Embed Size (px)
Citation preview

Behavioral Ecology Vol. 14 No. 6: 823–840DOI: 10.1093/beheco/arg072
Ecological and behavioral correlates ofcoloration in artiodactyls: systematic analysesof conventional hypotheses
C. J. Stoner, T. M. Caro, and C. M. GrahamDepartment of Wildlife, Fish, and Conservation Biology, University of California, Davis,CA 95616, USA
To test the generality of adaptive explanations for coat coloration in even-toed ungulates, we examined the literature forhypotheses that have been proposed for color patterns exhibited by this taxon, and we derived a series of predictions from eachhypothesis. Next, we collected information on the color, behavioral, and ecological characteristics of 200 species of even-toedungulates and coded this in binary format. We then applied chi-square or Fisher’s Exact probability tests that pitted presence ofa color trait against presence of an ecological or behavioral variable for cervids, bovids, and all artiodactyls. Finally, we reanalyzedthe data by using concentrated-changes tests and a composite molecular and taxonomic phylogeny. Hinging our findings onwhether associations persisted after controlling for shared ancestry, we found strong support for hypotheses suggesting even-toedungulates turn lighter in winter to aid in concealment or perhaps thermoregulation, striped coats in adults and spotted pelage inyoung act as camouflage, side bands and dark faces assist in communication, and dark pelage coloration is most common inspecies living in the tropics (Gloger’s rule). Whereas white faces, dark legs, white legs, dark tails, and white tails did not appear toassist in communication alone, legs and tails that were either dark or white (i.e., conspicuous) did seem to be linked withcommunication. There was moderate support for hypotheses that countershading aids concealment, that white faces area thermoregulatory device, and that white rumps are used in intraspecific communication. There was weak support for spots inadults and stripes in young providing camouflage and for dark leg markings being a form of disruptive coloration. We found littleor no evidence that overall coat color serves as background matching, that side bands are disruptive coloration devices, or thatwhite rumps help in thermoregulation. Concealment appears the principal force driving the evolution of coloration in ungulateswith communication, and then thermoregulation, playing less of a role. Key words: bovids, cervids, communication, concealment,countershading, disruptive coloration, pattern blending, phylogeny, thermoregulation, ungulates. [Behav Ecol 14:823–840(2003)]
The adaptive significance of coloration in ungulates hasnever been addressed systematically. Indeed, for mam-
mals, many of the current hypotheses about coloration arestill the same ones formulated more than 100 years ago(Beddard, 1892; Buxton 1923; Mottram, 1915, 1916; Poulton,1890; Roosevelt, 1911; Thayer, 1909; Wallace, 1889) and havereceived remarkably little attention ever since (but seeOrtolani, 1999). Hypotheses to explain the function ofcoloration in mammals can be grouped into three broadcategories: concealment, communication, and regulation ofphysiological processes (Cott, 1940).
Animals can remain concealed when their overall colorresembles, or matches, the natural background in which theylive (Endler, 1978). This phenomenon of backgroundmatching, also known as general color resemblance, includescrypsis in which overall body color resembles the generalcolor of the habitat, or pattern blending, in which colorpatterns on the body match color patterns of light and dark inthe environment (Cott, 1940; Poulton, 1890). Backgroundmatching may be subtle in that an individual may change itspelage or skin coloration annually if backgrounds changebetween seasons (also termed variable background match-ing), or may vary with age if the background environments ofimmatures and adults differ in predictable ways. Alternatively,
animals may achieve concealment through disruptive colora-tion by sporting contrasting colors or irregular marks thatbreak up the body’s outline (Cott, 1940). Lastly, animalsmay attain concealment by having a pelage that lightenstoward the ventral surface. This counteracts the sun’s effect oflightening the dorsum and shading the ventrum when itshines from above (Kiltie, 1988, 1989; Thayer, 1896, 1909).All of these types of coloration are found in even-toedungulates.
When coloration is conspicuous, as in the black andwhite faces of oryx species, intraspecific communication(Wickler, 1968) or communication between prey andpredators is thought to be involved (Rowland, 1979). Forexample, patches of color may be used to signal danger toconspecifics (Alvarez et al., 1976) or to signal young to followtheir mothers (Leyhausen, 1979). Alternatively, they may beused to signal to predators, the classic example beingaposematic coloration in species that have noxious defenses,such as skunks (Spilogale putorius) (Guilford, 1990; Johnson,1921). Many ungulates exhibit conspicuously coloredheads, legs, and anal regions (Guthrie, 1971; Meyer andKranzle, 1999).
Finally, coloration may be related to thermoregulation orprotection against ultraviolet rays (Hamilton, 1973). Thus,dark-colored animals may be expected to absorb solarradiation better than light-colored animals do and mighttherefore be found in colder climates (Burtt, 1979). Even-toedungulates exhibit a range of hues from black through grayand red to pale. In contrast to birds, none of the majorhypotheses for coloration in mammals pertains to matechoice (see Badyaev and Hill, 2000).
Address correspondence to C.J. Stoner. E-mail: [email protected].
Received 17 August 2001; revised 16 January 2002; accepted 21January 2003.
� 2003 International Society for Behavioral Ecology
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

Within these three broad categories, a number of morespecific hypotheses have been advanced for overall bodycoloration and specific parts of the body in artiodactyls(Table 1). Most of these functional hypotheses for colorationwere formulated by using one or a handful of ungulate species,and are based on qualitative arguments. Although they may wellapply to the species in question, the extent to which they are
generally applicable to other species in the order Artiodactylais completely unknown. The purpose of this study was to testthese conventional hypotheses systematically in order todetermine their generality within the order. Our main focuswas on overall body coloration and color of specific parts ofthe body rather than a constellation of body parts such as‘‘contrasting markings’’ (see Geist, 1978) because hypotheses
Table 1
Summary of suggested hypotheses for the adaptive significance of coloration in even-toed ungulates andexamples
Hypothesis,Example Source
Overall body coloration
Uniform coloration and background matching 1, 2Reddish coat matches rocky habitats in markhor
Uniform coloration and thermoregulation 2Pale coat minimizes heat exposure in hartebeest in desert
Lighter coats in winter and background matching 3Seasonal color changes reduces conspicuousness in red deer in snowy arctic
Spots on adults and pattern blending 3Dappled coat provides camouflage for giraffe in dappled light of woodland
Spots on adults and reduced aggression 4Dappled coat assists in lowering aggression in axis deer living in groups
Stripes on adults and pattern blending 5, 6Striped coat in greater kudu blends in with riverine habitat
Stripes on adults and reduced aggression 7Striped markings in group-living eland lower aggressive interactions
Spots on young and pattern blending 5, 8Dappled coat in young mule deer provides camouflage in dappled light of woodland
Stripes on young and pattern blending 1Striped coat in young red river hog aids concealment in grassy vegetation
Side bands and disruptive coloration 6Dark stripe breaks up springbok’s form in open environment
Side bands and maintaining gregariousness 4Side bands assist Thomson’s gazelle in maintaining group cohesion
Countershading and concealment 3, 9Light ventrum in the diurnal dibitag counteracts shading from the sun
Body parts
Dark face markings and communication 2, 10Black face assists in signaling to group members in oryx
White face markings and communication 11White face assists in signaling to group members in vicuna
White face markings and thermoregulation 12White face helps pronghorn reflect heat in desert
Conspicuous face and communication 13Black and white face in impala assists in signaling to conspecifics
Dark leg markings and disruptive coloration 14Dark leg markings in lechwe break up form in open environments
Dark leg markings and communication 2, 6, 12Black knees assist in signaling to group members in addax
White leg markings and communication 12, 15, 16White legs in bontebok act as visual signals in daylight
Conspicuous leg markings and communication 4White legs in okapi signal to stalking predators
Dark tail and communication 1Black tail in Grant’s gazelle signals to coursing predators
White tail and communication 1, 6, 17White tail in blue duiker is prominent at night
Conspicuous tail and communication 18White tail in white-tailed deer signals to other group members
White rump and communication 1, 6, 19White rump in Soemmering’s gazelle signals to other group members
White rump and thermoregulation 2, 6White rump in bighorn sheep acts to deflect heat in desert
Sources: 1Kingdon, 1982; 2Spinage, 1986; 3Cott, 1940; 4J. Wolff, personal communication; 5Thayer, 1909;6Estes, 1991; 7Lobao Tello and van Gelder, 1975; 8Linnell and Andersen, 1998; 9Fogden and Fogden,1974; 10Hasson, 1986; 11Poulton, 1890; 12Geist, 1987; 13Guthrie and Petocz, 1970; 14Roosevelt and Heller,1914; 15Kingdon, 1997; 16Danilkin, 1996; 17Bildstein, 1983; 18Geist, 1978; 19Ward, 1979.
824 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

have, in general, been formulated for specific areas of thebody in this order and our data set allowed us to test suchhypotheses at this relatively fine-grained level. The value ofthe comparative approach used here is that it allows us toinvestigate the adaptive significance of morphological traits byusing a very large number of species, and make conclusionsthat have wide generality. Similar comparative analyses oncoloration in birds have been successful in opening up debateon coloration signaling unprofitability to predators (Bakerand Parker, 1979), on linking coloration to lekking (Hoglund,1989), and in explaining patches of white plumage in waders(Brooke, 1998).
METHODS
Species
Recent discoveries of new species in the order Artiodactyla(see Groves and Schaller, 2000), a growing number ofphylogenetic studies focusing on this order, and controversiesregarding the distinctness of numerous species and sub-species have led to continual changes in artiodactyl taxono-mies. We started analyses based on the species listed in Nowak(1999) and refined this list to exclude (1) purely domesticat-ed species; (2) species that are, or were at one time, extinct inthe wild; and (3) species for which very little morphological orbehavioral data could be found. Purely domestic species, suchas domestic cattle (Bos taurus), domestic goats (Capra hircus),and domestic sheep (Ovis aries), were excluded because theirpelage coloration may reflect artificial selection rather thanadaptations to natural environments. The llama (Lama glama)and alpaca (Lama pacos) were similarly excluded as Nowak(1999) and Franklin (1982) note that these are generallythought to be domestic descendants of the wild guanaco(Lama guanicoe). Dromedary camels (Camelus dromedarius),were excluded because some taxonomists (see Corbet andHill, 1991, 1992) believe they never existed in the wild (seeNowak, 1999). Although we included some species that arefound both in the wild and in domestic or feral populations(e.g., wild boars [Sus scrofa], Bactrian camels [Camelusbactrianus], banteng and domestic Bali cattle [Bos javanicus],guar [Bos guarus], and yaks [Bos grunniens]), we focused onthe coloration or ecological characteristics of wild populationsfor our analyses. One such species, the water buffalo (Bubalusbubalus), was excluded because some taxonomists feel thisscientific name refers only to domestic populations of thespecies (see Corbet and Hill, 1991, 1992 and discussion inNowak, 1999).
To be conservative, we excluded species that are currently,or were at one time, extinct in the wild: the cave goat,Myotragus balearicus; three gazelle species, Gazella arabica,Gazella rufina, and Gazella saudiya; the blue buck, Hippotragusleucophaeus; and Schomburgki’s deer, Cervus schomburgki. Wesimilarly excluded Pere David’s deer, Elaphurus davidianus,European bison, Bison bonasus, and the Arabian oryx, Oryxleucoryx, because at one time they existed only in captiveenvironments (see Nowak, 1999). Thus, the species retainedin our analyses are currently, and have presumably alwaysbeen, extant in the wild.
Finally, we excluded species for which little information onpelage coloration or behavior was available. For example,information regarding Pseudonovibos spiralis is based primarilyon horns and accounts from local hunters. Gazella bilkis andNaemorhedus baileyi were also excluded, as their descriptionsare based on rare specimens collected in the past (seeGrzimeck, 1990; Nowak, 1999). As a result of this selectionprocess, our analyses were based on a total of 200 artiodactylspecies (see Supplementary Material online).
