Embed Size (px)
Citation preview

Diagnosis and Prognosis of Male Infertility in Mammal: The Focusingof Tyrosine Phosphorylation and Phosphotyrosine ProteinsWoo-Sung Kwon, Md Saidur Rahman, and Myung-Geol Pang*
Department of Animal Science & Technology, Chung-Ang University, Anseong, Gyeonggi-do 456-756, Republic of Korea
ABSTRACT: Male infertility refers to the inability of a man to achieve a pregnancyin a fertile female. In more than one-third of cases, infertility arises due to the malefactor. Therefore, developing strategies for the diagnosis and prognosis of maleinfertility is critical. Simultaneously, a satisfactory model for the cellular mechanismsthat regulate normal sperm function must be established. In this regard, tyrosinephosphorylation is one of the most common mechanisms through which severalsignal transduction pathways are adjusted in spermatozoa. It regulates the variousaspects of sperm function, for example, motility, hyperactivation, capacitation, theacrosome reaction, fertilization, and beyond. Several recent large-scale studies haveidentified the proteins that are phosphorylated in spermatozoa to acquirefertilization competence. However, most of these studies are basal and have notpresented an overall mechanism through which tyrosine phosphorylation regulatesmale infertility. In this review, we focus of this mechanism, discussing most of thetyrosine-phosphorylated proteins in spermatozoa that have been identified to date.We categorized tyrosine-phosphorylated proteins in spermatozoa that regulate male infertility using MedScan Reader (v5.0) andPathway Studio (v9.0).
KEYWORDS: diagnosis, prognosis, male infertility, tyrosine phosphorylation, sperm protein
1. INTRODUCTION
Infertility is a common problem in humans and animalsworldwide that is caused by reproductive factors in male,female, or both. Male infertility is involved in approximately40% of infertility cases.1 The WHO Laboratory Manual for theExamination of Human Semen and Sperm-Cervical MucusInteraction2 is widely used to evaluate semen samples inlaboratories and hospitals. Consequently, conventional meth-ods for the prediction of male fertility based on sperm featuressuch as total and progressive motility,3 morphology,4,5 andDNA quality6 have been studied extensively in the males ofdomestic species, including bulls,7,8 pigs,9 and rams.10 However,conventional semen analysis cannot precisely diagnose fertilityor infertility or determine prognosis in the case of abnormalsperm function or molecular defects in spermatozoa.11
Therefore, research related to sperm function and male fertilitymust establish more accurate diagnostic and prognosticmethods for the management of male infertility and/orsubfertility.Spermatozoa are terminally differentiated and highly
specialized cells. A mature spermatozoon is the specific outputof the testes that can fertilize a functionally mature oocyte inthe female reproductive tract during sexual reproduction. Thesperm maturation process is predominantly regulated bymultiple signaling cascades associated with tyrosine phosphor-ylation.12−19 Several reports have postulated that, as the mostcommon post-translational modification, tyrosine phosphor-ylation extensively regulates sperm motility,20−22 hyperactiva-tion,17,23 chemotaxis,24,25 capacitation, the acrosome reac-
tion,14,18,26−28 and sperm-zona pellucida-binding.17,29 It thenfacilitates the subsequent fertilization of an oocyte. Therefore,understanding how tyrosine phosphorylation regulates theseevents in spermatozoa during successful fertilization by fertilemales is of paramount importance.Previous in vitro and in vivo studies have shown that
mammalian spermatozoa must undergo a series of complex,stage-specific events for successful fertilization.30 Spermatozoaare produced in the testis, and basic maturation occurs duringtheir epididymal passage.31,32 Consequently, spermatozoabecome hyperactivated, with characteristic amplified asym-metrical flagellar beating, which pushes them toward the oocytein the female reproductive tract.33−35 The next step iscapacitation, which is a comparatively more complex and lesswell understood maturational process. Only capacitated spermcan undergo the acrosome reaction, a process that enables asperm to penetrate and fertilize an egg.17,18,36,37 Recent studieshave demonstrated that all these aforementioned events areregulated by multiple signaling cascades and that tyrosinephosphorylation is involved in almost every step. Yanagima-chi38 has reported that spermatozoa from the caput epididymisare unable to fertilize eggs because they lack protein tyrosinephosphorylation activity. This finding suggests that spermato-zoa gain their tyrosine phosphorylation capability during
Special Issue: Proteomics of Human Diseases: Pathogenesis,Diagnosis, Prognosis, and Treatment
Received: May 28, 2014Published: September 16, 2014
Reviews
pubs.acs.org/jpr
© 2014 American Chemical Society 4505 dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−4517

epididymal maturation near the time of fertilization. Si andOkuno39 identified four major tyrosine-phosphorylated pro-teins in sperm flagella that are correlated with motility andhyperactivation. Simultaneously, Visconti and colleagues40,41
successfully evaluated the effect of protein tyrosine phosphor-ylation on capacitation and the acrosome reaction. In addition,Rajesh and Naz42 have reported a relationship between tyrosinephosphorylation and sperm−zona-binding. Therefore, one can,without considering additional signaling cascades, assume thatmanipulation of tyrosine phosphorylation might offer astraightforward approach for regulating male fertility.Recent studies have identified several proteins that are
phosphorylated in spermatozoa.16,17,43 Additionally, Schumach-er et al.44 identified 99 proteins in normozoospermic humanspermatozoa, and the phosphorylation status for most of themhas been determined. Therefore, attempts should continue toisolate additional proteins that are phosphorylated inspermatozoa from the time of their production in the testisuntil fertilization to understand the signaling cascadesassociated with tyrosine phosphorylation and male fertility.We recently reported that blocking or manipulating thefunction of proteins in spermatozoa, such as ubiquinol−cytochrome c reductase core protein 2 (UQCRC2),13 voltage-
dependent anion channels proteins (VDACs),14 argininevasopressin receptor 2 (AVPR2),15 and actin-related protein2/3 complex,45 alters motility, capacitation, the acrosomereaction, and fertilization by tyrosine phosphorylation. There-fore, studies with similar designs might identify novel proteinsthat are tyrosine-phosphorylated or affect tyrosine phosphor-ylation to alter male fertility. The current review summarizesthe basic mechanism and importance of tyrosine phosphor-ylation in the regulation of sperm function. In addition, wefocus on the sperm proteome related to tyrosine phosphor-ylation identified and illustrated by MedScan Reader (v5.0) andPathway Studio (v9.0), respectively, and its potentialimplications for the diagnosis and prognosis of male infertility.
2. PROTEIN PHOSPHORYLATION AND TYROSINEPHOSPHORYLATION IN SPERMATOZOA
Phosphorylation is a well-studied post-translational modifica-tion of proteins in which protein kinases phosphorylate atserine, threonine, or tyrosine residues by covalently adding aphosphate group.46,47 This phenomenon has been describednot only in eukaryotes but also in bacteria and viruses.36
Phosphorylation occurs through the addition of a phosphategroup to a protein (or other organic molecule), can induce
Figure 1. Tyrosine phosphorylation and related events in mammalian spermatozoa (from production to sperm−zona interaction). Abbreviations:BSA, bovine serum albumin; GF, growth factor; IL, interleukin; P4, progesterone; PAF, platelet aggregation factor; TP, tyrosine phosphorylation;ZP, zona proteins.
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174506

major conformational changes, and may predispose theseproteins to either activation or inactivation. Phosphorylationplays vital roles in almost every cell function, includingmetabolism, transcription, cell-cycle progression, differentia-tion, cytoskeleton arrangement, apoptosis, intercellular com-munication, ionic current modulation, receptor regulation, andneuronal and immunological functions, among others.17,42,46−48
Therefore, phosphoproteomics has been established as a branchof proteomics that focuses solely on the identification andcharacterization of proteins that can be phosphorylated(Figures 2, 3, and 5).Mature spermatozoa are widely known to be transcriptionally
and translationally silent27,49 or barely capable of translation.50
Therefore, post-translational modification is potentially im-portant for the regulation of several physiological functions ofspermatozoa required for fertilization, such as motility,hyperactivation, capacitation, the acrosome reaction, andsperm−egg interaction.51−55 A literature review showed thatserine/threonine and tyrosine phosphorylation of proteins havebeen reported in spermatozoa and that the latter is criticallyimportant and might be the primary inducer of signaltransduction pathways.42 Despite enormous advances insperm phosphoproteomic research,44,56 the mechanisms ofphosphorylation, especially tyrosine phosphorylation, inmammalian spermatozoa remain poorly understood. Capacitat-ing human spermatozoa reportedly shed their sperm cytoplasm,which contains several growth factors/cytokines, includinginterleukin-2 (IL-2),57,58 interleukin-6 (IL-6),59 interleukin-8(IL-8),60 colony stimulating factor-1 (CSF-1),61 interferon-g(IFN-g),62 tumor necrosis factor-a (TNF-a),63 epidermalgrowth factor (EGF),64 Ta1,65 and thymosin β4 (Tβ4),66 inaddition to several unidentified factors. Several of these factorsare also present in the cervical mucus of the femalereproductive tract.61 In addition, bovine serum albumin(BSA), which is frequently used in medium to capacitatesperm, has growth factor-like activity.66 In fact, these moleculestrigger the phosphorylation of various membrane proteins andreceptors after binding to the sperm membrane.67 A model ofsperm tyrosine phosphorylation, including the several matura-tional events induced by this process, is depicted in Figure 1.
