Embed Size (px)
Citation preview

Aquaculture and Fisheries Management 1993, 24, 295-304
Essential fatty acid requirements of larval gilthead seabream, Sparus aurata (L.)
C. RODRIGUEZ & J. A. PEREZ Departamento de Biologia Animal, Universidad deLa Laguna, Tenerife, Spain
M. S. IZQUIERDO Departamento de Biologia, Fac. Ciencias del Mar, Universidad deLas Palmas, Las Palmas, Spain
J. MORA & A. LORENZO Departamento de Biologia Animal, Universidad de LaLaguna, Tenerife, Spain
H. FERNANDEZ-PALACIOS Centro de Tecnologia Pesquera, Telde, Las Palmas,Spain
Abstract. Two experiments were carried out to study the effect of «-3 HUFA levels in rotiferson survival, growth, activity and fatty acid composition of gilthead bream, Sparus aurata (L.),larvae. From the third to the 15th day after hatching, gilthead bream larvae received one of thethree kinds of rotifers containing different percentages of n-3 HUFA. Moisture, crude lipid,saponifiable matter and fatty acid composition of total lipids of rotifers and larvae weredetermined.
A good correlation was found between larval growth and n-3 HUFA levels in rotifers.Larval survival was also significantly improved by the elevation of the n-3 HUFA levels inrotifers. A high occurrence of hydrops was registered in larvae fed with EFA-deficient rotifers.The n-3 HUFA levels in the larvae were increased by the elevation of n-3 HUFA contents inrotifers. However, n-9 fatty acids in the larvae remained almost constant, regardless of thedifferent 18:1 n-9 contents in rotifers. Therefore, the ratio of oleic acid to n-3 HUFA, known tobe an indicator of the EFA deficiency in fish, was reduced by the elevation of the n-3 HUFAlevels in rotifers.
Introduction
Lipids in marine teleost eggs and larvae play an important role in biomembrane formationwas well as in energy production during development (Tocher & Sargent 1984; Tocher,Fraser, Sargent & Gamble 1985). Analysis ofthe chemical composition of eggs and the earlylarval stages shows that (n-3) highly unsaturated fatty acids (HUFA) are some of the maincomponents of the lipids of several marine species such as red sea bream, Pagrus major(Temminck & Schlegel) (Kimata 1983; Izquierdo, Watanabe, Takeuchi, Arakawa &Kitajima 1989) and gilthead sea bream, Sparus aurata (L.) (Mourente & Odriozola 1990),suggesting the importance of these fatty acids during larval development. Falk-Petersen,Sargent, Fox, Falk-Petersen, Huag & Kjorsvik (1989) found that n-3 HUFA in Atlantichalibut, Hippoglossus hippoglossus (JL.), eggs are catabolized markedly following fertili-zation and can be regarded as a major energy source during this period, whereas saturates areused in preference to /i-3 HUFA during the later embryonic stages.
Correspondence: Dr M.S. Izquierdo, Dpto. de Biologia, Fac. Ciencias del Mar, Univ. Las Palmas, 35017 LasPalmas, Spain.
295

296 C. Rodriguez et al.
Because of the importance of these fatty acids for marine fish larvae, a principal factor inthe dietary value of rotifers as live food is their content of n-3 HUFA (Watanabe et al.1978a,b,c). Few studies have been cpnducted on the essential fatty acid (EFA) requirementsof marine fish larvae (Le Milinaire, Gatesoupe & Stephan 1983; Franicevic, Lisac, Buble,L6ger & Sorgeloos 1987; Koven, Kissil & Tandler 1989; Izquierdo, Watanabe, Takeuchi,Arakawa & Kitajima 1990; Ostrowski & Divakaran 1990; Izquierdo, Arakawa, Takeuchi,Haroun & Watanabe 1992). Some of them indicate that EFA requirements differ not onlybetween species but also between growth stages (Gatesoupe & Le Milinaire 1985; Izquierdoet al. 1989). Watanabe, Izquierdo, Takiuchi, Satoh & Kitajima (1989) also founddocosahexaenoic acid to be superior to eicosapentaenoic acid as EFA for larval red seabream. N-3 HUFA are essential for the growth of gilthead bream larvae (Koven, Tandler,Kissil, Sklau, Friezlander & Havel 1990) but the effect of these fatty acids on larval survival isstill unclear.