Data collection
Information on overall body coloration, seasonal colorationchange, and coloration of underparts was obtained fromdescriptions in the literature; face, leg, rump, and tailcoloration was determined from photographs (Table 2 andappendix). Information on ecological characteristics wastaken from descriptions in the literature and was placed intocategories; latitude characteristics were determined fromrange maps (Table 3). With a few exceptions, we principallylooked for associations between coloration and ecologicalrather than behavioral variables because behavioral hypothe-ses for a color marking, such as ‘‘follow-me’’ signals, aredifficult to test. Such signals might apply between mother andyoung (Schaller, 1967), between members of small groups onthe move (Hickman, 1979), or between members of largesocial groups; they are more amenable to tests through directobservation.
For all variables, a value of one was assigned to speciesdemonstrating a given trait, zero to species that did notdisplay the trait, and a ‘‘?’’ for species for which little or noinformation on the variable was available (supplementaryappendix online). Variables were scored present or absent asvariation in pelage characteristics and ecological and behav-ioral characters was unknown for most species. Categorieswere not mutually exclusive. For example, a species demon-strating light red coloration would be assigned a value of onein both the ‘‘pale’’ and ‘‘red’’ color categories.
Analyses
Nonparametric testsTo identify general trends between coloration traits andecological variables, we first applied chi-squared tests sepa-rately to the two largest families, cervids (N ¼ 39 species) andbovids (N ¼ 125 species), to identify associations that werepresent within particular families. When sample sizes wereinadequate to perform chi-squared statistics, we instead usedFisher’s Exact probability tests. We then performed theseanalyses for all the artiodactyls (N ¼ 200 species) in order toinclude important families such as the Suidae and Giraffidaeand to determine whether associations held across the wholeorder. For each test, we included only those species for whichinformation was available for the coloration and ecologicalvariables of interest. Only significant results are reported inthe text (p , .1 because we were searching for associationsusing very coarse behavioral and ecological measures). We feltit important to present cross-species comparisons in light ofthe current controversy over the applicability of phylogenet-ically controlled methods (Irschick et al., 1997; Price, 1997),although in our discussion we attach far more weight toresults that controlled for shared ancestry (see below).
Phylogenetic comparisonsCross-species comparisons fail to account for the fact thatspecies values are nonindependent: shared character statesmay reflect common ancestry rather than independentadaptations (Harvey and Pagel, 1991). To control for thepotential effects of shared ancestry, we analyzed the samehypotheses as in the nonparametric tests by using a compositephylogenetic tree and Maddison’s concentrated-changes test(Maddison, 1990) as implemented in MacClade (Maddisonand Maddison, 1992). Concentrated-changes tests have beenreported to detect such correlated changes statistically basedon fewer character state changes compared with that of othermethods (Cooper, 2002; Cooper et al., 2002; Ridley, 1983;Sillen-Tullberg, 1993). Although concentrated-changes testsrely on the topology of a given phylogeny rather than on
Stoner et al. • Color of artiodactyls 825
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

incorporating branch lengths estimates like more rigorousmethods (see Nee et al., 1996; Pagel, 1994; Read and Nee,1995), we deemed these tests appropriate for our analysesowing to the scarcity of branch length and divergence timeestimates for ungulate taxa (Perez-Barberıa and Gordon 1999,2000, 2001; Perez- Barberıa et al. 2002).
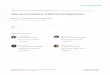
Figure 1 demonstrates the hypothesized phylogenetic treeon which analyses were based. Owing to the lack of consensusover the placements of Artiodactyla taxa in the literature, webased this composite tree primarily on the findings of recentmolecular studies and supplemented these with taxonomiesand studies based on morphological traits. The generalpositions of the Cervidae, Bovidae, Giraffidae, Antilocapridae,Tragulidae, Hippopotamidae, Suidae, and Camelidae werebased on cladistic analyses by Gatesy et al. (1999). Additionalsources were used to resolve the genus Moschus (Su et al.,1999), the Tragulidae (based on taxonomy in Grubb, 1993),Suidae (Groves, 1981; Randi et al., 1996; Thenius, 1970),Peccaridae (Theimer and Keim, 1998), and Camelidae (basedon Gatesy et al., 1999: Figures 11 and 12).
Cervidae branch. The general positions of the Cervinae-Muntiacus, Hydropotes-Alces-Capreolus, and Rangifer-New Worldodocoileinae clades were inferred from Cronin et al. (1996),Douzery and Randi (1997), Gatesy et al. (1999), and Randiet al. (1998). Resolution within Cervinae-Axis-Dama was basedon Emerson and Tate (1993) and Randi et al. (2001), theElaphodus-Muntiacus clade was based on Amato et al. (2000),and the placements of Hydropotes, Alces and Capreolus were
based on Randi et al. (1998). The placements of Hippocamelus,Pudu, Blastoceros, and Ozotoceros (in relation to Rangifer,Odocoileus, and Mazama) were based on Webb (2000).
Bovidae branch. All information gained from the simulta-neous analysis of data sets in Gatesy and Arctander (2000) wasretained in the Bovidae tree. Gatesy and Arctander (2000) wasused to infer the basic branches within Bovidae andsupplemented with information from the following sources.Resolution within the bovini and tragelaphini tribes was basedon Schreiber et al. (1999) (Bulbus, Bos), Gatesy and Arctander(2000) (Syncerus, Psuedorx, Boselaphus), Hassanin and Douzery(1999a,b) (Tetracerus quadricornis in clade with Boselaphustragocamelus), and Mathee and Robinson (1999) (Tragelaphus,Taurotragus). Resolution within the Antilopini clade was basedon Gatesy et al. (1999), Mathee and Robinson (1999),Rebholz and Harley (1999) (general positions of Rachicerusand Madoqua clades), and Brashares et al. (2000) (resolutionwithin the Rachicerus and Madoqua clades). Resolution withinthe Gazella-Antilope clade was based on Rebholz and Har-ley’s (1999) parsimony cladogram. Placements of Litocraniuswalleri, Ammodorcas, Antidorcus, Dorcatragus were based onBrashares et al. (2000). Within the Alcelaphini, the position ofthe Neotragus/Aepyceros clade (Hassanin and Douzery, 1999b;Mathee and Robinson, 1999) was based on Gatesy andArctander (2000). Resolution within Neotragus was taken fromBrashares et al. (2000). Positions of Connochaetes, Damaliscus,Acelaphus, and Sigmoceros were taken from Gatesy andArctander (2000), whereas Brashares et al. (2000) provided
Table 2
Descriptions of coloration variables (see Appendix for sources)
Variables Descriptions
Overall body colorationa
Pale White, sandy, light brown, pale gray, light gray, buffy gray, light buff, light red, fawn,tawny, or yellowish-brown coloration
Red Red, rusty, chestnut, tawny, rufous, ochraceous, cinnamon, or auburn tonesGray Gray or grayish colorationDark Black, blackish, or dark brown coloration
Seasonal coloration change
Lighter in winter Pelage becomes white or lighter seasonally
Underparts
Countershading Ventral coloration described as paler than upper parts, white, dingy-white, whitish,or tinged with white
Color patterns
Presence of spots in young Black or white spots on dorsal surface, rear or legsPresence of spots on adults Black or white spots on dorsal surface, rear or legsPresence of stripes (vertical or horizontal) on young Black or white stripes on dorsal surface, rear or legsPresence of stripes (vertical or horizontal) on adult Black or white stripes on dorsal surface, rear or legsPresence of side band Prominent line along side of body
Tail
Light Markings on tail white or lighter than coloration of rumpDark Markings on black or darker than coloration of rump
Rump
White markings Presence of rump patch or white markings on rump
Face
Conspicuous light markings White or light markings on face that contrast with the overall body colorationConspicuous dark markings Dark or black markings on face that contrast with the overall body coloration
Legs
Conspicuous light markings White or light markings on legs that contrast with the overall body colorationConspicuous dark markings Dark or black markings on legs that contrast with the overall body coloration
a Sexually dichromatic species were scored as both colors.
826 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

the topography within Damaliscus. Within the Caprini-Rupi-caprini-Ovibovini clade, the general placement of Ovis,Oreamnos, Capra, Hemitragus (species assumed to be mono-phyletic), Pseudois (assumed to be monophyletic), Pantholops,Ovibos, Rupicapra, Nemorhaedus (assumed to be monophyletic),and Capricornis was inferred from Gatesy and Arctander(2000). Resolution within Ovis was based on Hassanin et al.(1998), Ludwig and Knoll (1998), Ludwig and Fisher (1998),and Hiendleder et al. (1998). The placement of Budorcastaxicolor is based on Hassanin and Douzery (1999a,b). Thetopography of Capra species is based on Hassanin et al. (1998)and Manceau et al. (1999). The placement of Ammotragus withRupicapra was based on Hassanin and Douzery (1999b),whereas the topography of Capricornis species was based onHassanin et al. (1998). In the tribe Hippotragini, theplacement of Oryx, Addax, and Hippotragus was based onGatesy and Arctander (2000). The general placements ofRedunca, Kobus, Pelea, and Cephalophus branches in theRedunci-Cephalophini tribes were based on Gatesy andArctander (2000). Relationships within these genera werederived from additional sources: Brasheres et al. (2000)(Redunca, Cephalophus), Gatesy and Arctander (2000) (Kobus),Hassanin and Douzery (1999b), and Mathee and Robinson(1999) (placement of Oreotragus in clade with Cephalophus).
Some species were excluded from the composite phylogenybecause relatively little information could be found regard-ing their phylogenetic position (e.g., Bos sauveli, Tragelaphus
buxtoni, Cephalophus rufilatus, Cephalophus natalensis, Cephalo-phus nigrifrons, Modoqua piacentii), or great disagreements existregarding their placements (e.g., Saiga saiga, Oribi oribi, andProcapra species). To resolve polytomies in the composite tree(as MacClade requires a fully dichotomized tree for concen-trated-changes tests), we used an online library database(BIOSIS) to count the number of citations retrieved for eachspecies in the polytomy (Nunn C, personal communication).Species with the least number of citations (indicating thosefor which less information is known) were excluded from thepolytomies until a fully dichotomized tree remained. Thesespecies included Ovis vignei, Ovis nivicola, Hemitragus jayakari,Capra wallie, Capra falconeri, Capra pyrenaica, and Oryx dammah.