3. DIAGNOSIS AND PROGNOSIS OF MALEINFERTILITY INVOLVED IN TYROSINEPHOSPHORYLATION
Phosphorylation is among the most common regulatorymechanisms for protein function, regulating cell functions byinducing conformational changes in proteins via allostericmodification. Tyrosine phosphorylation triggers several phys-iological events in spermatozoa, such as hyperactivation,capacitation, and the acrosome reaction, that are necessaryfor fertilization both in vitro and in vivo; thus, it can alter malefertility (Figure 1).17 However, the mechanism through whichtyrosine phosphorylation/tyrosine-phosphorylated proteinsregulate intracellular signaling in spermatozoa and the relevanceof such signaling for the diagnosis and prognosis of maleinfertility remain poorly understood. We discuss this topicbelow.
3.1. Sperm Motility and Hyperactivation
The capability of a spermatozoon to move progressively towardan egg is defined as motility. Motility is a factor in successfulpregnancies because only motile spermatozoa can fertilize anoocyte; poorly motile or immotile spermatozoa cannot.23,68
Hyperactivation is a vigorous swimming pattern in spermato-zoa34 characterized by increased amplitude and asymmetricalflagellar bending that aids the release of spermatozoa fromoviductal storage to boost them through the mucus of theoviductal lumen and the matrix of the cumulus oophorus forfertilization.23,68 Hyperactivation of spermatozoa is believed tobe a capacitation-associated phenomenon (described in thenext subsection); however, abundant experimental data suggestthat these phenomena are independent.69−71 The originalconcept of sperm hyperactivation was described in 1970 by theremarkable findings of Yanagimachi in golden hamsterspermatozoa in an in vitro setting.68,72 Despite extensiveresearch on sperm motility, little is known of the molecularmechanism underlying hyperactivation. A literature reviewshowed that the acquisition of sperm motility and hyper-activation may be related to the tyrosine phosphorylation ofseveral sperm proteins along with numerous other fac-tors.17,55,73,74 In this section, we summarize the publishedstudies on the possible correlations among sperm motility,hyperactivation, tyrosine phosphorylation, and male fertility.In 1999, Mahony and Gwathmey showed that treating
spermatozoa with genistein, a tyrosine kinase inhibitor,significantly decreased tyrosine phosphorylation levels ofsperm tail proteins, thus blocking subsequent hyperactivation.73
A similar finding was reported in more detail in anotherstudy.39 Four major tyrosine-phosphorylated proteins inflagellar extracts (∼90, ∼80, ∼62, and ∼48 kDa) that werecorrelated with sperm motility and hyperactivation have beenidentified.39 In addition, the incubation of the spermatozoa witheither protein kinase A (PKA) or a protein tyrosine kinaseinhibitor efficiently prevents both hyperactivation and thetyrosine phosphorylation of flagellar proteins, whereas proteinphosphatase inhibitors reduce protein dephosphorylation andthe subsequent increase in hyperactivation.39 Conversely,protamines are the major nuclear proteins in spermatozoa.The nucleus of human spermatozoa comprises two protaminetypes: protamine 1 and protamine 2.75 Changing the expressionof sperm protamines reportedly plays a key role in theregulation of sperm motility and fertilization.75,76 Growingevidence suggests that protamines are involved in thecondensation of sperm chromatin and might remodel thesperm head shape, responsible for fast swimming velocity, andtherefore generate more efficient sperm.76,77 Aleem et al.78
reported that tyrosine phosphorylation occurs in rat spermprotamine during transit to the cauda epididymis and thuscould regulate fertilization. These independent experimentsestablish a strong correlation among motility, hyperactivation,tyrosine phosphorylation, and fertilization.Most studies on sperm motility have focused on the effects of
cyclic adenosine monophosphate (cAMP) and calcium orpH.23,79−82 Cyclic adenosine monophosphate has the potentialto stimulate sperm motility via activation of a cAMP-dependentprotein kinase.55,83 Cyclic adenosine monophosphate isbelieved to induce protein phosphorylation in spermatozoaand subsequently enhance sperm motility. Therefore, tyrosinephosphorylation of sperm proteins may be one of the mostcritical regulatory mechanisms for hyperactivation in sperma-tozoa. In addition, Nassar et al.55 have reported that theincubation of spermatozoa with pentoxifylline significantlyenhances hyperactivation, which is positively correlated withsperm tail protein phosphorylation. However, the authors didnot discuss the effects of protein phosphorylation/proteintyrosine phosphorylation on hyperactivated sperm motility.
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174507
A-kinase anchoring proteins (AKAPs) are tyrosine phosphor-ylation related proteins that form a major component of thefibrous sheath.43 One study reported that mice lacking AKAP4and their wild-type counterparts have a similar number of viablespermatozoa; however, the motility of the AKAP4-deficientspermatozoa decreased by nearly 10% and accompanied bysluggish flagellar motion and low amplitude, and these mutantssubsequently failed to progress.84 The authors suggested thatthe decreased motility and hyperactivation were related to ashortened, absent, or substantially reduced fibrous sheath.However, AKAP4-deficient spermatozoa may also reduce thecyclic adenosine monophosphate (cAMP)-mediated phosphor-ylation of flagellar proteins, which prevents the vigorousflagellar movement with a high-amplitude waveform thatcharacterizes hyperactivated motility.72,84 Sperm motility islost in the absence of AKAP4 because signal transduction andthe activity of several glycolytic enzymes associated with the
fibrous sheath function improperly.84 In addition, AKAP82, itsprecursor pro-AKAP82, and a fibrous sheath protein of 95 kDa(FSP95) are reportedly the most prominent tyrosine-phosphorylated proteins involved in the regulation of spermmotility and hyperactivation.17,85,86
Recently, we reported that the treatment of mousespermatozoa with nutlin-3a, a small molecule antagonist ofthe mouse double minute 2 repressor (MDM2), downregulatesUQCRC2 and the ∼100 and 60 kDa tyrosine-phosphorylatedproteins, which correlates with reduced sperm motility,hyperactivation, and early embryonic development during invitro fertilization.13 This study provided evidence that spermmotility and hyperactivation are partially regulated by tyrosinephosphorylation. However, an interaction between UQCRC2and tyrosine phosphorylation have to be demonstrated.Moreover, another study has proposed that Deamino [Cys 1,D-ArgS] vasopressin (dDAVP), an AVPR2 agonist, significantly
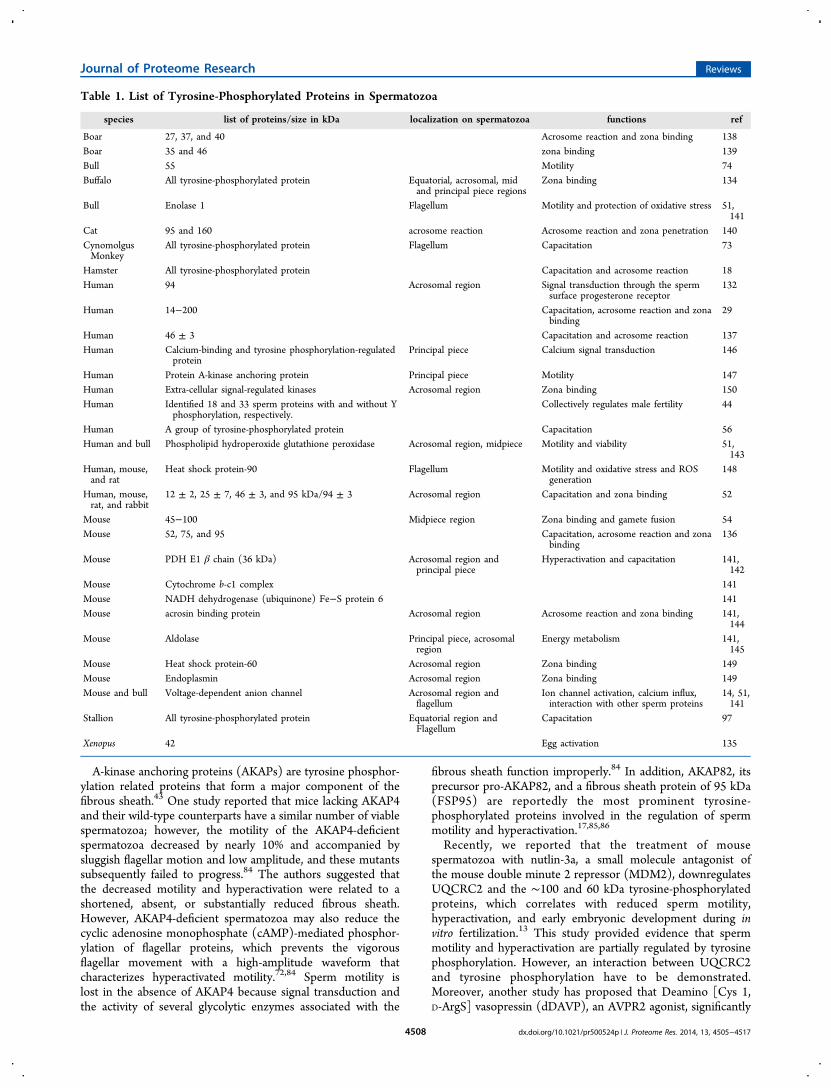
Table 1. List of Tyrosine-Phosphorylated Proteins in Spermatozoa
species list of proteins/size in kDa localization on spermatozoa functions ref
Boar 27, 37, and 40 Acrosome reaction and zona binding 138Boar 35 and 46 zona binding 139Bull 55 Motility 74Buffalo All tyrosine-phosphorylated protein Equatorial, acrosomal, mid
and principal piece regionsZona binding 134
Bull Enolase 1 Flagellum Motility and protection of oxidative stress 51,141
Cat 95 and 160 acrosome reaction Acrosome reaction and zona penetration 140CynomolgusMonkey
All tyrosine-phosphorylated protein Flagellum Capacitation 73
Hamster All tyrosine-phosphorylated protein Capacitation and acrosome reaction 18Human 94 Acrosomal region Signal transduction through the sperm
surface progesterone receptor132
Human 14−200 Capacitation, acrosome reaction and zonabinding
29
Human 46 ± 3 Capacitation and acrosome reaction 137Human Calcium-binding and tyrosine phosphorylation-regulated
proteinPrincipal piece Calcium signal transduction 146
Human Protein A-kinase anchoring protein Principal piece Motility 147Human Extra-cellular signal-regulated kinases Acrosomal region Zona binding 150Human Identified 18 and 33 sperm proteins with and without Y
phosphorylation, respectively.Collectively regulates male fertility 44
Human A group of tyrosine-phosphorylated protein Capacitation 56Human and bull Phospholipid hydroperoxide glutathione peroxidase Acrosomal region, midpiece Motility and viability 51,
143Human, mouse,and rat
Heat shock protein-90 Flagellum Motility and oxidative stress and ROSgeneration
148
Human, mouse,rat, and rabbit
12 ± 2, 25 ± 7, 46 ± 3, and 95 kDa/94 ± 3 Acrosomal region Capacitation and zona binding 52
Mouse 45−100 Midpiece region Zona binding and gamete fusion 54Mouse 52, 75, and 95 Capacitation, acrosome reaction and zona
binding136
Mouse PDH E1 β chain (36 kDa) Acrosomal region andprincipal piece
Hyperactivation and capacitation 141,142
Mouse Cytochrome b-c1 complex 141Mouse NADH dehydrogenase (ubiquinone) Fe−S protein 6 141Mouse acrosin binding protein Acrosomal region Acrosome reaction and zona binding 141,
144Mouse Aldolase Principal piece, acrosomal
regionEnergy metabolism 141,
145Mouse Heat shock protein-60 Acrosomal region Zona binding 149Mouse Endoplasmin Acrosomal region Zona binding 149Mouse and bull Voltage-dependent anion channel Acrosomal region and
flagellumIon channel activation, calcium influx,interaction with other sperm proteins
14, 51,141
Stallion All tyrosine-phosphorylated protein Equatorial region andFlagellum
Capacitation 97
Xenopus 42 Egg activation 135
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174508

