Embed Size (px)
Citation preview

Developmental Cell
Article
Distinct Regulatory Cascades Govern Extraocularand Pharyngeal Arch Muscle Progenitor Cell FatesRamkumar Sambasivan,1,2 Barbara Gayraud-Morel,1,2 Gerard Dumas,1,2 Clemire Cimper,2 Sylvain Paisant,1,2
Robert Kelly,3 and Shahragim Tajbakhsh1,2,*1Institut Pasteur, Stem Cells & Development, CNRS URA 25782Department of Developmental Biology, 25 rue du Dr. Roux, 75724 Paris Cedex 153Developmental Biology Institute of Marseilles-Luminy, UMR 6216 CNRS
Universite de la Mediterranee, Campus de Luminy, Case 907, 13288 Marseille Cedex 9, France
*Correspondence: [email protected]
DOI 10.1016/j.devcel.2009.05.008
SUMMARY
Genetic regulatory networks governing skeletal myo-genesis in the body are well understood, yet theirhierarchical relationships in the head remain unre-solved. We show that either Myf5 or Mrf4 is neces-sary for initiating extraocular myogenesis. WhereasMrf4 is dispensable for pharyngeal muscle progen-itor fate, Tbx1 and Myf5 act synergistically for gov-erning myogenesis in this location. As in the body,Myod acts epistatically to the initiating cascades inthe head. Thus, complementary pathways, governedby Pax3 for body, and Tbx1 for pharyngeal muscles,but absent for extraocular muscles, activate the coremyogenic network. These diverse muscle progeni-tors maintain their respective embryonic regulatorysignatures in the adult. However, these signaturesare not sufficient to ensure the specific muscle pheno-types, since the expected differentiated phenotypeis not manifested when satellite cells are engraftedheterotopically. These findings identify novel geneticnetworks that may provide insights into myopathieswhich often affect only subsets of muscles.
INTRODUCTION
During development, instructive signaling and positional cues
impinge on gene regulatory networks that act in a hierarchical
fashion for the specification, commitment, and differentiation
of stem and progenitor cells. Transcriptional cascades that
govern lineage commitment are crucial for orchestrating organ-
ogenesis and development. Muscle stem cell self-renewal and
commitment are controlled by a genetic network including the
paired-box transcription factors Pax3 and Pax7, and basic
helix-loop-helix muscle regulatory factors (MRFs) Myf5, Mrf4,
Myod, and Myogenin (Kassar-Duchossoy et al., 2004; Relaix
et al., 2005; Rudnicki et al., 1993; Seale et al., 2000; Tajbakhsh
et al., 1997). Genes in the core regulatory network comprising
Myf5, Mrf4, and Myod act as the obligate determinants for di-
recting myogenic cell fate (Kassar-Duchossoy et al., 2004; Rud-
nicki et al., 1993), whereas Myogenin functions as an essential
differentiation factor (Hasty et al., 1993; Nabeshima et al., 1993).
810 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier
The genetic linkage between Myf5 and Mrf4 had obscured the
role of Mrf4 in earlier studies (Braun and Arnold, 1995; Olson
et al., 1996; Rudnicki et al., 1993; Tajbakhsh et al., 1996,
1997). Using an allelic series of Myf5 mutants that differentially
affect Mrf4 expression, it was shown that Mrf4 and Myf5 act
upstream of Myod in the myogenic program of somites (Kas-
sar-Duchossoy et al., 2004). Myogenesis in the head is less
well investigated (Grifone and Kelly, 2007), and it is generally
thought that Myf5 and Myod act redundantly and in parallel for
initiating head muscle cell fate, while the role of Mrf4 is unknown.
In vertebrates, muscle progenitor cell (MPC) specification
occurs first in somites (Sambasivan and Tajbakhsh, 2007) and
these MPCs make body muscles. Notably, MPCs in the head
are derived from cranial mesoderm (Noden and Francis-West,
2006), and some signaling cascades act distinctly in somitic
and cranial mesoderm (Hacker and Guthrie, 1998; Mootoosamy
et al., 2002; Tzahor et al., 2003). Whereas extraocular muscles
(EOMs) develop in the periocular region, muscles of the face,
jaw, and neck develop from progenitor cells in the mesoderm
of the pharyngeal arches (PAs). EOMs govern eye movements
and have unique properties including expression of specific
Myosin heavy-chain isoforms (Porter, 2002; Sartore et al., 1987).
Pax3 and Pax7 play critical roles in somitic progenitors (Sam-
basivan and Tajbakhsh, 2007). Notably, Pax3:Pax7 double-
mutant somites are severely hypoplastic (Relaix et al., 2005)
where muscle stem and progenitor cells are eventually lost by
apoptosis; therefore, these genes regulate cell survival as well
(see Sambasivan and Tajbakhsh, 2007). Remarkably, Pax3:
Myf5(Mrf4) mutants (Mrf4 compromised in cis) uncouple myo-
genesis in the head from that in the body, where in the latter,
muscles do not form due to a failure to activate Myod (Tajbakhsh
et al., 1997). Therefore, Pax3 acts in a genetic pathway parallel to
Myf5 and Mrf4, and rescues Myod-mediated myogenesis in their
absence.
Although these regulatory networks have been largely estab-
lished for somite derived muscles, the hierarchical interactions
conferring muscle cell fate in the head have been elusive (Grifone
and Kelly, 2007; Sambasivan and Tajbakhsh, 2007). Resolving
these genetic relationships is critical since different myopathies
affect only subgroups of muscles in the body or head (Emery,
2002), yet the underlying mechanism of these clinical observa-
tions remains unresolved. Given that overall head muscle devel-
opment appears to be unhindered in Pax3:Myf5(Mrf4) mutants
and that Pax3 is not expressed in head MPCs or muscles,
Inc.

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
Pax7, which is expressed in adult head progenitors (Horst et al.,
2006; Nathan et al., 2008), or other transcriptional regulators,
may cooperate with the core MRF network in this location. The
sine oculus (Six) and eyes absent (Eya) genes are key regulators
as Six1:Six4 and Eya1:Eya2 double nulls are deficient in limb
MPCs (Grifone et al., 2005, 2007). However, they do not rescue
Myod activation in Pax3:Myf5(Mrf4) mutants (Tajbakhsh et al.,
1997), suggesting that they act outside the core (MRFs) and
complementary (Pax3) network.
Interestingly, Musculin (Msc or MyoR), Tcf21 (Capsulin), Pitx2,
and Tbx1 play important roles in head muscle development (Gri-
fone and Kelly, 2007; Noden and Francis-West, 2006). Embryos
double null for the transcriptional repressors Msc and Tcf21 fail
to develop a subset of first-arch-derived muscles (Lu et al.,
2002). The bicoid-related homeodomain gene Pitx2 and the T-
box containing gene Tbx1 are expressed widely in the devel-
oping head, as well as in the body (Chapman et al., 1996; Kita-
mura et al., 1999; L’Honore et al., 2007). In Pitx2 null mice,
first-arch-derived muscles do not develop normally (Dong
et al., 2006; Shih et al., 2007), EOMs are lost (Gage et al.,
1999; Kitamura et al., 1999), and their progenitors mislocalized
(Dong et al., 2006). In Tbx1 mutants, myogenesis in the first
arch is impaired and muscles derived from other arches are
absent due primarily to a loss of these structures (Jerome and
Papaioannou, 2001; Kelly et al., 2004). A random complement
of hypomorphic first-arch-derived muscles are observed at later
stages in Tbx1 null embryos, revealing a requirement for Tbx1 in
ensuring robust bilateral activation of the myogenic program in
the mandibular arch (Kelly et al., 2004). Therefore, a complex
circuitry governs head myogenesis (PA versus EOM) and the
genetic relationships between these regulators, and with the
core MRFs, remain unresolved.
Using genetic manipulations and cell transplantations in mice,
we show that head and body myogenic regulatory cascades
have co-opted different combinations of a core and complemen-
tary pathway. Unlike all other skeletal muscles in the embryo,
EOMs are entirely dependent on Myf5/Mrf4. We show also
that, like Pax3 in the body, Tbx1 operates complementary to
Myf5 and acts genetically upstream of Myod in the PA. Interest-
ingly, the molecular signature of adult progenitors reflects their
developmental history, yet heterotopically transplantated adult
head satellite cells do not retain their original phenotype upon
differentiation. We propose that multiple distinct genetic
programs regulate overall skeletal muscle development in the
mouse.
RESULTS
An Obligate Requirement for Myf5 or Mrf4 to InitiateEOM Cell FateTo investigate developmental myogenesis in the vertebrate
head, we focused first on the patterning of EOMs in mice. How
MPCs are born, migrate, and pattern the EOMs is unknown.
Myf5nlacZ/+ and Myf5nlacZ/nlacZ embryos of different stages were
stained using X-gal for whole mount visualization of EOMs. A
single bilateral EOM anlage which is apparent at E10.5 (see
Figure S1A available online), appears to initiate the other EOMs
after splitting and patterning as evidenced at E11.5 and E12.5
(Figures 1A, 1C, 1E, S1C, S1D, and S1E). By E13.5 all EOMs
Deve
are properly positioned (data not shown). In Myf5nlacZ/nlacZ
embryos (Mrf4 also inactivated in cis; Kassar-Duchossoy et al.,
2004), the initial EOM anlage was formed and expanded in the
mutants, but surprisingly, it failed to pattern and began to
atrophy by E11.5 (Figures 1B and 1D; Figure S1B). This initial
growth, followed by a decline, indicates that the MPCs are
born but are eventually lost in these mutants. This is not the
case for any of the other MPCs (somites, limbs, pharyngeal
arches; Tajbakhsh et al., 1996, 1997; data not shown). Staining
with antibodies against activated Caspase-3 and TUNUL
labeling, which mark apoptotic cells, revealed that the mutant
anlagen had 5–6-fold more apoptotic cells compared to the
control (Figures 1F and 1H–1I0, and data not shown). Costaining
with an anti-b-galactosidase antibody to demarcate this struc-
ture and Ki67 or Cyclin D1 to identify cycling cells, showed no
significant difference in the proliferation of mutant progenitors
(Figures 1G and 1J–1K0; data not shown). Thus, disruption of
EOM patterning as well as loss of EOM progenitors by apoptosis
occurs in Myf5nlacZ/nlacZ mutants.
