Embed Size (px)
Citation preview

Physiological and Molecular Plant Pathology (1997) 50, 67–83
Effects of chloroplast targeted potato virus Y coat
protein on transgenic plants
M. N and P. H. B*
Department of Plant, Soil and Entomological Sciences, Di�ision of Plant Pathology, Uni�ersity of Idaho, Moscow,ID 83844-2339, U.S.A.
(Accepted for publication August 1996)
Studies of several plant viruses have shown that virus coat protein (CP) may play a role insymptom development. In order to determine how and where potato virus Y (PVY)-CP maycontribute to symptom development, two sets of plants were generated, one expressing PVY-CP,and one transformed with a chimeric construct consisting of PVY-CP fused to the transit peptide(TP) of pea ribulose bisphosphate carboxylase small subunit (rbcs) to target PVY-CP to thechloroplast stroma. Numerous transgenic plants developed chlorotic or white mosaic symptomsafter transformation with the TP-CP construct, but not after transformation with PVY-CP alone.Both western blot analysis of stroma and thylakoid protein fractions and immunocytochemistryrevealed that the imported PVY-CP was preferentially associated with thylakoid membranes inTP-CP+ plants. Light and electron microscopy showed that PVY-CP disrupted chloroplastdevelopment and cell structure in TP-CP+ transformants. Virus challenge of non-transformedCP+ and a range of TP-CP+ tobacco plants indicated that high level PVY-CP expressing TP-CP+
plants were more susceptible to PVY. These observations suggest that PVY-CP action inchloroplasts may contribute to symptom development in PYV-infected plants.
# 1997 Academic Press Limited
INTRODUCTION
Potyviruses cause a wide range of symptoms in their hosts, dependent on the virus–
host combination as well as environmental conditions. The precise manner by which
plant viruses cause disease is unknown. Since a common symptom is chlorosis, as is
typical when mosaic or mottle symptoms are observed, it is reasonable to assume that
virus infection has some bearing on chloroplast function. Several mechanisms
underlying virus pathogenicity have been proposed, including the possibility that ac-
cumulation of one or more viral products in the chloroplast leads to symptom
development [for reviews see 5, 46]. The interaction of viruses or specific viral gene
products with host chloroplasts has been observed in a variety of virus–host
combinations [5, 28, 46].
Disruption of chloroplast development or function in virus-infected plants could be
*To whom correspondence should be addressed.Abbreviations used in text : CP, coat protein; LM, light microscopy; PVY, potato virus Y; TEM,
transmission electron microscopy; TMV, tobacco mosaic virus ; TP, transit peptide ; TVMV, tobacco veinmottling virus.
0885–5765}97}02006717 $25.00}0}pp960071 # 1997 Academic Press Limited

68 M. Naderi and P. H. Berger
due to a variety of factors. Among these may be the result of an interaction(s) of virus
coat protein and host chloroplasts. Here, we have focused on the coat protein of potato
virus Y (PVY), and have done so for two reasons : (1) we previously observed that not
only genomic RNA but also two PVY gene products, coat protein (CP) and the helper
component-proteinase protein, are found inside chloroplasts of infected plant cells
[15]. Moreover, PVY-CP is not distributed uniformly in chloroplasts but is almost
exclusively associated with isolated thylakoid membranes [15]. We have also observed
that viral RNA isolated from chloroplasts of infected plants is infectious (Gunasinghe
and Berger, unpublished data). Further, there can be a specific reduction or inhibition
of photosystem II activity in potyvirus-infected plants [29, 30] or a reduction in
chloroplast numbers and size [44]. (2) While it is likely, and has been demonstrated in
several cases, that virus gene products other than coat protein correlate or are somehow
involved in alterations of chloroplast function [4,11, 18, 19, 27, 32, 33, 41], there is
considerable evidence indicating that CPs of a number of viruses are involved in
chlorosis induction [7, 16, 31, 34, 40].
In order to investigate the effects of PVY-CP in different compartments of the cell,
we have generated several transgenic tobacco lines. One series produces and
accumulates CP in the cytoplasm, while another series accumulates CP in the
chloroplast stroma compartment by use of a stroma-specific transit peptide (TP). We
report here the effects of chloroplast targeted CP on host cells.
MATERIALS AND METHODS
Vector construction
An 800 bp SalI}EcoR I fragment encoding the PVY-CP gene was cloned into plasmid
pJIT117 [14] to create a transcriptional}translational fusion of PVY! coat protein and
the transit peptide (TP¯ 80 amino acids) of pea ribulose bisphosphate carboxylase
small subunit (rbcs). The TP-CP chimeric gene is flanked at the 5«-end with a CaMV
35S promoter (with duplicated enhancers) and at the 3«-end with a CaMV transcription
terminator}poly(A) addition signal. This expression cassette, as a Xho I}Sac I
fragment, was cloned into pKYLX [39] binary vector, flanked by another transcription
terminator}poly(A) addition signal (rbcs). The resultant plasmid was designated
pMNLX1.0. The PVY-CP gene without the transit peptide sequence and with an in-
frame, translationally optimal initiation codon was cloned into another vector,
designated pKYLX68.1.
Plant transformation
Nicotiana tabacum cv. Xanthi, a systemic host for PVY!, was used throughout this study.
Binary vectors were mobilized into Agrobacterium tumefaciens LBA4404 using the method
of An [1]. Nicotinia tabacum plants were transformed with the engineered Agrobacterium
using the leaf disk method and transformants selected on modified Linsmaier–Skoog
media [39] using kanamycin concentrations of 25, 50 and 300 mg l−". Plants
transformed with PVY-CP without the transit peptide sequences were selected on a
medium containing 300 mg l−" kanamycin. Shoots were transferred to shooting media
without antibiotics and, if roots developed, transferred to soil and grown in growth

PVY coat protein and chloroplasts 69
chambers at 25 °C with a 16 h photoperiod. R"seeds were germinated in the presence
of 50 mg l−" kanamycin under the above conditions on modified Linsmaier–Skoog
media [39].
Protein isolation and western blot analysis
Total leaf proteins were extracted as described previously [15] and stored at ®20 °Cuntil use. For western blot analysis, total soluble proteins from each sample were
separated by 12% SDS PAGE [23], electrophoretically transferred to nitrocellulose,
and probed with anti-PVY sera as described previously [15].
Chloroplast isolation and separation of stroma and thylakoid protein fractions
Intact chloroplasts were isolated as described by Fish and Jagendorf [12] as modified
by Reinero and Beachy [34]. Chloroplasts were resuspended in resuspension buffer,
thermolysin (200 µg ml−") and CaCl#(0±5 m) added, and incubated on ice for 1 h [3].
Chloroplasts were washed twice in five volumes of resuspension buffer. Thylakoid and
stromal protein fractions were obtained as described [37]. Proteins from both fractions
were precipitated with nine volumes of cold acetone, equalized on the basis of OD#)!
,
and stored at ®20 °C for later use in SDS PAGE and western blotting.
RNA isolation and RNA gel blot analysis
Total RNA was isolated as described by Sambrook et al. [38]. RNA accumulation was
determined by northern blot hybridization using 10 µg of total RNA per lane. The blot
was exposed to Kodak XAR-5 film for 12 h at ®80 °C.
Light and electron microscopy
Developmentally similar tobacco leaf tissue was fixed and infiltrated with LR White
resin and polymerized in gelatin capsules at 60 °C for 24 h, as described elsewhere by
Tranbarger et al. [43], except that 2% osmium tetroxide postfixation was used for
tissues for transmission electron microscopy (TEM) but not for immunolocalization.
Plant tissue was selected for collection at approximately the same time of day (approx.
