Embed Size (px)
Citation preview

Disturbed Object Processing in 5xFAD and 3xTG Mouse Models of Alzheimer’s Disease: Going Beyond “Object Recognition”
by
Samantha Danielle Creighton
A Thesis
presented to
The University of Guelph
In partial fulfilment of requirements
for the degree of
Master of Science
in
Psychology and Neuroscience
Guelph, Ontario, Canada
© Samantha Danielle Creighton, August, 2016

ABSTRACT
DISTURBED OBJECT PROCESSING IN 5XFAD AND 3XTG MOUSE MODELS
OF ALZHEIMER’S DISEASE: GOING BEYOND “OBJECT RECOGNITION”
Samantha Danielle Creighton Advisor:
University of Guelph, 2016 Dr. Boyer Winters
Object recognition not a unitary process, and there are many uncharacterized
facets of object processing that have relevance to Alzheimer’s disease (AD). To elucidate
the specific nature of object processing deficits in transgenic mouse models of AD, we
systematically evaluated performance on tasks that manipulate different types of object
information: object identity (i.e., object recognition: OR), spatial processing (object
location; OL), temporal processing (temporal order; TO), and multisensory perception
(multisensory object oddity; MSO) in 12-month-old male and female 5xFAD and 3xTG
mice. 5xFAD mice were impaired on OR, OL, TO and MSO. Conversely, 3xTG females
had intact short-term OR, and when spatial cues were minimized both males and females
had intact short-term OR. 3xTG mice had impaired OL, TO and MSO. Results reveal
dissociations across AD strain, sex, and type of object processing, and should help to
clarify the relationship between specific aspects of AD pathology and object-related
information processing.

iii
ACKNOWLEDGEMENTS
I would like to thank my advisor, Dr. Boyer Winters for his support and guidance. I
would also like to thank my advisory committee members Dr. Elena Choleris and Dr.
Craig Bailey, for their valuable feedback. Finally, I would like to thank everyone in the
Winters lab for their help. In particular, I would like to thank Daniel Palmer for this
assistance with object processing experiments.

iv
Table of Contents
Abstract Acknowledgments iii
Table of Contents iv List of Tables v List of Figures vi
List of Abbreviations vii Introduction 1
Alzheimer’s Disease Neuropathology 1 Episodic Memory Deficits in Alzheimer’s Disease 8 Modeling Alzheimer’s Disease in Rodents 10
Modeling Recognition Memory in Rodents 13 Recognition Memory Deficits in Rodent Models of AD 18
Methods 22 Animals 22 Behavioural Testing 23
Experiment 1: Open-field & Y-Apparatus Object Recognition 25 Experiment 2: Object Location 26
Experiment 3: Temporal Order 26 Experiment 4: Multisensory Object Oddity 27 Behavioural Data Analysis 28
Results 29 Experiment 1: 29
Open-field Object Recognition 29 Y-Apparatus Object Recognition 30
Experiment 2: Object Location 34
Experiment 3: Temporal Order 36 Experiment 4: Multisensory Object Oddity 37
Discussion 40 Object Recognition 41 Object Location 45
Temporal Order 48 Multisensory Object Oddity 50
General Discussion 51 References 57 Appendix 121

v
List of Tables
Table 1: Transgenic Mouse Models of Alzheimer’s Disease
Table 2: Object Recognition Deficits in Transgenic Mouse Models of Alzheimer’s Disease
Table 3: Correlation Between Total Sample Exploration and Novelty Preference Index
Table 4: Object Processing Results Summary

vi
List of Figures
Figure 1: Object processing test battery.
Figure 2: 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) open field object recognition.
Figure 3: 5xFAD and 3xTG open-field object recognition total exploration.
Figure 4: 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) Y-apparatus object recognition.
Figure 5: 5xFAD and 3xTG Y-apparatus object recognition total exploration.
Figure 6: 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) object location.
Figure 7: 5xFAD and 3xTG object location total exploration.
Figure 8: 5xFAD and 3xTG performance on short-term (3min) object temporal order.
Figure 9: 5xFAD and 3xTG object temporal order total exploration.
Figure 10: 5xFAD and 3xTG performance on object oddity. Figure 11: 5xFAD and 3xTG object oddity total exploration.

vii
List of Abbreviations
Aβ - β-Amyloid
ACh - Acetylcholine
AD - Alzheimer’s Disease
aMCI - Amnesic Mild Cognitive Impairment
APP - Amyloid Precursor Protein
ChATi - Cholinesterase Inhibitors
CMOR - Cross-modal object recognition
CNS - Central Nervous System
DMS - Delayed Matching to Sample
DNMS - Delays Non-Matching to Sample
DR - Discrimination Ratio
HPC - Hippocampus
mPFC - Medial Prefrontal Cortex
MSO - Multisensory Object Oddity
MTL - Medial Temporal Lobe
OFC - Orbitofrontal Cortex
OL - Object Location
OR - Object Recognition
PPC - Posterior Parietal Cortex
PRh - Perirhinal Cortex
PS1 - Presenilin 1

viii
PS2 - Presenilin 2
SOR - Spontaneous Object Recognition
TG - Transgenic
TO - Temporal Order
VRM - Visual Recognition Memory
WT - Wild-type

1
Introduction
Alzheimer’s disease (AD) is a fatal incapacitating neurodegenerative disorder
characterized by a progressive dementia that is correlated with brain pathology. Alois
Alzheimer first described AD as a novel disease distinguishable from senile dementia by
rapid early-onset cognitive decline that co-occurred with an atrophic brain defined by the
deposition of plaques and tangles of fibrils in the cerebral cortex (Alzheimer, 1906, 1911;
Maurer, Volk, & Gerbaldo, 1997; Stelzmann, Schnitzlein, & Murtagh, 1995). AD is now
characterized by the neuropathological accumulation of β-amyloid peptide and
hyperphosphorylated tau protein, oxidative stress and inflammation in the brain (Cuello,
2005; Duyckaerts, Delatour, & Potier, 2009; Honda & Casadesus, 2004; McLaurin,
Yang, Yip, & Fraser, 2000). While the specific etiology of the sporadic (early onset) form
of AD is unknown, the rare familial (late onset) AD is triggered by autosomal dominant
mutations in genes implicated in amyloid processing, specifically the amyloid precursor
protein (APP) gene and presenilin genes (PS1, PS2) (Vilatela, López-López, & Yescas-
Gómez, 2012; Armstrong, 2013). The accumulation of AD pathology is associated with
cognitive decline, including early deficits in executive function and declarative memory.
Alzheimer’s Disease Neuropathology
Excessive production of β-amyloid causes the formation of extracellular amyloid
plaques. β-amyloid (Aβ) has numerous physiological functions, including neural stem
cell development, neuronal survival, growth, repair, synaptic excitability, synaptic
transmission, long-term potentiation (Dawkins & Small, 2014; Turner, O’Connor, Tate,
& Abraham, 2003; Whitson, Selkoe, & Cotman, 1989), and is important for memory

2
formation (Garcia-Osta & Alberini, 2009; Morley, Farr, Banks, Johnson, & Louis, 2010).
In AD, Aβ accumulates intracellularly and extracellularly following accelerated
processing and abnormal cleavage of the membrane bound APP protein in the
amyloidogenic pathway by β- and γ-secretases (McLaurin et al., 2000; Sisodia & St
George-Hyslop, 2002). Specifically, β-secretase (β-site APP-cleaving enzyme, or BACE)
cleavage of APP, favoured by mutations in the APP gene, yields APPsβ and a β-stub
(McLaurin et al., 2000; Sisodia & St George-Hyslop, 2002). The β-stub is subsequently
cleaved by γ-secretase, to produce Aβ peptides (soluble Aβ40 and fibrillogenic Aβ42
[mutations in PS promote the overproduction of pathogenic Aβ42]) (McLaurin et al.,
2000; Sisodia & St George-Hyslop, 2002; Thinakaran & Koo, 2008). Aβ42 accumulation
is favoured by mutations in the APP gene that produce a longer Aβ peptide via
potentiation of β-secretase, inhibition of α-secretase, and γ-secretase slice alteration
(Hardy, 1997). Mutations in PS1 and PS2 also favour a long pathogenic Aβ peptide,
potentially through competitive γ-secretase binding or altered γ-secretase trafficking in
the endoplasmic reticulum (Haass, 1997; Selkoe, 1998). Aβ peptides may have
numerous intracellular consequences, such as lysosome up-regulation, mitochondrial
dysfunction, dysregulation of CRE-directed gene expression and potentially tau
phosphorylation (Cuello, 2005). As Aβ accumulates, peptides aggregate to form amyloid
fibrils which can rupture the vascular membrane of neuronal cells leading to cell death
and the additional accumulation of extracelluar Aβ (Friedrich et al., 2010). β- and γ-
secretases may also cleave membrane bound APP, resulting in increased extracellular
concentrations of Aβ. Extracellular Aβ peptides and fibrils are associated with synaptic
dysfunction, depletion of glutamatergic and cholinergic tone, oxidative stress, altered

3
glucose metabolism and ultimately neuronal death (Bossy-Wetzel, Schwarzenbacher, &
Lipton, 2004; Cuello, 2005; Laursen, Mørk, Plath, Kristiansen, & Bastlund, 2013; Nizzari
et al., 2012; Rossor et al., 2015; Thinakaran & Koo, 2008).
Hyperphosphorylation of tau leads to the formation of intracellular
neurofibrillary tangles. Physiologically, tau proteins bind to tubulin and are crucial to the
stability of microtubules in central nervous system (CNS) axons. In AD tau
serine/threonine residues are phosphorylated by proline and non-proline directed proline
kinases which causes tau to dissociate and destabilizes microtubules (Avila, 2006; Wang,
Xia, Iqbal, 2012). Moreover, phosphorylated tau binds and aggregates with other tau
proteins to form neurofibrillary tangles, neuropil threads, and dystrophic neurites
(Alonso, Iqbal, 1996; Gong, Liu, Iqbal, & Iqbal, 2006; Maccioni, Vera, Dominguez, &
Avila, 1989; Wang et al., 2012). Intracellularly, phosphorylation and aggregation of tau
inhibits axonal trafficking and triggers apoptosis (Alonso et al., 1996; Bancher, Braak,
Fischer, & Jellinger, 1993; Gong et al., 2006; Wang et al., 2012).
Ultimately, AD is a multifactorial disease in which pathology, including amyloid
plaques and neurofibrillary tangles, is associated with extensive neuronal dysfunction, a
decrease in synaptic responsiveness, inflammation, oxidative stress, cell death and brain
atrophy (Braak & Braak, 1991, 1995; Griffin et al., 1998; Jacobsen et al., 2006; Selkoe,
1991). There are several models concerning the causal pathological element in the AD
neurodegenerative cascade.

4
The Cholinergic Hypothesis of Alzheimer’s Disease. The cholinergic hypothesis
suggests that the degeneration of basal forebrain cholinergic neurons and consequent
disruption of cholinergic neurotransmission in the cortex, striatum and hippocampus is
related to Aβ accumulation, hyperphosphorylation of tau and cognitive dysfunction in
AD patients (Bartus, 2000; Bartus, Iii, Beer, & Lippa, 1982; Francis, Palmer, Snape, &
Wilcock, 1999). A critical role of acetylcholine (ACh) in AD is supported by abnormal
cholinergic activity in the late stages of AD and a critical role of ACh in memory.
Specifically, late AD is associated with decreased choline uptake in the hippocampus
(Rylett, Ball, & Colhoun, 1983); decreased choline acetyltransferase in the cortex,
hippocampus and amygdala (Bowen, Smith, White, & Davison, 1976; Davies &
Maloney, 1976; Perry, Gibson, Blessed, Perry, & Tomlinson, 1977); decreased release of
ACh in the cortex (Nilsson, Nordberg, Hardy, Wester, & Winblad, 1986); and loss of
cholinergic neurons in the basal forebrain (Whitehouse, Price, Struble, Clark, 1982). The
cholinergic hypothesis has led to the approval of cholinesterase inhibitors (ChATi) for the
mitigation of cognitive symptoms in moderate AD (Birks, 2006; Cummings, 2003;
Lanctôt et al., 2003; Raschetti et al., 2005). However, in patients with mild-cognitive
impairment and early AD the level and activity of choline acetyltransferase is not
decreased and cholinergic neurons in the basal forebrain are conserved (Davis et al.,
1999; Dekosky et al., 2002; Gilmor et al., 1999). Thus, it is unlikely that deficits in
cholinergic neurotransmission directly cause AD neurodegeneration.
The Tau Hypothesis of Alzheimer’s Disease. The tau hypothesis postulates a
critical role for inflammation and excessive tau hyperphosphorylation in the formation of
neurofibrillary tangles, abnormalities in signaling pathways, neurotoxicity and

5
neurodegeneration in AD patients (Maccioni, Farias, Morales, & Navarrete, 2010).
Indeed, neurofibrillary tangles may be unrelated to or precede Aβ plaques (Braak &
Braak, 1991; Tabaton et al., 1989), hyperphosphorylated tau and neurofibrillary tangles
correlate with impairment in episodic memory in human AD patients (Ghoshal et al.,
2002; Maccioni et al., 2006), and pharmaceutical agents that target tau restore spatial
memory in transgenic mice (Min et al., 2015).
The Amyloid Cascade Hypothesis of Alzheimer’s Disease. Although controversial,
it has been suggested that Aβ accumulation causes hyperphosphorylation of tau in AD
(Cuello, 2005; Gamblin et al., 2003; Hardy, & Higgins, 1992; Hardy, 1997; Hardy &
Allsop, 1991; Selkoe, 1999). Specifically, the amyloid cascade hypothesis states that
abnormal production and cleavage of Aβ is the causative pathological feature of AD that
facilitates a hierarchical sequence of amyloid plaque formation, inflammation, oxidative
stress, tau hyperphosphorlation, neuronal death and dementia (Hardy et al., 1992; Hardy
& Allsop, 1991; Hardy & Selkoe, 2002; Lemere & Masliah, 2010). A causal role for Aβ
in AD was suggested based on the discovery that Aβ is the primary component of
amyloid plaques in patients with AD and Down syndrome, the localization of the APP
gene to chromosome 21 and the high prevalence of AD in individuals with Down
syndrome (Hardy & Allsop, 1991; Hardy & Higgins, 1992; Hardy & Selkoe, 2002). In
further support of the amyloid hypothesis: Aβ is toxic in vitro (Iversen, Mortishire-Smith,
Pollack, & Shearman, 1995); sporadic and familial AD are associated with genetic
mutations that alter Aβ production, processing and clearance (Schellenberg & Montine,
2012); overexpression of human APP in mice may produce amyloid plaques (Terai et al.,

6
2001; Webster, Bachstetter, Nelson, Schmitt, & Van Eldik, 2014), memory deficits
(Chen, Chen, Knox, Inglis, Bernard, Martin, Justice, McConlogue, et al., 2000), and in
some transgenic models tau hyperphosphorylation (Kanno, Tsuchiya, & Nishizaki,
2014b); and amyloid plaques increase the risk of AD (Chen et al., 2014). However, the
amyloid cascade hypothesis has several limitations: many older individuals have
extensive amyloid pathology without cognitive impairment (Villemagne et al., 2011);
anti-amyloid treatments in human patients reduce plaques but do not prevent further
cognitive decline or neurodegeneration (Holmes et al., 2008); transgenic mice with
overexpression of Aβ, but not APP, do not have behavioural deficits (Kim et al., 2013);
neurofibrillary tangles may develop prior to (Schönheit, Zarski, & Ohm, 2004) or
independently of amyloid plaques (Arriagada, Growdon, Hedleywhyte, & Hyman, 1992;
Bouras, Hof, Giannakopoulos, Michel, & Morrison, 1994; Price, Davis, Morris, & White,
1991); and tangles strongly correlate with neurodegeneration and cognitive decline
(Arriagada et al., 1992; Giannakopoulos et al., 2003).
There is also substantial support for a critical role of neuroinflammation
(Cameron & Landreth, 2010; McGeer, Schulzer, & McGeer, 1996; Meraz-Ríos, Toral-
Rios, Franco-Bocanegra, Villeda-Hernández, & Campos-Peña, 2013; Mosher & Wyss-
Coray, 2011), oxidative stress and mitochondrial dysfunction (Lin & Beal, 2006; Schrag
et al., 2013; Swerdlow, Burns, & Khan, 2014; Valla et al., 2006), altered calcium
homeostasis and excitotoxicity (Bezprozvanny & Mattson, 2008; Demuro, Parker, &
Stutzmann, 2010; Green & LaFerla, 2008; Szwagierczak, Bultmann, Schmidt, Spada, &
Leonhardt, 2010), DNA damage (Bucholtz & Demuth, 2013; Lovell & Markesbery,

7
2007; Obulesu & Rao, 2010; Weissman, de Souza-Pinto, Mattson, & Bohr, 2009), and
loss of cell cycle control (Arendt, Brückner, Mosch, & Lösche, 2010; Busser,
Geldmacher, & Herrup, 1998; McShea, Harris, Webster, Wahl, & Smith, 1997; Yang,
Mufson, & Herrup, 2003) in AD. Thus, while there are many alternative theories
regarding the cause of AD dementia, Aβ and tau likely play a significant role in the
neurodegenerative cascade.
Early and extensive AD neuropathology in the medial temporal lobe (MTL).
Although the clinical staging of AD pathology is heterogeneous, histopathological and
imaging studies report pathological changes that begin in the MTL prior to AD diagnosis
and progress to widespread cortical and sub-cortical regions (Braak & Braak, 1991, 1995;
Hyman, Hoesen, Damasio, & Clifford, 1984; Scahill, Schott, Stevens, Rossor, & Fox,
2002; Schönheit et al., 2004; Whitwell et al., 2007). Braak & Braak (1991, 1995)
describe an Aβ and tau deposition pattern that begins in limbic regions, specifically the
hippocampus (HPC), association cortices, basal forebrain, thalamus and hypothalamus
and spreads to neocortex and various subcortical nuclei. It has been suggested that AD
pathology deposits in a non-random fashion following signaling pathways via cell-to-cell
transmission of Aβ and tau ( Hyman et al., 1984; Saper, Wainer, & German, 1987;
Steiner, Angot, & Brundin, 2011). The distribution of brain atrophy is consistent with
patterns of Aβ and tau deposition. Specifically, gray matter atrophy is present in the HPC,
amygdala, entorhinal cortex and fusiform gyrus prior to AD diagnosis, atrophy of the
MTL becomes quite extensive as mild cognitive impairment advances, and at AD

8
diagnosis atrophy extends to frontal and parietal lobes (Scahill et al., 2002; Whitwell et
al., 2007).
Episodic Memory Deficits in Alzheimer’s Disease
The degree of AD neuropathology is correlated with the severity of cognitive
deficits (Bancher et al., 1993; Riley, Snowdon, & Markesbery, 2002). Amyloid plaque
deposition is correlated with cognitive decline in early stages of AD, while
neurofibrillary tangles strongly correlate with cognitive deficits throughout AD (Braak &
Braak, 1991, 1995; Nelson et al, 2013; Nelson, Braak, & Markesbery, 2007). Deficits in
episodic memory, executive function, and perceptual processing may be apparent prior to
AD diagnosis (Bäckman, Jones, Berger, Laukka, & Small, 2004; Perry & Hodges, 1999;
Riley et al., 2002; Snowden et al., 2011).
Episodic memory for life events is a subtype of declarative memory (the
conscious memory for facts and events; Squire & Zola, 1996) and is facet of cognitive
function affected early in AD (Barbeau et al., 2008; Didic et al., 2013; Didic, Ranjeva,
Barbeau, Confort-gouny, & Le, 2010; Libon et al., 1998; Pike et al., 2007; Stoub et al.,
2006). Visual recognition memory is a subclass of episodic memory that is dependent on
the integrity of MTL structures and is commonly evaluated using delayed matching-to-
sample (DMS) and delayed nonmatching-to-sample (DNMS) tasks in rodents, non-
human primates and humans. Generally, in D(N)MS tasks subjects encode a sample
stimulus, and following a retention delay, in a forced-choice test participants must

9
indicate the stimulus that matches the sample stimulus (DMS) or that does not match the
sample stimulus (DNMS) (Huppert & Piercy, 1976).
Functional degeneration of medial temporal lobe structures such as the HPC,
entorhinal cortex, and perirhinal cortex (PRh) is associated with impairments in episodic
memory, including visual recognition memory (Barbeau et al., 2008; Didic et al., 2010).
Tests of visual recognition memory (VRM), including familiarity-based recognition and
delayed matching to sample paradigms, have been used to evaluate episodic memory in
patients with cognitive decline. Barbeau et al. (2008) evaluated VRM in patients with
amnesic mild cognitive impairment (aMCI) to assess the relationship between cortical
gray matter atrophy and memory impairment. VRM was evaluated using a delayed
matching to sample recognition memory (DMS48) paradigm. In DMS48 patients
incidentally learn 48 coloured drawings and, following a 1h long-term retention delay,
are evaluated on their ability to identify a learned drawing amongst distractors. aMCI
patients impaired in DMS48 had gray matter loss in the MTL and temporal-parietal
regions, including the PRh (Barbeau et al., 2008). Didic et al. (2010) evaluated VRM in
patients with amnesic mild cognitive impairment to determine if VRM deficits, using
DMS48, are associated with metabolic abnormalities in the MTL. Patients with impaired
VRM had decreased bilateral MTL metabolism, including regions of the HPC, and VRM
deficit correlated with MTL metabolism (Didic et al., 2010). Similarly, Wolk et al. (2008,
2011) evaluated recognition memory for associative (word-pairs) or featural information
(font colour) and found aMCI patients to be impaired on both tests of familiarity and
recognition memory in which the HPC and PRh play a dissociable role (such that the

10
HPC is required for recollection and the PRh is required for familiarity). Importantly,
familiarity recognition is spared in healthy aging (Davidson & Glisky, 2002; Yonelinas et
al., 2002); thus loss of familiarity recognition may be a marker for pathogenic aging.
Indeed, performance on visual recognition memory tests in aMCI patients is predictive of
AD diagnosis (Didic et al., 2013, 2010; Wolk, Signoff, & DeKosky, 2008).
In order to complement and extend the understanding of episodic memory deficits
in AD patients it may be advantageous to model AD and tests of visual recognition
memory in experimental animals in order to characterize cognitive deficits, identify new
therapeutic targets and evaluate the therapeutic potential of pharmaceutical agents.
Modeling Alzheimer’s Disease in Rodents
Most rodents do not develop Alzheimer’s pathology with age. Popular rodent
models of AD, however, recapitulate key features of amyloid and tau pathology via
genetic manipulation or injection of Aβ into the CNS (Ashe & Zahs, 2010; Bilkei-Gorzo,
2014; LaFerla & Green, 2012; Lecanu & Papadopoulos, 2013; Van Dam & De Deyn,
2011; Webster et al., 2014). Accumulation of Aβ and tau hyperphosphorylation is
followed by molecular and cellular cascades that contribute to neurodegeneration and
behavioural decline. In addition to behavioural profiling, rodent models of AD are
valuable for the characterization of oxidative stress, inflammation, abnormal
mitochondrial function, immune responses and other molecular abnormalities that may
identify novel substrates and increase applicability of findings to therapeutic drug
discovery.

11
Notably, no rodent model perfectly recapitulates neuropathological staging and
cognitive decline seen in the human disease. Many transgenic strains model the less
common familial form of AD via overexpression of APP and/or PS1 genes. 5xFAD and
3xTG are distinct and complementary transgenic mouse models of familial AD, and both
will be evaluated in the current study (see Table 1).
5xFAD. The 5xFAD transgenic mice were developed with three mutations in the
APP gene and two mutations in the PS1 gene. 5xFAD transgenic mice have a more
aggressive and earlier onset of amyloid pathology compared to other transgenic strains.
Specifically, Aβ42 is almost exclusively produced and accumulates intracellularly and
extracellularly in young mice at 1.5 and 2 months of age in the HPC and deep cortex,
respectively (Oakley et al., 2006). Amyloidosis is followed by loss of cholinergic and
noradrenergic neurons and tau hyperphosphorylation in old mice at approximately 12
months of age (Devi & Ohno, 2010; Kalinin et al., 2012; Kanno et al., 2014). While the
very early onset may compromise the translational validity of the 5xFAD strain, the
reduction in animal care expense and the severe amyloid pathology has made this model
popular. Behavioural deficits in spatial memory in spontaneous Y-maze alteration present
at 4 months of age (Oakley et al., 2006), followed by impaired contextual fear memory
and spatial learning in the Morris water maze by 6 months of age (Kimura & Ohno, 2009;
Ohno et al., 2006).
3xTG. The 3xTG model has high face validity because both the human amyloid
plaque and neurofibrillary tangle pathologies are recapitulated. In 3xTG mice, APP and

12
PS1 are overexpressed to induce amyloid pathology, and the MAPT gene is mutated to
induce tauopathy (Oddo, Caccamo, Shepherd, et al., 2003). Intracellular Aβ accumulation
has been observed at 3 to 4-months-of-age, and extracellular amyloid plaques develop at
approximately 6-months-of-age (Oddo, Caccamo, Shepherd, et al., 2003).
Hyperphosphorylation of tau is observed in the HPC at 6-months-of-age, followed by
hyperphosphorylation of tau in cortical regions and eventually, unlike the 5xFAD model,
the formation of neurofibrillary tangles (Oddo, Caccamo, Shepherd, et al., 2003; Rohn et
al., 2008). With age there is cholinergic and noradrenergic neuronal death, increased
microglia activity, inflammation, and alterations in glucose metabolism (Da Cruz et al.,
2012; Manaye et al., 2013; Mastrangelo & Bowers, 2008; Nicholson et al., 2010; Oddo,
Caccamo, Shepherd, et al., 2003; Sy et al., 2011). However, unlike the human disease,
HPC neurodegeneration is not reported and amyloidosis begins in the cortex (Manaye et
al., 2013; Oddo, Caccamo, Shepherd, et al., 2003). Young 3xTG mice have deficits on
the what-where-which episodic object memory task as early as 3 months of age (Davis,
Easton, Eacott, & Gigg, 2013), followed by impaired long-term Morris water maze and
contextual fear retention at 6 months of age (Billings, Oddo, Green, McGaugh, &
LaFerla, 2005).
Rodent AD models have been vital to enhancing our understanding of AD.
Assessing transgenic models that embody complementary aspects of the human disease –
one of the primary goals of this thesis - may elucidate behavioural deficits associated
with specific elements of AD pathology.

13
Modeling Recognition Memory in Rodents
A rodent variation of the DNMS task is spontaneous object recognition (SOR;
Ennaceur & Delacour, 1988). The SOR task does not require extensive training or
reward; therefore memory can be evaluated in a manner similar to daily human
interaction with objects (Dere, Huston, & De Souza Silva, 2007; Ennaceur & Delacour,
1988). Tests of spontaneous object recognition exploit rodents’ innate preference to
approach and explore novel stimuli (Winters, Saksida, & Bussey, 2008). The
neurobiological mechanisms required for object recognition are contingent on the specific
nature of the task. In the most common form of SOR, rodents are presented with two
identical objects during a sample phase and, following a retention delay, during a choice
phase rodents are presented with an object from the sample phase (familiar) and a new
stimulus (novel). The novelty preference is manifested by greater exploration of novel
stimuli or spatial locations compared to familiar stimuli (Ennaceur & Delacour, 1988;
Winters et al., 2008).
When evaluated in this fashion SOR is reliant on functional integrity of
cholinergic, glutamatergic and serotonergic signaling in structures including the PRh,
HPC and the medial prefrontal cortex (mPFC; (Barker & Warburton, 2011; Dere et al.,
2007; Winters et al., 2008).
There are many ways in which objects may be processed that have relevance to
AD. Interaction with objects may engage memory for specific object features, spatial
memory processing, temporal memory processing, and multisensory and perceptual

14
processing. By modifying the nature of the SOR paradigm it is possible to tax different
forms of object processing that have not entirely overlapping neural mechanisms.
Spontaneous object recognition and feature processing. The SOR paradigm has
been predominately performed using an open-arena and is used to assess rodents’ ability
to distinguish between a novel and familiar object. When tested in an open-field
apparatus, spatial and contextual information from within the apparatus and around the
testing room could potentially influence memory encoding and performance in the SOR
task. Previously, there has been debate concerning the contributions of the PRh and HPC
to object recognition memory. More recent experimental evidence from rats suggests the
PRh is necessary for SOR, while the HPC is not necessary for object recognition per se
(Barker & Warburton, 2011; Forwood, Winters, & Bussey, 2005; Winters, Forwood,
Cowell, Saksida, & Bussey, 2004; Winters et al., 2008), but can be involved depending
on the nature of encoding conditions. Specifically, while damage to the HPC has
occasionally been shown to impair SOR in rats in the open field (Clark, Zola, & Squire,
2000; Mumby, Gaskin, Glenn, Schramek, & Lehmann, 2002), Winters et al. (2004)
established a functional double dissociation between the necessity of the PRh and HPC
for object and spatial memory. Bilateral excitotoxic lesions of the PRh or HPC in male
rats impaired SOR and spatial memory in the 8 arm radial maze, respectively. Critically,
as it is difficult to determine what cues rodents use to perform the SOR task, object
recognition was tested in a Y-shaped apparatus with high opaque walls in an effort to
minimize influence of spatial and contextual information. Forwood et al. (2005) provided
further support for dissociable roles of the PRh and HPC in SOR such that rats with

15
bilateral excitotoxic lesions of the HPC were impaired on a spatial non-matching to
sample task but not SOR in the Y-apparatus. Similarly, Barker & Warburton (2011)
evaluated recognition memory using a variety of object paradigms for rats that required
feature or object identity information, spatial information, and temporal information
processing. Results suggested that the HPC is required for tests of object processing only
when spatial and/or temporal object information is involved. Thus, it has been suggested
that the PRh is necessary for memory of object identity, while HPC is not involved in
object recognition but rather is important for recognition memory for spatial and temporal
information. To my knowledge a similar double dissociation between the function of the
PRh and HPC in object and spatial memory has not been demonstrated in mice. It appears
that mice process objects differently, as the performance of rats and mice may
systematically differ in learning and memory paradigms (Cohen et al., 2013; Dere et al.,
2007), and inactivation of the HPC has been shown to impair SOR in mice (DeVito &
Eichenbaum, 2010; Hammond, Tull, & Stackman, 2004); it is important to note,
however, that findings from this study may also be confounded by spatial information
including object configuration and the open-field testing apparatus. Yet, the HPC does
appear to be necessary for object identity processing in mice, as SOR is intact in control
mice but impaired following inactivation of the HPC, by muscimol, when spatial and
contextual information was made irrelevant by presenting objects in unique contexts
(Cohen et al., 2013). SOR performance is also known to rely on the functional integrity
of the PRh in mice (Romberg et al., 2013), consistent with rat, monkey, and human
findings (Meunier, Bachevalier, Mishkin, & Murray, 1993; Murray & Mishkin, 1998;
Murray & Bussey, 1999; Winters et al., 2004; Winters, Saksida, & Bussey, 2010; Zola-

16
Morgan, Squire, Amaral, & Suzuki, 1989). Thus, while the open field version of SOR
may be more likely to recruit the HPC in mice, both open field and Y-apparatus SOR
evaluate forms of object memory considered to model facets of human declarative
memory, and both rely on MTL brain regions that are highly relevant to AD.
Spontaneous object recognition and spatial processing. Spatial memory may be
evaluated using the spontaneous object location (OL) task. The SOR paradigm can be
slightly modified to assess OL such that during the choice phase rodents are presented
with two copies of the sample object but one of which is placed in a novel location (Dix
& Aggleton, 1999; Murai, Okuda, Tanaka, & Ohta, 2007), to selectively evaluate
spatial/contextual object associations. OL is thought to be dependent on the HPC.
Specifically, lesion and inactivation of the HPC in rats and mice has been shown to
impair OL (Assini, Duzzioni, & Takahashi, 2009; Barker & Warburton, 2011; Mumby &
Pinel, 1994), while the PRh is not necessary for OL performance (Barker & Warburton,
2011). Additionally, lesion of the fornix and cingulate cortex have also been shown to
impair OL in rats (Ennaceur, Neave, & Aggleton, 1997).
Spontaneous object recognition and temporal processing. Temporal processing
may be evaluated using the temporal order (TO) task. In TO rodents are exposed to two
distinct sets of identical stimuli in two different sample phases and following a delay are
tested on their ability to differentiate between the more remote and more recently
presented stimuli. As rodents have an innate preference for novelty, if temporal
processing is intact rodents will preferentially explore the stimuli that were presented less

17
recently (more novel). Evidence from lesion studies in rats suggests a critical role of the
PRh, HPC, mPFC and functional connectivity between the PRh and mPFC in TO
memory tasks (Barker, Bird, Alexander, & Warburton, 2007; Barker & Warburton, 2011;
Hannesson, Howland, & Phillips, 2004; Mitchell & Laiacona, 1998).
Spontaneous object recognition and multisensory integration. The object
recognition paradigm may also be adapted for the evaluation of multisensory integration.
For example, the cross-modal object recognition task (CMOR) may be used to evaluate
tactile-to-visual multisensory integration where rodents use a previously formed tactile
object representation to distinguish between novel and familiar visual stimuli (Winters &
Reid, 2010). In CMOR the sample phase is run under red light, restricting visual
exploration, such that only tactile object information is available. Following a retention
delay, the choice phase is run in white light with objects placed behind transparent
barriers, such that only visual information is available. The CMOR task has been shown
to be dependent on functional connectivity of the PRh, posterior parietal cortex (PPC),
and regions of the PFC, including the orbitofrontal cortex (OFC; Cloke, Jacklin, &
Winters, 2014; Reid, Jacklin, & Winters, 2012, 2014; Winters & Reid, 2010).
More recently, our lab has developed a multisensory object oddity task (MSO),
that is also likely dependent on integrity of the PRh, PPC, and OFC network for
multisensory binding (Cloke et al., 2014; Reid et al., 2012, 2014). Standard object oddity
tasks are advantageous for the evaluation of visual perception and are similarly dependent
on the functional integrity of the PRh (Bartko, Winters, Cowell, Saksida, & Bussey,

18
2007; Forwood, Bartko, Saksida, & Bussey, 2007). Object oddity paradigms are often run
without a retention delay; therefore the task has minimal mnemonic demand and provides
insight into the basic perceptual ability of rodents. The MSO task can be used to assess
multimodal perception under tactile-visual or olfactory-visual conditions. In this task
rodents are simultaneously presented with pairs of objects with shared combinations of
tactile-visual (or olfactory-visual) features and one dissimilar (‘odd’) object comprising a
unique combination of features. If perception is intact, rodents will preferentially explore
the odd object.
Recognition Memory Deficits in Rodent Models of Alzheimer’s Disease
Tests of object processing are advantageous to the study of AD because
performance is reliant on signaling pathways and brain regions affected by Alzheimer’s
pathology in humans. An extensive literature suggests that rodent models of AD are
impaired on object recognition (OR) when evaluated in an open-field apparatus. Deficits
in object recognition have been observed in several transgenic AD mice: APP (Ambrée et
al., 2009; Arrieta-Cruz, Pavlides, & Pasinetti, 2010; Balducci et al., 2011; Bardgett,
Davis, Schultheis, & Griffith, 2011; Beggiato et al., 2014; Cho et al., 2011; Dewachter et
al., 2002; Dobarro, Gerenu, & Ramírez, 2013; Dodart et al., 1999; Escribano et al., 2009;
Francis et al., 2012; Fukumoto et al., 2014; Galeano et al., 2014; Görtz et al., 2008; Greco
et al., 2010; Hernandez, Kayed, Zheng, Sweatt, & Dineley, 2010; Hillen et al., 2010; Jin
et al., 2014; Kauppinen et al., 2011; Knowles et al., 2013; Martín-Moreno et al., 2012;
Mouri et al., 2007; Nishida et al., 2006; Ohta, Arai, Akita, Ohta, & Fukuda, 2012; Polito
et al., 2014; Ribes, Torrente, Vicens, Colomina, & Domingo, 2012; Richter et al., 2008;

19
Saydoff et al., 2013; Simón et al., 2009; Sivilia et al., 2013; Verret et al., 2013; L. Zhang
et al., 2006, 2014); PS1/PS2 models (Huang, 2003; P. Wu et al., 2008; Zufferey et al.,
2013); APP/PS1 (Barbero-Camps, Fernández, Martínez, Fernández-Checa, & Colell,
2013; Donkin et al., 2010; Frye & Walf, 2008; Hafez et al., 2012; Howlett et al., 2004;
McClean & Hölscher, 2014; McClean, Parthsarathy, Faivre, & Hölscher, 2011; Mori,
Koyama, Guillot-Sestier, Tan, & Town, 2013; Scholtzova et al., 2008; Roach et al., 2004;
Vargas, Fuenzalida, & Inestrosa, 2014; Webster, Bachstetter, & Van Eldik, 2013; Woo et
al., 2010; Yao et al., 2013; Zhang et al., 2014; Zhang et al., 2013; Zhang et al., 2012);
5xFAD (Giannoni et al., 2013; Joyashiki, Matsuya, & Tohda, 2011; Seo et al., 2014;
Tohda, Nakada, Urano, Okonogi, & Kuboyama, 2011; Tohda, Urano, Umezaki, Nemere,
& Kuboyama, 2012; Wang et al., 2013) and 3xTG (Arsenault, Julien, Tremblay, &
Calon, 2011; Blanchard et al., 2010; Chen et al., 2014; Feld et al., 2014; Filali et al.,
2012; Guzmán-Ramos et al., 2012; Kazim et al., 2014; Onishi et al., 2011; St-Amour et
al., 2014).
However, the specific nature of the OR deficit in AD mice is controversial
(Grayson et al., 2014; Simón et al., 2009; Taglialatela, Hogan, Zhang, & Dineley, 2009),
and some studies have failed to report OR impairments (Cheng, Low, Logge, Garner, &
Karl, 2014; Davis, Eacott, Easton, & Gigg, 2013; Davis, Easton, et al., 2013; Fragkouli,
Tsilibary, & Tzinia, 2014; Good, Hale, & Staal, 2007; Gulinello et al., 2009; Hale &
Good, 2005; Karl, Bhatia, Cheng, Kim, & Garner, 2012; Yassine et al., 2013).
Differential performance on OR tasks may be related to procedural differences that alter
the conceptual nature of the task and thus tax distinct behavioural processes.

