Embed Size (px)
Citation preview

Vol. 32, No. 3INFECTION AND IMMUNITY, June 1981, p. 1227-12330019-9567/81/061227-07$02.00/0
Oligonucleotide Fingerprinting of Ribonucleic Acids ofInfectious Bronchitis Virus Strains
J. P. CLEWLEY, J. MORSER, R. J. AVERY,* AND B. LOMNICZIt
Department of Biological Sciences, University of Warwick, Coventry CV4 7AL, United Kingdom
A total of 11 distinct oligonucleotide fingerprints were obtained in studies ofthe ribonucleic acids of 13 isolates of infectious bronchitis virus. Different sero-
types had distinct fingerprints, but so did varieties within a serotype, allowing a
greater degree of strain differentiation than was previously possible. Some con-
clusions can be drawn from the fingerprints concerning theories of origin andspread of infectious bronchitis virus.
Infectious bronchitis virus (IBV), the proto-type of the family Coronaviridae (29) causespathological conditions of the respiratory tract,the reproductive tract, and the kidneys of chick-ens (7). The disease was first described in theUnited States (27), but by the early 1960s, it hadbeen identified all over the world. Early workshowed that more than one serotype of IBVexisted (18). Since then, a large number ofstrains have been isolated, and attempts havebeen made to classify these strains by using aserological approach. On the basis ofimmunoflu-orescence data (22, 23) and virus neutralizationtests (5, 16, 17), carried out mostly with referencestrains, at least eight serotypes were established(16). However, in a study wherein large numbersof field strains were examined and comparedwith reference strains, it became apparent thatit was not feasible to classify IBV isolates byusing a serological approach (9). In an attemptto circumvent this problem, two groups of strainswere examined by cluster analysis with Euclid-ean distance as the measure of similarity, andtwo main groups were established on the basisof neutralization tests (8). It was also found that12 strains fell into one or the other of two cate-gories of protein patterns differing in the mobil-ity of a small glycoprotein on polyacrylamidegels (4, 24). It is of interest that reference strainsplaced in different groups by the cluster analysis(8) also showed different protein patterns (24).Under circumstances when live vaccine strains
are used against a disease or when the relativelyrapid emergence of new field strains occurs (asin the case of infectious bronchitis), a reliablemethod for strain identification is essential. Theincreasing understanding of the molecular biol-ogy of coronaviruses during the past few years(26) prompted us to compare the ribonucleic
t Present address: Veterinary Medical Research Instituteof the Hungarian Academy of Sciences, Budapest P.f. 18,Hungary.
acids (RNAs) of different IBV strains by oligon-ucleotide fingerprinting, which has been foundto be useful in establishing the genetic basis ofa serological classification of rhabdoviruses, pi-cornaviruses, bunyaviruses, and alphaviruses (J.P. Clewley and D. H. L. Bishop, in C. R. Howard,ed., Newer Techniques in Practical Virology, inpress). Previous studies have shown that viruseswhich cannot be antigenically differentiated usu-ally give comparable but distinguishable oligon-ucleotide fingerprints. When few differences areseen, the viruses are called variants of one an-other. In other cases where many differencesoccur between the fingerprints of two isolatesclassified by other methods as the same virustype, the viruses are called varieties of the sametype.
In this study, the RNAs of strains of IBVbelonging to the same serotype and strains rep-resenting different serotypes have been com-pared by oligonucleotide fingerprinting.
