Embed Size (px)
Citation preview

REVIEWS–A PEER REVIEWED FORUM
Flipping the Switch: How a Sperm Activatesthe Egg at FertilizationJohn Parrington,1 Lianne C. Davis,1 Antony Galione,1 and Gary Wessel2*
Sperm interaction with an egg in animals was first documented 160 years ago in sea urchins by AlphonseDerbes (1847) when he noted the formation of an “envelope” following the sperm’s “approach” to the egg.The “envelope” in sea urchins is an obvious phenotype of fertilization in this animal and over the past 35years has served to indicate a presence of calcium released from cytoplasmic stores essential to activate theegg. The mechanism of calcium release has been intensely studied because it is a universal regulator ofcellular activity, and recently several intersecting pathways of calcium release have been defined. Here weexamine these various mechanisms with special emphasis on recent work in eggs of both sea urchins andmice. Developmental Dynamics 236:2027–2038, 2007. © 2007 Wiley-Liss, Inc.
Key words: fertilization; sperm; calcium, egg
Accepted 22 May 2007
INTRODUCTION
A new cellular potential is formed bythe union of an egg and a sperm. Thesperm supplies the paternal genome,and in most species the centriole, andthe stimulus that activates the fertil-ized egg to develop into an embryo.Egg activation is a multi-faceted pro-cess that includes the initiation of theblock to polyspermy (Wong and Wes-sel, 2006), resumption of meiosis andentry into the embryonic cell cycle,and changes in the expression of genesand their protein products within thezygote and early embryo (Whitting-ham, 1980; Whitaker and Steinhardt,
1982; Jaffe, 1983; Yanagimachi,1994). The mechanism whereby thesperm activates the egg at fertiliza-tion has been a subject of interest forover a century, ever since pioneerssuch as Jacques Loeb and Frank Lilliebegan studying the process at the Ma-rine Biological Laboratory at WoodsHole. The sea urchin has played a cen-tral role as a model organism forstudying egg activation from thosefirst studies to the present day, and isa key emphasis of this review (Fig. 1).
It was Loeb who first proposed, in1898, that egg activation was a chem-ical process involving changes in the
concentration of ions within the egg(see Loeb, 1899), a bold idea given thatmany people at that time were stilltalking of embryo development interms of “vital forces” (Pauly, 1987).Loeb’s proposal was based upon hisdiscovery that sea urchin eggs couldbe induced to begin developing parthe-nogenetically, without the need for asperm, simply by altering the propor-tion of ions in the solution bathing theeggs. Later, in 1902, Loeb suggestedthat the sperm initiated such a changein ionic composition in the egg bymeans of a “sperm factor” that wasintroduced into the egg during gamete
ABBREVIATIONS C2 conserved region 2 of PKC cADPR cyclic ADP ribose DAG diacyl glycerol DAG diacyl glycerol IP3 inositoltrisphosphate NAADP nicotinic acid adenine dinucleotide phosphate NO nitric oxide NOS nitric oxide; synthase PH pleckstrin homologyPIP2 phosphatidylinositol (4,5)-bisphosphate PLC phospholipase C SFK Src family kinase SH2 Src homology domain 2.
1Department of Pharmacology, University of Oxford, Oxford, United Kingdom2Department of Molecular and Cellular Biology and Biochemistry, Brown University, Providence, Rhode IslandGrant sponsors: National Science Foundation; National Institutes of Health; Medical Research Council; Wellcome Trust.*Correspondence to: Gary Wessel, Department of Molecular and Cellular Biology and Biochemistry, Brown University, 185Meeting Street, Box G, Providence, RI 02912. E-mail: [email protected]
DOI 10.1002/dvdy.21255Published online 24 July 2007 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 236:2027–2038, 2007
© 2007 Wiley-Liss, Inc.

fusion. A different model of egg acti-vation was put forward by Frank Lil-lie. He suggested that the process wastriggered by an interaction between asperm ligand and a receptor on theegg surface (Lillie, 1913). The impor-tance of these pioneering scientists isdemonstrated by the fact that thesetwo alternative models are still thedominant frameworks for our under-standing of the process of egg activa-tion today.
Despite Loeb’s pioneering sugges-tion that ionic changes were a keycomponent of egg activation, it wasnot until much later that one ion inparticular, calcium (Ca2�), was shownto play the initiating role. The impor-tance of Ca2� was suggested duringthe 1930s by several investigators,most notably Daniel Mazia (Mazia,1937), Lewis Victor Heilbrunn (see,e.g., Heilbrunn, 1952), and FloydMoser (Moser, 1939). However, defin-itive evidence that a change in intra-cellular egg Ca2� was the trigger foregg activation only emerged in the1970s. A key step forward came withstudies in which eggs were treatedwith the Ca2� ionophore A23187(Steinhardt and Epel, 1974; Stein-hardt et al., 1974). This triggered eggactivation, demonstrating that artifi-cially raising intracellular egg Ca2�
induces events associated with egg ac-tivation (Steinhardt et al., 1974; Ful-ton and Whittingham, 1978). It wassubsequently shown that a rise inCa2� within the egg accompanies fer-tilization (Ridgway et al., 1977; see,e.g., Stricker, 1999; Whitaker, 2006),and that blocking this rise in Ca2�
with chelating agents inhibited eggactivation (Whitaker and Steinhardt,1982; Kline, 1988; Kline and Kline,1992). While many of these studieswere first carried out on sea urchineggs, it is now well established thatCa2� acts as the trigger for egg acti-vation in practically all species(Stricker, 1999; Whitaker, 2006). De-spite being universally employed, theexact form of the Ca2� signal at fertil-ization varies between animal groups:whereas a single wave of Ca2� crossesthe egg in sea urchins, frogs, and fish(Fig. 2), the sperm triggers a series ofperiodic increases that have beentermed Ca2� oscillations in mammals,nemertean worms, and ascidians(Stricker, 1999; Whitaker, 2006).
