Embed Size (px)
Citation preview

Fates of blood:Studies on stem cell differentiation potential
and B lymphocyte generation
in chimeric cattle
Mikael Niku
Department of Basic Veterinary Sciences
University of Helsinki
Finland
Academic dissertation
To be presented, with the permission of the Faculty of Veterinary Medicine,
for public examination in the Walter Auditorium, Agnes Sjöbergin katu 2, Helsinki,
on 12 January 2007, at 12 noon.
Helsinki 2006

Supervisors: Antti Iivanainen, DVM, PhDUniversity of Helsinki, Finland
LarsAxel Lindberg, DVM, PhDUniversity of Helsinki, Finland
Reviewers: Hanna Mikkola, MD, PhDUniversity of California, USA
Wayne Hein, BVSc (Hons), PhDAgResearch, New Zealand
Opponent: Hannu Sariola, MD, PhDUniversity of Helsinki, Finland
ISBN: 952103579X (paperback)
ISBN: 9521035803 (PDF, http://ethesis.helsinki.fi/)
ISSN: 17957079
University Printing House
Helsinki 2006

“Omnis cellula e cellula.”
Rudolf Virchow and Robert Remak, 1855

Abstract
Stem cells exist in most adult tissues. Some of these somatic stem cells may be more
plastic than expected, not limited to generating new cells for the tissue of their origin.
While such findings suggest a revolution in regenerative medicine, they remain
controversial. Most stem cell research is based on transplantation experiments of
isolated and often cultured cells in mice, and the results may be affected by the
manipulation of the animals and the transplanted cells.
Here, naturally chimeric twin cattle were used to investigate the differentiation
potential of stem cells in a nonmanipulated large mammal. Due to conjoined placental
circulations, blood of the bovine twins is mixed for most of the fetal period, and
circulating stem cells are effectively exchanged. We developed powerful methods for
tracing their progeny in various tissues of freemartin cattle, females born as a twin to a
bull. While from 10% to 90% of the hematopoietic system in freemartins was donor
derived, donor contribution to nonhematopoietic tissues was in most cases minor.
Thus, hematopoietic stem cells and other cell types circulating in fetal blood do not
generate significant numbers of nonhematopoietic cells in the development, growth
and physiological turnover of bovine tissues. However, they may be important in tissue
repair and regeneration, as suggested by increasing numbers of donorderived cells in
newly forming granulation tissue.
Chimeric cattle were also used to investigate the generation of bovine B
lymphocytes, which occurs differently than in the commonly studied human and
mouse, and is poorly understood. The results indicate that the ileal Peyer’s patch
determines the peripheral B cell pool in young cattle, and is likely responsible for the
production of the preimmune antibody repertoire by postrearrangement strategies.
To facilitate a direct analysis of bovine stem cells, the first antibodies against bovine
CD34 were generated. The CD34 glycoprotein is commonly used as a marker for
hematopoietic progenitors and endothelial cells in the human and mouse. CD34 mRNA
was found to be alternatively spliced and widely expressed in cattle tissues. Using the
new antibody, the protein was detected in most blood vessel endothelia, primitive
hematopoietic cells and some nonhematopoietic cell types.

Acknowledgements
This study was carried out at the Department of Basic Veterinary Sciences, Division of
Veterinary Anatomy, at the Faculty of Veterinary Medicine, University of Helsinki.
Financial support from the University of Helsinki, the Finnish Veterinary Foundation,
the Academy of Finland, the Ministry of Agriculture and Forestry, the Anna and Karl
Eklund Foundation and the Mercedes Zachariassen Foundation is gratefully
acknowledged.
I warmly thank the following persons, in the approximate order of appearance:
My parents, for passing on the genetic material and for encouraging me to pursue an
academic career.
The KoHu guild of international adventurers, for being the best.
Jaakko and UllaMaija Lumme, who largely are to blame for my decision to become
a biologist. Jaska, The Geneticist, has been my idol since I was 12. On the
expeditions to Varanger and elsewhere, he convinced me that biologists truly are
supreme beings. Ultsi, my legendary biology teacher, showed me that they may also
have a big warm heart.
Seppo Vainio, for introducing me to the wonderful world of developmental biology.
Antti Iivanainen, my humane and understanding supervisor, for all his ideas, and
for letting me work and learn independently (which I can say almost completely
without the usual sarcasm).
LarsAxel Lindberg, for the opportunity to work at the Department and for generous
support.
Leif C. Andersson, for allowing us to use the Haartman lab for cell culture and other
work, and for his jolly optimism. I’m sorry we never submitted to Nature (we did try
Science, though).
Tuire Pankasalo (and Hanna Valtonen), for cutting more than 28700 tissue sections,
and for important friendship; Kirsi Lahti, for skilfully staining most of those
sections, among countless other things; Pirjo Puroranta, especially for all the

Western blots; and Merja Salmitie, for the initial observation of Peyer’s patch
chimerism.
Tiina PessaMorikawa, Runar Ra, Satu Kuure and Kirsi Sainio, for inspiring
discussions and for the methodological advice provided.
Anna Ekman, Annamari Alitalo, Pauli Turunen, Lotta Ilmonen, Sanna Sirkiä, and
Marisa Palmberg, for the spirit and enthusiasm only bright young minds can bring.
The staff at the Vainio abattoir, for generous and valuable technical assistance and
for providing a cosy work atmosphere.
Jukka Pösö, for kind assistance in accessing and mining the Faba cattle database.
Eeva Jokinen and the whole VIELO team, for adding to my days some art, which I
would have missed.
All of the veterinary students, for refreshing breaks from research work and for
letting me learn by teaching (which is the second best way, after making mistakes,
and certainly more enjoyable).
Hanna Mikkola and Wayne Hein for thoroughly reviewing my thesis and for kind
and constructive comments; and Carol Ann Pelli for carefully editing the English of
the manuscript.
Finally, I am deeply grateful to my family, Mari Elisa, Iikka and Kuutti. You have
shown me what is truly important and have carried me over the moments of
frustration. To Mari Elisa, I am also indebted for reviewing the manuscript of this
thesis, making it significantly less painful an experience for the hypothetical reader.
Helsinki, 6th December 2006

Contents
1. List of original publications ..................................................................................... 1
2. Abbreviations........................................................................................................... 2
3. Introduction ............................................................................................................. 3
4. Review of the literature............................................................................................ 4
4.1. Somatic stem cells ............................................................................................ 4
4.1.1. “The stem cell is the origin of life”............................................................ 44.1.2. Hematopoietic stem cells .......................................................................... 54.1.3. Mesenchymal stem cells ........................................................................... 94.1.4. Epithelial stem cells ................................................................................ 114.1.5. Neuronal stem cells................................................................................. 134.1.6. Somatic stem cell plasticity..................................................................... 154.1.7. What makes a stem cell? ......................................................................... 20
4.2. The freemartin................................................................................................ 24
4.3. Generation of B lymphocytes in ruminants.................................................... 26
4.3.1. Production of the ruminant antibody repertoire ..................................... 274.3.2. The ileal Peyer’s patch ............................................................................ 29
5. Objectives of the study........................................................................................... 32
6. Materials and methods........................................................................................... 33
6.1. Animals, tissues and cells .............................................................................. 33
6.1.1. Animal welfare........................................................................................ 34
6.2. In situ hybridization ....................................................................................... 34
6.2.1. Genomic in situ hybridization................................................................. 346.2.2. Tyramide amplified mRNA in situ hybridization.................................... 356.2.3. Probes...................................................................................................... 35
6.3. Immunostaining ............................................................................................. 36
6.3.1. Antibodies and antigen retrieval ............................................................. 366.3.2. Immunohistochemistry ........................................................................... 366.3.3. Immunofluorescence............................................................................... 37
6.4. Combined ISH and IH double staining........................................................... 37
6.5. Microscopy and photomicrography................................................................ 37
6.6. Cell counting and statistics............................................................................. 38
6.6.1. Counting of donorderived nonhematopoietic cells in tissues............... 386.6.2. Counting of donorderived cells in leukocyte fractions........................... 38
6.7. PCRbased IgH analysis .................................................................................. 39
6.7.1. Laser capture microdissection................................................................. 396.7.2. Hybridoma culture.................................................................................. 396.7.3. Preparation of genomic DNA templates .................................................. 396.7.4. PCR amplification and analysis of products............................................ 39
6.8. Leukocyte fractionation .................................................................................. 40

6.9. Generation of a polyclonal antibovine CD34 antibody .................................. 40
6.9.1. Cloning of the fulllength CD34 cDNA.................................................... 406.9.2. Expression and purification of polypeptide fragments............................ 406.9.3. Antibody production............................................................................... 41
6.10. Western blot.................................................................................................. 41
6.11. Analysis of the bovine CD34 gene and mRNA expression............................ 42
6.11.1. Expression of CD34 mRNA.................................................................... 426.11.2. Assaying single nucleotide polymorphism............................................ 42
7. Results.................................................................................................................... 43
7.1. Detection of bull cells ..................................................................................... 43
7.2. Selection of phenotypic markers .................................................................... 43
7.3. Bullderived cells in freemartin tissues .......................................................... 43
7.4. Bullderived cells in freemartin lymphocyte populations............................... 45
7.5. Generation of a polyclonal antibovine CD34 antibody .................................. 47
7.5.1. Cloning of bovine CD34 cDNA................................................................ 477.5.2. Expression of CD34 polypeptide fragments............................................. 477.5.3. Production and characterization of a polyclonal antibody...................... 47
7.6. Expression of CD34 mRNA and protein in bovine tissues.............................. 48
8. Discussion .............................................................................................................. 50
8.1. Differentiation potential of somatic cells in adult cattle ................................. 50
8.1.1. Evaluation of methods............................................................................. 508.1.2. Implications of bullderived cells in freemartin tissues .......................... 51
8.2. Role of the ileal Peyer’s patch in generation of the B cell pool in cattle ......... 55
8.3. Generation of an antibovine CD34 antibody.................................................. 56
8.4. Expression of CD34 mRNA and protein in bovine tissues.............................. 57
9. Conclusions............................................................................................................ 59
10. References ............................................................................................................ 60
11. Original publications............................................................................................ 77

1
1. List of original publications
This thesis is based on the following publications:
I Niku M., Ekman A., PessaMorikawa T., Iivanainen A. (2006)
Identification of major cell types in paraffin sections of bovine tissues.
BMC Vet. Res. 2, 5.
II Niku M., Ilmonen L., PessaMorikawa T., Iivanainen A. (2004) Limited
contribution of circulating cells to the development and maintenance of
nonhematopoietic bovine tissues. Stem Cells 22, 1220.
III Niku M., PessaMorikawa T., Andersson L.C., Iivanainen A. (2002)
Oligoclonal Peyer's patch follicles in the terminal small intestine of cattle.
Dev. Comp. Immunol. 26, 689695.
IV PessaMorikawa T., Niku M., Iivanainen A. (2004) Persistent differences in
the level of chimerism in B versus T cells of freemartin cattle. Dev. Comp.
Immunol. 28, 7787.
V Niku M., PessaMorikawa T., Ra R., Ekman A., Iivanainen A. (2006)
Expression of CD34 mRNA and protein in bovine tissues. Submitted to
Vet. Immunol. Immunopathol.
These publications are referred to in the text by their Roman numerals.
Some unpublished material is presented (V).
Reprints are published in the printed version of this thesis with the permission of the
copyright holders. Publications I and II are freely available in full text on the respective
journals’ websites.
2
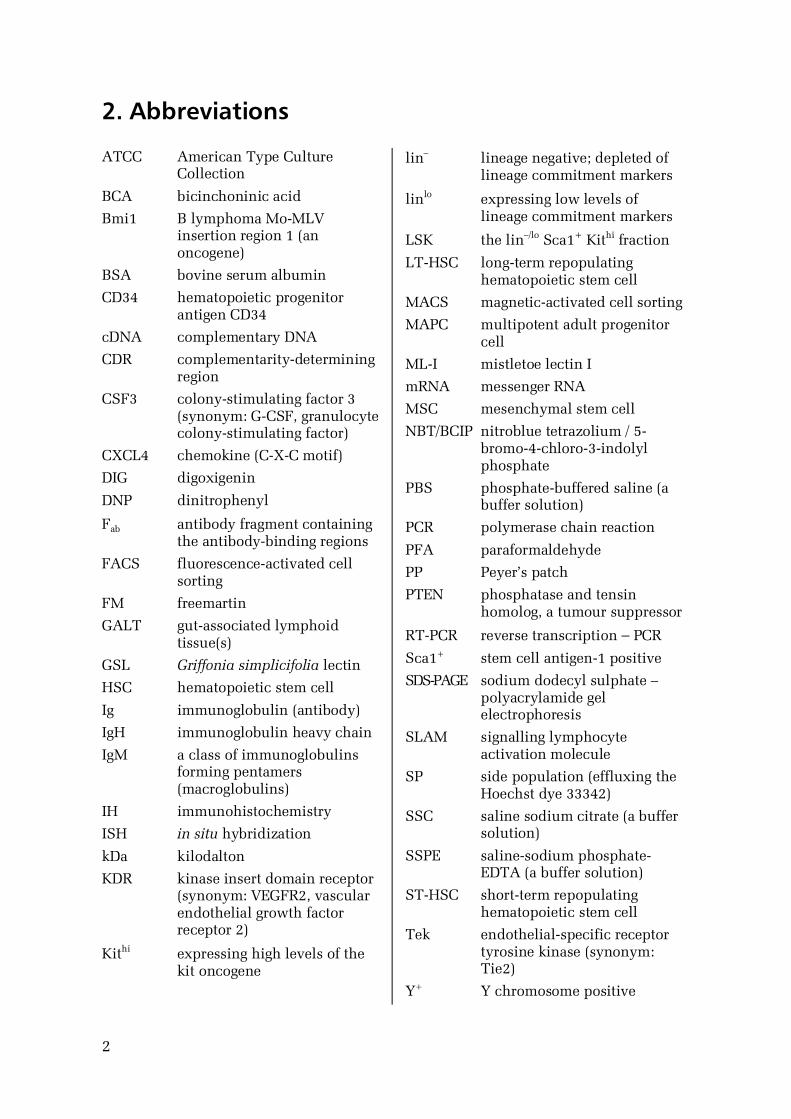
2. Abbreviations
ATCC American Type CultureCollection
BCA bicinchoninic acid
Bmi1 B lymphoma MoMLVinsertion region 1 (anoncogene)
BSA bovine serum albumin
CD34 hematopoietic progenitorantigen CD34
cDNA complementary DNA
CDR complementaritydeterminingregion
CSF3 colonystimulating factor 3(synonym: GCSF, granulocytecolonystimulating factor)
CXCL4 chemokine (CXC motif)
DIG digoxigenin
DNP dinitrophenyl
Fab antibody fragment containingthe antibodybinding regions
FACS fluorescenceactivated cellsorting
FM freemartin
GALT gutassociated lymphoidtissue(s)
GSL Griffonia simplicifolia lectin
HSC hematopoietic stem cell
Ig immunoglobulin (antibody)
IgH immunoglobulin heavy chain
IgM a class of immunoglobulinsforming pentamers(macroglobulins)
IH immunohistochemistry
ISH in situ hybridization
kDa kilodalton
KDR kinase insert domain receptor(synonym: VEGFR2, vascularendothelial growth factorreceptor 2)
Kithi expressing high levels of thekit oncogene
lin– lineage negative; depleted oflineage commitment markers
linlo expressing low levels oflineage commitment markers
LSK the lin–/lo Sca1+ Kithi fraction
LTHSC longterm repopulatinghematopoietic stem cell
MACS magneticactivated cell sorting
MAPC multipotent adult progenitorcell
MLI mistletoe lectin I
mRNA messenger RNA
MSC mesenchymal stem cell
NBT/BCIP nitroblue tetrazolium / 5bromo4chloro3indolylphosphate
PBS phosphatebuffered saline (abuffer solution)
PCR polymerase chain reaction
PFA paraformaldehyde
PP Peyer’s patch
PTEN phosphatase and tensinhomolog, a tumour suppressor
RTPCR reverse transcription – PCR
Sca1+ stem cell antigen1 positive
SDSPAGE sodium dodecyl sulphate –polyacrylamide gelelectrophoresis
SLAM signalling lymphocyteactivation molecule
SP side population (effluxing theHoechst dye 33342)
SSC saline sodium citrate (a buffersolution)
SSPE salinesodium phosphateEDTA (a buffer solution)
STHSC shortterm repopulatinghematopoietic stem cell
Tek endothelialspecific receptortyrosine kinase (synonym:Tie2)
Y+ Y chromosome positive

3
3. Introduction
Stem cells have been a focus of intense research and publicity for the last decade. They
are changing our understanding of development, physiology and pathophysiology
(Joseph and Morrison, 2005; Clarke and Fuller, 2006). In addition to the embryo, adult
tissues also contain stem cells; even the central nervous system is continuously
renewed by them, contrary to longheld dogma (Gross, 2000). Intriguingly, some of
these somatic stem cells may be more plastic than expected, not limited to generating
cells for the tissue of their origin (Blau et al., 2001). Such discoveries suggest a
revolution in regenerative medicine, stimulating public interest (Weissman, 2000). Yet,
stem cells are also a controversial issue in many ways. Some of the exciting findings
have been problematic to reproduce (Goodell, 2003). In the research models typically
used, the manipulation of cells and animals may affect the results. The interpretation of
experimental data is further complicated by poorly understood biological processes like
cell fusion (Wagers and Weissman, 2004).
The present work aimed to dissect the differentiation potential of stem cells in a
large, longlived mammal, where tissue maintenance and regenerative processes might
differ from the commonly studied small rodents. Cattle twins provide a unique
chimeric model without artificial manipulation. Blood of bovine twins is mixed for
most of the fetal period, and circulating progenitors, including hematopoietic stem
cells, are effectively exchanged (Lillie, 1917; Owen, 1945). The progeny of donor
derived cells may then be traced by genetic markers (Wilkes et al., 1981). However,
technical limitations have previously prevented an effective analysis of their
contribution to nonhematopoietic tissues. We have now developed methods making
this possible.
Chimeric cattle also were instrumental in early immunological research (Brent,
1997). Here, they offered a view on the generation and maintenance of the bovine B
lymphocyte pool, a process strikingly different from that of the more commonly studied
mouse and poorly understood.
In this thesis, I review the current literature on somatic stem cells, cattle twins, and
the production of ruminant B lymphocytes. I then present our research on the fates of
circulating cells, on the generation of B cells in cattle and on the expression of the
hematopoietic progenitor antigen CD34 in bovine tissues.

4
4. Review of the literature
4.1. Somatic stem cells
4.1.1. “The stem cell is the origin of life”
A stem cell is defined as being capable of selfrenewal and multilineage differentiation
on a singlecell basis (Weissman, 2000). The zygote, the fertilized egg, is the ultimate
stem cell; it is totipotent, able to generate all cells of the body, as well as the placenta
and the fetal membranes (Harvey, 1651; Seidel, 1952).
The early mammalian embryo forms a hollow sphere of cells, and in most species an
inner cell mass is positioned on one side of the ring (Eakin and Behringer, 2004). These
inner cells are pluripotent; each may produce any cell of the body, but not the placental
structures (Gardner and Rossant, 1979). They can be isolated and cultured as
embryonic stem cells, which are able to proliferate indefinitely, and can be induced to
differentiate into specific cell types (Evans and Kaufman, 1981).
As the development of the individual proceeds, cells gradually differentiate, losing
potential and acquiring specialized functions. Our bodies mostly consist of terminally
differentiated cells, such as mature neurons, erythrocytes, or keratinocytes of the skin,
which are unable to proliferate (Lipinski and Jacks, 1999). Yet, stem cells also exist in
adult mammals. Lost cells are in many tissues continuously replaced by somatic stem
cells (Fuchs and Segre, 2000). These typically are tissuespecific, generating cells for
the tissue in which they reside. Somatic stem cells are therefore considered multipotent,
capable of generating several cell types, oligopotent or even unipotent, depending on the
case (Sell, 2004).
The actual stem cells typically cycle relatively rarely (Cheshier et al., 1999;
Bickenbach and Mackenzie, 1984). An asymmetric cell division renews the stem cell
and creates a committed daughter (Ho, 2005). The daughter cell has restricted self
renewal capacity and potency, but often proliferates rapidly before the terminal
differentiation. These progenitor or precursor cells produce the huge numbers of new
cells needed for tissue maintenance, such as an estimated trillion blood cells daily in
man (Akashi et al., 2000; Raff, 2003; Ogawa, 1993). Therefore, they are also called
transit amplifiers.

Somatic stem cells
5
A stem cell is the beginning of an individual, and stem cells renew the tissues
throughout life. Thus, the stem cell indeed is the origin of life, as phrased poetically by
Sell (2004). It can also be an origin of death: cancer appears to be largely a stem cell
disease (Clarke and Fuller, 2006), and stem cells may have a major role in aging (van
Zant and Liang, 2003). In this review, however, I focus on somatic stem cells in a
healthy animal. I introduce some of the best characterized stem cell types and methods
for identifying or enriching them. Evidence of somatic stem cell plasticity and currently
emerging general principles in stem cell biology are discussed.
4.1.2. Hematopoietic stem cells
Hematopoietic stem cells (HSCs) are the most studied stem cells, and widely used in
the clinic for treatment of leukemias, other cancers, and congenital immunodeficiencies
(Kondo et al., 2003). Their existence was proposed already in 1906 by Alexander
Maximov, in his hematopoietic theory (Maximow, 1906). Proof arrived decades later,
when the use of nuclear weapons inspired the investigations of radiation effects on
animals, and HSC failure was shown to be a cause of radiation death (Weissman, 2000).
Lethally irradiated animals could be rescued by spleen or bone marrow transplants
from normal animals (Jacobson et al., 1951; Lorenz et al., 1952; Nowell et al., 1956;
Ford et al., 1956). These were soon shown to contain multipotent hematopoietic
progenitors (Till and McCulloch, 1961; Becker et al., 1963).
HSC is a multipotent somatic stem cell: a single HSC is able to generate all blood
cell lineages (Osawa et al., 1996). A subset of HSCs is capable of lifelong selfrenewal,
and these are called longterm hematopoietic stem cells (LTHSCs; Jones et al., 1990;
Smith et al., 1991). They give rise to shortterm HSCs (STHSCs), which are also
multipotent but have a reduced selfrenewal potential (Smith et al., 1991; Osawa et al.,
1996; Morrison et al., 1997). STHSCs generate nonrenewing multipotent progenitors
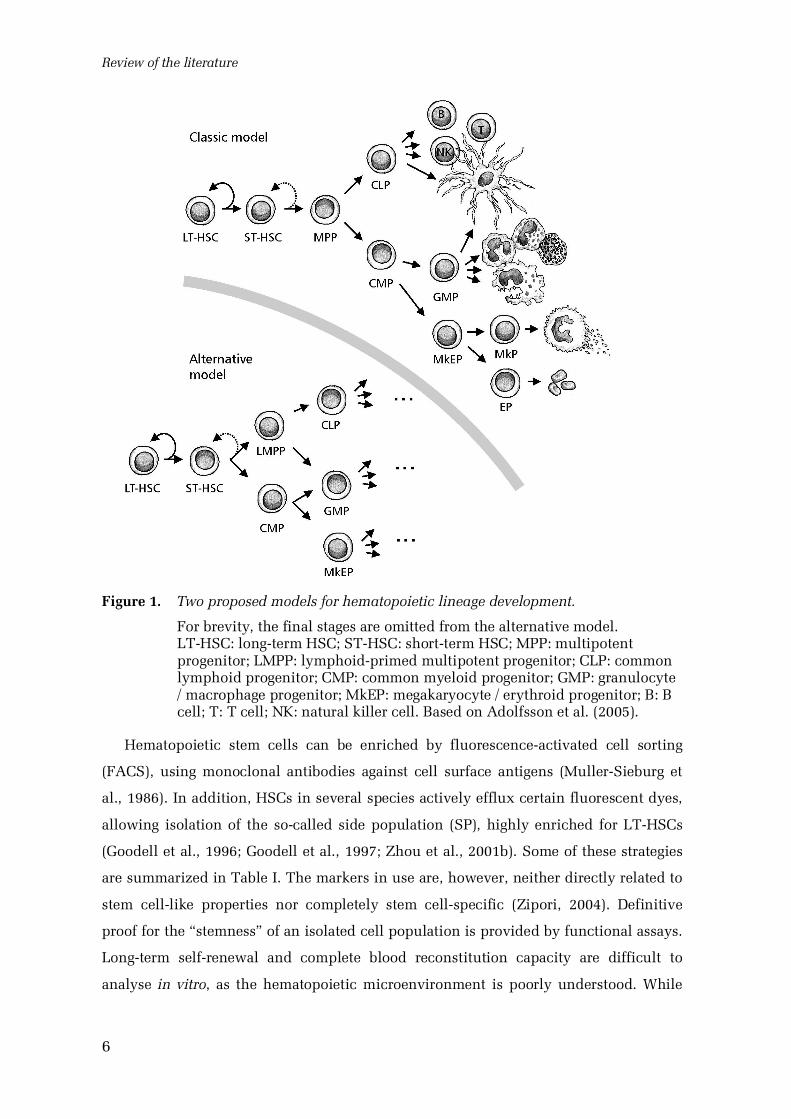
(Morrison et al., 1997) or, according to a recent alternative model (Fig. 1), common
myeloid progenitors and lymphoidprimed multipotent progenitors lacking significant
megakaryocyte and erythrocyte potentials (Adolfsson et al., 2005). Mature blood cell
lineages are finally produced through further committed progenitor stages.
Review of the literature
6
Figure 1. Two proposed models for hematopoietic lineage development.
For brevity, the final stages are omitted from the alternative model.LTHSC: longterm HSC; STHSC: shortterm HSC; MPP: multipotentprogenitor; LMPP: lymphoidprimed multipotent progenitor; CLP: commonlymphoid progenitor; CMP: common myeloid progenitor; GMP: granulocyte/ macrophage progenitor; MkEP: megakaryocyte / erythroid progenitor; B: Bcell; T: T cell; NK: natural killer cell. Based on Adolfsson et al. (2005).
Hematopoietic stem cells can be enriched by fluorescenceactivated cell sorting
(FACS), using monoclonal antibodies against cell surface antigens (MullerSieburg et
al., 1986). In addition, HSCs in several species actively efflux certain fluorescent dyes,
allowing isolation of the socalled side population (SP), highly enriched for LTHSCs
(Goodell et al., 1996; Goodell et al., 1997; Zhou et al., 2001b). Some of these strategies
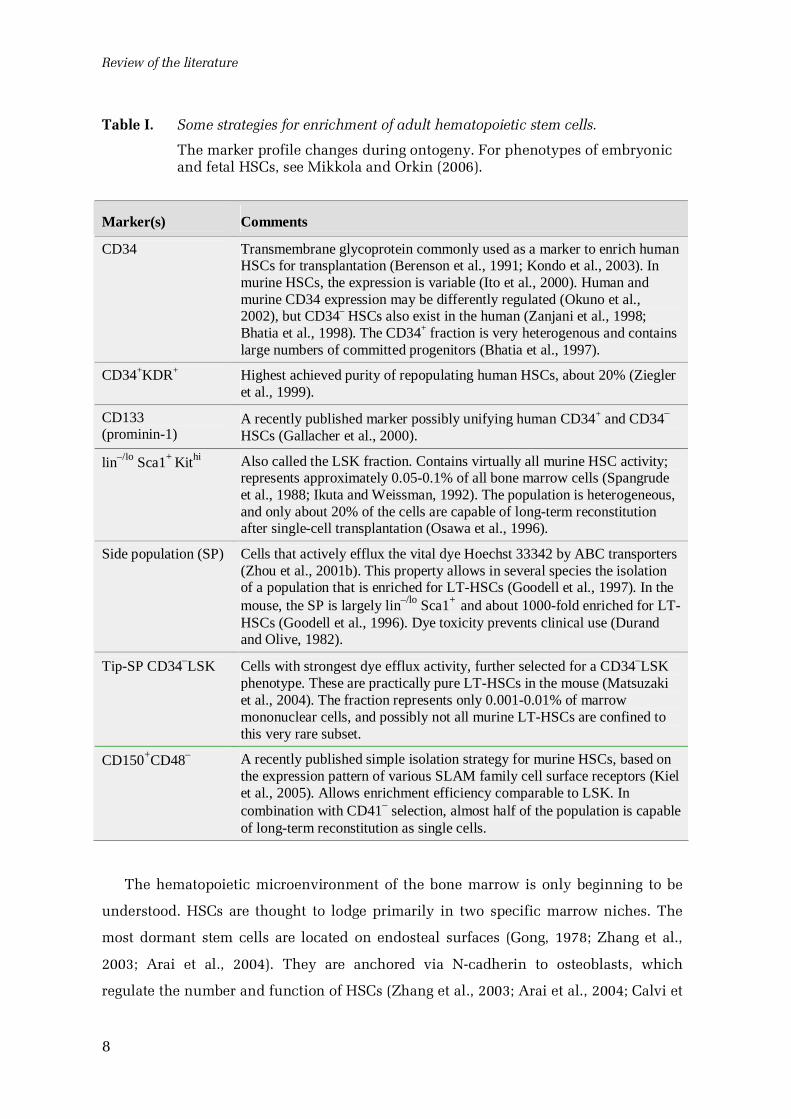
are summarized in Table I. The markers in use are, however, neither directly related to
stem celllike properties nor completely stem cellspecific (Zipori, 2004). Definitive
proof for the “stemness” of an isolated cell population is provided by functional assays.
Longterm selfrenewal and complete blood reconstitution capacity are difficult to
analyse in vitro, as the hematopoietic microenvironment is poorly understood. While