10:00 am). Sections were prepared on an LKB 8800 Ultratome III (Gaithersburg,
MD, U.S.A.) ultramicrotome for light microscopy (LM) or TEM. For light microscopy,
sections 2 µm thick were affixed to gelatin-coated glass microscope slides and stained
with methylene blue. Stained sections were photographed under bright-field
illumination using a JENAVAL (aus JENA, Germany) transmitted-light research
microscope. For TEM, thin sections were picked up onto formvar-coated nickel grids,
and stained with uranyl acetate and potassium permanganate. For TEM immuno-
cytochemistry, anti-PVY sera was used as described by Grimes et al. [13]. Electron
micrographs were taken with a Zeiss EM10A electron microscope.
Virus challenge tests
R"progeny seed was surface sterilized as described above and plated on tissue culture
media containing 50 µg ml−" kanamycin. Plantlets that germinated and survived to the
three leaf stage, and thus were homo- or heterozygous for the gene of interest, were

70 M. Naderi and P. H. Berger
transplanted to soil and mechanically inoculated with PVY. Inoculum was prepared
by grinding PVY-infected tobacco in phosphate buffer (0±05 , pH 7±0) at 1:5 (w}v),
aliquoting this suspension and freezing aliquots at ®80 °C until use, when it was mixed
with carborundum. Ten plants per line were inoculated, and samples for ELISA tests
were collected just prior to inoculation and at 5 day intervals thereafter. Virus titre was
determined by relative amounts of helper component produced using anti-PVY HC
(kindly provided by D. Thornbury, University of Kentucky).
RESULTS
Symptom de�elopment in TP-CP+ plants
Leaf discs of N. tabacum cv. Xanthi were infected with A. tumefaciens harbouring
pMNLX1.0 (Fig. 1) and transformants selected on different concentrations of
kanamycin up to 300 mg l−". When only PVY-CP (pKYLX68.1) lacking the rbcs
transit peptide (CP+) was transformed into tobacco, nearly all CP+ transformants were
green and phenotypically normal, regardless of which kanamycin concentration was
used (Table 1). Two of these lines were chosen for comparative purposes, based on
western analysis (see below). The few white shoots that did develop initially became
green and developed normally after transfer onto non-selective media (data not shown)
and appeared to be non-transformed, antibiotic sensitive plants which had escaped
selection during transformation. Antibiotic resistant TP-CP+ plantlets or calli could
only be obtained using kanamycin concentrations of 50 mg l−" or less. Many TP-CP+
transformants were chlorotic or pale green (Table 1). Chlorotic plants stopped growing
after two to five leaves developed. Some of these chlorotic TP-CP+ plantlets were
transferred onto non-selective media where they remained chlorotic (Table 1),
indicating that this phenotype was not the characteristic response of non-transformed
plants to toxic levels of kanamycin.
Some primary TP-CP+ pale green transgenic plants started showing white mosaic
(white and green areas) symptoms 4 weeks after transfer to soil. These symptoms were
generally pronounced on the adaxial surface, but the abaxial leaf surface had no
apparent symptoms (data not shown). When white spot symptoms progressed beyond
the upper cell layers, leaves became deformed (data not shown). Figure 2 shows a
kanamycin resistant R"progeny TP-19 from plants with these symptoms. These plants
showed symptoms on both adaxial and abaxial leaf surfaces, and on both selective and
non-selective media, indicating that white mosaic symptoms were not due to side-
effects of kanamycin treatment. These disease-like symptoms, therefore, were not due
to culture conditions but rather to the presence of the TP-CP allele. In addition, the
more rapid and pronounced development of symptoms in the R"progeny compared to
the primary transformants may indicate that the phenotype penetrance increases with
gene dosage. Moreover, white mosaic symptoms are more pronounced along the leaf
vein system (Fig. 2). This could be explained by higher activity of 35S promoter along
the vein system in transgenic plants [2]. Many TP-CP+ transgenic plantlets that
survived initial selection at 50 mg l−" kanamycin were severely chlorotic and unable to
develop further. No shoot regeneration was observed when kanamycin at 300 mg l−"
was used. Since the number of integrated genes tends to increase as selection stringency

PVY coat protein and chloroplasts 71
F. 1. Organization of the binary vector pMNLX1.0 which was used to target PVY-CP intothe stroma compartment of tobacco chloroplasts. RB, right T-DNA border ; LB, left T-DNAborder ; P35 S#, cauliflower mosaic virus 35 S promoter with duplicated enhancers ; TP, transitpeptide of ribulose bisphosphate carboxylase small subunit from pea; CP, PVY coat protein;TCaMV, cauliflower mosaic virus 35 S transcriptional terminator ; Trbcs, ribulose bisphosphatecarboxylase transcriptional terminator.
T 1Effects of transformation of tobacco with PVY coat protein with (TP-CP) or without (CP) the rbcs transitpeptide (TP). Numbers of transformants based on number of calli after selection on 50 mg l−" kanamycin
for TP-CP or 300 mg l−" for CP
TP-CP
chlorotic
Experiment No. transformants No. %
1 41 12 292 74 66 89Total 115 78 68
CP
1 40 2 52 50 4 8Total 90 6 7

72 M. Naderi and P. H. Berger
increases [45], it is possible that higher concentrations of CP in chloroplasts were
detrimental or perhaps lethal to the plant cell. CP+ plants could be obtained at high
kanamycin concentrations and these did not develop any symptoms and remained
normal.
F. 2. White mosaic symptom development on TP-CP+ transformant which were selected on50 mg l−" kanamycin. R1 seedlings from the white mosaic TP-CP+ transformant TP-19.
Expression of TP-CP and PVY-CP transcripts
We were unable to maintain the most severely affected TP-CP+ plants. However,
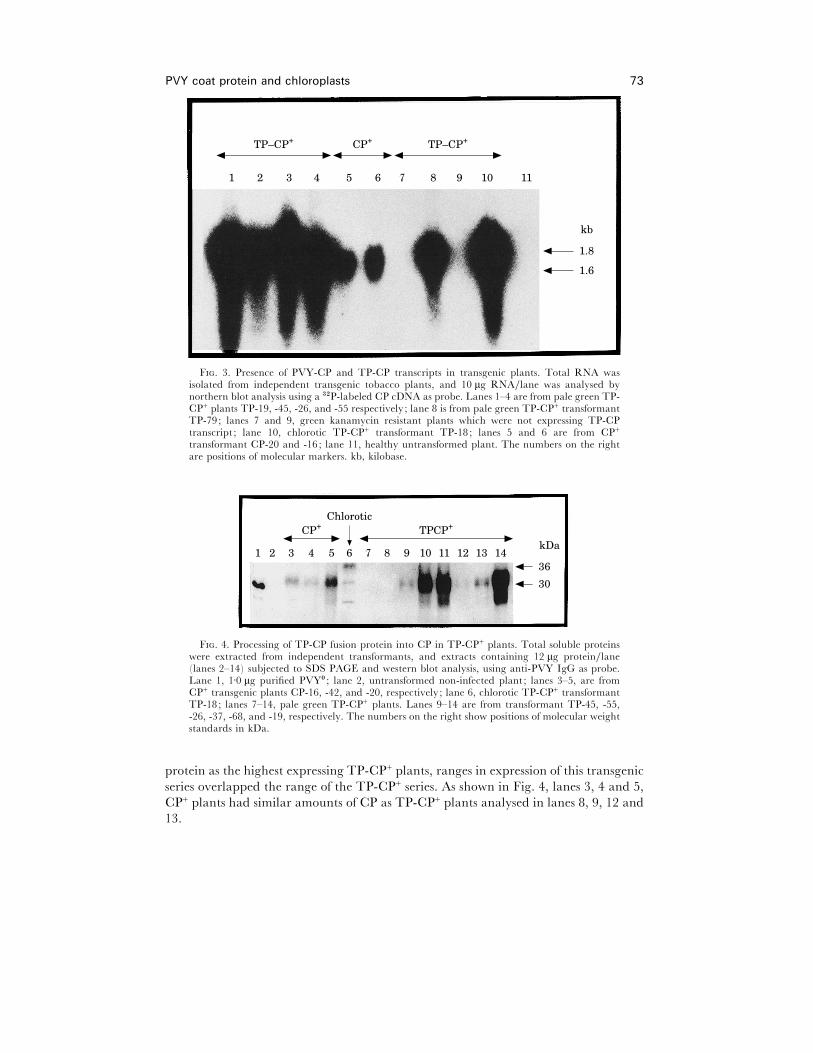
northern blot analysis (Fig. 3) showed that chlorotic (Fig. 3, lane 10), pale green TP-
CP+ (Fig. 3, lanes 1–4 and 8), and green CP+ plants (Fig. 3, lanes 5 and 6) expressed
TP-CP and PVY-CP transcripts corresponding to their expected sizes (PVY-CPE1±6 kb and TP-CPE 1±8 kb, respectively). In order to show that the RNA was being
translated in the plants, protein extracts were analysed using anti-PVY serum (Fig. 4).