CALCIUM CONDUIT,SOLUBLE SPERM FACTOR,OR SURFACEINTERACTION?
Even though Ca2� has been known tobe the trigger of egg activation for sev-eral decades, the mechanism wherebythe sperm induces a change in the con-centration of Ca2� within the eggremains to be fully resolved. Threemajor hypotheses dominate under-standing of this process: (1) the Ca2�
bomb or conduit model; (2) the solublesperm factor model; and (3) the mem-brane receptor model (Fig. 3; Whi-taker and Swann, 1993; Evans andKopf, 1998; Parrington, 2001). Thefirst model was based around the ideathat the sperm introduced a bolus ofCa2� during gamete fusion (Jaffe,1983, 1991), later modified to the sug-gestion that channels in the surfacemembrane of the sperm allowed it toact as a conduit for Ca2� entry (Cretonand Jaffe, 2001). Ca2� introduced inthis way would then act to triggerCa2� induced Ca2� release (CICR)within the egg (Jaffe, 1991). Althoughattractive in its straightforwardness,and consistent with the finding thatsperm-egg fusion in both the sea ur-chin and the mouse always precedesegg Ca2� release (McCulloh andChambers, 1992; Lawrence et al.,1997), a number of studies have nowcast doubt upon the validity of thismodel. Counterevidence includesdemonstration that injection of Ca2�
into sea urchin, Ascidian, or mamma-lian eggs fails to induce further Ca2�
release (Whitaker and Swann, 1993;Swann and Ozil, 1994), and that inthe mouse, no elevation on the localcytoplasmic Ca2� concentration oc-curs at gamete fusion, which would beexpected if the sperm acted as a Ca2�
conduit (Jones et al., 1998).The second model, which proposes
that egg activation is triggered by asoluble factor released during gametefusion from the sperm into the egg,was very controversial for many yearsdue to the failure to identify such afactor at the molecular level. In mam-mals, this situation has greatlychanged in the last few years with thediscovery that the mammalian spermfactor appears to be a novel type ofsperm-specific phospholipase C, PLC�(Saunders et al., 2002). While it has
now become generally accepted thatPLC� is likely to be the physiologicalagent of egg activation in mammals(Swann et al., 2006), many questionsremain about both its mechanism ofaction and whether such a sperm PLCplays a role during egg activation inother animals.
The third hypothesis, the mem-brane receptor model, was the domi-nant hypothesis for many years. Pro-posing that egg activation is triggeredby an interaction between a ligandupon the sperm and a receptor on theegg (Jaffe, 1990; Schultz and Kopf,1995; Evans and Kopf, 1998), thismodel had the virtue that such sur-face-mediated interactions are a nor-mal feature of cell signaling in somaticcells. From the start, this model wascentered around the idea that the sig-naling cascade triggered by such aninteraction would involve activation ofan egg PLC (Jaffe, 1990; Schultz andKopf, 1995; Evans and Kopf, 1998).While studies in echinoderms andsome other animals have pointed to acentral role in egg activation for PLC�(Townley et al., 2006), what remainsfar less clear is whether this is theconsequence of a surface-mediated in-teraction between a sperm ligand andan egg receptor.
THE PHOSPHOINOSITIDESIGNALLING PATHWAY
The PLC family of proteins are cyto-plasmic enzymes that cleave phospha-tidylinositol (4,5)-bisphosphate (PIP2)at the plasma membrane to yield ino-sitol triphosphate (IP3) and diacylglycerol (DAG), each of which regulatea variety of cell processes (Rhee,2001). IP3 may then bind its receptor,the IP3 receptor, in the endoplasmicreticulum to release Ca2� into the cy-toplasm (see Fig. 5). At least six dif-ferent subfamilies of PLC have beenidentified, including �, �, �, �, andmore recently also � and � (Rhee,2001; Saunders et al., 2002; Nakaharaet al., 2005). All PLC isoforms containseparate X and Y domains that cometogether to form the active site re-sponsible for PIP2 cleavage (Fig. 4). Inaddition to the X and Y domains, theyalso contain various combinations ofregulatory domains, including PH do-mains (pleckstrin homology), SH2 do-mains (Src homology 2), and C2 do-
2028 PARRINGTON ET AL.
mains (constant or conserved region 2of PKC; Fig. 4).
The regulatory domains of PLCtarget different isoforms to their re-spective activators or substrates viaprotein–protein or protein–lipid inter-actions. The mechanism of PLC activa-tion varies according to subfamily, andis dependent on the specific combina-tion of regulatory domains present ineach isoforms (see, e.g., Roux et al.,2006). PH domains bind polyphosphoi-nositides such as PIP2 and PIP3, andthus serve to target the protein to theplasma membrane (Harlan et al., 1994;Rameh and Cantley, 1999). PH do-mains can also mediate interaction withand activation by the �� subunits ofheterotrimeric G proteins (Camps et al.,1992; Smrcka and Sternweis, 1993; Leeet al., 1994). SH2 domains interact withreceptor protein tyrosine kinases aswell as with non-receptor tyrosine ki-nases like Src (Weiss, 1993; Noh et al.,1995). Finally, C2 domains bind Ca2�
and confer Ca2�-mediated phospholip-id-binding properties to the proteinsthat contain them (see below). Isoformsof PLC�, PLC�, and PLC� have beenidentified in eggs, while isoforms ofPLC�, �, �, and most recently �, havebeen described in sperm (Dupont et al.,1996; Mehlmann et al., 1998; Wang etal., 1998; Wu et al., 2001; Parrington etal., 2002; Saunders et al., 2002).
