios. Parece, por tanto, que la circulación de Ca en
sistemas arbustivos mediterráneos está menos restringida que en el resto de
los sistemas.
La circulación de P en matorrales es inferior a pinares mediterráneos y a
encinares. Sin embargo normalizando la producción de desfronde en estos tres
sistemas encontraríamos unos aportes similares en todas ellas, que serían a su
vez inferiores a los de bosques caducifolios.
Para comparar de una manera más precisa el comportamiento de los
distintos ecosistemas terrestres respecto al nivel de circulación de
nutrientes vía desfronde se ha considerado oportuno aplicar el índice de
eficiencia de Vitousek (1982) para aquellos nutrientes que utilicen
preferentemente el flujo de desfronde para retornar al suelo (N, P y Ca).
Eficiencia en el uso de los nutrientes.
Vitousek (1982) propone un índice de eficiencia en el uso de nutrientes
que consiste en calcular la cantidad de materia seca caída por unidad de
nutriente perdido en el desfronde. Este índice permite utilizar de forma
comparable la información sobre el retorno de materia seca y nutrientes por
esta vía y con él pueden establecerse en qué medida la cantidad de desfronde
está acoplada con la circulación de los distintos nutrientes en la hojarasca.
Los resultados que obtenía este autor indicaban que la circulación de N está
más estrechamente ligada a la cantidad de hojarasca que la del P y Ca. La
utilización del índice de eficiencia es ya un clásico en cualquier estudio
referente al retorno de elementos por desfronde. Sobre la representación
básica de las casi 100 estaciones correspondientes a diferentes ecosistemas
que apareció en el trabajo de Vitousek se han ido añadiendo poco a poco más
puntos de los que varios corresponden a sistemas mediterráneos (Verdú 1984,
Núñez 1989, Lledó 1990).
212
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
En las figuras 3.33 a, b y c, se han representado las eficiencias de uso
respecto al N, P y Ca frente a la circulación de estos nutrientes en el
desfronde de las cuatro especies estudiadas y de distintos tipos de
ecosistemas. Los datos de coniferas utilizadas corresponden básicamente a
pinares mediterráneos y a plantaciones jóvenes (de la zona templada). Los
datos de caducifolios corresponden tanto a la zona mediterránea como a la
templada. Los valores de la zona tropical están tomados de la recopilación de
Bruijnzeel (1985), los de fijadores de Vitousek (1982), Wheeler et al. (1987)
y Schwintzer (1984), los matorrales de Gray y Schlesinger (1981), Merino
(1986), Núñez (1989) y Colé y Rapp (1981).
Respecto al N, (fig. 3.33 a) P. pinaster, P. nigra y C. laurifolius
resultan ser especies muy eficientes con unas tasas que difieren muy poco
entre ellas. Por el contrario A. decorticans presenta una eficacia muy
inferior pero que queda totalmente encajada dentro del rango de bosques o
sistemas dominados por fijadores simbiontes de N2. Estos resultados vienen a
coincidir con los obtenidos para la retraslocación, lo que resulta lógico ya
que la reabsorción sería uno de los principales mecanismos para controlar o
reducir el retorno de N aumentando así la eficiencia en el uso de éste
(Vitousek 1982). Los pinos estudiados se sitúan entre los más eficientes
dentro de las coniferas representadas. En la figura 3.33. c se aprecian dos
grupos dentro de los sistemas arbustivos mediterráneos. El grupo de mayor
eficiencia corresponde a matorrales de cistáceas (Núñez 1989, Merino 1986,
presente trabajo) además de un matorral australiano. El segundo grupo de una
eficiencia considerablemente menor está formado por sistemas arbustivos del
tipo del chaparral, garriga y brezales, estos últimos con eficiencias
prácticamente idénticas a los encinares. Todos los sistemas fijadores de
nitrógeno poseen eficiencia muy bajas y similares independientemente de cual
sea la tasa de circulación de N que varía en un rango considerable.
Respecto a la eficiencia en el uso del P existe una diferencia clara
entre el pinar y el matorral estudiado en este trabajo. P. pinaster y P.
nigra poseen eficacias altas que resultan en la mayoría de los casos
superiores a las de otras coniferas representadas en la figura 3.33. b. Por el
contrario A. decorticans y C. laurifolius se sitúan entre las menos eficientes
213
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
300
250
200
150
100
50
Peso seco:N en dasíron
• ) -
Pn •+•
ci • a
X 0 4 , » a
V x* +
/W*
de
a ? a
*
* 4 » » • • # * »
0
* * I
*
) 20
* conifera*
° caduclfolloa
40 60 80 100
Kg/ha.año N
matorral medlt
encinares
120
fijadoras
tropicales
140
3500
3000
2500
2000
1500
1000
500
°930 39C0: P
4 «. ><
-PnPp -, 4
1
9fl
4
•
desfronde
. a
• 4 *. . *
1
ft X
1 -
•
1
* X 4
*
coniferas a encinares
4 8 8
Kg/ha.año P
* matorral medt
* tropicales
10
* caduclfolloa
300
250
200
150
100
50
Peso seco: Ca en desfronde
50 100 150
Kg/ha.año Ca
O coniferas + caduclfolloa * mator/enclna
200
tropicales
12
• o
+ ' ?! H ° »
Pit + Pp
-Cl n a*a
* a a * * +
4 Q °
. *
t
i -1 1 1
250
Figura 3.33. Eficiencias en el uso del N, P y Ca según el modelo de Vitousek
(1982) para distintos tipos de ecosistemas terres t res .
214
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde
de todos los matorrales mediterráneos estudiados asociados con otro jaral
estudiado por Núñez (1989) y el chaparral de Ceanothus. Los niveles de
eficiencia de este grupo son muy similares a los encontrados en encinares.
Existe un pequeño grupo de sistemas arbustivos que posee, sin embargo
eficiencias que superan incluso a las de las coniferas más eficientes. Este
grupo esta formado por garriga y brezal. Resulta curioso que estos dos tipos
de vegetación sean precisamente las que menor eficacia presentaban respecto al
N.
Las cuatro especies estudiadas presentan eficiencias de uso de Ca más
similares que las correspondientes al P. A. decorticans y P. nigra resultan
las más eficientes, P. pinaster ocupa una posición intermedia mientras que C.
laurtfolius posee el valor más bajo. Los resultados de retraslocación para
esta especie ya indicaban un uso no demasiado ahorrativo del P por parte de la
jara.(ver apartado 3.3.6).
El rango de eficiencia encontrado en distintas coniferas es amplio. P.
pinaster y P. nigra se sitúan en una posición intermedia. Tanto matorrales
como encinares están caracterizados a la vista de la figura 3.33.C por
eficiencias bajas y muy similares, sea cual sea el nivel de circulación de Ca
en hojarasca exceptuando A. decorticans y un matorral australiano que poseen
eficiencias algo superiores al resto de sistemas arbustivo.
215
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
216
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 4 Retorno de nutrientes.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