Several polypeptides were recognized by the antiserum that had mobilities
corresponding to unprocessed TP-CP fusion protein (PVY-CPE 30 kDaTPE6 kDa), mature PVY-CP, and smaller peptides (Fig. 4, lane 6). The smaller peptides
were probably break-down products of PVY-CP. Since rbcs TP-mediated protein
import and processing into chloroplasts is normally very efficient and rapid (Fig. 4,
lanes 9–14) such that unprocessed protein is often undetected in transgenic plants
[6, 8, 14], we believe the unprocessed product, which was apparent only in samples
from chlorotic TP-CP+ plants, might be an indication of the poor physiological state
of these plantlets and their chloroplasts.
Western blot analysis demonstrated that PVY-CP was synthesized in CP+ transgenic
plants (Fig. 4, lanes 3–5). While none of the CP+ plants analysed accumulated as much
PVY coat protein and chloroplasts 73
1 11102 3 4 5 6 7 8 9
TP–CP+ TP–CP+CP+
1.8
1.6
kb
F. 3. Presence of PVY-CP and TP-CP transcripts in transgenic plants. Total RNA wasisolated from independent transgenic tobacco plants, and 10 µg RNA}lane was analysed bynorthern blot analysis using a $#P-labeled CP cDNA as probe. Lanes 1–4 are from pale green TP-CP+ plants TP-19, -45, -26, and -55 respectively; lane 8 is from pale green TP-CP+ transformantTP-79; lanes 7 and 9, green kanamycin resistant plants which were not expressing TP-CPtranscript ; lane 10, chlorotic TP-CP+ transformant TP-18; lanes 5 and 6 are from CP+
transformant CP-20 and -16; lane 11, healthy untransformed plant. The numbers on the rightare positions of molecular markers. kb, kilobase.
1 102 3 4 5 6 7 8 9
CP+ TPCP+
36
30
kDa11 12 13 14
Chlorotic
F. 4. Processing of TP-CP fusion protein into CP in TP-CP+ plants. Total soluble proteinswere extracted from independent transformants, and extracts containing 12 µg protein}lane(lanes 2–14) subjected to SDS PAGE and western blot analysis, using anti-PVY IgG as probe.Lane 1, 1±0 µg purified PVY! ; lane 2, untransformed non-infected plant ; lanes 3–5, are fromCP+ transgenic plants CP-16, -42, and -20, respectively; lane 6, chlorotic TP-CP+ transformantTP-18; lanes 7–14, pale green TP-CP+ plants. Lanes 9–14 are from transformant TP-45, -55,-26, -37, -68, and -19, respectively. The numbers on the right show positions of molecular weightstandards in kDa.
protein as the highest expressing TP-CP+ plants, ranges in expression of this transgenic
series overlapped the range of the TP-CP+ series. As shown in Fig. 4, lanes 3, 4 and 5,
CP+ plants had similar amounts of CP as TP-CP+ plants analysed in lanes 8, 9, 12 and
13.
74 M. Naderi and P. H. Berger
F. 5. For legend see opposite.

PVY coat protein and chloroplasts 75
Effects on chloroplast structure and de�elopment
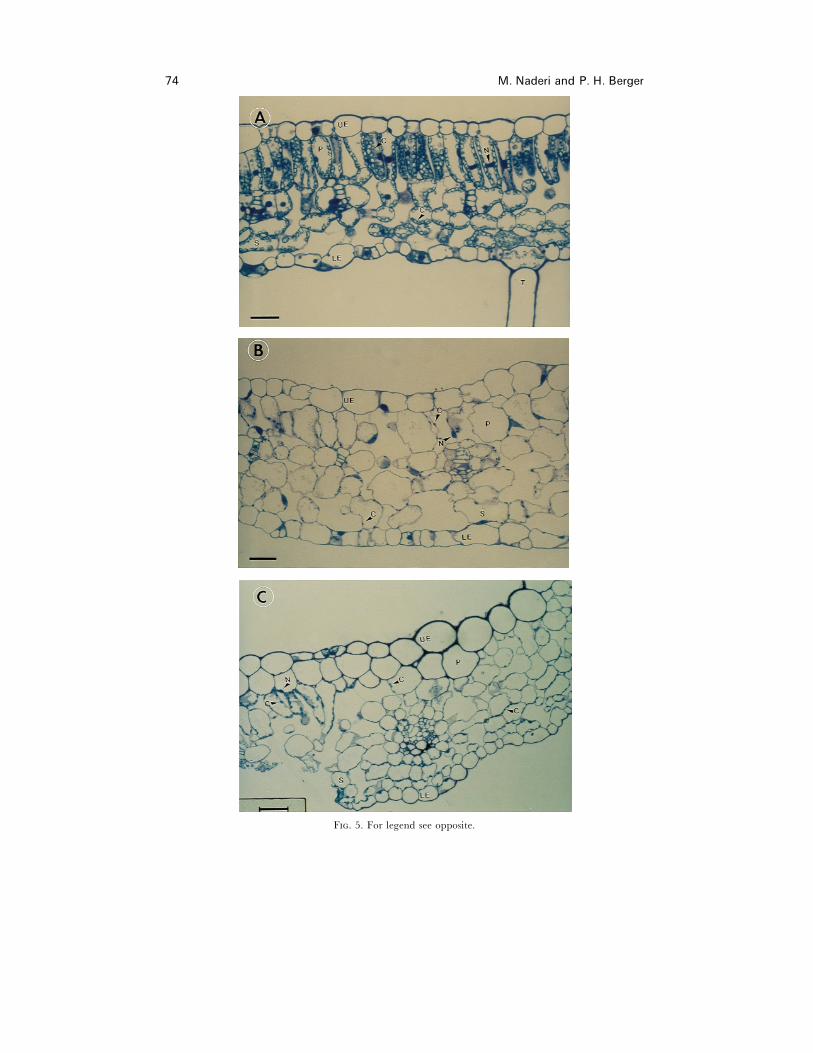
Cell morphology did not change in CP+ plants (data not shown), but light microscopy
studies indicated that chloroplast-directed PVY-CP affected cell and chloroplast
development (Fig. 5). Figure 5 represents typical cellular changes which were observed
in different TP-CP+ plants with mosaic symptoms. Cells were enlarged, cell shape
altered, and the number and size of chloroplasts decreased in chlorotic TP-CP+ plants
(Fig. 5B). TP-CP+ plant TP-19 was selected to show the differences between adjacent
green and white areas in plants showing symptoms. Cell enlargement, cell shape
alteration, and a decrease in number and size of chloroplasts were more pronounced
in white areas of TP-CP+ transformant TP-19 with white mosaic symptoms than in
nearby green areas (Fig. 5C). More chloroplasts could be seen, for example, as the field
of view moved away from white (centre) areas towards green (left) areas (Fig. 5C).