decreases sperm motility (%), fertilization, and blastocystformation when tyrosine phosphorylation levels of the ∼55,∼23, ∼22, ∼21, and ∼18 kDa proteins decrease.15 Given theaforementioned studies, it is tempting to hypothesize thattyrosine phosphorylation is a mechanism by which motility andhyperactivation in spermatozoa can be controlled withsubsequent fertility. However, further studies are required todetermine the exact mechanism through which particularproteins regulate protein tyrosine phosphorylation as well asspermatozoa motility, hyperactivation, and subsequent fertilityand early embryonic development. Therefore, understandingthe molecular basis of these processes requires characterizationof the phosphoproteins involved in the relevant signaltransduction pathways. In addition, the large number ofreported tyrosine-phosphorylated proteins should be morethoroughly characterized. We have generated a schematicrepresentation of the various signaling pathways involved intyrosine phosphorylation and relationship of tyrosine phos-phorylation to other protein functions. These proteins exhibitsignificant modifications that induce sperm hyperactivation andchemotaxis via the regulation of tyrosine phosphorylation(Table 1, Figure 2).
3.2. Capacitation and Acrosome Reaction
Mature spermatozoa cannot fertilize oocytes even if they aremotile and physiologically normal.87 In internally fertilizinganimals, mature spermatozoa achieve fertilization competenceonly after they enter the female genital tract;38 the alterations of
spermatozoa once inside the tract is known as capacitation.36,37
Only capacitated spermatozoa can undergo the acrosomereaction upon binding to the zona pellucida, at which pointthey become capable of penetrating and fertilizing an egg.13−15
Capacitation confers a series of metabolic and structuralmodifications to the spermatozoa that are accompanied bychanges in membrane fluidity; HCO−3, Ca2+, and cAMP levels;PKA activity; and tyrosine phosphorylation of pro-teins.14,15,18,88−91 Studies have demonstrated that tyrosinephosphorylation is upregulated during capacitation.92−94
Other studies have consistently reported a positive correlationbetween capacitation and protein tyrosine phosphorylation invarious mammalian species, including humans, that is correlatedwith fertilization and male fertility.18,40,42,95−101 The relation-ship between capacitation and tyrosine phosphorylation wasdiscovered before the beginning of the 21st century; however,the basic molecular mechanism underlying tyrosine phosphor-ylation-mediated control of capacitation, the acrosome reaction,and male fertility is still unclear. Our review of the literatureshowed that four major signaling pathways modulating tyrosinephosphorylation-guided capacitation and the acrosome reactionhave been reported, namely the cAMP/PKA-dependentpathway,8−12,102−104 receptor tyrosine kinase pathway,105,106
nonreceptor protein tyrosine kinase pathway,17 and G-proteincoupled receptor pathway.105−107 The cAMP/PKA-dependentpathway is the most extensively studied and is more specific tosperm cells; therefore, we discuss in detail the relationship ofthis pathway with tyrosine phosphorylation, male fertilization,
Figure 2. Interactions related to tyrosine phosphorylation-regulated motility and hyperactivation that ultimately affect male infertility in mammalianspermatozoa. Abbreviations: ADCY10, adenylate cyclase 10; AKAP, A-kinase anchor proteins; AKAP3, A-kinase (PRKA) anchor protein 3; AKAP4,A-kinase (PRKA) anchor protein 4; ATP, adenosine triphosphate; AVP, arginine vasopressin; AVPR2, arginine vasopressin receptor 2; CABYR,calcium binding tyrosine-(Y)-phosphorylation regulated; cAMP, cyclic adenosine monophosphate; CATSPER2c cation channel, sperm associated 2;CD55, CD55 molecule; decay accelerating factor for complement; CNR1, cannabinoid receptor 1; CRISP1, cysteine-rich secretory protein 1; IP3,inositol trisphosphate; ODF2, outer dense fiber of sperm tails 2; PI3K, phosphatidylinositol 3-kinase; PKA, cAMP-dependent protein kinase, proteinkinase A; PKC, protein kinase C; PLC, phospholipase C; PP1, protein phosphatase 1; PRKAR2B, protein kinase, cAMP-dependent, regulatory, typeII, beta; PRKCA, protein kinase C, alpha; ROPN1, rhophilin associated tail protein 1; ROPN1L, rhophilin associated tail protein 1-like; ROS,reactive oxygen species; SLC9A10, solute carrier family 9, member 10; SRC, v-src sarcoma (Schmidt-Ruppin A-2) viral oncogene homologue(avian); ZP3, zona pellucida glycoprotein 3 (sperm receptor).
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174509

and embryonic development (depicted hypothetically in Figure3).17
Previous studies have reported that tyrosine phosphoryla-tion-dependent acquisition of capacitation and the acrosomereaction in mammalian spermatozoa are achieved through theregulation of a pathway mediated by cAMP (an intracellularsecond messenger) in which PKA plays a fundamentalrole.18,40,41,97,99,108−111 Adenosine triphosphate activates mem-brane bound soluble adenylyl cyclase (sAC) to generate cAMP,and the cAMP in spermatozoa activates PKA, which regulatesprotein tyrosine phosphorylation.13−15,18,40,41 Most previousstudies of cAMP have focused on its role in the regulation ofsperm motility; however, its roles in capacitation and theacrosome reaction remain unknown.38,111 Alternatively, PKAmight stimulate capacitation and the acrosome reaction ofspermatozoa through serine/threonine phosphorylation.112 Theoptimal level at which cAMP/PKA regulates steady-state levelsof tyrosine phosphorylation in sperm under conditionsconducive to capacitation, the acrosome reaction, andfertilization will likely be a subject of intense study in thenear future.Using the mouse as an experimental model, we and other
researchers have demonstrated that several factors includingCa2+,113,114 HCO3
−,115,116 BSA,117 heparin,118,119 glucose,reactive oxygen species (ROS),120,121 intracellular pH(pHi),
110,114 and upregulation or blockage of de novo proteinfunctions regulate tyrosine phosphorylation-dependent capaci-tation and the acrosome reaction in spermatozoa.13−15
Therefore, determination of the appropriate concentrations ofsuch factors is critical when inducing sperm capacitation in vitro
with chemically defined media. Furthermore, low concen-trations of nitric oxide (NO)-releasing compounds inducecapacitation in human115 and cryopreserved bull spermato-zoa.116 Interestingly, NO synthase inhibitors meaningfullydecrease this process.117,118 Jagan Mohanarao and Atreja119
hypothesized that NO-triggered capacitation in buffalospermatozoa is associated with an increase in protein tyrosinephosphorylation. The authors identified a group of proteinsthat undergo tyrosine phosphorylation in response to NO-mediated signaling cascades during capacitation.119 Simulta-neously, previous studies have shown that Ca2+ and HCO3
−
have a similar effect, as both ions trigger the activity ofmammalian sperm cAMP and subsequently induce capacitationfollowing increased levels of tyrosine phosphorylation.113,120,121
However, the exact mechanism through which these ionsstimulate tyrosine phosphorylation remains to be elucidated.Furthermore, the addition of BSA together with Ca2+ andHCO3
− to the medium reportedly stimulates protein tyrosinephosphorylation in spermatozoa before capacitation.122 There-fore, it is tempting to hypothesize that the absence of any ofthese components from the medium might prevent proteintyrosine phosphorylation and capacitation. BSA regulatescapacitation by removing cholesterol from the sperm plasmamembrane.114 Ions that have similar functions in spermatozoaremain to be elucidated. Therefore, cholesterol release as asignaling event might be highly correlated with tyrosinephosphorylation. This unique manner of signal transductiondefinitely deserves further investigation.Recently, several researcher have demonstrated that free
radicals (especially ROS) can affect male fertility via lipid
Figure 3. Possible mechanism underlying hyperpolarization, capacitation, and the acrosome reaction in mammalian spermatozoa. Protein kinase-A(PKA)-stimulated tyrosine phosphorylation induces capacitation and the acrosome reaction. PKA might also regulate the phosphorylation of spermproteins at Ser and Thr residues to regulate these sperm functions. The main hypothesis has been modified according to Visconti and Kopf.114
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174510

peroxidation and DNA damage.123,124 However, no reports areavailable on the mechanism through which free radicals affectcapacitation and the acrosome reaction. Leclerc et al.125 havefound that ROS upregulate the tyrosine phosphorylation levelsof several proteins. A similar effect of ROS has been describedby Aitken et al.126 Therefore, it seems reasonable tohypothesize that ROS affect capacitation, the acrosomereaction, and subsequent fertilization. However, the effect ofROS on cAMP or other capacitation-related signaling remainsto be determined. In addition, pHi is thought to play a criticalrole in capacitation and the acrosome reaction. Studies havesuggested that changes in pHi mainly affect capacitation via thePKA-dependent pathway110 and thus modulate protein tyrosinephosphorylation.110,127
In vitro capacitation can be achieved by incubatingspermatozoa in medium containing heparin or oviductalfluid.128,129 Heparin affects tyrosine phosphorylation byupregulating cAMP synthesis, thus affecting sperm capacita-tion.129 By contrast, glucose has the opposite effect oncapacitation. In bovine sperm, glucose inhibits heparin-inducedcapacitation in vitro through a mechanism involving cAMPmetabolism and the reduction of pHi.