4.1. INTRODUCCIÓN.
Retorno de nutrientes
El desfronde y la lixiviación mediante la trascolación neta constituyen
las dos principales vías de retorno de nutrientes al suelo procedentes de la
biomasa aérea del bosque. La escorrentía cortical contiene soluciones muy
concentradas de nutrientes procedentes del lavado de las cortezas pero éstas
tienen un área de influencia pequeña restringiéndose a las zonas de suelo más
próximas al tronco. La deposición de aerosoles o partículas sobre la
vegetación durante los períodos secos es lavada por el agua de lluvia pero no
constituye un retorno propiamente dicho sino una entrada externa.
El retorno vía desfronde varía estacionalmente dependiendo del tipo de
bosque y de la época de la abcisión (Waring y Schlesinger 1985). Así mismo, el
mayor o menor desarrollo de los mecanismos de retraslocación inmediatos a la
caída fenológica suponen modificaciones en las restitución de algunos
elementos al suelo (básicamente N y P). El contenido en nutrientes de la
fracción foliar del desfronde es fundamental para la determinación del retorno
vía detritus dada su importancia porcentual en este flujo sobre todo en
sistemas jóvenes como el que nos ocupa. La trascolación varía dependiendo de
la composición específica y del régimen de lluvias. En zonas mediterráneas la
variación y magnitud del flujo de nutrientes vía trascolación está
condicionada a la distribución irregular de precipitaciones en esta zona
concentradas esencialmente en primavera y otoño. i
En general, el desfronde es la vía más importante desde el punto de vista
cuantitativo en el retorno de nutrientes como el N, P, Ca y Mg mientras que el
lixiviado es el responsable principal del retorno de K (Waring y Schlesinger
1985, Colé y Rapp 1981). El Na tiene origen predominantemente externo que
entra al sistema entre un 40-100% por vía seca (Parker 1983)
Conocido ya el aporte de nutrientes que supone el flujo del desfronde
(capítulo anterior) abordaremos en una primera parte de este capítulo el
estudio del retorno vía trascolación neta estudiada en esta misma cuenca por
Domingo (1991). La segunda parte interelaciona ambos flujos, analizando el
217
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
retorno y relacionándolo con las entradas al sistema mediante la deposición
global. Otro objetivo del capítulo es, discriminar en la medida de lo posible
qué parte de nutrientes retornados vía trascolación tienen origen fuera o
dentro del sistema. Para ello, dado que no se disponía de ningún tipo de
aproximación directa o indirecta a partir de estudios previos (Domingo 1991),
se ha aplicado el modelo de Lovett y Lindberg (1984), para el cálculo de las
tasas de deposición seca e intercambio en las cuatro especies.
4.2 Retorno de nutrientes por trascolación neta.
4.2.1. Características de la trascolación en las especies estudiadas.
Los principales aspectos de la trascolación en la cuenca han sido
estudiados por Domingo (1991) y Domingo et al. (1991). Entre ellos se contaban
el evaluar la cantidad de lluvia que es canalizada por este flujo y el papel
de la cubierta de cada una de las especies mayoritarias en modificar la
química de la precipitación. Un último aspecto consistía en la cuantificación
de los aportes de nutrientes al suelo por esta vía. En las tablas 4.1 a 4.3 se
resumen algunas de las características de la trascolación en las principales
especies. Se ha seleccionado la información que puede resultar de mayor
interés en relación al papel de la vegetación en el aporte de nutrientes:
concentraciones en trascolación neta (tabla 4.1), coeficientes de
enriquecimiento iónico (tabla 4.2) y aportes brutos (tabla 4.3).
El estudio de Domingo (1991) pone de manifiesto diferencias específicas
importantes respecto a la capacidad de la vegetación para modificar el agua de
lluvia. C. laurifolius es la especie que la modifica en menor medida, los
pinos se encuentran en una posición intermedia, mientras que las
modificaciones más importantes las sufre el agua de precipitación al atravesar
la cubierta de A. decorticans. También se observan diferencias destacables
entre el pinar y el matorral con respecto a algunos iones. Así, existen
tendencias opuestas respecto al P-P04 y N-N03. Mientras que el pinar posee
coeficientes de enriquecimiento negativos para los fosfatos, el comportamiento
inverso se observa en el matorral. Los nitratos son retenidos en la cubierta
218
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
l a f i l a 4 . 1 . concentraciones netas en la trascolación, según el modelo (C): TRU = ( [TR] * VTB - CDG] * (Vn- - V p r ) ) / VT
Ha* K* Ca*"2 Mg*2 <HH4)+ H*
0*eq/O " Deposición global 25.99 13.83 111.54 30.26 36.86 2.34
P. pinaster 37.31 18.79 188.27 51.44 -8.40 -2.20
P. nigra 47.33 21.60 208.46 41.64 -11.69 -1.96
A. decorticans 47.19 107.94 156.23 55.70 15.84 -1.99
C. laurifolius 15.81 69.46 63.50 31.76 -18.45 -.26
(NOj)" (S04 )"2 Cl" <POA>"3 (HCOj)"
(Aeq/l)
Deposición global 17.67 52.89 30.15 2.39 136.13
P. pinaster 32.27 49.61 69.15 -1.60 88.36
P. nigra 15.66 48.22 56.98 -2.08 102.11
A. decorticans -16.57 49.54 108.79 7.20 101.76
C. laurifol ius -17.54 15.39 90.03 11.96 -50.09
T a b l a 4 . 2 . Tasas de enriquecimiento relativo (E) , según el modelo (8 ) , en la trascolación de cada especie. E * C(Aporte»)TR . <Aportes)0G] / (Aportes)DG
Ha* K* Ca+2 Mg*2 r n n . i + u* run^" rcn %*2 r l - ,„„ , - 3 •< /¿eq/0 — -- - ( m g / l )
(HH4) H (MOj)" CS04rZ Cl" ( P 0 4 ) ' 3 (HCOj)" org.M t . » . « « » » W » » M M I l I I l l t I 1 3 J l l I l J J , i i , l l : 1 , s , M , 1 M a M 1 J K B . , a a i l I M 1 M 5 ¡ M M J M I S i . . . . „ „ j .
P, Pjnaster 1.22 1.15 1.44 1.45 -0.21 -0.82 1.55 0.79 1.96 -0.59 0.54 0.06
p . m'qrc i.*i i . i° 1.« 1.04 -0.39 -o.83 0.62 o.65 1.47 -0.86 0.51 0.43
A. decorticans. 1.33 5.86 1.02 1.35 0.28 -0.69 -0.75 0.67 2.69 2.23 0.52 1.04
C. taurifoHua 0.32 3.18 0.30 0.61 -0.40 -0.15 -0.71 0.12 1.86 3.16 -0.31 -0.68 I 3 S 3 3 S 3 3 S
219
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 4 . 3 . APORTES BRUTOS (por hectárea IDEAL) EN TRASCOLACIOH
(nra) - ---Ka/ha - -especia voluean «a* K* Ca+Z Mg*Z (*H4) H* (MOj)" ( » 4 ) " 2 C l " (TO4 ) "3 (HCOjf a org
P. pinastcr 865.80 13.36 11.73 54.86 9.06 5.29 4.166-3 28.20 45.85 31.84 3.12E-1 129.13 2.12 P. n ig ra 849.24 14.52 11.95 55.23 7.55 4.09 4.056-3 17.92 42.11 26.60 1.09E-1 126.31 2.85
A. decorticana 762.75 14.03 37.41 45.42 8.71 8.59 7.37E-3 2.72 42.63 39.72 2.47E+0 127.41 4.08 C. l a u r i f o l i u s 652.16 7.96 22.77 29.20 5.96 4.05 2.02E-2 3.15 28.57 30.81 3.18E-0 57.72 .64
3 n i t a u i i i t i M u u n t i t n s t s s x i i i i i i i i u a j t s s i s i s i i n u i t u t u t t u a i i i s i n x i i i i i t i t s s i M t u i i i i t i u a a t u x f 3 i t u s s x 3 ,
Deposición Global 1007.77 6.02 5.45 22.53 3.71 6.70 2.36E-2 11.04 25.60 10.77 7.64E-1 83.71 2.51 s t u u i i a i j n u i u u u i H t i n n u f a i i m t i t M H i u H a H U i a i i a u u i s g a s m a i n t u i u t u i a w M u u t i x i u u i u i i i i u a ,
Aportas bruto* (Kg/ha) en todo el periodo de nuestreo (Abr88-SepS9) por por TRASC0LACI0N suponiendo que toda la hectárea estuviese cubierta homogéneamente por cada una de las especies separadamente.
arbustiva mientras que se lixivian, aunque en pequeñas cantidades en el pinar.
El agua de trascolación se enriquece con K en todas las especies aunque lo
hace con mucha mayor intensidad en las especies arbustivas. La j a ra es la
única especie en la que se ha detectado una disminución ligera de Ph
manifestada por la tasa de enriquecimiento ligeramente positiva para los
protones. El enriquecimiento de NH4 y de nitrógeno orgánico observado en A.
decorticans puede ser atribuíble a la contribución de flora o fauna epífita
(Domingo et al. 1991).
Respecto a los aportes de nutrientes a la cuenca, Domingo (1991) comenta
que son importantes y superiores a otras zonas de la vertiente mediterránea
sobre todo para aquellos de origen mayoritariamente externo en la cuenca
"Nacimiento", básicamente C03H y Ca así como S04, Cl y Na, lo que demuestra,
según este autor el importante papel de la cubierta vegetal como captadora de
nutrientes de origen atmosférico. Suponiendo que la cuenca estuviera sólo
cubierta por el matorral, el suelo recibiría menos aportes de nitratos y
mayores de fosfato y potasio que si lo estuviera con pinar. En este último
caso el suelo recibiría por el contrario cantidades sensiblemente mayores de
bicarbonato y calcio, lo que parece indicar una mayor y efectividad en la
captación de deposición seca por parte del pinar. (Domingo 1991).
220
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
4.2.2. Tasas de deposición seca e intercambio en el flujo de trascolación
neta: Aplicación del modelo de Lovett y Lindberg.
Caracter ís t icas y limitaciones del modelo.
La cantidad de precipitación es un factor determinante en la composición
química de la trascolación (Kazda 1987, Bellot 1989, Domingo 1991). Además, la
duración del período seco que precede cada precipitación influye también de
manera importante en la química del agua que atraviesa la cubierta forestal
(Lovett y Linberg 1984, Bellot 1989) sobre todo en zonas cercanas a fuentes
emisoras. Basándose en estas dos premisas, Lovett y Lindberg (1984) construyen
un modelo simple basado en regresiones múltiples para calcular las tasas de
deposición seca y de los procesos de intercambio a nivel de copas para cada
elemento presente en la trascolación y escorrentía cortical pudiéndose
escribir como sigue:
(TC+EC)= bl*D + b2* P
donde TC+EC son los aportes netos por trascolación y escorrentía cortical, bl
es la tasa media de deposición seca y b2 la tasa media de intercambio. D es el
número de días secos entre eventos lluviosos y P la cantidad de precipitación
(en mm).
, El modelo considera el papel de la vegetación en la captación de
deposición seca y en distintos procesos de absorción, intercambio o exudación
de elementos que pueden ocurrir a nivel de sus superficies o en la fauna o
flora epífita asociadas a éstas.
El modelo presupone que los eventos lluviosos son independientes y que
una determinada lluvia lava totalmente las copas. También se asume que el
intercambio a nivel de copas ocurre solo en solución acuosa de una forma
proporcional al flujo de agua. Es necesario, que se reduzca al mínimo la
contaminación de los colectores de polvo procedente de suelo o de restos de
material vegetal. Para ello es recomendable utilizar dispositivos de apertura
221
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
automática en períodos ie lluvia (Bellot 1989). La simplificación de los
procesos de interacción i.uvia- cubierta vegetal en un modelo tan sencillo
hace que aparezcan una serie de problemas. Lovett y Linberg (1984) citan entre
los principales el hecho de que el material lixiviado en períodos secos (por
ejemplo: exudados vegetales) sea considerado como deposición seca o que
deposición seca que se va lavando de una forma lenta durante la lluvia afecte
a ambos coeficientes. Además, las tasas bl y b2 son tasas netas, es decir,
resultado de procesos que a veces pueden ir en sentidos opuetos y de los que
sólo puede apreciarse el resultado final con la consiguiente pérdida de
información. Tampoco se contempla la variabilidad temporal de las tasas, la
que sería imprescindible para establecer relaciones entre los cambios
fenológicos y ambientales con respecto a la química del agua.
Teniendo en cuenta todas estas consideraciones, los resultados obtenidos
en el área de estudio habrán de tomarse con muchísimas precauciones sobre todo
en relación a dos puntos:
1. El período de estudio de los flujos de trascolación abarca solamente
dieciocho meses en el que se detectaron 24 eventos lluviosos. Aunque Lovett y
Linberg (1984) consideran 20 lluvias como número mínimo para obtener
resultados fiables a partir de este modelo.su aplicación a la cuenca
Nacimiento puede ser más problemática debido a las características más
irregulares de los eventos lluviosos.
2. Los colectores de trascolación no tienen dispositivo de cierre
automático y no están elevados debido a la poca altura sobre el suelo de las
primeras ramas. Por ello la contaminación a par t i r del suelo de los colectores
puede tener importancia aunque los individuos se encuentren en zonas del
bosque relativamente cerradas y protegidas.
Aplicación del modelo a las especies estudiadas.
A la hora de aplicar el modelo a las especies estudiadas hemos tenido
solamente en cuenta los aportes por trascolación neta de cada lluvia
muestreada. La escorrentía cortical, debido a su efecto restringido a una
222
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
pequeña superficie, tiene una escasísima importancia cuantitativa al calcular
las tasas para una hectárea de suelo y por tanto no los hemos considerado en
este estudio. El período muestreado abarca desde Abril 1988 a Septiembre de
1989. Los valores del volumen de lluvia, dias secos y aportes netos de cada
ion aparece en los apéndices 4.1 a 4.4. Para poder efectuar mejor las
comparaciones entre las distintas especies, los aportes netos de trascolación
están referidos a un 1002 de cobertura de cada especie.
Los parámetros para las ecuaciones de regresión aparecen en las tablas
4.4 a 4.7. Se indica el valor del coeficiente de regresión (r ) y su
significación, los valores de bl y b2 en (g/ha.día seco y g/ha mm)
respectivamente junto con su nivel de significación. Excepto en los casos que
se indica, el término independiente no es significativamente distinto de cero,
por lo que las ecuaciones se han calculado igualando este término a cero
aumentando así los grados de libertad del modelo.
Se han obtenido regresiones significativas para la mayoría de los iones
(p<0.005) excepto para el NH4 en las cuatro especie, el PCH en P. pinaster, P.
nigra y A. decorticans y protones en P. nigra y A. decorticans. Existen
algunas ecuaciones débilmente significativas (p<0.05) con coeficientes de
regresión entre 0.24 y 0.30 (protones en P. pinaster y j a ra y N03 en P. nigra)
cuya fiabilidad es, también dudosa. En general para todos estos elementos
existe un considerable número de lluvias con tracolaciones netas negativas es
decir, la trascolación contiene un contenido menor de esos nutrientes que¡ la
lluvia, indicando una reabsorción foliar o inmovilización por flora o fauna
epífita. Lovett y Lindberg (1984) tampoco encuentran relaciones significativas
para aquellas elementos con predominio de trascolaciones netas negativas.
Estos autores argumentan que el modelo parece poco satisfactorio para los
nutrientes con este tipo de comportamiento, lo que parece también confirmarse
con nuestros resultados.
Los elementos químicos cuyo comportamiento en trascolación parece
explicarse mejor por el modelo son el K y el NO3 en las especies arbustivas
mientras que es la trascolación neta de los cationes Ca y 'Mg y los aniones
SO4 y Cl la mejor explicada en el pinar.
223
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 4.4. Ecuaciones múltiples ajustadas según el modelo de Lovett y Lindberg (1984). para Jos aportes netos en agua de trascolación de P.pinaster. Se indican el coeficiente de regresión para cada una de las ecuaciones junto con su nivel de significación. Así mismo se indica la significación de los coeficientes bl y b2. (*): p<0.05, (**): p<0.01, (*••): p<0.001.
ELEMENTO
Na
K
Ca
Mg
NH4
H
N03
S04
Cl
P04
C03H
(Bl).DEP.SECA (j/ha.día seco)
ns 5.79
•
12.48
• 27.40
ns 3.19
ns 1.51
ns 0.17E"1
26.5
• •
33.9
•* 25.47 '
ns 0.38
•
62.61
(B2).INTERCAMBIO (j/ha.mrn)
• •
5.31
•
6.17
• • • 21.14
• • • 6.72
ns -1.57
•
-0.19E"1
ns 0.27
•
11.39
•
9.82
ns -0 .55
ns 16.48
P
***
***
»**
• * •
NS
•
*«*
»»»
***
NS
**
2
r
0.54
0 .58
0.71
0 .80
-
0.24
0 .53
0.70
0 .65
-
0 .48
Tasas de deposición seca.
Las tasas de deposición seca (coeficiente bl, tablas 4.4 a 4.7) son
significativas para la mayoría de los iones a excepción de los protones y del
N03 en P. nigra, A. decorticans y C. laurifolius. Por el contrario el
224
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
Tabla 4.5. Ecuaciones múltiples ajustadas según el modelo de Lovett y Lindberg (1984) para los aportes netos en agua de trascolación de P.nigra. Se indican el coeficiente de regresión para cada una de las ecuaciones junto a su nivel de significación. Así mismo se muestra la significación de los coeficientes bl y b2. (*) p<0.05; (**) p<0.01; (•**) p<0.001.
ELEMENTO
Na
K
Ca
Mg
NH4
H
N03
S04
Cl
P04
C03H
Bl (DEP.SECA) (t/ha.dí» seco)
• •
19.23
• • •
33.65
• • •
72.03
• • • 9.01
ns 4.86
ns 7.92
• • • 72.19
• • • 48.47
• • • 191.4
B2 (INTERCAMBIO) ((/ha.mm)
ns 2.44
ns -4 .6
ns 1.50
ns 0.37
• -3 .22
ns 9.26
ns -5 .8
ns 1.63
• -46
P
*»»
***
***
***
NS
NS
*
»»*
*»*
NS
***
2 r
0.56
0.58
0.77
0:64
0 .20
0 .04
0 .30
0.74
0.61
0 .08
0.58
coeficiente correspondiente a los N03 en P. pinaster resulta altamente
significativo. En esta última especie tampoco son significativas las tasas del
Na y Mg. Otro tanto ocurre con el P04 en la jara. En general son los elementos
que presentan menores tasas, incluso negativas, los que no resultan
significativos. Un coeficiente negativo puede ser el resultado de captación de
225
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
deposición seca por parte de flora-fauna epífita y en estos casos parece que
el modelo no resulta satisfactorio.
Lovett y Lindberg (1984) apuntan la irregularidad temporal que existe en
las tasas de deposición seca y a que se necesita un número considerable de
lluvias para alcanzar resultados significativos. Este patrón irregular sea
probablemente más patente en el aréa de estudio si tenemos en cuenta factores
como: escasa cubierta vegetal próxima a zonas degradadas con fuerte erosión,
proximidad a fuentes emisoras de partículas como son las canteras de mármol de
Macael y distribución irregular de los períodos secos a lo largo del año.
Todos estos factores pueden actuar en mayor o menor grado determinando una
fuerte variabilidad en las tasas de deposición seca anterior a cada lluvia
explicando la baja o nula significación de algunos coeficientes de deposición
seca encontrados en este trabajo.
La magnitud de los coeficientes bl significativos en cada especie pueden
indicarnos el papel de las distintas cubiertas vegetales como captadoras de
deposición seca. El orden de magnitud es el siguiente:
P. pinaster: N03 > S04 > Cl - C03H > K > Ca
P.nigra: SO4-CO3H > K > Ca > Mg- Cl > Na
A. decorticans: CO3H > Cl > K > Ca > S04 > Na > Mg
C: laurifolius: Cl- CO3H > K > SCH- Ca > Na > Mg
Las dos especies arbustivas se comportan de manera similar, ya que tanto
la ordenación de los iones como las tasas calculadas son similares en ambos
casos (tabla 4.6 y 4.7). Ambos matorrales presentan pues similares eficiencias
en la captación de partículas vía deposición seca.
No ocurre así en el pinar, en que se aprecian diferencias tanto en la
ordenación iónica como en la magnitud de las tasas (tablas 4.4 y 4.5). Las dos
especies de pino presentan muchas similitudes en la geometría de sus
individuos y en la morfología y estructura de las hojas. Por ello, es un tanto
extraño que se detecten diferencias, a veces importantes respecto a la
deposición seca de algunos iones. Una de las causas probables es la diferente
226
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrlentei
Tabla 4.6. Ecuaciones múltiples ajustadas según el modelo de Lovett y Lindberg (1984) para los aportes netos en el agua de trascolación de C. laurifolius. Se indican el coeficiente de regresión para cada una de las ecuaciones junto con su nivel de significación. Asimismo se muestra la significación de los coeficientes bl y b2. (*) p<0.05; (**) p<0.01; (•**) p<0.001.
ELEMENTO
Na
K
Ca
Mg
NH4
H
N03
S04
Cl
P04
C03H1
Bl(DEP.SECA) (g/ha.día teco)
• • 11.68
*• 27.1
• 16.62
• 4.39
ni
2.06
ns -0.017
ns -0 .82
• 17.87
• • • 61.86
ns 4.23
• • • 56.26
B2UNTERCAMBIO) ((/ha.mm)
ns 1.22
• 10.78
• 11.98
•
2 .93
*
-3.15
• 0 .024
*•• -4 .84
ns 3.51
ns 6.69
• 2.83
• -15.1
P
***
*•*
*•#
»**
NS
*
**«
**
»»»
*»
**
2
r
0.54
0.72
0.56
0 .60
0.21
0.25
0.68
0.48
0.73
0.39
0.52
1: término Independiente significativamente distinto de cero.
localización de estas especies en la cuenca. La zona repoblada con el pino
resinero se sitúa en las laderas y en las zonas más próximas a la vaguada. En
esta situación los individuos están más expuestos al viento sur y protegidos
de los del norte (Domingo 1991). Por el contrario P.nigra se sitúa en las
cotas superiores, más descubiertas y expuestas al viento de componente N
227
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 4.7. Ecuaciones múltiples ajustadas según el modelo de Lovett y Lindberg (1984) para los aportes netos en agua de trascolación de A. decorticans. Se indican el coeficiente de regresión para cada una de las ecuaciones junto con su nivel de significación. Así mismo se muestra la significación de los coeficientes bl y b2. (*) p<0.05; (*•) p< 0.01; (***) p< 0.001.
ELEMENTO
Na
K
Ca
Mg
NH4
H
N03
S04
Cl
P04
C03H
(Bl) DEP.SECA (j/ha.dí* seco)
•
11.96
• •
32.53
• 22.01
• •
6.79
ns -3 .8
• 17.65
• • 50.85
• • 65.68
(B2) INTERCAMBIO / { / h i . m m )
ns -1.02
• 10.51
ns 0.53
ns 0.89
• • -3 .63
ns 1.58
ns 6.21
• • -35 .5
P
*
»««
*
• • *
NS
NS
**•
*
»**
NS
*#
2 r
0.30
0.70
0.32
0.58
0.06
0 .05
0.61
0.36
0.55
0.01
0.42
(Domingo 1991). Los mayores aportes por deposición seca en P.nigra pueden ser
debidos en primer lugar a su crecimiento en zona llanas de elevada altitud
menos protegidas. Además, las partículas procedentes de las canteras de mármol
próximas (ricas en CO H y Ca fundamentalmente) son transportadas por vientos
de componente norte contribuyendo también a explicar tasas de deposición seca
228
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
del orden de dos y medio-tres veces más altas en P.nigra. Sin embargo para el
Cl, con un origen marino en el aréa de estudio (Domingo 1991) las diferencias
se reducen considerablemente ya que este elemento se transporta por vientos
del sur que afectan de manera más directa a P.pinaster. En esta especie
destaca también la elevada contribución del NO3 a la deposición seca con un
coeficiente altamente significativo que resulta mayor que el de ningún otro
elemento.
Tasas de intercambio a nivel de copas.
Los coeficientes de intercambio (b2) para los distintos iones en la
especies estudiadas aparecen en la tablas 4.4 a 4.7. Los valores no son
directamente comparables con los de bl ya que las unidades son distintas.
Algunos de los iones con origen predominantemente externo (Na, Cl y SO4) no
poseen coeficientes significativos de intercambio en P. nigra, C. laurifolius
y A. decorticans. Son, por tanto la vía seca y la deposición húmeda las rutas
preferentes de entrada al suelo para estos elementos. P.nigra tampoco posee
valores significativos respecto al K, Ca, H y N03 siendo así la especie para
la cual el modelo explica peor los procesos de intercambio que se dan en las
copas, bien debido a que éstos tienen una menor importancia o a que presentan
mayor irregularidad. En P. pinaster tampoco se encontraron tasas
significativas para la alcalinidad y nitratos y en A. decorticans para Ca, Mg
y H.
La ordenación de las tasas de intercambio significativas en cada una de
las especies es la siguiente:(en negrita: tasas negativas).
P.pinaster: Ca > S04 > Cl > Mg-K > Na > H
P.nigra: NO3 >C03H
A.decorticans: K > NO3 > CO3H
C.laurifolius: K > Ca > Mg-P04 > H > NO3 > CO3H
Los coeficientes negativos encontrados para el N03 en la especies
arbustivas son altamente significativos indicando absorción de nitrato a
229
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
part ir del agua de lluvia, característica también comentada por Domingo (1991)
y Domingo et. al. (1991) a par t i r de los coeficientes de enriquecimiento de NO3
en estas especies. El mecanismo de absorción foliar está causado probablemente
por un gradiente de concentración ya que se ha encontrado una fuerte
correlación negativa entre aportes de NO3 en lluvia y en trascolación neta de
ambas especies (r=0.85 en la j a r a y r= 0.84 en el gaznacho, en ambos casos
p<0.001, n =22).EL bicarbonato es también retenido por la copas de los
matorrales. Los coeficientes de enriquecimiento para este anión encontrados
por Domingo (1991) y Domingo et al. (1991) son positivos, pero ello se debe al
efecto superpuesto de la deposición seca que, como vimos en el apartado
anterior, juega también un papel importante.
El lixiviado de nutrientes de las copas a part ir del agua de trascolación
(coeficientes b2 positivos) sería en realidad el verdadero aporte de la
vegetación "per se" al suelo dado que el lavado de deposición seca constituye
una entrada externa aunque canalizada por la vegetación. Biológicamente, tal
como lo define Parker(1983) el lixiviado ("leaching") es un proceso pasivo que
transfiere sustancias químicas desde el pool interno de la planta. Esta
transferencia viene determinada por el paso del agua de lluvia que actúa como
solución externa y por el gradiente de concentración que se establece entre
ésta y el pool interno.
Con el modelo aplicado no se han encontrado demasiadas tasas de lixiviado
significativas sobre todo en A. decorticans y P. nigra. Sin embargo existen
algunos resultados que interesa comentar.
Las mayores tasas de lixiviado se obtienen para el K en el matorral
(lOg/ha.mm). las tasas en P. pinaster son ligeramente inferiores (6.17
g/ha.mm) pero en este caso no es el K quien ocupa el primer lugar. Existe
unanimidad en la bibliografía respecto a la elevada movilidad del K y por
tanto a su facilidad para el lixiviado por agua de lluvia, sobre todo teniendo
en cuenta sus relativamente altos contenidos en la fracción foliar. Según
Parker (1983) de un 60-90% del K en trascolación neta provendría de lixiviado.
El lixiviado de Ca es también muy importante en C. laurifolius ocupando
230
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
la primera posición (11.9 g/ha.mm) junto al K. En P. pinaster este ion ocupa
también la primera posición con tasas que prácticamente duplican las
encontradas en la jara . En el pinar, existe un pool interno importante de Ca
debido a la presencia de acículas de varios años en el árbol y ricas en Ca.
Por su parte, la fracción foliar de C. laurifolius es también muy rica en este
elemento. Aunque las entradas de Ca por deposición global son altas en la
cuenca, los contenidos foliares son suficientemente elevados para que se
establezca un gradiente de concentración a favor del lixiviado. Aunque el Ca
suele considerarse como un elemento no demasiado móvil debido a su carácter
estructural (Stachurski 1987) existen varios estudios citados por Parker
(1983) que sugieren la existencia de almacenes móviles de Ca en hojas. En el
mismo trabajo se indica que un máximo de 60-80% del Ca en trascolación
provendría de lixiviado sobre todo en zonas lejanas a posibles fuentes
emisoras.
La tasa de lixiviado de S04 es también importante en P. pinaster (11.4
g/ha.día). Lovett y Lindberg (1984) así como Bellot (1989) y Schaefer et al.
(1988) también han encontrado una contribución importante del lixiviado para
este ion. La procedencia del sulfato en estos trabajos así como en el presente
estudio tiene un doble origen (deposición seca y lixiviado). La importancia
porcentual de este último puede ser considerablemente mayor en áreas no
industrializadas lejanas de fuentes emisoras (lindberg y Garten 1988). De
todos modos la fisiología del lixiviado del S04 y su movilización desde las
hojas está aún poco estudiada (Parker 1983).
Las elevadas tasas de lixiviado encontradas para el Cl y Na en P.
pinaster pueden ser resultado de un error del modelo [deposición seca que se
lava lentamente] ya que el cloruro tiene un índice de movilidad menor del 1%
(Tuckey en Parker 1983), y su concentración en hojas es poco importante. El
Na, a pesar de ser un elemento muy móvil se encuentra en muy bajas
concentraciones en la hoja en relación al agua de lluvia. Para ambos elementos
se considera un origen predominante de deposición seca (Parker 1983) y es muy
posible que las elevadas tasas de intercambio encontradas a part i r del modelo
sean una sobrestima del lixiviado real.
231
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
•4.2.3 Contribución de la deposición seca, y lixiviado a los apor tes ct
nutrientes al suelo.
Considerando las tasas calculadas a par t i r del modelo, la precipitaciór
anual y los días secos anuales durante el período de muestreo de los flujos
hidrológico (718 mm y 280 días secos), pueden calcularse los aportes (Kg/ha) a
la cuenca vía deposición seca y lixiviado.
En la tabla 4.8 se muestran por especies, los aportes de cada vía y su
contribución a la trascolación neta total. Los resultados están corregidos por
la cobertura de cada especie en la cuenca (para los pinos la cobertura
considerada es la de una ha representativa del área repoblada con cada uno de
ellos). Los valores que aparecen sin negrita son los calculados a par t i r de
coeficientes del modelo que no son significativos y que por tanto no reflejan
de manera fiable el comportamiento del ion correspondiente.
Los iones de procedencia externa que entran al sistema vía deposición
seca son para todas las especies los cloruros, bicarbonatos y sulfatos. Este
último elemento tiene mucha menor importancia cuantitativa en los matorrales.
Es destacable la entrada de No3 por deposición seca en P. pinaster, mientras
que para las otras especies no tiene importancia significativa. También
resultan interesantes las elevadas entradas de Ca y bicarbonato en P. nigra
quizá facilitadas por una favorable exposición a vientos de componente norte
que transportan estos elementos procedentes de las canteras de mármol. Un
75-827. de las entradas de Na por trascolación y un 76-92% de las de Cl en P.
nigra, A. decorticans y C. laurifolius provienen de deposición seca, casi con
toda seguridad de origen marino. No ocurre lo mismo en P. pinaster, en que la
vía de intercambio calculada es importante, aunque ya hemos comentado que este
parece ser un caso de deposición seca que el modelo estima como intercambio.
El K que se considera siempre de origen interno parece tener aquí un
componente de deposición seca importante en P. pinaster, A. decorticans y C.
laurifolius que se acerca al 507.. Otros estudios han destacado también esta
característica. Bellot (1989) encuentra similares porcentajes de deposición
seca y lixiviado en encina, mientras que Lovett y Lindberg (1984) lo hacen en
las caducifolias Quercus alba y Quercus prunus pero solamente durante la
232
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente!
a.
.8 a I
a
5 n í M S i • * i •
» sO 1/1 o 0» «— «# í»
«na «"i s 5 j
5 3 K/H\K i • » f • •
e» o e m io e> •— •" m •* •Ó >* ni Jj|
3Í s B ¡o n i "•»
3 3 8 S S . • • / i ! .
N f l • * ' lO */s
5 fe ft Ó fe
* • S iw
a o S •» So
' ~ N* 12 a
itlli
á a a s s Í Í 3 S 3
I a • • •
>0 N.
3 s n s s « • • • •
o o *• /o> ;o . i • ; • / «
•. n V CM
-~ o «- o O ui m O o B • • • • •
o> e in uj
• • • * •
•* «- \r\ u> «#
• • • , • • «v M -* in
1 o £ 2 « 3
-«
3 fe fc 8 5 • • • • •
4 a m c •>
t n t ~ t n u*i m IO oo • »
- •* M *
t • • • •
»• N eo 2
¡* 5 fio?
^ 8 ^ 2 ^ *
4 n « N a *- T- Rl oo «-• • • • i
t a 8 11 S vn N <- t ¡ «•• nj «* ^ e
~ •' ~ 6 ú
üiH
S 5 & fc {Q 3 SÍ
rn <* » 10 n i * i i f l • • » oo
' ' ' 3 a
&j S » ¡c R •* «-- •• « ^ •
n j r»»
S N 5 « í ~ " ~ R i
' "" 5 "•'
A M M W ; N . \ fO
s 8 a e s ~ ui « " '
• f i • i
3 *
' " í «I
I ; I
5 S S 5 6 ' " ¿ '¿ é
1 " c 5 £ o — t - X
233
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
estación de reposo. Quizá la explicación más lógica sea la de una
contaminación de los colectores a partir de la vegetación o a exudados de K a
partir de las hojas en períodos secos. Para C. laurifolius y P. plnaster
parece existir una fuente interna importante de Ca mientras que en P. nigra y
A. decorticans el origen del Ca es externo en más de un 90%. El S04 en P.
pinaster tiene doble origen con una importancia similar en cada vía. Bellot
(1989) y Schaefer et al. (1988) han indicado también la existencia de una
importante fuente interna de S04 en encina y Abies balsamea respectivamente.
Parker (1983) y Lindberg y Garten (1989) destacan la variabilidad existente
respecto a al contribución de la vía seca respecto al lixiviado de S04 en
diferentes estudios. En principio parece que la vía seca ha de tener mucha
importancia sobre todo en áreas industrializadas. Pero estos autores concluyen
que el lixiviado puede ser también notable y del orden de un 50% en algunos
casos. Sin embargo, es difícil predecir la contribución de la vía interna dado
que es muy poco lo que se conoce acerca de los mecanismos foliares de
liberación de esta sustancia.
No se aprecia lixiviado de N03 en las especies arbustivas, sino una
absorción o retención de este elemento por parte de las superficies foliares.
La vía seca no es significativa para este elemento en el matorral. Estos
resultados coinciden con los encontrados por Domingo (1991) en relación a los
coeficientes de enriquecimientos negativos encontrados en la trascolación de
ad y C. laurifolius. Por su parte son bastante los trabajos que describen
absorción de No3 por parte de las superficies vegetales (López 1989, Bellot
1989, Shaefer et al. 1988, Nihlgard, Horntredt en Hasselrot y Grennfelt
(1987).
Domingo (1991) calcula coeficientes de enriquecimiento de 0.32 para los
protones en el agua que trascola a través de la ja ra . La aplicación del modelo
de Lovett y Lindberg indica también una ligera acidificación del agua
manifestada por las tasas de lixiviado positivas y significativas encontradas
para esta especie. Por el contrario, la vía seca carece de importancia
significativa. En P. pinaster se observa sin embargo una tendencia a la
neutralización de la acidez de la lluvia manifestada por las tasas negativa y
234
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
significativas de intercambio en esta especie.
En relación al bicarbonato, se han encontrado tasas de intercambio
significativas y negativas en tres de las especies estudiadas (pn A.
decorticans y C. laurifolius) , tasas que por otro lado son de una magnitud
considerable (entre 3.01 y 15.36 Kg/ha.año). El bicarbonato constituye la
entrada iónica más importante tanto por deposición húmeda (Domingo et al.
1991, Domingo 1991) como por vía seca como ya hemos visto. En esta
circustancias, es muy probable que el bicarbonato funcione como fuente de
protones destinados a intercambiarse con otros procedentes del interior de la
planta. Por tanto más que una absorción por parte de la planta lo que puede
ocurrir es una disociación en CO y protones.
4.2.4. Importancia de la vía hidrológica en el aporte de nutrientes al suelo.
Una vez conocidas las tasas de deposición seca y lixiviado en las que
puede compartimentalizarse el flujo de trascolación neta así como los aporte
por deposición global, puede estimarse el porcentaje de contribución de cada
vía hidrológica a los aportes totales al suelo forestal (no se incluyen los
portes por escorrentía cortical)
Suponiendo un suelo cubierto al 100% por cada una de las especies
estudiadas para facilitar la comparación interespecífica, se han representado
para cada ion los porcentajes relativos de deposición húmeda, seca e
intercambio (figuras 4.1 y 4.2). Las entradas anuales por precipitación se han
tomado de Domingo (1991). Aunque estos aportes corresponden a la deposición
global, suponemos, según las hipótesis de este mismo autor, que la deposición
seca recogida por los colectores de lluvia es mínima comparada con la captada
por la propia vegetación.
Para el pinar, (figura 4.1), los iones predominantemente externos cuya
entrada proviene tanto de la vía seca como de la húmeda son los sulfatos,
cloruros, bicarbonatos y sodio. En P. pinaster, no parece existir ningún ion
para el que la vía de entrada preferente sea la de la deposición seca, sino
que lo que se observa es una importancia similar de la vía seca y húmeda o
235
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
1ÍT '4Á9
Na K Ca Mg H N03 S 0 4 Cl Ale
na k ca mg no3 so4 cl ale
Figura 4.1. Contribución (%) de cada una de las vas hidrológicas (deposición
global, deposición seca y lixiviado) al aporte de nutrientes al suelo por
parte de las especies de pino. La figura superior corresponde a P.pinaster y
la inferior a P.nigra. Se supone una cobertura del 100% para cada especie.
bien una predominancia de la húmeda. El caso del Cl y del Na en esta especie
son un tanto especiales ya que el modelo sobrestima claramente las tasas de
lixiviado. Si aplicamos los porcentajes de deposición seca que aparecen en la
bibliografía (Parker 1983, López 1989) la contribución de la vía seca sería
superior a la húmeda.
236
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
En P. nlgra parece existir una predominancia clara de la deposición seca
en los iones de origen predominantemente marino así como para el Ca, de
origen predominantemente continental. Iones con doble entrada externa y
contribución similar de cada vía son los sulfatos, el magnesio y los
bicarbonatos. Esta mayor eficacia de P. nigra en la captación de la deposición
seca puede ser debida, como ya hemos comentado, más a una localización
favorable para la captación de partículas o aerosoles, que a diferencias en la
geometría de las dos especies de pino o a características estructurales de sus
hojas. A la hora de comparar estas dos especies debe hacerse algún comentario
acerca del diferente origen de los nitratos. En P. pinaster las tasas de
deposición seca son elevadas y altamente significativas. Lo mismo ocurre para
la de intercambio en P. nigra. Este diferente comportamiento resulta algo
extraño en especies tan afines pero parece apoyarse con algunos resultados
encontrados por Domingo (1991). Así, este autor encuentra correlación
significativa entre la concentración neta de nitrato en trascolación y los
días secos interlluvias en P. pinaster y no en P. nigra. Por otro lado, existe
una correlación negativa entre las concentraciones de nitrato en trascolación
neta y en el agua de precipitación en P. nigra. Esta relación indicaría la
existencia de mecanismos de lixiviado interno mediante difusión por gradiente
de concentración en el pino laricio. Sin embargo en P. pinaster no se ha
encontrado este tipo de relación (Domingo 1991).
La vía de intercambio para el K, Ca y Mg solo tiene importancia
significativa en P. pinaster y no en P. nigra. Los protones en esta especie
son retenidos funcionando probablemente como intercambiadores iónicos,
mientras que en P. nigra la fuente de protones podrían ser los iones
bicarbonato que utilizan como vía de entrada tanto la deposición seca como la
húmeda. El Mg y sulfato en esta especie tienen también procedencia externa y
penetran tanto por vía seca como húmeda.
En C. laurifolius (figura 4.2), los nutrientes de origen externo que
entran a formar parte del sistema son básicamente los cloruros, sulfatos y
nitratos así como el Cl y el Na. Para estos dos últimos iones existe una
preferencia por la vía seca. Los demás utilizan fundamentalmente la vía
húmeda. Ca, Mg y H tienen su origen tanto en fuentes externas como internas.
237
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Los mayores porcentajes (53 7. del Ca, 587. de los H+ y 427. de Mg provienen de
la deposición global, aunque la contribución interna alcanza el 30 7. para el
Ca, 367. para el Mg y 23.6 7. para los protones.
Según este modelo la deposición seca de K parece contribuir de manera
importante en ambas especies con contribuciones muy similares a las del
lixiviado. La mayoría de resulatdos de bibliografía coinciden en valores mucho
más altos del lixiviado (superiores al 60%) en relación a la deposición seca
dado el predominante origen interno del K, su movilidad y su escasez en la
baja atmósfera y en el agua de precipitación. Es muy posible que exudados de
la propia vegetación o deposición seca de origen interno y no de la baja
atmóferahaya podido depositarse en las superficies foliares aumentando la
contribución de este flujo. Otra posibilidad es la contaminación de los
colectores por restos vegetales. Ambas hipótesis pueden ser posibles si
tenemos en cuenta la cercanía de las hojas y ramas de estas especies al suelo
y el tipo de colector empleado en la recogida.
En las dos especies arbustivas parte del bicarbonato y nitrato, iones que
penetran fundamentalmente vía húmeda, es captado por la cubierta vegetal. Los
mecanismos implicados podrían ser diferentes para ambos iones. Existe un buen
número de autores que constata la reabsorción de parte del nitrato de la
lluvia (Lemée 1974, Bellot 1989, López 1989 en especies de Quercus, Cronan y
Reiners 1983, Shaefer et ai . 1988 en coniferas, Bellot 1989 en el arbusto
Phyllirea inedia), aunque los mecanismo implicados en ella no sean del todo
conocidos (Domingo 1991). No ocurre lo mismo con los bicarbonatos. Como ya se
ha comentado no es muy probable que este ion sea reabsorbido sino disociado en
C02 y H sobre todo en la j a ra en la que ésta reacción podría verse favorecida
por los ph más bajos del agua que trascola a través de su cubierta (Domingo
1991).
Las diferencias más destacables entre las vías hidrológicas utilizadas
para el retorno de nutrientes por parte de los dos arbustos son la no
importancia significativa de lixiviado de Ca y Mg en A. decorticans (tabla
4.8). Los hongos adheridos a la superficie de las hojas de j a ra quizá podría
ser una posible fuente de este catión. Destaca tanbién en la j a r a una ligera
238
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
Na K Ca Ms H N03 S04 CI Ale P04
Na K Ca Mg N03 S04 Cl Ale
Figura 4.2. Contribución (%) de cada una de las vas hidrológicas (deposición
global, deposición seca y lixiviado) al aporte de nutrientes al suelo por
parte de las especies de matorral. La figura superior corresponde a
C.laurifoliusy la inferior a A.decorticans. Se supone una cobertura del 100%
para cada especie.
acidificación del agua de lluvia, aunque poco importante. La fuente de estos
protones es según el modelo aplicado, el lixiviado. Por su parte la pérdida
significativa vía lixiviado de P-P04 es una característica diferencial de la
j a ra con el resto de especies que, por otro lado, se enmarca dentro de lo que
parece ser una constante, ya habitual en esta especie: el uso poco
conservativo del P (capítulo 2, 3 y 5).
239
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
4.3. Retorno conjunto por desfronde y trascolación neta.
4.3.1. Introducción.
La vegetación utiliza las vía hidrológicas (trascolación y escorrentía
cortical) y la de desfronde para retornar nutrientes al suelo. Sin embargo, el
retorno así calculado está incluyendo la deposición seca que no constituye una
entrada del propio sistema sino de procedencia exterior. En este apartado se
pretende estudiar el papel de cada especie en el retorno de nutrientes
discriminando en la manera de los posible las entradas puramente externas de
las internas (lixiviado, desfronde). Una segunda aproximación es evaluar la
importancia del retorno de nutrientes al suelo incluyendo los aportes por
deposición global. Para ello hemos utilizado una diagrama sencillo propuesto
por Parker (1983) con el fin de estimar el origen predominante de cada ion.
Estos diagramas serán construidos para dos situaciones:
Aproximación específ ica .
Supondremos que la cuenca posee únicamente un tipo de vegetación
representada por cada una de las cuatro especies estudiadas y con cobertura
dei 1007. para cada una de ellas. Normalizando los datos de esta forma pueden
realizarse comparaciones entre el comportamiento de las distintas especies y
evaluar también la relación matorral-pinar.
i
Aproximación por regiones.
Supondremos una situación real en la que: Un sector de la cuenca está
repoblada con P. pinaster con un porcentaje de cobertura del 53.5% coincidente
con una población arbustiva mixta de A. decorticans y C. laurifolius con 17% y
40% de cobertura respectivamente (RPP). Otra zona de la cuenca (RPN) se
encuentra repoblado con P. nigra con un 46.5% de cobertura y con matorral
mixto de A. decorticans (43.3.% de cobertura) y C. laurifolius (21.7.%). En
este caso, mientras que el recubrimiento del pinar en cada sector es muy
similar (tabla 1.6), no ocurre lo mismo con cada una de las especies
240
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
arbustivas. En la zona de P. pinaster el arbusto predominante es la j a ra y en
la de P. nigra el A. decorticans. A lo largo del trabajo se han podido
constatar importante diferencias respecto a desfronde, descomposición y
trascolación entre estas dos especies. Esta aproximación pretende establecer
si, dada la estructura vegetal actual en ambos sectores, existen diferencias
destacables respecto al retorno que pueden ser achacables a la predominancia
de una u otra especie, o bien, si las diferencias interespecíficas respecto a
este flujo se compensan con esa determinada estructura de la vegetación.
4.3.2. Características del retorno en las distintas especies.
En la tabla 4.9 se muestran los valores (en Kg/ha) de retorno anual en
las cuatro especies. Los resultados de las especies de pino se refieren al
igual que en el desfronde a la superficie repoblada y los del matorral al
conjunto de la cuenca. Los valores de desfronde corresponden a la media anual
encontrado en este trabajo (capítulo 3). La aportes por trascolación neta
están calculados a a part ir de los datos de Domingo (1991) teniendo solamente
en cuenta los aportes netos positivos. A part ir de ellos y tomando los
porcentajes de deposición seca y lixiviado calculados en el apartado anterior
puede estimarse la contribución de la vía externa e interna en el flujo
hidrológico de trascolación. Los porcentajes de intercambio negativos se han
tomado en estos cálculos como de un 0% ya que no retornan nutrientes al suelo.
En la tabla se indican también qué valores están calculados a part i r de
coeficientes de deposición seca o intercambio no significativos (valores que
no aparecen en negrita). En estos casos, ios flujos calculados a partir de
ellos han de tomarse con cierta precaución.
De la tabla 4.9 se deduce que el flujo de trascolación neta es la
principal vía de retorno para el K y Na con porcentajes que oscilan entre el
51-167. y el 85-92% respectivamente. La contribución de esta vía respecto al
retorno de Mg es también notable con una contribución del orden del (28-41%).
Por su parte, el desfronde retorna al suelo mayoritariamente N y P. En el
matorral de un 95-1007. del N es devuelto al suelo por esta vía mientras que en
el pinar se obtienen porcentajes algo inferiores (87-91%). Concretamente en el
caso de P. pinaster la menor contribución del desfronde se debe a la
241
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
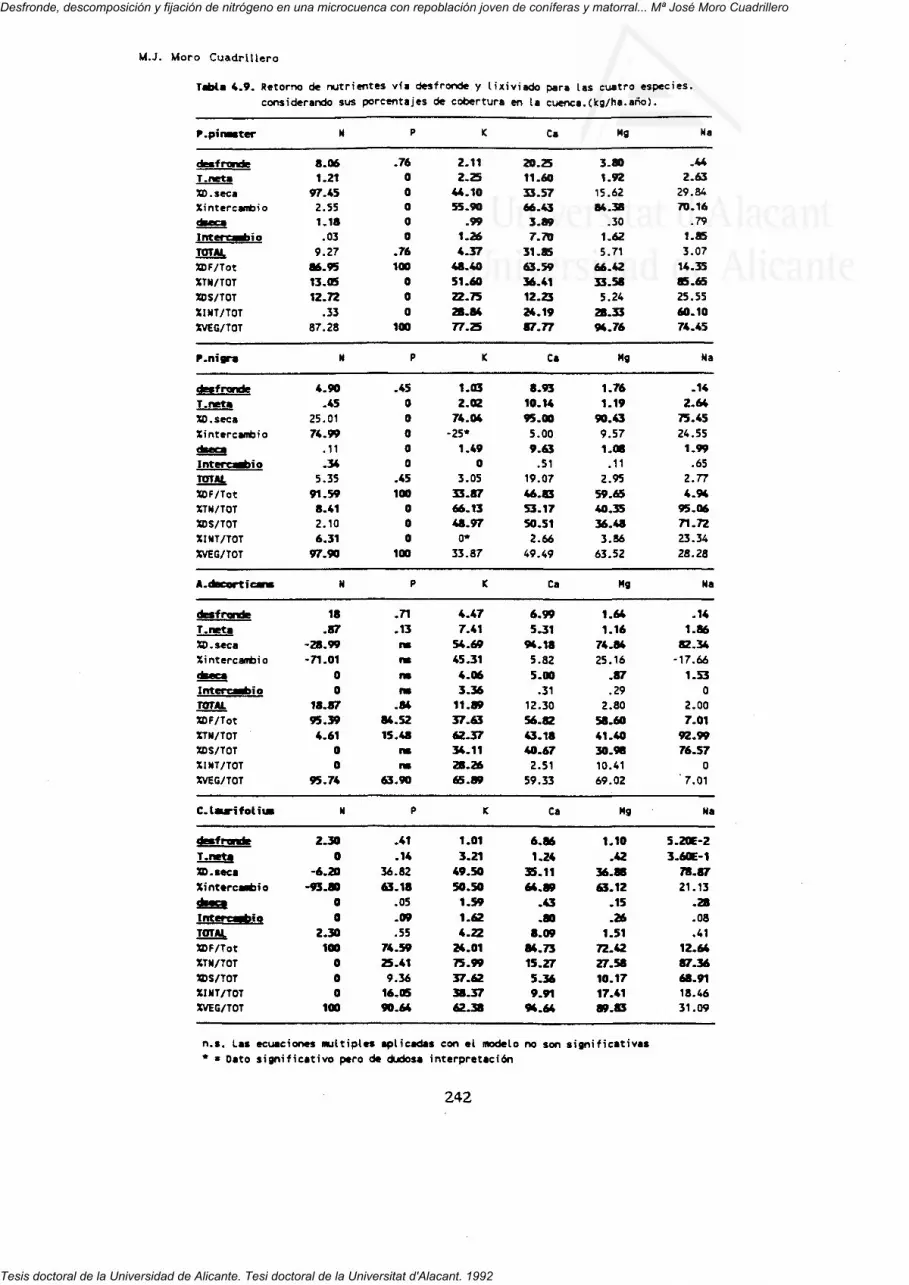
M.J. Moro Cuadrillero
labia 4.9. Retorno de nutrientes vfa desfronde y lixiviado para las cuatro especies, considerando sus porcentajes de cobertura en la cuenca.(ka/ha.año).
•.pinaater N P K Ca Mg Na
desfronde T.neta XD.seca Xintercambio daeca Int t i r—t i io TQTAJ.
XDF/Tot
XTN/TOT
XDS/TOT
XINT/TOT
XVEG/TOT
8.06 1.21
97.45 2.55 1.18 .03
9.27 86.95 13.05 12.72
.33 87.28
.76 0 0 0 0 0
.76 100
0 0 0
100
2.11 2.25
44.10 55.90
.99 1.26 4.37
48.40 51.60 22.75 28.84 77.25
20.25 11.60 33.57 66.43 3.89 7.70
31.85 63.59 36.41 12.23 24.19 87.77
3.80 1.92
15.62 84.38
.30 1.62 5.71
66.42 33.58
5.24
28.33 94.76
.44 2.63
29.84 70.16
.79 1.85 3.07
14.35 85.65 25.55 60.10 74.45
P.nigrs N P IC Ca Mg Na
deafronde 4.90 .45 1.03 8.93 1.76 .14 T.neta .45 0 2.02 10.14 1.19 2.64 XD.seca 25.01 0 74.04 95.00 90.43 75.45 Xintercambio 74.99 0 -25» 5.00 9.57 24.55 daeca .11 0 1.49 9.63 1.08 1.99 Intercambio .34 0 0 .51 .11 .65 Tom 5.35 .45 3.05 19.07 2.95 2.77 XOF/Tot 91.59 100 33.87 46.83 59.65 4.94 XTN/TOT 8.41 0 66.13 53.17 40.35 95.06 XOS/TOT 2.10 0 48.97 50.51 36.48 71.72 XINT/TOT 6.31 0 0* 2.66 3.86 23.34 XVEG/TOT 97.90 100 33.87 49.49 63.52 28.28
A.dBCorticana N P K Ca Mg Na
desfronde T.neta XD.seca Xintercambio daeca Intfn —«lia TOTAL
XDF/Tot
XTN/TOT
XOS/TOT
XINT/TOT
XVEG/TOT
18 .87
-28.99 -71.01
0 0
18.87 95.39 4.61
0 0
95.74
.71
.13 na na na na
.84 84.52 15.48
na na
63.90
4.47 7.41
54.69 45.31 4.06 3.36
11.89 37.63 62.37 34.11 28.26 65.89
6.99 5.31
94.18 5.82
5.00 .31
12.30 56.82 43.18 40.67
2.51 59.33
1.64 1.16
74.84 25.16
.87
.29 2.80
58.60 41.40 30.98 10.41 69.02
.14 1.86
82.34 -17.66
1.53 0
2.00 7.01
92.99 76.57
0 7.01
C.taurifoliua N P K Ca Mg Na
dBBfrtnx
T T " * *
XD.seca Xintercambio a» acá I n t e r n a » »
TOTAL
XDF/Tot
XTN/TOT
XOS/TOT
XINT/TOT
XVEG/TOT
2.30 0
-6.20 -93.80
0 0
2.30 100
0 0 0
100
.41
.14 36.82 63.18
.05
.09
.55 74.59 25.41 9.36
16.05 90.64
1.01
3.21 49.50 50.50
1.59 1.62 4.22
24.01 75.99 37.62 38.37 62.38
6.86 1.24
35.11 64.89
.43
.80 8.09
84.73 15.27 5.36 9.91
94.64
1.10 .42
36.88 63.12
.15
.26 1.51
72.42 27.58 10.17 17.41 89.83
5.20E-2 3.60E-1
78.87 21.13
.28
.08
.41 12.64 87.36 68.91 18.46 31.09
n.s. Las ecuaciones múltiples aplicadas con el modelo no son significativas * * Dato significativo pero de dudosa interpretación
242
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
importante tasa de lavado de deposición seca (tabla 4.4 y 4.9). En el pinar el
desfronde constituye la única ruta significativa de retorno al suelo de P con
contribuciones del 100% ya que en ambas especies el P04 de la lluvia es
absorbido por las cubiertas tanto de P. pinaster como de P. nigra. Sin embargo
en el matorral la trascolación neta aporta de un 15-257. del fósforo total.
La importancia de ambas vías en el retorno de Ca varía entre especies
según la mayor o menor importancia del lavado de la deposición seca (tabla
4.9). El desfronde contribuye de forma destacable en la j a ra (85%). En A.
decorticans y P. nigra, el lavado de deposición seca contribuye con un 41-507.
del retorno total de Ca y, en consecuencia, la importancia de la vía orgánica
disminuye a niveles comprendidos entre el 46-57%. En P. pinaster el desfronde
retorna el 64% de Ca pero con una contribución importante tanto del Ca de
origen externo (127.) como del lixiviado (24.%).
A. decorticans retorna al suelo una mayor cantidad de N, P y K que el
resto de las especies (18.8, 0.84 y 11.9 Kg/ha.año respectivamente). Este
mismo comportamiento para el N y K era observado en el desfronde (capítulo 3)
de esta especie. Al comparar ambos flujos se observa una fuerte
intensificación en el retorno de K causado por una importante transferencia a
través de la trascolación neta (62% del retorno total). Por el contrario, para
el N la intensidad de retorno es prácticamente idéntica a la encontrada en
desfronde. En esta especie el N-N03 es absorbido o retenido por las copas
(tasas negativas de intercambio y deposición seca) mientras que el N-NH4 y el
Nitrógeno orgánico enriquecen el agua de lluvia, (tabla 4.2) transfiriéndose
en pequeñas cantidades que se reflejarían en los 0.87 Kg/ha.año aportados por
trascolación neta.
Los cationes Ca, Mg y Na son aportados en mayor cantidad por P. pinaster,
en relación a las demás especies, comportamiento similar al observado en
desfronde. Como era de esperar la vía hidrológica es la que intensifica el
retorno de Na (85.6% del retorno total) aportándolo en una cantidad cinco
veces superior que la perdida en hojarasca. Sin embargo el origen externo o
interno del Na trascolado no queda claro con las tasas calculadas a part ir del
modelo que, como ya comentamos, no reflejan el comportamiento esperable de
243
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
este ion. Para el Ca y el Mg la contribución de la vía húmeda es algo menos
importante (33 y 36% del retorno total) y se reparte entre lixiviado (5 y 127.)
y deposición seca (28 y 24%) (tabla 4.9).
El P-P04 supone un 25% del retorno total de fósforo en la jara . De esta
cantidad aproximadamente un 63% procede de lixiviado. Este comportamiento
encaja con la utilización poco conservativa de este nutriente ya observada
para los flujos de descomposición y desfronde en esta especie. En A.
decorticans se observa una tendencia similar aunque el procentaje retornado
por trascolación es menos importante (15.5%). Las especies de pino, con tasas
de enriquecimiento negativas en la trascolación (Domingo 1991) y con
importantes eficiencias de retraslocación se comportan por el contrario como
especies conservativas en el uso del P.
En el capítulo 3 se comentaron las similitudes en las cantidades de
algunos elementos aportadas por el desfronde de C. laurifolius y P. nigra.
Estas pequeñas diferencias se intensifican en el retorno total al incluir la
vía hidrológica sobre todo respecto al N, P y Ca. Para los dos primeros
elementos ambas especies tienen comportamientos opuestos. P. nigra lixivia N03
por trascolación y absorbe PO4 mientras que C. laurifolius lixivia P04 y
absorbe N tanto en forma N-NH4 y N-N03 (tabla 4.2). Respecto al Ca, los
elevados aportes encontrados en P. nigra provienen en un 53% de la
trascolación mayoritariamente de origen externo (tabla 4.9) mientras que en la
j a r a la importancia de la vía húmeda sólo supone un 17%.
El Ca es un elemento de origen predominantemente interno dada su íntima
asociación a estructuras celulares. Se considera que la vía principal de
transferencia para este ion es el desfronde. No obstante Parker (1983) comenta
la importancia que puede tener la vía seca en aquellos zonas cercanas a
fuentes emisoras. Este parece ser el caso de la cuenca estudiada cercana a
zonas calizas y a explotaciones de mármol. En la cuenca "Nacimiento", el Ca es
uno de los elementos mayoritarios en la deposición global y en deposición seca
calculada por el modelo de Lovett y Lindberg. No es de extrañar que los
porcentajes de contribución de la vía desfronde a los aportes de Ca sean
inferiores a muchos resultados de la bibliografía sobre todo en el caso de P.
244
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
nigra y A. decorticans en los que las tasas de deposición seca de Ca
calculadas por el modelo suponen un 50% y un 40 7. respectivamente del retorno
total.
4.3.3. Contribución de los aportes internos (lixiviado + Desfronde).
Después de haber considerado las vías de entrada y trasferencia de
nutrientes al suelo atravesando la cubierta vegetal, deberíamos considerar que
porcentaje de contribución a estos aportes tiene la vegetación " per se". Es
decir, cuales son los principales nutrientes que se aportan por vía interna y
qué diferencias interespecíficas pueden establecerse a este respecto. La
transferencia interna está ligada tanto a diferencias específicas (genéticas,
fisiológicas) y a su interacción con las condiciones concretas en las que
viven. El retorno por vía interna constituye un buen indicador del flujo de
cada nutriente dentro del sistema y de su tasas de renovación cuando lo
relacionamos con el almacén o pool de nutriente almacenados en la biomasa
(Gray 1983).
En la tabla 4.9 se ha calculado el porcentaje que supone este flujo
respecto al total de nutrientes que retornan al suelo vía vegetación
(%veg/7.tot). Aunque ya hemos comentado ampliamente que la partición de la
trascolación neta calculada en este trabajo es una mera aproximación, sí puede
darnos ciertas indicaciones acerca de esta circulación interna. A part ir de
los resultados obtenidos pueden hacerse las siguientes consideraciones:
- Todos los nutrientes estudiados a excepción del Na penetran al suelos
forestal predominantemente por vía interna.
- En general, los mayores porcentajes de contribución se obtienen para el
N y P y en algunos casos para el Ca.
- En el matorral y el pino laricio, la vegetación aporta entre un 95-
100% del N con una mínima importancia del lixiviado en el matorral, mientras
que en P. nigra la lixiviación aporta un 6.5 % de los aportes internos en
forma de nitrato.
245
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
- El desfronde contribuye en un 100% al retorno de P en el pinar ya qu«
el P04 disuelto es absorbido a nivel de copas. En C. laurifolius, la
lixiviación de P04 aumenta la importancia de la vía interna considerablemente
alcanzándose un porcentaje de retorno interno del 90.6%. En A. decorticans, el
papel de la vegetación puede haber estado subestimado ya que no se conoce que
porcentaje de la trascolación neta de N-NH4 y Nitrógeno orgánico corresponden
al lixiviado.
- El K, ion de origen interno móvil y fácilmente lixiviable de las
superficies vegetales (Parker 1983, Stachurski 1987) retorna fundamentalmente
por esta vía en C. laurifolius (38%) en relación al desfronde (24%). En A.
decorticans, al hojarasca sigue siendo el flujo principal (37%),aunque sólo
ligeramente por encima del lixiviado (28.3%). En conjunto, la contribución de
la vegetación es similar en ambos arbustos (62.4% en C. laurifolius y 65.9% en
A. decorticans). Como ya comentamos,probablemente en esta dos especies ciertas
cantidades de K estimadas como deposición seca puedan ser exudados de origen
interno. En P. pinaster, la vía interna supone un 77% del potasio retornado
del cual un 25% es lixiviado. En P. nigra, el valor obtenido para este
elemento es tan sólo de un 34% que corresponde en su totalidad a la hojarasca.
No se han encontrado tasas significativas de lixiviado de K en esta
especie.Los valores obtenidos indicaban incluso una ligera retención lo que no
parece explicable desde el punto de vista biológico.
- Existen dos tendencias respecto a la restitución del Ca al suelo por
parte de las especies estudiadas. Por un lado, la circulación interna es la
mayor responsable del retorno de este elemento en C. laurifolius y P. pinaster
(94% y 88%), especies en la que tanto el desfronde como el lixiviado tienen
una contribución superior. En P. nigra y A. decorticans, en los que la vía
externa tiene una mayor contribución relativa, el Ca retornado por hojarasca
es menor que en P. pinaster y C. laurifolius y por tanto la circulación
interna tiene menor importancia (49.5% y 59.3% respectivamente. El mismo
comportamiento se observa para el Mg. Las contribuciones internas más elevadas
se obtiene en el pino resinero y en la j a ra y (94% y 89%) y la menores en P.
nigra y A. decorticans (63.5% y 69.2%).
246
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
- El Na tiene un origen claramente externo en A. decorticans para la que
la vía interna sólo supone un 17% de contribución. Porcentajes algo superiores
se han obtenido para P. nigra y C. laurifolius (287. y 317. respectivamente). En
esta últimas especies, las tasas de lixiviado a partir de las que se han
obtenido los porcentajes no son significativas, por lo que es muy posible que
estén sobrestimadas. La contribución relativa de los aportes internos de Na
está claramente sobrestimada, a pesar de que las tasas de intercambio fuesen
significativas. Probablemente estemos ante un caso de clara deposición seca
que el modelo de Lovett y Lindberg estima como intercambio.
4.3.4. Tasas de circulación de nu t r i en tes
La tasa de circulación de nutrientes por desfronde y trascolación puede
expresarse como el cociente entre los aportes debido al desfronde y al
lixiviado dividido por la mineralomasa acumulada en la vegetación (Gray 1983).
Obviamente el cociente está afectado por la magnitud de nutrientes acumulados
en la biomasa. Al excluir en este cálculo la deposición seca, estas tasas de
circulación serían las puramente internas. Incluyendo la deposición seca, las
tasas de turn-over de elementos predominantemente externos (Na
fundamentalmente) disminuyen considerablemente. En la tabla 4.10 se han
calculado las tasas de circulación internas para los elementos analizados.
Para el cálculo de algunas de ellas (las marcadas con asterisco en la tabla),
la contribución del lixiviado ha sido calculada a part ir de la bibliografía
(Parker 1983. López 1989) dado que los resultados obtenidos en este trabajo a
part i r del modelo de Lovett y Lindberg (1984) no son significativos ó resultan
muy poco interpretables desde el punto de vista biológico ya que parecen
sobrestimar claramente la contribución interna de Na y subestimar la de K.
Para el pinar, hemos aplicado un coeficiente de lixiviado de 657. que se
encuentra entre los valores mínimos encontrados en la bibliografía (López
1989, Müller en Parker 1983, Parker 1983) dado que parece las especies de pino
presentan, por lo general lixiviados menos intensos de este elemento. Para el
Na el porcentaje aplicado es el mínimo dado por Parker (1983):un 10%.
247
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
El orden de nutrientes según la mayor o menor rapidez de circulación es
la siguiente:
P. pinaster: Ca •> Mg > Na > N > P > K
P. nigra: Na > Ca > Mg > K > N > P
A. decorticans: Ca > Mg > K > P > N > Na
C. laurifolius: Mg > K > Ca-Na > P > N
Se observa en general una mayor rapidez de circulación en el Ca y el Mg y
una circulación más lenta en el N, P y K. Este orden coincide a grandes rasgos
con el encontrado por Gray (1983) en especies del matorral californiano. C.
laurifolius tiene un comportamiento algo diferente a las demás especies
respecto al K, elemento para el que se han obtenido elevadas tasas de
circulación en relación a los demás nutrientes. El Na no parece seguir una
pauta concreta y se encuentra en muy distintas posiciones según la especie que
se considere. N y/o P suelen tener por lo general tasas de circulación
similares al K aunque el origen sea diferente. Mientras que para el K la vía
hidrológica es predominante para N y P el predominio corresponde a la vía de
detritus. Esta pauta también fue apuntada por Ferrés (1984).
El porcentaje de turn-over de nutrientes en las especies vegetales
declina conforme la biomasa y la acumulación de nutrientes crece (Waring y
Schlesinger 1985). En este sentido, hay que considerar que, en nuestro
sistema, sobre todo en el caso de los pinos, no se ha alcanzado el cierre de
la bóveda y consiguientemente la estabilización de la producción primaria neta
y del índice foliar. Flujos como desfronde y lixiviado son dependientes del
área foliar (Waring y Schlesinger 1985) y estarán en esta situación
incrementándose hasta llegar a la estabilización de ese área foliar. No es por
tanto extraño encontrar en las especies estudiadas altas tasas de ciclado que
pueden parecer elevadas cuando las comparamos con valores de bibliografía
referidos a sistemas más maduros (Colé y Rapp 1981 en bosque caducifolios y
conifera y Ferrés (1984) en encinares), pero que son totalmente lógicas al
considerar la estructura actual del pinar . en la cuenca. Otra cuestión
importante es que, que el Ca y el Mg ocupan en este trabajo los primeros
lugares respecto a su velocidad de circulación, mientras que ocupan las
248
i
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
z
00 2 :
U
¡*¡
• n n n n
• vD W (M CM
00
n T I
if) n CM o\
CM
CM
1-t *-H ^ T i T i ^ 1
^ X ~
I 0
« I *> N
s = 5 «J
« •o # 4
TJ 1.
•O 0.
0
* fl t . 0
c a 0
•O
> X
_ -f 0
•0
c 0 i .
<~ a 0 0
O w 0-
n CTv
T i 1/) «J1
n o •4<
vO
CM
CM
If)
n
o CM
0\
CM CM TI
o c
CJ
I* o u <U •o
3
O < « 1
•-* 3
1/)
CM
C- Ti o t/i 00
\r> O Ti
1 LO 00 Ti
CM
if)
n
CM
cr. CM
CM
CM
T *
O n
0> TI
CM TI
TI CT>
O TI CM'
O T i
o if)
CM
CM
00
o o n cr>
.. • 2 « t. S
TJ <J • «J 9 N C C _ . 0
i f s e o
•o t. 0 o «j
1 S o •o
H
c TJ
*
O TJ
— « > o
- 1 o TJ 01
T i t - t T i O
L, <U
* J
W m c - H
Q.
m L, 60
• 1
c
3 m ü
* "M •4-1
« 1
O O <u •a
3 • 1
- H 0 v, •M 1* 3 m —H
ü
249
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
últimas plazas en trabajos de otros autores que t ratan con otros tipos de
bosque como por ejemplo el encinar (Ferrés 1984). La ordenación de los
nutrientes en función de su velocidad de ciclado es también un parámetro que
puede variar en función de la edad del sistema (Colé y Rapp 1981). No obstante
en los bosques de coniferas maduras incluidas en la recopilación de estos
mismos autores, el N y el P constituyen, al igual que en el pinar estudiado,
los elementos con menores tasas de circulación aunque difieran
considerablemente en los valores absolutos.
De la tabla 4.10a pueden señalarse, en primer lugar, diferencias
importantes en la circulación de nutrientes entre las dos especies de pino.
Sin embargo estos porcentajes no son estrictamente comparables ya que las
correspondientes tasas de turn-over de la materia orgánica son distintas entre
los dos pinos. Normalizando los datos teniendo en cuenta el flujo de materia
orgánica en cada especie, obtenemos los resultados de la tabla 4.10b. En ella
puede apreciarse una estrecha similitud en el comportamiento de los dos pinos
respecto a las tasas de circulación interna. La mayores tasas corresponden al
Ca y Mg, elementos que serán reciclados predominantemente vía descomposición,
mientras que las menores corresponden a elementos móviles e indispensables
para la planta (N, P y K), elementos para los que el pinar presenta un
comportamiento más conservativo sobre todo en el caso del P. Las tasas de
circulación normalizada en las coniferas estudiadas por Colé y Rapp (1981) son
del orden de 4-7 veces mayores a las de la materia seca en contraste con los
resultados obtenidos en P. pinaster y P. nigra para los que existe una mayor
similitud entre circulación de nutrientes-circulación de materia.
En relación al matorral, (tabla 4.10a) el comportamiento respecto a los
distintos nutrientes es similar al pinar, destacando la mayor magnitud en los
porcentajes de turn-over en los arbustos, lo que resulta lógico dado que se
t r a t a de especies con ciclos de vida más cortos. Normalizando las tasas, se
aprecian diferencias importantes entre C. laurifolius y A. decorticans que
confirman una vez más el diferente comportamiento de los dos arbustos
mayoritarios en la cuenca "Nacimiento". Para N y Ca A. decorticans presenta
tasas claramente superiores que C. laurifolius. También son más elevadas sus
250
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
tasas de circulación al Mg y K y similares para el Na. En A. decorticans, el
calcio y el magnesio son los elementos de flujo más rápido mientras que el N
y el P son los más conservados. El potasio, debido en parte a la importante
contribución del lixiviado en las especies arbustivas presenta un flujo más
rápido de circulación que las especies de pino. A pesar de la existencia de un
flujo más lento de circulación del N y P en A. decorticans, las diferencias
con respecto al Ca y Mg son pequeñas. Esto parece indicar que, de forma
global, A. decorticans es la especie que presenta una tasa de circulación más
elevada para todo los nutrientes incluso para aquellos que podrían ser
limitantes (N y P).
Sólo N en C. laurifolius, P en el pinar y K en P.pínaster poseen tasas de
circulación normalizadas ligeramente inferiores a las de la materia seca
(tabla 4.10b) lo que supone un comportamiento altamente conservativo para
estos elementos. En general, las tasas de circulación de N y P en el pinar y
del N en el matorral son muy similares a los de la materia seca. A.
decorticans posee tasas 1.7 veces más rápidas de N con respecto al peso seco
mientras que la correspondiente al P es del orden del doble. En la j a ra el P
circula 1.4 veces más rápidamente que la masa. Las tasas de circulación del Ca
y Mg son en todos los casos bastante superiores a las de la materia seca
oscilando entre 1.7 y 3.5 veces las de Ca y entre 2 y 3 veces el Mg. Respecto
al K, existen diferencias según se considere el pinar o el matorral. Mientras
que la circulación de K en P. pinaster es muy similar a la de la materia
orgánica y la de P. nigra 1.6 veces mayor, las tasas normalizadas encontradas
en el matorral doblan con creces la correspondiente al peso. La causa
principal de estas diferencias radica en la mayor intensidad del lixiviado en
las especies arbustivas, aspecto ya comentado con anterioridad. Estos
resultados son del mismo orden que los encontrados por Gray (1983) en el
chaparral perenne californiano exceptuando las de N para las que obtiene tasas
ligeramente superiores a las nuestras (1.41). Para el K, Gray encuentra tasas
similares a las especies arbustivas de este estudio (2.08) e inferiores a las
del pinar, mientras que los valores para Ca y Mg son similares siendo estos
dos elementos para los que también se obtienen las mayores velocidades de
circulación en el chaparral.
251
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Al normalizar los flujos se observa que C. laurifolius pasa a presentar
tasas de circulación superiores a A. decorticans en relación al K y Mg. La
tasa para estos dos elementos en la j a ra es la más alta de todas la especies.
La tasa de circulación de fósforo (1.42) está muy por encima de la del N
(0.69) elemento altamente conservado en esta especie, (tabla 4.10b). Esta
característica parece confirmar la baja eficiencia de la j a r a en la
utilización del P y la alta del N, ya comentada en varias ocasiones a lo largo
de este trabajo. Destaca también en C. laurifolius la baja tasa de circulación
de Ca, sobre todo teniendo en cuenta los altos contenidos de Ca en hoja viva y
en hojarasca. La explicación al bajo flujo de circulación se ha de buscar en
los bajos contenidos de Ca que presenta el restos de fracciones de la biomasas
en C. laurifolius (ramas y ramillas) (8 mg/g) frente al (10 mg/g) de hojas.
Evidentemente, resulta extraño que el elevado contenido de Ca en las hojas no
se vea correspondido con mayores porcentajes de este nutriente en fracciones
leñosas que son las que acumulan básicamente el Ca. Habría que buscar una
explicación a estos elevados niveles de Ca en la fracción foliar de C.
laurifolius. La hipótesis más plausible es que parte del Ca analizado en las
hojas no sea interno sino de origen externo. En este sentido, observando las
microfotografías electrónicas de hojas de j a r a se observa una gran cantidad de
partículas depositadas adheridas a su superficie que no desaparecen al lavar
las hojas (Domingo 1991). Este autor comenta que probablemente sean las
resinas que impregnan las hojas de j a ra las responsables \ d e esta eficaz
adherencia. Es muy posible que parte de esta partículas contengan calcio y que
sea la deposición seca la responsable de los altos contenidos foliares de este
elemento. Observando las mismas microfotografías se aprecian así mismo la
presencia de numerosos hongos básicamente en el envés que tampoco desaparecen
con el lavado (Domingo 1991). Algunos autores han constatado la capacidad de
hifas de ciertos hongos para acumular Ca (Klemmedson et al. 1985, Cromack y
Monk 1975). Evidentemente, parte del Ca también puede provenir de estos
organismos.
252
i
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente*
4.3.5. Origen de los iones que penetran en el suelo fores ta l .
Las distintas especies que cubren el suelo forestal pueden ejercer una
marcada influencia en el predominio de alguna de las vías de penetración de
nutrientes (Bellot 1989, Bellot y Escarré 1991). El origen de los distintos
elementos que llegan al suelo puede visualizarse mediante un diagrama ternario
sencillo propuesto por Parker (1983). Este autor utiliza esquemas triangulares
en el que cada lado de un triángulo equilátero constituye una vía de
penetración pero de origen distinto (desfronde, trascolación neta y deposición
global). El desfronde representa la vía interna, la deposición global la
externa mientras que la trascolación neta participa de ambas. Cada región del
diagrama ilustra el diferente movimiento de los nutrientes según sea la
combinación de sus puntos de entrada al suelo. La región cercana al vértice
superior indica el predominio de entrada vía atmosférica, la más próxima a la
base del triángulo representa el reciclado de nutrientes bien por
descomposición (izquierda) o lixiviado (derecha) (figura 4.3). La parte
izquierda exterior a los límites del triangulo delimita la región dominada por
la reabsorción foliar.
Aproximación específica. (Supuesto 1).
Las figuras 4.4 a 4.8 contemplan una primera situación hipotética en la
que el suelo de la cuenca estaría cubierto al 1007. por cada una de las
especies estudiadas. Con este supuesto, puede conocerse la influencia
diferencial de cada una de las especies sobre las vías de penetración de
nutrientes al suelo. Los datos utilizados para la realización de estos
diagramas se muestran en las tabla 4.11 Los aportes por desfronde
corresponden a la caída anual media encontrada en este trabajo, los de
trascolación y deposición global están calculados en base anual a part i r de
los datos de Domingo (1991) para el periodo Abril 1988/Septiembre 1989. Una
consideración a destacar es el importante volumen de deposición global
recogido en el periodo de estudio (718 mm) valor que supera en más de 200 mm
la media anual para esta zona. En consecuencia los nutrientes aportados por
esta vía aunque reales, son claramente una sobrestimación de las posibles
entradas a part i r de volúmenes de lluvia más próximos a los valores medios.
253
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillaro
100% Desfronde 0%
Figura 4 .3 . Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal, según Parker (1983).
La importancia relativa de las t res vías de penetración de elementos al
suelo no variarían esencialmente en la cuenca bien estuviera ésta cubierta de i
P. pinaster o bien de P. nigra (figuras 4.4 y 4.5) tal como lo refleja la
situación de los iones en el triángulo. La única diferencia de posición algo
marcada la constituye el N, elemento que se tranfiere vía desfronde de forma
similar en las dos especies, pero no tanto en las dos rutas de la vía
hidrológica. En P. nigra la deposición global contribuye con un porcentaje
mayor que en P. pinaster al retorno de N al suelo. En consecuencia, y dadas
las similitudes en los porcentajes de desfronde la trascolación neta sólo
supone el 0.79% de los aporte en P. nigra frente al 7.5% de P. pinaster. Por
ello, en esta situación en concreto las trasferencias hidrológicas de N se
verían algo más favorecidas bajo cubierta de P. pinaster dada la menor
254
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
Figura 4.4. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal a través de P.pinaster suponiendo un 1007. de cobertura para
esta especie. Datos tomados de la tabla 4.11.
influencia que P. nigra. ejerce respecto al enriquecimiento en nitrógeno del i
agua de lluvia al atravesar sus copas. Según los resultados de la aplicación
del modelo de Lovett y Lindberg (apartado 4.2), los aportes de N en
trascolación en el pino resinero se deben fundamentalmente a una mayor
eficacia en la captación de deposición seca de nitrato respecto al laricio,
pero sería necesario comprobar si ello se debe a diferencias en determinadas
características específicas entre los dos pinos, o bien, a una más favorable
localización física de los individuos de P. pinaster en el área de estudio.
Los demás iones, presentan comportamientos similares en ambas especies y
su localización en el diagrama se sitúa dentro de los límites apuntados por
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
100% Desfronde 0%
Figura 4.5. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal a través de P.nigra suponiendo un 100% de cobertura para esta
especie. Datos tomados de la tabla 4.11.
Parker (1983) en distintos bosques. Así, en la cuenca Nacimiento cubierta por
pinar de cualquiera de las dos especies, el Ca y Mg se reciclarían básicamente
mediante la descomposición y el K vía lixiviado. El Na presenta un origen
claramente externo situada en la zona definida por Parker como deposición
seca. Los resultados para los cationes son similares a los encontrados por
Bellot (1989) y López (1989) en Quercus ilex en la Sierra de Prades. La
localización del P en el diagrama para las dos especies de pino (figuras 4.4 y
4.5) queda representado en la zona de reabsorción foliar dadas las
trascolaciones netas negativas que se producen en ambas especies y la clara
dominancia del desfronde en el retorno al suelo de este nutriente.
256
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
100% Desfronde 0%
Figura 4.6. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal a través de C.laurifolius suponiendo un 100% de cobertura para
esta especie. Datos tomados de la tabla 4.11.
Si la cuenca estuviese cubierta por matorral monoespecífico bien de A.
decorticans o C. lawifolius (figuras 4.6 y 4.7), la importancia relativa de
las diferentes vías de penetración de nutrientes no diferirían en lo básico
suponiendo una u otra situación a excepción del N y Na. La localización en el
diagrama de los iones para ambas especies arbustivas es similar a la
encontrada por Parker (1983) para un conjunto amplio de bosques. Solamente el
N en la j a ra parece quedar excluido de la región que de forma "ideal" delimita
este autor para el N.
La localización de algunos elementos básicos para la nutrición vegetal
más próxima a la base del triángulo (figuras 4.6 y 4.7) que la encontrada en
257
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
100% Desfronde 0%
Figura 4.7. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal a través de A.decorticans suponiendo un 1007. de cobertura para
esta especie. Datos tomados de la tabla 4.11.
el pinar, indica una menor influencia de la vía externa (deposición global) i
frente a las vías de componente interno. Por tanto una cubierta arbustiva aceleraría el retorno de estos nutrientes esenciales vía vegetación
El Ca y Mg serían reciclados bajo cubierta arbustiva de cualquiera de las
dos especies vía descomposición dado la mayor importancia relativa del
desfronde para estos nutrientes. La importancia del desfronde en C.
laurifolius es superior a la de A. decorticans para Ca y Mg. Existe un ligero
desplazamiento del Ca hacia la derecha del triángulo en A. decorticans (figura
4.7) indicando una contribución nada despreciable de la trascolación neta.
Estas entradas podrían corresponder a lavado de deposición seca dada la
258
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente»
Tabla «.11.
P.pínaster
D. global
Desfronde
T. neta
TOTAL
%dg %df Xtn
P.nigra
0. global
Desfronde
T. neta
TOTAL
%dg %df %tn
Flujos de depoi
supuesto de un
N
6.50
14.98
1.76
23.24
27.97
64.46
7.57
N
6.50
10.54
.13 17.17
37.86
61.38
.76
A.decorticans N
D. global
Desfronde
T. neta
TOTAL
%dg %df %tn
6.50
51.72
1.06
59.28
10.96
87.25
1.79
C.laurifoliua N
0. global
Desfronde
T. neta
TOTAL
%dg %df %tn
6.50
8.27
-3.51
11.26
57.71
73.45
-31.16
;ición global. desfronde y
10OX de cobertura en cada
P
.17 1.41
-.10
1.48
11.52
95.46
-6.98
P
.17
.97 -.15
.99 17.21
97.98
-15.19
P
.17 2.04
.37 2.58
6.59
79.07
14.34
P
.17 1.48
.52 2.17
7.84
68.18
23.98
K
3.63
3.93
4.19
11.75
30.93
33.43
35.64
K
3.63
2.22
4.33
10.19
35.67
21.79
42.54
<
3.63
12.85
21.31
37.79
9.61
34.01
56.38
<
3.63
3.65
11.55
18.83
19.30
19.37
61.33
trascolacion neta en el
especie
Ca
15.02
37.64
21.55
74.21
20.24
50.72
29.04
Ca
15.02
19.20
21.80
56.02
26.81
34.28
38.91
Ca
15.02
20.08
15.26
50.36
29.82
39.87
30.30
Ca
15.02
24.67
4.45
44.13
34.03
55.89
10.08
(Cg/ha.año).
Mg
2.47
7.06
3.57
13.10
18.89
53.88
27.24
Mg
2.47
3.78
2.56
8.82
28.05
42.92
29.03
Mg
2.47
4.72
3.33
10.53
23.50
44.83
31.67
Mg
2.47
3.94
1.50
7.91
31.26
49.78
18.96
Na
4.01
.82 4.89
9.73
41.26
8.43
50.31
Na
4.01
.29 5.67
9.97
40.24
2.95
56.81
Na
4.01
.40 5.34
9.76
41.14
4.12
54.74
Na
4.01
.19 1.29
5.49
73.05
3.40
23.54
259
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
situación en la que se encuentra el Ca en el diagrama que por otro lado
concuerda con los resultados de la aplicación del modelo de Lovett y Lindberg
(1984) al gaznacho (apartado 4.2.2.). La proximidad de fuentes cercanas
emisoras de partículas de carbonato calcico favorecerían esta entrada tanto
por trascolación como por deposición global (Domingo 1991).
El K es reciclado vía lixiviado en ambas situaciones de cubierta
arbustiva mientras que el P se recicla vía descomposición fundamentalmente
(figuras 4.6 y 4.7).Es de hacer notar también la interesante contribución del
lixiviado en ambas especies (14.3% en A. decorticans y 24% en C. laurifolius)
(tabla 4.11) La entrada de Na procede del exterior en ambos tipos de matorral
con predominio de deposición seca en A. decorticans y de la deposición global
en C. laurifolius.
La localización del N en A. decorticans se encuentra muy próxima a la
zona de reabsorción foliar. Aunque el valor de trascolación tenga signo
positivo, es necesario indicar que se t ra ta de un balance entre las distintas
formas de presentarse el nitrógeno en el agua. Las formas nítricas y orgánica
son absorbidas a nivel foliar en esta especie (tabla 4.2), bien por las
propias hojas ó por fauna o flora epífita asociada (Domingo 1991). El
nitrógeno amoniacal es el único lixiviado o lavado en A. decorticans . De ahí
los resultados de signo positivo pero próximos a la región de captación
foliar. La situación del N en C. laurifolius hacia la parte superior del
diagrama indica una fuerte influencia de la deposición global en la entrada de
N frente al desfronde. Si se observa la tabla 4.11, el retorno vía detritus
representa solamente un 21% más de la entrada por deposición global. La ja ra
absorbe nitrógeno del agua de trascolación en sus t res formas (orgánico,
amoniacal y nítrico), de tal manera que un 50% de las entradas aportadas por
la lluvia quedarían retenidas en la cubierta de este matorral si este cubriera
por completo la cuenca, (tabla 4.11). Por ello, es en último término el
desfronde el máximo responsable de los aportes de N al suelo en esta
situación.
La trasferencia de P en los dos supuestos de cubiertas arbustivas para la
cuenca "Nacimiento " difiere sensiblemente con la pauta encontrada para este
260
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente*
elemento en Arbutus unedo y Phyllirea media en la Sierra de Prades para una
situación, también hipotética, de densidades extremadamente altas de esas
especies (Bellot 1989). En el madroño y labiérnaga el P se sitúa en la zona de
absorción foliar o bien muy próxima a ella. En el matorral estudiado en este
trabajo existe claramente un reciclado vía descomposición. Respecto al N, cabe
destacar la mayor intensidad de reabsorción foliar que se detecta en la j a ra
frente a Arbutus unedo y Phyllirea media del trabajo de Bellot (1989).
En resumen, las principales diferencias que podrían apreciarse en la
cuenca "Nacimiento" en función de las diferentes coberturas monoespecíficas de
las cuatro especies estudiadas serían las siguientes:
- En todos los supuestos es el desfronde la principal vía de trasferencia
en relación al N y P. Para el primer elemento este flujo tiene una
contribución relativa mayor en el matorral respecto al pinar, mientras que
para el P ocurre a la inversa. C.laurifolius es capaz de retener buena parte
del N procedente del agua de lluvia con lo que es, en definitiva, el desfronde
la principal vía de trasferencia al suelo en el hipotético jaral . Bajo
cubierta de A. decorticans, también existe absorción de N procedente de la
deposición global en sus formas nítrica y orgánica. Sin embargo el agua de
lluvia se enriquece con cantidades superiores de N-NH4 provocando un balance
de signo positivo en favor de la lixiviación pero con valores absolutos muy
pequeños.
- En una cuenca cubierta al 100% por A. decorticans y G. laurifolius, el
P se reciclaría básicamente por descomposición (figuras 4.6 y 4.7). Para
ambas especies también el lixiviado aporta contribuciones nada despreciables,
sobre todo en la jara, respecto a la trasferencia de nutrientes al suelo. Sin
embargo, bajo cubierta de pinar, la única fuente destacable de este nutriente
sería el desfronde debido a que la practica totalidad del fósforo que entra
por deposición global es absorbido por las copas (figuras 4.4 y 4.5).
- Ca y Mg se transfieren preferentemente vía desfronde en las cuatro
situaciones aunque para el matorral de A. decorticans y en menor medida en el
pinar de P. nigra un porcentaje importante proceda de la trascolación neta,
261
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
probablemente vía deposición seca. El reciclado de ambos nutrientes se
produciría fundamentalmente vía descomposición en los cuatro supuestos.
- En todos los supuestos, la trascolación neta juega un papel fundamental
para la trasferencia de K pero ésta es mucho más patente bajo cubierta
arbustiva.
- En todos los casos, el Na tiene un claro origen externo. La captación
de deposición seca tendría una mayor importancia si la cuenca estuviese
cubierta por cualquiera de los dos pinos ó por A. decorticans que si lo
estuviera por el jaral .
Aproximación por regiones. (Supuesto 2).
Consideraremos ahora una situación real en la cuenca que hemos dividido
en las dos grandes áreas definidas según la especie de pino que las cubre RPP
y RPN). Con este planteamiento pretendemos evaluar la contribución real de
cada una de las especies en estas dos regiones. El área 1 (RPP) correponde a
la zona repoblada con P. pinaster que abarca los sectores 2, 6 y 7 con una
superficie total de 11.1 Ha. El aréa 2 (RPN) corresponde a la repoblación de
P. nigra con una superficie de 22.8 Ha correspondiente a los sectores 1, 3 y
4. En ambas localizaciones está presente el matorral de C. laurifolius y A.
decorticans pero existe una clara diferenciación en cuanto a la estructura de
las dos especies en cada una de las localizaciones sobre todo en relación a la
densidad y porcentaje de cobertura. Sin embargo respecto al pinar las
diferencias no son tan acusadas. Los datos relativos a la densidad, cobertura,
así como la contribución de los flujos de deposición global, desfronde y
trascolación neta pueden observarse en la tabla 4.12 para el aréa 1 y en la
tabla 4.13 para el área 2. En las figuras 4.8 y 4.9 se han construido los
diagramas de Parker para ambas situaciones a par t i r de los datos de las tablas
anteriores.
En ninguno de los dos supuestos existe una especie claramente dominante
sobre las otras dos. Por ello en una primera visualización de los dos
diagramas y comparándolos con los de las figuras (4.4 a 4.7), se observa un
262
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutriente!
Tabla 4.12. Flujos de deposición global, desfronde y trascolación neta (kg/ha.año)
en RPP teniendo en cuenta el porcentaje de cobertura (*) de cada especie.
P.pinaster N P < ca Mg Na
DGCKg/ha.año
DF TN TOTAL
%dg %df Xtn
6.50
8.06
.94 15.50
41.94
52 6.06
.17
.76 -.06
.87 19.47
86.83
-6.30
3.63
2.11
2.25
8.00
45.42
26.42
28.16
15.02
20.25
11.60
46.87
32.05
43.21
24.74
2.47
3.80
1.92
8.19
30.21
46.36
23.43
4.01
.44 2.63
7.09
56.63
6.22
37.15
A.decorticans
DGCKg/ha.año
0F TN TOTAL
%dg %df %tn
6.50
8.95
.19 15.64
41.56
57.22
1.21
.17
.35
.06
.59 28.96
60.13
10.90
3.63
2.22
3.69
9.54
38.07
23.30
38.63
15.02
3.47
2.64
21.13
71.07
16.44
12.49
2.47
.82
.58 3.87
63.97
21.11
14.92
4.01
.07
.92 5.01
80.16
1.39
18.45
C.laurífolius
DGCKg/ha.año
DF TN TOTAL
%dg %df %tn
6.50
3.31
-1.39
8.42
77.20
39.31
-16.51
.17
.59
.Z^
.97 17.50
60 21
.88
.62
3.63
1.46
4.62
9.71
37.41
15.02
47.56
15.02
9.87
1.78
26.66
56.33
37.00
6.67
2.47
1.58
.60 4.65
53.20
33.89
12.91
4.01
.07
.52 4.61
87.14
1.62
11.23
(*) X Cobertura: PP:53 X ; AO: 17.3 X ; CL: 40 X
263
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 4.13. Flujos de deposición g loba l , t rascolación neta y desfronde (Kg/ha.año) en RPN teniendo en cuenta el porcentaje de cobertura de cada especie.(*)
P.nígra
DG(Kg/ha.año DF TN TOTAL
Xdg %df %tn
A.decorticam
DG(Kg/ha.ano DF TN TOTAL
Xdg %df %tn
C. laur i fo l i iB
DGCKg/ha.año DF TN
TOTAL
%dg Xdf Xtn
N
6.50 4.90
.05 11.45
56.77 42.79
.44
6.50 22.40
.45 29.35
22.15 76.32
1.53
6.50 1.80 - .76 7.54
86.26 23.83
-10.09
P
.17
.45 - .07
.55
30.80 81.52
-12.32
.17
.88
.16 1.21
14.01 72.80 13.19
.17
.32
.11
.60
28.29 53.40 18.31
<
3.63 1.03 2.02 6.68
54.39 15.45 30.16
3.63 5.57 9.23
18.42
19.72 30.21 50.07
3.63 .79
2.51
6.93
52.43 11.42 36.15
Ca
15.02 8.93
10.14 34.09
44.06 26.20 29.74
15.02 8.69 6.61
30.32
49.53 28.67 21.79
15.02 5.35
.96 21.34
70.39 25.08 4.52
Mg
2.47 1.76 1.19 5.42
45.60 32.45 21.95
2.47 2.04 1.44 5.96
41.50 34.28 24.22
2.47 .85 .33
3.65
67.70 23.39 8.91
Na
4.01 .14
2.64 6.79
59.15 2.02
38.83
4.01 .17
2.31 6.50
61.75 2.68
35.57
4.01 .04 .28
4.33
92.59 .94
6.47
(*) X de cobertura: PN: 46.5X ; A0: 43.3 X; CL: 21.7 X.
264
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrientes
N D
D D
N
&/Na\
/ N a \
JZa > it
Mg* a Na Mg
• K
K * Ca
> , ^,
P. pin áster *
C.laurifolius o
A.decorticans*
\ t f
Yc> \ XX V> \<>
K Y? Mg
P *
100% Desfronde
Figura 4.8. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal en la RPP de la cuenca "Nacimiento". Datos tomados de la tabla
4.12.
notable desplazamiento de la mayoría de los iones hacia la mitad superior del
triángulo de tal forma que casi todos los elementos a excepción del P quedan
incluidos en la región correspondiente a entradas de origen externo.
Evidentemente las especies más afectadas por esta situación son las que
contribuyen con una menor importancia estructural (densidad, cobertura) a cada
una de las áreas. En la zona 1 (RPP) este desplazamiento es muy acusado en A.
decorticans y en la zona 2 (RPN) en C. laurifolius.
En RPP, P.pinaster es la única especie que ejerce un control bastante
importante sobre los cationes Ca y Mg debido a la importancia relativa que aún
sigue ejerciendo el desfronde (tabla.4.12). Estos iones quedan incluidos en la
265
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
P.nigra *
C.laurifo/ius O
A.decorticans *
100% Desfronde Figura 4.9. Diagrama para comparar las vías de penetración de nutrientes al
suelo forestal en la RPN de la cuenca "Nacimiento". Datos tomados de la tabla
4.13.
zona del triángulo que representa el reciclado vía descomposición. Por su
parte, la contribución del retorno de P vía desfronde e incluso vía
trascolación neta en C. laurifolius es aún manifiesta con lo que este
nutriente sigue localizándose en la región que representa el reciclado aunque
en una posición mucho más superior a la que se contemplaba en la figura
4. Respecto al K, la trascolación neta sigue siendo la vía principal de
entrada en C. laurifolius. El P en P. pinaster se incluye en la región de
absorción foliar, pero ésta ha pasado de suponer el 59% de las entradas de P
en lluvia a un 35%
En el RPN, los cambios con respecto a la situación hipotética de un 100%
266
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Retorno de nutrlentei
de cobertura no son excesivamente drásticos en A. decorticans sobre todo en
relación a algunos nutrientes importantes (P, K y N). Los dos primeros se
mantienen incluidos en la región que delimita el reciclado, (aunque con rutas
diferentes cada uno) mientras que el N, a pesar de una mayor influencia de la
entrada externa se sigue aportando en desfronde con contribuciones relativas
del 76% superiores a las otras especies incluidas en el RPN (437. en P. nigra y
24% en C. laurífolius) (tabla 4.13) El desplazamiento de los iones en C.
laurifolius es drástico en la zona 2 si exceptuamos la contribución aún
notable de la trascolación neta respecto al K y el desfonde respecto al P. En
P. nigra únicamente el P responde a la situación modélica de Parker.
Podemos concluir a la vista de estos últimos resultados que la
repoblación de pinar en el momento actual no constituye una vía de
trasferencia excesivamente importante en relación a la entrada total de
nutrientes al suelo sobre todo en el caso de P. nigra. El mayor control parece
ejercerse sobre el Ca y Mg en P. pinaster. En ambas zonas la deposición global
supone una importante fuente de nutrientes. Hay que tener en cuenta que
durante el período de estudio el flujo de entrada externa ha sido mucho más
elevado de lo normal en la zona dados los altos volúmenes de precipitación.
Por ello, en años más secos, la situación descrita aquí podría variar
sustancialmente en favor de una mayor importancia porcentual de los flujos
puramente internos en esta especies.
En relación al matorral hay que destacar el papel ejercido por A.
decorticans respecto a la circulación de elementos tan importantes como P, N y
K. Con tan sólo coberturas del 40% (Área 2) esta especie se constituye en la
principal vía de trasferencia al suelo de estos nutrientes. Por su parte la
cubierta de C. laurifolius sólo ejerce una influencia notable en el área 1
dónde constituye la principal vía de trasferencia de P y K al suelo de la
cuenca.
267
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
268
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 5 Descomposición.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
5.1 INTRODUCCIÓN.
El término general de descomposición se utiliza para describir un
numeroso grupo de procesos interelacionados que ocurren a nivel del suelo y
que en muchos aspectos son aún pobremente conocidos (Waring y Schlesinger
1985). La descomposición en ecosistemas ter res t res es la mayor fuente de
energía y nutrientes para el suelo y los organismos que lo habitan. Constituye
la principal vía de reciclado de nutrientes disponibles para la absorción
vegetal (uptake) y para la formación de la materia orgánica del suelo. A pesar
de su importancia, uno de los aspectos menos conocido del proceso es el papel
que juegan los distintos grupos de organismos descomponedores (Lousier y
Parkinson 1976, Takeda 1988). En síntesis, la descomposición consiste en un
proceso dinámico que comporta la rotura, pérdida de masa y cambio químico de
un sustrato orgánico, concretamente de la hojarasca y otros detritus.
Swift et al. (1979) describen t res principales procesos que participan en
la descomposición y que están regulados por un numeroso grupo de factores
tanto bióticos como abióticos: <AI
» •«
1 , 7 A
c
L
MU
0 0
»c
" i
(8)
I, M V . » t,
71 g <3 Fia. 2.2 (A) Decomposition of a resource (R) over a short period of time ('i to t*). Tne three component processes, cataboJám (K), comminution ( O and Icaching (L)result in chemkaJ chantes, e.g. mincfaiisation fiyinf rúe toinorfantc forma (fN) and the resyntnesá of decomposcr (íssues (DO) and humus (HLT>. physicaJ changes. e.g. reduction of particle size of chernically unchanged litter (/fC) and the removal of soluble resource maceríals in unchanged form to other sites (R0- (B) Decomposition of a resource over a longer period of time (¡i to O , resulting in complete mineralisation.
269
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
LIXIVIADO.- Proceso abiótico mediante el cual la fracción soluble del
sustrato (recurso) es liberada por la acción del agua.
CATABOLISMO.- Proceso enzimático de transformación de las moléculas
complejas del recurso en otras más simples. A largo plazo, el resultado final
de este proceso sería la oxidación completa de las moléculas (CO , NO , PO ).
A más corto plazo el resultado es una mezcla de productos totalmente oxidados
(mineralizados), con otros que sólo lo están parcialmente y que pueden ser
utilizados por los descomponedores, incorporados al suelo para formar el humus
o bien lixiviados.
FRAGMENTACIÓN.- Proceso físico que puede acompañar o no a los anteriores.
Es llevado a cabo básicamente por la actividad alimentaria de la fauna
descomponedora.
Estos t res procesos actúan de forma simultánea y, aunque en ciertas
etapas predomine alguno de ellos en la practica resulta difícil diferenciar su
efecto individual.
La tasa y patrón de descomposición están reguladas por muchos factores.
Según Swift et al. (1979) éstos pueden agruparse en dos categorías: 1°,
factores físico-químicos tanto ambientales como del propio suelo y 2 , calidad
del sustrato. En relación con este último, autores básicamente escandinavos
(Berg 1986, Berendse et al. 1987, Berg y Staaf 1980) han destacado que
mientras el nivel de nutrientes, sobre todo aquellos más limitantes para el
crecimiento microbiano (N, P, S), regulan los primeros estadios de la
descomposición, las últimas fases son controladas por la dinámica de la
lignina.
La metodología empleada en los estudios sobre descomposición es variada.
La incubación de la hojarasca en bolsas con distintos tamaños de malla es la
más frecuentemente empleado ya que permite la comparación de datos entre
distintas especies y localidades. Además es un método sencillo y barato. Para
los estudios faunísticos y microbiológicos son preferidos aquellos métodos de
incubación que no confinan la hojarasca. Una segunda metodología consiste en
la medida de la masa y de nutrientes en el desfronde y el mantillo estimando
el porcentaje perdido de esos parámetros mediante un sencillo cociente. Otros
270
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
métodos menos utilizados son los isótopos radiactivos o la descomposición de
sustratos estándar. Estas y otras técnicas están revisadas en Swift et al.
(1979).
Las condiciones climáticas en ecosistemas mediterráneos podrían hacer
esperables bajas tasas de descomposición. Durante la estación húmeda invernal,
la actividad microbiana está limitada por las bajas temperaturas, mientras que
en el periodo estival lo está por la baja humedad de suelo (Gray y Schlesinger
1981). Los mecanismos de retraslocación de nutrientes como el N y el P están
bien desarrollados en especies mediterráneas (Read y Mitchell 1983),
favoreciendo su inmovilización en la hojarasca y no su mineralización. Además
el predominio de vegetación esclerófila (con elevados porcentajes de
compuestos recalcitrantes difícilmente descomponibles) también favorece el
acumulo de materia orgánica en el suelo. Sin embargo Gray y Schlesinger (1981)
comentan que la descomposición anual en estos sistemas, es más alta de lo que
podría esperarse en el contexto de las condiciones antes citadas. A este
respecto citan varios trabajos para los que el tiempo de residencia tanto de
materia orgánica como de nutrientes en la hojarasca no difiere de manera
significativa con los encontrados para bosque templados de similar latitud.
Parece que la poblaciones decomponedoras pueden responder de forma rápida e
intensa durante los cortos periodos en que las condiciones son favorables para
la descomposición en la zonas mediterráneas. De todos modos son relativamente
pocos los estudios de descomposición que se han abordado es zonas
mediterráneas para llegar a unas conclusiones generales. Existen trabajos de
este tipo en el chaparral californiano (Schlesinger y Hasey 1981, Schlesinger
1985,) en bosques de Quercus ilex en el sureste francés (Lossaint y Rapp
1971), en el scrub australiano (Meggs y Pearson citado en Gray y Schlesinger
1981). En España, se han realizado estudios en el macizo del Montseny (Verdú
1984), en Cistus la.dan.ifer y otras arbustivas de Extremadura (Núñez 1989,
Lavado et ai. 1988), en distintos bosques y dehesas de la provincia de
Salamanca (Garrido et al. 1989, Escudero et al. 1985), en la depresión central
catalana (Rosich et al. 1989), en bosques caducifolios del Moncayo (Carceller
et al. 1989), en bosques de pino silvestre y acebo en los Pirineos (Alvera
1990) y en repoblaciones de Pinus halepensis en la provincia de Alicante
(Pastor, com. personal).
271
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
El objetivo principal de este capítulo es la realización de un estudio
comparativo sobre la dinámica de la descomposición de hojarasca en las cuatro
especies mayoritarias de la cuenca "Nacimiento". Para ello se aborda en primer
lugar el estudio de pérdida de peso en la hojarasca y su modeiización
utilizando ecuaciones exponenciales simples y dobles. En la segunda parte del
capítulo se t ratan los aspectos referentes a la dinámica de acumulación y
liberación de los nutrientes durante el proceso de descomposición a lo largo
del periodo de estudio en las cuatro especies.
5.2 MATERIALES Y MÉTODOS.
El desfronde foliar se recolectó en la época de máximo caída fenológica
en las especies estudiadas. Las hojas de A. decorticans y C. LaurifoLius se
recogieron en Agosto de 1988 y las de los pinos a finales de Septiembre del
mismo año. Se aprovechó para ello el material caído en las trampas de
desfronde durante esos meses.
Las hojarasca de cada una de las especies se secó al aire hasta peso
constante. Tres submuestras de cada especie se secaron en la estufa a 75 para
calcular la relación peso seco/peso secado al aire y se utilizaron para
análisis químicos.
Para abordar la evolución del peso y nutrientes en la hojarasca se ha
empleado el método de incubación de ésta en el campo mediante bolsas
fabricadas con malla de fibra de vidrio. Esta metodología, introducida por
Bocock et al. (1960) ha sido utilizado no solamente para material foliar sino
también para ramas e incluso suelo (Bockeim y Leide 1986). Es un método útil
para propósitos comparativos y ha sido ampliamente empleado para el análisis
de la descomposición en diversos ecosistemas (Read y Mitchell 1983). El método
se basa en introducir cantidades conocidas del material a estudiar en bolsas
de nylon o fibra de vidrio con malla de tamaño variable que a su vez se
colocan en la superficie del suelo forestal. Periódicamente se recoge un
número determinado de estas bolsas y los contenidos son analizados para
cuantificar la pérdida de masa y de nutrientes. El tamaño de malla apropiado
ha sido siempre motivo de controversia ya que se debe intentar que el poro no
272
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
sea excesivamente pequeño para impedir la entrada a organismos descomponedores
o fragmentadores, ni demasiado grande que permita las pérdidas de fragmentos o
la entrada de material externo. La mayor parte de ios autores utilizan Imm de
poro como el tamaño de compromiso que equilibra lo máximo posible estas dos
tendencias (Sharma y Ambasht 1987, Schlesinger 1985, Stohlgren 1988).
En este estudio se han utilizado bolsas de 15 x 20 cm fabricadas con
fibra de vidrio con malla de lmm y cerradas mediante una lengüeta. Se
rellenaron 30 bolsas con hojarasca secada al aire de cada una de las especies.
En las destinadas a P. pinaster, P. nigra. y A. decorticans se introdujeron 5 g
de hojarasca. Sin embargo, para la j a ra se colocaron 10 g. La razón es que
esta especie tiene hojas más grandes y con solamente 5 g en las bolsas, éstas
quedaban muy sueltas y separadas. En el campo, las hojas de jara se encuentran
de manera mucho más agrupada en la superficie del suelo. Por ello, se optó por
introducir una cantidad superior en las bolsas de esta especie.
Para las incubaciones en el campo, se utilizaron las mismas parcelas en
donde se realizaban los muéstreos de trascolación (Domingo 1991). Una de las
parcelas se localiza en la parte alta y más llana de la cuenca y contiene a
tres de las especies (P. pinaster, C. laurifolius y A. decorticans). Se
escogió un individuo al azar de cada una de estas especies bajo cuya cubierta
se colocaron las treinta bolsas formando un cuadrado de 6X5 bolsas. La parcela
de P. pinaster se localiza a una menor altitud (1600 msnm) y tiene una mayor
pendiente. En ésta parcela se procedió de la misma manera que en la de la
parte alta. El comienzo de las incubaciones en el matorral se inició el 28
Agosto de 1988 y en el pinar el 25 de Octubre del mismo año. Los muéstreos
finalizaron en Diciembre de 1990. Sin embargo la evolución de los nutrientes
sólo se analizó hasta Mayo 1990.
Con una periodicidad bimensual (con algunas excepciones), se recogían dos
bolsas al azar que se introducían en bolsas de papel y se transportaban al
laboratorio. Las muestran eran cuidadosamente sacadas de las bolsas eliminando
en la medida de lo posible las partículas de suelo y otros materiales
adheridos. Posteriormente eran secadas en la estufa a 75" y pesadas apreciando
hasta 0.001 g.
273
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Análisis químico.
Después de pesar las muestras éstas eran tr i turadas individualmente en un
molinillo convencional. Una submuestra de 0.2 g era utilizada para determinar
el N total mediante el método Kjlheldal (Alien et al. 1974). Otra submuestra
de 0.5 g se utiliza para el analizar el contenido en cenizas, P y cationes
(Na, K, Ca y Mg). La acenización se realiza por via semihúmeda en mufla a
450°. Las cenizas se disolvieron utilizando 5 mi de C1H 207. y 5 mi de NO H 5N 3 2
enrasando con agua desionizada hasta 50 mi para preparar las soluciones
madres. El P se analizó mediante el método colorimétrico molibdato-
metavanadato amónico (Alien et al 1974) en un espectrofotómetro U.V. Los
cationes se determinaron con espectrofotometría de absorción atómica (Alien et
al 1974, Ximenez 1980). En los patrones de Ca se añadió un 0.1 7. de lantano
con el fin de evitar interferencias. Para un número considerable de muestras
escogidas al azar, se realizaron replicados a distintos niveles (muestra
molida, solución madre y repetición de análisis químicos).
5.3 RESULTADOS.
5.3.1. PERDIDA DE PESO.
5.3.1.1. Generalidades.
Pérdida de peso en las bolsas.
i
Los cambios y la evolución de la hojarasca contenida en las bolsas para
las cuatro especies puede observarse en las tablas 5.1a y 5.1b y figuras 5.1a
y 5.1b. Los valores son medias de los dos replicados de cada especie en cada
muestreo. Al final del periodo de incubación (770 días), las acículas de los
pinos habían perdido porcentajes no demasiado similares de peso seco inicial
(25.5 7. en P. pinaster y 34.7 7. en P. nigra). Las especies arbustivas, sin
embargo, muestran diferencias todavía más notables en este aspecto. A los 738
días A. decorticans había perdido ya un 70 7. de la masa inicial lo que le
acerca al comportamiento de especies caducifolias caracterizadas por hojas
menos consistentes. El peso perdido por la ja ra fue más moderado, el 38Z a los
738 días, valor que le acerca más al comportamiento de otras arbustivas
274
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
mediterráneas con hojas con un mayor grado de esclerofilia y a P. nigra.
A los 239 días, la pérdida de peso en las especies de pino es idéntica
(12.9 %), (figura 5.1a, tabla 5.1a). A part ir de ese momento, aparecen
oscilaciones en los porcentajes de pérdida de peso de una especie con respecto
a la otra alternándose periodos en los que los valores son superiores en un
pino con periodos en el que lo son en el otro. Puede comprobarse
estadísticamente si existe una tendencia significativa a que los valores
observados para una especie, (expresados en este caso como porcentaje de masa
inicial remanente) se encuentren por encima/debajo de los de la otra.
Aplicando el test t de muestras emparejadas para todo el periodo se ha
constatado que ambas curvas no difieren desde el punto de vista estadístico
(t= 2.00, ns). Por lo tanto, para el conjunto del ensayo existe una
compensación de estas oscilaciones en los valores observados.
Por su parte, en el matorral, los valores de pérdida de peso son siempre
superiores en A. decorticans respecto a C. Laurifolius (o inferiores si
hablamos de % de peso seco inicial remanente). El test t de muestras
emparejadas indica claramente una diferencia significativa entre ambas curvas
(t=13.4, p< 0.001).
Tasas diarias de pérdida de peso.
La tasa de pérdida de masa diaria fue calculada en base al porcentaje de
peso seco inicial que va quedando cada día, (tabla 5.2), observándose
fluctuaciones en las tasas de descomposición entre periodos de recogida para
cada especie. Algunos autores han detectado patrones cíclicos en estas tasas
que han resultado estar relacionadas con condiciones ambientales tales como
precipitación y temperatura media, (Witkamp y Van der Drift en Klemmedson et
al. (1985), Klemendson et al. 1985). En las especies estudiadas, las tasas
diarias de descomposición no parecen seguir un patrón cíclico o estacional
claro (figuras 5.2a y 5.2b). Solo en el pino laricio parece existir una cierta
tendencia a que las tasas sean superiores en los meses de Noviembre (primer y
segundo año) y Diciembre (tercer año) (figura 5.2b).
Los picos de máxima descomposición diaria se producen en el primer
275
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
o
o o
o
o
oo
03 oo 03
U1 O
03
R oo
00
s 5 £
o
ff «— m
a «O 00 LA « -
o o m fM > N Í O o T - r - r - N S «O
o> O CO co
^ o c o N . f o o m o o c o r v . i r i i n c o ^ r g r \ j o * o r ^
>o r-- «— ro oo oo oo eo oo h»
s in N oo js* N. N- N.
O* U1 C\J N. lf\ *T m N S S -O N >0 "O
> «- m o *- *T *-
£ R v- 00
c a.
00
N tn i/i o> in in o o ro m «— in
oo oo oo oo oo oo co -
< o
O í - « - T - f - N K ( O M I O K 1 tO n <- <" *- ^ N N M fO M * f < i n m i n i n , O ^ N N
o o « - « — ( M r v ^ r s r ^ o - o ^ o » * — «-<— m K í i n i n o ^ ^ r u í M
276
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición
o u m •o
2
&
00 sr
8¡
in i-
IA «—
«- M
o> N. eo ro co r>-
» < >» o » . - ©• «- «-
c» o o
- f T -
*- O
O» T -
- §
o o
° ° «; ^- ^ oo
co
N- 00 O. O* co in
S M B rv» o» fs-
rvj co o u"» o *- in (N- N . fs. *Q fs«- N- NO
O- O-O CO CO O CK S . N-
ifl in co eo »• *-
>*• ^ í N- N. *- * - O O o o co co "P r* O -O m IÍI m m
<o <o co r\j (\J ro NO >0 r*
CO 00 co co co
o o o o o o * - ^ * e M r \ j ^ j - ^ j ' - 0 ' O O ^ o v r - r - ^ r O M i n i n ^ t > ( \ j w
277
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Figura 5.1. Masa remanente (en 7. de peso seco inicial) en las especies
estudiadas (valores medios en cada muestreo).
278
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.2 Tasas medias de pérdida de peso diario (% respecto al peso inicial): k, en las cuatro especies durante el período de muestreo.
mespin
10
11
2
4
6
9
11
1
3
5
9
12
kPP
0
-.2845
-.0335
-.0002
-.0218
-.0103
-.0466
-.0607
-.0745
-.0193
.0366
-.0442
kPN
0
-.2605
-.0572
' .0208
-.0225
-.0460
-.0838
-.0189
.0511
-.0842
-.0239
-.0674
mesmat
8
10
11
2
4
6
9
11
1
3
5
9
12
kCL
0
-.191
-.166
-.032
-.062
-.032
-.079
.055
-.076
.041
-.092
-.017
-.065
kAD
0
-.431
-.707
.001
-.033
-.085
.028
-.066
-.152
-.067
-.078
.025
,
mespin: meses correspondientes a las recogidas en el pinar. mes mar: ídem para el matorral. PP: P. pinaster; PW: P. nigra: CL: C. laurifolius: AD: A. decorticans
período de recogida (pinar) o en los dos primeros periodos (matorral). Al
comienzo de la incubación, los compuestos fácilmente lixiviables se pierden
rápidamente (Swift et al. 1979). En nuestro estudio, la lixiviación en los
primeros estadios de descomposición podría haberse visto favorecida por las
precipitaciones otoñales que coinciden con los primeros meses de muestreo
trayendo como consecuencia una altas tasas de descomposición en esos momentos.
Para algunos de los muéstreos existe una ganancia de peso (valores
positivos de la tabla 5.2). Este incremento de masa no es, por supuesto, un
incremento en el peso de la hojarasca incial, sino que puede derivar de
distintas fuentes de contaminación. Klemmedson et al. (1985) citan varias
referencias en las que se han encontrado como causas a estos incrementos la
279
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
a,
b.
i I k, P. pinaster k, P. nigra
-0,2 -
C.faurifolius k, A. decorticans
Figura 5.2. Tasas medias diarias de pérdida de peso (k) a lo largo del período
de muestreo (en 7. de peso seco inicial), a: Pinar, b: Matorral.
280
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
acumulación de polvo, aumento de la biomasa de heterótrofos asociados a la
hojarasca, contaminación por hojarasca reciente etc. En este estudio las
contaminaciones más acusadas se produjeron en Marzo de 1990 para P. nigra y C.
laurifolius (+0.057. y +0.04 7.psi/día) y en Noviembre pra C. lawifolius (+0.05
7. psi /dia) .
Variabilidad de la pérdida de peso en las bolsas.
La variación en la pérdida de masa se incrementa con el tiempo tal y como
lo refleja el aumento en el coeficiente de variación (figuras 5.3a y 5.3b,
tabla 5.3). Los coeficientes de variación son una manifestación de la
variación de las condiciones locales a las que se ve sometida la hojarasca en
el trascurso del proceso de descomposición en cada boisa y ha sido observado
por otros autores (Klemmedson et al. 1985, Dziadowiek 1987, Takeda 1988).
Según este último autor, este aumento en la variabilidad puede estar causado
por los efectos acumulativos que provocan las condiciones cada vez más
heterogéneas del microhabitat en el que ocurre el proceso de descomposición.
Parece que la pérdida inicial de masa en los primeros estadios del proceso
está estrechamente relacionado con las "calidad " química del sustrato, (Berg
1986, Berg and Lundmark (1987). Dado que al principio existen unas condiciones
de mayor homogeneidad en la "calidad" de este sustrato para las distintas
réplicas, la variabilidad en la pérdida de peso entre estos replicados en un
momento determinado de esta fase inicial será menor. El incremento de
variabilidad con el tiempo puede representar la contribución cada vez más
importante de agentes bióticos en el curso del proceso y puede considerarse
como un índice de la actividad de la microflora, microfauna y microganismos
(Takeda 1988).
En todas las especies estudiadas el CV aumenta considerablemente a partir
del primer año (figuras 5.3a y 5.3b). Los pinos registran valores máximos de
CV de 5.4% en P. pinaster y 8.6 7. en P. nigra pero que en todo caso resultan
inferiores a los registrados en el matorral (10.47. de C. laurifolius y 15.37.
de A. decorticans). Por otro lado, los CV medios para cada especie a partir
del primer año (392 días) son bastante superiores en A. decorticans (8.297.)
que en las otras tres especies (3.57. en P. pinaster, 2.96 7. en P. nigra y 3.8
en C. laurifolius). Esta característica parece estar en consonancia con la
281
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
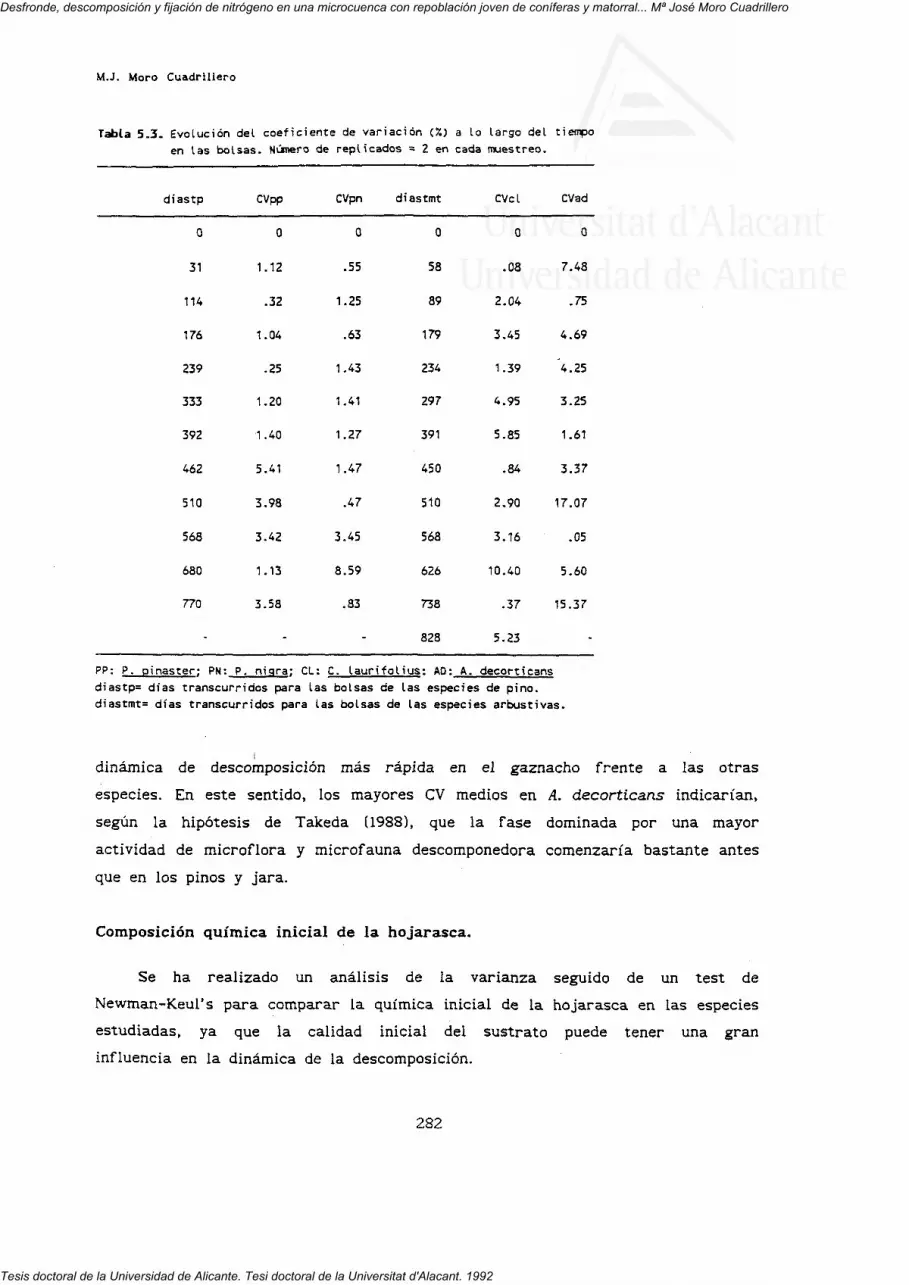
M.J. Moro Cuadrillero
Tabla 5 .3 . Evolución del coef ic iente de var iación (%) a lo largo del tiempo en las bolsas. Número de replicados = 2 en cada muestreo.
diastp
0
31
114
176
239
333
392
462
510
568
6S0
770
-
CVpp
0
1.12
.32
1.04
.25
1.20
1.40
5.41
3.98
3.42
1.13
3.58
.
CVpn
0
.55
1.25
.63
1.43
1.41
1.27
1.47
.47
3.45
8.59
.83
.
diastmt
0
58
89
179
234
297
391
450
510
568
626
738
828
CVcl
0
.08
2.04
3.45
1.39
4.95
5.85
.84
2.90
3.16
10.40
.37
5.23
CVad
0
7.48
.75
4.69
'4.25
3.25
1.61
3.37
17.07
.05
5.60
15.37
99: 9. p inaster: PN: 9. n igra : CL: C. l a u r i f o l i u s : AD: A. decorticans diastp= días transcurr idos para las bolsas de las especies de pino. diastmt= días transcurr idos para las bolsas de las especies arbust ivas.
i
dinámica de descomposición más rápida en el gaznacho frente a las otras
especies. En este sentido, los mayores CV medios en A. decorticans indicarían,
según la hipótesis de Takeda (1988), que la fase dominada por una mayor
actividad de microflora y microfauna descomponedora comenzaría bastante antes
que en los pinos y jara.
Composición química inicial de la hojarasca.
Se ha realizado un análisis de la varianza seguido de un test de
Newman-Keul's para comparar la química inicial de la hojarasca en las especies
estudiadas, ya que la calidad inicial del sustrato puede tener una gran
influencia en la dinámica de la descomposición.
282
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

