Embed Size (px)
Citation preview

SUMMARY
The responses of relative growth rate (% day–1) andpigment content (chlorophyll a, phycocyanin and phyco-erythrin) to temperature, irradiance and photoperiodwere analyzed in culture in seven freshwater red algae:Audouinella hermannii (Roth) Duby, Audouinellapygmaea (Kützing) Weber-van Bosse, Batrachospermumambiguum Montagne, Batrachospermum delicatulum(Skuja) Necchi et Entwisle, ‘Chantransia’ stages of B. delicatulum and Batrachospermum macrosporumMontagne and Compsopogon coeruleus (C. Agardh)Montagne. Experimental conditions included tempera-tures of 10, 15, 20 and 25°C and low and high irradi-ances (65 and 300 µmol photons m–2 s–1, respectively).Long and short day lengths (16:8 and 8:16 LD cycles)were also applied at the two irradiances. Growth effectsof temperature and irradiance were evident in mostalgae tested, and there were significant interactionsamong treatments. Most freshwater red algae had thebest growth under low irradiance, confirming the pref-erence of freshwater red algae for low light regimens. Ingeneral there was highest growth rate in long days andlow irradiance. Growth optima in relation to tempera-ture were species-specific and also varied between lowand high irradiances for the same alga. The most significant differences in pigment content were relatedto temperature, whereas few significant differencescould be attributed to variation in irradiance and photo-period or interactions among the three parameters. Theresponses were species-specific and also differed forpigments in distinct temperatures, irradiances andphotoperiods in the same alga. Phycocyanin was gener-ally more concentrated than phycoerythrin and phyco-biliproteins were more concentrated than chlorophyll a.The highest total pigment contents were found in twospecies typical of shaded habitats: A. hermannii andC. coeruleus. The expected inverse relationship of pig-ment with irradiance was observed only in C. coeruleus.In general, the most favorable conditions for growthwere not coincident with those with highest pigmentcontents.
Key words: chlorophyll, ecophysiology, freshwater redalgae, growth, irradiance, photoperiod, phycocyanin,phycoerythrin, temperature.
INTRODUCTION
Light regimen, including fluctuations in intensity,quality and photoperiod, is one of the major factorsaffecting the distribution and seasonality of freshwaterred algae (Sheath 1984). Temperature also influenceslatitude, altitude and drainage basin distribution pat-terns, as well as seasonality, of freshwater red algae(Sheath and Hambrook 1990). Despite the importanceof these environmental factors, few experimentalstudies have been conducted to evaluate their eco-physiological role in freshwater red algae.
The effects of light and temperature on pigmentcontent have been little studied in freshwater red algae.Rider and Wagner (1972) observed that the chlorophyllcontent of Batrachospermum turfosum [as B. vagum(Roth) C. Agardh] decreased two- to threefold whenplants were exposed for 1 month to green or far redlight, in comparison to white, blue or red wavelengths.They also found a greater chlorophyll production in low-intensity ranges (< 55 µmol photons m–2 s–1). Thirb andBenson-Evans (1983) reported alterations in pigmentcontent in Lemanea sp. in response to changes in bothlight quantity and quality. The highest concentrations of chlorophyll and phycoerythrin were obtained at lowirradiance (ca 25 µmol m–2 s–1) and in red light,whereas minimum values were found at high irradiance(ca 94 µmol m–2 s–1) and in yellow light. The effect oftemperature and suspended solids on the pigment concentration in Lemanea sp. was tested by Thirb andBenson-Evans (1984, 1985). They reported a trend ofdecreasing chlorophyll a and phycoerythrin content with
Phycological Research 2001; 49: 103–114
Effects of temperature, irradiance and photoperiod ongrowth and pigment content in some freshwater red algae incultureMarcelo Ribeiro Zucchi and Orlando Necchi Jr*São Paulo State University, Zoology and Botany Department, Rua Cristóvão Colombo, 2265 – 15054-000, São José doRio Preto, SP, Brazil
*To whom correspondence should be addressed.Email: [email protected] editor: D. J. Garbary.Received 11 July 2000; accepted 11 December 2000.
increased temperatures, and an opposite trend withincreased concentrations of suspended solids, exceptfor a reduction at the highest level tested. Kaczmarczykand Sheath (1991) showed that total irradiance con-trolled most pigment alterations in Batrachospermumboryanum Sirodot, and that ratios of phycocyanin tophycoerythrin tended to decrease as irradianceincreased in laboratory studies. Kaczmarczyk andSheath (1992) presented data on pigment content(total pigment, chlorophyll a, phycocyanin and phyco-erythrin) for 24 populations of seven freshwater redalgal species under natural conditions with varyingdegrees of shading. Despite these investigations, it isclear that more taxa and additional environments needto be studied (Kaczmarczyk and Sheath 1991). In addition, laboratory experiments are required to betterunderstand adaptations to light and temperature inthese algae.
Phycobiliprotein composition has been studied infew freshwater red algae (Gantt et al. 1986). Two spec-trally distinct c-phycocyanin forms were found in Comp-sopogon coeruleus (C. Agardh) Montagne by Gantt et al.(1986), along with B-phycoerythrin. Glazer et al.(1994) reported an unusual B-phycoerythrin inC. coeruleus, which completely lacks phycourobilinchromophores and in which the α-, β- and γ-subunitsall carry phycoerythrobilins. Glazer et al. (1997)described a new type of phycoerythrin in acrochaetioidfreshwater red algae, characterized by the presence ofone phycocyanobilin combined with one phycourobilinand one phycoerythrobilin on the β-subunit.
Growth studies of freshwater red algae are few.Karsten et al. (1993) reported the growth pattern inrelation to a range of salinities and irradiances inBostrychia moritziana (Kützing) J. Agardh, includingtwo freshwater isolates, which showed the best growthat low salinities (4–5‰) and were low-light adapted(12–60 µmol photons m–2 s–1). Glazer et al. (1994)measured growth rates of C. coeruleus in various salin-ities and found that the alga grows and reproduces insalinities ranging from 5 to 35‰. Leukart and Hanelt(1995) evaluated the thallus extension under five distinct irradiance levels in Batrachospermum helmin-thosum Bory. Growth was maximal at low irradiances(20–50 µmol photons m–2 s–1) and inhibited at100 µmol photons m–2 s–1.
Considering the scarcity of experimental investiga-tions on growth and pigment content in freshwater redalgae, this study was initiated to evaluate the responsesto temperature, irradiance and photoperiod in somefreshwater red algae under culture conditions.
MATERIALS AND METHODS
Seven freshwater red algae were used in this study(Table 1). These taxa encompass a range of morpho-logical types (Sheath and Hambrook 1988): tufts(Audouinella and ‘Chantransia’ stages), non-mucilaginousfilaments (Compsopogon) and mucilaginous filaments(Batrachospermum). The three orders of Rhodophytawidespread in freshwater habitats are represented:Compsopogonales (Bangiophyceae), Acrochaetiales and
104 M. R. Zucchi and O. Necchi Jr
Table 1. Location, date of isolation and maintenance conditions in culture of the freshwater red algal isolates used in the experiments.
