Embed Size (px)
Citation preview

Effects of Preceding Stimulation on Brain Activation in Response to ColonicDistention in HumansTOYOHIRO HAMAGUCHI, PHD, MICHIKO KANO, MD, PHD, MOTOYORI KANAZAWA, MD, PHD, MASATOSHI ITOH, MD, PHD,KAZUHIKO YANAI, MD, PHD, AND SHIN FUKUDO, MD, PHD
Objective: It has been suggested that the pattern of distension (moderate following mild and vice versa) might influence brainactivation and the experience of hypersensitivity, offset analgesia, and anticipation. Nevertheless, how the pattern of stimulationaffects sensitization and/or desensitization to visceral stimulation remains unknown. Methods: In 45 nonclinical healthy participants(12 women, 33 men; 20Y26 years old), brain processing of visceral sensation induced by colonic distension was examined using H2
15Opositron emission tomography. Subjective feelings regarding the stimuli were also measured. The descending colon was stimulatedusing six patterns of three bag pressures (0, 20, and 40 mmHg). To evaluate the neural sensitization to visceral stimulation arising fromthe precedence effect, the effects of a 20- or 40-mm Hg distention after a sham or 20- or 40-mm Hg distension were analyzed usingstatistical parametric mapping. The level of significance was set at a voxelwise level of p G .0001, with cluster extent sizes of k 9 50.Results: The midbrain, insula, and cerebellum, were more strongly activated by a 20-mm Hg distension with a preceding 40-mm Hgdistention than by a 20-mm Hg distention without a preceding stimulation (p G .0001). Conversely, a sham stimulation after theexperience of an intense stimulation activated the midcingulate cortex, compared with a sham stimulation without the experience ofactual visceral stimulation (p G .0001). Conclusions: By directly comparing different patterns of visceral stimuli, preceding visceralstimuli may affect neural sensitization and/or desensitization in humans, including elevated midbrain, insula, and midcingulate cortex.Key words: visceral sensitization, hyperalgesia, visceral perception, emotion, brain-gut interaction.
IBS = irritable bowel syndrome; PET = positron emissiontomography; ACC = anterior cingulate cortex; fMRI = functionalmagnetic resonance imaging; OFC = orbitofrontal cortex; PAG =periaqueductal gray; rCBF = regional cerebral blood flow.
INTRODUCTION
I rritable bowel syndrome (IBS) is a functional gastrointestinaldisorder characterized by chronic abdominal pain and ab-
normal bowel habituation (1,2). The symptoms of IBS areoften aggravated by stress, which alters colonic motility andvisceral perception (3Y5). Most patients with IBS exhibit hy-persensitivity in response to the distention of the gut lumen(6Y8). Several causative mechanisms explaining visceral hy-persensitivity have been proposed including inflammationinjury, stress, and abnormal pain sensitization (9,10).
The order of distension (moderate preceded by mild andvice versa) has been suggested to influence brain activationand the experience of hypersensitivity, offset analgesia, andanticipation. Repetitive colonic stimuli may induce visceralhyperalgesia in patients with IBS (8,11). In patients with IBS,repetitive stimulation of the sigmoid visceral afferents re-sults in the development of central sensitization, which mani-fests as hyperalgesia and increased viscerosomatic referralduring rectal distention as well as spontaneous visceral hyper-algesia in the absence of applied stimuli (12). A combinationof previous inflammation and repetitive colorectal distentionhas been shown to result in colon hypersensitivity in rats (10).
A neural concept explaining these mechanisms is the devel-opment of hyperexcitability in the neurons of the dorsal horn,which occurs either in response to peripheral tissue irritationor in response to descending influences in the central visceralpain pathway (13). Inflammatory mediators such as bradyki-nin, prostaglandin, 5-hydroxytryptamine, and histamine cansensitize nociceptors to heat. Sensitization to inflammatorymediators has been observed not only for cutaneous receptors(14,15) but also for visceral polymodal receptors (16,17). Thissensitization might only involve input from nociceptors be-cause the mechanical pain thresholds after a cutaneous injuryare of the same order as those of nociceptors (16). Neuralsensitization might also arise because of enhanced connectiv-ity between low-threshold mechanoreceptors (18). Previousreports suggest that tissue inflammation increases the sensi-tivity of the colon to mechanical stimuli, leading to enhancedresponses to colorectal distension (19,20). This enhancementinvolves the activation of spinal N-methyl-D-aspartate andnonYN-methyl-D-aspartate receptors, but not metabotropic re-ceptors (20,21).
Offset analgesia may also provide a set of novel end pointsto assess pathophysiological changes over time, inasmuch asthis phenomenon has been proposed to function as a temporalsharpening mechanism (22). Offset analgesia occurs duringdynamic noxious stimuli when a small incremental decrease inthe noxious stimulus intensity evokes a disproportionatelylarge decrease in the perceived pain ratings. The capsaicin-heatsensitization model has been shown to produce a centrallysensitized state in healthy humans, and the alteration of offsetanalgesia by neural changes induced during capsaicin-heatsensitization has been hypothesized (23). Rectal distentionactivates sensory and/or sensorimotor regions of the cortex(24). By contrast, the descending colon, which is purely in-nervated by the autonomic nervous system (25), is highlysensitive to stimuli in patients with IBS (8,26). Visceral hy-persensitivity thus differs from somatic hypersensitivity, whichis commonly associated with preceding stimuli.
Psychosomatic Medicine 75:453Y462 (2013) 4530033-3174/13/7505Y0453Copyright * 2013 by the American Psychosomatic Society
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
From the Departments of Behavioral Medicine (T.H., M.K., M.K., S.F.) andPharmacology (K.Y.), Graduate School of Medicine, and Cyclotron RadioIsotope Center (M.I.), Tohoku University, Aoba, Sendai, Japan.
Address correspondence and reprint requests to Shin Fukudo, MD, PhD,Department of Behavioral Medicine, Graduate School of Medicine, TohokuUniversity, 2-1 Seiryo, Aoba, Sendai 980-8575, Japan. E-mail: [email protected]
Received for publication December 21, 2011; revision received February21, 2013.
