Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Effects of the Nesting Environment on Embryonic Development, Sex Ratio, andHatching Success in Podocnemis unifilis (Testudines: Podocnemididae) in an Areaof Várzea Floodplain on the Lower Amazon River in BrazilAuthor(s): Marina T. Pignati , Luana F. Fernandes , Priscila S. Miorando , Paulo D. Ferreira , and JuarezC. B. PezzutiSource: Copeia, 2013(2):303-311. 2013.Published By: The American Society of Ichthyologists and HerpetologistsDOI: http://dx.doi.org/10.1643/CE-11-146URL: http://www.bioone.org/doi/full/10.1643/CE-11-146
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

Effects of the Nesting Environment on Embryonic Development, Sex Ratio,
and Hatching Success in Podocnemis unifilis (Testudines: Podocnemididae)
in an Area of Varzea Floodplain on the Lower Amazon River in Brazil
Marina T. Pignati1, Luana F. Fernandes2, Priscila S. Miorando3,Paulo D. Ferreira4, and Juarez C. B. Pezzuti5
The nesting site selected by a female turtle influences the temperature and humidity of the incubating eggs, whichaffects the development, survival, and sex of the embryo and the subsequent developmental phases. The present studyevaluated the effects of environmental variables on the duration of incubation, hatching success, and sex ratio ofhatchlings of Podocnemis unifilis in an area of varzea swamp on the lower Amazon River in Brazil, during the 2007 and2009 nesting seasons. Nests were located, marked, and monitored, and physical and environmental characteristics weremeasured. The temperature of the nests was measured, and hatchlings were collected to determine the sex ratio. Themean temperature, vegetation cover, and nesting date influenced the duration of the incubation period, and the grainsize of the substrate influenced hatchling survival. The sex ratio differed between the years and was female-biased in2007 and male-biased in 2009, emphasizing the need for further long-term studies of the reproductive cycle inchelonians.
THE selection of nesting sites has been analyzed in anumber of reptilian species, chelonians in particular(Caut et al., 2006). In chelonians, the characteristics
of the habitat where the nest is located and the microhabitatwithin the nest are related, and have specific consequencesfor the development of the embryo (Weisrock and Janzen,2000). In the majority of turtle species, site choice by thefemale will influence incubation temperatures, which will,in turn, determine the sex of the embryos (Mrosovsky, 1980)and the timing of the later stage of development (McKnightand Gutzke, 1993), which in turn may affect post-hatchingsurvival and thermoregulation of juveniles and adults (Rhenet al., 1999). Understanding the thermal conditions ofnatural nests is essential for the analysis of the influence ofnesting-site choice on the reproductive success of cheloni-ans (Doody et al., 2003).
The discovery of the effect of incubation temperatures onsex differentiation in chelonians (Yntema, 1976) highlight-ed the importance of the nesting site. Because the femalesignore their nests after oviposition, embryonic developmentand reproductive success are entirely dependent on theenvironmental conditions to which the eggs are exposed(Bujes and Verrastro, 2009). The nest site must provideincubation temperatures within an appropriate range forboth the survival of the hatchlings and development of a sexratio compatible with the population structure (FerreiraJunior, 2009). Incubation conditions will vary according tothe mineral composition and granulometry of the substrate,humidity, depth of the nest, ambient temperatures, andvegetation cover (Janzen, 1994a; Souza and Vogt, 1994;Ferreira Junior and Castro, 2003). These factors will in turndetermine a number of different developmental parameters,such as sex ratio, incubation duration, and hatching success.
The Yellow-spotted River Turtle, Podocnemis unifilisTroschel 1848, has temperature-dependent sex determina-
tion and a high pivotal temperature of near 32uC (Souza andVogt, 1994), which is typical for neotropical podocnemidids(Alho et al., 1985; Paez et al., 2009; Lubiana and FerreiraJunior, 2009). The temperature transition zone for thegeneration of males and females in different proportions(Pieau, 1996) is wide (Gomes, 2011), and this allows thefemales to use riverbanks with different granulometric andtopographic characteristics and vegetation cover for nesting(Escalona and Fa, 1998; Fachın-Teran and Von Mulhen,2003; Ferreira Junior and Castro, 2003). In Brazil, P. unifilis isfound in the Amazon and Tocantins-Araguaia basins, a vastarea with varying climatic characteristics, which havedifferent effects on local nesting conditions.
Knowledge of the proportion of males and females isessential in studies of the life history of species, and isimportant in chelonians (Congdon et al., 2000) due to thedifficulties of monitoring and the absence of sexualdimorphism in hatchlings and juveniles, which makes itimportant to estimate the sex ratio at hatching. Informationon the initial stages of life is important, because it serves as abasis to evaluate the population in the different life stages,especially for species that, like P. unifilis, are subject tointense predation of adult females during nesting (Vanzo-lini, 1967; Pritchard, 1979; Alho and Padua, 1982; Pritchardand Trebbau, 1984). In addition to sex, estimates ofhatching success are essential for studies on the populationdemographics. Since the predation on and flooding of nestsof P. unifilis may have drastic effects, the nesting time andplace will play important roles in the reproductive success offemales. Since P. unifilis utilizes nesting sites with a widerange of environmental conditions, the variables associatedwith the time and place of nesting should influence thetemperature and hatching success, affecting the sex andsurvival of the hatchlings. Because the weather conditions(temperature, rainfall, and flooding of nests) vary between
1 Post-graduate Program in Zoology, Universidade Federal do Para e Museu Paraense Emılio Goeldi/CZO, Av. Perimetral 1901, Terra Firme,Belem, Para 66077-530, Brazil; E-mail: [email protected]. Send reprint requests to this address.
2 Secretaria Municipal de Meio Ambiente de Muana, Muana, Para 68825-000, Brazil; E-mail: [email protected] Post-graduate Program in Aquatic Ecology and Fisheries/PPGEAP, Universidade Federal do Para, Belem, Para 66075-650, Brazil; E-mail:
[email protected] Universidade Vila Velha, Vila Velha, Espırito Santo 29102-770, Brazil; E-mail: [email protected] Nucleo de Altos Estudos Amazonicos (NAEA), Universidade Federal do Para, Belem, Para 66075-650, Brazil; E-mail: [email protected]: 7 October 2011. Accepted: 3 December 2012. Associate Editor: J. F. Schaefer.F 2013 by the American Society of Ichthyologists and Herpetologists DOI: 10.1643/CE-11-146
Copeia 2013, No. 2, 303–311

nesting seasons, it is expected that the sex ratio and survivalof the offspring will change over the years, minimizing theeffects of abnormally hot or rainy years in the long term. Weevaluated the effects of environmental variables on sex ratio,incubation duration, and hatching success for nests in thevarzea floodplain of the lower Amazon River near Santarem,in the Brazilian state of Para. This information is importantfor management and conservation projects, at a time whenthe Amazon region is a frontier for production of hydro-electricity, leading to dramatic changes in the fluvialenvironment, with unknown consequences for the residentchelonians.