To prepare for phylogenetic analyses, we first usedMacClade to map the coloration (dependent) and ecological(independent) variables onto the phylogenetic tree using thematrix depicting each species’ character states (one, zero, or‘‘?’’; supplementary appendix online). From the distributionof character states across all species, MacClade reconstructsthe evolutionary history of a given trait throughout the tree byusing parsimony. This makes it possible to count the numberof evolutionary gains (change in a character state froma zero to a one) and losses (change from a one to a zero) ineither the coloration or ecological variables (Maddison andMaddison, 1992). In those instances in which character re-construction was ambiguous (i.e., the tree contained areas inwhich both zero and one were equally parsimonious), within
Table 3
Descriptions of ecological and behavioral variables (see Appendix for sources)
Variables Descriptions
Latitude categories
Tropics Part of range falls in tropic latitudes (0�10 degrees from equator)Subtropics Part of range falls in tropic latitudes (11�30 degrees)Transitional Part of range falls in tropic latitudes (31�50 degrees)Subarctic Part of range falls in tropic latitudes (51�60 degrees)Arctic Part of range falls in tropic latitudes (more than 60 degrees)
Environmental categories
Open Grasslands, deserts, or tundra habitatsClosed Swamp/riverine, or dense forest habitats
Habitat categories
Grassland Occupies prairie, savannah, meadows, or steppe grasses habitatsBushland Occupies scrub, bushland, riparian, thicket, or shrub vegetation habitatsWoodland Occupies woods, woody areas, or woodlandsLight forest/woodland All species included in the woodland category, plus those occupying open, sparse, or light forestsDense forest Occupies alpine, tropical, boreal, deciduous, mixed, timberland, or dense forestsDesert Found in deserts or semidesertsRocky Occupies rocky areas such as talus, boulders, rocky outcrops, crevices, or cliffsTundra Found in tundraSwamp/riverine Occupies swamp, marsh, bogland, moorland, reedbeds or riverine habitats
Activity pattern categories
Diurnal Primarily active during the dayCrepuscular/nocturnal Primarily active during the night, or in the morning or late/afternoon evening
Group size categories
Solitary Primarily found alone or in pairsIntermediate groups Primarily found in groups of 2�50 individualsLarge groups Forms aggregations of more than 50 individuals
Hider/follower behavior
Hider Young lie concealed for more than 1 week after birthFollower Young follow mothers within 1 week of birth
Hunting style of principal predators
Stalker Predator usually observes/follows/stalks prey before attackingCourser Predator usually runs down/exhausts prey
Stoner et al. • Color of artiodactyls 827
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022
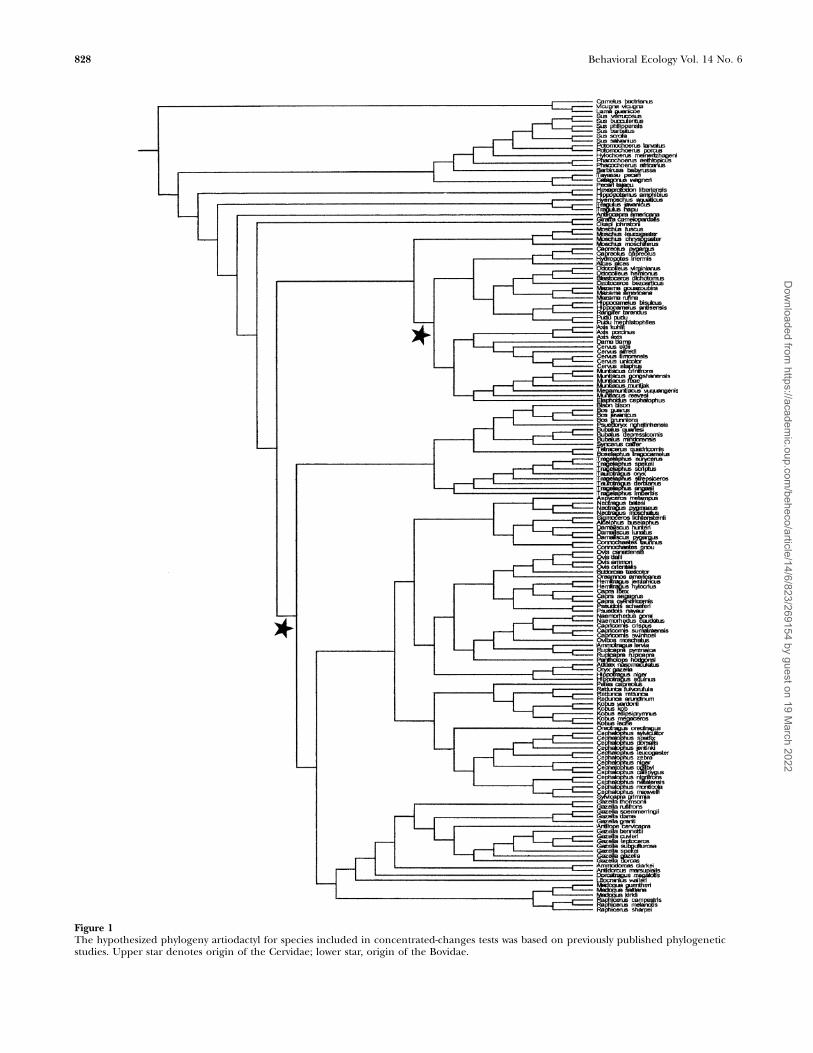
Figure 1The hypothesized phylogeny artiodactyl for species included in concentrated-changes tests was based on previously published phylogeneticstudies. Upper star denotes origin of the Cervidae; lower star, origin of the Bovidae.
828 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

MacClade we used ‘‘most parsimonious reconstructionmode,’’ which includes all reconstructions produced by thedeltran and acctran options, to generate all possible recon-structions of the character. Because multiple ambiguities forthe same character can lead to many possible reconstructions,we selected the first and the last reconstructions for the firstanalysis (following the method of Ortolani 1999; Ortolani andCaro 1996). We examined the first and last reconstructions asthese are the reconstructions with the maximum number ofgains and fewest losses, and minimum number of gains andmost losses, respectively.
Next, we used concentrated-changes tests to test theprobability that gains in the coloration variable were asso-ciated with the ecological variable more than could be ex-pected by chance, and losses in the coloration variableoccurred in the presence of the ecological trait less than ex-pected by chance. Thus, we examined whether the presenceof the ecological trait facilitated the maintenance of the colortrait over evolutionary time. The null hypothesis, testedagainst a distribution derived through simulation and10,000 replicates, was that gains and losses in a given colora-tion variable were randomly distributed on the tree withrespect to the ecological variable. When ambiguity in char-acter reconstruction existed, the multiple reconstructionsresulted in two or four probability values for each test,depending on whether one or both of the dependent andindependent variables displayed ambiguity. Because, inconcentrated-changes tests (but not the nonparametric tests),a dependent variable could be matched to an independentvariable more than once because of multiple phylogeneticreconstructions, we applied a standard Bonferroni correction.This is a more conservative way to apply this test than by usinga sequential Bonferroni (Rice, 1989). Thus, if four tests hadbeen conducted between a coloration and ecological variable,we multiplied each p value by four and reported it only if theproduct was less than 0.1. Following the method of Rice(1989), we limited application of the Bonferroni correction tomultiple tests of the same pair of dependent and independentvariables rather than across one dependent and severalindependent variables because each independent variable ex-hibited a different phylogenetic reconstruction and was usedto test a different hypothesis. If several concentrated-changestests were conducted for a given prediction, we report only themost significant p value corrected using the standardBonferroni technique.
For the sake of convenience, even-toed ungulates isa general term we use loosely to refer to all species that weanalyzed, cervids refers to Cervidae, bovids to the Bovidae,and artiodactyls to all species in the supplementary appendixonline. As a short hand, ‘‘cross-species comparisons’’ refer tospecies counts analyzed by using chi-square tests or Fisher’sexact probability tests that do not control for phylogeny,whereas ‘‘phylogenetically controlled tests’’ refer to the resultsof concentrated changes tests using the reduced species list,termed CCT. For Fisher’s and chi-square tests, true p valuesare given, but for CCT, p values are given after Bonferronicorrections. We consider an association worth discussing if anyof the phylogenetic reconstructions was significant aftera Bonferroni correction.
RESULTS
Coloration for concealment
Uniform colorationTo discover whether ungulate coloration aids in concealment,we first tested whether uniform pelage color typically matcheshabitat background. Cross-species comparisons showed that
pale bovids and artiodactyls were likely to be found in openenvironments (N ¼ 125 species, v2 ¼ 9.28, p , .01; N ¼ 194,v2 ¼ 13.26, p , .01, respectively), but results failed to reachsignificance when confounding effects of shared ancestrywere taken into account. Pale bovids and artiodactyls werealso associated with one habitat category, deserts, but only incross-species comparisons (N ¼ 125, v2 ¼ 6.91, p ¼ .03; N ¼194, v2 ¼ 10.19, p , .01, respectively). Next, we tested grayand red pelage coloration against rocky habitats and foundgray bovids (N ¼ 125, v2 ¼ 8.20, p ¼ .02) and artiodactyls (N ¼194, v2 ¼ 8.80, p ¼ .01) were significantly more likely to live inthis habitat, but again, these results disappeared usingphylogenetic tests. Surprisingly, red pelage was associatedwith living in habitats that were not rocky (bovids: N ¼ 125,v2 ¼ 9.80, p , .01). Dark bovids and artiodactyls were foundsignificantly more often in closed environments (N ¼ 125,v2 ¼ 4.37, p , .0001; N ¼ 195, v2 ¼ 33.2, p , .0001,respectively), and dark cervids, bovids, and artiodactyls werefound in dense forest habitats (Fisher’s test: N ¼ 38, p ¼ .02;N ¼ 125, v2 ¼ 16.16, p , .01; N ¼ 195, v2 ¼ 28.23, p , .0001,respectively), although not using CCTs. An associationbetween dark bovids and swamp habitats approached signif-icance (N ¼ 125, v2 ¼ 5.30, p ¼ .08).
Lighter coats in winterIf seasonal pelage coloration contributes to concealment,species displaying a lighter winter coat should occupy habitatswhich become lighter seasonally. Cervids, bovids, and artio-dactyls were likely to turn lighter in winter if they lived inarctic latitudes (Fisher’s test: N ¼ 39, p ¼ .07; N ¼ 125, v2 ¼2.72, p , .01; N ¼ 197, v2 ¼ 24.79, p , .0001, respectively;CCTs for both bovids and artiodactyls: p , .001). Bovids andartiodactyls with lighter winter coats were also associated withtundra habitats (N ¼ 125, v2 ¼ 12.72, p , .01; N ¼ 197, v2 ¼24.37, p , .0001, respectively), and this association held truefor all three taxonomic groups using CCTs (p ¼ .07, p , .01,p , .001, respectively). In short, there was strong backing forungulates taking on lighter coats in arctic and tundra regions.
Spotted coatsIf spotted coats facilitate crypsis through pattern blending,having the appearance of dappled light would be favorable fordiurnal, solitary, forest-dwelling species and those that hidetheir young. Cervids with spotted adult coats were diurnal(CCT: p ¼ .06) but were less likely to be solitary (Fisher’s test:N ¼ 37, p ¼ .03). Conversely, bovids with spotted young andspotted adult coats were likely to be solitary (N ¼ 125, v2 ¼.79, p ¼ .03, N ¼ 125, v2 ¼ 5.17, p ¼ .08, respectively), as werespecies with spotted young across all artiodactyls (N ¼ 193,v2 ¼ 13.90, p , .01; CCT: p ¼ .02). Spotted adult bovids wereassociated with light forests (N ¼ 125, v2 ¼ 9.97, p , .01; CCT:p ¼ .08). Cervids with spotted young were strongly tied tograssland/bushland habitats (CCT: p , .0001), bovid spottedyoung to light forests habitats (N ¼ 125, v2 ¼ 11.66, p , .01;CCT: p ¼ .06), and spotted young across all artiodactyls todense forests (N ¼ 196, v2 ¼ 13.62, p , .01). Species withspotted young were significantly more likely to hide after birthin all taxonomic groups (cervids, Fisher’s test: N ¼ 38, p ¼ .09;bovids: N ¼ 123, v2 ¼ 5.60, p ¼ .06; artiodactyls: N ¼ 194, v2 ¼23.40, p , .0001, CCT: p ¼ .05) (Figure 2). Indeed, spottedcervid, bovid, and artiodactyl young were significantly lesslikely to follow their mothers (Fisher’s: N ¼ 38, p ¼ .09; N ¼123, v2 ¼ 5.90, p ¼ .05; N ¼ 194, v2 ¼ 24.61, p , .0001,respectively). In short, there was patchy support for adultspotted coats helping with pattern blending, whereas inyoung, spotted coats were found in forest bovids, grasslandcervids, and hiding and solitary artiodactyls, strongly support-ing a concealment hypothesis.
