Embed Size (px)
Citation preview

Comparative Biochemistry and Physiology Part B 135(2003) 449–460
1096-4959/03/$ - see front matter� 2003 Elsevier Science Inc. All rights reserved.doi:10.1016/S1096-4959Ž03.00111-8
Cloning and nutritional regulation of aD6-desaturase-like enzyme inthe marine teleost gilthead seabream(Sparus aurata)�
I. Seiliez*, S. Panserat, G. Corraze, S. Kaushik, P. Bergot
Laboratory of Fish Nutrition, Unite mixte INRA-IFREMER, 64310 St-Pee-sur-Nivelle, France´ ´
Received 2 January 2003; received in revised form 9 April 2003; accepted 9 April 2003
Abstract
Marine fish are presumed to have a lower capacity than freshwater fish for the bioconvertion of 18C fatty acids into20–22C highly unsaturated fatty acids(HUFA). The present work investigated the first step of this pathway, theD6-desaturation, in gilthead seabream. A full-length desaturase-like cDNA was identified from total RNA extracted fromviscera of juvenile fish fed for 96 days on an experimental HUFA-free diet containing olive oil as the sole lipid source.The open reading frame encodes a 445-amino acid peptide that contains two membrane-spanning domains, threehistidine-rich regions, and a cytochromeb domain, which are characteristic ofD6- andD5-desaturases. Predicted protein5
sequence of seabream desaturase-like indicated a high percentage of identity with mammalianD6-desaturases(approx.65%). Northern analysis showed two transcripts of approximately 3.7 and 1.8 kb which were highly expressed in fishfed on HUFA-free diet and slightly expressed in fish fed on HUFA-rich diet. The fatty acid profile of the former groupwas characterized by high levels ofD6-desaturation products(18:2 n-9 and 20:2 n-9) with no detectable levels ofD5-desaturation product(20:3n-9). These results demonstrate for the first time the presence and nutritional modulation of aD6-desaturase-like cDNA in a marine fish.� 2003 Elsevier Science Inc. All rights reserved.
Keywords: Cloning; Desaturase; Fish; Gene expression; HUFA; Lipid; Nutrition; Teleost
1. Introduction
Fish, like mammals, require a dietary supply ofessential fatty acids of the n-3 and n-6 series thatthey are unable to synthesize(Watanabe, 1982;Sargent et al., 2002). Among teleosts, freshwaterspecies differ from marine species as regards theirfatty acid requirements(Sargent et al., 2002). The
� Genbank accession number for the gilthead seabreamD6-desaturase-like cDNA sequence is AY055749, and Genbankaccession number for the gilthead seabream partialD6-desa-turase-like gene sequence is AF525692*Corresponding author. Tel.:q33-5-59-51-59-51; fax:q
33-5-59-51-54-52.E-mail address: [email protected](I. Seiliez).
former have a recognized capacity to desaturate—elongate the fatty acids with 18 carbon atoms,linoleic (18:2n-6) and linolenic (18:3n-3) acids,into highly unsaturated fatty acids(HUFA) with20 and 22 carbon atoms such as arachidonic(AA),eicosapentaenoic(EPA) and docosahexaenoic(DHA) acids (Henderson and Tocher, 1987; Sar-gent et al., 2002). These HUFA play a major rolein eicosanoid production(AA and EPA), vision(DHA), brain development(DHA) and regulationof expression of several genes involved in lipidmetabolism(Cook, 1991; Spiegelman and Flier,1996; Clarke et al., 1997; Forman et al., 1997). Incontrast to freshwater fish, marine fish are assumedto have a deficient capacity to bioconvert 18C

450 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Table 1Formulation of the experimental diet
Ingredient HUFA(%) Free diet
Caseina 50Casein hydrolysateb 5Dextrinc 21Olive oild 10Mineral mixe 5Vitamin mixf 5Arginineg 2.5Methionineh 0.5Binderi 1
Casein. Prolabo nr. 22544292.a
Casein hydrolysate. Sigma C0626.b
Dextrin. Sigma D2256.c
Olive oil. Pujet, France.d
Mineral mix. National Research Council(1993).e
Vitamin mix. National Research Council(1993).f
Arginine (L-Arg-HCI). Sigma NA-5131.g
L-Methionine. Ajinomoto nr. 78350.h
Binder. Agar–agar. Prolabo nr. 20768235.i
precursors into HUFA and hence, require the pres-ence of preformed HUFA in their diet(Owen etal., 1975; Kanazawa et al., 1978; Henderson andTocher, 1987; Mourente and Tocher, 1994; Buzziet al., 1996; Sargent et al., 2002). Although longrecognized, the reasons for this deficiency inmarine species remain unclear. On the one hand,it could be due to a gene loss corresponding to anadaptation to the n-3 HUFA-rich marine food web,as also suggested for terrestrial carnivores(Sargentet al., 1995). On the other hand, the deficiency ofD6- and D5-desaturase activity in marine fishcould be due to a repression of desaturase activityby the high levels of HUFA usually present in thediet (Olsen et al., 1990). Inhibition of desaturasegene expression by dietary HUFA is known inmammals(Cho et al., 1999a,b) and has also beenobserved for theD6-desaturase-like gene of rain-bow trout(Seiliez et al., 2001).Following the cloning of D6-desaturase-like
cDNA in rainbow trout (Seiliez et al., 2001),cDNA sequences of several freshwater fish specieshave been reported(Danio rerio AF309556,Cypri-nus carpio AF309557, Oncorhynchus masouAB074149 and AB070444,Oreochromis niloticaAB069727, andSalmo salar AF478472), but noinformation is available about desaturase cDNApresence in a marine teleost. The present workaimed at cloning aD6-desaturase in a marineteleost, the gilthead seabream, in whichD6- andD5-desaturase activities have been reported in vivo(Mourente and Tocher, 1993, 1994, 1998) and incell culture(Tocher and Ghioni, 1999).