Mrf4 is expressed in EOM progenitors (Figures S1F–S1H00) and
since the Myf5nlacZ allele results in compromised Mrf4 expres-
sion, we wondered if the EOM defects in Myf5nlacZ/nlacZ mutants
were due to the combined loss of these genes. To address this
point, we examined an allelic series of Myf5 knockins which
progressively affect Mrf4 expression in MPCs: Myf5loxP/loxP (not
affected significantly), Myf5GFP-P/GFP-P (�50% reduced), and
Myf5nlacZ/nlacZ (fully compromised) (Kassar-Duchossoy et al.,
2004). At E14.5, a stereotyped EOM pattern is normally observed
by immunostaining for Myosin Heavy chain (MyHC; Figures 2A–
2C). Notably, we found a range of EOM phenotypes correspond-
ing to the extent of disruption of Mrf4 expression among the
Myf5 null alleles (Figures 2D–2F). In Myf5loxP/loxP mutants, all
anlage of EOMs were present, although some were hypoplastic
(Figure 2D). Myf5GFP-P/GFP-P mutants displayed an intermediate
phenotype, where some EOMs were severely reduced, and
others were missing (Figure 2E). Notably, Myf5nlacZ/nlacZ embryos
completely lacked all EOMs or had only residual myofibers at
birth (Figure 2F; data not shown). Accordingly, Mrf4nlacZ-P/nlacZ-P:
Myf5loxP/loxP mutants where both Mrf4 and Myf5 are inactivated
by gene targeting (Kassar-Duchossoy et al., 2004) phenocopied
the Myf5nlacZ/nlacZ mutant (Figure 2G).
To determine if Mrf4 is necessary for EOM development we
examined Mrf4�/� null mutants (Zhang et al., 1995). These
embryos have mild EOM defects (Figure 2H), comparable in
severity to Myf5loxP/loxP. To assess if delayed activation of
Myod or Myogenin eventually rescues extraocular myogenesis,
we dissected out eyeballs from adult mutants along with the
retinue of EOMs and immunostained them in whole-mount with
anti-MyHC antibody. As shown in Figures 2J–2L, newborn or
adult mutants display the defects that reflect those observed in
the early fetus. Thus, we conclude that either Myf5 or Mrf4 is
necessary for EOM progenitors to acquire their myogenic fate,
and unlike muscles elsewhere, Myod does not rescue EOM
development.
Lineage Progression and Cell Survival Are Distinctin EOM and Somitic MPCsWe reported previously that somitic MPCs are developmentally
arrested along the dermomyotome (DM) epithelium for about
lopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 811
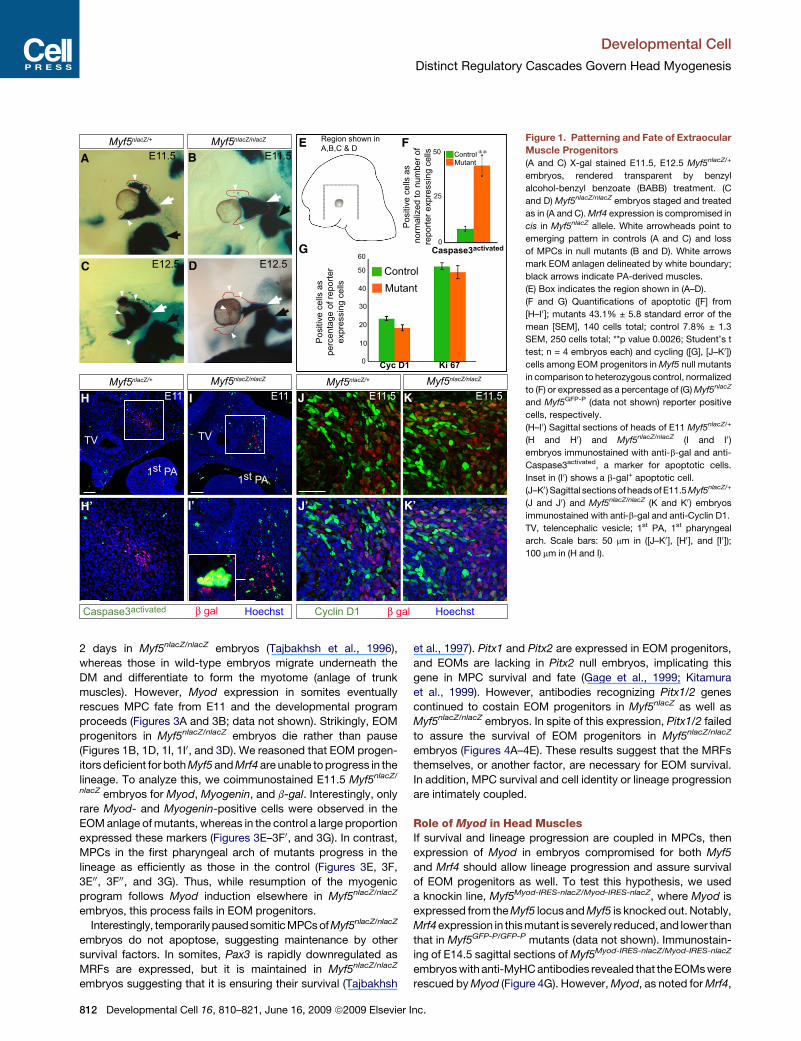
Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
A
G
B
C
J’ K’
D
J
H’ I’
E
KH I
F Figure 1. Patterning and Fate of Extraocular
Muscle Progenitors
(A and C) X-gal stained E11.5, E12.5 Myf5nlacZ/+
embryos, rendered transparent by benzyl
alcohol-benzyl benzoate (BABB) treatment. (C
and D) Myf5nlacZ/nlacZ embryos staged and treated
as in (A and C). Mrf4 expression is compromised in
cis in Myf5nlacZ allele. White arrowheads point to
emerging pattern in controls (A and C) and loss
of MPCs in null mutants (B and D). White arrows
mark EOM anlagen delineated by white boundary;
black arrows indicate PA-derived muscles.
(E) Box indicates the region shown in (A–D).
(F and G) Quantifications of apoptotic ([F] from
[H–I0]; mutants 43.1% ± 5.8 standard error of the
mean [SEM], 140 cells total; control 7.8% ± 1.3
SEM, 250 cells total; **p value 0.0026; Student’s t
test; n = 4 embryos each) and cycling ([G], [J–K0])
cells among EOM progenitors in Myf5 null mutants
in comparison to heterozygous control, normalized
to (F) or expressed as a percentage of (G) Myf5nlacZ
and Myf5GFP-P (data not shown) reporter positive
cells, respectively.
(H–I0) Sagittal sections of heads of E11 Myf5nlacZ/+
(H and H0 ) and Myf5nlacZ/nlacZ (I and I0)
embryos immunostained with anti-b-gal and anti-
Caspase3activated, a marker for apoptotic cells.
Inset in (I0) shows a b-gal+ apoptotic cell.
(J–K0 ) Sagittal sections of heads of E11.5 Myf5nlacZ/+
(J and J0 ) and Myf5nlacZ/nlacZ (K and K0) embryos
immunostained with anti-b-gal and anti-Cyclin D1.
TV, telencephalic vesicle; 1st PA, 1st pharyngeal
arch. Scale bars: 50 mm in ([J–K0], [H0], and [I0]);
100 mm in (H and I).
2 days in Myf5nlacZ/nlacZ embryos (Tajbakhsh et al., 1996),
whereas those in wild-type embryos migrate underneath the
DM and differentiate to form the myotome (anlage of trunk
muscles). However, Myod expression in somites eventually
rescues MPC fate from E11 and the developmental program
proceeds (Figures 3A and 3B; data not shown). Strikingly, EOM
progenitors in Myf5nlacZ/nlacZ embryos die rather than pause
(Figures 1B, 1D, 1I, 1I0, and 3D). We reasoned that EOM progen-
itors deficient for both Myf5 and Mrf4 are unable to progress in the
lineage. To analyze this, we coimmunostained E11.5 Myf5nlacZ/
nlacZ embryos for Myod, Myogenin, and b-gal. Interestingly, only
rare Myod- and Myogenin-positive cells were observed in the
EOM anlage of mutants, whereas in the control a large proportion
expressed these markers (Figures 3E–3F0, and 3G). In contrast,
MPCs in the first pharyngeal arch of mutants progress in the
lineage as efficiently as those in the control (Figures 3E, 3F,
3E00, 3F00, and 3G). Thus, while resumption of the myogenic
program follows Myod induction elsewhere in Myf5nlacZ/nlacZ
embryos, this process fails in EOM progenitors.
Interestingly, temporarily pausedsomitic MPCs of Myf5nlacZ/nlacZ
embryos do not apoptose, suggesting maintenance by other
survival factors. In somites, Pax3 is rapidly downregulated as
MRFs are expressed, but it is maintained in Myf5nlacZ/nlacZ
embryos suggesting that it is ensuring their survival (Tajbakhsh
812 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier
et al., 1997). Pitx1 and Pitx2 are expressed in EOM progenitors,
and EOMs are lacking in Pitx2 null embryos, implicating this
gene in MPC survival and fate (Gage et al., 1999; Kitamura
et al., 1999). However, antibodies recognizing Pitx1/2 genes
continued to costain EOM progenitors in Myf5nlacZ as well as
Myf5nlacZ/nlacZ embryos. In spite of this expression, Pitx1/2 failed
to assure the survival of EOM progenitors in Myf5nlacZ/nlacZ
embryos (Figures 4A–4E). These results suggest that the MRFs
themselves, or another factor, are necessary for EOM survival.
In addition, MPC survival and cell identity or lineage progression
are intimately coupled.
Role of Myod in Head MusclesIf survival and lineage progression are coupled in MPCs, then
expression of Myod in embryos compromised for both Myf5
and Mrf4 should allow lineage progression and assure survival
of EOM progenitors as well. To test this hypothesis, we used
a knockin line, Myf5Myod-IRES-nlacZ/Myod-IRES-nlacZ, where Myod is
expressed from the Myf5 locus and Myf5 is knocked out. Notably,
Mrf4expression in thismutant is severely reduced, and lower than
that in Myf5GFP-P/GFP-P mutants (data not shown). Immunostain-
ing of E14.5 sagittal sections of Myf5Myod-IRES-nlacZ/Myod-IRES-nlacZ
embryos with anti-MyHC antibodies revealed that the EOMs were
rescued by Myod (Figure 4G). However, Myod, as noted for Mrf4,
Inc.
