Embed Size (px)
Citation preview

GLOBAL BIOGEOCHEMICAL CYCLES, VOL. 1, NO. 3, PAGES 171-186, SEPTEMBER 1987
OCEANIC PRIMARY PRODUCTION ESTIMATES FROM
MEASUREMENTS OF SPECTRAL IRRADIANCE AND
PIGMENT CONCENTRATIONS
Robert R. Bidigare, • R.C. Smith,2 K.S. Baker,S and J. Marra •
Abstract. A major objective of biological oceanography today is to quantify the mean and the variance of phytoplankton production on a global basis. Synoptic satellite sensing of the world's ocean is essential to this effort
which requires contemporaneous sea surface data to provide complete water column information. Toward these objectives we present a spectrally dependent bio-optical model for the computation of in situ phytoplankton production. Using this model we show that in situ phytoplankton production can be accurately estimated from measurements of incident spectral
•Geochemical and Environmental Research
Group, Texas A&M University, College Station.
2University of California Marine Bio- Optics Group, Department of Geography, University of California, Santa Barbara.
SUniversity of California Marine Bio- Optics Group,Scripps Institution of Oceanography, University of California, San Diego.
•Lamont-Doherty Geological Observatory of Columbia University, Palisades, New York.
Copyright 1988 by the American Geophysical Union
Paper number 7J0728. 0886-6236/88/007J-0728510.00
irradiance and phytoplankton pigment concentrations. We also present estimates of photosynthetically absorbed radiation as a function of wavelength for a natural phytoplankton population. These complete spectral data provide insight into the wavelength-dependent utilization of radiant energy by phytoplankton and the influence of phytoplankton on the optical properties of the water column. We show that the model can be used for shipboard observations and that it may be especially useful for predicting production rates from data provided by untended buoys.
INTRODUCTION
The estimation of primary production and an increased understanding of the factors controlling marine biogeochemical cycling from regional to global scales are major goals of ocean scientists [National Academy of Sciences, 1984]. Phytoplankton, the major component responsible for primary production in the world's ocean, have a principal influence on elemental cycles. The abundance and distributions of phytoplankton are primarily dependent upon the availability of nutrients and radiant energy. The fluxes of these rate-limiting factors in the upper ocean are governed by the physical, biological and optical properties of the water column. As a consequence, an integrated research

I?P Bidigare et al.: Oceanic Primary Production Estimates
approach is required to define the causal links between physical, biological and optical processes.
Biological variability is poorly sampled by classical shipboard operations. This has led to the development of remote sensing techniques to estimate pigment biomass [Gordon et at., 1980a; Hovis et at., 1980; Smith and Baker, 1982; Shannon et at., 1983; McCtain et at., 1984] and primary production [Smith et al., 1982, t987a; Brown et at., 1985; Epptey et at., 1985; Platt, 1986]. Investigations of biological variability have been advanced by this development, but these approaches are limited since they only provide information for the upper attenuation length (i.e., the 37% light level) of the water column [Gordon and McCtuney, 1975; Smith, 1981] over long time scales. Thus, there are strong reasons for multiplatform sampling strategies [Smith et at., t987a] where ships and moorings provide complementary information on the vertical structure of the water column over shorter
time scales.
Knowledge of the absorption properties of natural phytoplankton assemblages can be used to calculate "photosynthetically absorbed radiation," an important parameter for modeling phytoplankton production rates [cf. Morel, 1978; Kirk, 1983]. However, many of the previous attempts to estimate phytoplankton absorption in natural waters have been confounded by the presence of covarying detrital material [Kiefer and SooHoo, 1982; Lewis et al., 1985a, b], and the fact that phytoplankton pigment composition changes with depth and physiological state [Falkowski et al., 1985; Bidigare et al., 1985; SooHoo et al., 1986; Sathyendranath et al., 1987]. To establish links between these bio-
optical interactions, a series of stations were occupied in the Sargasso Sea to examine the temporal and spatial variability of spectral irradiance, algal pigments and primary production in the water column. These data were
incorporated into a spectrally dependent bio-optical model, and the results demonstrate that primary production rates can be accurately estimated from incident spectral irradiance measurements and the distributions of algal pigments.
BIO-OPTICAL MODEL
The rate of phytoplankton photosynthesis depends ultimately upon the
rate of photon capture from incident solar radiation. The effectiveness of utilizing photons depends upon two coefficients: the absorption coefficient, which represents the ability of the pigments to absorb photons, and the quantum yield of photosynthesis, which represents the efficiency with which absorbed radiation is used to fix carbon. A simple model [Kirk, 1983] of photosynthesis based on these coefficients is
P(z) - •(z) 12,000 PHAR(z) (•)
where P(z) is the daily rate of photosynthesis (mg C m -s d -1) at a specific depth (z); •(z) is the in situ quantum yield (mol C Einstein-l); 12,000 is a factor to convert moles of carbon to
milligrams of carbon; and PHAR(z) is the photosynthetically absorbed radiation (Einstein m -s d-l). In the approach describ•_dhere, PHARwas calculated from the spectral quantum irradiance associated with radiant energy, Q(z,%), and the wavelength-dependent absorption properties of the resident phytoplankton at a specific depth:
PHAR(z) -- 700 5
7Q(z,•) Zai(• ) Ci(z ) d• (2) 400 i= 1
where ai(% ) are the specific spectral absorption coefficients for the major absorbing pigment groups (m 2 mg -1) and Ci(z ) are the pigment concentrations (mg m -•) at a specific depth (z). While variations in the in vivo absorption properties of the individual pigments are relatively minor [Haardt and Maske, 1987; Mitchell, 1987], quantum yield can vary up to tenfold as a function of growth irradiance [Tyler, 1975; Kishino et al. , 1986] and thus changes with depth. Kiefer and Mitchell [1983] , working with laboratory data, determined that a Michaelis-Menten function could be used to
model the dependence of quantum yield on growth irradiance. However, their model is not spectrally dependent and is only valid for "white" light. Since spectral quality changes with depth , a new formulation of the Kiefer-Mitchell model
was required to provide accurate estimates of in situ quantum yield for use in equation (1). The following equation differs from the Kiefer-Mitchell model in
that the wavelength-dependent Q(%) and ai(• ) and the wavelength-independent •max and Pmax terms are treated separately and
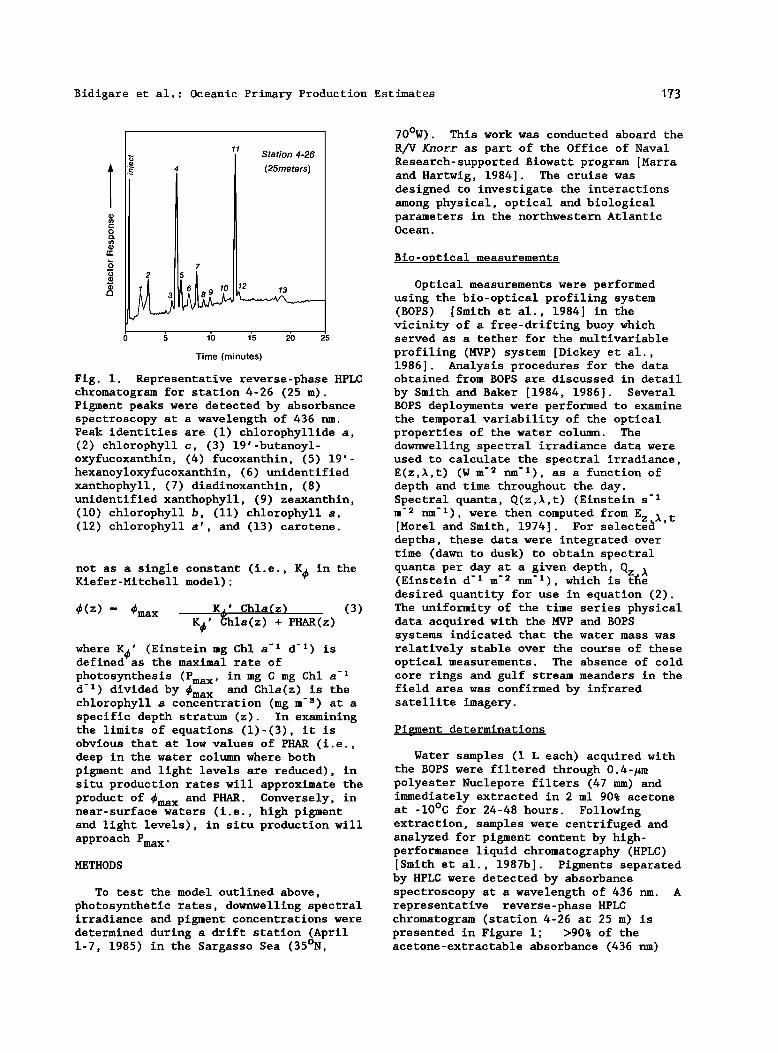
Bidigare et at.- Oceanic Primary Production Estimates 173
Station 4-26
(25meters)
•2 13
0 5 10 15 20 25
Time (minutes)
Fig. 1. Representative reverse-phase HPLC chromatogram for station 4- 26 (25 m). Pigment peaks were detected by absorbance spectroscopy at a wavelength of 436 nm. Peak identities are (1) chlorophyllide a, (2) chlorophyll c, (3) 19 w -butanoyl- oxyfucoxanthin, (4) fucoxanthin, (5) t9 w- hexanoyloxyfucoxanthin, (6) unidentified xanthophyll, (7) diadinoxanthin, (8) unidentified xanthophyll, (9) zeaxanthin, (10) chlorophyll b, (lt) chlorophyll a, (12) chlorophyll a • , and (13) carotene.