129 However, a similareffect has not been demonstrated in an in vivo trial.Furthermore, these cascades may not be mutually exclusive,and may include cross-talk among several molecules. Many keymolecules and receptors must still be identified to elucidatecompletely the molecular mechanism and signal transductioncascade involved in capacitation. A G-protein-coupled receptorpathway has not been included in this model.
Recent studies have established that manipulating thefunction of a sperm protein can regulate capacitation and theacrosome reaction and ultimately affect male fertility. Ourprevious studies have demonstrated that blocking the functionof UQCRC2 with nutlin-3a, a small molecule antagonist ofMDM2, lowers male fertility by reducing capacitation rate andthe acrosome reaction through downregulation of tyrosinephosphorylation (100 and 60 kDa).13 In another study, wereported that blocking VDACs with the specific inhibitor 4,4′-diisothiocyanostilbene-2,2′-disulfonic acid (DIDS) could con-trol capacitation, the acrosome reaction, and fertility by alteringtyrosine phosphorylation in ICR mice.14 Interestingly, Park etal.51 reported that VDAC2 and UQCRC2 are more highlyexpressed in low-fertility (<70% Nonreturn rate) bulls. Becausehigher expression levels of VDAC2 and UQCRC2 were foundin the lower-fertility genotype, the blocking of these proteinsshould improve fertility. Possible explanations for thediscrepancies in results are that a particular protein couldaffect capacitation, the acrosome reaction, and fertilizationwhen expressed at a certain level. Alternatively, increasedexpression levels of VDAC2 and UQCRC2 in less fertile males(bulls) could be a compensation for lowered fertility. Similarly,another study has proposed that exposure of mice spermatozoato the specific AVPR2 agonist dDAVP decreases the PKAsubstrate level (23 kDa) and tyrosine phosphorylation level (30kDa) dose dependently, subsequently affecting capacitation, theacrosome reaction, and embryo development.15 Therefore,further studies should attempt to evaluate the effect of severalsperm proteins on fertilization and their association withprotein tyrosine phosphorylation. The application of Pathway
Figure 4. Interactions related to tyrosine phosphorylation-regulated capacitation and the acrosome reaction that ultimately affect male infertility inmammalian spermatozoa. Abbreviations: ADCY10, adenylate cyclase 10; ATP, adenosine triphosphate; AVP, arginine vasopressin; AVPR2, argininevasopressin receptor 2; cAMP, cyclic adenosine monophosphate; CFTR, cystic fibrosis transmembrane conductance regulator (ATP-binding cassettesubfamily C, member 7); CYCS, cytochrome c, somatic; DMBT1, deleted in malignant brain tumors 1; EGF, epidermal growth factor; EGFR,epidermal growth factor receptor; GPD2, glycerol-3-phosphate dehydrogenase 2 (mitochondrial); GSN, Gelsolin; GSTM3, glutathione S-transferasemu 3; HDL, high-density lipoprotein; IP3, inositol trisphosphate; IZUMO1, izumo sperm-egg fusion 1; LY6G6C, lymphocyte antigen 6 complex,locus G6C; MAPK3, mitogen-activated protein kinase 3; MIF, macrophage migration inhibitory factor (glycosylation-inhibiting factor); PDHA1,pyruvate dehydrogenase (lipoamide) alpha 1; PKA, cAMP-dependent protein kinase, protein kinase A; PKC, protein kinase C; Spinkl, serineprotease inhibitor; Kazal type-like; SRC, v-src sarcoma (Schmidt-Ruppin A-2) viral oncogene homologue (avian); TPA, tetradecanoylphorbolacetate; ZP3, zona pellucida glycoprotein 3 (sperm receptor).
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174511

Studio helped us analyze the proteins implicated in tyrosinephosphorylation-mediated regulation of capacitation, theacrosome reaction, and male fertility (Table 1, Figure 4).
3.3. Sperm-Zona Binding and Postfertilization EggActivation
Tyrosine phosphorylation has been recognized as an importantphenomenon correlated with various maturational processes inspermatozoa (described earlier). Several studies have alsoreported a direct association among tyrosine phosphorylation,sperm−zona-binding capability, and postfertilization eggactivation. It has been reported that zonadhesin, a spermmembrane protein, binds postacrosomally to the zonapellucida, acting as a primary gamete-recognition compo-nent.130 An elegant study conducted by Herlyn and Zischler131
reported that, together with several conformational changes,phosphorylation motifs occur in zonadhesin (D domains) andare primarily responsible for its function in various animalspecies. Accordingly, tyrosine phosphorylation is the mostcommon post-translational modification/phosphorylation inmammalian spermatozoa.17,42 Therefore, it could be hypothe-sized that tyrosine phosphorylation has a critical role in thespecies-specific binding of zonadhesin to the zona pellucida.Naz et al.52 have reported that the incubation of humanspermatozoa with an antiphosphotyrosine antibody significantlyinhibits sperm binding and penetration of zona-free hamster
eggs. In another study, Naz and Ahmad53 demonstrated thattyrosine phosphorylation occurs in three human sperm proteins(95, 51, and 14−18 kDa) capable of binding zona pellucidaproteins and stimulating sperm-zona binding. Therefore, itseems reasonable to hypothesize that tyrosine phosphorylationof sperm proteins plays a dynamic role in sperm−zonainteraction and binding.An excellent study by Urner et al.54 showed that the level of
tyrosine phosphorylation increases during zona binding andgamete fusion in mice spermatozoa. Furthermore, the authorsshowed that, following binding to the oocyte zona pellucida andoolemma, tyrosine phosphorylation in the sperm flagellum isdelayed in the absence of glucose (described earlier).Therefore, this study provides a direct link between proteintyrosine phosphorylation and gamete interaction. In addition,Nassar et al.55 have reported that exposure of humanspermatozoa to pentoxifylline increases protein phosphoryla-tion levels in sperm tail and the capability of sperm to bind tothe zona pellucida. However, the molecular mechanismunderlying protein phosphorylation/tyrosine phosphorylation-stimulated binding remains to be elucidated and might be anexcellent target for research. Progesterone is well documentedto trigger the rapid influx of Ca2+ and acrosomal exocytosis inthe spermatozoa of various animal species. Tesarik et al.132 havereported a selective increase in the tyrosine phosphorylationlevels of a 94 kDa phosphoprotein in the presence of
Figure 5. Interactions related to tyrosine phosphorylation-regulated sperm−zona interaction and egg activation that ultimately affect male fertility inmammalian spermatozoa. Abbreviations: ADCY10, adenylate cyclase 10; AGT, angiotensinogen (serpin peptidase inhibitor, clade A, member 8);ALB, albumin; Amp1, amplitude of circadian rhythm 1; ATP, adenosine triphosphate; AVP, arginine vasopressin; AVPR2, arginine vasopressinreceptor 2; cAMP, cyclic adenosine monophosphate; CCNB2, cyclin B2; CRISP1, cysteine-rich secretory protein 1; CTSB, cathepsin B; EGF,epidermal growth factor; FYN, FYN oncogene related to SRC, FGR, YES; GSTM3, glutathione S-transferase mu 3; IL6, interleukin 6 (interferon,beta 2); IP3, inositol trisphosphate; MAPK1, mitogen-activated protein kinase 1; PECAM1, platelet/endothelial cell adhesion molecule; PI3K,phosphatidylinositol 3-kinase; PKA, cAMP-dependent protein kinase, protein kinase A; PLC, phospholipase C; PP1, protein phosphatase 1; ROS,reactive oxygen species; SHC1, SHC (Src homology 2 domain containing) transforming protein 1; SRC, v-src sarcoma (Schmidt-Ruppin A-2) viraloncogene homologue (avian); TPA, tetradecanoylphorbol acetate; VDAC, voltage-dependent anion channel; ZP3, zona pellucida glycoprotein 3(sperm receptor).