2. Materials and methods
2.1. Experimental fish and diet
Gilthead seabream(mean wet wt.s0.5 g) weretransported from a commercial hatchery(FermeMarine de Douhet, La Bree-les-Bains, France) to´the experimental INRA facility(Biarritz, France).They were reared for 96 days, at 208C, in 50-laquaria supplied with running seawater in a semi-closed system equipped with a pump filter andUV lamps. The photoperiod was regulated as a 12hy12 h dark–light cycle. An experimental dry diet(Table 1) was continuously delivered for 12 hy24h by a belt dispenser, to excess(approx. 10% offish biomass per day). This diet was formulatedwith purified ingredients and contained olive oil,rich in oleic acid(18:1n-9) and devoid of HUFA,
as the sole lipid source. This HUFA-free diet wasexpected to facilitate cloning by preventing orlimiting the inhibition of desaturase gene expres-sion by HUFA(Cho et al., 1999a).
2.2. Sampling and analysis
Fish were sampled, after a 24 h fasting period,at the beginning(initial fish) and at the end(HUFA deficient fish) of the rearing period.Following anesthesia with phenoxy-2-ethanol(1:3333), some fish were directly frozen(y80 8C) for analyses of total body lipids, andothers were dissected in two parts: viscera, expect-ed to be an active site of expression of desaturases,and muscle(posterior trunk without head). Tissueswere frozen in liquid nitrogen and stored aty80 8C. In addition, samples of fish fed on HUFA-rich commercial diet(control fish) with the samefinal body weight(1.5 g) as the fish fed on HUFAfree diet were collected in the fish farm and treatedin the same way.Total lipids from the HUFA-free diet and from
fish were extracted and gravimetrically measuredaccording to Folch et al.(1957) by using dichlo-romethane instead of chloroform as solvent. For abetter detection of fatty acids indicating desatura-tion, polar and neutral lipids of viscera wereseparated by elution on a silica column(Sep Paksilica cartridge, Millipore) according to Juanedaand Rocquelin(1985). Fatty acid methyl esters

451I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Table 2Primers used for the desaturase cDNA cloning by RT-PCR and RACE-PCR, for partial desaturase gene cloning by PCR and fordesaturase gene expression analysis by RT-PCR
Step Primer
Partial cDNA cloningRT-PCR Forward: 59–39 (ACCTGGGARGAGGTSCAGAA)
Reverse: 59–39 (TKGAGRTACGCATCCAGCCA)
Full-length cDNA cloningRACE-PCR 59RACE: 59–39 (GCTCTCCAATCAGCAACGGCTTCAG)
39RACE: 59–39 (DCCTTCAACGACTGGTTCAG)
Full-length ORF cloningRT-PCR Forward: 59–39 (ATGGACAGTGAACGCATCGG)a
Reverse: 59–39 (ACCATACCACTGCTGTTCAG)a
Partial gene cloningPCR1 Forward: 59–39 (AYCARCACMAGTACTTCTTC)
Reverse: 59–39 (GTSACCCANACAAACCAGTT)
PCR 2 Forward: 59–39 (AGCCACTGGTTTGTGTGGGT)Reverse: 59–39 (TKGAGRTACGCATCCAGCCA)
Gene expression analysisRT-PCR for the desaturase Forward: 59–39 (AYCARCACMAGTACTTCTTC)
Reverse: 59–39 (TCRATCTGRAAGTTGAGGTG)
RT-PCR for theb-actin Forward: 59–39 (ATCCTGACCCTGAAGTACCC)a
Reverse: 59–39 (CCATCCTTCCTCGGTATGGA)a
Gilthead seabream-specific desaturase primers; DsAyGyT; RsAyG; SsGyC; KsGyT; YsCyT; M-A yC, NsAyGyCyT.a
from total, polar and neutral lipids were preparedby acid-catalyzed transmethylation of total lipidsaccording to Shantha and Ackman(1990) andanalyzed using a Varian 3400 gas chromatographequipped with a DB Wax fused silica capillarycolumn(30 m=0.25 mm i.d., film thickness: 0.25mm, J&W Scientific, Folsom, CA, USA). Heliumwas used as carrier gas(1.4 mlymin). The thermalgradient was 100–1808C at 8 8Cymin, 180–2208C at 4 8Cymin, and then a constant temperatureof 220 8C during 25 min. Injector and flameionization detector temperatures were 2608C and250 8C, respectively. Fatty acid methyl esters wereidentified by comparison with known standardmixtures (Sigma, France) and quantified using acomputer.
2.3. RNA isolation, poly(A) mRNA purificationand reverse transcription
Total RNA was extracted from gilthead sea-bream tissues as described by Chomczinski andSacchi(1987). cDNA was obtained by annealing2 mg of total RNA with 1mg of random primersand incubating with AMV reverse transcriptase(Boehringer, Roche Molecular Biochemicals, Ger-many) at 42 8C for 1 h.