not as a single constant (i.e., K• in the Kiefer-Mitchell model)'
•(z)- •max K•' Chla(z) (3) 'Chla(z) + PHAR(z)
where K•' (Einstein mg Chl a TM d -z) is defined-as the maximal rate of
photosynthesis (Pmax' in mg C mg Chl a -z d -z) divided by •max and Chla(z) is the chlorophyll a concentration (mg m -s) at a specific depth stratum (z). In examining the limits of equations (1)-(3), it is obvious that at low values of PHAR (i.e., deep in the water column where both pigment and light levels are reduced), in situ production rates will approximate the product of •max and PHAR. Conversely, in near-surface waters (i.e. , high pigment and light levels), in situ production will approach Pmax'
METHODS
To test the model outlined above, photosynthetic rates, downwelling spectral irradiance and pigment concentrations were determined during a drift station (April 1-7, 1985) in the Sargasso Sea (35øN,
70øW). This work was conducted aboard the R/VKnorr as part of the Office of Naval Research-supported Biowatt program [Marra and Hartwig, 1984]. The cruise was designed to investigate the interactions among physical, optical and biological parameters in the northwestern Atlantic Ocean.
Bio-optical measurements
Optical measurements were performed using the bio-optical profiling system (BOPS) [Smith et al., 1984] in the vicinity of a free-drifting buoy which served as a tether for the multivariable
profiling (MVP) system [Dickey et al., 1986]. Analysis procedures for the data obtained from BOPS are discussed in detail
by Smith and Baker [1984, 1986]. Several BOPS deployments were performed to examine the temporal variability of the optical properties of the water column. The downwelling spectral irradiance data were used to calculate the spectral irradiance, E(z,%,t) (W m -2 nm-Z), as a function of depth and time throughout the day. Spectral quanta, Q(z,%,t) (Einstein s -z m -2 nm-Z), were then computed from E z • t [Morel and Smith, 1974]. For selecte• ' depths, these data were integrated over time (dawn to dusk) to obtain spectral
quanta per day at a given depth, Qz • 1 2 1 (Einstein d- m- nm- ), which is the
desired quantity for use in equation (2). The uniformity of the time series physical data acquired with the MVP and BOPS systems indicated that the water mass was relatively stable over the course of these optical measurements. The absence of cold core rings and gulf stream meanders in the field area was confirmed by infrared satellite imagery.
Pigment determinations
Water samples (1 L each) acquired with the BOPS were filtered through 0.4-•m polyester Nuclepore filters (47 mm) and immediately extracted in 2 ml 90% acetone at -10øC for 24-48 hours. Following extraction, samples were centrifuged and analyzed for pigment content by high- performance liquid chromatography (HPLC) [Smith et al., 1987b]. Pigments separated by HPLC were detected by absorbance spectroscopy at a wavelength of 436 nm. A representative reverse-phase HPLC chromatogram (station 4-26 at 25 m) is presented in Figure 1; >90% of the acetone-extractable absorbance (436 nm)

174 Bidigare et al.: Oceanic Primary Production Estimates
was identified and quantified using the HPLC method employed. Concentrations of phaeopigments (assumed to be primarily in the form of phaeophorbide a [Trees et al., 1986] were determined using the standard fluorometric method [Smith et al., 1981] since concentrations were below the limit
of HPLC quantification. Overestimation of phaeopigment concentration produced by the presence of chlorophyll b [Trees et al., 1985; Vernet and Lorenzen, 1987] averaged 27% in the upper 200 m.
Primary productivity
In situ primary production rates were directly measured using the 14 C_labeling technique [Strickland and Parsons, 1969] . Precautions were taken to avoid
contamination in sampling, isotope addition, and incubation [Fitzwater et al., 1982], although a completely "clean" protocol was not possible. Samples were transferred to 250 ml polycarbonate bottles, and after the addition of the isotopic tracer, were incubated in situ from a free-floating spar buoy at the depths from which they had been collected. Water samples for primary production were collected before dawn and incubated from dawn to dusk. At the end of the
incubation, the samples were filtered onto methyl-cellulose filters (Millipore HA), and the filters counted using liquid scintillation [Marra and Heinemann, 1984] .
Specific absorption coefficients
Since diatoms were a major biomass component at this station (F. Reid, personal commumication, 1987), we have utilized corresponding spectral absorption data available in the literature.
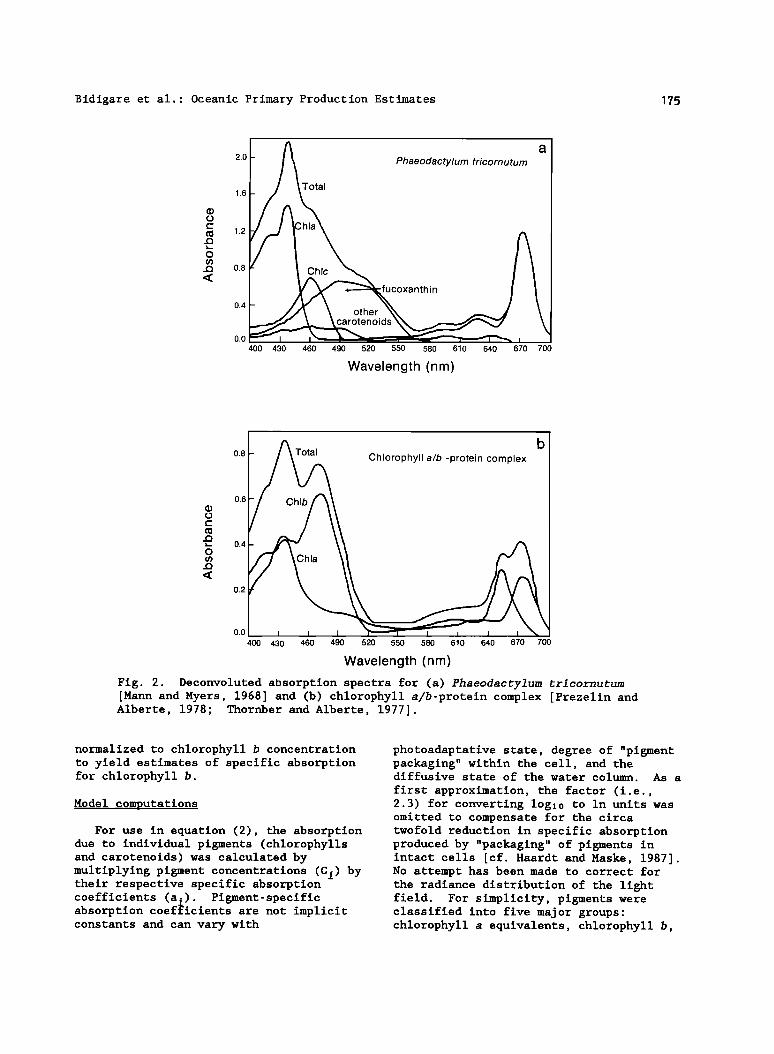
Specific absorption coefficients for the diatom pigments (chlorophyll a, fucoxanthin, chlorophyll c and nonphotosynthetic carotenoids) were determined from the deconvoluted "in vivo"
absorption spectra published for the major pigments isolated from Phaeodactylum tricornugum [Mann and Myers, 1968] . Their absorption spectra (m -1) were digitized (400-700 nm) in 2-nm increments (Figure 2a) and normalized to pigment content (mg m -s) to yield estimates of specific absorption, a i (m 2 mg-1). The concentration of chlorophyll a was estimated to be 13.9 •g m1-1 from their absorption spectrum of a 90% acetone
extract of this diatom [Mann and Myers, 1968] and an extinction coefficient of 87.67 L g-1 cm-1 [Jeffrey and Humphery, 1975]. Concentrations of the remaining pigments (chlorophyll c, fucoxanthin, diadinoxanthin and •-carotene) could not be easily determined from their available data and were estimated from accessory pigment- to-chlorophyll (ide) a ratios determined by HPLC for the same clone of P. •ricornu•um (Culture Collection of Marine Phytoplankton, Bigelow Laboratory for Ocean Sciences, West Boothbay Harbor, Maine). Since the chlorophyllase activity of diatoms is activated by harvesting and/or extraction techniques [Barrett and Jeffrey, 1964; Suzuki and Fuj ira 1986; Jeffrey and Hallegraeff, 1987] , it was assumed that the chlorophyllide a present in the P. •ricornutum extracts was
originally in the form of its parent compound, chlorophyll a. For this reason, accessory pigments were normalized to chlorophyll(ide) a (i.e. , chlorophyll a equivalents calculated as [chlorophyll a] + 1.45 [ chlorophyllide a] ). The HPLC determined accessory pigment-to- chlorophyll(ide) a ratios (w:w) were in good agreement with previously published values [Abaychi and Riley, 1979] for this species of phytoplankton. The concentrations of nonphotosynthetic carotenoids (i.e., "other carotenoids" in Figure 2a) were calculated by summing the diadinoxanthin and •-carotene concentrations [cf. Mann and Myers, 1968] .