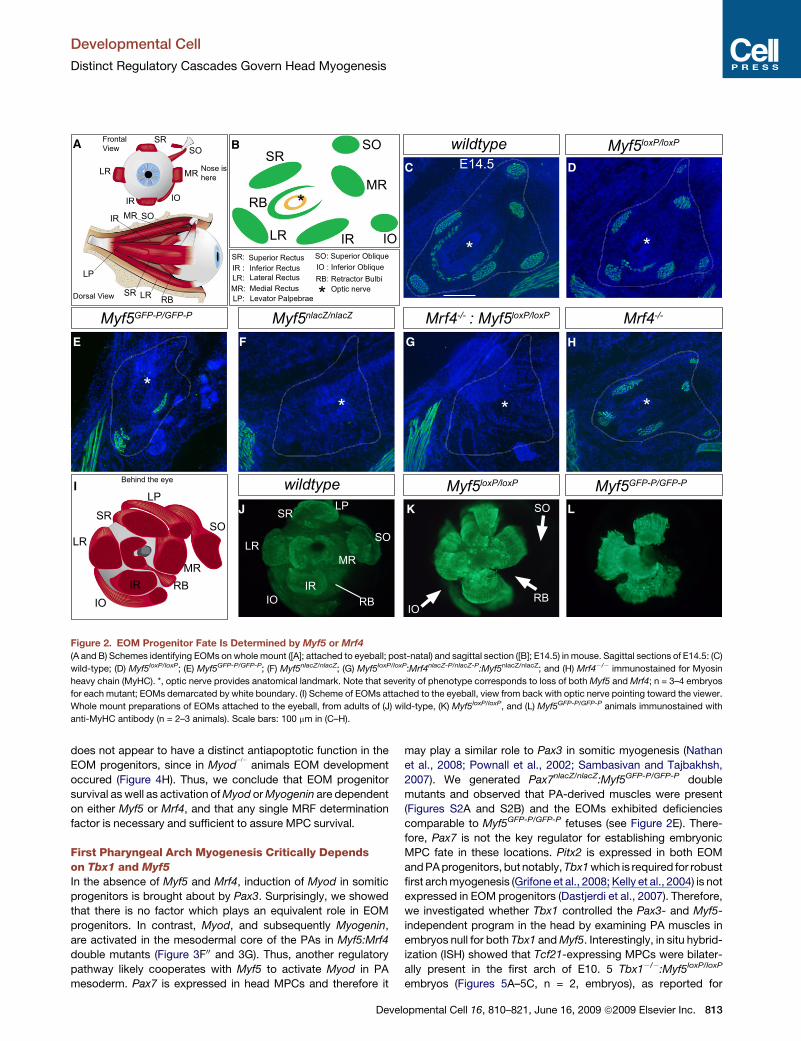
Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
Figure 2. EOM Progenitor Fate Is Determined by Myf5 or Mrf4
(A and B) Schemes identifying EOMs on whole mount ([A]; attached to eyeball; post-natal) and sagittal section ([B]; E14.5) in mouse. Sagittal sections of E14.5: (C)
wild-type; (D) Myf5loxP/loxP; (E) Myf5GFP-P/GFP-P; (F) Myf5nlacZ/nlacZ; (G) Myf5loxP/loxP:Mrf4nlacZ-P/nlacZ-P:Myf5nlacZ/nlacZ; and (H) Mrf4�/� immunostained for Myosin
heavy chain (MyHC). *, optic nerve provides anatomical landmark. Note that severity of phenotype corresponds to loss of both Myf5 and Mrf4; n = 3–4 embryos
for each mutant; EOMs demarcated by white boundary. (I) Scheme of EOMs attached to the eyeball, view from back with optic nerve pointing toward the viewer.
Whole mount preparations of EOMs attached to the eyeball, from adults of (J) wild-type, (K) Myf5loxP/loxP, and (L) Myf5GFP-P/GFP-P animals immunostained with
anti-MyHC antibody (n = 2–3 animals). Scale bars: 100 mm in (C–H).
does not appear to have a distinct antiapoptotic function in the
EOM progenitors, since in Myod�/�
animals EOM development
occured (Figure 4H). Thus, we conclude that EOM progenitor
survival as well as activation of Myod or Myogenin are dependent
on either Myf5 or Mrf4, and that any single MRF determination
factor is necessary and sufficient to assure MPC survival.
First Pharyngeal Arch Myogenesis Critically Dependson Tbx1 and Myf5
In the absence of Myf5 and Mrf4, induction of Myod in somitic
progenitors is brought about by Pax3. Surprisingly, we showed
that there is no factor which plays an equivalent role in EOM
progenitors. In contrast, Myod, and subsequently Myogenin,
are activated in the mesodermal core of the PAs in Myf5:Mrf4
double mutants (Figure 3F00 and 3G). Thus, another regulatory
pathway likely cooperates with Myf5 to activate Myod in PA
mesoderm. Pax7 is expressed in head MPCs and therefore it
Deve
may play a similar role to Pax3 in somitic myogenesis (Nathan
et al., 2008; Pownall et al., 2002; Sambasivan and Tajbakhsh,
2007). We generated Pax7nlacZ/nlacZ:Myf5GFP-P/GFP-P double
mutants and observed that PA-derived muscles were present
(Figures S2A and S2B) and the EOMs exhibited deficiencies
comparable to Myf5GFP-P/GFP-P fetuses (see Figure 2E). There-
fore, Pax7 is not the key regulator for establishing embryonic
MPC fate in these locations. Pitx2 is expressed in both EOM
and PA progenitors, but notably, Tbx1 which is required for robust
first arch myogenesis (Grifone et al., 2008; Kelly et al., 2004) is not
expressed in EOM progenitors (Dastjerdi et al., 2007). Therefore,
we investigated whether Tbx1 controlled the Pax3- and Myf5-
independent program in the head by examining PA muscles in
embryos null for both Tbx1 and Myf5. Interestingly, in situ hybrid-
ization (ISH) showed that Tcf21-expressing MPCs were bilater-
ally present in the first arch of E10. 5 Tbx1�/�:Myf5loxP/loxP
embryos (Figures 5A–5C, n = 2, embryos), as reported for
lopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 813
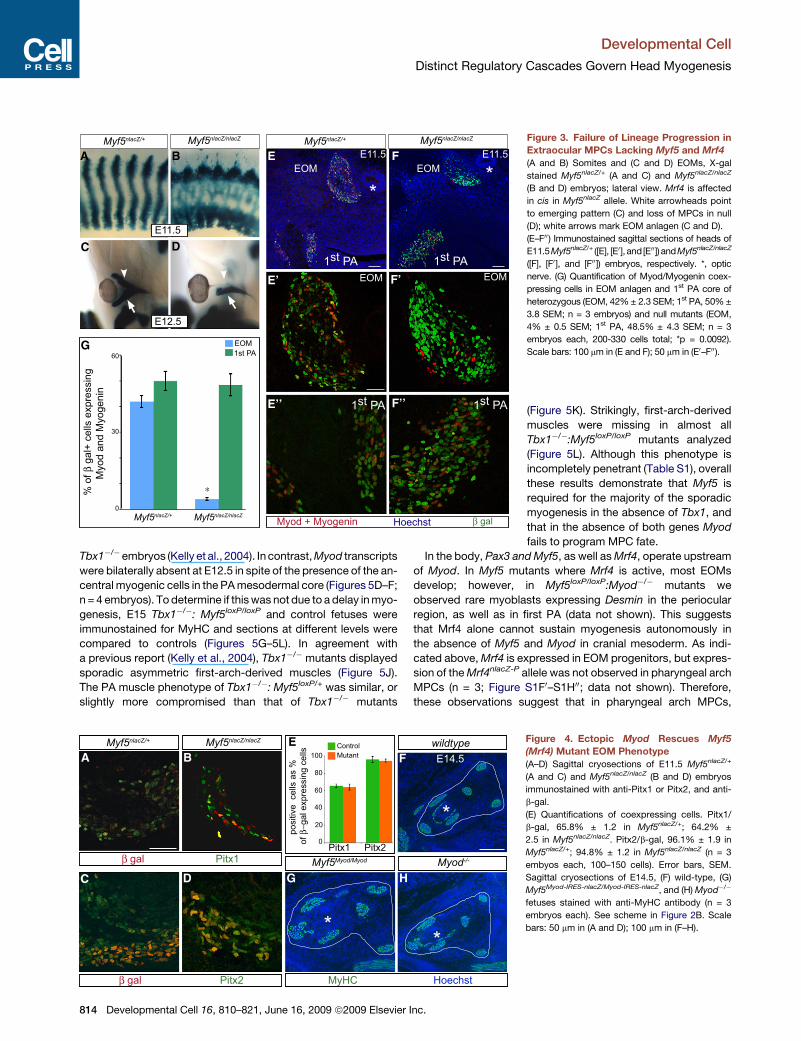
Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
Tbx1�/�embryos (Kelly et al., 2004). In contrast, Myod transcripts
were bilaterally absent at E12.5 in spite of the presence of the an-
central myogenic cells in the PA mesodermal core (Figures 5D–F;
n = 4 embryos). To determine if this was not due to a delay in myo-
genesis, E15 Tbx1�/�: Myf5loxP/loxP and control fetuses were
immunostained for MyHC and sections at different levels were
compared to controls (Figures 5G–5L). In agreement with
a previous report (Kelly et al., 2004), Tbx1�/� mutants displayed
sporadic asymmetric first-arch-derived muscles (Figure 5J).
The PA muscle phenotype of Tbx1�/�: Myf5loxP/+ was similar, or
slightly more compromised than that of Tbx1�/� mutants
A B
C D
E F
E’
E’’ F’’
F’
G
Figure 3. Failure of Lineage Progression in
Extraocular MPCs Lacking Myf5 and Mrf4
(A and B) Somites and (C and D) EOMs, X-gal
stained Myf5nlacZ/+ (A and C) and Myf5nlacZ/nlacZ
(B and D) embryos; lateral view. Mrf4 is affected
in cis in Myf5nlacZ allele. White arrowheads point
to emerging pattern (C) and loss of MPCs in null
(D); white arrows mark EOM anlagen (C and D).