THE ROLE OF THEPHOSPHOINOSITIDEPATHWAY DURING EGGACTIVATION
The idea that the phosphoinositidepathway (PI) plays a central role dur-ing egg activation has been supportedby a number of studies over the lasttwo decades. Biochemical measure-ments in sea urchin and frog eggs
showed that there is an increasedturnover of phospholipids and an in-crease in inositol trisphosphate (IP3)levels at fertilization (Turner et al.,1986; Stith et al., 1994). Further evi-dence of a role for the PI pathway isthe fact that the PLC inhibitorU73122 inhibits both Ca2� releaseand egg activation in sea urchins andmice (Dupont et al., 1996; Lee et al.,1998) while egg activation in mice is
Fig. 1. A: Top view of the sea urchin Strongy-locentrotus purpuratus, induced to shed eggs(orange mass in center). B: An adult opened toreveal the five lobes of ovary. C: An egg at themoment of activation showing the fusing spermon the left, along with the rising fertilizationenvelop. D: The same egg 45 sec followinginsemination showing the fully formed fertiliza-tion envelope, and the two pronuclei within theegg. Fig. 2. Two phases of Ca2� increase occur at fertilization, reflecting two spatially and temporally
different events. The first increase is the cortical flash due to a rapid influx of Ca2� across theplasma membrane. The second phase is due to Ca2� release from intracellular stores, whichpropagates as a wave across the egg from the sperm entry point. These different phases ofresponse rely on different Ca2� mobilizing messengers. The cortical flash is uniquely reliant uponNAADP, whereas IP3 and cADPR activate and prolong the Ca2� spike, respectively. cADPR ismostly produced after the spike upstroke; however, this is not entirely clear because of messengerredundancy.
Fig. 3. The three major models of egg activation. The conduit model suggests that Ca2� entersthe egg via a Ca2� channel opening in the sperm plasma membrane; the membrane receptor modelsuggests that a receptor activated by sperm binding relays a signal to initiate the Ca2� releasemechanism, and the soluble sperm factor model suggests that sperm deliver a molecule thatinitiates the Ca2� release mechanism. See text for additional details.
EGG ACTIVATION AT FERTILIZATION 2029

blocked by treatment with an anti-body raised against the IP3 receptor(Miyazaki et al., 1992). Additional ev-idence also shows that an egg PLCplays a central role during egg activa-tion in echinoderms (Townley et al.,2006), and that a sperm PLC� is thephysiological agent of egg activationin mammals (Swann et al., 2006).
THE IDENTIFICATION OFTHE MAMMALIAN SPERMFACTOR AS PLC
The first evidence that a solublesperm factor might be the trigger ofegg activation came from a study insea urchins, in which injection of a seaurchin sperm extract induced egg ac-tivation (Dale et al., 1985). A laterstudy identified a similar putative eggactivation factor in Ascidian sperm(Dale, 1988). It was not clear fromthese studies whether the activatingstimulus was a protein or a small mo-lecular weight Ca2� mobilizing mes-senger. However, evidence for the ex-istence of a soluble sperm protein thatcould mimic the ability to trigger apattern of Ca2� release similar to thatseen at fertilization and induce eggactivation emerged from a series ofstudies in which sperm extracts wereinjected into eggs of a variety of mam-mals, marine worms, and ascidians(Swann, 1990; Stricker, 1999; Kyo-zuka et al., 1998; Parrington, 2001).More recently, sperm extracts fromfrogs, chickens, and tilapia fish havealso been shown to trigger Ca2� oscil-lations in mouse eggs (Dong et al.,2000; Coward et al., 2003, 2005).
Many years of effort were spent try-ing to identify the sperm factor at themolecular level (Parrington, 2001). Abreakthrough came with the discoverythat the mammalian sperm factor wasa sperm-specific PLC that had verydistinctive enzymatic properties com-pared to other known PLCs (Jones etal., 1998, 2000; Parrington et al.,1999, 2002). This discovery was madepossible by the use of a cell-free egg-based Ca2� release assay, the sea ur-chin egg homogenate, which enabled abiochemical dissection of the spermfactor’s mechanism of action in a waythat would have been much more dif-ficult using an intact egg. The discov-ery of an apparent sperm-specific fac-tor initiated a search for a novel type
of PLC by analysis of cDNA data-bases, leading to the identification ofPLC� (Saunders et al., 2002). Confir-mation that PLC� was the long-sought-after mouse sperm factor camewith the finding that recombinantPLC� RNA and protein trigger Ca2�
oscillations when injected into amouse egg (Saunders et al., 2002;Kouchi et al., 2005). In addition, im-muno-depletion of PLC� from spermextracts abolished their ability tocause Ca2� release (Saunders et al.,2002). Further studies have shownthat the presence of PLC� in fraction-ated sperm extracts correlates withtheir ability to induce Ca2� oscilla-tions in the egg (Fujimoto et al., 2004;Kurokawa et al., 2005).
Current questions of interest aboutPLC� include its exact mode of actiononce within the fertilized egg. Recentstudies have shown that PLC� ap-pears to become sequestered in thepronuclei of the fertilized egg (Larmanet al., 2004; Yoda et al., 2004), sug-gesting a possible role in mediatingthe cell-cycle dependency of Ca2� os-cillations in the early embryo, but theexact functional significance of thispattern of localization remains to bedetermined. Of great interest is howPLC� is stored and packaged in thesperm, and whether it plays a role dur-ing egg activation in other animalgroups besides mammals. This latterpoint has support, as demonstrated in arecent study that identified a chickenorthologue of PLC� with similar spermspecificity and functional properties asmammalian PLC� (Coward et al., 2005),suggesting perhaps a conserved, wide-spread mechanism for egg activation ina number of vertebrates.
The definitive test remains as towhether PLC� is both necessary andsufficient for physiological egg activa-tion in mammals. A recent study usedRNA interference to create mousesperm partially deficient in PLC�.These sperm had partial deficienciesin their ability to trigger Ca2� oscilla-tions in mouse eggs and to cause eggactivation (Knott et al., 2005), sug-gesting that PLC� is necessary. How-ever, a concrete answer will requirethe creation of a mouse knockout inPLC�, or the identification of a humaninfertility patient with no functionalPLC�. The fact that some infertilemen appear to produce sperm defi-
cient in the ability to trigger egg acti-vation (Flaherty et al., 1998; Mahutteand Arici, 2003; Heindryckx et al.,2004; Yanagida, 2004) provides thepossibility to further study the linkbetween PLC� structure and functionin mammals.