MATERLALS AND METHODS
Viruses. The identification code, year of isolation,and geographical origin of the IBV strains used in thisstudy are summarized in Table 1. Strain Mass-C waskindly supplied by E. Neuvonen, State VeterinaryMedical Institute, Helsinki, Finland; strain Bea wassupplied by V. von Biilow, Bundesforschungsanstaltfur Viruskrankheiten der Tiere, Tubingen, West Ger-many; and NZ-A was supplied by J. E. Lohr,Staatliches Tierarztliches Untersuchungsamt, Aulen-dorf, West Germany. Strain LED was isolated in Hun-gary (19). All of the other strains were kindly suppliedby D. J. Alexander, Central Veterinary Laboratory,New Haw, Weybridge, United Kingdom. Strains Mass41, 927, HVI140, Conn, Holte, and Iowa 97 were sup-plied as chicken embryo kidney cell-adapted strains,whereas the remainder were egg grown and wereadapted to chicken embryo kidney cells during thesestudies by serial (usually not less than five) passages.The exact number of passages of the strains after theirisolation is unknown, but is probably in the order of10 to 20. Strain Bea is a highly passed strain whose
1227
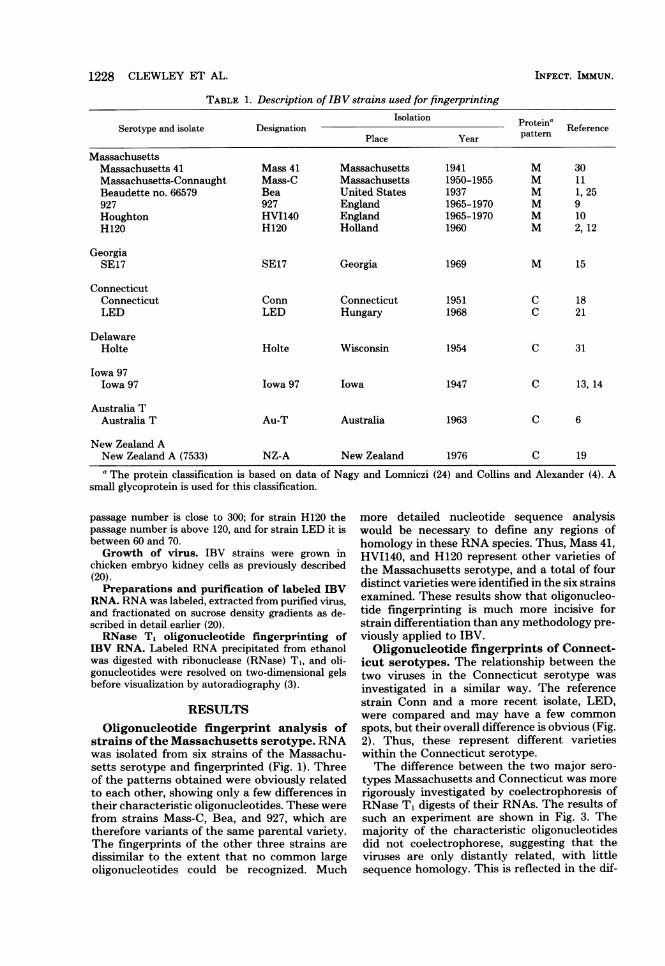
TABLE 1. Description of IBV strains used for fingerprintingIsolation Poen
Serotype and isolate Designation Protemn ReferencePlace Year patr
MassachusettsMassachusetts 41 Mass 41 Massachusetts 1941 M 30Massachusetts-Connaught Mass-C Massachusetts 1950-1955 M 11Beaudette no. 66579 Bea United States 1937 M 1, 25927 927 England 1965-1970 M 9Houghton HVI140 England 1965-1970 M 10H120 H120 Holland 1960 M 2, 12
GeorgiaSE17 SE17 Georgia 1969 M 15
ConnecticutConnecticut Conn Connecticut 1951 C 18LED LED Hungary 1968 C 21
DelawareHolte Holte Wisconsin 1954 C 31
Iowa 97Iowa 97 Iowa 97 Iowa 1947 C 13, 14
Australia TAustralia T Au-T Australia 1963 C 6
New Zealand ANew Zealand A (7533) NZ-A New Zealand 1976 C 19a The protein classification is based on data of Nagy and Lomniczi (24) and Collins and Alexander (4). A
small glycoprotein is used for this classification.