Somatic stem cells
7
several in vitro and xenograft models have been developed, only allograft
transplantation of candidate stem cell populations to lethally irradiated recipients is
considered an unequivocal test (Kondo et al., 2003). HSCs have thus been most
thoroughly characterized in mice. Even there, the precise phenotypes of the various
hematopoietic cell populations, and subsequently also the hematopoietic fate map,
remain somewhat uncertain, as shown in Fig. 1. In mammals other than mouse and
man, HSC phenotypes are poorly characterized, but they have been enriched using
lineage depletion (Leon et al., 2005) or Kit positivity (Le Guern et al., 2003).
In the developing embryo, the hematopoietic and cardiovascular organ systems are
the first to appear, being essential beyond the early postimplantation period (Copp,
1995). The earliest hematopoietic and endothelial progenitors are detected in the yolk
sac, at embryonic day 7.25 in the mouse (Ferkowicz and Yoder, 2005). They appear to
have a common precursor, the hemangioblast (Huber et al., 2004; Bailey et al., 2004).
The yolk sac first produces primitive erythrocytes, which are initially nucleated and
express embryonic globins; later, it also generates hematopoietic progenitors and
erythrocytes of the adult type (McGrath and Palis, 2005). In the embryo proper,
definitive HSCs are generated in the aortagonadmesonephros region from day 10.5
(DieterlenLièvre and Martin, 1981; Medvinsky and Dzierzak, 1996; de Bruijn et al.,
2000); the placenta has recently been recognized as another significant source (Gekas et
al., 2005; Ottersbach and Dzierzak, 2005). HSCs soon colonize the liver, which becomes
the main hematopoietic organ for most of the fetal life (Johnson and Moore, 1975;
Houssaint, 1981; Morrison et al., 1995; Ema and Nakauchi, 2000). Bone marrow is
seeded by circulating LTHSCs only shortly before birth, at day 17.5, migrating in a
chemotactic response to the stromal cellderived chemokine CXCL12 and the kit ligand
(Christensen et al., 2004). In the mouse, at least, the seeding occurs gradually, with low
numbers of HSCs constitutively circulating in the fetal bloodstream. Engraftment to the
marrow is dependent on the calciumsensing receptor, suggesting that it is guided by
the high extracellular calcium concentrations associated with bone modelling (Adams
et al., 2006).
In larger mammals, the timing of the changes in the hematopoietic environment is
somewhat different. Hematopoietic activity is seen in the bone marrow from 1015
weeks’ gestation in human fetuses (Charbord et al., 1996) and from the 4th month in
cattle (Rüsse, 1991).
Review of the literature
8
Table I. Some strategies for enrichment of adult hematopoietic stem cells.
The marker profile changes during ontogeny. For phenotypes of embryonicand fetal HSCs, see Mikkola and Orkin (2006).
Marker(s) Comments
CD34 Transmembrane glycoprotein commonly used as a marker to enrich humanHSCs for transplantation (Berenson et al., 1991; Kondo et al., 2003). Inmurine HSCs, the expression is variable (Ito et al., 2000). Human andmurine CD34 expression may be differently regulated (Okuno et al.,2002), but CD34– HSCs also exist in the human (Zanjani et al., 1998;Bhatia et al., 1998). The CD34+ fraction is very heterogenous and containslarge numbers of committed progenitors (Bhatia et al., 1997).
CD34+KDR+ Highest achieved purity of repopulating human HSCs, about 20% (Ziegleret al., 1999).
CD133(prominin1)
A recently published marker possibly unifying human CD34+ and CD34–
HSCs (Gallacher et al., 2000).
lin–/lo Sca1+ Kithi Also called the LSK fraction. Contains virtually all murine HSC activity;represents approximately 0.050.1% of all bone marrow cells (Spangrudeet al., 1988; Ikuta and Weissman, 1992). The population is heterogeneous,and only about 20% of the cells are capable of longterm reconstitutionafter singlecell transplantation (Osawa et al., 1996).
Side population (SP) Cells that actively efflux the vital dye Hoechst 33342 by ABC transporters(Zhou et al., 2001b). This property allows in several species the isolationof a population that is enriched for LTHSCs (Goodell et al., 1997). In themouse, the SP is largely lin–/lo Sca1+ and about 1000fold enriched for LTHSCs (Goodell et al., 1996). Dye toxicity prevents clinical use (Durandand Olive, 1982).
TipSP CD34–LSK Cells with strongest dye efflux activity, further selected for a CD34–LSKphenotype. These are practically pure LTHSCs in the mouse (Matsuzakiet al., 2004). The fraction represents only 0.0010.01% of marrowmononuclear cells, and possibly not all murine LTHSCs are confined tothis very rare subset.
CD150+CD48– A recently published simple isolation strategy for murine HSCs, based onthe expression pattern of various SLAM family cell surface receptors (Kielet al., 2005). Allows enrichment efficiency comparable to LSK. Incombination with CD41– selection, almost half of the population is capableof longterm reconstitution as single cells.
The hematopoietic microenvironment of the bone marrow is only beginning to be
understood. HSCs are thought to lodge primarily in two specific marrow niches. The
most dormant stem cells are located on endosteal surfaces (Gong, 1978; Zhang et al.,
2003; Arai et al., 2004). They are anchored via Ncadherin to osteoblasts, which
regulate the number and function of HSCs (Zhang et al., 2003; Arai et al., 2004; Calvi et

Somatic stem cells
9
al., 2003; Visnjic et al., 2004; Wilson and Trumpp, 2006). HSCs are also found in
association with the marrow sinusoidal endothelium (Kiel et al., 2005), where they
proliferate more actively (Wilson and Trumpp, 2006). Small numbers of HSCs are
normally present in the circulation, quickly migrating through blood and exiting within
minutes (Goodman and Hodgson, 1962; Wright et al., 2001). This may serve to maintain
hematopoietic homeostasis by distributing HSCs to unoccupied niches, but could also
be a means to delete unwanted stem cells (Abkowitz et al., 2003). Rare HSCs can also
be found in most other organs (Asakura and Rudnicki, 2002; Kotton et al., 2005;
McKinneyFreeman et al., 2002).
Most HSCs in adult bone marrow are quiescent at any one time (Cheshier et al.,
1999; Kiel et al., 2005). Yet, they are regularly recruited to cycle and can be rapidly
activated and mobilized in response to stress or injury (Wilson and Trumpp, 2006). The
hematopoietic cytokine CSF3 is commonly used in the clinic to mobilize HSCs to blood
for transplantation; interestingly, the process appears to be regulated by the
sympathetic nervous system (Katayama et al., 2006). Membranebound kit ligand
expressed by osteoblasts is essential for longterm maintenance of HSC activity (Lyman
and Jacobsen, 1998). Osteoblasts maintain stem cell quiescence through
Tek/angiopoietin1 signalling (Arai et al., 2004).
HSCs have been shown to generate differentiating progeny by asymmetric cell
division (Suda et al., 1984; Takano et al., 2004; Giebel et al., 2006), but the mechanisms
inducing the asymmetry are not understood (Wilson and Trumpp, 2006). Notch and
Wnt signalling are thought to promote selfrenewal and inhibit differentiation in an
integrated manner (Duncan et al., 2005; Reya and Clevers, 2005; Maillard et al., 2003).
Also the transcription factors HoxB4 and cmyc and the tumour suppressor PTEN have
been implicated as important regulators of HSC renewal and differentiation (Antonchuk
et al., 2002; Wilson et al., 2004; Zhang et al., 2006b).
4.1.3. Mesenchymal stem cells
Bone marrow also contains a nonhematopoietic stem cell system, the mesenchymal
stem cells (MSCs; Prockop, 1997; Väänänen, 2005). Cells generating clones of
mesenchymal lineages (fibroblasts, osteoblasts, chondroblasts and adipocytes) can be
enriched by simple plastic adhesion from the marrow and several other tissues
(Friedenstein et al., 1970; Friedenstein et al., 1987; Kuznetsov et al., 2001; Zuk et al.,

Review of the literature
10
2002). These cells are highly expandable in culture, growing up to 50 population
doublings, and multipotent on a singlecell basis (Friedenstein et al., 1987; Colter et al.,
2000; Pittenger et al., 1999; de Bari et al., 2006). They are thus regarded as stem cells,
although their stemlike properties have not been assayed as rigorously as those of
HSCs (Javazon et al., 2004). MSCs are poorly characterized and little is known of their
in vivo biology; they are of nevertheless of considerable clinical interest, due to the
favourable in vitro properties (Väänänen, 2005).
MSCs differ from HSCs in several ways. Turnover of mesenchymal tissues is very
slow compared with blood; thus, the production of differentiated cells is limited, and
also the selfrenewal capacity of MSCs in vivo is poorly elucidated (Dennis and
Charbord, 2002). MSC lineages appear to be less strictly defined than hematopoietic
lineages. The commitment to a specific blood cell lineage is thought to be irreversible
under normal conditions (Akashi et al., 2000). Fully differentiated osteoblasts,
adipocytes and chondrocytes, by contrast, may transdifferentiate to other mesenchymal
lineages (Song and Tuan, 2004). On the other hand, MSCs and HSCs may be
functionally connected. MSCs produce hematopoiesissupporting stroma (Dennis and
Charbord, 2002) and colocalize with HSCs throughout ontogeny (Mendes et al., 2005).
MSCs are found in the fetal circulation, suggesting that they might migrate through
blood like HSCs (Campagnoli et al., 2001).
No specific markers for MSCs are known, although they are generally negative for
hematopoietic and endothelial markers, and can be enriched by integrin 1 positivity
or using the monoclonal antibody STRO1 (Deschaseaux and Charbord, 2000; Simmons
and TorokStorb, 1991; Stewart et al., 2003; Gronthos et al., 2003; Javazon et al., 2004).
Therefore, MSCs are primarily identified by their in vitro differentiation capacity, and
all MSC populations investigated are heterogeneous (Javazon et al., 2004). MSC
cultures can be fractionated by FACS scatter analysis to small rapidly renewing cells,
which are highly clonogenic, and larger, granular, slowly proliferating cells (Colter et
al., 2000; Smith et al., 2004).
Recently, progenitor cells capable of generating mesenchymal and other lineages
have been isolated from bone marrow and other tissues (Reyes and Verfaillie, 2001;
Jiang et al., 2002a; Jiang et al., 2002b). These multipotent adult progenitors (MAPCs)
were obtained by prolonged culture of cells depleted for hematopoietic markers, in a
medium supplemented by epidermal growth factor, plateletderived growth factor BB,
and for the murine cells, leukemia inhibitory factor (Jiang et al., 2002a). MAPCs may

Somatic stem cells
11
represent a rare subset of MSCs, but could also be a product of the culture conditions
used to isolate them.
4.1.4. Epithelial stem cells
Epithelial stem cells have been primarily studied in the rapidly renewing epithelia of
the skin and the intestine (Rizvi and Wong, 2005). In contrast to the HSC niches, the
microanatomy of these epithelial stem cell systems is well defined, but less is known of
their cell biology. The stem cells are typically identified as retaining a label in the
chromosomal DNA, introduced by modified nucleotides or histone proteins (Tumbar et
al., 2004). In nonstem cells, the label is gradually diluted by DNA replication or is lost
by cell death, while stem cells retain the label because of their relative quiescence or,
possibly, due to selective chromatin segregation (Potten et al., 2002).
The epidermis is continuously renewed every one to three weeks, depending on
species and location (Potten, 1975a; Potten, 1975b). Cells proliferate at the basal layer
next to the basement membrane, then move towards the surface, gradually
differentiating into dying proteinaceous sacs, which are finally sloughed off (Alonso
and Fuchs, 2003). In normal epidermal homeostasis, the tissue is replenished by basal
layer stem cells, which probably comprise about 10% of all basal cells, and their
progeny, the rapidly proliferating transit amplifying cells (Levy et al., 2005; Ito et al.,
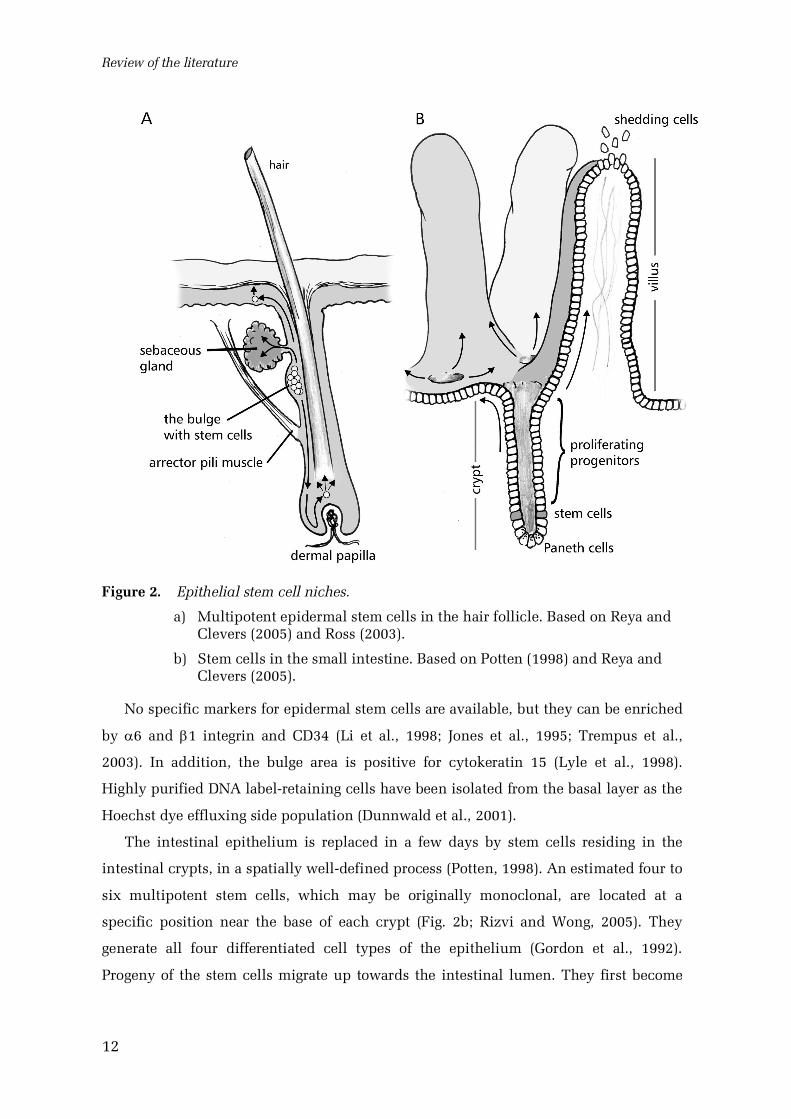
2005; Mackenzie, 1997). Still, the most multipotent epidermal stem cells are found in
the hair follicles (Fig. 2a; Moore and Lemischka, 2006). The follicle is a complex
epidermal appendage, consisting of a permanent upper portion and a cycling lower
portion which generates the hair (Alonso and Fuchs, 2003). The bulge region of the
outer root sheath, below the sebaceous gland and near the attachment to the arrector
pili muscle, is considered to be the follicular stem cell niche (Cotsarelis et al., 1990;
Alonso and Fuchs, 2003). A single cultured bulge stem cell is able to generate all
epithelial lineages of the hair follicle and sebaceous gland and also transiently
contributes to damaged interfollicular epidermis (Blanpain et al., 2004; Claudinot et al.,
2005; Levy et al., 2005; Ito et al., 2005). The stem cells are selfrenewing. They can be
cultivated for more than 120 doublings, theoretically producing far more epithelium
than necessary to cover every human being (Rochat et al., 1994). The progeny of a
single cultured cell can generate thousands of hair follicles, even in serial
transplantation to a secondary recipient (Claudinot et al., 2005; Blanpain et al., 2004).
Review of the literature
12
Figure 2. Epithelial stem cell niches.
a) Multipotent epidermal stem cells in the hair follicle. Based on Reya andClevers (2005) and Ross (2003).
b) Stem cells in the small intestine. Based on Potten (1998) and Reya andClevers (2005).
No specific markers for epidermal stem cells are available, but they can be enriched
by 6 and 1 integrin and CD34 (Li et al., 1998; Jones et al., 1995; Trempus et al.,
2003). In addition, the bulge area is positive for cytokeratin 15 (Lyle et al., 1998).
Highly purified DNA labelretaining cells have been isolated from the basal layer as the
Hoechst dye effluxing side population (Dunnwald et al., 2001).
The intestinal epithelium is replaced in a few days by stem cells residing in the
intestinal crypts, in a spatially welldefined process (Potten, 1998). An estimated four to
six multipotent stem cells, which may be originally monoclonal, are located at a
specific position near the base of each crypt (Fig. 2b; Rizvi and Wong, 2005). They
generate all four differentiated cell types of the epithelium (Gordon et al., 1992).
Progeny of the stem cells migrate up towards the intestinal lumen. They first become

Somatic stem cells
13
transit amplifying cells at the upper part of the crypt, then differentiate and stop
proliferating, and finally are exfoliated to the lumen (Potten, 1998). Paneth cells in the
small intestine follow an opposite course, settling at the crypt bases (Rizvi and Wong,
2005). The villi of the small intestine are covered by sectors of epithelium originating
from several neighbouring crypts (Bjerknes and Cheng, 1999).
Studies of the intestinal epithelium have been hindered by the lack of markers.
Recently, the RNA binding protein Musashi1 was reported to label stem and early
progenitor cells in the intestine (Potten et al., 2003; Kayahara et al., 2003).
The Wnt signalling cascade is essential in the regulation of epithelial proliferation
and differentiation in both the skin and the intestine (Reya and Clevers, 2005). It is
required for the maintenance of intestinal crypt progenitors (Korinek et al., 1998), and
persistent activation of the pathway is an initiating event in colorectal cancer (Reya and
Clevers, 2005). On the other hand, Wnt signals induce the differentiation of Paneth
cells (van Es et al., 2005). Through the EphB/ephrinB system, the Wnt pathway also
affects the sorting and positioning of intestinal epithelial cells (Batlle et al., 2002). In
the epidermis, Wnt signalling is thought to promote proliferation and hair follicle
development (Reya and Clevers, 2005). The bone morphogenetic protein (BMP)
signalling pathway is integrated with Wnt to negatively regulate stem cell proliferation
and stimulate differentiation (Moore and Lemischka, 2006). Notch signalling is also
implicated in the regulation of differentiation in both the epidermis and the intestinal
epithelium (Rizvi and Wong, 2005).
4.1.5. Neuronal stem cells
New neurons are continuously produced in the brains of adult vertebrates, contrary to
the longheld dogma (Gross, 2000; GarciaVerdugo et al., 2002). Adult neurogenesis was
originally suggested by DNA labelling experiments in the rodent hippocampus (Altman
and Das, 1965), and then shown to be associated with vocal learning in songbirds (for a
historical review, see Nottebohm, 2004). However, this theory was not generally
accepted until the 1990s (Ming and Song, 2005). Multipotent neural stem cells are now
known to exist in the hippocampus and in the walls of the lateral ventricles of adult
rodents and humans (Fig. 3; Reynolds and Weiss, 1992; Reynolds and Weiss, 1996;
Palmer et al., 1997; Eriksson et al., 1998; Kukekov et al., 1999; Ahn and Joyner, 2005).
They are capable of selfrenewal and clonal generation of neurons and glia in vitro and

Review of the literature
14
in vivo. In culture, the stem cells typically form floating aggregates of cells called
neurospheres (Reynolds and Weiss, 1992). In the hippocampus, new neurons are
generated in the subgranular zone of the dentate gyrus and then differentiate into
granular neurons (Altman and Das, 1965; Kaplan and Bell, 1984; Bayer, 1982; Stanfield
and Trice, 1988). The stem cells in the ventricular walls are located at the
subventricular zone, below the ependymal layer (Lois and AlvarezBuylla, 1993;
Chiasson et al., 1999). In rodents, their progeny migrates to the olfactory bulb, along the
rostral migratory stream (Lois and AlvarezBuylla, 1994; Sanai et al., 2004). In other
areas of the central nervous system, the generation of new neurons is controversial and
speciesdependent (Falk and Frisen, 2005; GarciaVerdugo et al., 2002). The cerebral
cortex of healthy adult humans showed no neuronal turnover in an ingenious study
applying 14C dating to cellular DNA (Spalding et al., 2005).
Figure 3. Neuronal stem cell niches in the murine brain.
Schematic sagittal view. Based on Uchida (2000) and Sidman et al. (1971).
In both germinal regions, neuronal stem cells are astroglialike cells expressing glial
fibrillary acidic protein (GFAP), a traditional marker of mature astroglia (Doetsch et al.,
1999a; Seri et al., 2001; Garcia et al., 2004). They also express nestin (Lendahl et al.,
1990) and prominin1 (Uchida et al., 2000). Yet, labelling of dividing cells remains the
principal method for reliably identifying them and their progeny (Ming and Song,
2005).
The neuronal stem cell niches appear to be associated with blood vessels (Doetsch,
2003). Hippocampal neurogenesis occurs around capillaries in proliferative clusters

Somatic stem cells
15
also containing endothelial precursors (Palmer et al., 2000). In the subventricular zone,
the proliferating cells are in contact with specialized extensions of the vascular basal
lamina (Mercier et al., 2002), and the stem cells occasionally also extend to the
ventricle (Doetsch et al., 1999b).
Neuronal stem cells are thought to generate several stages of transitamplifying cells,
which then differentiate into postmitotic neurons (AlvarezBuylla et al., 2001;
Kempermann et al., 2004). They acquire a neuronal phenotype, express neuronspecific
receptor proteins, establish synaptic connections, display typical electrophysiological
properties and respond to stimuli (Carlén et al., 2002; van Praag et al., 2002; Song et al.,
2002; Belluzzi et al., 2003; Carleton et al., 2003). The new neurons are thought to
differentiate in an activitydependent manner, possibly sensing activities of the local
neuronal network even before receiving synaptic innervation (Carleton et al., 2003).
The process resembles the neuronal development in the embryo (Esposito et al., 2005).
The functional significance of adult neurogenesis is still unclear, largely due to the
extreme complexity of the central nervous system. It seems to be associated with
learning and memory, although the process is too slow for acute benefits (Kempermann
et al., 2004). Neurogenesis is also stimulated by physical exercise or injury, and
possibly reduced in stress and depression (Ming and Song, 2005).
Major developmental signalling pathways regulate stem cells also in the central
nervous system (AlvarezBuylla and Lim, 2004). Wnt signalling is required for
neurogenesis, at least in the hippocampus (Lie et al., 2005), whereas Sonic hedgehog
and Notch are thought to maintain both stem cell populations (AlvarezBuylla and Lim,
2004). The tumour suppressors PTEN and p53 inhibit stem cell selfrenewal (Groszer et
al., 2006; Meletis et al., 2006).
4.1.6. Somatic stem cell plasticity
Somatic stem cells are traditionally thought to be restricted to particular fates,
producing only cell types found in the tissue of their origin (Lakshmipathy and
Verfaillie, 2005; Wagers and Weissman, 2004). This dogma has been challenged by
reports of stem cell plasticity, claiming that somatic stem cells are also capable of
generating cells of nonrelated tissues. Transplanted bone marrow cells have been
reported to produce in vivo skeletal and cardiac muscle (Ferrari et al., 1998; LaBarge
and Blau, 2002; Orlic et al., 2001), liver (Petersen et al., 1999; Theise et al., 2000),

Review of the literature
16
endothelium (Asahara et al., 1999), epithelia (Okamoto et al., 2002; Körbling et al.,
2002; Kale et al., 2003), neurons (Mezey et al., 2000; Brazelton et al., 2000; Mezey et al.,
2003; Priller et al., 2001) and even oocytes and male germ cells (Johnson et al., 2005;
Nayernia et al., 2006). Of the various bone marrow lineages, purified HSCs (typically
defined as the LSK fraction) also seem to contribute to muscle (Corbel et al., 2003;
Camargo et al., 2003), liver (Lagasse et al., 2000), endothelium (Grant et al., 2002; Bailey
et al., 2004), various epithelia (Krause et al., 2001) as well as astrocytes and
oligodendrocytes (Koshizuka et al., 2004). Similarly, MSCs were reported to
differentiate into cardiomyocytes (Toma et al., 2002) and astroglia (Kopen et al., 1999),
besides the various mesenchymal lineages. In addition to these bone marrowderived
stem cells, MAPCs isolated from several tissues generate hematopoietic cells, various
epithelia, and skeletal muscle (Jiang et al., 2002a; Jiang et al., 2002b; Muguruma et al.,
2003); skeletal muscle appears to produce hematopoietic cells (Jackson et al., 1999);
and neuronal stem cells may give rise to blood (Bjornson et al., 1999), muscle (Galli et
al., 2000) and, indeed, in chimeric embryos, to all germ layers (Clarke et al., 2000).
Plasticity of somatic stem cells could revolutionize our understanding of tissue
development and maintenance, and a multitude of clinical applications may be
envisioned (Herzog et al., 2003). Unfortunately, it has often been problematic to
reproduce the reported findings. Bone marrow transplants, purified HSCs and
parabiosis partners have repeatedly failed to yield neuronal cells in an intact or injured
brain, although their contribution to microglia is remarkable (Castro et al., 2002;
Roybon et al., 2006; Massengale et al., 2005). Instead, some of the bone marrow
hematopoietic populations were confusingly found to express proteins regarded as
neuronal markers. In another study, recipients of single HSCs showed no contribution
to most tissues, and only a single donorderived neuron was detected in more than 13
million total cells examined, despite a robust hematopoietic engraftment (Wagers et al.,
2002). HSCs also failed to generate cardiomyocytes (Murry et al., 2004), and no oocytes
were derived from bone marrow transplants or longterm parabiosis partners (Eggan et
al., 2006). Generally, reported frequencies of bone marrowderived engraftment vary
several orders of magnitude, and are usually low, around 0.52% of nonhematopoietic
cells in various tissues and experiments (Goodell, 2003). Concerning neuronal stem
cells, hematopoietic capacity was undetected in almost 130 · 106 neurosphere cells
transplanted to more than 100 recipient mice (Morshead et al., 2002). Hence, the