The purpose of the present study was to clarify the effect of n-3 HUFA levels in rotifers onsurvival, growth, activity and fatty acid composition of gilthead bream larvae.
Materials and methods
Rotifers, which had been cultured on baker's yeast, were enriched by the direct method(Watanabe, Ohta, Kitajima & Fujita 1982). A lipid emulsion was prepared of 8ml of lipids,91 ml of sea water, Ig of raw egg yolk and 0-30g of soybean lecithin. The lipids used were:oleic acid (Sigma Chemical Co. Ltd, St Louis, MO, USA) for treatment 1; 50% oleic acidcombined with 50% EPA 28, a triacilglycerol mixture containing 40% n-3 HUFA (NippaiCo. Ltd, Tokyo, Japan) for treatment 2; and EPA 28 for treatment 3. Thus, the n-3 HUFAlevels in the emulsions were calculated to be 0%, 20% and 40%, respectively. Rotifers, at aninitial density of 133-200 individuals/ml, were fed 100ml of lipid emulsion in 100-1 tanks filledwith filtered sea water. They were enriched for 18h at a water temperature of 19-5°C.Samples of rotifers were taken nine times during the experiment to analyse their n-3 HUFAcontents.
During larval feeding trials, sea water was supplied at a rate of 300-350 ml/min to preventthe consumption of starved rotifers by the larvae, as their weight and biochemicalcomposition changes during starvation (Scott & Baynes 1978; Rubio 1986). As the n-3HUFA content of rotifers fed Nannochloropsis or other microalgae seems to increaseconsiderably (Scott & Middleton 1979; Watanabe, Oowa, Kitajima, Fujita & Yone 1979),algae were not added to the experimental tanks for a better control of the n-3 HUFA level inrotifers.
Larval survival was determined by counting all the living fish at the end of the experiment.Growth rate was assessed from 30 living larvae from the initial sample and each replicate atthe end of the experiment, as well as the percentage of air-inflated swim bladder and theactivity test. This test was carried out with 30 larvae of each replicate by holding them out ofthe water in a scoop net for 3 s and moving them into lO-I tanks to check the fish response tothe stress (Kitajima, Arakawa, Fujita, Imada, Watanabe & Yone 1980). At the end of thefeeding trial, the remaining living larvae from each tank were collected to analyse theirbiochemical composition.
Essential fatty acids for larval Sparus aurata 297
Moisture, crude lipid, saponifiable matter and fatty acid composition of total lipids ofeach sample were analysed. Lipid extraction was carried out by the method described byFolch, Lees & Stanley (1957). Fatty acid mixtures were prepared from the crude lipids bysaponification with KOH. Unsaponifiable matter was weighed. Methyl esters of fatty acidswere prepared by transesterification with borontrifluoride in methanol as described byMetcalfe & Schmitz (1961).
Experimental conditions
Experiment 1. Two-day-old gilthead sea bream larvae measuring 3 12mm in total length,obtained from the broodstock of Instituto Espanol de Oceanografi'a in Tenerife, wererandomly distributed in three 500-1 tanks at a density of 100 larvae/1. From the third to the15th day after hatching, larvae received one of the three kinds of rotifers enriched with theemulsions containing 0%, 20% or 40% n-3 HUFA. During the feeding experiment watertemperature ranged between 19-5 and 21°C.
Experiment 2. Gilthead bream eggs were obtained by natural spawning from thebroodstock of Centro de Tecnologia Pesquera in Gran Canaria. Just before hatching theywere randomly divided into three triplicated groups and stocked in 100-1 tanks with acontinuous flow of water. About 10000 3-day-old larvae/tank were experimentally fed. Fromthe third to the 17th day after hatching, larvae were respectively fed with one of the threekinds of enriched rotifers. Water temperature ranged between 19-1 and 20-6°C during thefeeding experiment.
All biological and biochemical results were analysed by one-way ANOVA, Tukey {P s0-05) and Scheff6 (P s 0-01) tests. Linear regression analyses were performed on severalcombinations of rotifer-larva parameters.