20
Discrepancies in the mnemonic index, the length of the retention delay, the age of
behavioural testing and/or the choice of sex and transgenic model may also affect OR
data. And in several cases, due to the nature of the OR paradigm, object recognition
results cannot be interpreted with confidence, as control mice were not able to perform
the task (Maroof, Ravipati, Pardon, Barrett, & Kendall, 2014; Scullion, Kendall,
Marsden, Sunter, & Pardon, 2011; Stover, Campbell, Van Winssen, & Brown, 2015).
According to previous reports transgenic AD mice appear to have intact short-term OR
memory, as a novelty preference has been observed with 2-3min retention delays (Davis,
Easton, et al., 2013; Good & Hale, 2007; Middei, Daniele, Caprioli, Ghirardi, &
Ammassari-Teule, 2006; Taglialatela et al., 2009). Yet, there is evidence that OR is
impaired in transgenic AD mice when the retention delay is ≥ 5min (Ambrée et al., 2009;
Bardgett et al., 2011; Dewachter et al., 2002; Dodart et al., 1999; Gerenu, Dobarro,
Ramirez, & Gil-Bea, 2013; Giuliani et al., 2013; Greco et al., 2010; Hillen et al., 2010;
Spilman et al., 2014; Yuede et al., 2009). Further, OR performance is age-dependent,
such that OR deficits have been described in transgenic mice 3 to 15-months-of-age.
Therefore, additional research is necessary to clarify when OR is impaired in various AD
transgenic mouse models. Few studies have systematically examined OR with varied
retention delays in multiple transgenic AD models. The understanding of object
processing in AD is far from comprehensive. In addition to the familiarity of object
identity there are many other types of object processing that are altered by AD
neuropathology in human patients. Indeed, AD patients also have deficits in spatial
processing (Caterini, Sala, Spinnler, Stangalino, & Turnbull, 2002; Snowden et al.,
2011), temporal processing (Becker, Wess, Hunkin, & Parkin, 1993), sensory integration

21
(Drzezga et al., 2005; Wu et al., 2012), and spatial, but not featural, perception (Lee et
al., 2006). Accordingly, a few studies have described deficits in object spatial processing
in AD transgenic mice (Bergin & Liu, 2010; Bollen et al., 2013; Dodel et al., 2011; Frye
& Walf, 2008; Gulinello et al., 2009). Further, despite impairment in human AD patients
and the necessity of the HPC in temporal tasks, Hale & Good (2005) and Davis et al.
(2013) have failed to demonstrate temporal order memory impairments in transgenic
APP, Tg2576, and 3xTG mice. However, the retention delay in temporal tasks was not
sufficiently long to induce deficits in object recognition (2 min). Extending the retention
delay may induce a temporal order memory deficit in transgenic AD mice. Therefore, it is
insufficient to conclude that AD transgenic mice are impaired on “object recognition”
without a full characterization of behavioural deficits in object processing; there are
numerous aspects of object processing related to recognition and AD that remain to be
thoroughly analyzed.
The purpose of the current study was to perform a systematic analysis of object
identity, spatial, temporal, multisensory, and perceptual processing in 5xFAD and 3xTG
mouse models of AD. It is hypothesized that transgenic AD mice will have abnormal
performance on object processing paradigms at 12 months of age. By manipulating the
nature and retention delay of the object recognition paradigm we were able to alter
mnemonic demand and tax different facets of object processing with relevance to AD to
clarify the nature of object information processing deficits.

22
Methods
Animals
Male and female wild-type 5xFAD wild-type (B6SJLF1/J) and transgenic
(B6SJLTg(APPSwFlLon,PSEN1*M146L*L286V)6799Vas/Mmjax) and 3xTG wild-type
(B6129SF2/J) and transgenic (B6;129 Psen1tm1Mpm Tg(APPSwe,tauP301L)1Lfa/Mmjax)
mice were obtained from the Jackson Laboratory (Bar Harbor, ME, USA) at
approximately 2-months-of-age.
5xFAD transgenic mice have five mutations that overexpress familial AD genes:
human APP(695) with the Swedish (K670N, M671L), Florida (I716V), and London
(V717I) as well as two human PS1 mutations (M146L and L286V), inserted into exon 2
of the Thy gene and regulated by the Thy1 promoter to drive overexpression in the brain.
5xFAD mice have an extremely early accumulation of Aβ42 that advances to amyloid
plaque formation, and synaptic and neuronal loss (Oakley et al., 2006).
3xTG mice have familial AD transgene knockin of APPSwe and tauP301L that
recapitulate amyloid and tau pathology in the central nervous system. 3xTG mice were
generated by co-injection of APPSwe and tauP301L, under the control of Thy1.2
promoter, into presenilin-1 (M146V) knock-in mice embryos. The mutant transgenes
facilitate beta-amyloid deposition, amyloid plaque formation, hyperphosphorylation of
tau and eventually the formation of neurofibrillary tangles (Oddo, Caccamo, Shepherd, et
al., 2003).

23
To investigate object processing in transgenic AD mouse models we used 3xTG
transgenic (n = 21male, n = 35 female) and wild-type (n = 24 male, n = 35 female) and
5xFAD transgenic (n = 23 male, n = 33 female) and wild-type mice (n = 20 male, n = 33
female). Mice used in this study were previously tested in touchscreen-based operant
tasks, but were not exposed to objects. Sample sizes were selected to compensate for the
attrition rate. Mice were group housed in clear polyethylene cages (16 12 26 cm), with
corncob bedding, crink-l’Nest and cotton nest squares in a controlled environment (22
±2°C) on a 12h light/dark cycle (0800h lights on; 2000h lights off). Prior to the onset of
behavioural testing, food (Teklad Global 16% Protein Rodent Maintenance Diet, Harlan
Teklad, WI) and water were available ad libitum.
Behavioural testing commenced when mice were approximately 12-months-of-
age (when AD pathology is sufficiently developed). All behavioural testing was
conducted during the light phase of the light/dark cycle. All procedures adhered to the
guidelines of the Canadian Council on Animal Care and were approved by the Animal
Care Committee at the University of Guelph.
Behavioural Testing
Prior to behavioural testing all mice were extensively handled and habituated to
an empty testing apparatus for 10min on two consecutive days.

24
Apparatus, Objects & General Procedure. In all object processing paradigms, a
JVC everio camcorder was placed above the testing apparatus for recording and
subsequent analysis of behavioural data using ORscore. An unused set of objects was
used for each trial in a given cohort of mice. Objects were distinct, approximately 5-15cm
tall and made of glass, metal, or plastic. Objects had no apparent biological significance
to mice and were equally preferred within each pair. All objects were fixed to the floor of
the testing apparatus with white adhesive putty. Immediately prior to behavioural testing
mice were brought into the testing room in their home cage. To prevent ‘pre-exposure’ to
objects before the start of the testing phase, mice were placed into a bottomless cardboard
box (start box) located in the middle of the open-field or in the start arm of the Y-
apparatus. The trial began when the start box was opened or removed and the mouse
exited the start box/arm. Between behavioural trials objects were wiped with 50% ethanol
(to eliminate olfactory cues) and the testing apparatus was only wiped with a dry paper
towel.
Cohorts of mice were tested on several object processing paradigms. The first
cohorts of 5xFAD and 3xTG male and female were tested on open-field OR then Y-
apparatus OR (experiment 1). The second cohorts of 5xFAD and 3xTG male and female
mice were tested on object oddity (experiment 4) then object location (experiment 2). The
third cohort of 5xFAD and 3xTG females were tested on temporal order (experiment 3).

25
Experiment 1
Open-Field Object Recognition. The open-field SOR task (Figure 1a) assesses the
ability of rodents to distinguish a previously explored object from a novel object. Open-
field OR was conducted in a square open arena (45 45 30 cm) made entirely of white
corrugated plastic. When object recognition is evaluated in the open-field, mice are able
to see many contextual cues in the testing room; thus spatial information is readily
available. The object recognition task is comprised of a sample (learning) phase and
choice (test) phase. In the sample phase mice were presented with two identical objects in
the top corners of the arena 5cm from the wall for 10min. A retention delay of either
5min or 3h was used to alter the mnemonic demand of the task. Specifically, a 5min
delay was used to evaluate short-term memory and 3h delay to evaluate long-term
memory; all mice were tested with each delay on separate counterbalanced trials at least
48h apart. Following the retention delay, in the 2min choice phase mice were presented
with one object from the sample phase and a novel object. The location of the novel
object was counterbalanced between and within mice. If memory is intact, mice
preferentially explore the novel object.
Y-Apparatus Object Recognition. SOR was also evaluated in a Y-apparatus
(Figure 1b), which has walls 30.5cm high, and arms 15cm long and 7cm wide
constructed from white Plexiglas. The start arm of the Y-apparatus has a guillotine door
11cm from the back of the arm. When SOR is conducted in the Y-apparatus, spatial
information is minimized, allowing for systematic evaluation of object identity
processing (Winters et al., 2004, 2008). In the sample phase (10min) mice were presented

26
with two identical objects in the end of the arms. Following a retention delay (see above)
in the choice phase mice were presented with an object from the sample phase and a
novel object. The location of the novel object was counterbalanced between and within
mice. If memory is intact, mice preferentially explore the novel object.
Experiment 2
Object Location (OL). The OL task (Figure 1c) evaluates rodents’ ability to
distinguish between familiar and novel spatial locations. The OL task was conducted in
the same open-field arena used for open-filed object recognition testing (45 45 30 cm).
In the sample phase mice were presented with two identical objects for 10min. The
objects were placed in the top corners of the arena 5cm away from the walls. Following a
retention delay (see above), in the 2min choice phase, the identity of the two objects
remained unchanged, but one object was moved to an adjacent corner of the arena. The
relocated object was counterbalanced between and within mice. If memory is intact, mice
preferentially explore the object in the novel spatial location.
Experiment 3
Temporal Order. The temporal order task (Figure 1d) assesses rodents ability to
recognize the relative recency of object presentation (Gareth R I Barker & Warburton,
2011), or how recently an object has been presented. This task was conducted in the same
open-field arena (45 45 30 cm) used for OR and OL tasks. There are two sample phases
in this task (10min each). In each sample phase, mice were presented with two identical
objects in the top corners of the arena 5cm away from the walls; distinct objects were

27
used in sample phase 1 and 2. Following a retention delay (3min), in the choice phase
(2min) mice were presented with one object from each sample phase; thus both objects
were equally familiar, but one was more recently presented. The location of the more
remotely presented object in the choice phase was counterbalanced between and within
mice. If memory is intact, mice preferentially explore the object presented less recently
(i.e. the object from sample phase 1).
Experiment 4
Multisensory Oddity (MSO). Tactile-visual object oddity (Figure 1e) evaluates
multisensory perceptual integration in mice. The oddity task was conducted in a modified
trapezoid-like open-field (front wall 39cm, side wall 14cm, angled side wall 10cm, back
wall 28cm) such that the mice were restricted to a smaller section of the arena. In the
oddity task, mice were presented two pairs of objects for 10min that share the same
combinations of tactile and visual features (e.g., AB/AB and CD/CD), as well as a fifth
object that comprises a unique (i.e., ‘odd’) combination of those features (e.g., AD). The
location of the odd object was counterbalanced. Tactile object features were manipulated
using varieties of sandpaper with different grades, while visual object features were
manipulated using 2-D stickers with distinctive visual markings. Unimodal control trials
were also performed where only two tactile object features or two visual object features
were available; critically, for these control trials, the associative nature of the task
remained the same, but the stimulus dimensions being manipulated were not
multisensory. The location of the odd object was counterbalanced between mice. If
perception is intact, rodents explore the object with the same configurations of tactile and

28
visual features equally (AB/AB and CD/CD), resulting in a preference for the dissimilar
object (e.g. AD).
Behavioural Data Analysis
Exploration was defined as active sniffing within ~1cm of the object and/or
touching the object with the nose. ORscore was used to quantify object exploration
(seconds). Specifically, the experimenter viewed the mouse on a television screen and
pressed a key corresponding to a given stimulus type at the onset of an exploratory bout
and again at the end of a bout. The total 10min of sample and 2min of choice exploration
were quantified.
For all object memory tasks, the novelty preference was quantified by calculating
a discrimination ratio (DR) [(novel object exploration – familiar object exploration)/
(total object exploration)]. In the sample phase, as all objects should be equally novel, a
discrimination ratio of zero is expected. In the choice phase, a discrimination ratio
significantly greater than zero is indicative of intact memory. In the object oddity tasks,
the primary index of performance was quantified by calculating a preference for the odd
object (exploration of the odd object/ total object exploration). Mice that spent less than
3% of the choice duration (3.6s) and choice DR outliers (>2 SD mean) were excluded
from analysis.
Repeated measures ANOVAs were used, where appropriate, to analyze
discrimination ratios and total exploratory behaviour (at sample and choice), with
retention delay as a within-subjects factor and sex and genotype as between-subjects

29
factors. Independent samples t-tests, with the Bonferroni correction, were used to analyze
between group differences in the sample and choice phases. A significant increase in the
discrimination ratio from sample to choice was taken as indicative of intact memory and
was assessed using paired-samples t-tests. For multisensory object oddity a one-way
ANOVA with genotype as the between subjects factor was conducted on the oddity score
and total object exploration. Pearson product moment correlation coefficients were used
to examine the relationship between total sample exploration and novelty preference
index. All statistical analyses were conducted with a significance level of α = .05, unless
otherwise specified, using IBM SPSS statistics. Only significant effects on total
exploration are reported.
Results
Experiment 1: Open-Field & Y-Apparatus Object Recognition
1.1 5xFAD Open-Field Object Recognition
Recognition Memory during the Choice Phase. A three-way repeated measures
ANOVA demonstrated significant main effects of genotype F(1, 32) = 6.392, p = .017,
and delay F(1, 32) = 22.725, p < .001, as well as a significant genotype by delay
interaction F(1, 32) = 7.791, p = .009.
WT and TG 5xFAD females DR’s were not significantly different at 5min
t(13.651) = 1.601, p = .132, but were significantly different at 3h as indicated by a
significant independent samples t-test t(15) = 5.24, p < .001. Paired samples t-tests
between sample and choice DR indicated intact memory in WT females at 5min t(7) =

30
4.657, p = .002 and 3h t(7) = 3.928, p = .006, but impaired memory in TG females at
5min t(10) = .668, p = .519 and 3h t(8) = .335, p = .746. Independent samples t-tests
demonstrated WT and TG 5xFAD male DR’s did not significantly differ at 5min t(19) =
.677, p = .507, but were significantly different at 3h t(18) = 4.324, p < .001. Paired
samples t-tests between sample and choice DR indicated intact memory in WT males at
5min t(10) = 6.219, p < .001 and 3h t(9) = 4.346, p = .002, but impaired memory in
TG males at 5min t(9) = 2.070, p = .068 and 3h t(8) = .415, p = .689 (Figure 2).
Exploratory Behaviour during the Sample and Choice Phase. There was a
significant main effect of genotype F(1, 36) = 17.764, p < .001, as 5xFAD mice generally
explored less than WT in the sample phase. There was a significant main effect of
genotype for choice phase exploration F(1, 36) = 14.606, p = .001, again as TG mice
explored less overall than WT. There was a significant main effect of delay on
exploration during the choice phase F(1, 36) = 23.855, p < .001, such that mice generally
explored less at 3h than at 5min (see total exploration figures for general exploratory data
and specific comparisons for all experiments; Figure 3).
1.2 5xFAD Y-Apparatus Object Recognition
Recognition Memory during the Choice Phase. A three-way repeated measures
ANOVA demonstrated significant main effects of genotype F(1, 36) = 19.198, p < .001,
and delay F(1, 36) = 9.560, p = .004, as well as a significant genotype by delay
interaction F(1, 36) = 8.563, p = .006.

31
Independent samples t-tests demonstrated that WT and TG female DR’s
significantly differed at 5min t(16) = 3.289, p = .005 and were trending at 3h t(17) =
2.071, p = .054. Paired samples t-tests between sample and choice DR indicated
significantly intact memory in WT females at 5min t(7) = 6.195, p < .001, trending 3h
t(7) = 2.244, p = .060, but impaired memory in TG females at 5min t(10) = .407, p =
.692 and 3h t(10) = .870, p = .405. Independent samples t-tests demonstrated that WT and
TG male DR’s did not significantly differ at 5min t(19) = .838, p = .413, but were
significantly different at 3h t(19) = 3.556, p = .002. Paired samples t-tests between
sample and choice DR indicated intact memory in WT males at 5min t(10) = 3.253, p =
.009 and 3h t(10) = 2.787, p = .019, tending in TG males at 5min t(9) = 2.062, p = .069
and significantly impaired at 3h t(9) = 1.009, p = .339 (Figure 4).
Exploratory Behaviour during the Sample and Choice Phase. There was a
significant genotype by sex interaction on exploration during the choice phase F(1, 35) =
4.127, p = .050, but all other comparisons were non-significant (Figure 5).
Correlation Between Exploratory Behaviour during the Sample Phase and
Discrimination Ratio in the Choice Phase. There was a significant positive correlation
between total sample exploration and choice DR in TG females at 5min r(11) = .653, p =
.029 (see Table 3 for specific correlations between sample exploration and choice DR for
all experiments).

32
1.3 3xTG Open-field Object Recognition
Recognition Memory during the Choice Phase. A three-way repeated measures
ANOVA demonstrated significant main effects of genotype F(1, 36) = 41.616, p < .001,
and delay F(1, 36) = 4.839, p = .034.
WT and TG female DR’s significantly differed at 5min t(20) = 2.220, p = .038
and 3h t(20) = 3.489, p = .002 as indicated by significant independent samples t-tests.
Paired samples t-tests between sample and choice DR indicated intact memory in WT
females at 5min t(11) = 7.094, p < .001, 3h t(11) = 4.835, p - .001 and TG females at
5min t(9) = 4.096, p .003, but impaired memory at 3h t(9) = 1.230, p = .250.
Independent samples t-tests demonstrated WT and TG male DR’s significantly differed at
5min t(17) = 3.918, p = .001 and 3h t(16) = 3.000, p = .008. Paired samples t-tests
between sample and choice DR indicated intact memory in WT males at 5min t(10) =
7.619, p < .001 and 3h t(10)= 3.369, p = .007, and impaired memory in TG males at
5min t(6) = .413, p = .694 and 3h t(6) = .708, p = 505 (Figure 2).
Exploratory Behaviour During the Sample and Choice Phase. There were no
significant effects on total exploration during the sample phase. There was a significant
main effect of genotype F(1, 36) = 6.023, p = .019 on total exploration during the choice
phase, as TG explored less than WT. There was a significant main effect of delay on
exploration during the choice phase F(1, 36) = 6.350, p = .016, such that at 3h mice
generally explored less than at 5 min (Figure 3).

33
1.4 3xTG Y-Apparatus Object Recognition
Recognition Memory during the Choice Phase. A repeated measures ANOVA
demonstrated significant main effects of genotype F(1, 36) = 26.260, p < .001, and delay
F(1, 36) = 19.172, p < .001, as well as a significant genotype by delay interaction F(1,
36) = 21.012, p < .001.
Independent samples t-tests demonstrated that WT and TG female DR’s did not
differ at 5min t(20) = .334, p = .742, but were significantly different at 3h t(20) = 6.276,
p < .001. Paired samples t-tests between sample and choice DR indicated intact memory
in WT females at 5min t(11) = 4.013, p = .002, 3h t(11) = 7.740, p < .001 and TG
females at 5min t(9) = 8.264, p < .001, but impaired memory at 3h t(9) = .963, p = .361.
Independent samples t-tests demonstrated that WT and TG male DR’s did not differ at
5min t(16) = .067, p = .947, but were significantly different at 3h t(16) = 4.673, p < .001.
Paired samples t-tests between sample and choice DR indicated intact memory in WT
males at 5min t(10) = 4.453, p = .001, 3h t(10) = 6.851, p < .001 and TG males at 5min
t(6) = 4.321, p = .005, but impaired at 3h t(6) = .135, p = 897 (Figure 4).
Exploratory Behaviour During the Sample and Choice Phase. There was a
significant delay by sex by genotype interaction F(1, 35) = 4.184, p = .048, on
exploration during the sample phase. During the choice phase there was a significant
main effect of sex on exploration F(1, 36) = 4.732, p = .036, as female mice generally
explored less than males (Figure 5).

34
Correlation Between Exploratory Behaviour during the Sample Phase and
Discrimination Ratio in the Choice Phase. There was a significant negative correlation
between total sample exploration and choice DR in TG females at 5min r(10) = -.646, p =
.044 (Table 3).
Experiment 2: Object Location
2.1 5xFAD Object Location
Recognition Memory during the Choice Phase. A repeated measures ANOVA
demonstrated a significant main effect of genotype F(1, 40) = 4.528, p = 040 and a
significant genotype by sex interaction F(1, 40) = 4.945, p = .032.
Independent samples t-tests demonstrated a trending difference between WT and
TG female DR’s at 5min t(11.370) = 1.966, p = .074 and a significant difference at 3h
t(19) = 4.209, p < .001. Paired samples t-tests between sample and choice DR indicated
intact memory in WT females at 5min t(11) = 4.013, p = .002 and 3h t(9) = 2.663, p =
.026, impaired memory in TG females at 5min t(10) = .522, p = .613, and a trending
familiarity preference at 3h t(10) = 2.098, p = .062. Independent samples t-tests
demonstrated a non-significant difference between WT and TG male DR’s at 5min t(20)
= .946, p = .356 and 3h t(20) = -1.058, p = .303. Paired samples t-tests between sample
and choice DR indicated impaired memory in WT t(9) = -1.359, p = 207 and TG males
t(11) = -1.467, p = .170 at 5min. Paired samples t-tests between sample and choice DR
indicated impaired memory in WT males at 3h t(9) = -1.335, p = .215, but intact memory
in TG males t(11) = -3.099, p = .010 (Figure 6).

35
Exploratory Behaviour During the Sample and Choice Phase. There was a
significant main effect of delay on exploration during the sample phase F(1, 40) =
12.649, p = .001, a significant delay by genotype interaction F(1, 40) = 5.917, p = 020,
and a significant delay genotype by sex interaction F(1, 40) = 4.445, p = .041. There was
a significant main effect of genotype on total exploration during the choice phase F(1,
40) = 6.021, p = 019, a significant main effect of sex F(1, 40) = 11.210, p = .002, and a
significant genotype by sex interaction F(1, 40) = 4.319, p = 044 (Figure 7).
2.2 3xTG Object Location
Recognition Memory during the Choice Phase. A repeated measures ANOVA
demonstrated significant main effects of genotype F(1, 50) = 43.417, p < .001, and delay
F(1, 50) = 16.387, p < .001.
Independent samples t-tests demonstrated a significant difference between WT
and TG female DR’s at 5min t(27) = 3.247, p = .003 and 3h t(17.965) = 7.090, p < .001.
Paired samples t-tests between sample and choice DR indicated intact memory in WT
females at 5min t(13) = 5.264, p < .001 and 3h t(14) = 10.856, p < .001, and impaired
memory in TG females at 5min t(13) = 1.149, p = .271 and 3h t(13) = .576, p = .575.
Independent samples t-tests demonstrated a significant difference between WT and TG
male DR’s at 5min t(25) = 3.071, p = .005 and 3h t(23) = 2.738, p = .012. Paired samples
t-tests between sample and choice DR indicated intact memory in WT males at 5min
t(12) = 3.850, p = .002 and trending at 3h t(10) = 1.978, p = .076, and significantly

36
impaired memory in TG males at 5min t(13) = 1.251, p = .233 and a significant
familiarity preference at 3h t(12) = 2.518, p = .027 (Figure 6).
Exploratory Behaviour During the Sample and Choice Phase. There was a
significant main effect of delay on exploration during the sample phase F(1, 49) = 5.126,
p = .028, and a significant delay by genotype interaction F(1, 49) = 25.305, p < .001, on
exploration during the sample phase. There was a significant main effect of sex on
exploration during the choice phase F(1, 50) = 17.202, p < .001, such that females
generally explored more than males (Figure 7).
Experiment 3: Temporal Order
3.1 5xFAD Temporal Order
Recognition Memory during the Choice Phase. Independent sample t-tests
demonstrated a non-significant difference between WT and TG female DR’s at 3min
t(23) = 1.402, p = .174. Paired sample t-tests between sample and choice DR indicated
impaired memory in WT t(13) = -.453, p = .658 and TG t(10) = 1.370, p = .201 females
at 3min (Figure 8).
Exploratory Behaviour During the Sample and Choice Phase. There was a
significant effect of genotype on total exploration during sample phase one t(25) = 2.979,
p = .006, such that TG generally explored less than WT mice (Figure 9).

37
Correlation Between Exploratory Behaviour and Oddity Preference. There was a
significant positive correlation between total sample exploratory behaviour and choice
DR in WT females r(14) = .584, p = .028. There was a significant negative correlation
between total sample exploratory behaviour and choice DR in TG females r(11) = -.648,
p = .031 (Table 3).
3.2 3xTG Temporal Order
Recognition Memory during the Choice Phase. Independent samples t-tests
demonstrated a significant difference between WT and TG female DR’s at 3min t(17) =
2.239, p = .039. Paired samples t-tests between sample and choice DR indicated intact
memory in WT females at 3min t(7) = 5.589, p = .001, but impaired memory in TG
females t(10) = .022, p = .983 (Figure 8).
Exploratory Behaviour During the Sample and Choice Phase. There were no
significant effects of genotype on total exploration during the sample and choice phases
(Figure 9).
Experiment 4: Multisensory Object Oddity
4.1 5xFAD Multisensory Object Oddity
Oddity Preference. A three-way repeated measures ANOVA demonstrated a
significant main effect of task F(2, 78) = 3.740, p = .035 and a significant task by sex
interaction F(2, 78) = 4.302, p = .022.

38
Independent samples t-tests demonstrated a significant difference between WT
and TG females on the MSO task t(19) = 3.547, p = .002. One-sample t-tests indicated
MSO oddity scores above chance in WT females t(9) = 6.018, p < .001, but not in TG
females t(10) = .058, p = .955. Independent samples t-tests demonstrated a significant
difference between WT and TG females on the visual task t(19) = 3.547, p = .002. One-
sample t-tests demonstrated visual perception above chance performance in WT females
t(9) = 4.4092, p = .003 and TG females t(10) = 3.366, p = .007. Independent samples t-
tests demonstrated a non-significant difference between WT and TG females on the
tactile task t(19) = .303, p = .765. One-sample t-tests demonstrated tactile perception
above chance performance in WT females t(9) = 3.062, p = .014 and TG females t(10) =
4.502, p = 4.502, p = .001. Independent samples t-tests demonstrated a non-significant
difference between WT and TG males on the MSO task t(20) = .863, p = .398. One-
sample t-tests demonstrated impaired multisensory perception in WT t(9) = -.409, p =
.692 and TG males t(11) = -1.978, p = .073. Independent samples t-tests indicated no
difference between WT and TG males on the visual tsk t(20) = .189, p = .852. One-
sample t-tests demonstrated intact visual perception in WT males t(9) = 2.749, p = 023,
but impaired visual perception in TG males t(11) = 1.619, p = .134. Independent samples
t-tests demonstrated a non-significant difference between WT and TG males on the tactile
task t(20) = .778, p = .447. One-sample t-tests demonstrated impaired tactile perception
in WT t(9) = 2.123, p = 066 and TG males t(11) = 1.127, p = .286 (Figure 10).
Total Exploratory Behaviour. There was a significant main effect of task on
exploration during the sample phase F(2, 78) = 27.775, p < .001 and a significant task by

39
genotype interaction on exploration during the sample phase F(2, 78) = 3.628, p = .043.
There was a significant main effect of sex F(1, 39) = 22.560, p < .001 and genotype F(1,
39) = 12.781, p = .001 on exploration during the sample phase (Figure 11).
Correlation Between Exploratory Behaviour and Oddity Preference. There was a
significant negative correlation between total exploratory behaviour and MSO oddity
preference TG females r(11) = -.822, p = .002. There was a significant positive
correlation between total exploratory behaviour and visual oddity preference in TG males
r(12) = .737, p = .006 (Table 3).
4.2 3xTG Multisensory Object Oddity
Oddity Preference. A three-way repeated measures ANOVA demonstrated a
significant main effect of task , F(2, 104) = 4.795, p = .010 and a significant task by
genotype interaction F(1, 104) = 6.967, p = .001.
Independent samples t-tests demonstrated a significant difference between WT
and TG females on the MSO task t(27) = 3.218, p = .003. One-sample t-tests
demonstrated multisensory perception above chance performance in WT females t(14) =
5.333, p < .001, and in TG females t(13) = 2.156, p = .050. Independent samples t-tests
demonstrated a non-significant difference between WT and TG females on the visual task
t(27) = .295, p = .770. One-sample t-tests demonstrated visual perception above chance
performance in WT females t(14) = 5.627, p < .001, and in TG females t(13) = 4.027, p =
.001. Independent samples t-tests demonstrated a non-significant difference between WT

40
and TG females on the tactile task t(27) = 1.232 , p = .229. One-sample t-tests
demonstrated tactile perception above chance performance in WT females t(14) = 6.460,
p < .001, and in TG females t(13) = 3.788, p = .002. Independent samples t-tests
demonstrated a significant difference between WT and TG males on the MSO task t(25)
= 3.059, p = .005. One-sample t-tests demonstrated multisensory perception above
chance performance in WT males t(12) = 4,572, p =.001, but impaired multisensory
perception in TG males t(13) = .401, p = .695. Independent samples t-tests indicated no
difference between WT and TG males on the visual task t(25) = 1.901, p = .069. One-
sample t-tests demonstrated visual perception above chance performance in TG males
t(13) = 3.609, p = .003, but not WT males t(12) = 1.274, p = .227. Independent samples t-
tests demonstrated a non-significant difference between WT and TG males on the tactile
task t(25) = .858, p = .399. One-sample t-tests demonstrated visual perception above
chance performance in WT males t(12) = 4.535, p = .001 and TG males t(13) = 5.065, p <
.001 (Figure 10).
Total Exploratory Behaviour. There was a significant main effect of task on
exploration during the sample phase F(2, 104) = 18.988, p < .001, as mice generally
explored the objects more in the tactile oddity task (Figure 11).
Discussion
Results are consistent with a multifaceted impairment in object processing in
5xFAD and 3xTG transgenic mice. Memory for object identity, as evaluated using open-
field and Y-apparatus OR paradigms, is dissociable across transgenic AD strain and sex.