MATERIALS AND METHODS
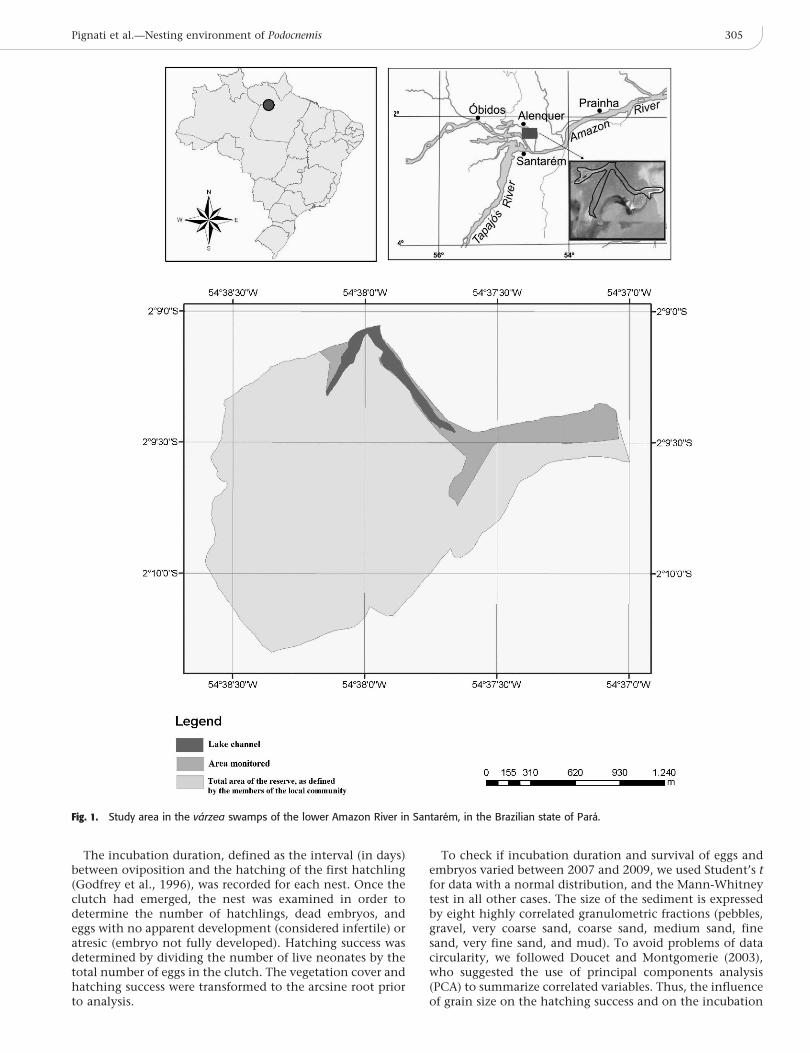
The data were collected at Taboleiro da Agua Preta, a riverterrace in the western part of the Brazilian state of Para,32 km northeast of Santarem (2u09912.50S, 54u37951.70W).The site is located at the edge of a floodplain lake on thelower Amazon River (Fig. 1). The varzea floodplains areflooded periodically (Ribeiro, 2007), with the highestprecipitation levels in April through July, and the lowestin October through December (data provided by CPRM-Belem). The varzea vegetation of this area is composed ofperiodically flooded marshes, interspersed with areas offorest. The substrate is alluvial and rich in organic matterdeposited by the annual floods (Ribeiro, 2007). This areaundergoes frequent modification from the constant processof sedimentation and erosion, which creates sandbanks anddestroys the beaches and margins of rivers and lakes,creating a phenomenon known locally as terras crescidas(deposition area) and terras caıdas (erosion area), respectively(Irion et al., 1997).
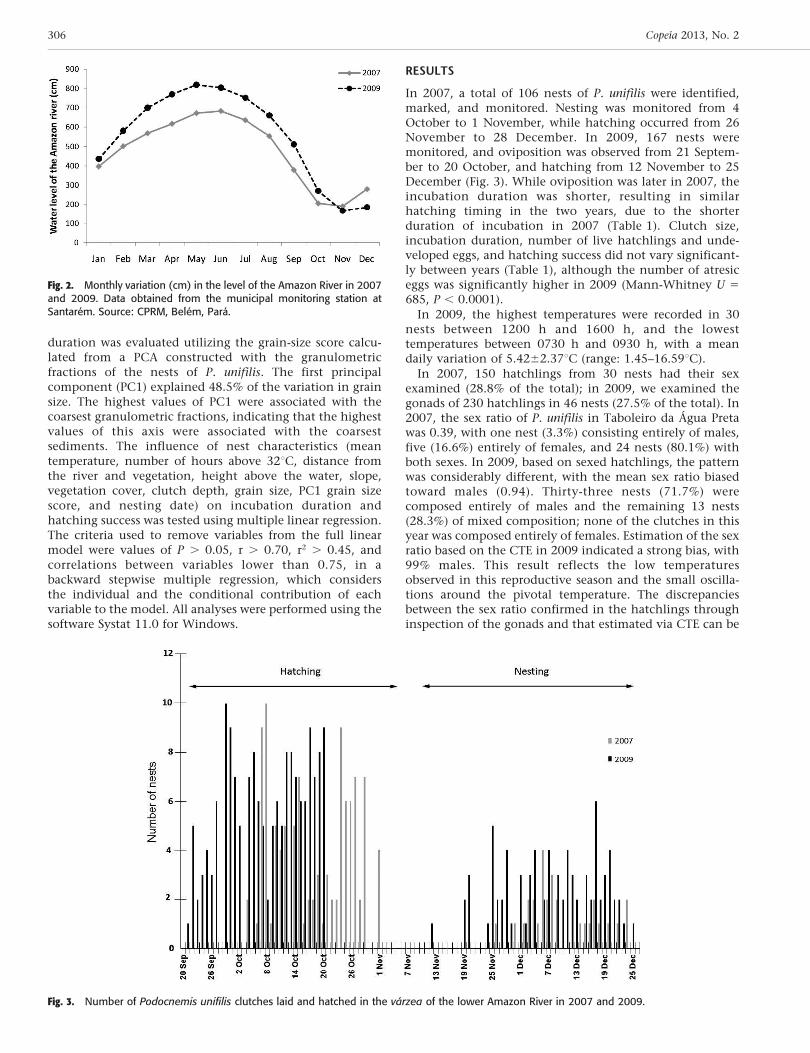
The Taboleiro da Agua Preta is located in an area of varzeaadministered by the residents of the Agua Preta community,where fishing is the primary economic activity. Themembers of the local community monitor and protect thestudy area in order to preserve the chelonian nesting siteand maintain stocks for harvesting and consumption duringthe high-water period. The terrace is covered by marshes andshrubs, with a predominance of Mimosa pigra (Giantsensitive tree), Echinochloa pyramidalis (Antelope grass),and Paspalum fasciculatum (Venezuela grass). The level ofthe Amazon River at Agua Preta fluctuated 493 cm in 2007(190–683 cm) and 653 cm in 2009 (166–819 cm; Fig. 2).