Results
Experiment 1
The n-3 HUFA levels in rotifers were 0-19%, 2-22% and 5-90% ofthe dry weight (Table 1).After 11 days of feeding, lower feeding and swimming activities were observed in fishreceiving the lowest amount of n-3 HUFA. Fish growth was low in this treatment, but theelevation of n-3 HUFA levels in rotifers improved the growth in the other two treatments.
Besides the poor growth found in fish receiving the lowest amount of n-3 HUFA, a highpercentage of hydrops was found in these larvae. The affected larvae showed subepidermic
Table 1. Growth of gilthead sea bream larvae after 14 days of feeding in experiment 1*
Variable
n - 3 HUFA in rotifers (% dry weight)Total length (mm)
1
0-194-45"
Treatment
2
2-224-57"
3
5-9590=
* Means with the same superscript letter are not significantly different (P < 0-001), n = 30.
298 C. Rodriguez et al.
and cephalic water accumulations which could be related to the n-3 HUFA deficiency. Thispathology has been described by Yamashita (1981) for red sea bream.
Possibly owing to poor egg quality and handling of the initial larvae, high larval mortalitywas observed in all the treatments during the first days of feeding. Therefore, no relationshipbetween n-3 HUFA levels in rotifers and larval survival could be found.
Biochemical composition of rotifers and larvae followed the same trends as in experiment2 and will be discussed later.
Experiment 2
The n-3 HUFA levels in rotifers were 0-07%, 3-24% and 4-98% of the dry weight (Table 2).These treatments clearly affected the growth and survival of fish. After 7 days of feeding asmall difference was apparent in larval growth rate between the treatments, but they were notsignificantly different (P s 0-05). However, at the end of the experiment, after 14 days offeeding, there were differences in total length at a level of P :s 0-05 between each of the threetreatments (Table 2).
After 14 days of feeding, a high mortality appeared in the replicates of larvae fed withrotifers containing only 0-07% n-3 HUFA. Survival rate was the lowest in these fish and thehighest in all the replicates receiving the highest amount of n-3 HUFA, being significantlydifferent from treatment 1 (P :£ 0 05).
Although it was observed that larvae were more active when they were fed higher levels ofn-3 HUFA in rotifers, there were no significant differences (P ^ 0-05) among the treatments,in terms of either survival after the activity test or percentages of air-inflated swim bladder.
The rotifers' contents of «-3 fatty acids and n-3 HUFA increased significantly (P < 0 001)with.the elevation of the n-3 HUFA contents in the lipid emulsions (Table 3). In contrast, thelevels ofn-9 and monounsaturated fatty acids decreased significantly (P ^ 0 001). There wasa significant (P s 0-0001, r = 0-84, r = 0-92) correlation between n-3 and n-9 and betweenn-3 and monounsaturated fatty acids. Saturated and n-6 fatty acid composition of rotifersremained steady.
In total lipids of larvae (Table 4) it was observed at the end of the experiment thatsaturated and monounsaturated fatty acids were similar in all the fish groups and to the levelsat the beginning of the experiment, despite the great variations in the rotifers' contents ofthese fatty acids.
Table 2. Growth, survival and activity of gilthead bream larvae in experiment 2*
Variable
n - 3 HUFA in rotifers (% dry weight)Total length (mm)''
at day 7at day 14
Survival rate (%)*Survival rate after activity test (%)*
1
0-07
4-23°4-62"
6-5«
Treatment
2
3-24
4-24"4.79c798 ' '7-65«
3
4-98
4-25='5-32"
1476'8-00«
* Means with the same letter are not significantly different (P < 0-001).+ n = 30.