2.4. Cloning of the partial gilthead seabreamdesaturase cDNA
D6-Desaturase sequences from human(Genbankaccession number AF126799), rat (Genbankaccession number AB021980), mouse(Genbankaccession number AF126798), zebrafish(Genbankaccession number AF309556), carp (Genbankaccession number AF309557), and rainbow trout(Genbank accession number AF301910) werecompared using the Clustal-W multiple alignmentalgorithm (Higgins and Sharp, 1989). Primerswere chosen corresponding to the most conservedcoding regions ofD6-desaturase at positions 64–83 and 1309–1328 of the human gene. Thesequences of primers are shown in Table 2. RT-PCR was performed on visceral total RNA extract-ed from fish fed on HUFA-free diet. cDNA(2 ml)was amplified by PCR using 100 pmol of theprimers, in a reaction mixture containing 2 mMMgCl , 50 mM KCl, 20 mM Tris–HCl, 0.25 mM2
dNTP, and 2.5 U of Taq polymerase(Boehringer,Roche Molecular Biochemicals, Germany). Thirty-five cycles of denaturation at 948C for 20 s,annealing at 578C for 20 s, and extension at 728C for 30 s were performed. PCR products were

452 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
subjected to electrophoresis in 1% agarose gelsand fragments of the expected size range werepurified (Micropure System, Amicon, USA). Thepurified DNA fragments were inserted into thepCR�II-TOPO plasmid and used for transforma-tion of One Shot� competent cells(Invitrogen,Carlsbad, CA, USA). Inserts were detected byEcoRI digestion of the extracted plasmid DNA.Clones with inserts were sequenced(Cybergene,`Evry, France).
2.5. Reverse transcription (RT), rapid amplifica-tion of the cDNA ends-polymerase chain reaction(RACE-PCR) and molecular cloning of PCRfragments
The 59 and 39 cDNA extremities were deter-mined by the RACE-PCR method as detailed inthe manufacturer protocol(Gene Racer kit, Invi-trogen, USA). Using the reverse transcription sys-tem, cDNA was synthesized by incubating 1mgof hepatic polyA mRNA from fish fed on acomplete practical diet, with AMV reverse tran-scriptase at 428C for 1 h using either the oligodTprimer (39RACE) or a primer chosen fromsequence information of the cloned partial giltheadseabream desaturase cDNA(59RACE) (Table 2).The RACE-PCR reactions were carried out at 558C (39RACE) and 59 8C (59RACE) annealingtemperatures using primers designed from partialdesaturase sequence data(Table 2). PCR productswere then cloned and sequenced as describedabove.
2.6. Cloning of the full length gilthead seabreamdesaturase open reading frame (ORF)
A cDNA fragment of 1.7 kb comprising the fulllength gilthead seabream desaturase ORF wascloned and sequenced. The method was asdescribed above and the sequences of primers arealso shown in Table 2.
2.7. Cloning of the partial gilthead seabreamdesaturase gene
As an additional control, gilthead seabreamdesaturase-like gene was partially cloned. Thegenomic DNA was extracted from gilthead sea-bream liver using the Wizard Genomic DNA�
purification kit (Promega, Madison, USA). For thedesaturase gene cloning, the method was as
described above using exonic primers(Table 2) at53 8C (PCR1) and 57 8C (PCR2) annealingtemperatures. The two PCR reactions were per-formed showing nucleotide sequence overlap inexon 9.
2.8. Sequence analysis
Nucleotide sequences(excluding the primersequences) were compared with DNA sequencesfrom the Genbank database using the basic localalignment search tool (BLAST) algorithm(Altschul et al., 1990). Sequence alignments andpercentage of amino acid conservation wereassessed with the Clustal-W multiple alignmentalgorithm (Higgins and Sharp, 1989) using thecloned gilthead seabream sequence and vertebrateD6 and D5-desaturase sequences. These aminoacid sequence alignments were used to construct aphylogenetic tree with Fitch–Margoliash and LeastSquares Methods with Evolutionary Clock(KITSCH). Analysis was bootstrapped with 1000replicates using the Phylogeny Inference Package(PHYLIP) version 3.5c.
2.9. Gilthead seabream desaturase gene expres-sion: Northern analysis
Twenty micrograms of extracted total RNA sam-ples were electrophoresed in 1% agarose gelscontaining 5% formaldehyde and transferred ontonylon membrane(Hybond-N , Amersham, UK).q
After transfer, RNA blots were stained with meth-ylene blue to locate 28S and 18S rRNAs and toascertain the amount of loaded RNAs. Membraneswere hybridized withw Px-DNA probes labeled32
by random priming(Stratagene, USA) recognizingthe partial gilthead seabream desaturase cDNAdescribed above. Membranes were then washedonce for 20 min at 358C with 2=SSC, 0.1%SDS, once for 20 min at 298C with 1=SSC,0.1% SDS and finally, for 15 min at room temper-ature with 0.2=SSC, 0.1% SDS. After washing,the membranes were exposed to X-ray film.
2.10. Gilthead seabream desaturase gene expres-sion: RT-PCR analysis
cDNA synthesized from viscera and muscle offish fed on a diet lacking HUFA were amplifiedby PCR using specific primers chosen for theseabream desaturase andb-actin (positive control)

453I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Table 3Total lipid content(% wet wt.) and fatty acid composition(percentage of total fatty acid) of the HUFA-free diet and fish(whole body)
HUFA-free diet Initial fish HUFA deficient fish Control fish
Total lipid 7.8"0.1 4.4"0.1b 7.0"0.1a 6.2"0.4a
Fatty acidsC14:0 0.2"0.1 6.6"1.1a 3.8"0.9a 6.8"2.3a
C16:0 13.3"0.6 21.6"0.7a 14.8"1.2a 20.5"3.3a
C18:0 3.2"0.2 3.4"0.1a 2.4"0.2b 2.8"0.1a
C20:0 0.3"0.1 0.2"0.0a 0.1"0.0a 0.2"0.0a
Total saturated 17.0"0.3 31.8"1.7a 21.2"1.9a 30.3"5.0a
C16:1 n-7 1.1"0.1 6.7"0.1a 9.4"0.9a 8.6"1.7a
C18:1 n-9 72.5"0.3 17.4"1.8b 41.5"1.7a 20.1"0.1b
C18:2 n-9 nd ndb 13.8"0.1a ndb
C20:2 n-9 nd ndb 0.5"0.1a ndb
C20:3 n-9 nd nd nd ndTotal n-9 FA 72.5"0.3 17.4"1.8b 55.9"1.9a 20.1"0.1b
C18:2 n-6 4.9"0.1 3.3"0.2a 2.6"0.1b 3.4"0.1a
C18:3 n-6 nd 0.1"0.1b 0.8"0.0a tb
C20:3 n-6 t 0.1"0.0a 0.1"0.0a 0.1"0.0a
C20:4 n-6 nd 0.7"0.0a 0.2"0.0b 0.5"0.1a
Total n-6 FA 4.9"0.1 4.2"0.1a 3.3"0.8a 4.1"0.1a
C18:3n-3 0.6"0.0 0.7"0.1b 0.3"0.0c 0.9"0.0a
C18:4n-3 nd 1.5"0.3a 0.2"0.0b 1.7"0.0a
C20:5 n-3 t 5.6"0.0a 0.7"0.1b 5.4"0.8a
C22:5 n-3 nd 0.8"0.1a 0.1"0.0b 0.9"0.4a
C22:6 n-3 nd 10.0"1.0b 1.6"0.4b 10.6"3.8a
Total n-3 FA 0.6"0.0 18.6"0.6a 2.9"0.5b 19.6"5.0a
Data are means"S.D. with ns2 (two samples from a pool of 15 fish). S.D.s0.0 implies S.D.-0.05. Values in the same rowfollowed by different letters are significantly different(P-0.05). Nd, not detected;t, trace(-0.05%).