a Descomposición.
b.
10 c.v.
0 50 100 150 200 250 300 350 400 450 500 550 600 650 700 750 800
Días transcurridos.
-&— P. pinaster --*•- P. nigra
20
15
10
C.V.
—
-
Ilí—
*
y*'
,*-•
i i
*---
i
" • - * ' '
i i
V *
*
; •
I -L*—
/ \ /
A \
i i
A
i s í i
O 50 100 150 200250300350400450500550600650 700 750800850
Días transcurridos
—e_ C. Laurifolius ••*•- A. decorticans
Figura 5.3. Evolución del coeficiente de variación (7.) del peso de la
hojarasca en las bolsas de pinar (a) y de matorral (b), a lo largo del período
de estudio.
283
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Mediante el ANOVA se ha constatado que existen diferencias significativas
(p<0.001) entre especies para el N, P, K y contenido de cenizas (F=117.23,
24.02, 78.66 y 40.88 respectivamente) y con p< 0.005 para Ca y Na (F= 13.25 y
13.87 respectivamente). La concentración de Mg no difiere de manera
significativa en las especies estudiadas (F= 0.33, ns). Aplicando a posterior
el test de Newman-Keul's puede discriminarse entre qué especies se producen
estas diferencias, (tabla 5.4). Los resultados indican que la hojarasca de A.
decorticans tiene concentraciones medias de N y K superiores (1.595% de N y
0.668 de K) al resto de las especies. C. laurifolius posee concentraciones
significativamente más altas de P, Ca y cenizas (0.147., 2.067. y 8.26 7.
respectivamente) a las demás especies. P. pinaster y P. nigra, con niveles de
nutrientes bastante bajos solo se diferencian de una manera significativa
rspecto al Na en el que P. pinaster presenta concentraciones más elevadas
(0.022%). Las especies de matorral no presentan diferencias en los niveles de
Na y éstos son significativamente superiores a los del pinar. Las
concentraciones de K en A. decorticans y C. laurifolius (0.338% y 0.689%) y de
cenizas (8.267. y 7.077.) aún siendo distintas entre ambas especies son,
superiores a las encontradas en el pinar.
En relación con la química inicial de la hojarasca, estos resultados
indican que las dos especies de pino pueden considerarse como un grupo
homogéneo con bajos niveles de los principales macronutrientes cuyos cuyos
contenidos son por lo demás muy similares. Respecto al matorral, no puede
hablarse de homogeneidad entre ambas hojarascas sino maás bien todo los
contrario. Quizá la única característica que compartan se refiere a que en
general los niveles de nutrientes son superiores a las del pinar. Existen
diferencias muy patentes entre ellas sobre todo en lo que se refiere al nivel
inicial de nutrientes importantes en la dinámica de la descomposición, (N y P)
por lo que parecen esperables comportamientos distintos en el patrón de
pérdida de peso y en la mineralización de estos nutrientes durante el proceso.
En la tabla 5.4 se muestran también los valores del índice C/N inicial
calculado suponiendo que la materia orgánica de las hojas contiene un 507. de C
(Alvera 1990, Schwintzer 1984). El valor superior corresponde a P. pinaster
(198) ya que es el que posee concentraciones más bajas de N, P. nigra y
C.laurifolius ocupan posiciones intermedias con valores bastante parecidos de
284
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición
•o <u E
4-»
c 01
t_
3 C O í_ U n j
ü t -
o
2 01
o —i
l ü
E 01
E
a
a>
í
01 0)
c •0)
i (0
(O w o
^ X s ^
co u c
c ' O
u ÍO L.
4-»
c 01 o c o u
•* l O
18
1 »
. 1 ü ) 01
y i 01
u el «1 cu o (_ * r f
ra 3 ( J
CA (D
cu "0
(0 ( J Uí (0
(_ í ü
o - C
ó ;u "5 O )
5! 01
o L.
i c
«1
o —* (0
c 1 3 —* s 10
-o m u
c LU
• O V a
ro tt
c
01 o «4
c •*• «¿ 01
"O 01
C
i A
> as u
c O í
U)
• Z
*»% O)
u
1 —* 01
"O
ao
o • * ÍO
O» •
o NO
r— >o o f\l
«- O «— o
(0 " .
_ o* 03 «£•
•tf N-~
^ ~ O ~
a -* ~o o-
^% &•
CM ^
O -O o
¿ ^
o tn a 03
N. ro
^* m (M « • "
«J Í O Í O
Í O O
O* ca -o
o 2 -r o
*- o -¿r o o
<0 f \ J
m r\i
>-» o. o o
10 r\J r\i ro
^\ ro o
<0
ro ^ T ro
* • * < \ j ro o
*— L n O-u->
^ x
O N
o V -
c '5 a.
a c a.
285
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
este índice (155 en el laricio y 145 en la jara) . A. decorticans posee valores
mucho más bajos (31.34) que las demás especies dados sus niveles de N
significativamente superiores. La existencia una reabsorción de N
inmediatamente antes de la abcisión (ya comentada en el capítulo 3) más
patente en el matorral implica que los valores obtenidos sean más altos que
los esperables si la hojarasca con que se realiza el estudio hubiese sido
recogida en otro momento del año no coincidente con el periodo de máximo
desfronde, o si se hubiesen tomado hojas verdes de la propia planta. Así, los
porcentajes de N en hoja viva con respecto a la concentración de este elmento
en hojarasca en la época de abcisión son del orden del doble en los pinos y el
gaznacho y del triple en la jara.
Se han realizado análisis de correlación entre el índice C/N y el peso
medio remanente observado (en porcentaje de peso seco inicial) al final del
primer año (391) días y al final del segundo año (740-770 días) (figura 5.4).
Las relaciones encontradas son las siguientes:
PR final 2° = 21.98 + 0.27 (C/N) r=0.998;p<0.005
PR final 1° = 41.66 + 0.209 (C/N) r=0.978;p<0.05
Existe una relación positiva y significativa entre ambas variables pero
según puede verse en las figuras 5.4 a y b sólo la que contempla el conjunto
de los dos años puede ser considerada como aceptable dado el mejor ajuste de
los valores observados. Puede decirse pues, que las tasas de descomposición
dismunuyen de una manera significativa en orden inverso a los índices C/N
sobre todo para el conjunto del periodo de estudio.
5.3.1.2 Modelos para describir la dinámica de la descomposición.
Introducción.
La principal aproximación ai análisis de los datos de descomposición
consiste en el ajuste de éstos a determinados modelos matemáticos que
describan la dinámica de la pérdida de peso respecto al tiempo. Una crítica e
286
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
A.
80
60
40
20
n
%PS¡ final periodo
-
-
n
i
4-*
r=0.998, p<0.005
i >
X
50 100
C/N 150 200
PP pn el
Relación peso remanente e índice C/N
ad
B. 00
80
60
40
20
n
%PS¡ 1er año
a
i
+ *
r= 0.978, p< 0.05
1 — i
X
50 100
C/N 150 200
PP pn el a ad
Relación peso remanente 1 año y C/N
Figura 5.4. Relación entre el peso seco remanente e índice C/N en las cuatro
especies estudiadas. 5.4a: final del período de estudio. 5.4b: final del
primer año.
287
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
interesante revisión de los principales modelos empleados puede encontrarse en
Wieder & Lang (1982) y en Ezcurra & Becerra (1987). Según Wieder & Lang
(1982), a la hora de modelizar el proceso de la descomposición es necesario
que las ecuaciones empleadas sean realistas no sólo en términos matemáticos
sino biológicos y concluye después de una intenso análisis de los principales
características de cada tipo de ecuación que los modelos que mejor cumplen
estos requerimientos son los exponenciales. Otros modelos como los lineares,
potenciales o cuadráticos presentan varios problemas en su aplicación por lo
que no son recomendables si lo que se pretende es una interpretación biológica
del proceso de pérdida de peso durante la descomposición (Wieder & Lang 1982).
Los modelos de descomposición basados en ecuaciones exponenciales son
fundamentalmente tres: exponencial simple, exponencial doble y asintótico.
El modelo exponencial simple fue inicialmente propuesto por Jenny et al.
(1949) y discutido con detalle por Olson (1963). El modelo puede expresarse
como:
X = Xo e"kt
t
donde X es el porcentaje de masa inicial remanente a tiempo = t., X es
la masa inicial y k es la tasa de descomposición.
El modelo presupone que la tasa absoluta de descomposición (primera
derivada de la función = -ktX ) disminuye linearmente conforme lo hace la
cantidad de sustrato y que la tasa relativa de descomposición (-k) es una
constante. Biológicamente estos supuestos matemáticos tendrían una
explicación. Conforme transcurre el proceso, los componentes relativamente
lábiles se pierden con rapidez mientras que el material más recalcitrante lo
hace con una menor tasa. Así, al t rancurrir el tiempo, la proporción relativa
de recalcitrante aumenta y la tasa absoluta de descomposición disminuye
mientras que la tasa relativa se mantiene constante (Wieder y Lang 1982).
Este tipo de modelo ha sido empleado de una manera generalizada por
diversos autores fundamentalmente por dos motivos: Primeramente su simplicidad
ya que sólo un parámetro, k, caracteriza la pérdida de masa y en segundo lugar
288
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
la linearización de la función que permite la comparación de los parámetros
entre distintos grupos de datos.
El modelo exponencial doble considera la compartimentalización de la
hojarasca o sustrato en dos tipos de componentes, uno lábil de fácil
descomposición y otro refractario de descomposición más lenta y considera para
cada uno de ellos una tasa de pérdida de peso, kl y k2. El modelo se
expresaría como:
X - Xi.e_lt l t + (1-Xi). e"k2t
t
donde Xt es la proporción de masa inicial que queda a tiempo t, kl y k2 son
las tasas de descomposición de la fracción lábil y refractaria
respectivamente y Xi y (1-Xi) los porcentajes de masa inicial representados
por la fracción lábil y refractaria respectivamente. Con esta ecuación pueden
calcularse pues la cantidad de fracción lábil y refractaria del sustrato
inicial y sus constantes de descomposición.
Este tipo de ecuaciones han sido menos empleadas (Bunell y Tait (1974),
Lousier y Parkinson (1976), O'Conell (1987)b, Ezcurra y Becerra (1987),
Escudero et al. (1987). La no linearización del modelo que impide
comparaciones estadísticas directas entre los parámetros de distintas
ecuaciones de este tipo es la principal causa de su menor uso entre los
autores. '
Por su parte el modelo asintótico es un caso particular de la ecuación
exponencial simple en la que K2 es igual a cero. Esto implica que el material
refractario no se descompone o que lo hace con una tasa que no es
significativamente distinta de cero.Esta suposición es muy cuestionable desde
un punto de vista biológico y por lo tanto la ecuación sólo debería ser
aplicada para algunas condiciones concretas o manipulaciones experimentales
como por ejemplo la eliminación de la fauna descomponedora de compuestos más
recalcitrantes o en cierto tipo de condiciones ambientales extremas (Wieder y
Lang 1982). Este modelo sí puede linearizarse con la ventajas de comparación
de parámetros que esto lleva consigo.
289
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Aplicación del modelo exponencial simple y doble a las especies estudiadas.
PARÁMETROS OBTENIDOS Y HABILIDAD DE LOS AJUSTES.
Se ha aplicado a los datos de pérdida de peso en hojarasca de las cuatro
especies los modelos exponenciales simple y doble. Se ha excluido el modelo
asintótico pues no parece asumible en estas condiciones que la tasa de
descomposición de la fracción refractaria sea nula. En todo caso, si este
modelo fuese el más adecuado, se habría llegado a él directamente al obtener
un valor de cero para Kr en el proceso de cálculo de los parámetros de la
ecuación exponencial doble (Escudero et al. 1987).
El ajuste del modelo exponencial simple se ha realizado de dos formas.
Por un lado linearizando la ecuación calculando la regresión entre el tiempo
transcurrido (años) y el logaritmo neperiano del porcentaje del peso
remanente. Algunos autores como Chauvet (1987) han recomendado el uso de datos
sin transformar en el modelo exponencial simple. Por tanto, el otro
procedimiento consistió en obtener directamente los parámetros del modelo
utilizando un programa de regresión no linear (Hintze 1988). El ajuste del
modelo exponencial doble se efectuó también utilizando este último programa.
Para determinar la bondad del ajuste de los modelos a los datos
observados, hemos seguido el mismo procedimiento que Escudero et al. (1987) y
Ezcurra y Becerra (1987). Este consiste en dividir la parte de variación no
explicada por el modelo (sumatorio de los residuales al cuadrado) entre dos
componentes. Por un lado el error puro (PE) que corresponde a la variabilidad
inherente a los datos dentro de cada grupo (suma de desviaciones de la media
al cuadrado), y por otro el componente denominado error debido a la regresión
(RE). El mejor ajuste sería aquel en el que la ecuación pasara por el valor
medio de los puntos de cada muestreo, de forma que la variación no explicada
por el modelo fuera igual al PE y por tanto RE=0. El ajuste se evalúa
calculando la razón de los cuadrados medios del RE y PE que sigue una
distribución de la F:
cuadrados medios de RE F= ____
cuadrados medios de PE
290
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
por tanto cuanto menor sea F mejor será el ajuste del modelo a los datos.
En la tabla 5.5 se muestran los parámetros obtenidos para cada uno de los
modelos en las cuatro especies, así como el valor de la razón de cuadrados
medios (RMS). Podemos comprobar en dicha tabla como para las cuatro especies
estudiadas el modelo que presenta un mejor ajuste es el exponencial doble que
puede verse representado en las figuras 5.5.a y 5.5. b. Sin embargo el ajuste
no es igualmente bueno en todas las especies, pues mientras que el valor de F
no es significativo en el caso de P. nigra y C. laurifolius, sí que alcanza la
significación en P. pinaster y A. decorticans.
En general, los valores de RMS son superiores a los hallados por Escudero
et al. (1987) en encina y roble. Sin embargo, el periodo de estudio de estos
autores es de solamente un año mientras que en este trabajo el periodo fue de
dos años. Por tanto es posible que el peor ajuste al modelo exponencial doble
se deba a los datos del último año, cuando la descomposición es más variable y
está más afectada por los factores locales como vimos anteriormente. Para
comprobar esta posibilidad, hemos ajustado una ecuación exponencial doble a la
pérdida de peso únicamente durante el primer año en esta dos especies (P.
pinaster y A. decorticans). En el caso de P. pinaster el modelo exponencial
doble se ajusta mucho mejor a los datos del primer año. (WL=9.9677.; KL= 22.77;
KR=0.049, RMS = 0.411), mientras que en el caso de A. decorticans el ajuste
empeora (WL=49.67.; KL= 6.114; KR= 0.085; RMS= 10.19). Esto implica que
efectivamente la heterogeneidad de los muéstreos en el segundo año empeora el
ajuste en el pino. Sin embargo en A. decorticans hay que buscar otra causa.
Analizando los datos y la figura 5.6 se llega a la conclusión de que es el
tercer muestreo (a los 89 días transcurridos) el que proporcionó valores
anormalmente bajos e inferiores a los de los dos muéstreos consecutivos.
DINÁMICA DE LA DESCOMPOSICIÓN SEGÚN EL MODELO DOBLE.
En la tabla 5.6 se muestran los parámetros resultantes de la aplicación
del modelo exponencial doble a las cuatro especies estudiadas. También se
indica la vida media de la hojarasca (Olson 1963) y los T (957.) según Jenny et
al. (1949).
291
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
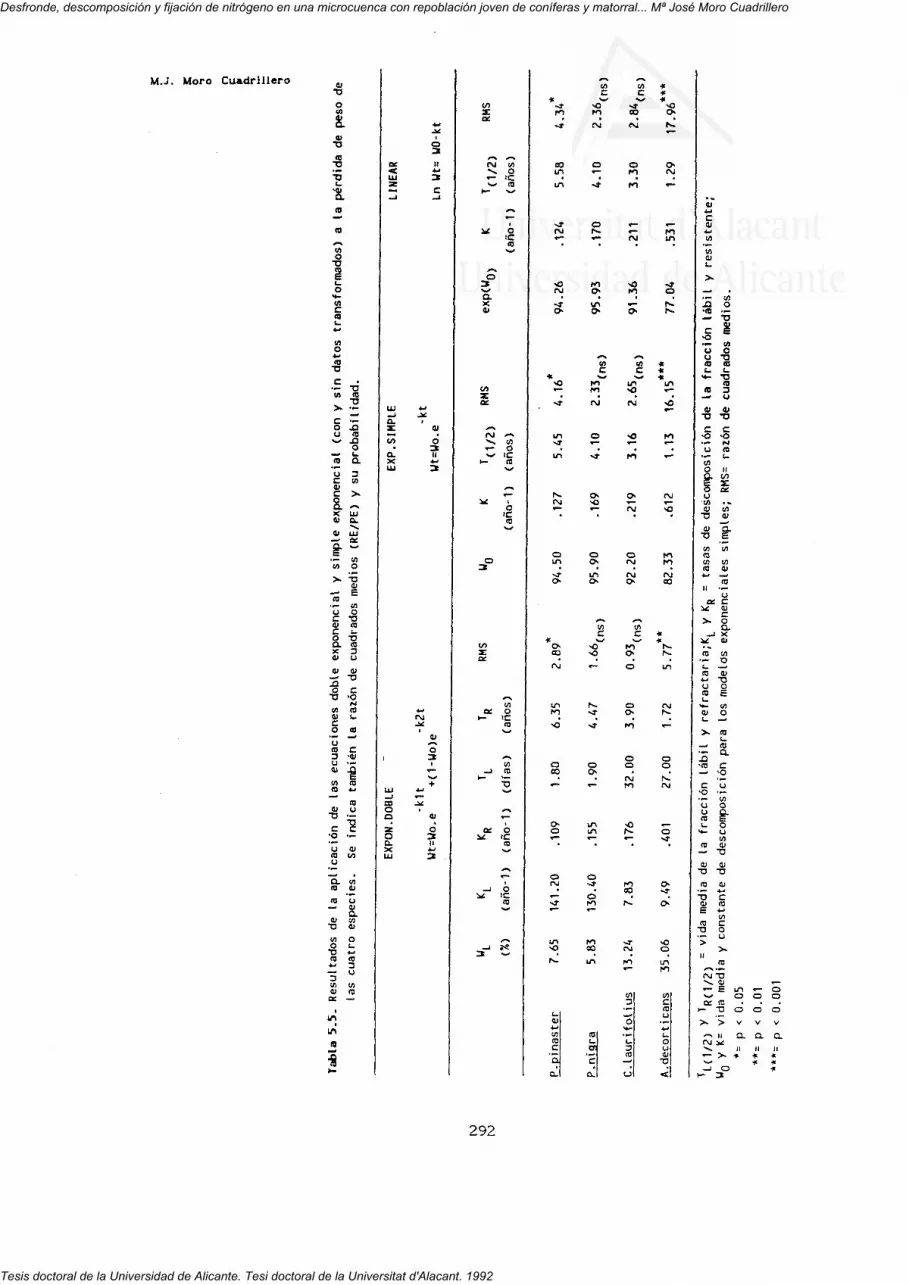
M.J. Moro Cuadrillero
o w 2L
3
e l
* - « - <\l
fr~
O. X ai
m • -
-o c
c U1
o 1C
a -
>-~ - l
m u c 01
c o a X Cl
o .O o -o
"O ai E tr¡ o •o (D l_
•o to 3 u 41
-o c •o
c '41
r\i «—
•a
o-o
<£ O i ¿ IC
O
- s. tu •D V) O "• <U • M
3 W <U o:
m in
a
1
V) Oí
o L.
4 - i
(0 3 <J
WJ (U
-*
o
o o»
«- f i
c Q
292
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
oo;
80
60
40
9Speso seco inicial
* *^SL.
1 1
+ 4: "
i
+
X
i
X X
X
1
+
X
+
+
X
1
X
x~~
+
0 100 200 300 400 500 600 700 800
t¡empo(d¡as)
+ C.laurifolius x A.decorticans
5,5a
100 % peso seco inicia
400 tiempo(cüas)
+ P.pinaster x P.nigra
800
5.5b
Figura 5.5. Ajustes mediante el modelo exponencial doble para las cuatro
especies estudiadas.
293
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
%peso seco inicial
0,8 - \ .
0,6 - ^"~"^--^- -_______^ .
0,4 -
0,2 -
0 I . . • ' 0 0,2 0,4 0,6 0,8 1
años
• valor observado
Figura 5.6. Aplicación del modelo exponencial doble a la descomposición de las
hojas de Adenocarpus decorticans durante el primer año de estudio.
El porcentaje de la fración lábil (W ), también denominada de los
solubles verdaderos (O'Conell 1987) varía entre especies. Los valores más
bajos corresponden a las acículas (7.657. en P. pinaster y 5.83 en P. nigra),
la j a ra posee un 13% de solubles mientras que el valor más alto se encontró en
A. decorticans (35%) lo que, por otra parte es esperable puesto que esta
especie es la que presenta hojas menos consistentes. Los porcentajes de
material más difícilmente descomponible varía en consecuencia en sentido
inverso y este parámetro (Wr) junto a su correspondiente Kr probablemente
influirá de modo determinante la dinámica de descomposición a más largo plazo
en las cuatro especies. Se han realizado análisis de correlación entre el
porcentaje de componente recalcitrante y la masa remanente tanto al final del
periodo como al final del primer año. Las bajas correlaciones encontradas (r=
0.96 en ambos casos) sí bien significativas a nivel de p< 0.05, indican que el
porcentaje de la fracción refractaria por sí misma no puede considerarse en
este estudio como un buen predictor de la pérdida de peso. Sin embargo
utilizando el cociente entre fracción refractaria (7.) y la concentración de N
294
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.6. Porcentajes y tasas de descomposición de las fracciones lábil y resistente;
vida inedia y T (95%) de la hojarasca de las cuatro especies estudiadas.
¡¡picTi í; r k7 T l ( 0 5 ) T r ( Q 5 ) Tl(95%) Tr(95X) (%) (año ) (año ') (días) (anos) (días) (años)
P.pinaster 7.65 141.20 .109 1.8 6.4 7.8 27.5
P.nigra 5.83 130.40 .155 1.9 4.5 8.4 19.4
C . lau r i fo l i us 13.24 7.83 .176 32.0 3.9 140.0 17.0
A. decorticans 35.06 9.49 .401 27.0 1.7 115.0 7.5
T l ( 0 5) y T r (0 5 ) : v ' ^ a m e ^ ' a ^e ^a f racción l áb i l y r e f rac ta r i a , respectivamente; K, y K : tasas de descomposición de la f racción l áb i l y res is tente.
inicial(Z) la correlación mejora sensiblemente sobre todo la que contempla
todo el periodo de muestreo :(r= 0.997 p<0.005) para todo el periodo y r=0.988
(p<0.05) para el primer año {figura 5.7).
Las tasas de descomposición del componente lábil (KL) son
extraordinariamente altas en el pinar, con lo que la pequeña cantidad de
solubles presentes en estas especies se descompone en muy pocos días. El
TU/2) calculado para los dos pinos es de aproximadamente dos días y el T(95%)
de aproximadamente 8 días (tabla 5.6). Los componentes lábiles del gaznacho y
la ja ra poseen también notable KL, (9.5 en A. decorticans y 7.8 en C.
laurifolius) aunque muy inferiores al pinar. Al cabo de aproximadamente 30
días ha desaparecido la mitad de la cantidad inicial de solubles en las dos
especies. Aplicando la ecuación de Jenny et al. (1949), el 957. de este
295
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
100 %PSi 1er año
100 200 % Wr/%N
300
x pp + pn * el a ad
Relación peso remanente 1 año y [ref/N]
400
%PSi final periodo
100
PP
200 % Wr/%N
pn el
300
ad
400
Relación peso remanente final y [ref/N]
Figura 5.7. Relación entre peso seco remanente y cociente componente
refractario/nitrógeno en las cuatro especies estudiadas. 5.7a: final del
primer año. 5.7b: final del período de estudio.
296
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