Somatic stem cells
17
plasticity of somatic stem cells is clearly a controversial issue (Goodell, 2003; Raff,
2003; Wagers and Weissman, 2004).
The outcomes of the experiments obviously vary depending on the type and status
of the transplant recipient. Tissue damage generally promotes engraftment (Petersen et
al., 1999; LaBarge and Blau, 2002; Okamoto et al., 2002); this is in many studies
intentional, but poorly defined damage is also induced by common transplantation
procedures (Prise et al., 2005). The transplanted cells are to a variable degree affected
by the manipulation involved in their isolation and possible cultivation (Morshead et
al., 2002).
Some of the findings may even be due to detection artefacts; the risk is increased in
the analysis of exceedingly rare events. The transplanted cells can be traced in recipient
tissues by labels such as the Y chromosome, a detectable genetic polymorphism, or a
transgene, typically expressing green fluorescent protein (GFP) or galactosidase.
Colocalization of the label with various phenotypic markers, most commonly using
immunostaining, then reveals the fates of donorderived cells (Herzog et al., 2003). In
ordinary light microscopy, superimposed cells may produce an illusion of marker
colocalization (Goodell, 2003). Any labelling method used to track stem cells and their
progeny is subject to potential false positives: in situ hybridization to genomic markers
could be unspecific, autofluorescence may confuse interpretation of GFP signals and
endogenous galactosidase activity occurs in some conditions (Herzog et al., 2003).
Regarding derivatives of human bone marrow transplants, false positives may be caused
by male cells existing in female tissues in the absence of transplantation (Stevens et al.,
2004; Khosrotehrani et al., 2004). These may be of fetal origin, but are found quite often
even in women who have not given birth to a son (Yan et al., 2005). False negatives
may be caused by unstable or lowlevel transgene expression (Torensma and Figdor,
2004). In addition, antibodies (or even antigens!) used to assay the phenotypes of
labelled cells are often not completely specific (Goodell, 2003).
Still, detection artefacts cannot account for all of the observations suggestive of
somatic cell plasticity. Confocal microscopy with 3D reconstruction and double
staining with hematopoietic markers have been used to resolve potential cell overlays
(LaBarge and Blau, 2002; Jiang et al., 2002a). Specificity of labels can be ensured with
careful controls or by the verification of results with several different labelling systems
(Bailey et al., 2004). Unambiguous morphological properties of cells or functional
analysis support the immunostaining data in some works (Priller et al., 2001; Lagasse et

Review of the literature
18
al., 2000). A stepwise progression of bone marrow cells to muscle, in response to
various stimuli, has also been documented (LaBarge and Blau, 2002).
Several biological explanations may account for the apparent plasticity, depending
on the experimental setup (Fig. 4). These have been directly addressed in few studies
(Raff, 2003; Wagers and Weissman, 2004; Lakshmipathy and Verfaillie, 2005).
True plasticity would involve transdifferentiation (Fig. 4a) or dedifferentiation (Fig.
4b) of somatic cells. Transdifferentiation refers to a direct lineage change to a
completely different cell type, while dedifferentiation implies a conversion of a tissue
specific cell to a more primitive stem cell, followed by redifferentiation along a new
pathway (Wagers and Weissman, 2004). These processes have not been unambiguously
documented in mammals (Wagers and Weissman, 2004). Transitamplifying precursors
may be converted to stem cells in the brain (Doetsch et al., 2002) and intestinal
epithelium (Potten, 1998); also, HSCs express a multitude of genes related to non
hematopoietic tissues (Akashi et al., 2003). Theoretically, plasticity clearly should be
possible. Nuclei of even extremely specialized cells like lymphocytes can sometimes be
reprogrammed to enable cloning of an animal (Hochedlinger and Jaenisch, 2002), and
regeneration involving plasticity of differentiated cells occurs readily in urodele
amphibians (Brockes and Kumar, 2002).
Unfortunately, in many of the cases presented above, the transplanted cell
populations are far from homogeneous. Unfractioned bone marrow preparations are
likely to contain several kinds of stem and progenitors cells, instead of a single highly
plastic type (Fig. 4c; Wagers and Weissman, 2004). Yet, in several studies, single
purified cells have been transplanted, indicating true multipotency and attributing this
capacity to actual stem cells at least indirectly (Krause et al., 2001; Jiang et al., 2002a;
Bailey et al., 2004).
Alternatively, rare multipotent stem cells may contaminate the isolated tissue
specific stem cells (Fig. 4d; Wagers and Weissman, 2004). Contaminating HSCs seem to
account for the reported hematopoietic capacity of skeletal muscle (McKinneyFreeman
et al., 2002). In addition, as presented in section 4.1.3., highly potent MAPCs have been
derived from many tissues, although their actual existence in vivo is unclear (Jiang et
al., 2002b). In transplantations of single stringently purified LSK HSCs, multilineage
differentiation obviously indicates actual plasticity of HSCs defined by the currently
recognized criteria; even this population is, however, known to be heterogeneous
(Osawa et al., 1996).

Somatic stem cells
19
Figure 4. Potential mechanisms for apparent somatic stem cell plasticity.
a) Transdifferentiation.
b) Dedifferentiation.
c) Multiple stem cells (a heterogeneous population isolated).
d) A rare multipotent stem cell among tissuespecific stem cells.
e) Cell fusion.
Based on Herzog et al. (2003) and Wagers and Weissman (2004).
Finally, bone marrowderived cells have been shown to contribute to several tissues
by cell fusion (Fig. 4e). In culture, bone marrow cells fuse to embryonic stem cells,
adopting their phenotype (Terada et al., 2002). Under selective pressure, bone marrow
derived hepatocytes in vivo arise primarily by fusion (Wang et al., 2003). In another
studies, bone marrow cells fused to liver, Purkinje cells and cardiac muscle in
irradiated recipients, with no actual transdifferentiation events detected (Alvarez
Dolado et al., 2003), and to intestinal stem cells (Rizvi et al., 2006). Fusion to Purkinje
cells was shown to produce stable heterokaryons, where the bone marrow cell nucleus
is reprogrammed (Weimann et al., 2003). Cell fusion is a wellknown property of
macrophages (Vignery, 2005), and indeed, HSCs appear to contribute to skeletal muscle
through myeloid intermediates rather than directly (Camargo et al., 2003). On the other
hand, fusion events are rare in the absence of selection (Wagers and Weissman, 2004).
Observations of HSC and neuronal stem cell plasticity have also been reported in the
absence of cell fusion (Jang et al., 2004; Bailey et al., 2004; Wurmser et al., 2004).
Taken together, the significance of somatic stem cell plasticity is still unclear, and
understanding the often apparently contradictory observations requires further
research. To convincingly demonstrate an actual transdifferentiation event, several
stringent criteria are proposed: the investigated cells should be minimally manipulated
and cultured; clonal analysis should be performed; the transdifferentiated cells must be

Review of the literature
20
unambiguously identified, phenotyped and shown to be functionally engrafted to
tissue; nuclear reprogramming should be assessed, and cell fusion must be excluded
(Wagers and Weissman, 2004). However, clinical applications may be feasible, even if
transdifferentiation is not a physiologically relevant process.
4.1.7. What makes a stem cell?
The basis of “stemness” is not understood, despite intensive research (Mikkers and
Frisen, 2005). The commonly used definitions of a stem cell are ambiguous: some
differentiated cells are able to selfrenew (Luckey et al., 2006), and some stem cells,
such as those of the germline, are unipotent, generating a single type of progeny (de
Rooij and Grootegoed, 1998).
Stem cells do share some characteristics. Several signalling pathways, including
Wnt and Notch, are common to multiple stem cell types, as presented above. In
addition, specific small RNAs are emerging as important stem cell regulators (Zhang et
al., 2006a). Stem cells in various tissues have been enriched by their dye efflux ability,
as the side population. The enhanced membrane transport may be involved in
protection of longlived stem cells from cytotoxic substances (Challen and Little, 2006).
Stem cells also seem to carefully protect their genomic DNA. Telomerase is commonly
expressed by stem and progenitor cells, preserving chromosome ends from degradation
(Harrington, 2004). Evidence is also accumulating for the immortal strand hypothesis;
asymmetrically dividing stem cells apparently retain the original template strands,
passing the newly synthesized chromatids to the committed daughters (Cairns, 1975;
Merok et al., 2002; Potten et al., 2002; Smith, 2005; Karpowicz et al., 2005; Shinin et al.,
2006). In contrast, some forms of DNA repair may actually be downregulated;
potentially mutated stem cells are preferentially replaced, in agreement with a strict
control of genomic stability (Cairns, 2002).
Despite such general features, no unambiguous molecular markers of stem cells are
available, let alone markers common to all stem cell types (Zipori, 2004). In search of a
stem cell signature, transcriptomes of embryonic, hematopoietic, and neuronal stem
cells have been compared by three independent groups using DNA array technology
(Ivanova et al., 2002; RamalhoSantos et al., 2002; Fortunel et al., 2003). All groups
found more than 200 genes upregulated in all stem cell types, thus representing
putative “stemness” genes. These gene sets were, however, strikingly different; only a

Somatic stem cells
21
single gene, integrin 6, was identified in all three studies, although a multitude of
genes enriched for embryonic or neuronal stem cells were reported in good agreement
(Fortunel et al., 2003). While variations in stem cell purification methods and
shortcomings in the current DNA arrays partially explain the discrepancies, it appears
that specific stemness genes are expressed at a low level or only transiently, or may not
exist at all (Fortunel et al., 2003; Zipori, 2004). Instead, stemness may be established by
subtle differences in the expression of many genes, or different stem cells may use
distinct genetic programs for selfrenewal and differentiation (Fortunel et al., 2003).
Figure 5. Stem cell: a blank state (a) or priming for multipotentiality (b)?
Adapted from Zipori (2004) with permission.
HSCs show a highly promiscuous gene expression pattern, expressing multiple
genes active in specific hematopoietic lineages, and even genes characteristic of non
hematopoietic cells (Akashi et al., 2003). As the cells differentiate to various committed
progenitors, the expression pattern is progressively restricted. HSCs actively express
genes associated with energy metabolism and translation (Hüttmann et al., 2006). They
thus appear to be quiescent only with regard to proliferation, otherwise prepared to
rapidly respond to environmental stimuli. Stem cells may in fact be characterized by a
wideopen chromatin structure (Mikkers and Frisen, 2005). Rather than representing a
blank state that gradually obtains properties of differentiated cells (Fig. 5a), stem cells
could be primed for multipotentiality by expression of many genes, which are then
selectively repressed or enhanced during differentiation (Fig. 5b; Zipori, 2004). The
importance of chromatin modification in the specification of stemness is illustrated by
the requirement of Bmi1 in selfrenewal of hematopoietic and neuronal stem cells, as
well as many murine and human cancers (ValkLingbeek et al., 2004). This polycomb
group member is involved in epigenetic regulation.

Review of the literature
22
Difficulties in defining a stem cell, heterogeneity of isolated stem cell populations,
and observed plasticity have led some authors to suggest that stemness is a regulatable
state rather than a discrete entity (Zipori, 2004; Blau et al., 2001; Fig. 6). Perhaps any
cell is able to function as a stem cell, given the correct environment (although
differentiated cells are less likely to do so)!
Figure 6. Route stem cell.
A stem cell landscape inspired by the initial reports of plasticity. Stem cellsare envisioned here as travelling in the bloodstream, entering diverse tissuesand generating appropriate cell types in response to local cues.Reproduced from Blau et al. (2001) with permission. Original artwork by N.Gewertz and B. Colyear.
Indeed, the niche appears essential in stem cell biology, as exemplified above for
various stem cell types. The niche is a specialized microenvironment sheltering the
stem cells, consisting of neighbouring cells and extracellular substance (Moore and
Lemischka, 2006; Fuchs et al., 2004; Spradling et al., 2001). By interactive signalling,
the niche maintains a balance of stem cell quiescence and activity, regulating self
renewal and differentiation. Best characterized in the Drosophila ovary, asymmetry and

Somatic stem cells
23
subsequent differentiation may be commonly achieved by directional cell division,
where one daughter remains attached to the niche, and the other is separated, as
schematically shown in Fig. 7 (Spradling et al., 2001). Little is known, though, of the
strategies actually used by mammalian somatic stem cells (Morrison and Kimble, 2006).
Figure 7. Symmetric and asymmetric division in the niche.
Symmetric divisions expand the stem cell population; asymmetric divisionsgenerate differentiating daughter cells and renew the stem cells. Based onSpradling et al. (2001).
The state and history of the cell are still likely to affect the interpretation of
environmental signals (Mikkers and Frisen, 2005). This is a central principle in
developmental biology (Gilbert, 2006). As previously presented, intestinal stem cells
and Paneth cells reside in intimate proximity, yet certain signals seem to affect them in
almost opposite ways. Stem cells may be described as being halted in the progression
towards differentiation, an apparently inherent tendency of mammalian cells (Fig. 8;
Mikkers and Frisen, 2005; Clarke and Fuller, 2006). This status is maintained by
complex interactions between the stem cell and the niche (Mikkers and Frisen, 2005).

Review of the literature
24
Figure 8. Stem cells are halted in the progression towards differentiation.
Adapted from Mikkers and Frisén (2005) with permission. Artwork assistedby Iikka Niku.
4.2. The freemartin
Cattle provide a unique large animal model for stem cell research: most bovine twins
are naturally chimeric as a result of conjoined fetal circulations. It is thus unnecessary
to apply the typical transplantation procedures, where poorly defined radiation damage
affects the results (Prise et al., 2005). Isolation and in vitro cultivation of transplanted
cells are also avoided, minimizing alterations caused by manipulation (Morshead et al.,
2002).
Vascular anastomosis occurs in about 92% of cattle twins, due to fused chorions
(Fig. 9; Marcum, 1974; Lillie, 1917). The anastomoses form early in the development,
from the 10 to 15mm crownrump stage (Lillie, 1917; Bissonnette, 1924; Ohno and
Gropp, 1965). This corresponds to about 3035 days postcoitum or Carnegie stages 15
18; the limb buds and forelimb plates are visible, many internal organs develop rapidly,
and the genital ridges appear (Ohno and Gropp, 1965; Rüsse, 1991; Butler and Juurlink,
1987; Bissonnette, 1924). The blood is thus effectively mixed between the fetuses for
most of the 280day gestation. Postnatally, the twins permanently share identical

The freemartin
25
composite blood types, indicating that HSCs are exchanged and successfully engraft in
the recipient (Owen, 1945). The proportions of donorderived nucleated cells in blood
and bone marrow in different twins are reported to be randomly distributed between
less than 5% and more than 95% (Wilkes et al., 1981). The twins are tolerant to tissue
allografts from each other (Anderson et al., 1951; Billingham et al., 1952). This
serendipitous discovery led to further research in mice and chickens, unravelling the
basis of acquired immunological tolerance (Billingham et al., 1953); the findings earned
Peter Medawar a Nobel prize in 1960 (Brent, 1997).
In other species, chimerism caused by vascular anastomosis is rare, although it has
been reported in at least sheep, goats and pigs (Marcum, 1974). Lowlevel
microchimerism appears to be surprisingly common in humans (Boklage, 2006).
However, this may be primarily caused by cell exchange between temporarily fused
early embryos, rather than through vascular connections between fetuses or between a
fetus and the mother.
Donorderived cells can be readily identified in a female animal born as a twin to a
male, as only bull cells contain a Y chromosome. In cattle, a female calf born as a twin
to a bull is usually infertile due to disturbances in the development of the reproductive
organs. These are probably caused by the antiMüllerian hormone produced by the
male fetus (Jost, 1970; Vigier et al., 1977; Vigier et al., 1987); the bovine twins were a
key research model in the early days of reproductive endocrinology (Lillie, 1917; Capel
and Coveney, 2004). Otherwise, the animals develop normally (Padula, 2005).
The infertile heifers are called freemartins. The “free” may derive from the Scottish
farrow, for a cow not giving milk or without a calf, or from the AngloSaxon fearr, a bull
(Forbes, 1946). The “martin” possibly originates from the Gaelic and Irish mart,
meaning cow, heifer, ox, cow for slaughter, or beef. The term was often applied to cattle
killed at the time of the Martinmas festival in November, to preserve the meat for
winter; the nonproductive freemartins were probably selected for slaughter. In the
scientific literature, freemartins were first described more than two hundred years ago
by John Hunter (1779):
“ It is a known fact, and, I believe, is understood to be universal, that when acow brings forth two calves, and that one of them is a bullcalf, and the other acow to appearance, the cowcalf is unfit for propagation; but the bullcalfbecomes a very proper bull. They are known not to breed: they do not evenshew the least inclination for the bull, nor does the bull ever take the leastnotice of them.”

Review of the literature
26
Diagnostic methods for freemartinism range from clinical examination to Y
chromosomespecific PCR (Padula, 2005). Composition of freemartin tissues has been
analysed by cytogenetic techniques (Wilkes et al., 1981). Still, due to limitations of
these methods, little is known of the contribution of bullderived cells to the non
hematopoietic tissues.
Figure 9. Cattle twins.
1: arterial anastomosis; 2: cotyledon with venous connection with bothsides; 3: opened amniotic sacs; 4: clitoris of the freemartin. Reproducedfrom Lillie (1917) with permission of WileyLiss, Inc. a subsidiary of JohnWiley & Sons, Inc. Copyright 1917 John Wiley & Sons, Inc.
4.3. Generation of B lymphocytes in ruminants
Our current understanding of the immune system is mostly based on mouse research,
and to some extent on the human system. However, it turns out that these species are
not representative of all mammals (Reynolds, 1997). A striking lesson is provided by
the ontogeny of the antibodyproducing B lymphocytes in ruminants. In this part of the
review, I discuss the strategies ruminants use for the generation of their antibody
repertoire, and the major role of the ileal Peyer’s patch in this process.

Generation of B lymphocytes in ruminants
27
4.3.1. Production of the ruminant antibody repertoire
Antibodies are able to specifically recognize almost any substance due to tremendous
variation in their antigenbinding sites (Janeway et al., 2001). Every immunoglobulin
(Ig) variant cannot be directly encoded in the genome because they are far more
numerous than all of our genes. Instead, vertebrates use various alternative strategies to
generate the necessary diversity (Ratcliffe, 2002).
The variable regions of Igs are assembled from several gene segments by
rearrangements of genomic DNA in developing B lymphocytes (Fig. 10). In the mouse
and human, multiple slightly different copies of each gene segment are available,
allowing a high number of possible combinations. Additional diversity is generated in
the joints between gene segments by addition and removal of nucleotides (Janeway et
al., 2001). This preimmune repertoire is produced in the absence of antigenic
stimulation (Griebel and Hein, 1996). In postnatal mice and humans, the process occurs
in bone marrow, continuing more or less throughout life (Nunez et al., 1996; Min et al.,
2006). Since B cells arise there from uncommitted progenitors (see Fig. 1), the bone
marrow is regarded as a primary or central lymphoid tissue (Janeway et al., 2001). Igs
are finetuned later, as B cells are exposed to antigens, to create the immune repertoire.
In the process of somatic hypermutation, point mutations are introduced at a high rate
into the variable region genes. This takes place in secondary or peripheral lymphoid
tissues, such as the lymph nodes (Janeway et al., 2001).
In chicken and in several mammalian species, gutassociated lymphoid tissues
(GALT) are essential for the generation of B lymphocytes (Reynolds, 1997; Weill and
Reynaud, 1998). Actually, B cells were first identified as bursadependent lymphocytes
in the chicken, produced in the cloacaassociated bursa of Fabricius (Glick et al., 1956).
Lymphoid follicles in the ileal Peyer’s patch appear to function analogously in
ruminants, the bestcharacterized of which are sheep (Yasuda et al., 2006). Cattle are
speculated to be similar.
The GALT species seem to rely less on the combinatorial diversity than the mouse
and human. Instead, their preimmune Ig repertoire is generated largely in GALT by
antigenindependent somatic hypermutation or gene conversion, where pseudogene
sequences replace parts of the functional Ig genes (Ratcliffe, 2002).
Sheep were originally reported to have a very limited selection of Ig variable region
gene segments, even fewer of which are actually used (Reynaud et al., 1991). Igs are
subsequently diversified by somatic hypermutation, where point mutations are

Review of the literature
28
introduced to the variable regions at an extremely high rate. The process starts antigen
independently in the sterile fetus and continues after birth (Reynaud et al., 1995).
However, more Ig gene segments have later been identified in the ovine genome,
suggesting that the combinatorial mechanisms contribute more greatly to Ig diversity
than previously appreciated (Jenne et al., 2003). While somatic hypermutation does
play a role, reassessment of the original data suggests that it occurs mostly after birth,
when the animal is exposed to foreign antigens.
Figure 10. Construction of an immunoglobulin heavy chain.
A simplified view; the actual number of available gene segments is largerand speciesdependent. V: variable segment, D: diversity segment, J: joiningsegment, C: constant segment specifying the antibody isotype. Based onJaneway et al. (2001) and Tizard (2004).
Cattle are thought to generate their antibody repertoire using strategies similar to
sheep, although available data are even more limited (Zhao et al., 2006; Kaushik et al.,

Generation of B lymphocytes in ruminants
29
2002). A small number of utilized Ig gene segments have been detected (Sinclair et al.,
1997; Saini et al., 1997; Berens et al., 1997). Both somatic hypermutation and gene
conversion have been proposed to be involved in the Ig diversification (Parng et al.,
1996; Berens et al., 1997; Lucier et al., 1998). CDR3, one of the highly variable
complementaritydetermining regions of Igs, shows marked length heterogeneity in
cattle Ig heavy chains (Berens et al., 1997; Sinclair et al., 1997; Saini et al., 1997; Saini
and Kaushik, 2002). In a substantial proportion of bovine antibodies, the CDR3 is
longer than found in any other species (Saini et al., 1999), at least partially due to
exceptionally long diversity (D) segments in the germline genome (Shojaei et al., 2003).
This is thought to provide additional material for somatic hypermutation in the
diversification of bovine immunoglobulins (Kaushik et al., 2002).
4.3.2. The ileal Peyer’s patch
Lymphoid organs called Peyer’s patches (PPs) exist in the walls of the mammalian
small intestine (Yasuda et al., 2006; Griebel and Hein, 1996). They are accumulations of
lymphoid follicles, with considerable variation in structure and distribution between
species. In ruminants, pigs, horses, dogs and humans, most of the follicles are
contained in a single ileal PP, while in jejunum multiple discrete accumulations are
found (Griebel and Hein, 1996). PPs are generally responsible for mucosal immunity. In
the mammalian GALT species, however, the large ileal PP is important for the entire
peripheral B cell pool (Reynolds, 1997).
The lymphoid follicles in the PPs extend from the lamina propria to the submucosa
of the intestine (Fig. 11). In the ileal PP, they are larger and more cylindrical than in the
jejunum (Reynolds and Morris, 1983b). A dome covered by a specialized follicle
associated epithelium protrudes towards the mucosa; this is more prominent in the
follicles of jejunal PPs. Each follicle is encapsulated by connective tissue and
surrounded by lymphatic sinuses draining from the intestinal villi (Reynolds and
Morris, 1983b; Lowden and Heath, 1992). The ileal follicles contain mostly B cells
expressing surface IgM, supported by a rich network of stromal cells (Hein et al., 1989;
Griebel and Hein, 1996; Reynolds, 1997).
The ileal PP develops relatively late in the ruminant fetus, as compared with jejunal
PPs and other lymphoid organs. In sheep, it forms by day 110 of the 150day gestation
(Reynolds and Morris, 1983b). In cattle, the first ileal lymphocyte clusters are seen at

Review of the literature
30
day 150, and the follicular structures can be distinguished by 180185 days’ gestation
(Ishino et al., 1991; Beyaz and Asti, 2004; Doughri et al., 1972). The PP is seeded by B
cells, which have already rearranged one Ig light chain allele and express surface IgM
(Jenne et al., 2006; Reynaud et al., 1991; Griebel et al., 1992). The B cells are thought to
originate at least partially from the spleen, although they can also be produced in other
locations (Lucier et al., 1998; Press et al., 2001). The essential population appears to
exist already at day 63, when an injection of antiIgM antibody is able to perturb later
PP development (Press et al., 1996).
By observing Ig light chain rearrangement patterns, sheep ileal PP follicles were
discovered to be oligoclonal, indicating that each follicle is founded by a small number
of B cells (Reynaud et al., 1995). Igs are then diversified by somatic hypermutation.
Based on sequence genealogies, diversification of the light chain variable regions has
been shown to occur in the ileal PP also in cattle; some of the diversity is, however,
already present in the fetal spleen (Lucier et al., 1998).
Growing until 68 weeks after birth, the ileal PP is up to 2.5 meters long in young
lambs and contains about 105 lymphoid follicles; it accounts for up to 1.2% of body
weight, more than the T cellproducing thymus (Reynolds and Morris, 1983b). The
ovine ileal PP starts to involute at about 3 months postnatally and is mostly replaced by
connective tissue by 18 months of age (Reynolds and Morris, 1983b). The jejunal PPs
remain even in old animals.
The follicular B cells in the ileal PP proliferate rapidly already in the fetus. About
5% of them enter metaphase each hour, equalling the production of 3.6 · 109 new cells;
this is approximately ten times the mitotic rate in the thymus (Reynolds, 1986;
Reynolds, 1987). Uncontrolled, the mass of such an expanding population would
exceed that of the whole animal within a week (Reynolds, 1997). About 5% of the B
cells leave the PP and are distributed to all lymphoid tissues, with no preference for the
mucosal sites (Pabst and Reynolds, 1987; Reynolds et al., 1991). Most of the rest are
removed by apoptosis (Pabst and Reynolds, 1986; Pabst and Reynolds, 1986; Motyka
and Reynolds, 1991). A poorly understood selection process may be involved in the
deletion of B cells (Yasuda et al., 2006).
Bovine ileal PP follicles were recently characterized by serial analysis of gene
expression (Neill et al., 2006). Conforming to earlier studies in sheep, the data indicate
active protein synthesis, IgMdominant expression of Ig genes and apoptotic activity.
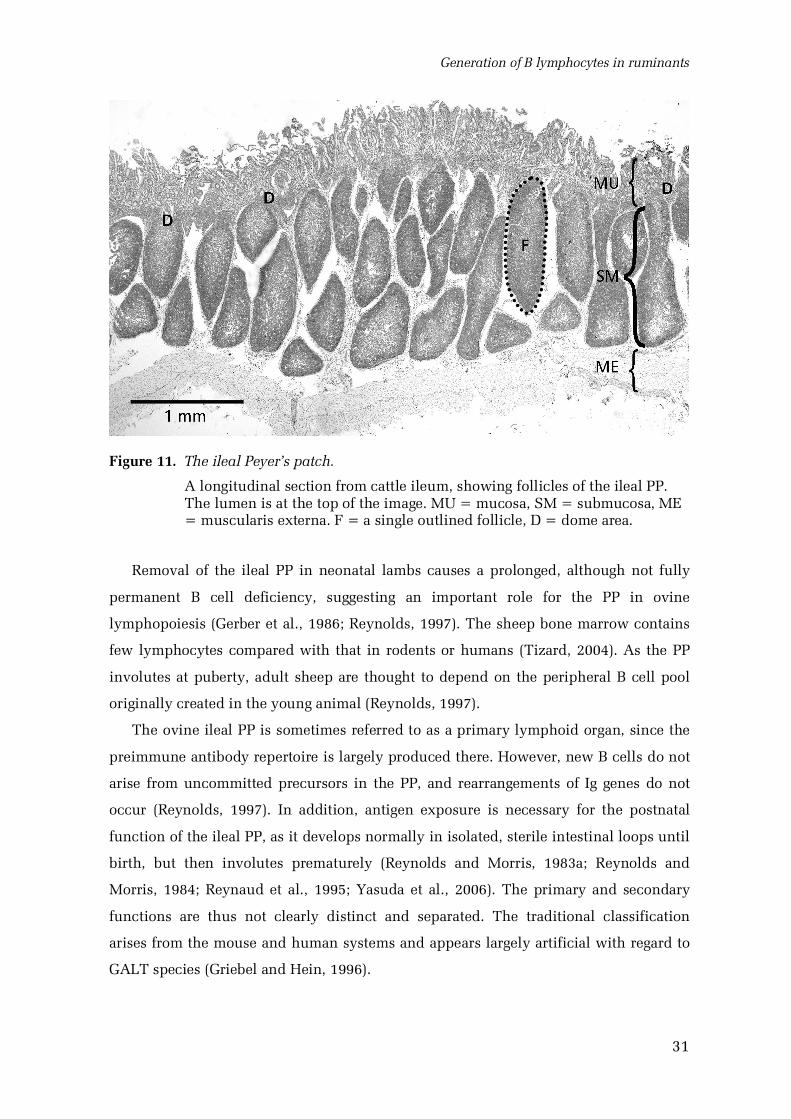
Generation of B lymphocytes in ruminants
31
Figure 11. The ileal Peyer’s patch.
A longitudinal section from cattle ileum, showing follicles of the ileal PP.The lumen is at the top of the image. MU = mucosa, SM = submucosa, ME= muscularis externa. F = a single outlined follicle, D = dome area.
Removal of the ileal PP in neonatal lambs causes a prolonged, although not fully
permanent B cell deficiency, suggesting an important role for the PP in ovine
lymphopoiesis (Gerber et al., 1986; Reynolds, 1997). The sheep bone marrow contains
few lymphocytes compared with that in rodents or humans (Tizard, 2004). As the PP
involutes at puberty, adult sheep are thought to depend on the peripheral B cell pool
originally created in the young animal (Reynolds, 1997).
The ovine ileal PP is sometimes referred to as a primary lymphoid organ, since the
preimmune antibody repertoire is largely produced there. However, new B cells do not
arise from uncommitted precursors in the PP, and rearrangements of Ig genes do not
occur (Reynolds, 1997). In addition, antigen exposure is necessary for the postnatal
function of the ileal PP, as it develops normally in isolated, sterile intestinal loops until
birth, but then involutes prematurely (Reynolds and Morris, 1983a; Reynolds and
Morris, 1984; Reynaud et al., 1995; Yasuda et al., 2006). The primary and secondary
functions are thus not clearly distinct and separated. The traditional classification
arises from the mouse and human systems and appears largely artificial with regard to
GALT species (Griebel and Hein, 1996).