Control tobacco plants had 16±2 chloroplasts per palisade parenchyma cell, while TP-
CP+ plant TP-19, as an example, had 5±2 chloroplasts per palisade parenchyma cell.
In equivalent areas of tissue sections, TP-CP+ plants had 38% fewer cells.
Since we observed chlorosis (Fig. 2), and light microscopy indicated that cell size and
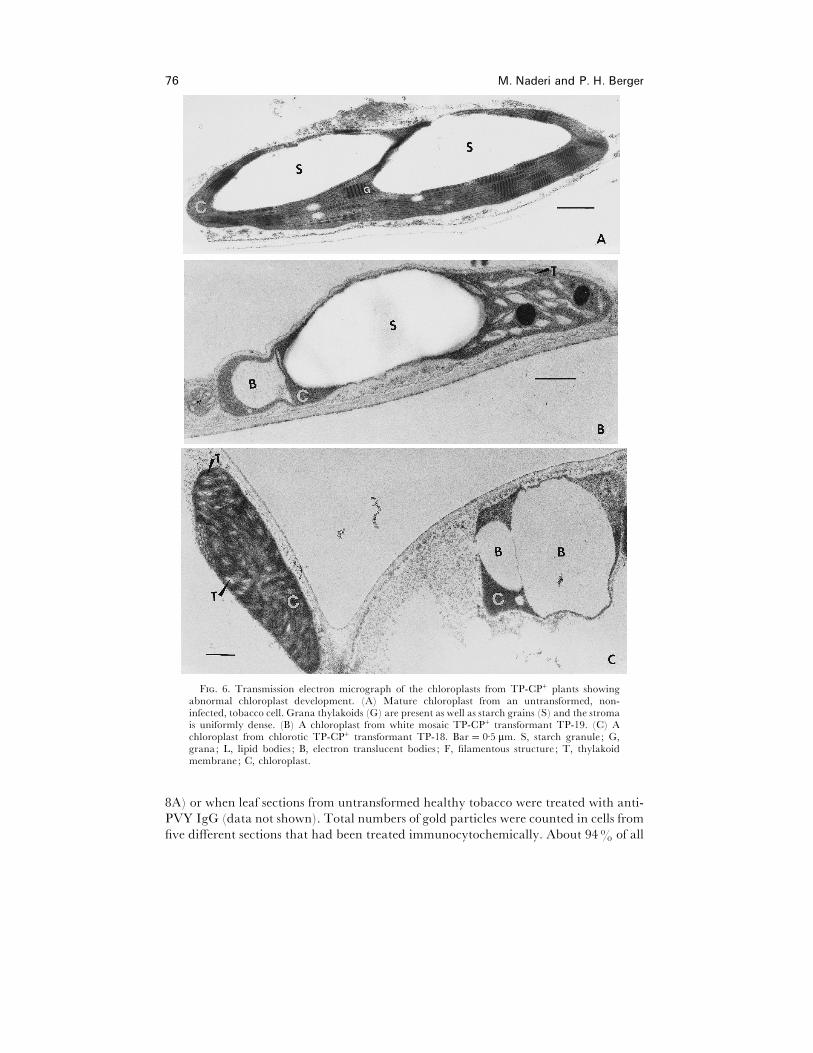
chloroplast number had decreased in TP-CP+ plants, we used TEM to investigate the
ultrastructure of TP-CP+ plants. Fig. 6 shows a TEM analysis of TP-CP+ transformants
TP-18 (Fig. 6C) and TP-19 (Fig. 6B). Thylakoid membrane stacking was abnormal
and the number of starch granules per chloroplast reduced in the transformants (Fig.
6B, C) in comparison with untransformed plants (Fig. 6A). Expression of TP-CP
resulted in the formation of filamentous or rod shaped structures in some chloroplasts
(Fig. 8A and C). The chloroplasts in Fig. 6B were typical for those observed in the
border area between green (left) and white (right) sections in white mosaic TP-CP+
transformant TP-19. Thylakoid membrane stacking appeared abnormal in chloroplasts
from green areas of TP-CP+ transformant TP-19 with white mosaic symptoms (data
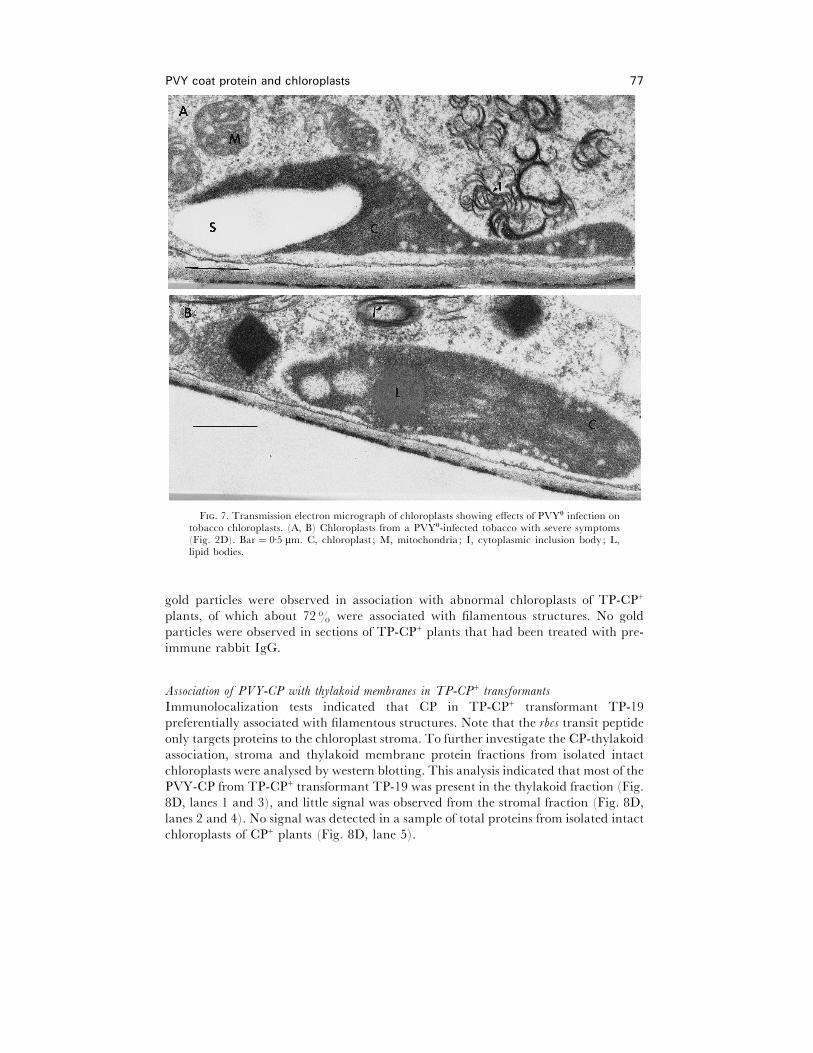
not shown). The ultrastructure of chloroplasts from PVY-infected plants was also
changed (Fig. 7A and B). In contrast to TP-CP+ plants, there were no apparent
ultrastructural alterations in chloroplast structure of CP+ plants (data not shown).