Essential fatty acids for larval Sparus aurata 299
Table 3. Lipid content and fatty acid composition (% area) of total lipids of rotifers in experiment 2
Treatment
Fatty acids 1
12:014:014:115:115:0
16:016:1/1 -16:1/1 -16:1/1 -17:017:1
18:018:1/1 -18:1/1 -18:1/1 -18:1/1 -18:2/1 -19:018:2/1 -18:3/1 -18:3/1 -18:2/1 -18:4/1 -18:4/1 -
20:1/1-20:1/1-20:2/1 -20:3/1 -20:2/1 -20:4/1 -20:4/1 -20:5/1 -
22:1/1-22:1/1 -22:4/1 -22:5/1 -22:6n -
975
119759
696331
119996633
119933
n-3 HUFAn - 3n - 6n - 9SaturatedMonoenoicsTotal lipids*
0-12 0-25 0-362-22 2-70 3-940-50 044 0-546-13 0-14 0-160-17 0-11 0-21
5-53 5-03 6-042-14 1-30 0-91
16-00 16-08 16-220-39 0-31 0-320-35 0-56 1-060-98 0-86 0-91
2-03 1-77 1-950-79 1-02 0-85
45-26 29-30 16-813-06 3-54 2-90015 0-38 0-170-58 0-50 0-452-10 1-78 1-488-55 6-73 5-200-50 0-44 0-401-39 1-37 0-971-45 1-02 1460-11 2-23 4-290-00 0-26 0-55
0-39 0-72 0-691-07 1-32 1-080-64 0-60 0-50O-Il 0-02 0-020-07 0-17 0-100-23 0-80 1-010-01 0-80 1-180-27 9-71 16-13
0-13 0-10 0-260-00 O-Il 0-090-02 0-13 0-330-04 0-89 1-410-08 3-13 • 4-47
0-40 14-52 23-191-95 17-77 28-94
10-25 9-07 7-3050-32 33-71 20-5912-52 12-19 15-0971-00 55-62 41-9124-48 30-46 32-46
' As % dry weight of rotifers.
300 C. Rodriguez et al.
Table 4. Lipid content and fatty acid composition (% area) of total lipids of larvae in experiment 2
Fatty acids
12:014:014:115:115:0
16:016:1/1-916:1/1-716:1/1-517:017:1
18:018:1/1- 1118:1/1-918:1/1-718:2/1 - 919:018:2n - 618:3n - 918:3n - 620:018:3/1 - 318:4/1 - 318:4/1 - I
20:1/1 - 1120:ln - 920:2/1 - 920:2/1 - 620:4/1 - 620:3/1 - 320:4/1 - 320:5/1 - 3
22:1/1-1122:1/1 - 922:4/1 - 922:4/1 - 322:5/1 - 322:6/1 - 3
/I - 3 HUFA/ 1 - 3/ j - 6/ i - 9SaturatedMonoenoicsTotal lipids*/I - 3 HUFA*18:1/1-9//!-3 HUFA
Eggs
0-334-860-190-00063
21-743-069-350-300-420-18 ..
4-340-00
16-262-580-940-375-540-163-060-751-390-870-24
0-000-410-000-000-840-000-475-45
0-000-000-000-001-06
10-35
17-3319-599-44
20-9933-4432-1513-132-280-94
Initial larvae
Hatched
0-163-170-140-000-38
18-450-47
10-320-210-290-53
4-180-00
23-780-000-540-006-010-180-560-000-720-770-14
0-220-670-180-161-180-090-606-16
0-000-160-210-281-91
16-15
25-1926-68
7-9126-3726-6335-9730-831-110-94
3 day-old
0-513-450-000-000-41
15-020-563-49
11-240-570-71
1-860-00
16-612-920-610-007-280-003-950-000-821-040-00
0-000-000-000-001-640-000-478-15
0-820-000-000-001-39
16-02
26-0327-8912-8717-7821-8235-6426-76
6-970-64
Final
1
1-033-260-000-970-00
15-821-314-518-830-001-22
3-440-00
13-442-442-991-10
10-240-85
11-000-003-590-000-00
0-000-000-000-001-750-000-002-43
0-000-000-000-002-023-88
8-3311-9222-9919-4424-6531-5019-831-651-61
larvae of treatment
2
0-884-480-001-130-00
19-193-32
10-280-831-101-13
4-300-00
14-673-241-910-876-200-007-590-002-740-860-00
0-000-000-000-002-010-000-008-21
0-000-000-000-000-005-03
13-2416-8415-8019-9030-8233-4719-792-62M l
3
0-002-340-630-600-00
14-521-578-590-880-541-42
6-040-79
16-064-660-000-984-640-005-680-001-341-550-00
0-990-000-000-002-040-000-00
11-18
0-000-000-000-002-08
10-09
23-3526-2412-3617-6324-4234-7719-754-610-69
As % of dry weight of fish.