DOI: 10.1097/PSY.0b013e3182926682
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.
The anticipation of intense visceral stimuli induces theactivation of the orbitofrontal cortex (OFC), prefrontal cortex,perigenual anterior cingulate cortex (ACC), temporal cortex,thalamus, lentiform nucleus, and periaqueductal gray (PAG)region (27). This response resembles brain activation by theactual rectal stimulus (27). A previous study showed that pla-cebos affect the midbrain, which contains endogenous opoids,during analgesic anticipation (28). In that study (28), antici-pation reduced the ACC activity during the earlier phase ofpain and the thalamic and insula activity during the later phaseof pain. We previously reported that the main effect of a shamstimulation (0 mmHg) to the descending colon, compared withthe baseline condition, consisted of the activation of the OFC,inferior parietal cortex, and thalamus in healthy participants(29). In this previous study, the intensity of the colonic dis-tention was randomly varied (0, 20, or 40 mm Hg). Thisstimulation protocol might have produced intense anticipationof a higher stimulation intensity, which occurred in a totallyunpredictableway. Such anticipation and/or anxiety could haveproduced brain activation independent of that induced by co-lonic distention because the anticipation of visceral stimulation isconsidered a unique stressor capable of triggering an intero-ceptive neural response (30).
Despite the reported phenomena and concepts describedearlier, how the order of stimulation (moderate preceded bymild and vice versa) affects sensitization and/or desensitiza-tion to visceral stimulation remains unknown. We hypothesizedthat brain activation arising from colonic distention with anintense initial stimulation would differ from that arising fromintense stimulation preceded by mild stimulation. We alsohypothesized that brain activation in response to sham colo-nic distention occurring before an intense stimulation woulddiffer from that occurring after intense stimulation. Thus, wesuspected that the brain might behave differently in responseto different patterns of visceral stimulation.
METHODSParticipantsForty-five individuals (12 women, 33 men; 20Y26 years old, all right
handed) participated in this study. This study used a database of brain-gutimages obtained at Tohoku University. Therefore, some participants had beenincluded in previous reports from our institution (29,31,32), but the hypothesisof the present study was completely different from that of previous studies.
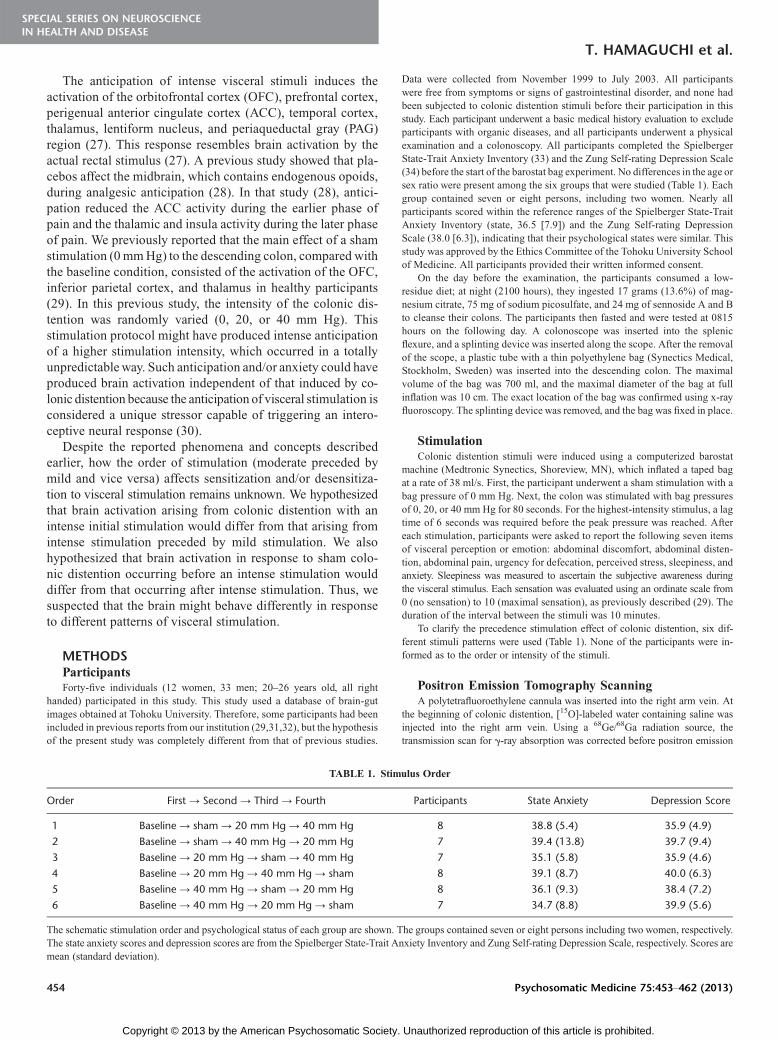
Data were collected from November 1999 to July 2003. All participantswere free from symptoms or signs of gastrointestinal disorder, and none hadbeen subjected to colonic distention stimuli before their participation in thisstudy. Each participant underwent a basic medical history evaluation to excludeparticipants with organic diseases, and all participants underwent a physicalexamination and a colonoscopy. All participants completed the SpielbergerState-Trait Anxiety Inventory (33) and the Zung Self-rating Depression Scale(34) before the start of the barostat bag experiment. No differences in the age orsex ratio were present among the six groups that were studied (Table 1). Eachgroup contained seven or eight persons, including two women. Nearly allparticipants scored within the reference ranges of the Spielberger State-TraitAnxiety Inventory (state, 36.5 [7.9]) and the Zung Self-rating DepressionScale (38.0 [6.3]), indicating that their psychological states were similar. Thisstudy was approved by the Ethics Committee of the Tohoku University Schoolof Medicine. All participants provided their written informed consent.
On the day before the examination, the participants consumed a low-residue diet; at night (2100 hours), they ingested 17 grams (13.6%) of mag-nesium citrate, 75 mg of sodium picosulfate, and 24 mg of sennoside A and Bto cleanse their colons. The participants then fasted and were tested at 0815hours on the following day. A colonoscope was inserted into the splenicflexure, and a splinting device was inserted along the scope. After the removalof the scope, a plastic tube with a thin polyethylene bag (Synectics Medical,Stockholm, Sweden) was inserted into the descending colon. The maximalvolume of the bag was 700 ml, and the maximal diameter of the bag at fullinflation was 10 cm. The exact location of the bag was confirmed using x-rayfluoroscopy. The splinting device was removed, and the bag was fixed in place.