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174512

progesterone. Because progesterone induces spermatozoabinding to and penetration of the oocyte plasma membrane,133
it is reasonable to hypothesize that tyrosine phosphorylation isthe signal that regulates progesterone-mediated spermactivation and fertilization. However, the molecular basis ofthis relationship remains to be determined. A similar affiliationbetween tyrosine phosphorylation and zona binding wasreported in a recent study by Kadirvel et al.134 These authorsshowed a strong correlation between the protein tyrosinephosphorylation pattern and zona-binding capability in vitro incapacitated (heparin induced) and frozen−thawed (cryocapa-citated) buffalo spermatozoa. In addition, they observed adifference in zona-binding capability between frozen−thawedand in vitro capacitated spermatozoa; however, an explanationwas not provided. By contrast, studies in Xenopus laevis (clawedfrog) eggs by Sato and colleagues135 have demonstrated proteintyrosine phosphorylation and dephosphorylation pathways insperm-induced egg activation. However, similar studies inmammalian spermatozoa have not yet been reported. Wegenerated an illustration of the sperm protein signalingpathways to facilitate understanding of the mechanismsunderlying tyrosine phosphorylation, egg activation, andsperm−zona binding (Table 1, Figure 5).
4. SUMMARY AND FUTURE PROSPECTSConventional semen analysis has several drawbacks in thediagnosis and prognosis of male infertility, subfertility, or both.Therefore, the development of new approaches is extraordi-narily important. In this regard, sperm functions and themechanism of male fertility must be fully understood. Thetyrosine phosphorylation of sperm proteins is a centralregulator of many key activities in spermatozoa, all of whichare required for fertilization. Despite previous research efforts,several questions remain to be addressed for a more completeview of tyrosine phosphorylation-regulated sperm functions andtheir roles in male fertility. Our literature review yielded manyreports that tyrosine phosphorylation in spermatozoa isregulated by several factors; however, the underlyingmechanism is less clearly defined. Moreover, future studiesshould focus on the detection and isolation of other factors thatmay affect tyrosine phosphorylation as well as male (in)fertility.Manipulation of sperm protein function using special agonistsor antagonists may regulate tyrosine phosphorylation inspermatozoa.5−7 Although recently applied proteomic ap-proaches have identified several sperm-specific proteins, theirfunctions in tyrosine phosphorylation and their association withfertility are unclear. Therefore, future studies should attempt toprovide a robust understanding of tyrosine phosphorylationand male infertility. Such findings will assist in the developmentof more accurate tools for the diagnosis and prognosis of maleinfertility.
■ AUTHOR INFORMATIONCorresponding Author
*Telephone: +82.31.670.4841. Fax: +82.31.675.9001. E-mail:[email protected]
The authors declare no competing financial interest.
■ ACKNOWLEDGMENTSThis study was supported by the Cooperative ResearchProgram for Agricultural Science and Technology Develop-
ment (Project No. PJ008415) of the Rural DevelopmentAdministration, Republic of Korea.
■ REFERENCES(1) Sharlip, I. D.; Jarow, J. P.; Belker, A. M.; Lipshultz, L. I.; Sigman,M.; Thomas, A. J.; Schlegel, P. N.; Howards, S. S.; Nehra, A.;Damewood, M. D.; Overstreet, J. W.; Sadovsky, R. Best practicepolicies for male infertility. Fertil. Steril. 2002, 77, 873−882.(2) World Health Organization. WHO Laboratory Manual for theExamination of Human Semen and sperm-cervical mucus interaction, 4thed.; Cambridge University Press: Cambridge, 1999; pp 128.(3) Dowsett, K. F.; Pattie, W. A. Characteristics and fertility ofstallion semen. J. Reprod. Fertil. Suppl. 1982, 32, 1−8.(4) Kenney, R. M.; Kingston, R. S.; Rajamannon, A. H.; Ramberg, C.F. Stallion semen characteristics for predicting fertility. Proc. Ann. Mt.Am. Assoc. Equine Pract. 1971, 53−67.(5) Jasko, D. J.; Lein, D. H.; Foote, R. H. Determination of therelationship between sperm morphologic classifications and fertility installions: 66 cases. J. Am. Vet. Med. Assoc. 1990, 197, 389.(6) Morrell, J. M.; Johannisson, A.; Dalin, A. M.; Hammar, L.;Sandebert, T.; Rodriguez-Martinezet, H. Sperm morphology andchromatin integrity in Swedish warmblood stallions and theirrelationship to pregnancy rates. Acta Vet. Scand. 2008, 50, 1−7.(7) Kastelic, J. P.; Thundathil, J. C. Breeding soundness evaluationand semen analysis for predicting bull fertility. Reprod. Domest. Anim.2008, 43, 368−373.(8) Lee, M. S.; Rahman, M. S.; Kwon, W. S.; Chung, H. J.; Yang, B.S.; Pang, M. G. Efficacy of four synchronization protocols on the estrusbehavior and conception in native Korean cattle (Hanwoo).Theriogenology 2013, 80, 855−861.(9) Gadea, J. Sperm factors related to in vitro and in vivo porcinefertility. Theriogenology 2005, 63, 431−444.(10) Sanchez-Partida, L. G.; Windsor, D. P.; Eppleston, J.; Setchell, B.P.; Maxwell, W. M. Fertility and its relationship to motilitycharacteristics of spermatozoa in ewes after cervical, transcervical,and intrauterine insemination with frozen-thawed ram semen. J.Androl. 1999, 20, 280−288.(11) Pizzol, D.; Ferlin, A.; Garolla, A.; Lenzi, A.; Bertoldo, A.;Foresta, C. Genetic and molecular diagnostics of male infertility in theclinical practice. Front. Biosci. 2014, 19, 291−303.(12) Kumar, R.; Singh, V. K.; Atreja, S. K. Glutathione-S-transferase:Role in buffalo (Bubalus bubalis) sperm capacitation and cryopre-servation. Theriogenology 2014, 81, 587−598.(13) Shukla, K. K.; Kwon, W. S.; Rahman, M. S.; Park, Y. J.; You, Y.A.; Pang, M. G. Nutlin-3a decreases male fertility via UQCRC2. PLoSOne 2013, 8, e76959.(14) Kwon, W. S.; Park, Y. J.; Mohamed, el-S. A.; Pang, M. G.Voltage-dependent anion channels are a key factor of male fertility.Fertil. Steril. 2013, 99, 354−361.(15) Kwon, W. S.; Park, Y. J.; Kim, Y. H.; You, Y. A.; Kim, I. C.; Pang,M. G. Vasopressin effectively suppresses male fertility. PLoS One 2013,8, e54192.(16) Kota, V.; Dhople, V. M.; Shivaji, S. Tyrosine phosphoproteomeof hamster spermatozoa: role of glycerol-3-phosphate dehydrogenase 2in sperm capacitation. Proteomics 2009, 9, 1809−1826.(17) Naz, R. K.; Rajesh, P. B. Role of tyrosine phosphorylation insperm capacitation/acrosome reaction. Reprod. Biol. Endocrinol. 2004,2, 75.(18) Visconti, P. E.; Stewart-Savage, J.; Blasco, A.; Battaglia, L.;Miranda, P.; Kopf, G. S.; Tezon, J. G. Roles of bicarbonate, cAMP, andprotein tyrosine phosphorylation on capacitation and the spontaneousacrosome reaction of hamster sperm. Biol. Reprod. 1999, 61, 76−84.(19) Baldi, E.; Luconi, M.; Bonaccorsi, L.; Krausz, C.; Forti, G.Human sperm activation during capacitation and acrosome reaction:role of calcium, proteinphosphorylation and lipid remodellingpathways. Front. Biosci. 1996, 15, 189−205.(20) Goodla, L.; Morrell, J. M.; Yusnizar, Y.; Stalhammar, H.;Johannisson, A. Quality of bull spermatozoa after preparation bysingle-layer centrifugation. J. Dairy. Sci. 2014, 97, 2204−2212.