41
Specifically, 5xFAD males and females were impaired on open-field OR when the
retention delay was 5min or 3h. However, when spatial and contextual cues were
minimized, using a modified Y-apparatus, 5xFAD males and females are impaired at
5min and 3h. However, the 5xFAD female WT were unable to perform open-field OR at
3h. 3xTG males were impaired on open-field OR at 5min and 3h, whereas females were
selectively impaired at 3h. 3xTG males and females were selectively impaired on Y-
apparatus OR at 3h. Conversely, memory for the spatial location of objects, assessed
using the object location paradigm, was impaired in 5xFAD females and 3xTG females
with 5min and 3h delays. 3xTG males had impaired OL at 5min but a familiarity
preference at 3h. 5xFAD WT and TG males had impaired OL at 5min, whereas only WT
males had impaired OL at 3h. Temporal processing was also impaired in WT and
transgenic 5xFAD females and transgenic 3xTG females at 3min. Lastly, multisensory
perception was impaired in 5xFAD females, as well as 3xTG males and females, despite
intact basic visual and tactile object perception. 5xFAD WT and TG males had impaired
multisensory perception. 5xFAD TG males were also impaired on visual and tactile
object perception and 5xFAD WT males had impaired tactile perception (Table 4). It is
important to note that when WT performance is impaired it is difficult to make
conclusions about object processing in TG mice.
Object Recognition
In agreement with previous research, we have demonstrated impaired object
recognition when tested in an open-field with a retention delay ≥ 5min (Ambrée et al.,
2009; Bardgett et al., 2011; Dewachter et al., 2002; J. Dodart et al., 1999; Gerenu et al.,

42
2013; Giuliani et al., 2013; Greco et al., 2010; Hillen et al., 2010; Spilman et al., 2014;
Yuede et al., 2009). However, when we evaluated OR in the Y-apparatus behavioural
deficits appeared to be less severe in some cases, as indicated by intact short-term
memory in 3xTG males.
Better OR performance in the Y-apparatus is likely related to facilitated
processing of the objects themselves. There are several significant differences between
WT and TG exploration in the open-field that were not observed in the Y-apparatus,
particularly in the 5xFAD strain. Although object exploration during the sample phase
can positively correlate with DR’s (5xFAD female 5min Y-apparatus OR; 5xFAD WT
male visual oddity; Albasser, Davies, Futter, & Aggleton, 2009), in our recognition
experiments object exploration does not consistently correlate with choice DR’s. Indeed,
increased object exploration does not necessarily improve object encoding (Akkerman et
al., 2012; Gaskin et al., 2010). Gaskin et al. (2010), for example, found that the degree of
preference for the novel object does not correlate with total object exploration during the
sample phase, but rather a minimum amount of exploration is required to observe a
novelty preference. Therefore, increased exploration may not explain better OR
performance in the Y-apparatus. Still, the Y-apparatus appears to increase focus on
objects and has less potentially distracting or interfering contextual stimuli than the open-
field. Perhaps transgenic AD mice have insufficient mnemonic resources for object and
environmental information in the open-field. Indeed, familiarity with the testing
environment can influence DR’s (Besheer & Bevins, 2000; Gaskin et al., 2010;
Wilkinson, Herrman, Palmatier, & Bevins, 2006); Wilkinson et al. (2006), for example,

43
demonstrated that rats familiarized with the test environment had greater exploration of
the novel object.
Enhanced OR memory at short retention delays and when spatial information is
limited may also be indicative of the severity of AD neuropathology in the HPC.
Shortening the retention delay and limiting spatial information may alter the necessity of
the HPC in OR memory. Hammond et al., (2004) temporarily inactivated the HPC and
evaluated short (5min) and long-term OR memory in mice. OR was intact at 5min but
impaired at 24h. This finding suggests the HPC is critically involved in OR at long
retention delays, whereas at short delays the PRh is likely sufficient to support OR
memory. Furthermore, a contextually rich open-field OR paradigm may encourage
increased processing of the spatial environment and require functional integrity of the
HPC as well as the PRh (Barker & Warburton, 2011; Forwood et al., 2005; Winters et al.,
2004). Although Aβ, hyperphosphorylated tau and neuronal loss are present in the HPC
and association cortices areas, including the PRh (Braak & Braak, 1991, 1995; Davis,
Easton, et al., 2013; Oakley et al., 2006; Oddo, Caccamo, Shepherd, et al., 2003; Scahill
et al., 2002; Whitwell et al., 2007) by 12 months of age, our findings, and others (Davis,
Easton, et al., 2013), suggest that HPC-dependent object processing may be more
sensitive to AD pathology. Interestingly, 5xFAD mice appear to be more impaired in
SOR than 3xTG, as shown by a delay-independent impairment in OR paradigms.
Behavioural deficits may be more severe in the 5xFAD model because 3xTG mice do not
have hippocampal neuronal loss and have a less aggressive Aβ pathology (Kastyak-

44
Ibrahim et al., 2013; Li, Ebrahimi, & Schluesener, 2013; Manaye et al., 2013; Oakley et
al., 2006; Oddo, Caccamo, Shepherd, et al., 2003).
AD prevalence, incidence and Aβ plaque deposition is higher in women than men
(Corder et al., 2004; Rocca, Amaducci, & Schoenberg, 1986; Ruitenberg, Ott, Van
Swieten, Hofman, & Breteler, 2001). Similarly, 5xFAD mice have an early and
aggressive Aβ pathology that is more severe in female mice (Bhattacharya, Haertel,
Maelicke, & Montag, 2014; Oakley et al., 2006; Reid & Darvesh, 2015). Since Aβ
pathology correlates with impaired synaptic plasticity and memory (Chapman et al.,
1999; Oddo, Caccamo, Shepherd, et al., 2003; Puoliväli et al., 2002; Shankar et al.,
2008), perhaps a more aggressive amyloid pathology is sufficient to block the beneficial
effects of short retention delays and limited spatial information on OR memory in female
5xFAD mice. Conversely, 3xTG females had a less severe OR deficit than males.
Specifically, short-term OR memory was intact in 3xTG females in open-field and Y-
apparatus OR, whereas 3xTG males had short-term open-field OR deficits that were not
observed at short retention delays in the Y-apparatus by limiting spatial information and
increasing object exploration. Our finding is consistent with sexual dimorphism in
working memory, such that 3xTG males have a more severe behavioural deficit
beginning at two-months-of-age (Stevens & Brown, 2015). Although a more aggressive
amyloid pathology has been reported in 3xTG females (Carroll et al., 2010; Clinton et al.,
2007; Hirata-Fukae et al., 2008), there may be differential effects of organizational and
activational sex hormones on AD pathology (Carroll et al., 2007, 2010). Perinatal female
sex hormones increase adult 3xTG vulnerability to Aβ (Carroll et al., 2010). Conversely,

45
activational estrogens are believed to be neuroprotective against Alzheimer’s pathology.
Ovariectomy-induced depletion of sex hormones in 3xTG females increases Aβ
accumulation and impairs HPC-dependent memory, an effect that can be attenuated by
estrogen treatment (Carroll & Pike, 2008; Carroll et al., 2007; Levin-allerhand,
Lominska, Wang, & Smith, 2002; Zheng et al., 2002). Interestingly, tauopathy correlates
more strongly with behavioural deficits in human AD patients (Giannakopoulos et al.,
2003; Guillozet, Weintraub, Mash, & Mesulam, 2003) and mnemonic improvement
requires alteration of both Aβ and tau pathology in 3xTG mice (Oddo, Caccamo, et al.,
2006a). Sexual dimorphism in 3xTG tauopathy has not been reported (Hirata-Fukae et
al., 2008); however adult progesterone and estrogen treatment reduces tau
hyperphosphorylation in 3xTG females and 3xTG males, respectively (Carroll et al.,
2007; Rosario, Carroll, & Pike, 2010). Thus, it is possible that female adult sex hormones
are protective against AD neuropathology and object recognition deficits in 3xTG
females but not 5xFAD females. It is also possible that differences in male and female
longevity contributed to sex differences in 3xTG mice. Specifically, 3xTG TG females
have a longer lifespan than TG males (Rae & Brown, 2015).
Object Location
Processing of object spatial location was also compromised in transgenic AD
mice. Impaired object location memory in 5xFAD and 3xTG mice is consistent with
abnormal spatial processing of objects (abnormal canonical orientation and mental
rotation; Caterini et al., 2002) and spatial memory (Amieva et al., 2005; Blackwell,
Vesey, Semple, Robbins, 2003; Lee et al., 2014; Toledo-Morrell & Dickerson, 2000) in

46
human AD patients, as well as impaired contextual fear conditioning (Espana et al., 2010;
Jacobsen et al., 2006; Kimura & Ohno, 2009; Medeiros et al., 2014); Morris water maze
(Billings, Green, McGaugh, & LaFerla, 2007; Billings et al., 2005; Chen et al., 2000;
Clinton et al., 2007; Gulinello et al., 2009; Kotilinek et al., 2002; Lesné et al., 2006;
Medeiros et al., 2014; Puoliväli et al., 2002; Schneider, Baldauf, Wetzel, & Reymann,
2014; Zhang et al., 2013); and object location (Bollen et al., 2013; Davis, Easton, et al.,
2013; Dodel et al., 2011; Frye & Walf, 2008; Good et al., 2007; Gulinello et al., 2009;
Kornecook, McKinney, Ferguson, & Dodart, 2010; Kroker et al., 2014; Ma et al., 2013;
Masciopinto et al., 2012; Middei et al., 2006) memory in transgenic AD mice.
Paradoxically, OL was selectively intact in 5xFAD males at 3h. Short- but not
long-term memory deficits have been reported in rodents, and suggest STM and LTM
have some distinct non-overlapping neurobiological mechanisms (Barker, 2006;
Izquierdo, Medina, Vianna, Izquierdo, & Barros, 1999; Morice et al., 2008). Given the
OL impairment at 5min, the inability of WT males to perform OL at 3h and impaired
long-term memory on all other object processing tasks, it may be problematic to conclude
spatial memory is intact in 5xFAD males at 3h. It is possible other non-mnemonic factors
may have contributed to task performance. A significant OL familiarity preference was
observed in transgenic 3xTG males at 3h. A familiarity preference may imply intact
memory, and could reflect abnormities in habituation, dishabituation or novelty
preference. Abnormal dishabituation, but not habituation, and neophobia to a novel
context have been observed in APP transgenic mice (Hsiao et al., 1995; Sanchez et al.,
2012). However, we occasionally observe familiarity preferences in object recognition

47
data that do not consistently reflect mnemonic abilities. For example, a preference for the
familiar side-to-be is present in the sample phase of open-field OR in 5xFAD females at
3h. Further, the familiarity preference was not observed at the 5min delay. In this case, it
is possible the familiarity preference is due to a sample DR greater than zero.
Impaired OL across strain and sex suggests that spatial processing of objects may
be more difficult for transgenic AD mice than the processing of object features. Indeed,
previous findings with APP/PS1 and 3xTG mice have demonstrated impaired OL but not
OR (Davis, Easton, et al., 2013; Frye & Walf, 2008; Gulinello et al., 2009), which may
be related to the development of OL impairments prior to standard OR (Middei et al.,
2006). At 12 months of age Aβ and hyperphosphorylated tau are present in the HPC of
5xFAD and 3xTG mice (Kanno, Tsuchiya, & Nishizaki, 2014a; Oakley et al., 2006;
Oddo, Caccamo, Shepherd, et al., 2003), while 5xFAD mice also have hippocampal
synaptic loss (Oakley et al., 2006). As OL is a HPC-dependent task this finding provides
additional support for susceptibility of HPC-dependent processes to AD pathology
(Assini et al., 2009; Oakley et al., 2006; Oddo, Caccamo, Shepherd, et al., 2003).
However, 5xFAD male WT mice were unable to perform the OL task at both retention
delays and 3xTG WT males were unable to perform OL at 3h, and it is possible that some
OL deficits in males may be related to general aging rather than AD pathology. Indeed,
female rodents can outperform males on OL tasks (Saucier, Shultz, Keller, Cook, &
Binsted, 2008), and HPC dependent OL is impaired in aged mice (Wang, Li, Dong, Lv,
& Tang, 2009; Wimmer, Hernandez, Blackwell, & Abel, 2012).

48
Temporal Order
We also report that temporal processing of object information is impaired in AD
models. 5xFAD and WT female mice were unable to perform the temporal order task,
whereas transgenic 3xTG females failed to differentiate between objects presented in
sequential order. Impaired temporal object processing is in agreement with impaired
temporal context memory in human AD patients (Becker et al., 1993; Kopelman, 1989).
Specifically, AD patients were impaired at identifying when news and autobiographical
events occurred in time and how recently an object or image had been presented.
Previous examination of temporal order memory in AD transgenic mice has failed to
demonstrate an impairment in temporal processing (Davis, Easton, et al., 2013; Hale &
Good, 2005). However, differences in strain, age of testing and retention delay might
account for discrepancies in mnemonic impairment. Hale & Good (2005) evaluated
temporal order in Tg2576 male mice (a model of Aβ pathology) at 14 months of age with
a 2min retention delay. Unlike the 3xTG model, Tg2576 does not recapitulate tauopathy
and a slightly shorter retention delay should have reduced the mnemonic demand. Davis
et al. (2013) assessed temporal order in 3xTG female mice at 4.5 months of age with a 5
min retention delay. Although mnemonic deficits can be detected at 4.5-months-of-age in
3xTG mice (Billings et al., 2005), Aβ and tau pathologies do not develop until 6 and 12
months of age, respectively (Oddo, Caccamo, Shepherd, et al., 2003). Therefore,
impaired temporal processing at 12 months of age, as reported here, is likely related to a
more developed AD pathology (Oakley et al., 2006; Oddo, Caccamo, Shepherd, et al.,
2003).

49
The TO impairment reported here in 3xTG mice reveals a distinct behavioural
impairment in temporal processing of objects. Given the need for a shorter retention
delay (WT and TG could not perform TO with 5min retention delays; data not shown),
and the fact that a deficit in the standard OR paradigm would predict preferential
exploration of the first (more remote) object rather than equal preference for both objects,
TO appears to be more difficult than basic OR and it is unlikely that impairments are due
to an inability to recognize the objects per se. Thus, object processing required for TO
may be differentially affected by AD pathology. Since the HPC, PRh, and mPFC network
is necessary for temporal object processing, TO deficits reemphasize the vulnerability of
the MTL to AD pathology and suggests impaired connectivity between the HPC+PRh
and HPC+mPFC (Barker & Warburton, 2011; Hannesson et al., 2004; Mitchell &
Laiacona, 1998). Consistent with this idea, AD neuropathology causes degeneration of
myelin (Xu et al., 2001) and may damage neural networks (Bartzokis, 2004). Extensive
Aβ and tau pathology in the perforant pathway (the principle pathway relaying input from
entorhinal cortex to the HPC; Steward, 1976; Van Hoesen, Pandya, & Butters, 1972),
CA1 and subiculum (HPC output; Rosene & Van Hoesen, 1977; Swanson, Wyss, &
Cowan, 1978) in AD patients may effectively disconnect the HPC from cortical
structures (Hyman et al., 1984; Hyman, Van Hoesen, Kromer, & Damasio, 1986; Wang
et al., 2006). Alzheimer’s patients have reduced functional connectivity between the
prefrontal cortex and HPC and impaired memory when performing a delayed match to
sample paradigm (Grady, Furey, Pietrini, Horwitz, & Rapoport, 2001). Aβ also disrupts
connectivity in transgenic AD mice (Hsai et al., 1999), and disconnection between the
cortex and HPC is present in PDAPP mice. Specifically aged PDAPP mice have atrophy

50
of the corpus callosum, fornix and HPC (Gonzalez-Lima, Berndt, Valla, Games, &
Reiman, 2001). In 3xTG mice, white matter atrophy in the entorhinal cortex and HPC is
evident by 2 months of age (Desai et al., 2009), and may disrupt the HPC, PRh and
mPFC network required for temporal memory.
Multisensory Object Oddity
Multisensory integration is a neglected facet of object processing in the field of
AD research. Notably, the current results represent the first demonstration of impaired
multisensory processing in transgenic models of AD. Here, we have shown a selective
impairment on visual-tactile perceptual binding in 5xFAD female and 3xTG mice,
whereas visual-only and tactile-only object perception were intact. Both TG and WT
5xFAD male mice were impaired on visual-tactile and tactile-only object perception.
Further, 5xFAD male TG were also impaired on visual-only object perception.
Previous findings with human patients also suggest abnormal multisensory
integration in AD (Wu et al., 2012). Control, MCI and AD patients performed audio-
visual tasks in which they responded to either audio, visual, or audio-visual stimuli. AD
patients could accurately perform multimodal audio-visual trials but reaction times were
significantly longer, suggesting that AD pathology attenuates the well-established
beneficial effects of multisensory integration. Delbeuck et al. (2007) have also
demonstrated impaired audio-visual speech perception in AD patients. Specifically, AD
patients, unlike age-matched controls, failed to combine dissimilar audio and visual
speech information.

51
Multisensory perception involves the binding of information across sensory
modalities, which can require functional connectivity of modality-specific sensory
processing and multisensory areas (Cloke et al., 2014; Hindley, Nelson, Aggleton, &
Vann, 2014; Jacklin, Cloke, Potvin, Garrett, & Winters, 2016; Winters & Reid, 2010).
Consistent with multisensory impairment, Aβ plaques and neurofibrillary tangles disrupt
cortical connectivity, specifically areas that receive input from multimodal areas (de
LaCoste & White, 1993; Delbeuck, Van der Linden, & Collette, 2010). Indeed, Golob et
al. (2001) demonstrated abnormal sensory cortical evoked potentials elicited by pairs of
audio-visual stimuli. Specifically, only when a visual stimulus was presented prior to an
auditory stimulus was a refractory effect not present in AD patients. This finding suggests
a disconnection between, but not within, visual and auditory cortices (Delbeuck et al.,
2010; Golob et al., 2001). In aged APP/PS1 mice cortico-cortical fibers, including those
originating in the visual cortex, are preferentially disrupted compared to fibers originating
from subcortical structures (Delatour, Blanchard, Pradier, & Duyckaerts, 2004). Although
transgenic AD mice have distinct pathological profiles, impaired connectivity between
cortical regions may impair binding of visual and tactile object information, but not
unimodal visual or tactile information, in 5xFAD and 3xTG mice.
General Discussion
Memory deficits in AD are widespread and are related to impaired working
memory (Baddeley, Logie, Bressi, Sala, & Spinnler, 1986; Baddeley, Bressi, Della Sala,
Logie, & Spinnler, 1991), as well as the inability to learn, encode and store new

52
information (Delis et al., 1991; Golby et al., 2005; Granholm & Butters, 1988; Hodges,
Salmon, & Butters, 1990; Pihlajamaki, O’Keefe, O’Brien, Blacker, & Sperling, 2011;
Sperling et al., 2003; Weintraub, Wicklund, & Salmon, 2012; White & Ruske, 2002). For
example, AD patients have difficulty performing multiple tasks at once and recalling
word lists, geometric shapes, and face-name pairs. Recently, selective HPC-dependent
memory encoding deficits have also been reported in young APP/PS1 mice (Roy et al.,
2016). However, as AD pathology progresses behavioural deficits become more severe
and various mnemonic processes may be compromised.
The current results suggest that 12-month-old 5xFAD and 3xTG mice do not have
universal deficits in object perception and memory. Specifically, we have observed
impairments in cognitive processes, ranging from object identity and spatial memory to
multisensory object feature binding. Given that basic unimodal tactile and visual object
perception are intact across strains as well as short-term OR memory in 3xTG mice, basic
encoding of objects is likely intact. Although we have observed short-term object
processing impairments for object identity, location and temporal recency, these deficits
may be attributed to increased susceptibility of complex conjunctive object
representations and short-term or working memory to AD pathology. HPC, PRh, mPFC
networks that are required for hierarchical object representations in which the PRh is
important for storage of complex conjunctive object feature representations, whereas the
HPC and mPFC combine object representations with space, context and time (Barker &
Warburton, 2011; Cowell, Bussey, & Saksida, 2006, 2010) are vulnerable to AD
pathology (Gonzalez-Lima et al., 2001; Grady et al., 2001). This network is affected in

53
5xFAD and 3xTG mice. In 3xTG mice, by 12 months of age Aβ is prevalent in the
frontal cortex, entorhinal cortex, and hippocampus and hyperphosphorylated tau is
evident in the HPC (Janelsins et al., 2005; Oddo, Caccamo, Kitazawa, Tseng, & LaFerla,
2003; Oddo, Caccamo, et al., 2006b; Oddo, Caccamo, Shepherd, et al., 2003; Oddo,
Vasilevko, et al., 2006). An association between type of object processing and retention
delay was not generally observed in 5xFAD mice. It could be argued that broad delay-
independent impairments in 5xFAD mice are due to a more aggressive AD pathology.
Specifically, by 12 months of age 5xFAD mice have a very extensive Aβ pathology in the
frontal and entorhinal cortices and HPC, hyperphosphorylation of tau in the HPC, as well
as synaptic, neuronal and glial loss in the cortex and HPC (Bhattacharya et al., 2014;
Giannoni et al., 2013; Kanno et al., 2014a; Oakley et al., 2006). Critically, WT 5xFAD
mice were impaired on several object processing paradigms, ergo behavioural
impairments may also be related to accelerated aging and other abnormalities in the
5xFAD background strain. Indeed, the longevity of the 5xFAD WT background strain
SJL/J may be lower than the 3xTG background strain 129S1/SV1mJ (D’Antona et al.,
2010; Jaszberenyi et al., 2012; Rae & Brown, 2015). A more robust impairment was
observed in 5xFAD WT males and may be related to a shorter lifespan (Rae & Brown,
2015).
Due to the widespread damage from AD pathology in aged 5xFAD and 3xTG
mice that includes Aβ deposition and neuronal loss in the frontal cortex and HPC,
impaired short-term and working memory have relevance to object processing deficits.
For example, the putative visuo-spatial sketchpad is a human working memory scheme

54
involved in the temporary storage and manipulation of visual information, allowing the
time-limited maintenance of object features necessary to recognize a recently
encountered object (Baddeley, 2003). In our rodent object processing paradigms, working
memory may be required during the choice phase to maintain information about the
relative familiarity of objects.
Abnormal cholinergic function could be a central mechanism of object processing
dysfunction in transgenic AD mice. Cholinergic transmission is altered in these strains
(Da Cruz et al., 2012; Machová et al., 2008; Watanabe et al., 2009), and ACh is critical to
various facets of object processing. Specifically, ACh is implicated in working memory
(Fadda, Melis, & Stancampiano, 1996; Seeger et al., 2004), object recognition memory
(Aigner, Walker, & Mishkin, 1991; Bartko, Winters, Saksida, & Bussey, 2014; Prado et
al., 2006; Tinsley et al., 2011; Warburton et al., 2003; Winters et al., 2006), object
location (Murai et al., 2007), and temporal order (De Castro et al., 2009). There is also
evidence that ACh may be involved in synaptic plasticity (Bashir, 2003; Massey, Bhabra,
Cho, Brown, & Bashir, 2001; Warburton et al., 2003), which suggests a function of ACh
in object memory consolidation (Abe, Ishida, & Iwasaki, 2004). Furthermore, drugs that
increase cholinergic transmission attenuate object recognition deficits in transgenic AD
mice (Francis et al., 2012; Medeiros et al., 2014; Zhang et al., 2012).
It is important to consider non-mnemonic confounds on task performance because
some object processing impairments reported here were delay-independent. As object
oddity was generally intact in visual-only and tactile-only trials (excluding 5xFAD

55
males), basic object perception appears to be intact. However, abnormal emotional,
motor, and attention behaviour has been reported in transgenic AD mice. Attention
appears to be abnormal in transgenic AD mice. Unpublished data from our group, and
others (Romberg, Mattson, Mughal, Bussey, & Saksida, 2011), has shown impaired
attention on the 5-choice serial reaction time test in 3xTG and 5xFAD male and female
mice. Alterations in exploratory behaviour are consistent with previous studies (Arsenault
et al., 2011) and may relate to abnormal anxiety-like behaviour, and motor function in
transgenic AD mice. Aged 3xTG mice have increased anxiety- like behaviours including
increased restlessness, startle responses, and freezing (Espana et al., 2010; García-Mesa
et al., 2012; St-Amour et al., 2014; Sterniczuk, Antle, Laferla, & Dyck, 2010), whereas
motor behaviour is more complex with accounts of hypoactivity (García-Mesa et al.,
2012) or normal locomotion in the open-field (St-Amour et al., 2014), as well as
enhanced performance on other motor tasks (Filali et al., 2012; Stover et al., 2015).
Conversely, 5xFAD mice have reduced anxiety- like behaviours and motor deficits
(Jawhar, Trawicka, Jenneckens, Bayer, & Wirths, 2012; Schneider et al., 2014).
However, abnormal exploratory behaviour did not consistently correlate with OR
memory in the current study. Differences between WT and TG exploration, where TG
explored objects less than WT, were present in the OR, OL and MSO open-field
paradigms. In OR, exploration differences were not present in the Y-apparatus, but
similar behavioural patterns were observed. Thus, abnormal exploratory behaviour does
not consistently explain OR deficits in the current study.

56
In conclusion, object recognition is not a unitary cognitive function. In the current
study, the systematic characterization of different facets of object processing (object
identity, spatial location, temporal relationships, and multisensory features), reliant on the
integrity of distinct neurobiological substrates, across strain and sex has helped to clarify
the nature of behavioural deficits and the regional susceptibility to AD pathology in
transgenic mouse models.

57
References
Abe, H., Ishida, Y., & Iwasaki, T. (2004). Perirhinal N-methyl-D-aspartate and
muscarinic systems participate in object recognition in rats. Neuroscience Letters,
356(3), 191–194. doi:10.1016/j.neulet.2003.11.049
Abildayeva, K., Berbée, J. F. P., Blokland, A., Jansen, P. J., Hoek, F. J., Meijer, O., …
Mulder, M. (2008). Human apolipoprotein C-I expression in mice impairs learning
and memory functions. Journal of Lipid Research, 49(4), 856–69.
doi:10.1194/jlr.M700518-JLR200
Aigner, T. G., Walker, D. L., & Mishkin, M. (1991). Comparison of the effects of
scopolamine administered before and after acquisition in a test of visual recognition
memory in monkeys. Behavioral and Neural Biology, 55(1), 61–67.
doi:10.1016/0163-1047(91)80127-Z
Akkerman, S., Blokland, A., Reneerkens, O., van Goethem, N. P., Bollen, E., Gijselaers,
H. J. M., … Prickaerts, J. (2012). Object recognition testing: Methodological
considerations on exploration and discrimination measures. Behavioural Brain
Research, 232(2), 335–347. doi:10.1016/j.bbr.2012.03.022
Albasser, M. M., Davies, M., Futter, J. E., & Aggleton, J. P. (2009). Magnitude of the
object recognition deficit associated with perirhinal cortex damage in rats: Effects of
varying the lesion extent and the duration of the sample period. Behavioral
Neuroscience, 123(1), 115–124. doi:10.1037/a0013829
Alonso Vilatela, M. E., López-López, M., & Yescas-Gómez, P. (2012). Genetics of
Alzheimer’s disease. Archives of Medical Research, 43(8), 622–31.
doi:10.1016/j.arcmed.2012.10.017

58
Alonso, a C., Grundke-Iqbal, I., & Iqbal, K. (1996). Alzheimer’s disease
hyperphosphorylated tau sequesters normal tau into tangles of filaments and
disassembles microtubules. Nature Medicine, 2(7), 783–787. doi:10.1038/nm0796-
783
Alzheimer, A. (1911). Uber eigenartige Krankheitsfalle des spateren Alters (On certain
peculiar diseases of old age).
Ambrée, O., Richter, H., Sachser, N., Lewejohann, L., Dere, E., de Souza Silva, M. A.,
… Schäbitz, W.-R. (2009). Levodopa ameliorates learning and memory deficits in a
murine model of Alzheimer’s disease. Neurobiology of Aging, 30(8), 1192–204.
doi:10.1016/j.neurobiolaging.2007.11.010
Amieva, H., Jacqmin-Gadda, H., Orgogozo, J.-M., Le Carret, N., Helmer, C., Letenneur,
L., … Dartigues, J.-F. (2005). The 9 year cognitive decline before dementia of the
Alzheimer type: a prospective population-based study. Brain : A Journal of
Neurology, 128(Pt 5), 1093–1101. doi:10.1093/brain/awh451
Arendt, T., Brückner, M. K., Mosch, B., & Lösche, A. (2010). Selective cell death of
hyperploid neurons in Alzheimer’s disease. The American Journal of Pathology,
177(1), 15–20. doi:10.2353/ajpath.2010.090955
Armstrong, R. (2013). Review article What causes alzheimer’s disease? Folia
Neuropathologica, 3(3), 169–188. doi:10.5114/fn.2013.37702
Arriagada, P. V, Growdon, J. H., Hedleywhyte, E. T., & Hyman, B. T. (1992).
Neurofibrillary Tangles But Not Senile Plaques Parallel Duration and Severity of
Alzheimers-Disease. Neurology, 42(3), 631–639.
Arrieta-Cruz, I., Pavlides, C., & Pasinetti, G. M. (2010). Deep Brain Stimulation in

59
Midline Thalamic Region Facilitates Synaptic Transmission and Shortterm Memory
in a Mouse Model of Alzheimer’S Disease. Translational Neuroscience, 1(3), 188–
194. doi:10.2478/v10134-010-0023-x
Arsenault, D., Julien, C., Tremblay, C., & Calon, F. (2011). DHA improves cognition and
prevents dysfunction of entorhinal cortex neurons in 3xTg-AD mice. PloS One, 6(2),
e17397. doi:10.1371/journal.pone.0017397
Ashe, K. H., & Zahs, K. R. (2010). Review Probing the Biology of Alzheimer ’ s Disease
in Mice. Neuron, 66(5), 631–645. doi:10.1016/j.neuron.2010.04.031
Assini, F. L., Duzzioni, M., & Takahashi, R. N. (2009). Object location memory in mice:
Pharmacological validation and further evidence of hippocampal CA1 participation.
Behavioural Brain Research, 204(1), 206–211. doi:10.1016/j.bbr.2009.06.005
Avila, J. (2006). Tau phosphorylation and aggregation in Alzheimer’s disease pathology.
FEBS Letters, 580(12), 2922–7. doi:10.1016/j.febslet.2006.02.067
Bäckman, L., Jones, S., Berger, a. K., Laukka, E. J., & Small, B. J. (2004). Multiple
cognitive deficits during the transition to Alzheimer’s disease. Journal of Internal
Medicine, 256(3), 195–204. doi:10.1111/j.1365-2796.2004.01386.x
Baddeley, A. (2003). Working memory: looking back and looking forward. Nature
Reviews Neuroscience, 4(10), 829–839. doi:10.1038/nrn1201
Baddeley, A. D., Bressi, S., Della Sala, S., Logie, R., & Spinnler, H. (1991). The Decline
of Working Memory in Alzheimer’S Disease. Brain, 114(6), 2521–2542.
doi:10.1093/brain/114.6.2521
Baddeley, A., Logie, R., Bressi, S., Sala, S. Della, & Spinnler, H. (1986). Dementia and
Working Memory. The Quarterly Journal of Experimental Psychology, 38, 603–