De«composición.
componente tarda 115 días en desaparecer en gaznacho y 140 en la jara .
Las tasas de descomposición de material más resistente (Kr) presentan una
mayor similitud que las KL en t res de las especies estudiadas (pinos y jara) .
Así, P. pinaster tiene una Kr de 0.109, P. nigra de 0.155 y C. laurifolius de
0.17. Para A. decorticans se ha calculado una tasa entre 2-3 veces superior a
las de las otras t res especies. Esta característica de A. decorticans, junto a
su menor proporción de componente recalcitrante en relación a pinos y ja ra
será determinante de la mayor rapidez en la pérdida de peso de este componente
en A. decorticans y en general de toda su dinámica de descomposición una vez
pasados los primeros estadios dominados por la pérdida de la otra fracción, la
soluble. La figura 5.5 y los valores obtenidos para Tíl/2) (1.72 años) y
T(957.) (7.5 años) así lo manifiestan.
P. pinaster es la especie que representa la dinámica de descomposición
más lenta para este componente. Se han calculado (tabla 5.6) un T (95%) de
27.5 años y un T(l/2) de 6.35. La jara y el pino laricio prácticamente
concluyen su descomposición (T (957.)) a los 17 y 19.3 años respectivamente con
T(l/2) correspondientes de 3.9 y 4.5 años. Parecen, pues, existir más
similitudes a este respecto entre P. nigra y C. Laurifolius que entre los
pinos entre sí. Una de las posibles causas que podría influir en esta
diferente dinámica de descomposición a largo plazo entre los dos pinos
(dejando a parte posibles diferencias estructurales en sus respectivas
hojarascas) se refiere al espacio físico que cada una de ellas ocupa en la
cuenca y por tanto a las posibles diferencias edáficas y microclimáticas que
pueden operar en cada caso. El pino laricio se localiza en la zona alta de la
cuenca caracterizada por ser casi llana, con mayor profundidad de suelo, mayor
grado de humedad y buena parte del invierno cubierta por una capa de nieve que
puede actuar de amortiguador de la bajas temperaturas favoreciendo la
descomposición (Stohlgren 1988).
P. pinaster ocupa zonas de ladera, más degradadas, de mayor pendiente,
con suelo menos profundo y más disgregado y con una menor retención del agua
de lluvia. Es probable que las condiciones microclimáticas de la parte alta
favorezcan la descomposición de las acículas en el laricio mientras que las
que actúan en las laderas influyan negativamente en la pérdida de peso del P.
297
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
pinaster. No obstante, sería necesario un estudio de campo con acículas de
ambas especies colocadas en las dos localizaciones para poder sacar
conclusiones a este respecto.
Según el modelo exponencial doble se ha calculado la masa remanente (en
porcentaje de peso seco inicial) al final del primer y segundo año para cada
especie (tabla 5.7).
Al final del primer año, la pérdida de peso calculada para A. decorticans
fue del 56.5% de su masa inicial, valor muy superior a las otras especies. La
j a ra perdió un 27.3 % y el pinar lo hizo con porcentajes similares de su masa
incial (17.2% en P. pinaster y 19.3% en P. nigra). Al final del segundo año,
el porcentaje de peso perdido disminuye bastante en todos los casos. El
descenso más acusado se produjo para A. decorticans que pierde un 75% menos
que el primer año, la j a ra un 56 % menos, P. pinaster un 50 % mientras que el
peso perdido en P. nigra fue de sólo un 40 % menor. Por tanto, a part i r del
primer año, P. nigra es la especie que pierde proporcionalmente más peso
respecto al primer año. A los 770 días los valores de masa remanente de los
dos pinos se alejan ligeramente en relación a los encontrados al final de los
365 días. Parece, pues, que las diferencias en la descomposición de ambas
especies de pino aparecen o se hacen más marcadas a partir del primer año.
(ver también figura 5.5).
La asumción implícita del modelo doble exponencial consiste en que dos
de los parámetros de esta ecuación (WL y Wr) representan el porcentaje inicial
del componente lábil y refractario respectivamente (Lousier y Parkinson 1976).
Sin embargo, algunos autores han señalado que el modelo no tiene en cuenta los
trasvases de un compartimento a otro a lo largo del proceso, por ejemplo,
durante la síntesis de biomasa microbiana (Wieder y Lang 1982). Ezcurra y
Becerra (1987) comentan que la separación de la hojarasca en sólo dos
compartimentos que no estén perfectamente definidos o medidos desde un punto
de vista químico puede no ser una asumción realista.
Pero además existen por lo menos otros tres características que pueden
hacer variar de una manera a veces patente los porcentajes de lábil y
refractario calculados a part ir de las exponenciales dobles (y también
298
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.7. Masa remanente ( en X del peso seco inicial)
al final del primer y segundo año calculadas
a partir del modelo exponencial doble.
ESPECIE % PSI % PSI
(1er año) (2° año)
P.pinaster 82.77 74.18
P.nigra 80.64 69.06
C.laurifolius 72.73 60.95
A. decorticans 43.50 29.13
asintóticos).
1- Estado inicial de la hojarasca en la muestras.
Las características de la fracción lábil hacen que este compartimento sea
muy suceptible al lixiviado (Swift et al. 1979) y por tanto su cantidad
inicial y su dinámica posterior pueden variar dependiendo del estado inicial
de la hojarasca y de las codiciones climáticas (sobre todo existencia de
lluvias) a que se vea sometido en las primeras fases de exposición. Así, los
porcentajes de fracción lábil obtenidas en este estudio (tabla 5.5) se
refieren a una hojarasca inicial que llevaba en el suelo un máximo de 30 días
en el transcurso de los cuales pudieron haber ocurrido procesos de lixiviado
sobre todo si se registraron lluvias. Concretamente, se utilizó hojarasca de
299
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
matorral caída entre el 18 Julio y 19 Agosto de 1988 en que se procedió a su
recogida para iniciar el ensayo. En este periodo no se registró ninguna
precipitación y la época del año tampoco es la más favorable para el lixiviado
(stress hídrico estival). Por tanto no es esperable que la hojarasca recién
caída tenga porcentajes de componente lábil muy distintos al W calculado a
partir de la ecuación. No hemos encontrado referencias bibliográficas que
contemplen la doble compartimentalización de hojarasca arbustiva.
Comparando con los valores de W que aparecen en la tabla 5.8 y que se
refieren en todos los casos a especies arbóreas las mayores similitudes
se encuentran con la j a ra y la hojarasca de Quercus rotundí folia
colocada en primavera (Escudero et al. 1987).
En el caso del pinar, nuestro estudio tuvo como sustrato acículas caídas
entre 19 Agosto y 22 Septiembre 1988. En este periodo sí se registro una
lluvia otoñal el 14 de Septiembre con un volumen de 23 mm que pudo provocar
lixiviado en las acículas recién caídas. Por tanto, los valores de W para las
acículas de pino al comienzo del ensayo probablemente sean más pequeños que
los que existen en realidad en la pinada recién caída. Como puede verse en la
figura 5.5, la fase inicial de la curva correspondiente a la una rápida
disminución de peso o fase de lixiviado (Gosz et al. 1973), no presenta en
ninguno de los dos pinos una rama vertical demasiado acusada como la que
correspondería si el contenido en elementos solubles iniciales fuera más alta.
Una falta casi completa de rama vertical es lo que encuentran Klemmedson et
al. (1985) utilizando como sustrato hojarasca de Pinus ponderosa de una edad
máxima de tres meses para sus estudios de descomposición.
Deberíamos añadir que no tiene demasiado sentido hacer comparaciones
sobre los valores de W entre las especies arbustivas estudiadas con respecto
a las de pino ya que las condiciones climáticas previas a las que fueron
sometidos los correspondientes sustratos no fueron exactamente las mismas. Sin
embargo si cabrían las comparaciones entre las arbustivas y pinar entre sí.
Esta calidad inicial del recurso ha de ser un factor muy importante a
tener en cuenta tanto a la hora de extrapolar los datos de nuestro estudio
como a la hora de compararlos con datos bibliográficos ya que no existe un
criterio unificado respecto a la edad de la hojarasca que se toma como
300
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
sustrato base para colocar en las bolsas. Hay autores que recogen las hojas
frescas del propio árbol (Enright y Ogden 1987), y, aunque en la mayoría de
trabajos se colocan trampas de recogida, el tiempo transcurrido entre
colocación y recogida también es variable.
2. Posición en el espacio de las muestras y momento del año en que se
colocan.
En un estudio sobre la dinámica de descomposición en Quercus rotundi folia
Escudero et al. (1987), señalan que los parámetros del modelo exponencial
doble ajustados a sus datos llegaban a ser considerablemente distintos según
la posición en el espacio en que se colocaran las muestras, (bajo árbol, borde
árbol o en suelo sin cubierta). En los t res casos se utilizaban replicados de
hojas por tanto no puede hablarse de distinto estado inicial de éstas. En el
mismo estudio se colocaban tandas de hojas en distintos momentos del año y se
estudiaba su evolución. Se comprobaba, por ejemplo, que las diferencias
químicas de las hojas de primavera y verano no eran suficientes para explicar
los diferentes patrones de pérdida de peso y por tanto de los parámetros que
definían los modelos de descomposición. Todo ello lleva a los autores a
concluir que los factores ambientales llegan a enmascarar o se solapan con los
debidos a la calidad del sustrato y que debido a ello los parámetros de las
ecuaciones pueden ser muy distintos en unas condiciones que en otras. Además
concluyen que nos se puede asignar de una manera estricta un concreto
componente químico (por ejemplo lábil ó refractario) a un determinado
parámetro matemático.
Como conclusión, los valores de W , Wr han de tomarse sólo como
orientativos de la doble compartimentalización de la hojarasca y sólo
aplicables estrictamente para unas condiciones dadas. En ningún momento deben
de utilizarse como unos parámetros que caracterizen la hojarasca de una
especie dada. Las extrapolaciones o comparaciones que se deriven de la
utilización de estos parámetros han de tomarse con extrema precaución.
Como se ha comentado antes son bastante pocos los datos que en
bibliografía pueden encontrase sobre la dinámica de descomposición de
hojarasca ajustada a ecuaciones exponenciales que supongan la doble naturaleza
301
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
del sustrato (exponenciales dobles o asintóticas). Las dobles son utilizada
por O'Conell, 1987b) en Eucalyptus diversicolor en Australia, Escudero et a,
(1987) en distintas especies de Quercus en España, Ezcurra y Becerra (1987) e:
Liquidambar en México y Sharma & Ambasht (1987) en Alnus nepalensis de 1¡
India. Las asintóticas han sido utilizadas por Chauvet (1987), Gourbiére i
Corman (1987) en Abies alba en Francia.
En la tabla 5.8 pueden contemplarse estos resultados recogidos de la
bibliografía sobre aplicación de modelos exponenciales dobles y asintóticos a
hojarasca de alguna especies arbórea. Existe una gran heterogeneidad en los
valores de los parámetros. Los factores comentados en las líneas anteriores
pueden tener aún más peso que las posibles diferencia específicas. Así mismo,
el tipo de ecuación utilizada o el tiempo de muestreo pueden también modificar
el cálculo de los parámetros.
Los valores de W para los pinos de la cuenca "Nacimiento" se encuentran
muy por debajo de cualquiera de los valores correspondientes presentes en la
tabla 5.8. Por lo menos parte de estas diferencias podrían ser explicadas por
el posible lixiviado de acículas al que ya nos referimos anterioremente. Los
porcentajes de lábil en j a ra y gaznacho se aproximan más a los de la
bibliografía. Si sólo se toman las referencias á ecuaciones exponenciales A.
decorticans tendría el más elevado porcentaje de lábil de las especies citadas
en la tabla 5.8 pero, dada su estructura foliar mucho menos esclerófila a las
especies de la bibliografía, {Eucalyptus, Quercus) deberían esperarse aun
valores más elevados.
DINÁMICA DE LA DESCOMPOSICIÓN SEGÚN EL MODELO EXPONENCIAL SIMPLE
1. Tasas de descomposición (k).
El modelo exponencial simple linearizado permite la comparación directa
de las pendientes (constantes relativas de descomposición (k)) entre distintas
especies y períodos de muestreo (primer, segundo año y período total). Los
parámetros de las ecuaciones aplicadas para cada especie y período pueden
observarse en las tablas 5.9 y la representación de los ajustes en la figura
5.8. Hemos aplicado un tes t - t para comparar las tasas relativas de
descomposición entre especies y períodos (primer año, segundo año y período
302
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.8. Parámetros obtenidos por diversos autores en la aplicación de varios modelos exponenciales
y asintóticos a la dinámica de pérdida de peso durante la descomposición.
ESPECIE
Duración
(días) AUTOR
Quercus rotundifolia
Primavera bajo
borde
Otoño
bajo
borde
Invierno
bajo
borde
13.7
30.0
26.4
80.9
51.5
43.7
86.2
64.4
73.6
19.0
48.5
56.3
-1 -0.57
-0.58
-0.13
-0.16
-0.28
-0.080
-0.068
-0.037
0.000
0.000
0.000
365 365
365 365
365 365
Escudero et al (1987)
Liquidambar styraciflua
-0.20 -0.006 90 Ezcurra y Becerra (1987)
Eucalyptus diversicolor
40 años
6 años
26.1 73.9 -2.66 -0.190 574
23.9 76.1 -7.59 -0.310 574
O'Conell (1987)
Abies alba
con mal la
sin mal la
62.7 37.3 -1.058 0.000 1062
55.7 44.2 -.949 0.000 1062
Gourbiére et Corman (1987)
total). Los resultados se expresan en la tabla 5.10.
Para el período total de muestreo, la hojarasca de cada especie se
descompone con una tasa, k, que es significativamente distinta a las de las
demás especies. El orden de las especies según la velocidad de descomposición
es de :
A. decorticans > C. laurifolius > P. nigra > P. pinaster
303
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 5.9. Parámetros calculados aplicando el modelo exponencial simple a cada una
de las cuatro especies estudiadas. 5.9a: primer año, 5.9b: segundo año, 5.9c: todo el
período. Se indican el coeficiente de regresión (r ), número de datos (n),
probabilidad (p) y tiempo de semidescomposición en años (TQ ¿).
X = masa remanente (%) a tiempo = 0; K.= constante de descomposición.
5.9a(1eraño):E1
P.pinaster
P.niqra
C. l a u r i f o t i i K
A.decorticans
X0
94.33
95.90
95.00
84.12
-K1
-.129
-.169
-.313
-.824
n
12
12
12
12
r2
.66
.80
.85
.75
P<
.001
.001
.001
.001
T<0.5)
5.37
4.10
2.21
.84
5.9b(20año):E2 -K1 P< '(0.5)
P.pinaster
P.niqra
C. laurifol in<!
A.decorticans
86.74
95.72
85.83
87.50
-.074
-.168
-.172
-.602
12
12
14
12
.30
.72
.65
.71
ns
.001
.001
.001
4.12
4.03
1.15
5.9c(Todo el periodo):Er -K1 (0.5)
P.pinastpr
P.niqra
C. l au r i fo l ¡LIC
A.decort i ran*
94.25
95.87
91.35
77.05
-.127
-.169
-.211
-.535
24
24
26
24
.83
.92
.90
.85
.001
.001
.001
.001
5.40
4.10
3.28
1.30
304
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
%peso seco inicia 100f
200 400 600 800
t¡empo(d¡as)
+ C.laurifolius x A.decort icans
1000
5.8a
% peso seco inicial lOOf-
90
80 -
70
60 200 400
t¡empo(dias)
+ P.pinaster x P.nigra
600, 800
5.8b
Figura 5.8. Ajustes para las cuatro especies según el modelo exponencial
simple.
305
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5-10. Valores de K para las cuatro especies a dis t intos tiempos y resultados de la aplicación del t e s t - t . Para la misma columna, los valores seguidos del mismo número no son significativamente d i s t in tos ; para la misma f i l a , los valores seguidos de una misma letra no son significativamente d i s t in tos . ()= número de datos. ns: no significativamente diferente de 0.
P.pinaster P.nigra C . lau r i fo l i us A.decorticans
K1
h
<t
1
.129 a
(12)
ns
.074 ns
(12)
1
.127 a
(24)
1
.169 a
(12)
1
.168 a
(12)
1
.169
b (24)
1
.313 d
(12)
2
.172 a
(12)
2
.211 d
(24)
1
.824 c
(12)
1,2
.602 b
(12)
2
.535 c
(26)
Durante el primer año, los resultados del tes t - t indican que no existen
diferencias significativas entre las k de los dos pinos, (t=0.994, ns). Por
tanto en esta primera fase de la descomposición, la hojarasca de P. pinaster y
P. nigra presentan comportamientos similares. La diferencia significativa de
sus correspondientes tasas para el conjunto del período (t= 14.46, p< 0.05)
induce a pensar que esta diferencia viene sólo determinada por la dinámica que
siguen las aciculas de ambas especies durante el segundo año.
Para el resto de parejas de k entre especies para el primer año, los
resultados del tes t - t indican idénticos resultados a los ya comentados para
todo el período exceptuando que el nivel de significación es algo menor (lo
que puede deberse en buena parte al menor número de datos manejados). La
relación más significativa (t=4.56, p<0.001) la presentan P. pinaster y A.
306
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
decorticans para las que se han calculado las k más baja y más alta
respectivamente (-0.82 y -0.12). Por su parte C. laurifolius con A.
decorticans, P. nigra y P. pinaster y A. decorticans con P. nigra presentan
diferencias significativas con p< 0.01.
Durante el segundo año, la relación masa remanente frente al tiempo en P.
pinaster no resultó significativa (r = 0.30 p= 0.07); por tanto en este
periodo P. pinaster no puede compararse estadísticamente con las demás
especies. A pesar de todo, esta ecuación no significativa calculaba una k de
-0.074. que probablemente hubiese sido significativamente más baja que la
encontrada para el mismo año en P. nigra (k=-.168) lo que confirmaría que
estas dos especies se alejan en cuanto a su dinámica de descomposición sólo a
partir del segundo año.
La hojarasca de A. decorticans sigue siendo la más fácilmente
descomponible durante el segundo año (K=-0.60) y significativamente más alta
que las demás especies (tabla 5.10). Por su parte, no se encontraron
diferencias significativas entre la pérdida de peso de P. nigra y C.
Laurifolius para este periodo (t= -0.06, ns). En el apartado anterior ya se
había comentado que existían similitudes entre las dinámicas de estas dos
especies, más incluso que entre los dos pinos entre sí en algunos aspectos y
esto parece constatarse por lo menos para el segundo año de muestreo.
Para cada especie en particular, la variación de la constante de
decomposición en distintos periodos ha sido también comprobada con el
tes t - t (tabla 5.10). A este respecto sólo se han encontrado diferencias en las
especies arbustivas y no entre las dos especies de pino. No existen
diferencias significativas en las velocidades relativas de decomposición para
A. decorticans entre el primer y segundo año (t= -1.16, ns) aunque la tasa
calculada para el primer año aparentemente es bastante más elevada (k=-0.82)
que el segundo (K=-0.60). Quizá el menor número de datos y los errores
estándar algo elevados impiden que se alcance la significación. Sin embargo sí
existe diferencia entre primer año y periodo total (t= -2.6 p< 0.05) siendo la
k calculada para el conjunto de los 770 días de -0.53 que, a su vez, no
resulta distinta (t= - 0.619, ns) a la k del segundo año (k =-0.60).
307
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
En C. Laurifolius, existen diferencias significativas entre las
velocidades de descomposición del primer año con respecto al segundo y al
periodo total, siendo más altas las del primer año (k = 0.313). En el primer
caso, (t= -2.60, p< 0.05), en el segundo (t =-2.93 p<0.01). No existen, sin
embargo diferencias entre las tasas calculadas para el segundo año y el
periodo total (t= 1.20, ns).
2. - Vida media de la hojarasca y pérdida de peso.
Dado que no existen diferencias significativas entre las k a distintos
tiempos para el pinar, los tiempos de semidescomposición (T ) calculados
aplicando cualquiera de las t res tasas ( 1 , 2 y total) son prácticamente
idénticos (tabla.5.9 a,b y c). Así, la hojarasca de P. pinaster tiene una vida
media de unos 5.4 años y P. nigra de 4.1 años. Para la j a ra los T llegan a
ser bastante distintos si aplicamos Kl que si lo hacemos con K2 o con Kt ya
que existen diferencias significativas entre K1-K2 y Kl-Kt. En este caso,
parece pues más realista utilizar el dato de vida media de hojarasca calculado
a part ir de constante del segundo año. Además, al no existir diferencias
significativas entre K2 y Kt, los tiempos de semidescomposición pueden
calcularse indistintamente con una u otra tasa. Igualmente parece más correcto
calcular el T a partir de las K2 o Kt en A. decorticans que con Kl. Para
estas tasas, los valores de semidescomposición en C. Laurifolius oscilarían
entre 3.3 y 4 años y en A. decorticans entre 1.15 y 1.3 años (tabla 5.9 b y
c).
El peso remanente al final del primer y segundo año de estudio (en
porcentaje del peso seco inicial acumulativo) se muestra en la tabla 5.11. Las
parejas de valores que aparecen en la tabla corresponden a los porcentajes
calculados a part ir de la ecuación exponencial linearizada para cada año (El ó
E2) y a par t i r de la ecuación para todo el periodo (Et). También se añadido el
nivel de significación del tes t - t a las k de esas parejas de ecuaciones.
Al final del primer año, los porcentajes de masa remanente de hojarasca
de pino alcanzan el 83% en P. pinaster y 817. en P. nigra. Lógicamente los
valores son muy similares dada la semejanza en la dinámica de descomposición
de estas especies en el primer año que ya hemos comentado. Por tanto durante
308
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.11. Peso seco remanente (% del peso seco inicial) tras el primer y
segundo año calculados según el modelo exponencial simple.
E1 Et p E2 Et
P.pinaster
P.nigra
C.laurifolius
A.decorticans
83.40
81.01
69.48
36.87
83.26
81.04
74.12
45.12
ns - 73.50 ns -
68.33
60.88
22.26
73.50
65.58
60.30
26.41
E1, E2 y Et corresponden a las ecuaciones exponenciales simples para el primer,
segundo año y periodo total respectivamente.
ios primeros 365 días los pinos pierden alrededor del 20% de su masa inicial. 3or las mismas razones comentadas en el cálculos de los tiempos de
semidescomposición, la masa perdida por los arbustos variará si empleamos El o
ít ya que tanto la jara como el gaznacho tienen tasas significativamente más
•ápidas el primer año. Consideramos como valores más próximos a la realidad
os datos obtenidos a partir de la ecuación de ese primer año ya que los
atenidos por la Et están también influidos por la valores de pérdida de peso
el segundo año. Así, la jara ha perdido al cabo del primer año un 30.57. de su
eso inicial. En el gaznacho, la pérdida de peso duplica a la de C.
aurifolias y triplica a la de los pinos ya que solamente algo menos de un 407.
el peso seco inicial remanece en esta especie.
Para el segundo año, los valores de K2 y Kt no difieren
309
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
significativamente en ninguna de las especies por lo que los porcentajes de
peso seco inicial remanente son prácticamente idénticos aplicando una u otra
ecuación (tabla 5.11). La hojarasca que más masa conserva a los 770 días es p.
pinaster (73.57.) seguido de P. nigra (68.5%) y C. laurifolius (60.37.) mientras
que se ha perdido ya un 75 % del peso inicial de A. decorticans. Quizá lo más
destacable de este segundo año es la tendencia a la estabilización de las
dinámicas de descomposición. Durante ese año todas las especies pierden
alrededor del un 10% de peso. P. nigra es la especie con un mayor pérdida
relativa (12.5%) y C. laurifolius la menor (97.). Por tanto, la cantidad de
masa perdida disminuye notablemente en relación al primer año en todas las
especies, (ver también apartado referente a la aplicación de ecuaciones
exponenciales dobles). La variación más brusca se produjo en A. decorticans
que pierde el segundo año un 82% menos de su peso que el primero; la menor en
P. nigra que sólo perderá el 37% menos.
Según estos resultados, parecen existir dos tendencias más o menos
marcadas en las especies estudiadas; una, la que siguen las especies
arbustivas con disminuciones en su masa muy acusadas el primer año, sobre todo
en el caso de A. decorticans seguidas de un periodo de estabilización (en el
segundo año y probablemente siguientes) en que las significativamente menores
tasas relativas de descomposición determinan que la masa perdida descienda a
valores de entre le 70-80% menores a las del primer año. El pinar por el
contrario, parece mantener una mayor continuidad, sin cambios demasiado
bruscos, en lo que se refiere a su dinámica de descomposición a lo largo de
los dos años. De hecho en P. nigra las Kl y K2 no son distintas. Si se elimina
la fase inicial de la curva de descomposición correspondientes a la caída
rápida de peso (dos primeras recogidas) (tabla.5.12) puede observarse que las
k' son prácticamente idénticas a las correspondientes k en el pinar. El test t
confirma estos resultados ya que las diferencias entre k y k' en P. pinaster y
P. nigra no son significativas. En el matorral, las constantes de
descomposición sin tener en cuenta el lixiviado inicial resultan inferiores a
las tasas (k) que sí lo incluyen. Sin embargo, aplicando el test t tampoco se
han detectado diferencias significativas entre k y k'. Esto parece indicar que
en las especies arbustivas la fase de lixiviado inicial no resulta tan
determinante para la dinámica de descomposición a más largo plazo.
310
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
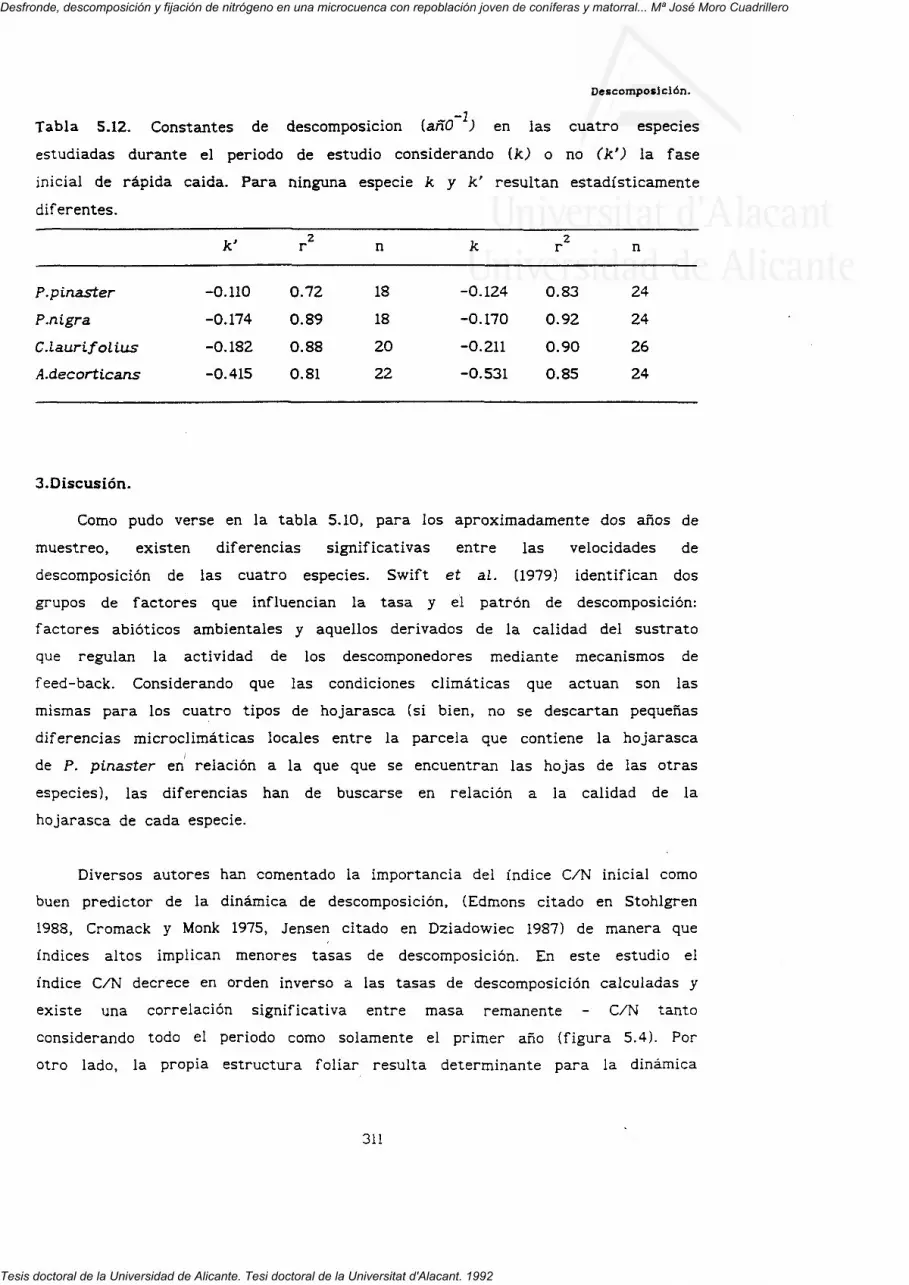
Descomposición.
Tabla 5.12. Constantes de descomposición (añO ) en las cuatro especies
estudiadas durante el periodo de estudio considerando (k) o no (k') la fase
inicial de rápida caida. Para ninguna especie k y k' resultan estadísticamente
diferentes.
P.pinaster
P.nigra
C.laurifolius
A.decorticans
k'
-0.110
-0.174
-0.182
-0.415
2 r
0.72
0.89
0.88
0.81
n
18
18
20
22
k
-0.124
-0.170
-0.211
-0.531
2 r
0.83
0.92
0.90
0.85
n
24
24
26
24
3.Discusión.
Como pudo verse en la tabla 5.10, para los aproximadamente dos años de
muestreo, existen diferencias significativas entre las velocidades de
descomposición de las cuatro especies. Swift et al. (1979) identifican dos
grupos de factores que influencian la tasa y el patrón de descomposición:
factores abióticos ambientales y aquellos derivados de la calidad del sustrato
que regulan la actividad de los descomponedores mediante mecanismos de
feed-back. Considerando que las condiciones climáticas que actúan son las
mismas para los cuatro tipos de hojarasca (si bien, no se descartan pequeñas
diferencias microclimáticas locales entre la parcela que contiene la hojarasca
de P. pinaster en relación a la que que se encuentran las hojas de las otras
especies), las diferencias han de buscarse en relación a la calidad de la
hojarasca de cada especie.
Diversos autores han comentado la importancia del índice C/N inicial como
buen predictor de la dinámica de descomposición, (Edmons citado en Stohlgren
1988, Cromack y Monk 1975, Jensen citado en Dziadowiec 1987) de manera que
índices altos implican menores tasas de descomposición. En este estudio el
índice C/N decrece en orden inverso a las tasas de descomposición calculadas y
existe una correlación significativa entre masa remanente - C/N tanto
considerando todo el periodo como solamente el primer año (figura 5.4). Por
otro lado, la propia estructura foliar resulta determinante para la dinámica
311
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
de la descomposición (Carlyle 1986, Enright y Ogden 1987, Swift et al. 1979).
En las especies bajo estudio, los pinos son claros representantes de hojas 2 2
esclerófilas con densidades foliares de 42 mg/cm y 46mg/cm respectivamente,
la j a ra posee hojas malacófilas con densidades foliares inferiores (15.6 2 2
mg/cm ) mientras que A. decorticans con densidad foliar de 14.4 mg/cm puede
considerarse como moderadamente mesófila.
Las hojas esclerófilas son más ricas en compuestos refractarios pues
contienen más proporción de tejidos esclerenquimatosos, ligninas y celulosas
insoluoles mientras que al pasar a hojas con esclerofilia más moderadas o
mesofilia éstas contienen progresivamente una mayor proporción relativa de
componentes solubles y lixiviables (Read y Mitchell 1983). El índice de
esclerofilia, bien calculado como densidad foliar o como la relación entre
fibra/proteína (Loveless en Cromack y Monk 1975 y Rundel 1988), puede
considerarse como un buen indicador de la proporción reclacitrante/lábil en la
hojarasca. Los valores de densidad foliar en las especies estudiadas
concuerdan con los valores calculados mediante el modelo exponencial doble en
relación a la prporción de compuestos lábiles y recalcitrantes: los pinos
poseen mayores coeficientes de componente refractario (tabla 5.6) y mayores
densidades foliares, estos porcentajes disminuyen en la j a ra (86%) y más aún
en el A. decorticans (657.). El orden de las cuatro especies según la cantidad
de material difícilmente descomponible y según la densidad foliar coincide y
es: P. nigra > P. pinaster > C. laurifolius > A. decorticans.
Muchos autores han constatado el buen papel predictor del contenido de
ADF (acid detergent fiber) y sobre todo del contenido de lignina inicial en la
dinámica de descomposición sobre todo para las últimas etapas (Berg 1986,
Stohlgren 1988, Cromack y Monk 1975). Aunque en este trabajo no se realizaron
análisis para determinar el contenido en celulosas, ligninas o adf en la
hojarasca, dados los resultados para el componente recalcitrante obtenido a
partir del modelo doble y su relación con la densidad foliar, podría
justificarse su utilización como posible predictor de la dinámica de
descomposición. Sin embargo la baja correlación encontrada entre WR y masa
remanente (r= 0.96) indica que el porcentaje de componente refractario por sí
solo no es un buen predictor.
312
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Se han encontrado relaciones inversas entre grado de esclerofilia y
velocidad de descomposición en varias especies componentes del bosque de kauri
en Nueva Zelanda (Enright y Ogden 1987). Las hojas de las especies estudiadas
en este bosque son clasificadas en cuatro categorías, esclerófilas duras y
suaves y mesófilas fuertes y suaves. El rango de pérdida de peso al final del
primer año de descomposición para los esclerófilos duros varía entre el 15.3 y
29.37., el de los esclerófilos blandos entre le 35 y 41 % y el de los mesófilos
duros entre el 50 y 61%. Los pinos de nuestro estudio, se englobarían en el
primer grupo con valores cercanos al límite inferior del rango dado por estos
autores. La jara, con pérdidas del 32%, se encontraría a caballo entre los
esclerófilos duros y blandos. A. decorticans con pérdidas observadas de un 51%
a los 391 días se englobaría en la categoría de mesófilos duros. Sin embargo,
no parece que las diferencias de esclerofilia entre las dos especies de
matorral estudiadas sean capaces de explicar las diferencias tan notables que
existen respecto a las tasas de descomposición sobre todo en el primer año
(0.8 para A. decorticans y 0.3 para C. laurifolius). La cantidad de componente
refractario es solo un 32% superior en A. decorticans respecto a C.
laurifolius y la densidad foliar solo un 7%. La explicación a las diferencias
de ambas tasas ha de buscarse más bien en diferencias entre los niveles de
nutrientes iniciales. En este sentido los valores C/N pueden ser más
orientativos. El C/N de C. laurifolius es muy superior al de las hojas de A.
decorticans (145 frente a 31.4). El hecho de que el gaznacho sea una planta
fijadora de nitrógeno permite que sus hojas posean elevados contenidos de este
elemento. A pesar que durante la época de máximo desfronde estival se produce
un considerable porcentaje de retraslocación de N en A. decorticans las
concentraciones foliares en hojarasca poseen valores medios del 1.5 % frente
al 0.34 % de la jara. En consecuencia el C/N es mucho más bajo en la
leguminosa lo que puede ser decisivo para su dinámica de descomposición.
Por tanto al considerar la calidad de la hojarasca como parámetro
relacionado con la dinámica de descomposición han de contemplarse dos
características: una la heterogeneidad química de la hojarasca referida a una
mayor o menor riqueza en componentes difíciles de descomponer y otra el nivel
de nutrientes iniciales en ella. En general, ha sido establecido por diversos
autores (Maheswaran y Attiwill 1987, Berg y Lundmark 1987, Berg 1986) que
estos dos factores actúan de forma interdependiente pero con una importancia
313
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
diferencial según la etapa del proceso que se considere. Así, mientras que los
nutrientes, sobre todo aquellos que se relacionan con el metabolismo
microbiano (N, P, S) tienen un mayor peso específico en las fases iniciales
del proceso, (Berg 1986) el contenido en lignina, máximo representante de
compuestos recalcitrantes determina la dinámica de descomposición a más largo
plazo (Stohlgren 1988, Maheswaran y Attiwill 1987, Berg 1986).
El índice [lignína]/[N] inicial de la hojarasca combina ambas
características foliares comentadas arriba y ha sido empleado por algunos
autores que han constatado que puede ser el mejor índice predictor de las
tasas de descomposición en algunos casos (Stohlgren 1988, Aber y Melillo y
Melillo et al. en Stohlgren 1988). En este trabajo se ha utilizado la razón
entre el componente refractario y la concentración de N inicial (7.Wr/7SN) ya
que no se disponían de datos de lignina. Al aplicarlo frente a la masa
remanente, se encuentran relaciones significativas incluso mejores que las
encontradas con el C/N especialmente en el primer año (figura 5.7). Esto
parece confirmar la influencia de estas dos características foliares: nivel de
N en las hojas y contenido en componentes refractarios en la dinámica general
de la descomposición. También Cromack y Monk (1975) encuentran relaciones
significativas entre la pérdida de peso tanto respecto al C/N como al índice
de esclerofilia.
Los estudios de descomposición suelen ser bastante heterogéneos en lo que
refiere a número de años de seguimiento. En la mayoría de los trabajos en los
que se calculan parámetros como K, To.s etc para periodos superiores al año se
suele hacer referencia a ios valores que toman éstos durante el primer año
para poder comparar con otros autores. Los valores calculados de Kl a partir
de exponenciales simples para los pinos reflejan velocidades de descomposición
inferiores a los rangos encontrados en la bibliografía para coniferas de zonas
templadas. Stohlgren (1988b) encuentra valores de k en coniferas que oscilan
entre 0.18 y 0.62 año-1. A part ir de los T citados por Verdú (1984) también
para coniferas templadas, hemos calculado con la formula de Olson (1963) las
tasas de descomposición, oscilando éstas entre 0.19 y 0.72 año"2. Dejando a un
lado las diferencias climáticas y latitudinales obvias, también es cierto que
las concentraciones iniciales en la mayoría de los elementos son superiores
para los pinos de zonas templadas (Klemmedson et al. 1985, Stohlgren 1988b).
314
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Estas diferencias de concentraciones pueden agudizarse para algunos elementos
importantes (N.P) mediante mecanismos de retraslocación que se encuentran bien
desarrollados en las vegetales que viven en zonas mediterráneas (Read y
Mítchell 1983). La existencia de estos procesos para P. pinaster y P. nigra
fueron ya comentados en el capítulo 3 y su consecuencia es un aumento de los
índices C/N y C/P en la hojarasca recién caída. También en zonas mediterráneas
el máximo desfronde se produce inmediatamente antes o durante el periodo más
seco (estival), (Núñez 1989, Verdú 1984, Escudero eí al. 1985) con lo que los
componentes solubles y lixiviables de la hojarasca se ven menos sometidos al
lavado que si ésta cayera en otras épocas del año lo que también puede afectar
considerablemente a las tasa de pérdida de peso.
Respecto a coniferas de zonas mediterráneas, nuestros valores también
resultan ligeramente inferiores a los encontrados en la bibliografía. Verdú
(1984) encuentra en la Castanya una tasa de descomposición de 0.24 par Abies
alba. Santa Regina y Gallardo (1989) calculan una K de 0.2 en Pinus sylvestris
en la Sierra de Béjar (Salamanca). Garrido eí al. (1989) encuentran pérdidas
de peso para el primer año de descomposición del 407. en Pinus pirinea y de 30 7.
en P. pinaster también en Salamanca mientras que P. pinaster y P. nigra de
nuestro trabajo lo hacen con porcentajes algo más bajos (alrededor del 207. del
peso inicial). El buscar una explicación a todas estas diferencias
posiblemente carezca de sentido pues son numerosos los factores que pueden
influir y reducido el número de datos para comparar. Entre estos factores
podrían señalarse aparte del específico otros como diferencias en parámetros
ambientales entre zonas de estudio, microclimas locales, tipo de suelo,
diversidad y riqueza de microflora y microfauna, cobertura vegetal, edad del
bosque, distribución de la precipitación en el tiempo de muestreo,momento del
año del comienzo de los ensayos etc.
La jara posee para el primer año tasas de decomposición de 0.31 año ,
incluidas dentro del rango hallado para otras esclerófilas mediterráneas.
Verdú (1984) calcula una k de 0.34 para la encina en la Castanya mientras que
Rosich eí al. (1989) encuentran un valor de 0.40 para la misma especie en la
Depresión Central Catalana. En especies de matorral californiano, Schlesinger
(1985) obtiene tasas de 0.36 año para Ceanhotus megacarpus y de 0.34- 0.4
para Salvia mellifera. En otro estudio similar, Schlesinger y Hasey (1981)
315
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
encuentran valores de K de 0.22 año para las mismas especies. Núñez (1989)
calcula una tasa de 0.54 para Cistus la.dan.ifer en Extremadura mientras que
Lavado et al. (1988) encuentran tasas superiores (0.61 año ) en el mismo
lugar para el mirto (Myrtus communis).
El comportamiento del gaznacho en cuanto a la dinámica de descomposición
se aleja algo del patrón encontrado para estos arbustos mediterráneos
esclerófilos con constantes de descomposición muy altas tanto para el primer
año (0.80) como para el segundo (0.6). Como ya comentamos, la condición de
fijadora de nitrógeno le permite alcanzar altos niveles de nitrógeno en hoja
viva y senescente, y consecuentamente bajos C/N. Esta características parecen
estar compartidas con otras especies fijadoras leñosas, (leguminosas o no)
(Schwintzer 1984). En este trabajo, se comentan algunos parámetros encontrados
en plantas fijadoras en relación con su descomposición, por ejemplo C/N, 7.
lignina, %N etc. En la tabla 5.13 aparecen algunos de estos resultados. Puede
verse como la hoja de Alnus glutinosa con un 3.06% de N en hojarasca
prácticamente se descompone durante el primer año (70-90 7. de pérdida). Alnus
rubra con un 1.527. de N pierde en total durante el primer año un 557. del peso
inical. Tanto la concentración de N como la pérdida de peso en esta especie es
muy similar a la encontrada para A. decorticans (1.51 7. de N y un 517. de peso
perdido). En estas especies así como en Robinia pseudoacacia que también
aparece en la tabla, las concentraciones iniciales de N, y C/N parecen
determinantes para la velocidad de descomposición. Sin embargo la pérdida de
peso en Myrica gale es sorprendentemente más baja que las demás fijadoras
(Schwintzer 1984) teniendo en cuenta sus bajos valores de C/N y altos 7. de N
inicial. El autor explica estas diferencias a part ir del porcentaje de lignina
encontrado para Myrica gale, muy superior al de las otras fijadoras (tabla
5.13). En este caso, la lignina se constituye como el factor dominante y
regulador de la descomposición de esta especie enmascarando el efecto del
nivel de nutrientes inicial (7.N ó C/N).
La relativamente alta velocidad de descomposición de A. decorticans
consecuencia de los factores estructurales y de composición elemental de su
hojarasca ya comentados, hacen que sus constantes de descomposición para el
primer año se encuentren muy próximoas a algunas especies caducifolías. En
este sentido Dziadowiec (1987) da valores para el primer año de 0.91 en tilo
316
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.13. Comparación de algunos parámetros importantes en la
descomposición de hojarasca de fijadoras leñosas (según Schwintzer 1984).
ESPECIE Biomasa perdida en primer año
>70-90
55
20-23
40-41
N(%) inicial
3.06
2.57
1.52
2.10
0.90
1.69
1.67
1.55
C/N inicial
16.3
19.5
31.5
23.8
55.6
28.9
24.2
32.3
% lignina inicial
21
10
10
40
25
26
Alnas glutinosa
Al ñus rubra
Ceanothus spp.
Myrica gale
Robinia pseudoacacia
1.06 en carpe y 0.63 en roble (en el gaznacho obteníamos 0.8 año ). Con
periodos de estudio superiores al año de muestreo se han encontardo tasas de
0.51 en j4cer saccharum similar al 0.53 año del gaznacho.
La pérdida de peso durante el primer año y más concretamente del periodo
inicial en el que normalmente existe una brusca e intensa disminución de masa
puede sesgar el valor final de la tasa de descomposición calculada para un
periodo mas largo (Dziadowiek 1987). De las cuatro especies estudiadas sólo
los arbustos presentan caídas de peso importantes en los tres primeros meses.
De hecho la kl es bastante superior a K2 en ambas especies. Sin embargo la
tasa calculada para todo el periodo sin contar esos primeros 89 días no
resulta significativamente distinta a la obtenida para todo el periodo en
ninguna de las dos especies (tabla 5.12). Sin embargo otros autores sí que han
encontrado diferencias importantes. Stohlgren (1988) elimina los 0.7 años
317
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
iniciales para el cálculo de la tasa de descomposición para los 3.6 años que
dura su estudio encontrando una reducción media en esta tasa de 347.. con lo
que los parámetros que se deducen a partir de ella (T957., T ) varían también
de una forma notable. Gosz et al. (1973) calculan la tasa de descomposición en
especies del bosque de Hubbard Brook para el periodo completo (12 meses) y
eliminando el primer de fuerte lixiviado (11 meses) encontrando una reducción
media del 307.. Existe en estos casos una fuerte influencia de ese primer
periodo en el resultado final.
Otros autores han comparado las constantes de descomposición entre el
primer año y las de años subsiguientes (Stolhgren 1988, Schlesinger 1985),
encontrando a veces fuertes reduciones en ellas, ya a partir del segundo año.
En este estudio, no se han encontrado diferencias estadísticamente
significativas para las especies de pino entre ninguna de las t res tasas
calculadas (Kl, K2 y Kt). Estos resultados reflejan una relativa constancia en
la pérdida de peso a lo largo de los dos años de estudio. Como se vio en la
figura 5.1.a., no se manifiesta de una manera patente una fase inicial de
brusca caída de peso en ninguno de los pinos. El comienzo del ensayo para la
hojarasca de pino se produjo en Octubre cuando ya habían comenzado la lluvias
otoñales y las condiciones son,pues, más óptimas para ese lixiviado inicial
que, a pesar de ello no se produjo o lo hizo con una baja intensidad. Los
motivos podrían buscarse en la utilización de hojas senescentes recién caídas
y no verdes como algunos otros autores (Dziadowiec 1987), la fuerte
retraslocación antes de la caída de hojarasca, el bajo nivel de compuestos
lixiviables,' y la posible pérdida de peso desde la caída hasta que fueron
recogidas la acículas para iniciar el ensayos entre otros.
En las especies arbustivas sí que se encontraron diferencias entre primer
y segundo año debidas básicamente a una mayor caída en los t res primeros
meses. De hecho kl es en ambas especies bastante superior a K2 aunque en A.
decorticans no alcance la significación por los motivos ya comentados
anteriormente. Schlesinger (1985) apunta también disminuciones en la k de 727.
para Ceanhotus entre la tasa del primer año y de los tres años en el chaparral
californiano. Sin embargo, las tasas calculadas para todo el periodo en el
matorral no resultan estadísticamente distintas a la del segundo año. Este
último resultado parece indicar que tampoco para las especies arbustivas puede
318
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
hablarse de una influencia determinante de «se primer periodo inicial en la
dinámica global de pérdida de peso.
5.3.2. DINÁMICA DE LOS NUTRIENTES EN EL PROCESO DE DESCOMPOSICIÓN.
5.3.2.1 Introducción.
Los nutrientes contenidos en las diferentes fracciones de un vegetal son
retornados al suelo mayoritariamente vía desfronde. La posterior liberación
durante la descomposición completa el ciclo de nutrientes entre el sistema
suelo-planta. Los mecanismos de retraslocación, muy desarrollados sobre todo
en especies mediterráneas (Schlesinger &. Hasey 1981, Read y Mitchell 1983),
pueden hacer disminuir considerablemente los contenidos de algunos nutrientes
(sobre todo N y P) en la hojarasca (Swift et al. 1979). El índice C/elemento
en el curso del proceso de descomposición refleja la relación entre la
dinámica de un nutriente y la del peso seco. Si el índice aumenta, el elemento
está siendo liberado (lixiviado o mineralizado). Si ei índice disminuye, el
elemento se acumula bien de manera biológica o físico- química. Si el índice
permanece constante la dinámica de ese elemento sigue un patrón similar al del
peso seco (Lousier y Parkinson 1978).En este sentido, pueden calcularse
valores críticos C/elemento por encima o debajo de los cuales un elemento es
retenido o mineralizado. A pesar de la importancia del C/elemento, otros
factores afectan a la mineralización durante la descomposición: calidad del
recurso y ambiente físico son parámetros que pueden afectar a los organismos
descomponedores y que también determinan la tasa con la que un elemento dado
estará disponible para el vegetal. (Swift et al. 1979).
5.3.2.2 Evolución de las concentraciones químicas.
Los cambios en la concentración química para los distintos elementos en
cada una de las especies aparecen en las figuras 5.9 a 5.14. El Mg y el K son
los elementos cuya concentración disminuye de una manera clara en todas la
especies en el transcurso del proceso. El descenso en la concentración de K es
muy marcada en ios primeros meses de muestreo manteniéndose después en unos
niveles prácticamente constantes a excepción de P. nigra en dónde la
319
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
concentración alimenta en los últimos muéstreos. A. decorticans y c.
LaurifolLus poseen concentraciones de K significativamente diferentes en la
hojarasca inicial (ver apartado 5.3.1.1) (0.69 7. en A. decorticans y 0.347. en
Cistus lawifolius); sin embargo, durante la descomposición estas
concentraciones disminuyen llegando a unos valores finales prácticamente
idénticos (0.067.). En las dos especies de pino ocurre algo parecido: las
concentraciones finales de K se sitúan alrededor del 0.257., pero en este caso
hay que tener en cuenta que sus respectivas cconcentraciones iniciales para
este elemento no difieren de manera significativa. La concentración de Mg
tiene un patrón de disminución más suave, progresivo y continuo que el del K.
N y Ca aumentan su concentración en todas la especies. En el matorral las
concentraciones de Ca se mantienen próximas al valor inicial oscilando por
encima y debajo de éste. La concentración de N aumenta progresivamente
conforme se va perdiendo la materia orgánica y por tanto la cantidad de C.
El Na es el elemento que presenta mayores oscilaciones de su
concentración durante el proceso, oscilaciones que resultan más marcadas en P.
nigra y A. decorticans. A. decorticans, P. pinaster y C. laurifolius tienden a
disminuir su concentración de Na. En el pino resinero y en la j a ra la
concentración de Na se estabiliza a partir de los 333 días y 234 días
respectivamente. La concentración de Na en la hojarasca de P. nigra tiende a
aumentar con el tiempo.
El P presenta un comportamiento bastante diferente según la especie que
se considere. En P. nigra la concentración disminuye los primeros 333 días
para después aumentar durante la fase final. En P. pinaster existe un descenso
acusado durante el primer mes, le sigue una fase de aumento progresivo y poco
importante y aumenta bruscamente durante el último periodo. Probablemente esta
subida final pueda deberse a una contaminación. La concentración de P en A.
decorticans aumenta paulatinamente durante el primer año para después mantener
unos niveles casi constantes. La ja ra posee concentraciones iniciales de P
significativamente superiores a las demás especies (ver apartado 5.3.1.1.).
Esta concentración disminuye rápidamente durante los primeros meses de
exposición para después mantenerse en niveles prácticamente constantes. La
disminución de concentración de P en esta especie es muy acusada pasando de
0.147. a 0.0477..
320
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
o o co 8
o o
o o co
o o
o o
S co
o o
2
O o
co co
o o
o o
o o o
o
2 [_
co d o" o
- o o o
z ,
Figura 5.9. Evolución de las concentraciones químicas en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Nitrógeno.
321
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
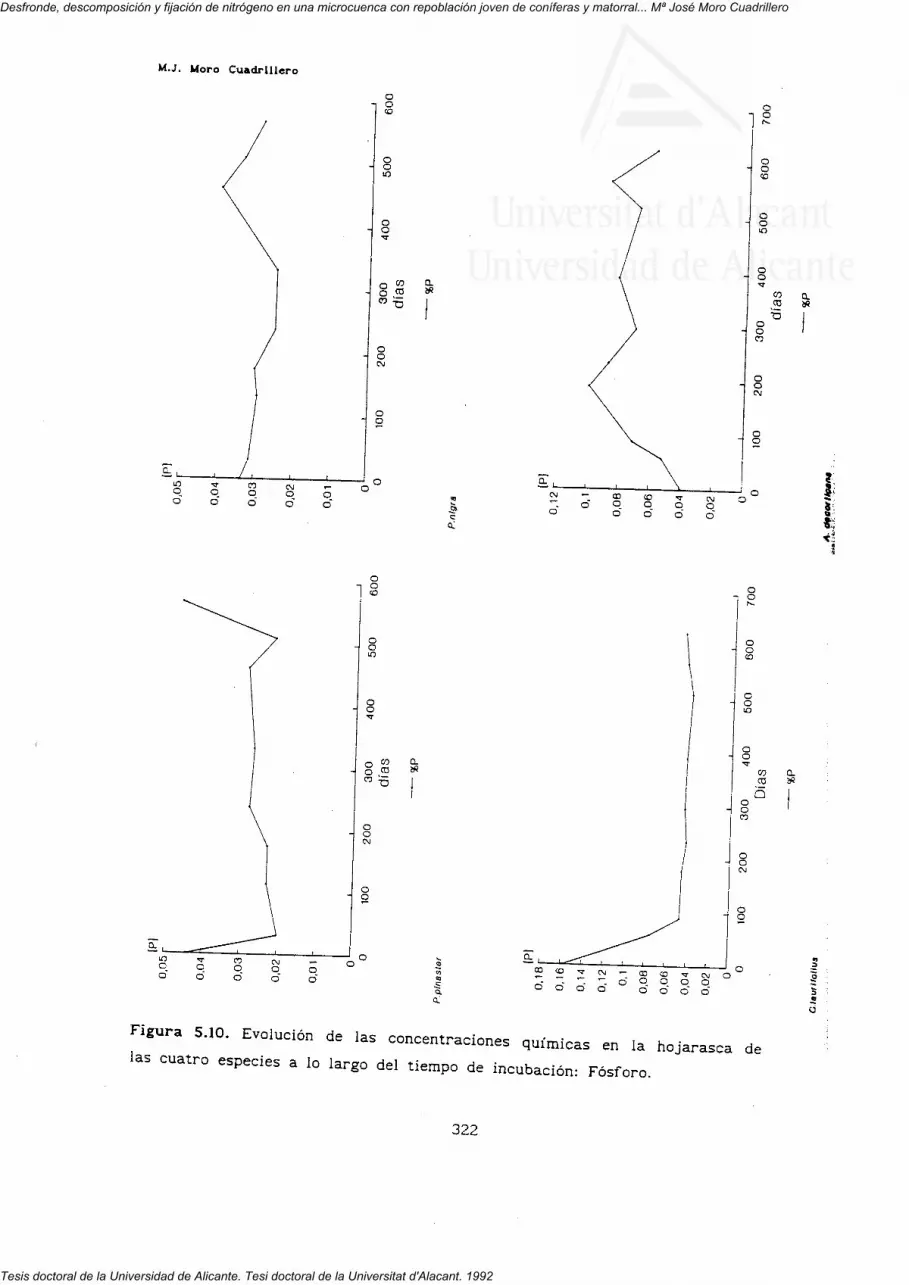
M.J. Moro Cuadrillero
o o
O O
o o
O <B EL
03 O O
<0 O O
T O
o"
tM O o"
I l tí
Q.
a.
Figura 5.10. Evolución de las concentraciones químicas en la hojarasca de
las cuatro especies a lo largo del tiempo de incubación: Fósforo.
322
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
o o
O O ¡a
o o
• l o »
o o
* *
o o
§ to o d
o d § § d §
*: *
co o d
o d
O d
CSJ
o d
Figura 5.11. Evolución de las concentraciones químicas en la hojarasca de
las cuatro especies a lo largo del tiempo de incubación: Potasio.
323
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
2 L
O 03 <0 S
ü .
CO
& CO Ü
Q.
U3 C\í
CD
Figura 5.12. Evolución de las concentraciones químicas en la hojarasca de
las cuatro especies a lo largo del tiempo de incubación: Calcio.
324
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Deicompo»lclón.
a
2 L m n o"
CTJ d
m CVJ
o
(N d
w *-d
,-o" ia o d
O W
d
Figura 5.13. Evolución de las concentraciones químicas en la ho ja rasca de
las cua t ro especies a lo largo del tiempo de incubación: Magnesio.
325
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
a z
TT
^ O
o o o
<— o n
03 O O
o
(O o o o
• o o o
r\j O O O
z
o C\J o o
1N O o
¡a
o o
*— o o
lO o o o
03
Z b
z i*
§ o o" §
o o
o o o"
Figura 5.14. Evolución de las concentraciones químicas en la hojarasca de
las cuatro especies a lo largo del tiempo de incubación: Sodio.
326
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Para establecer la relación entre la evolución del peso seco total y la
concentración de los distintos nutrientes hemos ensayado distintas ecuaciones
de regresión. Estas relaciones, sin embargo, no son determinantes para
establecer después la evolución de los contenidos de nutrientes ya que en este
último caso, existen otro factores que tienen una mayor importancia que la
propia evolución de la materia orgánica como son : el grado de unión de los
elementos al material vegetal y la mayor o menor demanda de los nutrientes por
parte de los heterótrofos (Dziadowiec 1987, Gosz et al. 1973, Swift eí al.
1979).
En general, la disminución de la materia orgánica suele ir acompañada de
aumentos correlativos en la concentración de los elementos exceptuando
aquellos que se pierden con mayor velocidad que la propia materia orgánica y
en los que la concentración disminuye. Se han encontrado correlaciones
inversas significas entre la masa remanente (en porcentaje de peso seco
inicial) y concentración para N en todas las especies, P en A. decorticans y
P. pinaster y Ca en P. pinaster y P. nigra. Las ecuaciones mejor ajustadas
(tabla 5.14) fueron lineales para N en las dos especies de matorral y para el
P en A. decorticans, potenciales negativas para el N en P. pinaster y
exponencial negativa para el Ca en ambos pinos y el N en P. nigra.
Los elementos que presentan correlaciones positivas significativas entre
masa remanente y concentración son K en matorral y P. pinaster, Mg en todas
las especies y Na para P. pinaster y A. decorticans. Los mejores ajustes se
consiguieron con ecuaciones lineales en el caso del Mg en matorral y P. nigra
y potenciales para Na y K en todos los casos (tabla 5.14).
5.3.2.3 Evolución del contenido en nutr ientes .
Introducción.
La tasa de liberación de nutrientes en la hojarasca durante la
descomposición depende sólo en un pequeño grado de la tasa de pérdida de peso
(Dziadowiec 1987). Los factores que deciden la dinámica y el curso del proceso
de liberación (o retención) de nutrientes son la forma en que se presenta un
determinado elemento en el material vegetal y la demanda de microorganismos y
327
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 5.14. Relación entre el porcentaje de peso remanente y la concentración
de los distintos nutrientes a lo largo del proceso de descomposición en lag
bolsas de las cuatro especies. Sólo aparecen las ecuaciones mejor ajustadas.
P.pinaster:
LN(% PSi)» 4.28 + LN [N] » (-0.164) r2= 0.70, p<0.001
LN(% PSi)- 4.97 + LN [K] * (0.168) r2= 0.51, p<0.005
LN(% PSi)= 5.08 + LN [Na]* (0.135) r2= 0.47, p<0.005
LNÍJÍ PSi)» 4.57 • e-5-02*tp3 r
2= 0>25, p<0.05
LN(% PSi)» 4.89 * e-°-2 9 5* C C a I
r2= 0.78, p<0.001
P. nigra:
LN(% PSi)« 4.78 * e -°- 9 0 9* t N ] r
2= 0.61, p<0.001
LN(0/. PSi)= 4.72 * e -°- 2 2 3* [ C a l r
2= 0.41, p<0.005
(%PSi) = 63.95 + 93.22 * [Mg] r2= 0.77, p<0.001
A. decorticans: (%PSi)= 156.4 - 45.18 * [N] r2= 0.87, p<0.001
(%PSi)= 94.71 - 596 * [P] r2= 0.29, p<0.05
C/.PSi)= 2.49 + 230.8, * [Mg] r2= 0.65, p<0.001
LN(%PSi)= 4.67 + 0.40 * LN[K] r2= 0.78, p<0.001
LN(%PSi)= 6.10 + 0.50 * LN[Na] r2= 0.22, p<0.05
C. laurifolius:
(JíPSi)» 119.24 - 82.6 * [N] r2= 0.87, p<0.001
(%PSi)= 63.26 + 242.1 * [P] r2= 0.60, p<0.001
(%PSi)= 40.69 + 148.8 * [Mg] r2= 0.75, p<0.001
LN(%PSi)= 4.84 + 0.212 * LN[K] r2= 0.58, p<0.001
328
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
otros descomponedores para un elemento dado (Dziadowiec 1987, Bockheim y Leide
1986, Carlyle 1986).
Los nutrientes que aparecen mayoritariamente en forma ionizada (K, Mg)
son liberados de la hojarasca mucho más rápidamente que lo que podría
esperarse a partir de la dinámica de la pérdida de peso durante ese mismo
periodo. El lixiviado juega un importante papel en el proceso de liberación de
estos elementos. Los nutrientes básicos para la síntesis de biomasa microbiana
(N, P) y aquellos que están intimamente unidos a estructuras celulares
persistentes (por ejemplo, paredes celulares) como es el caso del Ca, son
liberados durante la descomposición de manera más lenta que lo que cabría
esperar a part ir de las pérdidas de peso. En las fases iniciales del proceso,
que según muchos autores viene a coincidir con el primer año de decomposición,
nutrientes como el N y P pueden ser totalmente retenidos en la biomasa de
microorganismos o incluso acumulados. El Ca no suele ser liberado hasta que la
paredes celulares son desintegradas, por ejemplo por hifas de basidiomicetos
(Dziadowiek 1987) y el curso de su liberación está en estrecha consonacia con
la evolución de la pérdida de peso (Lousier y Parkinson 1978, Gosz eí al.
1973).
Se han barajado índices críticos,(Lutz y Chandler 1946), sobre todo C/N y
C/P por debajo de los cuales comienza una mineralización neta y por encima se
mantiene la inmovilización neta. Sin embargo, si bien estos índices
preestablecidos pueden ser útiles como valores orientativos, muchos trabajos
han demostrado que los valores críticos pueden ser extremadamente variables
dependiendo de muchos factores. Solamente para las coniferas Stolhgren
(1988) hace referencia a valores críticos de C/N que van desde 109 a 19.
En definitiva la base teórica que justifica estos valores críticos está
en el hecho de que una mineralización neta sólo comenzará cuando el elemento
liberado en la hojarasca exceda al requerido por los organismos. Esto ocurre
cuando el C/elemento del sustrato se aproxima o cae por debajo del de la
biomasa microbiana o descomponedora. Cuando el C/elemento excede al de los
microorganismos aparecerá una inmovilización neta.
329
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Resultados.
Los cambios en las cantidades de los distintos nutrientes en la hojarasca
en decomposición están expresados como porcentajes de las cantidades iniciales
de cada uno de ellos y pueden observarse en las figuras 5.15 a 5.20. y en las
tablas 5.15a a 5.15d. Valores por encima del 100% indican un incremento
absoluto (inmovilización + acumulación) del nutriente considerado. Valores
inferiores al 100% indican una disminución del pool del nutriente considerado.
La evolución del contenido en nutrientes sigue a grandes rasgos el mismo
patrón que la de las concentraciones químicas (figuras 5.15 a 5.20.) con dos
excepciones claras: la dinámica del N y Ca en A. decorticans. Así mientras que
la concentración de N sigue un patrón progresivamente asecendente, su
contenido total sigue una evolución totalmente opuesta (figura 5.9d y figura
5.15d). La concentración de Ca en A. decorticans oscila por encima y debajo
del valor inicial (figura 5.12d), sin sufrir grandes cambios. El contenido
total, sin embargo sigue un patrón continuo y descendente a lo largo de todo
el proceso (figura 5.18d).
El orden de liberación de nutrientes en las bolsas es el siguiente:
P.pinaster: K > Mg > Na > P - Ca > N
P. nigra : K > Mg > P > Na > Ca > N
C. laurifolius:K > P > Mg > Na > Ca > N
A. decorticans: K > Mg > Na > Ca > P > N
En todas las especies, el K es elemento mayormente movilizado, mientras
que el N ocupa siempre la última plaza. Para este nutriente, todas las
especies exceptuando el gaznacho presentan inmovilizaciones netas. Para los
demás elementos existen variaciones de una especie a otra, a veces muy
acusadas, como en el caso del P. Lo que en general puede observarse es que K y
Mg son los elementos más fácilmente liberados seguidos por el Na en algunos
casos mientras que Ca y N ocupan las últimas posiciones. El P no presenta una
pauta común respecto a su velocidad de liberación en las especies estudiadas.
En C. laurifolius este nutriente tiene una dinámica muy similar a la del K
ocupando la segunda plaza. En A. decorticans, si bien existe una liberación
330
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 15.a. Cistus laurifolius.
DÍAS N P K Ca Mg Na PESO
0
58
89
179
234
297
391
450
510
568
626
100
117.56
113.34
135.31
146.17
132.81
149.39
-
143.35
155.49
145.86
100
42.96
25.21
23.49
20.61
20.95
18.31
-
16.35
19.36
19.12
100
29.38
16.29
18.99
14.69
15.88
14.31
-
12.19
14.00
11.82
100
85.75
82.96
59.23
67.55
79.09
73.75
-
65.52
66.38
66.39
100
66.33
58.58
42.48
43.26
46.22
34.42
-
33.53
35.53
33.34
100
65.78
155.07
94.26
67.36
62.96
57.21
-
57.56
57.93
48.50
100
88.92
83.79
80.94
77.54
75.51
68.09
71.35
66.81
69.18
63.86
Tabla 15b. A. decorticans.
DÍAS N P K Ca Mg Na PESO
0
58
89
191
234
297
391
450
520
568
626
100
96.07
73.36
77.92
78.05
70.49
72.44
-
63.04
' 59.53
48.19
100
98.44
96.07
133.62
113.62
81.93
101.64
-
62.62
71.22
49.84
100
60.37
7.58
9.94
9.28
6.60
6.94
-
3.60
2.95
2.31
100
75.48
63.06
51.12
46.83
43.39
46.34
-
33.52
23.91
24.33
100
74.19
42.48
29.43
23.71
21.38
27.79
-
18.09
17.06
13.55
100
69.64
86.42
52.99
74.10
32.06
34.48
-
41.59
21.28
17.75
100
74.99
53.07
53.15
51.33
45.95
48.60
44.72
35.58
31.71
26.11
Tablas 5.15a y 5.15b. Valores medios de mineralomasa remanente para cada nutriente
(en % de peso seco inicial) en las especies arbustivas.
331
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadril lero
Tabla 5.15 c. P. p inaster.
DÍAS N P y K C a M9 Na PESO
0
31
114
176
239
333
392
462
510
568
Tabla 5.15.d.
DÍAS
0
31
133
176
239
333
392
462
510
568
100
92.825
106, .972
97.900
156.314
131.
142,
260
.723
.518
.127
237.303
P.nigra.
N
100
81.
102.
121.
111,
103.
110.
106.
124.
.14
.13
.18
.17
.43
.90
.76
.05
100
61. .61
67.45
69 .78
82.67
77.63
76.
56
118
.89
.75
.14
P
100
86.
76.
79.
64.
61.
91,
80.
64.
.11
.85
.79
.76
.64
.41
.45
.37
100
41.752
34.291
28.074
29.919
21.899
-
19.729
24.066
22.122
<
100
67.99
59.34
57.93
49.88
50.45
-
54.87
70.61
52.38
100
97.466
95.i
99.
650
709
103.551
104
119
106
121
(
.672
.787
.641
.061
:a
100
85
80
• 95
113
86
98
101.
113.
.88
.17
.07
.68
.19
.96
.57
.48
100
70.
64.
58.
53.
52.
39.
28.
29.
915
354
238
624
899
-
881
855
405
Mg
100
81
61
62
54
40
36
39
30
.22
.68
.57
.55
.50
-
.85
.69
.81
100
70.343
35.440
34.780
86.665
48.809
-
34.447
29.844
29.189
Na
100
250.85
102.75
177.98
98.01
122.41
-
137.08
155.20
130.07
100
91.182
88.400
88.389
87.019
86.046
83.295
79.048
75.471
74.351
PESO
100
91.92
87.18
88.47
87.05
82.73
77.78
76.46
78.91
74.03
Tabla 5.15 c y d. Valores medios de mineralomasa remanente (en % del contenido i n i c i a l ) para los dos especies de pino.
332
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
neta de P ésta es inferior al resto de nutrientes exceptuando al N. En el
pinar, la liberación de P se sitúa en posición intermedia, pero mientras que
al final del periodo de estudio el P es acumulado en P. pinaster no ocurre lo
mismo en el pino laricio donde se observa una liberación neta (si bien esta es
pequeña).
La mineralomasa de la hojarasca al final del periodo del que se disponen
de datos de concentración química (568 días en el pinar y 626 en el matorral)
puede observarse enla última fila de las tablas 5.15a a 5.15d. Existe
retención y acumulación para P, Ca y N en P. pinaster; Na, Ca y N en P. nigra
y N en la jara. En A. decorticans todos los elmentos presentan en mayor o
menor grado mineralización neta.
Es interesante destacar que, aunque el orden de liberación para las dos
especies de pino es bastante similar, no lo son las cantidades finales
observadas. Solamente los contenidos de Ca y Mg presentan similitudes. Sin
embargo la cantidad de de N remanente al final del periodo es muy alto en P.
pinaster un 1407. más alto que el contenido inicial frente al 24 7. de P. nigra.
Nitrógeno.
La cantidad absoluta de N en las bolsas se incrementa con el tiempo en el
pinar y en la j a ra mientras que disminuye en A. decorticans; por tanto se
produce una inmovilización- retención en las t res primeras especies y una
mineralización neta en el gaznacho.
Los contenidos en N en P. pinaster oscilan entre el 96-1067. durante el
primer otoño-invierno (primeros 114 días). Existe una liberación neta durante
el primer mes que supone el 7.27. del contenido inicial resultado probable de
lixiviado ayudado por las lluvias otoñales. A part ir de la primavera (176
días) se detecta ya una retención de N importante que aumenta
considerablemente hacia el final de periodo de estudio, (figura 5.15a).
Esta acumulación final llega a ser un 1407. superior al contenido inicial.
En los dos últimos muéstreos realizados para esta especie (510 y 568 días), el
contenido medio de N presenta unos elevados errores estándar debido a la
disparidad de concentraciones en cada uno de los dos replicados. En las
333
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
primera fecha, las concentraciones medias de N son de 1.22 7. y 0.48% y en la
segunda 1.157. y 0.427.. Nos inclinamos a pensar que los valores más bajos se
aproximan más a la realidad mientras que los valores que rondan el 1% pueden
haberse derivado de una contaminación local de la hojarasca contenida en las
bolsas. Si sólo consideramos el replicado de 7. de N inferior en cada uno de
los muéstreos obtenemos contenidos de N de 152 7. para los 510 días y de 1327.
en los 568 días, valores en principio más realistas y más de acuerdo con los
encontrados en la bibliografía.
En P. nigra, se observa también una importante disminución en la cantidad
de N durante los primeros 30 días mucho más pronunciada que la descrita para
P. pinaster perdiéndose un 197. del contenido inicial. Igual que en la especie
anterior la causa presumible es el lixiviado. Este es el único momento en el
que la curva que sigue el N en P. nigra queda por debajo de la de materia seca
(figura 5.15b). A partir de aquí, se observa una progresiva retención a lo
largo del proceso. El contenido final de N es un 24 % superior al contenido
inicial.
El nitrógeno permanece retenido en la hojarasca de la j a ra desde los
primeros 58 días. La inmovilización aumenta progresivamente a lo largo del
tiempo de exposición. Al final del periodo la cantidad de N excede en un 45.87.
a la inicial. No se ha detectado en ningún momento liberación de este elemnto,
ni siquiera en los primeros periodos de recogida.
La mineralización neta de N en A. decorticans comienza ya en las etapas
tempranas de descomposición. Tras los primeros 58 días A. decorticans pierde
ya un 307. de su contenido inicial de N. Esta primera fase probablemnte se deba
a lixiviado más que a factores biológicos. La liberación continua
progresivamente en los meses sucesivos. Al final del periodo A. decorticans ha
perdido un 55 7. del contenido inicial de N. A pesar de la concentraciones de N
progresivamente ascendentes en A. decorticans, la evolución del contenido de
este elemento es descendente. En este caso la rápida pérdida de peso es el
factor dominante en la evolución del contenido en N. En la figura 5.15a puede
apreciarse que la materia orgánica (peso seco) y el N siguen una dinámica muy
similar si bien la curva de N se sitúa siempre por encima de la del peso seco.
La correlación es de r= 0.98 (p<0.001) entre ambas curvas.
334
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
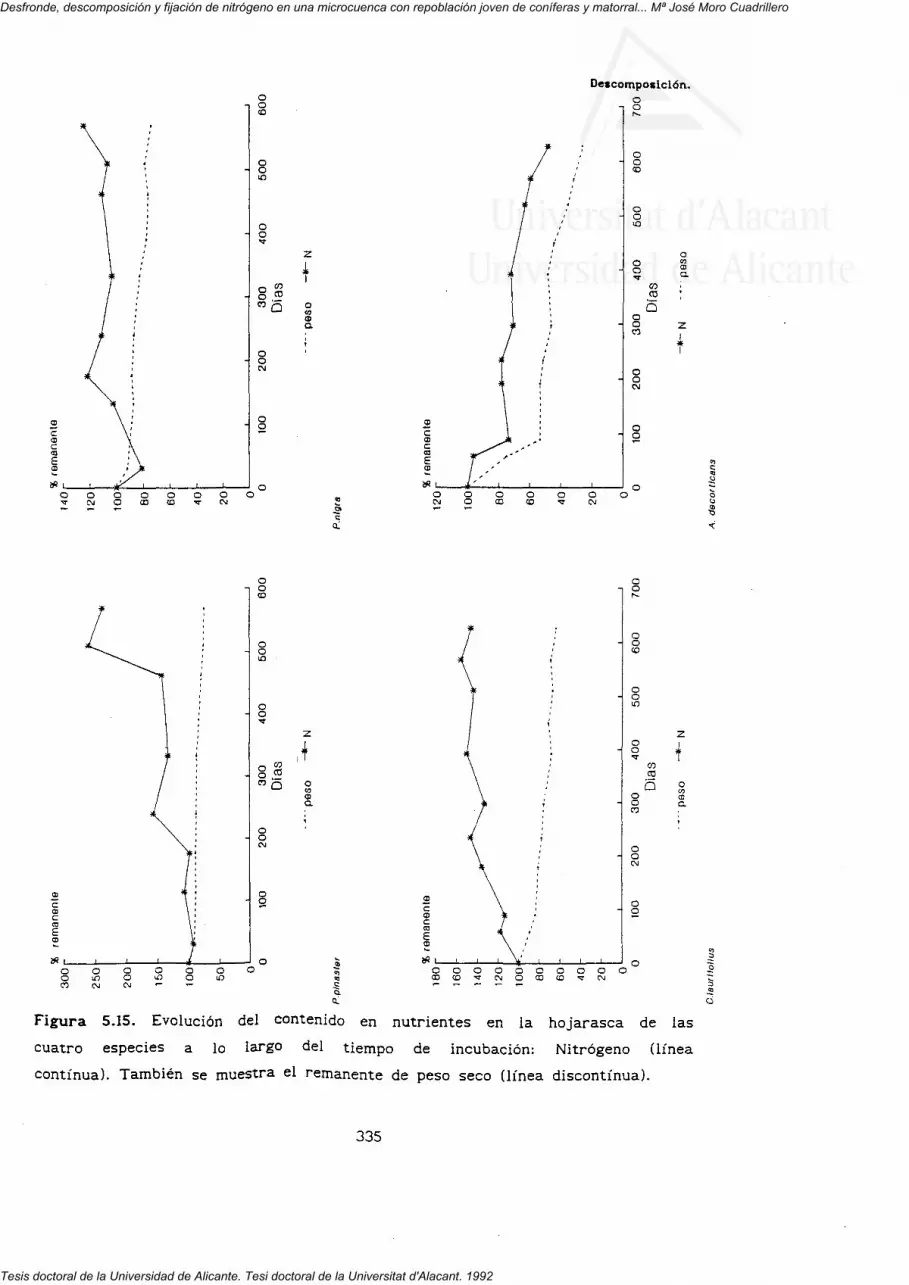
Descompoíiclón. o
Q
o o
o co
o o ÍM
o o en
o m CM
o o M
O iO
*~ ó o *~
o o
Figura 5.15. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Nitrógeno (línea
continua). También se muestra el remanente de peso seco (línea discontinua).
335
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
El valor C/N inicial en A. decorticans es el más bajo de las t res
especies: 31/1. Sin considerar el periodo inicial de lixiviado (los primeros
58 días) el índice C/N ha descendido a 25.5/1 (tomando como 7. de C un 50%).
Fósforo.
El contenido de P en la hojarasca del pinar sigue comportamientos
diferentes en ambas especies. Al final del periodo de seguimiento existe una
inmovilización neta en P. pinaster y mineralización neta en P. nigra (tabla
5.15 c y d y figuras 5.16a y b). En ambas especies coexisten periodos de
retención con otros de liberación de P.
Durante los 31 primeros días de exposición se observó un descenso en la
cantidad absoluta de P en la hojarasaca de ambos pinos, presumiblemente debido
a lixiviación. Este descenso es más acusado en el pino resinero que pierde
durante el primer mes un 38 7. del P total inicial. En esta especie comienza a
continuación un periodo de inmovilización que se mantiene en niveles de
aproximadamente 75-807. del P inicial. Durante esta fase la curva sigue una
evolución muy parecida a la del peso seco total. A los 518 días se observa un
cierto grado de mineralización, seguido de un considerable aumento en la
cantidad de P que sobrepasará al contenido inicial en un 18 7. a los 568 días.
En P. nigra la liberación de P se produce progresivamente durante el primer
año, siendo siempre estas tasas de liberación superiores a las del peso seco
total (figura 5.16b). Durante el invierno 89-90 (462 días) se detecta un
periodo de inmovilización transitorio, seguido de liberación de P hasta el
final del periodo. La hojarasca de esta especie pierde al final de los 568
días un 35.6 7. de la cantidad inicial de este elemento.
La evolución del contenido en P en A. decorticans puede dividirse en dos
tramos bien diferenciados. El primer año está dominado por la inmovilización
neta e incluso la acumulación si bien, existe una pequeña liberación en los
tres primeros meses que no supera el 47.. Esta fase viene seguida de un periodo
de mineralización neta durante el segundo año. Las tasas de liberación en esta
última fase no superan a las de la materia orgánica (peso seco total) (figura
5.16d).
336
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