cDNA sequences(Table 2). The PCR reaction wascarried out in a final volume of 25ml containing1.5 mM MgCl and 4 pmol of each primer, 2ml2
cDNA and 1 U of Taq polymerase(Boehringer,Roche Molecular Biochemicals, Germany). Num-ber of cycles was 35, each consisting of 20 s forhybridization (at 52 8C for the desaturase and at57 8C for theb-actin), 30 s for elongation(at 728C) and 20 s for denaturation(at 94 8C).
2.11. Data analysis
The results of lipid contents and fatty acidprofiles are expressed as mean"standard deviation(S.D.). Diet-related differences were analyzedusing one-way ANOVA. The Newman–Keuls mul-tiple range test was used to compare means whena significant difference was found. All the statis-tical analyses were performed with the computingprogramSTAT-ITCF (ITCF, 1988) and differenceswere considered significant whenP-values were-0.05.
3. Results
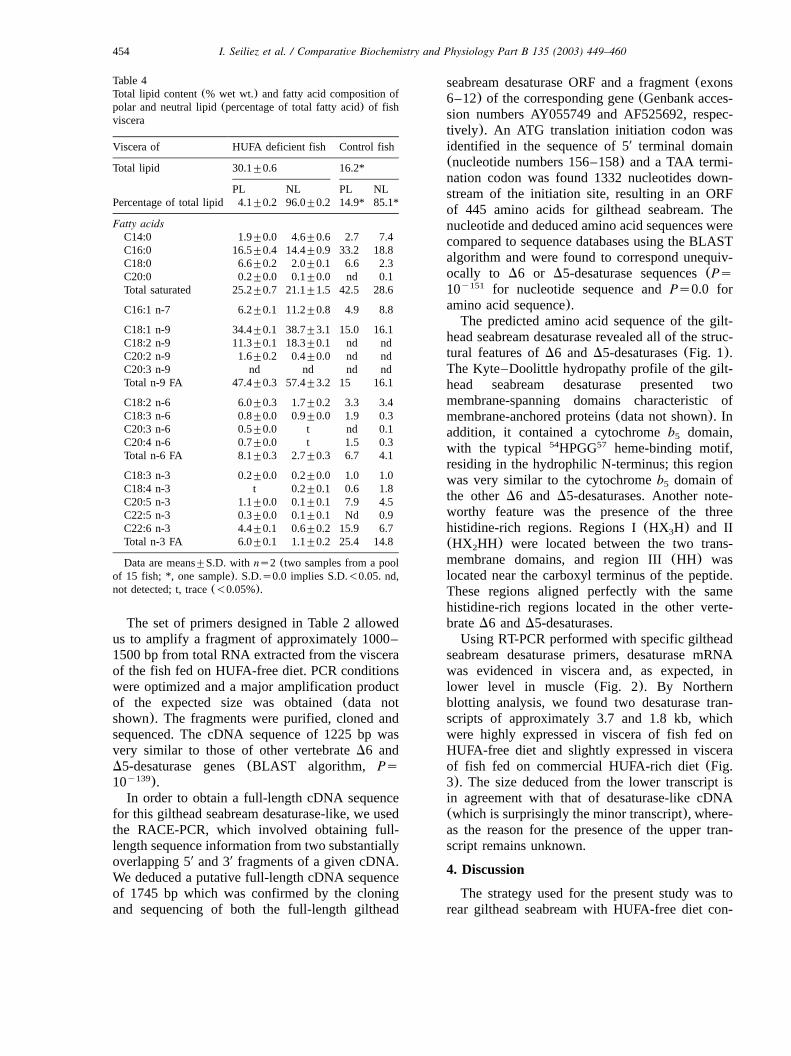
Feeding on HUFA-free diet resulted in markedchanges of the fatty acid profile of deficient fish(Table 3). Compared to initial and control fish feda HUFA rich diet, HUFA deficient fish showedhigher levels of 18:1n-9(17.4% and 20.1% oftotal fatty acids vs. 41.5%, respectively) anddecreased levels of n-3 FA(18.6% and 19.6% oftotal fatty acids vs. 2.9%, respectively). Levels ofEPA were more depressed than those of AA andDHA as indicated by the changes in the AAyEPAratio (from 0.1 to 0.3) and the EPAyDHA ratio(from 0.6 to 0.4). Fish fed on HUFA-free dietshowed also higher levels of 18:3n-6 and notice-able levels of 18:2n-9 and 20:2n-9 in body totallipids (13.8 and 0.5% of total fatty acids) whichwere absent in initial fish and in the deficient diet.These two fatty acids were also prominent both inpolar lipids (11.3 and 1.6%) and neutral lipids(18.3 and 0.4%) of viscera in HUFA-deficient fish(Table 4). In contrast, no 20:3n-9 was detected inthe same fish(Tables 3 and 4).