(E–F00) Immunostained sagittal sections of heads of
E11.5Myf5nlacZ/+ ([E], [E0], and[E00]) andMyf5nlacZ/nlacZ
([F], [F0 ], and [F00]) embryos, respectively. *, optic
nerve. (G) Quantification of Myod/Myogenin coex-
pressing cells in EOM anlagen and 1st PA core of
heterozygous (EOM, 42% ± 2.3 SEM; 1st PA, 50% ±
3.8 SEM; n = 3 embryos) and null mutants (EOM,
4% ± 0.5 SEM; 1st PA, 48.5% ± 4.3 SEM; n = 3
embryos each, 200-330 cells total; *p = 0.0092).
Scale bars: 100 mm in (E and F); 50 mm in (E0–F00).
(Figure 5K). Strikingly, first-arch-derived
muscles were missing in almost all
Tbx1�/�:Myf5loxP/loxP mutants analyzed
(Figure 5L). Although this phenotype is
incompletely penetrant (Table S1), overall
these results demonstrate that Myf5 is
required for the majority of the sporadic
myogenesis in the absence of Tbx1, and
that in the absence of both genes Myod
fails to program MPC fate.
In the body, Pax3 and Myf5, as well as Mrf4, operate upstream
of Myod. In Myf5 mutants where Mrf4 is active, most EOMs
develop; however, in Myf5loxP/loxP:Myod�/� mutants we
observed rare myoblasts expressing Desmin in the periocular
region, as well as in first PA (data not shown). This suggests
that Mrf4 alone cannot sustain myogenesis autonomously in
the absence of Myf5 and Myod in cranial mesoderm. As indi-
cated above, Mrf4 is expressed in EOM progenitors, but expres-
sion of the Mrf4nlacZ-P allele was not observed in pharyngeal arch
MPCs (n = 3; Figure S1F0–S1H00; data not shown). Therefore,
these observations suggest that in pharyngeal arch MPCs,
*
Myod-/-
wildtype
Pitx1β gal
Pitx2β gal
A
C D
EF
G
B
H
E14.5
**
MyHC Hoechst
posi
tive
cel
ls a
s %
of β
−gal
exp
ress
ing
cells
Pitx1 Pitx20
100
20
40
60
80
ControlMutant
Myf5nlacZ/+
Myf5nlacZ/nlacZ
Myf5Myod/Myod
Figure 4. Ectopic Myod Rescues Myf5
(Mrf4) Mutant EOM Phenotype
(A–D) Sagittal cryosections of E11.5 Myf5nlacZ/+
(A and C) and Myf5nlacZ/nlacZ (B and D) embryos
immunostained with anti-Pitx1 or Pitx2, and anti-
b-gal.
(E) Quantifications of coexpressing cells. Pitx1/
b-gal, 65.8% ± 1.2 in Myf5nlacZ/+; 64.2% ±
2.5 in Myf5nlacZ/nlacZ. Pitx2/b-gal, 96.1% ± 1.9 in
Myf5nlacZ/+; 94.8% ± 1.2 in Myf5nlacZ/nlacZ (n = 3
embyos each, 100–150 cells). Error bars, SEM.
Sagittal cryosections of E14.5, (F) wild-type, (G)
Myf5Myod-IRES-nlacZ/Myod-IRES-nlacZ, and (H) Myod�/�
fetuses stained with anti-MyHC antibody (n = 3
embryos each). See scheme in Figure 2B. Scale
bars: 50 mm in (A and D); 100 mm in (F–H).
814 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc.
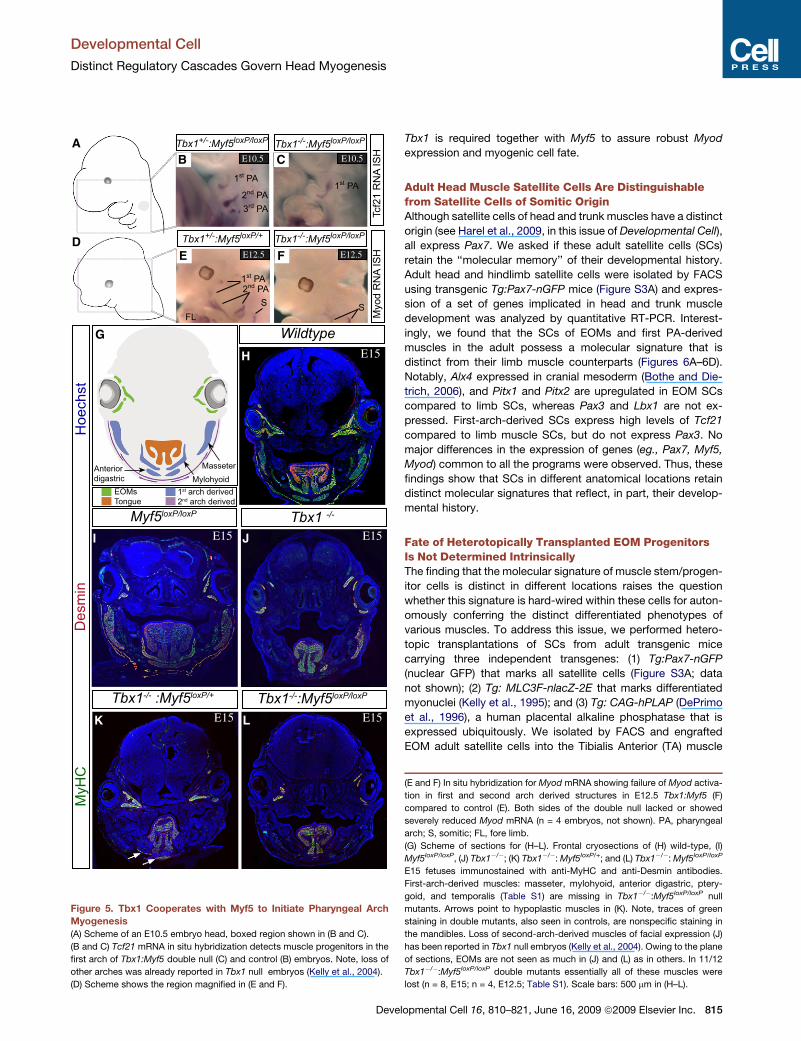
Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
1st PA
2nd PA1st PA
E F
Myo
d R
NA
ISH
E10.5 E10.5
E12.5 E12.5
3rd PA
Tbx1+/-
:Myf5loxP/loxP
Tbx1-/-
:Myf5loxP/loxP
Tbx1-/-
:Myf5loxP/loxP
Tbx1+/-
:Myf5loxP/+
Tbx1-/-:Myf5
loxP/loxPTbx1
-/- :Myf5
loxP/+
Tbx1 -/-
E15 E15
E15
E15
Wildtype
H
I J
Des
min
MyH
C
Hoe
chst
E15K L
Myf5loxP/loxP
CB
G
Tcf2
1 R
NA
ISH
SS
EOMsTongue
1st arch derived2nd arch derived
A
D
MasseterMylohyoid
Anterior digastric
1st PA2nd PA
FL
Figure 5. Tbx1 Cooperates with Myf5 to Initiate Pharyngeal Arch
Myogenesis
(A) Scheme of an E10.5 embryo head, boxed region shown in (B and C).
(B and C) Tcf21 mRNA in situ hybridization detects muscle progenitors in the
first arch of Tbx1:Myf5 double null (C) and control (B) embryos. Note, loss of
other arches was already reported in Tbx1 null embryos (Kelly et al., 2004).
(D) Scheme shows the region magnified in (E and F).
Deve
Tbx1 is required together with Myf5 to assure robust Myod
expression and myogenic cell fate.
Adult Head Muscle Satellite Cells Are Distinguishablefrom Satellite Cells of Somitic OriginAlthough satellite cells of head and trunk muscles have a distinct
origin (see Harel et al., 2009, in this issue of Developmental Cell),
all express Pax7. We asked if these adult satellite cells (SCs)
retain the ‘‘molecular memory’’ of their developmental history.
Adult head and hindlimb satellite cells were isolated by FACS
using transgenic Tg:Pax7-nGFP mice (Figure S3A) and expres-
sion of a set of genes implicated in head and trunk muscle
development was analyzed by quantitative RT-PCR. Interest-
ingly, we found that the SCs of EOMs and first PA-derived
muscles in the adult possess a molecular signature that is
distinct from their limb muscle counterparts (Figures 6A–6D).
Notably, Alx4 expressed in cranial mesoderm (Bothe and Die-
trich, 2006), and Pitx1 and Pitx2 are upregulated in EOM SCs
compared to limb SCs, whereas Pax3 and Lbx1 are not ex-
pressed. First-arch-derived SCs express high levels of Tcf21
compared to limb muscle SCs, but do not express Pax3. No
major differences in the expression of genes (eg., Pax7, Myf5,
Myod) common to all the programs were observed. Thus, these
findings show that SCs in different anatomical locations retain
distinct molecular signatures that reflect, in part, their develop-
mental history.
Fate of Heterotopically Transplanted EOM ProgenitorsIs Not Determined IntrinsicallyThe finding that the molecular signature of muscle stem/progen-
itor cells is distinct in different locations raises the question
whether this signature is hard-wired within these cells for auton-
omously conferring the distinct differentiated phenotypes of
various muscles. To address this issue, we performed hetero-
topic transplantations of SCs from adult transgenic mice
carrying three independent transgenes: (1) Tg:Pax7-nGFP
(nuclear GFP) that marks all satellite cells (Figure S3A; data
not shown); (2) Tg: MLC3F-nlacZ-2E that marks differentiated
myonuclei (Kelly et al., 1995); and (3) Tg: CAG-hPLAP (DePrimo
et al., 1996), a human placental alkaline phosphatase that is
expressed ubiquitously. We isolated by FACS and engrafted
EOM adult satellite cells into the Tibialis Anterior (TA) muscle
(E and F) In situ hybridization for Myod mRNA showing failure of Myod activa-
tion in first and second arch derived structures in E12.5 Tbx1:Myf5 (F)
compared to control (E). Both sides of the double null lacked or showed
severely reduced Myod mRNA (n = 4 embryos, not shown). PA, pharyngeal
arch; S, somitic; FL, fore limb.
(G) Scheme of sections for (H–L). Frontal cryosections of (H) wild-type, (I)
Myf5loxP/loxP, (J) Tbx1�/�; (K) Tbx1�/�: Myf5loxP/+; and (L) Tbx1�/�: Myf5loxP/loxP
E15 fetuses immunostained with anti-MyHC and anti-Desmin antibodies.