Essential fatty acids for larval Sparus aurata 301
On the other hand, n-6 fatty acids levels in final larvae decreased proportionally to then-3 HUFA contents in rotifers, mainly because of the reduction in the 18:3n-6 levels in fish,although this fatty acid remained almost constant in the rotifers. A negative correlation wasfound between the n-6 and n-3 fatty acid contents in the larvae, indicating an incorporationof n-3 fatty acids in the tissue lipids. The n-3 HUFA contents of larvae fed 0-07% n-2, HUFAwere very low (11-30%), half of that in the initial larvae (24-91%). There was a certainamount oill'-^n-Ti in these larvae although rotifers contained hardly any 22:6n-3. However,the n-3 HUFA levels in the other groups were increased by the elevation of n-3 HUFAcontents in rotifers. In larvae fed with rotifers containing the highest percentage of n-3HUFA, the levels of these fatty acids were similar to those of the initial larvae. When thelarval n-3 HUFA contents in both experiments were compared, it was observed that theelevation of n-3 HUFA levels in rotifers affects the n-3 HUFA content in fish.
The ratio of oleic acid to n-3 HUFA, known to be an indicator of the EFA deficiency infish, was also reduced by the elevation of the n-3 HUFA levels in rotifers.
Discussion and conclusions
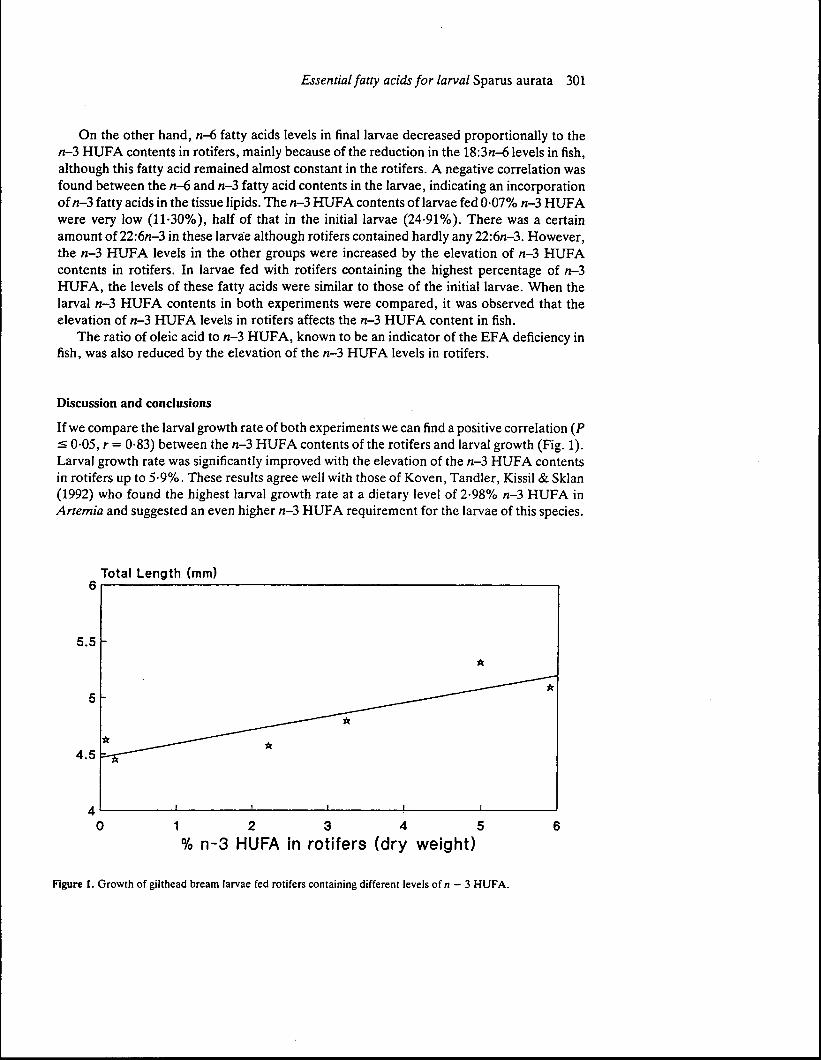
If we compare the larval growth rate of both experiments we can find a positive correlation (P:< 0-05, r = 0-83) between the n-3 HUFA contents of the rotifers and larval growth (Fig. 1).Larval growth rate was significantly improved with the elevation of the n-3 HUFA contentsin rotifers up to 5-9%. These results agree well with those of Koven, Tandler, Kissil & Sklan(1992) who found the highest larval growth rate at a dietary level of 2-98% n-3 HUFA inArtemia and suggested an even higher n-3 HUFA requirement for the larvae of this species.