passage number is close to 300; for strain H120 thepassage number is above 120, and for strain LED it isbetween 60 and 70.Growth of virus. IBV strains were grown in
chicken embryo kidney cells as previously described(20).Preparations and purification of labeled IBV
RNA. RNA was labeled, extracted from purified virus,and fractionated on sucrose density gradients as de-scribed in detail earlier (20).RNase T, oligonucleotide fingerprinting of
IBV RNA. Labeled RNA precipitated from ethanolwas digested with ribonuclease (RNase) T,, and oli-gonucleotides were resolved on two-dimensional gelsbefore visualization by autoradiography (3).
RESULTS
Oligonucleotide fingerprint analysis ofstrains ofthe Massachusetts serotype. RNAwas isolated from six strains of the Massachu-setts serotype and fingerprinted (Fig. 1). Threeof the patterns obtained were obviously relatedto each other, showing only a few differences intheir characteristic oligonucleotides. These were
from strains Mass-C, Bea, and 927, which are
therefore variants of the same parental variety.The fingerprints of the other three strains are
dissimilar to the extent that no common largeoligonucleotides could be recognized. Much
more detailed nucleotide sequence analysiswould be necessary to define any regions ofhomology in these RNA species. Thus, Mass 41,HVI140, and H120 represent other varieties ofthe Massachusetts serotype, and a total of fourdistinct varieties were identified in the six strainsexamined. These results show that oligonucleo-tide fingerprinting is much more incisive forstrain differentiation than any methodology pre-viously applied to IBV.Oligonucleotide fingerprints of Connect-
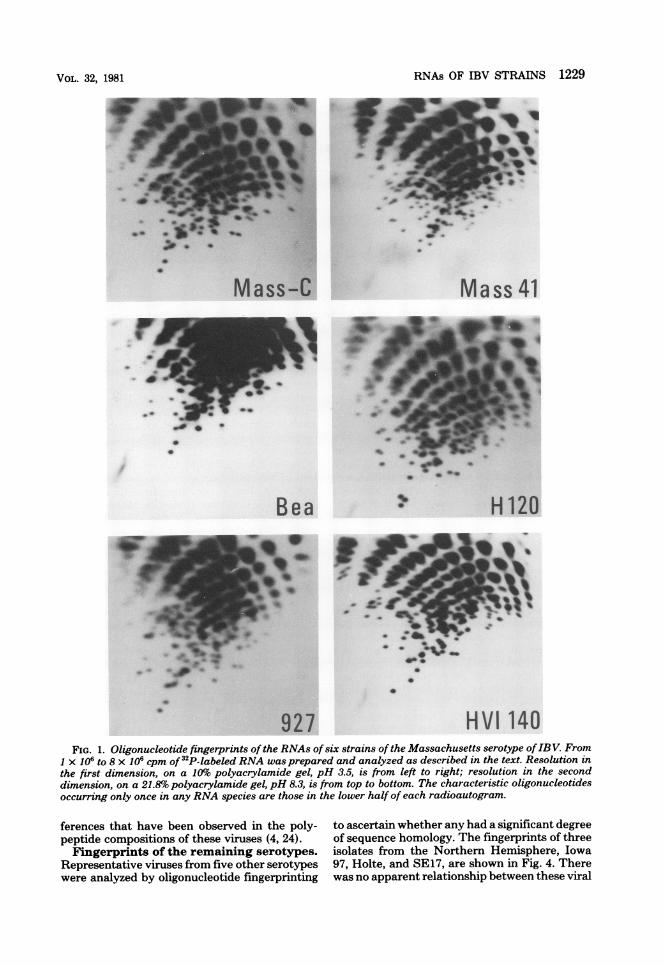