Photoperiod 12:12 LD
Location Isolation date Temperature Irradiance Species and isolate number(°C) (µmol photons
m–2 s–1)
Australia: Melbourne, Victoria, near 21.iii.1996 15 45 Audouinella hermannii (Roth) Duby, Warburton, Yarra River, 37°49′S, 21.vi.1996 isolate no. 38144°58′E (reinoculated)
USA: Amherst, Massachusetts 20.i.1966 20 125–160 Audouinella pygmaea (Kützing)University of Massachusetts, Botany 04.iii.1996 Weber-van Bosse, isolate no. 55Department, 42°23′N, 72°31′W (reinoculated)
Brazil: São Paulo State, Itirapina, Route 30.v.1995 20 140 ‘Chantransia’ stage of BatrachospermumSP-310, km 213–214, Córrego do delicatulum (Skuja) Necchi et Entwisle, Feijão, 22°09′S, 47°47′W isolate no. 1
Brazil: São Paulo State, São José 16.viii.1995 20 140 ‘Chantransia’ stage of Batrachospermum do Rio Preto, Córrego da Lagoa, macrosporum Montagne, isolate no. 1320°49′S, 49°20′W
Brazil: Santa Catarina, São Pedro 19.iii.1996 20 170 Batrachospermum ambiguum Montagne,de Alcântara, Salto, 27°34′S, isolate no. 3548°50′W
Brazil: São Paulo State, São José 19.ix.1995 20 165–190 Batrachospermum delicatulum (Skuja)do Rio Preto, Córrego da Lagoa, Necchi et Entwisle, isolate no. 2320°49′S, 49°20′W
Brazil: São Paulo State, Pontes 01.viii.1997 20 85 Compsopogon coeruleus (C. Agardh)Gestal, Cachoeira de São Montagne, isolate no. 58Roberto, Rio Preto, 20°11′S,49°41′W

Batrachospermales (Florideophyceae). ‘Chantransia’stages had two distinct origins (Necchi and Zucchi1997): (i) a population isolated from monosporic fieldplants, in culture reproducing by monosporangia andalso producing juvenile Batrachospermum gameto-phytes attached to apices of filaments (isolate no. 1);and (ii) a population isolated from carpospore germina-tion, in culture reproducing by monosporangia and alsoproducing juvenile Batrachospermum gametophytes(isolate no. 13). Isolation into culture followed proce-dures described previously (Necchi and Zucchi 1997).
Experiments were initiated from cultures in expo-nential growth. Details on each isolate and culturemaintenance are summarized in Table 1. All experi-ments and maintenance of isolates were carried out in20:1 water/soil culture medium (Necchi and Zucchi1997) with initial pH and specific conductance of6.9–7.0 and 915–1050 µS cm–1, respectively. Incuba-tions were conducted in RI 12–555 Revco (Asheville,NC, USA) incubators with illumination from above supplied by two to four cool-white fluorescent lamps(15 W; Phillips, São Paulo, Brazil). Irradiances withinthe incubators were adjusted by moving the shelves orusing a neutral black filter. Measurements were made(in distilled water) with a Li-Cor (Lincoln, NB, USA)LI-189 quantameter with a spherical quantum sensorLI-193 SA. Temperature and irradiance experimentsfollowed a 4 × 2 factorial design, with temperatures of10, 15, 20 and 25°C (± 0.3°C) at low (65 ± 2 µmolphotons m–2 s–1) and high (300 ± 5 µmol photonsm–2 s–1) irradiances. In photoperiod and irradianceexperiments a 2 × 2 factorial design was applied: longday (16:8 LD cycle) and short day (8:16) and the twoirradiances specified above, all at 20°C.
The duration of growth experiments was 30 days forBatrachospermum and Compsopogon and 45 days forAudouinella and ‘Chantransia’ stages. These periodswere determined from preliminary growth curves, estab-lished by measurements made at 5-day intervals over60 days at 20°C and 160 µmol photons m–2 s–1. Growthwas based on initial and final fresh weights. Plantlength (thallus extension) was initially utilized (with aruler under a stereoscope) but was found to be unreli-able because of error in disregarding lateral branches.Plant segments were grown from initial fresh weights of1.0–3.5 mg, and the relative growth rate was calculatedby the formula: R (% day–1) = 100 (ln Wt – ln W0) t–1,where W0 = initial fresh weight, Wt = final fresh weight,t = time (days) (Kain 1987). Plants were blotted dryand then weighed (resolution 0.1 mg). Eight replicateswere used in experiments, each consisting of a testtube (16 × 150 mm) with 10 mL of culture medium.Position of tubes within each set was changed at 5-dayintervals to minimize irradiance differences among thereplicates.
At the end of the growth period, five replicates wereused for pigment determination and the remaining
three were used for morphological analyses (data notpresented). Plants were frozen for 7–15 days beforepigment extraction.