The specific absorption coefficients (400-700 nm) for chlorophyll b were calculated by subtracting the chlorophyll a contribution to the absorption spectrum published for the light harvesting chlorophyll a/b-protein complex [Thornber and Alberte, 1977], and normalizing to chlorophyll b concentration. The "total" absorption spectrum of this complex was partitioned into contributions by chlorophyll a and b (Figure 2b). The concentrations of chlorophyll a and b representated by this spectrum were calculated with the chlorophyll a-specific molar extinction coefficient (158 L mmo1-1 cm -1) and chlorophyll b:a ratio (circa 1:1) published for this complex [Kan and Thornber, 1976] . The absorption due to chlorophyll a was estimated from the spectral properties of the P700- chlorophyll a-protein complex [Thornber, 1969; Prezelin and Alberte, 1978]. After subtraction, the remaining absorption was
Bidigare et al.: Oceanic Primary Production Estimates 175
2.0
1.6
(D
• 1.2 ..{3
o
=• 0.8
0.4
0.0
A Phaeodactylum tricornutum a
400 430 460 490 520 550 580 610 640 670 700
Wavelength (nm)
b
0.0 , 400 430 460 490 520 550 580 610 640 670 700
Wavelength (nm)
Fig. 2. Deconvoluted absorption spectra for (a) Phaeodactylum tricornutum [Mann and Myers, 1968] and (b) chlorophyll a/b-protein complex [Prezelin and Alberte, 1978; Thornber and Alberte, 1977] .
normalized to chlorophyll b concentration to yield estimates of specific absorption for chlorophyll b.
Model computations
For use in equation (2), the absorption due to individual pigments (chlorophylis and carotenoids) was calculated by multiplying pigment concentrations (Ci) by their respective specific absorption coefficients (ai). Pigment-specific absorption coefficients are not implicit constants and can vary with
photoadaptative state, degree of "pigment packaging" within the cell, and the diffusive state of the water column. As a
first approximation, the factor (i.e., 2.3) for converting logz0 to in units was omitted to compensate for the circa twofold reduction in specific absorption produced by "packaging" of pigments in intact cells [cf. Haardt and Maske, 1987]. No attempt has been made to correct for the radiance distribution of the light field. For simplicity, pigments were classified into five major groups' chlorophyll a equivalents, chlorophyll b,
176 Bidigare et al.- Oceanic Primary Production Estimates
Chlorophyll Concentration Carotenoid Concentration (ng L -• ) (ng L -• )
0 1 O0 200 300 400 500 0 1 O0 200 300 400 0
20
40
60
100
10¾ / ,,o T f o III o
"ø1/ 1 200
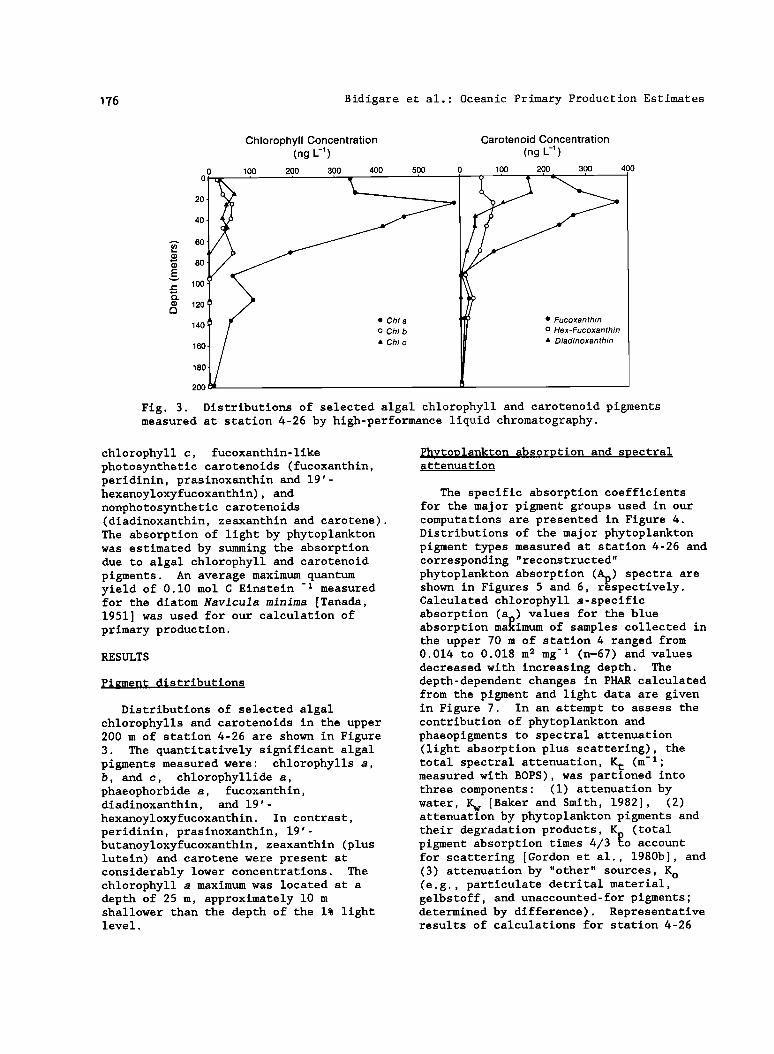
Fig. 3. Distributions of selected algal chlorophyll and carotenoid pigments measured at station 4-26 by high-performance liquid chromatography.
chlorophyll c, fucoxanthin-like photosynthetic carotenoids (fucoxanthin, peridinin, prasinoxanthin and 19'- hexanoyloxyfucoxanthin), and nonphotosynthetic carotenoids (diadinoxanthin, zeaxanthin and carotene). The absorption of light by phytoplankton was estimated by summing the absorption due to algal chlorophyll and carotenoid pigments. An average maximum quantum yield of 0.10 mol C Einstein -• measured for the diatom Navicula minima [Tanada, 1951] was used for our calculation of primary production.
RESULTS
Pigment distributions
Distributions of selected algal chlorophylis and carotenoids in the upper 200 m of station 4-26 are shown in Figure 3. The quantitatively significant algal pigments measured were: chlorophylis a, b, and c, chlorophyllide a, phaeophorbide a, fucoxanthin, diadinoxanthin, and 19'- hexanoyloxyfucoxanthin. In contrast, peridinin, prasinoxanthin, 19'- butanoyloxyfucoxanthin, zeaxanthin (plus lutein) and carotene were present at considerably lower concentrations. The chlorophyll a maximum was located at a depth of 25 m, approximately 10 m shallower than the depth of the 1% light level.