454 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Table 4Total lipid content(% wet wt.) and fatty acid composition ofpolar and neutral lipid(percentage of total fatty acid) of fishviscera
Viscera of HUFA deficient fish Control fish
Total lipid 30.1"0.6 16.2*
PL NL PL NLPercentage of total lipid 4.1"0.2 96.0"0.2 14.9* 85.1*
Fatty acidsC14:0 1.9"0.0 4.6"0.6 2.7 7.4C16:0 16.5"0.4 14.4"0.9 33.2 18.8C18:0 6.6"0.2 2.0"0.1 6.6 2.3C20:0 0.2"0.0 0.1"0.0 nd 0.1Total saturated 25.2"0.7 21.1"1.5 42.5 28.6
C16:1 n-7 6.2"0.1 11.2"0.8 4.9 8.8
C18:1 n-9 34.4"0.1 38.7"3.1 15.0 16.1C18:2 n-9 11.3"0.1 18.3"0.1 nd ndC20:2 n-9 1.6"0.2 0.4"0.0 nd ndC20:3 n-9 nd nd nd ndTotal n-9 FA 47.4"0.3 57.4"3.2 15 16.1
C18:2 n-6 6.0"0.3 1.7"0.2 3.3 3.4C18:3 n-6 0.8"0.0 0.9"0.0 1.9 0.3C20:3 n-6 0.5"0.0 t nd 0.1C20:4 n-6 0.7"0.0 t 1.5 0.3Total n-6 FA 8.1"0.3 2.7"0.3 6.7 4.1
C18:3 n-3 0.2"0.0 0.2"0.0 1.0 1.0C18:4 n-3 t 0.2"0.1 0.6 1.8C20:5 n-3 1.1"0.0 0.1"0.1 7.9 4.5C22:5 n-3 0.3"0.0 0.1"0.1 Nd 0.9C22:6 n-3 4.4"0.1 0.6"0.2 15.9 6.7Total n-3 FA 6.0"0.1 1.1"0.2 25.4 14.8
Data are means"S.D. with ns2 (two samples from a poolof 15 fish; *, one sample). S.D.s0.0 implies S.D.-0.05. nd,not detected; t, trace(-0.05%).
The set of primers designed in Table 2 allowedus to amplify a fragment of approximately 1000–1500 bp from total RNA extracted from the visceraof the fish fed on HUFA-free diet. PCR conditionswere optimized and a major amplification productof the expected size was obtained(data notshown). The fragments were purified, cloned andsequenced. The cDNA sequence of 1225 bp wasvery similar to those of other vertebrateD6 andD5-desaturase genes(BLAST algorithm, Ps10 ).y139
In order to obtain a full-length cDNA sequencefor this gilthead seabream desaturase-like, we usedthe RACE-PCR, which involved obtaining full-length sequence information from two substantiallyoverlapping 59 and 39 fragments of a given cDNA.We deduced a putative full-length cDNA sequenceof 1745 bp which was confirmed by the cloningand sequencing of both the full-length gilthead
seabream desaturase ORF and a fragment(exons6–12) of the corresponding gene(Genbank acces-sion numbers AY055749 and AF525692, respec-tively). An ATG translation initiation codon wasidentified in the sequence of 59 terminal domain(nucleotide numbers 156–158) and a TAA termi-nation codon was found 1332 nucleotides down-stream of the initiation site, resulting in an ORFof 445 amino acids for gilthead seabream. Thenucleotide and deduced amino acid sequences werecompared to sequence databases using the BLASTalgorithm and were found to correspond unequiv-ocally to D6 or D5-desaturase sequences(Ps10 for nucleotide sequence andPs0.0 fory151
amino acid sequence).The predicted amino acid sequence of the gilt-
head seabream desaturase revealed all of the struc-tural features ofD6 andD5-desaturases(Fig. 1).The Kyte–Doolittle hydropathy profile of the gilt-head seabream desaturase presented twomembrane-spanning domains characteristic ofmembrane-anchored proteins(data not shown). Inaddition, it contained a cytochromeb domain,5
with the typical HPGG heme-binding motif,54 57
residing in the hydrophilic N-terminus; this regionwas very similar to the cytochromeb domain of5
the otherD6 and D5-desaturases. Another note-worthy feature was the presence of the threehistidine-rich regions. Regions I(HX H) and II3
(HX HH) were located between the two trans-2
membrane domains, and region III(HH) waslocated near the carboxyl terminus of the peptide.These regions aligned perfectly with the samehistidine-rich regions located in the other verte-brateD6 andD5-desaturases.Using RT-PCR performed with specific gilthead
seabream desaturase primers, desaturase mRNAwas evidenced in viscera and, as expected, inlower level in muscle (Fig. 2). By Northernblotting analysis, we found two desaturase tran-scripts of approximately 3.7 and 1.8 kb, whichwere highly expressed in viscera of fish fed onHUFA-free diet and slightly expressed in visceraof fish fed on commercial HUFA-rich diet(Fig.3). The size deduced from the lower transcript isin agreement with that of desaturase-like cDNA(which is surprisingly the minor transcript), where-as the reason for the presence of the upper tran-script remains unknown.