First-arch-derived muscles: masseter, mylohyoid, anterior digastric, ptery-
goid, and temporalis (Table S1) are missing in Tbx1�/�:Myf5loxP/loxP null
mutants. Arrows point to hypoplastic muscles in (K). Note, traces of green
staining in double mutants, also seen in controls, are nonspecific staining in
the mandibles. Loss of second-arch-derived muscles of facial expression (J)
has been reported in Tbx1 null embryos (Kelly et al., 2004). Owing to the plane
of sections, EOMs are not seen as much in (J) and (L) as in others. In 11/12
Tbx1�/�:Myf5loxP/loxP double mutants essentially all of these muscles were
lost (n = 8, E15; n = 4, E12.5; Table S1). Scale bars: 500 mm in (H–L).
lopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 815
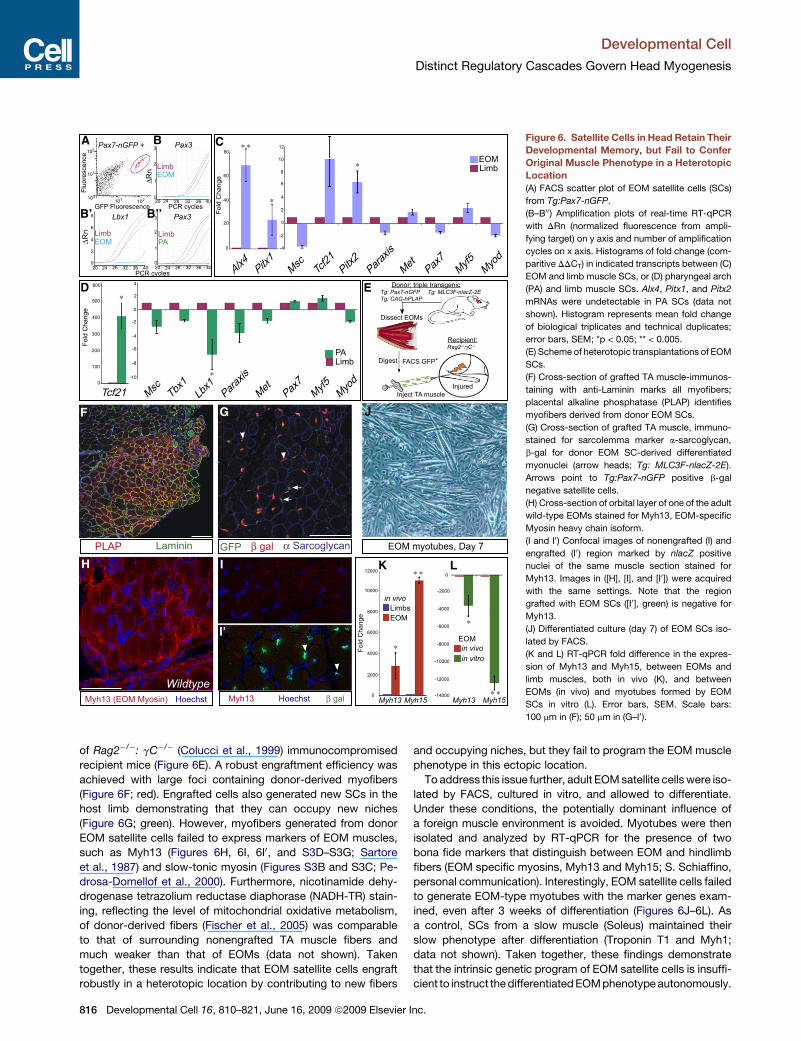
Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
A C
D
F G
H
I’
I
B’ B’’
B
J
K L
E
Figure 6. Satellite Cells in Head Retain Their
Developmental Memory, but Fail to Confer
Original Muscle Phenotype in a Heterotopic
Location
(A) FACS scatter plot of EOM satellite cells (SCs)
from Tg:Pax7-nGFP.
(B–B00) Amplification plots of real-time RT-qPCR
with DRn (normalized fluorescence from ampli-
fying target) on y axis and number of amplification
cycles on x axis. Histograms of fold change (com-
paritive DDCT) in indicated transcripts between (C)
EOM and limb muscle SCs, or (D) pharyngeal arch
(PA) and limb muscle SCs. Alx4, Pitx1, and Pitx2
mRNAs were undetectable in PA SCs (data not
shown). Histogram represents mean fold change
of biological triplicates and technical duplicates;
error bars, SEM; *p < 0.05; ** < 0.005.
(E) Scheme of heterotopic transplantations of EOM
SCs.
(F) Cross-section of grafted TA muscle-immunos-
taining with anti-Laminin marks all myofibers;
placental alkaline phosphatase (PLAP) identifies
myofibers derived from donor EOM SCs.
(G) Cross-section of grafted TA muscle, immuno-
stained for sarcolemma marker a-sarcoglycan,
b-gal for donor EOM SC-derived differentiated
myonuclei (arrow heads; Tg: MLC3F-nlacZ-2E).
Arrows point to Tg:Pax7-nGFP positive b-gal
negative satellite cells.
(H) Cross-section of orbital layer of one of the adult
wild-type EOMs stained for Myh13, EOM-specific
Myosin heavy chain isoform.
(I and I0) Confocal images of nonengrafted (I) and
engrafted (I0 ) region marked by nlacZ positive
nuclei of the same muscle section stained for
Myh13. Images in ([H], [I], and [I0]) were acquired
with the same settings. Note that the region
grafted with EOM SCs ([I0], green) is negative for
Myh13.
(J) Differentiated culture (day 7) of EOM SCs iso-
lated by FACS.
(K and L) RT-qPCR fold difference in the expres-
sion of Myh13 and Myh15, between EOMs and
limb muscles, both in vivo (K), and between
EOMs (in vivo) and myotubes formed by EOM
SCs in vitro (L). Error bars, SEM. Scale bars:
100 mm in (F); 50 mm in (G–I0).
of Rag2�/�: gC�/� (Colucci et al., 1999) immunocompromised
recipient mice (Figure 6E). A robust engraftment efficiency was
achieved with large foci containing donor-derived myofibers
(Figure 6F; red). Engrafted cells also generated new SCs in the
host limb demonstrating that they can occupy new niches
(Figure 6G; green). However, myofibers generated from donor
EOM satellite cells failed to express markers of EOM muscles,
such as Myh13 (Figures 6H, 6I, 6I0, and S3D–S3G; Sartore
et al., 1987) and slow-tonic myosin (Figures S3B and S3C; Pe-
drosa-Domellof et al., 2000). Furthermore, nicotinamide dehy-
drogenase tetrazolium reductase diaphorase (NADH-TR) stain-
ing, reflecting the level of mitochondrial oxidative metabolism,
of donor-derived fibers (Fischer et al., 2005) was comparable
to that of surrounding nonengrafted TA muscle fibers and
much weaker than that of EOMs (data not shown). Taken
together, these results indicate that EOM satellite cells engraft
robustly in a heterotopic location by contributing to new fibers
816 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier
and occupying niches, but they fail to program the EOM muscle
phenotype in this ectopic location.
To address this issue further, adult EOM satellite cells were iso-
lated by FACS, cultured in vitro, and allowed to differentiate.
Under these conditions, the potentially dominant influence of
a foreign muscle environment is avoided. Myotubes were then
isolated and analyzed by RT-qPCR for the presence of two
bona fide markers that distinguish between EOM and hindlimb
fibers (EOM specific myosins, Myh13 and Myh15; S. Schiaffino,
personal communication). Interestingly, EOM satellite cells failed
to generate EOM-type myotubes with the marker genes exam-
ined, even after 3 weeks of differentiation (Figures 6J–6L). As
a control, SCs from a slow muscle (Soleus) maintained their
slow phenotype after differentiation (Troponin T1 and Myh1;
data not shown). Taken together, these findings demonstrate
that the intrinsic genetic program of EOM satellite cells is insuffi-
cient to instruct the differentiated EOM phenotype autonomously.
Inc.

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
DISCUSSION
Skeletal muscles have evolved the striking ability to adopt
selected elements of a core regulatory network in concert with
a complementary genetic pathway to assure self-renewal and
the acquisition of cell fate in different anatomical locations. These
pathways have been studied extensively in the body, yet the hier-
archical relationships of key regulators in cranial-derived MPCs,
and their relevance to the core MRF network, remained unre-
solved. We show that MPCs in the two major groups of head
muscles, the extraocular muscles and those in the pharyngeal
arches, have adopted surprisingly different modes of regulation
which are distinct from that governing MPC fate in the body.
Complementary studies using lineage tracing strategies show
that cranial satellite cells have a distinct origin from those in the
body (Harel et al., 2009, this issue). We show that skeletal muscle
stem cells in the adult retain their ontological molecular signature,
but these stem cells cannot autonomously assure the specific
muscle phenotypes in an ectopic location. These findings
provide a framework for investigating myopathies that are
restricted to subsets of muscles.
Initiation of EOM Cell Fate Is Governed by Myf5 or Mrf4
The myotome in the mouse, which is formed from the input of
MPCs that are born principally in the dorsal and ventral dermo-
myotome (DM) lips (Cossu et al., 1996; Tajbakhsh et al., 1996),
is a single undivided entity that patterns, splits, and displaces
into different muscle masses as this structure enlarges. Pharyn-
geal arch-derived and limb muscles follow a similar ontology.
Previous reports suggested that some EOM muscles in the chick
arise as independent anlage (Noden et al., 1999; von Scheven
et al., 2006). Observations with Myf5nlacZ animals suggest that
individual EOMs arise from a single, apparently undivided anlage
in mice, although this requires further investigation. We propose
that the EOM subsets are then patterned by local migrations of
progenitors away from this source to their more distal positions.
The mechanisms guiding the patterning process in the EOMs
remain to be explored.
Mrf4 drives the first embryonic wave of trunk myogenesis in
Myf5: Myod mutants (Kassar-Duchossoy et al., 2004), but its
role in head muscle development had not been elucidated. We
demonstrate that Mrf4 and Myf5 determine EOM progenitor
cell fate to form this distinct subset of head muscles of cranial
mesoderm origin. Apparently, both the prechordal and paraxial
head mesoderm contribute to EOMs in avians (Couly et al.,
1992; Noden and Francis-West, 2006; Wachtler et al., 1984).
Although data are lacking in the mouse concerning this point,
in either case, there is an obligate requirement for Myf5 or Mrf4
for initiating EOM fate.