Phycobiliproteins (phycocyanin and phycoerythrin)were extracted in 0.1 mol L–1 phosphate buffer (pH 6.8)and quantified according to the spectrophotometrictechnique of Beer and Eshel (1985), with a B 342-IIMicronal (São Paulo, Brazil) spectrophotometer (1 nmresolution).
Chlorophyll a was extracted by resuspending thepellet resulting from phycobiliprotein extraction in 90% alkaline acetone, and determination followed the spectrophotometric technique (Wetzel and Likens1991). Pigment extraction and reading were conductedunder dim light to avoid degradation.
Numerical data were submitted to analysis of vari-ance (two-way ANOVA, Zar 1999) in order to test simul-taneously the effect of two variables (temperature +irradiance or photoperiod + irradiance) on growth andpigment content for each alga exposed to distinct treatments.
RESULTS
Growth
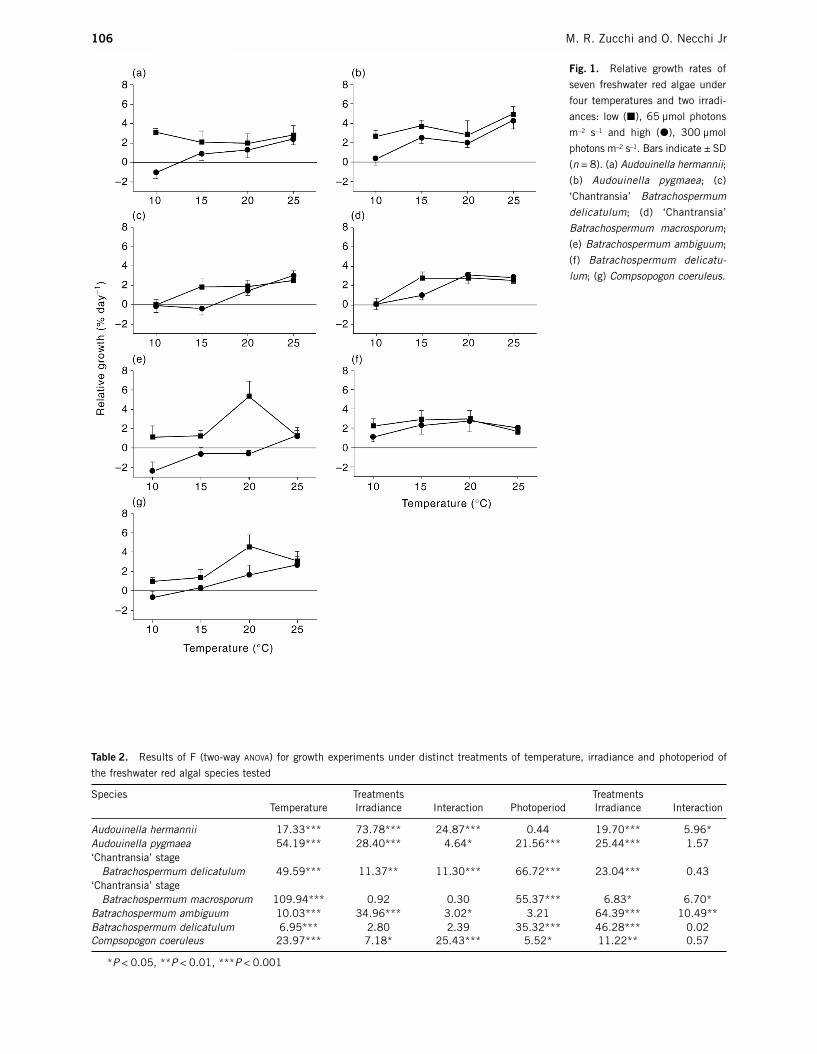
In general, higher growth rates were present in mostalgae under low irradiances (Fig. 1). The only excep-tions to this pattern were cases in which the bestgrowth under high irradiance coincided with the highesttemperature (25°C) (Fig. 1). In contrast, the tempera-ture optima differed either among the algae andbetween low and high irradiances for the same alga. Inthe ‘Chantransia’ stages the peaks were observed at thesame temperature regardless of irradiance: 20°C for‘Chantransia’ of Batrachospermum macrosporum Mon-tagne and 25°C for ‘Chantransia’ of Batrachospermumdelicatulum (Skuja) Necchi et Entwisle. Audouinellahermannii (Roth) Duby had the highest growth at 10°Cunder low irradiance, and at 25°C under high irradi-ance. Batrachospermum ambiguum Montagne andC. coeruleus presented the highest values at 20°C and 25°C under low and high irradiances, respectively.For Audouinella pygmaea (Kützing) Weber van-Bossethe growth patterns were similar under both irradiances,with the highest values at 25°C, whereas in B. delicat-ulum the differences were generally minor with peaks at 15–20°C and 20°C in low and high irradiances. For most algae, growth was negligible, or even negative,at 10°C under high irradiances and at 15°C for some (B. ambiguum, C. coeruleus and the ‘Chantransia’stage of B. delicatulum). ANOVA (two-way) revealedhighly significant differences in temperature effects in all species. In addition, differences between irradi-ance treatments were found in most algae, except for B. delicatulum and the ‘Chantransia’ stage of B. macrosporum (Table 2). A significant interaction
105Freshwater red algae growth and pigment
106 M. R. Zucchi and O. Necchi Jr
Fig. 1. Relative growth rates of
seven freshwater red algae under
four temperatures and two irradi-
ances: low (�), 65 µmol photons
m–2 s–1 and high (�), 300 µmol
photons m–2 s–1. Bars indicate ± SD
(n = 8). (a) Audouinella hermannii;
(b) Audouinella pygmaea; (c)
‘Chantransia’ Batrachospermum
delicatulum; (d) ‘Chantransia’
Batrachospermum macrosporum;
(e) Batrachospermum ambiguum;
(f) Batrachospermum delicatu-
lum; (g) Compsopogon coeruleus.
Table 2. Results of F (two-way ANOVA) for growth experiments under distinct treatments of temperature, irradiance and photoperiod of
the freshwater red algal species tested
Species Treatments TreatmentsTemperature Irradiance Interaction Photoperiod Irradiance Interaction
Audouinella hermannii 17.33*** 73.78*** 24.87*** 0.44 19.70*** 5.96*Audouinella pygmaea 54.19*** 28.40*** 4.64* 21.56*** 25.44*** 1.57‘Chantransia’ stage
Batrachospermum delicatulum 49.59*** 11.37** 11.30*** 66.72*** 23.04*** 0.43‘Chantransia’ stage
Batrachospermum macrosporum 109.94*** 0.92 0.30 55.37*** 6.83* 6.70*Batrachospermum ambiguum 10.03*** 34.96*** 3.02* 3.21 64.39*** 10.49**Batrachospermum delicatulum 6.95*** 2.80 2.39 35.32*** 46.28*** 0.02Compsopogon coeruleus 23.97*** 7.18* 25.43*** 5.52* 11.22** 0.57
*P < 0.05, **P < 0.01, ***P < 0.001
between these parameters was also present in all butthese two algae.
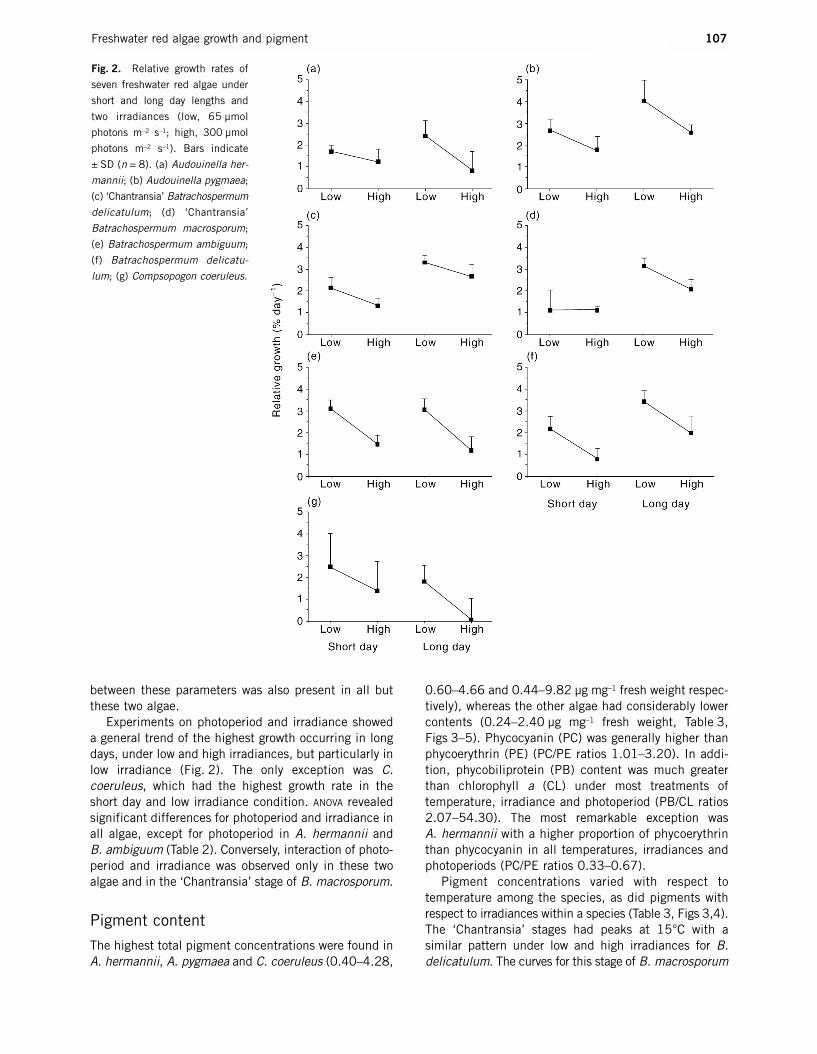
Experiments on photoperiod and irradiance showeda general trend of the highest growth occurring in longdays, under low and high irradiances, but particularly inlow irradiance (Fig. 2). The only exception was C.coeruleus, which had the highest growth rate in theshort day and low irradiance condition. ANOVA revealedsignificant differences for photoperiod and irradiance inall algae, except for photoperiod in A. hermannii and B. ambiguum (Table 2). Conversely, interaction of photo-period and irradiance was observed only in these twoalgae and in the ‘Chantransia’ stage of B. macrosporum.