4. Discussion
The strategy used for the present study was torear gilthead seabream with HUFA-free diet con-

455I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Fig. 1. Alignment of the deduced amino acid sequences from gilthead seabream desaturase-like cDNA to humanD6-desaturase(GenBank Accession no. AF126799) and humanD5-desaturase(GenBank Accession no. AF199596). Stars correspond to the aminoacids conserved between the desaturases compared. The putative cytochromeb domain is underlined. Putative transmembrane domains5
are shown in shaded areas, and three putative histidine-rich regions are in bold.
taining olive oil as sole lipid source in order toprevent the inhibition of desaturase gene expres-sion by HUFA. As expected, feeding an oleic acidbased diet resulted in a decreased level of n-3HUFA and the appearance of n-9 polyunsaturatedfatty acids, typical of essential fatty acid deficien-cies(Sargent et al., 1995; Buzzi et al., 1996). Thefatty acid profile obtained was very similar to thatof cells of gilthead seabream cultured in fetal calfserum having low levels of n-6 and n-3 HUFAand high levels of 18:2n-9(Tocher and Ghioni,1999). High levels of 18:2n-9 and 20:2n-9 were
also reported in juvenile grey mullet(initialweights0.7 g) fed on a fat-free diet for 84 daysin sea water, 18:2n-9 being higher in neutral(11.7% total fatty acids) than in polar lipids(8.5%) and 20:2n-9 showing the reverse order(3.1and 4.6%, respectively) (Argyropoulou et al.,1992). A similar preferential distribution wasnoticed presently in gilthead seabream viscerallipids. The high levels of 18:2n-9 can be inter-preted as the consequence of an importantD6-desaturase activity in fish fed on deficient dietsand in gilthead seabream cells cultured in HUFA-

456 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Fig. 2. Tissue specificity of desaturase-like andb-actin gene expression in gilthead seabream fed on HUFA-free diet. Analysis by RT-PCR.y, negative control(PCR without cDNA); WM, molecular weight marker, 5–2.000 bp(Sigma, Missouri, USA). The exactlengths of the amplified fragments were determined following the known desaturase-like andb-actin (Genbank accession numberAF384096) cDNA sequences.
Fig. 3. Desaturase-like gene expression in viscera of gilthead seabream fed on diet with(control fish) or without(HUFA deficient fish)HUFA. Desaturase-like mRNA levels were determined by Northern analysis(20mg total RNA per lane). 18S rRNA served as internalcontrol of sample loading.
deficient medium(Tocher and Ghioni, 1999). Theincrease of 18:3n-6 might be a further indicationof a D6-desaturase activity in these fish. Noconclusion can be drawn about the formation of18:4n-3 due to the relatively high level of thisfatty acid in initial fish and to the negligible levelof 18:3n-3 in the HUFA-free diet. The presence of20:2n-9 attests that 18:2n-9 is elongated. Absenceof 20:3n-9, despite the presence of precursor20:2n-9, suggests that no detectableD5-desaturaseactivity occurred in the experimental fish studied.Possibly, longer deficiency period or differentnutritional conditions would be necessary to clear-ly demonstrate theD5-desaturase activity. Such an
activity has been reported in gilthead seabream,both in vivo and in vitro, albeit being low com-pared to D6-desaturase activity(Mourente andTocher, 1993; Tocher and Ghioni, 1999).At the end of the growth period, RT-PCR was
performed on total RNA extracted from the visceraof fish fed on HUFA-free diet and a fragmentencoding a 445 amino acid peptide was cloned.The high similarity(from 57% to 65%) betweenthe gilthead seabream peptide sequence and thecorresponding sequences fromD5- andD6-desa-turase gene family previously characterized inother vertebrates is a strong indication that thissequence corresponds to a desaturase(Fig. 4). The

457I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Fig. 4. Phylogenetic relationship between the amino acid sequences of gilthead seabream desaturase-like and 15 available animaldesaturases. The amino acid sequences were derived from coding sequences deposited at the GenBank under the following accessionnumbers(in parentheses): D6-desaturases from mouse(AF126798), rat (AB021980) and human(AF126799); D5-desaturase frommouse(AB072976), rat (AF320509) and human(A F199596); FADS-3 from mouse(NM_021890) and human(AF084560); D6-desaturase-like from carp(AF309557), masou salmon(AB074149 and AB070444), rainbow trout(AF301910), salmon(AF478472)and tilapia(AB069727); andD5yD6-desaturase from zebrafish(AF309556). Sequences were aligned using the CLUSTAL W algorithmand were analyzed by phylogenetic analysis using the Fitch–Margoliash and Lest Squares Methods with Evolutionary Clock(KITSCH).The numbers on the branches indicate the bootstrapping value for the node. Sequences marked with an asterisk are not functionallycharacterized.
peptide contained all of the structural features(twomembrane-spanning domains, three histidine-richregions and a cytochromeb domain) which are5
characteristic ofD6 andD5-desaturases(Fig. 1)(Napier et al., 1998; Michaelson et al., 1998a; Aki
et al., 1999; Cho et al., 1999a,b; Watts and Browse,1999; Leonard et al., 2000; Marquardt et al., 2000;Sato et al., 2001; Matsuzaka et al., 2002). In thesame way, partial data obtained for gilthead sea-bream desaturase gene(Fig. 5 and Genbank acces-

458 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Fig. 5. ExonyIntron boundaries of gilthead seabreamD6-desaturase-like gene and humanD6-desaturase,D5-desaturase and FADS-3genes(exon 6–12). Locations of exonyintron boundaries are shown by thin bars. Stars correspond to the nucleotides conserved betweenthe desaturases compared.