StimulationColonic distention stimuli were induced using a computerized barostat
machine (Medtronic Synectics, Shoreview, MN), which inflated a taped bagat a rate of 38 ml/s. First, the participant underwent a sham stimulation with abag pressure of 0 mm Hg. Next, the colon was stimulated with bag pressuresof 0, 20, or 40 mm Hg for 80 seconds. For the highest-intensity stimulus, a lagtime of 6 seconds was required before the peak pressure was reached. Aftereach stimulation, participants were asked to report the following seven itemsof visceral perception or emotion: abdominal discomfort, abdominal disten-tion, abdominal pain, urgency for defecation, perceived stress, sleepiness, andanxiety. Sleepiness was measured to ascertain the subjective awareness duringthe visceral stimulus. Each sensation was evaluated using an ordinate scale from0 (no sensation) to 10 (maximal sensation), as previously described (29). Theduration of the interval between the stimuli was 10 minutes.
To clarify the precedence stimulation effect of colonic distention, six dif-ferent stimuli patterns were used (Table 1). None of the participants were in-formed as to the order or intensity of the stimuli.
Positron Emission Tomography ScanningA polytetrafluoroethylene cannula was inserted into the right arm vein. At
the beginning of colonic distention, [15O]-labeled water containing saline wasinjected into the right arm vein. Using a 68Ge/68Ga radiation source, thetransmission scan for F-ray absorption was corrected before positron emission
TABLE 1. Stimulus Order
Order First Y Second Y Third Y Fourth Participants State Anxiety Depression Score
1 Baseline Y sham Y 20 mm Hg Y 40 mm Hg 8 38.8 (5.4) 35.9 (4.9)
2 Baseline Y sham Y 40 mm Hg Y 20 mm Hg 7 39.4 (13.8) 39.7 (9.4)
3 Baseline Y 20 mm Hg Y sham Y 40 mm Hg 7 35.1 (5.8) 35.9 (4.6)
4 Baseline Y 20 mm Hg Y 40 mm Hg Y sham 8 39.1 (8.7) 40.0 (6.3)
5 Baseline Y 40 mm Hg Y sham Y 20 mm Hg 8 36.1 (9.3) 38.4 (7.2)
6 Baseline Y 40 mm Hg Y 20 mm Hg Y sham 7 34.7 (8.8) 39.9 (5.6)
The schematic stimulation order and psychological status of each group are shown. The groups contained seven or eight persons including two women, respectively.The state anxiety scores and depression scores are from the Spielberger State-Trait Anxiety Inventory and Zung Self-rating Depression Scale, respectively. Scores aremean (standard deviation).
T. HAMAGUCHI et al.
454 Psychosomatic Medicine 75:453Y462 (2013)
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

tomography (PET) scanning. The PET scanning room was darkened, and theparticipants, while awake, were instructed to keep their eyes closed for thewhole period of the scan (70 seconds). Regional cerebral blood flow (rCBF) wasmeasured in each participant during four scans (70 seconds each) using a PETscanner in a three-dimensional sampling mode (HEADTOME V, SET-2400W;Shimadzu, Kyoto, Japan). The scanner produced 63 horizontal slices with aseparation of 3.125 mm, an axial field of view of 200 mm, an in-plane resolution
of 590 mm, and an axial resolution of 3.9 mm at full width at half maximum(35). There was a 10-minute interval between successive scans.
Data AnalysisThe data were processed using Statistical Parametric Mapping Software
(SPM2; Wellcome Department of Cognitive Neurology, London, UK) runningon a Matlab 6.5 operating platform (36,37). The PET images were changed
Figure 1. Statistical analysis design. A, Intense distention with preceding mild distention versus without preceding distention. B, Mild distention with precedingintense distention versus without preceding distention. C, Sham distention with preceding intense distention versus without preceding distention. D, Sham distention withpreceding mild distention versus without preceding distention. Participants were combined from stimulus order (see Table 1) for this comparison. To view image in color,please visit: www.psychosomaticmedicine.org.
Figure 2. Statistical (Z) maps of rCBF during 40-mmHg distention for [with preceding 20-mmHg distention40-mm Hg j baseline]j [without preceding distention40-mm
Hg j baseline]. Specific regions of the right orbital gyrus were detected (x, y, z = 20, 40,j22; Z = 4.83). The maps were overlaid on a sagittal (left) and verticofrontal (right)view of a single-participant MRI anatomical image using SPM2 and show the locations of areas with significantly higher rCBF. Coordinates are relative to the anteriorcommissure in the interaural (x), anterior-posterior (y), and superior-inferior (z) directions (puncorrected G .0001 for multiple comparisons, voxel threshold with an > levelZ = 3.48). The color calibration bar of each image represents the critical Z value magnitude of the activation areas. R indicates the right side of the brain image. To viewimage in color, please visit: www.psychosomaticmedicine.org.
BRAIN ACTIVATION DURING COLONIC DISTENTION
Psychosomatic Medicine 75:453Y462 (2013) 455
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

from DICOM to ANALYZE formatting for analysis, and the data were pre-processed using SPM2. The images were then normalized to the MontrealNeurological Institute space and smoothed spatially using a Gaussian filter of12 � 12 � 12 mm. The rCBF values were expressed in milliliters per deciliterper minute and were adjusted according to the individual global CBF valueusing an analysis of covariance and scaled to a mean of 50. The contribution ofeach parameter of interest to the changes in rCBF was estimated using SPM2,according to a general linear model at the voxel level.
We used a repeated (distention and baseline) factorial (with and withoutpreceding distention) contrasting analysis of variance analysis for comparison.To estimate the neural sensitization to visceral stimulation by the precedenceeffect, the statistical subtraction images were generated (Fig. 1). We also re-analyzed the data for subcortical areas such as the midbrain (including thePAG) using a volume of interest (VOI) analysis in SPM2 to detect smallerdefined structures.