Localization of PVY-CP in abnormal chloroplasts from TP-CP+ transformants
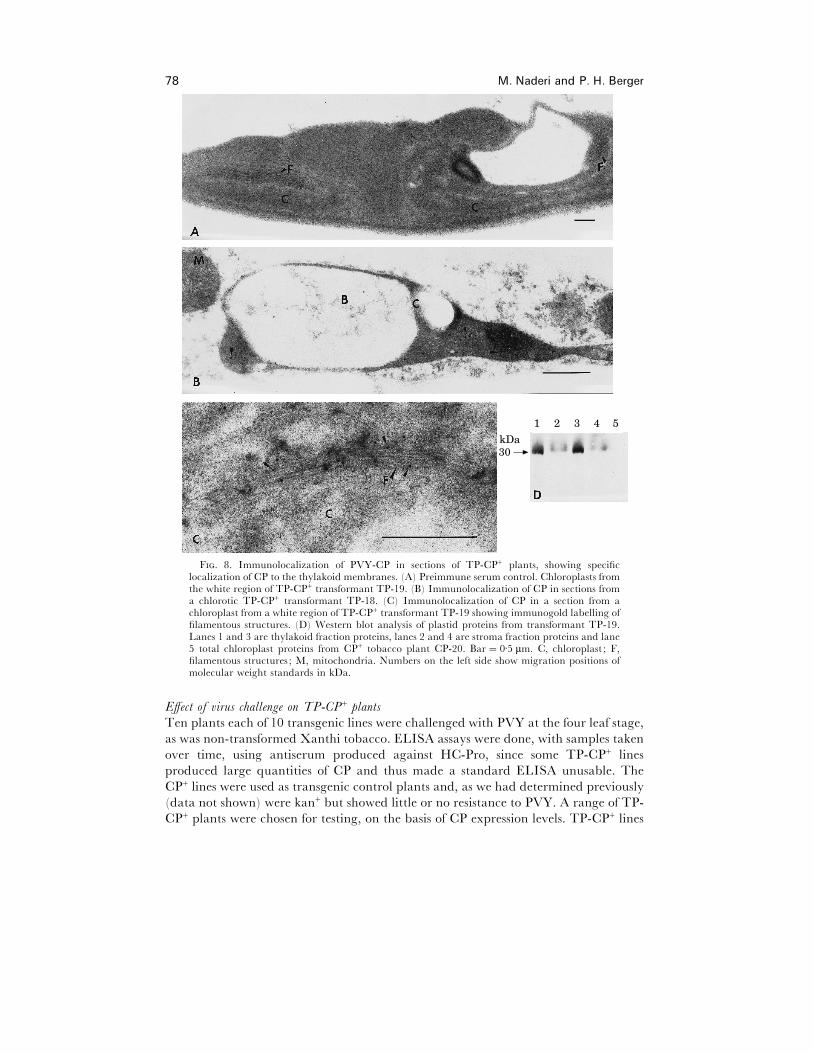
The presence and subcellular localization of CP in chloroplasts of TP-CP+ plants was
determined by immunocytochemistry and western blot analysis using anti-PVY IgG
(Fig. 8B–D). TEM immunocytochemistry of chlorotic transformant TP-18, and white
mosaic transformant TP-19 indicated that PVY-CP was localized in abnormal
chloroplasts (Fig. 8B and C). PVY-CP was localized mainly in filamentous structures
(Fig. 8C). No labelling was observed in normal chloroplasts from green areas of white
mosaic TP-CP+ plants (data not shown), indicating that chloroplast abnormalities
were correlated with the presence of CP inside these organelles. No labelling was
observed when sections from chlorotic tissue were treated with preimmune serum (Fig.
F. 5. Abnormal cell and chloroplast development associated with chloroplast-directed PVY-CP. (A) Leaf cross section from an untransformed non-infected tobacco. (B) Leaf cross-sectionfrom chlorotic TP-CP+ transformant TP-18. (C) Leaf cross-section from white mosaic area of aTP-CP+ transformant TP-19. Bar¯ 40 µm. UE, upper epidermis ; P, palisade parenchyma; S,spongy mesophyll. ; C, chloroplast ; N, nucleus ; LE, lower epidermis ; T, trichome.
76 M. Naderi and P. H. Berger
F. 6. Transmission electron micrograph of the chloroplasts from TP-CP+ plants showingabnormal chloroplast development. (A) Mature chloroplast from an untransformed, non-infected, tobacco cell. Grana thylakoids (G) are present as well as starch grains (S) and the stromais uniformly dense. (B) A chloroplast from white mosaic TP-CP+ transformant TP-19. (C) Achloroplast from chlorotic TP-CP+ transformant TP-18. Bar¯ 0±5 µm. S, starch granule ; G,grana; L, lipid bodies ; B, electron translucent bodies ; F, filamentous structure ; T, thylakoidmembrane; C, chloroplast.
8A) or when leaf sections from untransformed healthy tobacco were treated with anti-
PVY IgG (data not shown). Total numbers of gold particles were counted in cells from
five different sections that had been treated immunocytochemically. About 94% of all
PVY coat protein and chloroplasts 77
F. 7. Transmission electron micrograph of chloroplasts showing effects of PVY! infection ontobacco chloroplasts. (A, B) Chloroplasts from a PVY!-infected tobacco with severe symptoms(Fig. 2D). Bar¯ 0±5 µm. C, chloroplast ; M, mitochondria; I, cytoplasmic inclusion body; L,lipid bodies.
gold particles were observed in association with abnormal chloroplasts of TP-CP+
plants, of which about 72% were associated with filamentous structures. No gold
particles were observed in sections of TP-CP+ plants that had been treated with pre-
immune rabbit IgG.
Association of PVY-CP with thylakoid membranes in TP-CP+ transformants
Immunolocalization tests indicated that CP in TP-CP+ transformant TP-19
preferentially associated with filamentous structures. Note that the rbcs transit peptide
only targets proteins to the chloroplast stroma. To further investigate the CP-thylakoid
association, stroma and thylakoid membrane protein fractions from isolated intact
chloroplasts were analysed by western blotting. This analysis indicated that most of the
PVY-CP from TP-CP+ transformant TP-19 was present in the thylakoid fraction (Fig.
8D, lanes 1 and 3), and little signal was observed from the stromal fraction (Fig. 8D,
lanes 2 and 4). No signal was detected in a sample of total proteins from isolated intact
chloroplasts of CP+ plants (Fig. 8D, lane 5).
78 M. Naderi and P. H. Berger
1 2 3 4 5
30kDa
F. 8. Immunolocalization of PVY-CP in sections of TP-CP+ plants, showing specificlocalization of CP to the thylakoid membranes. (A) Preimmune serum control. Chloroplasts fromthe white region of TP-CP+ transformant TP-19. (B) Immunolocalization of CP in sections froma chlorotic TP-CP+ transformant TP-18. (C) Immunolocalization of CP in a section from achloroplast from a white region of TP-CP+ transformant TP-19 showing immunogold labelling offilamentous structures. (D) Western blot analysis of plastid proteins from transformant TP-19.Lanes 1 and 3 are thylakoid fraction proteins, lanes 2 and 4 are stroma fraction proteins and lane5 total chloroplast proteins from CP+ tobacco plant CP-20. Bar¯ 0±5 µm. C, chloroplast ; F,filamentous structures ; M, mitochondria. Numbers on the left side show migration positions ofmolecular weight standards in kDa.