60
618. doi:10.1080/14640748608401616
Balducci, C., Mehdawy, B., Mare, L., Giuliani, A., Lorenzini, L., Sivilia, S., … Nisticò,
R. (2011). The γ-secretase modulator CHF5074 restores memory and hippocampal
synaptic plasticity in plaque-free Tg2576 mice. Journal of Alzheimer’s Disease :
JAD, 24(4), 799–816. doi:10.3233/JAD-2011-101839
Bancher, C., Braak, H., Fischer, P., & Jellinger, K. a. (1993). Neuropathological staging
of Alzheimer lesions and intellectual status in Alzheimer’s and Parkinson's disease
patients. Neuroscience Letters, 162(1-2), 179–182. doi:10.1016/0304-
3940(93)90590-H
Barbeau, E. J., Ranjeva, J. P., Didic, M., Confort-Gouny, S., Felician, O., Soulier, E., …
Poncet, M. (2008). Profile of memory impairment and gray matter loss in amnestic
mild cognitive impairment. Neuropsychologia, 46(4), 1009–1019.
doi:10.1016/j.neuropsychologia.2007.11.019
Barbero-Camps, E., Fernández, A., Martínez, L., Fernández-Checa, J. C., & Colell, A.
(2013). APP/PS1 mice overexpressing SREBP-2 exhibit combined Aβ accumulation
and tau pathology underlying Alzheimer’s disease. Human Molecular Genetics,
22(17), 3460–76. doi:10.1093/hmg/ddt201
Bardgett, M. E., Davis, N. N., Schultheis, P. J., & Griffith, M. S. (2011). Ciproxifan, an
H3 receptor antagonist, alleviates hyperactivity and cognitive deficits in the APP
Tg2576 mouse model of Alzheimer’s disease. Neurobiology of Learning and
Memory, 95(1), 64–72. doi:10.1016/j.nlm.2010.10.008
Barker, G. R. I. (2006). The Different Effects on Recognition Memory of Perirhinal
Kainate and NMDA Glutamate Receptor Antagonism: Implications for Underlying

61
Plasticity Mechanisms. Journal of Neuroscience, 26(13), 3561–3566.
doi:10.1523/JNEUROSCI.3154-05.2006
Barker, G. R. I., Bird, F., Alexander, V., & Warburton, E. C. (2007). Recognition
memory for objects, place, and temporal order: a disconnection analysis of the role
of the medial prefrontal cortex and perirhinal cortex. The Journal of Neuroscience :
The Official Journal of the Society for Neuroscience, 27(11), 2948–2957.
doi:10.1523/JNEUROSCI.5289-06.2007
Barker, G. R. I., & Warburton, E. C. (2011). When is the hippocampus involved in
recognition memory? The Journal of Neuroscience : The Official Journal of the
Society for Neuroscience, 31(29), 10721–31. doi:10.1523/JNEUROSCI.6413-
10.2011
Bartko, S. J., Winters, B. D., Cowell, R. A., Saksida, L. M., & Bussey, T. J. (2007).
recognition and perceptual oddity tasks Perirhinal cortex resolves feature ambiguity
in configural object recognition and perceptual oddity tasks, 821–832.
doi:10.1101/lm.749207
Bartko, S. J., Winters, B. D., Saksida, L. M., & Bussey, T. J. (2014). Different roles for
M1 and M2 receptors within perirhinal cortex in object recognition and
discrimination. Neurobiology of Learning and Memory, 110, 16–26.
doi:10.1016/j.nlm.2014.01.002
Bartus, R. (2000). On neurodegenerative diseases, models, and treatment strategies:
lessons learned and lessons forgotten a generation following the cholinergic
hypothesis. Experimental Neurology, 163, 495–529. doi:10.1006/exnr.2000.7397
Bartus, R., Iii, R., Beer, B., & Lippa, A. (1982). The Cholinergic Hypothesis of Geriatric

62
Memory Dysfunction. Science, 217(4558), 408–417.
Bartzokis, G. (2004). Age-related myelin breakdown: A developmental model of
cognitive decline and Alzheimer’s disease. Neurobiology of Aging, 25(1), 5–18.
doi:10.1016/j.neurobiolaging.2003.03.001
Bashir, Z. I. (2003). On long-term depression induced by activation of G-protein coupled
receptors. Neuroscience Research, 45(4), 363–367. doi:10.1016/S0168-
0102(03)00002-6
Becerril-Ortega, J., Bordji, K., Fréret, T., Rush, T., & Buisson, A. (2014). Iron overload
accelerates neuronal amyloid-β production and cognitive impairment in transgenic
mice model of Alzheimer’s disease. Neurobiology of Aging, 35(10), 2288–301.
doi:10.1016/j.neurobiolaging.2014.04.019
Becker, J. T., Wess, J., Hunkin, N. M., & Parkin, A. J. (1993). Use of temporal context
information in Azheimer’s disease. Neuropsychologia. doi:10.1016/0028-
3932(93)90042-X
Beggiato, S., Giuliani, a, Sivilia, S., Lorenzini, L., Antonelli, T., Imbimbo, B. P., …
Ferraro, L. (2014). CHF5074 and LY450139 sub-acute treatments differently affect
cortical extracellular glutamate levels in pre-plaque Tg2576 mice. Neuroscience,
266, 13–22. doi:10.1016/j.neuroscience.2014.01.065
Berbée, J. F. P., Vanmierlo, T., Abildayeva, K., Blokland, A., Jansen, P. J., Lütjohann,
D., … Mulder, M. (2011). Apolipoprotein CI knock-out mice display impaired
memory functions. Journal of Alzheimer’s Disease : JAD, 23(4), 737–47.
doi:10.3233/JAD-2010-100576
Bergin, D. H., & Liu, P. (2010). Agmatine protects against beta-amyloid25-35-induced

63
memory impairments in the rat. Neuroscience, 169(2), 794–811.
doi:10.1016/j.neuroscience.2010.05.004
Besheer, J., & Bevins, R. A. (2000). The role of environmental familiarization in novel-
object preference. Behavioural Processes, 50(1), 19–29. doi:10.1016/S0376-
6357(00)00090-5
Bezprozvanny, I., & Mattson, M. M. P. (2008). Neuronal calcium mishandling and the
pathogenesis of Alzheimer’s disease. Trends in Neurosciences, 31(9), 454–463.
doi:10.1016/j.tins.2008.06.005.Neuronal
Bhattacharya, S., Haertel, C., Maelicke, A., & Montag, D. (2014). Galantamine slows
down plaque formation and behavioral decline in the 5XFAD mouse model of
Alzheimer’s disease. PLoS ONE, 9(2), 1–12. doi:10.1371/journal.pone.0089454
Bilkei-Gorzo, A. (2014). Genetic mouse models of brain ageing and Alzheimer’s disease.
Pharmacology & Therapeutics, 142(2), 244–57.
doi:10.1016/j.pharmthera.2013.12.009
Billings, L. M., Green, K. N., McGaugh, J. L., & LaFerla, F. M. (2007). Learning
Decreases A *56 and Tau Pathology and Ameliorates Behavioral Decline in 3xTg-
AD Mice. Journal of Neuroscience, 27(4), 751–761.
doi:10.1523/JNEUROSCI.4800-06.2007
Billings, L., Oddo, S., Green, K., McGaugh, J., & LaFerla, F. (2005). Intraneuronal Abeta
causes the onset of early Alzheimer’s disease-related cognitive deficits in transgenic
mice. Neuron, 45(5), 675–88. doi:10.1016/j.neuron.2005.01.040
Birks, J. (2006). Cholinesterase inhibitors for Alzheimer’s disease. The Cochrane
Library, (1). doi:10.1002/14651858.CD005593.

64
Blackwell, Andrew D; Sahakian, Barbara J; Vesey, Robyn; Semple, James M; Robbins,
T. W. et al. (2003). Detecting Dementia: Novel Neuropsychological Markers of
Preclinical Alzheimer’s Disease - ProQuest. Dementia and Geriatric Cognitive
Disorders, 35–41.
Blanchard, J., Wanka, L., Tung, Y.-C., Cárdenas-Aguayo, M. D. C., LaFerla, F. M.,
Iqbal, K., & Grundke-Iqbal, I. (2010). Pharmacologic reversal of neurogenic and
neuroplastic abnormalities and cognitive impairments without affecting Aβ and tau
pathologies in 3xTg-AD mice. Acta Neuropathologica, 120(5), 605–21.
doi:10.1007/s00401-010-0734-6
Bollen, E., Vanmierlo, T., Akkerman, S., Wouters, C., Steinbusch, H. M. W., &
Prickaerts, J. (2013). 7,8-Dihydroxyflavone improves memory consolidation
processes in rats and mice. Behavioural Brain Research, 257, 8–12.
doi:10.1016/j.bbr.2013.09.029
Bongers, G., Leurs, R., Robertson, J., & Raber, J. (2004). Role of H3-receptor-mediated
signaling in anxiety and cognition in wild-type and Apoe-/- mice.
Neuropsychopharmacology : Official Publication of the American College of
Neuropsychopharmacology, 29(3), 441–9. doi:10.1038/sj.npp.1300352
Bossy-Wetzel, E., Schwarzenbacher, R., & Lipton, S. a. (2004). Molecular pathways to
neurodegeneration. Nature Medicine, 10, S2–S9. doi:10.1038/nm1067
Bouras, C., Hof, P. R., Giannakopoulos, P., Michel, J. P., & Morrison, J. H. (1994).
Regional distribution of neurofibrillary tangles and senile plaques in the cerebral
cortex of elderly patients: a quantitative evaluation of a one-year autopsy population
from a geriatric hospital. Cerebral Cortex, 4(2), 138–150.

65
doi:10.1093/cercor/4.2.138
Bowen, D. M., Smith, C. B., White, P., & Davison, a N. (1976). Neurotransmitter-related
enzymes and indices of hypoxia in senile dementia and other abiotrophies. Brain : A
Journal of Neurology, 99(3), 459–96. doi:10.1093/brain/99.3.459
Braak, H., & Braak, E. (1991). Neuropathological stageing of Alzheimer-related changes.
Acta Neuropathologica, 82(4), 239–259. doi:10.1007/BF00308809
Braak, H., & Braak, E. (1995). Staging of Alzheimer’s disease-related neurofibrillary
changes. Neurobiology of Aging, 16(3), 271–278. doi:10.1016/0197-4580(95)00021-
6
Brownlow, M. L., Joly-Amado, A., Azam, S., Elza, M., Selenica, M.-L., Pappas, C., …
Morgan, D. (2014). Partial rescue of memory deficits induced by calorie restriction
in a mouse model of tau deposition. Behavioural Brain Research, 271, 79–88.
doi:10.1016/j.bbr.2014.06.001
Bucholtz, N., & Demuth, I. (2013). DNA-repair in mild cognitive impairment and
Alzheimer’s disease. DNA Repair, 12(10), 811–816.
doi:10.1016/j.dnarep.2013.07.005
Busser, J., Geldmacher, D. S., & Herrup, K. (1998). Ectopic Cell Cycle Proteins Predict
the Sites of Neuronal Cell Death in Alzheimer’s Disease Brain. J. Neurosci., 18(8),
2801–2807.
Cameron, B., & Landreth, G. E. (2010). Inflammation, Microglia and AD. Neurobiology
of Disease, 37(3), 503–509. doi:10.1016/j.nbd.2009.10.006.Inflammation
Carroll, J. C., & Pike, C. J. (2008). Selective estrogen receptor modulators differentially
regulate Alzheimer- like changes in female 3xTg-AD mice. Endocrinology, 149(5),

66
2607–2611. doi:10.1210/en.2007-1346
Carroll, J. C., Rosario, E. R., Chang, L., Stanczyk, F. Z., Oddo, S., LaFerla, F. M., &
Pike, C. J. (2007). Progesterone and estrogen regulate Alzheimer- like
neuropathology in female 3xTg-AD mice. The Journal of Neuroscience, 27(48),
13357–13365. doi:10.1523/JNEUROSCI.2718-07.2007
Carroll, J. C., Rosario, E. R., Kreimer, S., Villamagna, A., Gentzschein, E., Stanczyk, F.
Z., & Pike, C. J. (2010). Sex differences in B-amyloid accumulation in 3xTg-AD
mice: Role of neonatal sex steroid hormone exposure. Brain Research, 1366, 233–
245. doi:10.1016/j.brainres.2010.10.009
Caterini, F., Sala, S. Della, Spinnler, H., Stangalino, C., & Turnbull, O. H. (2002). Object
recognition and object orientation in Alzheimer’s disease. Neuropsychology, 16(2),
146–155. doi:10.1037//0894-4105.16.2.146
Chapman, P. F., White, G. L., Jones, M. W., Cooper-Blacketer, D., Marshall, V. J.,
Irizarry, M., … Hsiao, K. K. (1999). Impaired synaptic plasticity and learning in
aged amyloid precursor protein transgenic mice. Nature Neuroscience, 2(3), 271–
276. doi:10.1038/6374
Chen, G., Chen, K. S., Knox, J. H., Inglis, J., Bernard, A., Martin, S. J., … Morris, R. G.
M. (2000). A learning deficit related to age and b-amyloid plaques in a mouse model
of Alzheimer’s disease. Nature, 408(1998), 975–979. doi:10.1038/35050103
Chen, G., Chen, K. S., Knox, J., Inglis, J., Bernard, A., Martin, S. J., … Morris, R. G. M.
(2000). A learning deficit related to age and b-amyloid plaques in a mouse model of
Alzheimer ’ s disease, 2940(1998), 975–979.
Chen, X., Li, M., Wang, S., Zhu, H., Xiong, Y., & Liu, X. (2014). Pittsburgh compound

67
B retention and progression of cognitive status - a meta-analysis. European Journal
of Neurology, 21(8), 1060–1067. doi:10.1111/ene.12398
Chen, Y., Liang, Z., Tian, Z., Blanchard, J., Dai, C., Chalbot, S., … Gong, C. (2014).
intracerebroventricular streptozotocin exacerbates Alzheimer-like changes of 3xTg-
AD mice. Molecular Neurobiology, 49(1), 547–62. doi:10.1007/s12035-013-8539-y
Cheng, D., Low, J. K., Logge, W., Garner, B., & Karl, T. (2014a). Chronic cannabidiol
treatment improves social and object recognition in double transgenic
APPswe/PS1∆E9 mice. Psychopharmacology, 231(15), 3009–17.
doi:10.1007/s00213-014-3478-5
Cheng, D., Low, J. K., Logge, W., Garner, B., & Karl, T. (2014b). Novel behavioural
characteristics of female APPSwe/PS1ΔE9 double transgenic mice. Behavioural
Brain Research, 260, 111–8. doi:10.1016/j.bbr.2013.11.046
Cherry, J. D., Liu, B., Frost, J. L., Lemere, C. a, Williams, J. P., Olschowka, J. a, &
O’Banion, M. K. (2012). Galactic cosmic radiation leads to cognitive impairment
and increased aβ plaque accumulation in a mouse model of Alzheimer’s disease.
PloS One, 7(12), e53275. doi:10.1371/journal.pone.0053275
Cho, S.-H., Sun, B., Zhou, Y., Kauppinen, T. M., Halabisky, B., Wes, P., … Gan, L.
(2011). CX3CR1 protein signaling modulates microglial activation and protects
against plaque-independent cognitive deficits in a mouse model of Alzheimer
disease. The Journal of Biological Chemistry, 286(37), 32713–22.
doi:10.1074/jbc.M111.254268
Clark, R. E., Zola, S. M., & Squire, L. R. (2000). Impaired recognition memory in rats
after damage to the hippocampus. The Journal of Neuroscience, 20(23), 8853–8860.

68
doi:20/23/8853 [pii]
Clinton, L. K., Billings, L. M., Green, K. N., Caccamo, A., Ngo, J., Oddo, S., … LaFerla,
F. M. (2007). Age-dependent sexual dimorphism in cognition and stress response in
the 3xTg-AD mice. Neurobiology of Disease, 28, 76–82.
doi:10.1016/j.nbd.2007.06.013
Cloke, J. M., Jacklin, D. L., & Winters, B. D. (2014). The neural bases of crossmodal
object recognition in non-human primates and rodents: A review. Behavioural Brain
Research, 1–13. doi:10.1016/j.bbr.2014.09.039
Cohen, S., Munchow, A., Rios, L., Zhang, G., Ásgeirsdóttir, H., & Stackman, R. W.
(2013). The rodent hippocampus is essential for non-spatial object memory. Curr
Biol, 23(17), 1685–1690. doi:10.1016/j.biotechadv.2011.08.021.
Corder, E. H., Ghebremedhin, E., Taylor, M. G., Thal, Dietmar, R., Ohm, T. G., & Braak,
H. (2004). The Biphasic Relationship between Regional Brain Senile Plaque and
Neurofibrillary Tangle Distributions: Modification by Age, Sex, and APOE
Polymorphism. Annals of the New York Academy of Sciences, 1019, 24–28.
Cowell, R. A., Bussey, T. J., & Saksida, L. M. (2006). Why does brain damage impair
memory? A connectionist model of object recognition memory in perirhinal cortex.
The Journal of Neuroscience, 26(47), 12186–12197.
doi:10.1523/JNEUROSCI.2818-06.2006
Cowell, R. A., Bussey, T. J., & Saksida, L. M. (2010). Components of recognition
memory: Dissociable cognitive processes or just differences in representational
complexity? Hippocampus, 20(11), 1245–1262. doi:10.1002/hipo.20865
Cuello, a C. (2005). Intracellular and extracellular Abeta, a tale of two neuropathologies.

69
Brain Pathology (Zurich, Switzerland), 15(1), 66–71.
Cummings, J. L. (2003). Use of Cholinesterase Inhibitors in Clinical Practice: Evidence-
Based Recommendations. The American Journal of Geriatric Psychiatry, 11(2),
131–145. doi:10.1097/00019442-200303000-00004
D’Antona, G., Ragni, M., Cardile, A., Tedesco, L., Dossena, M., Bruttini, F., … Nisoli,
E. (2010). Branched-chain amino acid supplementation promotes survival and
supports cardiac and skeletal muscle mitochondrial biogenesis in middle-aged mice.
Cell Metabolism, 12(4), 362–372. doi:10.1016/j.cmet.2010.08.016
Da Cruz, M. T. G., Jordo, J., Da Silva, K. a., Ayala-Grossoa, C. a., Ypsilantia, A., Weng,
Y. Q., … Auberta, I. (2012). Early increases in soluble amyloid-β levels coincide
with cholinergic degeneration in 3xTg-AD mice. Journal of Alzheimer’s Disease,
32(2), 267–272. doi:10.3233/JAD-2012-100732
Davidson, P. S. R., & Glisky, E. L. (2002). Neuropsychological correlates of recollection
and familiarity in normal aging. Cognitive, Affective & Behavioral Neuroscience,
2(2), 174–186. doi:10.3758/CABN.2.2.174
Davies, P., & Maloney, A. J. F. (1976). Selective Loss of Central Cholinergic Neurons in
Alzheimer’S Disease. The Lancet, 308(8000), 1403. doi:10.1016/S0140-
6736(76)91936-X
Davis, K. E., Easton, A., Eacott, M. J., & Gigg, J. (2013). Episodic-like memory for
what-where-which occasion is selectively impaired in the 3xTgAD mouse model of
Alzheimer’s disease. Journal of Alzheimer’s Disease : JAD, 33(3), 681–98.
doi:10.3233/JAD-2012-121543
Davis, K., Eacott, M., Easton, A., & Gigg, J. (2013). Episodic-like memory is sensitive to

70
both Alzheimer’s-like pathological accumulation and normal ageing processes in
mice. Behavioural Brain Research, 254, 73–82. doi:10.1016/j.bbr.2013.03.009
Davis, K., Mohs, R., Marin, D., Purohit, D., Perl, D., Lantz, M., … Haroutunian, V.
(1999). Cholinergic markers in elderly patients with early signs of Alzheimer
disease. Journal of American Medical Association, 281(15), 1401–1406.
Dawkins, E., & Small, D. H. (2014). Insights into the physiological function of the β-
amyloid precursor protein: Beyond Alzheimer’s disease. Journal of Neurochemistry,
129(5), 756–769. doi:10.1111/jnc.12675
De Castro, B. M., Pereira, G. S., Magalh??es, V., Rossato, J. I., De Jaeger, X., Martins-
Silva, C., … Prado, M. A. M. (2009). Reduced expression of the vesicular
acetylcholine transporter causes learning deficits in mice. Genes, Brain and
Behavior, 8(1), 23–35. doi:10.1111/j.1601-183X.2008.00439.x
de LaCoste, M.-C., & White Iii, C. L. (1993). The role of cortical connectivity in
Alzheimer’s disease pathogenesis: A review and model system. Neurobiology of
Aging, 14, 1–16. doi:http://dx.doi.org/10.1016/0197-4580(93)90015-4
Dekosky, S. T., Ikonomovic, M. D., Styren, S. D., Beckett, L., Wisniewski, S., Bennett,
D. A., … Mufson, E. J. (2002). Upregulation of choline acetyltransferase activity in
hippocampus and frontal cortex of elderly subjects with mild cognitive impairment.
Annals of Neurology, 51(2), 145–155. doi:10.1002/ana.10069
Delatour, B., Blanchard, V., Pradier, L., & Duyckaerts, C. (2004). Alzheimer pathology
disorganizes cortico-cortical circuitry: Direct evidence from a transgenic animal
model. Neurobiology of Disease, 16(1), 41–47. doi:10.1016/j.nbd.2004.01.008
Delbeuck, X., Collette, F., & Van der Linden, M. (2007). Is Alzheimer’s disease a

71
disconnection syndrome?. Evidence from a crossmodal audio-visual illusory
experiment. Neuropsychologia, 45(14), 3315–3323.
doi:10.1016/j.neuropsychologia.2007.05.001
Delbeuck, X., Van der Linden, M., & Collette, F. (2010). Alzheimer’s disease as a
disconnection syndrome. Neuropsychology Review, 20(2), 191–208.
doi:10.1007/s11065-010-9128-8
Delis, D. C., Massman, P. J., Butters, N., Salmon, D. P., Cermak, L. S., & Kramer, J. H.
(1991). Profiles of demented and amnesic patients on teh California Verbal Learning
Test: Implications for the assessment of memory disorders. Psychological
Assessment: A Journal of Consulting and Clinincal Psychology, 3(1), 19–26.
Demuro, A., Parker, I., & Stutzmann, G. E. (2010). Calcium signaling and amyloid
toxicity in Alzheimer disease. Journal of Biological Chemistry, 285(17), 12463–
12468. doi:10.1074/jbc.R109.080895
Dere, E., Huston, J. P., & De Souza Silva, M. a. (2007). The pharmacology,
neuroanatomy and neurogenetics of one-trial object recognition in rodents.
Neuroscience and Biobehavioral Reviews, 31(5), 673–704.
doi:10.1016/j.neubiorev.2007.01.005
Desai, M. K., Sudol, K. L., Janelsins, M. C., Mastrangelo, M. A., Frazer, M. E., &
Bowers, W. J. (2009). Triple-transgenic Alzheimer’s disease mice exhibit region-
specific abnormalities in brain myelination patterns prior to appearance of amyloid
and tau pathology. Glia, 57(1), 54–65. doi:10.1002/glia.20734
Devi, L., & Ohno, M. (2010). Phospho-eIF2α level is important for determining abilities
of BACE1 reduction to rescue cholinergic neurodegeneration and memory defects in

72
5XFAD mice. PLoS ONE, 5(9). doi:10.1371/journal.pone.0012974
DeVito, L. M., & Eichenbaum, H. (2010). Distinct contributions of the hippocampus and
medial prefrontal cortex to the “what-where-when” components of episodic-like
memory in mice. Behavioural Brain Research, 215(2), 318–325.
doi:10.1016/j.bbr.2009.09.014
Dewachter, I., Reverse, D., Caluwaerts, N., Ris, L., Kuipe, C., Haute, C. Van Den, …
Leuven, F. Van. (2002). Neuronal Deficiency of Presenilin 1 Inhibits Amyloid
Plaque Formation and Corrects Hippocampal Long-Term Potentiation But Not a
Cognitive Defect of Amyloid Precursor Protein [ V717I ] Transgenic Mice, 22(9),
3445–3453.
Didic, M., Felician, O., Barbeau, E. J., Mancini, J., Latger-Florence, C., Tramoni, E., &
Ceccaldi, M. (2013). Impaired visual recognition memory predicts Alzheimer’s
disease in amnestic mild cognitive impairment. Dementia and Geriatric Cognitive
Disorders, 35(5-6), 291–299. doi:10.1159/000347203
Didic, M., Ranjeva, J., Barbeau, E., Confort-gouny, S., & Le, Y. (2010). Impaired Visual
Recognition Memory in Amnestic Mild Cognitive Impairment is Associated with
Mesiotemporal Metabolic Changes on Magnetic Resonance Spectroscopic Imaging,
22, 1269–1279. doi:10.3233/JAD-2010-101257
Dix, S. L., & Aggleton, J. P. (1999). Extending the spontaneous preference test of
recognition: Evidence of object-location and object-context recognition.
Behavioural Brain Research, 99(2), 191–200. doi:10.1016/S0166-4328(98)00079-5
Dobarro, M., Gerenu, G., & Ramírez, M. J. (2013). Propranolol reduces cognitive
deficits, amyloid and tau pathology in Alzheimer’s transgenic mice. The

73
International Journal of Neuropsychopharmacology / Official Scientific Journal of
the Collegium Internationale Neuropsychopharmacologicum (CINP), 16(10), 2245–
57. doi:10.1017/S1461145713000631
Dodart, J., Meziane, H., Mathis, C., Ungerer, A., Pasteur, U. L., Bales, K. R., …
Foundation, E. L. (1999). Behavioral Disturbances in Transgenic Mice
Overexpressing the V717F p-Amyloid Precursor Protein, 113(5), 982–990.
Dodart, J.-C., Bales, K. R., Gannon, K. S., Greene, S. J., DeMattos, R. B., Mathis, C., …
Paul, S. M. (2002). Immunization reverses memory deficits without reducing brain
Abeta burden in Alzheimer’s disease model. Nature Neuroscience, 5(5), 452–7.
doi:10.1038/nn842
Dodel, R., Balakrishnan, K., Keyvani, K., Deuster, O., Neff, F., Andrei-Selmer, L.-C., …
Bacher, M. (2011). Naturally occurring autoantibodies against beta-amyloid:
investigating their role in transgenic animal and in vitro models of Alzheimer’s
disease. The Journal of Neuroscience, 31(15), 5847–54.
doi:10.1523/JNEUROSCI.4401-10.2011
Donkin, J. J., Stukas, S., Hirsch-Reinshagen, V., Namjoshi, D., Wilkinson, A., May, S.,
… Wellington, C. L. (2010). ATP-binding cassette transporter A1 mediates the
beneficial effects of the liver X receptor agonist GW3965 on object recognition
memory and amyloid burden in amyloid precursor protein/presenilin 1 mice. The
Journal of Biological Chemistry, 285(44), 34144–54. doi:10.1074/jbc.M110.108100
Drzezga, A., Grimmer, T., Peller, M., Wermke, M., Siebner, H., Rauschecker, J. P., …
Kurz, A. (2005). Impaired cross-modal inhibition in Alzheimer disease. PLoS
Medicine, 2(10), e288. doi:10.1371/journal.pmed.0020288

74
Duyckaerts, C., Delatour, B., & Potier, M. C. (2009). Classification and basic pathology
of Alzheimer disease. Acta Neuropathologica. doi:10.1007/s00401-009-0532-1
Ennaceur, A., & Delacour, J. (1988). A new one-trial test for neurobiological studies of
memory in rats. 1: Behavioral data. Behavioural Brain Research, 31(1), 47–59.
doi:10.1016/S0166-4328(05)80315-8
Ennaceur, A., Neave, N., & Aggleton, J. (1997). Spontaneous object recognition and
object location memory in rats: The effects of lesions in the cingulate cortices, the
medial prefrontal cortex, the cingulum bundle and the fornix. Experimental Brain
Research, 113(3), 509–519. doi:10.1007/PL00005603
Escribano, L., Simón, A.-M., Pérez-Mediavilla, A., Salazar-Colocho, P., Del Río, J., &
Frechilla, D. (2009). Rosiglitazone reverses memory decline and hippocampal
glucocorticoid receptor down-regulation in an Alzheimer’s disease mouse model.
Biochemical and Biophysical Research Communications, 379(2), 406–10.
doi:10.1016/j.bbrc.2008.12.071
Espana, J., Gimenez-Llort, L., Valero, J., Minano, A., Rabano, A., Rodriguez-Alvarez, J.,
… Saura, C. A. (2010). Intraneuronal B-Amyloid Accumulation in the Amygdala
Enhances Fear and Anxiety in Alzheimer’s Disease Transgenic Mice. Biological
Psychiatry, 67(6), 513–521. doi:10.1016/j.biopsych.2009.06.015
Fadda, F., Melis, F., & Stancampiano, R. (1996). Increased hippocampal acetylcholine
release during a working memory task. European Journal of Pharmacology, 307(2),
1–2. doi:10.1016/0014-2999(96)00289-0
Farías, G., González-Billault, C., & Maccioni, R. B. (1997). Immunological
characterization of epitopes on tau of Alzheimer’s type and chemically modified tau.