Myod activation is mediated by Pax3, either directly or indi-
rectly, to initiate MPC fate in somites of Myf5:Mrf4 double
mutants. Interestingly, we show here that in the same mutants
neither Myod nor Myogenin rescue EOM development. Pitx2
acts upstream of MRFs in cranial MPCs and Pitx2 null embryos
lack EOMs (Gage et al., 1999; Kitamura et al., 1999); however,
the precise role of Pitx2 in inducing MRFs remained to be ad-
dressed. We show that Pitx2 fails to bring about the myogenic
rescue by Myod or Myogenin in EOM progenitors in the absence
of Myf5 and Mrf4, although some residual fibers were present in
Deve
these double mutants. Thus, Myod is epistatic to Myf5 and Mrf4
in EOM progenitors. We propose that unlike other skeletal
muscle progenitors (see below), EOM progenitors lack a Myf5/
Mrf4-independent pathway to activate myogenesis via Myod
(Figure 7).
Survival of Muscle Progenitors Is Coupledto CommitmentThe rapid apoptotic death of EOM progenitors in the absence of
Myf5 and Mrf4 is unusual among all MPCs in the embryo. In
Myf5:Mrf4:Myod triple mutants, some somitic MPCs also
undergo apoptosis, but several days after their birth (Kablar
et al., 2003; S.T., unpublished data). In both cases, MPCs fail
to commit and this may be linked to their death. Interestingly,
expression of Myod using a Myf5Myod allele, in the absence of
Myf5 and Mrf4, prevents the loss of EOM progenitors and
rescues EOMs. Thus, any one determination factor is sufficient
to ensure MPC survival. Other factors acting upstream may
also maintain MPC viability in the absence of MRFs. Interestingly
however, Myod plays a proapoptotic role in committed precur-
sors by inducing apoptosis in MPCs if they fail to differentiate,
and this requires p21/Cip1 (Peschiaroli et al., 2002). Additionally,
Myod null myoblasts are more viable than wild-type after trans-
plantion (Asakura et al., 2007). These opposing functions of the
MRFs need to be investigated in a developmental context.
Distinct Regulation of Pharyngeal Arch MPC Fateand Myogenic Cell PopulationsThe report that Pax3 activates somitic myogenesis through
a parallel program independent of Myf5 and Mrf4 (Tajbakhsh
et al., 1997) raised the possibility that another gene may do so
in cranial MPCs. We show that Pax7 in the head does not play
the early role of Pax3 in the body. The bilateral loss of first PA
muscles in Tbx1�/�:Myf5loxP/loxP double mutants suggests that
these genes are required together to initiate myogenic cell fate,
although Tbx1 may act genetically upstream of Myf5 as well
(Kelly et al., 2004). Apart from mesoderm, Tbx1 is expressed in
PA endoderm and ectoderm (Chapman et al., 1996) and hence
the analysis of the non-cell-autonomous role of this gene on
MPCs (Dastjerdi et al., 2007; Grifone and Kelly, 2007) needs to
be extended. Furthermore, unlike Pitx2 mutants, 2nd and 3rd
pharyngeal arches themselves are virtually absent in Tbx1 null
mutants, therefore the genetic hierarchy in the MPCs in these
arches remains an open question.
As reported for Pax3:Myf5(Mrf4) (Tajbakhsh et al., 1997) and
Msc:Tcf21 (MyoR:Capsulin, respectively) null mutants (Lu
et al., 2002), we note an incomplete penetrance and residual
myogenesis in some Tbx1�/�:Myf5loxP/loxP fetuses. This could
be due to compensation by Myod, another factor, or the genetic
background. Pitx2, expressed in the surface ectoderm and
mesodermal core of the first arch, is necessary for the specifica-
tion of the first arch itself, as the entire arch is severely reduced
in Pitx2 null mice (Dong et al., 2006; Shih et al., 2007). Elimi-
nating Pitx2 expression specifically in the mesoderm results in
a severe reduction of Myogenin expression (Dong et al.,
2006). Furthermore, Pitx2 and Tbx1 are known to regulate
each other (Nowotschin et al., 2006; Shih et al., 2007) and could
potentially act in parallel (Dong et al., 2006; Lamolet et al.,
2001). Therefore, we speculate that Pitx2 is a likely candidate
lopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 817

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
for activating myogenesis in a smaller pool of arch MPCs in
Tbx1:Myf5 null mutants, resulting in some sporadic myogenesis
in the first arch. Taken together, we conclude that the combined
activities of Tbx1 and Myf5 are crucial for PA myogenesis
(Figure 7).
Our genetic analyses reveal that in EOM and PA MPCs, as in
somitic progenitors, Myod acts epistatically to Myf5. However,
on a cellular level, a Myf5-independent population was reported,
as identified by the failure of Myf5Cre to eliminate the entire MPC
pool after the conditional activation of a diptheria toxin gene
(Gensch et al., 2008; Haldar et al., 2008). Our data indicate that
a Myf5-independent population, if present in EOM, must be
largely dependent on Mrf4, or it is insignificant in EOMs. In the
first arch, we presume that this population would be largely
dependent on Tbx1, either in pharyngeal endoderm or meso-
derm, for Myod activation. We propose that the epistatic rela-
tionship between Myf5 and Myod is common for the majority
of muscle progenitors in the embryo. How cell nonautonomous
events could account for the rescue of myogenesis in the
absence of Myf5 and Mrf4 requires further investigation to deci-
pher the upstream decisions that decide which of the core MRFs
is selectively employed. Finally, we would speculate that two to
four genetic layers separate mesoderm formation from muscle
stem cell specification.
Figure 7. Core and Complementary Genetic
Networks Regulating EOM and PA Muscle
Fate
EOMs have dispensed with a complementary
regulatory pathway, therefore Myod expression
is not rescued in absence of Myf5 and Mrf4, and
EOMs do not form. In PA muscles, Tbx1 cooper-
ates with Myf5; in their combined absence, Myod
is not activated, and PA muscles are essentially
all missing. In the body (Kassar-Duchossoy et al.,
2004) and EOMs, Mrf4 determines embryonic
but not fetal MPC fate. Pitx2 may cooperate with
Tbx1 at this regulatory step in the PA. Pax3 acts
as the complementary pathway for body myogen-
esis to rescue Myod expression (Tajbakhsh et al.,
1997). With this model, the genetic distance
between mesoderm and muscle stem cells can
be envisioned. We speculate that two to four
layers separate these landmarks.
Adult MPC Molecular Signature andStem Cell Behavior in HeterotopicTransplantationsPax7 expressing stem/progenitor cells
appear in developing muscles throughout
the embryo around E11.5, after the estab-
lishment of a muscle anlage. A develop-
mental continuity exists between this
pool and adult SCs which are also depen-
dent on Pax7 for survival (Sambasivan
and Tajbakhsh, 2007). Hence, we were
surprised to observe that EOM and first-
arch-derived SCs bear distinct molecular
signatures compared to those in the
limbs. Remarkably, this distinct identity
partially reflects their developmental ontology. It would be inter-
esting to determine to what extent this occurs with stem cells in
other tissues.
Observations that skeletal muscles have specific functional
characteristics and differences in their ability to regenerate rai-
ses the question to what extent these phenotypes are conferred
by the stem/progenitor populations (Pavlath et al., 1998; Zammit
et al., 2006). Although heterotopic transplantation of temporalis
muscle of cat to the hind limb resulted in the regenerate
acquiring the donor jaw muscle identity (Hoy and Hughes,
1988), the cell autonomous role of SCs in conferring the donor
muscle phenotype remained unaddressed. Here we demon-
strate that SCs isolated directly from EOMs and transplanted
into a hind limb muscle engrafted robustly; however, EOM-
specific myosin expression was not observed in this ectopic
environment. In this context, we note that there are contradictory
reports regarding the intrinsic cue of limb muscle progenitors to
generate slow or fast fiber types. Whereas mouse fetal or post-
natal prequiescent myoblasts are not commited to form
a specific fiber type (Hughes and Blau, 1992; Robson and
Hughes, 1999), avian embryonic myoblasts appeared to be
restricted to slow or fast phenotypes (Nikovits et al., 2001).
Intriguingly, we report that the EOM SCs when differentiated
in vitro fail to express EOM-specific myosin heavy chain
818 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc.

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
markers. Thus, we conclude that the EOM phenotype is not
dictated by the intrinsic cues of the stem/progenitors alone.
Trunk muscles function chiefly in locomotion and posture,
whereas the head muscles have varied functions such as eye
movements, feeding, and facial expression. These varied
requirements may have determined their distinct phenotypic
properties. We speculate that these properties reflect a develop-
mental underpinning. The distinct set of phenotypic characteris-
tics, in turn, is likely to underlie the differential involvement of
head muscles in a variety of myopathic diseases. Thus, under-
standing the differences in developmental programs and their
influence on the adult muscle phenotype is paramount to
building a framework to address the etiology of distinct
myopathic diseases.
EXPERIMENTAL PROCEDURES
Animals
Animals were handled as per European Community guidelines. Mouse lines
with mutant alleles, Myf5loxP, Myf5GFP-P, Myf5nlacZ, and Mrf4nlacZ:Myf5loxP,
were described previously (Kassar-Duchossoy et al., 2004; Tajbakhsh et al.,
1996). Myod�/� (Rudnicki et al., 1993); Mrf4�/� (kindly provided by E. Olson;
Zhang et al., 1995); Myf5Myod-IRES-nlacZ/Myod-IRES-nlacZ (provided by D. Gomes,
J. Perreau, G.D., D. Rocancourt, M. Buckingham, and S.T.); Tg: CAG-hPLAP
(courtesy of D. Daegelen; DePrimo et al., 1996); Tg: MLC3F-nlacZ-2E (Kelly
et al., 1995). For transplantations, Rag2�/�: gC�/� mice (Colucci et al., 1999)
were obtained from the Pasteur animal facilities. Mice carrying the Tbx1tm1pa
allele (referred to as Tbx1�) were genotyped as described; (Jerome and
Papaioannou, 2001). For generation of Tg: Pax7-nGFP line and Pax7nlacZ lines,
see the Supplemental Experimental Procedures. Mouse mutants were
genotyped and interbred as described (Kassar-Duchossoy et al., 2004).
In Situ RNA Hybridization, Immunofluorescence, and Imaging
ISH was performed as described (Kassar-Duchossoy et al., 2004; Tajbakhsh
et al., 1997) with Myod or Mrf4 riboprobes. Comparisons were done between
age-matched littermates hybridized under the same conditions. Images were
acquired with a Zeiss Axiocam numerical camera and Axiovision software.