A PLC� homologue has not been iden-tified in echinoderms or ascidians, sothe controversy over the universality ofa sperm-delivered PLC� as a masterregulator of Ca2� release and egg acti-vation remains to be resolved. Instead,evidence suggests that egg activation inechinoderms and in some vertebrates,like amphibians, is mediated by a dis-tinct mechanism, with an egg PLC�playing a central role.
THE CENTRAL ROLE OFEGG PLC� DURING EGGACTIVATION INECHINODERMS
In species where a PLC� has not beenidentified, other members of the PLCfamily of proteins, mainly PLC�, areessential mediators of Ca2� release(Runft and Jaffe, 2000; Runft et al.,2002, 2004; Giusti et al., 2003). Fertil-ization in both echinoderms and frogs,for example, is associated with an in-crease in the activity of PLC� (Fig. 5)(Rongish et al., 1999; Sato et al.,2000). Most of the functional evidenceimplicating PLC� in the release ofCa2� in invertebrate eggs has reliedon the use of dominant-negative SH2domains. SH2 domains, of whichPLC� contains two, bind Src familykinases (SFKs), a group of tyrosinekinases that are well-known activa-tors of PLCs. Injection of PLC� SH2domains into Ascidian and echino-derm eggs competes away Src bindingand inhibits Ca2� release at fertiliza-tion, suggesting that PLC� is an im-portant activator of the IP3 pathwayin invertebrates (Carroll et al., 1997;Giusti et al., 1999; Runft et al., 2002).In frogs, PLC� associates with an SFKknown as Xyk and becomes tyrosinephosphorylated shortly after fertiliza-tion (Sato et al., 2000). While injectionof PLC� SH2 domains into vertebrateeggs, including frogs and mammals,has no inhibitory effect on Ca2� re-lease pathways (Mehlmann et al.,1998; Runft et al., 1999, 2002), injec-tion of recombinant PLC�1 can induceCa2� release in mouse oocytes (Mehl-
2030 PARRINGTON ET AL.
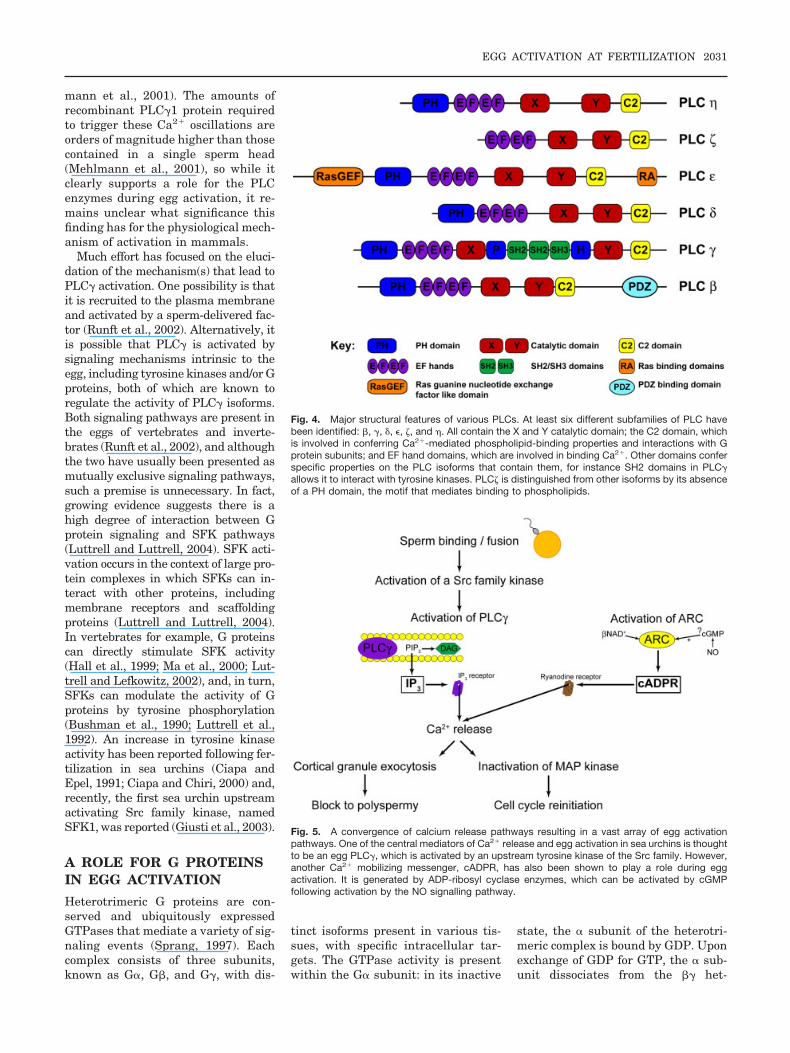
mann et al., 2001). The amounts ofrecombinant PLC�1 protein requiredto trigger these Ca2� oscillations areorders of magnitude higher than thosecontained in a single sperm head(Mehlmann et al., 2001), so while itclearly supports a role for the PLCenzymes during egg activation, it re-mains unclear what significance thisfinding has for the physiological mech-anism of activation in mammals.