Stoner et al. • Color of artiodactyls 829
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022
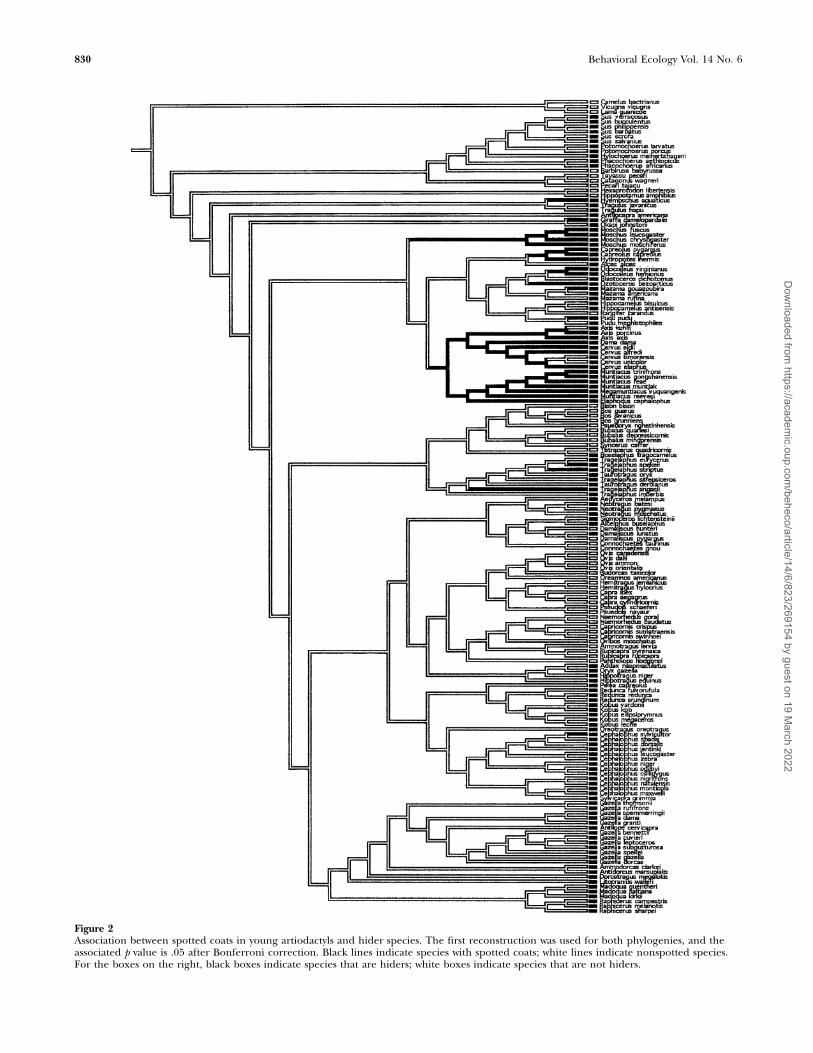
Figure 2Association between spotted coats in young artiodactyls and hider species. The first reconstruction was used for both phylogenies, and theassociated p value is .05 after Bonferroni correction. Black lines indicate species with spotted coats; white lines indicate nonspotted species.For the boxes on the right, black boxes indicate species that are hiders; white boxes indicate species that are not hiders.
830 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

Striped coatsSimilar analyses of striped patterns revealed that adultsdisplaying stripes were significantly associated only with lightforests in bovid and artiodactyl species (N ¼ 125, v2 ¼ 15.15,p , .001, CCT: p , .1; N ¼ 197, v2 ¼ 11.32, p , .01, CCT:p ¼ .09, respectively) (Figure 3). We found marginalassociations between bovids with striped young and lightforests (N ¼ 125, v2 ¼ 15.15, p , .001, CCT: p , .1) andartiodacyls with striped young and dense forests (N ¼ 197, v2
¼ 4.88, p ¼ .09). There was a tendency for artiodactyls withstriped young to be solitary (N ¼ 194, v2 ¼ 5.33, p ¼ .07), butthis did not hold after controlling for phylogeny. Bovids andartiodactyls with striped young were chiefly those thatexhibited a hiding strategy (N ¼ 123, v2 ¼ 5.87, p ¼ .05; N¼ 195, v2 ¼ 11.60, p , .01, respectively), and they did notexhibit following behavior (N ¼ 123, v2 ¼ 6.34, p ¼ .04; N ¼195, v2 ¼ 12.21, p , .01, respectively); however, these resultsdid not hold up after controlling for phylogeny. In short,striped coats in adults were found in light forests, and stripedyoung were associated with hiding species.
Dark side bands and dark leg markingsIf side bands act as disruptive coloration by acting to break upthe body shape, they may aid in concealment for species thatare diurnal or are found in open and sunny environments inwhich other forms of crypsis are limited. Artiodactyls withprominent side bands were principally diurnal (N ¼ 187, v2 ¼6.41, p ¼ .04) and found in open environments (N ¼ 194,v2 ¼ 14.73, p , .001), as were bovids (N ¼ 124, v2 ¼ 11.84,p , .01). There were strong associations between side bandsand desert-dwelling for both bovids (N ¼ 124, v2 ¼ 10.00,p , .01) and artiodactyls (N ¼ 194, v2 ¼ 14.71, p ¼ .0006).Reindeer (Rangifer tarandus), the only cervid with side bands,is found in tundra (Fisher’s: N ¼ 38, p ¼ .03). None of theanalyses of side bands remained significant after controllingfor phylogeny.
Dark leg markings, which may serve as another form ofdisruptive coloration, should similarly be associated with openenvironments. We found that artiodactyls with dark legs werelikely to live in deserts (N ¼ 196, v2 ¼ 8.36, p ¼ .0.02; CCT:p ¼ .07), although there was no association with other openhabitats such as tundra.
CountershadingAs predicted if countershading acts to conceal shadows caston the lower body, countershaded bovids were diurnal (CCT:p ¼ .06) and strongly associated with desert habitats (CCT: p ¼.01). Comparisons across all countershaded artiodactylssimilarly revealed an association with desert-dwelling (CCTs:p ¼ .03). Conversely, there were fewer countershaded speciesliving in tundra than expected by chance (bovids: N ¼ 125,v2 ¼ 12.11, p , .01; artiodactyls: N ¼ 195, v2 ¼ 10.03, p , .01).
Coloration for communication
Markings may be most effective as visual signals if they areassociated with conditions in which they are most visible (e.g.,diurnal activity patterns and open habitats), associated withgregarious species (for intraspecific communication), orassociated with particular predator types (for interspecificcommunication).
Spotted and striped coats in adultsSpotted adult pelages might provide a mechanism to reduceintraspecific aggression because spotted coats resemble thoseof young in many species and might therefore act as a signalof subordination. Spotted cervids were more likely to be
found in intermediate-sized groups (two to 50 individuals;Fisher’s test: N ¼ 37, p , .01), but there were no associationsbetween spotted coats in adults and living in intermediate-sized or large (more than 50) groups after controlling forphylogeny. There were no significant associations betweenstriped coats in adults and intermediate or large group sizes.
Side bandsIf side bands are used in communication between conspe-cifics, perhaps to maintain gregariousness, they should befound in group-living species. Artiodactyls with side bandslived in intermediate-sized (N ¼ 192, v2 ¼ 8.56, p , .01) andlarge groups (N ¼ 192, v2 ¼ 15.91, p , .0001; CCT: p ¼ .01),providing support for this hypothesis.
Face markingsDark visual signals for intra or interspecific communicationmight be most noticeable in diurnal species occupying openhabitats. Cross-species comparisons revealed that cervids withdark faces tended to be diurnal (N ¼ 36, v2 ¼ 6.77, p ¼ .03),but there was no evidence that dark-faced species in any of thethree clades lived in open environments. Dark-faced bovidsand artiodactyls lived in intermediate (CCTs: p , .01; p , .01,respectively) and large-sized social groups (CCTs: p ¼ .02; p ¼.02, respectively) (Figure 4), as is expected if dark faces areused to communicate with conspecifics. Although dark-facedartiodactyls were apparently subject to predation by stalkingpredators (N ¼ 188, v2 ¼ 8.79, p ¼ .01), there were alsoindications that bovids and artiodactyls with dark faces werepursued by coursers after controlling for shared ancestry(CCTs: p , .01; p , .01, respectively).
If white face markings are used in communication, theymight be prominent signals for either diurnal or nocturnalspecies. Artiodactyls with white faces were likely to be diurnal(CCT: p ¼ .01) but not nocturnal, and to live in intermediate-sized social groups (CCT: p ¼ .06). Bovids with white faceswere likely to be pursued by coursers (N ¼ 119, v2 ¼ 5.92,p ¼ .06), although this result was not upheld using phylo-genetic tests.
Because any conspicuous markings might provide aneffective signal, rather than dark or white facial markingsper se, we repeated the analyses for species with either dark orwhite markings, or a combination of the two. Cervids andartiodactyls with conspicuous faces inhabited grassland/bushland habitats (CCTs: p ¼ .03; p ¼ .03, respectively), butno other associations were significant. Bovids with conspicu-ous faces were less likely to be found in large groups thanexpected by chance (N ¼ 125, v2 ¼ 6.05, p ¼ .01). In short,there was rather little support for conspicuous faces having anobvious communicatory role.
Leg markingsArtiodactyls with dark leg markings were associated withdeserts (N ¼ 196, v2 ¼ 8.36, p ¼ .02; CCT: p ¼ .07), wherethis color pattern is presumably prominent. Artiodacylswith dark legs are found in large social groups (CCT:p , .1), as expected if such markings facilitate intraspecificcommunication; there was no association with stalking orcoursing predators for any taxonomic group.
White leg markings were associated with diurnality inartiodactyls (CCT: p ¼ .04), but we found no associationsbetween these markings, open habitats, gregariousness, orpredator types.
We repeated leg marking analyses combining species witheither dark or white leg coloration and found that both bovidsand artiodactyls with conspicuous legs were diurnal (CCTs:p ¼ .02; p ¼ .08, respectively) and principally lived in deserts
Stoner et al. • Color of artiodactyls 831
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022
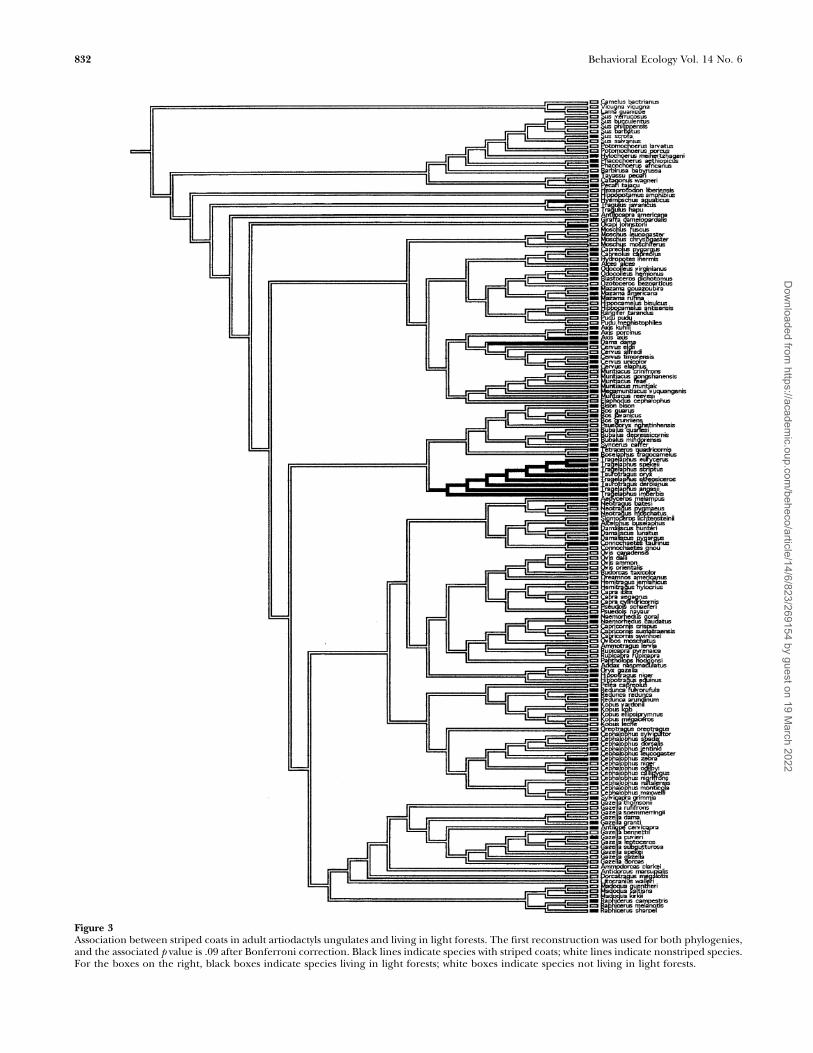
Figure 3Association between striped coats in adult artiodactyls ungulates and living in light forests. The first reconstruction was used for both phylogenies,and the associated p value is .09 after Bonferroni correction. Black lines indicate species with striped coats; white lines indicate nonstriped species.For the boxes on the right, black boxes indicate species living in light forests; white boxes indicate species not living in light forests.