The level of significance was set at a voxel level of p G .0001, uncorrectedfor multiple comparisons, with an extent threshold of 50 voxels as the regionof significant correction. The results were reported in Talairach coordinates(38) and activated Brodmann areas (BAs). The Talairach Daemon database(39) was used to complement the transformation of the coordinates in Talairachspace and to determine the precise cortical-activated regions. We used thealgorithm-estimated Talairach coordinates that corresponded with the Mon-treal Neurological Institute brain (40). Subjective visceral perception andemotion were compared among the groups (with prior stimuli and withoutprior stimuli) and conditions (baseline and stimulation) using the generalizedestimating equations (GEEs) (41) in SPSS 19.0 (IBM, Tokyo, Japan). After theLevene test for equality of variances, the independent-samples t test was usedfor post hoc test. We set the significance level at p G .05.
RESULTSChanges in rCBF During Equivalent Colonic DistentionWith or Without Preceding StimuliA significantly higher rCBF was observed in the right or-
bital gyrus (BA11, Z = 4.83) during a 40-mm Hg distentionoccurring after a 20-mm Hg distention (Orders 1 and 4) thanduring a 40-mm Hg distention occurring without prior expe-rience of visceral stimuli (Orders 5 and 6, puncorrected G .0001;Fig. 2 and Table 2). No voxels with suprathreshold results wereobserved when the 40-mm Hg distention with prior 20-mm Hg
distention results were subtracted from the 40-mm Hg disten-tion without preceding distention results.
The main effect of colonic distention at 20 mm Hg after theexperience of 40-mm Hg distention (Orders 2 and 6) was asignificantly higher rCBF in the midbrain, including the PAGregion (Z = 4.68), the left insula (BA13, Z = 4.39), and thecerebellum (Z = 3.63), compared with colonic distention at20 mm Hg without the prior experience of visceral stimuli(Orders 3 and 4; puncorrected G .0001; Fig. 3AYC and Table 2).No voxels with suprathreshold results were observed whenthe 20-mm Hg distension with prior 40-mm Hg results wassubtracted from the 20-mm Hg distension without precedingdistention results.
Sham distention (0 mm Hg) occurring after a 40-mm Hgdistension (Orders 4 and 5) produced a higher rCBF in the rightcingulate gyrus (BA24, Z = 4.11) than a sham distentionwithout the prior experience of visceral stimuli (Orders 1 and2, puncorrected G .0001; Fig. 4 and Table 2). No voxels withsuprathreshold results were observed when the sham disten-sion with prior 20-mm Hg distension results was subtractedfrom the sham distension without preceding distention results.No areas with a significantly higher rCBF were observed whensham distention with a preceding 20-mm Hg distension wascompared with sham distension without a preceding 20-mm Hgdistension.
Figure 5 shows the PAG region, which was further exam-ined in the VOI analysis to investigate sensitization. A sig-nificant difference in rCBF in the PAG was found when the20-mm Hg distention with prior 40-mm Hg distention resultswas compared with the 20-mm Hg without prior stimuli results(two-way analysis of variance, F(3,116) = 19.342, p G .0001).Post hoc comparisons using the Bonferroni test indicated thatthe mean PET signal obtained during the 20-mm Hg distentionwith prior 40-mm Hg distention (1.57 [0.24]) was significantlyhigher than that obtained during the 20-mm Hg distentionwithout prior stimuli (j0.27 [0.30], p G .001) and the baseline
TABLE 2. Neural Activation Resulting From Experienced Visceral Stimulation
Main Effect of Condition Region Side BACoordinates Z
valuex y z
(20 Y 40 j baseline) 9 (40 j baseline) Orbital gyrus R 11 20 40 j22 4.83
(40 mm Hg of order 1 and 4 9 order 5 and 6)
(40 Y 20 j baseline) 9 (20 j baseline) Cerebellum, uvula L j30 j82 j26 4.68
(20 mm Hg of order 2 and 6 9 order 3 and 4) Midbrain (including PAG) 2 j30 j18 4.63
Insula L 13 j38 j4 j10 4.39
(40 Y sham j baseline) 9 (sham j baseline) Cingulate gyrus R 24 8 10 30 4.11
(Sham of order 4 and 5 9 order 1 and 2)
(20 Y sham j baseline) 9 (sham j baseline) No suprathreshold of voxels
(Sham of order 3 and 6 9 order 1 and 2)
Coordinates refer to the location in stereotaxic space. The table shows the maxima of the SPM2 defined at each value. We used a repeated (distention and baseline)factorial (with and without preceding distention) contrasting two-way repeated-measures analysis of variance analysis for comparison. Height threshold: Z = 3.84,uncorrected p G .0001, extent threshold k = 50 voxels. BA = Brodmann area; PAG = periaqueductal gray. Bold characters indicate that they were compared with anequivalent stimulus (minus the baseline, respectively).
T. HAMAGUCHI et al.
456 Psychosomatic Medicine 75:453Y462 (2013)
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

signal (j0.63 [0.19], p G .001). No significant change betweenthe baseline results (j0.50 [0.20]) and the 20-mm Hg disten-tion without prior stimuli (j0.27 [0.30]) was seen (p = .48).Data represent means (standard error).