D
5.5
5
4.5
A
Total Length (mm)
-
_ - - — — — — ^
— - — • A
1 1 r
1 2 3 4 5% n-3 HUFA in rotifers (dry weight)
Figure 1. Growth of gilthead bream larvae fed rotifers containing different levels of /i - 3 HUFA.

302 C. Rodriguez et al.
However, the results differ from the lower n-3 HUFA requirements (0-05% in rotifers)found by Koven etal. (1990) for larval gilthead bream. This difference could be explained bythe lower rotifer total lipid levels (11-1%) in that experiment compared with those of thepresent experiment (32-46% total lipids in rotifers). An increased requirement for dietaryEFA may occur as the lipid levels in the diet increase (Watanabe 1982). Takeuchi &Watanabe (1977) have found that the elevated dietary lipid levels increase the requirement ofrainbow trout, Oncorhynchus mykiss (Walbaum), for 18:3n-3, an EFA for this fish species.
Although survival was not so high as in our common rearing conditions (about 30%), thiscould be due to the lack of Nannochloropsis in the rearing tanks (Herndndez-Cruz,Femdndez-Palacios, Salhi, Vergara & Izquierdo in press).
The type of activity test used in these experiments has been extensively cited in theliterature (Kitajima et al. 1980; Watanabe, Kitajima & Fujita 1983; Izquierdo et al. 1989) buta more precise method to measure larval activity is necessary to correlate larval activity withn-3 HUFA deficiency in gilthead sea bream larvae.
No differences were found in the percentages of air inflated swim bladder between thetreatments, which may be associated with other factors (Kitajima, Tsukashiyama, Fujita,Watanabe & Yone 1981; Koven etal. 1990; Chatain 1982).
Although the 18:ln-9 levels in rotifers changed dramatically among treatments, becauseoleic acid was used to dilute the n-3 HUFA-rich oil, n-9 fatty acids in the larvae remainedalmost constant and even at a similar level to that in the initial larvae. These results suggestthat 18:1 n-9 may be selectively catabolized and used as an energy source during this period oflarval development.
The high levels of 18:3/i-6 in fish lipids, especially in larvae of treatment 1, suggests adesaturation process from the precursor 18:2/i-6 which was present at high concentrations inboth rotifers and larvae.
The results of this experiment have shown that there is a significant correlation betweenlarval growth and the n-3 HUFA levels in rotifers. Larval survival was significantly improvedby the elevation of the n-3 HUFA levels in rotifers. These levels also affected significantlyn-3 HUFA contents and oleic acid to n-3 HUFA ratio in the larvae. The occurrence ofhydrops in larval gilthead bream fed with very low levels of n-3 HUFA seems to be associatedwith an EFA deficiency in these fish.
Acknowledgments
We express here our sincere thanks to Mrs J.R. Cejas Pulido, Instituto Espafiol deOceanografia, for her valuable suggestions and for providing us with the necessary materialsfor experiment 1.
This study is part of the project 'Estudio de los requerimientos de ^cidos grasosesenciales de las larvas de dorada (Sparus aurata)' financially supported by GobiernoAut6nomo de Canarias.
References
Chatain B. (1982) Contribution k I'^tude de I'elevage larvaire de la dorade japonaise (Pagrus major). Thfese de 3°cycle, Univ. d'aix-Marseille II.