icut serotypes. The relationship between thetwo viruses in the Connecticut serotype wasinvestigated in a similar way. The referencestrain Conn and a more recent isolate, LED,were compared and may have a few commonspots, but their overall difference is obvious (Fig.2). Thus, these represent different varietieswithin the Connecticut serotype.The difference between the two major sero-
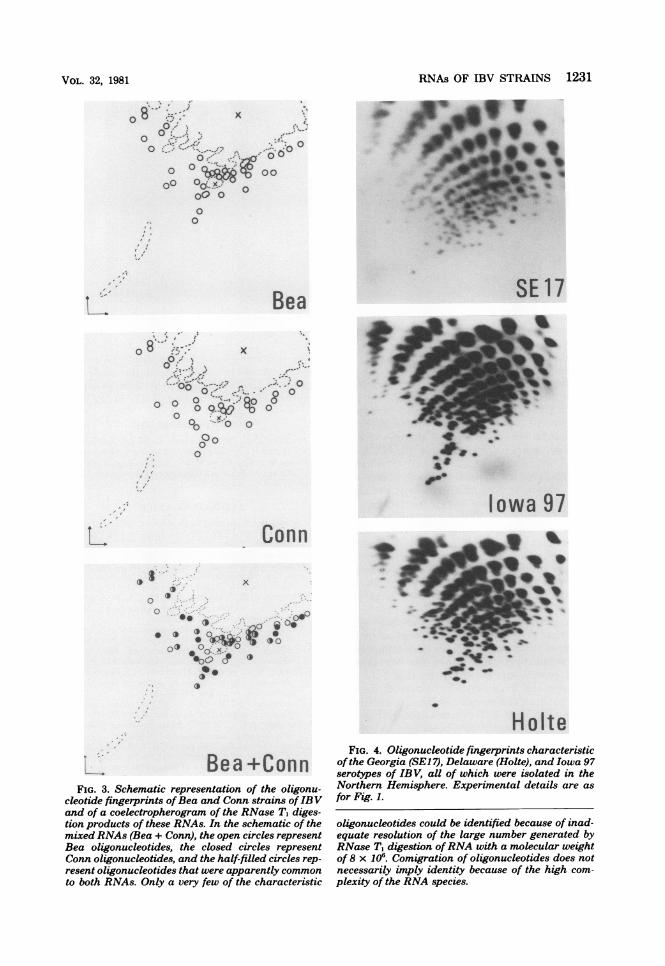
types Massachusetts and Connecticut was morerigorously investigated by coelectrophoresis ofRNase T1 digests of their RNAs. The results ofsuch an experiment are shown in Fig. 3. Themajority of the characteristic oligonucleotidesdid not coelectrophorese, suggesting that theviruses are only distantly related, with littlesequence homology. This is reflected in the dif-
1228 CLEWLEY ET AL. INFECT. IMMUN.
RNAs OF IBV STRAINS 1229
.V7wd
Mass-C
-*_
. wa
0
0 .00
Bea 9.
9.4
41
-01-,-i.I
0
927 HVI 140FIG. 1. Oligonucleotide fingerprints of the RNAs of six strains of the Massachusetts serotype ofIB V. From
1 X 106 to 8 x 106 cpm of 32P-labeled RNA was prepared and analyzed as described in the text. Resolution inthe first dimension, on a 10%o polyacrylamide gel, pH 3.5, is from left to right; resolution in the seconddimension, on a 21.8%o polyacrylamide gel, pH 8.3, is from top to bottom. The characteristic oligonucleotidesoccurring only once in any RNA species are those in the lower half of each radioautogram.
ferences that have been observed in the poly-peptide compositions of these viruses (4, 24).Fingerprints of the remaining serotypes.
Representative viruses from five other serotypeswere analyzed by oligonucleotide fingerprinting
to ascertain whether any had a significant degreeof sequence homology. The fingerprints of threeisolates from the Northern Hemisphere, Iowa97, Holte, and SE17, are shown in Fig. 4. Therewas no apparent relationship between these viral
Mass 41
H120
VOL. 32, 1981
. .. Id--91 40ft-
IMPdo .