Visceral Perception and Emotional Changes DuringEquivalent Stimuli With or Without Preceding StimuliGEE of visceral perception and emotion showed a significant
condition (40-mm Hg distention with preceding 20-mm Hgdistention versus 40-mm Hg distention without preceding dis-tention) � stimulus (baseline versus 40 mm Hg) interactioneffect (p G .01 for abdominal discomfort and p = .03 for ab-dominal distention; Fig. 6A). On the other hand, there is no
significant condition � stimulus interaction of visceral per-ception and emotion between after a 20-mm Hg distension witha preceding 40-mm Hg distension and a 20-mm Hg distensionwithout a prior distention by GEE (Fig. 6B). GEE changes invisceral perception and emotion revealed a significant condi-tion � stimulus interaction in abdominal pain (p G .01),sleepiness (p G .01), and anxiety (p G .01) comparing a shamdistention with a preceding 40-mm Hg distention and without apreceding distention (Fig. 6C). Comparing a sham distentionwith a preceding 20-mm Hg distension and that without dis-tention, GEE of visceral perception and emotion showed asignificant condition � stimulus interaction in abdominal dis-comfort (p G .01), abdominal pain (p G .01), and urgency for
Figure 3. Statistical (Z) maps of rCBF during 20-mmHg distention for [with preceding 40-mmHg distention20-mm Hg j baseline]j [without preceding distention20-mm
Hg j baseline]. Note the significantly higher activation in the midbrain, including the PAG region (x, y, z = 2, j30, j18; Z = 4.63) (A), the left insula (BA13; x, y, z =j38, j4, j10; Z = 4.39) (B), and the left cerebellum (x, y, z = j30, j82, j26; Z = 4.68) (C). The maps were overlaid on a sagittal (left) and verticofrontal (right)view of an MRI anatomical image using SPM2. See the legend of Figure 2 for graphical details. To view image in color, please visit: www.psychosomaticmedicine.org.
BRAIN ACTIVATION DURING COLONIC DISTENTION
Psychosomatic Medicine 75:453Y462 (2013) 457
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

defecation (p G .01; Fig. 6D). No significant differences invisceral perception and emotion were found between precedingstimulation and no stimulation by post hoc test.
DISCUSSIONThe data collected by the direct comparison of different
patterns of equivalent visceral stimuli show that visceral sen-sitization is associated with neural activity in a specific area ofthe midbrain, insular cortex, cerebellum, cingulate cortex, andOFC. The brain behaves differently in response to different
patterns of visceral stimulation. We also found a different vis-ceral perception and emotion between preceding stimuli andwithout preceding stimuli. The determinants of visceral hy-persensitivity may include the combined effects of intrinsicfactors and the program of the stimuli.
The right OFC was activated by a 40-mm Hg distentionoccurring after a 20-mm Hg stimulation. The OFC is consid-ered to play important roles in goal-directed behavior byadjusting the motivational level toward a certain (current orfuture) outcome with a particular motivational significancebased on reward and punishment expectancy processes (42,43).
Figure 4. Statistical (Z) maps of rCBF during sham distention for [with preceding 40-mm Hg distentionsham j baseline]j [without preceding distentionsham j baseline].The right cingulate gyrus (BA24; x, y, z = 8, 10, 30; Z = 4.11) was found to have higher levels of activation under the conditions of sham distention after the experienceof 40-mm Hg distention than for sham distention without prior stimuli. See the legend of Figure 2 for graphical details. To view image in color, please visit:www.psychosomaticmedicine.org.
Figure 5. Changes in rCBF in the PAG according to the predefinedVOI approach. TheVOI analysiswas performed in the PAG using the coordinates x, y, z = 2,j30,j18.The horizontal section of the brain image indicates the PAG region. The vertical axis indicates the adjusted rCBF. See Table 2 and Figure 3A. Data represent mean(standard error). To view image in color, please visit: www.psychosomaticmedicine.org.
T. HAMAGUCHI et al.
458 Psychosomatic Medicine 75:453Y462 (2013)
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

The activation of the OFC is correlated with sensory-specificsatiety for a liquid food in humans (44). A study using func-tional magnetic resonance imaging (fMRI) indicated thatpleasant and painful stimuli produce greater activation of theOFC, relative to the somatosensory cortex, than effectivelyneutral stimuli (45). Patients with OFC damage do not showan impairment in spatial working memory but have impulsiveand inappropriate behavior (46). Therefore, it may be that ac-tivation of the OFC in this study is involved in processes thatoccur in advance of visceral pain if warning stimuli precedencethat pain is imminent. Affective and motivational responses tovisceral stimulation are examples of such processes.
The midbrain, including the PAG, insula, and cerebellum,wasmore strongly activated by a 20-mmHg distension occurring
after a 40-mm Hg distention than by a 20-mm Hg distentionoccurring without the experience of actual distention. As in-dicated by a VOI analysis of the PAG, this study demonstratedthe involvement of PAG activation in the desensitization tovisceral stimuli. Abdominal pain during a 20-mm Hg disten-tion occurring after a 40-mm Hg distention tended to be lowerthan that during a 20-mm Hg distention without precedingstimuli, but a significant difference was not observed. Thisfinding is very similar to that for the offset analgesia that isobserved when thermal stimuli are applied to the skin (47).Offset analgesia is typically demonstrated by an immediate changein stimulus intensity (47), not by a change after 10 minutes,as performed in our study. Therefore, that methodological dif-ference should be recognized.
Figure 6. Comparisons of subjective visceral perception and emotion in response to an equivalent intensity of colonic distention with or without a preceding distension(baseline versus stimulation). GEE of changes in the ordinate scale revealed a significant condition� stimulus interaction (*p G .05, **p G .01). A, 40-mm Hg distensionwith preceding 20-mm Hg distension (n = 16) versus 40-mm Hg distention without prior stimulation (n = 15). B, 20-mm Hg distension with a preceding 40-mm Hgdistension (n = 14) versus 20-mm Hg distension without prior stimulation (n = 15). C, Sham distension with a preceding 40-mm Hg distension (n = 16) versus shamdistension without a prior stimulation (n = 15). D, Sham distension with a preceding 20-mmHg distension (n = 14) versus sham distension without a prior stimulation(n = 15). The vertical axis indicates the visceral perception and emotion. Data represent mean (standard error). B = baseline; S = sham distention; 20 = 20 mm Hgdistention; 40 = 40 mm Hg distention.