Much effort has focused on the eluci-dation of the mechanism(s) that lead toPLC� activation. One possibility is thatit is recruited to the plasma membraneand activated by a sperm-delivered fac-tor (Runft et al., 2002). Alternatively, itis possible that PLC� is activated bysignaling mechanisms intrinsic to theegg, including tyrosine kinases and/or Gproteins, both of which are known toregulate the activity of PLC� isoforms.Both signaling pathways are present inthe eggs of vertebrates and inverte-brates (Runft et al., 2002), and althoughthe two have usually been presented asmutually exclusive signaling pathways,such a premise is unnecessary. In fact,growing evidence suggests there is ahigh degree of interaction between Gprotein signaling and SFK pathways(Luttrell and Luttrell, 2004). SFK acti-vation occurs in the context of large pro-tein complexes in which SFKs can in-teract with other proteins, includingmembrane receptors and scaffoldingproteins (Luttrell and Luttrell, 2004).In vertebrates for example, G proteinscan directly stimulate SFK activity(Hall et al., 1999; Ma et al., 2000; Lut-trell and Lefkowitz, 2002), and, in turn,SFKs can modulate the activity of Gproteins by tyrosine phosphorylation(Bushman et al., 1990; Luttrell et al.,1992). An increase in tyrosine kinaseactivity has been reported following fer-tilization in sea urchins (Ciapa andEpel, 1991; Ciapa and Chiri, 2000) and,recently, the first sea urchin upstreamactivating Src family kinase, namedSFK1, was reported (Giusti et al., 2003).
A ROLE FOR G PROTEINSIN EGG ACTIVATION
Heterotrimeric G proteins are con-served and ubiquitously expressedGTPases that mediate a variety of sig-naling events (Sprang, 1997). Eachcomplex consists of three subunits,known as G�, G�, and G�, with dis-
tinct isoforms present in various tis-sues, with specific intracellular tar-gets. The GTPase activity is presentwithin the G� subunit: in its inactive
state, the � subunit of the heterotri-meric complex is bound by GDP. Uponexchange of GDP for GTP, the � sub-unit dissociates from the �� het-
Fig. 4. Major structural features of various PLCs. At least six different subfamilies of PLC havebeen identified: �, �, �, �, �, and �. All contain the X and Y catalytic domain; the C2 domain, whichis involved in conferring Ca2�-mediated phospholipid-binding properties and interactions with Gprotein subunits; and EF hand domains, which are involved in binding Ca2�. Other domains conferspecific properties on the PLC isoforms that contain them, for instance SH2 domains in PLC�allows it to interact with tyrosine kinases. PLC� is distinguished from other isoforms by its absenceof a PH domain, the motif that mediates binding to phospholipids.
Fig. 5. A convergence of calcium release pathways resulting in a vast array of egg activationpathways. One of the central mediators of Ca2� release and egg activation in sea urchins is thoughtto be an egg PLC�, which is activated by an upstream tyrosine kinase of the Src family. However,another Ca2� mobilizing messenger, cADPR, has also been shown to play a role during eggactivation. It is generated by ADP-ribosyl cyclase enzymes, which can be activated by cGMPfollowing activation by the NO signalling pathway.
EGG ACTIVATION AT FERTILIZATION 2031

erodimer, and each component is freeto interact with and regulate the ac-tivity of distinct downstream targets.GTP hydrolysis results in the inacti-vation of the � subunit, which thenre-associates with the �� heterodimerto reconstitute the inactive heterotri-meric complex.
The involvement of G proteins inCa2� release and egg activationevents at fertilization has been sus-pected since PLC was shown to induceCa2� release via the IP3 pathway (Fig.5). Various data support a role for Gprotein signaling in the induction ofCa2� release and egg activation, butthe exact mechanism of action, or thecontribution of each G protein, re-mains poorly understood. Early stud-ies showed that sperm activates Gproteins that stimulate the productionof IP3, leading to Ca2� release andcortical granule exocytosis in sea ur-chins (Turner et al., 1986). Activationof G proteins via the non-hydrolysableGTP analog GTP-�S was also shownto cause Ca2� release and corticalgranule exocytosis in mammalian oo-cytes and in frog and echinoderm eggs(Cran et al., 1988; Jaffe et al., 1988;Miyazaki, 1988; Crossley et al., 1991;Fissore and Robl, 1994). In contrast,injection of the G protein antagonistGDP-�S inhibited Ca2� transients inhamster and rabbit oocytes (Miyazaki1988; Fissore and Robl, 1994). Lateron, the G protein activator AlF4 wasshown to induce cortical granule exo-cytosis in mouse oocytes (Tahara etal., 1996).
The function of G proteins was ad-dressed in a more specific manner us-ing pertussis and cholera toxins,which inhibit G�i and activate G�s,respectively. In mammals, neitherpertussis nor cholera toxin had an ef-fect on Ca2� release (Miyazaki, 1988;Williams et al., 1992; Moore et al.,1994). This suggested that if G� pro-teins were involved, they must belongto a pertussis and cholera toxin insen-sitive subfamily, which includes G�qand G�11. In frogs, neither G�q norG�11 are essential for egg activation(Runft et al., 2002), but in sea urchins,G�q and G�s contribute to Ca2� re-lease pathways (Voronina and Wessel,2004). Furthermore, it has been estab-lished that the effects of G�q and G�ssignaling are transduced by the re-lease of G�� subunits rather than by
activation of specific G� subunits (Vo-ronina and Wessel, 2004).
THE ROLE OF NITRICOXIDE AND CYCLIC ADPRIBOSE DURINGFERTILIZATION
Despite the prominence of the PI sig-naling pathway in models of egg acti-vation, increasing evidence suggeststhat other intracellular Ca2� signal-ing pathways also play importantroles. The important Ca2� mobilizingmessenger cyclic ADP-ribose (cADPR)was first identified using the sea ur-chin egg homogenate assay (Clapperet al., 1987). Heparin, an IP3 receptorantagonist, does not prevent the Ca2�
wave during sea urchin fertilization(Rakow and Shen, 1990; Crossley etal., 1991), an event that is shaped bycADPR (Fig. 2) (Galione et al., 1993;Lee et al., 1993). Further studies insea urchin egg homogenates revealedthat cADPR mediates Ca2� releasevia the ryanodine receptor (Fig. 5)(Galione et al., 1991; Lee, 1993; Lee etal., 1995) (Fig. 5). The timing ofcADPR production suggested that itmight have a role during the laterphase of the Ca2� transient (Leckie etal., 2003). In line with this result, mi-croinjection of an antagonist ofcADPR, 8-amino-cADPR, decreasedthe duration of the fertilization Ca2�
transient, suggesting that cADPRplays a major part in sustaining theCa2� signal. This is in contrast to IP3
production, which precedes, and coin-cides with, the fertilization Ca2�
wave, but which is short lived (Ciapaand Whitaker, 1986; Carroll et al.,1999; Thaler et al., 2004).