Pigment content
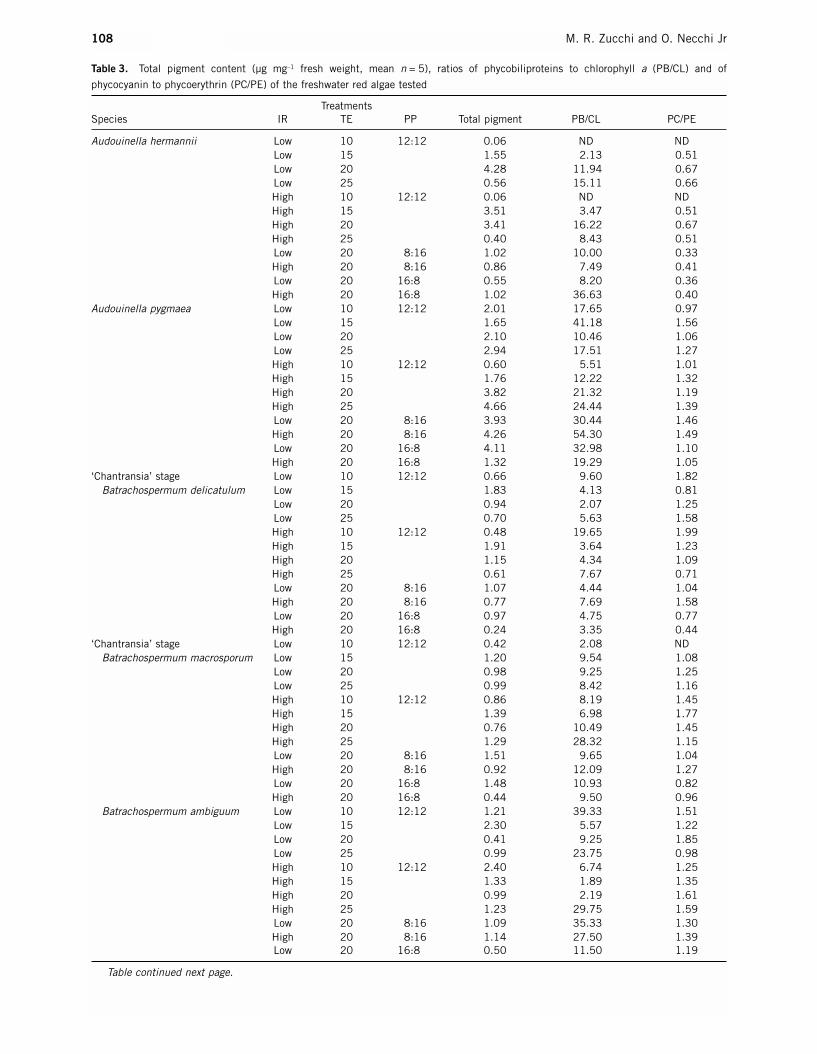
The highest total pigment concentrations were found inA. hermannii, A. pygmaea and C. coeruleus (0.40–4.28,
0.60–4.66 and 0.44–9.82 µg mg–1 fresh weight respec-tively), whereas the other algae had considerably lowercontents (0.24–2.40 µg mg–1 fresh weight, Table 3,Figs 3–5). Phycocyanin (PC) was generally higher thanphycoerythrin (PE) (PC/PE ratios 1.01–3.20). In addi-tion, phycobiliprotein (PB) content was much greaterthan chlorophyll a (CL) under most treatments of temperature, irradiance and photoperiod (PB/CL ratios2.07–54.30). The most remarkable exception was A. hermannii with a higher proportion of phycoerythrinthan phycocyanin in all temperatures, irradiances andphotoperiods (PC/PE ratios 0.33–0.67).
Pigment concentrations varied with respect to temperature among the species, as did pigments withrespect to irradiances within a species (Table 3, Figs 3,4).The ‘Chantransia’ stages had peaks at 15°C with asimilar pattern under low and high irradiances for B.delicatulum. The curves for this stage of B. macrosporum
107Freshwater red algae growth and pigment
Fig. 2. Relative growth rates of
seven freshwater red algae under
short and long day lengths and
two irradiances (low, 65 µmol
photons m–2 s–1; high, 300 µmol
photons m–2 s–1). Bars indicate
± SD (n = 8). (a) Audouinella her-
mannii; (b) Audouinella pygmaea;
(c) ‘Chantransia’ Batrachospermum
delicatulum; (d) ‘Chantransia’
Batrachospermum macrosporum;
(e) Batrachospermum ambiguum;
(f) Batrachospermum delicatu-
lum; (g) Compsopogon coeruleus.
108 M. R. Zucchi and O. Necchi Jr
Table 3. Total pigment content (µg mg–1 fresh weight, mean n = 5), ratios of phycobiliproteins to chlorophyll a (PB/CL) and of
phycocyanin to phycoerythrin (PC/PE) of the freshwater red algae tested
TreatmentsSpecies IR TE PP Total pigment PB/CL PC/PE
Audouinella hermannii Low 10 12:12 0.06 ND NDLow 15 1.55 2.13 0.51Low 20 4.28 11.94 0.67Low 25 0.56 15.11 0.66High 10 12:12 0.06 ND NDHigh 15 3.51 3.47 0.51High 20 3.41 16.22 0.67High 25 0.40 8.43 0.51Low 20 8:16 1.02 10.00 0.33High 20 8:16 0.86 7.49 0.41Low 20 16:8 0.55 8.20 0.36High 20 16:8 1.02 36.63 0.40
Audouinella pygmaea Low 10 12:12 2.01 17.65 0.97Low 15 1.65 41.18 1.56Low 20 2.10 10.46 1.06Low 25 2.94 17.51 1.27High 10 12:12 0.60 5.51 1.01High 15 1.76 12.22 1.32High 20 3.82 21.32 1.19High 25 4.66 24.44 1.39Low 20 8:16 3.93 30.44 1.46High 20 8:16 4.26 54.30 1.49Low 20 16:8 4.11 32.98 1.10High 20 16:8 1.32 19.29 1.05
‘Chantransia’ stage Low 10 12:12 0.66 9.60 1.82Batrachospermum delicatulum Low 15 1.83 4.13 0.81
Low 20 0.94 2.07 1.25Low 25 0.70 5.63 1.58High 10 12:12 0.48 19.65 1.99High 15 1.91 3.64 1.23High 20 1.15 4.34 1.09High 25 0.61 7.67 0.71Low 20 8:16 1.07 4.44 1.04High 20 8:16 0.77 7.69 1.58Low 20 16:8 0.97 4.75 0.77High 20 16:8 0.24 3.35 0.44
‘Chantransia’ stage Low 10 12:12 0.42 2.08 NDBatrachospermum macrosporum Low 15 1.20 9.54 1.08
Low 20 0.98 9.25 1.25Low 25 0.99 8.42 1.16High 10 12:12 0.86 8.19 1.45High 15 1.39 6.98 1.77High 20 0.76 10.49 1.45High 25 1.29 28.32 1.15Low 20 8:16 1.51 9.65 1.04High 20 8:16 0.92 12.09 1.27Low 20 16:8 1.48 10.93 0.82High 20 16:8 0.44 9.50 0.96
Batrachospermum ambiguum Low 10 12:12 1.21 39.33 1.51Low 15 2.30 5.57 1.22Low 20 0.41 9.25 1.85Low 25 0.99 23.75 0.98High 10 12:12 2.40 6.74 1.25High 15 1.33 1.89 1.35High 20 0.99 2.19 1.61High 25 1.23 29.75 1.59Low 20 8:16 1.09 35.33 1.30High 20 8:16 1.14 27.50 1.39Low 20 16:8 0.50 11.50 1.19
Table continued next page.