75
Molecular and Cellular Biochemistry, 168(1-2), 59–66.
doi:10.1023/A:1006838626730
Farr, S. a, Erickson, M. a, Niehoff, M. L., Banks, W. a, & Morley, J. E. (2014). Central
and Peripheral Administration of Antisense Oligonucleotide Targeting Amyloid-B
Protein Precursor Improves Learning and Memory and Reduces Neuroinflammatory
Cytokines in Tg2576 (ABPPswe) Mice. Journal of Alzheimer’s Disease : JAD,
40(4), 1005–16. doi:10.3233/JAD-131883
Feld, M., Krawczyk, M. C., Sol Fustiñana, M., Blake, M. G., Baratti, C. M., Romano, A.,
& Boccia, M. M. (2014). Decrease of ERK/MAPK overactivation in prefrontal
cortex reverses early memory deficit in a mouse model of Alzheimer’s disease.
Journal of Alzheimer’s Disease : JAD, 40(1), 69–82. doi:10.3233/JAD-131076
Filali, M., Lalonde, R., Theriault, P., Julien, C., Calon, F., & Planel, E. (2012). Cognitive
and non-cognitive behaviors in the triple transgenic mouse model of Alzheimer’s
disease expressing mutated APP, PS1, and Mapt (3xTg-AD). Behavioural Brain
Research, 234(2), 334–42. doi:10.1016/j.bbr.2012.07.004
Forwood, S. E., Bartko, S. J., Saksida, L. M., & Bussey, T. J. (2007). Rats spontaneously
discriminate purely visual, two-dimensional stimuli in tests of recognition memory
and perceptual oddity. Behavioral Neuroscience, 121(5), 1032–1042.
doi:10.1037/0735-7044.121.5.1032
Forwood, S. E., Winters, B. D., & Bussey, T. J. (2005). Hippocampal lesions that abolish
spatial maze performance spare object recognition memory at delays of up 48 hours.
Hippocampus, 15(3), 347–355. doi:10.1002/hipo.20059
Fragkouli, A., Tsilibary, E. C., & Tzinia, A. K. (2014). Neuroprotective role of MMP-9

76
overexpression in the brain of Alzheimer’s 5xFAD mice. Neurobiology of Disease,
70, 179–89. doi:10.1016/j.nbd.2014.06.021
Francis, B. M., Kim, J., Barakat, M. E., Fraenkl, S., Yücel, Y. H., Peng, S., … Mount, H.
T. J. (2012). Object recognition memory and BDNF expression are reduced in
young TgCRND8 mice. Neurobiology of Aging, 33(3), 555–63.
doi:10.1016/j.neurobiolaging.2010.04.003
Francis, B. M., Yang, J., Hajderi, E., Brown, M. E., Michalski, B., McLaurin, J., …
Mount, H. T. J. (2012). Reduced tissue levels of noradrenaline are associated with
behavioral phenotypes of the TgCRND8 mouse model of Alzheimer’s disease.
Neuropsychopharmacology, 37(8), 1934–44. doi:10.1038/npp.2012.40
Francis, P., Palmer, A., Snape, M., & Wilcock, K. (1999). The cholinergic hypothesis of
Alzheimer ’ s disease : a review of progress. J Neurol Neurosurg Psychiatry, 66,
137–147.
Friedrich, R. P., Tepper, K., Rönicke, R., Soom, M., Westermann, M., Reymann, K., …
Fändrich, M. (2010). Mechanism of amyloid plaque formation suggests an
intracellular basis of Abeta pathogenicity. Proceedings of the National Academy of
Sciences of the United States of America, 107(5), 1942–1947.
doi:10.1073/pnas.0904532106
Frydman-Marom, A., Levin, A., Farfara, D., Benromano, T., Scherzer-Attali, R., Peled,
S., … Ovadia, M. (2011). Orally administrated cinnamon extract reduces B-amyloid
oligomerization and corrects cognitive impairment in Alzheimer’s disease animal
models. PLoS ONE, 6(1), 1–11. doi:10.1371/journal.pone.0016564
Frye, C. a, & Walf, A. a. (2008). Effects of progesterone administration and

77
APPswe+PSEN1Deltae9 mutation for cognitive performance of mid-aged mice.
Neurobiology of Learning and Memory, 89(1), 17–26.
doi:10.1016/j.nlm.2007.09.008
Fukumoto, H., Takahashi, H., Tarui, N., Matsui, J., Tomita, T., Hirode, M., …
Miyamoto, M. (2010). A noncompetitive BACE1 inhibitor TAK-070 ameliorates
Abeta pathology and behavioral deficits in a mouse model of Alzheimer’s disease.
The Journal of Neuroscience, 30(33), 11157–11166.
doi:10.1523/JNEUROSCI.2884-10.2010
Fukumoto, K., Mizoguchi, H., Takeuchi, H., Horiuchi, H., Kawanokuchi, J., Jin, S., …
Suzumura, A. (2014). Fingolimod increases brain-derived neurotrophic factor levels
and ameliorates amyloid β-induced memory impairment. Behavioural Brain
Research, 268, 88–93. doi:10.1016/j.bbr.2014.03.046
Galeano, P., Martino Adami, P. V, Do Carmo, S., Blanco, E., Rotondaro, C., Capani, F.,
… Morelli, L. (2014). Longitudinal analysis of the behavioral phenotype in a novel
transgenic rat model of early stages of Alzheimer’s disease.2. Frontiers in
Behavioral Neuroscience, 8, 321. doi:10.3389/fnbeh.2014.00321
Gamblin, T. C., Chen, F., Zambrano, A., Abraha, A., Lagalwar, S., Guillozet, A. L., …
Cryns, V. L. (2003). Caspase cleavage of tau: linking amyloid and neurofibrillary
tangles in Alzheimer’s disease. Proceedings of the National Academy of Sciences of
the United States of America, 100(17), 10032–10037. doi:10.1073/pnas.1630428100
García-Mesa, Y., Giménez-Llort, L., López, L. C., Venegas, C., Cristòfol, R., Escames,
G., … Sanfeliu, C. (2012). Melatonin plus physical exercise are highly
neuroprotective in the 3xTg-AD mouse. Neurobiology of Aging, 33(6), 1124.e13–

78
1124.e29. doi:10.1016/j.neurobiolaging.2011.11.016
Garcia-Osta, A., & Alberini, C. M. (2009). Amyloid beta mediates memory formation.
Learning & Memory (Cold Spring Harbor, N.Y.), 16(4), 267–72.
doi:10.1101/lm.1310209
Gaskin, S., Tardif, M., Cole, E., Piterkin, P., Kayello, L., & Mumby, D. G. (2010). Object
familiarization and novel-object preference in rats. Behavioural Processes, 83(1),
61–71. doi:10.1016/j.beproc.2009.10.003
Gerenu, G., Dobarro, M., Ramirez, M. J., & Gil-Bea, F. J. (2013). Early cognitive
sitmulation compensates for memory and pathological changes in Tg2576 mice.
Biochimica et Biophysica Acta, 1832(6), 837–47. doi:10.1016/j.bbadis.2013.02.018
Ghoshal, N., Garcia-Sierra, F., Wuu, J., Leurgans, S., Bennett, D. A., Berry, R. W., &
Binder, L. I. (2002). Tau Conformational Changes Correspond to Impairments of
Episodic Memory in Mild Cognitive Impairment and Alzheimer’s Disease.
Experimental Neurology, 177(2), 475–493. doi:10.1006/exnr.2002.8014
Giannakopoulos, P., Herrmann, F. R., Bussière, T., Bouras, C., Kövari, E., Perl, D. P., …
Hof, P. R. (2003). Tangle and neuron numbers, but not amyloid load, predict
cognitive status in Alzheimer’s disease. Neurology, 60(9), 1495–1500.
doi:10.1212/01.WNL.0000063311.58879.01
Giannoni, P., Gaven, F., de Bundel, D., Baranger, K., Marchetti-Gauthier, E., Roman, F.
S., … Claeysen, S. (2013). Early administration of RS 67333, a specific 5-HT4
receptor agonist, prevents amyloidogenesis and behavioral deficits in the 5XFAD
mouse model of Alzheimer’s disease. Frontiers in Aging Neuroscience,
5(December), 96. doi:10.3389/fnagi.2013.00096

79
Gilmor, M. L., Erickson, J. D., Varoqui, H., Hersh, L. B., Bennett, D. A., Cochran, E. J.,
… Levey, A. I. (1999). Preservation of Nucleus Basalis Neurons Containing Choline
Acetyltransferase and the Vesicular Acetylcholine Transporter in the Elderly with
Mild Cognitive Impairment and Early Alzheimer’s Disease. The Journal of
Comparative Neurology, 411, 693–704. doi:10.1016/j.neurobiolaging.2010.08.015
Giuliani, A., Beggiato, S., Baldassarro, V. a, Mangano, C., Giardino, L., Imbimbo, B. P.,
… Ferraro, L. (2013). CHF5074 restores visual memory ability and pre-synaptic
cortical acetylcholine release in pre-plaque Tg2576 mice. Journal of
Neurochemistry, 124(5), 613–20. doi:10.1111/jnc.12136
Golby, A., Silverberg, G., Race, E., Gabrieli, S., O’Shea, J., Knierim, K., … Gabrieli, J.
(2005). Memory encoding in Alzheimer’s disease: An fMRI study of explicit and
implicit memory. Brain, 128(4), 773–787. doi:10.1093/brain/awh400
Golob, E. J., Miranda, G. G., Johnson, J. K., & Starr, A. (2001). Sensory cortical
interactions in aging, mild cognitive impairment, and Alzheimer’s disease.
Neurobiology of Aging, 22(5), 755–763. doi:10.1016/S0197-4580(01)00244-5
Gong, C. X., Liu, F., Grundke-Iqbal, I., & Iqbal, K. (2006). Dysregulation of protein
phosphorylation/dephosphorylation in Alzheimer’s disease: A therapeutic target.
Journal of Biomedicine and Biotechnology, 2006, 1–11.
doi:10.1155/JBB/2006/31825
Gonzalez-Lima, F., Berndt, J. D., Valla, J. E., Games, D., & Reiman, E. M. (2001).
Reduced corpus callosum, fornix and hippocampus in PDAPP transgenic mouse
model of Alzheimer’s disease. Neuroreport, 12(11), 2375–2379.
doi:10.1097/00001756-200108080-00018

80
Good, M. a, & Hale, G. (2007). The “Swedish” mutation of the amyloid precursor protein
(APPswe) dissociates components of object-location memory in aged Tg2576 mice.
Behavioral Neuroscience, 121(6), 1180–91. doi:10.1037/0735-7044.121.6.1180
Good, M. a, Hale, G., & Staal, V. (2007). Impaired “episodic-like” object memory in
adult APPswe transgenic mice. Behavioral Neuroscience, 121(2), 443–8.
doi:10.1037/0735-7044.121.2.443
Görtz, N., Lewejohann, L., Tomm, M., Ambrée, O., Keyvani, K., Paulus, W., & Sachser,
N. (2008). Effects of environmental enrichment on exploration, anxiety, and
memory in female TgCRND8 Alzheimer mice. Behavioural Brain Research, 191(1),
43–8. doi:10.1016/j.bbr.2008.03.006
Grady, C. L., Furey, M. L., Pietrini, P., Horwitz, B., & Rapoport, S. I. (2001). Altered
brain functional connectivity and impaired short-term memory in Alzheimer’s
disease. Brain : A Journal of Neurology, 124, 739–756. doi:10.1093/brain/124.4.739
Granholm, E., & Butters, N. (1988). Associative encoding and retrieval in Alzheimer’s
and Huntington's disease. Brain and Cognition, 7(3), 335–347. doi:10.1016/0278-
2626(88)90007-3
Grayson, B., Leger, M., Piercy, C., Adamson, L., Harte, M., & Neill, J. C. (2014).
Assessment of disease-related cognitive impairments using the novel object
recognition (NOR) task in rodents. Behavioural Brain Research.
doi:10.1016/j.bbr.2014.10.025
Greco, S. J., Bryan, K. J., Sarkar, S., Zhu, X., Smith, M. a, Ashford, J. W., … Casadesus,
G. (2010). Leptin reduces pathology and improves memory in a transgenic mouse
model of Alzheimer’s disease. Journal of Alzheimer’s Disease : JAD, 19(4), 1155–

81
67. doi:10.3233/JAD-2010-1308
Green, K. N., & LaFerla, F. M. (2008). Linking Calcium to AB and Alzheimer’s Disease.
Neuron, 59(2), 190–194. doi:10.1016/j.neuron.2008.07.013
Griffin, W. S., Sheng, J. G., Royston, M. C., Gentleman, S. M., McKenzie, J. E., Graham,
D. I., … Mrak, R. E. (1998). Glial-neuronal interactions in Alzheimer’s disease: the
potential role of a “cytokine cycle” in disease progression. Brain Pathology, 8(1),
65–72. doi:10.1111/j.1750-3639.1998.tb00136.x
Guillozet, A. L., Weintraub, S., Mash, D. C., & Mesulam, M. M. (2003). Neurofibrillary
tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol,
60(5), 729–736. doi:10.1001/archneur.60.5.729
Gulinello, M., Gertner, M., Mendoza, G., Schoenfeld, B. P., Oddo, S., LaFerla, F., …
Faber, D. S. (2009). Validation of a 2-day water maze protocol in mice. Behavioural
Brain Research, 196(2), 220–7. doi:10.1016/j.bbr.2008.09.002
Guzmán-Ramos, K., Moreno-Castilla, P., Castro-Cruz, M., McGaugh, J. L., Martínez-
Coria, H., LaFerla, F. M., & Bermúdez-Rattoni, F. (2012). Restoration of dopamine
release deficits during object recognition memory acquisition attenuates cognitive
impairment in a triple transgenic mice model of Alzheimer’s disease. Learning &
Memory, 19(10), 453–60. doi:10.1101/lm.026070.112
Haass, C. (1997). Presenilins-Genes for Life and Death. Neuron, 18(5), 687–690.
Hafez, D. M., Huang, J. Y., Richardson, J. C., Masliah, E., Peterson, D. a, & Marr, R. a.
(2012). F-spondin gene transfer improves memory performance and reduces
amyloid-β levels in mice. Neuroscience, 223, 465–72.
doi:10.1016/j.neuroscience.2012.07.038

82
Hale, G., & Good, M. (2005). Impaired visuospatial recognition memory but normal
object novelty detection and relative familiarity judgments in adult mice expressing
the APPswe Alzheimer’s disease mutation. Behavioral Neuroscience, 119(4), 884–
91. doi:10.1037/0735-7044.119.4.884
Hammond, R. S., Tull, L. E., & Stackman, R. W. (2004). On the delay-dependent
involvement of the hippocampus in object recognition memory. Neurobiology of
Learning and Memory, 82(1), 26–34. doi:10.1016/j.nlm.2004.03.005
Hannesson, D. K., Howland, J. G., & Phillips, A. G. (2004). Interaction between
perirhinal and medial prefrontal cortex is required for temporal order but not
recognition memory for objects in rats. The Journal of Neuroscience, 24(19), 4596–
4604. doi:10.1523/JNEUROSCI.5517-03.2004
Hardy, J. (1997). Amyloid, the presenilins and Alzheimer’s disease. Trends in
Neurosciences, 20(4), 154–159. doi:10.1016/S0166-2236(96)01030-2
Hardy, J., & Allsop, D. (1991). Amyloid deposition as the central event in the aetiology
of Alzheimer’s disease. Trends in Pharmacological Sciences, 12(C), 383–388.
doi:10.1016/0165-6147(91)90609-V
Hardy, J., & Higgins, G. (1992). Alzheimer ’ s Disease : The Amyloid Cascade
Hypothesis Published by : American Association for the Advancement of Science
Alzheimer ’ s Disease : The Amyloid Cascade Hypothesis. Science, 256(5054), 184–
185.
Hardy, J., & Selkoe, D. J. (2002). The amyloid hypothesis of Alzheimer’s disease:
progress and problems on the road to therapeutics. Science (New York, N.Y.),
297(5580), 353–356. doi:10.1126/science.1072994

83
Harris, J. a, Devidze, N., Halabisky, B., Lo, I., Thwin, M. T., Yu, G.-Q., … Mucke, L.
(2010). Many neuronal and behavioral impairments in transgenic mouse models of
Alzheimer’s disease are independent of caspase cleavage of the amyloid precursor
protein. The Journal of Neuroscience, 30(1), 372–381.
doi:10.1523/JNEUROSCI.5341-09.2010
Heneka, M. T., Ramanathan, M., Jacobs, A. H., Dumitrescu-Ozimek, L., Bilkei-Gorzo,
A., Debeir, T., … Staufenbiel, M. (2006). Locus Ceruleus Degeneration Promotes
Alzheimer Pathogenesis in Amyloid Precursor Protein 23 Transgenic Mice. Journal
of Neuroscience, 26(5), 1343–1354. doi:10.1523/JNEUROSCI.4236-05.2006
Hernandez, C. M., Kayed, R., Zheng, H., Sweatt, J. D., & Dineley, K. T. (2010). Loss of
alpha7 nicotinic receptors enhances beta-amyloid oligomer accumulation,
exacerbating early-stage cognitive decline and septohippocampal pathology in a
mouse model of Alzheimer’s disease. The Journal of Neuroscience, 30(7), 2442–
53. doi:10.1523/JNEUROSCI.5038-09.2010
Hillen, H., Barghorn, S., Striebinger, A., Labkovsky, B., Müller, R., Nimmrich, V., …
Ebert, U. (2010). Generation and therapeutic efficacy of highly oligomer-specific
beta-amyloid antibodies. The Journal of Neuroscience, 30(31), 10369–79.
doi:10.1523/JNEUROSCI.5721-09.2010
Hindley, E. L., Nelson, A. J. D., Aggleton, J. P., & Vann, S. D. (2014). Dysgranular
retrosplenial cortex lesions in rats disrupt cross-modal object recognition. Learning
& Memory (Cold Spring Harbor, N.Y.), 21(3), 171–9. doi:10.1101/lm.032516.113
Hirata-Fukae, C., Li, H. F., Hoe, H. S., Gray, A. J., Minami, S. S., Hamada, K., …
Matsuoka, Y. (2008). Females exhibit more extensive amyloid, but not tau,

84
pathology in an Alzheimer transgenic model. Brain Research, 1216(3), 92–103.
doi:10.1016/j.brainres.2008.03.079
Hodges, J. R., Salmon, D. P., & Butters, N. (1990). Differential impairment of semantic
and episodic memory in Alzheimer’s and Huntington's diseases: a controlled
prospective study. Journal of Neurology, Neurosurgery, and Psychiatry, 53(12),
1089–95. doi:10.1136/jnnp.53.12.1089
Holmes, C., Boche, D., Wilkinson, D., Yadegarfar, G., Hopkins, V., Bayer, A., … Nicoll,
J. a R. (2008). Long-term eff ects of Aβ 42 immunisation in Alzheimer’s disease :
follow-up of a randomised, placebo-controlled phase I trial. Lancet, 372, 11–14.
doi:10.1016/S0140-6736(08)61075-2
Honda, K., & Casadesus, G. (2004). Oxidative Stress and Redox Active Iron in
Alzheimer’s Disease. Annals of the New York Academy of Sciences, 1012, 179–182.
Retrieved from http://onlinelibrary.wiley.com/doi/10.1196/annals.1306.015/full
Howlett, D. R., Richardson, J. C., Austin, A., Parsons, A. a, Bate, S. T., Davies, D. C., &
Gonzalez, M. I. (2004). Cognitive correlates of Abeta deposition in male and female
mice bearing amyloid precursor protein and presenilin-1 mutant transgenes. Brain
Research, 1017(1-2), 130–6. doi:10.1016/j.brainres.2004.05.029
Hsai, A. Y., Masliah, E., McConlogue, L., Yu, G.-Q., Tatsuno, G., Hu, K., … Mucke, L.
(1999). Plaque-independent disruption of neural circuits in Alzheimer’s disease
mouse models. Proceedings of the National Academy of Sciences of the United
States of America, 96(6), 3228–33. doi:10.1073/pnas.96.6.3228
Hsiao, K. K., Borchelt, D. R., Olson, K., Johannsdottir, R., Kitt, C., Yunis, W., …
Carlson, G. (1995). Age-related CNS disorder and early death in transgenic FVB/N

85
mice overexpressing Alzheimer amyloid precursor proteins. Neuron, 15(5), 1203–
1218. doi:10.1016/0896-6273(95)90107-8
Huang, S.-M., Mouri, A., Kokubo, H., Nakajima, R., Suemoto, T., Higuchi, M., … Iwata,
N. (2006). Neprilysin-sensitive synapse-associated amyloid-beta peptide oligomers
impair neuronal plasticity and cognitive function. The Journal of Biological
Chemistry, 281(26), 17941–51. doi:10.1074/jbc.M601372200
Huang, X. (2003). Behavioral and neurochemical characterization of transgenic mice
carrying the human presenilin-1 gene with or without the leucine-to-proline
mutation at codon 235. Experimental Neurology, 183(2), 673–681.
doi:10.1016/S0014-4886(03)00242-5
Hyman, B., Hoesen, G., Damasio, A., & Clifford, L. (1984). Alzheimer’s Disease : Cell-
Specific Pathology Isolates the Hippocampal Formation Published by : American
Association for the Advancement of Science Stable URL :
http://www.jstor.org/stable/1693749 . Science, 225(4667), 1168–1170.
Hyman, B. T., Van Hoesen, G. W., Kromer, L. J., & Damasio, a R. (1986). Perforant
pathway changes and the memory impairment of Alzheimer’s disease. Annals of
Neurology, 20(4), 472–81. doi:10.1002/ana.410200406
Iversen, L. L., Mortishire-Smith, R. J., Pollack, S. J., & Shearman, M. S. (1995). The
toxicity in vitro of beta-amyloid protein. Biochemical Journal, 311(Pt 1), 1–16.
Izquierdo, I., Medina, J. H., Vianna, M. R. M., Izquierdo, L. A., & Barros, D. M. (1999).
Separate mechanisms for short- and long-term memory. Behavioural Brain
Research, 103(1), 1–11. doi:10.1016/S0166-4328(99)00036-4
Jacklin, D. L., Cloke, J. M., Potvin, A., Garrett, I., & Winters, B. D. (2016). The

86
Dynamic Multisensory Engram: Neural Circuitry Underlying Crossmodal Object
Recognition in Rats Changes with the Nature of Object Experience. Journal of
Neuroscience, 36(4), 1273–1289. doi:10.1523/JNEUROSCI.3043-15.2016
Jacobsen, J. S., Wu, C.-C., Redwine, J. M., Comery, T. A., Arias, R., Bowlby, M., …
Bloom, F. E. (2006). Early-onset behavioral and synaptic deficits in a mouse model
of Alzheimer’s disease. Proceedings of the National Academy of Sciences of the
United States of America, 103(13), 5161–6. doi:10.1073/pnas.0600948103
Janelsins, M. C., Mastrangelo, M. a, Oddo, S., LaFerla, F. M., Federoff, H. J., & Bowers,
W. J. (2005). Early correlation of microglial activation with enhanced tumor
necrosis factor-alpha and monocyte chemoattractant protein-1 expression specif-
ically within the entorhinal cortex of triple transgenic Alzheimer’s disease mice.
Journal of Neuroinflammation, 2(1), 23. doi:10.1186/1742-2094-2-23
Jardanhazi-Kurutz, D., Kummer, M. P., Terwel, D., Vogel, K., Dyrks, T., Thiele, A., &
Heneka, M. T. (2010). Induced LC degeneration in APP/PS1 transgenic mice
accelerates early cerebral amyloidosis and cognitive deficits. Neurochemistry
International, 57(4), 375–82. doi:10.1016/j.neuint.2010.02.001
Jaszberenyi, M., Rick, F. G., Szalontay, L., Block, N. L., Zarandi, M., Cai, R.-Z., &
Schally, A. V. (2012). Beneficial effects of novel antagonists of GHRH in different
models of Alzheimer’s disease. Aging-Us, 4(11), 755–767. Retrieved from <Go to
ISI>://WOS:000312289900008
Jawhar, S., Trawicka, A., Jenneckens, C., Bayer, T. a., & Wirths, O. (2012). Motor
deficits, neuron loss, and reduced anxiety coinciding with axonal degeneration and
intraneuronal AB aggregation in the 5XFAD mouse model of Alzheimer’s disease.

87
Neurobiology of Aging, 33(1), 196.e29–196.e40.
doi:10.1016/j.neurobiolaging.2010.05.027
Jin, G., Wang, L.-H., Ji, X.-F., Chi, T.-Y., Qi, Y., Jiao, Q., … Zou, L.-B. (2014).
Xanthoceraside rescues learning and memory deficits through attenuating beta-
amyloid deposition and tau hyperphosphorylation in APP mice. Neuroscience
Letters, 573, 58–63. doi:10.1016/j.neulet.2014.04.032
Jing, J., Jun, Y., Jung, S., Keun, T., Ashrafu, K., Chang, H., & Hong, W. (2013).
Protective Effects of Ferulic Acid in Amyloid Precursor Protein Plus Presenilin - 1
Transgenic Mouse Model of Alzheimer Disease, 36(1), 140–143.
Joyashiki, E., Matsuya, Y., & Tohda, C. (2011). Sominone improves memory
impairments and increases axonal density in Alzheimer’s disease model mice,
5XFAD. The International Journal of Neuroscience, 121(4), 181–90.
doi:10.3109/00207454.2010.541571
Kalinin, S., Polak, P. E., Lin, S. X., Sakharkar, A. J., Pandey, S. C., & Feinstein, D. L.
(2012). The noradrenaline precursor L-DOPS reduces pathology in a mouse model
of Alzheimer’s disease. Neurobiology of Aging, 33(8), 1651–1663.
doi:10.1016/j.neurobiolaging.2011.04.012
Kanno, T., Tsuchiya, A., & Nishizaki, T. (2014a). Hyperphosphorylation of Tau at
Ser396 occurs in the much earlier stage than appearance of learning and memory
disorders in 5XFAD mice. Behavioural Brain Research, 274, 302–306.
doi:10.1016/j.bbr.2014.08.034
Kanno, T., Tsuchiya, A., & Nishizaki, T. (2014b). Hyperphosphorylation of Tau at
Ser396 occurs in the much earlier stage than appearance of learning and memory

88
disorders in 5XFAD mice. Behavioural Brain Research, 274C, 302–306.
doi:10.1016/j.bbr.2014.08.034
Kariv-Inbal, Z., Yacobson, S., Berkecz, R., Peter, M., Janaky, T., Lütjohann, D., …
Michaelson, D. M. (2012). The isoform-specific pathological effects of apoE4 in
vivo are prevented by a fish oil (DHA) diet and are modified by cholesterol. Journal
of Alzheimer’s Disease : JAD, 28(3), 667–83. doi:10.3233/JAD-2011-111265
Karl, T., Bhatia, S., Cheng, D., Kim, W. S., & Garner, B. (2012). Cognitive phenotyping
of amyloid precursor protein transgenic J20 mice. Behavioural Brain Research,
228(2), 392–7. doi:10.1016/j.bbr.2011.12.021
Kastyak-Ibrahim, M. Z., Di Curzio, D. L., Buist, R., Herrera, S. L., Albensi, B. C., Del
Bigio, M. R., & Martin, M. (2013). Neurofibrillary tangles and plaques are not
accompanied by white matter pathology in aged triple transgenic-Alzheimer disease
mice. Magnetic Resonance Imaging, 31(9), 1515–1521.
doi:10.1016/j.mri.2013.06.013
Kauppinen, T. M., Suh, S. W., Higashi, Y., Berman, A. E., Escartin, C., Won, S. J., …
Swanson, R. a. (2011). Poly(ADP-ribose)polymerase-1 modulates microglial
responses to amyloid β. Journal of Neuroinflammation, 8(1), 152.
doi:10.1186/1742-2094-8-152
Kazim, S. F., Blanchard, J., Dai, C.-L., Tung, Y.-C., LaFerla, F. M., Iqbal, I.-G., & Iqbal,
K. (2014). Disease modifying effect of chronic oral treatment with a neurotrophic
peptidergic compound in a triple transgenic mouse model of Alzheimer’s disease.
Neurobiology of Disease, 71, 110–30. doi:10.1016/j.nbd.2014.07.001
Kim, J., Chakrabarty, P., Hanna, A., March, A., Dickson, D. W., Borchelt, D. R., …

89
Janus, C. (2013). Normal cognition in transgenic BRI2-Aβ mice. Molecular
Neurodegeneration, 8, 15. doi:10.1186/1750-1326-8-15
Kimura, R., & Ohno, M. (2009). Impairments in remote memory stabilization precede
hippocampal synaptic and cognitive failures in 5XFAD Alzheimer mouse model.
Neurobiology of Disease, 33(2), 229–235. doi:10.1016/j.nbd.2008.10.006
Knowles, J. K., Simmons, D. a, Nguyen, T.-V. V, Vander Griend, L., Xie, Y., Zhang, H.,
… Longo, F. M. (2013). Small molecule p75NTR ligand prevents cognitive deficits
and neurite degeneration in an Alzheimer’s mouse model. Neurobiology of Aging,
34(8), 2052–63. doi:10.1016/j.neurobiolaging.2013.02.015
Kopelman, M. D. (1989). Remote and autobiographical memory, temporal context
memory and frontal atrophy in Korsakoff and Alzheimer patients.
Neuropsychologia, 27(4), 437–460. doi:10.1016/0028-3932(89)90050-X
Kornecook, T. J., McKinney, a P., Ferguson, M. T., & Dodart, J.-C. (2010). Isoform-
specific effects of apolipoprotein E on cognitive performance in targeted-
replacement mice overexpressing human APP. Genes, Brain, and Behavior, 9(2),
182–92. doi:10.1111/j.1601-183X.2009.00545.x
Kotilinek, L. a, Bacskai, B., Westerman, M., Kawarabayashi, T., Younkin, L., Hyman, B.
T., … Ashe, K. H. (2002). Reversible memory loss in a mouse transgenic model of
Alzheimer’s disease. The Journal of Neuroscience, 22(15), 6331–6335.
doi:20026675
Kroker, K. S., Mathis, C., Marti, A., Cassel, J.-C., Rosenbrock, H., & Dorner-Ciossek, C.
(2014). PDE9A inhibition rescues amyloid beta-induced deficits in synaptic
plasticity and cognition. Neurobiology of Aging, 35(9), 2072–8.

90
doi:10.1016/j.neurobiolaging.2014.03.023
LaFerla, F. M., & Green, K. N. (2012). Animal models of Alzheimer disease. Cold
Spring Harb Perspect Med, 2(11), 1–13. doi:10.1101/cshperspect.a006320
Lanctôt, K. L., Herrmann, N., Yau, K. K., Khan, L. R., Liu, B. A., LouLou, M. M., &
Einarson, T. R. (2003). Efficacy and safety of cholinesterase inhibitors in
Alzheimer’s disease: a meta-analysis. CMAJ : Canadian Medical Association
Journal = Journal de l’Association Medicale Canadienne, 169(6), 557–64.
doi:za2963e q8za9 q8zb8 q8zc9 q8zd4 q8zeb q8zf5 q8zg5
Laursen, B., Mørk, A., Plath, N., Kristiansen, U., & Bastlund, J. F. (2013). Cholinergic
degeneration is associated with increased plaque deposition and cognitive
impairment in APPswe/PS1dE9 mice. Behavioural Brain Research, 240, 146–52.
doi:10.1016/j.bbr.2012.11.012
Lecanu, L., & Papadopoulos, V. (2013). Modeling Alzheimer ’ s disease with non-
transgenic rat models. Alzheimer’s Research & Therapy, 1–9.
Lee, A. C. H., Buckley, M. J., Gaffan, D., Emery, T., Hodges, J. R., & Graham, K. S.
(2006). Differentiating the roles of the hippocampus and perirhinal cortex in
processes beyond long-term declarative memory: a double dissociation in dementia.
The Journal of Neuroscience, 26(19), 5198–203. doi:10.1523/JNEUROSCI.3157-
05.2006
Lee, J. Y., Kho, S., Yoo, H. Bin, Park, S., Choi, J. S., Kwon, J. S., … Jung, H. Y. (2014).
Spatial memory impairments in amnestic mild cognitive impairment in a virtual
radial arm maze. Neuropsychiatric Disease and Treatment, 10, 653–660.
doi:10.2147/ndt.s58185

91
Lemere, C. A., & Masliah, E. (2010). Can Alzheimer disease be prevented by amyloid-B
immunotherapy? Nature Reviews Neurology, 6(2), 108–119.
doi:10.1038/nrneurol.2009.219.Can
Lesné, S., Koh, M. T., Kotilinek, L., Kayed, R., Glabe, C. G., Yang, A., … Ashe, K. H.
(2006). A specific amyloid-beta protein assembly in the brain impairs memory.
Nature, 440(7082), 352–357. doi:10.1038/nature04533
Levin-allerhand, J. A., Lominska, C. E., Wang, J., & Smith, J. D. (2002). 17α-estradiol
and 17β-estradiol treatments are effective in lowering cerebral amyloid-β levels in
AβPPSWE transgenic mice. Journal of Alzheimer’s Disease, 4, 449–457.
Li, C., Ebrahimi, A., & Schluesener, H. (2013). Drug pipeline in neurodegeneration
based on transgenic mice models of Alzheimer’s disease. Ageing Research Reviews,
12(1), 116–40. doi:10.1016/j.arr.2012.09.002
Libon, D. J., Bogdanoff, B., Cloud, B. S., Skalina, S., Giovannetti, T., Gitlin, H. L., &
Bonavita, J. (1998). Declarative and procedural learning, quantitative measures of
the hippocampus, and subcortical white alterations in Alzheimer’s disease and
ischaemic vascular dementia. Journal of Clinical and Experimental
Neuropsychology, 20(1), 30–41. doi:10.1076/jcen.20.1.30.1490
Lin, M. T., & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in
neurodegenerative diseases. Nature, 443(7113), 787–795. doi:nature05292
[pii]\r10.1038/nature05292
Lovell, M. A., & Markesbery, W. R. (2007). Oxidative DNA damage in mild cognitive
impairment and late-stage Alzheimer’s disease. Nucleic Acids Research, 35(22),
7497–7504. doi:10.1093/nar/gkm821

92
Ma, T., Trinh, M. a, Wexler, A. J., Bourbon, C., Gatti, E., Pierre, P., … Klann, E. (2013).
Suppression of eIF2α kinases alleviates Alzheimer’s disease-related plasticity and
memory deficits. Nature Neuroscience, 16(9), 1299–305. doi:10.1038/nn.3486
Maccioni, R. B., Farias, G., Morales, I., & Navarrete, L. (2010). The Revitalized Tau
Hypothesis on Alzheimer’s Disease. Archives of Medical Research, 41(3), 226–231.
doi:10.1016/j.arcmed.2010.03.007
Maccioni, R. B., Lavados, M., Guillón, M., Mujica, C., Bosch, R., Farías, G., & Fuentes,
P. (2006). Anomalously phosphorylated tau and Aβ fragments in the CSF correlates
with cognitive impairment in MCI subjects. Neurobiology of Aging, 27(2), 237–244.
doi:10.1016/j.neurobiolaging.2005.01.011
Maccioni, R. B., Vera, J. C., Dominguez, J., & Avila, J. (1989). A discrete repeated
sequence defines a tubulin binding domain on microtubule-associated protein tau.
Archives of Biochemistry and Biophysics, 275(2), 568–579. doi:10.1016/0003-
9861(89)90403-7
Machová, E., Jakubík, J., Michal, P., Oksman, M., Iivonen, H., Tanila, H., & Doležal, V.
(2008). Impairment of muscarinic transmission in transgenic APPswe/PS1dE9 mice.
Neurobiology of Aging, 29(3), 368–378. doi:10.1016/j.neurobiolaging.2006.10.029
Manaye, K. F., Mouton, P. R., Xu, G., Drew, A., Lei, D. L., Sharma, Y., … Turner, S.
(2013). Age-related loss of noradrenergic neurons in the brains of triple transgenic
mice. Age, 35(1), 139–147. doi:10.1007/s11357-011-9343-0
Maroof, N., Ravipati, S., Pardon, M. C., Barrett, D. a, & Kendall, D. a. (2014).
Reductions in Endocannabinoid Levels and Enhanced Coupling of Cannabinoid
Receptors in the Striatum are Accompanied by Cognitive Impairments in the

93
ABPPswe/PS1DE9 Mouse Model of Alzheimer’s Disease. Journal of Alzheimer’s
Disease : JAD, 42(1), 227–45. doi:10.3233/JAD-131961
Martín-Moreno, A. M., Brera, B., Spuch, C., Carro, E., García-García, L., Delgado, M.,
… de Ceballos, M. L. (2012). Prolonged oral cannabinoid administration prevents
neuroinflammation, lowers β-amyloid levels and improves cognitive performance in
Tg APP 2576 mice. Journal of Neuroinflammation, 9(1), 8. doi:10.1186/1742-2094-
9-8
Masciopinto, F., Di Pietro, N., Corona, C., Bomba, M., Pipino, C., Curcio, M., … Sensi,
S. L. (2012). Effects of long-term treatment with pioglitazone on cognition and
glucose metabolism of PS1-KI, 3xTg-AD, and wild-type mice. Cell Death &
Disease, 3(12), e448. doi:10.1038/cddis.2012.189
Massey, P. V., Bhabra, G., Cho, K., Brown, M. W., & Bashir, Z. I. (2001). Activation of
muscarinic receptors induces protein synthesis-dependent long-lasting depression in
the perirhinal cortex. European Journal of Neuroscience, 14(1), 145–152.
doi:10.1046/j.0953-816X.2001.01631.x
Mastrangelo, M. a, & Bowers, W. J. (2008). Detailed immunohistochemical
characterization of temporal and spatial progression of Alzheimer’s disease-related
pathologies in male triple-transgenic mice. BMC Neuroscience, 9, 81.
doi:10.1186/1471-2202-9-81
Matsuo, K., Okamoto, H., Kawai, Y., Quan, Y. S., Kamiyama, F., Hirobe, S., …
Nakagawa, S. (2014). Vaccine efficacy of transcutaneous immunization with
amyloid B using a dissolving microneedle array in a mouse model of Alzheimer’s
disease. Journal of Neuroimmunology, 266(1-2), 1–11.