Precisely how sperm-induced eggactivation leads to an increase in eggcADPR levels is at present unknown.cADPR is believed to be synthesizedfrom �NAD� by ADP ribosyl cyclase.Most studies have focused on a role forcGMP and nitric oxide (NO) in medi-ating the increase in cADPR levels atfertilization via the activation of ADP-ribosyl cyclase. Injection of cGMP intosea urchin eggs resulted in a Ca2�
transient most closely resembling thatof both fertilization (Whalley et al.,1992) and cADPR-induced prolonga-tion of the Ca2� signal (Galione et al.,1993; Lee et al., 1993; Leckie et al.,2003). NO, by virtue of its activation
of soluble guanylyl cyclase and hencecGMP production, could also evokeCa2� signals in sea urchin eggs viathe cADPR signaling pathway (Will-mott et al., 1996). Furthermore, inhib-itors of ADP-ribosyl cyclase such asnicotinamide, and inhibitors of cGMP-dependent protein kinases inhibitedcGMP-induced Ca2� release (Sethi etal., 1996). These results indicate thatcGMP mobilizes Ca2� in sea urchineggs by initiating a signaling pathwaypromoting cADPR synthesis (Fig. 5).
An important question still to be an-swered is where NO functions in thehierarchy of messengers involved inactivation of the egg. While activationof Ca2� release via the NO pathwayand the ryanodine receptor does notlead to full egg activation in mamma-lian eggs, one report did suggest thatNO may be the initiator of Ca2� re-lease in sea urchins (Kuo et al., 2000).This was based on the observationthat microinjection of NO donors or ofnitric oxide synthase (NOS), the en-zyme responsible for NO production,was able to activate the egg, whereasmicroinjection of NO scavengers in-hibited egg activation after fertiliza-tion. A more recent study, where lev-els of Ca2� and NO were measuredsimultaneously, shows that NO-medi-ated Ca2� release through the ryano-dine receptor is a late event in thefertilization response in sea urchins(Leckie et al., 2003), suggesting thatNO is not responsible for the initiationof the Ca2� wave. Instead, these au-thors propose that activation of theNO pathway is needed to maintainthe duration of the Ca2� transient ini-tially elicited via the IP3 pathway.Nevertheless, the overall resultsclearly show that, at least in inverte-brates, NO-mediated signaling is animportant component of normal Ca2�
dynamics at fertilization.Whether the NO pathway has any
role in Ca2� release in vertebrates re-mains to be conclusively established.One study using an antibody that rec-ognizes type 2 and 3 ryanodine recep-tors, both of which are present in themouse egg, concluded that Ca2� re-lease via these two receptor subtypeswas not necessary for Ca2� release inthis species (Ayabe et al., 1995).Whether the type 1 ryanodine recep-tor could have an overlapping functionthough, it is not clear. A different
2032 PARRINGTON ET AL.

study that again simultaneously mea-sured Ca2� and NO levels in the eggshowed that NO has no role at least inthe initiation of Ca2� release (Hyslopet al., 2001). Whether or not NO playsa role in the propagation of the Ca2�
wave, as it seems to do in inverte-brates (Leckie et al.. 2003), is still un-known.
An important unresolved questionis what signaling mechanisms lie up-stream of NO. NO is produced by thefamily of nitric oxide synthases(NOS). Upon activation, these en-zymes catalyze the conversion of theamino acid L-arginine into NO andcitrulline. Three distinct isoforms ofNOS have been identified in mamma-lian tissues, including neuronal NOS(nNOS), which functions in thenervous system, endothelial NOS(eNOS), which functions in the cardio-vascular system, and inducible NOS(iNOS), which functions in the im-mune system (Moncada et al., 1991;Dinerman et al., 1993; Kerwin et al.,1995). The relative contribution ofeach isoform to Ca2� release path-ways at fertilization remains unclear.Two distinct NOS cDNAs that are ex-pressed in testes and ovary have beenidentified in sea urchins (Cox et al.,2001; Sodergren, 2006).
One proposed mechanism of activa-tion for NOS proteins is via PKC-me-diated phosphorylation (Matsubara etal., 2003), which places them down-stream of PLC activation. Recall thatcleavage of PIP2 by PLC not only pro-duces IP3, which induces Ca2� releasevia the IP3 receptor, but also DAG,which activates PKC (Fig. 5) (Liu andHeckman, 1998; Rhee, 2001). Thus,upon activation by DAG, PKC canphosphorylate NOS, which in turnproduces NO and stimulates Ca2� re-lease via the ryanodine receptor (Fig.5). The fact that PLCs produce bothIP3 and DAG upon cleavage of PIP2
again places them at center stage be-cause it suggests that PLCs are regu-latory nodes that can induce Ca2� re-lease via both the IP3 and NOpathways. Though it is tempting tospeculate that a single protein mayinitiate all egg activation events, it isalso likely that signal transductionpathways are complex networks of in-teractions that integrate and trans-duce diverse cellular signals. One
point of convergence for diverse cellu-lar signals is NOS.
NOS can be activated not only byPKC-mediated phosphorylation, butalso by Ca2� and calmodulin (Matsub-ara et al., 2003). In fact, binding tocalmodulin via the highly conservedcalmodulin-binding domains is essen-tial for NOS function (Cox et al., 2001;Li and Poulos, 2005) and differentNOS isoforms bind calmodulin withdifferent affinities. While nNOS andeNOS, which are expressed in a con-stitutive manner, bind calmodulin ina Ca2�-dependent manner, iNOS,which is expressed upon induction ofthe immune response, binds calmodu-lin constitutively at physiologicalCa2� levels of about 100 nM (Li andPoulos, 2005).