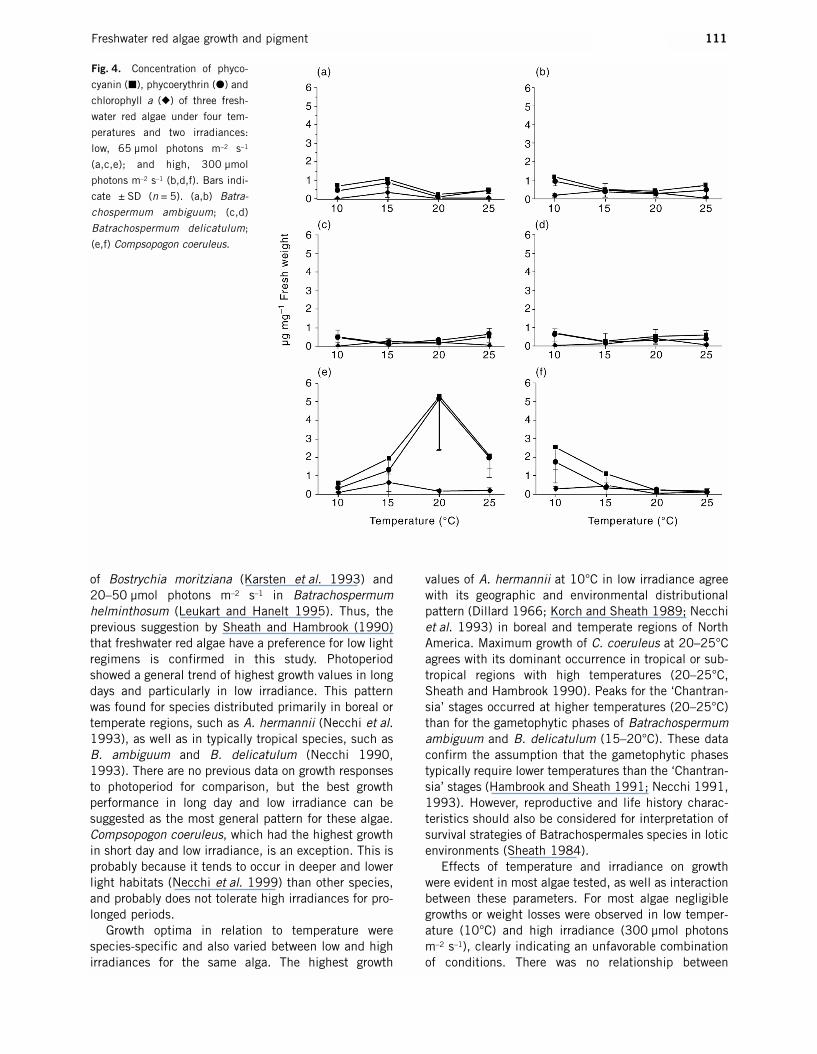
differed between irradiances, and the highest chloro-phyll concentration was at 10°C in low irradiance. In A. hermannii maximum chlorophyll and phycobili-proteins occurred at 15°C and 20°C respectively inboth low and high irradiances. In contrast, there was asharp increasing trend with temperature for the threepigments under high irradiance with peaks at 25°C forphycobiliproteins and at 20°C for chlorophyll under lowirradiance in A. pygmaea. Peaks occurred at 15°C forall pigments under low irradiance in B. ambiguum butdiffered for phycobiliproteins under high irradiance(10°C). In the gametophytic phase of B. delicatulum,phycoerythrin and phycocyanin peaked at 25°C andchlorophyll at 15°C under low irradiance; however, athigh irradiance the peaks were at 10°C for phyco-biliproteins and at 20°C for chlorophyll. It is noteworthythat the highest relative proportions of chlorophyll(PB/CL ratios 1.11–1.95) occurred under high irradi-ance at 15–20°C or in low irradiance at 15°C inB. ambiguum and B. delicatulum. In C. coeruleus themaxima for chlorophyll occurred at 15°C in low andhigh irradiances, whereas for phycobiliproteins pronounced peaks occurred at 20°C in low irradiance,but they peaked at 10°C and had a decreasing trendwith temperature under high irradiance.
ANOVA revealed significant differences among treat-ments of temperature for most species and pigments
(Table 4). The most notable exception was the‘Chantransia’ stage of B. macrosporum with no differ-ences for phycobiliproteins. In contrast, no significantdifferences were found between treatments of irradi-ance for the algae tested, except for phycobiliproteinsin C. coeruleus and chlorophyll in B. ambiguum. Simi-larly, no interactions between the two parameters wereobserved, except for phycobiliproteins in C. coeruleusand chlorophyll in B. delicatulum and the ‘Chantransia’stage of B. macrosporum.
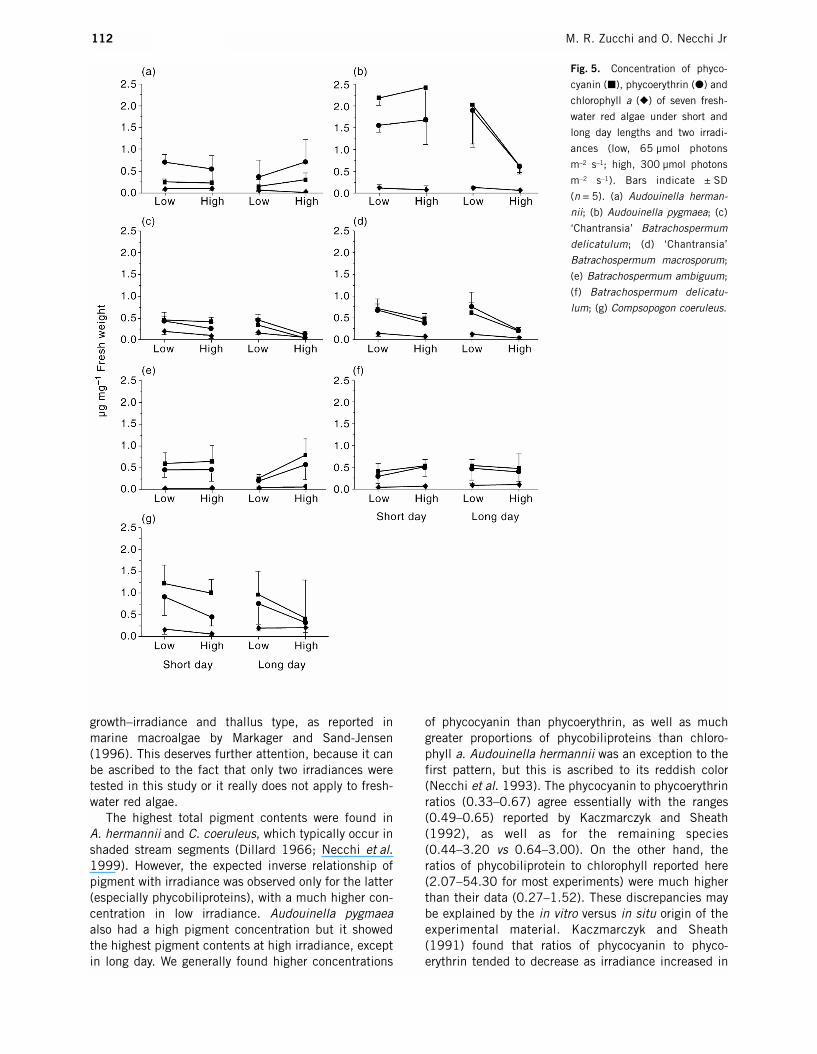
Responses in relation to photoperiod and irradiancevaried among the species and also for pigments in the same alga (Table 3, Fig. 5). Four algae had a sharpdecrease in total pigment concentrations in long day and high irradiance: the ‘Chantransia’ stages, A. pygmaea and C. coeruleus. In C. coeruleus, how-ever, the highest chlorophyll values occurred in thistreatment. The highest total pigment contents wereobserved under the following conditions: short day andlow irradiance in C. coeruleus and the ‘Chantransia’stages, short day and high irradiance in A. pygmaea,long day and low irradiance in B. delicatulum, and longday and high irradiance in B. ambiguum; equal con-tents were found in short day and low irradiance or longday and high irradiance in A. hermannii. The highestphycobiliprotein values were observed under the fol-lowing conditions: long day and high irradiance in
109Freshwater red algae growth and pigment
Table 3. Continued
TreatmentsSpecies IR TE PP Total pigment PB/CL PC/PE
High 20 16:8 1.44 17.00 1.39Batrachospermum delicatulum Low 10 12:12 1.02 33.00 0.98
Low 15 0.59 1.11 0.72Low 20 0.74 2.52 1.94Low 25 1.25 24.00 1.31High 10 12:12 1.36 33.00 1.03High 15 0.62 3.13 1.23High 20 1.24 1.95 1.56High 25 1.04 10.55 1.50Low 20 8:16 0.71 22.67 1.34High 20 8:16 1.09 17.17 1.06Low 20 16:8 1.12 13.00 1.26High 20 16:8 0.97 8.70 1.18
Compsopogon coeruleus Low 10 12:12 1.08 8.82 1.77Low 15 3.89 5.17 1.49Low 20 9.82 53.56 0.86Low 25 4.24 20.20 1.05High 10 12:12 4.58 15.35 1.47High 15 1.90 3.42 3.20High 20 0.50 11.50 0.92High 25 0.44 2.38 1.58Low 20 8:16 2.28 14.20 1.34High 20 8:16 1.49 20.29 2.38Low 20 16:8 1.91 8.55 1.28High 20 16:8 0.93 3.43 1.25
IR, irradiance (Low, 65 µmol photons m–2 s–1; high, 300 µmol photons m–2 s–1); ND, no data; PP, photoperiod (light:dark); TE, temperature (°C). Photoperiod 12 : 12 if not otherwise specified.