Falk-Petersen S., Sargent J.R., Fox C , Falk-Petersen I.-B., HaugT. & Kjorsvik E. (1989) Lipids in Atlantic Halibut{Hippoglossushippoglossus) eggs from planktonicsamples in Northern Norway. Marine Biology 101,553-556.

Essential fatty acids for larval Sparus aurata 303
Folch J., Lees M. & Stanley G.H.S. (1957) A simple method for the isolation and purification of total lipids fromanimal tissues. Journal of Biological Chemistry 226, 497-509.
Franicevic V., Lisac D., Buble J., L6ger Ph. & Sorgeloos P. (1987) International study on Artemia. XLIL The effectof the nutritional quality of Artemia on the growth and survival of sea bass (Dicentrarchus labrax L.) larvae in acommercial hatchery. In: Proceedings of the Conference on Production in Marine Hatcheries, Rovinj-Zadar(Yugoslavia) 10-28 February (1986). Ed. Loix B. Medrap, Tunis.
Gatesoupe F.J. & Le Milinaire C. (1985) Dietary value adaptation of live food organisms for covering the nutritionalrequirements of marine fish larvae. .Coll. Fr. -Jp. Oceanogr.- Marseille 16-21 Sept. 85, 8, 51-63.
Herndndez-Cruz CM., Ferndndez-Palacios H., Salhi M., Vergara J.M. & Izquierdo M.S. (in press) Nutritionalvalue of rotifers fed different enrichment diets for gilthead seabream larvae. Informes Ticnicos de ScientiaMarina (in press).
Izquierdo M.S., Watanabe T., Takeuchi T., Arakawa T. & Kitajima C. (1989) Requirement of larval Red SeaBream Pagrus major for essential fatty acids. Nippon Suisan Gakkaishi 55(5), 859-867.
Izquierdo M.S., Watanabe T., Takeuchi T., Arakawa T. & Kitajima C. (1990) Optimal EFA levels in Artemia tomeet the EFA requirements of Red Seabream ( Pagrus major). In: Proceedings Third International Symposiumon Feeding and Nutrition in Fish, Toba, August 28-September 1, Japan, 1989 (ed. by H. Takeda & I.Watanabe), pp. 221-232. Japan Translation Centre, Tokyo.
Izquierdo M.S., Arakawa T., Takeuchi T., Haroun R. & Watanabe T. (1992) Effect of n-3 HUFA levels in Artemiaon growth of larval Japanese Flounder {Paralicthys olivaceus). Aquaculture 105, 733-82.
Kimata M. (1983) Changes of fatty acid composition during early development in red sea bream Chrysophys major(Temminck et Shiegel) egg and larva. Tokay Daigaku Kayo Gakubu 16, 213-223.
Kitajima C , Arakawa T., Fujita S., Imada C , Watanabe T. & Yone Y. (1980) Dietary value for red sea breamlarvae of rotifer Brachionus plicatilis cultured with a new type of yeast. Nippon Suisan Gakkaishi 46, 43-46.
Kitajima C , Tsukashiyama Y., Fujita S., Watanabe T. & Yone Y. (1981) Relationship between uninflated swimbladders and lordic deformity in hatchery reared seabream, Pagrus major. Nippon Suisan Gakkaishi 47,1289-1294.
Koven W. M.. Kissil G. W. & Tandler A.(1989) Lipid and n-3 requirement of Sparus aurata larvae during starvationand feeding. Aquaculture 79, 185-191.
Koven W.M., Tandler A., Kissil G.Wm., Sklan D., Friezlander O. & Harel M. (1990) The effect of dietary (n-3)polyunsaturated fatty acids on growth, survival and swim bladder development in Sparus aurata larvae.Aquaculture 9\, 131-141.
Koven W.M., Tandler A., Kissil G.Wm. & Sklan D. (1992) The importance of n-3 highly unsaturated fatty acids forgrowth in larval Sparus aurata and their effect on survival, lipid composition and size distribution. Aquaculture104,91-104.
Le Milinaire C , Gatesoupe F.J. & Stephan G. (1983) Approche du besoin quantitative en acides ras longspolyinsatur^es de la series n-3 chez la larve du Turbot {Scophthalmus maximus). Comptes rendus, Academiedes Sciences, Paris 2(111), 917-920.