94
doi:10.1016/j.jneuroim.2013.11.002
Maurer, K., Volk, S., & Gerbaldo, H. (1997). Auguste D and Alzheimer’s disease.
Lancet, 349(9064), 1546–1549. doi:10.1016/S0140-6736(96)10203-8
McClean, P. L., & Hölscher, C. (2014). Lixisenatide, a drug developed to treat type 2
diabetes, shows neuroprotective effects in a mouse model of Alzheimer’s disease.
Neuropharmacology, 86, 241–58. doi:10.1016/j.neuropharm.2014.07.015
McClean, P. L., Parthsarathy, V., Faivre, E., & Hölscher, C. (2011). The diabetes drug
liraglutide prevents degenerative processes in a mouse model of Alzheimer’s
disease. The Journal of Neuroscience, 31(17), 6587–94.
doi:10.1523/JNEUROSCI.0529-11.2011
McGeer, P. L., Schulzer, M., & McGeer, E. G. (1996). Arthritis and anti-inflammatory
agents as possible protective factors for Alzheimer’s disease: a review of 17
epidemiologic studies. Neurology, 47(2), 425–432. doi:10.1212/WNL.47.2.425
McLaurin, J., Yang, D., Yip, C. M., & Fraser, P. E. (2000). Review: modulating factors
in amyloid-beta fibril formation. Journal of Structural Biology, 130(2-3), 259–270.
doi:10.1006/jsbi.2000.4289
McShea, A., Harris, P. L., Webster, K. R., Wahl, A. F., & Smith, M. A. (1997).
Abnormal expression of the cell cycle regulators P16 and CDK4 in Alzheimer’s
disease. The American Journal of Pathology, 150(6), 1933–9.
Medeiros, R., Castello, N. A., Cheng, D., Kitazawa, M., Baglietto-Vargas, D., Green, K.
N., … Laferla, F. M. (2014). a7 nicotinic receptor agonist enhances cognition in
aged 3xTg-AD mice with robust plaques and tangles. American Journal of
Pathology, 184(2), 520–529. doi:10.1016/j.ajpath.2013.10.010

95
Meraz-Ríos, M. A., Toral-Rios, D., Franco-Bocanegra, D., Villeda-Hernández, J., &
Campos-Peña, V. (2013). Inflammatory process in Alzheimer’s Disease. Frontiers
in Integrative Neuroscience, 7(August), 59. doi:10.3389/fnint.2013.00059
Meunier, M., Bachevalier, J., Mishkin, M., & Murray, E. a. (1993). Effects on visual
recognition of combined and separate ablations of the entorhinal and perirhinal
cortex in rhesus monkeys. The Journal of Neuroscience, 13(12), 5418–5432.
Middei, S., Daniele, S., Caprioli, a, Ghirardi, O., & Ammassari-Teule, M. (2006).
Progressive cognitive decline in a transgenic mouse model of Alzheimer’s disease
overexpressing mutant hAPPswe. Genes, Brain, and Behavior, 5(3), 249–56.
doi:10.1111/j.1601-183X.2005.00160.x
Min, S.-W., Chen, X., Tracy, T. E., Li, Y., Zhou, Y., Wang, C., … Gan, L. (2015).
Critical role of acetylation in tau-mediated neurodegeneration and cognitive deficits.
Nature Medicine, 21(10), 1154–1162. doi:10.1038/nm.3951
Mitchell, J. B., & Laiacona, J. (1998). The medial frontal cortex and temporal memory:
Tests using spontaneous exploratory behaviour in the rat. Behavioural Brain
Research, 97(1-2), 107–113. doi:10.1016/S0166-4328(98)00032-1
Mori, T., Koyama, N., Guillot-Sestier, M.-V., Tan, J., & Town, T. (2013). Ferulic acid is
a nutraceutical β-secretase modulator that improves behavioral impairment and
alzheimer- like pathology in transgenic mice. PloS One, 8(2), e55774.
doi:10.1371/journal.pone.0055774
Mori, T., Rezai-Zadeh, K., Koyama, N., Arendash, G. W., Yamaguchi, H., Kakuda, N.,
… Town, T. (2012). Tannic acid is a natural β-secretase inhibitor that prevents
cognitive impairment and mitigates Alzheimer- like pathology in transgenic mice.

96
The Journal of Biological Chemistry, 287(9), 6912–27.
doi:10.1074/jbc.M111.294025
Morice, E., Andreae, L. C., Cooke, S. F., Vanes, L., Fisher, E. M. C., Tybulewicz, V. L.
J., & Bliss, T. V. P. (2008). Preservation of long-term memory and synaptic
plasticity despite short-term impairments in the Tc1 mouse model of Down
syndrome. Learning & Memory, 15(7), 492–500. doi:10.1101/lm.969608
Morley, J. E., Farr, S. A., Banks, W. A., Johnson, S. N., & Louis, S. (2010). A
Physiological Role for Amyloid- β Protein : Enhancement of Learning and Memory,
19, 441–449. doi:10.3233/JAD-2010-1230
Mosher, K. I., & Wyss-Coray, T. (2011). Microglial Dysfunction in Brain Aging and
Alzheimer’s Disease. Biochemical Pharmacology, 88(4), 594–604.
doi:10.1038/nature13314.A
Mouri, A., Noda, Y., Hara, H., Mizoguchi, H., Tabira, T., & Nabeshima, T. (2007). Oral
vaccination with a viral vector containing Abeta cDNA attenuates age-related Abeta
accumulation and memory deficits without causing inflammation in a mouse
Alzheimer model. FASEB Journal : Official Publication of the Federation of
American Societies for Experimental Biology, 21(9), 2135–48. doi:10.1096/fj.06-
7685com
Mumby, D. G., Gaskin, S., Glenn, M. J., Schramek, T. E., & Lehmann, H. (2002).
Hippocampal Damage and Exploratory Preferences in Rats: Memory for Objects,
Places, and Contexts. Learning & Memory, 9(2), 49–57. doi:10.1101/lm.41302
Mumby, D. G., & Pinel, J. P. J. (1994). Rhinal cortex lesions and object recognition in
rats. Behavioral Neuroscience, 108(1), 11–18. doi:10.1037//0735-7044.108.1.11

97
Murai, T., Okuda, S., Tanaka, T., & Ohta, H. (2007). Characteristics of object location
memory in mice: Behavioral and pharmacological studies. Physiology and Behavior,
90(1), 116–124. doi:10.1016/j.physbeh.2006.09.013
Murray, E. a, & Mishkin, M. (1998). Object recognition and location memory in
monkeys with excitotoxic lesions of the amygdala and hippocampus. The Journal of
Neuroscience, 18(16), 6568–6582.
Murray, E. a., & Bussey, T. J. (1999). Perceptual-mnemonic functions of the perirhinal
cortex. Trends in Cognitive Sciences, 3(4), 142–151. doi:10.1016/S1364-
6613(99)01303-0
Nelson, P., Alafuzoff, I., Bigio, E., Bouras, C., Braak, H., Cairns, N., … Beach, T.
(2013). Correlation of Alzheimer Disease Neuropathologic Changes With Cognitive
Status: A Review of the Literature. J Neuropathol Exp Neurol, 71(5), 362–381.
doi:10.1097/NEN.0b013e31825018f7.Correlation
Nelson, P. T., Braak, H., & Markesbery, W. R. (2007). Neuropathology and cognitive
impairment in Alzheimer Disease: A complex but coherent rel. Aging, 3(6), 379–
389. doi:10.1097/NEN.0b013e3181919a48.Neuropathology
Nicholson, R. M., Kusne, Y., Nowak, L. a., Laferla, F. M., Reiman, E. M., & Valla, J.
(2010). Regional cerebral glucose uptake in the 3xTG model of Alzheimer’s disease
highlights common regional vulnerability across AD mouse models. Brain
Research, 1347, 179–185. doi:10.1016/j.brainres.2010.05.084
Nilsson, L., Nordberg, A., Hardy, J., Wester, P., & Winblad, B. (1986). Physostigmine
restores3H-acetylcholine efflux from Alzheimer brain slices to normal level. Journal
of Neural Transmission, 67(3-4), 275–285. doi:10.1007/BF01243353

98
Nishida, Y., Yokota, T., Takahashi, T., Uchihara, T., Jishage, K., & Mizusawa, H.
(2006). Deletion of vitamin E enhances phenotype of Alzheimer disease model
mouse. Biochemical and Biophysical Research Communications, 350(3), 530–6.
doi:10.1016/j.bbrc.2006.09.083
Nizzari, M., Thellung, S., Corsaro, A., Villa, V., Pagano, A., Porcile, C., … Florio, T.
(2012). Neurodegeneration in Alzheimer disease: Role of amyloid precursor protein
and presenilin 1 intracellular signaling. Journal of Toxicology, 2012.
doi:10.1155/2012/187297
Oakley, H., Cole, S. L., Logan, S., Maus, E., Shao, P., Craft, J., … Vassar, R. (2006).
Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in
transgenic mice with five familial Alzheimer’s disease mutations: potential factors
in amyloid plaque formation. The Journal of Neuroscience, 26(40), 10129–40.
doi:10.1523/JNEUROSCI.1202-06.2006
Obulesu, M., & Rao, D. M. (2010). DNA damage and impairment of DNA repair in
Alzheimer’s disease. The International Journal of Neuroscience, 120(6), 397–403.
doi:10.3109/00207450903411133
Oddo, S., Caccamo, A., Kitazawa, M., Tseng, B. P., & LaFerla, F. M. (2003). Amyloid
deposition precedes tangle formation in a triple transgenic model of Alzheimer’s
disease. Neurobiology of Aging, 24(8), 1063–1070.
doi:10.1016/j.neurobiolaging.2003.08.012
Oddo, S., Caccamo, A., Shepherd, J. D., Murphy, M. P., Golde, T. E., Kayed, R., …
Laferla, F. M. (2003). Triple-Transgenic Model of Alzheimer’s Disease with
Plaques and Tangles : Intracellular AB and Synaptic Dysfunction. Neuron, 39, 409–

99
421.
Oddo, S., Caccamo, A., Tran, L., Lambert, M. P., Glabe, C. G., Klein, W. L., & LaFerla,
F. M. (2006a). Temporal profile of amyloid-B (AB) oligomerization in an in vivo
model of Alzheimer disease: A link between AB and tau pathology. Journal of
Biological Chemistry, 281(3), 1599–1604. doi:10.1074/jbc.M507892200
Oddo, S., Caccamo, A., Tran, L., Lambert, M. P., Glabe, C. G., Klein, W. L., & LaFerla,
F. M. (2006b). Temporal profile of amyloid-B (AB) oligomerization in an in vivo
model of Alzheimer disease: A link between AB and tau pathology. Journal of
Biological Chemistry, 281(3), 1599–1604. doi:10.1074/jbc.M507892200
Oddo, S., Vasilevko, V., Caccamo, A., Kitazawa, M., Cribbs, D. H., & LaFerla, F. M.
(2006). Reduction of soluble AB and tau, but not soluble AB alone, ameliorates
cognitive decline in transgenic mice with plaques and tangles. Journal of Biological
Chemistry, 281(51), 39413–39423. doi:10.1074/jbc.M608485200
Ohno, M., Chang, L., Tseng, W., Oakley, H., Citron, M., Klein, W. L., … Disterhoft, J.
F. (2006). Temporal memory deficits in Alzheimer’s mouse models: Rescue by
genetic deletion of BACE1. European Journal of Neuroscience, 23(1), 251–260.
doi:10.1111/j.1460-9568.2005.04551.x
Ohta, H., Arai, S., Akita, K., Ohta, T., & Fukuda, S. (2012). Effects of NK-4 in a
transgenic mouse model of Alzheimer’s disease. PloS One, 7(1), e30007.
doi:10.1371/journal.pone.0030007
Onishi, T., Iwashita, H., Uno, Y., Kunitomo, J., Saitoh, M., Kimura, E., … Takizawa, M.
(2011). A novel glycogen synthase kinase-3 inhibitor 2-methyl-5-(3-{4-[(S )-
methylsulfinyl]phenyl}-1-benzofuran-5-yl)-1,3,4-oxadiazole decreases tau

100
phosphorylation and ameliorates cognitive deficits in a transgenic model of
Alzheimer’s disease. Journal of Neurochemistry, 119(6), 1330–40.
doi:10.1111/j.1471-4159.2011.07532.x
Paper, O., Biscaro, B., Lindvall, O., Tesco, G., Ekdahl, C. T., & Nitsch, R. M. (2012).
Inhibition of microglial activation protects hippocampal neurogenesis and improves
cognitive deficits in a transgenic mouse model for Alzheimer’s disease. Neuro-
Degenerative Diseases, 9(4), 187–198. doi:10.1159/000330363
Parachikova, A., Vasilevko, V., Cribbs, D. H., LaFerla, F. M., & Green, K. N. (2010).
Reductions in amyloid-beta-derived neuroinflammation, with minocycline, restore
cognition but do not significantly affect tau hyperphosphorylation. Journal of
Alzheimer’s Disease : JAD, 21(2), 527–42. doi:10.3233/JAD-2010-100204
Perry, E., Gibson, P., Blessed, G., Perry, R., & Tomlinson, B. (1977). Neurotransmitter
enzyme abnormalities in senile dementia. Journal of the Neurological Sciences,
34(2), 247–265. doi:10.1016/0022-510X(77)90073-9
Perry, R., & Hodges, J. (1999). Attention and executive deficits in Alzheimer ’ s disease
A critical review. Brain, 383–404.
Pfankuch, T., Rizk, A., Olsen, R., Poage, C., & Raber, J. (2005). Role of circulating
androgen levels in effects of apoE4 on cognitive function. Brain Research, 1053(1-
2), 88–96. doi:10.1016/j.brainres.2005.06.028
Pihlajamaki, M., O’Keefe, K., O’Brien, J., Blacker, D., & Sperling, R. (2011). Failure of
repetition suppression and memory encoding in aging and Alzheimer’s disease.
Brain Imaging Behaviour, 5(1), 36–44. doi:10.1007/s11682-010-9110-3.Failure
Pike, K. E., Savage, G., Villemagne, V. L., Ng, S., Moss, S. a., Maruff, P., … Rowe, C.

101
C. (2007). β-amyloid imaging and memory in non-demented individuals: Evidence
for preclinical Alzheimer’s disease. Brain, 130(11), 2837–2844.
doi:10.1093/brain/awm238
Platt, B., Drever, B., Koss, D., Stoppelkamp, S., Jyoti, A., Plano, A., … Riedel, G.
(2011). Abnormal cognition, sleep, eeg and brain metabolism in a novel knock-in
alzheimer mouse, plb1. PLoS ONE, 6(11). doi:10.1371/journal.pone.0027068
Polito, L., Chierchia, A., Tunesi, M., Bouybayoune, I., Kehoe, P. G., Albani, D., &
Forloni, G. (2014). Environmental Enrichment Lessens Cognitive Decline in APP23
Mice Without Affecting Brain Sirtuin Expression. Journal of Alzheimer’s Disease :
JAD, 42(3), 851–64. doi:10.3233/JAD-131430
Polydoro, M., Acker, C. M., Duff, K., Castillo, P. E., & Davies, P. (2009). Age-
dependent impairment of cognitive and synaptic function in the htau mouse model
of tau pathology. The Journal of Neuroscience, 29(34), 10741–9.
doi:10.1523/JNEUROSCI.1065-09.2009
Prado, V. F., Martins-Silva, C., de Castro, B. M., Lima, R. F., Barros, D. M., Amaral, E.,
… Prado, M. A. M. (2006). Mice Deficient for the Vesicular Acetylcholine
Transporter Are Myasthenic and Have Deficits in Object and Social Recognition.
Neuron, 51(5), 601–612. doi:10.1016/j.neuron.2006.08.005
Price, J. L., Davis, P. B., Morris, J. C., & White, D. L. (1991). The distribution of tangles,
plaques and related immunohistochemical markers in healthy aging and Alzheimer’s
disease. Neurobiology of Aging, 12(4), 295–312. doi:10.1016/0197-4580(91)90006-
6
Puoliväli, J., Wang, J., Heikkinen, T., Heikkilä, M., Tapiola, T., van Groen, T., & Tanila,

102
H. (2002). Hippocampal A beta 42 levels correlate with spatial memory deficit in
APP and PS1 double transgenic mice. Neurobiology of Disease, 9(3), 339–347.
doi:10.1006/nbdi.2002.0481
Rae, E. A., & Brown, R. E. (2015). The problem of genotype and sex differences in life
expectancy in transgenic AD mice. Neuroscience and Biobehavioral Reviews, 57,
238–251. doi:10.1016/j.neubiorev.2015.09.002
Raschetti, R., Maggini, M., Sorrentino, G. C., Martini, N., Caffari, B., & Vanacore, N.
(2005). A cohort study of effectiveness of acetylcholinesterase inhibitors in
Alzheimer’s disease. European Journal of Clinical Pharmacology, 61(5-6), 361–
368. doi:10.1007/s00228-005-0946-1
Reid, G. A., & Darvesh, S. (2015). Butyrylcholinesterase-knockout reduces brain
deposition of fibrillar β-amyloid in an Alzheimer mouse model. Neuroscience, 298,
424–435. doi:10.1016/j.neuroscience.2015.04.039
Reid, J., Jacklin, D., & Winters, B. (2012). Crossmodal object recognition in rats with
and without multimodal object pre-exposure: no effect of hippocampal lesions.
Neurobiology of Learning and Memory, 98(3), 311–9.
doi:10.1016/j.nlm.2012.09.001
Reid, J., Jacklin, D., & Winters, B. (2014). Delineating prefrontal cortex region
contributions to crossmodal object recognition in rats. Cerebral Cortex, 24(8),
2108–2119. doi:10.1093/cercor/bht061
Ribes, D., Torrente, M., Vicens, P., Colomina, M. T., & Domingo, J. L. (2012).
Recognition Memory and b -amyloid Plaques in Adult Tg2576 Mice are not
Modified After Oral Exposure to Aluminum. Alzheimer Dis Assoc Disord, 26(2),

103
179–185.
Richter, H., Ambrée, O., Lewejohann, L., Herring, A., Keyvani, K., Paulus, W., …
Sachser, N. (2008). Wheel-running in a transgenic mouse model of Alzheimer’s
disease: protection or symptom? Behavioural Brain Research, 190(1), 74–84.
doi:10.1016/j.bbr.2008.02.005
Riley, K. P., Snowdon, D. a., & Markesbery, W. R. (2002). Alzheimer’s neurofibrillary
pathology and the spectrum of cognitive function: Findings from the Nun Study.
Annals of Neurology, 51(5), 567–577. doi:10.1002/ana.10161
Rocca, W. A., Amaducci, L. A., & Schoenberg, B. S. (1986). Epidemiology of clinically
diagnosed Alzheimer’s disease. Annals of Neurology, 19(5), 415–24.
doi:10.1002/ana.410190502
Rohn, T. T., Vyas, V., Hernandez-Estrada, T., Nichol, K. E., Christie, L.-A., & Head, E.
(2008). Lack of pathology in a triple transgenic mouse model of Alzheimer’s disease
after overexpression of the anti-apoptotic protein Bcl-2. The Journal of
Neuroscience, 28(12), 3051–3059. doi:10.1523/JNEUROSCI.5620-07.2008
Romberg, C., Mattson, M. P., Mughal, M. R., Bussey, T. J., & Saksida, L. M. (2011).
Impaired attention in the 3xTgAD mouse model of Alzheimer’s disease: rescue by
donepezil (Aricept). The Journal of Neuroscience, 31(9), 3500–7.
doi:10.1523/JNEUROSCI.5242-10.2011
Romberg, C., McTighe, S. M., Heath, C. J., Whitcomb, D. J., Cho, K., Bussey, T. J., &
Saksida, L. M. (2012). False recognition in a mouse model of Alzheimer’s disease:
rescue with sensory restriction and memantine. Brain : A Journal of Neurology,
135(Pt 7), 2103–14. doi:10.1093/brain/aws074

104
Romberg, C., Yang, S., Melani, R., Andrews, M., Horner, A., Spillantini, M., … Saksida,
L. (2013). Depletion of Perineuronal Nets Enhances Recognition Memory and
Long-Term Depression in the Perirhinal Cortex. The Journal of Neuroscience,
33(16), 7057–7065. doi:10.1523/JNEUROSCI.6267-11.2013
Rosario, E. R., Carroll, J., & Pike, C. J. (2010). Testosterone regulation of Alzheimer- like
neuropathology in male 3xTg-AD mice involves both estrogen and androgen
pathways. Brain Research, 1359, 281–290. doi:10.1016/j.brainres.2010.08.068
Rosene, D., & Van Hoesen, G. W. (1977). Hippocampal Efferents Reach Widespread
Areas of Cerebral Cortex and Amygdala in the Rhesus Monkey. Science, 198(4314),
315–317.
Rossor, A. M. N., Iversen, L. L., Reynolds, G. P., Mountjoy, C. Q., Roth, M., Roth, M.,
… Reynolds, G. P. (2015). Neurochemical Characteristics Of Early And Late Onset
Types Of Alzheimer ’ s Disease All use subject to JSTOR Terms and Conditions
CLINICAL Neurochemical characteristics of early and late onset types of Alzheimer
’ s disease. British Medical Journal, 288(6422), 961–964.
Roy, D. S., Arons, A., Mitchell, T. I., Pignatelli, M., Ryan, T. J., & Tonegawa, S. (2016).
Memory retrieval by activating engram cells in mouse models of early Alzheimer’s
disease. Nature, 531, 508–512. doi:10.1038/nature17172
Ruitenberg, A., Ott, A., Van Swieten, J. C., Hofman, A., & Breteler, M. M. B. (2001).
Incidence of dementia: Does gender make a difference? Neurobiology of Aging,
22(4), 575–580. doi:10.1016/S0197-4580(01)00231-7
Ryan, D., Koss, D., Porcu, E., Woodcock, H., Robinson, L., Platt, B., & Riedel, G.
(2013). Spatial learning impairments in PLB1triple knock-in Alzheimer mice are

105
task-specific and age-dependent. Cellular and Molecular Life Sciences : CMLS,
70(14), 2603–19. doi:10.1007/s00018-013-1314-4
Rylett, R. J., Ball, M. J., & Colhoun, E. H. (1983). 121 Evidence for High Affinity
Choline Transport in Synaptosomes Prepared From Hippocampus and Neocortex of
Patients With Alzheimerʼs Disease. Journal of Neuropathology and Experimental
Neurology, 42(3), 345. doi:10.1097/00005072-198305000-00128
Sanchez, P. E., Zhu, L., Verret, L., Vossel, K. A., Orr, A. G., Cirrito, J. R., … Mucke, L.
(2012). Levetiracetam suppresses neuronal network dysfunction and reverses
synaptic and cognitive deficits in an Alzheimer’s disease model. Proceedings of the
National Academy of Sciences, 109(42), E2895–E2903.
doi:10.1073/pnas.1121081109
Saper, C. B., Wainer, B. H., & German, D. C. (1987). Axonal and transneuronal transport
in the transmission of neurological disease: potential role in system degenerations,
including Alzheimer’s disease. Neuroscience, 23(2), 389–398. doi:10.1016/0306-
4522(87)90063-7
Saucier, D. M., Shultz, S. R., Keller, A. J., Cook, C. M., & Binsted, G. (2008). Sex
differences in object location memory and spatial navigation in Long-Evans rats.
Animal Cognition, 11(1), 129–137. doi:10.1007/s10071-007-0096-1
Saydoff, J. a, Olariu, A., Sheng, J., Hu, Z., Li, Q., Garcia, R., … von Borstel, R. (2013).
Uridine prodrug improves memory in Tg2576 and TAPP mice and reduces
pathological factors associated with Alzheimer’s disease in related models. Journal
of Alzheimer’s Disease : JAD, 36(4), 637–57. doi:10.3233/JAD-130059
Scahill, R. I., Schott, J. M., Stevens, J. M., Rossor, M. N., & Fox, N. C. (2002). Mapping

106
the evolution of regional atrophy in Alzheimer’s disease: unbiased analysis of fluid-
registered serial MRI. Proceedings of the National Academy of Sciences of the
United States of America, 99(7), 4703–4707. doi:10.1073/pnas.052587399
Schellenberg, G. D., & Montine, T. J. (2012). The genetics and neuropathology of
Alzheimer’s disease. Acta Neuropathologica, 124(3), 305–323. doi:10.1007/s00401-
012-0996-2
Schneider, F., Baldauf, K., Wetzel, W., & Reymann, K. G. (2014). Behavioral and EEG
changes in male 5xFAD mice. Physiology and Behavior, 135, 25–33.
doi:10.1016/j.physbeh.2014.05.041
Scholtzova, H., Wadghiri, Y. Z., Douadi, M., Sigurdsson, E. M., Li, Y.-S., Quartermain,
D., … Wisniewski, T. (2008). Memantine leads to behavioral improvement and
amyloid reduction in Alzheimer’s-disease-model transgenic mice shown as by
micromagnetic resonance imaging. Journal of Neuroscience Research, 86(12),
2784–91. doi:10.1002/jnr.21713
Schönheit, B., Zarski, R., & Ohm, T. G. (2004). Spatial and temporal relationships
between plaques and tangles in Alzheimer-pathology. Neurobiology of Aging, 25(6),
697–711. doi:10.1016/j.neurobiolaging.2003.09.009
Schrag, M., Mueller, C., Zabel, M., Crofton, A., Kirsch, W. M., Ghribi, O., … Perry, G.
(2013). Oxidative stress in blood in Alzheimer’s disease and mild cognitive
impairment: A meta-analysis. Neurobiology of Disease, 59, 100–110.
doi:10.1016/j.nbd.2013.07.005
Scullion, G. a, Kendall, D. a, Marsden, C. a, Sunter, D., & Pardon, M.-C. (2011). Chronic
treatment with the α2-adrenoceptor antagonist fluparoxan prevents age-related

107
deficits in spatial working memory in APP×PS1 transgenic mice without altering β-
amyloid plaque load or astrocytosis. Neuropharmacology, 60(2-3), 223–34.
doi:10.1016/j.neuropharm.2010.09.002
Seeger, T., Fedorova, I., Zheng, F., Miyakawa, T., Koustova, E., Gomeza, J., … Wess, J.
(2004). M2 Muscarinic Acetylcholine Receptor Knock-Out Mice Show Deficits in
Behavioral Flexibility, Working Memory, and Hippocampal Plasticity. Journal of
Neuroscience, 24(45), 10117–10127. doi:10.1523/JNEUROSCI.3581-04.2004
Selkoe, D. (1991). The Molecular of Alzheimer’s Pathology Disease Review. Neuron, 6,
487–498.
Selkoe, D. (1998). The cell biology of β-amyloid precursor protein and presenilin in
Alzheimer’s disease. Trends in Cell Biology, 8(11), 447–453. doi:10.1016/S0962-
8924(98)01363-4
Selkoe, D. (1999). Translating cell biology into therapeutic advances in Alzheimer’s
disease. Nature, 399(6738 Suppl), A23–A31. doi:10.1038/399a023
Seo, J., Giusti-Rodríguez, P., Zhou, Y., Rudenko, A., Cho, S., Ota, K. T., … Tsai, L.-H.
(2014). Activity-dependent p25 generation regulates synaptic plasticity and Aβ-
induced cognitive impairment. Cell, 157(2), 486–98. doi:10.1016/j.cell.2014.01.065
Shankar, G. M., Li, S., Mehta, T. H., Garcia-Munoz, A., Shepardson, N. E., Smith, I., …
Selkoe, D. J. (2008). Amyloid-beta protein dimers isolated directly from
Alzheimer’s brains impair synaptic plasticity and memory. Nature Medicine, 14(8),
837–42. doi:10.1038/nm1782
Simón, A. M., de Maturana, R. L., Ricobaraza, A., Escribano, L., Schiapparelli, L.,
Cuadrado-Tejedor, M., … Frechilla, D. (2009). Early changes in hippocampal Eph

108
receptors precede the onset of memory decline in mouse models of Alzheimer’s
disease. Journal of Alzheimer’s Disease : JAD, 17(4), 773–86. doi:10.3233/JAD-
2009-1096
Simón, A.-M., Schiapparelli, L., Salazar-Colocho, P., Cuadrado-Tejedor, M., Escribano,
L., López de Maturana, R., … Frechilla, D. (2009). Overexpression of wild-type
human APP in mice causes cognitive deficits and pathological features unrelated to
Abeta levels. Neurobiology of Disease, 33(3), 369–78.
doi:10.1016/j.nbd.2008.11.005
Sisodia, S. S., & St George-Hyslop, P. H. (2002). gamma-Secretase, Notch, Abeta and
Alzheimer’s disease: where do the presenilins fit in? Nature Reviews. Neuroscience,
3(4), 281–290. doi:10.1038/nrn785
Sivilia, S., Lorenzini, L., Giuliani, A., Gusciglio, M., Fernandez, M., Baldassarro, V. A.,
… Giardino, L. (2013). Multi-target action of the novel anti-Alzheimer compound
CHF5074: in vivo study of long term treatment in Tg2576 mice. BMC
Neuroscience, 14, 44. doi:10.1186/1471-2202-14-44
Snowden, J. S., Thompson, J. C., Stopford, C. L., Richardson, A. M. T., Gerhard, A.,
Neary, D., & Mann, D. M. a. (2011). The clinical diagnosis of early-onset
dementias: diagnostic accuracy and clinicopathological relationships. Brain : A
Journal of Neurology, 134(Pt 9), 2478–92. doi:10.1093/brain/awr189
Sperling, R. A., Bates, J. F., Chua, E. F., Cocchiarella, A. J., Rentz, D. M., Rosen, B. R.,
… Albert, M. S. (2003). fMRI studies of associative encoding in young and elderly
controls and mild Alzheimer’s disease. Journal of Neurology, Neurosurgery, and
Psychiatry, 74(1), 44–50. doi:10.1136/jnnp.74.1.44

109
Spilman, P., Descamps, O., Gorostiza, O., Peters-Libeu, C., Poksay, K. S., Matalis, A., …
Bredesen, D. E. (2014). The multi- functional drug tropisetron binds APP and
normalizes cognition in a murine Alzheimer’s model. Brain Research, 1551, 25–44.
doi:10.1016/j.brainres.2013.12.029
Squire, L. R., & Zola, S. M. (1996). Structure and function of declarative and
nondeclarative memory systems. Proceedings of the National Academy of Sciences
of the United States of America, 93(24), 13515–13522.
doi:10.1073/pnas.93.24.13515
St-Amour, I., Paré, I., Tremblay, C., Coulombe, K., Bazin, R., & Calon, F. (2014). IVIg
protects the 3xTg-AD mouse model of Alzheimer’s disease from memory deficit
and Aβ pathology. Journal of Neuroinflammation, 11, 54. doi:10.1186/1742-2094-
11-54
Steiner, J. a, Angot, E., & Brundin, P. (2011). A deadly spread: cellular mechanisms of α-
synuclein transfer. Cell Death and Differentiation, 18(9), 1425–1433.
doi:10.1038/cdd.2011.53
Stelzmann, R. a., Schnitzlein, H. N., & Murtagh, F. R. (1995). An English translation of
Alzheimer’s 1907 paper, “uber eine eigenartige erkankung der hirnrinde.” Clinical
Anatomy, 8(6), 429–431. doi:10.1002/ca.980080612
Sterniczuk, R., Antle, M. C., Laferla, F. M., & Dyck, R. H. (2010). Characterization of
the 3xTg-AD mouse model of Alzheimer’s disease: Part 2. Behavioral and cognitive
changes. Brain Research, 1348, 149–155. doi:10.1016/j.brainres.2010.06.011
Stevens, L. M., & Brown, R. E. (2015). Reference and working memory deficits in the
3xTg-AD mouse between 2 and 15-months of age: A cross-sectional study.

110
Behavioural Brain Research, 278, 496–505. doi:10.1016/j.bbr.2014.10.033
Steward, O. (1976). Topographic organization of the projections from the entorhinal area
to the hippocampal formation of the rat. The Journal of Comparative Neurology,
167(3), 285–314. doi:10.1002/cne.901670303
Stoub, T. R., deToledo-Morrell, L., Stebbins, G. T., Leurgans, S., Bennett, D. a, & Shah,
R. C. (2006). Hippocampal disconnection contributes to memory dysfunction in
individuals at risk for Alzheimer’s disease. Proceedings of the National Academy of
Sciences of the United States of America, 103(26), 10041–10045.
doi:10.1073/pnas.0603414103
Stover, K. R., Campbell, M. a., Van Winssen, C. M., & Brown, R. E. (2015). Early
detection of cognitive deficits in the 3xTg-AD mouse model of Alzheimer’s disease.
Behavioural Brain Research, 289, 29–38. doi:10.1016/j.bbr.2015.04.012
Swanson, L. W., Wyss, J. M., & Cowan, W. M. (1978). An autoradiographic study of the
organization of intrahippocampal association pathways in the rat. The Journal of
Comparative Neurology, 181(4), 681–715. doi:10.1002/cne.901810402
Swerdlow, R. H., Burns, J. M., & Khan, S. M. (2014). The Alzheimer’s Disease
Mitochondrial Cascade Hypothesis: Progress and Perspectives. October (Vol.
1842). doi:10.1016/j.surg.2006.10.010.Use
Sy, M., Kitazawa, M., Medeiros, R., Whitman, L., Cheng, D., Lane, T. E., & LaFerla, F.
M. (2011). Inflammation induced by infection potentiates tau pathological features
in transgenic mice. American Journal of Pathology, 178(6), 2811–2822.
doi:10.1016/j.ajpath.2011.02.012
Szwagierczak, A., Bultmann, S., Schmidt, C. S., Spada, F., & Leonhardt, H. (2010).