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174513

(21) Boerke, A.; van der Lit, J.; Lolicato, F.; Stout, T. A.; Helms, J. B.;Gadella, B. M. Removal of GPI-anchored membrane proteins causesclustering of lipid microdomains in the apical head area of porcinesperm. Theriogenology 2014, 81, 613−624.(22) Sebkova, N.; Ded, L.; Vesela, K.; Dvorakova-Hortova, K.Progress of sperm IZUMO1 relocation during spontaneous acrosomereaction. Reproduction 2013, 147, 231−240.(23) Rahman, M. S.; Kwon, W. S.; Pang, M. G. Calcium influx andmale fertility in the context of the sperm proteome: an update. Biomed.Res. Int. 2014, 841615.(24) De Lisa, E.; Salzano, A. M.; Moccia, F.; Scaloni, A.; Di Cosmo,A. Sperm-attractant peptide influences the spermatozoa swimmingbehavior in internal fertilization in Octopus vulgaris. J. Exp. Biol. 2013,15, 2229−2237.(25) Baldi, E.; Luconi, M.; Muratori, M.; Marchiani, S.; Tamburrino,L.; Forti, G. Nongenomic activation of spermatozoa by steroidhormones: facts and fictions. Mol. Cell. Endocrinol. 2009, 308, 39−46.(26) Mannowetz, N.; Naidoo, N. M.; Choo, S. A.; Smith, J. F.;Lishko, P. V. Slo1 is the principal potassium channel of humanspermatozoa. Elife 2013, 2, e01009.(27) Rahman, M. S.; Lee, J. S.; Kwon, W. S.; Pang, M. G. Spermproteomics: road to male fertility and contraception. Int. J. Endocrinol.2013, 2013, 360986.(28) Lishko, P. V.; Kirichok, Y. The role of Hv1 and CatSperchannels in sperm activation. J. Physiol. 2010, 588, 4667−4672.(29) Naz, R. K. Involvement of protein tyrosine phosphorylation ofhuman sperm in capacitation/acrosome reaction and zona pellucidabinding. Front. Biosci. 1996, 1, 206−213.(30) Revelli, A.; Ghigo, D.; Moffa, F.; Massobrio, M.; Tur-Kaspa, I.Guanylate cyclase activity and sperm function. Endocr. Rev. 2002, 23,484−494.(31) Visconti, P. E.; Westbrook, V. A.; Chertihin, O.; Demarco, I.;Sleight, S.; Diekman, A. B. Novel signaling pathways involved in spermacquisition of fertilizing capacity. J. Reprod. Immunol. 2002, 53, 133−150.(32) Vadnais, M. L.; Galantino-Homer, H. L.; Althouse, G. C.Current concepts of molecular events during bovine and porcinespermatozoa capacitation. Arch. Androl. 2007, 53, 109−123.(33) Bohmer, M.; Van, Q.; Weyand, I.; Hagen, V.; Beyermann, M.;Matsumoto, M.; Hoshi, M.; Hildebrand, E.; Kaupp, U. B. Ca2+ spikesin the flagellum control chemotactic behavior of sperm. EMBO J.2005, 24, 2741−2752.(34) Ho, H. C.; Suarez, S. S. Hyperactivation of mammalianspermatozoa: function and regulation. Reproduction 2001, 122, 519−526.(35) Suarez, S. S.; Dai, X. Hyperactivation enhances mouse spermcapacity for penetrating viscoelastic media. Biol. Reprod. 1992, 46,686−491.(36) Austin, C. R. Observations on the penetration of the sperm inthe mammalian egg. Aust. J. Sci. Res. B 1951, 4, 581−596.(37) Chang, M. C. Fertilizing capacity of spermatozoa deposited intothe fallopian tubes. Nature 1951, 168, 697−698.(38) Yanagimachi, R. Mammalian fertilization: The physiology ofreproduction; Knobil, E., Neill, J. D., Eds.; Raven Press Ltd.: New York,1994; pp 189−317.(39) Si, Y.; Okuno, M. Role of tyrosine phosphorylation of flagellarproteins in hamster sperm hyperactivation. Biol. Reprod. 1999, 61,240−246.(40) Visconti, P. E.; Bailey, J. L.; Moore, G. D.; Pan, D.; Olds-Clarke,P.; Kopf, G. S. Capacitation of mouse spermatozoa. I. Correlationbetween the capacitation state and protein tyrosine phosphorylation.Development 1995, 121, 1129−1137.(41) Visconti, P. E.; Moore, G. D.; Bailey, J. L.; Leclerc, P.; Connors,S. A.; Pan, D.; Olds-Clarke, P.; Kopf, G. S. Capacitation of mousespermatozoa. II. Protein tyrosine phosphorylation and capacitation areregulated by a cAMP-dependent pathway. Development 1995, 121,1139−1150.
(42) Naz, R. K.; Rajesh, P. B. Role of tyrosine phosphorylation insperm capacitation/acrosome reaction. Reprod. Biol. Endocrinol. 2004,2, 75.(43) Baker, M. A.; Smith, N. D.; Hetherington, L.; Taubman, K.;Graham, M. E.; Robinson, P. J.; Aitken, R. J. Label-free quantitation ofphosphopeptide changes during rat sperm capacitation. J. Proteome Res.2010, 9, 718−729.(44) Schumacher, J.; Ramljak, S.; Asif, A. R.; Schaffrath, M.; Zischler,H.; Herlyn, H. Evolutionary conservation of mammalian spermproteins associates with overall, not tyrosine, phosphorylation inhuman spermatozoa. J. Proteome Res. 2013, 12, 5370−5382.(45) Lee, J. S.; Park, Y. J.; Kim, J.; Rahman, M. S.; Kwon, W. S.;Yoon, S. J.; You, Y. A.; Pang, M. G. Effect of Arp2/3 complex onsperm motility and membrane structure in bovine. Reprod. Dev. Biol.2013, 37, 169−174.(46) Hunter, T. Signaling2000 and beyond. Cell 2000, 100, 113−127.(47) Pawson, T. Specificity in signal transduction: from phosphotyr-osine-SH2 domain interactions to complex cellular systems. Cell 2004,16, 191−203.(48) Johnson, L. N. The regulation of protein phosphorylation.Biochem. Soc. Trans. 2009, 37, 627−641.(49) Baker, M. A.; Nixon, B.; Naumovski, N.; Aitken, R. J. Proteomicinsights into the maturation and capacitation of mammalianspermatozoa. Syst. Biol. Reprod. Med. 2012, 58, 211−217.(50) Gur, Y.; Breitbart, H. Mammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomes. Genes Dev. 2006,20, 411−416.(51) Park, Y. J.; Kwon, W. S.; Oh, S. A.; Pang, M. G. Fertility-relatedproteomic profiling bull spermatozoa separated by Percoll. J. Proteome.Res. 2012, 11, 4162−4168.(52) Naz, R. K.; Ahmad, K.; Kumar, R. Role of membranephosphotyrosine proteins in human spermatozoal function. J. CellSci. 1991, 99, 157−165.(53) Naz, R. K.; Ahmad, K. Molecular identities of human spermproteins that bind human zona pellucida: nature of sperm-zonainteraction, tyrosine kinase activity, and involvement of FA-1. Mol.Reprod. Dev. 1994, 39, 397−408.(54) Urner, F.; Leppens-Luisier, G.; Sakkas, D. Protein tyrosinephosphorylation in sperm during gamete interaction in the mouse: theinfluence of glucose. Biol. Reprod. 2001, 64, 1350−1357.(55) Nassar, A.; Mahony, M.; Morshedi, M.; Lin, M. H.; Srisombut,C.; Oehninger, S. Modulation of sperm tail protein tyrosinephosphorylation by pentoxifylline and its correlation with hyper-activated motility. Fertil. Steril. 1999, 71, 919−923.(56) Ficarro, S.; Chertihin, O.; Westbrook, V. A.; White, F.; Jayes, F.;Kalab, P.; Marto, J. A.; Shabanowitz, J.; Herr, J. C.; Hunt, D. F.;Visconti, P. E. Phosphoproteome analysis of capacitated human sperm.Evidence of tyrosine phosphorylation of a kinase-anchoring protein 3and valosin-containing protein/p97 during capacitation. J. Biol. Chem.2003, 278, 11579−11589.(57) Paradisi, R.; Capelli, M.; Mandini, M.; Bellavia, E.; Focacci, M.;Flamigni, C. Interleukin-2 in seminal plasma of fertile and infertilemen. Arch. Androl. 1995, 35, 35−41.(58) Rajasekaran, M.; Hellstrom, W. J.; Naz, R. K.; Sikka, S. C.Oxidative stress and interleukins in seminal plasma duringleukocytospermia. Fertil. Steril. 1995, 64, 166−171.(59) Naz, R. K.; Kaplan, P. Interleukin-6 enhances the fertilizingcapacity of human sperm by increasing capacitation and acrosomereaction. J. Androl. 1994, 15, 228−233.(60) Shimoya, K.; Matsuzaki, N.; Tsutsui, T.; Taniguchi, T.; Saji, F.;Tanizawa, O. Detection of interleukin-8 (IL-8) in seminal plasma andelevated IL-8 in seminal plasma of infertile patients with leukospermia.Fertil. Steril. 1993, 59, 885−888.(61) Naz, R. K.; Stanley, E. R. Enhanced levels of colony stimulatingfactor-1 (CSF-1) in sera and seminal plasma of antisperm antibody-positive infertile men. Arch. Androl. 1995, 35, 5−11.(62) Naz, R. K.; Kaplan, P. Increased levels of interleukin-6 inseminal plasma of infertile men. J. Androl. 1994, 15, 220−227.