De«compo«lclón.
o. i
o 1 o
03 C0
Figura 5.16. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Fósforo (línea continua).
También se muestra el remanente de peso seco (línea discontinua).
337
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La dinámica del P en la ja ra sigue un patrón bien distinto a las de las
otras t res especies siendo su principal característica la intensidad y rapidez
de su liberación (figura 5.16c). La mineralización neta de P ocurre desde los
primeros días de exposición si bien, la dinámica del proceso sigue dos fases
bien diferenciadas. Una primera fase de liberación rápida acontece en los 89
días t r a s el comienzo del ensayo. Durante este periodo un 74.79 7. del P
inicial es liberado. Esta fase puede estar muy influenciada por las elevadas
concentraciones de P en la hojarasca de C. laurifolius que producen un índice
C/P bajo (357) que puede permitir la mineralización neta de este elemento.
Aunque el P no es un nutriente muy lixiviable, este proceso también podría
jugar su papel durante la primera fase. Esta etapa viene seguida de un periodo
de estabilización caracterizado por una liberación muy lenta durante la que
sólo se pierde un 4.37 7. del P inicial. La liberación de P es mucho más rápida
que la pérdida de peso. En la figura 5.16, la curva que describe el P se sitúa
siempre por debajo de la del peso seco. Al final del periodo de estudio (626
días) la mineralomasa remanente de fósforo es tan sólo del 19.12 7.. La
cantidad mineralizada es, por tanto, muy superior a las de las otras t res
especies. En este sentido, cabe recordar que las concentraciones de P en la
hojarasca de ja ra eran significativamente superiores a las del pinar y
gaznacho y que por tanto se obtenían valores bajos del índice C/P en la jara ,
el cual, a su vez, puede ser un factor determinante en el proceso de
mineralización neta del P en esta especie.
Potasio.
Como era de esperar, el comportamiento general de este elemento para
todas las especies es la de un nutriente que se libera con una gran rapidez
durante los estadios iniciales (dos primeros muéstreos) para después
mantenerse en niveles muy inferiores a los iniciales de una manera más o meno
estable (figura 5.17). Es conocida la facilidad del K para ser lavado. A la
vista de los resultados, el lixiviado de K fue intenso para todas las especies
favorecido, sin duda por las lluvias otoñales durante el otoño-invierno 88-89.
Las pérdidas de K son muy superiores a las del peso seco. Al final del
periodo de estudio sólo el 22.12 7. de K inicial en P. pinaster, 527. en P.
nigra, 11.87. en C. Laurifolius y 2.31 7. en A. decorticans permanecían en la
hojarasca. Destaca la pérdida relativamente lenta del potasio en P. nigra en
338
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
m
Q
Figura 5.17. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Potasio (línea continua).
También se muestra el "remanente de peso seco (línea discontinua).
339
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
que se observa, además, una etapa de inmovilización hacia el final del periodo
de exposición (516 días) que hace aumentar considerablemente la cantidad
absoluta remanente de K al terminar el estudio.
Las especies que inicialmente contaban con mayores concentraciones de K
en la hojarasca son las que presentan una dinámica más rápida en la liberación
de éste. Existe una correlación negativa aunque no significativa entre la
concentración inicial de K y la cantidad absoluta remanente de K al final del
periodo.
Calcio.
El calcio presenta una mineralización neta relativamente rápida y
continua en el gaznacho aunque como vimos anteriormente las concentraciones
químicas de este elemento aumentaban progresivamente con el tiempo de
exposición (figura 5.12d). La descomposición de la materia orgánica, expresada
aquí como peso seco total, parece ser el principal factor que controla la
liberación de calcio. De hecho, la figura 5.18d muestra la similitud entre la
dinámica del calcio y la del peso seco. Se ha encontrado una correlación
significativa entre ambas curvas (r= 0.96 p< 0.001). Al cabo de los 626 días,
sólo un 24.3 % de Ca es retenido en la hojarasca de esta especie, valor muy
similar al peso seco remanente para este mismo periodo (26.11%).
El Ca es también liberado desde el comienzo de la exposición en la otra
especie arbustiva téistus laurifolius), pero su dinámica no presenta la
continuidad apreciada en A. decorticans. Existe un brusco descenso en la
cantidad de Ca durante los primeros 179 días. Posteriormente, ésta aumenta
progresivamente (inmovilización) para disminuir después (liberación neta)
hasta el final del periodo. En este punto, la pérdida de Ca es de un 33.67.
valor que resulta bastante similar al peso seco perdido en el mismo intervalo
(36.14%). La dinámica del Ca en la j a ra parece seguir un comportamiento
similar al del peso seco si exceptuamos el periodo comprendido entre los 179 y
234 días (figura 5.18c).
La inmovilización y acumulación neta de Ca es la característica más
destacada de la dinámica de este elemento en el pinar. En P. pinaster, después
de un período en el que se aprecia una ligera liberación de Ca (de un 5 %
340
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
o S 05 Q
o
o £ " O
o o
Figura 5.18. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Calcio (línea continua).
También se muestra el remanente de peso seco (línea discontinua).
341
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
aproximadamente, figura 5.18a), comienza una fase de inmovilización que se
prolongará hasta el final del periodo de estudio. En el último muetreo, i a
cantidad de Ca sobrepasaba en un 217. la cantidad inicial. La curva del
contenido en Ca en esta especie durante la descomposición se sitúa en todo
momento por encima de la del peso seco.
P. nigra presenta una dinámica algo más irregular que P. pinaster
respecto a la evolución del contenido en Ca. Se observa una primera fase de
liberación (figura 5.18b) que transcurre durante los primeros 133 días. La
pérdida de Ca en ese momento es del 207.. A part i r de esa fase, la cantidad
absoluta de Ca en hojarasca aumenta, aunque de forma algo irregular ya que
existen periodos transitorios de liberación (333 días). El contenido final de
Ca es de un 13 7. superior al que existía al comienzo de los ensayos.
Magnesio.
El magnesio presenta un patrón muy similar al del K, sin embargo su
liberación en la hojarasca se realiza de forma más lenta que la de aquel. En
todos las especies las tasas de pérdida de Mg son más altas que las
correspondientes al peso seco. Debido a la intensa pérdida de materia orgánica
durante la descomposición en A. decorticans, las fase iniciales y finales del
proceso de mineralización del Mg se aproximan mucho a las de la evolución del
peso total en esta especie (figura 5.19d).
Al final del periodo de estudio, la cantidad absoluta remanente de Mg es
muy similar en los pinos (29.47. en P. pinaster, 30.8 7. en P. nigra) y en la
jara (33.4%) (tabla 15a, c y d). A. decorticans pierde una mayor cantidad de
mg permaneciendo al final del estudio tan sólo un 13.5 7. del contenido inicial
de este elemento en su hojarasca.
Sodio.
Existe una fuerte irregularidad en el comportamiento de este elemento
tanto temporalmente (periodos de muestreo) como interespecificamente (figura
5.20a a 5.20d). Así, aunque en todas las especies el resultado final sea una
mineralización neta de sodio han de tenerse en cuenta las siguientes
consideraciones:
342
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Figura 5.19. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Magnesio (línea
continua). También se muestra el remanente de peso seco (línea discontinua).
343
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
- Se observa una fuerte acumulación de Na en P. nigra para la mayoría de
los periodos de muestreo. El contenido en este elemnto presenta oscilaciones
sobre todo durante los primeros 239 días.
- No se observa acumulación neta de Na en P. pinaster. Existe una
considerable liberación durante los primeros t res meses y medio (64.57.). La
liberación progresa después mucho más lentamente con periodos de
inmovilizaciones transitorias (figura 5.20a). Al cabo de los 568 días sólo
un 29.2 % de Na permanece en la hojarasca.
- El Na es rápidamente mineralizado en A. decorticans. Al cabo de los 626
días sólo el 11.17. del contenido inicial permanece en la hojarasca. Durante
los primeros 58 días se constata una pérdida inicial atribuida a procesos de
lixiviación dado el carácter relativamente móvil de este elemento. A este
periodo inicial se suceden fases transitorias de inmovilización/liberación sin
un patrón definido.
- La jara es la especie que libera más lentamente el Na. Después de una
lixiviación rápida en la que la hojarasca pierde un 34.27. del contenido
inicial de sodio se sucede una fase de intensa acumulación en el que fueron
detectadas cantidades un 557. superiores al contenido inicial. Posteriormente,
una última fase de progresiva liberación conduce a contenidos finales de Na en
la hojarasca del 48.5 7. de la cantidad inicial.
- Existe una correlación significativa pero débil entre el contenido
final de Na en las cuatro especies y sus concentraciones iniciales de Na en
hojarsca (r= 0.88)
5.3.2.4. Aplicación del modelo exponencial simple a algunos nutrientes.
Algunos autores han aplicado ecuaciones exponenciales negativas simples
para describir la dinámica de los nutrientes durante la descomposición
(Bockheim y Leide 1986, Attiwill 1968). El modelo no se aplica a los elementos
que presentan inmovilización o acumulación. La validez del modelo es también
cuestionable para aquellos elementos cuya dinámica se basa en una fase de
retención seguida de una fase de liberación (Bockheim y Leide, 1986).
344
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
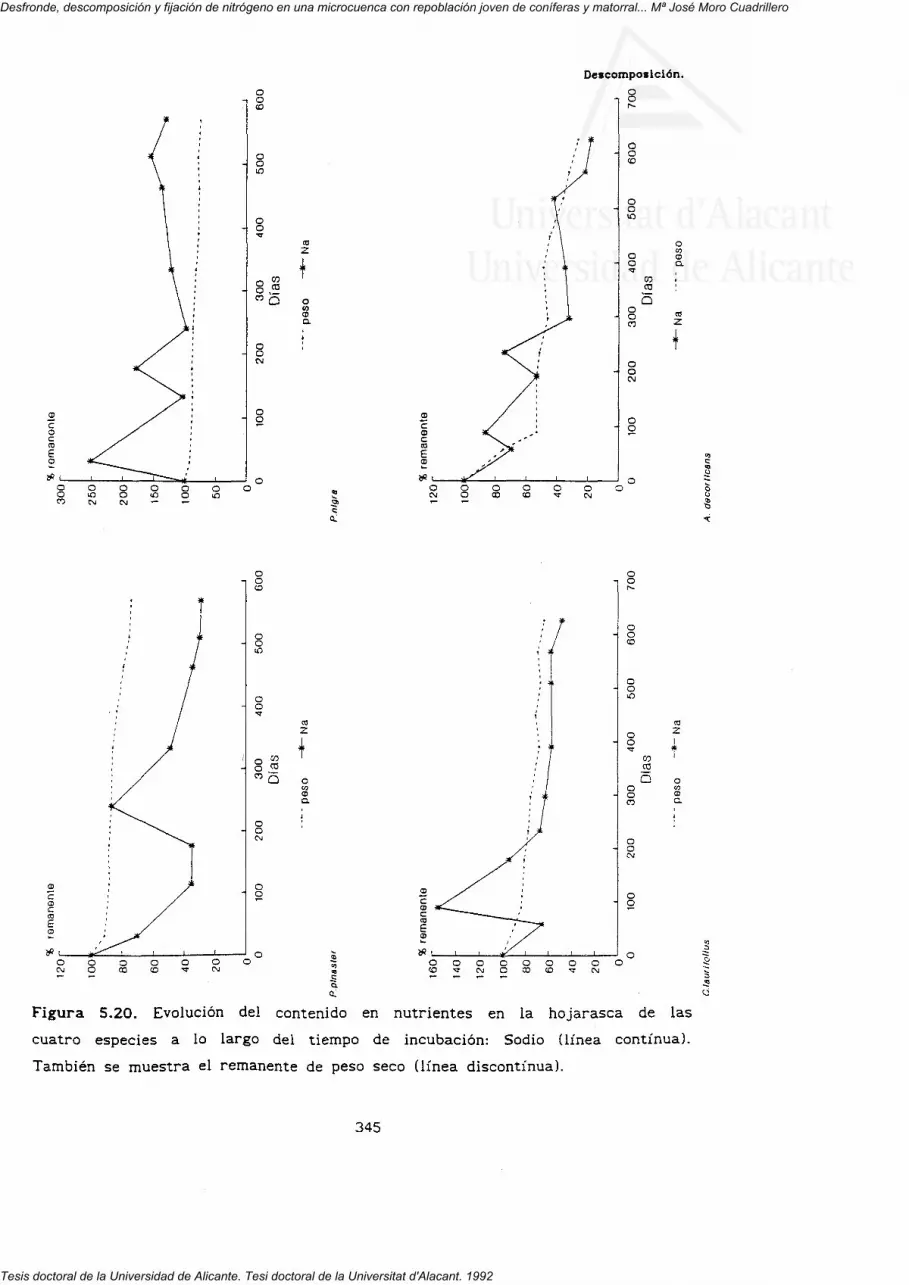
Descomposición.
Figura 5.20. Evolución del contenido en nutrientes en la hojarasca de las
cuatro especies a lo largo del tiempo de incubación: Sodio (línea continua).
También se muestra el remanente de peso seco (línea discontinua).
345
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
En nuestro estudio, el modelo puede resultar válido para aquellos
nutrientes que presentan un patrón de clara liberación desde los momentos
iniciales de la exposición (ver apartado 5.3.2.3). Consideraremos el K y Mg en
todas las especies, el Ca en el matorral, el P en la j a ra y el N y Na en A.
decorticans.
Las ecuaciones exponenciales resultantes son en todos los casos
significativas a nivel p<0.001 exceptuando la obtenida para el K en P. nigra
p< 0.05 y para el Ca en C. laurifolius p<0.005. Los valores de k resultantes
aparecen en la tabla 5.16. La menor significación obtenida para la ecuación
que relaciona la masa remanente de K en P. nigra frente al tiempo se debe al
comportamiento de este elemento durante los estadios finales de exposición en
los que se observa inmovilización (figura 5.17b). De ahí que los valores de la
constante de descomposición para este elemento resulten ser mucho más bajos
(k=-0.199) de los esperables a la vista de la figura 5.17b. En este sentido,
la aplicación del modelo exponencial negativo para describir la dinámica del K
en esta especie es también cuestionable.
Desde el punto de vista interespecífico el orden que ocupa cada especie
respecto a la tasa de liberación de K es la siguiente:
A. decorticans > C. Laurifolius > P. pinaster » P. nigra
Como ya hemos comentado, el valor de kK para P. nigra está muy
influenciado por la fase final de inmovilización. Excluyendo los muéstreos
correspondientes a ese período, la constante de descomposición calculada sería
de -0.65.
Respecto al Mg, el orden que ocupan las especies es :
A. decorticans > P. nigra - P. pinaster > C. laurifolius
Respecto al Ca:
A. decorticans > C. laurifolius
346
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5.16. Ecuaciones r e su l t an tes de apl icar el modelo exponencial simple
(linearizado) en aquellos nu t r i en tes que presentan una dinámica de l iberación
neta . La variable dependiente es tá expresada como porcenta je de masa remanente
de un nu t r i en te determinado. El t iempo e s t á expresado en años.
P.nigra:
G feQ*t 7
LN(7.Mg)= 4.46 • e r = 0.76, p<0.001 -O 20*t 2
LN(7.K) = 4.26 * e r = 0 .23, p<0.05
P.pinaster:
LN(7.K)= 3.95 * e" r = 0.62, p<0.001 O ɻ8*t 7
LN(7.Mg)= 4.44 * e r = 0.90, p<0.001
C. laurifolius:
LN(7.K)= 3.54 * e - 0 ' 7 5* 1 r 2 = 0.50, p<0.005 -O 71* t ?
LN(7.Ca)= 4.49 * e r = 0.42, p<0.005
LN(7.Mg)= 4.25 * e"0 '51*1 r 2 = 0.73, p<0.001
LN(7.P)= 3.78 * e -0-67*1 r
2= 0 . 5 1 , p<0.001
A. decorticans:
LN(7.N)= 4.54 * e"0 '31*1 r 2 = 0.81, p<0.001
LN(7.K)= 3.61 • e"1-69*1 r 2 = 0.72, p<0.001
LN(7.Mg)= 4.16 * e"0 '96*1 r 2 = 0.77, p<0.001 —O 68* t 7
LN(7.Ca)= 4.40 « e r = 0.86, p<0.001 -O <53*t 7
LN(7.Na)= 4.56 * e r = 0.72, p<0.001
347
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Se ha aplicado un tes t - t para comprobar si estas constantes de
descomposición son diferentes desde el punto de vista estadístico. Los
resultados del test aparecen en la tabla 5.17.
Se observa que A. decorticans tiene constantes de descomposición
significativamente superiores al resto de especies para el K (-1.69). Jara y
pino resinero no muestran diferencias significativas en cuanto a K (-0.74 y
-0.68 respectivamente, t= -0.35, n.s.) mientras que el pino laricio tiene
tasas de liberación de este elemento significativamente inferiores a las de
las demás especies (-0.199).
Respecto al Mg, A. decorticans es la especie con una mayor tasa de
liberación de este nutriente (-0.957), pero ésta sólo es significativamente
mayor que la de la jara con una kMg de -0.516 (p<0.005, t=3.08). Los pinos no
difieren entre sí respecto a kmg (-0.68 y -0.69 respectivamente) (t= 0.066,
n.s.). Además, ninguna de las especies de pino tiene diferencias
significativas con ninguno de los dos arbustos.
La tasa de liberación de Ca es significativamente mayor (-0.678) en A.
decorticans que en C. laurifolius (p<0.001, t=5.3).
La mayor o menor velocidad de liberación para los distintos elementos
dentro de una misma especie también se ha testado utilizando el tes t - t . Los
resultados de su aplicación están marcados con un número en la tabla 5.17.
En el gaznacho, la tasa de pérdida de N es significativamente inferior
(kN=-0.313) a la de los otros elemntos, mientras que el K posee velocidades de
liberación significativamente superiores (-1.69) a las de los otros
nutrientes. Ca, Mg y Na no difieren entre sí respecto a sus velocidades de
liberación.
En la jara , el Ca presenta una tasa de liberación significativamente más
baja (kCa=-0.21) que la del K.P.y Mg. Las tasas calculadas para estos tres
últimos elementos no son significativamente diferentes.
No existen diferencias significativas en las constantes de descomposición
para el K y Mg en P. pinaster (t= 0.027, n.s.), sin embargo sí existen
348
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Tabla 5-17. Valores de k para los nutr ientes que presentan una dinámica de l iberación neta y resultados del t e s t - t . Para una misma columna, los valores seguidos por el mismo numero no son signi f icat ivamente d i s t i n t o s . Para una misma f i l a , los valores seguidos de una misma le t ra no son signi f icat ivamente d i s t i n t o s . El número de datos es de 20 para las especies arbustivas y de 18 para el p inar. Todas las ecuaciones son s ign i f i ca t i vas con p<0.001 excepto la correspondiente a l K en P.nigra (p<0.05) y e l Ca en C . lau r i f o l i us (p<0.005).
N
P
K
Ca
Mg
Na
P.pinaster
1
.688
a
1
.684
ab
P, .nigra
1
.199
c
2
.691
ab
C. lauri folius
1
.667
1
.749
a
2
.210
b
1
.516
a
A. .decorticans
3
.313
1
1.690
b
2
.678
a
2
.957
b
2
.932
diferencias para estos mismos elementos en P. nigra, siendo más altas las
correspondientes al Mg (t= 3.71, p< 0.005).
5.3.2.5. Discusión.
Nitrógeno.
El N junto con el P constituye un macronutriente esencial para la
nutrición de los vegetales y ha recibido una considerable atención en los
estudios de descomposición (Bockheim y Leide 1986, Berg y Staaf 1981).
349
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Debido a este papel de nutriente esencial, el N tiende a ser conservado
por la planta y su concentración en hojarasca no suele ser alta. En coniferas
son conocidos los bajos niveles de N y en general de la mayoría de los
nutrientes en hoja viva, lo cual se reflejará también en bajas concentraciones
en hojarasca. Además los fenómenos de retrascolación inmediatamente antes de
la abcisión juegan un importante papel en la disminución de la concentración
de N en hojarasca. Según Read y Mitchell (1983), esta reabsorción puede ser
particularmente intensa en vegetales mediterráneos perennes. La traslocación
puede incrementar en buena medida el C/N inicial en las hojas caídas y por
tanto aumentar la conservación de N en el ecosistema al reducir la tasa de
descomposición microbiana (Read y Mitchell 1983).
La disminución progresiva del C en el material foliar durante el proceso
de descomposición causa un incremento en las concentraciones de N (Gosz eí al.
1973) haya o no haya liberación de N (Berg y Staaf (1981). Este aumento
progresivo de la concentración de N ha sido encontrado en las cuatro especies
estudiadas, así como en los trabajos consultados de la bibliografía
(Dziadowiec 1987, Lousier y Parkinson 1978, Maheswaran y Attiwill 1987,
Klemmedson eí al. 1985, Verdú 1984, Gosz eí al. 1973).
Berg y Staaf (1981) describen la dinámica del N durante la descomposición
como un proceso que sigue tres etapas consecutivas: una fase corta de
liberación (aproximadamente los 100 primeros días) atribuible a lixiviado sin
intervención de factores biológicos (Gosz eí al. 1973, Dziadowiec 1987), una
fase de inmovilización y/o acumulación (hasta unos 500 días) debida a
utilización microbiana del recurso y una última fase consistente en una
liberación neta de este nutriente.
La fase de lixiviado ha sido constatada en las especies de pino
estudiadas, con especial incidencia en P. nigra cuya hojarasca libera durante
el primer mes un 19 7. del contenido inicial. Varios autores han constatado la
existencia de esta fase inicial, sobre todo aquellos trabajos en los que el
comienzo del seguimiento de la descomposición coincide con el periodo
lluvioso. Gosz eí al. (1973) detecta lixiviados de N de aproximadamente un 20%
durante el primer mes de exposición en Fagus grandifolia en Hubbard Brook,
Maheswaran y Attiwill (1987) cita un 157. de lixiviado en Eucalyptus microcarpa
350
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
en Australia y Bockheim y Leide (1986) de un 57. de pérdida en Pinus resinosa
en Wisconsin también en los primeros treinta días. Sin embargo, en otras
especies, (Dziadowiec 1987, Gosz eí a i . 1973, Schlesinger 1985) no se han
encontrado lixiviados iniciales sino inmovilización o acumulación neta desde
el comienzo tal y como ocurre en este estudio con la ja ra .
La mayoría de los autores hacen referencia a la inmovilización de N e
incluso a su acumulación (aumentos en las cantidades absolutas) a lo largo de
la descomposición. Estos procesos aparecen también en t res de las especies
estudiadas: P. pinaster, P. nigra y C. laurifolius. En coniferas, Dziadowiec
(1987) cita retenciones de N en acículas de P.sylvestris en Polonia.
Klemmedson eí al. (1985) en Pinus ponderosa encuentra inmovilizaciones durante
los primeros 21 meses, Verdú (1984) en hojas de abeto al final del primer año
en el Montseny, Fahey (1983) en Pinus contorta durante los dos primeros años,
Alvera (1990) en Pinus sylvestris de Pirineos, Stohlgren (1988) en Sequoia
gigantea, Pinus lambertiana y Calocedrus decurrens de Sierra Nevada
(California). En no coniferas, Schlesinger (1985) también describe
inmovilizaciones en hojas de Ceanothus y Salvia en el chaparral californiano,
Verdú (1984) en Quercus ilex del bosque de La Castanya, y Alvera (1990) en
Ilex aquifolium en los Pirineos. En cuanto a especies caducas la retenciones
de N han sido descritas por Gosz eí ai . (1973) para varias especies arbóreas
caducas de Hubbard Brook durante el primer año, Lousier y Parkinson (1978) en
dos especies de Populus hasta los 8-12 meses, Dziadowiec (1987) en carpe, tilo
y roble durante el primer año. O'Conell (1987) y Maheswaran y Attiwill (1987)
encuentran pautas similares para Eucaiyptus diversicoLor y £. microcarpa en
Australia.
La inmovilización de N parece, pues, una pauta muy general en la dinámica
del N sea cual fuere el tipo de hoja que se considere. Según Gosz eí a i .
(1973), Carlyle (1986) y Swift eí ai. (1979), este proceso de inmovilización
tiene relación con la demanda de N por parte de los heterótrofos que supera en
los primeros estadios de descomposición a la liberación de N por parte de la
hojarasca con lo que éste es inmediatamente retenido en la biomasa microbiana.
Además, en el pinar estudiado y en la ja ra no sólo se observan
inmovilizaciones sino aumentos absolutos en la cantidad de N que superan al
351
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
contenido inicial. Estos aumentos no pueden explicarse por el aumento de
concentración de N en el transcurso del proceso, sino que han de explicarse a
part ir de fuentes externas que determinen en mayor o en menor grado una
importación de N. La mayoría de los trabajos citados en el párrafo anterior
describen también fases más o menos largas de acumulación neta de N. La
fijación atmosférica de N por parte de microorganismos, trascolación y
precipitación, polvo, invasión de hifas fúngicas, excrementos de insectos y
contaminación a part i r de hojarasca recién caída pueden ser algunas de las
principales causas que provocan incrementos absolutos en la masa de N (Lousier
y Parkinson 1978, Fahey 1983, Gosz et al. 1973). Estos factores pueden actuar
preferencialmente en unos lugares o condiciones más que en otras. Gosz et al.
(1973) cita a la precipitación y trascolación como causantes principales de
acumulación durante el invierno, mientras que en primavera son mayormente
atribuidas a factores biológicos. En las coniferas estudiadas podría esperarse
una mayor incidencia de factores biológicos, como excrementos de insectos, ya
que las mayores acumulaciones se producen en la primavera del primer y segundo
año. En la jara, la acumulación se produce de forma progresiva aunque las
mayores tasas se producen durante el primer invierno. Para esta especie quizá
exista una mayor influencia de factores meteorológicos. Durante el invierno
88-89, que coincide con el incremento más fuerte en la acumulación de N en C.
laurifolius, la nieve cubrió durante una buena parte del tiempo la parcela de
estudio. Estas precipitaciones en forma de nieve son más ricas en NH que las
de agua o barro (Domingo 1991).
A la fase de acumulación-inmovilización le sigue, según el modelo de Berg
y Staaf, una fase de progresiva liberación que no se aprecia en P. pinaster,
P. nigra y C. laurifolius del presente trabajo. El comienzo de esta última
etapa parece depender en buena medida del valor C/N. Estos autores consideran
que la mineralización neta de N comienza cuando el valor de C/N de la
hojarasca desciende a niveles similares a los de los heterótrof os
descomponedores. Se ha denominado C/N "crítico" aquel por debajo del cual
comienza la fase de mineralización neta. En el pinar y en la j a ra parece claro
que la no existencia de la fase de mineralización neta puede deberse a que no
se han alcanzado aún los valores críticos al acabar el estudio. El valor
crítico dado por Lutz y Chandler (1946) de 20 /1 - 30/1 ha sido criticado por
bastantes autores. Stohlgren (1988) consideran que éste índice puede ser muy
352
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
variable dependiendo de las especies y de las condiciones. Existen bastantes
trabajos, sobre todo aquellos cuya duración es superior al año en los que se
han observado las fases inmovilización neta-mineralización neta y en los que,
por lo tanto, pueden calcularse los C/N críticos. Los resultados confirman esa
variabilidad para este índice. En Stohlgren (1988) se han calculado C/N
críticos de 45-49/1 para Abies concolor, 67-77/1 para Pinas lambertiana en
Sierra Nevada (California). Se han encontrado también índices críticos C/N de
37-40/1 en Pinus ponderosa (Klemmedson et al. 1985) de Arizona a los 21 meses
de exposición, de 45/1 en Populus tremuloides y 47/1 Populus balsamífera a los
8-12 meses de exposición (Lousier y Parkinson 1978), de 31-34/1 para varias
especies planifolias de Hubbard Brook a los 12 meses (Gosz et al. 1973), de
18/1 a los 200 días en Tilia cordata y de 25/1 a los 360 días en Picea
estudiadas en un bosque polaco (Dziadowiec 1987). Por su parte, Stohlgren
(1988) hace referencia al rango de valores críticos C/N encontrados en
Coniferas que muestran una gran variabilidad: 19-109/1.
Como hemos comentado, t res de las especies estudiadas no han alcanzado
aún el valor crítico para mineralizar de una forma neta el N. Los valores C/N
en el último muestreo (tomando un 507. de C) son de 63.4 para P. pinaster, 103
para P. nigra y 77.2 para C. laurifolius, valores bastante superiores a la
mayoría de C/N críticos encontrados en la bibliografía.
En general, se acepta que altas concentraciones de N inicial juntamente
con elevadas tasas de descomposición influyen para que se alcance antes el
valor crítico C/N (Gosz et al. 1973). En este estudio, ninguna de las t res
especies (C. laurifolius, P. pinaster y P. nigra) poseen valores elevados en
el material de partida (los valores de N están entre el 0.2 -0.3 7.) que
además, son inferiores en general a las encontradas en bibliografía. Por otro
lado estas especies tampoco se caracterizan por poseer unas elevadas tasas de
descomposición. Resulta lógico pensar en estos dos factores como posibles
causas por las que aún no se ha llegado a la etapa de mineralización neta a
pesar de que el tiempo de exposición haya sobrepasado el año y medio. Otros
autores tampoco han detectado mineralización neta de N. O'Conell (1987) para
Eucalyptus diversicolor no observa mineralización neta al final de los dos
años. Lo mismo ocurre para Eucalyptus microcarpa a los 16 meses (Maheswaran y
Attiwill 1987) y para Pinus contorta en Wyoming (Fahey 1983) en donde después
353
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
de dos años sigue sin aparecer ia liberación neta. En este caso, los autores
atribuyen el largo periodo de inmovilización neta a las bajas concentraciones
iniciales de N {0.67.) que por otro lado son superiores a las encontradas en p.
pinaster, P. nigra y C. laurifolius de nuestro trabajo.
Un comportamiento totalmente opuesto ai de P. pinaster, P. nigra y c.
laurifolius es el observado en A. decorticans. En esta especie no se observa
inmovilización neta de N sino un mineralización neta desde los estadios
iniciales de exposición, aunque es probable que el descenso detectado en los
primeros 80 días se deba a lixiviado y no a una mineralización. Al final de
los 626 días que dura el seguimiento de este nutriente, en la hojarasca del
gaznacho sólo permanecía un 48% del N original. Algo muy similar ocurre en
plantaciones de distinta edad de otra especie fijadora, Alnus nepalensis en el
Himalaya (Sharma y Ambasht 1987). En este estudio, el N se pierde de forma
neta inmediatamente después de que las bolsas fueron colocadas en el suelo con
lo que al final de los 24 meses, el N remanente oscilaba entre el 37-54% según
la edad de la plantación. La hojarasca de esta especie de aliso presenta
aumentos en la concentración de N al avanzar el tiempo al igual que en A.
decorticans. Este mismo autor cita patrones parecidos para el N en la
hojarasca de otros Alnus.
Un factor decisivo en esta rápida liberación de N tanto en A. decorticans
como en Alnus nepalensis (Sharma y Ambasht 1987) son las altas concentraciones
de N del material de partida. Las concentraciones de N inicial son de 1.59% en
A. decorticans y de 2.8 % en Alnus nepalensis. Esto implica unos bajos C/N
iniciales que, a la vista de los resultados pueden ser considerados por debajo
del valor crítico. Por lo tanto el N liberado de estas dos especies está en
exceso en relación a la demanda heterótrofa. En leguminosas y otras fijadoras
actinorrizas se encuentran normalmente bajos valores de C/N inicial, pero esto
no implica necesariamente el comienzo de una mineralización neta. Schwintzer
(1984) constata una casi nula mineralización de N en Myrica gale presentando
esta especie, por el contrario, un patrón de inmovilización. Esta planta es
una activa fijadora actinorriza cuya hojarasca posee altos contenidos de N
reflejados por el bajo valor de C/N :28.9 y por ello debería esperarse una
aparición temprana de la fase de mineralización neta. Sin embargo esto no
ocurre así y el autor lo ha relacionado con la existencia en Myrica de unas
354
í
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
tasas de desaparición de materia orgánica mucho más bajas a las de otras
fijadoras, debidas al alto porcentaje de lignina en esta especie. Como
comentamos anteriormente, la velocidad de descomposición de la materia
orgánica es el otro factor determinante para alcanzar en mayor o menor tiempo
el C/N crítico (Gosz et al . 1973). En Myrtca gale, parece claro el efecto de
este último factor que detrmina la dinámica del N, no así el C/N inicial. En
el caso de A. decorticans y Alnus nepalensis (Sharma y Ambasht 1987) las tasas
de descomposición son considerablemente altas ya que A. decorticans pierde al
cabo de 21 meses un 13.87. de su peso seco total mientras que para las
plantaciones de Alnus estos valores oscilan entre el 55-71% de la materia
orgánica a los 24 meses. En estas especies ambos factores resultan, pues,
favorables para la mineralización del N pero, como hemos visto esta pauta no
puede generalizarse a todas las fijadoras de N.
Sea cual sea la dinámica particular de una determinada especie respecto
al N, este elemento es el que se pierde de una manera más lenta en todos los
trabajos consultados. En sistemas mediterráneos, la dinámica de mineralización
de nitrógeno contituye una de los procesos clave en el ciclo de este elemento
(Rapp 1990) con influencia determinante en la nutrición mineral. La
conservación o la lenta liberación de un nutriente tan fundamental en estos
sistemas regula el flujo de transferencia del N desde el suelo a la planta
(Nuñez 1989) a la vez que se minimizan sus pérdidas por lixiviado.
Fósforo.
El P sigue en importancia al N como macronutriente en la nutrición
vegetal (Larcher 1977). La cantidad de P disponible en la solución del suelo
es, sin embargo, baja con lo que la descomposición es de importancia capital
para abastecer de P a esta solución de suelo.
En contraste con el N, que normalmente sigue la dinámica descrita por
Berg y Staaf (1981), el P es un elemento con un comportamiento bastante
variable respecto a su dinámica en el proceso de descomposición. Un ejemplo
claro lo tenemos con las cuatro especies estudiadas. En el pinar t ras una fase
corta de lixiviado, P. pinaster inmoviliza el P mientras que P. nigra lo
355
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
libera. A. decorticans posee un patrón con una primera fase de inmovilización
con incrementos netos en la cantidad de P, seguida de una mineralización neta.
Por su parte, C. laurifolius libera el P de una forma rápida e intensa para
después mantener durante el resto del periodo unos niveles bajos más o menos
constantes. La bibliografía también refleja esta variabilidad. En hojas de
roble Dziadowiec 1987 encuentra un lixiviado inicial seguido de acumulación
neta para posteriormente ser liberado de nuevo siguiendo una dinámica similar
a la de P. nigra del presente trabajo. Schlesinger (1984) destaca periodos de
acumulación neta en los tres años de estudio en hojas de Ceanothus y Salvia
detectando pequeñas periodos de lixiviado inicial. El P en Eucalytus
diversicolor (O'Conell 1987) es retenido en la hojarasca de esta especie en
incluso acumulado hasta niveles superiores en un 607. al contenido inicial, sin
observarse liberación neta al final de los dos años. También se detecta
inmovilización en Eucalytus microcarpa (Maheswaran y Attiwill 1987) aunque no
acumulación ya que los contenidos de P nunca superan a los iniciales. Verdú
(1984) describe inmovilizaciones transitorias de P en acículas de abeto y
hojas de haya no apreciándose mineralización neta al final del primer año. Por
su parte Gosz et ai . (1973) encuentra para Acer saccharum y Fagus grandifolia
lixiviados iniciales de P seguidos de incrementos absolutos en la cantidad de
P y en las concentraciones químicas. Para una especie de abedul, este autor
destaca la acumulación de P desde el comienzo de la exposición. Stohlgren 1988
estudiando distintas coniferas de Sierra Nevada (California) describe una
pronunciada liberación en la sequoia e inmovilizaciones temporales en otras
coniferas.'
Otros autores, (Fahey 1983) en Pinus contorta de Wyoming, Gosz et al.
(1973) en ciertas planifolias y Dziadowiec (1987) en una especie de roble
describen la dinámica del P con una fase inicial de acumulación seguida de
liberación neta en un patrón similar al que se ha encontrado en A.
decorticans.
Por su parte mineralizaciones netas desde el comienzo de los ensayos han
sido detectados para Alnus nepalensis en cuya especie no se detecta ningún
periodo de inmovilización transitoria (Sharma y Ambasht 1987), en Pinus
lambertiana (Stohlgren 1988) y en distintos caducifolios de Polonia
(Dziadowiec 1987) en los que las tasas de pérdida de P son inferiores a las
356
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
del peso seco. Así mismo, para dos especies de Populas Lousier y Parkinson
(1978) encuentran una estrecha correlación positiva entre la pérdida de P y la
del peso seco indicando así que la mineralización neta comienza desde el
principio. Por su parte, las acículas de Pinus ponderosa en bosque de Arizona,
después de una fase de lixiviado inicial pierden P de forma neta y progresiva
a lo largo de los 24 meses de estudio (Klemrnedson et al. 1985).
La dinámica del P al igual que ocurre con el N parece estar en estrecha
consonancia con los índices C/P. Se han calculado valores críticos de C/P para
distintas especies a partir de los cuales comienza la fase de mineralización
neta. Enright y Ogden (1987) calcula un valor crítico de alrededor de 650/1 en
varias especies componentes del bosque de kauri en Nueva Zelanda. Por su parte
Gosz et al. (1973) en el bosque de Hubbard Brook calcula valores críticos de
480/1, Sharma y Ambasht (1987) dan un valor de 637/1 en Alnas nepalensis y
Dziadowiec 1987 de 400/1 en acículas de Pinos sylvestris en un bosque polaco.
El valor crítico calculado para la mineralización de P en A. decorticans es de
609/1, valor dentro del rango de los resultados encontrados por los autores
citados anteriormente.
C. laurifolias, con un valor C/P inicial bajo: (357/1) comienza a liberar
P de forma neta y rápida desde el principio de la exposición. Otros autores
han encontrado pautas similares para especies cuya hojarasca posee, al igual
que la ja ra valores bajos de C/P inicial. Enright y Ogden (1987) estudiando
las especies del bosque de kauri en Nueva Zelanda destaca que aquellas
especies con C/P iniciales de alrededor de 300/1 mineralizan el P de una forma
intensa y rápida. Lousier y Parkinson (1978) encuentran C/P iniciales de 234/1
y 233/1 para dos especies de Populus en un bosque templado frío de Alberta y
una rápida mineralización de P. En todos estos casos a la fase de pérdida
rápida se sucede una fase de meseta en la que el P es inmovilizado. Nos
inclinamos a pensar que los índices iniciales C/P en estas especies están por
debajo del nivel crítico, permitiendo por ello la pérdida rápida de P.
Probablemente sean los valores de C/P alcanzados inmediatamente antes de la
fase de meseta los que estén más cerca de los valores críticos que no los
iniciales. Estos valores serían 657/1 para la ja ra y 400/1 en las especies del
género Pópalas estudiados por Lousier y Parkinson (1978).
357
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Stohlgren (1988) encuentra una estrecha correlación positiva entre la
concentración de P inicial en varias coniferas de un bosque de Sierra Nevada
(California) y la pérdida de peso de este elemento al cabo de los 3.6 años que
dura su estudio. Swift eí al. (1979) también destacan la relación entre la
concentración inicial de P y la tasa de mineralización de este nutriente. En
nuestro trabajo, C. laurifolius con concentraciones de P significativamente
más altas de P que el resto de especies es también la que mayor cantidad de
este elemento pierde a lo largo del proceso.
Schlesinger (1985) encuentra relaciones inversas entre la materia
orgánica remanente y la concentración de P indicando según este autor
retención de este elemento. Tales relaciones se han encontrado también para A.
decorticans y P. pinaster en el presente trabajo. Sin embargo los periodos de
incremento absoluto no pueden explicarse con los aumentos de concentración
sino que han de buscarse en fuentes externas. En A. decorticans y P. pinaster
estos incrementos se producen básicamente en primavera. Gosz eí al. (1973)
hace referencia al papel que en este sentido pueden jugar la contaminación por
polen o flores en los meses de primavera y verano. Los incrementos absolutos
de P durante los meses invernales que se detectan en P. nigra pueden provenir
de los horizontes orgánicos del suelo (Gosz eí al. 1973) ya que tanto las
precipitaciones como la trascolación u escorrentía cortical en el área de
estudio aportan cantidades muy pequeñas de P (Domingo 1991).
A modo de resumen, parece claro que las concentraciones iniciales de P
influyen en la dinámica de éste elemento. La demanda heterótrofa de P es
elevada con lo que cantidades iniciales de P por debajo de esos requerimientos
implicarían una inmovilización. La disminución progresiva del C/P a lo largo
del proceso llevará a la obtención de valores críticos por debajo de los
cuales la mineralización neta es ya posible. En aquellos casos en que la
cantidad de P inicial está por encima de los requerimientos la mineralización
neta puede comenzar. En estos caso el P es liberado rápidamente hasta alcanzar
el nivel crítico para los heterótrofos apareciendo entonces una inmovilización
neta.
358
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
Potasio.
El K se pierde en las especies estudiadas de forma rápida e intensa
durante el primer otoño-invierno para después mantenerse en niveles bajos más
o menos constantes durante el restos del periodo exceptuando en P. nigra para
el que se constata una ligera inmovilización en la última fase. Todos los
trabajos bibliográficos coinciden en este patrón de pérdida rápida seguida de
mantenimiento. Además, el K ocupa el primer lugar (con algunas excepciones) en
la ordenación de los nutrientes según su velocidad de liberación en todas la
series estudiadas. La movilidad de este elemento, que aparece en vegetales de
forma ionizada, facilita el lixiviado y por tanto su liberación (Dziadowiec
1987). Ya que el contenido inicial de K en hojarasca se encuentra por encima
de los requerimientos heterótrofos, éste puede ser liberado y dada su
suceptibilidad al lixiviadora pérdida se produce muy rápidamente (Gosz et al.
1973). Además, la liberación de K por parte de la hojarasca no es la única
fuente importante de K al suelo sino que el lavado de copas lixivia también
considerables cantidades de K (Parker 1983, Domingo 1991 en nuestra zona de
estudio). Por ello el K se convierte en uno de los elemntos menos limitantes
para las poblaciones heterótrofas del suelo.
La existencia de una fase de meseta, tanto en las especies estudiadas
como en los trabajos consultados sugiere que se han alcanzado los valores
críticos C/K para los organismos descomponedores y que por tanto, la
inmovilización excede a la liberación en esta última fase (Gosz et al. 1973,
Lousier y Parkinson 1978, Swift 1979). En esta etapa pueden aparecer incluso
aumentos absolutos en la cantidad de K tal y como lo describen Gosz et al.
(1973), Lousier y Parkinson (1978) y detectado también en P. nigra en el
presente trabajo. Las posibles causas se han buscado en un incremento en la
población o actividad de los heterótrofos, tracolación o a incrementos en las
tasas de intercambio de K en el material que se descompone (Gosz et al. 1973).
La disminución en las concentraciones de K a lo largo del periodo es otra
característica común en ías especies estudiadas y en los trabajos que comentan
este aspecto (Lousier y Parkinson 1978, Verdú 1984, Gosz et al. 1973). Si
bien, las concentraciones químicas de partida pueden ser bien diferentes para
distintas hojarascas en un mismo sitio, parece existir una tendencia a que los
359
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

U.J. Moro Cuadrillero
valores de concentración mínimos alcanzados sean similares (Gosz et al. 1973).
En las especies arbustivas estudiadas resulta interesante el que A.
decorticans que posee concentraciones iniciales de K significativamente más
altas que C. lawifolius (0.68% frente a 0.337.) alcance un valor de
concentración mínimo idéntico al de la j a ra durante el proceso de
descomposición (0.068%). Las concentraciones de K mínimas alcanzadas por las
especies de pino son diferentes a las del matorral pero muy similares entre
ellas (0.026 y 0.035%) aunque en este caso hay que tener en cuenta que los
pinos no difieren significativamente respecto a la concentración inicial de K.
Gosz et al. (1973) encuentra que para concentraciones iniciales de K de 0.43%
y 0.31 en Betula algheniensis y Fagus grandifolia se alcanzan valores mínimos
similares (0.13% y 0.07%) respectivamente. Este autor comenta que esas
concentraciones mínimas deben representar la cantidad de K que es fijada o
inmovilizada. La misma tendencia ha sido descrita en Lousier y Parkinson
(1978) para dos especies de Populus en Alberta y por Enright y Ogden (1987) en
distintas especies del bosque de kauri.
La dinámica del K en nuestras especies se puede ajustar de manera
significativa a un modelo exponencial simple manteniéndose la curva siempre
por debajo de la del peso seco. En P. nigra, debido a la acumulación final de
K, el modelo, si bien significativo no se ajusta bien a la dinámica real del
proceso. Bockheim y Leide (1986) utiliza también este tipo de modelo. La
constante de descomposición del K calculada por este autor para una plantación
de Pinus resinosa es de -1.39 al cabo de un año de descomposición. Este valor
resulta superior a la encontrada para P. pinaster (-0.68) y C. laurifolius
(0.75) y bastante similar a la de A. decorticans (-1.69). Los valores tan
elevados del trabajo de Bockheim y Leide (1986) probablemente vengan causados
por la menor duración del ensayo (1 año) frente al nuestro (568-626 días).
Calcio.
Varios autores citados en Lousier y Parkinson (1978) han indicado que el
Ca es menos lixiviable que el K o Mg probablemente debido al papel del Ca como
componente estructural de las paredes celulares (Attwill (1968) citado por
Fahey (1983) y la menor solubilidad de las sales de Ca de las células (Lousier
360
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
y Parkinson 1978). Dado ese carácter estructural, la descomposición es
mayoritariamente responsable de la pérdida de Ca. Es por tanto esperable una
dinámica similar a la del peso seco (Lousier y Parkinson 1978).
La figura 5.18c muestra una estrecha relación entre pérdida de peso y de
Ca en C. laurifolius (exceptuando durante un corto periodo) y sobre todo en A.
decorticans (fig 5.18d). Para los pinos, sólo las primeras fases pueden
asemejarse a las correspondientes de la materia orgánica. Después, las curvas
de Ca en los dos pinos quedan siempre por encima de las del peso seco
indicando inmovilización de este elemento, [poner correlación]
Varios autores han comprobado la similitud entre la dinámica Ca y la del
peso seco (Schlesinger 1985, Gosz et al. 1973, Enright y Ogden 1987). Sin
embargo muchos otros han descrito inmovilizaciones y acumulaciones de Ca a
veces muy intensas de manera que las tasas de pérdida del nutriente quedan muy
por debajo de las de la materia orgánica. Así, son abundantes los trabajos que
citan inmovilizaciones y acumulaciones en especies de pino (Stohlgren 1988,
Cromack y Monk 1975, Klemmedson et al. 1985 y Fahey 1983) así como también en
planifolios caducos (Lousier y Parkinson 1978). En este estudio ambas especies
de pino siguen esta pauta en la que se aprecian incrementos absolutos de Ca en
relación al contenido inicial más patentes en P. pinaster (tabla 5.15 c y d y
figs 5.18a y b).
La concentración de Ca aumenta paulatinamente en el pinar aunque con
alguna oscilaciones. Esta característica ha sido observada por otros autores
(Verdú 1984, Lousier y Parkinson 1978, Klemmedson et al. 1985). Sin embargo
las concentraciones de Ca en el matorral se mantienen muy próximas al valor de
concentración inicial durante la mayor parte del periodo.
El Ca contenido en la hojarasca es superior a la demanda de la fauna del
suelo (Reichle 1969). Según Gosz et al. (1973), las concentraciones de Ca
iniciales (y por tanto los C/Ca) no parecen tan determinantes en la dinámica
de este elemento frente a lo que ocurre por ejemplo con N o P. Parece que el
tipo de hoja y la velocidad de descomposición de la materia orgánica resultan
más importantes en este aspecto. Enright y Ogden (1987) observa una mayores
tasas de liberación de Ca en las especies mesófilas frente a las esclerófilas.
361
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Dziadowiec (1987) encuentra para acículas de Pinus sylvestris altas tasas de
descomposición (60% de peso perdido en un año) y una liberación neta de Ca
desde el comienzo del ensayo a pesar de la esclerofilia de esta especie y de
sus bajos contenidos de Ca en su hojarasca. En nuestro estudio la hojarasaca
de A. decorticans presenta las mayores tasas de liberación de Ca a pesar que
sus contenidos de Ca no son los más elevados. Sin embargo las hojas del
gaznacho son las más mesófilas y las que poseen mayor tasa de descomposición.
La j a r a que presenta contenidos significativamente más altos de Ca pero
velocidades más bajas de descomposición que A. decorticans pierde éste muy
lentamente. Parece confirmarse que estos dos factores son los que determinan
en estos casos la mayor o menor liberación de Ca. Sin embargo en la
bibliografía aparecen casos en los que se considera a la concentración de Ca
inicial como el factor fundamental. Dziadowiec 1987 cita como posible causa de
la rápida liberación de Ca en hojas de tilo sus altos contenidos iniciales de
Ca. Probablemente todos estos factores puedan influir así como también las
relaciones del Ca con otros elementos por ejemplo N/Ca (Dziadowiec 1987).
Los aumentos en las concentraciones de Ca que también aparecen en el
pinar de estudio, han sido explicadas en algunos casos por la ingestión
preferencial por parte de invertebrados comedores de hojas de aquellos tejidos
más pobres en Ca (Lousier y Parkinson 1978). Sin embargo estos aumentos de
concentración no explican el incremento absoluto en el peso del Ca. Las
inmovilizaciones de Ca encontrados en P. pinaster y P. nigra y en otros muchos
trabajos pueden no estar relacionados con la demanda metabólica por
heterótrofos tanto hongos (Fahey 1983) como fauna del suelo (Gosz ' et al. 1973)
ya que según estos autores el Ca no es usualmente limitante para su
desarrollo. Sin embargo otros autores (Cromack y Monk 1975, Klemmedson et al.
1985) sí han comentado la evidencia de que ciertos microorganismos pueden
concentrar considerables cantidades de Ca quizá suficientes para producir
inmovilización de este nutriente en la hojarasca. Cromack y Monk 1975 cita un
trabajo de Foster (1949) en el que se comenta que puede existir una demanda de
Ca y de otros cationes para formar oxalatos a partir del ácido oxálico
producido en el metabolismo de ciertos microorganismos fúngicos. Klemmedson et
al. (1985) destaca que pequeñas invasiones de estos hongos pueden contribuir a
los incrementos en la cantidad de Ca observados en la hojarasca de Pinus
ponderosa. Nuestro trabajo y la mayoría de la bibliografía que estudia la
362
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Deicomposlclón.
beración del Ca y otros elementos en la hojarasca carece casi por completo
i información referente a los descomponedores por tanto todas estas
cplicaciones no pasan de ser meras especulaciones que deberían ser
improbadas experimentalmente. Fuentes externas como precipitación o
•ascolación, que en nuestra área de estudio son especialmente ricas en Ca
)omingo 1991), pueden participar en los incrementos en la cantidad de Ca
jservados en nuestras especies. Además, se ha constatado (Thomas 1969 citado
Dr Lousier y Parkinson 1978) que la mayor parte del Ca lixiviado por las
jpas permanece retenido en los horizontes orgánicos H y L sin pasar al suelo
ineral.
agnesio.
La mayoría de los autores sitúan a este elemento junto con el K dada la
militud que presentan en su dinámica de descomposición (fase de liberación
ípida seguida de mantenimiento). El Mg es un elemento lixibiable pero no
orto como el K con lo que se libera más lentamente que aquel. Los resultados
:bliográficos coinciden en que la liberación del Mg es más lenta y
sneralmente más paulatina que la del K (Dziadowiec 1987, Fahey 1983, Gosz eí
'. (1973), Stohlgren 1988). Estos resultados coinciden también con el patrón
icontrado para el Mg en las especies estudiadas.
Los dinámica de liberación de Mg es en algunos casos más rápida que la
ú peso seco o materia orgánica (Gosz eí ai. (1973), Maheswaran y Attiwill
)87, Cromack y Monk 1975, presente trabajo). Estos resultados difieren con
is encontrados por otros muchos autores en los que la dinámica del Mg se
próxima mucho más a la de la materia seca o a la de otros elementos menos
oviles como Ca y P. (Enright y Ogden 1987, Lousier y Parkinson 1978,
ziadowiec 1987, Schlesinger (1985), O'Conell (1987).
Al igual que con el K, Gosz eí ai. (1973) destaca que las concentraciones
ínimas de Mg alcanzadas en la hojarasca pueden corresponder a la demanda de
>s heterótrofos calculando con ellas el C/Mg entre 900/1 y 1350/1. Destaca
imbién que las concentraciones mínimas son similares en dos especies que
enen contenidos de Mg de partida muy diferentes. Para las especies
363
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
estudiadas las concentraciones mínimas fueron 0.15 para los pinos, 0.20 en la
jara y 0.16 en el gaznacho que darían valores críticos C/Mg de 333, 250 y
312,5 repectivamente, valores muy por debajo de las encontardas por Gosz et
al. (1973). En todas las especies, excepto en la ja ra estas concentraciones
mínimas corresponden al último muestreo con lo que es probable que esta
concentración pueda seguir disminuyendo por algún tiempo y por tanto el valor
crítico se alcanzaría para cocientes C/Mg más elevados.
Dada la dinámica del Mg en las especies estudiadas, es posible aplicar el
modelo exponencial simple. La constante de descomposición encontrada para el
Mg por Bockheim y Leide (1986) en Pinus resinosa es de - 0.45 muy similar a la
de la j a ra (-0.51), ligeramente más baja que las de las especies de pino
estudiadas (-0.68, -0.69) y muy inferior a la de A. decorticans (-0.957).
Sodio.
El papel de este elemento en la descomposición resulta bastante
desconocido y para la mayor parte de las especies no parece esencial (Gosz et
al. 1973). Quizá sea ésta una de la razones por la que son pocos los trabajos
que hacen referencia a él. Además la precisión en la detección analítica de la
muestras es menor que para otros elementos dadas sus bajas concentraciones.
Esto t rae como consecuencia una mayores errores estándar y una menor
fiabilidad de los resultados. En este trabajo y en otros (Klemmedson et al.
1985) las oscilaciones, tanto en la concentración de Na como en sus cantidades
absolutas son más marcadas que para otros elementos. Estos resultados pueden
estar perfectamente influenciados por esa variabilidad analítica más que por
la variación espacial -temporal debida a la descomposición con lo que las
posibles causas biológicas de estas oscilaciones han de tomarse con las
debidas precauciones.
El patrón de liberación de este elemento es variable en las especies
estudiadas así como en los trabajos bibliográficos. Al final del periodo de
exposición, P. pinaster y A. decorticans pierden cantidades considerables de
Na (707. en P. pinaster y 74 7. en A. decorticans), C. laurifolius pierde
cantidades más moderadas (527.) mientras que en P. nigra esta cantidad es mu
364
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Descomposición.
pequeña (57.). Algunos autores encuentran una liberación rápida de Na sólo
superada por el K y a veces por el Mg atribuible a la facilidad de lixiviado
del Na. Así, Enright y Ogden (1987) encuentra este patrón para distintas
especies del bosque de kauri, Lousier y Parkinson (1978) en dos especies de
Populus, Verdú (1984) en una especie de abeto y Maheswaran y Attiwill (1987) y
O'Conell (1987) para dos especies de Eucalyptus en Australia. En este trabajo
el Na se libera de forma neta en A. decorticans y P. pinaster si bien existen
interrupciones durante las cuales aparecen aumentos en la cantidad de Na, que
en todo caso no superan al contenido inicial. El Na ocupa el tercer lugar
(después del K y Mg en esta dos especies. Otros autores atribuyen al Na una
menor movilidad (Guha y Mitchell (1966) citado en Gosz et al. (1973). Hay
autores que encuentran periodos de acumulación neta por encima del contenido
inicial de Na (Gosz et al. 1973, Lousier y Parkinson 1978) tal y como acontece
en C. lawifolius y P. nigra en este trabajo.
En algunos casos parece existir una relación entre la concentración
inicial de Na y su contenido al final del estudio. Esta relación queda
reflejada en la correlación significativa entre la concentración de Na inicial
en las cuatro especies y la masa remanente al final del estudio. La misma
relación encuentra Enright y Ogden (1987) para especies componentes del bosque
de kauri en Nueva Zelanda.
C. laurifolius y P. nigra presentan periodos de acumulación de Na con
valores superiores a los del contenido inicial. Esta fase es transitoria en C.
laurifolius mientras que en P. nigra la acumulación se mantiene prácticamente
a lo largo de todo el periodo alcanzando valores que pueden llegar a ser un
1507. más altos que el contenido inicial. Se han descrito posibles causas para
esta acumulación neta de Na entre ellas la tracolación y/o precipitación (Gosz
et al. 1973, Lousier y Parkinson 1978) y la invasión de microfauna rica en Na
(Cromack et al. en Lousier & Parkinson 1978).
Orden de liberación.
Un buen número de estudios han ordenado los distintos nutrientes de
acuerdo a su orden de liberación en la hojarasca durante la descomposición. En
365
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 5.18. Comparación de las series de movilidad de los nutrientes estudiados para diferentes tipos de hojarasca.
ESPECIE LOCALIDAD ORDEN ANOS AUTOR
Pinas resinosa
Quercus robar
C'arpiñas beta las
Tilia cordata
Pinas sylvestris
Pinas sylvestris
Ilex aqaifoliam
Eacalytas diversicolor
Eacalyptas microcarpa
Qaercas ilex
Fagas sylvatica
Abies
Ceanhotus, Salvia
Varias
Pinas pinaster
Pinas nigra
Adenocarpas decorticans
Cistas laarifolias
U.S.A.
Polonia
i i
•i
i i
P i r i n e o s
•i
A u s t r a l i a
A u s t r a l i a
La Castanya
"
u
C a l i f o r n i a
Nueva Zelanda
Almería
rt
• i
• i
K>Mg>P>N>Ca
K>Mg>P>Ca>N
K>Mg>P>Ca>N
K>Mg>Ca>P>N
K>Mg>P>N>Ca
Mg>K>Ca>N>P
K>Mg>Ca>N>P
Na>K>Mg>Ca>N>P
K>Na>Mg>P>N>Ca
K>Na>Mg>P>Ca>N
Na>K>Mg>Ca>N>P
Na-K>Mg>P>N>Ca
K>Mg>Ca>P>N
K-Na>Ca-Mg>P
K>Mg>Na>P-Ca>N
K>Mg>P>Na>Ca>N
K>Mg>Na>Ca>P>N
K>P>Mg>Na>Ca>N
1
2
i i
i i
i l
1
•i
2
2
1
t i
II
M
"
1.
"
II
II
J
2
)
Bockheim y
Lei de 1986
- Dz iadowlec 19£
• Alvera 1990
O'Connell 1987
Maheswaran y
A t t i w i l l 1987
1
j
• Verdú 1984
S c h l e s i n g e r 19t
Enr ight y Ogdei
1987
^
• P r e s e n t e
t r a b a j o
la tabla 5.18 se han recogido algunas de estas series a partir de la
bibliografía consultada. Además en Lousier y Parkinson 1978 y Alvera (1990)
puede encontrarse también una buena recopilación de estas series de
366
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

liberación.
Descompoiiclón.
El K es el elemento más fácilmente liberado en todas las series excepto
en algún caso en que este lugar es ocupado por Na o Mg. El Mg suele ocupar la
segunda plaza mientras que C a y N o P y N s e sitúan en las últimas posiciones.
Todos estos resultados concuerdan con los encontrados en el presente trabajo.
En raras ocasiones el P ocupa las primeras plazas tal y como ocurre en C.
laurifolius de este estudio. No obstante se ha encontrado el P en segundo
lugar en algunos trabajos citados por ALvera (1990) como los de Edmonds (1980)
para Pseu.dotsu.ga menzienssi y el de Staff (1980) en haya.
367
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
368
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 6.
Fijación de Nitrógeno en Adenocarpus decorticans.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
6.1. INTRODUCCIÓN.
El gaznacho (Adenocarpus decorticans Boiss) es un matorral perenne nativo
de la región Ibero-Mauritana. La subespecie decorticans se encuentra solamente
en las montañas Béticas del sur de España (Rivas-Martínez y Belmonte 1989).
Fitosociológicamente, A. decorticans decorticans ha sido incluida en la serie
meso-supramediterránea filábrido-nevadense silicícola de la encina y se ha
considerado como un componente principal de la asociación climácica
Adenocarpus-quercetum-rotundifoliae (Losa et a l . 1986). Debido a la
degradación del bosque natural de encinas en gran parte de su área de
distribución, esta especie se encuentra frecuentemente en el sotobosque de las
plantaciones de pinos, acompañado por Cistus laurifolius.
La presencia cuantitativa importante de A. decorticans en el área de
estudio hizo que nos planteásemos abordar el estudio de su posible capacidad
de fijación de nitrógeno y en caso afirmativo incluir la cuantificación de
este flujo de entrada dentro del programa que estábamos llevando a cabo. Para
ello iniciamos una búsqueda bibliográfica y nos pusimos en contacto con el
profesor Bermudez de Castro de la UC de Madrid.
Las citas bibliográficas sobre A. decorticans y en general de todo el
género Adenocarpus son escasísimas y casi ninguna de ellas hace referencia a
la fijación (Moro et al . 1989). Solamente en la recopilación de Alien & Alien
(1981) encontramos una cita antigua de Dawson (1900) en la que se indicaba la
existencia de nodulos en esta especie, cuestión que, por otra parte ya
habíamos tenido ocasión de observar en el campo. En ningún momento se
comentaba la posible fijación efectiva de estos nodulos, por lo que decidimos
comprobarlo nosotros mismos.
A primeros de Junio de 1988 se tomaron nodulos de individuos escogidos al
azar en una de las zonas de la cuenca y se comprobó la fijación efectiva de
esta especie mediante la técnica de la reducción de acetileno (Moro et al.
1989) aunque todo parecía indicar que las cantidades fijadas eran bastante
pequeñas. A pesar de ello no animamos a continuar con el seguimiento de la
fijación por lo menos durante un año completo. La técnica utilizada (ARA) no
era la más adecuada para cuantificar las entradas de nitrógeno por la vía de
369
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
fijación debido a una serie de limitaciones que ya comentaremos en el apartado
de métodos pero resulta muy útil para estimar la variabilidad estacional y
diaria. La inaccesibilidad a otro tipo de técnicas como la del N nos hizo
decidirnos por la técnica del ARA, por otra parte muy empleada por lo grupos
de trabajo que se dedican a este tema.
Aunque existen alrededor de 18000 especies de leguminosas (Alien y Alien
1981), se han investigado principalmente las formas herbáceas cultivadas, a
pesar de que el aporte de cantidades adicionales de nitrógeno debido a las
leguminosas leñosas puede ser relevante para los ciclos de nitrógeno de los
ecosistemas. Además, los estudios sobre patrones estacionales de ARA se han
centrado casi exclusivamente en regiones templadas (Wheeler et al. 1981,
Pizelle 1984, Boring y Swank 1984, Wheeler et al. 1987) o boreales (Shulman et
al. 1988), en los que las condiciones hídricas y de temperatura son bastante
diferentes a las de los ecosistemas mediterráneos (Nahal 1981). Incluso, los
pocos estudios realizados en la región mediterránea española se han centrado
es especies riparias, como Alnus glutinosa (Bermúdez de Castro et al. 1976,
Schmitz y Bermúdez de Castro 1982, Pérez Hernández et al. 1989), donde la
disponibilidad de agua es obviamente mayor que en hábitats no riparios. Por
tanto, falta información sobre los patrones temporales de fijación de
nitrógeno en áreas mediterráneas semi-áridas.
En este capítulo emprendemos el estudio de los patrones estacionales de
actividad reductora del acetileno (ARA) en una leguminosa mediterránea que
habita en un área montañosa en la que la precipitación anual media es inferior
a los 500 mm. También se ha estudiado el patrón diario de ARA en dos momentos
extremos del ciclo anual en cuanto a disponibilidad hídrica: la sequía estival
y las lluvias otoñales. Por último se hace un intento de cuantificar la
cantidad de nitrógeno fijado anualmente por parte de esta especie, aunque como
ya comentaremos los márgenes de error de los resultados son muy amplios.
6.2. MATERIAL Y MÉTODOS.
En la cuenca Nacimiento se escogieron dos parcelas representativas, una
en la parte baja de la cuenca, a 1540 m.s.n.m., y otra en la parte alta, a
370
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
1740 m.s.n.m. El muestreo se limitó a individuos jóvenes no-reproductores, con
un diámetro basal menor que 4 cm, ya que los nodulos de individuos mayores son
difíciles de obtener en cantidades adecuadas, debido a sus largas raíces y a
una escasa nodulación. Además, la perturbación que conllevaría este tipo de
muéstreos podía ser considerable. Se estableció un límite de 20 cm para el
muestreo de nodulos, ya que éstos se encuentran en mayor densidad por encima
de esos niveles.
Los nodulos se colectaron mensualente en ambas parcelas desde final de
Octubre de 1988 hasta finales de Septiembre de 1989, excepto en Noviembre de
1988, cuando las nevadas impedían el acceso a la parcela superior. En Julio y
a principios de Octubre de 1989, se realizaron dos muéstreos a lo largo de un
día con el fin de determinar los patrones diarios de actividad de la
nitrogenasa. Las incubaciones de los nodulos se realizaron al mediodía en el
muestreo estacional, y a las 1, 4, 7, 10, 13, 16, 19 y 22 horas en los
muéstreos diarios. Aunque las pruebas con nodulos unidos a la ra íz son
preferibles cuando sea posible, la distribución irregular de los nodulos en
las raíces hacen que esta aproximación difícilmente sea viable en estudios en
el campo.
Los individuos fueron escogidos aleatoriamente en cada parcela. Los
nodulos presentes en cada individuo eran escindidos de la raíz y lavados con
agua desionizada. Para evitar la pérdida de actividad de la nitrogenasa, las
pruebas de reducción de acetileno se iniciaban rápidamente, en los 15-30
minutos siguientes a la escisión de los nodulos de la raíz. La incubación de
las muestras fue realizada en una atmósfera de aire con un 107. de acetileno,
utilizando viales de vidrio de 12 mi con tapón de goma que se enterraban cerca
de la superficie del suelo. Después de una hora (Hogberg y Kvarnstrom 1982,
Pérez et al. 1989, Silvester et al. 1985, Sharma 1988, Wheeler et ai . 1981, la
actividad era inhibida utilizando H SO 0.5 N. En cada parcela se realizaron 5 2 4
replicados de muestras con nodulos y un frasco con raíces como control.
También se registró el número de nodulos por m , la temperatura del suelo a 10
cm y la temperatura del aire. Los viales eran sellados con silicona y
transportados al laboratorio.
371
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La actividad de la nitrogenasa fue estimada con el método de reducciói
del acetileno (ARA) (Hardy et al. 1973). Las muestras del gas fueroi
analizadas con un cromatógrafo de gases KONIC KNF-2000 equipado con un<
columna PORAPAK R que proporciona un bajo tiempo de retención y una alt«
resolución de los picos (Müller y Bermúdez de Castro 1987). El ARA es ur
método indirecto de evaluar la actividad nitrogenásica, sensible,
reproducible y rápido, pero que no proporciona valores absolutos de
fijación de nitrógeno, sino que indica la velocidad de fijación en un
momento dado y en una condiciones determinadas. Se puede decir que no
proporciona la cantidad real de nitrógeno fijado, sino la actividad potencial
de la simbiosis para fijar nitrógeno atmosférico. Sin embargo es un meétodo
adecuado para comparar actividades entre lugares, tiempos, condiciones
ambientales o especies, siempre que se mida en similares condiciones de
incubación. Uno de los problemas principales del método ARA es la variación
del factor de conversión teórico C H : N , por lo que debe tenerse cuidado si 2 2 2
los resultados del ARA se emplean para estimar las cantidades de nitrógeno
fijadas. Además, se conoce que el método analítico está sometido a diversos
tipos de errores y complicaciones (Hansen et al. 1987a, Herridge 1982, Minchin
et al. 1983). De todas maneras, el ARA es una técnica sensible y relativamente
sencilla (Hardy et al. 1973) que permite obtener una medida satisfactoria de
la actividad de la nitrogenasa y que es ampliamente empleada en estudios
comparativos.
6.3. RESULTADOS.
6.3.1. Nodulación.
Los nodulos de A.decortlcans son por lo general aparentes, de color
pálido y alcanzan una longitud de 1 cm en su madurez. Se encuentran aislados o
agrupados en pequeños racimos con pocos lóbulos nodulares. Su distribución en
el suelo es irregular, encontrándose habitualmente entre 10 y 15 cm de
profundidad, con mayor frecuencia en las raíces finas laterales (Moro et al.
1992). Las primeras lluvias y la suavización de las temperaturas en Septiembre
t r a s la sequía estival, parecen favorecer la explosión de nuevos nodulos,
puesto que la densidad de éstos con tamaño comprendido entre 1 y 5 mm alcanza
372
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación d« nitrógeno.
valores máximos entre Septiembre y Octubre, a la vez que se detectan ya
algunos pequeños racimos. Los nodulos aislados, mayores que los de los meses
anteriores, se encuentran a partir de Diciembre, a c o m p a ñ a d o s por pequeños
racimos. De Marzo a Junio se observa un aumento en el tamaño y densidad tanto
en racimos como en nodulos aislados sin que ninguno de los dos tipos presente
signos de senescencia. Durante estos meses es cuando se obtienen también los
valores más altos del ARA (Moro eí al. 1992). A finales de Junio, comienza un
período estival caracterizado por la escasez de lluvias y las altas
temperaturas que se extiende hasta Agosto. Este estrés ambiental juega un
importante papel en la senescencia de los nodulos manifestada por la
deshidratación de éstos y las bajas tasas de fijación encontradas. Esta
senescencia llega a afectar a un 907. del total de nodulos muestreados en
Julio. La nodulación media anual fue pobre en ambas parcelas de estudio. En la
parcela de 1540 m la nodulación fue de 0.088 ±0.022 g peso seco de nodulo por 2
m , mientras que en la parcela de 1740 m fue de 0.12 ±0.05 g peso seco de 2
nodulo por m . Los valores máximos se dectaron en Junio en ambas parcelas 2
(parcela 1540 m: 0.17 g p.s. de nódulo/m , parcela 1740 m: 0.23 g p.s. de 2
nódulo/m ).
6.3.2. Patrón estacional.
La tabla 6.1 y la figura 6.1 muestran los resultados de la actividad de
la nitrogenasa (nmol C H / g p.s. de nodulo-hora) desde Octubre de 1988 hasta 2 4
Septiembre de 1989 en la parcela de 1540 m, y hasta Octubre de 1989 en la
parcela 1740 m. También se muestran la temperatura de,l suelo y del aire, así
como la precipitación mensual. El test t de muestras emparejadas muestra que
no existen diferencias significativas entre parcelas en el patrón anual de
actividad de la nitrogenasa. Además, la existencia de una correlación
significativa entre las parcelas indica un similar patrón estacional (r= 0.81,
p<0.05). Tampoco, las medias mensuales del ARA son estadísticamente diferentes
entre parcelas en ningún mes, aunque en Octubre la diferencia es casi
significativa (F = 4.35, p= 0.07), no alcanzándose la significación debido
al elevado error estándard en la parcela de menor altitud.
Aplicando el análisis de varianza se comprobó que los valores mensuales
373
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Tabla 6.1. Medias mensuales de actividad HB I O „•* , actividad de la mtrogenasa (nmol CU/ g. p . s<
2 4 nodulo-hora), error «standard (SE), temperatura del suelo y del aire CC) v
nodulaci6n ( g . p . s . n ó d u l o / ^ e n l a s d o s ^ ^ ^ ^ ^ ^ ^ J
precipitaren proceden de la estación pluviometría localizada e n la m i s m a
0 N D J F M A M J J A S
0 N D J F M A M J J A S 0
ARA
1089.82 731.62 52.19 0 0
167.89 1395.79 1316.23 1797.97 188.14 (-)
1766.62
ARA
96.86 (-) 49.83 0 0
170.74 1792.99 2497.87 1973.17 128.52 (-)
1193.62 2187.90
Parcela 1540
SE
541.52 138.10 10.02 0 0 44.98 930.30 736.48 413.10 18.14 (-)
441.40
Temp. suelo
13 2.8 1.2 1 0 5.6 6.5 7.4 9.5 17.5 18 14
Parcela 1740
SE
21.62 (-) 4.43 0 0 51.73 745.76 655.62 608.57 38.23 (-)
200.62 446.37
Temp. suelo
11.4 1 2 0 0 3 4.7 11 9 17 16 12.7 11.8
m.
Temp. aire
16 1 7 5 .5
12 16.5 15.4 13 25 26 17.4
m.
Temp. aire
15.5 0 5 0 0 8 15 12.9 13 27 26 15.8 15.1
Precipit. 1/m2
66.50 107.75 7.5
155.65 69.23 98.17 31.86 67.48 23.53 0 0
259.08
Precipit. 1/m2
66.50 107.75 7.50
155.65 69.23 98.17 31.86 67.44 23.53 0 0
259.08 122.44
Nodulos^ g P-s./m"
.041
.072
.210 (-) (-) .055 .066 .023 .174 .132 (-) .020
Nodulos^ g P-s./m
.035 (-) .131 (-) (-) .137 .169 .107 .228 .118 (-) .093 .095
374
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
medios del ARA son heterogéneos dentro de cada parcela (parcela 1540 m:F = 10,31
2.44, p<0.05; parcela 1740 m: F = 5.44, p<0.001). Por tanto, se aplicó el 10,36
test a posteriori LSD de Fisher, que mostró que los meses se podían asociar
en dos grupos con diferentes actividades. En la parcela de 1540 m los meses de
invierno (Diciembre, Enero, Febrero) junto con Julio fueron meses de baja
actividad. Marzo, el mes de transición entre invierno y primavera, también se
incluye en este grupo. Por otro lado se agrupaban los meses con alta actividad
de la nitrogenasa (Septiembre y Junio). No se encontraron diferencias
significativas entre estos grupos y los meses restantes. En la parcela de 1740
m todos los meses, con la excepción de Septiembre, se distribuyeron en dos
grupos significativamente diferentes: uno con bajas tasas de fijación que
incluye los meses invernales junto con Julio, Marzo y Octubre, y otro con
altas tasas de fijación que incluye los meses de primavera (Abril, Mayo y
Junio). Septiembre se sitúa en una posición intermedia y solo se diferencia
del mes de Mayo, que es el de mayor actividad anual.
La condición de planta perenne de A.decorticans permite suponer un
suministro continuo de fotosintato a los nodulos a lo largo de todo el año.
Sin embargo, no se detectó actividad de la nitrogenasa en Enero ni Febrero en
ninguna de las parcelas (figura 6.1, tabla 6.1). Las bajas temperaturas del
suelo consecuencia de las heladas y de las nieves que se produjeron durante
buena parte del invierno, podrían haber inhibido la actividad enzimática. A
ello hay que añadir el "sepultamiento" bajo la nieve durante un largo período
de tiempo y los daños mecánicos que ésta pudo haber provado en las partes
verdes de los individuos. Una combinación de estos factores podría explicar la
supresión del ARA.
En la primavera, el progresivo incremento del ARA coincide con la
elevación de la temperatura del aire y del suelo, la emergencia y crecimiento
de nuevos brotes foliares, y los últimos estados de crecimiento y maduración
nodular. Desde Abril hasta Junio los rangos de actividad fueron de entre 1316
y 1797 nmol C H / g p.s. nodulo'hora en la parcela a 1540 m, y de entre 1792 y
2497.8 nmol C H / g p.s. nodulo-hora en la parcela a 1740 m, alcanzándose en
ambas parcelas los valores máximos en el mes de Junio. No se detectan
en ningún momento descensos en el ARA atribuíbles a la demanda de
375
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
ARA 10000
1000
100
1740 m plot Temp.(aC)
Oct Nov Dec Jan FeD Mar Apr May Jun Jul Aug Sep
months 250
200
150
100
50
Precipitaron (mm)
n Oct Nov Oec Jan Feb Mar Apr May Jun Jul Aug Sep
ARA 10000
1000
100
1540 m plot Temp.CC)
Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep
months — nmol C2H4/g aw nod.h ~*~Soil Temp. —i—Air Temp.
Figura 6.1. Variación estacional de la actividad de la nitrogenasa en nodulos
de A. decorticans (± S.E.) y de la temperatura del aire y del suelo, en las
dos parcelas de estudio. Se muestra también la precipitación mensual.
376
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
Fijación de nitrógeno.
9.23 8.14 1.06 14.14 29.12 1.30 8.98 9.12
3 3 3 2 2 2 3 2
Tabla 6 . 2 . Var iac ión d i a r i a de l a a c t i v i d a d r educ to ra de l Ace t i l eno (ARAMnmol
CH/ g p . s . nodu lo•hora ) , en nodulos de A. decorticans.
J u l i o 1989
Hora ARA SE n
1 113.04 4 65.79 7 41.04
10 69.08 13 199.90 16 116.67 19 70.12 22 208.55
Octubre 1989
Hora ARA SE n
1 749.48 4 733.24 7 947.30
10 2470.88 13 1975.33 16 1676.12 19 590.48 22 510.25
fotosintato por parte de flores o frutos en formación dado que los
individuos que se muestrean no son aún reproductivamente activos.
El ARA disminuyó bruscamente en Julio, en el que los niveles de actividad
obtenidos están comprendidos entre los 100 y 2000 nmol C H / g p.s. 2 4
nodulo-hora, es decir, alrededor del 107. de la actividad en el mes anterior.
Durante este período alrededor de un 907. de los nodulos se encuentran en
estado senescente.
6.3.3. Patrón diario.
La tabla 6.2 y la figura 6.2 muestran los resultados del ARA diario para
los nodulos de A. decorticans en Julio y Octubre de 1989. El tes t de la t
mostró que la media diaria de Octubre es significativamente más alta que en
Julio (t= 4.35, p<0.001, g.l.= 47). Además el patrón diario del ARA en ambos
639.30 509.48 160.20 853.32 545.79 526.00 236.13 294.59
4 4 3 3 4 3 4 4
377
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
July 1989 250'
A A A
200
150
100
TemD.CC)
7 10 13 16 19 22
time of óay
October 1989.
7 10 13 16
time of day nM C2H4/gr(dw).h+ SE - * - S o i l Temp.
Temp.CC)
• Air Temp.
Figura 6.2. Patrón diario de actividad de la nitrogenasa (± SE) en nodulos de
A. decorticans en la parcela de 1740 m, en Julio y Octubre de 1989. Se muestra
también el fotoperíodo en la parte superior de las gráficas.
378
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nltrófeno.
meses fue diferente, ya que su correlación no resultó significativa (r=0.018).
En Octubre, el patrón diario de ARA mostró diferencias significativas entre
los períodos de luz y oscuridad (t= 3.19, g.l.= 27, p<0.01), de forma que la
actividad de la nitrogenasa fue mayor durante el día (1783.61 nmol C H / g
p.s. nodulo-hora) que durante la noche (645 nmol C H / g p.s. nódulo-hora). 2 4
Sin embargo, en Julio no se encontraron diferencias significativas entre ambos
períodos (t= 1.01, g.l.=18, n.s.), (figura 6.2). Los valores medios de ARA en
Julio oscilaron entre 41 y 298.5 nmol C H / g p.s. nodulo-hora, mientras que
en Octubre se alcanzaron valores entre 510 y 2470 nmol C H / g p.s. 2 4
nodulo-hora. En Octubre la actividad se vio favorecida por las temperaturas
suaves, una mayor humedad del suelo consecuencia de las primeras lluvias
otoñales, y la emergencia de nuevos nodulos. En Julio, las elevadas
temperaturas y las escasez de lluvias produjeron la senescencia de los
nodulos, y consecuentemente la actividad de la nitrogenasa fue menor. La
temperatura del suelo permaneció prácticamente constante a lo largo del
período de muestreo en ambos meses (figura 6.2).
6.4. DISCUSIÓN.
Se ha mostrado que la altitud afecta al ARA en la especie actinorriza
Alnus nepalensis a lo largo de un gradiente de altitud de entre 1000 y 2500 m
(Sharma 1988), principalmente debido a la variación de la temperatura del
suelo. Por el contrario, en Almería no hemos detectado diferencias en los
niveles de actividad atribuibles a la altitud. Puesto que nuestro estudio está
restringido a los limites altitudinales de la cuenca, con sólo una diferencia
de 200 m entre parcelas, la temperatura del suelo no difería substancialmente
(figura 6.1) y por lo tanto el patrón del ARA fue similar.
El comportamiento estacional de la actividad nitrogenasa ha sido
relacionado con distintos factores. La actividad de los nodulos se ha
relacionado con el suministro y mobilización de fotosintatos, y por tanto,
dependiente de la tasa de fotosíntesis neta (Carlyle 1986). Gordon y Wheeler
(1978) han encontrado una correlación positiva entre la tasa media de
fotosíntesis neta y la tasa media de reducción de acetileno en clones de Alnus
glutinosa. La aparición y expansión de nuevas hojas que tiene lugar durante la
379
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
estación de crecimiento permite que alcanzar la mayor área fotosintética al
final de este período. Por tanto, el aumento del ARA durante la estación de
crecimiento parece ser una regla general para la mayoría de especies
fijadoras. Así ha sido observado en Cytisus scoparius en Oregon (Wheeler eí
al. 1987), en Alnus nepalensis en el Himalaya (Sharma 1988), en Acacia
pulchella y Acacia alata en el Suroeste de Australia (Hansen et al. 1987b),
y en Robinia pseudoacacia en Coweeta (USA), además de en las dos parcelas del
presente estudio. Sin embargo, en algunos estudios llevados a cabo con
individuos reproductores se han detectado disminuciones temporalea del ARA
durante la estación de crecimiento, los cuales han sido relacionados con el
inicio de la floración y la maduración de los frutos ya que estos procesos
implican un considerable aumento de los requerimientos de carbohidratos por
parte de las nuevas flores y frutos (Pizelle 1984, Wheeler et al. 1987). En
nuestro estudio, esta disminución no tuvo lugar, al igual que en otros
trabajos que empleaban individuos jóvenes, no reproductores (Helgerson et al.
1984).
El estrés hídrico ha sido descrito como una de las causas de la
senescencia de los nodulos (Boring y Swank 1984), lo que explica el alto
porcentaje de nodulos senescentes en nuestras parcelas en Julio, así como sus
bajas tasas de fijación. El clima mediterráneo se caracteriza entre otras
cosas por una elevada variabilidad interanual de la distribución e intensidad
de las precipitaciones, lo que a su vez afecta a la duración del período de
estrés hídrico (Specht y Molí 1983). Este período normalmente se extiende
desde Junio hasta Septiembre en el área de estudio (ICONA 1987), pero en el
año 1989, que fue excepcionalmente húmedo (Domingo 1991), la humedad acumulada
en el suelo pudo haber reducido el estrés hídrico sólo a los meses de Julio y
Agosto. Por tanto, en años de características medias en cuanto a
precipitaciones (450-500 mm), el período de baja actividad de la nitrogenasa
puede ser más largo que el observado en 1989. En la parcela de 1740 m, la
fijación media en Octubre de 1988 fue de 96.86 nmol C H / g p.s. nodulo-hora, 2 4
mientras que en el mismo mes de 1989 fue de 2187 nmol C H / g p.s.
nodulo-hora, un valor incluso mayor que el de primavera de ese mismo año.
Estas diferencias interanuales podrían explicarse por las precipitaciones de
Septiembre y Octubre, que fueron de 100 mm en 1988 y de 259 mm en 1989.
380
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
Además, la precipitación acumulada desde Enero hasta el día de muestreo fue de
483 mm en 1988 y de 704 mm en 1989. El efecto de la escasez de lluvias sobre
la reducción de la actividad de la nitrogenasa ha sido también encontrado en
Acacia pulchella y Acacia alata creciendo en climas mediterráneos en el oeste
de Australia (Hansen et al. 1987b).
Los nodulos jóvenes de Robinia pseudoacacia (Boring y Swank 1984) y de
Adenocarpus decorticans (Moro et al. 1992) aparecen después de la senescencia
estival. El crecimiento y desarrollo de los nodulos de Robinia ocurre en
invierno pero la fijación efectiva no comienza hasta la aparición de las
nuevas hojas en primavera. Debido a que A. decorticans es una planta perenne,
sus nuevos nodulos pueden comenzar la fijación inmediatamente después del
verano. En algunas especies de zonas templadas, como Cytisus scoparius en
Oregon, los valores del ARA decrecen gradualmente desde el verano hasta el
invierno (Wheeler et al. 1987). En el mismo estudio, estos autores encontraron
un pico de nuevos nodulos de pequeño tamaño (< 3 mm) en el período comprendido
entre Marzo y Mayo, lo que constituye un patrón estacional que difiere
claramente con el encontrado en este trabajo. Las características climáticas
de las áreas mediterráneas del sur de España, con temperaturas suaves y
mayores precipitaciones en otoño, producen una condiciones similares en la
primavera y el principio del otoño, que lógicamente deben condicionar la
fenología de las plantas. La elevada actividad otoñal de la nitrogenasa y la
emergencia de nuevos nodulos podrían representar una estrategia adaptativa de
algunas plantas fijadoras a las condiciones mediterráneas. Los resultados de
Pérez et al! (1989) también mostraron mayores tasas de fijación de nitrógeno
durante el otoño en nodulos de Alnus glutinosa de un bosque ripario de España
central. No obstante, se necesita más investigación en áreas mediterráneas
para establecer la generalidad de este patrón estacional. Al igual que en
otras leguminosas de zonas templadas (Sharma 1988,' Wheeler et al. 1987), el
ARA fue nulo durante el invierno en A. decorticans.
Los valores diarios de ARA en Octubre alcanzaban durante el día valores
significativamente mayores que durante la noche. La actividad enzimática está
estrechamente relacionada con el suministro de fotosintatos (Wheeler 1971),
aporte que se ve reducido durante la noche. No obstante, los nodulos pueden
381
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
utilizar reservas acumuladas para realizar la fijación (Boring y Swank 1984 en
Robinia, Schweitzer y Harper 1989 en Glycine). Esto podría explicar la
fijación durante la oscuridad encontrada en Octubre, aunque los valores
nocturnos fueran inferiores que los diurnos. En una plantación de Alnus del
Himalaya se encontró un patríon diario similar, pero con la diferencia de que
la actividad de la nitrogenasa se suspendía por completo durante la noche
(Sharma 1988). Boring y Swank (1984) también han afirmado que la disminución
de la temperatura del suelo puede producir un descenso substancial de la
actividad de los nodulos de Robinia en oscuridad. Pero en nuestra área de
estudio, la temperatura del suelo fue casi constante a lo largo de las 24
horas por lo que esta variable no puede explicar las diferencias entre los
períodos de luz y oscuridad en Adenocarpus y sería entonces la oscilación en
el suministro de fotosintatos la explicación más plausible. Sin embargo, no se
encontraron diferencias significativas entre períodos de luz y oscuridad en
Julio, cuando un gran porcentaje de los nodulos eran senescentes. Tampoco
Boring y Swank 1984J encontraron estas diferencias en nodulos inmaduros (< 3
mm) de Robinia en„Junio. Es posible que en estas dos clases de edad extremas,
jóvenes y senescentes, los patrones diarios de actividad difieran del de los
nodulos maduros. Por otra parte, la máxima actividad nocturna en el muestreo
de Julio se alcanzó a las 22 horas, declinando a continuación hasta las 7
horas, momento en el que el ARA alcanza el mínimo (figura 6.2). Puesto que la
temperatura del suelo fue casi constante, este descenso del ARA se podría
deber a un agotamiento progresivo de las reservas de carbohidratos usados en
la fijación de nitrógeno. Además, la temperatura del aire es mucho menor
después de la puesta del sol, por lo que el estrés hídrico disminuye también.
Este factor, podría contribuir a aumentar la eficiencia en el uso de las
reservas de fotosintato y explicar ciertos valores nocturnos superiores a
algunos diurnos.
6.5. ESTIMA DE LAS ENTRADAS POR FIJACIÓN DE NITRÓGENO EN A. decorticans.
Nuestra área de estudio se caracteriza por poseer suelos de baja
fertilidad y con pequeñas cantidades de materia orgánica (1.182), nitrógeno
(0.077.) y bajo CEC (5.25 meq/100 g) en los primeros 15 cm de suelo (datos
inéditos), valores similares a los referidos en ICONA (1987) para otros puntos
382
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
de la Sierra de los Filabres. Aunque las tasas de fijación de nitrógeno de las
leguminosas leñosas tienden a ser bajas (Carlyle 1986), la existencia de
poblaciones indígenas de estas leguminosas puede tener una influencia
significativa sobre el ciclo de nitrógeno en ecosistemas mediterráneos con
suelos pobres, que puede ser particularmente importante en las ter razas
creadas para las repoblaciones, como las de nuestra área de estudio, en los
que los horizontes originales del suelo han sido alterados. En este sentido,
el cálculo de la cantidad de nitrógeno fijado anualmente por A. decorticans
resulta interesante a la hora de conocer qué papel juega en el ciclo de
nutrientes de esta especie y del ecosistema en general.
Sin embargo, la transformación entre acetileno reducido- nitrógeno fijado
no es en absoluto recomendada por la mayoría de los autores (Herridge 1982,
Hansen et al . 1987, Müller y Bermudez de Castro 1987) debido a los numerosos
factores que influencian el factor de conversión teórico 3:1 (Hardy et
al. 1973) deducido a part ir de las ecuaciones :
N + 6 H+ > 2 NH 2 3
C H + 2 H+ » C H 2 2 2 4
Existen evidencias de que el método ARA puede sobrestimar o subestimar en
distintas proporciones la fijación real de nitrógeno (Hansen et al. 1987
Sellstedt et al . 1989, Turner y Gibson 1980). Una sobrestimación puede
provenir del reciclado de H mediante el enzima hidrogenasa que se encarga de
reutilizar parte del H producido como reacción secundaria cuando la
nitrogenasa utiliza N como sustrato (Hansen et al . 1987). Este autor cita
varias referencias en las que se constata esta disyunción entre el factor
teórico y real. Una subestimación puede tener su origen en un efecto
inhibitorio del C H sobre el funcionamiento de la nitrogenasa (Hansen et al. 2 2
1987, Minchin et al . 1983). Esta variación del factor teórico no sólo ocurre
entre distintos organismos sino en la misma especie según la cepa de Rhizobium.
que la infecte o las condiciones de incubación (Herridge 1982, Müller y
Bermudez de Castro 1987). En leguminosas se han encontrado factores entre
1.5:1 y 4:1 (Hansen et al. 1987, Turner y Gibson 1980, Müller y Bermudez de
Castro 1987) aunque se han citado valores entre 1.5:1 y 25:1 (Hardy et al .
383
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
1973).
El utilizar nodulos escindidos puede comportar una disminución de
fotosisntatos en el nodulo conforme progresa la incubación a pesar de que esta
sea corta (30-60 minutos). La pérdida de actividad puede suponer desde un bajo
porcentaje hasta un 85% de la correspondiente a nodulos intactos (Warembourg,
com. pers.). Por tanto, esta metodología subestima considerablemente el ARA y
consecuentemente la reducción de N (Hansen et al. 1987, Sellstedt et al.
1989). Por ello, en este trabajo, somos conscientes de que los valores
detectados pueden ser bastante inferiores a los que se habrían obtenido a
part ir de nodulos intactos. Sin embargo, dado el patrón de distribución de
éstos en el campo se hacía imposible la incubación del sistema raiz-nódulo "in
situ". Como comentamos en el apartado de metodología la elección de una hora
de incubación se debió a la predominancia de este tiempo de incubación por los
diversos autores consultados.
Todos estos problemas que influyen en la conversión acetileno-nitrógeno
fijado pueden solucionarse calibrando el ensayo del ARA a la manera descrita
por Hansen et al. (1987) o Burris (1974), en la que se utiliza paralelamente
el método de N para establecer el nitrógeno fijado, el ARA para determinar
la reducción de actileno, y la producción de H en atmófera tanto de aire como
de acetileno para establecer la importancia de la vía secundaria que puede
disminuir el rendimiento de la nitrogenasa. Desgraciadamente, al no disponer
de N , esta calibración no ha podido realizarse para este trabajo aunque
esperamos poder aplicarlo en un futuro.
El ARA, como medida de fijación de nitrógeno en hábitats naturales tiene
las mismas desventajas que cualquier otro ensayo de corta duración que se
utilice para el cálculo a más largo plazo. La variación debida a parámetros
como la radiación incidente, temperatura, humedad bien siguiendo un ciclo
nocturno-diurno, diario o estacional t rae consigo dificultades adicionales a
la hora de integrar adecuadamente los ensayos realizados en momentos puntuales
del año. Este problema ha quedado solventado, al menos en parte, en este
trabajo al cuantificar la actividad reductora de acetileno durante un ciclo
anual completo y dos ciclos diarios en momentos especiales del año (estrés
384
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Fijación de nitrógeno.
hídrico estival y primeras lluvias otoñales).
Resulta arriesgado la obtención de un valor medio de fijación anual de N 2
por parte del gaznacho, debido a los problemas comentados anteriormente y
sobre todo a la no realización del ensayo de calibración. No obstante hemos
calculado un rango aproximado calculado a par t i r de los valores medios
mensuales en ambas parcelas y el área de nodulación en cada mes. La variación
diaria a part ir de los resultados de Julio 1989 y Octubre 1989 no ha sido, sin
embargo tenida en cuenta ya que no podemos saber hasta que punto son
reproducibles estos resultados en otros períodos del año. Los valores
obtenidos utilizando el factor de conversión teórico de t res y el mínimo de
1.5 encontrado en leguminosas son de 0.25 Kg/ha. año y 0.5 Kg/ha año.
Suponiendo un 50% de pérdida de actividad al utilizar nodulos escindidos los
valores obtenidos se situarían entre 0.5 -1 Kg/ha. año. Los valores se sitúan
en el límite inferior dado por Carlyle para leguminosas en el que los valores
más bajos corresponden precisamente a leguminosas leñosas. La baja nodulación
es básicamente las responsable de esta pequeña entrada por fijación. Los
valores de nodulación para Cytisus scoparius en Oregon alcanzan 1.2 g
nódulo/m (Wheeler et al. 1987) diez veces superior a los nuestros y valores
de fijación de 10.12 Kg/ha. año. No conocemos la existencia de estimas en otras
leguminosas de este tipo en la zona mediterránea, lo que supone un seri
handicap a la hora de establecer si los valores de ARA encontrados en A.
decortlcans son habituales en estas especies. En otra especie de Adenocarpus
que habita en las Sierras de la provincia de Madrid, el área de nodulación
pude llegar a ser incluso inferior a la de nuestro trabajo (Bermúdez de
Castro, com. personal) pero por el momento no existen ninguna estima
cuantitativa. La baja nodulación también parece una característ ica
generalizada en leguminosas leñosas aunque no sean mediterráneas (Carlyle
1986). La baja fijación anual también puede ser átribuíble en parte a la
existencia de dos momentos críticos en que la actividad se reduce al mínimo
(meses de invierno con bajas temperaturas y estrés hídrico estival, figura
6.1). La inhibición de la actividad durante el invierno podría no ser tan
drástica en años con climatología menos extrema y temperaturas edáficas algo
más altas que la encontradas en el presente estudio lo que implicaría un
considerable aumento en la cantidad de nitrógeno fijado.
385
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
386
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Sección tercera. Aproximación a los
flujos biogeoquímicos.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Capítulo 7 Flujos internos
y biogeoquímicos
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
En los capítulos anteriores se han tratado aspectos sobre algunos de los
compartimentos y flujos de nutrientes más importantes del ciclo interno
(nutrientes movilizados en producción, desfronde, lixiviado, descomposición).
Así mismo, se dispone de datos sobre los flujos externos en la cuenca (balance
de entradas y salidas por precipitación y avenamiento (Domingo 1991), tasas de
deposición seca, y de fijación de N atmosférico). Para algunos de los flujos,
los intervalos de muestreo no son exactamente coincidentes mientras que
algunos han sido estimados por métodos indirectos (deposición seca y lixiviado
por ejemplo). Tampoco se dispone de información de la dinámica de nutrientes
en raices, roca madre y suelo mineral. A pesar de todas estas limitaciones,
pensamos que puede establecerse de una manera muy general una descripción
sobre la circulación de nutrientes en la cuenca, utilizando tanto la
aproximación específica como la sectorial.
FLUJOS INTERNOS.
Con los datos obtenidos en base anual de los distintos flujos internos,
puede realizarse un balance de masas (Waring y Schlesinger 1985, Colé y Rapp
1981) calculando una serie de parámetros que son combinación de varios de
estos flujos:
RETENCIÓN: nutrientes incorporados anualmente a estructuras perennes:
tronco ramas y tallos del año. A la vista de los resultados expuestos en el
capítulo 2, se considera que la pérdida de material perenne por desfronde es
despreciable. Para las especies de pino se considera también la retención
foliar calculada a partir de los nutrientes asociados al incremento de la
biomasa foliarícapt 2).
RETORNO: Cantidades de nutrientes devueltas anualmente al suelo vía
interna. Sería equivalente a la suma de desfronde y lixiviado.
REQUERIMIENTOS. Cantidades de nutrientes requeridas anualmente para
estructuras de nueva formación. Sería equivalente a los nutrientes asociados a
la producción del material perenne, a la producción de hojas del año y a la de
387
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
órganos reproductivos añadiendo igualmente el lixiviado vía trascolación neta.
ABSORCIÓN. Equivaldría a las cantidades absorbidas por las raices.
Calculado como la suma de retorno y de retención.
RETRASLOCACION. Diferencia entre requerimientos y absorción. Equivaldría
a la cantidad de nutrientes requeridos que no han sido absorbidos por las
raices y que la planta debe cubrir a part ir de una retraslocación desde
órganos viejos. Un signo positivo representa retraslocación mientras que el
signo negativo representa que los requerimientos están sobradamente cubiertos
con la absorción.
Estos parámetros han sido calculados para cada especie y región (Tablas
7.1 y 7.2) con la finalidad de valorar tanto el factor específico como la
utilización conjunta de nutrientes por la vegetación en una y otra área. El
balance requerimiento -absorción en cada área representaría una medida de la
eficacia en la utilización interna de nutrientes por parte de la vegetación
teniendo en cuenta su estructura particular en esa región. Por otro lado, las
especies estudiadas presentan para algunos nutrientes estrategias diferentes
respecto a la circulación interna. La mayor o menor dominancia de cualquiera
de ellas podría implicar cambios sustanciales en la transferencia de
nutrientes de una región con respecto a la otra. También puede ocurrir que el
factor específico se diluya cuando consideremos la circulación conjunta de
cada elemento en las dos grandes regiones. Esta última posibilidad parece ser
un hecho en la cuenca " Nacimiento" (ver apartados finales de Tabla 7.1 y
7.2). No se aprecian grandes diferencias entre una y otra zona. Estas son
además, meramente cuantitativas más que cualitativas. Así, existe
retraslocación de N, P y K tanto en RPP como en RPN mientras que en Ca, Mg y
Na no se detecta retraslocación. El porcentaje de los requerimientos de N y P
que son cubiertos por retraslocación son de similar orden en ambas zonas (N 20
y 167.; P: 16 y 19 7. y algo más diferentes para el K (20 y 12.77.).
Si se comparan las dos especies de pino (tabla 7.1 y 7.2) destacan los
elevados requerimientos en ambas especies en buena parte determinados por la
elevada productividad foliar que mantienen en esta etapa de la sucesión
388
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