Effect of �irus challenge on TP-CP+ plants
Ten plants each of 10 transgenic lines were challenged with PVY at the four leaf stage,
as was non-transformed Xanthi tobacco. ELISA assays were done, with samples taken
over time, using antiserum produced against HC-Pro, since some TP-CP+ lines
produced large quantities of CP and thus made a standard ELISA unusable. The
CP+ lines were used as transgenic control plants and, as we had determined previously
(data not shown) were kan+ but showed little or no resistance to PVY. A range of TP-
CP+ plants were chosen for testing, on the basis of CP expression levels. TP-CP+ lines
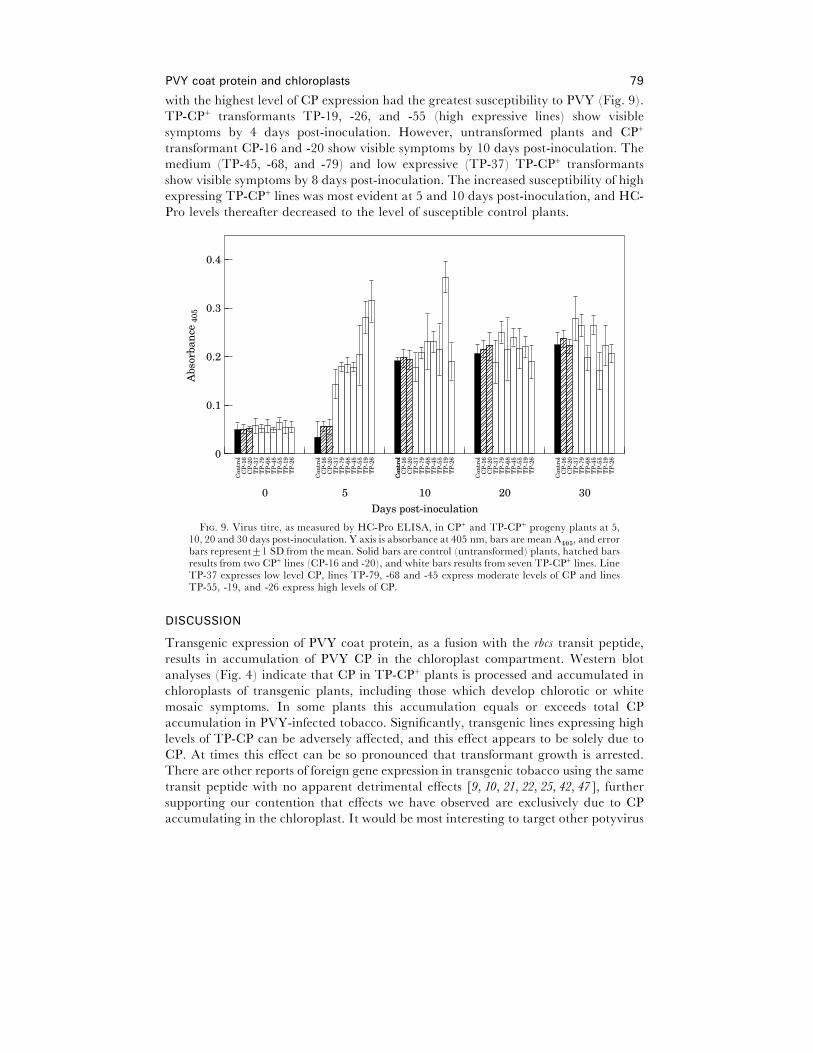
PVY coat protein and chloroplasts 79
with the highest level of CP expression had the greatest susceptibility to PVY (Fig. 9).
TP-CP+ transformants TP-19, -26, and -55 (high expressive lines) show visible
symptoms by 4 days post-inoculation. However, untransformed plants and CP+
transformant CP-16 and -20 show visible symptoms by 10 days post-inoculation. The
medium (TP-45, -68, and -79) and low expressive (TP-37) TP-CP+ transformants
show visible symptoms by 8 days post-inoculation. The increased susceptibility of high
expressing TP-CP+ lines was most evident at 5 and 10 days post-inoculation, and HC-
Pro levels thereafter decreased to the level of susceptible control plants.
0.4
0
Days post-inoculation
Abs
orba
nce
405
0.3
0.2
0.1
Con
trol
CP
-16
CP
-20
TP
-37
TP
-79
TP
-68
TP
-45
TP
-55
TP
-19
TP
-26
Con
trol
CP
-16
CP
-20
TP
-37
TP
-79
TP
-68
TP
-45
TP
-55
TP
-19
TP
-26
Con
trol
CP
-16
CP
-20
TP
-37
TP
-79
TP
-68
TP
-45
TP
-55
TP
-19
TP
-26
Con
trol
CP
-16
CP
-20
TP
-37
TP
-79
TP
-68
TP
-45
TP
-55
TP
-19
TP
-26
Con
trol
CP
-16
CP
-20
TP
-37
TP
-79
TP
-68
TP
-45
TP
-55
TP
-19
TP
-26
Con
trol
0 5 10 20 30
F. 9. Virus titre, as measured by HC-Pro ELISA, in CP+ and TP-CP+ progeny plants at 5,10, 20 and 30 days post-inoculation. Y axis is absorbance at 405 nm, bars are mean A
%!&, and error
bars represent³1 SD from the mean. Solid bars are control (untransformed) plants, hatched barsresults from two CP+ lines (CP-16 and -20), and white bars results from seven TP-CP+ lines. LineTP-37 expresses low level CP, lines TP-79, -68 and -45 express moderate levels of CP and linesTP-55, -19, and -26 express high levels of CP.
DISCUSSION
Transgenic expression of PVY coat protein, as a fusion with the rbcs transit peptide,
results in accumulation of PVY CP in the chloroplast compartment. Western blot
analyses (Fig. 4) indicate that CP in TP-CP+ plants is processed and accumulated in
chloroplasts of transgenic plants, including those which develop chlorotic or white
mosaic symptoms. In some plants this accumulation equals or exceeds total CP
accumulation in PVY-infected tobacco. Significantly, transgenic lines expressing high
levels of TP-CP can be adversely affected, and this effect appears to be solely due to
CP. At times this effect can be so pronounced that transformant growth is arrested.
There are other reports of foreign gene expression in transgenic tobacco using the same
transit peptide with no apparent detrimental effects [9, 10, 21, 22, 25, 42, 47], further
supporting our contention that effects we have observed are exclusively due to CP
accumulating in the chloroplast. It would be most interesting to target other potyvirus

80 M. Naderi and P. H. Berger
proteins to the chloroplasts and determine if similar effects are observed, particularly
HC-Pro, as we know this gene product is present in infected chloroplasts [15].
Detrimental effects due to TP-CP expression can be observed at the macroscopic and
microscopic levels. While the symptoms of PVY are not faithfully reproduced in
abnormal cases, the symptoms seen are reminiscent of virus infection. Light microscopy
(Fig. 5) and electron microscopy (Fig. 6) reveals that chloroplast targeted PVY-CP
disrupts chloroplast development and structure in cells. It also reduces chloroplast
numbers per cell, and these cells appear less differentiated. Mesophyll cells in yellow
areas of infected leaves with mosaic symptoms are commonly less differentiated with
fewer chloroplasts and fewer or no intracellular spaces [28].
Western analysis of fractionated chloroplast proteins from TP-CP plants indicates
that chloroplastic CP is associated with the thylakoid membrane fraction. This
observation is consistent with our previous observation that chloroplast-associated CP
from infected plants is almost exclusively associated with the thylakoid membrane
system, a situation very similar to what has been reported for tobacco mosaic virus
(TMV) [34], and consistent with our previous observations on PVY-infected tobacco
[15]. Our observation of abnormal thylakoids and lack of chlorophyll (chlorosis—Fig.