Nests of P. unifilis located along the channel of Agua Pretalake (340 ha) were monitored during the 2007 and 2009nesting seasons. An area of 25 ha was monitored in 2007,and 30 ha in 2009. In each year, the study plot wasexamined daily, between 0500 h and 0900 h, for signs ofnests, i.e., tracks made by nesting females and disturbedsubstrate indicating the location of the nest. Each nest wasmarked with a numbered stake and the date of ovipositionwas recorded; the location of each nest was determinedusing an Etrex H handheld GPS (Garmin). The depth of theclutch (distance between the surface and the first egg) wasmeasured on the day of oviposition, by opening the nestand using a measuring tape. The linear distances betweenthe nest and the edge of the water, and the nest and theclosest vegetation were measured on the day of oviposition.The height of the nests was measured from the lowest levelthat the river reached, which occurred on November 17 in2007 and November 20 in 2009. This procedure enabled themaximum height reached by nests to be compared between
the reproductive seasons, regardless of the variations in riverlevel.
The grain-size analysis was based on a 300 g sample ofsediment collected from the side of each monitored nest.The granulometric fractions were classified according to theFolk/Wentworth scale (Folk, 1974). The percentage ofvegetation cover above the surface of the nest was measuredusing a model C sphere densitometer (Lemmon, 1956), andthe slope of the substrate surface was measured using aclinometer. The latter three variables were recorded only in2009.
The sex ratio of P. unifilis was calculated by two auxiliaryand complementary methods: analysis of gonads and theincubation temperature. The gonad analysis was conductedin 2007, when five hatchlings were collected from each of 30randomly chosen nests, for verification of their gender andcalculation of the sex ratio (proportion of males/male+females). The hatchlings were killed and fixed in 10%
formalin (with 4 g of H3BO3 added per liter), and the sex wasdetermined based on the inspection of the gonads, follow-ing the procedure recommended by Malvasio et al. (2002).The specimens were preserved in 70% ethanol and depositedin the herpetological collection of the Goeldi Museum(MPEG) in Belem, Brazil, under catalog numbers MPEG 787–885 and MPEG 891–1035. In 2009, temperatures wererecorded in 30 nests using data loggers (Onset Corp)programmed to record the temperature, to the nearest0.1uC, at hourly intervals. The data loggers were maintainedin the center of the egg chamber for five days during thecritical period of thermosensitivity for the determination ofthe embryo’s sex (the middle third of the period ofembryonic development, according to Souza and Vogt,1994), that is, from the 25th day of incubation onward,considering an incubation period of around 60 days. In eachof the nests with data loggers, five hatchlings were alsocollected for analysis of the gonads, to determine the sex.
In the calculation of the sex ratio, the Constant Temper-ature Equivalent (CTE) of the middle third of the incubationperiod was also employed, which is preferable to the meantemperature in variable-temperature regimens (Georges etal., 1994, 2004), as in the case of nests of P. unifilis. For theestablishment of the CTE, it is necessary to know the rate ofembryonic development under constant temperature. Forthis, we used the equation established by Gomes (2011),who calculated the relationship between constant temper-ature and the incubation duration of P. unifilis, expressed by:
Duration of incubation daysð Þ~
182{ 3:727 incubation temperatureð Þ
The daily fraction of the embryonic development istherefore given by:
1=({3:727z182 incubation temperature)
The sum of all the daily fractions during the middle third ofthe incubation period was multiplied by the percentage ofthe total duration of incubation of the monitored period,obtaining the CTE of each nest. To estimate the sex ratio ofthe turtles in nests, the pivotal temperature of 31.8uC wasused; this was calculated by Gomes (2011), who incubatedeggs from a population in the Xingu River. With the aid ofsoftware TSD V.3.2.2 (Girondot, 1999; Godfrey et al., 2003)and the incubation data of Gomes (2011), the sex ratios ofeach nest were estimated, utilizing CTE.
304 Copeia 2013, No. 2
The incubation duration, defined as the interval (in days)between oviposition and the hatching of the first hatchling(Godfrey et al., 1996), was recorded for each nest. Once theclutch had emerged, the nest was examined in order todetermine the number of hatchlings, dead embryos, andeggs with no apparent development (considered infertile) oratresic (embryo not fully developed). Hatching success wasdetermined by dividing the number of live neonates by thetotal number of eggs in the clutch. The vegetation cover andhatching success were transformed to the arcsine root priorto analysis.
To check if incubation duration and survival of eggs andembryos varied between 2007 and 2009, we used Student’s tfor data with a normal distribution, and the Mann-Whitneytest in all other cases. The size of the sediment is expressedby eight highly correlated granulometric fractions (pebbles,gravel, very coarse sand, coarse sand, medium sand, finesand, very fine sand, and mud). To avoid problems of datacircularity, we followed Doucet and Montgomerie (2003),who suggested the use of principal components analysis(PCA) to summarize correlated variables. Thus, the influenceof grain size on the hatching success and on the incubation
Fig. 1. Study area in the varzea swamps of the lower Amazon River in Santarem, in the Brazilian state of Para.
Pignati et al.—Nesting environment of Podocnemis 305
duration was evaluated utilizing the grain-size score calcu-lated from a PCA constructed with the granulometricfractions of the nests of P. unifilis. The first principalcomponent (PC1) explained 48.5% of the variation in grainsize. The highest values of PC1 were associated with thecoarsest granulometric fractions, indicating that the highestvalues of this axis were associated with the coarsestsediments. The influence of nest characteristics (meantemperature, number of hours above 32uC, distance fromthe river and vegetation, height above the water, slope,vegetation cover, clutch depth, grain size, PC1 grain sizescore, and nesting date) on incubation duration andhatching success was tested using multiple linear regression.The criteria used to remove variables from the full linearmodel were values of P . 0.05, r . 0.70, r2 . 0.45, andcorrelations between variables lower than 0.75, in abackward stepwise multiple regression, which considersthe individual and the conditional contribution of eachvariable to the model. All analyses were performed using thesoftware Systat 11.0 for Windows.