'm$4

1230 CLEWLEY ET AL.
74
p
f 4i .. ,l al
I
Conn LEDFIG. 2. Oligonucleotide fingerprints of the RNAs of two strains of the Connecticut serotype of IBV.
Technical details are as described in the legend to Fig. 1.
RNAs.It was of interest to examine two strains from
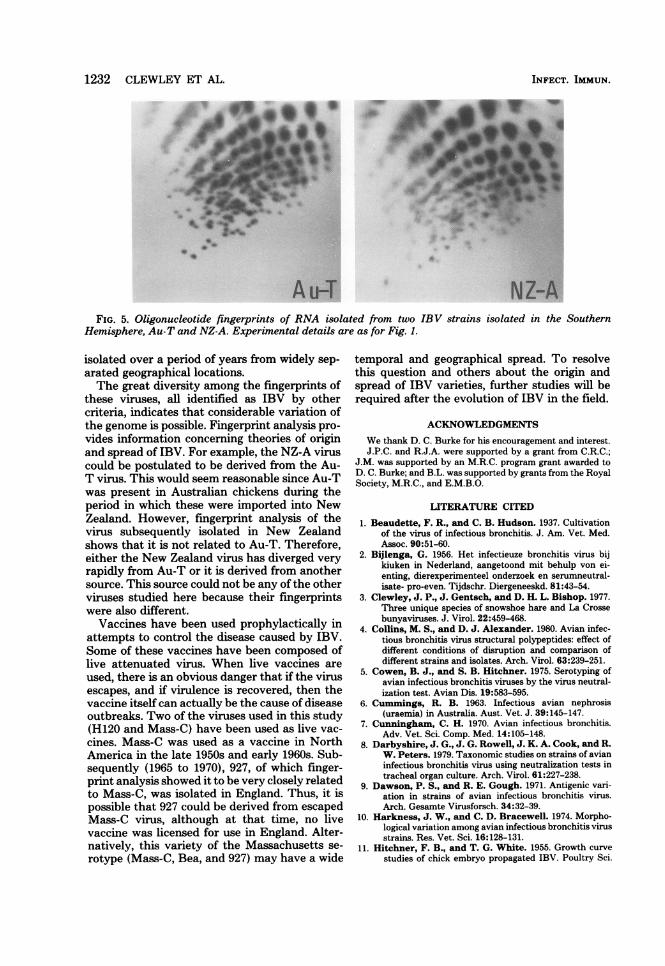
the Southern Hemisphere (NZ-A and Au-T) todetermine whether it was possible to shed anylight on the origin of these strains, since theywere isolated later than Iowa 97 and Holte (Ta-ble 1). The fingerprints of these RNAs are shownin Fig. 5. As far as it is possible to tell, NZ-A andAu-T are unrelated to each other and to Iowa97, Holte, and SE17, suggesting that NZ-A wasnot derived from the Australian strain, althoughNew Zealand allowed the importation of chick-ens from Australia before the NZ-A virus wasisolated.
In addition, none of these five strains showedany obvious similarity to the two other majorserotypes, Massachusetts and Connecticut.