2) suggests that their function is also probably impaired, which is consistent with
previous reports on TMV [17, 35] and a potyvirus that photosystem II is adversely
affected after virus infection [29, 30]. Since CP also associates with the thylakoids in the
absence of any other virus-encoded proteins, there must be an inherent property of CP
that directs this protein to the thylakoids, possibly in association with chloroplast
protein(s).
The presence of CP in chloroplasts (Fig. 8D) and the appearance of filamentous
structures, as observed by TEM (Fig. 8A and C), suggests that these may be aggregates
of CP, and thus with sufficient quantities of CP these molecules self-assemble or
aggregate spontaneously inside chloroplasts. Additionally, the approximate diameter
of these filamentous structures, as determined by comparison with the 10 nm gold
particles, is within the range of the diameter of PVY virions [20, 28]. Considering the
high level of CP expression in some TP-CP+ transformants and the fact that CP is
concentrated within the chloroplasts, the appearance of these potyvirus-like rods is not
surprising. Whether these particles have encapsidated any chloroplast RNA is
unknown, but assembly of potyvirus CP molecules into virus-like particles in the
absence of the viral RNA has been reported by Jagadish et al. [20].
TP-CP+ lines with the highest expression levels of TP-CP are more susceptible to
PVY. This observation indirectly supports the hypothesis that CP is toxic to the
chloroplast and CP that is already present in the organelle, after import of TP-CP, is
additive in terms of disease.
Our data indicate that PVY-CP action in host chloroplasts can affect this organelle
and even induce symptoms suggestive of virus infection, but it is unlikely that this is the
sole explanation for the symptoms we see in potyvirus-infected plants. For example,
Lindbeck et al. [24] showed that there is a correlation between the production of
mutated TMV coat protein and chloroplast degradation. Very little of this mutated
CP was detected in the chloroplasts of infected cells by immunolocalization, and the CP
was mainly localized to discrete-staining coat protein bodies in the cytoplasm. They
suggested that the mutated CP exerted its influence on chloroplasts from the cytoplasm

PVY coat protein and chloroplasts 81
instead of from inside of the chloroplasts. Recently, Masuta et al. [26] reported that
chlorosis induced by CMV Y satellite RNA (Y-satRNA) is attributed to a massive
reduction in chlorophyll content. The chlorosis induction by Y-satRNA was apparently
due to specific inhibition of chlorophyll accumulation rather than to chloroplast
degradation since chloroplast ultrastructure was not adversely affected. In the case of
tobacco vein mottling potyvirus (TVMV), the 3«-untranslated region of the genome
can have a direct effect on symptom severity [36].
This work could also have implications for the use of coat protein-mediated
resistance for crop protection. In lieu of a known mechanism for potyvirus CP import
into chloroplasts, if CP somehow enters chloroplasts of transgenic plants expressing
viral CP in the cytoplasm, one could speculate that the transgenic plants could lose
their acquired CP-mediated resistance. If enough CP enters host cell chloroplasts, there
could also be deleterious effects on photosynthesis. Of course, the quantity of CP
expressed in most transgenic plants is considerably less than we have seen with some
TP-CP+ plants, but lower levels of CP in chloroplasts could presumably have subtle
effects on plant vigour. Many groups generate transgenic plants which express viral CP
but do not show CP-mediated-resistance. Perhaps this could be due, in part, to import
of CP into chloroplasts of transgenic plants which is then manifested by susceptibility
of transformants or lack of resistance.
There are many aspects of PVY-host chloroplast interactions which remain
unresolved. Our work shows that the presence of PVY-CP inside chloroplasts induces
disease-like symptoms. Our data suggest that CP contains a thylakoid targeting
mechanism, but do not indicate how CP is imported into chloroplasts in infected
plants. Nor do we have any indication as to whether or not virus RNA translation
occurs in organello but we do know that PVY RNA is present in chloroplasts of infected
plants [15]. Further experimentation should address these questions as to how specific
chloroplast biochemical functions are affected by CP action inside the organelle. As
mentioned above, we have also observed the HC-pro protein in chloroplasts [15], but
have no information as to whether this gene product (or perhaps others from the PVY
genome) affects chloroplast function. The present study shows, nonetheless, that PVY-
CP is toxic to chloroplasts, at least under the conditions of our experiments, and may
be a factor in symptom development in infected plants.
We wish to thank A. Caplan for helpful suggestions, F. Bailey for assistance with
electron microscopy, P. J. Shiel for help with tissue culture media, S. Chowrira and
V. Francheschi for assistance with electron microscopy immunolocalization, and A.
Caplan, S. Wyatt, and M. Wiese for critical reading of the manuscript. This is Idaho
Agricultural Experiment Station publication No. 94728.
REFERENCES
1. An G. 1987. Binary Ti vectors for plant transformation and promoter analysis. Methods in Enzymology153 : 292–305.
2. Benfey PH, Chua N-H. 1989. Regulated genes in transgenic plants. Science.3. Cline K, Werner-Washburne M, Andrews J, Keegstra K. 1984. Thermolysin is a suitable protease
for probing the surface of intact pea chloroplasts. Plant Physiology 75 : 675–678.4. Collmer C, Howell S. 1992. Role of satellite RNA in the expression of symptoms caused by plant
viruses. Annual Re�iew of Plant Pathology 30 : 419–442.

82 M. Naderi and P. H. Berger
5. Culver, JN, Lindbeck AGC, Dawson WO. 1991. Virus-host interactions : induction of chlorotic andnecrotic responses in plants by tobamoviruses. Annual Re�iew of Plant Pathology 29 : 193–217.
6. Dahlin C, Cline K. 1991. Developmental regulation of the plastid protein import apparatus. Plant Cell
3 : 1131–1140.7. Dawson WO, Bubrick P, Grantham GL. 1988. Modification of the tobacco mosaic virus coat protein
gene affecting replication, movement and symptomatology. Phytopathology 78 : 783–789.8. de Boer D, Bakker H, Lever A, Bouma T, Salentijn E, Weisbeek P. 1991. Protein targeting
towards the thylakoid lumen of chloroplasts : proper localization of fusion protein is only observed invivo. EMBO Journal 10 : 2765–2772.
9. de Boer G, Cremers F, Teertstra R, Smits L, Hille J, Smeekens S, Weisbeek P. 1988. In �i�o
import of plastocyanin and a fusion protein into developmentally different plastids of transgenicplants. EMBO Journal 7 : 2631–2635.
10. Della-Cioppa G, Bauer SC, Taylor ML, Rochester DE, Klein BK, Shah DM, Fraley RT,Kishore GM. 1987. Targeting of a herbicide-resistant enzyme from Escherichia coli to chloroplasts ofhigher plants. Bio}technology 5 : 579–584.
11. Devic M, Jaegle M, Baulcombe D. 1989. Symptom production on tobacco and tomato is determinedby two distinct domains of the satellite RNA of cucumber mosaic virus (strain Y). Journal of General
Virology 70 : 2765–2774.12. Fish LE, Jagendorf AT. 1982. High rates of protein synthesis by isolated chloroplasts. Plant Physiology
70 : 1107–1114.13. Grimes HD, Overvoorde PJ, Ripp K, Francheschi VR, Hitz WD. 1992. A 62-kDa sucrose binding
protein is expressed and localized in tissues actively engaged in sucrose transport. Plant Cell 4 :1561–1574.
14. Guerineau F, Woolston S, Brooks L, Mullineaux P. 1988. An expression cassette for targetingforeign proteins into chloroplasts. Nucleic Acids Research 16 : 11380.