BRAIN ACTIVATION DURING COLONIC DISTENTION
Psychosomatic Medicine 75:453Y462 (2013) 459
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

The PAG is an important component of the descendingnoxious inhibitory system (48), which contains a high con-centration of opiate neurons with descending spinal afferents(49). One mechanism by which the PAG modulates visceralperception may involve the release of endogenous opioids. ThePAG contains significant quantities of all families of endoge-nous opioid peptides, and K-opioids are known to act by re-leasing PAG projection cells from GABAergic inhibition (50).Specific projections of the medial network to the PAG, hypo-thalamus, and amygdala presumably mediate integrated auto-nomic and antinociceptive responses to acute aversive stimuli(48,51). Activation in the PAG is significantly increased duringthe attentional modulation of pain intensity (49), and changesin pain responses attributable to changes in arousal or attentionresult from the action of modulatory networks that control thetransmission of nociceptive signals of the brain (52). Therefore,the results of this study suggest that mild (20-mm Hg) distentionafter the experience of intense (40-mm Hg) distention in thedescending colon might be sufficient to reduce visceral per-ception by activating the descending pain inhibitory systemin normal participants.
The central roles of the insula and somatosensory corticesin interoceptive attention endorse their proposed contributionsto subjective feeling states arising from representations ofbodily responses (53,54). The hypothesis that the insula me-diates somatic and visceral attention is further strengthenedby correlations between local gray matter volume and ques-tionnaire ratings of visceral awareness (53). Nevertheless, ourdata did not implicate the involvement of the insular cortex insupporting awareness of visceral responses that are enhancedin negative mood states.
The cerebellum was more activated by mild stimuli beforethe participant experienced intense stimuli than without stimuli.The cerebellothalamic projections from all the cerebellar nu-clei, including the fastigial nucleus, are targeted primarilyto the ventral lateral nucleus both contralaterally and ipsilat-erally (55,56). Interpolaris neurons have direct axonal pro-jections to the thalamus, cerebellum, and spinal cord (57).Bilateral lesions of the deep cerebellar nuclei prevent the learn-ing of an aversive task (51). Although this issue has rarely beendiscussed previously, the cerebellum may play a role in painprocessing.
In this study, a sham stimulation after the experience of a40-mm Hg visceral stimuli activated the right cingulate cortex(BA24), compared with a sham stimulation without the experi-ence of actual visceral stimulation. The cingulate cortex islocated on the medial surface of the human brain. By identi-fying the supplementary motor area (SMA) and the pre-SMA,three separate areas within the cingulate cortex can be identi-fied: a caudal cingulate cortex situated ventral to the SMA anda rostral cingulate cortex that can be further subdivided intoanterior and posterior subregions. Of note, fMRI BOLD acti-vations in specific areas of the cingulate cortex were associatedwith urges to perform certain behaviors (i.e., swallowing,micturition, and yawning) (58). The cingulate cortex is asso-ciated with the execution of simple movements and is also
activated in response to bodily stimulation, such as the deliveryof painful cutaneous heat and cold stimuli. Of note, althoughthe SMA itself has been linked to the conscious intention tomove (59) and the cingulate cortex and SMAmay be coactivatedduring movement execution, the SMA and cingulate cortexshould be considered functionally distinct from one another.
The insula and ACC have been considered as the input andoutput regions of a functional system that is engaged in cog-nitive, affective, and behavioral contexts (60). Consistent withthis proposal, there is now considerable evidence that theseregions are jointly active across a wide range of experimentalconditions. The anterior insula is connected functionally withthe anterior and midcingulate cortex, whereas the midregionand posterior region of the insula are connected only with theposterior region of the midcingulate cortex based on functionalconnectivity analyses performed using fMRI (61).
Although the results of this study are compatible with thekey roles of the OFC, cingulate cortex, and PAG in corticalmechanisms of visceral perceptional modulation, this inter-pretation was based on a PET image analysis. Thus, we cannotrule out the possibility that our experimental conditions dif-ferentially influenced this brain region through ascending mech-anisms. An interventional approach is needed to further testthe associations between OFC, ACC, and PAG activities andvisceral perception suppression. Future studies could examinethe effects of interactions between different visceral intensitiesand different orders of visceral stimuli or could use transcranialmagnetic stimulation to reversibly and selectively interfere withcortical function during visceral stimulation.
In conclusion, the results of this study support our twohypotheses, that is, a) that brain activation arising from co-lonic distention with a preceding intense stimulation woulddiffer from that arising from intense stimulation after mildstimulation and b) that brain activation in response to shamcolonic distention occurring before an intense stimulationwould differ from that occurring after an intense stimulation.These results suggest that preceding visceral stimuli may affectneural sensitization and/or desensitization in humans.
Source of Funding and Conflicts of Interest: This study wassupported by a Grant-in-Aid for Scientific Research from the Min-istry of Education, Culture, Sports, Science and Technology ofJapan and a Grant-in-Aid for Scientific Research from the Ministryof Health, Labour and Welfare of Japan. Shin Fukudo is the pri-mary investigator of these grants. The authors report no conflictsof interest.
REFERENCES1. Drossman DA, Sandler RS, McKee DC, Lovitz AJ. Bowel patterns among
subjects not seeking health care. Use of a questionnaire to identify apopulation with bowel dysfunction. Gastroenterology 1982;83:529Y34.
2. Drossman DA. Clinical research in the functional digestive disorders.Gastroenterology 1987;92:1267Y9.
3. DrossmanDA,McKeeDC, Sandler RS,Mitchell CM, Cramer EM, LowmanBC, Burger AL. Psychosocial factors in the irritable bowel syndrome. A
T. HAMAGUCHI et al.
460 Psychosomatic Medicine 75:453Y462 (2013)
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

multivariate study of patients and nonpatients with irritable bowel syndrome.Gastroenterology 1988;95:701Y8.
4. Fukudo S, Nomura T, Muranaka M, Taguchi F. Brain-gut response to stressand cholinergic stimulation in irritable bowel syndrome. A preliminarystudy. J Clin Gastroenterol 1993;17:133Y41.
5. Accarino AM, Azpiroz F, Malagelada JR. Attention and distraction: effectson gut perception. Gastroenterology 1997;113:415Y22.
6. Silverman DH, Munakata JA, Ennes H, Mandelkern MA, Hoh CK,Mayer EA. Regional cerebral activity in normal and pathological per-ception of visceral pain. Gastroenterology 1997;112:64Y72.
7. Mertz H. Role of the brain and sensory pathways in gastrointestinal sen-sory disorders in humans. Gut 2002;51(Suppl 1):i29Y33.