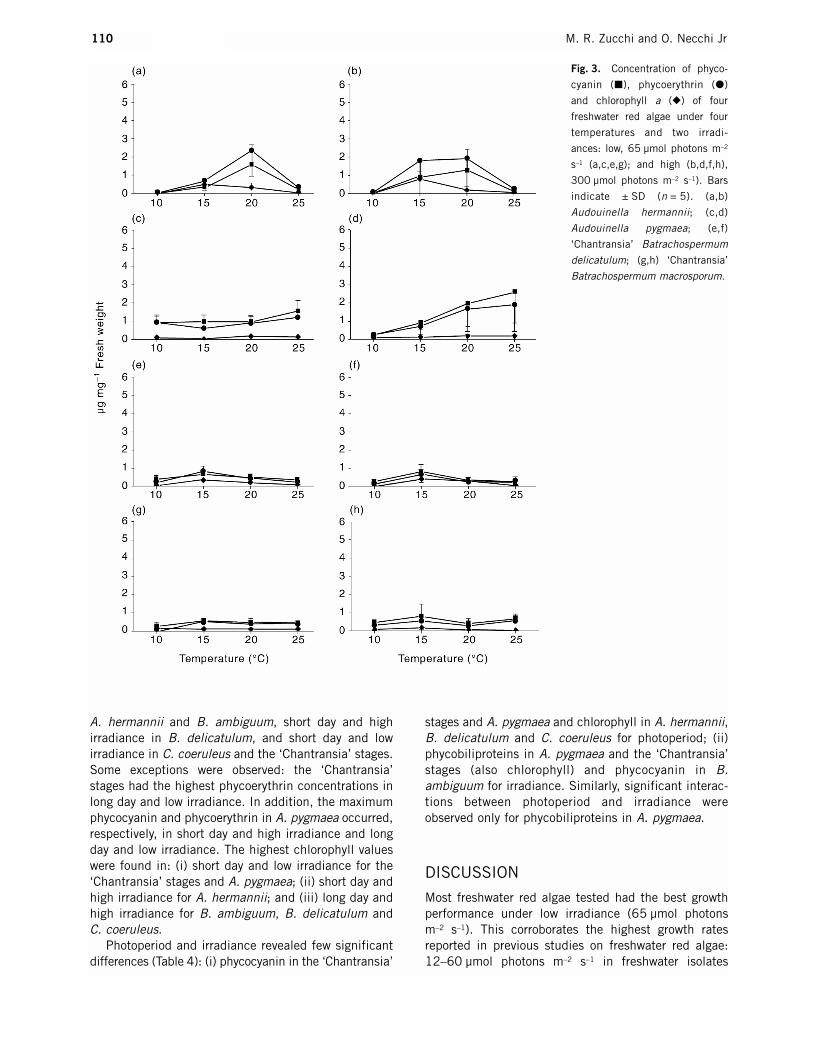
A. hermannii and B. ambiguum, short day and high irradiance in B. delicatulum, and short day and low irradiance in C. coeruleus and the ‘Chantransia’ stages.Some exceptions were observed: the ‘Chantransia’stages had the highest phycoerythrin concentrations inlong day and low irradiance. In addition, the maximumphycocyanin and phycoerythrin in A. pygmaea occurred,respectively, in short day and high irradiance and longday and low irradiance. The highest chlorophyll valueswere found in: (i) short day and low irradiance for the‘Chantransia’ stages and A. pygmaea; (ii) short day andhigh irradiance for A. hermannii; and (iii) long day andhigh irradiance for B. ambiguum, B. delicatulum and C. coeruleus.
Photoperiod and irradiance revealed few significantdifferences (Table 4): (i) phycocyanin in the ‘Chantransia’
stages and A. pygmaea and chlorophyll in A. hermannii,B. delicatulum and C. coeruleus for photoperiod; (ii)phycobiliproteins in A. pygmaea and the ‘Chantransia’stages (also chlorophyll) and phycocyanin in B.ambiguum for irradiance. Similarly, significant interac-tions between photoperiod and irradiance wereobserved only for phycobiliproteins in A. pygmaea.
DISCUSSION
Most freshwater red algae tested had the best growthperformance under low irradiance (65 µmol photonsm–2 s–1). This corroborates the highest growth ratesreported in previous studies on freshwater red algae:12–60 µmol photons m–2 s–1 in freshwater isolates
110 M. R. Zucchi and O. Necchi Jr
Fig. 3. Concentration of phyco-
cyanin (�), phycoerythrin (�)
and chlorophyll a (�) of four
freshwater red algae under four
temperatures and two irradi-
ances: low, 65 µmol photons m–2
s–1 (a,c,e,g); and high (b,d,f,h),
300 µmol photons m–2 s–1). Bars
indicate ± SD (n = 5). (a,b)
Audouinella hermannii; (c,d)
Audouinella pygmaea; (e,f)
‘Chantransia’ Batrachospermum
delicatulum; (g,h) ‘Chantransia’
Batrachospermum macrosporum.
of Bostrychia moritziana (Karsten et al. 1993) and20–50 µmol photons m–2 s–1 in Batrachospermumhelminthosum (Leukart and Hanelt 1995). Thus, theprevious suggestion by Sheath and Hambrook (1990)that freshwater red algae have a preference for low lightregimens is confirmed in this study. Photoperiodshowed a general trend of highest growth values in longdays and particularly in low irradiance. This pattern was found for species distributed primarily in boreal ortemperate regions, such as A. hermannii (Necchi et al.1993), as well as in typically tropical species, such asB. ambiguum and B. delicatulum (Necchi 1990,1993). There are no previous data on growth responsesto photoperiod for comparison, but the best growth performance in long day and low irradiance can be suggested as the most general pattern for these algae.Compsopogon coeruleus, which had the highest growthin short day and low irradiance, is an exception. This isprobably because it tends to occur in deeper and lowerlight habitats (Necchi et al. 1999) than other species,and probably does not tolerate high irradiances for pro-longed periods.