Immunofluorescence was performed as described (Kassar-Duchossoy et al.,
2004). Embryos were fixed for 1.5 hr in 4% paraformaldehyde and embedded
in 7% gelatin and 15% sucrose embedded for cryosectioning (12–16 mm).
Dissected muscles were either 2% paraformaldehyde + 0.1% Tween20 fixed
for 1.5 hr or unfixed and cryofrozen in isopentane bath (�30�C) for sectioning
(12 mm). Sagittal sections were ideal for scoring all EOMs at E15 and frontal
sections were chosen for assessing pharyngeal arch-derived muscles. Images
were acquired using an Leica SPE confocal and LAS software or Zeiss Axio-
plan and Axiovision software. Optical sections (1–1.5 mm) were reconstructed
using ImageJ (NIH). Images were assembled using Adobe Photoshop and
Adobe InDesign. Antibodies used in this study include Myosin heavy chain
(rabbit, kindly provided by G. Cossu; 1/750), cleaved activated Caspase3
(mouse, Cell Signaling; 1/100 ; marker of apoptosis; Cohen, 1997), b-galacto-
sidase (rabbit, kindly provided by O. Puijalon; 1/200), Myod (Dako; M3512; 1/
50), Myogenin (DSHB; F5D; 1/10), Myh13 (DSHB; 4A6; 1/5; Lucas et al., 1995),
slow tonic myosin heavy chain (DSHB; S46; 1/10), placental alkaline phospha-
tase (GeneTex, Inc.; 73609; 1/400), Laminin (Sigma; 9393; 1/1000) GFP (Ab-
cam; 13970; 1/300), Pitx1, Pitx2 (polyclonal; kindly provided by J. Drouin),
and cyclin D1 (Thermo Scientific; SP4; 1/250 ; marker of cycling cells; Stacey,
2003). Tissue was not fixed for immunostaining with 4A6 and S46 antibodies.
Transplantation Experiments
Mice were anesthetized with 0.5% Imalgene/2% Rompun. Satellite cells iso-
lated by FACS, were injected into the Tibialis anterior muscle (3,000–10,000
cells) that had been subjected to three consecutive cycles of freezing-thawing
by applying a cryocooled metallic rod (Gayraud-Morel et al., 2007) 48 hr before
transplantation. Grafted muscles were collected 3 weeks posttransplantation
for analysis.
Dev
Quantitative RT-PCR
Total RNA was extracted from cells isolated by FACS using the Qiagen
RNAeasy Micropurification Kit. From 400–600 ng of DNase-treated (Roche)
RNA, cDNA was prepared by random-primed reverse transcription (Super-
Script II, Invitrogen) and real-time PCR done using powerSYBR Green
Universal Mix or Taqman universal Master Mix (ABI Prism 7700 and StepOne-
Plus, Perkin-Elmer Applied Biosystems). GAPDH transcript levels were used
for the normalizations of each target (= DCT). At least three biological repli-
cates were used for each condition (D(DCT) method) (Schmittgen and Livak,
2008). Custom primers were designed using the Primer3Plus online software
(http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). Serial di-
lutions of total cDNA were used to calculate the amplification efficiency of
each primer set according to the equation: E = 10 � 1/slope. Primer dissocia-
tion experiments were performed to assure that no primer dimers or false am-
plicons would interfere with the results.
SUPPLEMENTAL DATA
Supplemental Data include three figures, two tables, and Supplemental Exper-
imental Procedures and can be found with this article online at http://www.cell.
com/developmental-cell/supplemental/S1534-5807(09)00210-X.
ACKNOWLEDGMENTS
We thank members of the lab and R. Grifone for helpful discussions; A. Jory for
help with establishing the Tbx1 colony; J. Drouin and P. Maire for reagents; M.
Nguyen-De Bernon for assistance with FACS, Pasteur Imaging platform
(PFID); F. Langa Vives and the Pasteur Transgenic Platform; D. Rocancourt,
M. Buckingham, E. Olson, S. Vincent, and D. Daegelen for mice; and S. Olivier
for artwork. Funding was granted by Institut Pasteur, AFM, FRM, ARC,
MyoRes (EU Framework 6), and EuroSyStem (EU Framework 7). R.K. is funded
by the AFM.
Received: December 21, 2008
Revised: April 9, 2009
Accepted: May 15, 2009
Published: June 15, 2009
REFERENCES
Asakura, A., Hirai, H., Kablar, B., Morita, S., Ishibashi, J., Piras, B.A., Christ,
A.J., Verma, M., Vineretsky, K.A., and Rudnicki, M.A. (2007). Increased survival
of muscle stem cells lacking the MyoD gene after transplantation into regener-
ating skeletal muscle. Proc. Natl. Acad. Sci. USA 104, 16552–16557.
Bothe, I., and Dietrich, S. (2006). The molecular setup of the avian head meso-
derm and its implication for craniofacial myogenesis. Dev. Dyn. 235, 2845–
2860.
Braun, T., and Arnold, H.H. (1995). Inactivation of Myf-6 and Myf-5 genes in
mice leads to alterations in skeletal muscle development. EMBO J. 14,
1176–1186.
Chapman, D.L., Garvey, N., Hancock, S., Alexiou, M., Agulnik, S.I., Gibson-
Brown, J.J., Cebra-Thomas, J., Bollag, R.J., Silver, L.M., and Papaioannou,
V.E. (1996). Expression of the T-box family genes, Tbx1-Tbx5, during early
mouse development. Dev. Dyn. 206, 379–390.
Cohen, G.M. (1997). Caspases: the executioners of apoptosis. Biochem. J.
326, 1–16.
Colucci, F., Soudais, C., Rosmaraki, E., Vanes, L., Tybulewicz, V.L., and Di
Santo, J.P. (1999). Dissecting NK cell development using a novel alymphoid
mouse model: investigating the role of the c-abl proto-oncogene in murine
NK cell differentiation. J. Immunol. 162, 2761–2765.
Cossu, G., Tajbakhsh, S., and Buckingham, M. (1996). How is myogenesis
initiated in the embryo? Trends Genet. 12, 218–223.
Couly, G.F., Coltey, P.M., and Douarin, N.M.L. (1992). The developmental fate
of the cephalic mesoderm in quail-chick chimeras. Development 114, 1–15.
Dastjerdi, A., Robson, L., Walker, R., Hadley, J., Zhang, Z., Rodriguez-Nieden-
fuhr, M., Ataliotis, P., Baldini, A., Scambler, P., and Francis-West, P. (2007).
elopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 819

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
Tbx1 regulation of myogenic differentiation in the limb and cranial mesoderm.
Dev. Dyn. 236, 353–363.
DePrimo, S.E., Stambrook, P.J., and Stringer, J.R. (1996). Human placental
alkaline phosphatase as a histochemical marker of gene expression in trans-
genic mice. Transgenic Res. 5, 459–466.
Dong, F., Sun, X., Liu, W., Ai, D., Klysik, E., Lu, M.F., Hadley, J., Antoni, L.,
Chen, L., Baldini, A., et al. (2006). Pitx2 promotes development of splanchnic
mesoderm-derived branchiomeric muscle. Development 133, 4891–4899.
Emery, A.E. (2002). The muscular dystrophies. Lancet 359, 687–695.
Fischer, M.D., Budak, M.T., Bakay, M., Gorospe, J.R., Kjellgren, D., Pedrosa-
Domellof, F., Hoffman, E.P., and Khurana, T.S. (2005). Definition of the unique
human extraocular muscle allotype by expression profiling. Physiol. Genomics
22, 283–291.
Gage, P.J., Suh, H., and Camper, S.A. (1999). Dosage requirement of Pitx2 for
development of multiple organs. Development 126, 4643–4651.
Gayraud-Morel, B., Chretien, F., Flamant, P., Gomes, D., Zammit, P.S., and
Tajbakhsh, S. (2007). A role for the myogenic determination gene Myf5 in adult
regenerative myogenesis. Dev. Biol. 312, 13–28.
Gensch, N., Borchardt, T., Schneider, A., Riethmacher, D., and Braun, T.
(2008). Different autonomous myogenic cell populations revealed by ablation
of Myf5-expressing cells during mouse embryogenesis. Development 135,
1597–1604.
Grifone, R., and Kelly, R.G. (2007). Heartening news for head muscle develop-
ment. Trends Genet. 23, 365–369.
Grifone, R., Demignon, J., Houbron, C., Souil, E., Niro, C., Seller, M.J.,
Hamard, G., and Maire, P. (2005). Six1 and Six4 homeoproteins are required
for Pax3 and Mrf expression during myogenesis in the mouse embryo. Devel-
opment 132, 2235–2249.
Grifone, R., Demignon, J., Giordani, J., Niro, C., Souil, E., Bertin, F., Laclef, C.,
Xu, P.X., and Maire, P. (2007). Eya1 and Eya2 proteins are required for hypaxial
somitic myogenesis in the mouse embryo. Dev. Biol. 302, 602–616.
Grifone, R., Jarry, T., Dandonneau, M., Grenier, J., Duprez, D., and Kelly, R.G.
(2008). Properties of branchiomeric and somite-derived muscle development
in Tbx1 mutant embryos. Dev. Dyn. 237, 3071–3078.
Hacker, A., and Guthrie, S. (1998). A distinct developmental programme for the
cranial paraxial mesoderm in the chick embryo. Development 125, 3461–3472.
Haldar, M., Karan, G., Tvrdik, P., and Capecchi, M.R. (2008). Two cell lineages,
myf5 and myf5-independent, participate in mouse skeletal myogenesis. Dev.
Cell 14, 437–445.
Harel, I., Nathan, E., Tirosh-Finkel, L., Zigdon, H., Guimaraes-Camboa, N.,
Evans, S.M., and Tzahor, E. (2009). Distinct origins and genetic programs of
head muscle satellite cells. Dev. Cell 16, this issue, 822–832.
Hasty, P., Bradley, A., Morris, J.H., Edmondson, D.G., Venuti, J.M., Olson,
E.N., and Klein, W.H. (1993). Muscle deficiency and neonatal death in mice
with a targeted mutation in the myogenin gene. Nature 364, 501–506.
Hoy, J.F.Y., and Hughes, S. (1988). Myogenic and neurogenic regulation of
myosin gene expression in cat jaw-closing muscle regenerating in fast and
slow limb muscle beds. J. Muscle Res. Cell Motil. 9, 59–72.