832 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022
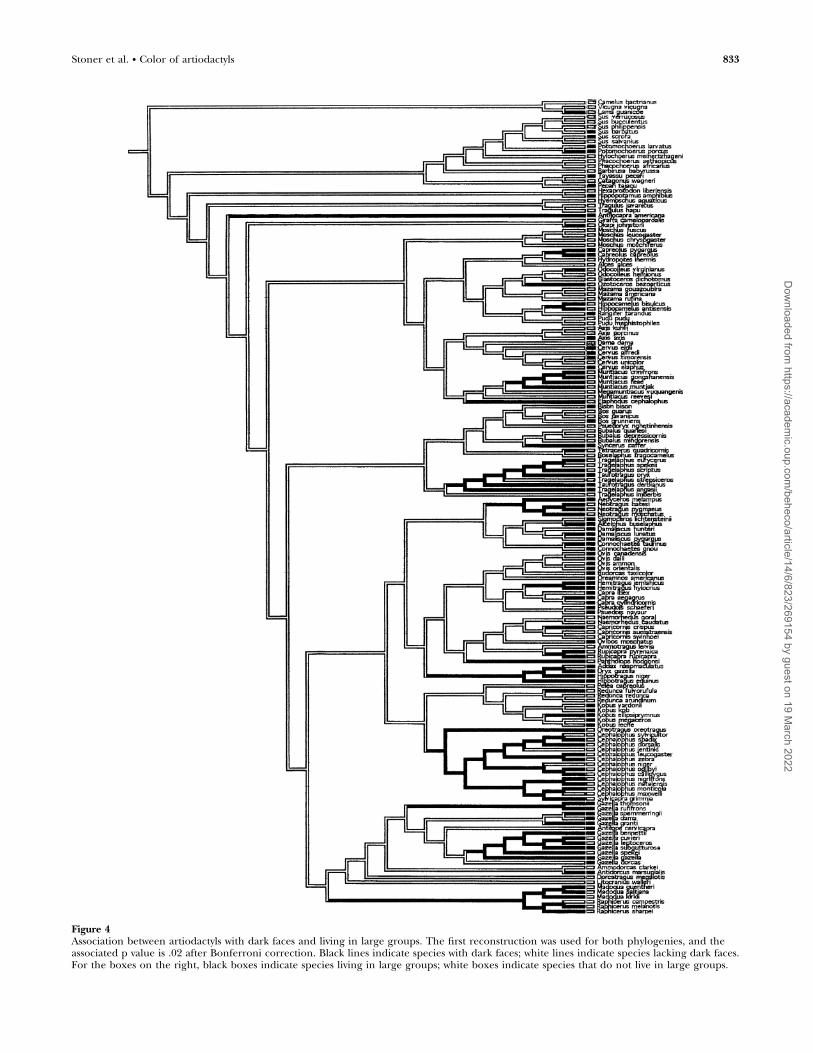
Figure 4Association between artiodactyls with dark faces and living in large groups. The first reconstruction was used for both phylogenies, and theassociated p value is .02 after Bonferroni correction. Black lines indicate species with dark faces; white lines indicate species lacking dark faces.For the boxes on the right, black boxes indicate species living in large groups; white boxes indicate species that do not live in large groups.
Stoner et al. • Color of artiodactyls 833
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

(CCTs: p ¼ .04; p ¼ .08, respectively). Artiodactyls withconspicuous legs were associated with also grassland/bush-land habitats (CCT: p ¼ .05). These findings suggest a role incommunication, but because no significant associations wereuncovered with group sizes or predator hunting style, it isunclear whether intra or interspecific communication isinvolved.
Tail markingsChi-square analyses gave some backing for dark tail markingsacting as visual signals. Dark tails were found among diurnalbovids (N ¼ 121, v2 ¼ 9.00, p ¼ .01) and artiodactyls (N ¼ 188,v2 ¼ 7.17, p ¼ .03). Bovids with dark tails tended to form largegroups (N ¼ 125, v2 ¼ 8.76, p ¼ .01), whereas artiodactyls withdark tails were associated with intermediate (N ¼ 192, v2 ¼8.33, p ¼ .02) and large groups (N ¼ 192, v2 ¼ 8.69, p ¼ .01).Dark-tailed bovids and artiodactyls were associated withstalking predators (N ¼ 119, v2 ¼ 9.47, p , .01; N ¼ 187,v2 ¼ 6.16, p ¼ .05, respectively), and there were no significantassociations with coursing predators. None of these resultswere upheld in phylogenetic tests, giving little support toconventional hypotheses for dark tail coloration.
White tails were associated with diurnality in bovids (N ¼121, v2 ¼ 7.01, p ¼ .03) but not with being diurnal or living inopen environments in other clades. There were few associa-tions between white tails and gregariousness, except white-tailed artiodactyls were weakly associated with large-sizedgroups (CCT: p ¼.1). There was an association between white-tailed bovids and stalking predators, but only in cross speciescomparisons (N ¼ 119, v2 ¼ 10.82, p , .01). Thus, there waslittle support for white tails having signal value.
In examining species with either black or white tails, wefound that bovids and artiodactyls with any conspicuous mar-kings were diurnal (N ¼ 121, v2 ¼ 9.34, p , .01, CCT: p , .01;N ¼ 188, v2 ¼ 7.40, p , .01, CCT: p ¼ .06, respectively).Artiodactyls with tail markings were associated with interme-diate-sized (N ¼ 193, v2 ¼ 3.09, p ¼ .08, CCT: p ¼ .07) andlarge (N ¼ 193, v2 ¼ 4.47, p ¼ .03, CCT: p ¼ .09) groups, aswere bovids (N ¼ 125, v2 ¼ 5.88, p ¼ .02). Conspicuous tailscould be involved in interspecific communication, as bovidswith conspicuous tails were attacked by stalking predators(N ¼ 119, v2 ¼ 12.48, p , .001), although cervids wereactually less likely to be attacked by stalkers (Fisher’s test:N ¼ 36, p , .01); significance for these analyses were lostafter controlling for phylogeny. In short, there was reason-ably strong support for conspicuous tails being associated withintraspecific communication.
White rumpsIn examining whether white rumps serve a communicatoryrole, we found white rumps were associated with diurnality inartiodactyls (N ¼ 188, v2 ¼ 5.15, p ¼ .07; CCT: p ¼ .09) andopen habitats in bovids (N ¼ 125, v2 ¼ 21.14, p , .0001; CCT:p ¼ .05) and artiodactyls (N ¼ 195, v2 ¼ 33.29, p , .0001;CCT: p , .0001). There were strong associations using cross-species comparisons between white rumps and living inintermediate-sized groups for cervids (Fisher’s test: N ¼ 36,p , .0001; CCT: p , .01), bovids (N ¼ 125, v2 ¼ 16.48, p ,.001), and artiodactyls (N ¼ 193, v2 ¼ 32.32, p , .0001) andliving in large groups (cervids: N ¼ 36, v2 ¼ 7.29, p ¼ .04;bovids: N ¼ 125, v2 ¼ 23.64, p , .0001; artiodactyls: N ¼ 193,v2 ¼ 33.37, p , .0001). Bovids and artiodactyls with whiterumps were likely to be pursued by coursers (N ¼ 119, v2 ¼13.88, p , .1; N ¼ 187, v2 ¼ 8.48, p ¼ .01, respectively), butabsence of significance in phylogenetically controlled testscasts doubt on their role in signaling to predators. Noassociations were found between species sporting white rumpsand those preyed on by stalking predators.
Coloration for thermoregulation
Uniform colorationBovids and artiodactyls with dark pelages were foundprincipally in the tropics (bovid CCT: p , .0001; artiodactylCCT: p ¼ .01), as expected from Gloger’s rule. Pale bovids andartiodactyls were associated with deserts in chi-square tests (N¼ 125, v2 ¼ 6.91, p ¼ .03; N ¼ 194, v2 ¼ 10.19, p , .01,respectively) but not after controlling for phylogeny.
Face markingsTo test whether white face markings serve to reduce heatstress, we checked whether they are found in species living inopen and warm environments. White-faced cervids were morelikely to live in open environments (Fisher’s test: N ¼ 39, p ¼.06; CCT: p ¼ .06). White faces were associated with living ingrassland/bushland in bovids (N ¼ 125, v2 ¼ 6.19, p ¼ .05)and in artiodactyls (N ¼ 196, v2 ¼ 4.82, p ¼ .09; CCT: p , .01).White faces were also found in desert bovids (N ¼ 125,v2¼ 6.19, p ¼ .05). In sum, white faces were associatedwith diurnality, open environments and grassland/bushlandhabitats. These findings provide evidence for a thermoregu-latory role.
White rumpsTurning to the idea that rumps may also deflect heat in even-toed ungulates, we found significant relationships betweenspecies having white rumps and those occupying deserts(cervids, Fisher’s test: N ¼ 38, p ¼ .08; bovids, N ¼ 125, v2 ¼24.37, p , .0001; artiodactyls, N ¼ 195, v2 ¼ 33.29, p , .0001)and grassland/bushland (bovids, N ¼ 119, v2 ¼ 7.92, p ¼ .05;artiodactyls, N ¼ 187, v2 ¼ 8.39, p ¼ .02). These associationswere not upheld when shared ancestry was taken into account,however.
DISCUSSION
After taking phylogeny into account, we found that manyaspects of coloration in ungulates are related to concealmentand are less involved in communication. Thermoregulationapparently played only a minor role in the evolution ofcoloration in this taxon. Although our results are preliminary,they point to predation pressure being a key selective forceacting on coloration in this order of mammals. Theimportance of mate choice was not assessed but is likely tobe of minor import, judging from the small number ofungulate species that are sexually dichromatic. If correct, thiscontrasts sharply with birds, although a phylogeneticallycontrolled analysis of sexual dichromatism and mating systemin ungulates would still be useful.
Before discussing our findings in more detail, we mustallude to five points that could have influenced our results.First, we know that our behavioral and especially our eco-logical categories are broad. Thus, dense forest constitutedalpine, tropical, boreal, deciduous, mixed dense forests, andtimberland, where lighting conditions are likely to beextremely variable (Endler, 1993) and would influence theextent to which prey might remain cryptic. Indeed, variationin lighting within subcategories of dense forest might even begreater than between dense forest and light forest/woodland,but it is impossible to measure this across the range ofenvironments in which species live. More refined categoriesmight increase our ability to detect associations.
Second, the large scope of the study meant that we lackeddetailed data on most species and were forced to use a coarsecategories that could be ascribed to most species, such ashabitat or group size. If analyses were restricted to a smallersample of well-known species, one might be able to use more
834 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

sensitive measures, such as ‘‘is’’ or ‘‘is not’’ preyed up bycheetahs (Acinonyx jubatus) and thereby be able to make farmore specific predictions about prey-predator signaling, forexample (see Caro, 1994).
Third, our attempts to tease out predictions that would lendsupport or cast doubt on conventional hypotheses may havebeen misplaced. Thus, it may be inappropriate to suggest thatif a color patch is used in communication, we would expect itto be found in diurnal or open country species; perhaps theywould be found in crepuscular species or forest-dwellingspecies too.
Fourth, our phylogenetic reconstruction was of lowresolution, as a complete molecular phylogeny is not avail-able. In the absence of such data, we constructed a compositetree based on the most recent molecular studies andsupplemented this with taxonomies based on morphologicaltraits. Certain species were additionally excluded because oflack of information or strong disagreements over theirphylogenetic position, and we resolved polytomies by usingthe arbitrary procedure of excluding species with fewestcitations. Nevertheless, this is the most comprehensive treebased primarily on molecular studies that is currently availablefor Artiodactyla. Although a more resolved tree might in thefuture change our results because concentrated changes testsare notoriously sensitive to inclusion or exclusion of species(Sillen-Tullberg, 1993), we think that this would not producemore significant associations. Nonetheless, future reanalysisof data in the supplementary appendix online by usinga thorough molecular phylogeny will definitely be worthwhile.