111
Sensitive enzymatic quantification of 5-hydroxymethylcytosine in genomic DNA.
Nucleic Acids Research, 38(19), e181. doi:10.1093/nar/gkq684
Tabaton, M., Mandybur, T. I., Perry, G., Onorato, M., Autilio-Gambetti, L., & Gambetti,
P. (1989). The widespread alteration of neurites in Alzheimer’s disease may be
unrelated to amyloid deposition. Annals of Neurology, 26(6), 771–778.
doi:10.1002/ana.410260614
Taglialatela, G., Hogan, D., Zhang, W.-R., & Dineley, K. T. (2009). Intermediate- and
long-term recognition memory deficits in Tg2576 mice are reversed with acute
calcineurin inhibition. Behavioural Brain Research, 200(1), 95–9.
doi:10.1016/j.bbr.2008.12.034
Terai, K., Iwai, a, Kawabata, S., Tasaki, Y., Watanabe, T., Miyata, K., & Yamaguchi, T.
(2001). beta-amyloid deposits in transgenic mice expressing human beta-amyloid
precursor protein have the same characteristics as those in Alzheimer’s disease.
Neuroscience, 104(2), 299–310. doi:S0306-4522(01)00095-1 [pii]
Thinakaran, G., & Koo, E. H. (2008). Amyloid precursor protein trafficking, processing,
and function. Journal of Biological Chemistry, 283(44), 29615–29619.
doi:10.1074/jbc.R800019200
Tinsley, C. J., Fontaine-Palmer, N. S., Vincent, M., Endean, E. P. E., Aggleton, J. P.,
Brown, M. W., & Warburton, E. C. (2011). Differing time dependencies of object
recognition memory impairments produced by nicotinic and muscarinic cholinergic
antagonism in perirhinal cortex. Learning & Memor,, 18(7), 484–492.
doi:10.1101/lm.2274911
Todd Roach, J., Volmar, C.-H., Dwivedi, S., Town, T., Crescentini, R., Crawford, F., …

112
Mullan, M. (2004). Behavioral effects of CD40-CD40L pathway disruption in aged
PSAPP mice. Brain Research, 1015(1-2), 161–8. doi:10.1016/j.brainres.2004.05.004
Tohda, C., Nakada, R., Urano, T., Okonogi, A., & Kuboyama, T. (2011). Kamikihi-to
(KKT) rescues axonal and synaptic degeneration associated with memory
impairment in a mouse model of Alzheimer’s disease, 5XFAD. The International
Journal of Neuroscience, 121(12), 641–8. doi:10.3109/00207454.2011.602809
Tohda, C., Urano, T., Umezaki, M., Nemere, I., & Kuboyama, T. (2012). Diosgenin is an
exogenous activator of 1,25D₃-MARRS/Pdia3/ERp57 and improves Alzheimer’s
disease pathologies in 5XFAD mice. Scientific Reports, 2, 535.
doi:10.1038/srep00535
Toledo-Morrell, L. de, & Dickerson, B. (2000). Hemispheric differences in hippocampal
volume predict verbal and spatial memory performance in patients with Alzheimer’s
disease. Hippocampus, 142, 136–142.
Turner, P. R., O’Connor, K., Tate, W. P., & Abraham, W. C. (2003). Roles of amyloid
precursor protein and its fragments in regulating neural activity, plasticity and
memory. Progress in Neurobiology (Vol. 70). doi:10.1016/S0301-0082(03)00089-3
Valla, J., Schneider, L., Niedzielko, T., Coon, K. D., Caselli, R., Sabbagh, M. N., …
Reiman, E. M. (2006). Impaired platelet mitochondrial activity in Alzheimer’s
disease and mild cognitive impairment. Mitochondrion, 6(6), 323–330.
doi:10.1016/j.mito.2006.10.004
Van Dam, D., & De Deyn, P. P. (2011). Animal models in the drug discovery pipeline for
Alzheimer’s disease. British Journal of Pharmacology, 164(4), 1285–1300.
doi:10.1111/j.1476-5381.2011.01299.x

113
Van Hoesen, G. W., Pandya, D., & Butters, N. (1972). Cortical Afferents to the
Entorhinal Cortex of the Rhesus Monkey. Science, 175(4029), 1471–1473.
Vargas, J. Y., Fuenzalida, M., & Inestrosa, N. C. (2014). In vivo activation of Wnt
signaling pathway enhances cognitive function of adult mice and reverses cognitive
deficits in an Alzheimer’s disease model. The Journal of Neuroscience , 34(6),
2191–202. doi:10.1523/JNEUROSCI.0862-13.2014
Vaucher, E., Fluit, P., Chishti, M. a, Westaway, D., Mount, H. T. J., & Kar, S. (2002).
Object recognition memory and cholinergic parameters in mice expressing human
presenilin 1 transgenes. Experimental Neurology, 175(2), 398–406.
doi:10.1006/exnr.2002.7915
Verret, L., Krezymon, A., Halley, H., Trouche, S., Zerwas, M., Lazouret, M., …
Rampon, C. (2013). Transient enriched housing before amyloidosis onset sustains
cognitive improvement in Tg2576 mice. Neurobiology of Aging, 34(1), 211–25.
doi:10.1016/j.neurobiolaging.2012.05.013
Villemagne, V., Pike, K., Chételat, G., Ellis, K., Mulligan, R., Bourgeat, P., … Rowe, C.
(2011). Longitudinal assessment of Aβ and cognition in aging and Alzheimer
disease. Ann Neurol., 69(1), 181–192. doi:10.1002/ana.22248.Longitudinal
Wang, D., Fu, Q., Zhou, Y., Xu, B., Shi, Q., Igwe, B., … Xiang, Y. K. (2013). β2
adrenergic receptor, protein kinase A (PKA) and c-Jun N-terminal kinase (JNK)
signaling pathways mediate tau pathology in Alzheimer disease models. The Journal
of Biological Chemistry, 288(15), 10298–307. doi:10.1074/jbc.M112.415141
Wang, J., Wright, H. M., Vempati, P., Li, H., Wangsa, J., Dzhuan, A., … Pasinetti, G. M.
(2013). Investigation of nebivolol as a novel therapeutic agent for the treatment of

114
Alzheimer’s disease. Journal of Alzheimer’s Disease : JAD, 33(4), 1147–56.
doi:10.3233/JAD-2012-120904
Wang, J., Xia, Y., Grundke-Iqbal, I., & Iqbal, K. (2012). Abnormal hyperphosphorylation
of tau: Sites, regulation, and molecular mechanism of neurofibrillary degeneration.
Advances in Alzheimer’s Disease, 3, 123–139. doi:10.3233/978-1-61499-154-0-123
Wang, L., Zang, Y., He, Y., Liang, M., Zhang, X., Tian, L., … Li, K. (2006). Changes in
hippocampal connectivity in the early stages of Alzheimer’s disease: Evidence from
resting state fMRI. NeuroImage, 31(2), 496–504.
doi:10.1016/j.neuroimage.2005.12.033
Wang, W., Li, S., Dong, H. ping, Lv, S., & Tang, Y. yuan. (2009). Differential
impairment of spatial and nonspatial cognition in a mouse model of brain aging. Life
Sciences, 85(3-4), 127–135. doi:10.1016/j.lfs.2009.05.003
Warburton, E. C., Koder, T., Cho, K., Massey, P. V, Duguid, G., Barker, G. R. I., …
Brown, M. W. (2003). Cholinergic Neurotrasnmission Is Essential for Perirhinal
Cortical Plasticity and Recognition Memory. Neuron, 38, 987–996.
doi:10.1016/s0896-6273(03)00358-1
Watanabe, T., Yamagata, N., Takasaki, K., Sano, K., Hayakawa, K., Katsurabayashi, S.,
… Fujiwara, M. (2009). Decreased acetylcholine release is correlated to memory
impairment in the Tg2576 transgenic mouse model of Alzheimer’s disease. Brain
Research, 1249, 222–228. doi:10.1016/j.brainres.2008.10.029
Webster, S. J., Bachstetter, A. D., Nelson, P. T., Schmitt, F. a, & Van Eldik, L. J. (2014).
Using mice to model Alzheimer’s dementia: an overview of the clinical disease and
the preclinical behavioral changes in 10 mouse models. Frontiers in Genetics,

115
5(April), 88. doi:10.3389/fgene.2014.00088
Webster, S. J., Bachstetter, A. D., & Van Eldik, L. J. (2013). Comprehensive behavioral
characterization of an APP/PS-1 double knock-in mouse model of Alzheimer’s
disease. Alzheimer’s Research & Therapy, 5(3), 28. doi:10.1186/alzrt182
Weintraub, S., Wicklund, A. H., & Salmon, D. P. (2012). The neuropsychological profile
of Alzheimer disease. Cold Spring Harbor Perspectives in Medicine, 2(4), 1–18.
doi:10.1101/cshperspect.a006171
Weissman, L., de Souza-Pinto, N. C., Mattson, M. P., & Bohr, V. A. (2009). DNA base
excision repair activities in mouse models of Alzheimer’s disease. Neurobiol Aging,
30(12), 2080–2081. doi:10.1016/j.surg.2006.10.010.Use
White, K. G., & Ruske, A. C. (2002). Memory deficits in Alzheimer’s disease: The
encoding hypothesis and cholinergic function. Psychonomic Bulletin & Review,
9(3), 426–437. doi:10.3758/BF03196301
Whitehouse, P., Price, D., Struble, R., Clark, A., Coyle, J., & DeLong, M. (1982).
Alzheimer’s Disease and Senile Dementia: Loss of Neurons in the Basal Forebrain.
Science, 215(4537), 1237–1239. doi:10.1038/020493a0
Whitson, J., Selkoe, D., & Cotman, C. (1989). Amyloid beta Protein Enhances the
Survival of Hippocampal Neurons in vitro, 243(4897), 1488–1490.
Whitwell, J. L., Przybelski, S. a, Weigand, S. D., Knopman, D. S., Boeve, B. F., Petersen,
R. C., & Jack, C. R. (2007). 3D maps from multiple MRI illustrate changing atrophy
patterns as subjects progress from mild cognitive impairment to Alzheimer’s
disease. Brain : A Journal of Neurology, 130(Pt 7), 1777–86.
doi:10.1093/brain/awm112

116
Wilkinson, J. L., Herrman, L., Palmatier, M. I., & Bevins, R. A. (2006). Rats’ novel
object interaction as a measure of environmental familiarity. Learning and
Motivation, 37(2), 131–148. doi:10.1016/j.lmot.2005.04.001
Wimmer, M. E., Hernandez, P. J., Blackwell, J., & Abel, T. (2012). Aging impairs
hippocampus-dependent long-term memory for object location in mice.
Neurobiology of Aging, 33(9), 2220–2224.
doi:10.1016/j.neurobiolaging.2011.07.007
Winters, B. D. (2006). Paradoxical Facilitation of Object Recognition Memory after
Infusion of Scopolamine into Perirhinal Cortex: Implications for Cholinergic System
Function. Journal of Neuroscience, 26(37), 9520–9529.
doi:10.1523/JNEUROSCI.2319-06.2006
Winters, B. D., Saksida, L. M., & Bussey, T. J. (2010). Implications of animal object
memory research for human amnesia. Neuropsychologia, 48(8), 2251–2261.
doi:10.1016/j.neuropsychologia.2010.01.023
Winters, B., Forwood, S., Cowell, R., Saksida, L., & Bussey, T. (2004). Double
dissociation between the effects of peri-postrhinal cortex and hippocampal lesions
on tests of object recognition and spatial memory: heterogeneity of function within
the temporal lobe. The Journal of Neuroscience, 24(26), 5901–8.
doi:10.1523/JNEUROSCI.1346-04.2004
Winters, B., & Reid, J. (2010). A distributed cortical representation underlies crossmodal
object recognition in rats. The Journal of Neuroscience, 30(18), 6253–61.
doi:10.1523/JNEUROSCI.6073-09.2010
Winters, B., Saksida, L., & Bussey, T. (2008). Object recognition memory:

117
neurobiological mechanisms of encoding, consolidation and retrieval. Neuroscience
and Biobehavioral Reviews, 32(5), 1055–70. doi:10.1016/j.neubiorev.2008.04.004
Wolk, D. a., Signoff, E. D., & DeKosky, S. T. (2008). Recollection and familiarity in
amnestic mild cognitive impairment: A global decline in recognition memory.
Neuropsychologia, 46(7), 1965–1978.
doi:10.1016/j.neuropsychologia.2008.01.017.Recollection
Woo, D.-C., Lee, S.-H., Lee, D.-W., Kim, S.-Y., Kim, G.-Y., Rhim, H.-S., … Choe, B.-
Y. (2010). Regional metabolic alteration of Alzheimer’s disease in mouse brain
expressing mutant human APP-PS1 by 1H HR-MAS. Behavioural Brain Research,
211(1), 125–31. doi:10.1016/j.bbr.2010.03.026
Wu, J., Yang, J., Yu, Y., Li, Q., Nakamura, N., Shen, Y., … Abe, K. (2012). Delayed
audiovisual integration of patients with mild cognitive impairment and Alzheimer’s
disease compared with normal aged controls. Journal of Alzheimer’s Disease, 32(2),
317–328. doi:10.3233/JAD-2012-111070
Wu, P., Shen, Q., Dong, S., Xu, Z., Tsien, J. Z., & Hu, Y. (2008). Calorie restriction
ameliorates neurodegenerative phenotypes in forebrain-specific presenilin-1 and
presenilin-2 double knockout mice. Neurobiology of Aging, 29(10), 1502–11.
doi:10.1016/j.neurobiolaging.2007.03.028
Xu, J., Chen, S., Ahmed, S. H., Chen, H., Ku, G., Goldberg, M. P., & Hsu, C. Y. (2001).
Amyloid-beta peptides are cytotoxic to oligodendrocytes. The Journal of
Neuroscience, 21(1), RC118. doi:20014845
Yang, Y., Mufson, E. J., & Herrup, K. (2003). Neuronal cell death is preceded by cell
cycle events at all stages of Alzheimer’s disease. The Journal of Neuroscience,

118
23(7), 2557–63. Retrieved from http://www.jneurosci.org/content/23/7/2557.abstract
Yao, Z.-G., Zhang, L., Liang, L., Liu, Y., Yang, Y.-J., Huang, L., … Qin, C. (2013). The
effect of PN-1, a Traditional Chinese Prescription, on the Learning and Memory in a
Transgenic Mouse Model of Alzheimer’s Disease. Evidence-Based Complementary
and Alternative Medicine : eCAM, 2013, 518421. doi:10.1155/2013/518421
Yassine, N., Lazaris, A., Dorner-Ciossek, C., Després, O., Meyer, L., Maitre, M., …
Mathis, C. (2013). Detecting spatial memory deficits beyond blindness in tg2576
Alzheimer mice. Neurobiology of Aging, 34(3), 716–30.
doi:10.1016/j.neurobiolaging.2012.06.016
Yonelinas, A. P., Kroll, N. E. a, Quamme, J. R., Lazzara, M. M., Sauvé, M.-J., Widaman,
K. F., & Knight, R. T. (2002). Effects of extensive temporal lobe damage or mild
hypoxia on recollection and familiarity. Nature Neuroscience, 5(11), 1236–1241.
doi:10.1038/nn961
Yuede, C. M., Zimmerman, S. D., Dong, H., Kling, M. J., Bero, A. W., Holtzman, D. M.,
… Csernansky, J. G. (2009). Effects of voluntary and forced exercise on plaque
deposition, hippocampal volume, and behavior in the Tg2576 mouse model of
Alzheimer’s disease. Neurobiology of Disease, 35(3), 426–32.
doi:10.1016/j.nbd.2009.06.002
Zhang, C., Cheng, Y., Wang, H., Wang, C., Wilson, S. P., Xu, J., & Zhang, H.-T. (2014).
RNA interference-mediated knockdown of long-form phosphodiesterase-4D
(PDE4D) enzyme reverses amyloid-β42-induced memory deficits in mice. Journal
of Alzheimer’s Disease : JAD, 38(2), 269–80. doi:10.3233/JAD-122236
Zhang, J., Guo, J., Zhao, X., Chen, Z., Wang, G., Liu, A., … Wang, C. (2013).

119
Phosphodiesterase-5 inhibitor sildenafil prevents neuroinflammation, lowers beta-
amyloid levels and improves cognitive performance in APP/PS1 transgenic mice.
Behavioural Brain Research, 250, 230–7. doi:10.1016/j.bbr.2013.05.017
Zhang, L., Shen, C., Chu, J., Zhang, R., Li, Y., & Li, L. (2014). Icariin decreases the
expression of APP and BACE-1 and reduces the β-amyloid burden in an APP
transgenic mouse model of Alzheimer’s disease. International Journal of Biological
Sciences, 10(2), 181–91. doi:10.7150/ijbs.6232
Zhang, L., Xing, Y., Ye, C.-F., Ai, H.-X., Wei, H.-F., & Li, L. (2006). Learning-memory
deficit with aging in APP transgenic mice of Alzheimer’s disease and intervention
by using tetrahydroxystilbene glucoside. Behavioural Brain Research, 173(2), 246–
54. doi:10.1016/j.bbr.2006.06.034
Zhang, R., Xue, G., Wang, S., Zhang, L., Shi, C., & Xie, X. (2012). Novel object
recognition as a facile behavior test for evaluating drug effects in AβPP/PS1
Alzheimer’s disease mouse model. Journal of Alzheimer’s Disease : JAD, 31(4),
801–12. doi:10.3233/JAD-2012-120151
Zhang, Z., Liu, X., Schroeder, J. P., Chan, C.-B., Song, M., Yu, S. P., … Ye, K. (2013).
7,8-Dihydroxyflavone Prevents Synaptic Loss and Memory Deficits in a Mouse
Model of Alzheimer’s Disease. Neuropsychopharmacology, 39(3), 638–650.
doi:10.1038/npp.2013.243
Zheng, H., Xu, H., Uljon, S. N., Gross, R., Hardy, K., Gaynor, J., … Duff, K. (2002).
Modulation of A(beta) peptides by estrogen in mouse models. Journal of
Neurochemistry, 80(1), 191–6. doi:10.1046/j.0022-3042.2001.00690.x
Zola-Morgan, S., Squire, L. R., Amaral, D. G., & Suzuki, W. a. (1989). Lesions of

120
perirhinal and parahippocampal cortex that spare the amygdala and hippocampal
formation produce severe memory impairment. The Journal of Neuroscience , 9(12),
4355–4370.
Zufferey, V., Vallet, P. G., Moeri, M., Moulin-Sallanon, M., Piotton, F., Marin, P., &
Savioz, A. (2013). Maladaptive exploratory behavior and neuropathology of the PS-
1 P117L Alzheimer transgenic mice. Brain Research Bulletin, 94, 17–22.
doi:10.1016/j.brainresbull.2013.01.011
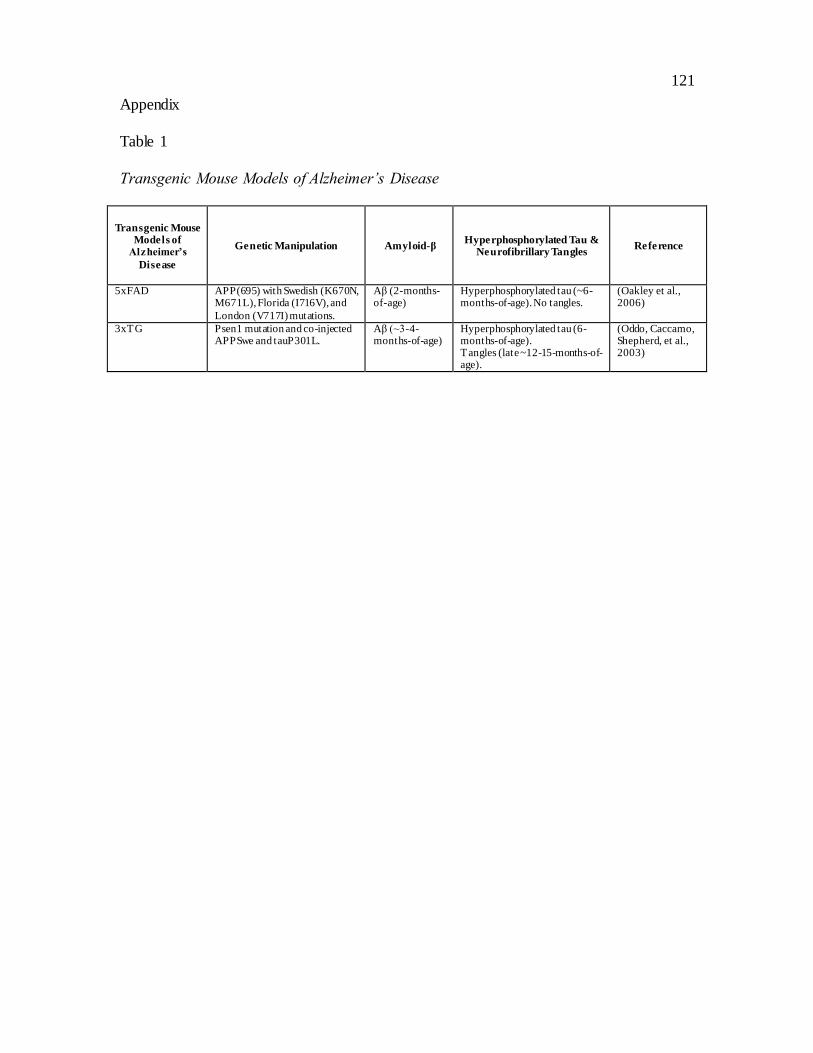
121
Appendix
Table 1
Transgenic Mouse Models of Alzheimer’s Disease
Transgenic Mouse Models of
Alzheimer’s
Disease
Genetic Manipulation Amyloid-β Hyperphosphorylated Tau &
Neurofibrillary Tangles Reference
5xFAD APP(695) with Swedish (K670N, M671L), Florida (I716V), and
London (V717I) mutations.
Aβ (2-months-of-age)
Hyperphosphorylated tau (~6-months-of-age). No tangles.
(Oakley et al., 2006)
3xTG Psen1 mutation and co-injected APPSwe and tauP301L.
Aβ (~3-4-months-of-age)
Hyperphosphorylated tau (6-months-of-age). Tangles (late ~12-15-months-of-age).
(Oddo, Caccamo, Shepherd, et al., 2003)

122
Table 2
Object Recognition Deficits in Transgenic Mouse Models of Alzheimer’s Disease
Transgenic
Strain
Sex Age Genetic,
Pharmacological, Surigcal or
Environmental Intervention
Inter-
Trial-Interval
Behavioural
Task
Behavioural
Result
Reference
J20 Male ~2
months of age
1h OR Tg intact OR. (Karl et
al., 2012)
J20 - 5-8
months of age
CX3CR1 GFP knock-in 24h OR APPTg intact
OR. CX3CR1 APP Tg impaired OR.
(Cho et
al., 2011)
J20 Male 6 months
of age
Poly(ADP-ribose) polymerase-1 (PARP-1;
nuclear protein that regulates cellular inflammatory responses)
24h OR Tg impaired OR. PARP-1
KO attenuated OR impairment in
Tg.
(Kauppinen et al.,
2011)
J20 Male & Female
4.5-6 months of age
Tropisetron (increase the ratio of the trophic, neurite-extending peptide sAPPα to the anti-trophic,
neurite-retractive peptide Aβ)
1h OR Tg impaired OR. Tropisetron attenuated
OR impairment in Tg.
(Spilman et al., 2014)
J20 & B254 Male 2-3, 5-7 months
of age
4h OR J20 and B254 impaired at 2-
3 and 5-7 months.
(Harris et al., 2010)
hAβPPswe-ind - 2, 4, 8 months of age
24h OR Tg impaired OR at 4 and 8 months.
(Simón et al., 2009)
hAPP - 4, 9-10 months
of age
Rosiglitazone (agonist at peroxisome proliferator-
activated receptor gamma (PPARc))
24h OR Tg impaired OR at 4 and
9-10 months. Rosiglitazone attenuated OR
impairment in Tg.
(Escribano et al.,
2009)
APP69K595N/M596L
Male 7 months of age
Xanthoceraside (triterpenoid saponin) and donepezil
24h OR Tg impaired OR at 1h and 24h.
Xanthoceraside and donepezil attenuated
OR impairment in Tg.
(Jin et al., 2014)
APPL/S Female 5.5-7.5 months
of age
LM11A-31 (p75^NTR ligand; neurotrophin
receptor ligand)
24h OR Tg impaired OR. LM11A-
31 attenuated OR impairment in Tg.
(Knowles et al.,
2013)
APP23 - 3.3
months of age
heterozygous neprilysin-
deficient background
24h OR APP23Tg
intact OR. Neprilysin deficiency induced OR
impairment in
(S.-M.
Huang et al., 2006)

123
APP23Tg.
APP23 Female 10 months of age
DSP-4 1h OR Tg impaired OR.
(Heneka et al., 2006)
PDAPP Male 8, 24 months of age
m226 Aβ antibody 3h OR Tg impaired OR at 8 and 24h months.
m226 attenuated OR
impairment in Tg at 24 months.
(J.-C. Dodart et al., 2002)
PDAPP Male 6-9, 13-15, 18-
21 months of age
10s, 1 min, 10min, 1h,
4h
OR Tg intact OR. (Chen, Chen,
Knox, Inglis, Bernard, Martin,
Justice, Mcconlogue, et al., 2000)
PDAPP Male 3, 6, 9-
10 months of age
3h OR Tg impaired
OR at 6, 9-10 months.
( Dodart
et al., 1999)
PDAPP Male &
Female
4, 10, 16
months of age
2,3,5,4'-tetrahydroxystilbene-2-O-
B-d-glucoside (TSG)
24h OR Tg impaired or at 4, 10, 16
months. TSG attenuated TG OR impairment at
10 and 16 months.
(Zhang et al., 2006)
APP/London Male & Female
4.5 months of age
A-887755 (Aβ globulomer specific antibody)
2.5h OR Tg impaired OR. A-887755
attenuated OR impairment in Tg.
(Hillen et al., 2010)
APPV7171 Male
& Female
4, 10
months of age
Icarrin (component of
herb-Epimedium)
24h OR Tg impaired
OR at 10 months Icarrin attenuated
OR impairment in Tg.
(Zhang et
al., 2014)
TgCRND8 Male ~3-4 months
of age
Levodopa ~1h OR (Note: Two object
tests over 2 days. First test used to habituate
mice to objects)
Tg impaired OR.
Levodopa attenuated OR impairment in
Tg.
(Ambrée et al.,
2009)
TgCRND8 Male ~5 months of age
Wheel runing 1h OR Tg impaired OR. Wheel running did
not alter Tg OR performance.
(Richter et al., 2008)
TgCRND8 - 3.5 months
of age
Dexefaroxan (a2-adrenoceptors antagonist)
and rivastigmine
3h OR Tg impaired OR.
Dexefaroxan
(Francis, Yang, et
al., 2012)

124
and rivastigmine attenuated OR deficit in
Tg.
TgCRND8 Male ~2 months of age
Deep brain stimulation (midline thalamic region)
1h OR Tg impaired OR. Deep brain stimulation
improved OR in Tg.
(Arrieta-Cruz et al., 2010)
TgCRND8 - 1, 2, 6-8 months of age
5min, 1h, 3h
OR Tg intact OR at 1 month. Tg impaired
OR with 5min or 3h retention
delay at 2 months. Tg impaired OR with 5min, 1h
and 3h retention delay at 6-8 months.
(Francis, Kim, et al., 2012)
TgCRND8 - 1, 2
months of age
Leptin 1h OR Tg impaired
OR at 1 and 2 months. Leptin attenuated
OR impairment in Tg.
(Greco et
al., 2010)
TgCRND8 Female ~3.8 months
of age
Environmental enrichment
90min OR Tg impaired OR.
Environmental enrichment improved OR in Wt and Tg.
(Görtz et al., 2008)
TgCRND8 - ~2
months of age
Memantine (N -methyl-D-
aspartic acid receptor antagonist) and sensory deprivation
1h False
recognition
Tg have false
recognition. Memantine and sensory deprivation
blocked false recognition in Tg.
(Romberg
et al., 2012)
TgCRND8 Female 14-16 months
of age
Nabs-Aβ (natural Aβ antibody)
30min OL Tg impaired OL. Nabs-Aβ
attenuated OL impairment in Tg.
(Dodel et al., 2011)
APP23 Male 7 months
of age
Environmental enrichment
24h OR Tg impaired OR.
Environmental enrichment attenuated OR deficit in
Tg.
(Polito et al., 2014)
Tg2576 Male & Female
~9 months of age
Voluntary or forced exercise
50min OR Voluntary exercise attenuated OR
impairment in Tg. (Note: did not evaluate
Wt).
(Yuede et al., 2009)

125
Tg2576 Female 6, 12 months of age
NK-4 (neurotrophic and antioxidant activity)
24h OR Tg impaired OR. NK-4 attenauted OR
impairment in Tg.
(Ohta et al., 2012)
Tg2576 Female ~14 months of age
Cognitive stimulation (food rewarded Lashley-like mazes)
1h, 24h OR Tg impaired OR at 1h and 24h.
Cognitive stimulation attenuated
OR deficit at 24h in Tg.
(Gerenu et al., 2013)
Tg2576 Male & Female
5 months of age
a7 nAChR knockout 24h OR Tg 2576 and a7KOTg2576 impaired OR.
(Hernandez et al., 2010)
Tg2576 - 12 months of age
T tpa-/- Mice (a-tocopherol transfer protein knockout)
24h, 28h OR Ttpa-/-Tg2576 mice impaired OR
at 24h and 48h.
(Nishida et al., 2006)
Tg2576 Male & Female
12-14 months of age
Ciproxifan (H3 receptor antagonist)
5min OR Tg impaired OR. Ciproxifan
attenuated OR impairment
(Bardgett et al., 2011)
Tg2576 Female 7 months
of age
CHF5074 (microglia modulator, non-steroidal
anti-inflammatory) and LY450139 (semagacestat) (gamma-secretase inhibitor)
4h OR Tg impaired OR.
CHF5074 attenuated OR impairment in
Tg.
(Giuliani et al.,
2013)
Tg2576 Female 7 months of age
24h OR Tg impaired OR.
(Beggiato et al., 2014)
Tg2576 Male & Female
5 months of age
FK506 (calcineurin inhibitor)
2min, 4h, 24h
OR Tg intact OR at 2min. Tg impaired OR
at 4h and 24h. FK506 attenuated OR
impairment in Tg.
(Taglialatela et al., 2009)
Tg2576 - 12 months of age
Nebivolol (B1 adrenergic receptor antagonist)
1h, 24h OR Nebivolol improved OR in Tg at 1h.
Nebivolol did not alter OR in Tg at 24h.
(Wang et al., 2013)
Tg2576 Female 18 months
of age
CHF5074 (non-steroidal anti-inflammatory
derivative, gamma secretase modulator)
4h OR Tg impaired OR.
CHF5074 attenuated OR impairment.
(Sivilia et al., 2013)
Tg2576 Female 6, 10,
13 months of age
Oral vaccine with a
recombinant adeno-associated viral vector carrying Aβ cDNA (AAV/Aβ)
24h OR Tg intact OR
at 6 months. Tg impairded OR at 10 and 13 months.
AAV/Aβ attenuated OR impairment in
TG at 13
(Mouri et
al., 2007)
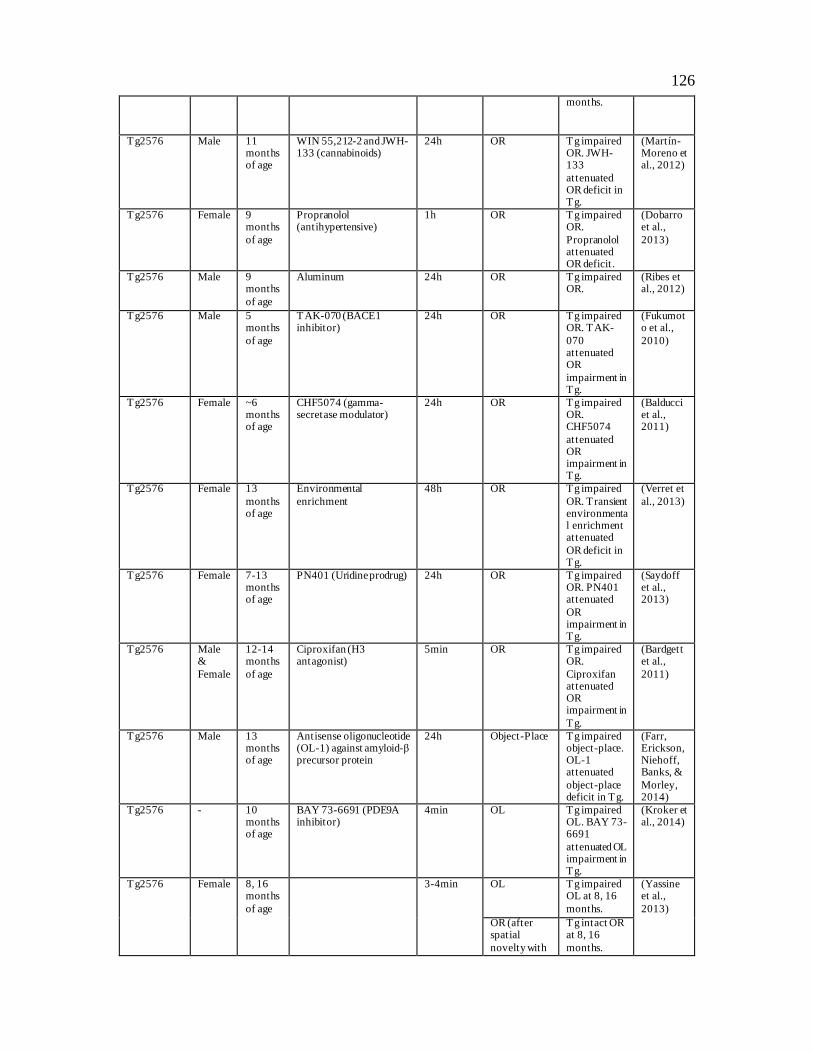
126
months.
Tg2576 Male 11 months of age
WIN 55,212-2 and JWH-133 (cannabinoids)
24h OR Tg impaired OR. JWH-133
attenuated OR deficit in Tg.
(Martín-Moreno et al., 2012)
Tg2576 Female 9 months
of age
Propranolol (antihypertensive)
1h OR Tg impaired OR.
Propranolol attenuated OR deficit .
(Dobarro et al.,
2013)
Tg2576 Male 9 months
of age
Aluminum 24h OR Tg impaired OR.
(Ribes et al., 2012)
Tg2576 Male 5 months
of age
TAK-070 (BACE1 inhibitor)
24h OR Tg impaired OR. TAK-
070 attenuated OR
impairment in Tg.
(Fukumoto et al.,
2010)
Tg2576 Female ~6 months of age
CHF5074 (gamma-secretase modulator)
24h OR Tg impaired OR. CHF5074
attenuated OR impairment in Tg.
(Balducci et al., 2011)
Tg2576 Female 13
months of age
Environmental
enrichment
48h OR Tg impaired
OR. Transient environmental enrichment attenuated
OR deficit in Tg.
(Verret et
al., 2013)
Tg2576 Female 7-13 months of age
PN401 (Uridine prodrug) 24h OR Tg impaired OR. PN401 attenuated
OR impairment in Tg.
(Saydoff et al., 2013)
Tg2576 Male &
Female
12-14 months
of age
Ciproxifan (H3 antagonist)
5min OR Tg impaired OR.
Ciproxifan attenuated OR impairment in
Tg.
(Bardgett et al.,
2011)
Tg2576 Male 13 months of age
Antisense oligonucleotide (OL-1) against amyloid-β precursor protein
24h Object-Place Tg impaired object-place. OL-1 attenuated
object-place deficit in Tg.
(Farr, Erickson, Niehoff, Banks, &
Morley, 2014)
Tg2576 - 10 months of age
BAY 73-6691 (PDE9A inhibitor)
4min OL Tg impaired OL. BAY 73-6691
attenuated OL impairment in Tg.
(Kroker et al., 2014)
Tg2576 Female
8, 16 months
of age
3-4min OL Tg impaired OL at 8, 16
months.
(Yassine et al.,
2013)
OR (after spatial
novelty with
Tg intact OR at 8, 16
months.