RESULTS
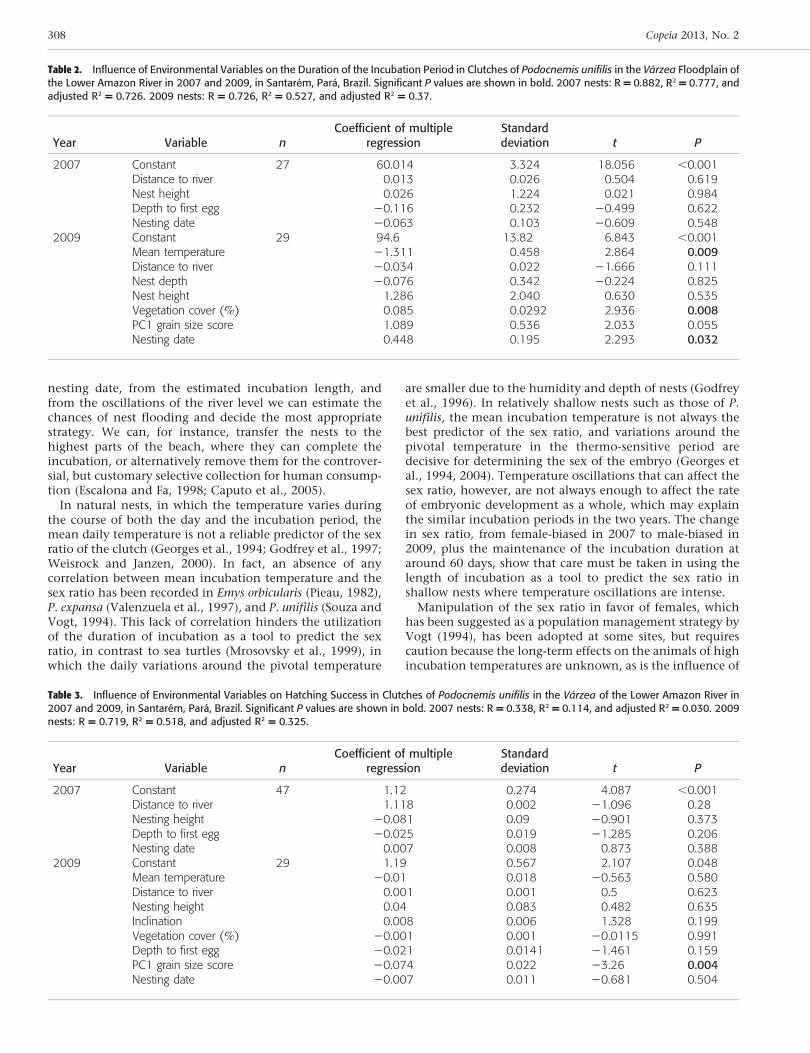
In 2007, a total of 106 nests of P. unifilis were identified,marked, and monitored. Nesting was monitored from 4October to 1 November, while hatching occurred from 26November to 28 December. In 2009, 167 nests weremonitored, and oviposition was observed from 21 Septem-ber to 20 October, and hatching from 12 November to 25December (Fig. 3). While oviposition was later in 2007, theincubation duration was shorter, resulting in similarhatching timing in the two years, due to the shorterduration of incubation in 2007 (Table 1). Clutch size,incubation duration, number of live hatchlings and unde-veloped eggs, and hatching success did not vary significant-ly between years (Table 1), although the number of atresiceggs was significantly higher in 2009 (Mann-Whitney U 5
685, P , 0.0001).
In 2009, the highest temperatures were recorded in 30nests between 1200 h and 1600 h, and the lowesttemperatures between 0730 h and 0930 h, with a meandaily variation of 5.4262.37uC (range: 1.45–16.59uC).
In 2007, 150 hatchlings from 30 nests had their sexexamined (28.8% of the total); in 2009, we examined thegonads of 230 hatchlings in 46 nests (27.5% of the total). In2007, the sex ratio of P. unifilis in Taboleiro da Agua Pretawas 0.39, with one nest (3.3%) consisting entirely of males,five (16.6%) entirely of females, and 24 nests (80.1%) withboth sexes. In 2009, based on sexed hatchlings, the patternwas considerably different, with the mean sex ratio biasedtoward males (0.94). Thirty-three nests (71.7%) werecomposed entirely of males and the remaining 13 nests(28.3%) of mixed composition; none of the clutches in thisyear was composed entirely of females. Estimation of the sexratio based on the CTE in 2009 indicated a strong bias, with99% males. This result reflects the low temperaturesobserved in this reproductive season and the small oscilla-tions around the pivotal temperature. The discrepanciesbetween the sex ratio confirmed in the hatchlings throughinspection of the gonads and that estimated via CTE can be
Fig. 2. Monthly variation (cm) in the level of the Amazon River in 2007and 2009. Data obtained from the municipal monitoring station atSantarem. Source: CPRM, Belem, Para.
Fig. 3. Number of Podocnemis unifilis clutches laid and hatched in the varzea of the lower Amazon River in 2007 and 2009.
306 Copeia 2013, No. 2

explained by the differences in temperature within the nestitself. Since the data loggers were placed in the middle of thenests, they did not include the entire microhabitat, so eggsin the upper part of the egg chamber might have beensubjected to higher temperatures or wider daily variations,which led to greater production of females. However, inspite of the differences, both methods showed goodconsistency of the results and a strong male bias in 2009.
In 2007, the incubation duration was not affected by anyof the variables evaluated (Table 2), whereas in 2009, nestslocated in areas with less plant cover and where meantemperatures were higher had a shorter incubation period.Early-deposited nests also had a shorter incubation period.
Hatching success was not affected by the distance of thenest from the river or vegetation, height above the river,slope of the beach, percentage of vegetation cover, initialnest depth, or days since first oviposition. In 2009, hatchingsuccess was negatively correlated with the PC1 grain-sizescores, indicating an increase in the survival of hatchlingsfrom nests with finer (Table 3), whereas an increasednumber of hours above the pivotal temperature negativelyaffected hatching success (F 5 4.4119, n 5 30, gl 5 1, P 5
0.0424).
DISCUSSION
The incubation duration recorded for P. unifilis at Agua Pretawas shorter than those reported for the species at other sitesin the Amazon and Orinoco basins, including ColombianAmazonia (Paez and Bock, 1998), the Capanaparo River inVenezuela (Thorbjarnarson et al., 1993), and the Javaes(Ferreira Junior and Castro, 2010) and Guapore (Soares,1996) rivers in Brazil. Of all these sites, the Taboleiro daAgua Preta is closest to the equator, with higher incubationtemperatures and, consequently, faster embryonic develop-ment and a shorter incubation duration. The incubationduration proved to be sensitive to the environmentalvariations; it was affected by the vegetation cover, nestingdate, and temperature, but did not vary between the nestingseasons. This indicates the importance of the time and nestlocation for embryonic development, because in theAmazon varzea floodplain the dry and rainy seasons varywidely year to year (Irion, 1986; Filizola et al., 2002). Sincethe incubation duration varied little, the time of nesting willdetermine the chances of hatching or flooding of nests. Thatis, the nests constructed near the beginning of the nestingseason have a greater chance to conclude hatching thanthose constructed at the end of the nesting season. Thelatter are flooded when the river begins to rise earlier(Ferreira Junior and Castro, 2010). Nests of P. unifilis located
in areas with less plant cover, similarly to the nests of P.expansa, which develops in more open areas such as beaches,have higher incubation temperatures and, consequently,shorter incubation periods (Pritchard and Trebbau, 1984;Ferreira Junior and Castro, 2003). In controlled experiments,Gutzke and Packard (1987) demonstrated that the meanincubation period in E. blandingii is 49 days at 31uC, but63 days at 26uC. This reinforces the findings of other studies,which showed that incubation is faster at higher tempera-tures (Yntema, 1976; Broderick et al., 2000; Godley et al.,2002).