Metcalfe L.D. & Schmitz A.A. (1961) The rapid preparation of fatty acid esters for gas chromatographic analysis.Analytical Chemistry 33, 363-364.
Mourente G. & Odriozola J.M. (1990) Effect of broodstock diets on total lipids and fatty acid composition of larvaeof gilthead seabream {Sparus aurata L.) during yolksac stage. Fish Physiology and Biochemistry 8, 103-110.
Ostrowski A.C. & Divakaran S. (1990) Survival and bioconversion of n-3 fatty acids during early development ofdolphin {Coryphaena hippurus) larvae fed oil-enriched rotifers. Aquaculture 89, 273-285.
Rubio E.A.R. (1986) Evolution de la composition au acides gras de I'ovocyte de la larve du turbot (fjeHa maximaL.) en fonction du regime alimentaire des reproducteurs et des larves, ainsi que la temperature d'incubation.Th^se de 3° cycle, Univ. Bretagne Occidentale, 183 pp.
Scott A.P. & Baynes S.M. (1978) Effect of algal diet and temperature on the biochemical composition of the rotiferBrachionus plicatilis. Aquaculture 14, 247-260.
Scott A.P. & Middleton C. (1979) Unicellular algae as food for turbot larvae {Scophthalmus maximus L.). Theimportance of dietary long chain polyunsaturated fatty acids. Aquaculture 18, 227-240.
Takeuchi T. & Watanabe T. (1977) Dietary levels of methyl laurate and essential fatty acid requirement of rainbowtrout. Bulletin of the Japanese Society of Scientific Fisheries 43, 893-898.
Tocher D.R.&SargentJ.R. (1984) Analysis of lipids and fatty acids in ripe roes of some northwest European marinefish. Lipids 19, 492-499.
Tocher D.R., Fraser A.J., Sargent J.R. & Gamble J.C. (1985) Lipid class composition during embryonic and earlylarval development in Atlantic Herring {Clupea harengus L.). Lipids 20(2), 84-89.
Watanabe T. (1982) Lipid nutrition in fish. Comparative Biochemistry and Physiology 73B, 3-15.

304 C. Rodriguez et al.
Watanabe T., Arakawa T., Kitajima C , Fukusho K. & Fujita S. (1978a) Proximate and mineral compositions ofliving feeds used in seed production of fish. Nippon Suisan Gakkaishi 44, 979-984.
Watanabe T., Arakawa T., Kitajima C. &FujitaS. (1978b) Nutritionalevaluationof proteins of living feeds used inseed production of fish. Nippon Suisan Gakkaishi 44, 985-988.
Watanabe T., Kitajima C , Arakawa T., Fukusho K. & Fujita S. (1978c) Nutritional quality of rotifer Brachionusplicatilis as a living feed from the viewpoint of essential fatty acids for fish. Nippon Suisan Gakkaishi 44,1109-1114.
Watanabe T., Oowa F., Kitajima C., Fujita S. & Yone Y. (1979) Relationship between the dietary value of rotifersBrachionus plicatilis and their content of w3 highly unsaturated fatty acids. Nippon Suisan Gakkaishi 45,883-889.
Watanabe T.jOhtaM., Kitajima C. & Fujita S. (1982) Improvement of dietary value of brine shrimp/4rtemiajfl/inafor fish larvae by feeding them on w3 highly unsaturated fatty acids. Nippon Suisan Gakkaishi 48,1775-1782.
Watanabe T., Kitajima C. & Fujita S. (1983) Nutritional values of live organisms used in Japan for mass propagationof fish: a review. Aquacutture 34, 115-143.
Watanabe T., Izquierdo M.S., Takeuchi T., Satoh S. & Kitajima C. (1989) Comparison between eicosapentaenoicand docosahexaenoic acids in terms of essential fatty acid efficacy in larval red sea bream. Nippon SuisanGakkaishi 55(9), 1635-1640.
Yamashita K. (1981) Hydrops of larval red sea bream Pagrus major. Japanese Journal of Ichthyology 28, 80-85.






