DISCUSSIONWe present here the RNase T1 oligonucleotide
fingerprints of RNA prepared from 13 virus iso-lates. All of these isolates were originally classi-fied as IBV by using pathological criteria. Wehave confirmed that all of the viruses studied inthis work have the virion density, genome size,and protein profile expected for IBV (data notshown). Nevertheless, at least 11 different fin-gerprints were recognized. These were given byMass 41, Mass-C, HVI140, and H120 from theMassachusetts serotype; Conn and LED fromthe Connecticut serotype; and five representa-tives of other serotypes, SE17 from the Georgiaserotype, Holte from the Delaware serotype,Iowa 97 from the Iowa 97 serotype, Au-T fromthe Australia T serotype, and NZ-A from theNew Zealand A serotype. Thus, different sero-types gave distinct fingerprints, but so did vari-eties within a serotype. Whether variants withsimilar fingerprints, such as Mass-C, Bea, and927, are antigenically closer to each other than
to other varieties of the same serotype withdifferent fingerprints is not yet established. Fur-ther generalizations about the genetic basis ofantigenic relationships among IBV isolateswould be premature and must await the intro-duction of more precise methods capable of rec-ognizing subtle antigenic differences amongstrains.One problem raised by this study is the ques-
tion of which isolate should be considered theprototype virus of the Massachusetts serotype.Three of the viruses from the Massachusettsserotype characterized in this study had vir-tually identical fingerprints. These are Mass-C,Bea, and 927. These fingerprints differed sub-stantially from the fingerprint of another viruswhich is currently regarded as the prototypevirus of the Massachusetts serotype (Mass 41).The differences are unlikely to have arisen dur-ing laboratory passage because the fingerprintof Bea produced in this work is identical to thatpreviously published (20). Thus, the limitednumber of passages between these viruses (fewerthan 10) has not allowed the accumulation ofsufficient mutations to change the pattern ofcharacteristic oligonucleotides. This agrees withresults from other viruses (Clewley and Bishop,in press). Stern and Kennedy (28) have recentlypublished a fingerprint of strain Beaudette(IBV42), obtained from the American Type Cul-ture Collection, which differs only slightly fromthe fingerprints we have obtained from Mass-C,Bea, and 927. IBV42 and strain Bea (66579) usedhere have been maintained as separate lines(25). Thus, these four viruses are all variants ofthe same parental virus.IBV was first recognized as a disease in the
United States in 1931 (27), but by the earlysixties, outbreaks had occurred all over theworld. The viruses examined in this study were
INFECT. IMMUN.
.
VOL. 32, 1981
0 <t
0o'0
o0 0
00.o
, .I
Q..5.
00
0 0
060c} --o o
0 0
o
RNAS OF IBV STRAINS 1231
a
S
Bea SE 17.,..''tzi;'I~~~
S .wtr
Iowa 97t :.L Conn
xI8.;DC3
0o . .10
0P 3
cbi
. ._Bea +ConnFIG. 3. Schematic representation of the oligonu-
cleotide fingerprints ofBea and Conn strains ofIBVand of a coelectropherogram of the RNase T, diges-tion products of these RNAs. In the schematic of themixed RNAs (Bea + Conn), the open circles representBea oligonucleotides, the closed circles representConn oligonucleotides, and the half-filled circles rep-resent oligonucleotides that were apparently commonto both RNAs. Only a very few of the characteristic
- * .s Iv
It
~ ~ ~ 1%
'a it0
HolteFIG. 4. Oligonucleotide fingerprints characteristic
of the Georgia (SEJ 7), Delaware (Holte), and Iowa 97serotypes of IBV, all of which were isolated in theNorthern Hemisphere. Experimental details are asfor Fig. 1.
oligonucleotides could be identified because of inad-equate resolution of the large number generated byRNase T1 digestion ofRNA with a molecular weightof 8 x 106. Comigration of oligonucleotides does notnecessarily imply identity because of the high com-plexity of the RNA species.
.1.. 0
8 0
1232 CLEWLEY ET AL.
Au-T NZ-AFIG. 5. Oligonucleotide fingerprints of RNA isolated from two IBV strains isolated in the Southern
Hemisphere, Au-T and NZ-A. Experimental details are as for Fig. 1.
isolated over a period of years from widely sep-arated geographical locations.The great diversity among the fingerprints of
these viruses, all identified as IBV by othercriteria, indicates that considerable variation ofthe genome is possible. Fingerprint analysis pro-vides information concerning theories of originand spread of IBV. For example, the NZ-A viruscould be postulated to be derived from the Au-T virus. This would seem reasonable since Au-Twas present in Australian chickens during theperiod in which these were imported into NewZealand. However, fingerprint analysis of thevirus subsequently isolated in New Zealandshows that it is not related to Au-T. Therefore,either the New Zealand virus has diverged veryrapidly from Au-T or it is derived from anothersource. This source could not be any of the otherviruses studied here because their fingerprintswere also different.