Nests in fine-grained sediments experience reducedthermal conductivity and therefore less temperature varia-tion (Souza and Vogt, 1994; Ferreira Junior and Castro,2006), avoiding the very high temperatures that would befatal to the embryos. Similarly to the present findings,hatching success in P. expansa (Ferreira Junior and Castro,2010), Chelonia mydas (Mortimer, 1990), and P. unifilis(Souza and Vogt, 1994; Fachın-Teran and Von Mulhen,2003) is higher in finer-grained sediments. Nests with longerperiods of high temperatures (above 32uC) showed lowerhatching success, a pattern also recorded in C. caretta(Godley et al., 2001). In Pelodiscus sinensis, there are twolethal temperature extremes—above 34uC and below 23uC—and hatching success is lower at temperatures close to theseextremes (Du and Ji, 2003).
The influence of plant cover on embryonic development(Escalona and Fa, 1998) indicates the importance ofenvironmental integrity and preservation of the riparianvegetation for the reproduction of P. unifilis. The influenceof plant cover on the incubation temperature and sex ratioof freshwater turtles has been shown for several species:more females are produced in more-open areas (Vogt andBull, 1984; Vogt and Flores-Villela, 1986; Janzen, 1994a;Foley et al., 2000).
The sex ratio of chelonian hatchlings may vary naturallyfrom year to year (Janzen, 1994b; Godfrey et al., 1996),which does not necessarily cause a problem in view of thelong reproductive period in these animals (Janzen andPaukstis, 1991). Felix-Silva (2009) recorded a similar varia-tion in P. unifilis at the Tucuruı Reservoir in Para. There isnevertheless a clear need for long-term monitoring in orderto provide a more systematic understanding of the influenceof climatic variation on reproductive parameters, which willbe important for the development of appropriate conserva-tion and management strategies. Should long-term studiesindicate that the incubation duration does not vary much ina certain population, this information can be useful forconservation projects by indicating the nests that are moreprone to flooding and that should be managed. From the
Table 1. Basic Characteristics of Nests of Podocnemis unifilis Monitored in the Varzea Floodplain of the Lower Amazon River in 2007 and 2009, inSantarem, Para, Brazil. The statistics (t or U) refer to comparisons between years. Significant P values are shown in bold.
Variable
Mean±SD (n; range)
Statistic P2007 2009
Incubation duration (days) 58.963.64 (29; 54–66) 6065.23 (78; 51–74) U 5 955.5 0.2187Clutch size (eggs) 30.3166.58 (104; 9–42) 28.766.53 (139; 6–48) t 5 1.886 0.0616Live hatchlings 21.769.28 (48; 1–38) 22.168.6 (78; 2–42) t 5 0.9315 0.3534Undeveloped eggs 4.3764.39 (48; 1–23) 4.0463.64 (114; 0–17) U 5 1803.5 0.0791Atresic eggs 4.9365.04 (48; 1–19) 0.9462.29 (78; 0–15) U 5 685 ,0.0001Hatching success 0.6760.25 (48; 0.02–1) 0.7560.21 (78; 0.06–1) t 5 221.0607 0.2909
Pignati et al.—Nesting environment of Podocnemis 307
nesting date, from the estimated incubation length, andfrom the oscillations of the river level we can estimate thechances of nest flooding and decide the most appropriatestrategy. We can, for instance, transfer the nests to thehighest parts of the beach, where they can complete theincubation, or alternatively remove them for the controver-sial, but customary selective collection for human consump-tion (Escalona and Fa, 1998; Caputo et al., 2005).
In natural nests, in which the temperature varies duringthe course of both the day and the incubation period, themean daily temperature is not a reliable predictor of the sexratio of the clutch (Georges et al., 1994; Godfrey et al., 1997;Weisrock and Janzen, 2000). In fact, an absence of anycorrelation between mean incubation temperature and thesex ratio has been recorded in Emys orbicularis (Pieau, 1982),P. expansa (Valenzuela et al., 1997), and P. unifilis (Souza andVogt, 1994). This lack of correlation hinders the utilizationof the duration of incubation as a tool to predict the sexratio, in contrast to sea turtles (Mrosovsky et al., 1999), inwhich the daily variations around the pivotal temperature
are smaller due to the humidity and depth of nests (Godfreyet al., 1996). In relatively shallow nests such as those of P.unifilis, the mean incubation temperature is not always thebest predictor of the sex ratio, and variations around thepivotal temperature in the thermo-sensitive period aredecisive for determining the sex of the embryo (Georges etal., 1994, 2004). Temperature oscillations that can affect thesex ratio, however, are not always enough to affect the rateof embryonic development as a whole, which may explainthe similar incubation periods in the two years. The changein sex ratio, from female-biased in 2007 to male-biased in2009, plus the maintenance of the incubation duration ataround 60 days, show that care must be taken in using thelength of incubation as a tool to predict the sex ratio inshallow nests where temperature oscillations are intense.
Manipulation of the sex ratio in favor of females, whichhas been suggested as a population management strategy byVogt (1994), has been adopted at some sites, but requirescaution because the long-term effects on the animals of highincubation temperatures are unknown, as is the influence of
Table 2. Influence of Environmental Variables on the Duration of the Incubation Period in Clutches of Podocnemis unifilis in the Varzea Floodplain ofthe Lower Amazon River in 2007 and 2009, in Santarem, Para, Brazil. Significant P values are shown in bold. 2007 nests: R = 0.882, R2 = 0.777, andadjusted R2 = 0.726. 2009 nests: R = 0.726, R2 = 0.527, and adjusted R2 = 0.37.