459I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
sion number: AF525692) suggest also the sameexonyintron organization, at least as regards humanexons 6–12 forD6, D5 and FADS3 genes(Mar-quardt et al., 2000).Finally, the presence of two desaturase-like tran-
scripts, presently demonstrated in gilthead sea-bream, has also been reported for rat and mouseD6-desaturases(Cho et al., 1999a,b), humanFADS3 (Marquardt, et al., 2000) and rainbowtrout D6-desaturase-like(Seiliez et al., 2001). Inall cases, the reason for these two different tran-scripts remains unknown; hybridizing the Northernblot with sequences from the 59, middle and 39regions of the mouseD6-desaturase ORF yieldedthe same outcomes with respect to the relativeabundance of the two transcripts and the dieteffects (Cho et al., 1999a). Both transcriptsappeared to be depressed in gilthead seabream fedon HUFA-rich diet compared to fish fed on HUFA-free diet, indicating a nutritional regulation similarto that reported for HUFA on mammalian desatu-rases(Cho et al., 1999a,b).The peculiar fatty acid composition of gilthead
seabream fed on HUFA-free diet over 96 days,showing D6 and no D5-desaturation, combinedwith the high expression of the desaturase-likegene in the same fish, suggests that the clonedcDNA is a D6-desaturase without detectableD5activity. These results in gilthead seabream contrastwith the properties of the zebrafishD6-like desa-turase, which was recently reported to be a non-specific D6yD5 desaturase with aD5 activityalmost as high asD6 activity (Hastings et al.,2001). Further investigation on the desaturationcapacities of fish is required to elucidate whetheror not fish possess a second desaturase genecorresponding to aD5 desaturase, as reported inmammals and in other organisms, including thenematodeCaenorhabditis elegans (Michaelson etal., 1998a) and the fungusMortierella alpina(Knutzon et al., 1998; Michaelson et al., 1998b).Ongoing projects devoted to fish genome with afreshwater(zebrafish) and a marine species(fugu)will likely help to answer this question.
Acknowledgments
This work was partially supported by a CIFRE(Ministry of Research and Ferme Marine de Dou-het) grant to the first author. We are grateful toDr J. Sargent and Dr D.R. Tocher for helpful
discussions and encouragement. We thank Y. Mer-cier and P. Aguirre for the growth trials withgilthead seabream. We wish to acknowledge L.Larroquet, E. Plagnes-Juan and M.J. Borthaire fortheir technical assistance. This research was fundedby EU (Q5RS-2000-30058) RAFOA (researchingalternatives to fish oil in aquaculture).
References
Aki, T., Shimada, Y., Inagaki, K., Higashimoto, H., Kawamoto,S., Shigeta, S., et al., 1999. Molecular cloning and functionalcharacterization of rat Delta 6 fatty acid desaturase. Bioch-em. Biophys. Res. Commun. 255, 575–579.
Argyropoulou, V., Kalogeropoulos, N., Alexis, M.N., 1992.Effect of dietary lipids on growth and tissue fatty acidcomposition of grey mullet(Mugil cephalus). Comp. Bioch-em. Physiol. A 101, 129–135.
Altschul, S., Gish, W., Miller, W., Myers, E., Lipman, S.,1990. Basic local alignment search tool. J. Mol. Biol. 215,403–410.
Buzzi, M., Henderson, R.J., Sargent, J.R., 1996. The desatur-ation and elongation of linoleic acid and eicosapentaenoicacid by hepatocytes and liver microsomes from rainbowtrout (Oncorhynchus mykiss) fed diets containing fish oil orolive oil. Biochim. Biophys. Acta 1299, 235–244.
Cho, H.P., Nakamura, M., Clarke, S.D., 1999. Cloning, expres-sion, and nutritional regulation of the mammalian Delta-6desaturase. J. Biol. Chem. 274, 471–477.
Cho, H.P., Nakamura, M., Clarke, S.D., 1999. Cloning, expres-sion, and fatty acid regulation of the human Delta-5 desa-turase. J. Biol. Chem. 274, 37335–37339.
Chomczinski, P., Sacchi, M., 1987. Single step method ofRNA isolation by acid guanidium thiocyanate phenol chlo-roform extraction. Anal. Biochem. 162, 156–159.
Clarke, S.D., Baillie, R., Jump, D.B., Nakamura, M.T., 1997.Fatty acid regulation of gene expression. Its role in fuelpartitioning and insulin resistance. Ann. N.Y. Acad. Sci.827, 178–187.
Cook, H.W., 1991. Fatty acid desaturation and chain elongationin eucaryotes. In: Vance, D., Vance, J.(Eds.), Biochemistryof Lipids, Lipoproteins and Membranes. Elsevier, Amster-damyNew York, pp. 141–169.
Folch, J., Lees, M., Sloane-Stanley, G.H., 1957. A simplemethod for the isolation and purification of total lipids fromanimal tissues. J. Biol. Chem. 226, 497–509.
Forman, B.M., Chen, J., Evans, R.M., 1997. Hypolipidemicdrugs, polyunsaturated fatty acids, and eicosanoids areligands for peroxisome proliferator-activated receptors alphaand delta. Proc. Natl. Acad. Sci. USA 94, 4312–4317.
Hastings, N., Agaba, M., Tocher, D.R., Leaver, M.J., Dick,J.R., Sargent, J.R., et al., 2001. A vertebrate fatty aciddesaturase with Delta 5 and Delta 6 activities. Proc. Natl.Acad. Sci. USA 98, 14304–14309.
Henderson, R.J., Tocher, D.R., 1987. The lipid compositionand biochemistry of freshwater fish. Prog. Lipid Res. 26,281–347.

460 I. Seiliez et al. / Comparative Biochemistry and Physiology Part B 135 (2003) 449–460
Higgins, D., Sharp, P., 1989. Fast and sensitive multiplesequence alignments on a microcomputer. Comput. Appl.Biosci. 5, 151–153.
ITCF, 1988. STAT-ITCF, 4th ed., Service des Etudes Statis-tiques, Paris, France.
Juaneda, P., Rocquelin, G., 1985. Rapid and convenient sepa-ration of phospholipids and non phosphorus lipids from ratheart using silica cartridges. Lipids 20, 40–41.