32
5. Objectives of the study
The present work aimed to characterize the differentiation potential of somatic stem
cells in a large, unmanipulated mammal to complement the data obtained from mouse
and human transplantation experiments. We studied naturally chimeric twin cattle,
where a substantial proportion of hematopoietic stem cells originates from the sibling.
The donor contribution to the immune system was also examined to investigate the role
of the ileal Peyer’s patch in the generation and maintenance of bovine B lymphocytes.
The following specific objectives were set:
1. To develop methods for detecting bullderived cells in the tissues of female
twin siblings (freemartins), while preserving tissue architecture (III).
2. To select and develop tools for identifying the phenotypes of bullderived
cells in freemartin tissues (I, V).
3. To use these methods to characterize the distribution and phenotypes of
bullderived cells in freemartin tissues (II).
4. To analyse the distribution of bullderived cells in various leukocyte
populations in freemartins to better understand the function of the ileal
Peyer’s patch in the generation and maintenance of the B cell pool in cattle
(III, IV).

33
6. Materials and methods
Detailed descriptions of methods, including primer and probe sequences, are available
in the original publications.
6.1. Animals, tissues and cells (IV)
Heterosexual cattle twins were located by veterinarian contacts or the cattle database
maintained by the Finnish Animal Breeding Association. The chimerism was
quantitated by Y chromosomespecific in situ hybridization to blood samples, as
described below. Blood samples were acquired from farms. Tissues were collected from
chimeric and normal cattle, which were either anaesthesized and exsanguinated at the
facilities of the Department of Basic Veterinary Sciences or slaughtered routinely at an
abattoir. Altogether 12 freemartins aged 20 days to 20 months were studied.
To obtain newly generated granulation tissue, blockshaped Cellspon cellulose
viscose sponges (Cellomeda, Finland; Viljanto and Kivikoski, 1962) were implanted
subcutaneously in the prescapular region of a 70dayold freemartin calf. Five sponges
were implanted at oneweek intervals. Operations were performed under surgical
anaesthesia, by A. Iivanainen. The calf was sacrificed and implants recovered one week
after the last operation.
Tissue samples were processed for paraffin sections or cryosections. For paraffin
sections, they were fixed either in 4% phosphatebuffered paraformaldehyde (PFA) for
24 h at +4 C or in 100% ethanol for 2 h at +4 C followed by 120 h at 20 C, embedded
in paraffin and cut to 2 to 4 m sections on Superfrost Plus slides (MenzelGläser,
Germany). For cryosections, the samples were frozen in liquid nitrogen, stored at 80 C
and cut to 10 m sections.
Leukocytes were collected from heparinized blood samples and bone marrow
suspensions by Ficoll gradient centrifugation. Erythrocytes were lysed where necessary.
Bone marrow was obtained from sternal pieces from slaughtered animals or as aspirates
from the wing of the ilium, taken under surgical anaesthesia prior to exsanguination.
For isolation of cells from the spleen, lymph nodes and thymus, tissue samples were
cut into small pieces in medium, and the released cells filtered through a Cell Strainer
(Becton Dickinson Labware, NJ) and centrifuged on a Ficoll gradient. Lymphocytes
from the ileal mucosa were isolated as described previously (Parsons et al., 1996).

Materials and methods
34
Cells were generally processed for cytocentrifuge preparations at 105 cells per slide,
fixed with ethanol or acetone, airdried and stored at 80ºC. Smears were prepared from
bone marrow aspirates.
6.1.1. Animal welfare (IV)
The use of animals was approved by the local authorities. Care was taken to minimize
any pain and suffering, and mostly postmortem material was used.
6.2. In situ hybridization
6.2.1. Genomic in situ hybridization (IIIV)
Optimized in situ hybridization (ISH) protocols were developed for the detection of the
Y chromosome in bovine tissue sections and cytocentrifuge preparates. For tissue
material, paraffin sections were used. The sections were deparaffinized and then
permeabilized by detergent treatment, microwave heating in 2×SSC pH 6, and protease
P6911 (for PFAfixed material; SigmaAldrich, MO) or detergent and protease only
(ethanolfixed material). Protease concentration (5250 g/ml) was optimized for each
tissue. The sections were then quickly fixed in PFA, dehydrated in graded ethanols,
and airdried. Genomic DNA was denatured by heating slides at +85 C after applying
hybridization solution and coverslips. Hybridization was carried out for 40 h in
humidified chambers at room temperature. The hybridization solution contained 7.515
nM digoxigenin (DIG)labelled oligonucleotide probe (see below), 50% formamide,
4×SSPE pH 7.4, 1 mg/ml ssDNA and 5×Denhardt's solution. The hybridized sections
were washed in 50% formamide, 4×SSPE and in 1.5×SSPE, at room temperature. DIG
labelled probe was detected by alkaline phosphataseconjugated antiDIGFab fragments
(Roche, Germany) and visualized using the NBT/BCIP chromogen (Roche). Sections
were lightly counterstained with Mayer's haematoxylin and mounted, usually with
Faramount (Dako Cytomation, Denmark).
In most cases, the slides were assembled in Shandon Coverplates (ThermoElectron),
with the exception of the hybridization steps. Normal bull and cow tissues and cells
were used as positive and negative controls in the analysis of freemartin material.
Cytocentrifuge preparates and bone marrow smears were processed essentially
similarly, with optimized pretreatments and an overnight hybridization.

In situ hybridization
35
6.2.2. Tyramide amplified mRNA in situ hybridization (V)
To analyse the expression of the CD34 gene in bovine tissues, ISH to mRNA was
performed. Fresh cryosections were used for this purpose. The sections were heated at
+60 C for 1 min and air dried. They were fixed in icecold 4% PFA pH 7.4 for 1 hour,
acetylated, incubated in 0.2 M HCl for 6 min, treated with 0.5 – 1 g/ml proteinase K for
30 min at room temperature, dehydrated, and air dried. The probes were diluted at 200
400 pg/ l in a hybridization buffer containing 50% formamide, 4×SSPE pH 7.4, 1
mg/ml ssDNA, and 5×Denhardt’s, denatured, and hybridized to the tissue sections at
+52 C overnight. The slides were washed in 4×SSPE; 4×SSPE, 50% formamide; and
1.5×SSPE, at +52 C. They were then assembled with Shandon Coverplates. The
sections were incubated in a blocking reagent and antiDIGperoxidase conjugate (both
Roche). Tyramide amplification was performed using the TSA Plus DNP alkaline
phosphatase system (Perkin Elmer, MA), with the NBT/BCIP substrate (Roche). The
sections were then counterstained with haematoxylin and mounted.
6.2.3. Probes (IIV)
The Y chromosome probe, designed by A. Iivanainen, was a degenerate 28mer
oligonucleotide [5'TT(A/C/T) TCA GCC CTG TGC C(C/T)T GG(A/C/G/T) (A/G)A(C/T)
TGT G3'] corresponding to the repetitive bovine btDYZ locus (Perret et al., 1990). As a
positive control, a previously described oligonucleotide probe (Modi et al., 1993) for the
bovine autosomal 1.709 satellite sequence (Skowronski et al., 1984) was applied. Both
probes contained a single DIG label and were synthesized by Proligo (France).
DIGlabelled antisense riboprobes and sense control probes were prepared by in
vitro transcription from plasmids containing the respective cDNA sequences, using T7
and SP6 polymerases and DIG11UTP (Roche, Germany). The CD34 probe consisted of
the sequence for the extracellular part (bases 57859). The DNA template was removed
by RQ1 DNAse (Promega, WI), and the probes purified by ethanolLiCl precipitation.
Products were checked by agarose gel electrophoresis and by a UV absorption spectrum
analysis using a Nanodrop spectrophotometer (Nanodrop Technologies, DL).

Materials and methods
36
6.3. Immunostaining
6.3.1. Antibodies and antigen retrieval (IV)
We evaluated the usefulness of 39 antibodies and lectins for the identification of major
cell types in paraffinembedded cattle tissues. Antibodies raised against bovine antigens
or known to be bovine crossreactive were used where possible. If no information on
bovine reactivity was available, antibodies with the broadest species crossreactivity
were selected. A comprehensive list of these markers can be found in openaccess
publication I. Some additional antibovine leukocyte antibodies were used in
publication IV, as described in Results. An antibovine CD34 antibody was generated as
a part of the work (V); all other antibodies and lectins are commercially available.
Immunostaining with each antibody was optimized with regard to tissue fixation
(PFA or ethanol) and antigen retrieval methods, as described in publication I. Each
antibody was tested with a standard test battery of retrieval procedures. This included
microwave heating in an acid, neutral or alkaline buffer (for PFAfixed material) and
protease treatments (for PFA and ethanolfixed material). The specificity and
sensitivity of the staining in various tissues were evaluated, and the fixation and
antigen retrieval methods yielding the best results selected for subsequent use.
6.3.2. Immunohistochemistry (IV)
Immunohistochemistry (IH) was performed using either the avidin biotin complex
(ABC) method or tyramide amplification, in most cases applying Shandon Coverplates.
Paraffinembedded sections were dewaxed, rehydrated, subjected to an optimized
antigen retrieval procedure and permeabilized with Tween20 in PBS. The sections
were then blocked for endogenous biotin, when necessary, and for nonspecific binding
with goat serum in PBS. They were incubated in the primary antibody overnight at
+4 C, in PBS containing 1% BSA, washed and incubated with goat biotinylated anti
mouse or antirabbit secondary antibody (Dako, Denmark) for 2 h at room temperature.
The ABC detection was performed using the Vectastain Elite ABC kit and the
diaminobenzidine substrate kit (both Vector Laboratories, CA) according to
manufacturer’s instructions. For tyramide amplification, sections were incubated in
avidin D conjugated peroxidase (Vector), in biotinylated tyramide (Hopman et al.,

Immunostaining
37
1998), again in avidinperoxidase and in diaminobenzidine substrate. The sections
were counterstained with Mayer’s haematoxylin and embedded with Faramount.
6.3.3. Immunofluorescence (V)
Immunofluorescence staining was performed essentially as described above for IH.
Secondary antibodies conjugated with Alexa Fluor dyes (Invitrogen, OR) were used.
Autofluorescence was suppressed where necessary by incubation in 0.1% Sudan Black
B (Merck, Germany) in 70% ethanol. For cytocentrifuge and smear preparates, no
antigen retrieval was performed, and antibody incubations were shortened. The slides
were embedded using Mowiol 4088 (Aldrich, Germany).
6.4. Combined ISH and IH double staining (II)
Combining ISH and IH was necessary to reliably analyse the phenotypes of bullderived
cells. This was problematic, as the ISH protocol destroyed many of the antigens, and
the IH reaction products caused false negatives in ISH. As a solution, tyramide
amplification was utilized in IH, depositing a covalently bound label to the target before
the colour reaction. Thus, the immunodetection could be started first. After the
tyramide reaction, ISH was performed, and thereafter the IH protocol was finished.
Diaminobenzidine (Vector Laboratories) was used as the peroxidase substrate. Sections
were counterstained with Mayer’s haematoxylin and mounted with Faramount (Dako).
For lectin histochemistry, the ISH protocol was beneficial. In combined genomic
ISH and lectin histochemistry, the lectin staining was therefore performed after ISH.
6.5. Microscopy and photomicrography
The stained preparates were viewed and documented using an Olympus BH2
microscope (Olympus Life and Material Science Europa, Germany) equipped with a
ColorView 12 digital camera and AnalySIS 3.0 image analysis software (both Soft
Imaging System, Münster, Germany), or a Leica DM4000 microscope (Leica
Microsystems, Germany) with an Olympus DP70 camera and Cell^P software (both
Olympus).

Materials and methods
38
6.6. Cell counting and statistics (II, IV)
To determine the frequency of Y chromosomecontaining (Y+) cells, several tissue
sections or cytocentrifuge preparates were hybridized, and the samples with the highest
Y+ cell frequencies were selected. They provided the most reliable data, as no false
positives were detected in any of the control female samples (see Results).
6.6.1. Counting of donorderived nonhematopoietic cells in tissues (II)
Y+ and Y– nuclei were counted in digitally photographed tissue sections doubly stained
for the Y chromosome and the hematopoietic marker CD45. The touch count tool of
AnalySIS 3.0 image analysis software was used. Nuclei in all cells negative for
hematopoietic markers in a field of 230 m × 185 m were counted. The process was
repeated for additional fields until the cumulative percentage of Y+ nuclei no longer
fluctuated more than 0.2 percentage units. If no nonleukocyte Y+ cells were found, the
whole section was viewed, and the total number of screened nuclei was estimated by
measuring the area viewed. It should be noted that for tissues with multinucleated
cells, such as the skeletal muscle, the Y+ count represents the proportion of donor
derived nuclei, rather than entire cells.
Numbers were corrected with counts obtained from similar bull tissues. All
statistical analyses were performed in SPSS 10.0 for Windows. MannWhitney U test
was used for estimating statistical significance of differences in cell frequencies.
Similarity of the implanted animal to the other freemartins was evaluated by comparing
scatterplots of cell counting results from different animals.
6.6.2. Counting of donorderived cells in leukocyte fractions (IV)
Y+ and Y– cells were counted in cytocentrifuge preparates processed for genomic ISH.
A total of 300500 cells were counted from each slide until the variation in the
cumulative percentage was less than 2 percentage units. Accuracy of counting was
controlled by analysing leukocytes from normal cows and bulls mixed in known
proportions.

PCRbased IgH analysis
39
6.7. PCRbased IgH analysis (III)
Genomic PCR was used to analyse the size of complementaritydetermining region 3
(CDR3) within the IgH locus, in individual ileal Peyer’s patch (PP) follicles. CDR3 size
can be used to mark a single B cell clone (Arnold et al., 1983). The assay is especially
feasible in cattle due to the large variation in the CDR3 size (Saini et al., 1999).
6.7.1. Laser capture microdissection
Individual ileal Peyer's patch follicles were laser microdissected from membrane
mounted, light greenstained, PFAfixed paraffin sections of freemartin ileum using a
PALM laser microscope system (P.A.L.M. Microlaser Technologies, Germany),
essentially as described previously (Bohm et al., 1997).
6.7.2. Hybridoma culture
For controls, two bovine B cell – mouse myeloma hybridoma lines ( BL5C2.870005 and
BL5C2.870009) were obtained from ATCC. These were cultured in Dulbecco's
modified Eagle's medium with 4.5 g/l glucose, supplemented with 10% horse serum.
6.7.3. Preparation of genomic DNA templates
The microdissected follicles were digested with proteinase K at +56 C for 57 days. The
enzyme was inactivated by heating. The crude digest was then used as PCR template.
For controls, genomic DNA was isolated from bovine peripheral blood leukocytes and
from the two B cell hybridoma lines.
6.7.4. PCR amplification and analysis of products
A region containing the hypervariable CDR3 segment of the rearranged bovine IgH
locus was PCR amplified. Primer cocktails were designed based on bovine IgH
sequences in GenBank. The 5' primer was specific to the framework 3 region of the IgH
gene, and the 3’ primer was specific to the framework 4.
The PCR products were analysed by agarose gel electrophoresis. The specificity of
products was confirmed by Southern blotting using a DIGlabelled internal framework
3 probe. Alkaline phosphataseconjugated antiDIGFab fragments (Roche) and the CDP

Materials and methods
40
Star substrate (Roche) were used for detection. Results were visualized using the
Molecular Imager phosphor imager (BioRad, Hercules, CA).
6.8. Leukocyte fractionation (II, V)
Magneticactivated cell sorting (MACS) was used to isolate various lymphocyte
fractions from Ficollisolated mononuclear cells and to deplete bone marrow cells for
lineage markers. MACS columns (Miltenyi Biotec Inc., Germany) were used according
to the manufacturer’s instructions for indirect separation. The efficiency of separation
and depletion was controlled by FACS after incubation of the cells with a fluorescently
labelled secondary antibody.
Granulocytes were isolated using Ficoll gradient centrifugation. They were obtained
from the cell fraction under Ficoll by lysis of the red blood cells.
Cell fractionations were performed by T. PessaMorikawa.
6.9. Generation of a polyclonal antibovine CD34 antibody (V)
6.9.1. Cloning of the fulllength CD34 cDNA
Fulllength CD34 cDNA was produced by RTPCR from total RNA of bone marrow
mononuclear cells. The total RNA was isolated by the singlestep guanidium
isothiocyanate method (Kingston et al., 1996).
The first strand was synthesized using MMuLV reverse transcriptase (Finnzymes,
Finland) and a primer specific for the 3’UTR (Zhou et al., 2001a). The product was
amplified in seminested PCR using DynaZyme EXT polymerase (Finnzymes) and
primers specific for the 5’UTR and 3’UTR of bovine CD34.
The cDNA was ligated into a SacII/SalI digested pBluescript II SK plasmid
(Stratagene, CA), and the recombinant plasmid was used for transforming Escherichia
coli. Five clones were sequenced at the DNA sequencing laboratory of the Viikki
campus. A fulllength sequence conforming to previously published data (Zhou et al.,
2001a) was then assembled from these clones.
6.9.2. Expression and purification of polypeptide fragments
The following three fragments of the CD34 polypeptide were selected for antigen
expression based on structure analysis with several prediction software programs: the

Generation of a polyclonal antibovine CD34 antibody
41
predicted extracellular domain, a part of the extracellular domain without most of the
predicted glycosylation sites and the predicted cytoplasmic domain.
The corresponding DNA sequences were PCR amplified from the cDNA plasmid and
ligated into a NcoI/XhoI digested pET22b expression plasmid (Novagen, WI)
containing the T7lac promoter and sequences for the pelB periplasmic localization
signal and Cterminal 6×Histag. The plasmids were then sequenced to exclude
potential PCR errors.
Protein expression was performed using E. coli strain BL21(DE3) (Novagen). The
partial extracellular fragment and the cytoplasmic fragment were selected for large
scale production, as the complete extracellular domain was found insoluble. Bacterial
cells were cultured in LuriaBertani medium with 100 g/ml ampicillin, and protein
expression induced with 1 mM isopropyl1thio dgalactoside. The cytoplasmic
fragment was collected from the supernatant, and the extracellular fragment from the
supernatant and the periplasmic space. Both fractions were dialysed to remove free
histidine and histidinecontaining oligopeptides.
The Histagged polypeptides were purified by metal affinity chromatography, using
the NiNTA Superflow nickelnitriloacetic acid resin (Qiagen, MD). The polypeptides
were bound to the resin, washed with buffer containing 120 mM imidazole and eluted
with 250 mM imidazole.
The fractions were quantitated with the BCA assay and analysed using SDSPAGE
electrophoresis. Identities of the polypeptides were confirmed by Nterminal
sequencing and mass spectrometry from reversedphase highperformance liquid
chromatography fractions, at the protein chemistry core facility of Viikki campus.
6.9.3. Antibody production
Rabbit polyclonal antibodies were raised against the CD34 polypeptide fragments. The
antibodies were produced by Sigma Genosys (Cambridge, UK) according to a standard
custom protocol. They were then purified by affinity chromatography on antigen
coupled to NHSactivated Sepharose (Amersham, Sweden).
6.10. Western blot (V)
Samples of homogenized bovine tissues were run on SDSPAGE gels, transferred to
polyvinylidine difluoride membranes and probed with the CD34 antibody. To test

Materials and methods
42
specificity, free polypeptide antigen was included in some experiments. After
incubation with a peroxidaseconjugated goat antirabbit antibody (Dako Cytomation),
the blots were developed by enhanced chemiluminescence substrates (ECL, Amersham)
and visualized using a Fuji LAS3000 camera (Fuji Photo Co., Ltd., Japan). T. Pessa
Morikawa was responsible for the Western blots.
6.11. Analysis of the bovine CD34 gene and mRNA expression (V)
6.11.1. Expression of CD34 mRNA
The possible alternative splicing of the CD34 mRNA in cattle was first examined by
bioinformatics. The tentative intron 7 splice sites in the bovine CD34 gene were
predicted by a CLUSTAL W (Thompson et al., 1994) alignment of the human and cattle
genomic sequences (Genbank accessions NT_021877 and NW_929130). This part of the
work was performed by A. Iivanainen.
The expression of CD34 mRNA in various bovine tissues was analysed using RT
PCR by R. Ra. To detect putative splice variants, a strategy published previously for
murine CD34 was used (Suda et al., 1992). Total RNA was reversetranscribed using the
MMuLV reverse transcriptase (Fermentas), and the products were PCR amplified with
the Dynazyme EXT DNA polymerase (Finnzymes). Primers specific for exons 7 and 8 of
the bovine CD34 gene, enclosing the possible exon 7 extension, were used in these
reactions. Products were then analysed by agarose gel electrophoresis.
In addition, the expression pattern of CD34 mRNA was analysed by in situ
hybridization, as described above.
6.11.2. Assaying single nucleotide polymorphism
A potential single nucleotide polymorphism (SNP) in the bovine CD34 gene was
suggested by a difference in our sequence to the previously published data. The
presence of the SNP was assayed by restriction fragment length in bovine genomic DNA
samples (9 different breeds, 49 animals in total). DNA was extracted from blood
samples using the Wizard genomic DNA purification kit (Promega, WI) or obtained
from the Nordic Gene Bank Farm Animals (the Japanese black cattle sample from
Professor Masahiro Yasuda, University of Miyazaki). The genomic DNA was then PCR
amplified using primers corresponding to bovine CD34 exons 4 and 6. The products
were digested using BshTI and analysed by agarose gel electrophoresis.