Fifth, we used stringent criteria to accept that an associationbetween a coloration pattern and an ecological or behavioralvariable was indeed significant by focusing attention onphylogenetic reconstructions that were significant only aftera standard and therefore conservative Bonferroni correction.With these points in mind, we now discuss specific associa-tions summarized in Table 4.
Overall body coloration
If analyses had been restricted to simple cross-speciescomparisons, there would have been strong support foroverall coat coloration being an adaptation for backgroundmatching or thermoregulation (Kingdon, 1982; Spinage,1986). Pale coats were found in open environments anddesert habitats, and dark coats were found in closed environ-ments and dense forests. Gray coats were found in rockyhabitats, which would support only a background-matchingrole. Nonetheless, none of these significant associations weremaintained once shared ancestry was taken into account.Moreover, on some of these tests, species counts revealed thatthere were actually more species not associated with thevariable of interest (pale bovids not found in deserts; grayartiodactyls not found on rocks), despite there being morepale species in deserts than non-pale species in deserts. Incontrast, parallel comparative analyses of overall bodycoloration in carnivores (Ortolani and Caro, 1996) andlagomorphs (Stoner et al., 2003) revealed that pale colorationwas associated with desert habitats, making the even-toedungulate results even more surprising. In conclusion, despiteseveral extraordinarily experienced naturalists having linkedoverall coat color to habitat on the basis of observationsmade on single species or guilds of ungulates, there isno comparative systematic support for background matchingacross the even-toed ungulates. Although a more resolvedphylogeny might, in the future, be more supportive ofbackground matching, we speculate that overall coat colora-tion, such as being pale, serves different functions in differentspecies, and that ascribing the function of all pale coats either
to background matching or to reflecting heat in hot climatesis too simplistic.
Cross taxonomic support for Gloger’s rule is equivocal. Thisstudy showed that dark even-toed ungulates live in the tropics;similarly, in canids, ursids, and herpestids, dark species arefound in tropical forests (Ortolani and Caro, 1996). Assumingmany tropical areas are humid, this result provides indirectsupport for Gloger’s rule, although the causal mechanismsmaintaining the association remain opaque. In contrast, darklagomorphs were not associated with tropical latitudes aftercontrolling for phylogeny (Stoner et al., 2003), so thegenerality of this rule in mammals is still open to question.
Ungulates that assume lighter coats in winter are associatedwith arctic latitudes and tundra habitats. This might be anadaptation to match their snowy background (Cott, 1940) ora thermoregulatory device, as white coats are translucent andlet heat and sunlight penetrate, which are then caught bydarker inner hairs. Dark outer hairs would only absorb heatremaining on the surface that is quickly carried away by wind(Wolff J, personal communication). Similar matches betweenwhite coats and living in the arctic have been found inphylogenetic comparisons in carnivores (Ortolani and Caro,1996) but not in lagomorphs (Stoner et al., 2003).
It seems obvious that deer are spotted to blend in withdappled light (Beddard, 1892; Schaller, 1967; Thayer, 1909),so we were surprised to find few significant associationsbetween species in which adults are spotted and living inforests or grassland. This was undoubtedly owing to sharedancestry because spotted species are closely related, particu-larly in the cervids. Nonetheless, spotted cervids were diurnal,and spotted bovids were found in light forests (although thereare only five species that are spotted). In sum, spotted coatsmay be an adaptation for concealment based on these twotests, but other predictions concerning habitats and beingsolitary were not supported. Among carnivores, dark spotswere associated with living in closed habitats and havingungulates as the main prey item, suggesting that, in that order,their function is aggressive camouflage (Ortolani, 1999).
There was definite support for striped coats aiding inconcealment (Cott, 1940; Estes, 1991) given that both bovidsand artiodactyls with such coats were more likely to inhabitlight forests after controlling for ancestry. Nonetheless, therewas no association with being diurnal, living in grassland, orbeing solitary and possessing a striped coat. In carnivores,however, vertically striped coats were associated with living ingrassland and with being terrestrial, bolstering the back-ground-matching hypothesis (Ortolani, 1999). In even-toedungulates, there was no evidence for spotted or striped coatsbeing involved in intraspecific communication because theywere not associated with gregariousness.
Many of the predictions regarding spotted coats in young ashelping in concealment were well supported (Cott, 1940;Thayer, 1909). Each of the three taxonomic groups was foundin forests or grasslands, and solitary artiodactyls wereparticularly likely to have spotted young (see Danilkin,1996). Extraordinarily, every one of the 30 species withspotted young exhibited a hiding strategy, demonstratinga tight link between this morphological character and thistype of antipredator behavior. Mothers of species that havespotted young all leave their neonates alone when they forage.Support for species with spotted young using the followerstrategy (Macdonald, 1984) was lacking.
Striped coats in young also aid in concealment. Bovids withstriped young were found in lightly forested habitats. Specieswhose young had striped coats were significantly more likelyto be hiders not followers, but based only on cross-species comparisons. Thus, the relationship between patternblending in spotted young and the hiding strategy was
Stoner et al. • Color of artiodactyls 835
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

replicated in striped young although without the same degreeof certainty.
It is difficult to interpret results regarding predictions aboutside bands acting as a form of disruptive coloration (Estes,1991). Although artiodactyls with side bands were diurnal andlived in open environments and in both desert and tundrahabitats, and although some of these findings were also foundseparately in cervids and bovids, none of these results held upafter controlling for phylogeny. The clearest result from cross-species analyses was that artiodactyls with side bands lived inopen environments (24 out of 24 species), but this is nota clear-cut expectation from the disruptive colorationhypothesis (e.g., disruptively colored woodcock, Scolopaxminor, chicks are found in leaf litter); thus, we cannot becertain that side bands serve a disruptive function. In contrast,artiodactyls with side bands were found in large groups,suggesting that they might be used in intraspecific commu-nication, perhaps allowing individuals to keep in sight of eachother.
There was a strong association between bovids andartiodactlys that were countershaded and inhabiting deserts.Because countershading is thought to aid in concealment byreducing shadow in well-lit environments (Cott, 1940; Fogdenand Fogden, 1974), the hypothesis is partially supporteddespite no significant associations with other open environ-ments. In contrast to even-toed ungulates, lagomorphs thatwere countershaded were not more likely to be found in openhabitats that included deserts (Stoner et al., 2003), so thewidespread importance of countershading in helping toconceal mammals is still questionable.
Body parts
Focusing on phylogenetically controlled comparisons, dark-faced bovids and artiodactyls were found in intermediate- andlarge-sized social groups, which implies an intraspecific
communicatory function (see Kingdon, 1982; Spinage,1986). The prediction that dark faces might be used to signalto stalking predators that they have been noticed (see Caro,1995) was supported only weakly (in a chi-square test onartiodactyls), but in contrast, there were highly significantassociations between dark faces and being pursued bycoursers, the biological significance of which is mysterious.Because even-toed ungulates are likely to flee from coursers,predators would be unlikely to see their faces. Perhaps theextent of blackness on the face signals ability to outdistancea predator, but this is pure speculation. In carnivores, darkpatches around the eyes were associated with crepuscularactivity, perhaps serving as a way to reduce glare (Ortolani andCaro, 1996).
White faces were associated with being diurnal (allartiodactyls) and living in open environments (cervids andartiodactyls). Species with white faces were found in grass-land/bushland habitats in all three clades. If, as we imagine,grassland habitats are sunny environments with relatively littleshade, this would lend (albeit weak) support to a thermoreg-ulatory role in which white faces are used to lowertemperature, as suggested by Geist (1987). There was weaksupport for white faces being used in intraspecific communi-cation based on patterns of grouping but no support forsignaling to predators (see Poulton, 1890). Similarly, therewas very little widespread support for conspicuous faces beingused in communication.
We found weak support for dark legs acting as disruptivecoloration (see Roosevelt and Heller, 1914) because artiodac-tyls with dark legs were found in deserts. Phylogeneticallycontrolled comparisons gave some support to a role for darklegs being used in intraspecific signaling, but only forartiodactyls and living in large groups. Hypotheses suggestingwhite legs might be an aid in communication (Danilkin, 1996;Geist, 1987; Kingdon 1982) received weak support. Conspic-uous leg markings were more obviously associated with
Table 4
Degree of support for associations tested in the present study
Concealment Communication Physiological
Coloration BM PB DC CS Intra Inter T
Overall body coloration
Uniform color None Yesa
Lighter in winter StrongSpotted adults Weak NoneStriped adults Strong NoneSpotted young StrongStriped young WeakSide bands None StrongCountershading Moderate
Body parts
Dark face Strong StrongWhite face Weak None ModerateConspicuous face None NoneDark legs Weak Weak NoneWhite legs WeakConspicuous legs StrongDark tail None NoneWhite tail Weak NoneConspicuous tail None NoneWhite rump Moderate None None
BM indicates background matching; PB, pattern blending; DC, disruptive coloration; CS countershading; Intra, intraspecific communication;Inter, interspecific communication; and T, themoregulation.
a Refers to Gloger’s rule.
836 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

diurnal communication in open habitats, but the type ofcommunication, to predators or conspecifics, was unknown.
Conventional hypotheses that dark tails are involved inintraspecific communication (Kingdon, 1982) or signal topredators were supported only in nonparametric tests.Backing for white tails acting in communication (Bildstein,1983; Kingdon, 1982) was similarly weak, and cross taxonomicanalyses did do not support the suggestion, based on singlespecies studies, that white tails are used to signal to predators(for a review, see Caro et al., 1995). However, our examinationof species with conspicuous tail markings (be they dark orwhite) revealed that these species are more likely to be diurnaland gregarious than those without distinct tail markings.Thus, conspicuous tails seem to be involved with intraspecificcommunication in even-toed ungulates. In lagomorphs,contrasting tail tips were similarly associated with sociality(Stoner et al., 2003) although not in carnivores (Ortolani andCaro, 1996), making it difficult to form general conclusionsabout mammals.
White rumps might either act as signals to conspecifics(Guthrie, 1971; Ward, 1979) or to predators (Caro, 1986;Fitzgibbon and Fanshawe, 1988), or be used in thermoregu-lation (Spinage, 1986; Estes, 1991). White-rumped species inall three ungulate clades were found in species inhabitingopen environments and, after controlling for phylogeny, inintermediate-sized social groups, lending weight to intraspe-cific communication. There was some evidence of signaling topredators because white-rumped species were pursued bycoursers, although not in concentrated changes tests. Thatwhite rumps were found in the open, in deserts, and ingrassland/bushland habitats lends credence to the thermo-regulation hypothesis, but we did not replicate all these resultsby using phylogenetic controls. The function of white rumpsremains an enigma, although intraspecific communicationand thermoregulation both remain promising candidates.
Conclusion
We uncovered strong support for the following color patternsserving to conceal even-toed ungulates: lightening in winter,striped coats in adults, and spotted coats in young. We foundsome support for a communicatory role of color patterns, thatis, side bands, dark faces, conspicuous tails, and white rumps.Finally, we found only moderate support for a color patternacting in a thermoregulatory capacity, notably dark coats andwhite faces. We made no attempt to explore the role of matechoice in driving coloration patterns, as no specific hypothesispertains to sexual selection in mammals, despite certainspecies, including some ungulates, being sexually dichromatic.
Our clearest findings were concerned with hypothesesabout concealment, particularly background matching andpattern blending, and our less conclusive findings tended toconcern issues of communication and thermoregulation. Inpart, it is easier to formulate clear-cut predictions abouthabitat characteristics than about social or predator variableswhen testing these hypotheses about communication, and it isnot always clear as to the way in which different habitats mightcontribute to heat stress (e.g., humid rain forests or parcheddeserts). Also, we suspect that several social variables may beintercorrelated (e.g., follower and social species, being socialand being pursued by coursers), and this will inevitablyconfound findings that rely on behavioral variables to testpredictions. That said, our findings suggest that somecoloration patterns in ungulates probably have differentfunctions in different species, and this is why the comparativedata set sometimes failed to support hypotheses based onstudies of one or two species. Despite these problems, wenonetheless feel that systematic comparative analyses are keyto making strong and supportable generalizations about the
adaptive significance of coloration in animals and that theyelevate the study of coloration from fascinating anecdotes totestable hypotheses about a wide array of species.