Growth optima in relation to temperature werespecies-specific and also varied between low and highirradiances for the same alga. The highest growth
values of A. hermannii at 10°C in low irradiance agreewith its geographic and environmental distributionalpattern (Dillard 1966; Korch and Sheath 1989; Necchiet al. 1993) in boreal and temperate regions of NorthAmerica. Maximum growth of C. coeruleus at 20–25°Cagrees with its dominant occurrence in tropical or sub-tropical regions with high temperatures (20–25°C,Sheath and Hambrook 1990). Peaks for the ‘Chantran-sia’ stages occurred at higher temperatures (20–25°C)than for the gametophytic phases of Batrachospermumambiguum and B. delicatulum (15–20°C). These dataconfirm the assumption that the gametophytic phasestypically require lower temperatures than the ‘Chantran-sia’ stages (Hambrook and Sheath 1991; Necchi 1991,1993). However, reproductive and life history charac-teristics should also be considered for interpretation ofsurvival strategies of Batrachospermales species in loticenvironments (Sheath 1984).
Effects of temperature and irradiance on growthwere evident in most algae tested, as well as interactionbetween these parameters. For most algae negligiblegrowths or weight losses were observed in low temper-ature (10°C) and high irradiance (300 µmol photonsm–2 s–1), clearly indicating an unfavorable combinationof conditions. There was no relationship between
111Freshwater red algae growth and pigment
Fig. 4. Concentration of phyco-
cyanin (�), phycoerythrin (�) and
chlorophyll a (�) of three fresh-
water red algae under four tem-
peratures and two irradiances:
low, 65 µmol photons m–2 s–1
(a,c,e); and high, 300 µmol
photons m–2 s–1 (b,d,f). Bars indi-
cate ± SD (n = 5). (a,b) Batra-
chospermum ambiguum; (c,d)
Batrachospermum delicatulum;
(e,f) Compsopogon coeruleus.
growth–irradiance and thallus type, as reported inmarine macroalgae by Markager and Sand-Jensen(1996). This deserves further attention, because it canbe ascribed to the fact that only two irradiances weretested in this study or it really does not apply to fresh-water red algae.
The highest total pigment contents were found in A. hermannii and C. coeruleus, which typically occur inshaded stream segments (Dillard 1966; Necchi et al.1999). However, the expected inverse relationship ofpigment with irradiance was observed only for the latter(especially phycobiliproteins), with a much higher con-centration in low irradiance. Audouinella pygmaeaalso had a high pigment concentration but it showedthe highest pigment contents at high irradiance, exceptin long day. We generally found higher concentrations
of phycocyanin than phycoerythrin, as well as muchgreater proportions of phycobiliproteins than chloro-phyll a. Audouinella hermannii was an exception to thefirst pattern, but this is ascribed to its reddish color(Necchi et al. 1993). The phycocyanin to phycoerythrinratios (0.33–0.67) agree essentially with the ranges(0.49–0.65) reported by Kaczmarczyk and Sheath(1992), as well as for the remaining species(0.44–3.20 vs 0.64–3.00). On the other hand, theratios of phycobiliprotein to chlorophyll reported here(2.07–54.30 for most experiments) were much higherthan their data (0.27–1.52). These discrepancies maybe explained by the in vitro versus in situ origin of theexperimental material. Kaczmarczyk and Sheath(1991) found that ratios of phycocyanin to phyco-erythrin tended to decrease as irradiance increased in
112 M. R. Zucchi and O. Necchi Jr
Fig. 5. Concentration of phyco-
cyanin (�), phycoerythrin (�) and
chlorophyll a (�) of seven fresh-
water red algae under short and
long day lengths and two irradi-
ances (low, 65 µmol photons
m–2 s–1; high, 300 µmol photons
m–2 s–1). Bars indicate ± SD
(n = 5). (a) Audouinella herman-
nii; (b) Audouinella pygmaea; (c)
‘Chantransia’ Batrachospermum
delicatulum; (d) ‘Chantransia’
Batrachospermum macrosporum;
(e) Batrachospermum ambiguum;
(f) Batrachospermum delicatu-
lum; (g) Compsopogon coeruleus.
laboratory studies on B. boryanum. In contrast, our datashowed no clear trend.
The most significant differences in pigment contentswere found for the temperature treatments. Few differ-ences could be attributed to irradiance and photope-riod, or the weak interactions among these threeparameters. Thirb and Benson-Evans (1984) found thatchlorophyll and phycoerythrin concentrations in theapical tips of the gametophytes and in filaments of the‘Chantransia’ stage of Lemanea sp. decreased withincreasing temperature (from 5 to 25°C) under low irra-diance (ca 25 µmol photons m–2 s–1). We found similarresults only in C. coeruleus but an opposite trend in A.pygmaea, both under high irradiance (300 µmolphotons m–2 s–1). However, for most algae tested theresponses were species-specific.
There was no correspondence between growth ratesand pigment contents either for individual species oramong them, indicating that the most favorable condi-tions for growth were generally not coincident withthose with highest pigment contents. Falkowski andRaven (1997) suggest some hypotheses to account forthe differences in growth rates among species that
seem to be applicable to the present study. Theseinclude: (i) different light harvesting properties, allow-ing some species to absorb available radiation moreeffectively than others; (ii) distinct photosynthesis/res-piration ratios, yielding variable energy supply tosupport growth; and (iii) differences in levels of organiccarbon reduction, which can explain growth rates. Par-allel studies on photosynthesis/respiration on thesesame algae are in progress. Together, these data willhelp clarify the ecophysiological characteristics offreshwater red algae and provide insight into the func-tional roles of these organisms in lotic ecosystems.
ACKNOWLEDGEMENTS
This research was supported by a FAPESP Grant-in-Aid(98/07537–8) and a National Council of Scientific andTechnological Development (CNPq) research grant(300379/86–2) to ONJ and a CAPES fellowship toMRZ. We are grateful to Maria Helena Carabolante forhelp in laboratory analyses; to John A. West, MelbourneUniversity, Australia, and Franklin D. Ott, USA, forkindly providing culture isolates.