43
7. Results
7.1. Detection of bull cells (IIIV)
The optimized Y chromosome in situ hybridization (ISH) protocol was shown to be
both sensitive and highly specific (III / Fig. 1). In bull tissues, approximately 4070% of
cells were strongly labelled, depending on tissue type, even in double staining
combined with immunohistochemistry (II). In the 2 to 4 m tissue sections, the Y
chromosomes were unavoidably excluded from some nuclei. No falsepositive signals
were seen in any of the female control samples (more than 1700 hybridizations using
20 different tissue types). In the counting of bullderived cells on cytocentrifuge
preparates, the results of control experiments with known proportions of bull and cow
cells varied less than 3 percentage units from the actual values (IV / Fig. 2). For
paraformaldehydefixed paraffin sections, effective hybridization required an enhanced
pretreatment with both microwave heating and protease.
7.2. Selection of phenotypic markers (I)
To facilitate the analysis of the phenotypes of the bullderived cells, various markers
were evaluated for immunohistochemistry (IH) and lectin histochemistry. A panel of 31
useful markers was selected. These allowed the identification of all major differentiated
cell types in cattle tissue sections, including epithelia, connective tissue, muscle and
nervous tissue, as well as cell proliferation and apoptosis (I). With paraformaldehyde
fixed material, heatinduced antigen retrieval at a specific pH was beneficial for most
antibodies. By selecting a suitable retrieval protocol, most of these markers could be
successfully applied to this type of material.
7.3. Bullderived cells in freemartin tissues (II, III)
To characterize the distribution of bullderived cells in freemartin tissues, a wide Y
chromosome ISH screening was first performed. Samples from 2074 different body
sites per animal were analysed from 12 freemartins. In total, more than 1650 freemartin
tissue sections were successfully hybridized. Y chromosomepositive (Y+) bullderived
cells were detected in all of the animals in all tissues examined. The frequency of Y+

Results
44
cells was highest in blood, bone marrow and lymphoid tissues, with considerable
variation present between individual animals. In blood mononuclear cells, the
proportion of Y+ cells varied from 10% to 90%. The variation was less pronounced in
other organs.
The distribution of the bullderived cells and double stainings with cell type
specific immunohistochemistry (primarily neuronal markers; unpublished
observations) suggested that a significant proportion of the Y+ cells were infiltrating
leukocytes. These are often difficult to distinguish from parenchymal cells in a
standard histological staining. To reliably label the leukocytes residing in the tissues, Y
chromosome ISH was combined with immunohistochemistry for the panleukocyte
marker CD45 (Thomas, 1989), as described in Methods. In the analysis of the central
nervous system, mistletoe lectin I (MLI) histochemistry was used to label bovine
microglial cells (HewickerTrautwein et al., 1996).
Using double staining, 14 different tissues were examined, representing all major
tissue types and derivatives of all embryonic layers. A total of 472 successfully double
stained freemartin tissue sections were viewed. CD45/MLI– Y+ cells were identified in
most tissues, but in 11 of the 12 calves they were mostly sporadic. Thus, the Y+ cells
were overwhelmingly of the hematopoietic lineage (II / Fig. 1AG). One of the calves
(designated FM10) was a striking exception, with high numbers of CD45/MLI– Y+ cells
in several tissues, as discussed below.
The proportion of Y chromosomepositive CD45/MLI– cells among all CD45/MLI–
cells was determined for 45 calves per tissue. With the exception of FM10, the calves
did not differ markedly in the relative frequency of CD45/MLI– Y+ cells, which
constituted less than 1% of all CD45/MLI– cells in most tissues investigated (II / Fig. 2).
No obvious correlation was observed in the frequency of Y+ cells between blood and
nonhematopoietic tissues, or between various nonhematopoietic tissues in a single
animal.
In 11 of the 12 freemartins, the highest numbers of CD45/MLI– Y+ cells were
detected in the mammary gland connective tissue (0.94 0.67%), liver (0.85 0.41%,
mostly not hepatocytes), and skeletal muscle (0.83 0.26% of nuclei). The average
frequency of CD45–Y+ cells was 0.1% or less in cardiac and smooth muscle, renal
epithelia, and the epidermis. Bullderived nonhematopoietic cells were markedly less
frequent in the ectodermderived epidermis and the central nervous system. In the
brain, MLI– Y+ cells were extremely rare, despite a robust contribution to the microglia

Bullderived cells in freemartin tissues
45
(notably, though, the lectin also labels most endothelia). In the cerebral cortex and the
underlying white matter, 0.0020 0.0023% of MLI– cells were Y+. No MLI–Y+ cells were
detected in the hippocampus or the olfactory bulb, among the approximately 2 105 cells
screened per tissue. No Y+ cells showing unambiguous neuronal morphology were seen
in more than 700 successfully hybridized sections of the freemartin central nervous
system. These included samples from the cerebral cortex, cerebellum, brainstem,
corpus callosum, hippocampus, olfactory bulb, thalamus, ventricular walls, hypophysis
and spinal cord, representing approximately 28 106 cells in total.
In FM10, the distribution of CD45/MLI– Y+ cells differed substantially from the
other calves. Higher numbers of CD45–Y+ cells were detected in the epithelium of small
intestine (19%, II / Fig. 1H), in the connective tissue of mammary gland (15%), and in
liver (4.3%), epithelium of mammary gland (4.0%), skeletal muscle (2.3%), cardiac
muscle (1.0%) and smooth muscle (0.5%). In the brain and renal epithelia, the
frequency did not differ notably from the other calves. Continuous strands of CD45–Y+
cells were seen in the villi of the intestinal mucosa (II / Fig. 1H). In the hematopoietic
system, the bull contribution to B cells was clearly weaker than to the total leukocyte
pool (data not shown).
Newly formed granulation tissue in subcutaneously implanted cellulose viscose
sponges contained relatively high numbers of CD45–Y+ cells (II / Fig. 1I and Fig. 2). The
frequency increased within an incubation period of 4 weeks. At 4 weeks, the frequency
of CD45–Y+ cells in the newly forming tissue was significantly higher than in most
other nonhematopoietic tissues of the same animal (P=0.12 for liver, P<0.02 for any
other tissue investigated). However, no obvious Y+ cell clusters indicating local
proliferation were seen. In the other tissues of the sponge recipient, the frequencies of
CD45/MLI– Y+ cells did not differ markedly from those in the 10 other calves of the
majority type.
7.4. Bullderived cells in freemartin lymphocyte populations (III, IV)
In the ileal Peyer's patch (PP) of freemartin cattle, a striking variation in the frequency
of Y+ cells was observed between individual follicles (Fig. 12 and III / Fig. 2A, B). Some
follicles were almost exclusively of the male genotype, while others were female. The
variation appeared discontinuous, with 2 or 3 discrete intermediate groups of follicles
identifiable based on the frequency of bullderived cells. The mosaic pattern was

Results
46
specific to the ileal PP follicles. In the jejunal PPs (III / Fig. 2C, D), the interfollicular
variation in the frequency of Y+ cells was very modest.
Figure 12. Y chromosomespecific in situ hybridization to the ileal Peyer’s patch of afreemartin calf.
A composite image assembled from several photomicrographs. Theintestinal lumen is at the top of the image. For a description of the structure,see Fig. 11.
To directly estimate the number of founder clones per follicle, the size distribution
of the CDR3 regions of the rearranged IgH genes in B cells isolated from individual
follicles were analysed. A small number of different PCR products were generated from
the ileal follicles, in comparison with reactions from blood cells (III / Fig. 3) or from the
jejunal or splenic follicles (data not shown).
To investigate the significance of the ileal PP for the bovine peripheral B
lymphocyte pool, we compared the frequencies of bullderived cells in freemartin B
cells, T cells and other leukocytes isolated from blood and various lymphoid tissues. In
2 freemartins (designated as FmC and FmD) of the 16 analysed, a striking difference in
the bull contribution was observed between blood B and T cells. The proportion of Y+
cells was only 8% and 9% among B lymphocytes, but as much as 90% and 63% in T
lymphocytes (IV / Fig. 1). Slightly less pronounced yet substantial differences were also
seen in two other calves. In the first two calves, FmC and FmD, the values remained
stable for a followup period of 9 months (IV / Fig. 3). They were also essentially the
same in lymphocytes isolated from the lymph nodes and spleen, and in T cells from the
thymus (IV / Fig. 5). This was reflected by Y chromosome ISH staining of lymphoid
tissue sections, where high numbers of Y+ cells were seen in T cellrich areas, while B
cell follicles were mostly devoid of them (IV). In the ileal PP of these animals, the great
majority of follicles were dominated by female cells. The bull contribution to
granulocytes and monocytes closely matched that of T cells (Fig. IV / Fig. 4). The twin

Generation of a polyclonal antibovine CD34 antibody
47
brother of one of these freemartins was also examined, and similar proportions of Y+
cells were found in the various leukocyte fractions and tissues (IV / Fig. 4).
7.5. Generation of a polyclonal antibovine CD34 antibody (V)
To facilitate a direct analysis of the bull contribution to various stem cell populations in
freemartins, we set out to generate antibodies against the bovine CD34, one of the most
widely used markers for hematopoietic stem and progenitor cells in other species
(Krause et al., 1996).
7.5.1. Cloning of bovine CD34 cDNA
RTPCR of bone marrow mononuclear cell total RNA yielded cDNA clones with a 1149
bp open reading frame. A sequence identical to the previously published bovine CD34
cDNA (Zhou et al., 2001a) was assembled, except for a difference of one nucleotide
(T781 C), changing one amino acid in the polypeptide (Trp261 Arg). This position
was identical in all of our clones. A C781 was identified in all samples analysed from 49
animals, representing 9 different cattle breeds (Finnish Ayrshire, Finnish Holstein
Friesian, Eastern Finncattle, Northern Finncattle, Western Finncattle, Icelandic cattle,
Doela cattle, Danish Red, and Japanese black cattle). This also is in agreement with the
cattle genomic data in GenBank (accession NW_929130).
7.5.2. Expression of CD34 polypeptide fragments
The partial extracellular domain and the cytoplasmic domain of the CD34 polypeptide
were successfully expressed in E. coli, while the complete extracellular domain was
insoluble. The Histagged polypeptides were adequately purified by metal affinity
chromatography, as shown by SDSPAGE (for the partial extracellular domain, see Fig.
13). Mass spectrometry indicated the expected fragment sizes, and Nterminal
sequencing confirmed that the periplasmic signal peptide introduced by the expression
vector was correctly cleaved.
7.5.3. Production and characterization of a polyclonal antibody
A useful rabbit polyclonal antibody against the bovine CD34 was obtained with the
partial extracellular domain of the polypeptide. The antibody was affinitypurified and

Results
48
tested in Western blots and immunostaining of cytocentrifuge preparates and tissue
sections.
The antibody, designated as boCD34ecf, recognized a protein of predicted size in
Western blots, showing broad and hazy bands characteristic of heavily glycosylated
membrane proteins (V / Fig. 4). An expected staining pattern, with positively labelled
blood vessels, was also seen in immunostained paraformaldehydefixed paraffin
sections (V / Fig. 5); a heatinduced antigen retrieval in an acid buffer and a mild
protease treatment were necessary. Some unspecific staining was observed, most
intensely in erythrocytes and the epidermis. The antibody did not label living cells.
Thus, it could not be used in fluorescence activated cell sorting.
Figure 13. Purification of the Histagged extracellular fragment of the CD34 polypeptideby metal affinity chromatography, as seen in SDSPAGE.
Crude culture medium (med), unbound fraction (ub) and fractions elutedwith 10, 20 and 250 mM imidazole are shown. STD: size standard (bandsizes shown in kDa).
7.6. Expression of CD34 mRNA and protein in bovine tissues (V)
We characterized the expression of CD34 mRNA in bovine tissues, and used the
boCD34ecf antibody to investigate the expression at the protein level.
CD34 mRNA was detected by RTPCR in all tissues analysed, including blood
leukocytes (V). Two products of expected sizes were obtained, confirming the existence
of two splice variants as predicted by analysis of genomic data. Tyramide amplified in
situ hybridization showed CD34 mRNA in vascular endothelia (V / Fig. 5O) and
fibroblastlike connective tissue cells.
In Western blots, the approximate molecular weight of the bovine CD34 was 105
115 kDa in the lung and lymph nodes, and 125132 kDa in lysates of the brain, muscle,
spleen and bone marrow cells enriched for lineage marker negative cells (V / Fig. 4). In

Expression of CD34 mRNA and protein in bovine tissues
49
the testis, two bands were present, with sizes of around 118 and 150 kDa. The
expression was weak in lymph nodes and the spleen. All of these bands were blocked
by free antigen (shown in V).
To further investigate the distribution of the CD34 protein in bovine tissues, double
immunofluorescence staining using boCD34ecf and the endothelial marker lectin GSL
IB4 was applied. Vascular endothelia were widely labelled by the antibody (V / Fig. 5).
The staining was generally strongest in blood capillary endothelium, which was CD34
positive in all tissues examined. The endothelium of hepatic sinuses was negative (V).
In larger blood vessels, the expression of CD34 was variable and generally weaker. The
endocardial endothelium was also weakly boCD34ecf positive. In contrast, all
histologically identifiable lymphatic vessels were negative for CD34 (V / Fig. 5D, E).
In cytocentrifuge preparates of bone marrow mononuclear cells, around 7.5% of the
cells were positive for boCD34ecf immunofluorescence (V). The proportion of labelled
cells was increased by enrichment for lineage negative cells (using antibodies against
CD3, IgM, CD14, CD41/61, IgG, granulocytes and red blood cells).
In addition to endothelia and primitive hematopoietic cells, certain other cell
populations were stained by the antibody. Smooth muscle in most vessel walls and
other tissues was strongly positive in the immunofluorescent staining with boCD34ecf.
Large numbers of fibroblastlike cells were labelled in connective tissues (V / Fig. 5B, D,
I). In the brain, cytoplasmic staining was seen in subpopulations of neurons, such as
the cerebellar Purkinje cells and rare neurons in the thalamus and olfactory bulb.
Thymic epithelioreticular cells were stained (shown in V). In addition,
immunofluorescence labelling was associated with spermatids in the seminiferous
tubules, but not with mature spermatozoa or earlier phases of spermatogenesis (V / Fig.
5I, L). In all of these tissues, the staining was blocked with the antigenic polypeptide.
For maximal specificity, the tissues were also stained using tyramide amplified
immunohistochemistry, which allows very dilute solutions of antibodies. By this
method, endothelia and fibroblasts were strongly labelled, but most smooth muscle was
negative or weakly stained, and no boCD34ecf staining could be seen in neurons (V /
Fig. 5A), epithelioreticulocytes and spermatids.
In bovine fetuses of the second trimester (110180 days of gestation), vascular
endothelia were generally intensely stained in immunofluorescence. Liver sinusoids
were negative, as in adult tissues. Large numbers of individual cells were labelled in
fetal liver (V / Fig. 5H).

50
8. Discussion
8.1. Differentiation potential of somatic cells in adult cattle
At the time that this work was started, transplanted bone marrow was reported to
generate nonhematopoietic tissues in mice (Ferrari et al., 1998; Petersen et al., 1999).
We set out to investigate somatic stem cell differentiation potential in an
unmanipulated large mammal. Chimeric cattle twins are a unique model for this
purpose; placental anastomosis allows mixing of blood for most of the fetal period,
resulting in highlevel hematopoietic chimerism (Lillie, 1917; Owen, 1945; IIIV). The
permanent nature of the chimerism proves the exchange of hematopoietic stem cells
(HSCs) by the strictest criteria applied in any model system, the longterm
repopulation, although no specific markers for cattle HSCs are available. In addition,
other stem cell populations circulating in the fetal blood are likely to be exchanged,
such as the mesenchymal stem cells (MSCs; Campagnoli et al., 2001); this was not
directly assayed in the present work. The siblingderived cells observed in twin cattle
thus are the progeny of HSCs and any other cells received during the fetal period.
Due to the probable heterogeneous nature of this originating population, the twins
cannot be used to confirm true plasticity of any somatic stem cell type. Rather, they
may provide information on the differentiation potential of circulating somatic stem
cells at a general level, or potentially reject the plasticity of any stem cell type. This
may seem a drawback compared with the commonly used mouse models. However, the
isolation of absolutely pure and homogenous stem cell populations is uncertain, a
matter of methodological definition, even in mice (Osawa et al., 1996; Wagers and
Weissman, 2004). Processes of tissue maintenance and regeneration in small shortlived
rodents may differ from those in large animals, and manipulations necessary for the
induction of artificial chimerism may affect the results. Thus, naturally chimeric cattle
twins valuably complement the data obtained in mouse experiments and clinical
human transplantations.
8.1.1. Evaluation of methods
Y chromosomespecific in situ hybridization (ISH) was shown to be a powerful method
for tracing the bullderived cells in freemartin tissues. In contrast to cytogenetic

Differentiation potential of somatic cells in adult cattle
51
techniques previously used (Wilkes et al., 1981), ISH is not restricted to dividing cells,
and tissue structure is preserved. The method was demonstrated to be highly specific
and sufficiently sensitive (II, III). The high signal intensity allowed rapid screening of
large numbers of cells. Furthermore, quantification of bullderived cells on
cytocentrifuge preparates was highly accurate (IV).
Identification of the phenotypes of donorderived cells is complicated by the narrow
range of markers available for cattle research, as compared with mouse or human tools.
In the present work, we were able to assemble a panel of antibodies and lectins
allowing identification of all major differentiated cell types in paraffin sections of
bovine tissues (I). While only some of them were fully exploited in the context of this
work, they provide a basis for further investigations. Few bovine stem cell markers are
available. Hence, we generated the first antibovine CD34 antibodies, as discussed
below.
Ordinary transmitted light microscopy was used to analyse the localization of the
genetic and phenotypic markers. Serial sectioning or confocal microscopy is generally
regarded as necessary for unambiguous marker colocalization. In this work, however,
the double staining for the Y chromosome and hematopoietic markers was used as
negative selection, to identify the nonhematopoietic bullderived cells (II). Possible
false positives thus did not affect the results essentially.
Cell fusion has recently been implicated as a major cause of apparent plasticity
(Wagers and Weissman, 2004). Multinucleated foreign body giant cells with both Y
chromosomepositive and negative nuclei were seen in the granulation tissue (II).
However, our attempts of assaying nuclear fusion events between bullderived cells and
female cells were unsuccessful due to technical limitations in the labelling of other
chromosomes (unpublished observations). Chromosomal probes suitable for such
experiments should be developed in the future.
8.1.2. Implications of bullderived cells in freemartin tissues
An abundance of bullderived (Y+) cells was observed in most freemartin tissues,
including the intestinal mucosa, liver, and brain (IIIV). However, double staining with
hematopoietic markers indicated that a vast majority of them were infiltrating
leukocytes (II). In most cases, clearly less than 1% of all nonhematopoietic (CD45/ML
I– Y+) cells in a tissue were of bull origin. The lack of donor contribution was especially

Discussion
52
striking in the central nervous system, although a significant proportion of microglia
were bullderived. This contrasts with several reports of bone marrow cells generating
neurons (Mezey et al., 2000; Brazelton et al., 2000; Priller et al., 2001). Less than
0.005% of all nonhematopoietic cells were Y+ in the cerebral cortex. The hippocampus
and the olfactory bulb are involved in the generation of new neurons in adult mammals
(Altman and Das, 1965; Lois and AlvarezBuylla, 1994); not a single bullderived non
hematopoietic cell was detected in these areas. No groups of nonhematopoietic Y+ cells
indicating local clonal expansion were seen in any tissues in the majority of calves.
One of the freemartins (FM10) showed an exceptionally high frequency of donor
derived cells in several tissues, especially in the intestinal mucosa and the mammary
gland, but not in the brain or the renal epithelia (II). In the intestinal villi, strands of
CD45–Y+ cells were observed, suggesting that some of the crypt stem cells were bull
derived. This was surprising, as the intestinal epithelium is thought to be derived from
the primitive gut endoderm (Trier and Moxey, 1979; Ponder et al., 1985). A similar
engraftment has, however, been reported with intravenously injected multipotent adult
progenitor cells (MAPCs) in immunodeficient mice (Jiang et al., 2002a). On the other
hand, the mammary gland is a highly plastic organ, theoretically a candidate target for
circulating stem cells. In the infertile freemartins, however, the glands obviously are
not engaged in the cyclic expansion and involution associated with pregnancy and
lactation.
The reasons for the remarkable donor contribution in the single freemartin (1 of 12)
are unknown. It was of the same breed as the majority of the other freemartins. While it
was the oldest animal examined (20 months), the difference in age to the closest cases
was modest (only 4 and 6 months). The animal was apparently healthy (unpublished
observations), but an earlier insult on the intestinal epithelium or other tissues could
have gone undetected. Genetic deficiencies in the freemartin cells could provide donor
cells with a competitive advantage in some tissues; in absence of such data, this
remains speculative. A very early formation of vascular anastomoses in the fetus might
result in an exchange of more multipotent primitive cells than available later, but this is
unlikely to occur, as discussed in study II. Instead, a rare fusion of early embryos could
result in substantial chimerism in random tissues. The Y+ cells in FM10 could originate
from either the surviving twin brother, or a third embryo lost before birth. In the first
case, the embryonic event would be impossible to retrospectively distinguish from the
effects of placental anastomosis, whereas triplets would be detectable by careful genetic

Differentiation potential of somatic cells in adult cattle
53
analysis. Intriguingly, spontaneous chimerism resulting from embryonic cell exchange
was recently proposed to have a 10% incidence in humans (Boklage, 2006). The high
bull contribution in FM10 may thus be explained by factors other than placental
anastomoses.
Our results indicate that HSCs (and any other somatic cell type circulating in the
fetal blood) generally do not produce significant numbers of nonhematopoietic cells in
the development, growth or turnover of bovine tissues. HSC plasticity does not appear
as a major physiological phenomenon, and the same is probably true for MSCs. It is
possible that a stronger contribution of donorderived cells would be seen in aged
animals. However, this is unlikely, as no obvious correlation was observed between age
and the level of chimerism in calves from 20 days to 20 months old (II).
In contrast, the progeny of circulating cells may be important in tissue repair and
regeneration. To begin to investigate such processes in cattle, we induced the formation
of granulation tissue in subcutaneous cellulose viscose implants (II). The proportion of
nonhematopoietic Y+ cells was markedly high in the newly forming granulation tissue,
and increased over time. Apparently, cells are recruited from the circulation or adjacent
tissues, from a population with a higher frequency of Y+ cells than in the intact
subcutaneous tissue. They probably do not proliferate extensively, as no clusters of
bullderived cells were seen. The origin of cells in granulation tissue is controversial,
with evidence for both local (Ross et al., 1970; Rangan, 1967) and systemic (Petrakis et
al., 1961) sources. Peripheral blood cells positive for vimentin, fibronectin, collagen,
CD45 and CD34 have been implicated in tissue repair (Abe et al., 2001). They could be
derived from the HSCs, a major proportion of which must be bullderived in
freemartins. In the present study, though, the leukocyte lineages were excluded due to
the CD45 expression, unless it is downregulated after extravasation. MSCs would be
another candidate source, although the frequency of Y+ cells in this population has not
been determined.
Notably, the relatively short followup time of 4 weeks does not allow us to
conclude that the donorderived cells stay in the repaired tissue permanently. Instead,
they may be accessory cells transiently promoting the repair process. Compatible with
this hypothesis, the frequency of Y+ cells around the fully healed ear tag holes was
comparable with that of the surrounding tissues (unpublished observations).
No comprehensive analysis with various cell typespecific markers was carried out
in this work due to the generally low frequencies of donorderived cells. Thus, the exact

Discussion
54
phenotypes of the CD45/MLI–Y+ cells in freemartin tissues were not determined. The
morphology and spatial organization of donorderived nonhematopoietic cells suggest,
however, that they are primarily fibroblastlike cells, pericytes, possibly myocytes
and/or associated satellite cells, and occassional epithelial cells (II). No cells with an
unambiguously endothelial phenotype were found. A majority of the donorderived
nonhematopoietic cells may represent interstitial cell populations, and integration into
parenchymal populations appears rare. The phenotypes of Y+ cells in the granulation
tissue will be characterized in the future.
The results from the present work are in agreement with several recent reports
questioning the plasticity of HSCs and other bone marrow cells (Castro et al., 2002;
Massengale et al., 2005; Roybon et al., 2006; Wagers et al., 2002; Murry et al., 2004). In
other studies, markedly higher contribution of transplants to nonhematopoietic tissues
was observed, as discussed in the literature review. Especially in the central nervous
system, engraftment rates in our material were strikingly lower than those reported in
some earlier experiments. These disparities may be explained by the effects of
transplantation procedures, such as radiation damage to the bloodbrain barrier,
variable criteria for defining the donorderived cells or species differences. The MAPCs,
convincingly documented as highly multipotent (Jiang et al., 2002a), may not have been
exchanged through fetal blood, although this would be expected due to the probable
MSC circulation. Alternatively, they could indeed be nonexistent in vivo, arising
through the culture methods used to isolate them.
Our results do, however, support the findings that injury stimulates the contribution
of bone marrow to nonhematopoietic tissues, at least in the short term (Petersen et al.,
1999; LaBarge and Blau, 2002; Okamoto et al., 2002). Due to the very restricted sample
size in this work, these processes in freemartins warrant further investigation.
The low level of chimerism in nonhematopoietic tissues of freemartins should
enable the transplantation and followup of such cells or tissues from their male twins,
naturally tolerated by the recipient. The cattle twins may thus be used as a large animal
model for investigations of nonhematopoietic stem cell systems.