TABLA 7.1. Flujos internos en las especis componentes de la región de P. pinaster. Todos los valores en (kg/ha.año)
P.pinaster N P K Ca Mg Na
PROHOJAS PROBROTES PROMADERA PROREP INCHOJA RETENCIÓN DESFRONDE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTOS RETRASLOCACION
13.95 2.63 2.09 .30
4.60 9.32
8 .03
8.03 17.35 20.40 3.06
1.58 .96 .24 .02 .50
1.70 .76 0
.76 2.46 2.88 .42
6.81 1.31 2.25 .04
2.25 5.81 2.11 1.26 3.37 9.18 12.10 2.93
3.43 1.36 2.94 .12
1.15 5.45 20.25 7.70 27.95 33.40 15.70 -17.70
2.52 .48 .74 .05 .80
2.02 3.80 1.62 5.42 7.44 5.62 -1.82
.35
.05
.19
.01
.08
.32
.44 1.80 2.24 2.56 .89
-1.67
A. decorticans
PROHOJAS PROBROTES PROMADERA PROREP RETENCIÓN DESFRONDE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTOS RETRASLOCACION RETRASLOCACION*
7.25 .97
1.11 2.39 2.08 8.79
0 8.79 10.87 11.72 .85
-2.08
.30
.06
.05
.09
.11
.35
.07
.42
.53
.57
.04 -.04
1.60 .49 .43 .77 .92
2.18 1.64 3.82 4.74 4.93 .19 .72
1.10 .14 .35 .63 .49
3.41 .15
3.56 4.05 2.37 -1.68 -.34
.52
.06
.08
.17
.14
.80
.14
.94 1.08 .97
-.11 -.00
.09
.03
.02
.02
.05
.07
.09
.16
.21
.25
.04
.05
(*)
C.
. Sin incluir
laurifoliuB
la fracci ón reproductiva en los requer imientos.
PROHOJAS PROBROTES PROMADERA PROREP RETENCIÓN DESFRONDE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTOS RETRASLOCACION
7.60 1.40 1.06 .56
2.46 3.31
0 3.31 5.77 10.62 4.85
.84
.21
.11
.06
.32
.59
.13
.72 1.04 1.35 .31
2.74 .95 .27 .36
1.22 1.46 2.30 3.76 4.98 6.62 1.64
5.88 1.79 1.93 .12
3.72 9.87 1.15 ' 11.02 14.74 10.87 -3.87
1.21 .24 .12 .08 .36
1.58 .37
1.95 2.31 2.02 -.29
.07
.02
.02
.01
.04
.08
.05
.13
.17
.16 -.01
TOTAL REGIÓN PP N P K Ca Mg Na
ABSORCIÓN 33.99 4.03 18.90 52.19 10.83 2.90 REQUERIMIENTOS 42.74 4.80 23.65 28.94 8.61 1.30 RETRASLOCACION 8.76 .77 4.76 -23.25 -2.22 -1.60
389
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
TABLA 7.2. Flujos interno* on l o„ T«in- ,nter"os en l « especs conponentes de la región de P. nigra. Todos los valores en Ckg/ha.año)
P- nigra N P * Ca Mg
PROHOJA PROBROT PROMAO PROREP INCHOJA RETENCIÓN DESFRONOE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTO RETRASLOCACION
16 2.58
2 .06
6.37 10.95 4.90 .34
5.24 16.19 21.20 5.01
1.62 .38 .15 .00 .63 1.16 .45 0
.45 1.61 2.17 .56
3.90 1.10 1.02 .01
1.47 3.59 1.03
0 1.03 4.62 6.18 1.56
3.10 1.35 2.59 .02
1.19 5.13 8.93 .51
9.44 14.57 12.12 -2.45
2.03 .64 .50 .01 .77
1.91 1.76 .11
1.87 3.78 3.34 -.44
.18
.04
.08
.00
.12
.24
.14
.65
.79 1.03 .06
-.97
A. decorticans
PROHOJA PROBROT PROMAO PROREP RETENCIÓN DESFRONDE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTO RETRASLOCACION RETRASLOCACION*
17.90 2.10 3.01 5.98 5.11 22.39
0 22.39 27.50 28.99 1.49 -4.49
.74
.12
.13
.24
.25
.88
.16 1.04 1.29 1.39 .10
-.14
3.96 1.08 1.16 1.92 2.24 5.56 4.18 9.74 11.98 12.30 .32
-1.60
2.74 .30 .95
1.57 1.25 8.69 .40
9.09 10.34 5.96 -4.38 -5.95
1.30 .14 .22 .44 .36
2.04 .36
2.40 2.76 2.46 -.30 -.74
(*). Sin incluir la fracción reproductiva en los requerimientos.
C. laurifoliuB
PROHOJA PROBROT PROMAD PROREP RETENCIÓN DESFRONDE LIXIVIADO RETORNO ABSORCIÓN REQUERIMIENTO RETRASLOCACION
4.24 .740 .85 .30 1.59 1.79
0 1.79 3.38 6.13 2.75
.46 .110 .09 .04 .20 .32 .07 .39 .59 .77 .18
1.51 .500 .22 .20 .72 .79
1.25 2.04 2.76 3.68 .92
3.25 .940 1.55 .06
2.49 5.35 .63
5.98 8.47 6.43 -2.04
.67 .130 , .09 .05 .22 .85 .21
1.06 1.28 1.15 -.13
.03 .014 .02 .01 .03 .04 .03 .07 .10 .10 0
TOTAL ÁREA P. MIG8A N P K Ca Mg Na
ABSORCIÓN 47.07 3.40 ^ ^
REQUERIMIENTO 56.32 4 33 3? 1, RETRASLOCACION o 25 '«, ,„„ 24"51 6-'5 .64
'* 2-80 -8-87 -.87 - 97
390
.09
.06
.04
.05
.10
.17
.24
.41
.51
.48 -.03 -.08
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
destinada no sólo al mantenimiento sino al incremento de la biomasa foliar. El
porcentaje de requerimientos destinados a las acículas oscila entre más del
707. para el N y el 22-25% para el Ca reflejando de forma combinada la riqueza
en hojas de los distintos elementos así como su importancia relativa en la
producción. En las dos especies se manifiestan mecanismos de retraslocación
para el N, P y K. Contrastan estos resultados con los obtenido por Colé y Rapp
(1981) en coniferas en las que el comportamiento medio de los distintos
bosques consiste en un equilibrio entre absorción-requerimientos para el N y
P y a ser negativo para el resto. Sin embargo Carlyle (1986) cita varias
referencias en las que por el contrario se han encontrado importantes tasas de
retraslocación en distintos bosques de coniferas. Los porcentajes de
retraslocación en los pinos estudiados son relativamente notables :3 y 5
Kg/ha.año (157. y 237. de los requerimientos) para el N y 0.42 y 0.56 Kg/ha.año
para el P (14 y 257. de los requerimientos). Los resultados obtenidos a par t i r
de diferencias de concentración N entre acículas de un año y caídas por
desfronde fenológico (capítulo 3) permitían calcular una eficacia en la
redistribución de N del 60% en P. nigra lo que supondría una retraslocación
foliar de 8 Kg /ha.año en esta especie. Con el mismo método se obtendrían
unos 7 Kg/ha.año en P. pinaster. Ambos valores son superiores a los obtenidos
mediante el balance de masas. Destaca la diferencia respecto al dato de
retraslocación en P. pinaster calculado de las dos formas. La probable
sobrestimación del peso del desfronde que ya hemos comentado en repetidas
ocasiones origina una clara disminución en el cálculo de la redistribución a
part i r de la diferencia entre requerimientos y absorción. La diferencia en la
utilización de N en las especies estudiadas frente a otros bosques de
coniferas puede explicarse en parte teniendo en cuenta las bajas cantidades de
N total en estos suelos (alrededor de 1.6 T/ha en" los 30 primeros cm del suelo
mineral) lo que podría constituir una limitación importante para el
crecimiento del pinar solventado en buena medida por el desarrollo de
mecanismos de redistribución interna. Circulaciones bajas de N en hojarasca
combinadas con un alto valor del cociente peso desfronde: N en desfronde
aparecen en las dos especies estudiadas (capt.3). Según Vitousek (1982), la
baja disponibilidad de N en suelo podría seleccionar este tipo de estrategia
para optimizar la utilización del nutriente considerado. A pesar de que esta
hipótesis haya sido ampliamente discutida en muchos trabajos y de que no
391
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
existe una evidencia general de que las especies que viven en sitio
infértiles sean más eficientes en el uso del nutriente limitante, el cas
estudiado sí podría constituir uno de estos casos.
En A. decorticans la retraslocación ha sido calculada considerando o n<
la fracción reproductiva como parte de los requerimientos. Algunos estudios n<
consideran esta fracción (Colé y Rapp 1981) pero en aquellos caso en que e
esfuerzo reproductivo sea cuantitativamente importante, el balance fina]
requerimientos - absorción puede variar de forma considerable. Sin considerar
la fracción reproductiva, no se detecta retraslocación para ninguno de los
elementos. En el caso del N esta característica podría resultar lógica dado
que en esta especie existe una fuente adicional de N mediante fijación
biológica. Sin embargo cuendo se incluye la fracción reproductiva, muy rica en
elementos móviles y esenciales (N, P y K), el balance se declina hacia signo
positivo (retraslocación) si bien la magnitud de ésta es muy pequeña si la
comparamos con el resto de especies estudiadas. En general se observa que para
todos los elementos, el balance entre requerimientos-absorción resulta muy
próxima a cero.
Para C. laurifolius se encuentran similares características a las de A.
decorticans, existiendo retraslocación respecto al N, P y K aunque de
magnitudes muy superiores. La traslocación de N supone un elevado porcentaje
de los requerimientos (44-45%) valor muy similar al encontrado por Gray (1983)
en el chaparral perenne californiano (44%). Este autor detecta también
retraslocación (que el denomina redistribución) para el P y el K. El
porcentaje de retraslocación de P en relación a los requerimientos no es
demasiado alto en la j a r a (23%) si la comparamos con el 56% obtenido por Gray
(1983). Este porcentaje supone para el conjunto de la cuenca una cantidad de
0.22 Kg/ha.año muy similar al que se puede calcular a par t i r de la diferencia
en concentraciones química entre hojas verdes y escindidas (0.17
Kg/ha.año). Como se ha comentado en anteriores capítulos C. laurifolius es una
especie que no utiliza de forma eficiente el P ya que a los valores elevado de
concentración foliar se le añade un contenido de este elemento en desfronde
fenológico muy similar al de hojas vivas. Por otro lado el fosfato de la
lluvia tampoco es aprovechado eficientemente puesto que no existe absorción
392
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
foliar y sí lixiviación. El porcentaje de reabsorción de K es del orden del
doble en C. laurifolius en relación al chaparral. Probablemente esta
diferencia se deba a que Gray( 1983) no contabiliza el lixiviado como parte de
los requerimientos y sí en la absorción. Si recalculamos de esta forma la
retraslocación en la jara , ésta no solamente disminuiría sino que no existiría
dado que la absorción superaría en unos 3 Kg/ha año a los requerimientos. Otro
tanto ocurriría en el gaznacho eliminando el lixiviado de K de los
requerimientos.
Balance a nivel de hojarasca.
Puede construirse un balance a nivel de hojarasca a la manera descrita
por Verdú (1984). La información combinada de la liberación de nutrientes
durante el proceso de descomposición (capítulo 5) y los aportes por la
fracción foliar del desfronde (capítulo 3) permite calcular la cantidad de
éstos que es inmovilizada o liberada por la hojarasca del suelo.
Posterioremente, se tienen en cuenta los aportes por lixiviado vía hidrológica
para deducir el balance del primer año de descomposición. Al disponer sólo de
datos de liberación de nutrientes en una año y medio, los valores han sido
únicamente calculados para el primer año de descomposición y para la fracción
foliar de desfronde. En las tablas 7.3 y 7.4 se ha construido este tipo de
balance para las dos grandes zonas de la cuenca. Los resultados se exponen
tanto para cada una de las especies como para su conjunción bien en RPP ó en
RPN.
Tras el primer año los dos pinos junto con la j a ra presentan
inmovilizaciones netas de N. A. decorticans libera N ya a part ir del primer
año. Para el resto de los elementos existe liberación en mayor y menor grado.
Los resultados varían poco al añadir el lixiviado excepto para el K en
matorrales y Ca en el pinar debido a una mayor importancia relativa del
lixiviado en estos casos. Comparando los resultados globales en las dos
regiones se aprecia una diferencia fundamental determinada por la mayor o
menor abundancia del gaznacho en las dos zonas. Así En RPP el balance neto
respecto al N es una inmovilización de 3.8 Kg/ha mientras que en RPN existe
mineralización de 1.61 Kg/ha en el primer año de descomposición. Para el resto
de los elementos las diferencias específicas se diluyen en el resultado global
393
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
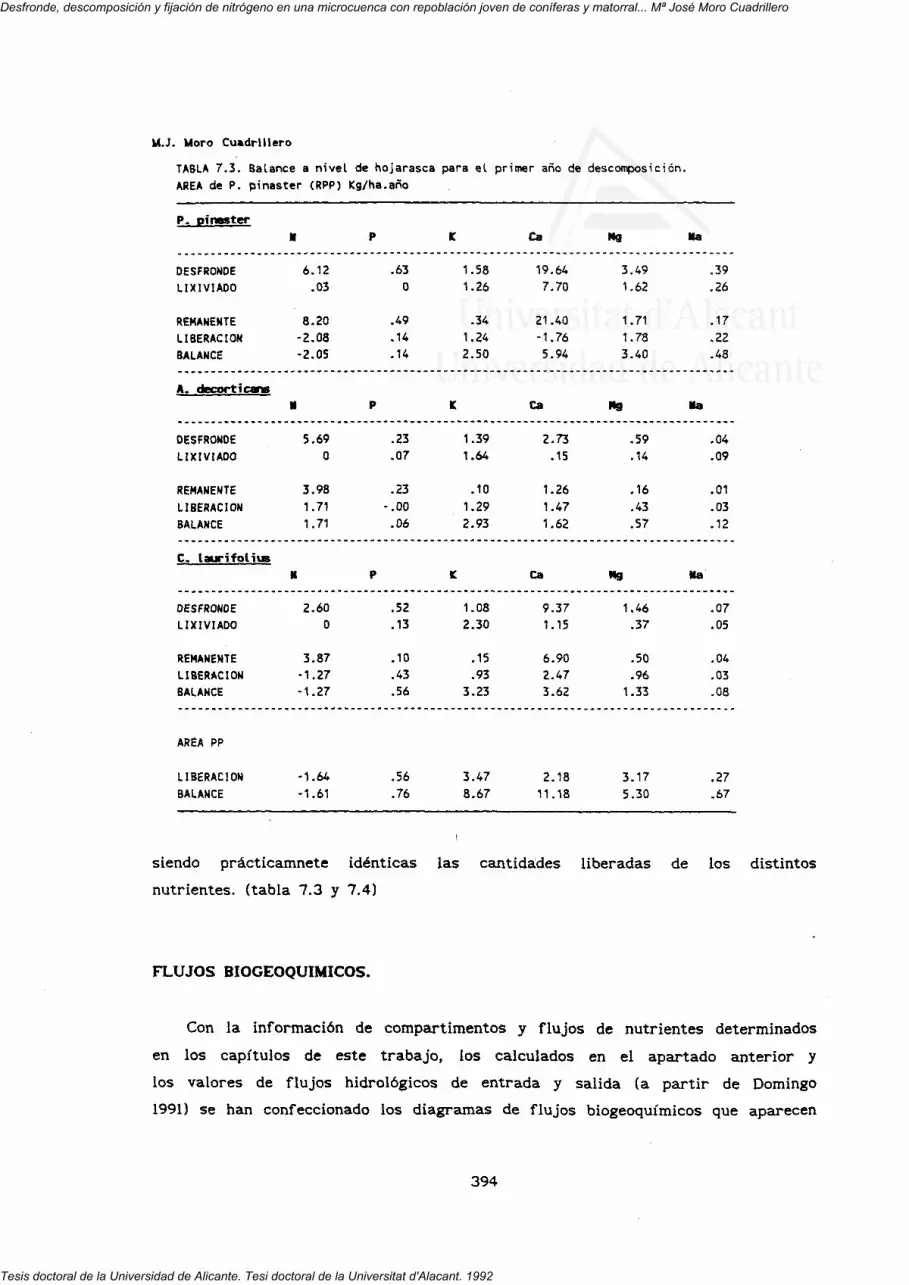
M.J. Moro Cuadrillero
TABLA 7.3. Balance a n ive l de hojarasca para e l primer año de descomposición. ÁREA de P. pinaster (RPP) Kg/ha.año
P. DÍnaster
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
A. decorticarw
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
C. taurifolius
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
ÁREA PP
LIBERACIÓN
BALANCE
H
6.12
.03
8.20
-2.08
-2.05
H
5.69
0
3.98
1.71
1.71
N
2.60
0
3.87
-1.27
-1.27
-1.64
-1.61
P
.63 0
.49
.14
.14
P
.23
.07
.23 -.00
.06
P
.52
.13
.10
.43
.56
.56
.76
K
1.58
1.26
.34 1.24
2.50
K
1.39
1.64
.10 1.29
2.93
K
1.08
2.30
.15
.93 3.23
3.47
8.67
Ca
19.64
7.70
21.40
-1.76
5.94
Ca
2.73
.15
1.26
1.47
1.62
Ca
9.37
1.15
6.90
2.47
3.62
2.18
11.18
Mg
3.49
1.62
1.71
1.78
3.40
«8
.59
.14
.16
.43
.57
Ng
1.46
.37
.50
.96 1.33
3.17
5.30
Ma
Ha
Ha
.39
.16
.17
.22
.48
.04
.09
.01
.03
.12
.07
.05
.04
.03
.08
.27
.67
siendo prácticamnete idénticas las cantidades liberadas de los distintos
nutrientes, (tabla 7.3 y 7.4)
FLUJOS BIOGEOQUIMICOS.
Con la información de compartimentos y flujos de nutrientes determinados
en los capítulos de este trabajo, los calculados en el apartado anterior y
los valores de flujos hidrológicos de entrada y salida (a par t i r de Domingo
1991) se han confeccionado los diagramas de flujos biogeoquímicos que aparecen
394
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
TABLA 7.4. Balance a nivel de hojarasca para el primer año de descomposición.
ÁREA de P. nigra (RPN). (Kg/ha.año)
P. niara
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
A. cJecorticans
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
C. laurífolius
DESFRONDE
LIXIVIADO
REMANENTE
LIBERACIÓN
BALANCE
ÁREA PN
LIBERACIÓN
BALANCE
M
4.14
.34
4.36
-.22
.12
II
14.50
0
10.15
4.35
4.35
H
1.40
0
2.08
-.68
-.68
3.45
3.79
P
.42
0
.29
.13
.13
P
.57
.16
.58
-.01
.15
P
.28
.07
.05
.23
.30
.35
.58
t
.86
0
.44
.42
.42
t
3.55
4.18
.25
3.30
7.48
K
.59
1.25
.08
.50
1.75
4.23
9.66
Ca
8.84
5
7.93
.91
5.91
Ca
6.95
.40
3.22
3.73
4.13
Ca
5.08
.63
3.76
1.32
1.95
5.96
11.99
Ng
1.68
.11
.66
1.02
1.13
Ng
1.51
.36
.42
1.09
1.45
Ng
.79
.21
.27
.52
.73
2.63
3.31
Na
-
Ha
Ha
.12
.26
.15
.03
.23
.09
.24
.03
.06
.30
.04
.03
.02
.01
.04
.04
.57
en las figuras 7.1. a 7.18. Se han escogido t res de los elementos estudiados
(N,P y Ca). N y P constituyen nutrientes esenciales para el Sistema y son los
que más fácilmente pueden resultar limitantes en sistemas mediterráneaos. Para
ellos, una pauta común en la cuenca "Nacimiento" es la reutilización mediante
traslocación y la retención vía hidrológica (Domingo 1991). En el otro extremo
está el Ca, elemento abundante en el suelo y cuyas entradas vía deposición
global son considerables en la zona de estudio. Este elemento es exportado de
la cuenca y no se han detectado mecanismos de retraslocación tendentes a
economizar su uso. Los esquemas se han realizado primero individualmente para
cada especie. En el matorral éstos sólo se construido para la zona RPN. A
395
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
continuación se realizan los diagramas conjuntos para las especies componentes
de cada área (RPP Y RPN). Por tanto, para cada elemento se contempla la
circulación individual y global de las especies y el efecto que unas u otras
pueden ejercer en el comportamiento general del sistema. Resulta interesante
para la gestión del bosque, conocer cómo se modificarían los flujos si el
matorral fuera eliminado y de qué manera afectaría este cambio al pinar. A
pesar de la información tan limitada de la que disponemos pueden plantearse ya
ciertas consideraciones que se irán viendo a los largo del capítulo.
La información sobre flujos y ciclos biogeoquímicos para cada especie y
elemento con la que se han confeccionado las figuras se muestra en las tablas
7.5 a 7.8. Se añade información adicional sobre el contenido en nutrientes del
mantillo y suelo mineral (primeros 30 cm). Los valores de mantillo se refieren
solamente a la fracción hoja del conjunto de las especies sin diferenciar su
grado de descomposición. Los valores se han obtenido a par t i r de 15 cuadrados
de 0.25 m seleccionados al azar en cada una de las dos grandes regiones
durante el año 1991. Dada la elevada variabilidad espacial de este almacén, el
número de muestras resulta claramente insuficiente pero aún así puede tomarse
como dato orientativo. Los resultados respecto al suelo mineral son parte de
un muestreo más amplio llevado a cabo por el equipo de Edafología de la U. B.
en la cuenca Nacimiento y en otra adyacente (Barranco del Agua). Los datos
presentados son los valores medios de contenido en nutrientes calculados en
muestras compuestas a part i r de extración con sonda en cada una de las dos
grandes regiones RPP y RPN. Los parámetros de nutrientes en suelo mineral son
los siguientes: N total, P asimilable y cationes de intercambio. No se
dispone, por tanto de valores de N potencialmente asimilable ni del contenido
total de P, K, Ca, Mg y Na. El cálculo de N disponible a part ir del total
utilizando porcentajes encontrados en la bibliografía resulta
extraordinariamente arriesgado dada su variabilidad temporal y espacial
incluso dentro de una misma zona. No obstante, hemos utilizado el intervalo
que da Cortina (1992) en zonas catalanas plantadas con poblaciones de P.
radiata de 17-19 años de edad y que oscila, entre el 4 y el 10% del N total
para los 30 primeros cm de suelo. Los valores son, por tanto meramente
especulativos.
396
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
TABLA 7.5. Compartimentos y flujos biogeoqufmicos en relación con P.pinaster
en su área de distribución.
FLUJOS (Kfl/ha.año)
N P K Ca Mg Na
D.GLOBAL
T.NETA
D.SECA
LIXIVIADO
ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
RETRASLOCACION
6.50
1.21
1.18
.03
.24
9.32
8
8.03
17.35
20.40
3.06
.17
0
0
0
-.06
1.70
.76
.76
2.46
2.88
.42
3.63
2.25
.99
1.26
0
5.81
2.11
3.37
9.18
12.10
2.93
15.02
11.60
3.89
7.70
0
5.45
20.25
27.95
33.40
15.70
-17.70
2.47
1.92
.30
1.62
0
2.02
3.80
5.42
7.44
5.62
-1.82
4.01
2.63
.80
1.80
0
.32
.44
2.20
2.50
.89
-1.60
COPARTUENTOS (KG/HA) O/ha MINERALOMASA 60.90 6.70 32.80 65.60 15.90 2.11
MANTILLO TOTAL 44.30 2.82 3.22 106 10.30 .53
intercambiables
Ntot P.asim K Ca Mg Na
SUELO MINERAL 1192 20 309 1877 197.50 167
TABLA 7.6 . Compartimentos y f l u j os biogeoqufmicos en re lación con P. nigra en su área de d i s t r i buc ión .
FLUJOS ( K a / h a . a ñ o N P K Ca Mg Na
0.GLOBAL
T.NETA
D.SECA
LIXIVIADO
ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTO
RETRASLOCACION
6.50
.45
.11
.34
-.40
10.95
4.90
5.24
16.19
21.20
5.01
.17
0
0
0
-.07
1.16
.45
.45
1.61
2.17
.56
3.63
2.02
1.49
--
3.59
1.03
1.03
4.62
6.18
1.56
15.02
10.14
9.63
.51
0
5.13
8.93
9.44
14.57
12.12
-2.45
2.47
1.19
1.08
.11
0
1.91
1.76
1.87
3.78
3.34
-.44
4.01
2.64
1.99
' .65
0
.24
.14
.79
1
.06
-.94
COTART nonos (Ka/ha) kG/HA
MINERALOMASA 74 7.65 21.40 48.90 12.80 1.20
MANTILLO TOTAL 42 2.04 2.81 103 9.11 .48 intercambiables
Ntot P.asim K Ca Mg Na
SUELO MINERAL 1673 33 337 2259 195.50 170
397
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
TABLA 7.7. Compartí «lento» y flujos biogeoqufmicos en relación con A. decorticans
en una de las áreas de distribución de la cuenca (Área de P. nigra).
FLUJOS(Ko/ha.sfio N P K Ca Mg Na
0.GLOBAL
T.NETA
LIXIVIADO
D.SECA
ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
TRASLOCACION
TRASLOCACION*
6.50
1.08
NS
- . 1 7
- .43
5.11
22.39
22.39
27.50
28.99
1.49
-4 .49
.17
.16
NS
NS
0
.25
.88
1.04
1.29
1.39
.10
- .14
3.63
9.22
4.18
5.05
0
2.24
5.56
9.74
11.98
12.30
.32 -1 .60
15.02
6.60
.38
6.22
0
1.25
8.69
9.09
10.34
5.96
-4 .38
-5.95
2.47
1.44
.36
1.08
0
.36
2.04
2.40
2.76
2.46
- .30 - .74
4.01
2.33 .24
1.90
-.10
.17
.41
.51
.48
- . 03 - . 08
asp«Tiierra5(Ka/iia) N
MINERALOMASA
MANTILLO TOTAL
SUELO MINERAL
59.60
42
Ntot
1673
P
1.98
2.04
P.asim
33
K
20
2.81
K
337
Ca
14.60
103
- in tercambiab les
Ca
Mg
4 .67
9.11
Mg
2259 195.50
Na
.60
.48
Na
170
TABLA 7.8. Compartimentos y flujos biogeoqufmicos en relación con C. taurifolius en una de las áreas de distribución de la cuenca (Área de P. nigra).
FLLUOS(Ca/ha-anD N P K Ca Mg Na
D.GLOBAL
T.NETA
D.SECA
LIXIVIADO ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS TRASLOCACION
6.50
0
0
0
- .75
1.59
1.79
1.79
3.38
6.13
2.75
.17
.11
.04
.07
0
.20
.32
.39
.59
.77
.18
3.63
2.50
1.24
1.26
0
.72
.79
2.04
2.76
3.68
.92
15.02 .97
.33
.62
0
2.49
5.35
5.98
8.47
6.43
-2.04
2.47
.33
.12
.20
0
.22
.85
1.06
1.28
1.15
- .13
4.01 .28
.22
.06
0
.03
.04
.07
.10
.10
0
OM>MTIIEI ITOS(ra/ l») N
MINERALOHASA 21.50
P
2.29
K
6.68
Ca
28.62
Mg
2.86
Na
.32
MANTILLO TOTAL 42 2.04 2.81 103 9.11 .48
i ntercambi ables
Ntot P.asim K Ca Mg Na
SUELO MINERAL 1673 33 337 2259 195.50 170
398
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
Nitrógeno.
La absorción de N por parte ambas especies de pino (figuras 7.1 y 7.2)
resulta elevada dados los altos requerimientos destinados mayoritariamente al
mantenimiento de una alta producción foliar. La retraslocación interna, más
desarrollada en P. nigra permite suplir la cantidad restante para satisfacer
los requerimientos. El acumulo de N en la biomasa aérea del pino laricio
supone un 47. de todo el N del sistema (biomasa aerea del pinar-mantillo-suelo
mineral), porcentaje que aumenta considerablemente si se considera el N
disponible en suelo y no el total. El coste nutricional es muy elevado para el
N en esta especie (4000g N / T biomasa), lo que implicaría "a priori" una
explotación del suelo para la incorporación de N a la biomasa, explotación que
queda en parte frenada mediante mecanismos de reutilización. Para P. pinaster
se obtienen resultados similares: el 4.7 7. del N total del sistema se acumula
en la biomasa aérea que presenta un coste nutricional de 3812 g de N/ T
biomasa.
La trascolación neta aporta vía lixiviado y lavado de deposición seca 0.45
Kg/ha.año (P. nigra) y 1.21 (P. pinaster) de N- N03 adicionales mientras que
parte del amonio de la lluvia es absorbido directamente por las superficies
foliares de ambas especies..
La existencia de retraslocación y absorción foliar parecen indicar
estrategias de conservación de N en estas especies que aumenta, por tanto, la
eficacia en el uso de éste. La conservación de N se manifiesta también en el
signo del balance hidrológico, que exporta anualmente sólo un 17% de las
entradas por deposición global lo que implica una retención de 5.35 Kg/ha.año
mayoritariamente en forma de compuestos asimilables (NH4 y N03) y en mucho
menor proporción de N orgánico (Domingo 1991).
El desfronde devuelve al mantillo un 6.7% (P. nigra) y un 13 % (P.
pinaster) del N acumulado en la biomasa aérea que será inmovilizado por lo
menos durante los dos primeros años de descomposición (capt. 5). Por su parte,
9.3 Kg de N (resinero) y 10.9 (laricio) quedan fuera de circulación por lo
menos a corto plazo al ser retenidos en la biomasa estructuras perennes y en
la fracción de la producción foliar destinada al incremento de biomasa.
399
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
N
$?/M,
^ BIOMASA 6 1 &*
¥¿$V< RETENCIÓN 9.3 '''\l¿ JL& 'l? TRASLOCACION \~+3~] l « _ >*V»# ,.-v REQUERIMIENTOS 20.4l¿.
ABSORCIÓN FOLIAR
0.24
LAUADO DEPOSICIÓN SECA 1.18
DER GLOBAL 6.5
•LIXIUIADO 0.83
TRASCOLACION 1 . 2 1
DESFRONDE 8
Sh \.V« ^ W > . , > S U r t U t o jUAS.Vtfl'/>-»<V^A¿ >\* ^ t .^O*,4r ^ ^ v W J s v :
» y v i v « o f \ o f 1
ABSORCIÓN 1 7 .
1 SUELO
DISPONIBLE
33
j
5 4 *
^ J ^ o -• O
nlNERALIZACIQN
MANTILLO 4 4 . 3
i r
SUELO MINERAL
C 4 7 - 1 1 9 3 ? AVENAMIENTO
1.15
1 1 9 2
F i g u r a 7.1. Flujos biogeoquímicos p a r a P.pinaster en su á r e a de dis t r ibución:
Nitrógeno. Flujos en Kg/ha -año . Compart imentos en Kg/ha.
400
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujos
N
Jw¡ VÁi «¿MV^ ™ . « , . « . „ . m „ J r r r - " J , — L A U A D O DEPOSICIÓN SECA0.11
• ABSORCIÓN FOLIAR 0.40
DEP. GLOBAL 6.5
•LIXIUIADO 0.34
TRASCOLACION 0 . 4 5
DESFRONDE 4 .9
^ M l ^ -üa i ^ . , A U > ^ i ^ t A Í rtusw^m^ \ j ^ ^M fon^y*^'^ uJ^ftJJSl.
'«'W °* • ^
t ABSORCIÓN 1S.2
SUELO DISPONIBLE
MINERALIZACION
MANTILLO 42
SUELO MINERAL
[67-1673?
AVENAMIENTO 1.15
1 6 7 3
Figura 7.2. Flujos biogeoquímicos p a r a P,nigra en su á r e a de dis tr ibución:
Nitrógeno. Flujos en Kg/ha -año . Compartimentos en Kg/ha.
401
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Cabe preguntarse si el suelo puede soportar este coste nutricional durante
años posteriores. Probablemente se esté en estos momentos en la etapa de mayor
demanda nutricional que disminuirá presumiblemente t ras el cierre de la bóveda
tal y como ocurre en otras plantaciones de pino (Cortina 1992). Son esperables
entoces fenómenos de autotala, estabilización o disminución de la
productividad foliar, aumento considerable de la contribución de la fracción
tronco frente a copa en la biomasa de las especies... Ello t rae rá consigo la
disminución de la cantidad de N destinada a la retención (puesto que no
existiría incremento foliar) y una disminución en la absorción radicular de
este elemento..
En relación a A. decorticans (figura 7.3), puede argumentarse que esta
especie sí constituye una especie explotadora del N edáfico dados los elevados
costes nutricionales (59 Kg/ha de N acumulados en tan sólo 5 T/ha de peso
seco). Los requerimientos anuales suponen 29 Kg/ha. año destinados a la
formación de estructuras todas ellas muy ricas en N (hojas, tallos incluso
madera). A ello debe añadirse el coste reproductivo, muy irregular durante el
periodo de muestreo pero que supone como media un 227. de los requerimientos
totales. El bajo porcentaje de retraslocación que se detecta en esta especie
t rae como consecuencia una fuerte absorción radicular (27 Kg/ha. año) extraídos
de una reserva muy limitada de N disponible en el suelo. Parte de esta
absorción puede ser suplida por el rápido retorno de N vía desfronde (un 387.
de la mineralomasa total acumulada) seguida de una rápida mineralización que
comienza inmediatamente después de que la hojarasca llegue al suelo. A par t i r
de ' los datos obtenido en el capítulo 5 se calcula que al cabo de poco más de
un año el 507. del N contenido en el desfronde foliar anual ha sido
mineralizado.
Por su parte se ha detectado absorción foliar de N-NO en esta especie
mientras que el N orgánico y el aminio se lavan o lixivian mediante
trascolación neta proporcionando cantidades adicionales de N al suelo.
Las entradas por fijación aún siendo bajas: (1 Kg/ha.año) como máximo, no
son despreciables dado que constituyen una entrada de N directamente
disponible a suelos potencialmente escasos en este elemento.
402
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
N
0.6
ABSORCIÓN FOLIAR DEP. GLOBAL 6.5
BIOMASA 5 9 . 6 3t> « \>0* RETENCIÓN 5.11
010,0 J n
LflUflDO DEPOSICIÓN SECA ?
-LIXIUIRDO ? o o
TRASCOLACION 1 . 0 8
DESFRONDE 22
FU. N:
IjUnifrlft-htf M M ^ I M ^ W IhWfrtffllfifftiT tofrVHK ^
C0.5-1 .21
* f 27 .5
ABSORCIÓN
SUELO DISPONIBLE
MINERALIZACION
MANTILLO 42
SUELO MINERAL
C67-1673? AVENAMIENTO
1.15
1673
Figura 7.3. Flujos biogeoquímicos para A.decorticans en una de las zonas de la
cuenca (RPN): Nitrógeno. Flujos en Kg/ha-año. Compartimentos en Kg/ha.
403
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
C. laurifolius (figura 7.4) es mucho más frugal con respecto al nitrógeno
y además eficiente en su uso. Los costes nutricionales son un 50% inferiores a
los de A. decorticans. Todas las formas de N presentes en la lluvia son
absorbidas por las superficies foliares de la j a ra lo que supone una entrada
directa de 0.75 Kg/ha.año (un 12% de todo el N presente en la deposición
global). La existencia de retraslocación se detecta no sólo al comparar las
concentraciones foliares en órganos verdes y senescentes sino al realizar el
balance Requerimientos-Absorción. El porcentaje que supone esta redistribución
respecto a los requerimientos anuales es de un 45% valor similar a encontrados
en otros arbustos mediterráneos (Gray 1983).
Un 8.5% de N acumulado en la biomasa del jaral es devuelta anualmente al
suelo mediante desfronde. Este N es, no obstante inmovilizado, al igual que
ocurre en el pinar, durante los primeros dos años (capítulo 5). Las
estructuras perennes también inmovilizan anualmente 1.6 Kg/ha.año que sólo
supone el 0.09 % del N total del suelo mineral mientras que la absorción
supone un 0.20% del N del suelo mineral y entre un 2-5 % del disponible.
Considerando ahora los flujos conjuntos de las t res especies presentes
en cada zona obtenemos los diagramas de la figura 7.5 y 7.6.
En RPN (figura 7.5), el 8.3 % del N total del sistema (155 Kg/ha) se
incorpora a la biomasa aérea en estos primeros trece años. Ello implica un
coste nutricional elevado determinado básicamente por la coexistencia de un
matorral con consumo de lujo para el N y un pinar que se encuentra en una
etapa de al ta demanda nutricional próxima ya al cierre de la bóveda.
Las entradas de N por deposición global y trascolación neta suponen en
esta zona 8 Kg/ha.año principalmente en forma disponible (N03 y NH4).
Descontando las salidas anuales de avenamiento la vía hidrológica aporta al
suelo 6.9 Kg/ha.año que suponen un porcentaje nada despreciable de la
absorción radicular (15 %). No obstante, la precipitación durante el periodo
de muestreo ha estado por encima de la media anual (450-500 mm), por lo que
estos aportes podrían verse fuertemente reducidos en año de mayor escasez de
404
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
N o
0£
0 " ^ 4 BIOMASA 2 1 . 5 o*0 « M>0 RETENCIÓN 1.6
TRASL0CACI0N
0 . 7 5 ABSORCIÓN FOLIAR DEP. GLOBAL
6.5
LAUADO DEPOSICIÓN SECA 0
. —LIXIUIADO 0
TRASCOLACION 0
DESFRONDE 1.79-
frxs W f w v w u r H j J rtl. .ts^th ' / .w CN ^ s > *>, ^ . v . o ^ i ^ í ^ W J s v :
° ^ V A * ^ 9 o / V o
I i iñ
ABSORCIÓN
1 SUELO
DISPONIBLE
4 i
4
. ^ T * O -" O
nlNERnUIZnCION i
MANTILLO 4 2
> r
SUELO MINERAL
C 6 7 - 1 6 7 J AVENAMIENTO 1.15
1 6 7 3
F i g u r a 7 .4 . Flujos biogeoquímicos p a r a C.laurifolias en una de las zonas de la
cuenca (RPN): Nitrógeno. Flujos en Kg/ha^año. Compart imentos en Kg/ha.
405
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
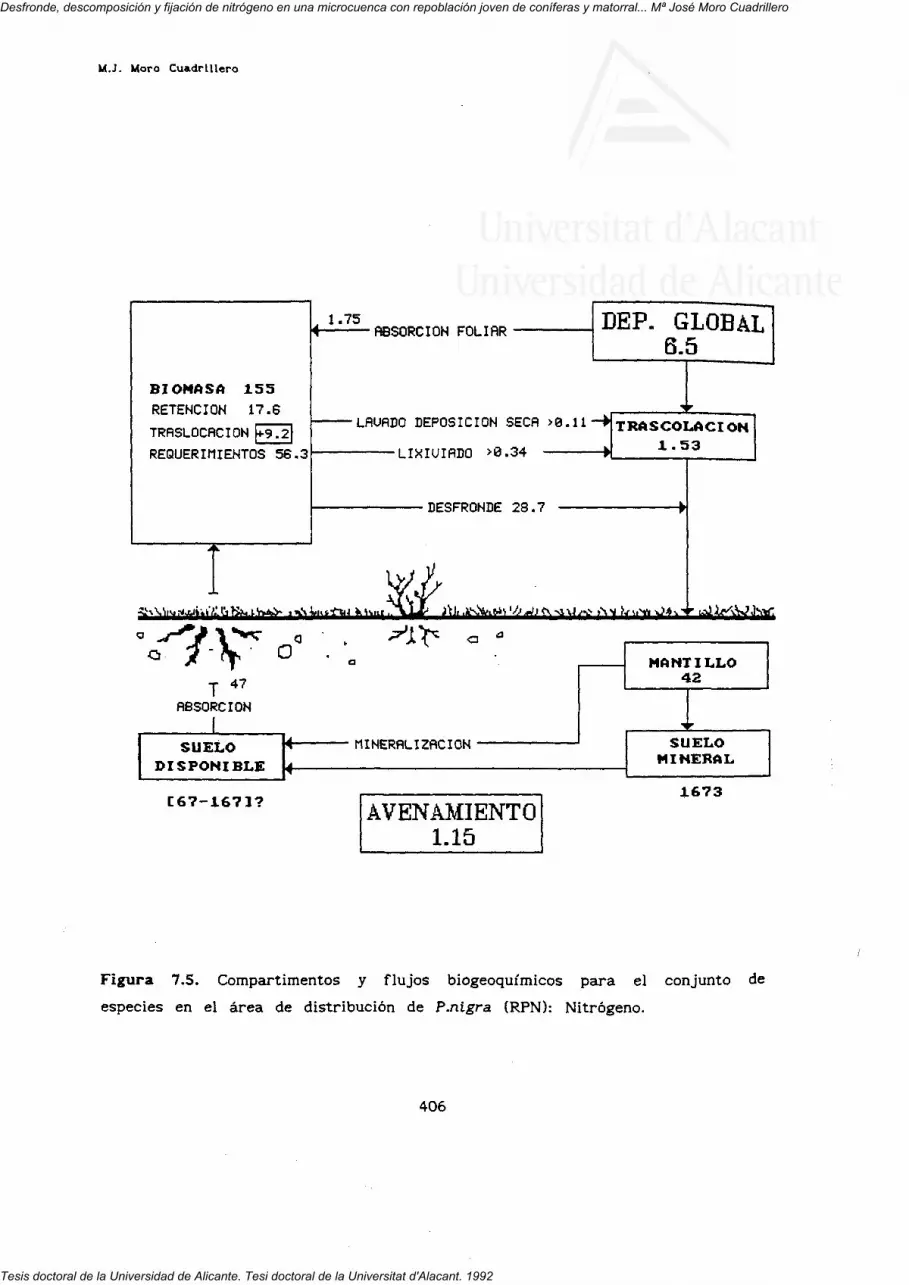
M.J. Moro Cuadrillero
„ 1.75 i ABSORCIÓN FOLIAR
BIOMASA 1 5 5 RETENCIÓN 17.6
TRASLOCACION 4-9.2
REQUERIMIENTOS 56.3
I
DEP. GLOBAL 6.5
LAUADO DEPOSICIÓN SECA > 8 . 1 1 -
LIXIUIADO >0.34 •
TRASCOLACIÓN 1 . 5 3
DESFRONDE 28.7
•^T.Mivi*j.JÍ*>V¿lj'K»<.tV>fr> i n S 4 » u f f t i J fc i\L.ii>$tntH,/t*»<\*¿\iso >\s ^si^ o ^ 4 * ioíle*&J 3s*r 3 ^V\V a o- /-V- °
> T 47 1
ABSORCIÓN
1 SUELO
D I S P O N I B L E
A
A i
^XV- <=> " a
rlINERALIZACION • "
MANTILLO 4 2
i r
SUELO MINERAL
C 6 7 - 1 6 7 1 ? AVENAMIENTO 1.15
1 6 7 3
Figura 7 .5 . Compart imentos y f lujos biogeoquímicos p a r a el conjunto de
especies en el á r e a de distr ibución de P.nigra (RPN): Nitrógeno.
406
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