Phytoplankton absorption and spectral attenuation
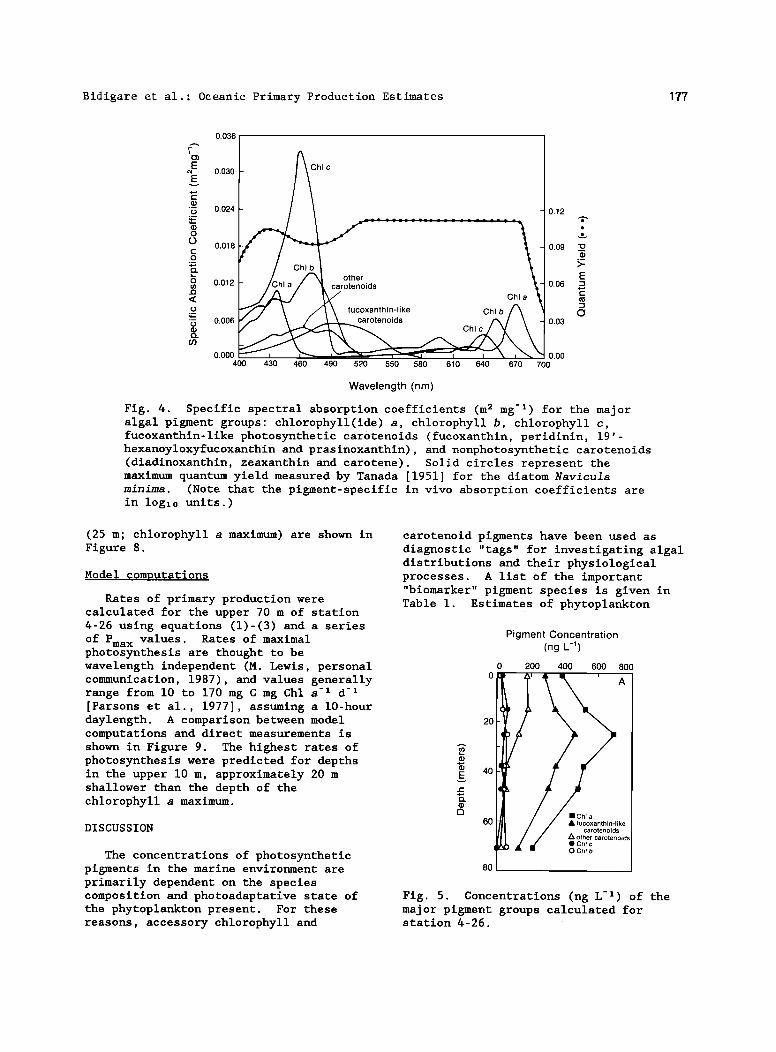
The specific absorption coefficients for the major pigment groups used in our computations are presented in Figure 4. Distributions of the major phytoplankton pigment types measured at station 4-26 and corresponding "reconstructed"
phytoplankton absorption (Ap) spectra are shown in Figures 5 and 6, respectively. Calculated chlorophyll a-specific
absorption (ap) values for the blue ß
absorption maximum of samples collected in the upper 70 m of station 4 ranged from 0.014 to 0.018 m 2 mg -• (n=67) and values decreased with increasing depth. The depth-dependent changes in PHAR calculated from the pigment and light data are given in Figure 7. In an attempt to assess the contribution of phytoplankton and phaeopigments to spectral attenuation (light absorption plus scattering), the total spectral attenuation, K t (m-•; measured with BOPS), was partioned into three components' (1) attenuation by water, K w [Baker and Smith, 1982], (2) attenuation by phytoplankton pigments and their degradation products, K p (total pigment absorption times 4/3to account for scattering [Gordon et al., 1980b], and (3) attenuation by "other" sources, K o (e.g., particulate detrital material, gelbstoff, and unaccounted-for pigments; determined by difference). Representative results of calculations for station 4-26
Bidigare et al.: Oceanic Primary Production Estimates 177
0.036
• 0.030 E
:_r,.] 0.024
O
(D 0.018 0
._
o o.o12
o 0.006
0.000
- 0.12 .;-.
ß
0.09 -o ._•
E 0.06
o 0.03
0.00 400 430 460 490 520 550 580 610 640 670 700
Wavelength (nm)
Fig. 4. Specific spectral absorption coefficients (m 2 mg -•) for the major algal pigment groups: chlorophyll(ide) a, chlorophyll b, chlorophyll c, fucoxanthin-like photosynthetic carotenoids (fucoxanthin, peridinin, 19'- hexanoyloxyfucoxanthin and prasinoxanthin), and nonphotosynthetic carotenoids (diadinoxanthin, zeaxanthin and carotene). Solid circles represent the maximum quantum yield measured by Tanada [1951] for the diatom Navicu2a mimima. (Note that the pigment-specific in vivo absorption coefficients are in log•0 units.)
(25 m; chlorophyll a maximum) are shown in Figure 8.
Model computations
Rates of primary production were calculated for the upper 70 m of station 4-26 using equations (1)-(3) and a series of Pmax values. Rates of maximal photosynthesis are thought to be wavelength independent (M. Lewis, personal communication, 1987), and values generally range from 10 to 170 mg C mg Chl a -• d -• [Parsons et al., 1977], assuming a 10-hour daylength. A comparison between model computations and direct measurements is shown in Figure 9. The highest rates of photosynthesis were predicted for depths in the upper 10 m, approximately 20 m shallower than the depth of the chlorophyll a maximum.
DISCUSSION
The concentrations of photosynthetic pigments in the marine environment are primarily dependent on the species composition and photoadaptative state of the phytoplankton present. For these reasons, accessory chlorophyll and
carotenoid pigments have been used as diagnostic "tags" for investigating algal distributions and their physiological processes. A list of the important "biomarker" pigment species is given in Table 1. Estimates of phytoplankton
Pigment Concentration (ng L -•)
0 200 400 600 800
0 , ' A
20
m 6ø I-Ill / / •, f-Uc'ø-xa nt h' n-h ke I III I / / caroteno•ds J J•l J / /' A other carot .... ds I
• • • eChlc J
T"'" ' ' OChlb I 80L I
Fig. 5. Concentrations (ng L -•) of the major pigment groups calculated for station 4- 26.
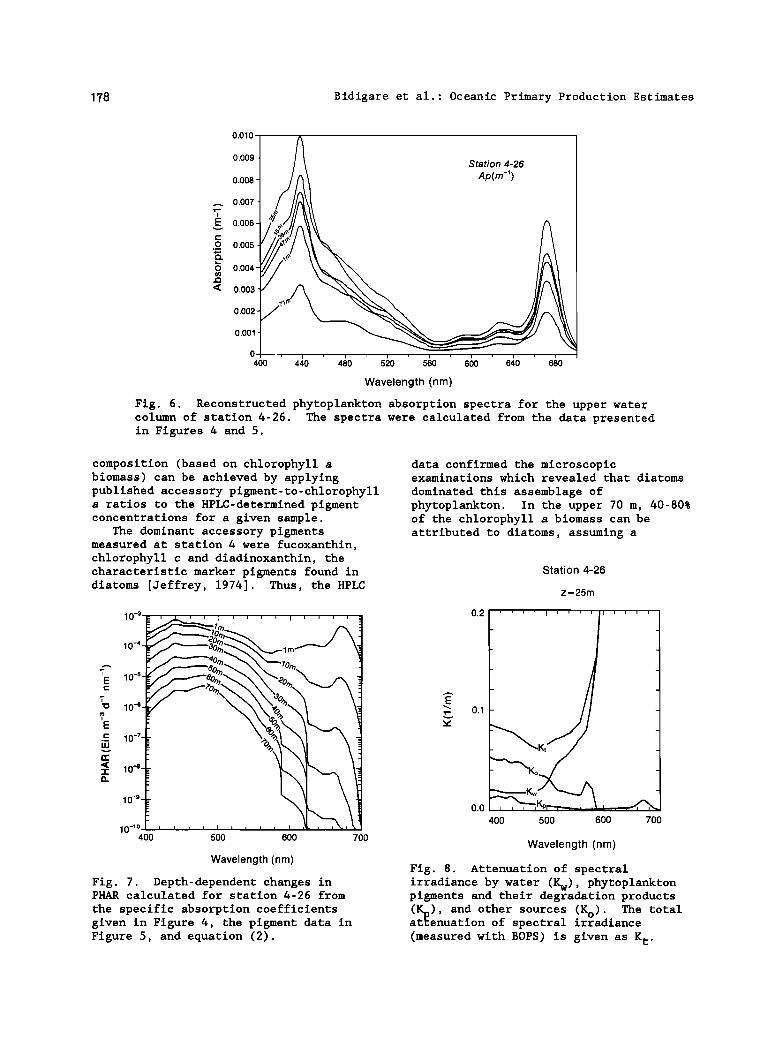
178 Bidigare et al.: Oceanic Primary Production Estimates
0.010 • ø'øø91 / / Station 4-26
o.oo, ] l/A\
0.004 •/ /
o.ooY
0.00 • • O• , ,
Wavelength (nm)
Fig. 6. Reconstructed phytoplankton absorption spectra for the upper water column of station 4-26. The spectra were calculated from the data presented in Figures 4 and 5.
composition (based on chlorophyll a biomass) can be achieved by applying published accessory pigment-to-chlorophyll a ratios to the HPLC-determined pigment concentrations for a given sample.