Vaccines have been used prophylactically inattempts to control the disease caused by IBV.Some of these vaccines have been composed oflive attenuated virus. When live vaccines areused, there is an obvious danger that if the virusescapes, and if virulence is recovered, then thevaccine itself can actually be the cause of diseaseoutbreaks. Two of the viruses used in this study(H120 and Mass-C) have been used as live vac-cines. Mass-C was used as a vaccine in NorthAmerica in the late 1950s and early 1960s. Sub-sequently (1965 to 1970), 927, of which finger-print analysis showed it to be very closely relatedto Mass-C, was isolated in England. Thus, it ispossible that 927 could be derived from escapedMass-C virus, although at that time, no livevaccine was licensed for use in England. Alter-natively, this variety of the Massachusetts se-rotype (Mass-C, Bea, and 927) may have a wide
temporal and geographical spread. To resolvethis question and others about the origin andspread of IBV varieties, further studies will berequired after the evolution of IBV in the field.
ACKNOWLEDGMENTSWe thank D. C. Burke for his encouragement and interest.J.P.C. and R.J.A. were supported by a grant from C.R.C.;
J.M. was supported by an M.R.C. program grant awarded toD. C. Burke; and B.L. was supported by grants from the RoyalSociety, M.R.C., and E.M.B.O.
LITERATURE CITED1. Beaudette, F. R., and C. B. Hudson. 1937. Cultivation
of the virus of infectious bronchitis. J. Am. Vet. Med.Assoc. 90:51-60.
2. Bijlenga, G. 1956. Het infectieuze bronchitis virus bijkiuken in Nederland, aangetoond mit behulp von ei-enting, dierexperimenteel onderzoek en serumneutral-isate- pro-even. Tijdschr. Diergeneeskd. 81:43-54.
3. Clewley, J. P., J. Gentsch, and D. H. L. Bishop. 1977.Three unique species of snowshoe hare and La Crossebunyaviruses. J. Virol. 22:459-468.
4. Collins, M. S., and D. J. Alexander. 1980. Avian infec-tious bronchitis virus structural polypeptides: effect ofdifferent conditions of disruption and comparison ofdifferent strains and isolates. Arch. Virol. 63:239-251.
5. Cowen, B. J., and S. B. Hitchner. 1975. Serotyping ofavian infectious bronchitis viruses by the virus neutral-ization test. Avian Dis. 19:583-595.
6. Cummings, R. B. 1963. Infectious avian nephrosis(uraemia) in Australia. Aust. Vet. J. 39:145-147.
7. Cunningham, C. H. 1970. Avian infectious bronchitis.Adv. Vet. Sci. Comp. Med. 14:105-148.
8. Darbyshire, J. G., J. G. Rowell, J. K. A. Cook, and R.W. Peters. 1979. Taxonomic studies on strains of avianinfectious bronchitis virus using neutralization tests intracheal organ culture. Arch. Virol. 61:227-238.
9. Dawson, P. S., and R. E. Gough. 1971. Antigenic vari-ation in strains of avian infectious bronchitis virus.Arch. Gesamte Virusforsch. 34:32-39.
10. Harkness, J. W., and C. D. Bracewell. 1974. Morpho-logical variation among avian infectious bronchitis virusstrains. Res. Vet. Sci. 16:128-131.
11. Hitchner, F. B., and T. G. White. 1955. Growth curvestudies of chick embryo propagated IBV. Poultry Sci.
INFECT. IMMUN.
00 10
0
v IF wI
.z.,.I
..jw. Iil? -d'..
"Af

RNAs OF IBV STRAINS 1233
34:590-594.12. Hoekstra, J., and B. Rispens. 1960. Infectieuze bron-
chitis bij pluimvee. I. Laboratorium-experimenten meteen sterkwerkende intstam. Tijdschr. Diergeneeskd. 85:398-403.