Role of the ileal Peyer’s patch in generation of the B cell pool in cattle
55
8.2. Role of the ileal Peyer’s patch in generation of the B cell pool incattle
In study III, we report that the bovine ileal Peyer’s patch follicles are oligoclonal. This
was clearly shown by the discontinuous variation in the proportion of Y+ cells in the
ileal Peyer’s patch follicles, together with the small number of different PCR products of
the rearranged IgH loci obtained from single follicles. Apparently, each follicle is
founded by a few B cells only, and later entry is prevented, as the oldest animals in the
study were about 1.5 years of age. Theoretically, a restricted CDR3 length
polymorphism could be produced by an intense selection process, but this would
probably not affect the distribution of bullderived cells in the observed manner. In
contrast, oligoclonality of lymphoid follicles was not observed in the jejunal PPs or
other lymphoid organs.
Surprisingly, a very uneven bull contribution to B and T lymphocytes was found in
some freemartins (IV). In the two animals studied in detail, the proportion of Y+ cells in
B cells was drastically lower than in T cells and other leukocyte populations. The
difference was uniform in all lymphocyte compartments and stable for at least 9
months.
The mechanisms establishing and maintaining the disparity are unclear. The
common dominance of either bull or cow cells throughout the hematopoietic system of
freemartins may be explained by subtle differences in the speed of early fetal
development. Fetal liver is colonized by HSCs around days 2732 of gestation (Rüsse,
1991). This coincides with the earliest vascular anastomoses detected at about day 30,
immediately accompanied by chimerism in the liver (Ohno and Gropp, 1965). Cells
from a slightly more advanced fetus may thus have the opportunity to colonize the
whole conjoined system. However, the timing alone does not readily explain the minor
donor contribution specifically to the B cell pool. The bullderived hematopoietic
progenitors may have had a weaker tendency to commit to the B lineage, or they could
have been less effective in the population of the PP follicles. Alternatively, they could
have been inferior in their capacity for rapid proliferation or for the survival of the
selection processes. Testing these hypotheses would require the isolation of
uncommitted progenitors in this type of freemartins and their brothers and a
comparison of their B cell generation potential in vitro or in transplantation models.
The observations are also compatible with a selfsufficient pool of cells committed
to the B lineage. The stability of the differences between lymphocyte populations may

Discussion
56
indicate that few B cells are continuously generated from HSCs or common lymphoid
progenitors in young cattle (see Fig. 1). As the majority of all other leukocytes
investigated were bullderived, the donor contribution to HSCs was probably
substantial. This would be expected also for de novo produced B cells, unless the B
lymphopoietic potency of the donorderived progenitors is persistently severely
inferior.
Our results indicate a special role for the ileal PP in the production of B
lymphocytes in cattle, and suggest that it may be responsible for the generation and
maintenance of the bovine peripheral B cell pool in juvenile animals. Moreover, the
limited number of IgH rearrangements in the PP follicles is compatible with the view
that cattle rely on postrearrangement mechanisms in the generation of antibody
diversity. Thus, cattle appear largely similar to sheep with regard to strategies for B
lymphopoiesis. For a detailed description of bovine B lymphocyte ontogeny, further
studies are necessary. The sites where ruminant B cells are originally generated from
uncommitted precursors remain unknown. The production of B cells in aged cattle has
also not yet been investigated.
8.3. Generation of an antibovine CD34 antibody (V)
Direct investigation of various stem cell populations requires specific markers, which
are scarcely available for cattle. To expand the tool kit, the first antibodies against
bovine CD34 were generated in the present work. This sialomucin is commonly used as
a marker for purification of human hematopoietic HSCs (Krause et al., 1996). CD34
enriched cells have also been reported to contribute to several nonhematopoietic
tissues (Krause et al., 2001). Although CD34 is hardly a perfect stem cell marker, as
discussed in the literature review, it certainly is one of the best known. Thus, it is a
reasonable starting point for stem cell investigations in cattle and provides a basis for
comparative studies. In addition, CD34 is useful as a vascular marker, being expressed
in endothelial cells from the earliest stages of development to adulthood (Young et al.,
1995).
Polypeptide fragments produced in E. coli were used as immunogens, although
bacteria are unable to produce the heavy glycosylation of the native CD34 protein
(Krause et al., 1996). The glycosylation is cell typespecific (Shailubhai et al., 1997), but
antibodies directed against the polypeptide backbone may recognize the protein in any

Expression of CD34 mRNA and protein in bovine tissues
57
cell. The extracellular polypeptide fragment yielded a polyclonal antibody, designated
boCD34ecf, which was found useful in Western blots, cytocentrifuge preparates, and
paraffin sections. However, it did not label living cells. While this is a major
shortcoming, such a reagent is still valuable for veterinary research and for the use of
cattle as a large animal model in stem cell and cardiovascular biology. In addition, the
polyclonal antibody will be helpful in the generation of monoclonal antibodies against
the glycosylated CD34 protein.
8.4. Expression of CD34 mRNA and protein in bovine tissues (V)
The expression of bovine CD34 was characterized at the mRNA level. The boCD34ecf
antibody was then used to investigate the previously unknown distribution of the CD34
protein in cattle tissues.
RTPCR analysis of the CD34 mRNA expression confirmed that two splice variants
are expressed in cattle, as predicted by the genomic data. The cytoplasmic domain is
truncated in the alternatively spliced variant by an exon extension introducing a stop
codon. This is the case also in human (Nakamura et al., 1993) and mice (Suda et al.,
1992), but has not been demonstrated previously for cattle (Zhou et al., 2001a). Both
isoforms were detected in all tissues investigated. The method applied did not provide
reliable quantitative information. However, any tissuelevel analysis would
nevertheless primarily reflect vascular densities; based on in situ hybridization, the
majority of CD34 mRNA is produced by endothelial cells. Studying the expression
patterns of the two splice variants specifically in situ will be interesting. They may be
produced by different cell types or in different cellular states, thus reflecting the poorly
known physiological roles of CD34.
In Western blots, several forms of the bovine CD34 protein with different apparent
molecular weights (105150 kDa) were detected, depending on the tissue. These
probably represent both alternatively spliced and variably glycosylated forms of the
protein.
Immunofluorescence staining showed the tissue distribution of the bovine CD34 to
be comparable with the other species investigated. The endothelium was positive in
most blood vessels, generally most intensely in the capillaries, arterioles and venules.
Hepatic sinusoids and lymphatic vessels were negative. BoCD34ecf staining was
enriched by lineage depletion of bone marrow mononuclear cells, suggesting CD34 as a

Discussion
58
marker of primitive hematopoietic cells also in cattle. Assaying CD34 expression in the
actual HSCs obviously requires the isolation of live cells and testing of their
differentiation and selfrenewal capacity, impossible with the current antibody.
The boCD34ecf antibody also stained fibroblast subpopulations, thymic
epithelioreticulocytes, some types of neurons and smooth muscle. These have
previously been reported to express the CD34 protein in mice and humans (Lin et al.,
1995; Miettinen et al., 1994). Only the fibroblast staining could be reproduced in
tyramide amplified immunohistochemistry, where a more specific staining is expected
due to highly diluted antibodies. We also were unable to detect CD34 mRNA in the
other cells and tissues by tyramide amplified in situ hybridization. Although the CD34
protein is known to be more longlived than the mRNA (Beauchamp et al., 2000), these
immunofluorescence results must be interpreted with caution. The antibody may
recognize a related, highly immunogenic epitope on another protein in the more
permissive conditions of nonamplified immunofluorescence stainings.
BoCD34ecf immunofluorescence was associated with late spermatids in the walls of
seminiferous tubules. The spermatid tails were apparently labelled, although it is not
possible to distinguish these from the surrounding Sertoli cells at the level of resolution
provided by standard epifluorescence microscopy. Mature spermatozoa in the lumen
were negative, as were the sections of the tubules without spermatids. The CD34 ligand
Lselectin (Baumheter et al., 1993) has been reported in rat Sertoli cells (Freeman et al.,
2002). It is tempting to speculate that CD34 may have a role in cell adhesion and
control of differentiation at the late stages of spermatogenesis, as it does in
hematopoietic progenitors and muscle satellite cells (Majdic et al., 1994; Healy et al.,
1995; Hu and Chien, 1998; Gordon et al., 2000; Beauchamp et al., 2000). However, the
findings could not be reproduced by tyramide amplified immunohistochemistry or
confirmed by in situ hybridization. Further studies are thus needed to validate the
immunofluorescence findings.

59
9. Conclusions
a) The chimeric freemartin cattle are a useful large animal model for somatic
stem cell research.
b) Y chromosome in situ hybridization, combined with immunohistochemistry,
is a powerful method for analysing the contribution of bullderived cells to
freemartin tissues.
c) Hematopoietic stem cells, and the progeny of other cell types circulating in
the fetal blood, do not generate significant numbers of nonhematopoietic
cells in the development, growth and physiological turnover of bovine
tissues. However, they may be important in tissue repair and regeneration.
d) The ileal Peyer’s patch has a dominant role in the generation and
maintenance of the bovine B lymphocyte pool, as it is thought to have in
sheep. Peripheral B cells are probably produced within the ileal Peyer’s
patch, rather than generated de novo from uncommitted bone marrow
precursors, at least in young animals. The oligoclonal nature of the Peyer’s
patch follicles suggests that cattle rely largely on postrearrangement
strategies in the diversification of immunoglobulins.
e) The first antibovine CD34 antibody was produced in this work. The
polyclonal antibody can be used to study cattle hematopoietic and
cardiovascular systems in tissue sections, cytocentrifuge preparates and
Western blot. The bovine CD34 mRNA is alternatively spliced, and both
splice variants are widely expressed. The CD34 protein was detected in most
blood vessel endothelia, primitive hematopoietic cells and also in some non
hematopoietic cells.

References
60
10. References
Abe,R., Donnelly,S.C., Peng,T., Bucala,R., and Metz,C.N. (2001). Peripheral blood fibrocytes:differentiation pathway and migration to wound sites. J. Immunol. 166, 75567562.
Abkowitz,J.L., Robinson,A.E., Kale,S., Long,M.W., and Chen,J. (2003). Mobilization ofhematopoietic stem cells during homeostasis and after cytokine exposure. Blood 102, 12491253.
Adams,G.B., Chabner,K.T., Alley,I.R., Olson,D.P., Szczepiorkowski,Z.M., Poznansky,M.C.,Kos,C.H., Pollak,M.R., Brown,E.M., and Scadden,D.T. (2006). Stem cell engraftment at theendosteal niche is specified by the calciumsensing receptor. Nature 439, 599603.
Adolfsson,J., Mansson,R., BuzaVidas,N., Hultquist,A., Liuba,K., Jensen,C.T., Bryder,D., Yang,L.,Borge,O.J., Thoren,L.A., Anderson,K., Sitnicka,E., Sasaki,Y., Sigvardsson,M., and Jacobsen,S.E.(2005). Identification of Flt3+ lymphomyeloid stem cells lacking erythromegakaryocyticpotential a revised road map for adult blood lineage commitment. Cell 121, 295306.
Ahn,S. and Joyner,A.L. (2005). In vivo analysis of quiescent adult neural stem cells respondingto Sonic hedgehog. Nature 437, 894897.
Akashi,K., He,X., Chen,J., Iwasaki,H., Niu,C., Steenhard,B., Zhang,J., Haug,J., and Li,L. (2003).Transcriptional accessibility for genes of multiple tissues and hematopoietic lineages ishierarchically controlled during early hematopoiesis. Blood 101, 383389.
Akashi,K., Traver,D., Miyamoto,T., and Weissman,I.L. (2000). A clonogenic common myeloidprogenitor that gives rise to all myeloid lineages. Nature 404, 193197.
Alonso,L. and Fuchs,E. (2003). Stem cells of the skin epithelium. Proc. Natl. Acad. Sci. U. S. A.100 Suppl 1, 1183011835.
Altman,J. and Das,G.D. (1965). Autoradiographic and histological evidence of postnatalhippocampal neurogenesis in rats. J. Comp. Neurol. 124, 319335.
AlvarezBuylla,A., GarciaVerdugo,J.M., and Tramontin,A.D. (2001). A unified hypothesis on thelineage of neural stem cells. Nat. Rev. Neurosci. 2, 287293.
AlvarezBuylla,A. and Lim,D.A. (2004). For the long run: maintaining germinal niches in theadult brain. Neuron 41, 683686.
AlvarezDolado,M., Pardal,R., GarciaVerdugo,J.M., Fike,J.R., Lee,H.O., Pfeffer,K., Lois,C.,Morrison,S.J., and AlvarezBuylla,A. (2003). Fusion of bonemarrowderived cells with Purkinjeneurons, cardiomyocytes and hepatocytes. Nature 425, 968973.
Anderson,D., Billingham,R.E., Lampkin,G.H., and Medawar,P.B. (1951). The use of skin graftingto distinguish between monozygotic and dizygotic twins in cattle. Heredity 5, 379397.
Antonchuk,J., Sauvageau,G., and Humphries,R.K. (2002). HOXB4induced expansion of adulthematopoietic stem cells ex vivo. Cell 109, 3945.
Arai,F., Hirao,A., Ohmura,M., Sato,H., Matsuoka,S., Takubo,K., Ito,K., Koh,G.Y., and Suda,T.(2004). Tie2/angiopoietin1 signaling regulates hematopoietic stem cell quiescence in the bonemarrow niche. Cell 118, 149161.
Arnold,A., Cossman,J., Bakhshi,A., Jaffe,E.S., Waldmann,T.A., and Korsmeyer,S.J. (1983).Immunoglobulingene rearrangements as unique clonal markers in human lymphoid neoplasms.N. Engl. J. Med. 309, 15931599.
Asahara,T., Masuda,H., Takahashi,T., Kalka,C., Pastore,C., Silver,M., Kearne,M., Magner,M., andIsner,J.M. (1999). Bone marrow origin of endothelial progenitor cells responsible for postnatalvasculogenesis in physiological and pathological neovascularization. Circ. Res. 85, 221228.
Asakura,A. and Rudnicki,M.A. (2002). Side population cells from diverse adult tissues arecapable of in vitro hematopoietic differentiation. Exp. Hematol. 30, 13391345.
Bailey,A.S., Jiang,S., Afentoulis,M., Baumann,C.I., Schroeder,D.A., Olson,S.B., Wong,M.H., andFleming,W.H. (2004). Transplanted adult hematopoietic stems cells differentiate into functionalendothelial cells. Blood 103, 1319.

61
Batlle,E., Henderson,J.T., Beghtel,H., van den Born,M.M., Sancho,E., Huls,G., Meeldijk,J.,Robertson,J., van de Wetering,M., Pawson,T., and Clevers,H. (2002). Betacatenin and TCFmediate cell positioning in the intestinal epithelium by controlling the expression ofEphB/ephrinB. Cell 111, 251263.
Baumheter,S., Singer,M.S., Henzel,W., Hemmerich,S., Renz,M., Rosen,S.D., and Lasky,L.A.(1993). Binding of Lselectin to the vascular sialomucin CD34. Science 262, 436438.
Bayer,S.A. (1982). Changes in the total number of dentate granule cells in juvenile and adultrats: a correlated volumetric and 3Hthymidine autoradiographic study. Exp. Brain Res. 46, 315323.
Beauchamp,J.R., Heslop,L., Yu,D.S., Tajbakhsh,S., Kelly,R.G., Wernig,A., Buckingham,M.E.,Partridge,T.A., and Zammit,P.S. (2000). Expression of CD34 and Myf5 defines the majority ofquiescent adult skeletal muscle satellite cells. J. Cell Biol. 151, 12211234.
Becker,A.J., McCulloch,E.A., and Till,J.E. (1963). Cytological demonstration of the clonal natureof spleen colonies derived from transplanted mouse marrow cells. Nature 197, 452454.
Belluzzi,O., Benedusi,M., Ackman,J., and LoTurco,J.J. (2003). Electrophysiologicaldifferentiation of new neurons in the olfactory bulb. J. Neurosci. 23, 1041110418.
Berens,S.J., Wylie,D.E., and Lopez,O.J. (1997). Use of a single VH family and long CDR3s in thevariable region of cattle Ig heavy chains. Int. Immunol. 9, 189199.
Berenson,R.J., Bensinger,W.I., Hill,R.S., Andrews,R.G., GarciaLopez,J., Kalamasz,D.F., Still,B.J.,Spitzer,G., Buckner,C.D., Bernstein,I.D., and . (1991). Engraftment after infusion of CD34+marrow cells in patients with breast cancer or neuroblastoma. Blood 77, 17171722.
Beyaz,F. and Asti,R.N. (2004). Development of ileal Peyer's patches and follicle associatedepithelium in bovine foetuses. Anat. Histol. Embryol. 33, 172179.
Bhatia,M., Bonnet,D., Murdoch,B., Gan,O.I., and Dick,J.E. (1998). A newly discovered class ofhuman hematopoietic cells with SCIDrepopulating activity. Nat. Med. 4, 10381045.
Bhatia,M., Wang,J.C., Kapp,U., Bonnet,D., and Dick,J.E. (1997). Purification of primitive humanhematopoietic cells capable of repopulating immunedeficient mice. Proc. Natl. Acad. Sci. U. S.A. 94, 53205325.
Bickenbach,J.R. and Mackenzie,I.C. (1984). Identification and localization of labelretaining cellsin hamster epithelia. J. Invest. Dermatol. 82, 618622.
Billingham,R.E., Brent,L., and Medawar,P.B. (1953). Actively acquired tolerance of foreign cells.Nature 172, 603606.
Billingham,R.E., Lampkin,G.H., Medawar,P.B., and Williams,H.L. (1952). Tolerance tohomografts, twin diagnosis, and the freemartin condition in cattle. Heredity 6, 201212.
Bissonnette,T.H. (1924). The development of the reproductive ducts and canals in the freemartinwith comparison of the normal. Am. J. Anat. 33, 267345.
Bjerknes,M. and Cheng,H. (1999). Clonal analysis of mouse intestinal epithelial progenitors.Gastroenterology 116, 714.
Bjornson,C.R., Rietze,R.L., Reynolds,B.A., Magli,M.C., and Vescovi,A.L. (1999). Turning braininto blood: a hematopoietic fate adopted by adult neural stem cells in vivo. Science 283, 534537.
Blanpain,C., Lowry,W.E., Geoghegan,A., Polak,L., and Fuchs,E. (2004). Selfrenewal,multipotency, and the existence of two cell populations within an epithelial stem cell niche.Cell 118, 635648.
Blau,H.M., Brazelton,T.R., and Weimann,J.M. (2001). The evolving concept of a stem cell: entityor function? Cell 105, 829841.
Bohm,M., Wieland,I., Schutze,K., and Rubben,H. (1997). Microbeam MOMeNT: noncontactlaser microdissection of membranemounted native tissue. Am. J. Anat. 151, 6367.
Boklage,C.E. (2006). Embryogenesis of chimeras, twins and anterior midline asymmetries. Hum.Reprod. 21, 579591.

References
62
Brazelton,T.R., Rossi,F.M., Keshet,G.I., and Blau,H.M. (2000). From marrow to brain: expressionof neuronal phenotypes in adult mice. Science 290, 17751779.
Brent,L. (1997). The discovery of immunologic tolerance. Hum. Immunol. 52, 7581.
Brockes,J.P. and Kumar,A. (2002). Plasticity and reprogramming of differentiated cells inamphibian regeneration. Nat. Rev. Mol. Cell Biol. 3, 566574.
Butler,H. and Juurlink,B.H.J. (1987). An atlas for staging mammalian and chick embryos. (BocaRaton: CRC Press).
Cairns,J. (1975). Mutation selection and the natural history of cancer. Nature 255, 197200.
Cairns,J. (2002). Somatic stem cells and the kinetics of mutagenesis and carcinogenesis. Proc.Natl. Acad. Sci. U. S. A. 99, 1056710570.
Calvi,L.M., Adams,G.B., Weibrecht,K.W., Weber,J.M., Olson,D.P., Knight,M.C., Martin,R.P.,Schipani,E., Divieti,P., Bringhurst,F.R., Milner,L.A., Kronenberg,H.M., and Scadden,D.T. (2003).Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 425, 841846.
Camargo,F.D., Green,R., Capetanaki,Y., Jackson,K.A., and Goodell,M.A. (2003). Singlehematopoietic stem cells generate skeletal muscle through myeloid intermediates. Nat. Med. 9,15201527.
Campagnoli,C., Roberts,I.A., Kumar,S., Bennett,P.R., Bellantuono,I., and Fisk,N.M. (2001).Identification of mesenchymal stem/progenitor cells in human first trimester fetal blood, liver,and bone marrow. Blood 98, 23962402.
Capel,B. and Coveney,D. (2004). Frank Lillie's freemartin: illuminating the pathway to 21stcentury reproductive endocrinology. J. Exp. Zoolog. A Comp. Exp. Biol. 301, 853856.
Carlén,M., Cassidy,R.M., Brismar,H., Smith,G.A., Enquist,L.W., and Frisen,J. (2002). Functionalintegration of adultborn neurons. Curr. Biol. 12, 606608.
Carleton,A., Petreanu,L.T., Lansford,R., AlvarezBuylla,A., and Lledo,P.M. (2003). Becoming anew neuron in the adult olfactory bulb. Nat. Neurosci. 6, 507518.
Castro,R.F., Jackson,K.A., Goodell,M.A., Robertson,C.S., Liu,H., and Shine,H.D. (2002). Failureof bone marrow cells to transdifferentiate into neural cells in vivo. Science 297, 1299.
Challen,G.A. and Little,M.H. (2006). A side order of stem cells: the SP phenotype. Stem Cells 24,312.
Charbord,P., Tavian,M., Humeau,L., and Peault,B. (1996). Early ontogeny of the human marrowfrom long bones: an immunohistochemical study of hematopoiesis and its microenvironment.Blood 87, 41094119.
Cheshier,S.H., Morrison,S.J., Liao,X., and Weissman,I.L. (1999). In vivo proliferation and cellcycle kinetics of longterm selfrenewing hematopoietic stem cells. Proc. Natl. Acad. Sci. U. S. A.96, 31203125.
Chiasson,B.J., Tropepe,V., Morshead,C.M., and van der Kooy,D. (1999). Adult mammalianforebrain ependymal and subependymal cells demonstrate proliferative potential, but onlysubependymal cells have neural stem cell characteristics. J. Neurosci. 19, 44624471.
Christensen,J.L., Wright,D.E., Wagers,A.J., and Weissman,I.L. (2004). Circulation andchemotaxis of fetal hematopoietic stem cells. PLos Biol. 2, E75.
Clarke,D.L., Johansson,C.B., Wilbertz,J., Veress,B., Nilsson,E., Karlstrom,H., Lendahl,U., andFrisen,J. (2000). Generalized potential of adult neural stem cells. Science 288, 16601663.
Clarke,M.F. and Fuller,M. (2006). Stem cells and cancer: two faces of eve. Cell 124, 11111115.
Claudinot,S., Nicolas,M., Oshima,H., Rochat,A., and Barrandon,Y. (2005). Longterm renewal ofhair follicles from clonogenic multipotent stem cells. Proc. Natl. Acad. Sci. U. S. A. 102, 1467714682.
Colter,D.C., Class,R., DiGirolamo,C.M., and Prockop,D.J. (2000). Rapid expansion of recyclingstem cells in cultures of plasticadherent cells from human bone marrow. Proc. Natl. Acad. Sci.U. S. A. 97, 32133218.

63
Copp,A.J. (1995). Death before birth: clues from gene knockouts and mutations. Trends Genet.11, 8793.
Corbel,S.Y., Lee,A., Yi,L., Duenas,J., Brazelton,T.R., Blau,H.M., and Rossi,F.M. (2003).Contribution of hematopoietic stem cells to skeletal muscle. Nat. Med. 9, 15281532.
Cotsarelis,G., Sun,T.T., and Lavker,R.M. (1990). Labelretaining cells reside in the bulge area ofpilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis.Cell 61, 13291337.
de Bari,C., Dell'Accio,F., Vanlauwe,J., Eyckmans,J., Khan,I.M., Archer,C.W., Jones,E.A.,McGonagle,D., Mitsiadis,T.A., Pitzalis,C., and Luyten,F.P. (2006). Mesenchymal multipotency ofadult human periosteal cells demonstrated by singlecell lineage analysis. Arthritis Rheum. 54,12091221.
de Bruijn,M.F., Speck,N.A., Peeters,M.C., and Dzierzak,E. (2000). Definitive hematopoietic stemcells first develop within the major arterial regions of the mouse embryo. EMBO J. 19, 24652474.
de Rooij,D.G. and Grootegoed,J.A. (1998). Spermatogonial stem cells. Curr. Opin. Cell Biol. 10,694701.
Dennis,J.E. and Charbord,P. (2002). Origin and differentiation of human and murine stroma.Stem Cells 20, 205214.
Deschaseaux,F. and Charbord,P. (2000). Human marrow stromal precursors are alpha 1 integrinsubunitpositive. J. Cell Physiol. 184, 319325.
DieterlenLièvre,F. and Martin,C. (1981). Diffuse intraembryonic hemopoiesis in normal andchimeric avian development. Dev. Biol. 88, 180191.
Doetsch,F. (2003). A niche for adult neural stem cells. Curr. Opin. Genet. Dev. 13, 543550.
Doetsch,F., Caille,I., Lim,D.A., GarciaVerdugo,J.M., and AlvarezBuylla,A. (1999a).Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97, 703716.
Doetsch,F., GarciaVerdugo,J.M., and AlvarezBuylla,A. (1999b). Regeneration of a germinallayer in the adult mammalian brain. Proc. Natl. Acad. Sci. U. S. A. 96, 1161911624.
Doetsch,F., Petreanu,L., Caille,I., GarciaVerdugo,J.M., and AlvarezBuylla,A. (2002). EGFconverts transitamplifying neurogenic precursors in the adult brain into multipotent stem cells.Neuron 36, 10211034.
Doughri,A.M., Altera,K.P., and Kainer,R.A. (1972). Some developmental aspects of the bovinefetal gut. Zentralbl. Veterinarmed. A 19, 417434.
Duncan,A.W., Rattis,F.M., DiMascio,L.N., Congdon,K.L., Pazianos,G., Zhao,C., Yoon,K.,Cook,J.M., Willert,K., Gaiano,N., and Reya,T. (2005). Integration of Notch and Wnt signaling inhematopoietic stem cell maintenance. Nat. Immunol. 6, 314322.
Dunnwald,M., TomanekChalkley,A., Alexandrunas,D., Fishbaugh,J., and Bickenbach,J.R.(2001). Isolating a pure population of epidermal stem cells for use in tissue engineering. Exp.Dermatol. 10, 4554.
Durand,R.E. and Olive,P.L. (1982). Cytotoxicity, mutagenicity and DNA damage by Hoechst33342. J. Histochem. Cytochem. 30, 111116.
Eakin,G.S. and Behringer,R.R. (2004). Diversity of germ layer and axis formation amongmammals. Semin. Cell Dev. Biol. 15, 619629.
Eggan,K., Jurga,S., Gosden,R., Min,I.M., and Wagers,A.J. (2006). Ovulated oocytes in adult micederive from noncirculating germ cells. Nature 441, 11091114.
Ema,H. and Nakauchi,H. (2000). Expansion of hematopoietic stem cells in the developing liverof a mouse embryo. Blood 95, 22842288.
Eriksson,P.S., Perfilieva,E., BjorkEriksson,T., Alborn,A.M., Nordborg,C., Peterson,D.A., andGage,F.H. (1998). Neurogenesis in the adult human hippocampus. Nat. Med. 4, 13131317.

References
64
Esposito,M.S., Piatti,V.C., Laplagne,D.A., Morgenstern,N.A., Ferrari,C.C., Pitossi,F.J., andSchinder,A.F. (2005). Neuronal differentiation in the adult hippocampus recapitulatesembryonic development. J. Neurosci. 25, 1007410086.
Evans,M.J. and Kaufman,M.H. (1981). Establishment in culture of pluripotential cells frommouse embryos. Nature 292, 154156.
Falk,A. and Frisen,J. (2005). New neurons in old brains. Ann. Med. 37, 480486.
Ferkowicz,M.J. and Yoder,M.C. (2005). Blood island formation: longstanding observations andmodern interpretations. Exp. Hematol. 33, 10411047.
Ferrari,G., CusellaDe Angelis,G., Coletta,M., Paolucci,E., Stornaiuolo,A., Cossu,G., andMavilio,F. (1998). Muscle regeneration by bone marrowderived myogenic progenitors. Science279, 15281530.
Forbes,T. (1946). The origin of freemartin. Bull. Hist. Med. 20, 461466.
Ford,C.E., Hamerton,J.L., Barnes,D.W., and Loutit,J.F. (1956). Cytological identification ofradiationchimaeras. Nature 177, 452454.
Fortunel,N.O., Otu,H.H., Ng,H.H., Chen,J., Mu,X., Chevassut,T., Li,X., Joseph,M., Bailey,C.,Hatzfeld,J.A., Hatzfeld,A., Usta,F., Vega,V.B., Long,P.M., Libermann,T.A., and Lim,B. (2003).Comment on " 'Stemness': transcriptional profiling of embryonic and adult stem cells" and "astem cell molecular signature". Science 302, 393.
Freeman,E.A., Jani,P., and Millette,C.E. (2002). Expression and potential function of Rho familysmall G proteins in cells of the mammalian seminiferous epithelium. Cell Commun. Adhes. 9,189204.
Friedenstein,A.J., Chailakhjan,R.K., and Lalykina,K.S. (1970). The development of fibroblastcolonies in monolayer cultures of guineapig bone marrow and spleen cells. Cell Tissue Kinet. 3,393403.
Friedenstein,A.J., Chailakhyan,R.K., and Gerasimov,U.V. (1987). Bone marrow osteogenic stemcells: in vitro cultivation and transplantation in diffusion chambers. Cell Tissue Kinet. 20, 263272.
Fuchs,E. and Segre,J.A. (2000). Stem cells: a new lease on life. Cell 100, 143155.
Fuchs,E., Tumbar,T., and Guasch,G. (2004). Socializing with the neighbors: stem cells and theirniche. Cell 116, 769778.
Gallacher,L., Murdoch,B., Wu,D.M., Karanu,F.N., Keeney,M., and Bhatia,M. (2000). Isolationand characterization of human CD34()Lin() and CD34(+)Lin() hematopoietic stem cells usingcell surface markers AC133 and CD7. Blood 95, 28132820.
Galli,R., Borello,U., Gritti,A., Minasi,M.G., Bjornson,C., Coletta,M., Mora,M., De Angelis,M.G.,Fiocco,R., Cossu,G., and Vescovi,A.L. (2000). Skeletal myogenic potential of human and mouseneural stem cells. Nat. Neurosci. 3, 986991.
Garcia,A.D., Doan,N.B., Imura,T., Bush,T.G., and Sofroniew,M.V. (2004). GFAPexpressingprogenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat.Neurosci. 7, 12331241.
GarciaVerdugo,J.M., Ferron,S., Flames,N., Collado,L., Desfilis,E., and Font,E. (2002). Theproliferative ventricular zone in adult vertebrates: a comparative study using reptiles, birds, andmammals. Brain Res. Bull. 57, 765775.
Gardner,R.L. and Rossant,J. (1979). Investigation of the fate of 45 day postcoitum mouse innercell mass cells by blastocyst injection. J. Embryol. Exp. Morphol. 52, 141152.
Gekas,C., eterlenLievre,F., Orkin,S.H., and Mikkola,H.K. (2005). The placenta is a niche forhematopoietic stem cells. Dev. Cell 8, 365375.
Gerber,H.A., Morris,B., and Trevella,W. (1986). The role of gutassociated lymphoid tissues inthe generation of immunoglobulinbearing lymphocytes in sheep. Aust. J. Exp. Biol. Med. Sci. 64( Pt 3), 201213.