The dominant accessory pigments measured at station 4 were fucoxanthin, chlorophyll c and diadinoxanthin, the characteristic marker pigments found in diatoms [Jeffrey, 1974]. Thus, the HPLC
1 , , , , i , i i i :
10-4 ? lrn •
10'
10' 10-
10 -• 400 500 600 7()0
Wavelength (nm)
Fig. 7. Depth-dependent changes in PHAR calculated for station 4-26 from
the specific absorption coefficients given in Figure 4, the pigment data in Figure 5, and equation (2).
data confirmed the microscopic examinations which revealed that diatoms
dominated this assemblage of phytoplankton. In the upper 70 m, 40-80% of the chlorophyll a biomass can be attributed to diatoms, assuming a
Station 4-26
z :25m
0.2 , , , , , , , , ] , [ , i [
_
o.•
o.o
400 500 600 700
Wavelength (nm)
Fig. 8. Attenuation of spectral irradiance by water (Kw) , phytoplankton pigments and their degradation products
(Kp), and other sources (Ko). The total attenuation of spectral irradiance (measured with BOPS) is given as K t.
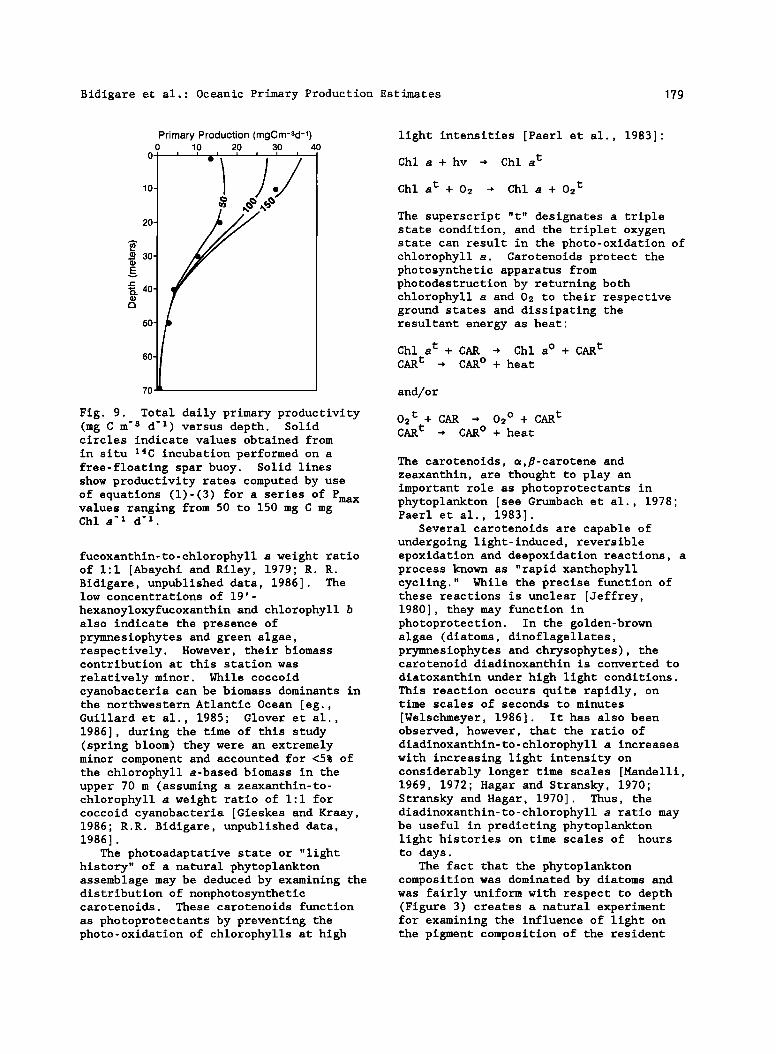
Bidigare et al.: Oceanic Primary Production Estimates 179
Primary Production (mgCm-3d -•) 0 10 20 30 40
0
10-
20-
v
50-
60-
7O
I I •1 I i I I
Fig. 9. Total daily primary productivity (mg C m -8 d -z) versus depth. Solid circles indicate values obtained from
in situ Z4C incubation performed on a free-floating spar buoy. Solid lines show productivity rates computed by use of equations (1)-(3) for a series of Pmax values ranging from 50 to 150 mg C mg Chl a -z d -z .
fucoxanthin-to-chlorophyll a weight ratio of 1:1 [Abaychi and Riley, 1979; R. R. Bidigare, unpublished data, 1986]. The low concentrations of 19'-
hexanoyloxyfucoxanthin and chlorophyll b also indicate the presence of prymnesiophytes and green algae, respectively. However, their biomass contribution at this station was
relatively minor. While coccoid cyanobacteria can be biomass dominants in the northwestern Atlantic Ocean [eg., Gulllard et al., 1985; Glover et al., 1986], during the time of this study (spring bloom) they were an extremely minor component and accounted for <5% of the chlorophyll a-based biomass in the upper 70 m (assuming a zeaxanthin-to- chlorophyll a weight ratio of 1:1 for coccoid cyanobacteria [Gieskes and Kraay, 1986; R.R. Bidigare, unpublished data, 1986].
The photoadaptative state or "light history" of a natural phytoplankton assemblage may be deduced by examining the distribution of nonphotosynthetic carotenoids. These carotenoids function
as photoprotectants by preventing the photo-oxidation of chlorophylis at high
light intensities [Paerl et al., 1983]'
Chl a + hv • Chl a t
Chl a t + 02 • Chl a + O2 t
The superscript "t" designates a triple state condition, and the triplet oxygen state can result in the photo-oxidation of chlorophyll a. Carotenoids protect the photosynthetic apparatus from photodestruction by returning both chlorophyll a and 02 to their respective ground states and dissipating the resultant energy as heat:
Chl a t + CAR • Chl a ø + CAR t CAR t • CAR ø + heat
and/or
02 t + CAR • 02 ø + CAR t CAR t w CAR ø + heat
The carotenoids, e,•-carotene and zeaxanthin, are thought to play an important role as photoprotectants in phytoplankton [see Grumbachet al., 1978; Paerl et al., 1983].
Several carotenoids are capable of undergoing light-induced, reversible epoxidation and deepoxidation reactions, a process known as "rapid xanthophyll cycling." While the precise function of these reactions is unclear [Jeffrey, 1980], they may function in photoprotection. In the golden-brown algae (diatoms, dinoflagellates, prymnesiophytes and chrysophytes), the carotenoid diadinoxanthin is converted to
diatoxanthin under high light conditions. This reaction occurs quite rapidly, on time scales of seconds to minutes
[Welschmeyer, 1986]. It has also been observed, however, that the ratio of diadinoxanthin-to-chlorophyll a increases with increasing light intensity on considerably longer time scales [Mandelli, 1969, 1972; Hagar and Stransky, 1970; Stransky and Hagar, 1970]. Thus, the diadinoxanthin-to-chlorophyll a ratio may be useful in predicting phytoplankton light histories on time scales of hours to days.
The fact that the phytoplankton composition was dominated by diatoms and was fairly uniform with respect to depth (Figure 3) creates a natural experiment for examining the influence of light on the pigment composition of the resident

180 Bidigare et al.: Oceanic Primary Production Estimates
Table 1. List of Important Phytoplankton Pigments Used as Diagnostic "Tags" and Their
Taxonomic/Phys iological Significance (But- fucoxanthin -- 19' -Butanoyloxyfucoxanthin and Hex-
fucoxanthin • 19' -Hexanoyloxyfucoxanthin)
Pigment Significance Reference
Chlorophyll a Chlorophyllide a Chlorophyllase-
containing diatoms Chlorophyll b Green algae Chlorophyll c Diatoms and/or
Fucoxanthin chrysophytes Peridinin Dinoflagellates But-fucoxanthin Chrysophytes
Hex- fucoxanthin
Hex- fucoxanthin
Hex-fucoxanthin
Pras inoxanthin
Zeaxanthin
Alloxanthin
Zeaxanthin
e,•-carotene
Diadinoxanthin
Phaeophorbide a
Phytoplankton biomass Jeffrey [1980] Jeffrey [1974]
Jeffrey [1974] Jeffrey [1974]
Jeffrey [1974] Liaaen-Jensen
[19853 Prymnesiophytes Gieskes and Kraay
[1986] Emiliania huxleyi Norgard et al.