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174514

(63) Hussenet, F.; Dousset, B.; Cordonnier, J. L.; Jacob, C.; Foliguet,B.; Grignon, G.; Nabet, P. Tumour necrosis factor alpha andinterleukin 2 in normal and infected human seminal fluid. Hum.Reprod. 1993, 8, 409−411.(64) Naz, R. K.; Kaplan, P. Effects of epidermal growth factor onhuman sperm cell function. J. Androl. 1993, 14, 240−247.(65) Naz, R. K.; Kaplan, P.; Goldstein, A. L. Thymosin alpha-1enhances the fertilizing capacity of human sperm cell: implication indiagnosis and treatment of male infertility. Biol. Reprod. 1992, 47,1064−1072.(66) Laursen, I.; Briand, P.; Lykkesfeldt, A. E. Serum albumin as amodulator on growth of the human breast cancer cell line, MCF-7.Anticancer Res. 1990, 10, 343−351.(67) Naz, R. K. Involvement of protein tyrosine phosphorylation ofhuman sperm in capacitation/acrosome reaction and zona pellucidabinding. Front. Biosci. 1996, 1, 206−213.(68) Suarez, S. S. Control of hyperactivation in sperm. Hum. Reprod.Update 2008, 14, 647−657.(69) Suarez, S. S. Hyperactivated motility in sperm. J. Androl. 1996,17, 331−335.(70) Olds-Clarke, P. Variation in the quality of sperm motility and itsrelationship to capacitation: Fertilization in Mammals; Bavister, B. D.,Cummins, J., Roldan, E. R. S., Eds.; Serono Symposia: Norwell, MA,1990; pp 91−99.(71) Yanagimachi, R. Mechanisms of fertilization in mammals:Fertilization and embryonic development in vitro; Mastroianni, L.,Biggers, J. D., Eds.; Plenum Press: New York, 1981; pp 81−182.(72) Yanagimachi, R. The movement of golden hamster spermatozoabefore and after capacitation. J. Reprod. Fertil. 1970, 23, 193−196.(73) Mahony, M. C.; Gwathmey, T. Protein tyrosine phosphorylationduring hyperactivated motility of cynomolgus monkey (Macacafascicularis) spermatozoa. Biol. Reprod. 1999, 60, 1239−1243.(74) Vijayaraghavan, S.; Trautman, K. D.; Goueli, S. A.; Carr, D. W.A tyrosine-phosphorylated 55-kilodalton motility-associated bovinesperm protein is regulated by cyclic adenosine 3′,5′-monophosphatesand calcium. Biol. Reprod. 1997, 56, 1450−1457.(75) Oliva, R. Protamines and male infertility. Hum. Reprod. Update2006, 12, 417−35.(76) Luke, L.; Campbell, P.; Varea Sanchez, M.; Nachman, M. W.;Roldan, E. R. Sexual selection on protamine and transition nuclearprotein expression in mouse species. Proc. Biol. Sci. 2014, 281,20133359.(77) Anderson, M. J.; Dixson, A. F. Sperm competition: motility andthe midpiece in primates. Nature 2002, 416, 496.(78) Aleem, M.; Padwal, V.; Choudhari, J.; Balasinor, N.; Gill-Sharma, M. K. Sperm protamine levels as indicators of fertilisingpotential in sexually mature male rats. Andrologia 2008, 40, 29−37.(79) Ickowicz, D.; Finkelstein, M.; Breitbart, H. Mechanism of spermcapacitation and the acrosome reaction: role of protein kinases. Asian J.Androl. 2012, 14, 816−821.(80) Marquez, B.; Suarez, S. S. Different signaling pathways in bovinesperm regulate capacitation and hyperactivation. Biol. Reprod. 2004,70, 1626−1633.(81) Ho, H. C.; Granish, K. A.; Suarez, S. S. Hyperactivated motilityof bull sperm is triggered at the axoneme by Ca2+ and not cAMP. Dev.Biol. 2002, 250, 208−217.(82) Ho, H. C.; Suarez, S. S. Hyperactivation of mammalianspermatozoa: function and regulation. Reproduction 2001, 122, 519−526.(83) Tash, J. S.; Hidaka, H.; Means, A. R. Axokinin phosphorylationby cAMP dependent protein kinase is sufficient for activation of spermflagellar motility. J. Cell Biol. 1986, 103, 649−655.(84) Miki, K.; Willis, W. D.; Brown, P. R.; Goulding, E. H.; Fulcher,K. D.; Eddy, E. M. Targeted disruption of the Akap4 gene causesdefects in sperm flagellum and motility. Dev. Biol. 2002, 248, 331−342.(85) Carrera, A.; Moos, J.; Ning, X. P.; Gerton, G. L.; Tesarik, J.;Kopf, G. S.; Moss, S. B. Regulation of protein tyrosine phosphorylationin human sperm by a calcium/calmodulin-dependent mechanism:
identification of A kinase anchor proteins as major substrates fortyrosine phosphorylation. Dev. Biol. 1996, 180, 284−296.(86) Mandal, A.; Naaby-Hansen, S.; Wolkowicz, M. J.; Klotz, K.;Shetty, J.; Retief, J. D.; Coonrod, S. A.; Kinter, M.; Sherman, N.; Cesar,F.; Flickinger, C. J.; Herr, J. C. FSP95, a testis-specific 95-kilodaltonfibrous sheath antigen that undergoes tyrosine phosphorylation incapacitated human spermatozoa. Biol. Reprod. 1999, 61, 1184−1197.(87) Yanagimachi, R. Intracytoplasmic sperm injection experimentsusing the mouse as a model. Hum. Reprod. 1998, 13, 87−98.(88) Fraser, L. R.; McDermott, C. A. Ca(2+)-related changes in themouse sperm capacitation state: a possible role for Ca(2+)-ATPase. J.Reprod. Fertil. 1992, 96, 363−377.(89) Luconi, M.; Krausz, C.; Forti, G.; Baldi, E. Extracellular calciumnegatively modulates tyrosine phosphorylation and tyrosine kinaseactivity during capacitation of human spermatozoa. Biol. Reprod. 1996,55, 207−216.(90) Kirichok, Y.; Navarro, B.; Clapham, D. E. Whole-cell patch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+
channel. Nature 2006, 439, 737−740.(91) Kaneto, M.; Krisfalusi, M.; Eddy, E. M.; O’Brien, D. A.; Miki, K.Bicarbonate-induced phosphorylation of p270 protein in mouse spermby cAMP-dependent protein kinase. Mol. Reprod. Dev. 2008, 75,1045−1053.(92) Chatterjee, M.; Nandi, P.; Ghosh, S.; Sen, P. C. Regulation oftyrosine kinase activity during capacitation in goat sperm. Mol. Cell.Biochem. 2010, 336, 39−48.(93) Baker, M. A.; Hetherington, L.; Curry, B.; Aitken, R. J.Phosphorylation and consequent stimulation of the tyrosine kinase c-Abl by PKA in mouse spermatozoa; its implications duringcapacitation. Dev. Biol. 2009, 333, 57−66.(94) Tartaglione, C. M.; Ritta, M. N. On the presence of 3H-GABAuptake mechanism in bovine spermatozoa. Anim. Reprod. Sci. 2008,108, 247−258.(95) Sidhu, K. S.; Mate, K. E.; Gunasekera, T.; Veal, D.;Hetherington, L.; Baker, M. A.; Aitken, R. J.; Rodger, J. C. A flowcytometric assay for global estimation of tyrosine phosphorylationassociated with capacitation of spermatozoa from two marsupialspecies, the tammar wallaby (Macropus eugenii) and the brushtailpossum (Trichosurus vulpecula). Reproduction 2004, 127, 95−103.(96) Galantino-Homer, H. L.; Florman, H. M.; Storey, B. T.;Dobrinski, I.; Kopf, G. S. Bovine sperm capacitation: assessment ofphosphodiesterase activity and intracellular alkalinization on capacita-tionassociated protein tyrosine phosphorylation. Mol. Reprod. Dev.2004, 67, 487−500.(97) Pommer, A. C.; Rutllant, J.; Meyers, S. A. Phosphorylation ofprotein tyrosine residues in fresh and cryopreserved stallionspermatozoa under capacitating conditions. Biol. Reprod. 2003, 68,1208−1214.(98) Tardif, S.; Dube, C.; Chevalier, S.; Bailey, J. L. Capacitation isassociated with tyrosine phosphorylation and tyrosine kinase-likeactivity of pig sperm proteins. Biol. Reprod. 2001, 65, 784−792.(99) Kulanand, J.; Shivaji, S. Capacitation-associated changes inprotein tyrosine phosphorylation, hyperactivation and acrosomereaction in hamster spermatozoa. Andrologia 2001, 33, 95−104.(100) Osheroff, J. E.; Visconti, P. E.; Valenzuela, J. P.; Travis, A. J.;Alvarez, J.; Kopf, G. S. Regulation of human sperm capacitation by acholesterol efflux-stimulated signal transduction pathway leading toprotein kinase A-mediated up-regulation of protein tyrosinephosphorylation. Mol. Hum. Reprod. 1999, 5, 1017−1026.(101) Kalab, P.; Peknicova, J.; Geussova, G.; Moos, J. Regulation ofprotein tyrosine phosphorylation in boar sperm through a cAMP-dependent pathway. Mol. Reprod. Dev. 1998, 51, 304−314.(102) Kong, M.; Diaz, E. S.; Morales, P. Participation of the humansperm proteasome in the capacitation process and its regulation byprotein kinase A and tyrosine kinase. Biol. Reprod. 2009, 80, 1026−1035.(103) Moseley, F. L.; Jha, K. N.; Bjorndahl, L.; Brewis, I. A.;Publicover, S. J.; Barratt, C. L.; Lefievre, L. Protein tyrosinephosphorylation, hyperactivation and progesterone-induced acrosome
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174515

reaction are enhanced in IVF media: an effect that is not associatedwith an increase in protein kinase A activation. Mol. Hum. Reprod.2005, 11, 523−529.(104) Ecroyd, H.; Asquith, K. L.; Jones, R. C.; Aitken, R. J. Thedevelopment of signal transduction pathways during epididymalmaturation is calcium dependent. Dev. Biol. 2004, 268, 53−63.(105) Maher, G. J.; Goriely, A.; Wilkie, A. O. Cellular evidence forselfish spermatogonial selection in aged human testes. Andrology 2014,2, 304−314.(106) Wan, H. T.; Mruk, D. D.; Li, S. Y.; Mok, K. W.; Lee, W. M.;Wong, C. K.; Cheng, C. Y. p-FAK-Tyr(397) regulates spermatidadhesion in the rat testis via its effects on F-actin organization at theectoplasmic specialization. Am. J. Physiol. Endocrinol. Metab. 2013, 305,687−699.(107) Adeoya-Osiguwa, S. A.; Gibbons, R.; Fraser, L. R. Identificationof functional alpha2- and beta-adrenergic receptors in mammalians-permatozoa. Hum. Reprod. 2006, 21, 1555−1563.(108) Maccarrone, M.; Barboni, B.; Paradisi, A.; Bernabo, N.;Gasperi, V.; Pistilli, M. G.; Fezza, F.; Lucidi, P.; Mattioli, M.Characterization of the endocannabinoid system in boar spermatozoaand implications for sperm capacitation and acrosome reaction. J. CellSci. 2005, 118, 4393−4404.(109) Schuel, H.; Burkman, L. J.; Lippes, J.; Crickard, K.; Mahony,M. C.; Giuffrida, A.; Picone, R. P.; Makriyannis, A. Evidence thatanandamide-signaling regulates human sperm functions required forfertilization. Mol. Reprod. Dev. 2002, 63, 376−387.(110) Galantino-Homer, H. L.; Visconti, P. E.; Kopf, G. S. Regulationof protein tyrosine phosphorylation during bovine sperm capacitationby a cyclic adenosine 3′5′-monophosphate-dependent pathway. Biol.Reprod. 1997, 56, 707−719.(111) Eddy, E. M.; O’Brien, D. A. The spermatozoon: The Physiologyof Reproduction; Knobil, E., Neill, J. D., Eds.; Raven Press: New York,1994; pp 29−77.(112) Leclerc, P.; de Lamirande, E.; Gagnon, C. Cyclic adenosine3′,5′monophosphate-dependent regulation of protein tyrosine phos-phorylation in relation to human sperm capacitation and motility. Biol.Reprod. 1996, 55, 684−692.(113) Visconti, P. E.; Muschietti, J. P.; Flawia, M. M.; Tezon, J. G.Bicarbonate dependence of cAMP accumulation induced by phorbolesters in hamster spermatozoa. Biochim. Biophys. Acta 1990, 1054,231−236.(114) Visconti, P. E.; Kopf, G. S. Regulation of proteinphosphorylation during sperm capacitation. Biol. Reprod. 1998, 59,1−6.(115) Zini, A.; De Lamirande, E.; Gagnon, C. Low levels of nitricoxide promote human sperm capacitation in vitro. J. Androl. 1995, 16,424−431.(116) Rodriguez, P. C.; O’Flaherty, C. M.; Beconi, M. T.; Beorlegui,N. B. Nitric oxide-induced capacitation of cryopreserved bullspermatozoa and assessment of participating regulatory pathways.Anim. Reprod. Sci. 2005, 85, 231−242.(117) Herrero, M. B.; de Lamirande, E.; Gagnon, C. Nitric oxideregulates human sperm capacitation and protein-tyrosine phosphor-ylation in vitro. Biol. Reprod. 1999, 61, 575−581.(118) Buzadzic, B.; Vucetic, M.; Jankovic, A.; Stancic, A.; Korac, A.;Korac, B.; Otasevic, V. New insights into male (in)fertility: theimportance of NO. Br. J. Pharmacol. 2014, Mar 7, doi: 10.1111/bph.12675 [Epub ahead of print].(119) Jagan Mohanarao, G.; Atreja, S. K. Identification of NOinduced and capacitation associated tyrosine phosphoproteins inbuffalo (Bubalus bubalis) spermatozoa. Res. Vet. Sci. 2012, 93, 618−23.(120) Garty, N.; Salomon, Y. Stimulation of partially purifiedadenylate cyclase from bull sperm by bicarbonate. FEBS. Lett. 1987,218, 148−152.(121) Hyne, R. V.; Garbers, D. L. Regulation of guinea pig spermadenylate cyclase by calcium. Biol. Reprod. 1979, 21, 1135−1142.(122) Cross, N. L.; Razy-Faulkner, P. Control of human spermintracellular pH by cholesterol and its relationship to the response ofthe acrosome to progesterone. Biol. Reprod. 1997, 56, 1169−1174.