Year Variable nCoefficient of multiple
regressionStandarddeviation t P
2007 Constant 27 60.014 3.324 18.056 ,0.001Distance to river 0.013 0.026 0.504 0.619Nest height 0.026 1.224 0.021 0.984Depth to first egg 20.116 0.232 20.499 0.622Nesting date 20.063 0.103 20.609 0.548
2009 Constant 29 94.6 13.82 6.843 ,0.001Mean temperature 21.311 0.458 2.864 0.009Distance to river 20.034 0.022 21.666 0.111Nest depth 20.076 0.342 20.224 0.825Nest height 1.286 2.040 0.630 0.535Vegetation cover (%) 0.085 0.0292 2.936 0.008PC1 grain size score 1.089 0.536 2.033 0.055Nesting date 0.448 0.195 2.293 0.032
Table 3. Influence of Environmental Variables on Hatching Success in Clutches of Podocnemis unifilis in the Varzea of the Lower Amazon River in2007 and 2009, in Santarem, Para, Brazil. Significant P values are shown in bold. 2007 nests: R = 0.338, R2 = 0.114, and adjusted R2 = 0.030. 2009nests: R = 0.719, R2 = 0.518, and adjusted R2 = 0.325.
Year Variable nCoefficient of multiple
regressionStandarddeviation t P
2007 Constant 47 1.12 0.274 4.087 ,0.001Distance to river 1.118 0.002 21.096 0.28Nesting height 20.081 0.09 20.901 0.373Depth to first egg 20.025 0.019 21.285 0.206Nesting date 0.007 0.008 0.873 0.388
2009 Constant 29 1.19 0.567 2.107 0.048Mean temperature 20.01 0.018 20.563 0.580Distance to river 0.001 0.001 0.5 0.623Nesting height 0.04 0.083 0.482 0.635Inclination 0.008 0.006 1.328 0.199Vegetation cover (%) 20.001 0.001 20.0115 0.991Depth to first egg 20.021 0.0141 21.461 0.159PC1 grain size score 20.074 0.022 23.26 0.004Nesting date 20.007 0.011 20.681 0.504
308 Copeia 2013, No. 2

a female bias on population structure. Podocnemidids showsperm storage, multiple paternity, and polyandrous behav-ior (Pearse et al., 2006; Fantin et al., 2008, 2010), which maybe beneficial for a number of reasons, especially where thequantity of sperm produced by a single male is insufficientto fertilize the clutch. Additional benefits would includeincreased fitness, and the maintenance of population sizeand genetic variability (Reichard et al., 2007; Fantin et al.,2008). Therefore, changing the sex ratio of clutches mayhave undesired consequences, especially from the geneticviewpoint. In addition, because the sex ratio may varybetween years, as found in the present study, long-termmonitoring of a number of sites will be necessary for a betterunderstanding of the role of temporal and regional fluctu-ations of this parameter in the structure and functioning ofthe species’ population.
Nests of P. unifilis with only one sex and/or a stronglybiased sex ratio were common, in spite of the wide range ofpivotal temperature variation. In Dermochelys coriacea (Che-valier et al., 1999) and C. caretta (Yntema and Mrosovsky,1982), a variation of around 1uC is enough to effectprofound changes in the sex ratio; but for P. unifilis, nestswith only one sex are predicted at temperatures of around28 and 35 C (Souza and Vogt, 1994; Gomes, 2011). On theother hand, this wide thermal tolerance indicates the greatenvironmental plasticity of P. unifilis for exploiting nestingareas and the most varied habitats (Pritchard and Trebbau,1984; Vogt, 2008).
The variation of sex ratio between the years indicates thenecessity of long-term monitoring in order to evaluate thepopulation dynamics and how P. unifilis is affected byenvironmental variables. The importance of the environ-mental characteristics (climate, vegetation, and geology) forthe embryonic development is unquestionable, but it is notyet possible to quantify and pinpoint the influence of eachof these elements on the sex ratio and hatching success. Ourdata indicate, however, that the annual variations originat-ing from the climate are of extreme importance forreproduction, exceeding the importance of plant coverand of the geological features of the nesting area.
If, on the one hand, the predominant production of acertain sex in a year has little or no effect on thepopulations of a long-lived organism such as P. unifilis,on the other hand, climatic changes such as those forecastby global warming can foster a gradual change in the sexratio of juveniles and subadults, and subsequently in thepopulation as a whole, with obvious negative consequencesas described above. Long-term monitoring involving di-verse populations, which might demonstrate the existenceof regional differences in climate even at the pivotaltemperature of the species, is thus recommended. There-fore, studies examining possible differences in this param-eter in subpopulations are also necessary. Integration ofthese pieces of information will enable the establishment ofmanagement measures that compensate for the impacts ofdisturbance on organisms whose sex ratio is determined byenvironmental variables.
ACKNOWLEDGMENTS
We are grateful to the Para State Scientific ResearchFoundation for financial support and the Institute forAmazonian Research for logistical support during fieldwork,and to the Brazilian National Research Council (CNPq) forproviding a post-graduate stipend (133325/2009-1). We also
thank R. Vieira and all the residents of the Agua Pretacommunity for their assistance during fieldwork, C. Costafor help with sexing of the hatchlings, and A. Silva for helpwith statistical analysis. Collection of the specimens wasauthorized by the Brazilian Institute for the Environmentand Renewable Natural Resources (IBAMA), under speciallicense number 10441-1.
LITERATURE CITED
Alho, C. J. R., T. M. S. Danni, and L. F. M. Padua. 1985.Temperature-dependent sex determination in Podocnemisexpansa (Testudinata: Pelomedusidae). Biotropica 17:75–78.
Alho, C. J. R., and L. F. M. Padua. 1982. Reproductiveparameters and nesting behavior of the Amazon turtlePodocnemis expansa (Testudinata: Pelomedusidae) in Bra-zil. Canadian Journal of Zoology 60:97–103.
Broderick, A. C., B. J. Godley, S. Reece, and J. R. Downie.2000. Incubation periods and sex ratios of green turtles:highly female biased hatchling production in the easternMediterranean. Marine Ecology Progress Series 202:273–281.
Bujes, C. S., and L. Verrastro. 2009. Nest temperature,incubation time, hatching, and emergence in the Hilaire’sside-necked turtle (Phrynops hilarii). Herpetological Con-servation and Biology 4:306–312.
Caputo, F. P., D. Canestrelli, and L. Boitani. 2005.Conserving the Terecay (Podocnemis unifilis, Testudines:Pelomedusidae) through a community-based sustainableharvest of its eggs. Biological Conservation 126:84–92.
Caut, S., E. Guirlet, P. Jouquet, and M. Girondot. 2006.Influence of nest location and yolkless eggs on thehatching success of leatherback turtle clutches in FrenchGuiana. Canadian Journal of Zoology 84:908–915.
Chevalier, J., M. H. Godfrey, and M. Girondot. 1999.Significant difference of temperature-dependent sex de-termination between French Guiana (Atlantic) and PlayaGrande (Costa-Rica, Pacific) leatherbacks (Dermochelyscoriacea). Annales des Sciences Naturelles—Zoologie etBiologie Animale 20:147–152.