[•974] Gyrodinium aureolum Tangen and
Bjornland [1981]
Prasinophytes Foss et al. [1984]
Cyanobacteria Gieskes and Kraay [1983a]
Cryptophytes Gieskes and Kraay [1983b]
Photoprotectant Paerl et al. [1983]
Photoprotectants Paerl et al. [1983]
Photoprotectant (?) Jeffrey [1980] Zooplankton grazing Jeffrey [1974]
phytoplankton. A comparison of the photosynthetic and nonphotosynthetic carotenoid to chlorophyll a equivalence ratios (w:w) for the upper water column of station 4 is given in Table 2. The samples were collected at 1-2 hours prior to local apparent noon on April 5 (station 4-26; sunny) and April 7 (station 4-45; overcast), respectively. While the photosynthetic carotenoid-to-chl a ratios were relatively constant with respect to depth and irradiance conditions (circa 0.60-0.70; w:w), the nonphotosynthetic carotenoid-to-chl a ratios showed two
distinct distributional patterns. On the sunny day (station 4-26), the ratios decreased monotonically from 0.45 at the surface to a value of 0.11 at depth. In contrast, on the overcast day (station 4- 45), the ratios were uniformly low (circa
0.11) and agreed well with ratios measured deep in the water column on the sunny day. These data strongly suggest that these nonphotosynthetic carotenoids may have a photoprotective function. Laboratory studies performed with diatom and green algal clones have shown that nonphotosynthetic carotenoids are labeled with a higher degree of specific radioactivity than are the photosynthetic accessory pigments [e.g., Anderson et al., 1960; Grumbach et al., 1978; Goericke and Welschmeyer, 1987]. The field measurements presented here support the results of the algal labeling experiments by demonstrating that the nonphotosynthetic carotenoids comprise a very dynamic pigment pool. These data also suggest that "photoprotective" adaptations by phytoplankton sampled

Bidigare et al.- Oceanic Primary Production Estimates 181
Table 2. Comparison of Photosynthetic and Nonphotosynthetic Carotenoid to
Chlorophyll a equivalence ratios (PC/Chl a and NPC/Chl a, respectively)
for Samples Collected in the Upper Water Column of Station 4
Depth PC/Chl a NPC/Chl a (w'w) (w'w)
Station 4-26 (Sunny)
1 m 0.72 0.45 15 m 0.65 0.35 25 m 0.66 0.18 38 m 0.67 0.10 47 m 0.64 0.10 71 m 0.62 0.11
Station 4-45 (Overcast)
1 m 0.71 0.14 15 m 0.68 0.11 34 m 0.65 0.11 61 m 0.67 0.10 81 m 0.67 0.12
during springtime in the Sargasso Sea occur on shorter time scales then does
vertical mixing. The estimated in vivo specific
absorption coefficients for chlorophyll a are in reasonable agreement with the values determined by Thornber [1969] for the chlorophyll a-protein (Chl-P) complex isolated from the blue-green alga, Phormidium luridum. The values reported here are 23 and 28% higher than those measured at the blue and red absorption maxima of this Chl-P complex, respectively. Calculated chlorophyll a-
specific absorption values g• ) for station 4 are consistent wi Pthose measured for diatom-rich waters (0.018- 0.022 m 2 mg TM [Prieur and Sathyendranath, 1981; Kiefer and SooHoo, 1982]) and laboratory cultures of diatoms (0.014- 0.027 m 2 mg TM [Dubinsky et al., 1986; Maske and Haardt, 1987; Mitchell, 1987]). In comparison, the theoretical estimates
of ap (435 nm) for large particles (16-32 •m) range from 0.015 to 0.028 m • mg -• [Morel and Bricaud, 1981]. Blue/red absorption band ratios (A4ss/A67•) calculated from the phytoplankton
absorption spectra reconstructed for station 4-26 ranged from 1.6 to 1.8. These ratios are well within the range of values (1.2-2.1) reported for diatoms grown under a variety of light conditions [SooHoo et al., 1986; Maske and Haardt, 1987; Mitchell, 1987].
A Pmax value of 100 mg C mg Chl a -• d -• yielded a good linear relationship between measured and calculated rates of primary production at all depths except the surface (Ymeas = l'02Xcalc - 0.54; r • = 0.945; n = 6). The discrepancy observed near the surface may in part be attributed to photorespiration and/or photoinhibition processes occurring in the fixed-depth bottle incubations. Alternatively, the fact that the computations for in situ quantum yield (equation (3)) were carried out using an average daily irradiance may have led to an overestimation of predicted production rates near the surface [cf. Platt et al., 1984]. Hogetsu et al. [1959] reported Pmax values of 9-17 mg C mg Chl a -• hr -1 for a summertime bloom of Skeletonema costaturn in Tokyo Bay. These values translate into daily values of 90- 170 mg C mg Chl a -• d -• (assuming 10 hours of daylight per day) and are consistent with the Pmax value of 100 mg C mg Chl a -• d TM used in the model presented here. The agreement between computed and directly measured productivity suggest that all the major pigment groups, except the photoprotective carotenoids, transfer their absorbed energy to photosynthesis reaction centers as effectively as does chlorophyll a.
These results are significant for several reasons. First, the approach described here is not biased by the presence of nonliving particulate matter, and our computation of PHAR corrects for depth-dependent variations in pigment composition resulting from photoadaptation and/or changes in phytoplankton composition. In general, it appears that the phytoplankton sampled at station 4 were responsible for a very small fraction of the light attenuation in the upper water column (Figure 8). This observation is consistent with measurements made in
other geographic areas which have shown that detrital particulates provide a large and variable contribution to light absorption in the water column [Yentsch, 1962; Kiefer and SooHoo, 1982; Kirk, 1983; Lewis et al., 1985a, b; Kishino et al., 1984; Kishino et al., 1986; Maske and Haardt, 1987; Mitchell, 1987]. In

182 Bidigare et al.: Oceanic Primary Production Estimates
general, these data suggest that the open ocean is very inefficient in light absorption and the presence of detrital material may be an important factor which limits rates of primary production there. This observation also has major implications regarding our ability to estimate phytoplankton absorption using satellite imagery.
Second, the fact that primary production can be modeled from the absorption characteristics of the phytoplankton represents a considerable advance in the ability to measure primary production in the ocean. Although there are no apparent problems with the incubation methodology used in this study for directly measuring primary productivity, incubations of any kind mean costly and time-consuming efforts on the part of oceanographers. Obtaining an es•timate of primary production from the knowledge of scalar quantities, as we have done here, permits enormous improvement in sampling coverage, which in turn allows expanded study of the environmental influences on phytoplankton production in the sea. To estimate primary productivity using the approach described here requires knowledge of the major pigment groups, their concentrations and specific absorption coefficients, spectral irradiance (quanta) as a function of depth and time of day, and an appropriate model describing the depth (irradiance) variability of quantum yield. We are now in the process of expanding our library of specific absorption coefficients to include the phycobilins which will permit the application of this model in cyanobac ter ia- dominated waters. In addition, we are examining in greater detail the influence of pigment packaging and radiance distribution on the specific absorption coefficients reported here. For routine application of this approach for shipboard estimates, only a single BOPS cast (circa 30 min) is required to acquire pigment samples and spectral attenuation data, and total radiant energy can be measured throughout the day using a shipboard sensor. Hence, the ship would be free to depart the area and perform additional casts to examine the spatial variability of primary productivity. This has a distinct advantage for work conducted in those oceanic regions which are highly variable (eg., frontal regions, and warm and cold core rings).
Third, the results show that it may be
possible to estimate primary production from an untended mooring. It should be possible to collect spectral irradiance data as a function of depth from moored sensors. If these data can be used to
estimate pigment data, for example, by means' of a bio-optical model [Smith and Baker, 1978; Baker and Smith, 1982], then the necessary parameters to estimate primary production as a function of depth can be obtained from an untended mooring.
Finally, these are among very few in situ determinations of photosynthetically absorbed radiation as a function of
wavelength for natural phytoplankton populations. These complete spectral data provide insight into how phytoplankton utilize radiant energy as a function of depth, how phytoplankton influence the spectral characteristics of optical properties, and how various populations of autotrophs are partitioned as a function of space and time.
Acknowledgments. This work was supported by the Office of Naval Research through the Biowatt Program (R.R.B. contract N00014-80-C-0113; R.C.S. and K.S.B. contract N00014-84-C-0382; and J.M. contract N00014-84-C-0132IIA). We thank the officers and crew of the R/V Knorr and our colleagues in the Biowatt program for their assistance at sea in obtaining the data and for helpful discussions. The bio- optical profiling system was supported by NASA grant NAWG-290-3 (R.C.S.). Valuable discussions regarding model formulation were provided by Marlon R. Lewis and Dale A. Kiefer. Biowatt contribution No. 02.
REFERENCES
Abaychi, J.K., and J.P. Riley, The determination of phytoplankton pigments by high-performance liquid chromatography, Anal. Chim. Acta, 107, 1-11, 1979.
Anderson, 3.M., U. Blass, and M. Calvin, Biosynthesis and possible relations among the carotenoids and between chlorophyll a and b, in Comparitive Biochemistry of Photoreactive Systems, edited by M. B. Allen, pp. 215-225, Academic, New York, 1960.