Kanazawa, A., Teshima, S.I., Ono, K., 1978. Relationshipbetween essential fatty acid requirements of aquatic animalsand the capacity for bioconversion of linoleic acid to highlyunsaturated fatty acids. Comp. Biochem. Physiol. B 63,295–298.
Knutzon, D.S., Thurmond, J.M., Huang, Y.S., Chaudhary, S.,Bobik Jr., E.G., Chan, G.M., et al., 1998. Identification ofDelta5-desaturase fromMortierella alpina by heterologousexpression in Bakers’ yeast and canola. J. Biol. Chem. 273,29360–29366.
Leonard, A.E., Kelder, B., Bobik, E.G., Chuang, L., Parker-Barnes, J.M., Thurmond, J.M., et al., 2000. cDNA cloningand characterization of human Delta 5 desaturase involvedin the biosynthesis of arachidonic acid. Biochem. J 347,719–724.
Marquardt, A., Stohr, H., White, K., Weber, B.H.F., 2000.¨cDNA cloning, genomic structure, and chromosomal local-ization of three members of the human fatty acid desaturasefamily. Genomics 66, 175–183.
Matsuzaka, T., Shimano, H., Yahagi, N., Amemiya-Kudo, M.,Yoshikawa, T., Hasty, A.H., et al., 2002. Dual regulation ofmouse Delta(5)- and Delta(6)-desaturase gene expressionby SREBP-1 and PPARalpha. J. Lipid Res. 43, 107–114.
Michaelson, L., Napier, J., Lewis, M., Griffiths, G., Lazarus,C., Stobart, K., 1998. Functional identification of a fattyacid delta5 desaturase gene fromCaenorhabditis elegans.FEBS Lett. 439, 215–218.
Michaelson, L.V., Lazarus, C.M., Griffiths, G., Napier, J.A.,Stobart, A.K., 1998. Isolation of a Delta5-fatty acid desa-turase gene fromMortierella alpina. J. Biol. Chem. 273,19055–19059.
Mourente, G., Tocher, D., 1993. Incorporation and metabolismof 14C-labelled polyunsaturated fatty acids in juvenile gilt-head seabreamSparus aurata L. in vivo. Fish Physiol.Biochem. 10, 443–453.
Mourente, G., Tocher, D.R., 1994. In vivo metabolism ofw1–14Cx linolenic acid(18:3(n-3)) and w1–14Cx eicosapentae-noic acid (20:5(n-3)) in a marine fish: time-course of thedesaturationyelongation pathway. Biochim. Biophys. Acta1212, 109–118.
Mourente, G., Tocher, D.R., 1998. The in vivo incorporationand metabolism ofw1–14Cx linolenate (18:3n-3) in liver,brain and eyes of juveniles of rainbow troutOncorhynchusmykiss L and gilthead seabreamSparus aurata L. FishPhysiol. Biochem 18, 149–165.
Napier, J.A., Hey, S.J., Lacey, D.J., Shewry, P.R., 1998.Identification of a Caenorhabditis elegans Delta 6-fatty-acid-desaturase by heterologous expression inSaccharomy-ces cerevisiae. Biochem. J 330, 611–614.
National Research Council, 1993. Nutrient requirements offish. National Academic Press, Washington, DC, USA.
Olsen, R.E., Henderson, R.J., McAndrew, B.J., 1990. Theconversion of linoleic acid and linoleic acid to longer chainpolyunsaturated fatty acids byTilapia (Oreochromis) nilo-tica in vivo. Fish Physiol. Biochem. 8, 261–270.
Owen, J.M., Adron, J.W., Middleton, C., Cowey, C.B., 1975.Elongation and desaturation of dietary fatty acids in turbotScophthalmus maximus L., and rainbow trout,Salmo gaird-nerii Rich. Lipids 10, 528–531.
Sargent, J.R., Bell, J.G., Bell, M.V., Henderson, R.J., Tocher,D.R., 1995. Requirement criteria for essential fatty acids. J.Appl. Ichthyol. 11, 183–198.
Sargent, J.R., Tocher, D.R., Bell, J.G., 2002. The lipids. In:Halver, J.E., Hardy, R.W.(Eds.), Fish Nutrition, Vol. III.Academic Press, New York, pp. 181–257.
Sato, M., Adan, Y., Shibata, K., Shoji, Y., Sato, H., Imaizumi,K., 2001. Cloning of rat delta 6-desaturase and its regulationby dietary eicosapentaenoic or docosahexaenoic acid. WorldRev. Nutr. Diet. 88, 196–199.
Seiliez, I., Panserat, S., Kaushik, S., Bergot, P., 2001. Cloning,tissue distribution and nutritional regulation of a Delta6-desaturase-like enzyme in rainbow trout. Comp. Biochem.Physiol. B 130, 83–93.
Shantha, N.C., Ackman, R.G., 1990. Nervonic acid versustricosanoic acid as internal standards in quantitative gaschromatographic analyses of fish oil longer-chain n-3 poly-unsaturated fatty acid methyl esters. J. Chromatogr. 533,1–10.
Spiegelman, B.M., Flier, J.S., 1996. Adipogenesis and obesity:rounding out the big picture. Cell 87, 377–389.
Tocher, D.R., Ghioni, C., 1999. Fatty acid metabolism inmarine fish: low activity of fatty acyl delta 5 desaturationin gilthead seabream(Sparus aurata) cells. Lipids 34,433–440.
Watanabe, T., 1982. Lipid Nutrition in Fish. Comp. Biochem.Physiol. 73B, 3–15.
Watts, J.L., Browse, J., 1999. Isolation and characterization ofa delta5-fatty acid desaturase fromCaenorhabditis elegans.Arch. Biochem. Biophys 362, 175–182.





