lluvias.
Flujo»
Por desfronde se devuelve un 187. del total acumulado (28 Kg/ha.año) de los
cuales 22,(75%) corresponden a A. decorticans. Las otras dos especies
reutilizan parte del N mediante mecanismos de traslocación, lo que implica en
definitiva que ese N no se transfiera a corto plazo a los otros
compartimentos. En este sentido la presencia del gaznacho puede resultar
especialmente favorable para transpasar considerables cantidades de N al
mantillo que, a su vez, son mineralizados de forma rápida. Así, el suelo
mineral adquiere de forma continua y rápida N mineral que queda a disposición
del sistema radicular de la vegetación, bien sea el propio Adenocarpus o
cualquiera de las otras especies. Como contrapartida, la hojarasca más
empobrecida del jaral y pinar constituye una reserva de N potencialmente
disponible a largo plazo (dado que la mineralización no comienza por lo menos
hasta pasados dos años de la caída). Además la materia orgánica de esta
hojarasca de pino y ja ra que pierde peso lentamente, tendría un efecto
positivo respecto a la retención de agua del suelo favoreciendo la disolución
de los nutrientes en general y por tanto su disponibilidad.
En RPP (figura 7.6), un 8.3 % del N total del sistema se acumula en la
biomasa aérea que además, tiene un coste nutricional elevado. Por vía
hidrológica están disponibles 7 Kg/ha año (N03-N, N-NH4) lo que supondría un
buen porcentaje de la absorción radicular (20%). La menor influencia de A.
decorticans en RPP se manifiesta sobre todo en la magnitud del N retornado por
esta especie que sólo supone en esta zona un 43% del desfronde total . Esto
permite disminuir considerablemente la magnitud de la absorción radicular pero
también las entradas de N fácilmente mineralizable al suelo.
Una cuestión que puede plantearse cuando se observan las figura 7.3 es
hasta que punto es positivo mantener en la cuenca el matorral de A.
decorticans que aunque retorne considerables cantidades de N fácilmente
mineralizables y fije nitrógeno atmosférico, tiene la mayor demanda
nutricional de las especies estudiadas. Desconocemos por completo el grado de
competencia por los nutrientes que puede establecerse entre el pinar y
Adenocarpus y es difícil sacar conclusiones al respecto. Pero en relación a lo
407
i
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
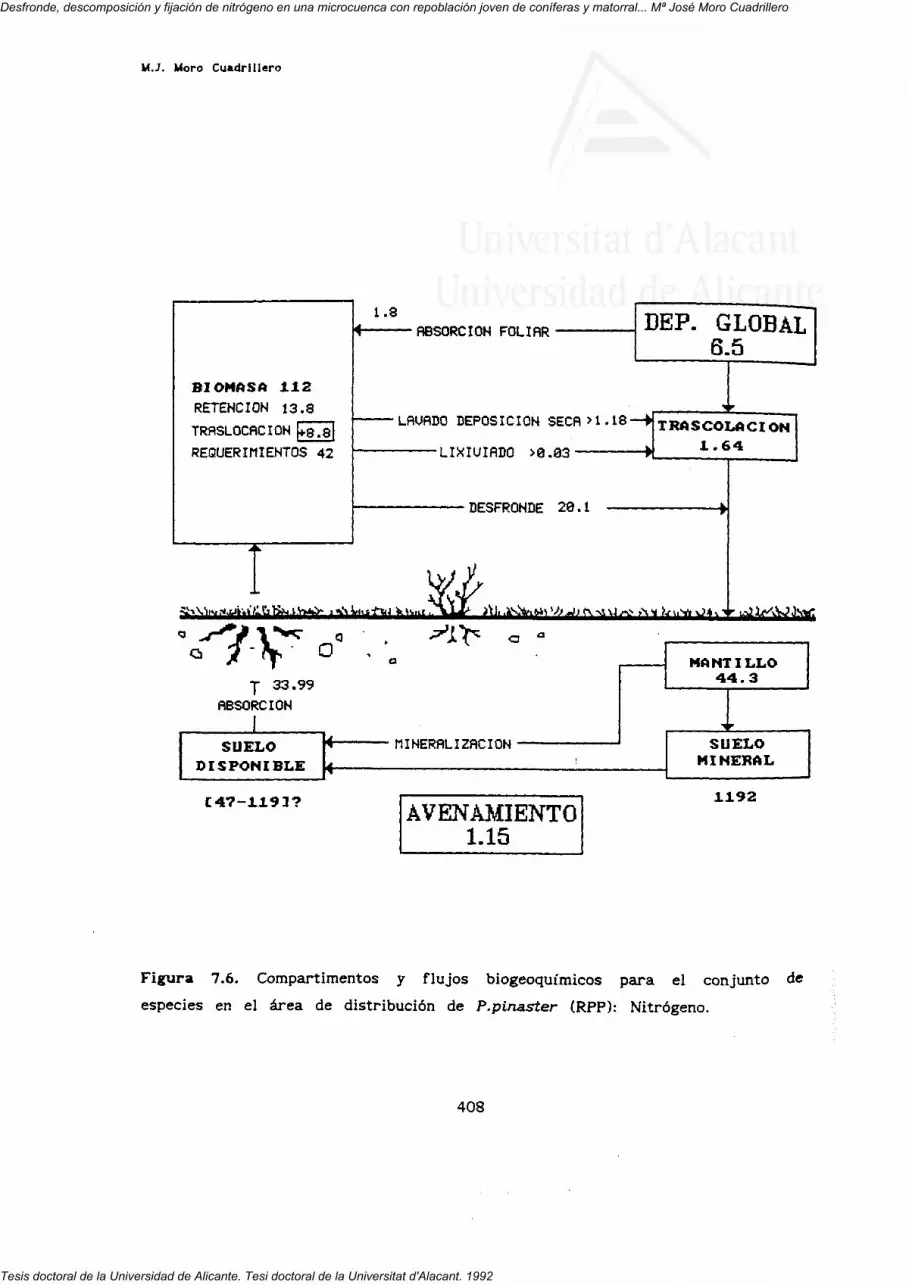
M.J. Moro Cuadrillero
BIOMASA 1 1 2 RETENCIÓN 13.8
TRASLOCACION +8.8
REQUERIMIENTOS 42
T
1.8 ABSORCIÓN FOLIAR DEP. GLOBAL
6.5
LAUADO DEPOSICIÓN SECA > 1 . 1 8 -
LIXIUIADO >0.83 •
TRASCOLACIÓN 1 . 6 4
DESFRONDE 2 9 . 1
T 33.99
ABSORCIÓN
I SUELO
DISPONIBLE
MINERALIZACION
MANTILLO 4 4 . 3
I SUELO
MINERAL
C 47-1 .19 3? AVENAMIENTO 1.15
1 1 9 2
Figura 7.6. Compartimentos y f lujos biogeoquímicos p a r a el conjunto de
especies en el á r ea de distr ibución de P.pinaster (RPP): Nitrógeno.
408
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
comentado en el párrafo anterior, la presencia de una especie que toma y
devuelve nutrientes de forma rápida en forma disponible parece en principio
ejercer más efectos positivos que negativos. Una tala en el momento actual
supondría la pérdida de esta importante reserva. Si el manejo provoca además
erosión de los horizontes orgánicos y de parte de suelo mineral
empobreciéndolos de un elemento ya escaso puede llegarse a una situación de
disminución de fertilidad precisamente en una etapa en que la demanda
nutricional del pinar se encuentra en los niveles máximos.
Fósforo.
La principal vía de entrada de P a los ecosistemas es la meteorización,
flujo no cuantificado en el presente trabajo. Las entradas vía hidrológica son
escasas (0.17 kg/ha.año) pero quedan prácticamente en su totalidad retenidas
por el sistema ya que no se ha detectado presencia de P disuelto en el agua de
avenamiento.
En P. nigra (figura 7.7) la trascolación neta tiene signo negativo, lo que
significa que parte del P04 de la lluvia es absorbido por las superficies
foliares o por flora-fauna epífita asociada. Si bien la magnitud de esta
absorción es pequeña (0.07 Kg/ha.año) supone un 41 7. del contenido en la
deposición global y un 37. de los requerimientos anuales de P en esta especie.
La inmovilización de P en el material perenne y acículas (0.53 Kg/ha.año)
supone un 37. del P asimilable y la absorción un 4.8 7.. Al igual que para el
nitrógeno, la reutilización de P por el pino laricio es altamente eficaz
estimándose en un 25 % de los requerimientos. El P retornado vía desfronde
constituye el 67. de la acumulación en biomasa del cual aproximadamente un 307.
se mineraliza durante el primer año de descomposición.
En P. pínaster (figura 7.8), la magnitud de los flujos es similar a P.
nigra.. Es de destacar que a pesar de que los requerimientos en el pino
resinero son sólo un 24% superiores a los del pino laricio, la absorción
radicular es del orden de un 347. más elevada lo que implica una menor
importancia de la redistribución interna en P. pinaster en relación a P. nigra
(un 137. de los requerimientos).
409
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
0.87 ABSORCIÓN FOLIAR
BIOMASA 7 . 6 RETENCIÓN 1.16
TRflSLOCflCION t-0 31 4
DEP. GLOBAL 0.17
W\Tf,.-v REQUERIMIENTOS2.17 , £ , LAUADO DEPOSICIÓN SECA 0.0
LIXIUIADO 0.0
TJMSCOLACION
DESFRONDE 0 .45-
f \ fe.!.-*» ,%su«-rtrf fcKKrHlJ J U .isifa>h ' ¿ w j f w i i / * As k».** o * , + •^Ir^Vw'Jsw: 3 -^aV\"v,r a Q / - Y ' °
-f 1.61 1
ABSORCIÓN
1 SUELO
DISPONIBLE
A
A 1
. *VF o -' <3
... .... niNERHLIZHCION i
MANTILLO 2 . 0 4
l r
SUELO MINERAL
3 3 AVENAMIENTO
0
F i g u r a 7.7. Flujos biogeoquímicos p a r a P.nigra en su á r e a de distr ibución:
Fósforo. Flujos en Kg/ha -año . Compart imentos en Kg/ha .
410
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
P
.J^CBIOMASA 6 . 7 í ^ . f í fy < RETENCIÓN 1.7 ' ' '4¡<J
3 * TRASLOCACION Í-0
ABSORCIÓN FOLIAR 0.06
LAUADO DEPOSICIÓN SECA 0
DEP. GLOBAL 0.17
LIXIUIADO 0
TRASCOLACION 0
DESFRONDE 0.76
i\l,.V<!tn.<A\[¿*l)<\^¡.\Íj<? >\* k M ^ o * ^ i¿lle<'-&Jis-*r.
» - ^ V \ * ^ =1 « / - y ° 1
T ABSORCIÓN 2 . 4 6
1 SUELO
DISPONIBLE
*
^
^l> o -' <a
"" nINfcRALIZnCION
MANTILLO 2 . 8 2
•< r
SUELO MINERAL
2 0 AVENAMIENTO
0
Figura 7.8. Flujos biogeoquímicos para P.pinaster en su área de distribución:
Fósforo. Flujos en Kg/ha-año. Compartimentos en Kg/ha.
411
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
La absorción foliar directa del fosfato de la lluvia es también una
característica que comparte esta especie con P. nigra y que a su
vez diferencia a los dos pinos del matorral en los que la trascolación neta es
positiva para este ion.
El P en C. laurífolius es menos eficazmente utilizado circulando de forma
rápida en esta especie. No existe absorción de PCH del agua de lluvia pero sí
lixiviación y lavado de deposición seca (figura 7.9). A pesar de la existencia
de retraslocación (22% de los requerimientos) se retorna por desfronde un 147.
del P total acumulado en la biomasa aérea. Este P sufre una mineralización muy
rápida durante el primer año (un 80% del P contenido en el desfronde foliar)
por lo que esta especie constituye una importante fuente de P rápidamente
disponible.
La circulación de P es también rápida en la otra especie arbustiva (figura
7.10). Vía trascolación neta se adicionan al suelo 0.16 Kg/ha.año de P04-P que
unidos a los 0.17 de la deposición global suponen un 23% del fósforo que esta
especie absorbe por la raiz. El retorno de P mediante el desfronde es también
notable: algo más de un 44 % del P acumulado en biomasa se adiciona anualmente
al mantillo, fósforo que se mineraliza lentamente durante el primer año y cuya
liberación en forma disponible se acelera durante el segundo de forma que
transcurridos unos 600 días (capítulo 5) un 60% del P retornado el año
anterior se encuentra en forma inorgánica. Los niveles de retraslocación son
muy bajos y pasan fácilmente a signo negativo en años con menor esfuerzo
reproductivo. Al igual que para el N, el P movilizado por parte de A.
decorticans entra en un circuito más completo (vegetacion-mantillo-suelo) en
relación a las otras especies en las cuales la reutilización del recurso
mineral mediante mecanismos internos impide en buena parte la transferencia a
corto plazo de un porcentaje más o menos alto del nutriente hacia otros
compartimentos.
Conjuntamente, el P acumulado por las t res especies en RPN (figura 7.11)
constituye un rico almacén en relación al P disponible en los treinta cm del
suelo (36%). Las entradas atmosféricas y de trascolación neta aportan un 12.5%
de las necesidades de absorción y éstas a su vez suponen alrededor del 10 %
412
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
8 ABSORCIÓN FOLIAR DEP. GLOBAL
0.17
o'O 9 \ L RETENCIÓN 0.2
LAUADO DEPOSICIÓN SECA 0.04-
O 0 n — LIXIUIflDO 0.07
TRASCOLACION 0 . 1 1
' -r^f I""* i o f \ O ' F
| a . 59 ABSORCIÓN
1 SUELO
D I S P O N I B L E
A
A ^
^l> o -
nlNERHLIZHCION
MANTILLO 2 . 0 4
1 r
SUELO , MINERAL
3 3
AVENAMIENTO 0
Figura 7.9. Flujos biogeoquímicos para C.laurifolius en una de las zonas
de la cuenca (RPN): Fósforo. Flujos en Kg/ha*año. Compartimentos en Kg/ha.
413
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
0 ABSORCIÓN FOLIAR DEP. GLOBAL
0.17
ero „ \L RETENCIÓN 8-25 O o
LAUADO DEPOSICIÓN SECA ?
-LIXIUIABO ?
TRASCOLACION 0 . 1 6
o o
DESFRONDE 0.88
f 1.3
ABSORCIÓN
SUELO DISPONIBLE
MINERALIZACION
MANTILLO 2.04
SUELO MINERAL
33 AVENAMIENTO
0
Figura 7.10. Flujos biogeoquímicos para A.decorticans en una de las zonas de
la cuenca (RPN): Fósforo. Flujos en Kg/ha-año. Compartimentos en Kg/ha.
414
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
0.07 i ABSORCIÓN FOLIAR
BIOMASA 1 1 . 8 7 RETENCIÓN 1.81
TRRSLOCRCION t-0.8
REQUERIMIENTOS 4.33
T
DEP. GLOBAL 0.17
LRURDO DEPOSICIÓN SECA >0.84 - •
LIXIUIRDO >0.07 •
TRASCOLACION 0 . 2 ?
1
DESFRONDE 1.65
3SUw.!«*iiuri«,K».iw¿. ,»sUrfui w s w . X j J i K . ^ V ^ ' ^ m N i v i ^ * * l ^ o * , 4 uJ^AJJ**: 0 ^J\^ < * /V °
T 3 . S 1
ABSORCIÓN
1 SUELO
DISPONIBLE
A
A *
. *$? o * a
nlNERRLIZRCIOH
MANTILLO 2 . 0 4
^ r
SUELO MINERAL
3 3 AVENAMIENTO
0
Figura 7.11. Compart imentos y f lujos biogeoquímicos p a r a el conjunto de
especies en el á r e a de distr ibución de P.nigra (RPN): Fósforo.
415
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
del P asimilable. Un 19 % de los requerimientos totales del sistema se
suplirían mediante mecanismos internos de retraslocación desarrollados
esencialmente por el pinar que permiten limitar la extracción de P del suelo.
La influencia de la vegetación arbustiva (tanto j a ra como gaznacho)
tendría un efecto positivo sobre el suelo dadas las rápidas tasas de
mineralización de P que aparecen en ambas especies sobre todo en C.
laurifolius. La eliminación del matorral t raer ía consigo una ralentización del
flujo de retorno y de mineralización. Suponiendo una tasa idéntica de
absorción radicular en P. nigra esta eliminación equivaldría a reducir la
extracción de P edáfico en un 54%. Como contrapartida, se eliminaría la
principal vía de circulación rápida de P llevada a cabo por los dos arbustos.
Así, el desfronde del estrato arbustivo proporciona al suelo una hojarasca
cuyas características permiten una rápida mineralización de P durante el
primer año y medio calculado en un 70 y 50% en la j a r a y el gaznacho
respectivamente. A esta rápida circulación vía orgánica puede unírsele también
el enriquecimiento positivo que sufre el agua de lluvia al atravesar la
cubierta de ambas especies. Por tanto, al igual que ocurría con el N, la tala
del matorral conllevaría la desaparición de un importante almacén conectado
con el sistema mantillo-suelo mediante flujos rápidos de circulación. Si bien
el pinar tiene sus propios mecanismos conservativos, la no renovación a corto
plazo del P disponible en suelo podría provocar una disminución de las
reservas y hacerse incluso limitante para su crecimiento de mantenerse las
mismas tasas de absorción.
La situación cambiaría muy poco al considerar el otro área (RPP) (figura
7.12) aunque existan ligeras diferencias cuantitativas, por lo que son válidas
las mismas consideraciones que para RPN.
Calcio.
A part i r de los depósitos de Ca intercambiable en el suelo mineral,
mantilllo y de las entradas por deposición global y su comparación con su
acumulación en la biomasa área tanto de forma individual como conjunta
(figuras 7.13 a 7.18), se deduce que este elemento no ha de ser limitante en
416
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
BIOMASA 1 0 . 5 RETENCIÓN 2 . 1 3
TRASLOCACION f0.8
REQUERIMIENTOS 4.8
T
. 8.86 4 ABSORCIÓN FOLIAR
DEP. GLOBAL 0.17
LAUADO DEPOSICIÓN SECA > 0 . 0 7 -
LIXIUIADO >0 .13 •
TRASCOLACION 0 . 2 6
DESFRONDE 1.7
^foftritfi'ifilftiihh'fr nhktfw hv iM\ i^w ' wm-^rr fn h\w yh t wilVfríftiti ° ^y \ * ^ q o / V 0
T 4 1 ^
ABSORCIÓN
1 SUELO
DISPONIBLE
A
4 i
^IT* o -a
nlNtRnLIZnCIUN
MANTILLO 2 . 8
•< r
SUELO MINERAL
2 0 AVENAMIENTO 0
Figura 7.12. Compartimentos y f lujos biogeoquímicos p a r a el conjunto de
especies en el á r e a de distr ibución de P.pinaster (RPP): Fósforo.
417
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
el sistema. La exportación hidrológica de la cuenca supera a las entradas pero
sólo en una baja proporción (un 117.) y su magnitud es una pequeñísima
proporción del Ca disponible los primeros cm del suelo (0.90 y 0.75 7. en RPN y
RPP respectivamente (figura 7.17 y 7.18). La vía de meteorización es
igualmente una fuente de entrada de Ca. Aunque no se ha calculado en el
presente trabajo, Lledó (1990) comenta que puede hacerse una aproximación
estimándola como la diferencia entre entradas al suelo menos salidas por
avenamiento suponiendo que el Ca disponible se mantenga constante. Según este
método este flujo aportaría unos 38 Kg/ha.año en RPN y 45 Kg/ha.año en RPP.
Existen considerables entradas de Ca en deposición global (15 Kg/ha.año),
buena parte vía deposición seca, entradas que son superiores a cualquier otro
ion presente en el agua de lluvia en la cuenca "Nacimiento" (Domingo 1991).
No existen en ninguna de las especies mecanismos que reutilizen o
aprovechen el Ca (retraslocación, absorción foliar) y la absorción supone un
excedente en relación a los requerimientos más patente en RPP en cuanto
funcionamiento global y en P. pinaster en cuanto a especie individual (figura
7.14) debido al elevado desfronde de esta especie que por otro lado es muy
rico en Ca.
Comparación de las cuatro especies.
Para concluir hemos considerado oportuno a modo de resumen, expresar
algunos de los compartimentos y flujos más característicos en las especies
estudiadas de una forma normalizada para facilitar la comparación. Para ello
hemos construido la tabla 7.9 en la que los valores de cada flujo se han
expresado en gramos de nutriente por kilogramo de biomasa aérea y año. También
se han calculado algunos cocientes indicativos del funcionamiento nutricional
de las especies estudiadas.
Los flujos que están relacionados con la incorporación de nutrientes al
suelo (desfronde, trascolación neta, retorno y los derivados de ellos
(absorción) son sistemáticamente más elevados en el matorral para los
elementos considerados. Las diferencias entre las especies arbustivas en este
tipo de flujos varían bastante dependiendo del tipo de nutriente. Las más
418
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujot
ABSORCIÓN FOLIAR DEP. GLOBAL 15
- t í " ^ < RETENCIÓN 5 - 1 3 ,,><,{<f
LAUADO DEPOSICIÓN SECA 9 «6
-LIXIUIALO 0.51
TRASCOLACIÓN 10.14
3 -rfV** a o / -V- o ABSORCIÓN
1 SUELO
DISPONIBLE
4
A
^
-^í* o * ' o
"" " ruNERnLIZnCIQN ""
MANTILLO 1 8 3
y
SUELO MINERAL
2259 AVENAMIENTO
17
Figura 7.13. Flujos biogeoquímicos para P.nigra en su área de distribución:
Calcio. Flujos en Kg/ha-año. Compartimentos en Kg/ha.
419
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Ca
BIOMASA 6 6 Í C RETENCIÓN 5 .5 , | '< i? ' ' "
p-C*Vj- < RETENCIÓN 5 .5 ' J Í V
tajT^ i/* TRASLOCAC ION p r r i í« 'ví'lf.v'í- REQUERIMIENTOS 15.7"".' ~
ABSORCIÓN FOLIAR DEP. GLOBAL 15
LAUADO DEPOSICIÓN SECA 3.89
LIXIUIADO 7 .7 •
DESFRONDE 20.25-
TRASCOLACIÓN 1 1 . 6
U . l s W ^ ^ t w o ^ ^ l ^ o * , + ^^xVjJsw:
o / ^ O t" 33 .4
ABSORCIÓN
SUELO DISPONIBLE
MINERALI2ACI0N
MANTILLO 1 0 6
SUELO MINERAL
1 8 7 7 AVENAMIENTO
17
F i g u r a 7.14. Flujos bíogeoquímicos p a r a P.pinaster en su á r e a de distr ibución:
Calcio. Flujos en Kg/ha-afio. Compart imentos en Kg/ha .
420
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
Ca O O O © "
a o ^ V o u . • ** i ' * / • J A o o
ABSORCIÓN FOLIAR DEP. GLOBAL 15
aí,0 °
BIOMASA 2 9 LAUflDO DEPOSICIÓN SECA 0.33-
rt „ \J„ RETENCIÓN 2.3 CÍO 0 *A0 oo —LIXIUIflDO 0.62
TRASCOLACION 0 . 9 7
DESFRONDE 5.35
ÍkiLbtaCÍ¿¿ UxMurfm Vi-.uiTvU' ^li.)í^»i^'/>wif\si\J^%> >\s ^^it v ) | s v L^íi^vWJ^j: 0 -<-/V\"v^ o o / - y O
T 8.5 ABSORCIÓN
1 SUELO
D I S P O N I B L E
A
A %
**XX* o -' O
"" " nlNfcRRLIZnCIQN
MANTILLO 1 0 3
•» r
S U E L O MINERAL
2259 AVENAMIENTO
17
Figura 7.15. Flujos biogeoquímicos p a r a C.laurifolius en una de las zonas de
la cuenca (RPN): Calcio. Flujos en Kg /ha -año . Compart imentos en Kg/ha .
421
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
0 ABSORCIÓN FOLIAR DEP. GLOBAL
15
BIOMASA 1 5 90 0 Mi/ RETENCION 1 -25
TRASLOCRCION
LAUADO DEPOSICIÓN SECA S.2
oo - —LIXIUIflDO 0.4
TRASCOLACION 6 . 6
a ^/X*^ a o / \ o
f 10.34 1
ABSORCIÓN
1 SUELO
DISPONIBLE
i f 4 i
^ T * a -' O MANTILLO
1 0 3
> ' SUELO
MINERAL
2 2 5 9 AVENAMIENTO
17
Figura 7.16. Flujos biogeoquímicos para A.decorticans en una de las zonas de
la cuenca (RPN): Calcio. Flujos en Kg/ha-año. Compartimentos en Kg/ha.
422
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujoi
BIOMASA 93 RETENCIÓN 8 .9
TRASLOCflCION -9
REQUERIMIENTOS 24.5
T
ABSORCIÓN FOLIAR DEP. GLOBAL 15
LflUflDO DEPOSICIÓN SECñ 16
LIXIUIñDO 1.5
TRASCOLACION 1 7 . 5
DESFRONDE 23
y 33.4
ABSORCIÓN
SUELO DISPONIBLE
MINERALIZACION
MANTILLO 183
SUELO MINERAL
2259 AVENAMIENTO
17
Figura 7.17. Compartimentos y flujos biogeoquímicos para el conjunto de
especies en el área de distribución de P.nigra (RPN): Calcio.
423
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992
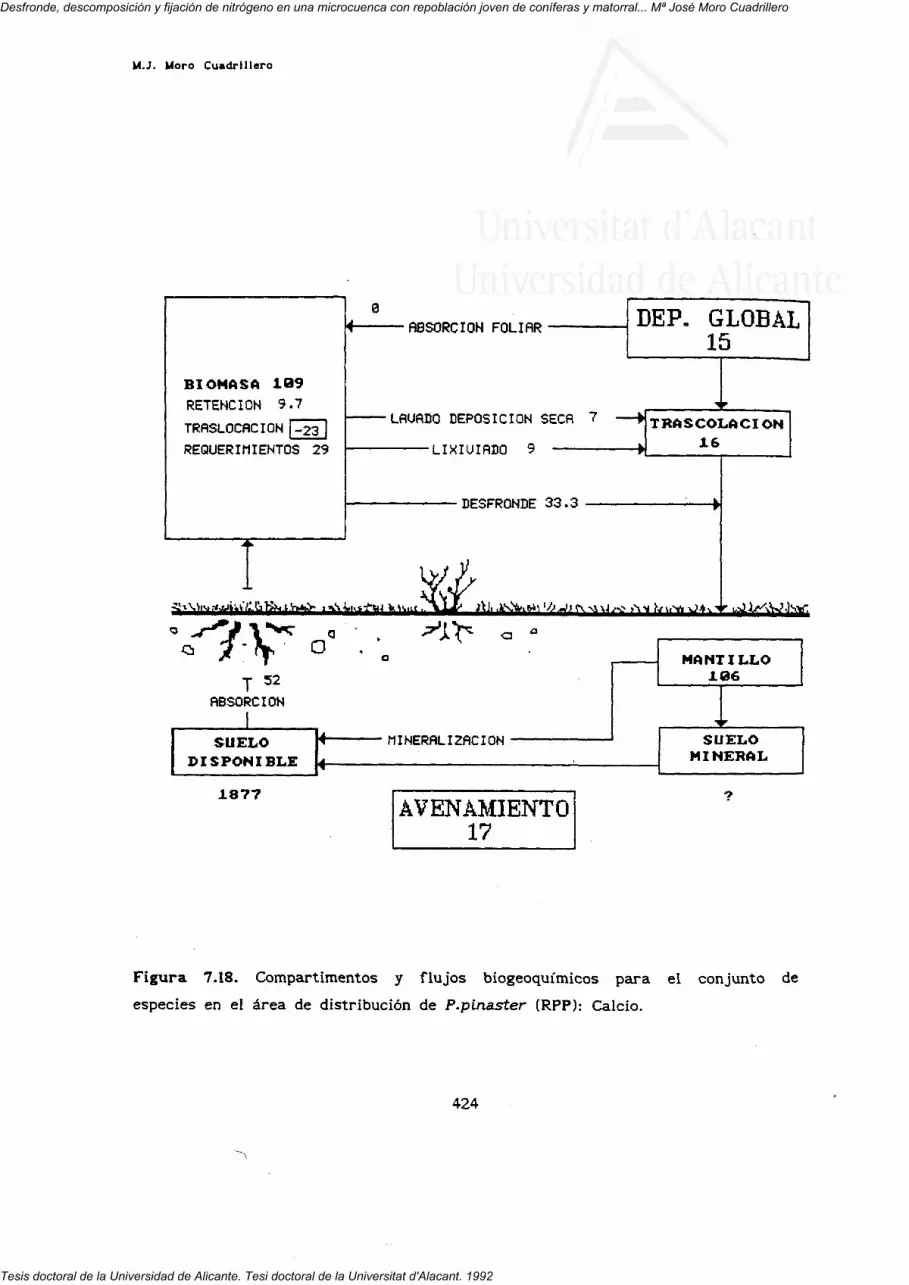
M.J. Moro Cuadrillero
BIOMASA 1 8 9 RETENCIÓN 9 . 7
TRASLOCACION -23
REQUERIMIENTOS 29
T
ABSORCIÓN FOLIAR DEP. GLOBAL 15
LAUADO DEPOSICIÓN SECA 7
LIXIUIADO 9
TRASCOLACIÓN 1 6
DESFRONDE 3 3 . 3
T 5 2
ABSORCIÓN
SUELO DISPONIBLE
MINERALIZACION
MANTILLO 1 0 6
SUELO MINERAL
1 8 7 7 AVENAMIENTO
17
Figura 7.18. Compartimentos y f lujos biogeoquímicos p a r a el conjunto de
especies en el á r ea de distr ibución de P.pinaster (RPP): Calcio.
424
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

TULA 7.9. Eficiencia de los flujos en cada especie,
g de NUTRIENTE/Kg de BIOMASA.AÑO
NITRÓGENO
T NETA
O.SECA
LIXIVIADO
ABS.FOLIAR
RETENCIÓN
OESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
RETRASLOCACION
MINERALOMASA
df/min
rtl/req
min/retorn
df/tn
min/reten
FOSFORO
T NETA
O.SECA
LIXIVIADO
ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
RETRASLOCACION
MINERALOMASA
df/min
rtl/req
min/retorn
df/tn
min/reten
PP PN
.073
.072
.002
.015
.565
.485
.487 1.051 1.236
.185 3.69
.026
.006
.020
.023
.640
.287
.306
.947 1.240
.293 4.33
.13 .07
.15 .24
7.58 14.12
6.61 10.89
6.54 6.76
PN
0 0 0
.003
.103
.046
.046
.149
.175
.025 .41
0 0 0
.004
.068
.026
.026
.094
.127
.033 .45
.11 .06
.15 .26
8.82 17
3.94 6.59
AD CL
.221 0
-.035 0
0
.088 .242 1.045 .513 4.579 .577 4.579 .577 5.624 1.090 5.928 1.977
.305 .887 12.188 6.935
.38 .08
.05 .45
2.66 12.01
20.73
11.66 13.52
AD CL
.033 --0
.051
.180
.213
.264
.284
.020
.405
.035
.013
.023 0
.064
.103
.126
.189
.247
.058
.739
.44 .14
.07 .23
1.90 5.87
5.50 2.91
7.92 11.62
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
TABLA 7.9. (CONT).
POTASIO PP PN AD CL
T NETA
D.SECA
LIXIVIADO ABS.FOLIAR
RETENCIÓN
DESFRONOE
RETORNO
ABSORCIÓN
REQUERIMIENTOS RETRASLOCACION
MINERALOMASA
df/min
rtl/req
min/retorn
df/tn
min/reten
CALCIO
T NETA
D.SECA LIXIVIADO
ABS.FOLIAR RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
RETRASLOCACION
MINERALOMASA
df/min
rtl/req
min/retorn
df/tn
min/reten
MAGNESIO
T NETA
D.SECA
LIXIVIADO
ABS.FOLIAR
RETENCIÓN
DESFRONDE
RETORNO
ABSORCIÓN
REQUERIMIENTOS
RETRASLOCACION
MINERALOMASA df/min
rtl/req min/retorn df/tn
mi n/reten
.136
.060
.076
0 .352
.128
.204
.556
.733
.177
1.99
.06
.24 9.73
.94 5.65
PP
.703
.236
.467
0 .330
1.227
1.694
2.024
.952
-1.073
3.98
.31 -1.13
2.35
1.75
12.03
PP
.116
.018
.098
0 .123
.230
.328
.451
.341
-.110
.96
.24 -.32
2.93
1.98
7.86
.118
.087 -.
.210
.060
.060
.270
.361
.091
1.25
.05
.25 20.78
.51 5.97
PN
.593
.563
.030
0 .300
.522
.552
.852
.709
-.143
2.86
.18 -.20
5.18
.88 9.53
PN
.070
.063
.006
0 .111
.103
.109
.221
.195
-.025
.75
.14 -.13
6.84
1.48
6.72
1.885
1.033
.855
0 .458
1.137
1.992
2.450
2.515
.065
4.090
.28
.03 2.05
.60 8.93
AD
1.350
1.272
.078
0 .256
1.777
1.859
2.115
1.219
-.896
2.986
.60 -.73
1.61
1.32
11.68
AD
.294
.221
.074
0
.074
.417
.491
.564
.503
-.061
.955
.44 -.12
1.95
1.42
12.97
.806
.400
.406
0
.231
.255
.658
.889
1.185
.297
2.155
.12
.25 3.27
.32 9.34
CL
.313
.106
.200
0
.803
1.726
1.929
2.732
2.074
-.658
9.232
.19 -.32
4.79
5.52
11.49
CL
.106
.039
.065
0
.072
.274
.342
.414
.372
-.042 07^
.30 -.11
2.70
2.58
12.88
426
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujoi
acusadas se refieren al N. A. decorticans retorna y absorbe respectivamente 8
y 5 veces más N que C. laurifolias. El gaznacho también desfronda, retorna y
absorbe más P, K y Mg que la jara . Las diferencias con respecto al K son del
orden de 2 a 4 veces mayores en A. decorticans y muy pequeñas en relación al P
y Mg. Para el Ca los valores son ligeramente superiores en la ja ra .
Los valores inferiores para los mismos flujos se han encontrado en P.
nigra. P. pinaster ocupa una posición intermedia entre el laricio y el
matorral. Las diferencias, a veces acusadas, entre los dos pinos respecto al
retorno y la absorción pueden ser un reflejo de distintos requerimientos
nutricionales para un determinado elemento. Así, el pino resinero posee
similares requerimientos por Kg de biomasa y año de N que el laricio y
superiores para P, Mg y sobre todo K lo que se refleja en las correspondientes
absorciones. Sin embargo ambas especies no difieren excesivamente en los
requerimientos de Ca pero ésto no se refleja de forma coherente al observar el
dato de absorción. Mientras que la formación de nuevas estructuras en P.
pinaster necesita solamente un 257. más de Ca que en P. nigra, la absorción
resulta un 577. superior en la primera especie. Dado que las retenciones son de
un orden similar (0.3 g de Ca/kg biomasa. año), la diferencia se encuentra
evidentemente en el elevado retorno existente en P.pinaster. Se ha comentado
en repetidas ocasiones que este elevado retorno se debe en buena parte a
causas accidentales. La llegada al suelo de un buen porcentaje de acículas
caídas en años anteriores añadidas al desfronde fenológico puede hacer
aumentar considerablemente la contribución de aquellos elementos que
precisamente tienden a concentrarse en el material de mayor edad (Ca y Mg).
Los nutrientes requeridos por Kg de biomasa y año son superiores en las
especies arbustivas frente al pinar. La fijadora necesita cantidades muy
superiores de N al resto de las especies incluida la jara. También es la que
mayor cantidad de P, K y Mg necesita. El Ca asociado a la PPN es superior en
C. laurifolius incluso respecto a las especies de pino. Los requerimientos de
Mg están bastante igualados en todas las especies.
La relación entre los nutrientes acumulados en biomasa aérea y su
incrementos anuales (retención) puede darnos una idea de la etapa sucesional
A 427
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en que se encuentran las especies. Los pinos están acumulando biomasa no sólo
perenne sino también foliar rica en nutrientes mientras que suponemos
condiciones en equilibrio (incremento biomasa foliar = 0) para el matorral. El
cociente MIN/RETEN es inferior en el pinar para todos los elementos excepto el
Ca indicando en este estrato una etapa sucesional mas temprana.
Los tiempos de renovación más altos (expresados en años como el cociente
entre MINERALOMASA/RETORNO) son los obtenidos en P. nigra para todos los
nutrientes seguidos generalamente por los de P.pinaster, especie en la que el
elevado retorno hace disminuir considerablemente los resultados obtenidos para
este parámetro en relación con P. nigra. El gaznacho, como era esperable,
renueva los nutrientes en un tiempo más corto que la demás especies siendo el
Ca y Mg los más rápidamente renovables (característica ya comentada en el cap
4) seguido del P K y N.
La interacción con el agua de lluvia permite también la observación de
diferencias específicas que fueron ya ampliamiente comentadas en el trabajo de
Domingo (1991) y Domingo et al (1991). Caben destacar las tasas superiores de
trascolación neta en el ganacho para todo los elementos excepto el fósforo,
las elevadas tasas de absorción foliar de N en todas su formas quínicas
(amonio, nitrato y orgánico) por parte de la jara , la existencia de absorción
foliar de P-P04 solamente en el pinar con valores prácticamente idénticos en
ambas especies, el mayor enriquecimiento en K que sufre el agua al atravesar
las cubiertas arbustivas y la tasa elevada de lavado de deposición seca de Ca
en A. decorticans, superior incluso a la del pinar al referirlo a unidad de
biomasa.
El esbozo del comportamiento biogeoquímico y con mas detalle del ciclo
interno de nutrientes que se ha presentado en este último capítulo ha sido un
intento de integrar toda la información disponible. Sin embargo dista mucho de
ser completo y exacto por varios motivos: No existe niguna información sobre
la biomasa subterránea, sobre la dinámica de las raices finas, N disponible en
el suelo y nutrientes totales en éste, meteorización etc. Por otro lado no
428
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Flujo»
coinciden exactamente en el tiempo los períodos de muestreo para algunos de
los flujos, la precisión con la que se muestrean los distintos compartimentos
o flujos no es la misma, las series temporales son cortas y un largo etc. Sin
embargo estos u otros problemas se encuentran en la mayoría de los trabajos
que sin embargo, realizan este tipo de esquemas para distintos ecosistemas.
El estudio que hemos llevado a cabo tiene la ventaja de que, todos los
resultados, independientemente de su mayor o menor precisión, se han obtenido
a partir de muéstreos "in situ" en el sentido que no se ha recurrido, salvo en
alguna ocasión puntual, a datos de bibliografía o a extrapolaciones de
parámetros calculados para otros lugares u otras especies que pueden
introducir, al aplicarlos a un área distinta de dónde se han obtenido, errores
considerables.
En el área de estudio han existido también una serie de llamémosle,
particularidades, que con todo seguridad han tenido mucha influencia en los
resultados. Citaremos la precipitación considerablemente más elevada que la
media en los años de muestreo, la problemática del desfronde, la
heterogeneidad de la cuenca y la existencia de un ecosistema tipo "mosaico" en
etapa juvenil cuyos parámetros pueden variar considerablemente en los próximos
años.
A pesar de esta problemática, pensamos que se han obtenido unos primeros
datos comparativos interesantes para especies representativas de coniferas,
fijadoras leguminosas y matorral colonizador en la zona mediterránea. A su
vez, la aproximación a nivel de cuenca permite evaluar de manera más o menos
precisa el comportamiento de un sistema tipo "mosaico" que si bien abundante
en toda España en general, ha sido muy poco estudiado en su integridad.
429
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
430
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y concludonet .
RESUMEN DE RESULTADOS.
CAPITULO 1.
La cuenca ha sido dividida en dos grandes áreas: Área de Pinus pinaster
(RPP) y área de Pinus nigra (RPN) definidas por el tipo de pino que las cubre
y por dos especies de matorral (Adenocarpus decortícans y Cistus laurifolius)
con estructura sensiblemente diferente en ambas áreas. Dentro de ellas pueden
establecerse distintos sectores que difieren en algún/os parámetros
estructurales. La no dominancia de un estrato arbóreo que no ha llegado a la
estabilización de las copas coexistiendo con un matorral de Adenocarpus y
Cistus que ocupa los espacios libres, dan a la vegetación el típico aspecto de
mosaico.
CAPITULO 2.
Las relaciones entre el peso de las diferentes fracciones en la biomasa
con respecto al diámetro del individuo en las especies estudiadas se ajustan a
ecuaciones potenciales (alométricas) siendo la alometría negativa en todos los
casos (coeficiente de alometría <3). Para la fracción foliar el coeficiente
alométrico no es significativamente diferente de 2 en ninguna de las especies.
Las mejores estimas se han obtenido para el peso total y peso de tronco en las
especies de pino y para las hojas en las especies arbustivas. Los coeficientes
alométricos respecto al peso de tronco en P. nigra son inferiores respecto a
los valores manejados en la bibliografía y parece estar causado por los
pequeños tamaños de los individuos utilizados (DBH< lOcm).
Existe una fuerte similitud en las pendientes de las ecuaciones
alométricas para una misma fracción entre las especies arbustivas. Dado que el
rango de diámetro basal manejado es muy parecido puede hablarse de una
relación tamaño peso muy similar en las dos especies.
La biomasa aérea ponderada para cada una de las dos grandes zonas de la
cuenca es similar: 22 t /ha en RPP y 25 t / ha en RPN. El pinar acumula como
media entre un 68 y 747. respectivamente de esta biomasa. A su vez, dentro de
cada zona se observa un cierto grado de heterogeneidad en la distribución de
ésta con sectores en que el pinar es claramente dominante (sector 1 y 6) y
431
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
sectores en los que el peso aéreo conjunto de los arbustos es superior al
pinar (sectores 3, 4 y 7).
La inmadurez de la plantación unida a una considerable longevidad de sus
acículas (cuatro años) son los principales factores que contribuyen al elevado
porcentaje que las acículas tienen respecto a la biomasa total (29 en
P.pinaster y 34 en P.nlgra). Los porcentajes relativos de las principales
fracciones son 29-25-45 para acículas-ramas tronco en P. pinaster y 34-29-37
en P. nigra resultados similares a otras plantaciones inmaduras de corta edad.
La fracción foliar del año supone, a su vez, un 30% de todas las acículas. En
el matorral las hojas acumulan un 23% y un 17% de la biomasa total en C.
Laurifolius y A. decorticans respectivamente.
Se describen pautas significativas en la evolución temporal de la
concentración de nutrientes en acículas y ramas de distinta edad.
Las concentraciones de Ca, N y K en las acículas de los pinos resultan
inferiores a la media de un amplio conjunto de pinares mediterráneos y
templados, pero sólo el K en las dos especies y Ca en el pino laricio están
por debajo del límite de confianza inferior. En ramas, los porcentajes de N, P
y K son más bajos y el Ca y el Mg mas altos que la media obtenida para
coniferas templadas pero siempre se encuentran dentro de los límites de
confianza para esta media. En tronco las concentraciones son en general
superiores a la media a excepción del P en ambas especies y el K en P.nigra.
Las concentraciones de N y Ca foliar en A.decorticans son respectivamente
más al ta y más baja que la media obtenida para arbustos mediterráneos pero de
orden similar a otras leguminosas también de la zona mediterránea. Los
contenidos de nutrientes en C.laurifolius se ajustan bien a los rangos
encontrados en matorrales mediterráneos a excepción del K, que resulta
inferior. Sin embargo los niveles de K sí quedan encajados dentro del rango
encontrado en cistáceas indicando una mayor frugalidad en el uso de este
elemento por el género Cístus.
Las especies estudiadas se sitúan perfectamente encajadas en las regiones
432
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
definidas para coniferas, jarales y leguminosas leñosas mediante la la
relación N y P foliar.
La mineralomasa acumulada en las especies mayoritarias es de 304 Kg/ha
año en RPP y 331 en RPN. Mientras que el matorral contribuye en algo menos de
un tercio a la biomasa total su mineralomasa supone entre el 40 y 50% del
total de nutrientes acumulados. El cociente peso nutrientes/biomasa es de
0.011 en P.pinaster, 0.009 en P.nigra y 0.020 en los dos arbustos.
El incremento radial en los dos pinos presenta una correlación positiva y
significativa con el DBH apoyando este tipo de comportamiento en poblaciones
arbóreas con individuos de tamaños pequeños. Esta característica, junto con
los altos valores absolutos del incremento radial indican que el rodal se
encuentra en una fase de rápida acumulación de biomasa.
La característica diferencial más destacable respecto a la producción
primaria neta del pinar es la elevada producción foliar así como los elevados
índices de eficiencia PH/PT o PT/BT. Las condiciones de estrés hídrico más
suavizadas en la zona de estudio y el momento sucesional en el que se
encuentra el pinar, son parámetros claves para explicar esta elevada
producción.
La producción de las especies arbustivas se sitúa dentro del rango de
otros sistemas de matorral mediterráneo, la eficiencia es mayor en A.
decorticans con una relación PPN/biomasa del 28-30/i frente a la j a ra (20-25%).
En las dos especies el esfuerzo invertido en la producción foliar está
alrededor del 507..
CAPITULO 3.
El desfronde medio anual alcanza en el sistema 2.5 T/ha.año (RPP) y 2.04
en (RPN), valores similares a la media para un amplio conjunto de ecosistemas
mediterráneos. Al estrato arbóreo corresponde un 64% en la primera zona y un
437. en la segunda. La fracción foliar es dominante con porcentajes respecto al
total de 837. en P. pinaster, 90% en P. nigra, 59% en A. decorticans y 82 % en
C. laurifolius. La fracción reproductiva tiene una contribución considerable
433
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
en el gaznacho (257.). El desfronde por individuo es 5 veces más aitn <**LO en A.
decorticans con respecto a C. laurifolius y del orden de 3 en P. pinaster
relación a P. nigra.
La cantidad de materia seca retornada por los dos pinos es en general
inferior a otros pinares mediterráneos y templados. El desfronde
del matorral resulta también inferior a sistemas arbustivos mediterráneos en
parte debido a sus porcentajes de recubrimiento inferiores a otros matorrales
dominantes o monoespecíficos de la bibliografía.
La cantidad de desfronde foliar y total en los arbustos se relaciona de
forma significativa y negativa con la temperatura ambiental. Eliminando tu
efecto, la humedad también se encuentra relacionada inversamente y de forma
significativa con el desfronde del gaznacho.
Tres de las especies estudiadas (P. pinaster, C. laurifolius y A.
decorticans) exhiben un patrón de abeisión foliar estival, característica
compartida con otras especies de pino y matorral mediterráneo. En P. ní¿ra,'~ia
abeisión es otoñal acercándolo en este aspecto, más a pinares templados que-*,
a mediterráneos. '>.
Las condiciones meteorológicas adversas de invierno provocan caídas
accidentales en ambos pinos y A. decorticans siendo menos marcada la
influencia sobre C. laurifolius.
i
La fuerte bimodalidad observada en el patrón estacional en los pinos es
ficticia ya que el pico invernal no corresponde en su mayor parte a un
desprendimiento foliar, sino a una caída de material escindido almacenado
sobre las ramas viejas.
Se observan pautas temporales en la composición química del desfronde
foliar más acusadas en las especies arbustivas. Existen claras disminuciones
en la concentración de N durante la abeisión fenológica en las cuatro
especies, de P en el gaznacho y el laricio y un aumento de concentración de Ca
en las especies arbustivas.
434
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
Las especies estudiadas con una mayor longevidad foliar (pinos)
retraslocan el N y P de forma gradual llegando a ser muy pequeña la
reabsorción en los momentos anteriores a la abcisión. Las especies arbustivas,
en concreto C. laurifolius, reabsorbe estos nutrientes con más intensidad en
los momentos inmediatos a la abcisión.
La eficacia en la retraslocación foliar de N calculada por distintos
métodos es superior en la j a ra (67-727.) seguida del pinar (40-60%) y de A.
decorticans (30-437.). Para el P la eficiencia puede variar sustancialmente
dependiendo del método empleado y sólo puede concluirse de forma clara la
menor eficiencia de la j a ra en relación a las demás especies.
Los aportes de nutrientes vía desfronde alcanzan valores similares en las
dos regiones (75 en RPP y 63 en RPN). El cociente nutrientes en
desfronde/nutrientes en biomasa es del 19 al 247. indicando un estadio joven
del sistema. C. laurifolius aporta cantidades semejantes a otros sistemas
arbustivos mediterráneos suponiendo un grado de recubrimiento similar mientras
que, en esas mismas condiciones, A. decorticans retornaría cantidades
superiores debido a las mayores concentraciones químicas de su desfronde para
la mayoría de nutrientes. El peso de nutrientes asociados al desfronde en P.
pinaster tienen una mayor similitud con pinares jóvenes que los de P. nigra
que resultan inferiores. La distribución en fracciones de los aportes muestra
un predominio de las hojas/acículas (79-83% de los aportes,). i
Para el conjunto de la cuenca, son similares los pesos de desfronde entre
el estrato arbóreo y el arbustivo. Sin embargo los matorrales aportan al suelo
un 47% más de nutrientes que el pinar. La relación peso nutrientes/ peso de
materia seca es en este flujo 0.017 en P. pinaster 0.015 en P. nigra 0.024 en
C. laurifolius y 0.034 en A. decorticans.
Aplicando el índice de eficiencia de Vitousek (1982), se observa que
respecto al N y el P, el pinar estudiado se sitúa entre las coniferas más
eficientes y en un lugar intermedio respecto al Ca. C. laurifolius se sitúa en
el grupo de matorrales más eficientes en el uso del N asociado a otros
435
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
matorrales de cistáceas españoles, mientras que A. decorticans se aeru
los sistemas fijadores simbiontes de N con una baja relación peso de mat •
seca/peso de N en desfronde. Respecto al P las especies arbustivas
localizan en el grupo de menor eficiencia de los matorrales mediterráneos
muy próximas a los encinares.
CAPITULO 4.
La aplicación del modelo de Lovett y Lindberg no resulta adecuado para
aquellos iones que en las especies estudiadas presentan trascolaciones netas
negativas para un elevado porcentaje de eventos lluviosos.
Las mayores tasas de deposición seca (vía externa) que resultan
significativas a part ir del modelo corresponden a los bicarbonatos y los
cloruros en las especies arbustivas, los sulfatos en las dos especies de pino
y los nitratos en P. pínaster.
Las mayores tasas de lixiviado significativas (vía interna) calculadas a
part ir del modelo corresponden al calcio y sulfato en P. pínaster, nitrato en
P. nigra y al K en las especies arbustivas. Se han encontrado tasas de
absorción foliar significativas para los bicarbonatos para el laricio y el
matorral y de nitrato en los arbustos.
Los iones predominantemente externos que entran al sistema por vía seca 6
húmeda son los sulfatos, cloruros, bicarbonatos y sodio. Existen diferencias
entre las dos especies de pino en relación a la captación de deposición seca
de algunos iones.
La contribución del lixiviado al flujo de retorno de nutrientes al suelo
sólo se manifiesta de forma marcada para el K en las especies estudiadas con
menor incidencia en el pinar. También es destacable el lixiviado de Ca y el Mg
en el pino resinero.
La vegetación es la vía principal de transferencia de todos los
nutrientes al suelo forestal con excepción del Na. Los mayores porcentajes de
contribución se encuentran para el N y P y en algunos casos el Ca y K.
436
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
Se observa en general una mayor rapidez en las tasas de circulación para
el Ca y Mg y una circulación más lenta en el N, P y K. A. decorticans es la
especie con que presenta tasas de circulación más elvadas para casi todos los
nutrientes. Las tasas normalizadas reflejan una estrecha similitud en el
comportamiento de los pinos no siendo así en las especies arbustivas.
La importancia relativa de las t res vías de penetración de nutrientes al
suelo forestal no variaría esencialmente en el supuesto de un 100% de
recubrimiento para cualquiera de las dos especies de pino. Las transferencias
de N se verían algo más favorecidas bajo la cubierta de P. pinaster.
En estas dos situaciones, la localización de los nutrientes en el triangulo se
sitúa dentro de los límites apuntados por Parker (1983) en distintos bosques.
En el supuesto de un 1007. de recubrimiento para cualquiera de las dos
especies arbustivas, la importancia relativa de las diferentes vías de
penetración no diferirían en lo básico a excepción del Na y N. La localización
de algunos elementos básicos para la nutrición vegetal (N, K y P) más próxima
a la base del triángulo que bajo cubierta de pinar, implica una mayor
aceleración del retorno para esos nutrientes bajo cubierta arbustiva.
Para cualquiera de las cuatro hipotéticas cubiertas monoespecíficas el
desfronde es la principal vía de trasferencia de N y P aunque los arbustos
retengan parte del N de la lluvia mediante absorción foliar y lixivien P-P04.
Ca y Mg se tranfieren preferentemente vía desfronde aunque en P. nigra y a.
decorticans un porcentaje importante proceda del lavado de deposición seca. En
todos los supuestos la trascolación neta juega un papel fundamental en la
transferencia de K, más patente bajo cubierta arbustiva. En todos los caso el
Na tiene un claro origen externo.
En cualquiera de las dos situaciones reales (RPP o RPN), la repoblación
de pinar no constituye en el momento actual una vía de transferencia
excesivamente importante en relación a la entrada total de nutrientes al suelo
sobre todo en RPN. En ambas zonas la deposición global supone una importante
fuente de nutrientes. El papel ejercido por A. decorticans en RPN es
437
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
destacable dado que con sólo coberturas del 407. se constituye en la princÍD I
vía de transferencia de N, P y K. La j a ra ejerce influencia notable en RPp
dónde es la principal vía de traansferencia de P y K al suelo de la cuenca.
CAPITULO 5.
Los porcentajes de pérdida de peso en la hojarasca de las especies
estudiadas al final de período de incubación (740- 770 días) es moderado en el
pinar y jaral (257. en P. pinaster, 35% en P. nigra y 387. en C. laurífolius) y
mucho más elevado en en A. decortícans (70%). Las curvas de pérdida de peso no
difieren significativamente entre las especies de pino y sí entre las especies
arbustivas. En todas las especies, sobre todo en el matorral, la variabilidad
en la pérdida de masa aumenta en el transcurso del segundo año.
La composición química inicial de la hojarasca muestra diferencias
significativas entre especies. La hojarasca de los dos pinos tiene composición
similar y bajos niveles de los principales macronutrientes en relación al
matorral mientras que la calidad de la hojarasca es mucho más heterogénea en
las especies arbustivas. Los contenidos de N y K en A. decortícans y de Ca, P
y cenizas en C. laurifolius son significativamente superiores al resto de las
especies. La masa remanente y el Índice C/N inicial en la hojarsca de las
especies estudiadas se relacionan de forma significativa y negativa.
De los t res modelos aplicados a los datos de pérdida de peso (exponencial
simple no linear, exponencial simple linear y exponencial doble), ha sido este
último el que ha proporcionado los mejores ajustes en todas las especies
siendo los valores del pino laricio y la j a ra los mejor explicados por este
tipo de modelo. Con él se calculan porcentaje de masa remanente al final de la
incubación de 74.27. en P. pinaster, 697. en P. nigra, 61 en C. laurifolius y
29.3 en A. decortícans.
El porcentaje de fracción resistente calculada en el modelo doble varía
en el siguiente orden de mayor a menor; P. pinaster > P. nigra > C.
laurifolius > A. decortícans coincidente con la variación de la densidad
foliar. Consecuentemente, la fracción lábil se ordena en sentido inverso. En
438
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
el gaznacho, la combinación de un menor porcentaje de componente resistente
junto a la mayor tasa de descomposición de éste resultan determinantes para
explicar su diferente dinámica de descomposición.
Se ha encontrado una correlación significativa y positiva entre masa
remanente y el cociente entre los porcentajes de componente refractario y
nitrógeno en la hojarasca inicial reflejando la relación existente entre
calidad de la hojarasca y la dinámica de la pérdida de peso.
Las comparaciones estadísticas de las constantes de descomposición inter
e intraespecíficas para distintos tiempos indican dos tendencias más o menos
marcada en las especies estudiadas: una, la que siguen las especies arbustivas
con disminuciones de masa muy acusadas el primer año seguidas de un período de
estabilización en el segundocon tasas relativas significativamente inferiores
a las del primero. Otra, la que sigue el pinar con una dinámica de
descomposición más continua a lo largo de los dos años. Para el período total
de muestreo la constante de descomposición de cada especie es
significativamente distinta del resto y varía en el siguiente orden: A.
decorticans, 0.53 > C. laurifolius, 0.21 > P. nigra, 0.17> P. pinaster, 0.13.
Las tasas de descomposición durante el primer año en los pinos estudiados
son inferiores a la de pinares templados y más similares en relación a
coniferas mediterráneas. La de C. laurifolius es totalmente concordante con
otras esclerófilas mediterráneas mientras que la hojarasca del gaznacho pierde
peso en cantidades similares a otras fijadoras de N.
Respecto a la liberación de nutrientes son el K y el Mg los más
fácilmente liberados mientras que N y Ca ocupan las últimas posiciones,
resultado concordante con la mayoría de series bibliográficas. El P sigue un
comportamiento variable según la especie aunque es destacable su alta
velocidad de liberación en la jara. Al final del periodo de seguimiento (algo
más de año y medio) se detecta retención y acumulación de P, Ca y N en P.
pinaster, Na, Ca y N en P. nigra y N en C. laurifolius. En A. decorticans
todos los elementos presentan en mayor o menor grado mineralización neta.
439
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
CAPITULO 6.
La fenología en los nodulos de A. decorticans sigue un patrón estacional
muy relacionado con las condiciones climáticas locales. La explosión de
nodulos jóvenes se registra entre Septiembre Octubre, la mayor densidad de
nodulos con altas tasas de fijación en primavera y la senescencia a finales de
junio marcada por el comienzo del estrés hídrico.
No se han detectado diferencias altitudinales respecto al patrón anual,
patrón estacional, ni a los valores mensuales del ARA (actividad reductora de
acetileno) entre las parcelas estudiadas. Los sólo 200 m de altitud entre la
cota alta y baja de la cuenca no parecen suficientes para detectar variacioens
significativas en la actividad nitrogenasa.
El patrón estacional se manifiesta en un aumento progresivo del ARA
durante la primavera con máximos en el mes de Mayo-Junio (1797-2497 nmol
C2H4/g p.s..hora). En verano la actividad disminuye a niveles mínimos
coincidiendo con el estrés hídrico y senescencia nodular. La actividad se
recupera en otoño pero se anula en los meses de heladas invernales
probablemente a causa de la inhibición de la nitrogenasa por la baja
temperatura edáfica.
La fijación media diaria es significativamente más alta durante el mes de
Octubre. También el patrón diario del ARA es diferente entre ambos meses. En
Octubre, la fijación durante el periodo de luz es significativamente más
alto que durante la oscuridad, mientras que no existen diferencias entre
períodos para el mes de Julio.
Aunque resulte arriesgado y con un amplio margen de error el calculo de
entradas de nitrógeno por fijación a part i r de los resultados del ARA, ésta
podría oscilar entre 0.5 - 1 Kg de N2 por hectárea y año, valor situado en el
límite inferior que Carlyle (1986) da para las fijadoras leguminosas que
precisamente corresponden a especies leñosas.
440
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Resumen y conclusiones.
CAPITULO 7.
Existe retraslocación de N, P y K en las cuatro especies estudiadas
indicando un uso conservativo de estos nutrientes lo que coincide los
resultados obtenidos a part i r del balance hidrológico (Domingo 1991).
A. decorticans es la especie más explotadora de los recursos edáficos de
la cuenca dados sus elevados costes nutricionales y sus mínimos porcentajes de
retraslocación. Sin embargo fija nitrógeno y es la única especie que
proporciona vía desfronde un retorno más elevado de N que, además se
mineraliza inmediatamente después de que la hojarasca llega al suelo. Tanto el
N como el P en el gaznacho entran en un circuito más completo
(vegetación-suelo-mantillo) en relación con las otras especies en las cuales
la reutilízación del recurso mineral mediante mecanismos internos impide en
buena parte la transferencia de éstos a corto plazo hacia otros
compartimentos.
La comparación normalizada para distintos compartimentos y flujos de
nutrientes permite concluir que los flujos relacionados con la incorporación
de nutrientes al suelo son sistemáticamente más altos en las especies
arbustivas, sobre todo en A. decorticans. Los tiempos de renovación más altos
se obtienen para P. nigra y los más bajos para A. decorticans. Los cocientes
MINERALOMASA/RETENCION son más altos en el pinar indicando una etapa
sucesional más temprana que en el matorral. La interacción con el agua de
lluvia permite también establecer algunas diferencias específicas.
441
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
CONCLUSIONES GENERALES.
No existen apenas diferencias en los valores globales de los flujos y
compartimentos de materia orgánica y nutrientes en las dos grandes áreas en
que se ha dividido la cuenca. Las diferencias específicas se atenúan al
considerar la estructura vegetal particular de cada zona.
Las características encontradas para los flujos de materia seca y
nutrientes estudiados en la cuenca vienen fundamentalmente determinados por la
inmadurez de la repoblación de pino y su coexistencia con una especie
arbustiva (A. decortícans) con altas tasas de circulación de nutrientes. En
este aspecto el papel de la j a ra es mucho más moderado.
En términos globales, la existencia de retraslocación para el N, P y K y
el signo del balance hidrológico para estos elementos indica un comportamiento
conservativo en el uso de estos nutrientes.
442
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Alien O.N. y Alien E.K. 1981. The Leguminosae. University of Wisconsin Press.
Alien S.E., Grimshaw H.M., Parkinson J.A., y Quarmby Ch. 1974. Chemical
analysis of ecological materials Oxford. Blackwell Scientific Publications.
Alvera B. 1990. Ciclos de elementos minerales en dos pinares pirenaicos. Tesis
doctoral. Universidad Complutense de Madrid.
Attiwill P.M. 1968. The loss of elements from decomposing li t ter. Ecology, 49:
142-145.
Avila A. 1988. Balanc d'aigua i nutrients en una conca d'alzinar al Montseny
Universidad Autónoma de Barcelona. Tesis doctoral.
Bellot F. 1978. El tapiz vegetal de la Península Ibérica Madrid. H BLUME.
Bellot J. 1989. Análisis de los flujos de deposición global, trascolación,
escorrentía cortical y deposición seca en el encinar mediterráneo de V'Avie
(Sierra de Prades, Tarragona) Alicante. Tesis Doctoral. Universidad de
Alicante.
Bellot J. y Escarré A. 1991. Chemical characteristics and temporal variations
of nutrients in throughfall an stemflow of three species in Mediterranean holm
oak forest. Forest Ecology and Management, 41: 125-135.
Berendse F., Berg B. y Bosatta E. 1987. The effect of lignin and nitrogen on
the decomposition of lit ter in nutrient-poor ecosystems: a theorettical
approach. Can. J. Bot., 65: 1116-1120.
Berendse F., Oudhof H. y Bol J. 1987. A comparative study on nutrient eyeling
in wet heathland ecosystems. Oecologia, 74: 174-184.
Berendse F., Oudhof H. y Bol J. 1987. A comparative study on nutrient eyeling
in wet heathland ecosystems. 1. Litter production and nutrient losses from the
plant. Oecologia, 74: 174-184.
443
v
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Berg B. 1986. Nutrient reléase from litter and humus in coniferous forest
soils: A mini review. Scand. J. For. Res., 1: 359-369.
Berg B. y Lundmark J.E. 1987. Decomposition of needle l i t ter in Pinus contorta
and Pinus sylvestris monocultures. A comparison. Scand. J. For Res, 2: 3-12.
Berg B. y McClaugherty C. 1987. Nitrogen reléase from li t ter in relation to
disappearance of lignin. Biogeochemistry, 4: 219-224.
Berg B. y Staaf H. 1981. Leaching, accumulation and reléase of nitrogen in
decomposing forest l i t ter. En: Clark F.E. and Rosswall T. (eds.), Terrestrial
nitrogen cycles. Estocolmo. Ecol. Bull. 3: 163-178.
Berg B. y Staaf H. 1980. Decomposition ra te and chemical changes of Scots Pine
needle li t ter. II. Influence of chemical composition. En: Persson T. (eds.),
Northern coniferous forests. An ecosystem study. pags.-.319-390. Stockholm.
Tundra Biome Steering Committee.
Bermúdez de Castro F., Miguel C. y Rodríguez Barrueco C. 1976. A study of the
capacity of soils to induce nodules in A. glutinosa (L.) and M. gale (L.) with
special reference to the specifity of the endophytes. An. Microbiol. (Inst.
Pasteur), 127: 307-315.
Bockheim J.G. y Leide J.E. 1986. Litter and forest floor dynamics in a Pinus
resinosa plantation in Wisconsin. Plant and Soil, 96: 393-406.
Bocock K., 1 , Gilbert O.J., Capstick C.K., Turner D.C., Ward J.S. y Woodman
M.J. 1960. Changes in leaf lit ter when placed on the surface of soil with
contrasting humus types. Journal of Soil Science, 11: 1-9.
Bocock K.L. y Gilbert O.J.W. 1957. The disappearence of leaf l i t ter under
different woodland conditions. Plant and Soil, 9: 179-184.
Boring L.R. y Swank W.T. 1984. Symbiotic nitrogen fixation in regenerating
Black Locust (Robinia pseudoacacia L.) stands. Forest Science, 30: 528-537.
444
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Bormann F.H. y Likens G.E. 1979. Pattern and Process in a forested ecosystem
N. York. Springer Veriag. 253 pp.
Bray J.R. y Gorham E. 1964. Litter production in Forest of the world. Adv.
Ecol. Res., 2: 101- 157.
Briggs S.V. y Maher M.T. 1983. Litter fall and leaf decomposition in a River
Red Gum (Eucalytus camaldulensis) Swamp. Aust. J. Bot., 31: 307-316.
Bruijnzeel L.A. 1985. Nutrient content of litterfall in coniferos forest
plantations in central Java, Indonesia. Journal of Tropical Ecology, 1:
353-372.
Bunnell F.L. y Tait D.E.N. 1974. Mathematical simulation models of
decomposition processes. En: Holding A.J. and Heal S.F. (eds.), Soil organisms
and decomposition in tundra. MacLean & Flanagan P.W.
Cañe 11 M.G.R. 1982. World forest biomass and primary production data London.
Academic Press.
Capel Molina J. 1976. Los mecanismos de precipitación en Almería y la
circulación en altura Almería. Caja Rural provincial de Almería.
Capel Molina J. 1981. Recopilación bibliográfica sobre naturaleza almeriense
(Climatología, Geología, Botánica y Zoología) Granada. Universidad de Granada
144pp.
Capel Molina J. 1977. El clima de la provincia de Almería Almería. Caja de
Ahorros de Almería.
Capel Molina J. 1986. El clima de la provincia de Almería Almería. Caja de
ahorros y Monte de Piedad de Almería.
Capel Molina J.J. 1984. El clima de zonas áridas. En: Instituto de Estudios
Almerienses, Seminario de Zonas Áridas. Almería 1982. Almería. Diputación
Provincial.
445
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
Carceller F., Santacecilia M.A. y Vallejo V.R. 1989. Primeros datos sobre el
ciclo de la materia orgánica en t res bosques caducifolios del Moncayo
(Zaragoza). Options Mediterranéennes- Serie A, 3: 183-186.
Carlyle J.C. 1986. Nitrogen cycling in forested ecosystems. Forestry
Abstracts, 47 (5): 307-336.
Chapín III F.S. 1980. The mineral nutrition of wild plants. Ann. Rev. Ecol.
Syst., 11: 233-260.
Chauvet E. 1987. Changes in the chemical composition of alder, poplar an
willow leaves during decomposition in a river. Hydrobiologia, 148: 35-44.
Clemente A. 1983. Componentes específico y estacional en la variación de
contenidos de elementos químicos de las especies y formas biológicas del
encinar mediterráneo Tesis de Licenciatura. Universidad de Alicante.
Colé D.W. y Rapp M. 1981. Elemental cycling in Forest Ecosystems. En: Reichle
D.E. (ed), Dinamics Properties of Forest Ecosystems. Cambridge. Cambridge
University Press.
Cortina J. 1992. Efectes de las plantacions de Pinus radiata A.Don sobre la
fertilitat del sol Universitat de Barcelona. Tesis Doctoral.
Cromack J.R. y Monk C.D. 1975. Litter. production, decomposition and nutrient
cycling an a mixed hardwood watershed and a white pine watersheed. En: Howell
F.J., Gentry J.B. and Smith M.H. ( eds j , Mineral cycling in southeastern
ecosystems. Washington DC. Technical Information Center, United Stated Energy
Research and Developement Administration.
Cronan C.S. y Reiners W.A. 1983. Canopy processing of acidic precipitation by
coniferous and hardwood forest in New England. Oecologia, 59: 216-223.
Dannaoui S. 1981. Production de litiere et restitution au sol d' elements
biogenes dans des peuplements mediterraneens de Pinus pinnea L and Pinus
446
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
brutia Ten. Ecología Mediterránea, 7 (2): 13- 25.
Dawson M. 1900. Further observations on the nature and functions of the
nodules of leguminous plants. Phil. Trans. R. Soc. London, 193: 51-67.
Del Arco J.M., Escudero A. y Garrido M.V. 1991. Effects of site
characteristics on nitrogen retranslocation from senescing leaves. Ecology,
72: 701-708.
Di Castri F., Goodall D.W. y Specht R.E. (eds) 1981. Ecosystems of the world
11. Mediterranean type shrublands. Elsevier Scientific Publishing Company.
643 pp.
Domingo F. 1991. Papel de la cubierta vegetal en los aportes de nutrientes y
salidas por avenamiento en una cuenca re forestada con coniferas en la Sierra
de Filabres (Almería) Alicante. Tesis doctoral. Universidad de Alicante.
Domingo F., Moro M.J., Bellot J. y Escarré A. 1991. Throughfall chemistry in
shrub and pine species studied in a Mediterranean catchment. Mitteilungsblat
des hydrographischen dienstes in Osterreich, 65/66: 192-196.
Dziadowiec H. 1987. The decomposition of plant l i t ter fall in a
oak-linden-hornbeam forest and an oak-pine mixed forest of the Bialowieza
National Park. Acta Societatis Botanicorum Poloniae, 56 (1): 169-185.
Enright N.J. y Ogden J. 1987. Decomposition of kitter from common woody
species of kauri (Agathis australis Salisb.) forest in northern New Zealand.
Australian Journal of Ecology, 12: 109-124.
Escarré A., Ferres Ll., López R., Martín J., Roda F. y Terrades J. 1987.
Nutrient use estrategy by evergreen-oak (Quercus ilex spp ilex) in NE Spain.
En: Tenhunen J.D. et al. (eds.), Plant response to stress. Berlín. Springer
Verlag.
Escudero A. y Del Arco J.M. 1987. Ecological significance of the phenology of
leaf abscission. Oikos, 49: 11- 14.
447
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadr i l lero
Escudero A., Del Arco J.M. y Garrido M.V. 1992. The efficiency of nitrogen
retranslocation from leaf biomasas in Quecus ilex ecosystems. Vegetatio,
99-100: 225-237.
Escudero A., García B., Gómez J.M. y Luis E. 1985. The nutrient cycling in
Quercus rotundifolia and Quercus pyrenaica ecosystems ("dehesas") of Spain.
Acta Oecologica. Oecol. Plant., 6 (20): 73-86.
Escudero A., Garrido M.V. y Matías M.D. 1987. Decay curves of leaf l i t ter from
evergreen and deciduous t ree species. Acta Oecologica. Oecol. Plant, 8 (22):
81-90.
Escudero A., Montalvo L, García B. y Gómez J.M. 1983. La circulación de
nutrientes en ecosistemas arbolados: Incidencia de los mecanismos fisiológicos
de control del flujo. Revista de la Real Academia de Ciencias Exactas, Físicas
y Naturales, 11 (4): 749-766.
Escudero Berian A., Garrido M.V. y Gómez Gutiérrez J.M. 1985. Descomposición
de la hojarasca de Quercus rotundifolia Lam y Quercus pyrenaica Willd en monte
adehesado. Anales de Edaf. y Agrobiol., 44 1-2: 65-79.
Ezcurra E. y Becerra J. 1987. Experimental decomposition of l i t ter from the
Tamaulipan cloud forest: A comparison of four simple models. Biotropica, 19
(4): 290-296.
Fahey T.J. 1983. Nutrient dynamics of aboveground detritus in lodgpole pine
(Pinus contorta ssp latifolia) ecosystems, southeastern Wyoming. Ecological
Monographs, 53 (1): 51-72.
Ferres Ll. 1984. Biomasa, producción y mineralomasas del encinar montano de la
Castanya (Montseny) Bejlaterra. U. Autónoma de Barcelona. Tesis Doctoral.
Ferres Ll., Roda F., Verdú A.M.C. y Terradas J. 1984. Circulación de
nutrientes en algunos ecosistemas del Montseny (Barcelona). Mediterránea. Ser.
Biol., 1: 139-166.
448
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Forrest W.G. y Ovington J.D. 1971. Variation in dry weight and mineral
nutrient contení of Pinus ra.dia.ta. progeny. Silvae Genet, 20: 174-179.
Gallardo J.F., Santa Regina I. y San Miguel C. 1989. Ciclos biogeoquímicos en
bosques de la Sierra de Béjar (Salamanca, España) 1. Producción de hojarasca.
Revue d'écologie et du biologie du sol, 26 (1): 35-46.
Garrido M.V., del Arco J.M. y Escudero A. 1989. Ciclo de nutrientes: Fenología
de la abscisión y dinámica de la descomposición en distintas especies
caducifolias y perennifolias. Options Mediterranéennes- Serie A, 3: 163-166.
Gholz H.L. 1982. Environmental limits on aboveground net primary production,
leaf área, and biomass in vegetation zones of the Pacific northwest. Ecology,
63: 469-481.
Gholz H.L. y Fisher R.F. 1985. Nutrient dynamics in Slash Pine plantations
ecosystems. Ecology, 66: 647-659.
Cordón J.C. y Wheeler C.T. 1978. Whole plant studies on photosynthetic and
acetylene reduction in Alnus glutinosa. New Phytol., 80: 179-186.
Gosz J.M., Likens G.E. y Bormman F.H. 1973. Nutrient reléase from descomposing
leaf and branch litter in the Hubbard Brook forest, New Hampshire. Ecological
Monographs, 43: 173-191.
Gosz J.M., Likens G.E. y Bormann F.H. 1972. Nutrient contení of l i t ter fall on
the Hubbard Brook experimental forest, New Hampshire. Ecology, 53, 5: 769-784.
Gourbiere F. 1986. Méthode d'étude simultanee de la décomposition et des
mycoflores des aiguilles de coniféres (Abies alba). Soil Biol. Biochem., 18
(2): 155-160.
Gourbiere F. y Corman A. 1987. Décomposition des aiguilles ú'Abies alba:
Hétérogéneité du substrat et de la mycoflore, role de Marasmius androsaceus.
Soil. Biol. Bichem, 19 (1): 69-75.
449
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadr i l lero
Gracia C. (ed) en prensa. Inventari ecológic i forestal de Catalunya 28 Pía de
l'Estany CREAF Barcelona.
Gracia C. (ed) en prensa. Inventari ecológic i forestal de Catalunya. Solsones
CREAF Barcelona.
Gray J .T. 1983. Nutrient use by evergreen and deciduous shrubs in southern
California. Journal of Ecology, 71: 21 - 41.
Gray J.T. y Schlesinger W.H. 1981. Nutrient cycling in Mediterranean type
Ecosystems pp 259- 284. En: Miller P.C. (eds.), Resource use by Chaparral and
Matorral. Ecol. Studies 39. Springer- Verlag.
Gray J.T. y Schlesinger W.H. 1981. Biomass, production and l i t terfal l in the
Coastal sage scrub of Shouthern Caifornia. Amer.J.Bot., 68 (1): 24- 33.
Hansen A.P., Pate J.S. y Atkins C.A. 1987. Relationshíps between acetylene
reduction activity, hydrogen evolution and nitrogen fixation in nodules of
Acacia spp.: experimental background to assaying fixation by acetylene
reduction on field conditions. J. Exp. Bot., 38, 186: 1-12.
Hansen A.P., Pate J.S. y Bell D.T. 1987. Nitrogen fixation in the Jarran
forest legumes. En: Proceedings of MEDECOS V: 67-67.
Hardy R.W.F., Burns C.R. y Holsten R.D. 1973. Applications of the acetylene
reduction assay for measurement of nitrogen fixation. Soil. Biol. Biochem., 5:
47-81.
Hasselrot B. y Grennfelt P. 1987. Deposition of air pollutants in a
wind-exposed forest edge. Water, air and soil pollution, 34: 135-143.
Helgerson O.T., Cordón J.C. y Perry D.A. 1984. N2 fixation by red alder {Alnus
rubra) and scotchbroom (Cytisus scoparius) planted under precommercially
thinned Douglas-fir (Pseudotsuga menziesii). Plant and Soil, 78: 221-223.
450
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Hendrickson O.Q., Burgess D.M. y Chatarpaul L. 1987. Biomass and nutrients in
Great Lakes-St. Lawrence forest species: Irnplications for whole t ree and
conventional harvest. Can. J. For. Res., 17: 210-218.
Herrera C.M. 1984. Tipos morfológicos y funcionales en plantas del matorral
mediterráneo del sur de España. Studia Oecologica, 5: 7-34.
Herridge D.F. 1982. Assessment of nitrogen fixation. En: Vincent J.M. (ed.),
Nitroxen fixation in legum.es. New York. Academic Press.
Hintze J.L. 1988. SOLO. BMDP Statístical Software. Inc. versión 2.0 Los
Angeles.
Hogberg P. y Kvarnstrom M. 1982. Nitrogen fixation by the woody legume
Leucaena leucocephala in Tanzania. Plant and Soil, 66: 21-28.
Hornung M., Roda F. y Langan S.I. (eds) 1990. A review of smalL catchment
studies in Western Europe producing hidrochemical budgets. Air pollution
research report 28 Bruselas. Commission of the European Communities.
ICONA 1987. Proyecto LUCDENE, Mapa de suelos. Macael 1013 Madrid. Ministerio
de Agricultura, Pesca y Alimentación.
i
Jenny H., Gessel S.P. y Binghman F.T. 1949. Comparative study of decomposition
ra tes of organíc matter in températe and tropical regions. Soil Sci., 68:
419-432.
Johnson D.W. y Todd D.E. 1990. Nutrient cycling in forests of Walker Branch
watershed, Tennessee: Roles of uptake and leaching in causing soil changes. J.
Environ. Qual., 19: 97-104.
Johnson D.W. y Todd T. 1990. Nutrient cycling in forest of Walker Branch
Watershed, Tennesse: Roles of uptake and leaching in causing soil changes.
Journal of Environmental Quality, 19: 97-104.
451
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadrillero
Jorgesen J.R., Wells C.G. y Metz L.J. 1980. Nutrient changes in decomposing
loblolly pine forest floor. Soil Sci. Soc. Am. J., 44: 1307-1314.
Kazda M. 1987. Sequencial stemflow sampling for estimation of washoff leaching
in beech stands. En: Harrison A.F., Ineson P. and Heal O.W. (eds.), Field
methods for studying nutrient cycling in terrestrial ecosystems. Elsevier.
Killingneck K.T. y Costigan S.A. 1988. Element resorption in a guild of
understory shrubs species: niche differentiation an resorption thresholds.
Oikos, 53: 366-374.
Klemmedson J.O., Meier CE. y Campbell R.E. 1985. Needle decomposition and
nutrient reléase in ponderosa pine ecosystems. Forest Sci., 31 (3): 647-660.
Kruger F.J., Mítchell D.T. y Jarvis J.U.M. (eds) 1983. Mediterranean Type
Ecosystems. Ecological Studies 43 Berlín. Springer Verlag.
Larcher W. 1977. Ecofisiología Vegetal Barcelona. Omega.
Lavado M., Núñez E. y Escudero J.C. 1989. Variaciones mensuales en el aporte
de biomasa al suelo por distintas especies de matorral mediterráneo. En:
Bellot J. (ed.), Jornadas sobre las bases ecológicas para la gestión en
ecosistemas terrestres. Zaragoza. CHIHEAM.
Lavado M., Núñez E. y Escudero J.C. 1988. Velocidad de descomposición anual de
hojas de dos especies de matorral mediterráneo: Myrtus communis L. y Pistacia
lentiscus L. En: Actas Congreso Mundial sobre el bosque y matorral
mediterráneo. Cáceres.
Lázaro Suau R. 1988. Los pisos bioclimáticos y los ombroclimas de la provincia
de Almería. Coord. A. Blanco de Pablos CSIC. Avances sobre la investigación en
climatología, pp 317-326.
Lemee G. 1974. Recherches sur les écosystémes des reserves biologiques de la
fóret de Fontenebleau IV. Entrées d'élements minéraux par les précipitations
et t ransfer í au sol par le pluviolessivage. Oecol. Plant, 9 (3): 187-200.
452
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Leonardi S., Rapp M. y La Rosa V. 1988. Repartition et dinamique de la
maatiére organique dans une fóret de Pinus laricio Poir. Ecología
Mediterránea, 14: 17 -29.
Lieth H. y Whittaker R.H. 1975. Primary productivity of the biosphere.
Springer Verlag.
Likens G.E., Bormann F.H., Pierce R.S., Eaton J.S. y Johnson N.M. 1977.
Biogeochemistry of a forested ecosystem N. York. Springer Verlag 148pp.
Lindberg S.E. and Garten C.T. 1989. Sources of sulphur in forest canopy
throughfall. Nature, 336 (6195): 148-151.
Lledó M.J. 1990. Compartimentos y flujos biogeoquímicos en una cuenca de
encinar del Monte Poblet Alicante. Universidad de Alicante. Tesis Doctoral.
Lledó M.J. 1982. Datos energéticos, estructurales y químicos de Pinus
halepensis Mili, en el pinar de Sierra Espuña (Provincia de Murcia) Tesis de
Licanciatura. Universidad de Alicante.
López Sánchez M.V. 1989. Estudio de los flujos hídricos, y de la contribución
de la deposición seca y lixiviación al aporte de nutrientes a un suelo
forestal, bajo encina (Q. ilex) y pino (P. sylvestris) Zaragoza. Master of
Science. I.A.M.Z.-C.I.H.E.A.M.
Losa J.M., Molero J. y Casares M. 1986. El paisaje vegetal de Sierra Nevada
(La cuenca alta del río Genil, Granada) Granada. Servicio de Publicaciones de
la Universidad de Granada.
Lossaint P. y Rapp M. 1971. Repartition de la matiere organique, productivite
et cycles des elements mineraux dans des ecosystemes de climat mediterraneeen.
En: Productivite des ecosystemes forestiers. Actes Coll Bruxelles 1969.
Lousier J.D. y Parkinson D. 1978. Chemical element dynamics in decomposing
leaf l i t ter. Can. J. Bot., 56: 2795-2812.
453
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadr i l le ro
Lousier J.D. y Parkinson D. 1976. Litter decomposition in a cool températe
deciduous forest. Can. J. Bot., 54: 419-436.
Lovett G.M. y Lindberg S.E. 1984. Dry deposition and canopy exchange in a
mixed Oak forest as determined by analysis of throughfall. Journal of Applied
Ecology, 21: 1013-1027.
Lutz H.J. y Chandler R.F. 1946. Forest Soils New York. John Wiley & Sons Inc.
Maheswaran J. y Attwill P.M. 1987. Loss of organic matter, elements and
organic matter in decomposing Eucalyptus microcarpa leaf l i t ter . Can. J. Bot.,
65: 2601-2606.
Márquez M.f Núñez E. y Escudero J.C. 1989. Dinámica del nitrógeno en un
gradiente de edad de Cistus ladanifer L. En: Bellot J. (ed.), Jornadas sobre
las bases ecológicas para la gestión en ecosistemas terrestres. Zaragoza.
CIHEAM.
Martín J. 1984. Variaciones estacionales en el contenido de nutrientes y
pigmentos en cuatro especies de espinar costero de Alicante Tesis Doctoral.
Universidad de Alicante.
Mayor X. 1990. El paper deis nutrients com a factors limitants de la prodúcelo
primaria de l'alzinar de la conca del Torrent de la Mina (Montseny) Barcelona.
Universitat autónoma de Barcelona. Treball de mestratge.
McClaugherty C. y Berg B. 1987. Cellulose, lignin and nitrogen concentrations
as ra te regulating factors in late stages of forest l i t ter decomposition.
Pedobiologia, 30: 101-112.
Merino J. 1986. Productividad. Ciclo de nutrientes en el área de Doñana. En:
Bases ecológiques per la gestió ambiental. Barcelona. Diputación de
Barcelona.
Merino O., Martin M.P., Martin A. y Merino J. 1990. Successional and temporal
454
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
changes in primary productivity in two mediterranean scrub ecosystems. Acta
Oecologica, 11 (1): 103- 112.
Miller P.C. (Ed.) 1981. Resource use by chaparral and matorral. Ecol. Studies
39 Nueva York. Springer Verlag.
Minchin F.R., Witty J.F., Shezhy J.E. y Muller M. 1983. A major error in the
acetylene reduction assay; decreases in nodular nitrogenase activity under
assay conditions. J. Exp. Bot., 34: 641-649.
Mooney H.A. 1981. Primary production in mediterranean-climate regions. En: Di
Castri F., Goodall D.W. and Specht R.L. (eds.), Ecosystems of the world 11.
Mediterranean type shrublands. Elsevier.
Moro M. J. Estudio comparativo de dos parcelas con pinar repoblado con y sin
tratamiento silvícola. En: "Funcionamiento biogeoquímico de cuencas con
vegetación natural, repoblaciones y cultivos; estima de pérdidas erosivas.
Memoria final. LUCDEME - ICONA.
Moro M.J., Domingo F., Pérez Corona E. y Bermúdez de Castro F. 1989.
Nodulación y fijación de nitrógeno atmosférico (ARA) en Adenocarpus
decorticans. En: IV Reunión Nacional de Fijación de Nitrógeno, pp. 219-220.
Sevilla. Universidad de Sevilla.
Moro M. J., Domingo F. y Bermúdez de Castro F. 1992. Acetylene reduction
activity (ARA) by the shrub legume A. decorticans Boiss. in Southern Spain
(Almería). Acta oecologica, 13 (3) 325-333.
Muller A. y Bermúdez de Castro F. 1987. Evaluación de la actividad reductora
del acetileno (ARA) por cromatografía de gases. Técnicas de Laboratorio, 135:
107-112.
Nahal I. 1981. The mediterranean climate from a biological viewpoint. En: di
Castri F., Goodall D.W. and Specht R.L. (eds.), Mediterranean type shrublands.
Ecosystems of the world 11. Amsterdam. Elsevier.
455
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
Naik M.L. y Shrivastava B.K. 1985. Leaf l i t ter in the forests of Surguja
(Madhya Pradesh). Journal of Tropical Forestry, 1 (11): 140- 144.
Nalsk M.L. y Shrivastava B.K. 1985. Leaf litter fall in the forests of Surguja
(Madhya Pradesh). Journal of Tropical Forestry, 1: 140-144.
Nuñez Oliveira E. 1989. Ecología del Jaral de Cistus ladanifer L. Tesis
doctoral Badajoz. Facultad de Ciencias. Universidad de Extremadura.
O'Conell A.M. 1987. Litter decomposition, soil respiration ans soil chemical
and biochemical properties at three contrasting sites in karri (Eucalyptus
diversicolor F. Muell) forests of south-western Australia. Australian Journal
of Ecology, 12: 31-40.
O'Conell A.M. 1987. Litter dynamics in karri (Eucalyptus diversicolor) forests
of South-Western Australia. Journal of Ecology, 75: 781-796.
O'Conell A.M. 1988. Decomposition of leaf lit ter in Karri (Eucalyptus
diversicolor) forest of varying age. Forest Ecology and Management, 24:
113-125.
Olson J.S. 1971. Primary productivity: Températe forests, specially American
deciduous types. En: Productivity of forest ecosystems. Act. Symp. Bruxelles
1969. 235-258.
Olson J.S. 1963. Energy storage and the balance of producers and decomposers
in ecological systems. Ecology, 44: 322-331.
O'Neill R.V. y De Angelis D.L. 1981. Comparative productivity and biomass
relations of forest ecosystems. En: Reichle D.E. (ed.), Dynamic properties of
forest ecosystems. Cambridge University Press.
Ovington J.D. 1959. Mineral contení of plantations of Pinus sylvestris L. Ann.
Bot. N. S, 23: 75-88.
456
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Parker G.G. 1983. Throughfall and stemflow in the forest nutrient cycle. Adv.
Ecol. Res., 13: 58-121.
Passana L. 1970. Composition minórale de diverses especies calcicoles et
calcifugues de la región mediterranéenne frangaise. Oecol. Plant, 5: 225-245.
Pastor A. y Martin J. 1989. Estudio de pinares en la provincia de Alicante.
Informe final. Consellería de Agricultura. Generalitat valenciana.
Pearson J.A., Knight D.H. y Fahey T. 1987. Biomass and nutrient accumulation
during stand development in Wyoming lodgepole pine forest. Ecology, 68:
1966-1973.
Pérez Hernández M.C., Gutiérrez Mañero F.J., Pozuelo González J.M. y Bermúdez
de Castro F. 1989. Entradas biológicas de nitrógeno en un bosquete ribereño.
Análisis del ARA. Options méditerranéennes Serie Séminaires, 3: 207-210.
Pinol J. 1990. Hidrología y biogeoquímica de conques forestades de les
Muntanyes de Prades Universidad de Barcelona. Tesis doctoral.
Pizelle G. 1984. Seasonal variation of the sexual reproductive growth and
nitrogenase activity (C2H2) in mature Alnus glutinosa. Plant and soil, 78:
181-188.
Polunin O. y Smythies B.E. 1977. Guía de campo de las flores de españa,
Portugal y sudoeste de Francia Barcelona. Ed. Omega.
Potter C.S., Ragsdale H.L. y Berish C.W. 1987. Resorption of foliar nutrients
in a regenerating southern Appalachian forest. Oecologia, 73: 268-271.
Puigdefabregas J. 1981. El pinar altoaragonés de Pinus sylvestris. Estructura
y producción primaria neta Pamplona. Universidad de Navarra. Tesis Doctoral.
Rapp M. 1967. Production de litiere et apport au sol d'elements mineraux et
d'azote dans un bois de pins d'Alep (Pinus halepensis Mili.). Oecol. Plant.,
457
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