Indirect evidence suggests that theNOS isoforms regulating NO produc-tion in eggs are nNOS or eNOS, orboth. First, nNOS and eNOS are in-hibited by the specific NOS antagonistNG-nitro-L-arginine (L-NAME), butiNOS is not (Li and Poulos, 2005). Insea urchin eggs, NOS activity can besuppressed using L-NAME (Kuo et al.,2000). Second, given that egg activa-tion is a Ca2�-dependent process, it islikely that NO production in the egg ismodulated by enzymes that are re-sponsive to Ca2� levels. Both nNOSand eNOS, for example, are sensitiveto Ca2� levels in a calmodulin-depen-dent manner (Michel and Feron,1997). Calmodulin can regulate theactivity of many other proteins at fer-tilization, including calmodulin-de-pendent kinase II (CaM kinase II).CaM kinase II activity undergoes os-cillations following fertilization inmouse eggs (Markoulaki et al., 2003,2004), and this activity is importantfor cortical granule exocytosis andprogression through the cell cycle,based on evidence that shows inhibi-tors of calmodulin or CaM kinase IIblock both latter processes (Abbottand Ducibella, 2001; Tatone et al.,2002). Notably, CaM kinase II has theability to regulate NOS function(Michel and Feron, 1997; Schneider etal., 2003). NOS proteins can also bephosphorylated by tyrosine kinaseslike Src (Li et al., 2004), suggestingpossible cross-talk between variousputative regulators of the Ca2� re-lease response in invertebrate eggs.Little is known about NOS function in
eggs, so a thorough understanding ofthe role of the NO pathway at fertili-zation will only be achieved once weidentify the various mechanisms bywhich NOS can be activated in thiscell.
A sea urchin homolog of the seventrans-membrane G protein-coupledreceptor for histamine (suH1R) wasrecently identified on the cell surfaceof sea urchin eggs that leads to NOproduction (Leguia and Wessel, 2006).Treatment of eggs with histaminecauses fluctuations in the resting lev-els of NO in the egg, while antagonistsor antibodies of H1R inhibit the rise ofNO normally observed at fertilization.Thus, inhibition of suH1R function de-creases the maintenance, but not theamplitude, of the Ca2� transient andsuggests that it is an integral part ofthe overall pathway leading to egg ac-tivation at fertilization in sea urchins.
The suH1R of the egg impinges onmultiple different signaling path-ways, including heterotrimeric G-pro-tein cascades, NOS activation, andCa2� release. It is unlikely that thephenotypes resulting from modifyingsuH1R activity affected only one of itstarget signaling pathway. For exam-ple, although Ca2� is clearly a targetof suH1R receptor activation, a result-ant phenotype must be interpreted inthe context of, at least, altered NOdynamics, and heterotrimeric G-pro-tein activities, the vast majority ofwhich are currently unknown. This isfurther complicated by the fact thateach animal egg has unique require-ments for each of these pathways, andminor alterations in the amplitude,timing, duration, and frequency ofeven one pathway such as Ca2�, canhave major consequences on fertiliza-tion and early development (Ozil etal., 2005). Thus, is it not possible toconclude that the decreased exocytosisof cortical granules observed whensuH1R is inhibited is solely the resultof the decreased Ca2� levels. This is-sue is further complicated in light ofrecent evidence showing that the G��subunits of heterotrimeric G proteinscan inhibit the Ca2�-induced exocy-totic machinery that mediates mem-brane fusion (Blackmer et al., 2005;Gerachshenko et al., 2005) perhaps bycompeting with the Ca2� sensor syn-aptotagmin for binding of the C-termi-nal tail of SNAP-25, and/or by
EGG ACTIVATION AT FERTILIZATION 2033

regulating fusion pore dynamics(Schneider, 2001). Added to this equa-tion are the effects of NO, which in-cludes nitrosylation of other receptorand signaling proteins, so that inap-propriate histamine receptor activa-tion or repression could lead to themisregulation of multiple cellular tar-gets, including Ca2�-sensing proteins(Tao and English, 2003), ryanodine re-ceptors (Xu et al., 1998), Ca2� chan-nels (Poteser et al., 2001), and fusionpore dynamics (Schneider, 2001;Thorn et al., 2004).
THE ROLE OF THE NAADPSIGNALING PATHWAY ATFERTILIZATION
A second important Ca2� signallingmessenger has been identified usingthe sea urchin egg homogenate assay,nicotinic acid adenine dinucleotidephosphate (NAADP) (Lee and Aarhus,1995; Perez Terzic et al., 1995). Stud-ies in the sea urchin egg first demon-strated that the NAADP-sensitiveCa2� store has a distinct sub-cellularlocalization from that of the IP3 orRyR/cADPR pathways (Lee and Aar-hus, 1995; Genazzani et al., 1997) andthat NAADP appears to target anacidic organelle such as the lysosome(Fig. 6). Stratification of egg or-ganelles showed that the NAADP-sen-sitive store segregated to the oppositepole from the endoplasmic reticularstores (Lee and Aarhus, 2000). Fur-thermore, the use of a lysosome-dis-rupting agent (glycylphenylalanine2-naphthylamide, GPN), inhibitedNAADP from mobilising Ca2� in bothmicrosomes and intact eggs, withoutaffecting the Ca2� released by IP3 orcADPR (Churchill et al., 2002). Addi-tional evidence for a lysosome-like or-ganelle NAADP-sensitive store wasprovided from fractionation of egg ho-mogenates, which showed that thestore co-distributed with lysosomalenzyme markers (Churchill et al.,2002). A key goal in the Ca2� signal-ling field is now to identify theNAADP receptor.