APPENDIX
Sources used to obtain morphological and ecologicalinformation for each species are as follows: Bauer, 1995,Chadwick, 1983; Chapman and Chapman, 1997; Clark, 1994;Cloudsley-Thompson, 1980; Dagg and Bristol-Foster, 1976;Danilkin, 1996; Estes, 1991, 1993, Grzimek, 1990; Heptneret al., 1989; Hoefs, 1985; Holmes, 1974; Huntingford andTurner, 1987; Johnson and Lockard, 1983; Jones et al., 1985;Kingdon, 1982; Lindsey et al., 1999; Macdonald, 1984;Mungall and Sheffield, 1994; Nabhan, 1993; Nowak, 1999;Miura et al., 1993; Oliver, 1993; Payne and Francis, 1985;Putman, 1988; Prior, 1987; Schaller, 1977, 1998; Shackleton,1999; Shrestha, 1997; Soma, 1987; Sowls, 1984, 1997; Spinage,1968, 1982; Spitz, 1991; Strahan, 1983; Stuart and Stuart,1993, 1997; Valdez, 1982; Valdez and Krausman, 1999; VanWormer, 1969; Walther, 1984; Walther et al., 1983; Wemmer,1987; Whitehead, 1972, 1993; Wilson, 2000.
We thank the University of California for supporting C.J.S. and theMcNair Scholars Program for supporting C.M.G., Kate Trimlett andJanelle Vargas for help with coding variables, Marisa Flores forrunning supplementary analyses, John Eadie and Charlie Nunn fordiscussion, John Byers for drawing our attention to the possibility ofwhite rumps being used in thermoregulation, Jerry Wolff forsuggesting we examine conspicuous coloration patterns and morebehavior, and Dave Westneat, Jerry Wolff, and an anonymous reviewerfor very helpful comments.
REFERENCES
Alvarez F, Braza F, Norzagaray A, 1976. The use of the rump patch inthe fallow deer (D. dama). Behaviour 56:298–308.
Amato G, Egan MG, Schaller GB, 2000. Mitochondrial DNA variationin Muntjac: evidence for discovery, rediscovery, and phylogeneticrelationships. In: Antelopes, deer, and relatives: fossil record,behavioral ecology, systematics, and conservation (Vrba ES, SchallerGB, eds). New Haven, Connecticut: Yale University Press; 285–295.
Badyaev AV, Hill GE, 2000. Evolution of sexual dichromatism:contribution of carotenoid- versus melanin-based coloration. BiolJ Linn Soc 69:153–172.
Baker RR, Parker GA, 1979. The evolution of bird coloration. PhilTrans R Soc Lond B 287:63–130.
Bauer EA, 1995. Elk: Behavior, ecology and conservation. Stillwater,Minnesota: Voyageur Press.
Beddard FE, 1892. Animal colouration. London: Hazell, Watson andViney.
Bildstein KL, 1983. Why white-tailed deer flag their tails. Am Nat121:709–715.
Brasheres JS, Garland T Jr, Arcese P, 2000. Phylogenetic analysis ofcoadaptation in behavior, diet, and body size in the Africanantelope. Behav Ecol 11:452–463.
Brooke M de L, 1998. Ecological factors influencing the occurrence of‘‘flash marks’’ in wading birds. Funct Ecol 12:339–346.
Burtt EH Jr, 1979. Tips on wings and other things. In: Behaviouralsignificance of color (Burtt EH Jr, ed). New York: Garland STPMPress; 75–110.
Buxton PA, 1923. Animal life in deserts: a study of the fauna inrelation to the environment. London: Edward Arnold.
Caro TM, 1986. The functions of stotting in Thomson’s gazelles(Gazella thomsoni): some tests of the predictions. Anim Behav34:663–684.
Caro TM, 1994. Ungulate antipredator behavior: preliminary andcomparative data from African bovids. Behaviour 128:189–228.
Caro TM, 1995. Pursuit-deterrence revisted. Trends Ecol Evol 10:500–503.
Stoner et al. • Color of artiodactyls 837
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

Caro TM, Lombardo SL, Goldizen AW, Kelly MJ, 1995. Tail-flaggingand other antipredator signals in white-tailed deer: new data andsynthesis. Behav Ecol 6:442–450.
Chadwick DH, 1983. A beast the color of winter: the mountain goatobserved. San Francisco: Sierra Club Books.
Chapman D, Chapman N, 1997. Fallow deer: their history, distribu-tion, and biology. Machynlleth: Coch-y-bonddu Books.
Clark JL, 1994. The great ark of the wild sheep. Norman: University ofOklahoma Press.
Cloudsley-Thompson JL, 1980. Tooth and claw: defensive strategies inthe animal world. London: M. Dent.
Cooper W Jr, 2002. Convergent evolution of plant chemicaldiscrimination by omnivorous and herbivorous scleroglossanlizards. J Zool 257:53–66.
Cooper WE Jr, Caldwell JP, Vitt LJ, Perez-Mellado V, Baird TA, 2002.Food-chemical discrimination and correlated evolution betweenplant diet and plant-chemical discrimination in lacertiform lizards.Can J Zool 80:655–663.
Corbet GB, Hill JE, 1991. A world list of mammalian species, 3rd ed.London: British Museum (Natural History).
Corbet GB, Hill JE, 1992. The mammals of the Indomalayan region:a systematic review. Oxford: Oxford University Press.
Cott HB, 1940. Adaptive colouration in animals. London: Methuen.Cronin MA, Stuart R, Pierson BJ, Patton JC, 1996. K-Casein gene
phylogeny of higher ruminants (Pecora, Artiodactyla). Mol PhyloEvol 6:295–311.
Dagg AI, Bristol-Foster J, 1976. The giraffe: its biology, behavior andecology. New York: Van Nostrand Reinhold.
Danilkin A, 1996. Behavioural ecology of Siberian and European roedeer. London: Chapman Hall.
Douzery E, Randi E, 1997. The mitochondrial control region ofCervidae: evolutionary patterns and phylogenetic content. Mol BiolEvol 14:1154–1166.
Emerson BC, Tate ML, 1993. Genetic analysis of evolutionaryrelationships among deer (subfamily Cervinae). J Hered 84:266–273.
Endler JA, 1978. A predator’s view of animal colour patterns. Evol Biol11:319–364.
Endler JA, 1993. The color of light in forest and its implications. EcolMonogr 63:1–27.
Estes RD, 1991. The behaviour guide to African mammals. LosAngeles: University of California Press.
Estes RD, 1993. The safari companion: a guide to watching Africanmammals. Post Mills, Vermont: Chelsea Green.
Fitzgibbon CD, Fanshawe JH, 1988. Stotting in Thomson’s gazelles: anhonest signal of condition. Behav Ecol Sociobiol 23:69–74.
Fogden M, Fogden P, 1974. Animals and their colors. New York:Crown.
Franklin WL, 1982. Biology, ecology and relationship to man of theSouth American camelids. Pymatuning Lab Ecol Spec Pub 6:457–489.
Gatesy J, Arctander P, 2000. Hidden morphological support for thephylogenetic placement of Pseudoryx nghinhensis with bovine bovids:a combined analysis of gross anatomical evidence and DNAsequences from five genes. Syst Biol 49:515–538.
Gatesy J, O’Grady P, Baker RH, 1999. Corroboration among data setsin simultaneous analysis: hidden support for phylogenetic relation-ships among higher level artiodactyl taxa. Cladistics 15:271–313.
Geist V, 1978. Life strategies, human evolution, environmental design:toward a biological theory of health. New York: Springer-Verlag.
Geist V, 1987. On the evolution of optical signals in deer:a preliminary analysis. In: Biology and management of the Cervidae(Wemmer C, ed). Washington, DC: Smithsonian Institute Press;235–255.
Groves CP, 1981. Ancestors for the pigs: taxonomy and phylogenyof the genus Sus, technical bulletin no. 3. Canberra, Australia:Department of Prehistory, Research School of Pacific Studies,Australian National University.
Groves CP, Schaller GB, 2000. The phylogeny and biogeographyof the newly discovered Annamite artiodactyls. In: Antelopes, deer,and relatives: fossil record, behavioral ecology, systematics, andconservation (Vrba ES, Schaller GB, eds). New Haven, Connecticut:Yale University Press; 261–282.
Grubb P, 1993. Order Artiodactyla. In: Mammal species of the world(Wilson DE, Reeder DM, eds). Washington, DC: SmithsonianInstitute Press; 377–414.
Grzimek B, 1990. Grzimek’s encyclopedia of mammals. New York:McGraw-Hill.
Guilford TC, 1990. The evolution of aposematism. In: Insect defences:adaptive mechanisms and strategies of prey and predators (EvansDL, Schmidt JO, eds). Albany: State University of New York Press;23–61.
Guthrie RD, 1971. A new theory of rump patch evolution. Behaviour38:132–145.
Guthrie RD, Petocz RG, 1970. Weapon automimicry among mammals.Am Nat 104:585–588.
Hamilton WJ III, 1973. Life’s color code. New York: McGraw-Hill.Harvey PH, Pagel MD, 1991. The comparative method in evolutionary
biology. Oxford: Oxford University Press.Hassanin A, Douzery EJP, 1999a. Evolutionary affinities of the
enigmatic saola (Pseudoryx nghetinhensis) in the context of themolecular phylogeny of Bovidae. Proc R Soc B 266:893–900.
Hassanin A, Douzery EJP, 1999b. The tribal radiation of the familyBovidae (Artiodactyla) and the evolution of mitochondrial cyto-chrome b gene. Mol Phylo Evol 13:227–243.
Hassanin A, Pasquet E, Vigne JD, 1998. Molecular systematics of thesubfamily Caprinae (Artiodactyla, Bovidae) as determined fromcytochrome b sequences. J Mammal Evol 5:217.
Hasson O, 1986. Predator decision-making and pursuit-deterrencesignals. Anim Behav 35:45–54.
Heptner VG, Nasimovich AA, Bannikov AG, 1989. Mammals of theSoviet Union (Hoffman RS, ed). New York: Brill.
Hickman GC, 1979. The mammalian tail: a review of functions.Mammal Rev 9:143–157.
Hiendleder S, Mainz K, Plante Y, Lewalski H, 1998. Analysis ofmitochondrial DNA indicates that domestic sheep are derivedfrom two different ancestral maternal sources: no evidencefor contributions from urial and argali sheep. J Hered 89:113–120.
Hoefs M, 1985. Wild sheep: distribution, abundance, managementand conservation of the sheep of the world and closely relatedmountain ungulates. Whitehorse: Northern Wild Sheep and GoatCouncil.
Hoglund J, 1989. Size and plumage dimorphism in lek-breeding birds:a comparative analysis. Am Nat 134:72–87.
Holmes F, 1974. Following the roe: a natural history of the roe deer.Edinburgh: J. Bartholomew.
Huntingford F, Turner AK, 1987. Animal conflict. New York:Chapman and Hall.
Irschick DJ, Vilt LJ, Zary PA, Losos JB, 1997. A comparison ofevolutionary radiations in mainland and Caribbean anolis lizards.Ecology 78:2191–2203.
Johnson CE, 1921. The hand-stand habit of the spotted skunk.J Mammal 2:87–89.
Johnson RL, Lockard FR, 1983. Mountain goats and mountain sheepof Washington, biological bulletin no. 18. Olympia: WashingtonDepartment of Game.
Jones JK, Armstrong DM, Choate JR, 1985. Guide to mammals of theplains states. Lincoln: University of Nebraska Press.
Kingdon J, 1982. East African mammals: bovids, vol. 3. New York:Academic Press.
Kingdon J, 1997. The Kingdon field guide to African mammals. NewYork: Academic Press.