127
same objects)
Tg2576 Male 16 months of age
2min OR Tg intact OR. (Good & Hale, 2007)
OL Tg impaired OL.
Object-in-Place
Tg impaired familiar
object and familiar location.
Tg2576 Male 7, 14 months
of age
3min spatial novelt (5 objects)
Tg impaired spatial
novelty at 7 months. Wt and Tg impaired
spatial novelty at 14 months.
(Middei et al., 2006)
Object novelty (after
OL with same objects; 5 objects)
Tg intact object novelty
at 7 months. Wt and Tg impaired object novelty
at 14 months.
Tg2576 Male & Female
14 months of age
2min, 30min, 24h
OR Tg intact OR at 2min, 30min. Wt and Tg may
be impaired at 24h.
(Hale & Good, 2005)
2min OL Tg impaired OL.
2min TO Tg intact recency.
Tg2576 Male 10-12 months
of age
2min OL Tg impaired OL.
(Good et al., 2007)
Episodic
Memory What-Where-When
Wt mice
preferred objects presented earlier in time
and in a different spatial location. Tg
only preferred objects presented less recently.
PS1 PS2
DKO
Male
& Female
8
months of age
4-month calorie restriction
(CR)
1h, 24h OR Tg impaired
OR at 1h and 24h. CR attenuated OR deficit in
Tg at 1h and 24h.
(Wu et al.,
2008)
PS1 L286V - 12-16 months of age
3h, 5h OR Tg facilitated OR at 3h. Tg impaired OR
(Vaucher et al., 2002)

128
at 5h.
PS1 L235P Male 6 months of age
5min OR Tg impaired OR.
(Huang, 2003)
APP/swe Male 7 months of age
7,8-DHF (TrkB agonist) 24h OL Tg mice impaired OL. 7,8-DHF
attenuated OL deficit .
(Bollen et al., 2013)
APP/PS1 Male & Female
10-12+ months of age
Camk2a-cre-Eif2ak3loxP/loxP transgene (PERK)
1h OL Tg impaired OL. Intact OL in APP/PS1
mice with PERK deletion.
(Ma et al., 2013)
hAPP/swe - 5, 8 months
of age
24h OR Tg intact OR at 5 months.
Tg impaired OR at 8 months.
(Simón et al., 2009)
TASTPM - 4,5,6,7,8
months of age
Fluparoxan (a2-adrenoceptor antagonist)
4h OR Tg and Wt and
fluparoxan mice had intact OR at
4; at 5 months OR was impaired in Tg mice and
fluparoxan Wt mice but intact in Wt and
fluparoxan Tg mice; at 6 months OR was impaired
in Wt and fluparoxan Tg mice but intact in
fluparoxan Wt mice and Tg mice.
(Scullion et al.,
2011)
TASTPM Male &
Female
3,4,6,8,10
months of age
4h OR Tg not impaired on
OR at 3 and 4 months; Tg impaired at
6,8 and 10 months
(Howlett et al.,
2004)
TASTPM Male & Female
5.5+ months
F- spondin (lentiviral vector-mediated overexpression of reelin
homolog) and green-fluorescent protein (GFP)
3h OR GFP treated Tg impaired OR. F-
spondin treated Tg intact OR.
(Hafez et al., 2012)
APP/PS1 Male 7+ months
of age
Sterol regulatory element-binding protein-2
(SREBP-2; involved in cholesterol homeostasis)
1h OR Tg mice overexpressin
g SREBP-2 impaired OR. Intact OR in Tg mice.
(Barbero-Camps et
al., 2013)
APP/PS1 Female 9+
months of age
GW3965 (liver X receptor
agonist) and ABCA1-/- (cholesterol transpoter ATP-binding cassette
4h OR Tg impaired
OR. GW3965 attenuated OR
(Donkin et
al., 2010)
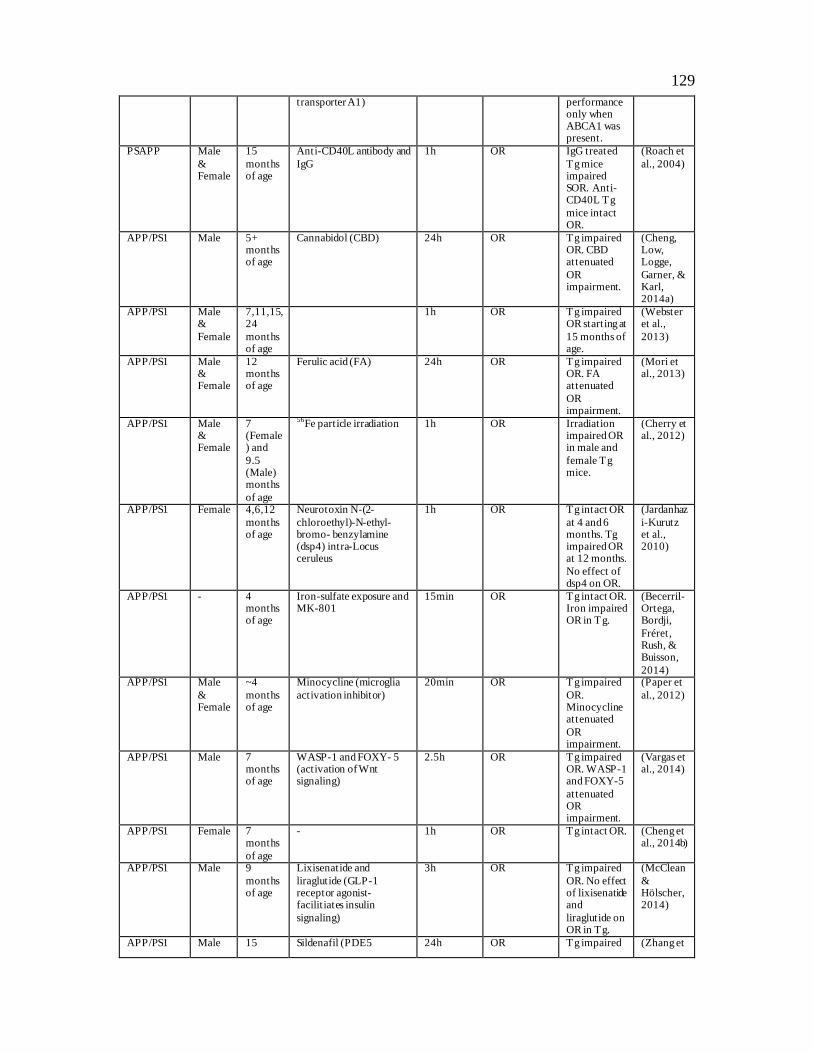
129
transporter A1) performance only when ABCA1 was present.
PSAPP Male
& Female
15
months of age
Anti-CD40L antibody and
IgG
1h OR IgG treated
Tg mice impaired SOR. Anti-CD40L Tg
mice intact OR.
(Roach et
al., 2004)
APP/PS1 Male 5+ months of age
Cannabidol (CBD) 24h OR Tg impaired OR. CBD attenuated
OR impairment.
(Cheng, Low, Logge,
Garner, & Karl, 2014a)
APP/PS1 Male &
Female
7,11,15,24
months of age
1h OR Tg impaired OR starting at
15 months of age.
(Webster et al.,
2013)
APP/PS1 Male & Female
12 months of age
Ferulic acid (FA) 24h OR Tg impaired OR. FA attenuated
OR impairment.
(Mori et al., 2013)
APP/PS1 Male & Female
7 (Female) and
9.5 (Male) months
of age
56Fe particle irradiation 1h OR Irradiation
impaired OR in male and
female Tg mice.
(Cherry et al., 2012)
APP/PS1 Female 4,6,12
months of age
Neurotoxin N-(2-
chloroethyl)-N-ethyl-bromo- benzylamine (dsp4) intra-Locus ceruleus
1h OR Tg intact OR
at 4 and 6 months. Tg impaired OR at 12 months.
No effect of dsp4 on OR.
(Jardanhaz
i-Kurutz et al., 2010)
APP/PS1 - 4 months of age
Iron-sulfate exposure and MK-801
15min OR Tg intact OR. Iron impaired OR in Tg.
(Becerril-Ortega, Bordji,
Fréret, Rush, & Buisson,
2014) APP/PS1 Male
& Female
~4
months of age
Minocycline (microglia
activation inhibitor)
20min OR Tg impaired
OR. Minocycline attenuated
OR impairment.
(Paper et
al., 2012)
APP/PS1 Male 7 months of age
WASP-1 and FOXY- 5 (activation of Wnt signaling)
2.5h OR Tg impaired OR. WASP-1 and FOXY-5
attenuated OR impairment.
(Vargas et al., 2014)
APP/PS1 Female 7 months
of age
- 1h OR Tg intact OR. (Cheng et al., 2014b)
APP/PS1 Male 9
months of age
Lixisenatide and
liraglutide (GLP-1 receptor agonist-facilit iates insulin
signaling)
3h OR Tg impaired
OR. No effect of lixisenatide and
liraglutide on OR in Tg.
(McClean
& Hölscher, 2014)
APP/PS1 Male 15 Sildenafil (PDE5 24h OR Tg impaired (Zhang et

130
months of age
inhibitor) and Rp-8-Br-PET-cGMPS (c-GMP-dependent protein kinase)
OR. Sildenafil, without Rp-8-Br-PET-
cGMPS, attenuated OR
impairment in Tg.
al., 2013)
APP/PS1 Male & Female
7 months of age
Donepezil (acetylcholinesterase inhibitor) and naltrindole
(opoid receptor antagonist)
1h OR Tg mice impaired OR. Donepezil
and naltrindole attenuated OR in Tg.
(Zhang et al., 2012)
APP/PS1 Female 12
months of age
Ferulic acid (anti-oxidant
and anti-inflammatory)
24h OR Tg mice
impaired OR. Ferulic acid attenuated OR
impairment in Tg.
(Jing et
al., 2013)
PSAPP Male & Female
6, 12 months of age
Flavonoid tannic acid (TA)
24h OR Tg impaired OR. Flavonoid
attenuated OR impairment in Tg.
(Mori et al., 2012)
APP/PS1 Male 7
months of age
Liraglutide (incretin
hormone glucagon-like peptide-1 analog)
3h OR Tg impaired
OR. Liraglutide attenuated OR deficit in
Tg.
(McClean
et al., 2011)
APP/PS1 Male & Female
8 months of age
Prescription number 1 (PN-1) (Chinese medicine) and donepezil
24h OR Tg impaired OR. PN-1 and donepezil attenuated
OR deficit in Tg.
(Yao et al., 2013)
APP/PS1 - 7 months of age
Memantine (NMDA receptor antagonist)
3h OR Tg impaired OR. Memantine
attenuated OR deficit in Tg.
(Scholtzova et al., 2008)
APP/PS1 - 8 months
of age
Aβ1–42-containing transcutaneous
immunization
1h OR Tg impaired OR. Aβ
immunization attenuated OR deficit in Tg.
(Matsuo et al., 2014)
APP/PS1 - 3-6
months of age
3h OR Tg impaired
OR.
(Dewachte
r et al., 2002)
APP/PS1 - 9-12 months of age
Progesterone (P4) 4h OL Tg impaired OL.
(Frye & Walf, 2008) OR P4 improved
OR in Wt and
Tg. APP/PS1 Male 4,6,8
months of age
80 min OL Tg intact OR
at 4 months. Tg and Wt impaired OR
at 6, 8 months.
(Maroof et
al., 2014)
OR Tg intact OL

131
at 4, 6, 8 months. Wt impaired OL at 6, 8
months.
5xFAD Female ~2 months of age
RS 67333 (5-HT4 receptor agonist)
1h OR Tg impaired OR. RS attenuated OR deficit in
Tg.
(Giannoni et al., 2013)
5xFAD Male 4-7 months of age
Kamikihi-to (KKT; traditional Japanese medicine)
30min OR Tg impaired OR. KKT attenuated OR deficit in
Tg.
(Tohda et al., 2011)
5xFAD Female 6-8 months
of age
diosgenin (steroidal sapogenin; Regenerates
neurite atrophy and syaptic loss) and memantine
24h OR Tg impaired OR.
Diosgenin attenuated OR deficict in Tg.
Memantine did not alter OR performance
in Tg.
(Tohda et al., 2012)
5xFAD Male 6-8 months of age
Sominone (steroidal sapogenin)
30min OR Tg impaired OR. Sominone attenuated
OR deficit in Tg.
(Joyashiki et al., 2011)
5xFAD Female 6.5 months of age
Neuronal overexpression of MMP-9 (amatrixmetalloproteinase
critically involved in neuronal plasticity, acts as α-secretase)
15min OR Tg impaired OR. MMP-9 attenuated
OR impairment in Tg.
(Fragkouli et al., 2014)
5xFAD - 12 months
of age
24h OR Tg impaired OR.
(Wang et al., 2013)
5xFAD - 6 months of age
Cinnamon extract - OR Tg impaired OR. Cinamon extract attenuated
OR deficit in Tg.
(Frydman-Marom et al., 2011)
Tg4510 - 6 months of age
Calorie restriction (CR) 5min OR Tg impaired OR. CR attenuated
OR impairment in Tg.
(Brownlow et al., 2014)
htau Male 4, 12 months
of age
5min, 30min
OR Tg intact OR at 4 months.
Tg impaired OR at 12 months.
(Polydoro, Acker,
Duff, Castillo, & Davies, 2009)
PLB1 4, 12
months of age
5min OR Tg impaired
OR at 4, 12 months
(Ryan et
al., 2013)
OL Tg impaired OL at 4, 12 months
PLB1 Male
& Female
8, 12
months of age
2min OR Tg impaired
OR at 8, 12 months.
(Platt et
al., 2011)

132
OL Tg impaired OL at 12 months.
3xTG Female 6 months
of age
Streptozotocin (STZ; a dibetogenic compound)
15min OR Tg impaired OR. STZ
exacerbated OR deficit in Tg.
(Chen et al., 2014)
3xTG - 5, 6 months
of age
PD098059 (A MEK inhibitor)
24h OR Tg impaired OR at 5
months. PD098059 attenuated OR
impairment in Tg at 6 months.
(Feld et al., 2014)
3xTG Female 12-14 months
of age
3h OR Tg impaired OR
(Filali et al., 2012)
3xTG Male ~11 months of age
2-methyl-5-(3-{4-[(S)-methylsulfinyl] phenyl}-1-benzofuran-5-yl)-1,3,4-oxadiazole (MMBO;
GSK-3 inhibitor)
5h OR Tg impaired OR. MMBO attenuated OR deficit in
Tg,
(Onishi et al., 2011)
3xTG - 12 months of age
Minocycline (anti-inflammatory)
1.5h, 24h OR Tg impaired OR. Minocycline improved OR
in TG.
(Parachikova, Vasilevko, Cribbs,
LaFerla, & Green, 2010)
3xTG Female 7+ months
of age
Peptide 6 (an 11-mer peptide which activates
ciliary neurotrophic factor)
15min OR Tg impaired OR. Peptide 6
attenuated OR impairment in Tg.
(Blanchard et al.,
2010)
3xTG Male 5, 10
months of age
Nomifensine (dopamine
reuptake inhibitor)
24h OR Tg intact OR
at 5 months. Tg impaired OR at 10 months.
Nomifensine attenuated OR deficit in
Tg at 10 months.
(Guzmán-
Ramos et al., 2012)
3xTG Female 15-16 months of age
P021 (ciliary neurotrophic factor)
24h OR Tg impaired OR. P021 attenuated
OR deficit in Tg.
(Kazim et al., 2014)
3xTG Female 12, 16 months
Immunoglobulin (IVIg) 1h OR Tg impaired OR at 12 and 16 months.
IVIg attenuated OR deficit at 16 months.
(St-Amour et al., 2014)
3xTG - 14
months of age
Docosahexaenoic acid
(DHA)
1h OR Tg impaired
OR. DHA attenuated OR impairment in
Tg.
(Arsenault
et al., 2011)
3xTG Male & Female
6.5 months of age
15min OR Wt and Tg impaired OR.
(Stover et al., 2015)

133
3xTG Male & Female
15-18 months of age
30-60s OR Tg intact OR. (Gulinello et al., 2009) 3min OL Tg impaired
OL.
3xTG Female 11 months
of age
2min OR Tg intact OR. (Davis et al., 2013)
OL Tg impaired OL.
9 months of age
5min What-Where Tg impaired egocentric What-Where.
Tg and Wt impaired allocentric What-Where
12
months of age
2min What-Which Tg intact
What-Which
6 months of age
2min, 5min, 10min,
15min, 30min
Episodic memory What-Where-
Which
Tg impaired What-Where-Which at all
delays. Wt impaired What-Where-Which at
15min and 30min.
14 months of age
2min Episodic memory What-Where-
When
What-Where-When intact in Wt and Tg,
but Tg impaired relative to Wt.
3xTG Female 4.5
months of age
5min TO Tg intact TO. (Davis et
al., 2013)
3, 6, 12 months of age
2min, 5min 10min, 15min,
30min
What-Where-Which
Tg impaired What-Where-Which at 3, 6,
12 months.
5-8 months of age
10min, 20min
What-Where-When
Tg intact What-Where-When
3xTG & PS1-KIM146V
Male & Female
12 months of age
Pioglitazone (PIO; TZD- aimed to improve insulin signaling)
24h OL PS1 impaired OL. PIO improved OL
in PS1. 3xTG impaired OL. PIO did not alter OL
performance in 3xTG.
(Masciopinto et al., 2012)
human APOC11/0
Female 12 months of age
1h OR Tg impaired OR.
(Abildayeva et al., 2008)
ApoE-/- Male 3-5
months of age
Thioperamide and
clobenpropit (H3 antagonists)
24h OR Tg intact OR.
H3 antagonists impaired OR in Tg.
(Bongers,
Leurs, Robertson, & Raber, 2004)
Apoc1–/– Female ~12
months of age
1h OR Tg impaired
OR.
(Berbée et
al., 2011)
ApoE3 and ApoE4 mice
Male 4+ months of age
Docosahexaenoic acid (DHA)
24h OR ApoE4 impaired OR. DHA
attenuated OR
(Kariv-Inbal et al., 2012)

134
impairment in ApoE4.
ApoE3, ApoE4, and Apoe-/-
Male 6-7 months
Castration 5min OR Castration impaired OR in Apoe-/-.
(Pfankuch, Rizk, Olsen,
Poage, & Raber, 2005)
OL Castration
impaired OL in ApoE4.
APP-Yac/apoE3-TR
(’apoE3’) and APP-Yac/apoE4-TR
(’apoE4’) mice
Female 6-7 months of age
24h OL Impaired OL in TG APP/apoE4,
but not APP/apoE3.
(Kornecook et al., 2010)

135
Table 3
Correlation Between Total Sample Exploration and Novelty Preference Index
Note. Pearson product-moment correlation coefficients between total sample exploration
and novelty preference index (discrimination ratio or oddity preference) are not significant unless otherwise reported. * p < .05, ** p < .001
Correlation Coefficient (r)
Experiment WT TG
Strain Task Sex Delay
5xFAD O -OR Female 5min .169 .400
3h -.206 .537
Male 5min .320 .298
3h .042 .024
Y-O R Female 5min -.154 .653*
3h .502 -.037
Male 5min .031 .014
3h .188 -.195
O L Female 5min .164 .143
3h .334 .047
Male 5min .545 -.052
3h .469 .195
Temporal O rder
Female 3min .584* .648*
Male 3min
MSO Female -.418 -.822**
Male -.286 -.051
Visual Oddity Female .169 -.252
Male .737** -.154
Tactile Oddity Female .124 -.100
Male .189 .073
3xTG O -OR Female 5min -.192 -.398
3h -.330 .363
Male 5min -.385 .587
3h -.121 -.367
Y-O R Female 5min .379 -.646*
3h -.006 -.492
Male 5min .106 -.338
3h .273 -.113
O L Female 5min .267 -.167
3h -.193 -.291
Male 5min .090 .039
3h -.135 -.200
Temporal
O rder
Female 3min -.146 -.220
Male 3min
MSO Female -.178 .347
Male .296 .532
Visual Oddity Female .235 .070
Male .500 -.042
Tactile Oddity Female .566* -.254
Male .091 .469

136
Table 4
Object Processing Results Summary
Note. Individual comparisons between sample DR and choice DR for mnemonic
paradigms as well as chance performance and sample OP for perceptual oddity tasks (
indicates intact performance, indicates impaired performance).
3xTG 5xFAD
WT TG WT TG
Female Male Female Male Female Male Female Male
O R O pen-
field OR 5min
3h
Y-O R 5min
3h
O L 5min
3h Familiarity Preference
TO 3min
Not completed
Not completed
Not completed
Not completed
O ddity MSO
Visual
Tactile
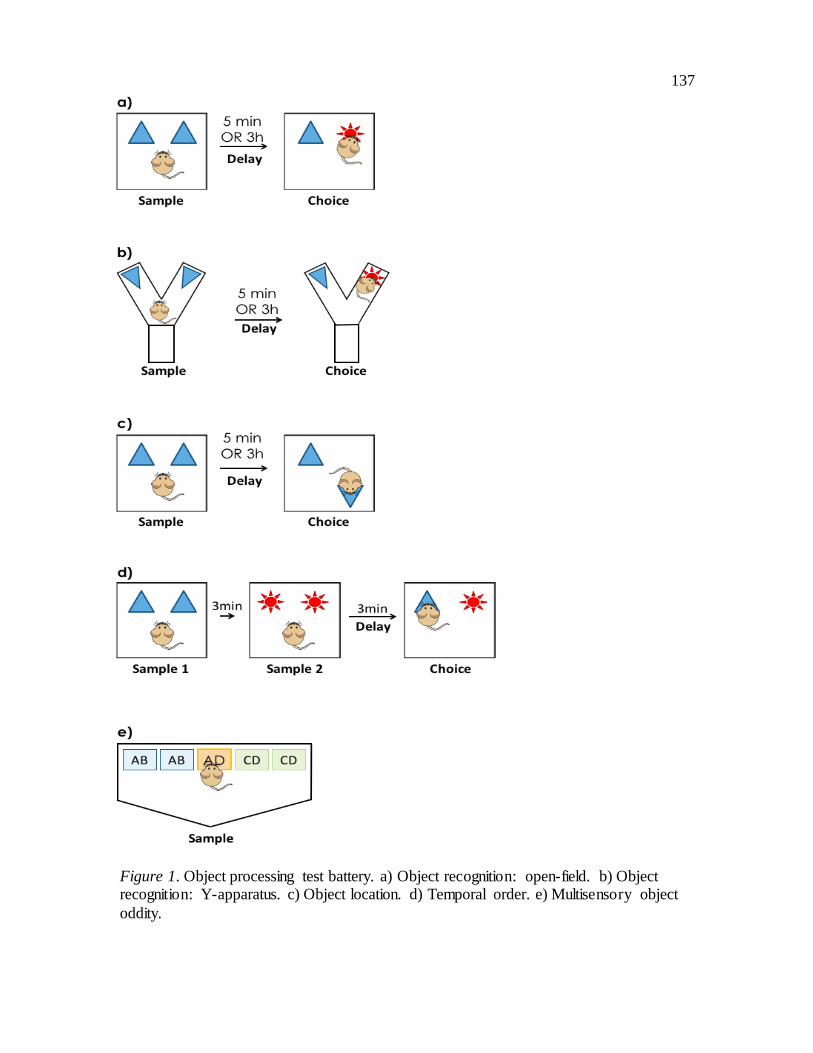
137
Figure 1. Object processing test battery. a) Object recognition: open-field. b) Object recognition: Y-apparatus. c) Object location. d) Temporal order. e) Multisensory object
oddity.

138
Figure 2. 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) open-field object recognition. (A) Female 5xFAD mice did not discriminate significantly
between novel and familiar objects in either retention delay condition. (B) Male 5xFAD mice did not discriminate significantly between novel and familiar objects in either retention delay condition. (C) Female 3xTG mice did not discriminate significantly
between novel and familiar objects in the 3h retention delay condition. (D) Male 3xTG mice did not discriminate significantly between novel and familiar objects in either
retention delay condition. *p < .05, **p < .01 ***p < .001
Female Male
5xFAD
3xTG
A B
C D
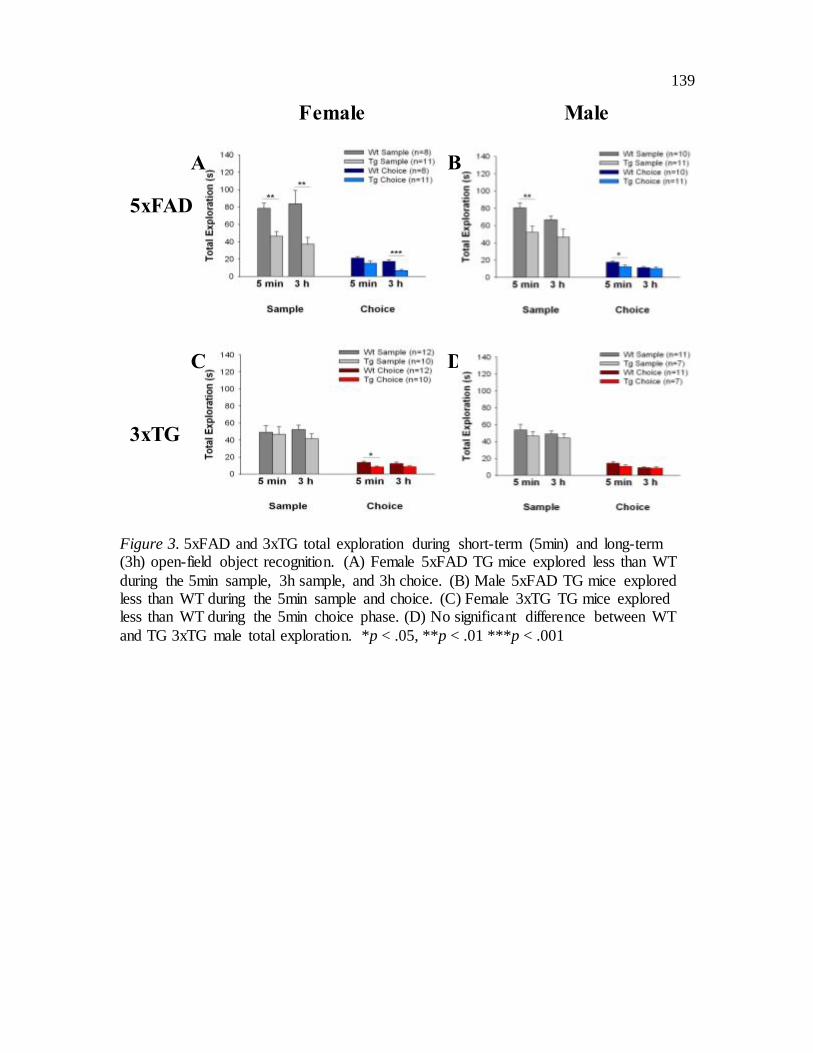
139
Figure 3. 5xFAD and 3xTG total exploration during short-term (5min) and long-term (3h) open-field object recognition. (A) Female 5xFAD TG mice explored less than WT
during the 5min sample, 3h sample, and 3h choice. (B) Male 5xFAD TG mice explored less than WT during the 5min sample and choice. (C) Female 3xTG TG mice explored less than WT during the 5min choice phase. (D) No significant difference between WT
and TG 3xTG male total exploration. *p < .05, **p < .01 ***p < .001
Female Male
5xFAD
3xTG
A B
C D

140
Figure 4. 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) Y-apparatus object recognition. (A) Female 5xFAD mice did not discriminate significantly
between novel and familiar objects in either retention delay condition. WT mice did not discriminate between not discriminate between novel and familiar object at 3h. (B) Male
5xFAD mice did not discriminate significantly between novel and familiar objects in either retention delay condition. (C) Female 3xTG mice did not discriminate significantly between novel and familiar objects in the 3h retention delay condition. (D) Male 3xTG
mice did not discriminate significantly between novel and familiar objects in the 3h retention delay condition. **p < .01 ***p < .001
Female Male
5xFAD
3xTG
A B
C D

141
Figure 5. 5xFAD and 3xTG total exploration during short-term (5min) and long-term (3h) Y-apparatus object recognition. (A) No significant difference between WT and TG
5xFAD female total exploration. (B) Male 5xFAD TG explored more than WT during the 3h choice phase. (C) No significant difference between WT and TG 3xTG female total exploration. (D) No significant difference between WT and TG 3xTG male total
exploration. *p < .05
Female Male
5xFAD
3xTG
A B
C D

142
Figure 6. 5xFAD and 3xTG performance on short-term (5min) and long-term (3h) object
location. (A) Female 5xFAD mice did not discriminate significantly between novel and familiar object spatial locations at 5min or 3h. (B) Male 5xFAD mice did not
discriminate between novel and familiar object spatial locations at 5min. WT mice did not discriminate between novel and familiar object spatial locations at 5min and 3h. (C) 3xTG females did not discriminate between novel and familiar object spatial locations at
5min or 3h. (D) 3xTG males did not discriminate between novel and familiar object spatial locations at 5min. WT mice did not discriminate between novel and familiar
object spatial locations at 3h. At 3h TG mice had a significant familiarity preference. *p < .05, **p < .01 ***p < .001
Female Male
5xFAD
3xTG
A B
C D

143
Figure 7. 5xFAD and 3xTG total exploration during short-term (5min) and long-term (3h) object location. (A) Female 5xFAD TG explored less than WT during the 3h choice
phase. (B) No significant difference between WT and TG 5xFAD male total exploration. (C) Female 3xTG TG explored less than WT during the 3h sample. (D) Male 3xTG TG explored more than WT during the 5min sample and choice. *p < .05, ***p < .001
Female Male
5xFAD
3xTG
B
D
A
C

144
Figure 8. Female 5xFAD and 3xTG performance on temporal order. (A) Female 5xFAD
TG and WT did not differentiate between novel and familiar stimuli at 3min. (B) 3xTG females were impaired on temporal order at 3min. *p < .05, **p < .01
Female
5xFAD
3xTG
A
B

145
Figure 9. Female 5xFAD and 3xTG total exploration during object temporal order. (A) Female 5xFAD TG explored less than WT during sample phase 1. (B) No significant difference between WT and TG 3xTG female total exploration. **p < .01
Female
5xFAD
3xTG
A
B

146
Figure 10. 5xFAD and 3xTG performance on object oddity. (A) 5xFAD females were
selectively impaired on MSO. (B) 5xFAD males were impaired on MSO, visual, and tactile oddity. WT males were impaired on MSO and tactile oddity. (C) 3xTG females were selectively impaired on MSO. (D) 3xTG males were selectively impaired on MSO.
WT males were impaired on visual oddity. *p < .05, **p < .01 ***p < .001
Female Male
5xFAD
3xTG
A B
C D

147
Figure 11. 5xFAD and 3xTG total exploration during object oddity. (A) Female 5xFAD TG explored more than WT during MSO, visual, and tactile oddity. (B) No significant difference between WT and TG 5xFAD males. (C) No significant difference between
WT and TG 3xTG females. (D) No significant difference between WT and TG 3xTG males. *p < .05, **p < .01
Female Male
5xFAD
3xTG
A B
C D





