U.J. Moro Cuadrillero
2: 325- 388.
Rapp M. 1983. Some problems of disturbance on the nutrient cycling ¡ n
ecosystems. En: Mooney H.A. and Godron M. (eds.), Disturbance ans ecosystems
components of response. Berlín. Springer Verlag.
Rapp M. y Cabanettes A. 1980. Biomasse, mineralomasse et productivité d'un
écosysteme a pins pignons (Pinus pinea L.) du li t toral méditerranéen. II.
Composition chimique et mineralomasse. Acta Oecol. Plant., 1: 151-164.
Rapp M., Leclerc M.C. y Lossaint P. 1979. The nitrogen economy in a Pinus
pinnea L stand. Forest Ecol. and Manag., 2: 221- 231.
Rapp M. y Lossaint P. 1981. Some aspects of mineral cycling in the garrigue of
Southern France. En: di , Castrí F., Goodall D.W. and Specht R.L. (eds.),
Ecosystems of the world 11. Mediterranean type Shrublands. Amsterdam.
Elsevier.
Read D.J. y Mítchell D.T. 1983. Decomposition and mineralization processes in
Mediterranean type Ecosystems an in a Heathland of similar s tructure. En:
Kruger F.J., Mitchell D.T. and Jarvis J.U.M. (eds.), Mediterranean Type
Ecosystems: The role of nutrients. New York. Springer Verlag.
Reichle D.E. 1971. Energy and nutrient metabolism of soil and l i t ter
invertebrates. En: Duvigneaud P. (eds.), Productivité des ecosystemes
forestiers. Actes Coll. Bruxelles. París. UNESCO.
Reiners W.A. y Lang G.E. 1987. Changes in litterfall along a gradient in
altitude. Journal of Ecology, 75: 629-638.
Rivas-Martínez S. y Belmonte D. 1989. Sinopsis de Adenocarpus DC
(Leguminosae). Opuse. Bot. Pharm. Complutensis, 5: 69-78.
Roda F. 1983. Biogeoquímica de les a'igues de pluja i de drenatge en alguns
ecosistemes forestáis del Montseny.Universidad Autónoma de Barcelona, tesis
doctoral.
458
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
Rodin L.E. y Bazilevich N.I. 1967. Production and mineral cycLing in
terrestrial vegetation London. Oliver Boyd. 288 pp.
Rundel P.W. 1988. Leaf structure and nutrition in mediterráneas climate
sclerophylls. En: Specht R.L. (eds.), Mediterranean-Type Ecosystems. Kluwer
Academic Publishers.
Sabate S. 1986. Evolució de l'estructura horitzonatal i ocupado de l'espai de
la garriga després del foc Universidad de Barcelona. Tesis doctoral.
Santa Regina I. y Gallardo J.F. 1989. Ciclos bíogeoquímicos en bosques de la
Sierra de Béjar (Provincia de Salamanca). Options Mediterranéennes- Serie A,
3: 147-149.
Santa Regina I., Gallardo J.F. y San Miguel C. 1989. Ciclos bíogeoquímicos en
bosques de Sierra de Béja (Salamanca España): 2. Retorno potencial de
bioelementos por medio de la hojarasca. Revue d'écologie et de Biologie du
sol, 26(2): 155-170.
Satoo T. 1982. Forest biomass Netherlands. Dr. Junk Publ.
Schaefer D.A., Reiners W.A. y Olson R.F. 1988. Factors controlling the
chemícal alteration of throughfall in a subalpine balsam fir canopy.
Environmental and Experimental Botany, 28: 175-189.
Schaefer D.A., Reiners W.A. y Olson R.K. 1988. Factors controlling the
chemical alteration of throughfall in a subalpine Balsam Fir canopy.
Environmental and experimental Botany, 28: 175-189.
Schlesinger W.H. 1985. Decomposition of chaparral shrub foliage. Ecology, 66
(4): 1353-1359.
Schlesinger W.H. y Hasey M.M. 1981. Decomposition of chaparral shrub foliage:
losses of organic and inorganic constituents from deciduous and evergreen
leaves. Ecology, 62: 762-774.
459
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadri l lero
Schmitz M.F. y Bermúdez de Castro F. 1982. Estudios en ecosistemas diatróficos
II. Alisedas del Noroeste de Guadalajara. Bol. Est. Cent. Ecol., 11: 23-36.
Schulman H.M., Lewis M.C., Tipping E.M. y Bordeleau L.M. 1988. Nitrogen
fixation by three species of leguminosae in the Canadian high arctic tundra.
Plant, Cell and Environment, 11: 721-728.
Schweitzer L.E. y Harper J.E. 1980. Effect of light, dark and temperature on
root nodule activity of soybeans. Plant Physiol, 65: 51-56.
Schwintzer C.R. 1984. Production, decomposítion, and nitrogen dinamics of
Myrica gale litter. Plant and Soil, 78: 245-258.
Schwintzer C.R. 1983. Primary productivity and nitrogen, carbón and biomass
distribution in a dense Myrica gale stand. Can. J. Bot., 61: 2943-2948.
Schwintzer C.R. 1984. Production, decomposítion and nitrogen dynamics of
Myrica gale litter. Plant and Soil, 78: 245- 258.
Sharma E. 1988. Altitudinal variation in nitrogenase activity of the Himalayan
alder naturally regenerating on landslide-affected sites. New Phytol., 108:
411-416.
i
Sharma E. y Ambasht R.S. 1987. Litterfall, decomposítion and nutrient reléase
in an age sequence of Alnus nepalensis plantation stands in the Eastern
Himalaya. Journal of Ecology, 75: 997- 1010.
Sharma S.C. y Pande P.K. 1988. Patterns of litter nutrient concentration in
some plantation ecosystems. Forest Ecol. and Manag., 29: 151- 163.
Shaver G.R. 1981. Mineral nutrient and nonstructural carbón utilization. En:
Miller P.C. (eds.), Resource use by chaparral and matorral. Ecol. Studies 39.
Berlín. Springer-Verlag.
Silvester W.B., Balboa O. y Martínez J.A. 1985. Nodulation and nitrogen
460
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
fixation in members of the Rhamnaceae (Colletia, Retanilla, Talguenea and
trevoa) growing in chilean matorral. Symbiosis, 1: 29-38.
Specht R.L. (ed) 1988. Mediterranean-type ecosystems. Kluwer Academic
Publishers.
Specht R.L. y Molí E. 1983. Mediterranean-Type heathlands and sclerophyllous
shrubland of the world: an overview. En: Kruger F.J., Mitchell D.T. and Jarvis
J.U.M. (eds.), Mediterranean type ecosystems. Ecological studies 43. Berlín.
Springer-Verlag.
Stachurski A. 1987. Nutrient control in throughfall waters of forest
ecosystems. Ekologia Polska, 35: 3-69.
Stohlgren T.J. 1988. Litter dynamics in two Sierran mixed conifer forests. 1
Litterfall and decomposition rates . Can. J. For. Res., 18: 1127-1135.
Stohlgren T.J. 1988. Litter dynamics in two Sierran mixed forests 2: Nutrient
reléase in decomposing leaf litter. Can. J. For. Res., 18: 1136-1144.
Swift M.J., Heal O.W. y Sanderson J.M. 1979. Decomposition in terrestr ia l
ecosystems. Studies in Ecology, 5: Blackwell Scientific Publications.
Takeda H. 1988. A 5 year study of pine needle l i t ter decomposition in relation
to mass loss and faunal abundances. Pedobiologia, 32: 221-226.
Terradas J., Prat N., Escarré A. y Margalef R. 1989. Historia Natural deis
Paisos Catalans. 14. Sistemes Naturals Barcelona. Fundació Enciclopedia
Catalana.
Turnbull C.R.A. y Madden J.L. 1983. Relátioships of litterfall to basal área
and climatic variables in cool températe forests of Southern Tasmania.
Australian Journal of Ecology, 8: 425- 431.
Turnbull C.R.A. y Madden J.L. 1983., Relationship of litterfall to basal área
and climatic variables in cool températe forests of southern Tasmania.
461
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

M.J. Moro Cuadr i l lero
Australian Journal of Ecology, 8: 425-431.
Verdu A.M.C. 1984. Circulado de nutrients en ecosistemes forestáis del
Montseny: Caiguda de virosta i descomposició de les fulles. Barcelona. Tesis
Doctoral. Universitat Autónoma de Barcelona.
Vitousek P. 1982. Nutrient cycling and nutrient use efficiency. The American
Naturalist, 119(4): 553- 572.
Vitousek P.M., Fahey T., Dale W.J. y Swift M.J. 1988. Elements interactions in
forest ecosystems: succession, allometry and input-output budget.
Biogeochemistry, 5: 7-34.
Voght K.A., Grier C.C. y Voght D.J. 1986. Production, turnover, and nutrient
dynamics of above- and belowground detritus of world forests. Adv. Ecol. Res.,
15: 303-377.
Waring R.H. y Schlesinger W.H. 1985. Forest Ecosystems. Concepts and
management London. Academic Press. 340 pp.
Weber M.G. 1987. Decomposition, l i t ter fall, and forest floor nutrient
dynamics in relation to fire in eastern Ontario jack pine ecosystems. Can. J.
For. Res, 17: 1496-1506.
West N.E. 1979. Formation, distribution and function of plant l i t ter in desert
ecosystems. En: Goodall D.W. and Perry R.A. (eds.), Arid-land ecosystems.
Structure, functioning and management. Cambridge. Cambridge University Press.
Wheeler C.T. 1971. The causation of the díurnal changes in nitrogen fixation
in the nodules of Alnus glutinosa. New Phytol., 70: 487-495.
Wheeler C.T., Helgerson O.T., Perry D.A. y Gordon J.C. 1987. Nitrogen fixation
and biomass accumulation in plant communities dominated by Cytisus scoparius
L. in Oregon and Scotland. Journal of Applied Ecology, 24: 231-237.
Wheeler C.T., McLaughlin M.E. y Steele P. 1981. A comparison of symbiotic
462
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Bibliografía
nitrogen fixation in Scotland in Alnus glutinosa and Alnus rubra. Plant and
Soil, 61: 169-188.
Whittaker R.H. y Woodwell G.M. 1968. Dimensión and production relation of
t rees and shrubs in the Brookhaven Forest, New York. J. of Ecology, 56: 1-25.
Whittaker R.H. y Woodwell G.M. 1971. Measurement of net priraary production of
forest. En: Duvigneaud P. (eds.), Productivity of forest ecosystems. Proc.
Brussels Symp. 1969. UNESCO.
Wieder R.K. y Lang G.E. 1982. A critique of the anlytical methods used in
examining decomposition data. Ecology, 63 (6): 1636-1642.
Yoda K., Kira T. y Hozumi K. 1963. Self-thinning in overcrowded puré stands
under cultivated and natural conditions. J. Biol. Osaka City University, 14:
107-129.
463
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndices.
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.1 Desfronde mensual (g/ntf) para P^inaster_en su superf
MES D[A AñO Jul hojas
cié repoblada.
f l o r 7 15 86 , J
1 0 . 7 6 T l o r P»» semil la
o 15 8 6 3 2 26.57 1g n -15
9 15 8 6 6 3 „ " 9 8 0 .28 1 0 15 «ó 93 • -
5.04 0 9.14 1 2 n
1 15 K7 1 « , r , „ ¿ ° .06
" " 9 3 7.43 n¿ " 1 5 " 12< 1 2 1 5 86 154 , u ° ° -03
8 7 1 8 5 4 5-70 09 is -« 2 15 8 7 216 7.21 o n ^
1 " 87 2 " 4 " '2 0 o ° t 15 87 275 6.39 o 0° ° I 15 87 305 2.54 46 o ° 7 15 87 336 3-27 rl 1354 "°3
l 15 87 366 7-" tVÍ « 2 -03
o 15 8 7 3 9 ' 48.64 \ 6 " ' * " ° 3
m 1S 87 428 33-63 'sí • 15 8? ™ 23.54 Q° °
15 87 *89 15.13 o Q° ° 12 15 V 519 20.18 15° Q° 2 15 ^ 581 59.73 6 08° 3 15 88 610 12.69 0
6 - ° 8 ° t 15 88 6^1 5.72 o Q° ° l 15 88 671 L O T 15° ° -06 t 15 88 7°2 .98 2l¡ o° 0
18 88 735 2.23 2 2 ! n ° ! 2° 88 768
1n9 22 38 801 28.83 '64 0° ^
25 88 83< 45.98 í 0° -12
11 20 88 860 5 99 is ° 12 22 88 892
1 ^ 8' 955 23"66 '06 15 87 ° t 28 89 1019 28.77 "03 n °
I 31 89 1°52 .86 o a 2 ! 8 9 1081
8 7 «9 115H -1 •, u 0
89 1120 2.63 1.90 I ' " 89 »» 50.8 ' 27 89 1171 „ « -i8 ° -21 89 1171 9.O5 •15 0 0
ñas .46
.28 .15
0 0
.09
.21
.55
.43
0 .03
.21 .64
0 0 0 0 0
.49
.46
.21
0 .09
0 .06 .06 .18 .03
0 .06 .43
0 0 0 0 0
reprod.tot .67
1.25 .55 .06 .03 .18
15.56 0 0 0
.49 14.31 15.29
.46
.31 0 0
.21 6.08
0 .06 .15
2.93 2.26
.55
.76 0
.15
.06 15.93
.03
.03 1.80 1.99 2.63
.15
to ta l
11.89 28.09 12.44 7.49 5.07 9.42
61.45 7.73 5.35 6.39 3.06
17.79 23.39 49.13 33.93 23.54 15.13 20.39 66.28 13.15 5.99 1.16 4.00 4.49
10.12 29.65 46.16 6.18
11.49 39.65 29.26
.89 2.81 4.62
53.44 9.23
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.2 Desfronde mensual (g/m2) para P nior, ig/m¿; para F- mqra_en su superficie repoblada.
«ES DÍA Añn ». -Ano ho jas f l o r „ , - -
37 " * • * « l ° : 7 4 3
P ' a2 1 8
SemHíaSQ ^ - P - t o t a l
8 86 3.653 .371 „ 0° •0 3 0 1 " 9 ^ 2.925
10 < 8 6 7 " 2 6 2 -0 8 9 3.074 030 ° -3 7 1 4.024 " " 9 ' 1 9 2 -134 . 6 9 8 -0
0350 ° 3.178 10.454
" ]l 8 6 6 - ° " 0 o n ° ^ 6 10.039
£ 2 1 - 1 ° 2
2 ? 8 7 3 0 - 5 0 2 -045 u. ,37 o 1 - 6 7 8 22-^80 2 J 8 7 10-781 o o n - ° 5 9 1 4"1 8 2 " .743 4 11 8 ? 3 - 5 3 4 ° 0 n ° ° 10.781
8 7 ^ 0 ^ J 0 0 3.534 6 í 8 7 -9 5° 0 0 o ° 1 - 4 2 6 3.222 t 8 7 -1 6 3 -015 o 0° - ° 4 5 0 . 9 9 5
8 í 8 7 -327 .030 o 0° ° -015 . 1 7 8
9 ? 8 7 3 - ^ 8 -045 2 . 9 1 1 0° ° -030 .356
10 8 7 9 - U 8 0 0 ° 2 "9 5 5 6-1°3
í 8 7 1 2 -3 1 1 -045 0 o ° ° 9-148 8 7 '-** 0 o 0 ° - ° 4 5 12-355
12 5 87 16.469 .045 0 n° -1 0* 0 9 . 3 4 1
3 8 8 3 1 ' 5 1 2 ° 0 030 ° "045 1 6 -«8 3 5 88 4.099 o 0 ^ "2 0 8 -030 31.734 5 8 8 2 - 9 7 0 0 0 o ° ° *-099
6 " 8 8 -5 9 4 0 0 n - ° 8 9 ° 3.059 7 ll 8 8 "7 1 3 - ^ 0 o ° ° "594 8 n 8 8 -3 4 2 -045 o 0° ° -149 .846 9 2° 8 8 • « -045 o 0° 0 -045 . 3 8 6
,0 2 8 8 1 1 -8 5 0 -252 o o ° - ° 4 5 1 - 0 4 0
í 25 8 8 25-171 0 0 ° ° -252 1 2 . 1 1 8 2° 8 8 19-231 .119 0 0° -0 5 9 0 25.230
32 2 2 8 8 25.334 0 0 n° ° -119 19.364
f J 8 9 4 ° - 1 1 0 -327 o n , ° ° 25"334 4 2 89 L619 o o 0°
1 " 6 4 8 -327 42.085
6 ll 8 9 1 - 1 2 9 0 0 n ° ° 1 '619 8 Zl 8 9 -8 0 2 O 0° ° 0 0 , . 1 2 9
9 ,7. 8 9 - 5 % 0 o J ° O . 8 0 2 9 11 8 9 8.865 .K9 0 n ° ° -594 9 27 89 2.272 „ ° ° -°59
0 0 o -149 9.073
0 2.272
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.3. Desfrorde mensual (g / m2) en C. l a u r i f o l i u s . .
MES DÍA AñO hojas flor 7 i5 ,. "ÜJaS . „ t l ° r _ '"*o corteza 15 86 5.26 •50 .03
" « 2 s:2 t f7 °2 11 15 86 -*i ' 'Z 12 15 " -33 ' 2S n l 15 87
15 8 7 "06 0 o n° 3 15 8 7 • < * 0 o n° 4 15 87 -13 0 S ° 5 15 «7 7 ° ° 0
15 87 . U
6 15 87 „ •: -U' 0 87 .21 •01 .01
7 15 87 ,n o* " 7 U ° 87 30.93 14 0
15 87 q -¡i . „ -u¿ 87 5.22 3-61 0 1-03 .03
9 15 87 2.13 10 • 15 «7 „, ° -35 -13 15 87 .87 11 15 87 „» „ "14 -06 87 .08
0 .14
12 15 87 ~ I U -09 87 .26 0 0 0 .22
2 15 88 3.06 03 "í¡ -17
3 15 » „, -° 3 -56 -69 15 88 .23 4 15 88
15 S8 .06 31 .01
0 .10 .14
5 15 «, -I' "U1 0 -12 01 o
6 15 88 .23 •"' ° -06
88 5.63 -18 0 10 25 8« , ,7 ° u -02 H8 2-14 35 1£
11 20 88 .44 'S 'i; -°6
12 22 «» ;; -22 -08 -06
22 88 56 .13 3 7 » .60 • -7 r 4 28 89 si •" -76 -1° 5 31 89
89 .51
• 08 0
09 1.39 1.7Q
6 29 89
98 ¿ 8899 ^ ^ - ' o "
1 1 8 9 17.43 2 25 / * 9 2 7 89 . i 5
2 ; • • "
.44
.08
.02
.31
.17
.23
.18
.06
.03
.30
.03 0 0 0 0 0
.06
.15
.51
.16
.14
.01
.07
.04
0 .20 .11 0
.09
.57
.23
.12
.12
.13
.14 04
rep.tot. .53 .16 .06
1.11 .67 .43 .09 0 0 0
.02
.14 3.61 1.06 .35 .14 0
.22
.58
.10
.01
.01
.01 1.70
.54
.18
.51
.30
.60
.86 1.48 .14 .47
5.10 2.68 .11
total
6.23 21.35 5.69 2.09 1.25 .99
2.16 .11 . .08 .43 .18 .35
34.56 6.29 2.61 1.07 .22 .81
4.85 .62 .59 .14 .36
6.36 24.15 6.03 2.81 .80
1.31 2.13 3.93 1.74 .93
26.44 20.42 .53
DFTOT 21.93 23.15 4.74 3.49 4.17 13.30
19
1.92 2.21 1.57 1.08
22.02 5.41 1.17 1.10 .97
1.78 1.45 .69
2.69 5.07 11.61 12.83 43.20 19.83 6.56 5.93 3.39 25.60 3.80 2.94 5.51 24.52 20.24 1.45
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.4. Desfronde mensual <g/m2) en A. decorticans.
MES 7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 2 3 4 5 6 7 8 9 10 11 12 3 4 5 6 8 9 9
DÍA 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 15 18 20 22 25 20 22 7 28 31 29 7 11 27
AñO 86 86 86 86 86 86 87 87 . 87 87 87 87 87 87 87 87 87 87 88 88 88 88 88 88 88 88 88 88 88 89 89 89 89 89 89 89
DFH 16.21 17.40 4.52 3.13 3.70 10.44 14.82
1.59 1.61
0 0
22.01 5.26 .71 .64 .7:
1.24 .68 .50
2.12 .06 .03
2.20 32.26 7.38 3.21 1.06 .62
3.48 2.51 2.15 4.34
18.83 17.76
.45
DFFL 5.11
.78 0 0 o o o
o .60
1.57 1.08
0 .12 .14 .09
0 0 0 0
.09 4.94
11.52 9.74 2-72 1.33 -42 .17
0 0 0 0
.83 4.91
.90
.87
DFFR
.10 3.83
0 0 0 0
.07
0 0 0 0 0 0
.03 0 0 0 o o o o o
.08 5.85
10.48 2.44
.30
.71
.89
.05
O
O
.53
37
03
DFSE
O
1
.10
.07
.02
O
O
O
O
O
O
O
O
O
.09
O
O
O
O
O
O
O
O
1.38
.51
.08
.26
O
O
O
O
O
O
.04
.06
DFRAM
.45
.14
.11
.29
.46
.86
.11
.33
O
O
O
o .02 .29 .26 .26 .43 .74 .14 .49 .05 .02 .81
1 .10 .34 .09
O 16.01
.97 .53 .26 .16 .66 .02
DFCTZ
.05 O O O O O O
O O o o o
.01
o .02
O .11 .02 .05
O .02 .03
O O
.03
.08
.04
.06 5.22
.27 .26 .08 .10 .51 .03
DFREP
5.22 5.60
.10
.07
.02 O
.07
O .60
1.57 1.08
O .12 .17 .17
O O O O
.09 4.94
11.52 9.82 9.95
12.31 2.93 4.74 2.71
.89
.05 O
.83 5.43 1.31 .96
DFTOT 21.93 23.15 4.74 3.49 4.17
13.30 19
1.92 2.21 1.57 1.08
22.02 5.41 1.17 1.10 .97
1.78 1.45 .69
2.69 5.07
11.61 12.83 43.20 19.83 6.56 5.93 3.39
25.60 3.80 2.94 5.51
24.52 20.24 1.45
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.5 a. Valores mensuales de la concentrac d e p- Pinaster
ion química (%) de la hojarasca
jul
7
S
9
10
11
12
1
2
3
4
5
6
7
8
9
10
11
12
2
3
4
5
6
1
32
63
93
124
154
185
216
244
275
305
336
366
397
428
458
489
519
581
610
641
671
702
hojarasca (g/m2.día)
.359
.857
.380
.248
.163
.305
1.474
.233
.176
.206
.085
.106
.249
1.569
1.085
.785
.488
.673
.963
.437
.184
.034
.032
.350
.330
.260
.260
.260
.330 .330 .340 .340 .320 .470 .400 .450 .300 .280 .270 .440 .290 .540 .450 .400 .390 .370
P(%)
.062
.057
.030 .030 .032
.036
.036
.084
.084
.013
.043
.040
.032
.048
.030
.030
.048
.016
.032
.031
.039
.040
.026
.125
.125
.112
.112 .082 .074 .074 .109 .109 .114 .160 .167 .167 .117 .102 .102 .034 .049 .044 .053 .079 .139 .052
Ca(%)
1.135 1.135
1 1
1.155 1.130 1.130 1.135 1.135 1.150 .820
1.050 1.050 1.030 1.180 1.180 1.192 1.020 1.210 1.166 1.112 1.237 1.363
Mg(%)
.210
.210 .220 .220 .190 .210 .210 .200 .200 .190 .190 .190 .200 .210 .215 .215 .169 .188 .156 .157 .219 .160 .188
Na(%)
.038
.038
.026
.026
.031
.027
.027 .032 .032 .029 .034 .027 .027 .018 .019 .019 .018 .020 .010 .020 .022 .019 .011
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.5 b. Valores mensuales de la concentración química (%) de la hojarasca de P. nigra.
j u l
7
"<%) P(%) K(%) Ca(%) Mg<%> Na(%)
8 „ -46° - ° 2 7 -0 8 6 1-°20 KO 9 « - 1 1 8 - 4 6 ° - ° 2 7 - 0 8 6 1-020 4 -° 6 3 " 2 3 4 - 3 3 0 -033 133 M* ' i " - 0 1 3
1 0 9 3 - 3 0 6 - 3 3 ° -033 ' 3 3 " 6 6 ' , - ° 1 3
« S -195 -41Q -039 :™ - ' ™ " - - --So0 -o0:; - s • - • - -
22 i: •*• -=° •'- : s .i -s -o°;:
4 Sí -126 - 5 3 ° - ° 4 5 -060 -370 7 o * - : 0 °3 5
28 - = 0 - 0 ° ; 5
5 • - - - . - o 0 ; :
5 £ -005 :2 •'- - :S5 - -£ 7 3 6 6 - 0 1 1 •"<> .072 .198 879 213 ' n S
' - •» - :S -S :£ ; -" s z i:: -s :°s :S - ° -2 58? lo» ' Í l •"" - ° 7 ' • « " • ' « -Mí 3 « ' t ' í -06' -681 • » 4 •«« : S; - - :¡£ :£ - -™ -' S :E :S ;s? •«• '•"• •-« •» •080 .860 .135 .009
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

Apéndice 3.5 c. Valores mensuales de la concentración quími
de C. laurifolius.
7
8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 2 3 4 5 6
jul
1 32 63 93 124 154 185 216 244 275 305 336 366 397 428 458 489 519 581 610 641 671 702
hojarasca (g/m2.dfa)
.175
.680
.180
.023
.013
.011
.061
.002
.002
.004
.005
.007
1.031
.168
.069
.029
.003
.009
.049
.008
.010
.002
.007
N(%)
.385
.385
.280
.280
1.200
1.210
1.210
1.100
1.100
1.100
1.100
.950
.420
.400
.400
.398
.938
.938
1.378
1.329
1.272
1.194
1.243
P(%)
.098
.098
.063
.063
.057
.166
.166
.100
.100
.100
.130
.130
.112
.059
.057
.057
.046
.046
.095
.100
.092
.079
.039
(%) de la hojarasca
K(%) Ca(%) Mg(0/0 Na(%)
.272
.137
.099
.099
.049
.250
.357
.357
.328
.238
.184
.400
.272
.137
.099
.099
.049
.250
.357
.328
.238
.184
.400
1.665
1.665
1.650
1.650
1.315
1.130
1.130
1.150
1.150
1.150
1.160
1.160
1.840
1.650
1.774
1.774
1.662
1.201
1.004
.923
1.138
1.108
.914
.250
.250
.300
.300
.180
.240
.240
.200
.200
.200
.195
.195
.291
.258
.204
.204
.163
.179
.220
.199
.219
.219
.217
.016
.016
.025
.025
.013
.013
.013
.017
.017
.017
.020
.020
.009
.010
.008
.008
.009
.008
.009
.009
.031
.020
.021
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

'*"""3¡ '• rn.™:: - - — — < » *..*,„...
7
Jul hojarasca N(%) P(%) K(%) (g/m2.día)
C a ( %) Mg(%) 1
m
Na<%)
,n - U Ó 1-590 046 ¡gn -260 -01* - m 1 - 6 0 0 -'oso • • ; ; ;
: 124 -H9 3.870 218 Un -250 -020 .348 2.820 ; 9 '"J
185 -478 2.6.0 54 '™ 1 -210 .023 2 m m - - „ -71° -46° -«O .016 3 244 -027 3 ... c
í 2 7 5 - ° 5 2 2.770 " jo Í l l -6 2° "250 .016 l 30S 0 . B - 1 5 ° m -4 4 2 -590 . 2 1 0 .0 0 Q 6 336 0 m m ^ l 366 -ni> 1-870 047 , , , m
l 3 9 ? -170 1-960 ' 0 5 6 -64 3 : - ° ° 6 -164 .006
10 4 2 8 " ° 2 3 2.250 089 ' 2 0
1 - W ° -1 7 2 -018 458 -021 2.250 089 1 L -956 -168 ^ 489 " ^ 3 " 2 9 " S -956 -168 -008
12 5 1 9 - ^ 3 " 2 9 -3 9 9 -8 0 2 -186 .006 2 581 -011 2.793 -34 11 - 8 ° 2
3 610 ni7 * — -6 3 4 -596
0 1 1 2.793 .134 a¡ f ¿ "1 8* -006 0 1 7 3.043 109 t í? -5 9 6
641 nzo , „„, -1UV -284 . 7 6 1
, — -017 3.043 109 "pT - Í 9 6 -203 -008 í " 1 •«« 2.906 22 i ' -761 -1 6 9 -013
6 7 1 -0 0 2 2.900 ' 1 2 2 • ' ! * -730 -209 .009 6 7 0 2 o m m " 1 2 2
m -3 8 6 -700 . 1 8 0 . 0 0 9
m
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

K > o u-i o co m r\j - j - o u-\ N- c\j ^ N- O o m c \ J > O i n c o r o
«— ••O ^O v}- sfr <*• f\J O T~ o «4- ro -3- o
fO » - N fO >0 m K )
O f \ l o r v j f ^ o o o o o o o o o *— ro o o
m f \ i * *
ÉT N r - W PJ » - r -
o j f O O > o o r o o r o c o o
N n i <- r - i -
eo 1/1 eo « -
ro
O O O O O O O O O O O r O O O f O N l
I I t I I I I I I I I I Mf O O O O O o o o o o O O O O O O O O O O I O O O S - O J O O O o
1 U U J U J U J U J U J U I U J o o o o o o o o o o o o o o o o
o o o o o r o o o ' + + + + + > + +
I I I I I I I I I I I I I I I I I I I I I I I I I I I o o o o o o o o o o o o o o o o o o
- o o
-o -o M *- eo fn o ro o rv
i - N i n i - C O O N O t - C O i n N I O W N C O ' O f l í o 3 o o o o o « - - * N O r ^ r o i n o r \ J o > 3 ' t — o * - o
N- eo CO 00
o
(\i ro T - r\i r\j * - » - m r - (M « -
c
Q.
o i n o o eo
>-* z
o <\J t n o M os í R r ^ r o i n t n o u - i L n o p o o o * - * — « — fO IM r- r- (\J
ÍC f -sO
N-O
N. i n
o i n
vi *0
o co
o o
vn OJ
r\j ro
m C\J
co ~ü
VQ V^ T-
m oo *•*• *o co
K> 00 N- "O O
m o -* *- ro N 0 O S - O O - J " ^ > d - r \ j T — O- f\l S 0»> m K1 O -r- *— C> O O N- O ^ Kt
C W T-o j c o r o r O T - ^ o c o L n
8. S
C 0 a 3 C 0 C 0 C 0 C 0 C 0 C 0 C 0 c 0 C 0 0 0 0 0 0 s í > 0 * O O O O O O O O c a o o c o o o c o c o c a a o c o c o c o c o c o c o c o o o c o o o o o c o c o c o c o c o
^ ^ r - < r i n m * o ^ o o o r \ j « - T - f \ ] m f o - ¿ - i n ^ o o o o
I <C *— ÍNJ *— r\j ro f *— Í M * - r - r v j r v r o m
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

CJ
< •
z
<
3 • *
—i
,— -
o
o SO
-* (NJ
O
C3
O ÍNJ
tn r\j
i
O
v t o
so t n
o
t n >j-
o
o
f^ ru
m r\j
rvi i
o
m LH
-.1" O
O
ro -d-
o> r\i
O r\l
o
t n o
r--co
<>>
o
ru
"*;
T —
ON
i
O
•d-M
«— fO
o
co m
o o
Í \ I
o
c -c>
^3--d-
o
o o
r\j •o
o
- o o
T —
CO
o
- r y>
r\i O
O
M f >
O
*—
O tn O
o •o
o
o
* 1 ^ í
m i^-
o o
- j -o
(M CO
> í
O o
r\j
*~
^r t n
IM r
O
-O t n
en V)
o
o t n
CO ÍO t_
•*-•
c <u
c/i o
-M a» c
w 0J
L .
<
o
OJ X
- J
,-» u K -1 ^
rtj z V . u
s
« - >» (M
~ o c s i n * a - e o o N - o
O. -o f 0 0 c O K i o » - r j t n O t n o o r ^ - ^ í r O ' —
ru *— «—
o o o o o o o o + + + + + + + + L U U J L U L U U J U J L U U J o o o o o - ~ —
o o o o o
o o r o o o o o o o Ú J l U U l U J I t l U l U J I U U I l i J o o o o o o o o o o o o o o o o o o o o
ÚJ 1Ú tu UJ l u UJ o o o o o o o o o o o o
o o o o o o
O O O r-
tn ro M tn
O O O O C M O O O O O O O O O O
O O O * — f \ J C \ l í \ J r \ J r \ i
r j o o
<: u
> í
• on ro ,— - t
t n •
o tn
<* v f o
•
v f v f *-
" O r\j
• í^ t n
tn V ) O S (V W m (V r - O K) *—
f \ J * 0 ^ * - * - « — «— ' < — «~ "— CM
O « - N . O CJ>
•o «- o *J o •O [O tO N IM •» m o -O o o r o o - j - O " — o t M o r o <3
o f\J
o m
in o
o p -
L/l r—
r\j T —
m o
r\i ur ^ en
o o •c—
o M o -a- ro o r- o
^O 00 N- CO CO o-, m m <t p-N. (N «- Kl 'J- o -J- -s- - Í
ru N- PJ o*
C O C O C O C O C O C O C O C O C O C O C O C O C O O O O O O ^ C N C N O O c o c o c o c o c o c o c g c o c o c o o o c o c o c o o o c o c o c o c o c o o o c o
> * - s 3 - N j - u - \ u i ' 0 , o o ^ o o r - T - í > j t — r o f o - ^ r i n ^ o ^ o o
i - ru r- r\i £%!•*- •— t— oj t\j r j m
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

in í\J s j >o O »-t\j ro O N- co co
r>- *--j- so o -o s»- o r-- eo
o o ro r*. o so
I o i n r o o o p s i o P s i » — o o o c o o r s . v j ' s o o i n v j - o o
o o * * u-» o ro «- ro o
ví- so rs. o - j - -d- o o -a-
«- «- psl eo t> O 10 1M sO O rO
M r- «-
O O O O O rO O O O O O O + + + + + + + + + + + i + III MI MI III III m ni III m III ni III III
O O O O O O O O O O O O O
o o o o o o o o o o o o o O O O O O O O O O O O P s l O O > -
O r O r O O O O r O O
o o o o o UÍ 111 UJ Lu
O O O v- O O
Os LO LO
- C
^ Oí V s ^
« c co CJ
•W ¿_
o u 01
T 3
•a:
< u z «t
V
z <
<
en o
P J
ro
*""
PU
o
o ps l
P J ,—
o
1— r \ j
ro o
i n m
r--ro
ro ^
ps l
o
o i n
o
so o
*— - j -
saca
ro i n
r -a-
rvi
i n s O
r~-ÍM
Psl
m psl
rO
ro o
s f
•»!)
N"> v j -
t\J c\i
o. o
*— s o
<M
s O
*~
CO N .
Psl s O
~ í
•O os
o sr
m ro
c—
sd-
o
ro o
r -u- i
O s
O
ro o psi
o o
en o
*— r \ j
o Psl
ro o-
s O
ro
o o
ro ^ í
i n r>-
i n o
ro
P J
o
ro v j -
ro
ro
v í -
sO
<>• psl
r~-v t
ro o
a i o
c/> o Psi u-i o ro os O PO «- >—
-— o co o o
*-" c <u y>
o 4» C
t—
c \ j
5 —1
,-* 3 (M
,
LO s ü
psi
«~
co cu
,-
r—
ro LA
s^-
s o CM
C0
o •sf -sT
u-t
S . • s í
Psl
*~
en
SO
*~ co
*~
LO
O
ro ro ro
sO
*~ ro rsi
o
SsT
co
,— *—
ro s í
ro i n
s O s O
tu ro
irt i n
co ro
co ro
s O O s
irt sO
ro
o u-t
rs.
co
f s .
ro ro Psl
LO s i )
- s í
*~ r» LTl
, „
ro
sO u» r~-rvj
N .
v i O
N í
ro
Psl -ST
Psl Os
i n s ü
f -
,-r~-i n rvj
-o -O
Os o
o eo
P J
o LO
*~
P J
N -
ro ro PU
8. o o a a c o a a c o a j o o a a a D c o c a c o c o o o o o o o o o o O c o c o c o c a c a c o c o c o c o c o c o c a c o c ú c a c a a o c o c o o o c o c a
<
• s 3 - s f " < r i n L n > a ' » o o o O ' c - í — ru ro -«r in >o o o o
•I O * - (\| «r- f\J t \ i r ~ T - fM r - f U í M f N J fO
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992

-o -o o o
N - T — OD CO < - 1/1 r - C Q s O C O O O
v- OJ r- «— i <— rsj T—
t \ j ro
• j - ro
in o o o PJ o o o «- to
« - ~» o o i n o o o o o co - J -
o « -o —
m r- >j-- o o p- i* . f - f- ir\
•o T - f\j i n o <o o •* «j-O f1^ O ^ f K>
IM 1/1 W N w í y
CO N- - * O I*- > - (\J P l i n ^ f N u-i o o ro T - co * -
I OJ Í\J NQ 00 o o ro ro
O in N ^ Í m o
o o o o o o o o o o o f o o r o o o o o o f i m o
U J U U U J U i U J U J U J L l J o o o o o o o o o o o o o o o o O O O O O O O O
a i U J tu a i uu tu o o o o o o o o o o o o
O O O O O O O O O O O O O i - O O O O O - r - f U O
i n eo «O * 0 » - • - r - o o « — o « - f o r o c o *— r u r ^ - o p o o o o * — w r\j r o ro <M w
r*- i n PO g-i m co N-o o o o ^o ro o
CJJ
z
ro o
•
ro o
1
*— i
C\J
i
á •
NI S3
O J
ro ro p j
o
«-
o
P J
CO
ro r -
CO
i
u-i p-tSi 1 -
•
^Z i
1
to o
'
f O
1
o f \J
PJ O
P J
P J
i
C V J O C O O O - ^ O C O r - C O O ^ O N - O ' O O O ' J - ' d '
r\j , - * - * -
o a o co -sf
(0 I -
Ü3 o u
o
o
rvi
o PJ
>* f O
p j
p -
,-
t n
^3-CO
<-
O
O
>o
ro
K )
ro •o
t>-
t >
vO
~*
<r
ro o ru
R
vO
>cf
T- O O o r o • * - J v r o T— > ^ n > S i / i w w y i SÍ re
t > o i n N Í ' - • J - r O O J U l r - C O C O ' -
<U - 1
o ^ m
8. C 0 C 0 C 0 C 0 Q 0 C 0 C 0 C 0 C 0 C 0 C 0 C 0 a « 0 > ' l > í > O > . c o e o c o c o c o c o c o c o c o c o c a c o c a c o c o o o c o
t n i n ^ o ^ o o ^ o O ' — *— r\j f i r o - í i n o o o o
"8 8. < f M N - s O ' — K l ^ O C O
( \ l T ~ •<— vo >o T - •<— r*- h - o -<r sj- N- o
Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral... Mª José Moro Cuadrillero
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d'Alacant. 1992





