(123) Guthrie, H. D.; Welch, G. R. Effects of reactive oxygen specieson sperm function. Theriogenology 2012, 78, 1700−1708.(124) Tremellen, K. Oxidative stress and male infertility-a clinicalperspective. Hum. Reprod. Update 2008, 14, 243−558.(125) Leclerc, P.; de Lamirande, E.; Gagnon, C. Regulation ofprotein-tyrosine phosphorylation and human sperm capacitation byreactive oxygen derivatives. Free Radical Biol. Med. 1997, 22, 643−656.(126) Aitken, R. J.; Paterson, M.; Fisher, H.; Buckingham, D. W.; VanDuin, M. Redox regulation of tyrosine phosphorylation in humanspermatozoa and its role in the control of human sperm function. J.Cell Sci. 1995, 108, 2017−2025.(127) Uguz, C.; Vredenburgh, W. L.; Parrish, J. J. Heparin-inducedcapacitation but not intracellular alkalinization of bovine sperm isinhibited by Rp-adenosine-3′,5′-cyclic monophosphothioate. Biol.Reprod. 1994, 51, 1031−1039.(128) Parrish, J. J.; Susko-Parrish, J.; Winer, M. A.; First, N. L.Capacitation of bovine sperm by heparin. Biol. Reprod. 1988, 38,1171−1180.(129) Miller, D. J.; Ax, R. L. Carbohydrates and fertilization inanimals. Mol. Reprod. Dev. 1990, 26, 184−198.(130) Hardy, D. M.; Garbers, D. L. A sperm membrane protein thatbinds in a species-specific manner to the egg extracellular matrix ishomologous to von Willebrand factor. J. Biol. Chem. 1995, 3, 26025−26028.(131) Herlyn, H.; Zischler, H. Sequence evolution, processing, andposttranslational modification of zonadhesin D domains in primates, asinferred from cDNA data. Gene 2005, 362, 85−97.(132) Tesarik, J.; Moos, J.; Mendoza, C. Stimulation of proteintyrosine phosphorylation by a progesterone receptor on the cellsurface of human sperm. Endocrinology 1993, 133, 328−335.(133) Publicover, S.; Barratt, C. Reproductive biology: Progester-one’s gateway into sperm. Nature 2011, 471, 313−314.(134) Kadirvel, G.; Kathiravan, P.; Kumar, S. Protein tyrosinephosphorylation and zona binding ability of in vitro capacitated andcryopreserved buffalo spermatozoa. Theriogenology 2011, 75, 1630−1639.(135) Sato, K.; Iwasaki, T.; Tamaki, I.; Aoto, M.; Tokmakov, A. A.;Fukami, Y. Involvement of protein-tyrosine phosphorylation anddephosphorylation in sperm-induced Xenopus egg activation. FEBSLett. 1998, 424, 113−118.(136) Leyton, L.; Saling, P. 95 kd sperm proteins bind ZP3 and serveas tyrosine kinase substrates in response to zona binding. Cell 1989,57, 1123−1130.(137) Kaplan, P.; Naz, R. K. The fertilization antigen-1 does not haveproteolytic/acrosin activity, but its monoclonal antibody inhibitssperm capacitation and acrosome reaction. Fertil. Steril. 1992, 58, 396−402.(138) Flesch, F. M.; Colenbrander, B.; van Golde, L. M.; Gadella, B.M. Capacitation induces tyrosine phosphorylation of proteins in theboar sperm plasma membrane. Biochem. Biophys. Res. Commun. 1999,262, 787−792.(139) Flesch, F. M.; Wijnand, E.; van de Lest, C. H.; Colenbrander,B.; van Golde, L. M.; Gadella, B. M. Capacitation dependent activationof tyrosine phosphorylation generates two sperm head plasmamembrane proteins with high primary binding affinity for the zonapellucida. Mol. Reprod. Dev. 2001, 60, 107−115.(140) Pukazhenthi, B. S.; Wildt, D. E.; Ottinger, M. A.; Howard, J.Inhibition of domestic cat spermatozoa acrosome reaction and zonapellucida penetration by tyrosine kinase inhibitors. Mol. Reprod. Dev.1998, 49, 48−57.(141) Arcelay, E.; Salicioni, A. M.; Wertheimer, E.; Visconti, P. E.Identification of proteins undergoing tyrosine phosphorylation duringmouse sperm capacitation. Int. J. Dev. Biol. 2008, 52, 463−472.(142) Kumar, V.; Rangaraj, N.; Shivaji, S. Activity of pyruvatedehydrogenase A (PDHA) in hamster spermatozoa correlatespositively with hyperactivation and is associated with spermcapacitation. Biol. Reprod. 2006, 75, 767−777.(143) Foresta, C.; Flohe, L.; Garolla, A.; Roveri, A.; Ursini, F.;Maiorino, M. Male fertility is linked to the selenoprotein phospholipid
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174516

hydroperoxide glutathione peroxidase. Biol. Reprod. 2002, 67, 967−971.(144) Tardif, S.; Guyonnet, B.; Cormier, N.; Cornwall, G. A.Alteration in the processing of the ACRBP/sp32 protein and spermhead/acrosome malformations in proprotein convertase 4 (PCSK4)null mice. Mol. Hum. Reprod. 2012, 18, 298−307.(145) Miki, K.; Qu, W.; Goulding, E. H.; Willis, W. D.; Bunch, D. O.;Strader, L. F.; Perreault, S. D.; Eddy, E. M.; O’Brien, D. A.Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specificglycolytic enzyme, is required for sperm motility and male fertility.Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 16501−16506.(146) Naaby-Hansen, S.; Mandal, A.; Wolkowicz, M. J.; Sen, B.;Westbrook, V. A.; Shetty, J.; Coonrod, S. A.; Klotz, K. L.; Kim, Y. H.;Bush, L. A.; Flickinger, C. J.; Herr, J. C. CABYR, a novel calcium-binding tyrosine phosphorylation-regulated fibrous sheath proteininvolved in capacitation. Dev. Biol. 2002, 242, 236−254.(147) Jha, K. N.; Shivaji, S. Identification of the major tyrosinephosphorylated protein of capacitated hamster spermatozoa as ahomologue of mammalian sperm a kinase anchoring protein. Mol.Reprod. Dev. 2002, 61, 258−270.(148) Ecroyd, H.; Jones, R. C.; Aitken, R. J. Tyrosinephosphorylation of HSP-90 during mammalian sperm capacitation.Biol. Reprod. 2003, 69, 1801−1807.(149) Asquith, K. L.; Baleato, R. M.; McLaughlin, E. A.; Nixon, B.;Aitken, R. J. Tyrosine phosphorylation activates surface chaperonesfacilitating sperm-zona recognition. J. Cell Sci. 2004, 117, 3645−3657.(150) Baldi, E.; Luconi, M.; Bonaccorsi, L.; Muratori, M.; Forti, G.Intracellular events and signaling pathways involved in spermacquisition of fertilizing capacity and acrosome reaction. Front. Biosci.2000, 5, 110−123.
Journal of Proteome Research Reviews
dx.doi.org/10.1021/pr500524p | J. Proteome Res. 2014, 13, 4505−45174517





