15. Gunasinghe UB, Berger PH. 1991. RNA and proteins of potato virus Y and their association withchloroplasts of infected tobacco. Molecular Plant Microbe Interactions 5 : 452–457.
16. Heaton LA, Lee TC, Wei N, Morris TJ. 1991. Point mutations in the turnip crinkle virus capsidprotein affect the symptoms expressed by Nicotiana benthamiana. Virology 183 : 143–150.
17. Hodgson RAJ, Beachy RN, Pakrasi HB. 1989. Selective inhibition of photosystem II in spinach bytobacco mosaic virus : an effect of the viral coat protein. FEBS Letters 245 : 267–270.
18. Holt CA, Hodgson RAJ, Coker FA, Beachy RN, Nelson RS. 1990. Characterization of the maskedstrain of tobacco mosaic virus : identification of the region responsible for symptom attenuation byanalysis of an infectious cDNA clone. Molecular Plant Microbe Interactions 3 : 417–423.
19. Jaegle M, Devic M, Longstaff M, Baulcombe D. 1990. Cucumber mosaic virus satellite RNA (Ystrain) : analysis of sequences which affect yellow mosaic symptoms on tobacco. Journal of General
Virology 71 : 1905–1912.20. Jagadish MN, Ward CW, Gough KH, Tulloch PA, Whittaker LA, Shukla DD. 1991. Expression
of potyvirus coat protein in Escherichia coli and yeast and its assembly into virus-like particles. Journal
of General Virology 72 : 1543–1550.21. Kavanagh TA, Jefferson RA, Bevan MW. 1988. Targeting a foreign protein to chloroplasts using
fusions to the transit peptide of a chlorophyll a}b protein. Molecular and General Genetics 215 : 38–45.22. Ko K, Cashmore AR. 1989. Targeting of proteins to the thylakoid lumen by the bipartite transit
peptide of the 33 kDa oxygen-evolving protein. EMBO Journal 8 : 3187–3194.23. Laemmli UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature 227 : 680–685.24. Lindbeck, AGC, Dawson WO, Thomson WW. 1991. Coat protein-related polypeptides from in �itro
tobacco mosaic virus coat protein mutants do not accumulate in the chloroplasts of directly inoculatedleaves. Molecular Plant Microbe Interactions 4 : 89–94.
25. Lubben TH, Gatenby AA, Ahlquist P, Keegstra K. 1989. Chloroplast import characteristics ofchimeric proteins. Plant Molecular Biology 12 : 13–18.
26. Masuta C, Suzuki M, Matsuzaki T, Honda J, Kuwata S, Takanami Y, Koiwai A. 1993. Brightyellow chlorosis by cucumber mosaic virus Y satellite RNA is specifically induced without severechloroplast damage. Physiological and Molecular Plant Pathology 42 : 267–278.
27. Masuta C, Takanami Y. 1989. Determination of sequence and structural requirements forpathogenicity of a cucumber mosaic virus satellite RNA (Y-satRNA). Plant Cell 1 : 1165–1173.
28. Matthews REF. 1991. Plant Virology. San Diego: Academic Press.29. Naidu RA, Krishnan M, Nayudu MV, Gnanam A. 1984. Studies on peanut green mosaic virus
infected peanut (Arachis hypogaea L.) leaves. II. Chlorophyll-protein complexes and polypeptidecomposition of thylakoid membranes. Physiological Plant Pathology 25 : 191–198.

PVY coat protein and chloroplasts 83
30. Naidu RA, Krishnan M, Ramanujam P, Gnanam, A, Nayadu MV. 1984. Studies on peanut greenmosaic virus infected peanut (Arachis hypogaea L.) leaves. I. Photosynthesis and photochemicalreactions. Physiological Plant Pathology 25 : 181–190.
31. Neeleman L, van der Kuyl AC, Bol J. 1991. Role of alfalfa mosaic virus coat protein gene in symptomformation. Virology 181 : 687–693.
32. Nishiguchi M, Kikuchi S, Kiho Y, Ohno T, Meshi T, Okada Y. 1985. Molecular basis of plant viralvirulence: the complete nucleotide sequence of an attenuated strain of tobacco mosaic virus. NucleicAcids Research 13 : 5585–5590.
33. Palukaitis P. 1988. Pathogenicity regulation by satellite RNAs of cucumber mosaic virus : minornucleotide sequence changes alter host responses. Molecular Plant Microbe Interactions 1 : 175–181.
34. Reinero A, Beachy RN. 1986. Association of TMV coat protein with chloroplast membranes in virus-infected leaves. Plant Molecular Biology 6 : 291–301.
35. Reinero A, Beachy RN. 1989. Reduced photosystem II activity and accumulation of viral coat proteinin chloroplasts of leaves infected with tobacco mosaic virus. Plant Physiology 89 : 111–116.
36. Rodrı!guez-Cerezo E, Klein PG, Shaw JG. 1991. A determinant of disease symptom severity is locatedin the 3«-terminal noncoding region of the RNA of a plant virus. Proceedings National Academy of Sciences,USA 88 : 9863–9867.
37. Roscoe TJ, Ellis RJ. 1982. Two-dimensional gel electrophoresis of chloroplast proteins. In: EdelmanM, Hallick RB, Chua N-H, eds. Methods in Chloroplast Molecular Biology. Amsterdam: ElsevierBiomedical Press, 1015–1028.
38. Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning. A Laboratory Manual. 2nd ed. ColdSpring Harbour Laboratory Press.
39. Schardl CL, Byrd AD, Benzion G, Altschuler MA, Hildebrand DF, Hunt AG. 1987. Design andconstruction of a versatile system for the expression of foreign genes in plants. Gene 61 : 1–11.
40. Shintaku MH, Zhang L, Palukaitis P. 1992. A single amino acid substitution in the coat protein ofcucumber mosaic virus induces chlorosis in tobacco. Plant Cell 4 : 751–757.
41. Sleat DE, Palukaitis P. 1992. A single nucleotide change within a plant virus satellite RNA alters thehost specificity of disease induction. Plant Journal 2 : 43–49.
42. Theg SM, Geske FJ. 1992. Biophysical characterization of a transit peptide directing chloroplastprotein import. Biochemistry 31 : 5053–5060.
43. Tranbarger TJ, Franceschi VR, Hildebrand DF, Grimes HD. 1991. The soybean 94-kiloDaltonvegetative storage protein is a lipoxygenase that is localized in paraveinal mesophyll cell vacuoles.Plant Cell 3 : 973–987.
44. Tu JC. 1967. Comparison of chloroplasts in maize dwarf mosaic virus-infected and healthy corn,sorghum, and Johnson grass. Phytopathology 57 : 834 (abstr.).
45. Vaeck M, Reynaerts A, Ho$ fte H, Jansens S, De Beuckeleer M, Dean C, Zabeau M, VanMontagu M, Leemans J. 1987. Transgenic plants protected from insect attack. Nature 328 : 33–37.
46. Van Loon LC. 1987. Disease induction by plant viruses. Ad�ances in Virus Research 33 : 205–255.47. Wong EY, Hironaka CM, Fischhoff DA. 1992. Arabidopsis thaliana small subunit leader and transit
peptide enhance the expression of Bacillus thuringiensis proteins in transgenic plants. Plant MolecularBiology 20 : 81–93.


























![[RECIPE] Korean Potato Pancake](https://img.pdfslide.net/doc/110x75/63136f91fc260b71020f1802/recipe-korean-potato-pancake.jpg)