8. Fukudo S, Kanazawa M, Kano M, Sagami Y, Endo Y, Utsumi A, Nomura T,Hongo M. Exaggerated motility of the descending colon with repetitivedistention of the sigmoid colon in patients with irritable bowel syndrome.J Gastroenterol 2002;37(Suppl 14):145Y50.
9. Gebhart GF. Pathobiology of visceral pain: molecular mechanisms andtherapeutic implications IV. Visceral afferent contributions to the patho-biology of visceral pain [review]. Am J Physiol Gastrointest Liver Physiol2000;278:G834Y8.
10. Saito-Nakaya K, Hasegawa R, Nagura Y, Ito H, Fukudo S. Corticotropin-releasing hormone receptor 1 antagonist blocks colonic hypersensitivityinduced by a combination of inflammation and repetitive colorectal dis-tension. Neurogastroenterol Motil 2008;20:1147Y56.
11. Mayer EA, Raybould HE. Role of visceral afferent mechanisms in func-tional bowel disorders. Gastroenterology 1990;99:1688Y704.
12. Munakata J, Naliboff B, Harraf F, Kodner A, Lembo T, Chang L,Silverman DH, Mayer EA. Repetitive sigmoid stimulation induces rectalhyperalgesia in patients with irritable bowel syndrome. Gastroenterology1997;112:55Y63.
13. Hobson AR, Khan RW, Sarkar S, Furlong PL, Aziz Q. Development ofesophageal hypersensitivity following experimental duodenal acidifica-tion. Am J Gastroenterol 2004;99:813Y20.
14. Campbell JN, Meyer RA. Sensitization of unmyelinated nociceptive af-ferents in monkey varies with skin type. J Neurophysiol 1983;49:98Y110.
15. Davis KD, Meyer RA, Campbell JN. Chemosensitivity and sensitization ofnociceptive afferents that innervate the hairy skin of monkey. J Neurophysiol1993;69:1071Y81.
16. Koda H, Mizumura K. Sensitization to mechanical stimulation by in-flammatory mediators and by mild burn in canine visceral nociceptors invitro. J Neurophysiol 2002;87:2043Y51.
17. Kumazawa T, Mizumura K, Minagawa M, Tsujii Y. Sensitizing effects ofbradykinin on the heat responses of the visceral nociceptor. J Neurophysiol1991;66:1819Y24.
18. Treede RD, Meyer RA, Raja SN, Campbell JN. Peripheral and centralmechanisms of cutaneous hyperalgesia. Prog Neurobiol 1992;38:397Y421.
19. Jaggar SI, Habib S, Rice AS. The modulatory effects of bradykinin B1and B2 receptor antagonists upon viscero-visceral hyper-reflexia in a ratmodel of visceral hyperalgesia. Pain 1998;75:169Y76.
20. Coutinho SV, Meller ST, Gebhart GF. Intracolonic zymosan producesvisceral hyperalgesia in the rat that is mediated by spinal NMDA and non-NMDA receptors. Brain Res 1996;736:7Y15.
21. Coutinho SV, Urban MO, Gebhart GF. Role of glutamate receptors andnitric oxide in the rostral ventromedial medulla in visceral hyperalgesia.Pain 1998;78:59Y69.
22. Yelle MD, Rogers JM, Coghill RC. Offset analgesia: a temporal contrastmechanism for nociceptive information. Pain 2008;134:174Y86.
23. Martucci KT, Yelle MD, Coghill RC. Differential effects of experimentalcentral sensitization on the time-course and magnitude of offset analgesia.Pain 2012;153:463Y72.
24. Hobday DI, Aziz Q, Thacker N, Hollander I, Jackson A, Thompson DG. Astudy of the cortical processing of ano-rectal sensation using functionalMRI. Brain 2001;124:361Y8.
25. Smout A, Akkermans L. Rectum, anus, and pelvic floor. In: Smout A,Akkermans L, editors. Normal and Disturbed Motility of the Gastroin-testinal Tract Petersfield. Petersfield: Biomedical Publishing Ltd; 1992:169Y210.
26. Fukudo S, Nomura T, Hongo M. Impact of corticotropin-releasing hor-mone on gastrointestinal motility and adrenocorticotropic hormone innormal controls and patients with irritable bowel syndrome. Gut 1998;42:845Y9.
27. Naliboff BD, Derbyshire SW, Munakata J, Berman S, Mandelkern M,Chang L, Mayer EA. Cerebral activation in patients with irritable bowel
syndrome and control subjects during rectosigmoid stimulation. PsychosomMed 2001;63:365Y75.
28. Wager TD, Rilling JK, Smith EE, Sokolik A, Casey KL, Davidson RJ,Kosslyn SM, Rose RM, Cohen JD. Placebo-induced changes in FMRI inthe anticipation and experience of pain. Science 2004;303:1162Y7.
29. Hamaguchi T, Kano M, Rikimaru H, Kanazawa M, Itoh M, Yanai K,Fukudo S. Brain activity during distention of the descending colon inhumans. Neurogastroenterol Motil 2004;16:299Y309.
30. Naliboff BD, Berman S, Chang L, Derbyshire SW, Suyenobu B, Vogt BA,Mandelkern M, Mayer EA. Sex-related differences in IBS patients: centralprocessing of visceral stimuli. Gastroenterology 2003;124:1738Y47.
31. Fukudo S, Kanazawa M, Mizuno T, Hamaguchi T, Kano M, Watanabe S,Sagami Y, Shoji T, Endo Y, Hongo M, Itoyama Y, Yanai K, Tashiro M,Aoki M. Impact of serotonin transporter gene polymorphism on brainactivation by colorectal distention. Neuroimage 2009;47:946Y51.
32. Kano M, Hamaguchi T, Itoh M, Yanai K, Fukudo S. Correlation betweenalexithymia and hypersensitivity to visceral stimulation in human. Pain2007;132:252Y63.
33. Spielberger CD, Gorsuch RL, Lushene RE, Vagg PR, Jacobs GA. Manualfor the State-Trait Anxiety Inventory STAI (From Y). Palo Alto, CA:Consulting Psychologist Press; 1983.