65
Giebel,B., Zhang,T., Beckmann,J., Spanholtz,J., Wernet,P., Ho,A.D., and Punzel,M. (2006).Primitive human hematopoietic cells give rise to differentially specified daughter cells upontheir initial cell division. Blood 107, 21462152.
Gilbert,S. (2006). Developmental biology. (Sunderland: Sinauer Associates).
Glick,B., Chang,T., and Jaap,G. (1956). The bursa of Fabricius and antibody production. PoultrySci. 35, 224225.
Gong,J.K. (1978). Endosteal marrow: a rich source of hematopoietic stem cells. Science 199,14431445.
Goodell,M.A. (2003). Stemcell "plasticity": befuddled by the muddle. Curr. Opin. Hematol. 10,208213.
Goodell,M.A., Brose,K., Paradis,G., Conner,A.S., and Mulligan,R.C. (1996). Isolation andfunctional properties of murine hematopoietic stem cells that are replicating in vivo. J. Exp.Med. 183, 17971806.
Goodell,M.A., Rosenzweig,M., Kim,H., Marks,D.F., DeMaria,M., Paradis,G., Grupp,S.A.,Sieff,C.A., Mulligan,R.C., and Johnson,R.P. (1997). Dye efflux studies suggest that hematopoieticstem cells expressing low or undetectable levels of CD34 antigen exist in multiple species. Nat.Med. 3, 13371345.
Goodman,J.W. and Hodgson,G.S. (1962). Evidence for stem cells in the peripheral blood of mice.Blood 19, 702714.
Gordon,J.I., Schmidt,G.H., and Roth,K.A. (1992). Studies of intestinal stem cells using normal,chimeric, and transgenic mice. FASEB J. 6, 30393050.
Gordon,M.Y., Marley,S.B., Davidson,R.J., Grand,F.H., Lewis,J.L., Nguyen,D.X., Lloyd,S., andGoldman,J.M. (2000). Contactmediated inhibition of human haematopoietic progenitor cellproliferation may be conferred by stem cell antigen, CD34. Hematol. J. 1, 7786.
Grant,M.B., May,W.S., Caballero,S., Brown,G.A., Guthrie,S.M., Mames,R.N., Byrne,B.J.,Vaught,T., Spoerri,P.E., Peck,A.B., and Scott,E.W. (2002). Adult hematopoietic stem cellsprovide functional hemangioblast activity during retinal neovascularization. Nat. Med. 8, 607612.
Griebel,P.J. and Hein,W.R. (1996). Expanding the role of Peyer's patches in Bcell ontogeny.Immunol. Today 17, 3039.
Griebel,P.J., Kennedy,L., Graham,T., Davis,W.C., and Reynolds,J.D. (1992). Characterization ofBcell phenotypic changes during ileal and jejunal Peyer's patch development in sheep.Immunology 77, 564570.
Gronthos,S., Zannettino,A.C., Hay,S.J., Shi,S., Graves,S.E., Kortesidis,A., and Simmons,P.J.(2003). Molecular and cellular characterisation of highly purified stromal stem cells derivedfrom human bone marrow. J. Cell Sci. 116, 18271835.
Gross,C.G. (2000). Neurogenesis in the adult brain: death of a dogma. Nat. Rev. Neurosci. 1, 6773.
Groszer,M., Erickson,R., ScriptureAdams,D.D., Dougherty,J.D., Le,B.J., Zack,J.A.,Geschwind,D.H., Liu,X., Kornblum,H.I., and Wu,H. (2006). PTEN negatively regulates neuralstem cell selfrenewal by modulating G0G1 cell cycle entry. Proc. Natl. Acad. Sci. U. S. A. 103,111116.
Harrington,L. (2004). Does the reservoir for selfrenewal stem from the ends? Oncogene 23,72837289.
Harvey,W. (1651). Exercitationes de generatione animalium: quibus accedunt quaedum departu, de membranis ac humoribus uteri et de conceptione. (London: Ent, Sir George).
Healy,L., May,G., Gale,K., Grosveld,F., Greaves,M., and Enver,T. (1995). The stem cell antigenCD34 functions as a regulator of hemopoietic cell adhesion. Proc. Natl. Acad. Sci. U. S. A. 92,1224012244.
Hein,W.R., Dudler,L., and Mackay,C.R. (1989). Surface expression of differentiation antigens onlymphocytes in the ileal and jejunal Peyer's patches of lambs. Immunology 68, 365370.

References
66
Herzog,E.L., Chai,L., and Krause,D.S. (2003). Plasticity of marrowderived stem cells. Blood 102,34833493.
HewickerTrautwein,M., Schultheis,G., and Trautwein,G. (1996). Demonstration of amoeboidand ramified microglial cells in pre and postnatal bovine brains by lectin histochemistry. Anat.Anz. 178, 2531.
Ho,A.D. (2005). Kinetics and symmetry of divisions of hematopoietic stem cells. Exp. Hematol.33, 18.
Hochedlinger,K. and Jaenisch,R. (2002). Monoclonal mice generated by nuclear transfer frommature B and T donor cells. Nature 415, 10351038.
Hopman,A.H., Ramaekers,F.C., and Speel,E.J. (1998). Rapid synthesis of biotin, digoxigenin,trinitrophenyl, and fluorochromelabeled tyramides and their application for In situhybridization using CARD amplification. J. Histochem. Cytochem. 46, 771777.
Houssaint,E. (1981). Differentiation of the mouse hepatic primordium. II. Extrinsic origin of thehaemopoietic cell line. Cell Differ. 10, 243252.
Hu,M.C. and Chien,S.L. (1998). The cytoplasmic domain of stem cell antigen CD34 is essentialfor cytoadhesion signaling but not sufficient for proliferation signaling. Blood 91, 11521162.
Huber,T.L., Kouskoff,V., Fehling,H.J., Palis,J., and Keller,G. (2004). Haemangioblast commitmentis initiated in the primitive streak of the mouse embryo. Nature 432, 625630.
Hunter,J. (1779). Account of the Free Martin. Philos. Trans. R. Soc. Lond. 69, 279293.
Hüttmann,A., Duhrsen,U., Heydarian,K., KleinHitpass,L., Boes,T., Boyd,A.W., and Li,C.L.(2006). Gene expression profiles in murine hematopoietic stem cells revisited: analysis of cDNAlibraries reveals high levels of translational and metabolic activities. Stem Cells 24, 17191727.
Ikuta,K. and Weissman,I.L. (1992). Evidence that hematopoietic stem cells express mouse ckitbut do not depend on steel factor for their generation. Proc. Natl. Acad. Sci. U. S. A. 89, 15021506.
Ishino,S., Kadota,K., Matsubara,Y., Agawa,H., and Matsui,N. (1991). Immunohistochemicalstudies on ontogeny of bovine lymphoid tissues. J. Vet. Med. Sci. 53, 877882.
Ito,M., Liu,Y., Yang,Z., Nguyen,J., Liang,F., Morris,R.J., and Cotsarelis,G. (2005). Stem cells inthe hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat.Med. 11, 13511354.
Ito,T., Tajima,F., and Ogawa,M. (2000). Developmental changes of CD34 expression by murinehematopoietic stem cells. Exp. Hematol. 28, 12691273.
Ivanova,N.B., Dimos,J.T., Schaniel,C., Hackney,J.A., Moore,K.A., and Lemischka,I.R. (2002). Astem cell molecular signature. Science 298, 601604.
Jackson,K.A., Mi,T., and Goodell,M.A. (1999). Hematopoietic potential of stem cells isolatedfrom murine skeletal muscle. Proc. Natl. Acad. Sci. U. S. A. 96, 1448214486.
Jacobson,L.O., Simmons,E.L., Marks,E.K., and Eldredge,J.H. (1951). Recovery from radiationinjury. Science 113, 510511.
Janeway,C., Travers,P., Walport,M., and Shlomchik,M. (2001). Immunobiology. The immunesystem in health and disease. (New York: Garland Publishing).
Jang,Y.Y., Collector,M.I., Baylin,S.B., Diehl,A.M., and Sharkis,S.J. (2004). Hematopoietic stemcells convert into liver cells within days without fusion. Nat. Cell Biol. 6, 532539.
Javazon,E.H., Beggs,K.J., and Flake,A.W. (2004). Mesenchymal stem cells: paradoxes ofpassaging. Exp. Hematol. 32, 414425.
Jenne,C.N., Kennedy,L.J., McCullagh,P., and Reynolds,J.D. (2003). A new model of sheep Igdiversification: shifting the emphasis toward combinatorial mechanisms and away fromhypermutation. J. Immunol. 170, 37393750.
Jenne,C.N., Kennedy,L.J., and Reynolds,J.D. (2006). Antibody repertoire development in thesheep. Dev. Comp. Immunol. 30, 165174.

67
Jiang,Y., Jahagirdar,B.N., Reinhardt,R.L., Schwartz,R.E., Keene,C.D., OrtizGonzalez,X.R.,Reyes,M., Lenvik,T., Lund,T., Blackstad,M., Du,J., Aldrich,S., Lisberg,A., Low,W.C.,Largaespada,D.A., and Verfaillie,C.M. (2002a). Pluripotency of mesenchymal stem cells derivedfrom adult marrow. Nature 418, 4149.
Jiang,Y., Vaessen,B., Lenvik,T., Blackstad,M., Reyes,M., and Verfaillie,C.M. (2002b). Multipotentprogenitor cells can be isolated from postnatal murine bone marrow, muscle, and brain. Exp.Hematol. 30, 896904.
Johnson,G.R. and Moore,M.A. (1975). Role of stem cell migration in initiation of mouse foetalliver haemopoiesis. Nature 258, 726728.
Johnson,J., Bagley,J., SkaznikWikiel,M., Lee,H.J., Adams,G.B., Niikura,Y., Tschudy,K.S.,Tilly,J.C., Cortes,M.L., Forkert,R., Spitzer,T., Iacomini,J., Scadden,D.T., and Tilly,J.L. (2005).Oocyte generation in adult mammalian ovaries by putative germ cells in bone marrow andperipheral blood. Cell 122, 303315.
Jones,P.H., Harper,S., and Watt,F.M. (1995). Stem cell patterning and fate in human epidermis.Cell 80, 8393.
Jones,R.J., Wagner,J.E., Celano,P., Zicha,M.S., and Sharkis,S.J. (1990). Separation of pluripotenthaematopoietic stem cells from spleen colonyforming cells. Nature 347, 188189.
Joseph,N.M. and Morrison,S.J. (2005). Toward an understanding of the physiological function ofmammalian stem cells. Dev. Cell 9, 173183.
Jost,A. (1970). Hormonal factors in the sex differentiation of the mammalian foetus. Philos.Trans. R. Soc. Lond. B Biol. Sci. 259, 119130.
Kale,S., Karihaloo,A., Clark,P.R., Kashgarian,M., Krause,D.S., and Cantley,L.G. (2003). Bonemarrow stem cells contribute to repair of the ischemically injured renal tubule. J. Clin. Invest112, 4249.
Kaplan,M.S. and Bell,D.H. (1984). Mitotic neuroblasts in the 9dayold and 11monthold rodenthippocampus. J. Neurosci. 4, 14291441.
Karpowicz,P., Morshead,C., Kam,A., Jervis,E., Ramunas,J., Cheng,V., and van der Kooy,D.(2005). Support for the immortal strand hypothesis: neural stem cells partition DNAasymmetrically in vitro. J. Cell Biol. 170, 721732.
Katayama,Y., Battista,M., Kao,W.M., Hidalgo,A., Peired,A.J., Thomas,S.A., and Frenette,P.S.(2006). Signals from the sympathetic nervous system regulate hematopoietic stem cell egressfrom bone marrow. Cell 124, 407421.
Kaushik,A., Shojaei,F., and Saini,S.S. (2002). Novel insight into antibody diversification fromcattle. Vet. Immunol. Immunopathol. 87, 347350.
Kayahara,T., Sawada,M., Takaishi,S., Fukui,H., Seno,H., Fukuzawa,H., Suzuki,K., Hiai,H.,Kageyama,R., Okano,H., and Chiba,T. (2003). Candidate markers for stem and early progenitorcells, Musashi1 and Hes1, are expressed in crypt base columnar cells of mouse small intestine.FEBS Lett. 535, 131135.
Kempermann,G., Jessberger,S., Steiner,B., and Kronenberg,G. (2004). Milestones of neuronaldevelopment in the adult hippocampus. Trends Neurosci. 27, 447452.
Khosrotehrani,K., Johnson,K.L., Cha,D.H., Salomon,R.N., and Bianchi,D.W. (2004). Transfer offetal cells with multilineage potential to maternal tissue. JAMA 292, 7580.
Kiel,M.J., Yilmaz,O.H., Iwashita,T., Yilmaz,O.H., Terhorst,C., and Morrison,S.J. (2005). SLAMfamily receptors distinguish hematopoietic stem and progenitor cells and reveal endothelialniches for stem cells. Cell 121, 11091121.
Kingston,R.E., Chomczynski,P., and Sacchi,N. (1996). Guanidine methods for total RNApreparation. In Current Protocols in Molecular Biology, Ausubel F.M., Brent R, Kingston R.E.,D.D.Moore, J.G.Seidman, J.A.Smith, and K.Struhl, eds. (Hoboken, NJ: John Wiley & Sons).
Kondo,M., Wagers,A.J., Manz,M.G., Prohaska,S.S., Scherer,D.C., Beilhack,G.F., Shizuru,J.A., andWeissman,I.L. (2003). Biology of hematopoietic stem cells and progenitors: implications forclinical application. Annu. Rev. Immunol. 21, 759806.

References
68
Kopen,G.C., Prockop,D.J., and Phinney,D.G. (1999). Marrow stromal cells migrate throughoutforebrain and cerebellum, and they differentiate into astrocytes after injection into neonatalmouse brains. Proc. Natl. Acad. Sci. U. S. A. 96, 1071110716.
Körbling,M., Katz,R.L., Khanna,A., Ruifrok,A.C., Rondon,G., Albitar,M., Champlin,R.E., andEstrov,Z. (2002). Hepatocytes and epithelial cells of donor origin in recipients of peripheralblood stem cells. N. Engl. J. Med. 346, 738746.
Korinek,V., Barker,N., Moerer,P., van Donselaar,E., Huls,G., Peters,P.J., and Clevers,H. (1998).Depletion of epithelial stemcell compartments in the small intestine of mice lacking Tcf4. Nat.Genet. 19, 379383.
Koshizuka,S., Okada,S., Okawa,A., Koda,M., Murasawa,M., Hashimoto,M., Kamada,T.,Yoshinaga,K., Murakami,M., Moriya,H., and Yamazaki,M. (2004). Transplanted hematopoieticstem cells from bone marrow differentiate into neural lineage cells and promote functionalrecovery after spinal cord injury in mice. J. Neuropathol. Exp. Neurol. 63, 6472.
Kotton,D.N., Fabian,A.J., and Mulligan,R.C. (2005). A novel stemcell population in adult liverwith potent hematopoieticreconstitution activity. Blood 106, 15741580.
Krause,D.S., Fackler,M.J., Civin,C.I., and May,W.S. (1996). CD34: structure, biology, and clinicalutility. Blood 87, 113.
Krause,D.S., Theise,N.D., Collector,M.I., Henegariu,O., Hwang,S., Gardner,R., Neutzel,S., andSharkis,S.J. (2001). Multiorgan, multilineage engraftment by a single bone marrowderivedstem cell. Cell 105, 369377.
Kukekov,V.G., Laywell,E.D., Suslov,O., Davies,K., Scheffler,B., Thomas,L.B., O'Brien,T.F.,Kusakabe,M., and Steindler,D.A. (1999). Multipotent stem/progenitor cells with similarproperties arise from two neurogenic regions of adult human brain. Exp. Neurol. 156, 333344.
Kuznetsov,S.A., Mankani,M.H., Gronthos,S., Satomura,K., Bianco,P., and Robey,P.G. (2001).Circulating skeletal stem cells. J. Cell Biol. 153, 11331140.
LaBarge,M.A. and Blau,H.M. (2002). Biological progression from adult bone marrow tomononucleate muscle stem cell to multinucleate muscle fiber in response to injury. Cell 111,589601.
Lagasse,E., Connors,H., Al Dhalimy,M., Reitsma,M., Dohse,M., Osborne,L., Wang,X.,Finegold,M., Weissman,I.L., and Grompe,M. (2000). Purified hematopoietic stem cells candifferentiate into hepatocytes in vivo. Nat. Med. 6, 12291234.
Lakshmipathy,U. and Verfaillie,C. (2005). Stem cell plasticity. Blood Rev. 19, 2938.
Le Guern,A.C., Giovino,M.A., Abe,M., Theodore,P.R., Qi,J., Down,J.D., Sachs,D.H., Sykes,M.,and Yang,Y.G. (2003). Stem cell activity of porcine ckit+ hematopoietic cells. Exp. Hematol. 31,833840.
Lendahl,U., Zimmerman,L.B., and McKay,R.D. (1990). CNS stem cells express a new class ofintermediate filament protein. Cell 60, 585595.
Leon,L.G., Ostronoff,L.K., Fermin,M.L., Fragio,C., Kremmer,E., Kolb,H.J., and Tejero,C. (2005). Invitro generation of mature neutrophils from canine Lin bone marrow cells. Vet. Immunol.Immunopathol. 107, 4150.
Levy,V., Lindon,C., Harfe,B.D., and Morgan,B.A. (2005). Distinct stem cell populationsregenerate the follicle and interfollicular epidermis. Dev. Cell 9, 855861.
Li,A., Simmons,P.J., and Kaur,P. (1998). Identification and isolation of candidate humankeratinocyte stem cells based on cell surface phenotype. Proc. Natl. Acad. Sci. U. S. A. 95, 39023907.
Lie,D.C., Colamarino,S.A., Song,H.J., Desire,L., Mira,H., Consiglio,A., Lein,E.S., Jessberger,S.,Lansford,H., Dearie,A.R., and Gage,F.H. (2005). Wnt signalling regulates adult hippocampalneurogenesis. Nature 437, 13701375.
Lillie,F.R. (1917). The freemartin; a study of the action of sex hormones in the foetal life ofcattle. J. Exp. Zool. 23, 371452.

69
Lin,G., Finger,E., and GutierrezRamos,J.C. (1995). Expression of CD34 in endothelial cells,hematopoietic progenitors and nervous cells in fetal and adult mouse tissues. Eur. J. Immunol.25, 15081516.
Lipinski,M.M. and Jacks,T. (1999). The retinoblastoma gene family in differentiation anddevelopment. Oncogene 18, 78737882.
Lois,C. and AlvarezBuylla,A. (1993). Proliferating subventricular zone cells in the adultmammalian forebrain can differentiate into neurons and glia. Proc. Natl. Acad. Sci. U. S. A. 90,20742077.
Lois,C. and AlvarezBuylla,A. (1994). Longdistance neuronal migration in the adult mammalianbrain. Science 264, 11451148.
Lorenz,E., Congdon,C., and Uphoff,D. (1952). Modification of acute irradiation injury in miceand guineapigs by bone marrow injections. Radiology 58, 863877.
Lowden,S. and Heath,T. (1992). Lymph pathways associated with Peyer's patches in sheep. J.Anat. 181 ( Pt 2), 209217.
Lucier,M.R., Thompson,R.E., Waire,J., Lin,A.W., Osborne,B.A., and Goldsby,R.A. (1998).Multiple sites of V lambda diversification in cattle. J. Immunol. 161, 54385444.
Luckey,C.J., Bhattacharya,D., Goldrath,A.W., Weissman,I.L., Benoist,C., and Mathis,D. (2006).Memory T and memory B cells share a transcriptional program of selfrenewal with longtermhematopoietic stem cells. Proc. Natl. Acad. Sci. U. S. A. 103, 33043309.
Lyle,S., ChristofidouSolomidou,M., Liu,Y., Elder,D.E., Albelda,S., and Cotsarelis,G. (1998). TheC8/144B monoclonal antibody recognizes cytokeratin 15 and defines the location of human hairfollicle stem cells. J. Cell Sci. 111 ( Pt 21), 31793188.
Lyman,S.D. and Jacobsen,S.E. (1998). ckit ligand and Flt3 ligand: stem/progenitor cell factorswith overlapping yet distinct activities. Blood 91, 11011134.
Mackenzie,I.C. (1997). Retroviral transduction of murine epidermal stem cells demonstratesclonal units of epidermal structure. J. Invest. Dermatol. 109, 377383.
Maillard,I., Adler,S.H., and Pear,W.S. (2003). Notch and the immune system. Immunity 19, 781791.
Majdic,O., Stockl,J., Pickl,W.F., Bohuslav,J., Strobl,H., Scheinecker,C., Stockinger,H., andKnapp,W. (1994). Signaling and induction of enhanced cytoadhesiveness via the hematopoieticprogenitor cell surface molecule CD34. Blood 83, 12261234.
Marcum,J. (1974). The freemartin syndrome. Anim. Breed. Abstr. 42, 227242.
Massengale,M., Wagers,A.J., Vogel,H., and Weissman,I.L. (2005). Hematopoietic cells maintainhematopoietic fates upon entering the brain. J. Exp. Med. 201, 15791589.
Matsuzaki,Y., Kinjo,K., Mulligan,R.C., and Okano,H. (2004). Unexpectedly efficient homingcapacity of purified murine hematopoietic stem cells. Immunity 20, 8793.
Maximow,A.A. (1906). Über experimentelle Erzeugung von KnochenmarksGewebe.Anatomischer Anzeiger 28, 609612.
McGrath,K.E. and Palis,J. (2005). Hematopoiesis in the yolk sac: more than meets the eye. Exp.Hematol. 33, 10211028.
McKinneyFreeman,S.L., Jackson,K.A., Camargo,F.D., Ferrari,G., Mavilio,F., and Goodell,M.A.(2002). Musclederived hematopoietic stem cells are hematopoietic in origin. Proc. Natl. Acad.Sci. U. S. A. 99, 13411346.
Medvinsky,A. and Dzierzak,E. (1996). Definitive hematopoiesis is autonomously initiated by theAGM region. Cell 86, 897906.
Meletis,K., Wirta,V., Hede,S.M., Nister,M., Lundeberg,J., and Frisen,J. (2006). p53 suppresses theselfrenewal of adult neural stem cells. Development 133, 363369.
Mendes,S.C., Robin,C., and Dzierzak,E. (2005). Mesenchymal progenitor cells localize withinhematopoietic sites throughout ontogeny. Development 132, 11271136.

References
70
Mercier,F., Kitasako,J.T., and Hatton,G.I. (2002). Anatomy of the brain neurogenic zonesrevisited: fractones and the fibroblast/macrophage network. J. Comp. Neurol. 451, 170188.
Merok,J.R., Lansita,J.A., Tunstead,J.R., and Sherley,J.L. (2002). Cosegregation of chromosomescontaining immortal DNA strands in cells that cycle with asymmetric stem cell kinetics. CancerRes. 62, 67916795.
Mezey,E., Chandross,K.J., Harta,G., Maki,R.A., and McKercher,S.R. (2000). Turning blood intobrain: cells bearing neuronal antigens generated in vivo from bone marrow. Science 290, 17791782.
Mezey,E., Key,S., Vogelsang,G., Szalayova,I., Lange,G.D., and Crain,B. (2003). Transplantedbone marrow generates new neurons in human brains. Proc. Natl. Acad. Sci. U. S. A.
Miettinen,M., Lindenmayer,A.E., and Chaubal,A. (1994). Endothelial cell markers CD31, CD34,and BNH9 antibody to H and Yantigens evaluation of their specificity and sensitivity in thediagnosis of vascular tumors and comparison with von Willebrand factor. Mod. Pathol. 7, 8290.
Mikkers,H. and Frisen,J. (2005). Deconstructing stemness. Embo Journal 24, 27152719.
Mikkola,H.K. and Orkin,S.H. (2006). The journey of developing hematopoietic stem cells.Development 133, 37333744.
Min,H., MontecinoRodriguez,E., and Dorshkind,K. (2006). Effects of aging on the commonlymphoid progenitor to proB cell transition. J. Immunol. 176, 10071012.
Ming,G.L. and Song,H. (2005). Adult neurogenesis in the mammalian central nervous system.Annu. Rev. Neurosci. 28, 223250.
Modi,W.S., Gallagher,D.S., and Womack,J.E. (1993). Molecular organization and chromosomallocalization of six highly repeated DNA families in the bovine genome. Anim. Biotechnol. 4,143161.
Moore,K.A. and Lemischka,I.R. (2006). Stem cells and their niches. Science 311, 18801885.
Morrison,S.J., Hemmati,H.D., Wandycz,A.M., and Weissman,I.L. (1995). The purification andcharacterization of fetal liver hematopoietic stem cells. Proc. Natl. Acad. Sci. U. S. A. 92, 1030210306.
Morrison,S.J. and Kimble,J. (2006). Asymmetric and symmetric stemcell divisions indevelopment and cancer. Nature 441, 10681074.
Morrison,S.J., Wandycz,A.M., Hemmati,H.D., Wright,D.E., and Weissman,I.L. (1997).Identification of a lineage of multipotent hematopoietic progenitors. Development 124, 19291939.
Morshead,C.M., Benveniste,P., Iscove,N.N., and van der Kooy,D. (2002). Hematopoieticcompetence is a rare property of neural stem cells that may depend on genetic and epigeneticalterations. Nat. Med. 8, 268273.
Motyka,B. and Reynolds,J.D. (1991). Apoptosis is associated with the extensive B cell death inthe sheep ileal Peyer's patch and the chicken bursa of Fabricius: a possible role in B cellselection. Eur. J. Immunol. 21, 19511958.
Muguruma,Y., Reyes,M., Nakamura,Y., Sato,T., Matsuzawa,H., Miyatake,H., Akatsuka,A., Itoh,J.,Yahata,T., Ando,K., Kato,S., and Hotta,T. (2003). In vivo and in vitro differentiation of myocytesfrom human bone marrowderived multipotent progenitor cells. Exp. Hematol. 31, 13231330.
MullerSieburg,C.E., Whitlock,C.A., and Weissman,I.L. (1986). Isolation of two early Blymphocyte progenitors from mouse marrow: a committed prepreB cell and a clonogenic Thy1lo hematopoietic stem cell. Cell 44, 653662.
Murry,C.E., Soonpaa,M.H., Reinecke,H., Nakajima,H., Nakajima,H.O., Rubart,M.,Pasumarthi,K.B., Virag,J.I., Bartelmez,S.H., Poppa,V., Bradford,G., Dowell,J.D., Williams,D.A.,and Field,L.J. (2004). Haematopoietic stem cells do not transdifferentiate into cardiac myocytesin myocardial infarcts. Nature 428, 664668.
Nakamura,Y., Komano,H., and Nakauchi,H. (1993). Two alternative forms of cDNA encodingCD34. Exp. Hematol. 21, 236242.