Horst, D., Ustanina, S., Sergi, C., Mikuz, G., Juergens, H., Braun, T., and Vor-
obyov, E. (2006). Comparative expression analysis of Pax3 and Pax7 during
mouse myogenesis. Int. J. Dev. Biol. 50, 47–54.
Hughes, S.M., and Blau, H.M. (1992). Muscle fiber pattern is independent of
cell lineage in postnatal rodent development. Cell 68, 659–671.
Jerome, L.A., and Papaioannou, V.E. (2001). DiGeorge syndrome phenotype in
mice mutant for the T-box gene, Tbx1. Nat. Genet. 27, 286–291.
Kablar, B., Krastel, K., Tajbakhsh, S., and Rudnicki, M.A. (2003). Myf5 and
MyoD activation define independent myogenic compartments during embry-
onic development. Dev. Biol. 258, 307–318.
Kassar-Duchossoy, L., Gayraud-Morel, B., Gomes, D., Rocancourt, D., Buck-
ingham, M., Shinin, V., and Tajbakhsh, S. (2004). Mrf4 determines skeletal
muscle identity in Myf5:Myod double-mutant mice. Nature 431, 466–471.
Kelly, R., Alonso, S., Tajbakhsh, S., Cossu, G., and Buckingham, M. (1995).
Myosin light chain 3F regulatory sequences confer regionalised cardiac and
820 Developmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier
skeletal muscle reporter gene expression in transgenic mice. J. Cell Biol.
129, 383–396.
Kelly, R.G., Jerome-Majewska, L.A., and Papaioannou, V.E. (2004). The
del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesis.
Hum. Mol. Genet. 13, 2829–2840.
Kitamura, K., Miura, H., Miyagawa-Tomita, S., Yanazawa, M., Katoh-Fukui, Y.,
Suzuki, R., Ohuchi, H., Suehiro, A., Motegi, Y., Nakahara, Y., et al. (1999).
Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart,
extra- and periocular mesoderm and right pulmonary isomerism. Development
126, 5749–5758.
Lamolet, B., Pulichino, A.M., Lamonerie, T., Gauthier, Y., Brue, T., Enjalbert, A.,
and Drouin, J. (2001). A pituitary cell-restricted T box factor, Tpit, activates
POMC transcription in cooperation with Pitx homeoproteins. Cell 104,
849–859.
L’Honore, A., Coulon, V., Marcil, A., Lebel, M., Lafrance-Vanasse, J., Gage, P.,
Camper, S., and Drouin, J. (2007). Sequential expression and redundancy of
Pitx2 and Pitx3 genes during muscle development. Dev. Biol. 307, 421–433.
Lu, J.R., Bassel-Duby, R., Hawkins, A., Chang, P., Valdez, R., Wu, H., Gan, L.,
Shelton, J.M., Richardson, J.A., and Olson, E.N. (2002). Control of facial
muscle development by MyoR and capsulin. Science 298, 2378–2381.
Lucas, C.A., Rughani, A., and Hoh, J.F. (1995). Expression of extraocular
myosin heavy chain in rabbit laryngeal muscle. J. Muscle Res. Cell Motil. 16,
368–378.
Mootoosamy, R.C., Dietrich, S., Gustafsson, M.K., Pan, H., Pinney, D.F., Liu,
Y., Lewandowski, A., Epstein, D.J., and Emerson, C.P., Jr. (2002). Distinct
regulatory cascades for head and trunk myogenesis. Development 129,
573–583.
Nabeshima, Y., Hanaoka, K., Hayasaka, M., Esumi, E., Li, S., Nonaka, I., and
Nabeshima, Y. (1993). Myogenin gene disruption results in perinatal lethality
because of severe muscle defect. Nature 364, 532–535.
Nathan, E., Monovich, A., Tirosh-Finkel, L., Harrelson, Z., Rousso, T., Rinon,
A., Harel, I., Evans, S.M., and Tzahor, E. (2008). The contribution of Islet1-ex-
pressing splanchnic mesoderm cells to distinct branchiomeric muscles
reveals significant heterogeneity in head muscle development. Development
135, 647–657.
Nikovits, W., Jr., Cann, G.M., Huang, R., Christ, B., and Stockdale, F.E. (2001).
Patterning of fast and slow fibers within embryonic muscles is established
independently of signals from the surrounding mesenchyme. Development
128, 2537–2544.
Noden, D.M., and Francis-West, P. (2006). The differentiation and morphogen-
esis of craniofacial muscles. Dev. Dyn. 235, 1194–1218.
Noden, D.M., Marcucio, R., Borycki, A.G., and Emerson, C.P., Jr. (1999).
Differentiation of avian craniofacial muscles: I. Patterns of early regulatory
gene expression and myosin heavy chain synthesis. Dev. Dyn. 216, 96–112.
Nowotschin, S., Liao, J., Gage, P.J., Epstein, J.A., Campione, M., and Morrow,
B.E. (2006). Tbx1 affects asymmetric cardiac morphogenesis by regulating
Pitx2 in the secondary heart field. Development 133, 1565–1573.
Olson, E.N., Arnold, H.-H., Rigby, P.W.J., and Wold, B.J. (1996). Know your
neighbors: three phenotypes in null mutants of the myogenic bHLH gene
MRF4. Cell 85, 1–4.
Pavlath, G.K., Thaloor, D., Rando, T.A., Cheong, M., English, A.W., and Zheng,
B. (1998). Heterogeneity among muscle precursor cells in adult skeletal
muscles with differing regenerative capacities. Dev. Dyn. 212, 495–508.
Pedrosa-Domellof, F., Holmgren, Y., Lucas, C.A., Hoh, J.F., and Thornell, L.E.
(2000). Human extraocular muscles: unique pattern of myosin heavy chain
expression during myotube formation. Invest. Ophthalmol. Vis. Sci. 41,
1608–1616.
Peschiaroli, A., Figliola, R., Coltella, L., Strom, A., Valentini, A., D’Agnano, I.,
and Maione, R. (2002). MyoD induces apoptosis in the absence of RB function
through a p21(WAF1)-dependent re-localization of cyclin/cdk complexes to
the nucleus. Oncogene 21, 8114–8127.
Porter, J.D. (2002). Extraocular muscle: cellular adaptations for a diverse func-
tional repertoire. Ann. N Y Acad. Sci. 956, 7–16.
Inc.

Developmental Cell
Distinct Regulatory Cascades Govern Head Myogenesis
Pownall, M.E., Gustafsson, M.K., and Emerson, C.P., Jr. (2002). Myogenic
regulatory factors and the specification of muscle progenitors in vertebrate
embryos. Annu. Rev. Cell Dev. Biol. 18, 747–783.
Relaix, F., Rocancourt, D., Mansouri, A., and Buckingham, M. (2005). A Pax3/
Pax7-dependent population of skeletal muscle progenitor cells. Nature 435,
948–953.
Robson, L.G., and Hughes, S.M. (1999). Local signals in the chick limb bud can
override myoblast lineage commitment: induction of slow myosin heavy chain
in fast myoblasts. Mech. Dev. 85, 59–71.
Rudnicki, M.A., Schneglesberg, P.N.J., Stead, R.H., Braun, T., Arnold, H.-H.,
and Jaenisch, R. (1993). MyoD or myf-5 is required for the formation of skeletal
muscle. Cell 75, 1351–1359.
Sambasivan, R., and Tajbakhsh, S. (2007). Skeletal muscle stem cell birth and
properties. Semin. Cell Dev. Biol. 18, 870–882.
Sartore, S., Mascarello, F., Rowlerson, A., Gorza, L., Ausoni, S., Vianello, M.,
and Schiaffino, S. (1987). Fibre types in extraocular muscles: a new myosin
isoform in the fast fibres. J. Muscle Res. Cell Motil. 8, 161–172.
Seale, P., Sabourin, L.A., Girgis-Gabardo, A., Mansourt, A., Gruss, P., and
Rudnicki, M.A. (2000). Pax7 is required for the specification of myogenic satel-
lite cells. Cell 102, 777–786.
Schmittgen, T.D., and Livak, K.J. (2008). Analyzing real-time PCR data by the
comparative C(T) method. Nat. Protoc. 3, 1101–1108.
Shih, H.P., Gross, M.K., and Kioussi, C. (2007). Cranial muscle defects of Pitx2
mutants result from specification defects in the first branchial arch. Proc. Natl.
Acad. Sci. USA 104, 5907–5912.
Deve
Stacey, D.W. (2003). Cyclin D1 serves as a cell cycle regulatory switch in
actively proliferating cells. Curr. Opin. Cell Biol. 15, 158–163.
Tajbakhsh, S., Rocancourt, D., and Buckingham, M. (1996). Muscle progenitor
cells failing to respond to positional cues adopt non-myogenic fates in myf-5
null mice. Nature 384, 266–270.
Tajbakhsh, S., Rocancourt, D., Cossu, G., and Buckingham, M. (1997). Rede-
fining the genetic hierarchies controlling skeletal myogenesis: Pax-3 and Myf-5
act upstream of MyoD. Cell 89, 127–138.
Tzahor, E., Kempf, H., Mootoosamy, R.C., Poon, A.C., Abzhanov, A., Tabin,
C.J., Dietrich, S., and Lassar, A.B. (2003). Antagonists of Wnt and BMP
signaling promote the formation of vertebrate head muscle. Genes Dev. 17,
3087–3099.
von Scheven, G., Alvares, L.E., Mootoosamy, R.C., and Dietrich, S. (2006).
Neural tube derived signals and Fgf8 act antagonistically to specify eye versus
mandibular arch muscles. Development 133, 2731–2745.
Wachtler, F., Jacob, H.J., Jacob, M., and Christ, B. (1984). The extrinsic ocular
muscles in birds are derived from the precordal plate. Naturwissenschaften 71,
379–380.
Zammit, P.S., Partridge, T.A., and Yablonka-Reuveni, Z. (2006). The skeletal
muscle satellite cell: the stem cell that came in from the cold. J. Histochem.
Cytochem. 54, 1177–1191.
Zhang, W., Behringer, R.R., and Olson, E.N. (1995). Inactivation of the
myogenic bHLH MRF4 gene results in up-regulation of myogenin and rib
anomalies. Genes Dev. 9, 1388–1399.
lopmental Cell 16, 810–821, June 16, 2009 ª2009 Elsevier Inc. 821





