34. Zung WW. A Self-Rating Depression Scale. Arch Gen Psychiatry1965;12:63Y70.
35. Fujiwara T, Watanuki S, Yamamoto S, Miyake M, Seo S, Itoh M, Ishii K,Orihara H, Fukuda H, Satoh T, Kitamura K, Tanaka K, Takahashi S.Performance evaluation of a large axial field-of-view PET scanner: SET-2400W. Ann Nucl Med 1997;11:307Y13.
36. Friston KJ, Ashbuner J, Poline JB, Frith CD, Frackowiak RSJ. Spatialregistration and normalization of images. Hum Brain Mapp 1995;2:165Y89.
37. Friston KJ, Holmes AP, Worsley KJ, Poline JP, Frith CD, Frackowiak RSJ.Statistical parametric maps in functional imaging: a general linear approach.Hum Brain Mapp 1995;2:189Y210.
38. Talairach J, Tournoux P. Co-Planar Stereotaxic Atlas of the Human Brain.New York: Thieme Medical Publishers; 1988.
39. Lancaster JL, Woldorff MG, Parsons LM, Liotti M, Freitas CS, Rainey L,Kochunov PV, Nickerson D, Mikiten SA, Fox PT. Automated Talairachatlas labels for functional brain mapping. Hum Brain Mapp 2000;10:120Y31.
40. Duncan J, Seitz RJ, Kolodny J, Bor D, Herzog H, Ahmed A, Newell FN,Emslie H. A neural basis for general intelligence. Science 2000;289:457Y60.
41. Zeger SL, Liang KY. Longitudinal data analysis for discrete and continu-ous outcomes. Biometrics 1986;42:121Y30.
42. O’Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Ab-stract reward and punishment representations in the human orbitofrontalcortex. Nat Neurosci 2001;4:95Y102.
43. Hikosaka K, Watanabe M. Long- and short-range reward expectancy in theprimate orbitofrontal cortex. Eur J Neurosci 2004;19:1046Y54.
44. Kringelbach ML, O’Doherty J, Rolls ET, Andrews C. Activation of thehuman orbitofrontal cortex to a liquid food stimulus is correlated with itssubjective pleasantness. Cereb Cortex 2003;13:1064Y71.
45. Francis S, Rolls ET, Bowtell R, McGlone F, O’Doherty J, Browning A,Clare S, Smith E. The representation of pleasant touch in the brain and itsrelationship with taste and olfactory areas. Neuroreport 1999;10:453Y9.
46. Berlin HA, Rolls ET, Kischka U. Impulsivity, time perception, emotion andreinforcement sensitivity in patients with orbitofrontal cortex lesions. Brain2004;127:1108Y26.
47. Derbyshire SW, Osborn J. Offset analgesia is mediated by activation in theregion of the periaqueductal grey and rostral ventromedial medulla.Neuroimage 2009;47:1002Y6.
48. Vogt BA, Sikes RW, Vogt LJ. Anterior cingulate cortex and the medial painsystem. In: Vogt BA, Gabriel M, editors. Neurobiology of Cingulate Cortexand Limbic Thalamus. Boston: Birkhauser; 1993:313Y44.
49. Tracey I, Ploghaus A, Gati JS, Clare S, Smith S, Menon RS, Matthews PM.Imaging attentional modulation of pain in the periaqueductal gray inhumans. J Neurosci 2002;22:2748Y52.
50. Fields HL, Basbaum AI. Central nervous system mechanism of painmodulation. In: Patrick DW, Ronald M, editors. Textbook of Pain. 4th ed.London: Churchill Livingstone; 1999:309Y29.
51. Steinmetz JE, Logue SF, Miller DP. Using signaled barpressing tasks to studythe neural substrates of appetitive and aversive learning in rats: behavioralmanipulations and cerebellar lesions. Behav Neurosci 1993;107:941Y54.
BRAIN ACTIVATION DURING COLONIC DISTENTION
Psychosomatic Medicine 75:453Y462 (2013) 461
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.

52. Petrovic P, Petersson KM, Ghatan PH, Stone-Elander S, Ingvar M. Pain-relatedcerebral activation is altered by a distracting cognitive task. Pain 2000;85:19Y30.
53. Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. Neural systemssupporting interoceptive awareness. Nat Neurosci 2004;7:189Y95.
54. Craig AD. How do you feel? Interoception: the sense of the physiologicalcondition of the body [review]. Nat Rev Neurosci 2002;3:655Y66.
55. Ilinsky IA, Kultas-Ilinsky K, Rosina A, Haddy M. Quantitative evalua-tion of crossed and uncrossed projections from basal ganglia and cere-bellum to the cat thalamus. Neuroscience 1987;21:207Y27.
56. Huh KH, Endo S, Olsen RW. Diazepam-insensitive GABAA receptors inrat cerebellum and thalamus. Eur J Pharmacol 1996;310:225Y33.
57. Hayashi H, Sumino R, Sessle BJ. Functional organization of trigem-inal subnucleus interpolaris: nociceptive and innocuous afferent inputs,
projections to thalamus, cerebellum, and spinal cord, and descendingmodulation from periaqueductal gray. J Neurophysiol 1984;51:890Y905.
58. Picard N, Strick PL. Imaging the premotor areas [review]. Curr OpinNeurobiol 2001;11:663Y72.
59. Desmurget M, Reilly KT, Richard N, Szathmari A, Mottolese C, Sirigu A.Movement intention after parietal cortex stimulation in humans. Science2009;324:811Y3.
60. Medford N, Critchley HD. Conjoint activity of anterior insular and anteriorcingulate cortex: awareness and response [review]. Brain Struct Funct2010;214:535Y49.
61. Taylor KS, Seminowicz DA, Davis KD. Two systems of resting stateconnectivity between the insula and cingulate cortex. Hum Brain Mapp2009;30:2731Y45.
T. HAMAGUCHI et al.
462 Psychosomatic Medicine 75:453Y462 (2013)
SPECIAL SERIES ON NEUROSCIENCEIN HEALTH AND DISEASE
Copyright © 2013 by the American Psychosomatic Society. Unauthorized reproduction of this article is prohibited.





