71
Nayernia,K., Lee,J.H., Drusenheimer,N., Nolte,J., Wulf,G., Dressel,R., Gromoll,J., and Engel,W.(2006). Derivation of male germ cells from bone marrow stem cells. Lab Invest 86, 654663.
Neill,J.D., Ridpath,J.F., and LieblerTenorio,E. (2006). Global gene expression profiling of bovineimmature B cells using serial analysis of gene expression. Anim. Biotechnol. 17, 2131.
Nottebohm,F. (2004). The road we travelled: discovery, choreography, and significance of brainreplaceable neurons. Ann. N. Y. Acad. Sci. 1016, 628658.
Nowell,P.C., Cole,L.J., Habermeyer,J.G., and Roan,P.L. (1956). Growth and continued function ofrat marrow cells in xradiated mice. Cancer Res. 16, 258261.
Nunez,C., Nishimoto,N., Gartland,G.L., Billips,L.G., Burrows,P.D., Kubagawa,H., andCooper,M.D. (1996). B cells are generated throughout life in humans. J. Immunol. 156, 866872.
Ogawa,M. (1993). Differentiation and proliferation of hematopoietic stem cells. Blood 81, 28442853.
Ohno,S. and Gropp,A. (1965). Embryological basis for germ cell chimerism in mammals.Cytogenetics. 4, 251261.
Okamoto,R., Yajima,T., Yamazaki,M., Kanai,T., Mukai,M., Okamoto,S., Ikeda,Y., Hibi,T.,Inazawa,J., and Watanabe,M. (2002). Damaged epithelia regenerated by bone marrowderivedcells in the human gastrointestinal tract. Nat. Med. 8, 10111017.
Okuno,Y., Iwasaki,H., Huettner,C.S., Radomska,H.S., Gonzalez,D.A., Tenen,D.G., and Akashi,K.(2002). Differential regulation of the human and murine CD34 genes in hematopoietic stemcells. Proc. Natl. Acad. Sci. U. S. A. 99, 62466251.
Orlic,D., Kajstura,J., Chimenti,S., Limana,F., Jakoniuk,I., Quaini,F., NadalGinard,B.,Bodine,D.M., Leri,A., and Anversa,P. (2001). Mobilized bone marrow cells repair the infarctedheart, improving function and survival. Proc. Natl. Acad. Sci. U. S. A. 98, 1034410349.
Osawa,M., Hanada,K., Hamada,H., and Nakauchi,H. (1996). Longterm lymphohematopoieticreconstitution by a single CD34low/negative hematopoietic stem cell. Science 273, 242245.
Ottersbach,K. and Dzierzak,E. (2005). The murine placenta contains hematopoietic stem cellswithin the vascular labyrinth region. Dev. Cell 8, 377387.
Owen,R.D. (1945). Immunogenetic consequenses of vascular anastomoses between bovine twins.Science 102, 400401.
Pabst,R. and Reynolds,J.D. (1986). Evidence of extensive lymphocyte death in sheep Peyer'spatches. II. The number and fate of newlyformed lymphocytes that emigrate from Peyer'spatches. J Immunol. 136, 20112017.
Pabst,R. and Reynolds,J.D. (1987). Peyer's patches export lymphocytes throughout the lymphoidsystem in sheep. J Immunol. 139, 39813985.
Padula,A.M. (2005). The freemartin syndrome: an update. Anim. Reprod. Sci. 87, 93109.
Palmer,T.D., Takahashi,J., and Gage,F.H. (1997). The adult rat hippocampus contains primordialneural stem cells. Mol. Cell Neurosci. 8, 389404.
Palmer,T.D., Willhoite,A.R., and Gage,F.H. (2000). Vascular niche for adult hippocampalneurogenesis. J. Comp. Neurol. 425, 479494.
Parng,C.L., Hansal,S., Goldsby,R.A., and Osborne,B.A. (1996). Gene conversion contributes to Iglight chain diversity in cattle. J. Immunol. 157, 54785486.
Parsons,K.R., Sopp,P., Jones,B.V., Bland,P., and Howard,C.J. (1996). Identification of a moleculeuniquely expressed on a gamma/delta TCR+ subset within bovine intestinal intraepitheliallymphocytes. Immunology 87, 6470.
Perret,J., Shia,Y.C., Fries,R., Vassart,G., and Georges,M. (1990). A polymorphic satellite sequencemaps to the pericentric region of the bovine Y chromosome. Genomics 6, 482490.
Petersen,B.E., Bowen,W.C., Patrene,K.D., Mars,W.M., Sullivan,A.K., Murase,N., Boggs,S.S.,Greenberger,J.S., and Goff,J.P. (1999). Bone marrow as a potential source of hepatic oval cells.Science 284, 11681170.

References
72
Petrakis,N.L., Lucia,S.P., and Davis,M. (1961). In vivo differentiation of human leukocytes intohistiocytes, fibroblasts and fat cells in subcutaneous diffusion chambers. Blood 17, 109118.
Pittenger,M.F., Mackay,A.M., Beck,S.C., Jaiswal,R.K., Douglas,R., Mosca,J.D., Moorman,M.A.,Simonetti,D.W., Craig,S., and Marshak,D.R. (1999). Multilineage potential of adult humanmesenchymal stem cells. Science 284, 143147.
Ponder,B.A., Schmidt,G.H., Wilkinson,M.M., Wood,M.J., Monk,M., and Reid,A. (1985).Derivation of mouse intestinal crypts from single progenitor cells. Nature 313, 689691.
Potten,C.S. (1998). Stem cells in gastrointestinal epithelium: numbers, characteristics and death.Philos. Trans. R. Soc. Lond. B Biol. Sci. 353, 821830.
Potten,C.S. (1975b). Epidermal cell production rates. J. Invest. Dermatol. 65, 488500.
Potten,C.S. (1975a). Epidermal transit times. Br. J. Dermatol. 93, 649658.
Potten,C.S., Booth,C., Tudor,G.L., Booth,D., Brady,G., Hurley,P., Ashton,G., Clarke,R.,Sakakibara,S., and Okano,H. (2003). Identification of a putative intestinal stem cell and earlylineage marker; musashi1. Differentiation 71, 2841.
Potten,C.S., Owen,G., and Booth,D. (2002). Intestinal stem cells protect their genome byselective segregation of template DNA strands. J. Cell Sci. 115, 23812388.
Press,C.M., McCullagh,P., and Landsverk,T. (2001). Effect of early fetal splenectomy on prenatalBcell development in sheep. Immunology 102, 131136.
Press,C.M., Reynolds,J.D., McClure,S.J., SimpsonMorgan,M.W., and Landsverk,T. (1996). Fetallambs are depleted of IgM+ cells following a single injection of an antiIgM antibody early ingestation. Immunology 88, 2834.
Priller,J., Persons,D.A., Klett,F.F., Kempermann,G., Kreutzberg,G.W., and Dirnagl,U. (2001).Neogenesis of cerebellar Purkinje neurons from genemarked bone marrow cells in vivo. J. CellBiol. 155, 733738.
Prise,K.M., Schettino,G., Folkard,M., and Held,K.D. (2005). New insights on cell death fromradiation exposure. Lancet Oncol. 6, 520528.
Prockop,D.J. (1997). Marrow stromal cells as stem cells for nonhematopoietic tissues. Science276, 7174.
Raff,M. (2003). Adult stem cell plasticity: fact or artifact? Annu. Rev. Cell Dev. Biol. 19, 122.
RamalhoSantos,M., Yoon,S., Matsuzaki,Y., Mulligan,R.C., and Melton,D.A. (2002). "Stemness":transcriptional profiling of embryonic and adult stem cells. Science 298, 597600.
Rangan,S.R. (1967). Origin of the fibroblastic growths in chicken buffy coat macrophagecultures. Exp. Cell Res. 46, 477487.
Ratcliffe,M.J. (2002). B cell development in gut associated lymphoid tissues. Vet. Immunol.Immunopathol. 87, 337340.
Reya,T. and Clevers,H. (2005). Wnt signalling in stem cells and cancer. Nature 434, 843850.
Reyes,M. and Verfaillie,C.M. (2001). Characterization of multipotent adult progenitor cells, asubpopulation of mesenchymal stem cells. Ann. N. Y. Acad. Sci. 938, 231233.
Reynaud,C.A., Garcia,C., Hein,W.R., and Weill,J.C. (1995). Hypermutation generating the sheepimmunoglobulin repertoire is an antigenindependent process. Cell 80, 115125.
Reynaud,C.A., Mackay,C.R., Muller,R.G., and Weill,J.C. (1991). Somatic generation of diversityin a mammalian primary lymphoid organ: the sheep ileal Peyer's patches. Cell 64, 9951005.
Reynolds,B.A. and Weiss,S. (1992). Generation of neurons and astrocytes from isolated cells ofthe adult mammalian central nervous system. Science 255, 17071710.
Reynolds,B.A. and Weiss,S. (1996). Clonal and population analyses demonstrate that an EGFresponsive mammalian embryonic CNS precursor is a stem cell. Dev. Biol. 175, 113.
Reynolds,J. (1997). The genesis, tutelage and exodus of B cells in the ileal Peyer's patch of sheep.Int. Rev. Immunol. 15, 265299.

73
Reynolds,J.D. (1986). Evidence of extensive lymphocyte death in sheep Peyer's patches. I. Acomparison of lymphocyte production and export. J Immunol. 136, 20052010.
Reynolds,J.D. (1987). Mitotic rate maturation in the Peyer's patches of fetal sheep and in thebursa of Fabricius of the chick embryo. Eur. J. Immunol. 17, 503507.
Reynolds,J.D., Kennedy,L., Peppard,J., and Pabst,R. (1991). Ileal Peyer's patch emigrants arepredominantly B cells and travel to all lymphoid tissues in sheep. Eur. J. Immunol. 21, 283289.
Reynolds,J.D. and Morris,B. (1983a). The influence of gut function on lymphoid cell populationsin the intestinal mucosa of lambs. Immunology 49, 501509.
Reynolds,J.D. and Morris,B. (1984). The effect of antigen on the development of Peyer's patchesin sheep. Eur. J. Immunol. 14, 16.
Reynolds,J.D. and Morris,B. (1983b). The evolution and involution of Peyer's patches in fetal andpostnatal sheep. Eur. J. Immunol. 13, 627635.
Rizvi,A.Z., Swain,J.R., Davies,P.S., Bailey,A.S., Decker,A.D., Willenbring,H., Grompe,M.,Fleming,W.H., and Wong,M.H. (2006). Bone marrowderived cells fuse with normal andtransformed intestinal stem cells. Proc. Natl. Acad. Sci. U. S. A. 103, 63216325.
Rizvi,A.Z. and Wong,M.H. (2005). Epithelial stem cells and their niche: there's no place likehome. Stem Cells 23, 150165.
Rochat,A., Kobayashi,K., and Barrandon,Y. (1994). Location of stem cells of human hair folliclesby clonal analysis. Cell 76, 10631073.
Ross,M., Kaye,G., and Pawlina,W. (2003). Histology. A text and atlas. (Baltimore: LippincotWilliams & Wilkins).
Ross,R., Everett,N.B., and Tyler,R. (1970). Wound healing and collagen formation. VI. The originof the wound fibroblast studied in parabiosis. J. Cell Biol. 44, 645654.
Roybon,L., Ma,Z., Asztely,F., Fosum,A., Jacobsen,S.E., Brundin,P., and Li,J.Y. (2006). Failure oftransdifferentiation of adult hematopoietic stem cells into neurons. Stem Cells 24, 15941604.
Rüsse,I. (1991). Lehrbuch der Embryologie der Haustiere. (Berlin: Parey).
Saini,S.S., Allore,B., Jacobs,R.M., and Kaushik,A. (1999). Exceptionally long CDR3H region withmultiple cysteine residues in functional bovine IgM antibodies. Eur. J. Immunol. 29, 24202426.
Saini,S.S., Hein,W.R., and Kaushik,A. (1997). A single predominantly expressed polymorphicimmunoglobulin VH gene family, related to mammalian group, I, clan, II, is identified in cattle.Mol. Immunol. 34, 641651.
Saini,S.S. and Kaushik,A. (2002). Extensive CDR3H length heterogeneity exists in bovine foetalVDJ rearrangements. Scand. J. Immunol. 55, 140148.
Sanai,N., Tramontin,A.D., QuinonesHinojosa,A., Barbaro,N.M., Gupta,N., Kunwar,S.,Lawton,M.T., McDermott,M.W., Parsa,A.T., ManuelGarcia,V.J., Berger,M.S., and AlvarezBuylla,A. (2004). Unique astrocyte ribbon in adult human brain contains neural stem cells butlacks chain migration. Nature 427, 740744.
Seidel,F. (1952). Die Entwicklungspotenzen einer isolierten Blastomere des Zweizellenstadiumsim Saugetierei. Naturwissenschaften 39, 355356.
Sell,S. (2004). Stem cells. In Stem cells handbook, S.Sell, ed. (Totowa: Humana Press), pp. 118.
Seri,B., GarciaVerdugo,J.M., McEwen,B.S., and AlvarezBuylla,A. (2001). Astrocytes give rise tonew neurons in the adult mammalian hippocampus. J. Neurosci. 21, 71537160.
Shailubhai,K., Streeter,P.R., Smith,C.E., and Jacob,G.S. (1997). Sulfation and sialylationrequirements for a glycoform of CD34, a major endothelial ligand for Lselectin in porcineperipheral lymph nodes. Glycobiology 7, 305314.
Shinin,V., GayraudMorel,B., Gomes,D., and Tajbakhsh,S. (2006). Asymmetric division andcosegregation of template DNA strands in adult muscle satellite cells. Nat. Cell Biol.
Shojaei,F., Saini,S.S., and Kaushik,A.K. (2003). Unusually long germline DH genes contribute tolarge sized CDR3H in bovine antibodies. Mol. Immunol. 40, 6167.

References
74
Sidman,R., Angevine,J., and Pierce,E. (1971). Atlas of the mouse brain and spinal cord.(Cambridge, Massachusetts: Harvard University Press).
Simmons,P.J. and TorokStorb,B. (1991). Identification of stromal cell precursors in human bonemarrow by a novel monoclonal antibody, STRO1. Blood 78, 5562.
Sinclair,M.C., Gilchrist,J., and Aitken,R. (1997). Bovine IgG repertoire is dominated by a singlediversified VH gene family. J. Immunol. 159, 38833889.
Skowronski,J., Plucienniczak,A., Bednarek,A., and Jaworski,J. (1984). Bovine 1.709 satellite.Recombination hotspots and dispersed repeated sequences. J Mol. Biol. 177, 399416.
Smith,G.H. (2005). Labelretaining epithelial cells in mouse mammary gland divideasymmetrically and retain their template DNA strands. Development 132, 681687.
Smith,J.R., Pochampally,R., Perry,A., Hsu,S.C., and Prockop,D.J. (2004). Isolation of a highlyclonogenic and multipotential subfraction of adult stem cells from bone marrow stroma. StemCells 22, 823831.
Smith,L.G., Weissman,I.L., and Heimfeld,S. (1991). Clonal analysis of hematopoietic stemcelldifferentiation in vivo. Proc. Natl. Acad. Sci. U. S. A. 88, 27882792.
Song,H.J., Stevens,C.F., and Gage,F.H. (2002). Neural stem cells from adult hippocampusdevelop essential properties of functional CNS neurons. Nat. Neurosci. 5, 438445.
Song,L. and Tuan,R.S. (2004). Transdifferentiation potential of human mesenchymal stem cellsderived from bone marrow. FASEB J. 18, 980982.
Spalding,K.L., Bhardwaj,R.D., Buchholz,B.A., Druid,H., and Frisen,J. (2005). Retrospective birthdating of cells in humans. Cell 122, 133143.
Spangrude,G.J., Heimfeld,S., and Weissman,I.L. (1988). Purification and characterization ofmouse hematopoietic stem cells. Science 241, 5862.
Spradling,A., DrummondBarbosa,D., and Kai,T. (2001). Stem cells find their niche. Nature 414,98104.
Stanfield,B.B. and Trice,J.E. (1988). Evidence that granule cells generated in the dentate gyrus ofadult rats extend axonal projections. Exp. Brain Res. 72, 399406.
Stevens,A.M., McDonnell,W.M., Mullarkey,M.E., Pang,J.M., Leisenring,W., and Nelson,J.L.(2004). Liver biopsies from human females contain male hepatocytes in the absence oftransplantation. Lab Invest 84, 16031609.
Stewart,K., Monk,P., Walsh,S., Jefferiss,C.M., Letchford,J., and Beresford,J.N. (2003). STRO1,HOP26 (CD63), CD49a and SB10 (CD166) as markers of primitive human marrow stromal cellsand their more differentiated progeny: a comparative investigation in vitro. Cell Tissue Res. 313,281290.
Suda,J., Suda,T., and Ogawa,M. (1984). Analysis of differentiation of mouse hemopoietic stemcells in culture by sequential replating of paired progenitors. Blood 64, 393399.
Suda,J., Sudo,T., Ito,M., Ohno,N., Yamaguchi,Y., and Suda,T. (1992). Two types of murine CD34mRNA generated by alternative splicing. Blood 79, 22882295.
Takano,H., Ema,H., Sudo,K., and Nakauchi,H. (2004). Asymmetric division and lineagecommitment at the level of hematopoietic stem cells: inference from differentiation in daughtercell and granddaughter cell pairs. J. Exp. Med. 199, 295302.
Terada,N., Hamazaki,T., Oka,M., Hoki,M., Mastalerz,D.M., Nakano,Y., Meyer,E.M., Morel,L.,Petersen,B.E., and Scott,E.W. (2002). Bone marrow cells adopt the phenotype of other cells byspontaneous cell fusion. Nature 416, 542545.
Theise,N.D., Nimmakayalu,M., Gardner,R., Illei,P.B., Morgan,G., Teperman,L., Henegariu,O.,and Krause,D.S. (2000). Liver from bone marrow in humans. Hepatology 32, 1116.
Thomas,M.L. (1989). The leukocyte common antigen family. Annu. Rev. Immunol. 7, 339369.
Thompson,J.D., Higgins,D.G., and Gibson,T.J. (1994). CLUSTAL W: improving the sensitivity ofprogressive multiple sequence alignment through sequence weighting, positionspecific gappenalties and weight matrix choice. Nucleic Acids Res. 22, 46734680.

75
Till,J.E. and McCulloch,E.A. (1961). A direct measurement of the radiation sensitivity of normalmouse bone marrow cells. Radiat. Res. 14, 213222.
Tizard,I. (2004). Veterinary immunology. An introduction. (Philadelphia: Saunders).
Toma,C., Pittenger,M.F., Cahill,K.S., Byrne,B.J., and Kessler,P.D. (2002). Human mesenchymalstem cells differentiate to a cardiomyocyte phenotype in the adult murine heart. Circulation 105,9398.
Torensma,R. and Figdor,C.G. (2004). Differentiating stem cells mask their origins. Stem Cells 22,250252.
Trempus,C.S., Morris,R.J., Bortner,C.D., Cotsarelis,G., Faircloth,R.S., Reece,J.M., andTennant,R.W. (2003). Enrichment for living murine keratinocytes from the hair follicle bulgewith the cell surface marker CD34. J. Invest. Dermatol. 120, 501511.
Trier,J.S. and Moxey,P.C. (1979). Morphogenesis of the small intestine during fetal development.Ciba Found. Symp. 70, 329.
Tumbar,T., Guasch,G., Greco,V., Blanpain,C., Lowry,W.E., Rendl,M., and Fuchs,E. (2004).Defining the epithelial stem cell niche in skin. Science 303, 359363.
Uchida,N., Buck,D.W., He,D., Reitsma,M.J., Masek,M., Phan,T.V., Tsukamoto,A.S., Gage,F.H.,and Weissman,I.L. (2000). Direct isolation of human central nervous system stem cells. Proc.Natl. Acad. Sci. U. S. A. 97, 1472014725.
Väänänen,H.K. (2005). Mesenchymal stem cells. Ann. Med. 37, 469479.
ValkLingbeek,M.E., Bruggeman,S.W., and van Lohuizen,M. (2004). Stem cells and cancer; thepolycomb connection. Cell 118, 409418.
van Es,J.H., Jay,P., Gregorieff,A., van Gijn,M.E., Jonkheer,S., Hatzis,P., Thiele,A., van denBorn,M., Begthel,H., Brabletz,T., Taketo,M.M., and Clevers,H. (2005). Wnt signalling inducesmaturation of Paneth cells in intestinal crypts. Nat. Cell Biol. 7, 381386.
van Praag,H., Schinder,A.F., Christie,B.R., Toni,N., Palmer,T.D., and Gage,F.H. (2002).Functional neurogenesis in the adult hippocampus. Nature 415, 10301034.
van Zant,G. and Liang,Y. (2003). The role of stem cells in aging. Exp. Hematol. 31, 659672.
Vigier,B., Prepin,J., Perchellet,J., and Jost,A. (1977). Développement de l'effet freemartin chez lefoetus de veau. Ann. Méd. Vét. 521536.
Vigier,B., Watrin,F., Magre,S., Tran,D., and Josso,N. (1987). Purified bovine AMH induces acharacteristic freemartin effect in fetal rat prospective ovaries exposed to it in vitro.Development 100, 4355.
Vignery,A. (2005). Macrophage fusion: the making of osteoclasts and giant cells. J. Exp. Med.202, 337340.
Viljanto,J. and Kivikoski,A. (1962). Local effect of neutral saltsoluble collagen on formation ofgranulation tissue a histological study with viscose cellulose implants on rats. Ann. Med. Exp.Biol. Fenn. 40, 118127.
Visnjic,D., Kalajzic,Z., Rowe,D.W., Katavic,V., Lorenzo,J., and Aguila,H.L. (2004). Hematopoiesisis severely altered in mice with an induced osteoblast deficiency. Blood 103, 32583264.
Wagers,A.J., Sherwood,R.I., Christensen,J.L., and Weissman,I.L. (2002). Little evidence fordevelopmental plasticity of adult hematopoietic stem cells. Science 297, 22562259.
Wagers,A.J. and Weissman,I.L. (2004). Plasticity of adult stem cells. Cell 116, 639648.
Wang,X., Willenbring,H., Akkari,Y., Torimaru,Y., Foster,M., AlDhalimy,M., Lagasse,E.,Finegold,M., Olson,S., and Grompe,M. (2003). Cell fusion is the principal source of bonemarrowderived hepatocytes. Nature 422, 897901.
Weill,J.C. and Reynaud,C.A. (1998). Galt versus bone marrow models of B cell ontogeny. Dev.Comp. Immunol. 22, 379385.
Weimann,J.M., Charlton,C.A., Brazelton,T.R., Hackman,R.C., and Blau,H.M. (2003).Contribution of transplanted bone marrow cells to Purkinje neurons in human adult brains.Proc. Natl. Acad. Sci. U. S. A. 100, 20882093.

References
76
Weissman,I.L. (2000). Translating stem and progenitor cell biology to the clinic: barriers andopportunities. Science 287, 14421446.
Wilkes,P.R., Wijeratne,W.V., and Munro,I.B. (1981). Reproductive anatomy and cytogenetics offreemartin heifers. Vet. Rec. 108, 349353.
Wilson,A., Murphy,M.J., Oskarsson,T., Kaloulis,K., Bettess,M.D., Oser,G.M., Pasche,A.C.,Knabenhans,C., Macdonald,H.R., and Trumpp,A. (2004). cMyc controls the balance betweenhematopoietic stem cell selfrenewal and differentiation. Genes Dev. 18, 27472763.
Wilson,A. and Trumpp,A. (2006). Bonemarrow haematopoieticstemcell niches. Nat. Rev.Immunol. 6, 93106.
Wright,D.E., Wagers,A.J., Gulati,A.P., Johnson,F.L., and Weissman,I.L. (2001). Physiologicalmigration of hematopoietic stem and progenitor cells. Science 294, 19331936.
Wurmser,A.E., Nakashima,K., Summers,R.G., Toni,N., D'Amour,K.A., Lie,D.C., and Gage,F.H.(2004). Cell fusionindependent differentiation of neural stem cells to the endothelial lineage.Nature 430, 350356.
Yan,Z., Lambert,N.C., Guthrie,K.A., Porter,A.J., Loubiere,L.S., Madeleine,M.M., Stevens,A.M.,Hermes,H.M., and Nelson,J.L. (2005). Male microchimerism in women without sons:quantitative assessment and correlation with pregnancy history. Am. J. Med. 118, 899906.
Yasuda,M., Jenne,C.N., Kennedy,L.J., and Reynolds,J.D. (2006). The sheep and cattle Peyer'spatch as a site of Bcell development. Vet. Res. 37, 401415.
Young,P.E., Baumhueter,S., and Lasky,L.A. (1995). The sialomucin CD34 is expressed onhematopoietic cells and blood vessels during murine development. Blood 85, 96105.
Zanjani,E.D., AlmeidaPorada,G.A., Livingston,A.G., Flake,A.W., and Ogawa,M. (1998). Humanbone marrow CD34 cells engraft in vivo and undergo multilineage expression that includesgiving rise to CD34+ cells. Exp. Hematol. 26, 353360.
Zhang,B., Pan,X., and Anderson,T.A. (2006a). MicroRNA: A new player in stem cells. J. CellPhysiol.
Zhang,J., Grindley,J.C., Yin,T., Jayasinghe,S., He,X.C., Ross,J.T., Haug,J.S., Rupp,D., PorterWestpfahl,K.S., Wiedemann,L.M., Wu,H., and Li,L. (2006b). PTEN maintains haematopoieticstem cells and acts in lineage choice and leukaemia prevention. Nature 441, 518522.
Zhang,J., Niu,C., Ye,L., Huang,H., He,X., Tong,W.G., Ross,J., Haug,J., Johnson,T., Feng,J.Q.,Harris,S., Wiedemann,L.M., Mishina,Y., and Li,L. (2003). Identification of the haematopoieticstem cell niche and control of the niche size. Nature 425, 836841.
Zhao,Y., Jackson,S.M., and Aitken,R. (2006). The bovine antibody repertoire. Dev. Comp.Immunol. 30, 175186.
Zhou,J.H., Hikono,H., Ohta,M., and Sakurai,M. (2001a). Cloning of bovine CD34 cDNA. J. Vet.Med. Sci. 63, 10511053.
Zhou,S., Schuetz,J.D., Bunting,K.D., Colapietro,A.M., Sampath,J., Morris,J.J., Lagutina,I.,Grosveld,G.C., Osawa,M., Nakauchi,H., and Sorrentino,B.P. (2001b). The ABC transporterBcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of thesidepopulation phenotype. Nat. Med. 7, 10281034.
Ziegler,B.L., Valtieri,M., Almeida Porada,G.A., De,M.R., Muller,R., Masella,B., Gabbianelli,M.,Casella,I., Pelosi,E., Bock,T., Zanjani,E.D., and Peschle,C. (1999). KDR receptor: a key markerdefining hematopoietic stem cells. Science 285, 15531558.
Zipori,D. (2004). The nature of stem cells: state rather than entity. Nat. Rev. Genet. 5, 873878.
Zuk,P.A., Zhu,M., Ashjian,P., De Ugarte,D.A., Huang,J.I., Mizuno,H., Alfonso,Z.C., Fraser,J.K.,Benhaim,P., and Hedrick,M.H. (2002). Human adipose tissue is a source of multipotent stemcells. Mol. Biol. Cell 13, 42794295.

77
11. Original publications





