Baker, K.S., and R.C. Smith, Bio- optical classification and model of natural waters, 2, Limnol. Oceanogr., 27, 500- 509, 1982.
Barrett, 3., and S.W. 3effrey,

Bidigare et al.: Oceanic Primary Production Estimates 18]
Chlorophyllase and formation of an atypical chlorophyllide in marine algae, Plant Physiol . , 39, 44-47, 1964.
Bidigare, R.R., J.D. Guffy, and J.M. Brooks, HPLC pigment investigations in the northwestern Atlantic Ocean (project BIOWATT) (abstract), Trans. AGU, 66, 1312- 1313, 1985.
Brown, O., R.H. Evans, H.R. Gordon, R.C. Smith, and K.S. Baker, Blooming off the U.S. coast: A satellite description, Science, 229, 163-167, 1985.
Dickey, T., D. Siegel, J. Marra, and M. Hamilton, Variability of physical and optical properties in the Sargasso Sea during the Biowatt I experiment, Trans. AGU, 67, 968, 1986.
Dubinsky, Z., P.G. Falkowski, and K. Wyman, Light harvesting and utilization by phytoplankton, Plant Cell Physiol., 27, 1335- 1349, 1986.
Eppley, R.W., E. Stewart, M.R. Abbott, and U. Heyman, Estimating ocean primary production from satellite chlorophyll: Introduction to regional differences and statistics for the
southern California bight, J. Plankton Res., ?, 57-70, 1985.
Falkowski, P.G., Z. Dub insky, and K. Wyman, Growth- irradiance relationships in phytoplankton, Limnol. Oceanogr., 30, 311- 321, 1985.
Fitzwater, S.E., G.A. Knauer, and 3.H. Martin, Metal contamination and its effect on primary production measurements, Limnol. Oceanogr. , 27, 544- 551, 1982.
Foss, P., R.R.L. Gulllard, and S. Liaaen-3ensen, Pras inoxanthin- -a chemosystematic marker for algae, Phytochemistry. , 23, 1629-1633, 1984.
Gieskes, W.W.C., and G.W. Kraay, Unknown chlorophyll a derivatives in the North Sea and tropical Atlantic Ocean revealed by HPLC analysis, Limnol. Oceanogr., 28, 757-766, 1983a.
Gieskes, W.W.C., and G.W. Kraay, Dominance of Cryptophyceae during the phytoplankton spring bloom in the central North Sea detected by HPLC analysis of pigments, Mar. Biol., 75, 179-185, 1983b.
Gieskes, W.W.C., and G.W. Kraay, Floristic and physiological differences between the shallow
and the deep nanoplankton community in the euphotic zone of the open tropical Atlantic revealed by HPLC analysis of pigments, Mar. Biol . , 91, 567-576,1986.
Glover, H.E. , L. Campbell, and B.B. Prezelin, Contribution of Synechococcus spp. to size- fractioned primary productivity in three water masses in the
northwest Atlantic Ocean, Mar. Biol., 91, 193-203, 1986.
Goericke, R. , and N. Welschmeyer, Pigment labeling and measurements of group-specific algal growth rates' Prospects and limitations, (abstract), in Abstracts of Papers for the 1987 Annual Meeting, American Society of Limnology and Oceanography, 1987.
Gordon, H.R., and W.R. McCluney, Estimation of the depth of sunlight penetration in the sea from remote sensing, Appl. Opt., 14, 413-416, 1975.
Gordon, H.R., D. Clark, 3.L Mueller, and W.A. Hovis, Phytoplankton pigments from Nimbus-7 coastal zone color scanner' Comparison with surface measurements, Science, 210, 63-66, 1980a.
Gordon, H.R., R. C. Smith, and 3.R.V. Zaneveld, Introduction to ocean optics, Proc. SPIE Ocean Opt. VI , 208, 14- 55, 1980b.
Crumbach, K.H., H.K. Lichtenthaler, and K.H. Erismann, Incorporation of 14CO 2 in photosynthetic pigments of Chlorella pyrenoidosa, Planta, 140, 37-43, 1978.
Gulllard, R.R.L., L.S. Murphy, P. Foss, and S. Liaaen-3ensen, Synechococcus spp. as likely z eaxanthin- dominant
ultraphytoplankton in the North Atlantic, Limnol. Oceanogr., 30, 412-414, 1985.
Haardt, H., and H. Maske, Specific in vivo absorption coefficient of chlorophyll a at 675 nm, Limnol. Oceanogr. , 32, 608-619, 1987.
Hagar, A. , and H. Stransky, Das Carotinoidmuster und die
Verbeitung des lichtinduzierten Xanthophyllcyclus in verschiedenen Algenklassen, Arch. Mikrobiol., ?3, 77-89, 1970.

184 Bidigare et al.: Oceanic Primary Production Estimates
Hogetsu, K., M. Sakamoto, and H. Sumikawa, On the high photosynthetic activity of Skeletonema costaturn under strong light intensity, Bot. MaE. , 72, 421-422, 1959.
Hovis, W.A., D.K. Clark, F. Anderson, R.W. Austin, W.H. Wilson, E.T. Baker, D. Ball, H.R. Gordon, J.L. Mueller, S.Z. E1- Sayed, B. Strun, R.C. Wrigley, and C.S. Yentsch, Nimbus-7 coastal zone color scanner: System description and initial imagery, Science, 210, 60-63, 1980.
Jeffrey, S.W., Profiles of photosynthetic pigments in the ocean using thin- layer chromatography, Mar. Biol., 26, 101-110, 1974.
Jeffrey, S.W., Algal pigment systems, in Primary Productivity in the Sea, edited by P.G. Falkowski, pp. 33-57, Plenum, New York, 1980.
3effrey, S.W., and G.M. Hallegraeff, Chlorophyllase distribution in 10 classes of phytoplankton: A problem for chlorophyll analysis, Mar. Ecol. ProEr. Ser., 35, 293- 304, 1987.
3effrey, S.W., and G.F. Humphery, New spectrophotometric equations for determining chlorophylis a, b, cl and c•_ in higher plants, algae, and natural phytoplankton, Blochem. Physiol. Pflanz, 167, 191-194, 1975.
Kan, K., and J.P. Thornher, The light-harvesting chlorophyll a/b complex of Chlamydomonas reinhardii, Plant Physiol., 57, 47-52, 1976.
Kiefer, D.A., and B.G. Mitchell, A simple, steady state description of phytoplankton growth base on absorption cross section and quantum efficiency, Limnol. OceanoEr. , 28, 770-776, 1983.
Kiefer, D.A., and J.B. SooHoo, Spectral absorption by marine particles of coastal waters of Baja California, Limnol. OceanoEr., 27, 492-499, 1982.
Kirk, J.T.O., LiEht and Photosynthesis in Aquatic Ecosystems, Cambridge University Press, New York, 1983.
Kishino, M., C.R. Booth, and N. Okami, Underwater radiant energy
absorbed by phytoplankton, detritus, dissolved organic matter, and pure water, Limnol. Oceanogr., 29, 340-349, 1984.
Kishino, M., N. Okami, M. Takahashi, and S. Ichimura, Light utilization efficiency and quantum yield of phytoplankton in a thermally stratified sea, Limnol. OceanoEr. , 31, 557-566, 1986.
Lewis, M.R., R.E. Warhock, B. Irwin, and T. Platt, Measuring photosynthetic action spectra of natural phytoplankton populations, J. Phycol., 21, 310-315, 1985a.
Lewis, M.R., R.E. Warhock, and T. Platt, Absorption and photosynthetic action spectra for natural phytoplanktons: Implications for production in the open ocean, Limnol. OceanoEr. , 30, 794-806, 1985b.
Liaaen-Jensen, S., Carotenoids of lower plants--Recent progress, Pure Appl. Chem., 57, 649-658, 1985.
Mandelli, E.F., Carotenoid conversion in light-dark cultures of the dinoflagellate Amphidinium klebsii, J. Phycol., 5, 382-384, 1969.
Mandelli, E.F., The effect of growth illumination on the pigmentation of a marine dinoflagellate, J. Phycol., 8, 367-369, 1972.
Mann, J.E., and J. Myers, On pigments, growth, and photosynthesis of Phaeodactylum tricornutum, J. Phycol., 4, 349- 355, 1968.
Marra, J., and E. Hartwig, Biowatt: A study of bioluminescence and optical variability in the sea, Trans. AGU, 65, 732-733, 1984.
Marra, J., and K. Heinemann, A comparison between noncontaminating and conventional incubation procedures in primary production measurements, Limnol. OceanoEr., 29, 389-392, 1984.