Congdon, J. D., R. D. Nagle, O. M. Kinney, M. Osentoski,H. W. Avery, C. R. S. Van Loben, and D. W. Tinckle.2000. Nesting ecology and embryo mortality: implicationsfor hatchling success and demography of Blanding’sturtles (Emydoidea blandingii). Chelonian Conservationand Biology 3:569–579.
Doody, J. S., A. Georges, and J. E. Young. 2003. Twice everysecond year: reproduction in the pig-nosed turtle, Car-ettochelys insculpta, in the wet-dry tropics of Australia.Journal of Zoology 259:179–188.
Doucet, S. M., and R. Montgomerie. 2003. Multiple sexualornaments in satin bowerbirds: ultraviolet plumage andbowers signal different aspects of male quality. BehavioralEcology 14:503–509.
Du, W. G., and X. Ji. 2003. The effects of incubationthermal environments on size, locomotor performanceand early growth of hatchling soft-shelled turtles, Pelo-discus sinensis. Journal of Thermal Biology 28:279–286.
Escalona, T., and J. E. Fa. 1998. Survival of nests of theterecay turtle (Podocnemis unifilis) in the Nichare—TawaduRivers, Venezuela. Journal of Zoology 244:303–312.
Fachın-Teran, A., and M. Von Mulhen. 2003. Reproduc-cion de la taricaya Podocnemis unifilis Troschel 1848(Testudines: Podocnemididae) en la varzea del medioSolimoes, Amazonas, Brasil. Ecologia Aplicada 2:125–132.
Pignati et al.—Nesting environment of Podocnemis 309

Fantin, C., I. P. Farias, L. A. S. Monjelo, and T. Hrbek.2010. Polyandry in the red-headed river turtle Podocnemiserythrocephala (Testudines, Podocnemididae) in the Bra-zilian Amazon. Genetics and Molecular Research 9:435–440.
Fantin, C., L. S. Viana, L. A. S. Monjelo, and I. P. Farias.2008. Polyandry in Podocnemis unifilis (Pleurodira; Podoc-nemididae), the vulnerable yellow-spotted Amazon Riverturtle. Amphibia–Reptilia 29:479–486.
Felix-Silva, D. 2009. Ecologia e Conservacao de Podocnemisunifilis Troschel 1848 (Testudines, Podocnemididae) noReservatorio da UHE Tucuruı, Para-Brasil. Unpubl. Ph.D.thesis, Department of Ecology, Universidade do Estado doRio de Janeiro, Rio de Janeiro, R.J., Brazil.
Ferreira Junior, P. D. 2009. Aspectos ecologicos da determi-nacao sexual em tartarugas. Acta Amazonica 39:139–154.
Ferreira Junior, P. D., and P. T. A. Castro. 2003. Geologicalcontrol of Podocnemis expansa and Podocnemis unifilisnesting areas in Rio Javaes, Bananal Island, Brazil. ActaAmazonica 33:445–468.
Ferreira Junior, P. D., and P. T. A. Castro. 2006. Thermalenvironment characteristics of Podocnemis expansa andPodocnemis unifilis nesting areas on the Javaes River,Tocantins, Brazil. Chelonian Conservation and Biology5:102–107.
Ferreira Junior, P. D., and P. T. A. Castro. 2010. Nestingecology of Podocnemis expansa (Schweigger, 1812) andPodocnemis unifilis (Troschel, 1848) (Testudines, Podocne-mididae) in the Javaes River, Brazil. Brazilian Journal ofBiology 70:85–94.
Filizola, N., J. L. Guyot, M. Molinier, V. Guimaraes, E. E.Oliveira, and M. A. Freitas. 2002. Caracterizacao hidrolo-gica da bacia Amazonica, p. 35–53. In: Amazonia: UmaPerspectiva Interdisciplinar. A. A. E. Rivas and C. E. C. Freitas(eds.). Editora da Universidade do Amazonas, Manaus.
Foley, M. A., A. S. Peck, R. G. Harman, and L. W.Richardson. 2000. Loggerhead turtle (Caretta caretta)nesting habitat on low-relief mangrove island in south-west Florida and consequences to hatchling sex ratios.Herpetologica 56:433–445.
Folk, R. L. 1974. Petrology of Sedimentary Rocks. HemphillPublication Company, Austin, Texas.
Georges, A., S. Doody, K. Beggs, and J. Young. 2004.Thermal models of TSD under laboratory and fieldconditions, p. 79–89. In: Temperature-Dependent SexDetermination in Vertebrates. N. Valenzuela and V. A.Lance (eds.). Smithsonian Books, Washington, D.C.
Georges, A., C. Limpus, and R. Stoutjesdijk. 1994.Hatchling sex in the marine turtle Caretta caretta isdetermined by proportion of development at a tempera-ture, not daily duration of exposure. Journal of Experi-mental Zoology 270:432–444.
Girondot, M. 1999. Statistical description of temperature-dependent sex determination using maximum likelihood.Evolutionary Ecology Research 1:479–486.
Godfrey, M. H., R. Barreto, and N. Mrosovsky. 1996.Estimating past and present sex ratios of sea turtles inSuriname. Canadian Journal of Zoology 74:267–277.
Godfrey, M. H., R. Barreto, and N. Mrosovsky. 1997.Metabolically generated heat of developing eggs and itspotential effect on sex ratio of sea turtle hatchlings.Journal of Herpetology 31:616–619.
Godfrey, M. H., V. Delmas, and M. Girondot. 2003.Assessment of patterns of temperature-dependent sex
determination using maximum likelihood model selec-tion. Ecoscience 10:265–272.
Godley, B. J., A. C. Broderick, F. Glen, and G. C. Hays.2002. Temperature-dependent sex determination of As-cension Island green turtles. Marine Ecology ProgressSeries 226:115–124.
Godley, B. J., A. C. Broderick, and N. Mrosovsky. 2001.Estimating hatchling sex ratios of loggerhead turtles inCyprus from incubation durations. Marine Ecology Prog-ress Series 210:195–201.
Gomes, A. S. 2011. Razao sexual de Podocnemis unifilis noParque Indıgena do Xingu, Mato Grosso Brasil. Unpubl.Master’s thesis, Universidade Vila Velha, Vila Velha, Brazil.
Gutzke, W. H. N., and G. C. Packard. 1987. The influenceof temperature on eggs and hatchling of Blanding’sturtles, Emydoidea blandingii. Journal of Herpetology21:161–163.
Irion, G. 1986. Quaternary geology of amazonian lowland,p. 494–498. In: Simposio do tropico umido, 1. Belem,Anais . . . EMBRAPA-CPATU, Belem.