Kiltie RA, 1988. Countershading: universally deceptive or deceptivelyuniversal? Trends Ecol Evol 3:21–23.
Kiltie RA, 1989. Testing Thayer’s countershading hypothesis: an imageprocessing approach. Anim Behav 38:542–552.
Leyhausen P, 1979. Cat behavior. New York: Garland STPM Press.Lindsey SL, Green ML, Bennett CL, 1999. The okapi: mysterious
animal of Congo-Zaire. Austin: University of Texas Press.Linnell JDC, Andersen R, 1998. Timing and synchrony of birth in
a hider species, the roe deer (Capreolus capreolus). J Zool 244:497–504.
Lobao Tello JLP, van Gelder RG, 1975. The natural history of nyalaTragelaphus angasi (Mammalia, Bovidae) in Mozambique. Am MusNat Hist 155:323–386.
Ludwig A, Fisher S, 1998. New aspects of an old discussion:phylogenetic relationships of Ammotragus and Pseudois within
838 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

the subfamily Caprine based on comparison of the 12S rDNAsequences. J Zool Syst Evol Res 36:173–178.
Ludwig A, Knoll J, 1998. Multivariate morphometrische analysen dergattund ovis linnaeus, 1758 (Mammalia, Caprinae). Z Saugetierkd63:210–219.
MacDonald D, 1984. The encyclopedia of mammals, vol. 2. London:George, Allen and Unwin.
Maddison WP, 1990. A method for testing the correlated evolution oftwo binary characters: are gains and losses concentrated on certainbranches of the phylogenetic tree? Evolution 44:539–557.
Maddison WP, Maddison DR, 1992. MacClade: analysis of phylogenyand character evolution. Sunderland, Massachusetts: SinauerAssociates.
Manceau V, Despres JB, Taberlet P, 1999. Systematics of the genusCapra inferred from mitochondrial DNA sequence data. Mol PhyloEvol 13:504–510.
Mathee CA, Robinson TJ, 1999. Cytochrome b phylogeny of the familyBovidae: resolution within the Alcelaphini, Antilopini, Neotragini,and Tragelaphini. Mol Phylo Evol 12:31–46.
Meyer W, Kranzle S, 1999. Zur Bedeutung von Farburg undZeichnung der Korperpole (Kopf und Analregion) bei Saugetieren.Zool Garten NF 3:159–187.
Miura S, Kaji K, Ohtaishi N, Koizumi T, Tokida K, Wu J, 1993. Socialorganization and mating behavior of the white-lipped deer in theQinghai-Xizang Plateau, China. In: Deer of China: biology andmanagement (Ohtaishi N, Sheng HI, eds). Amsterdam: Elsevier;220–234.
Mottram JC, 1915. Some observations on pattern-blending withreference to obliterative shading and concealment of outline. ProcZool Soc Lond 1915:679–692.
Mottram JC, 1916. An experimental determination of the factorswhich cause patterns to appear conspicuous in nature. Proc ZoolSoc Lond 1915:383–419.
Mungall EC, Sheffield WJ, 1994. Exotics on the range: the Texasexample. College Station: Texas A&M University Press.
Nabhan GP, 1993. Counting sheep: twenty ways of seeing desertbighorn (Nabhan GP, ed). Tucson: University of Arizona Press.
Nee S, Read AF, Harvey PH, 1996. Why phylogenies are necessary forcomparative analysis. In: Phylogenies and the comparative methodin animal behavior (Martins EP, ed). Oxford: Oxford UniversityPress; 399–411.
Nowak RM, 1999. Walker’s mammals of the world, 6th ed. Baltimore,Maryland: Johns Hopkins University Press.
Oliver WLR, 1993. Pigs, peccaries, and hippos: status survey andconservation action plan. Gland: IUCN.
Ortolani A, 1999. Spots, stripes, tail tips and dark eyes: predicting thefunction of carnivore colour patterns using the comparativemethod. Biol J Linn Soc 67:433–476.
Ortolani A, Caro TM, 1996. The adaptive significance of colorpatterns in carnivores: phylogenetic tests of classic hypotheses. In:Carnivore behavior, ecology, and evolution, vol. 2. (Gittleman J, ed).Ithaca, New York: Comstock; 132–188.
Pagel M, 1994. Detecting correlated evolution on phylogenies:a general method for the comparative analysis of discretecharacters. Proc R Soc Lond Ser B 255:37–45.
Payne J, Francis CM, 1985. A field guide to the mammals of Borneo.Kuala Lumpur: World Wildlife Fund Malaysia.
Perez-Barberıa FJ, Gordon IJ, 1999. The functional relationshipbetween feeding type and jaw and cranial morphology in ungulates.Oecologia 118:157–165.
Perez-Barberıa FJ, Gordon IJ, 2000. Differences in body mass and oralmorphology between the sexes in the Artiodactyla: evolutionaryrelationships with sexual segregation. Evol Ecol Res 2:667–684.
Perez-Barberıa FJ, Gordon IJ, 2001. Relationships between oralmorphology and feeding style in the Ungulata: a phylogeneticallycontrolled evaluation: Proc R Soc Lond B 268:1021–1030.
Perez-Barberıa FJ, Gordon IJ, Pagel M, 2002. The origins of sexualdimorphism in body size in ungulates. Evolution 56:1276–1285.
Poulton EB, 1890. The colours of animals. London: Kegan Paul,Trench, Trubner.
Price T, 1997. Correlated evolution and independent contrasts. PhilTrans R Soc B 352:519–528.
Prior R, 1987. Deer watch. London: David and Charles.Putman RJ, 1988. The natural history of deer. London: Christopher
Helm.
Randi E, Lucchini V, Diong CH, 1996. Evolutionary genetics of theSuiformes as reconstructed using mtDNA sequencing. J MammalEvol 3:163.
Randi E, Mucchi N, Claro-Hergueta F, Amelie B, Douzery EJP, 2001. Amitochondrial DNA control region phylogeny of the Cervinae:speciation in Cervus and implications for conservation. AnimConserv 4:1–11.
Randi E, Mucchi N, Pierpaoli M, Douzery EJP, 1998. New phylo-genetic perspectives on the Cervidae (Artiodactyla) are provided bythe mitochondrial cytochrome b gene. Proc R Soc Lond B 265:793–801.
Read AF, Nee S. 1995. Inference from binary comparative data.J Theor Biol 173:99–108.
Rebholz W, Harley E, 1999. Phylogenetic relationships in the bovidsubfamily Antilopinae based on mitochondrial DNA sequences.Mol Phylo Evol 12:87–94.
Rice WR, 1989. Analyzing tables of statistical tests. Evolution 43:223–225.
Ridley M, 1983. The explanation of organic diversity: the comparativemethod and adaptations for mating. Oxford: Oxford UniversityPress.
Roosevelt T, 1911. Revealing and concealing coloration in birds andmammals. Bull Am Mus Nat Hist 30:119–231.
Roosevelt T, Heller E, 1914. Life histories of African game animals.New York: Scribner’s.
Rowland WJ, 1979. The use of colour in intraspecific communication.In: The behavioral significance of color (Burtt EH Jr, ed). New York:Garland STPM Press; 379–421.
Schaller GB, 1967. The deer and the tiger. Chicago, Illinois: Universityof Chicago Press.
Schaller GB, 1977. Mountain monarchs: wild sheep and goats of theHimalaya. Chicago, Illinois: University of Chicago Press.
Schaller GB, 1998. Wildlife of the Tibetan steppe. Chicago, Illinois:University of Chicago Press.
Schreiber A, Seibold E, Notzold G, Wink M, 1999. Cytochrome b genehaplotypes characterize chromosomal lineages of Anoa, theSulawesi dwarf buffalo (Bovidae: Bubalus sp.). Am Genet Assoc90:165–176.
Shackleton DM, 1999. Hoofed mammals of British Columbia.Vancouver: UBC Press.
Shrestha TK, 1997. Mammals of Nepal: with reference to those ofIndia, Bangladesh, Bhutan, and Pakistan. Kathmandu: BimalaShrestha.
Sillen-Tullberg B, 1993. The effect of biased inclusion of taxa on thecorrelation between discrete characters in phylogenetic trees.Evolution 47:1182–1191.
Soma H, 1987. The biology and management of Capricornis andrelated mountain antelopes (Soma H, ed). New York: Croom Helm.
Sowls LK, 1984. The peccaries. Tucson: University of Arizona Press.Sowls LK, 1997. Javelinas and other peccaries: their biology,
management and use. College Station: Texas A&M University Press.Spinage CA, 1968. The book of the giraffe. London: Collins.Spinage CA, 1982. A territorial antelope: the Uganda waterbuck. New
York: Academic Press.Spinage CA, 1986. The natural history of antelopes. Beckenham,
Kent: Croom Helm Publishers.Spitz F, 1991. Proceedings of the International Symposium ‘‘Ongules/
Ungulates 91,’’ Toulouse, France: 2–6 September 1991 (Spitz F, ed).Toulouse: SFEPM-IRGM.
Stoner CJ, Bininda-Emonds ORP, Caro TM, 2003. The adaptivesignificance of coloration in lagomorphs. Biol J Linn Soc (79:309–328).
Strahan R, 1983. Complete book of Australian mammals: the nationalphotographic index of Australian wildlife. Sidney: Angus &Robertson.
Stuart C, Stuart T, 1993. Field guide to the mammals of southernAfrica, 2nd ed. Cape Town: Struik.
Stuart C, Stuart T, 1997. Field guide to the larger mammals of Africa.Cape Town: Struik.
Su B, Wang YX, Lan H, Wang W, Zhang Y, 1999. Phylogenetic study ofcomplete cytochrome b genes in musk deer (genus Moschus) usingmuseum samples. Mol Phylo Evol 12:241–249.
Thayer AG, 1909. Concealing coloration in the animal kingdom. NewYork: Macmillan.
Stoner et al. • Color of artiodactyls 839
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022

Thayer AH, 1896. The law which underlies protective coloration. Auk13:124–129.
Theimer TC, Keim P, 1998. Phylogenetic relationships of peccariesbased on mitochondrial cytochrome b DNA sequences. J Mammal79:566–572.
Thenius E, 1970. Zur evolution und verbreitungsgeschichte derSuidae (Artiodactyla, Mammalia). Z Saugetierkd 35:321–342.
Valdez R, 1982. The wild sheep of the world. Mesilla: Wild Sheep andGoat International.
Valdez R, Krausman PR, 1999. Mountain sheep of North America.Tucson: University of Arizona Press.
Van Wormer J, 1969. The world of the American Elk. Philadelphia,Pennsylvania: Lippincott.
Wallace AR, 1889. Darwinism. London: Macmillan.Walther FR, 1984. Communication and expression in hoofed animals.
Bloomington: Indiana University Press.Walther FR, Mungall EC, Grau GA, 1983. Gazelles and their relatives:
a study in territorial behavior. Westwood, New Jersey: Noyes.
Ward P, 1979. Colour for survival. London: Orbis.Webb SD, 2000. Evolutionary history of new world Cervidae. In:
Antelopes, deer, and relatives: fossil record, behavioral ecology,
systematics, and conservation (Vrba ES, Schaller GB, eds). New
Haven, Connecticut: Yale University Press; 38–64.Wemmer C, 1987. Biology and management of the Cervidae. In:
Research symposia of the National Zoological Park, Front Royal,
Virginia: 1–5 August 1982 (Wemmer C, ed). Washington, DC:
Smithsonian Institute Press; 577.Whitehead GK, 1972. Deer of the world. London: Constable.Whitehead GK, 1993. The Whitehead encyclopedia of deer. Stillwater,
Minnesota: Voyageur Press.Wickler W, 1968. Mimicry in plants and animals. London: Weidenfeld
and Nicholson.Wilson EO, 2000. Sociobiology: the new synthesis. Cambridge,
Massachusetts: Harvard University Press.
840 Behavioral Ecology Vol. 14 No. 6
Dow
nloaded from https://academ
ic.oup.com/beheco/article/14/6/823/269154 by guest on 19 M
arch 2022