Maske, H., and H. Haardt, Quantitative in vivo absorption spectra of phytoplankton: Detrital absorption and comparison with fluorescence excitation
spectra, Limnol. OceanoEr. , 32, 620-633, 1987.
McClain, C.R., L.J. Pietrafesa, and J.A. Yoder, Observations of gulf stream induced and wind driven

Bidigare et al.: Oceanic Primary Production Estimates 185
upwelling in the Georgia bight using ocean color and infrared imagery, J. Geophys. Res., 89, 3705-3723, 1984.
Mitchell, B.G., Ecological implications of variability in marine particulate absorption and fluorescence excitation spectra, Ph.D. dissertation, Department of Biological Sciences, Univ. of S. Calif. Los Angeles, 1987.
Morel, A., Available, useable, and stored radiant energy in relation to marine photosynthesis, Deep Sea Res., 25, 673-688, 1978.
Morel, A., and A. Bricaud, Theoretical results concerning light absorption in a descrete medium, and application to specific absorption of phytoplankton, Deep Sea Res., 28, 1375-1393, 1981.
Morel, A., and R.C. Smith, Relation between total quanta and total energy for aquatic photosynthesis, Limnol. Oceanogr., 19, 519-600, 1974.
National Academy of Sciences, Global Ocean Flux Study, Washington, D.C., 1984.
Norgard, S., W.A. Svec, and S. Liaaen-3ensen, Algal carotenoids and chemotaxonomy, Blochem. Syst. Ecol., 2, 7-9, 1974.
Paerl, H.W., 3. Tucker, and P. Bland, Carotenoid enhancement and its role in maintaining blue-green algal (Microcystis aeruginosa) surface blooms, Limnol. Oceanogr., 28, 847-857, 1983.
Parsons, T.R., M. Takahashi, and B. Hargrave, Biological Oceanographic Processes, Pergamon, New York, 1977.
Platt, T., Primary production of the ocean water column as a function
of surface light intensity: Algorithms for remote sensing, Deep Sea Res., 33, 149-163, 1986.
Platt, T., M. Lewis, and R. Geider, Thermodynamics of the pelagic ecosystem: Elementary closure conditions for biological production in the open ocean, in Flows of Energy and Materials in Marine Ecosystems, edited by 3.R. Fasham, pp. 49-84, Plenum, New York, 1984.
Prezelin, B.B., and R.S. Alberte, Photosynthetic characteristics and
organization of chlorophyll in marine dinoflagellates, Proc. Natl. Acad. Sci. U.S.A., ?5, 1801- 1804, 1978.
Prieur, L., and S. Sathyendranath, An optical classification of coastal and oceanic waters based
on the specific spectral absorption curves of phytoplankton pigments, dissolved organic matter, and other particulate materials, Limnol. Oceanogr., 26, 671-689, 1981.
Sathyendranath, S., L. Lazzara, and L. Prieur, Variations in the spectral values of specific absorption of phytoplankton, Limnol. Oceanogr., 32, 403-415, 1987.
Shannon, L.V., S.A. Mosterr, N.M. Walters, and F.P. Anderson, Chlorophyll concentrations in the southern Benguel current region as determined by satellite (Nimbus-7 CZCS), 3. Plankton Res., 5, 565- 584, 1983.
Smith, R.C., Remote sensing and the depth distribution of ocean chlorophyll, Mar. EcoI., 5, 359- 361, 1981.
Smith, R.C., and K.S. Baker, Optical classification of natural waters, Limnol. Oceanogr., 23, 260-267, 1978.
Smith, R.C., and K.S. Baker, Oceanic chlorophyll concentrations determined by satellite (Nimbus-7 coastal zone color scanner), Mar. BioI., 66, 269-279, 1982.
Smith, R.C., and K.S. Baker, The analysis of ocean optical data, Proc. SPIE Ocean Opt. VII, 489, 119-125, 1984.
Smith, R.C., and K.S. Baker, The analysis of ocean optical data II, Proc. SPIE Ocean Opt. VII, 63?, 95-107, 1986.
Smith, R.C., K.S. Baker, and P. Dustan, Fluorometric techniques for measurement of oceanic
chlorophyll in the support of remote sensing, Ref. 81-17, 14 PP., Scripps Inst. of Oceanogr., La 3olla, Calif., 1981.
Smith, R.C., R.W. Eppley, and K.S. Baker, Correlation of primary production as measured aboard ship in southern California coastal waters and as estimated from
satellite chlorophyll images, Mar. Biol., 66, 281-288, 1982.

186 Bidigare et al.: Oceanic Primary Production Estimates
Smith, R.C., C.R. Booth, and J.L. Star, Oceanographic bio-optical profiling system, Appl. Opt., œ$, 2791-2797, 1984.
Smith, R.C., O. Brown, F. Hoge, K.S. Baker, R.H. Evans, R.H. Swift, and W.E. Esaias, Multiplatform sampling (ship, aircraft and satellite) of a gulf stream warm core ring, App1. Opt., 26, 2068- 2081, 1987a.
Smith, R.C., R.R. Bidigare, B.B. Prezelin, K.S. Baker, and 3.M. Brooks, Optical characterization of primary productivity across a coastal front, Mar. Biol., in press, 1987b.
SooHoo, 3.B., D.A. Kiefer, D.3. Collins, and I.S. McDermid, In vivo fluorescence and absorption spectra of marine phytoplankton, I, Taxonomic characteristics and responses to photoadaptation, J. Plankton Res., 8, 197-214, 1986.
Stransky, H., and A. Hagar, Das Carotinoidmuster und die
Verbeitung des lichtinduzierten Xanthophyllcyclus in verschiedenen Algenklassen, II, Arch. Mikrobiol., 71, 164-190, 1970.
Strickland, 3.D.H., and T.R. Parsons, Practical Handbook for Seawater Analysis, Vol. 167, Fisheries Research Board of
Canada, Ottawa, 1969. Suzuki, R. , and Y. Fujita,
Chlorophyll decomposition in Skeletonema costaturn: A problem in chlorophyll determination in water samples, Mar. Ecol. Progr. Ser., 28, 81-85, 1986.
Tanada, T., The photosynthetic efficiency of carotenoid pigments in Navicula minima, Am. J. Bot., 38, 276- 283, 1951.
Tangen, K., and T. Bjornland, Observations on pigments and morphology of Gyrodinium aureolum Hulburt, a marine dinoflagellate containing 19' - hexanoyloxyfucoxanthin as the main carotenoid, J. Plankton Res., 3, 389-401, 1981.
Thornher, 3.P., Comparison of a chlorophyll a-protein complex isolated from a blue-green alga with chlorophyll-protein complexes obtained from green bacteria and
higher plants, Blochim. Biophys. Acta, 172, 230-241, 1969.
Thornher, 3.P., and R.S. Alberte, The organization of chlorophyll in vivo, in Photosynthesis I, edited by A. Trebst, and M. Avron, pp. 574•582, Springer-Verlag, New York, 1977.
Trees, C.C., M.C. Kennicutt, and 3.M. Brooks, Errors associated with the standard fluorimetric
determination of chlorophylis and phaeopigments, Mar. Chem., 17, 1- 12, 1985.
Trees, C.C., R.R. Bidigare, and 3.M. Brooks, Distribution of chlorophylis and phaeopigments in the northwestern Atlantic Ocean, J. Plankton Res., 8, 447-458, 1986.
Tyler, 3.E., The in situ quantum efficiency of natural phytoplankton populations, Limnol. Oceanogr, 18, 442-445, 1975.
Vernet, M., and C.3. Lorenzen, The presence of chlorophyll b and the estimation of phaeopigments in marine phytoplankton, J. Plankton Res., 9, 255-265, 1987.
Welschmeyer, N.A., Rapid xanthophyll cycling: An in situ tracer for mixing in the upper ocean, Trans. AGU, 67, 969, 1986.
Yentsch, C.S., Measurement of visible light absorption by particulate matter in the ocean, Limnol. Oceanogr., 9, 207- 217, 1962.
K.S. Baker, University of California Marine Bio-Optics Group, Scripps Institution of Oceanography, University of California, San Diego, La Jolla, CA 92037.
R.R. Bidigare, Geochemical and Environmental Research Group, Texas A&M University, College Station, TX 77843.
J. Marra, Lamont-Doherty Geological Observatory of Columbia University, Palisades, NY 10027.
R.C. Smith, University of California Marine Bio-Optics Group, Department of Geography, University of California, Santa Barbara, Santa Barbara, CA 93106.
(Received June 18, 1987; revised August 31, 1987; accepted August 31, 1987.)





