13. Hofstad, M. S. 1958. Antigenic differences among isolatesof avian infectious bronchitis virus. Am. J. Vet. Res. 19:740-743.
14. Hofstad, M. S., and H. W. Yoder, Jr. 1966. Avianinfectious bronchitis-virus distribution in tissues ofchickens. Avian Dis. 10:230-239.
15. Hopkins, S. R. 1969. Serological and immunologic prop-erties of a recent isolate of infectious bronchitis virus.Avian Dis. 13:356-362.
16. Hopkins, S. R. 1974. Serological comparisons of strainsof infectious bronchitis virus using plaque-purified iso-lants. Avian Dis. 18:231-239.
17. Johnson, R. B., and W. W. Marquardt. 1975. Theneutralizing characteristics of strains of infectious bron-chitis virus measured by the constant virus-variableserum method in chicken tracheal cultures. Avian Dis.19:82-90.
18. Jungherr, E. L, T. W. Chomiak, and R. E. Luginbuhl.1956. Immunologic differences in strains of infectiousbronchitis virus. Proc. Annu. Meet. U.S. LivestockSanit. Assoc. 60:203-209.
19. Lohr, J. E. 1976. Serological differences between strainsof infectious bronchitis virus from New Zealand, Aus-tralia and the United States. Avian Dis. 20:478-482.
20. Lomniczi, B., and L. Kennedy. 1977. Genome of infec-tious bronchitis virus. J. Virol. 24:99-107.
21. Lomniczi, B., and L. Stipkovits. 1968. Isolation of thevirus of infectious bronchitis. Magy. Allatorv. Lapja 23:600-603.
22. Lucio, B., and S. B. Hitchner. 1970. Differentiation anddetection of infectious bronchitis virus subtypes byimmunofluorescence. Avian Dis. 14:9-24.
23. Lukert, P. D. 1969. Differentiation of avian infectiousbronchitis virus serotype by immunofluorescence.Avian Dis. 13:847-852.
24. Nagy, E., and B. Lomniczi. 1979. Polypeptide pattern ofinfectious bronchitis virus serotype fall into two cate-gories. Arch. Virol. 61:341-345.
25. Purchase, H. G., C. H. Cunningham, and B. R. Bur-master. 1966. Identification and epizootiology of Infec-tious Bronchitis in a closed flock. Avian Dis. 10:111-121.
26. Robb, J. A., and C. W. Bond. 1980. Coronaviridae, p.193-247 In H. Fraenkel-Conrat and R. R. Wagner ed.Comprehensive virology, vol. 14. Plenum Press, NewYork.
27. Schalk, A. F., and M. C. Hawn. 1931. An apparentlynew respiratory disease of baby chicks. J. Am. Vet.Med. Assoc. 78:413-422.
28. Stern, D. F., and S. L. T. Kennedy. 1980. Coronavirusmultiplication strategy. I. Identification and character-ization of virus-specified RNA. J. Virol. 34:665-674.
29. Tyrrell, D. A. J., D. J. Alexander, J. D. Almfida, C.H. Cunningham, B. C. Easterday, D. J. Garwes, J.C. Hierholzer, A. Kapikian, M. R. MacNaughton,and K. McIntosh. 1978. Coronaviridae: second report.Intervirology 10:321-328.
30. Van Roekel, H., K. L. Balls, M. K. Clarke, 0. M.Olesiuk; and F. G. Sperling. 1950. Infectious bron-chitis. Mass. Agric. Exp. Stn. Bull. no. 460.
31. Winterfield, R. W., and S. B. Hitchner. 1962. Etiologyof an infectious nephritis-nephrosis syndrome of chick-ens. Am. J. Vet. Res. 23:1273-1279.
VOL. 32, 1981





