Irion, G., W. J. Junk, and J. A. S. N. Mello. 1997. The largeCentral Amazonian River floodplains near Manaus: geo-logical, climatological, hydrological and geomorphologi-cal aspects, p. 23–46. In: The Central Amazon Floodplain:Ecology of a Pulsing System. W. J. Junk (ed.). Springer,Berlin.
Janzen, F. J. 1994a. Vegetational cover predicts the sex ratioof hatchling turtles in natural nests. Ecology 75:1593–1599.
Janzen, F. J. 1994b. Climate change and temperature-dependent sex determination in reptiles. Proceedings ofthe National Academy of Sciences of the United States ofAmerica 91:7487–7490.
Janzen, F. J., and G. L. Paukstis. 1991. Environmental sexdetermination in reptiles: ecology, evolution, and exper-imental design. The Quarterly Review of Biology 66:149–179.
Lemmon, P. E. 1956. A spherical densiometer for estimatingforest overstory density. Forest Science 2:314–320.
Lubiana, A., and P. D. Ferreira Junior. 2009. Pivotaltemperature and sexual dimorphism of Podocnemis ex-pansa hatchlings (Testudines: Podocnemididae) fromBananal Island, Brazil. Zoologia 26:527–533.
Malvasio, A., A. M. Souza, E. S. Reis, and E. C. Farias. 2002.Morfologia dos orgaos reprodutores de recem-eclodidosde Podocnemis expansa (Schweigger, 1812) e P. unifilis(Troschel, 1848) (Testudines, Pelomedusidae). PublicacoesAvulsas do Instituto Pau Brasil de Historia Natural5:27–37.
McKnight, C. M., and W. H. N. Gutzke. 1993. Effects of theembryonic environment and of hatchling housing condi-tions on growth of young snapping turtles (Chelydraserpentina). Copeia 1993:475–482.
Mortimer, J. A. 1990. The influence of beach sandcharacteristics on the nesting behavior and clutch survivalof green turtles (Chelonia mydas). Copeia 1990:802–817.
Mrosovsky, N. 1980. Thermal biology of sea turtle.American Zoologist 20:531–547.
Mrosovsky, N., C. Baptistotte, and M. H. Godfrey. 1999.Validation of incubation duration as an index of the sexratio of hatchling sea turtles. Canadian Journal of Zoology77:831–835.
Paez, V. P., and B. C. Bock. 1998. Temperature effect onincubation period in the yellow-spotted river turtle,
310 Copeia 2013, No. 2

Podocnemis unifilis, in the Colombian Amazon. ChelonianConservation and Biology 3:31–36.
Paez, V. P., A. Restrepo, M. Vargas-Ramırez, and B. C.Bock. 2009. Podocnemis lewyana (Dumeril 1852)—Magda-lena River turtle, p. 024.1–024.6. In: Conservation Biologyof Freshwater Turtles and Turtoises: A Compilation Projectof the IUCN/SSC Tortoise and Freshwater Turtle Group.A. G. J. Rhodin, P. C. H. Pritchard, P. P. van Dijk, R. A.Sumure, K. A. Buhlmann, J. B. Iverson, and R. A.Mittermeier (eds.). Chelonian Research Monographs No. 5.
Pearse, D. E., R. B. Dastrup, O. Hernandez, and J. W. Sites,Jr. 2006. Paternity in an Orinoco population of endan-gered Arrau River Turtles, Podocnemis expansa (Pleurodira;Podocnemididae), from Venezuela. Chelonian Conserva-tion and Biology 5:232–238.
Pieau, C. 1982. Modalities of the action of temperature onsexual differentiation in field developing embryos of theEuropean pond turtle Emys orbicularis (Emydidae). TheJournal of Experimental Zoology 220:353–360.
Pieau, C. 1996. Temperature variation and sex determina-tion in reptiles. BioEssays 18:19–26.
Pritchard, P. C. H. 1979. Encyclopedia of Turtles. T.F.H.Publications Inc., Neptune, New Jersey.
Pritchard, P. C. H., and P. Trebbau. 1984. The Turtles ofVenezuela. Society for the Study of Amphibians andReptiles, Oxford, Ohio.
Reichard, M., S. C. Le Comber, and C. Smith. 2007.Sneaking from a female perspective. Animal Behaviour74:679–688.
Rhen, T., E. Willingham, J. Sakata, and D. Crews. 1999.Incubation temperature influence sex-steroid levels injuvenile red-eared slider turtles, Trachemys scripta, aspecies with temperature-dependent sex determination.Biology of Reproduction 61:1275–1280.
Ribeiro, N. V. 2007. Atlas da Varzea: Amazonia Brasil.Ibama, Manaus, Amazonas, Brazil.
Soares, M. F. G. S. 1996. Biometria de femea de Podocnemisexpansa (Testudines, Pelomedusidae) em atividade dedesova no rio Guapore, Rondonia, Brasil. Acta BiologicaLeopoldensia 18:93–101.
Souza, R. R. D., and R. C. Vogt. 1994. Incubationtemperature influences sex and hatchling size in theneotropical turtle Podocnemis unifilis. Journal of Herpetol-ogy 28:453–464.
Thorbjarnarson, J. B., N. Perez, and T. Escalona. 1993.Nesting of Podocnemis unifilis in the Capanaparo River,Venezuela. Journal of Herpetology 27:344–347.
Valenzuela, N., R. Botero, and E. Martinez. 1997. Fieldstudy of sex determination in Podocnemis expansa fromColombian Amazonia. Herpetologica 53:390–398.
Vanzolini, P. E. 1967. Notes on the nesting behaviorof Podocnemis expansa in the Amazon Valley (Testudines,Pelomedusidae). Papeis Avulsos de Zoologia 20:191–215.
Vogt, R. C. 1994. Temperature controlled sex determinationas a tool for turtle conservation. Chelonian Conservationand Biology 1:159–162.
Vogt, R. C. 2008. Amazon Turtles. Biblos, Lima, Peru.Vogt, R. C., and J. Bull. 1984. Ecology of hatchling sex ratio
in map turtle. Ecology 65:582–587.Vogt, R. C., and O. A. Flores-Villela. 1986. Determinacion
del sexo em tortugas por la temperatura de incubacion delos huevos. Ciencia 37:21–32.
Weisrock, D. W., and F. J. Janzen. 2000. Thermal andfitness-related consequences of nest location in PaintedTurtles (Chrysemys picta). Functional Ecology 13:94–101.
Yntema, C. L. 1976. Effects of incubation temperatures onsexual differentiation in the turtle, Chelydra serpentina.Journal of Morphology 150:453–462.
Yntema, C. L., and N. Mrosovsky. 1982. Critical periodsand pivotal temperatures for sexual differentiation inloggerhead sea turtles. Canadian Journal of Zoology60:1012–1016.
Pignati et al.—Nesting environment of Podocnemis 311



