113Freshwater red algae growth and pigment
Table 4. Results of F (ANOVA – two way) for pigment concentration experiments under distinct treatments of temperature, irradiance and
photoperiod of the freshwater red algal species tested
Treatments TreatmentsSpecies Temperature Irradiance Interaction Photoperiod Irradiance Interaction
Audouinella hermanniiPhycoerythrin 4.93* 0.10 0.04 0.25 0.30 2.31Phycocyanin 13.65*** 0.29 0.14 0.05 0.56 0.77Chlorophyll a 15.35*** 0.39 1.90 22.29*** 1.69 3.69
Audouinella pygmaeaPhycoerythrin 3.75* 0.58 1.93 2.47 6.30* 9.53**Phycocyanin 6.14* 1.57 2.60 13.19** 4.71* 9.28**Chlorophyll a 1.91 0.01 0.13 0.07 3.53 0.02
‘Chantransia’ stage Batrachospermum delicatulumPhycoerythrin 9.65*** 0.78 0.50 1.21 22.78*** 2.35Phycocyanin 9.38*** 0.51 1.19 13.37** 6.55* 4.29Chlorophyll a 42.64*** 0.04 0.85 1.73 22.38*** 0.01
‘Chantransia’ stage Batrachospermum macrosporumPhycoerythrin 1.34 0.01 1.10 0.24 17.01** 1.38Phycocyanin 2.00 1.73 0.56 7.22* 22.60*** 2.06Chlorophyll a 5.52* 0.30 3.73* 1.02 11.38** 0.05
Batrachospermum ambiguumPhycoerythrin 2.96* 0.01 2.43 0.40 3.20 2.99Phycocyanin 3.20* 0.31 2.40 0.53 4.80* 3.46Chlorophyll a 8.33*** 5.94* 1.06 3.66 1.51 0.67
Batrachospermum delicatulumPhycoerythrin 3.22* 0.21 0.47 0.14 0.58 1.80Phycocyanin 3.49* 0.76 0.40 0.30 0.02 0.92Chlorophyll a 13.25*** 0.84 3.83* 5.18* 2.10 0.10
Compsopogon coeruleusPhycoerythrin 3.76* 12.51** 8.47*** 0.42 4.27 0.01Phycocyanin 2.53 11.11** 10.92*** 2.71 2.16 0.43Chlorophyll a 7.79*** 0.84 1.49 5.65* 0.98 1.44
*P < 0.05, **P < 0.01, ***P < 0.001.

REFERENCES
Beer, S. and Eshel, A. 1985. Determining phycoerythrin andphycocyanin concentrations in aqueous crude extracts ofred algae. Aust. J. Mar. Freshwat. Res. 36: 785–92.
Dillard, G. E. 1966. The seasonal periodicity of Batracho-spermum macrosporum Mont. & Audouinella violacea(Kuetz.) Ham. In Turkey Creek, Moore County, North Car-olina. J. Elisha Mitchell Sci. Soc. 82: 204–7.
Falkowski, P. G. and Raven, J. A. 1997. Aquatic Photo-synthesis. Blackwell Science, Malden, 375 pp.
Gantt, E., Scott, J. and Lipschultz, C. 1986. Phycobiliproteincomposition and chloroplast structure in the freshwater redalga Compsopogon coeruleus (Rhodophyta). J. Phycol. 22:480–4.
Glazer, A. N., Chan, C. F., Karsten, U. and West, J. A. 1994.Salinity tolerance, biliproteins, and florisoside content ofCompsopogon coeruleus (Rhodophyta). J. Phycol. 30:457–61.
Glazer, A. N., Chan, C. F. and West, J. A. 1997. An unusualphycocyanobilin-containing phycoerythrin of severalbluish-colored, acrochaetioid, freshwater red algal species.J. Phycol. 33: 617–24.
Hambrook, J. A. and Sheath, R. G. 1991. Reproductiveecology of Batrachospermum boryanum Sirodot in a tem-perate headwater stream. Hydrobiologia 218: 233–46.
Kaczmarczyk, D. and Sheath, R. G. 1991. The effect of lightregime on the photosynthetic apparatus of the freshwaterred alga Batrachospermum boryanum. Cryptog. Algol. 12:249–63.
Kaczmarczyk, D. and Sheath, R. G. 1992. Pigment contentand carbon to nitrogen ratios of freshwater red algaegrowing at different light levels. Japn. J. Phycol. 40:279–82.
Kain, J. M. 1987. Seasonal growth and photoinhibition inPlocamium cartilagineum (Rhodophyta) off the Isle ofMan. Phycologia 26: 88–99.
Karsten, U., West, J. A. and Ganesan, E. K. 1993. Compara-tive physiological ecology of Bostrychia moritziana(Ceramiales, Rhodophyta) from freshwater and marinehabitats. Phycologia 32: 401–9.
Korch, J. E. and Sheath, R. G. 1989. The phenology ofAudouinella violacea (Acrochaetiaceae, Rhodophyta) in aRhode Island stream (USA). Phycologia 28: 228–36.
Leukart, P. and Hanelt, D. 1995. Light requirements forphotosynthesis and growth in several macroalgae from asmall soft-water stream in the Spessart Mountains,Germany. Phycologia 34: 528–32.
Markager, S. and Sand-Jensen, K. 1996. Implications ofthallus thickness for growth-irradiance relationships ofmarine macroalge. Eur. J. Phycol. 31: 79–87.
Necchi, O. Jr. 1990. Revision of the genus BatrachospermumRoth (Rhodophyta, Batrachospermales) in Brazil. Bibl.Phycol. 84: 1–201.
Necchi, O. Jr. 1991. The section Sirodotia of Batrachosper-mum (Rhodophyta, Batrachospermaceae) in Brazil. Algol.Stud. 62: 17–30.
Necchi, O. Jr. 1993. Distribution and seasonal dynamics ofRhodophyta in the Preto river basin, southeastern Brazil.Hydrobiologia 250: 81–90.
Necchi, O. Jr, Branco, C. C. Z. and Gomes, R. R. V. 1999.Microhabitat and plant structure of Compsopogoncoeruleus (Compsopogonaceae, Rhodophyta) populationsin streams from São Paulo State, southeastern Brazil.Cryptog. Algol. 20: 75–87.
Necchi, O. Jr, Sheath, R. G. and Cole, K. M. 1993. System-atics of the freshwater Audouinella (Acrochaetiaceae,Rhodophyta) in North America.1. The reddish species.Algol. Stud. 70: 11–28.
Necchi, O. Jr and Zucchi, M. R. 1997. Audouinella macro-spora (Acrochaetiaceae, Rhodophyta) is the ‘Chantransia’stage of Batrachospermum (Batrachospermaceae). Phycologia 36: 220–4.
Rider, D. E. and Wagner, R. H. 1972. The relationship oflight, temperature and current to the seasonal distributionof Batrachospermum (Rhodophyta). J. Phycol. 8: 323–31.
Sheath, R. G. 1984. The biology of freshwater red algae. In:Round, F. E. and Chapman, D. J. (Eds) Progress in Phyco-logical Research, Vol. 3. Biopress, Bristol, pp. 89–157.
Sheath, R. G. and Hambrook, J. A. 1988. Mechanical adap-tations to flow in freshwater red algae. J. Phycol. 24:107–11.
Sheath, R. G. and Hambrook, J. A. 1990. Freshwater ecology.In: Cole, K. M. and Sheath, R. G. (Eds) Biology of the Red Algae. Cambridge University Press, Cambridge, pp. 423–53.
Thirb, H. H. and Benson-Evans, K. 1983. The effect of dif-ferent light intensities and wavelengths on carpospore germination and the apical tips of the red alga LemaneaBory (1808). Nova Hedwigia 37: 669–82.
Thirb, H. H. and Benson-Evans, K. 1984. The effect of tem-perature on the growth of Lemanea thalli and carposporegermination. Arch. Hydrobiol. 103: 341–6.
Thirb, H. H. and Benson-Evans, K. 1985. The effect of sus-pended solids on the growth of apical tips of gametophyteplants of Lemanea and on carpospore germination andsubsequent colonisation. Arch. Hydrobiol. 103: 409–17.
Wetzel, R. G. and Likens, G. E. 1991. Limnological Analyses.Springer-Verlag, New York, 390 pp.
Zar, J. H. 1999. Biostatistical Analysis. Prentice Hall, NewJersey, 663 pp.
114 M. R. Zucchi and O. Necchi Jr




