NAADP also appears to play a roleduring fertilization in sea urchins. Mi-croinjection of NAADP in intact seaurchin eggs induced a Ca2� signalthat, as previously found in sea urchinegg homogenates, was not blocked byheparin or by prior desensitization to
IP3 or cADPR (Perez Terzic et al.,1995). Spatially resolved measure-ments of Ca2� in intact sea urchineggs (Churchill et al., 2003) and ma-tured starfish oocytes (Santella et al.,2000; Lim et al., 2001; Moccia et al.,2003) revealed that NAADP-inducedCa2� release is biphasic, consisting ofan initial Ca2� increase confined tothe perimeter of the egg that is fol-lowed by a global Ca2� increase,which extended through the egg cyto-plasm. Removal of extracellular Ca2�
or addition of the Ca2� channelblocker (Cd2�) did not affect thesecond Ca2� increase (consistentwith Ca2� release from intracellularstores), whereas the localized Ca2� in-crease at the plasma membrane wasabolished (Churchill et al., 2003).These data provide evidence thatNAADP may stimulate Ca2� entryacross the plasma membrane, in addi-tion to mobilizing intracellular Ca2�
stores (Patel, 2004).Sperm, like NAADP, also stimulate
a localized Ca2� increase at theplasma membrane, termed the corti-cal flash (Fig. 2), which involves acti-vation of voltage-sensitive Ca2� chan-nels (Shen and Buck, 1993) that mayaid in establishing the so-called fastblock to polyspermy (Jaffe, 1976; Mc-Culloh and Chambers, 1992). Inacti-vation of NAADP receptors abolishesthe cortical flash at fertilization, sug-gesting that NAADP and sperm-in-duced Ca2� entry are mediated bysimilar means (Churchill et al., 2003).Supramaximal, micromolar NAADPlevels were found in Lytechinus pictussea urchin sperm (Billington et al.,2002) and importantly these levelsmarkedly increase (4-fold) upon con-tact with egg jelly (Churchill et al.,2003). These data raise the possibil-ity that a concentrated source ofNAADP is delivered into the egg as abolus of messenger when the spermfuses with the egg. Measurement ofNAADP levels in fertilized eggs indi-cates that the increase in NAADP atfertilization occurs in two phases: aninitial surge, which may be sperm-derived, and a second later increasepeaking 4 –5 min post-fertilization(Churchill et al., 2003), probably re-sulting from synthesis of NAADPwithin the egg.
THE SEA URCHIN EGG ASA MODEL FOR EGGACTIVATION AND CALCIUMSIGNALING IN MAMMALS
Ultimately, the use of model organ-isms for studying processes such asfertilization and development as-sumes that findings made using suchorganisms will help further under-standing of these processes in our ownhuman bodies. Research into themechanism of egg activation in seaurchins was of key importance in thelight it shed on the central role ofCa2� as the trigger of this process. Atthe same time, the last few years haveled to a recognition that there may becritical differences in the signalingcomponents employed to generate aCa2� signal in sea urchins comparedto mammals. Thus, while it is nowwidely accepted that a key feature ofechinoderm egg activation is the acti-vation of an egg PLC� by a tyrosinekinase mechanism, such a mechanismdoes not appear to be conserved inmammalian eggs. Similarly, mamma-lian egg activation is thought to betriggered by a sperm phospholipase C,PLC�, yet sequencing of the sea urchingenome has failed to identify an or-tholog. These differences may reflectvery real divergences in the demandsmade upon the egg activation signal-ing machinery in echinoderms andmammals.
It would be unwise, however, tojudge the studies of signaling pro-cesses in the sea urchin egg purely interms of their importance for our un-derstanding of fertilization. The seaurchin egg has been essential for un-derstanding intracellular Ca2� signal-ing in general. Sea urchin egg homog-enates, for example, have allowed theinitial identification of the novel Ca2�
mobilizing messengers cADPR andNAADP, which play vital roles in theregulation of processes in the mam-malian body as diverse as insulin se-cretion by the pancreas, cardiacfunction, and nerve growth and differ-entiation.
ACKNOWLEDGMENTSThe authors appreciate the criticalreading of this manuscript by JulianWong, and acknowledge support fortheir laboratories from the National
2034 PARRINGTON ET AL.

Science Foundation and the NationalInstitutes of Health (G.M.W.), theMedical Research Council (J.P.), andthe Wellcome Trust (A.G. and J.P.).
REFERENCES
Abbott AL, Ducibella T. 2001. Calcium andthe control of mammalian cortical gran-ule exocytosis. Front Biosci 6:D792–806.
Ayabe T, Kopf GS, Schultz RM. 1995. Reg-ulation of mouse egg activation: presenceof ryanodine receptors and effects of mi-croinjected ryanodine and cyclic ADP ri-bose on uninseminated and inseminatedeggs. Development 121:2233–2244.
Billington RA, Ho A, Genazzani AA. 2002.Nicotinic acid adenine dinucleotide phos-phate (NAADP) is present at micromolarconcentrations in sea urchin spermato-zoa. J Physiol 544:107–112.
Blackmer T, Larsen EC, Takahashi M,Martin TF, Alford S, Hamm HE. 2005. Gprotein betagamma directly regulatesSNARE protein fusion machinery for se-cretory granule exocytosis. Nat Neurosci8:421–425.
Bushman WA, Wilson LK, Luttrell DK,Moyers JS, Parsons SJ.. 1990. Overex-pression of c-src enhances beta-adrener-gic-induced cAMP accumulation. ProcNatl Acad Sci USA 87:7462–7466.
Camps M, Carozzi A, Schnabel P, ScheerA, Parker PJ, Gierschik P. 1992.Isozyme-selective stimulation of phos-pholipase C-beta 2 by G protein betagamma-subunits. Nature 360:684–686.
Carroll DJ, Ramarao CS, Mehlmann LM,Roche S, Terasaki M, Jaffe LA. 1997.Calcium release at fertilization in star-fish eggs is mediated by phospholipaseCgamma. J Cell Biol 138:1303–1311.
Carroll DJ, Albay DT, Terasaki M, JaffeLA, Foltz KR. 1999. Identification of PL-Cgamma-dependent and -independentevents during fertilization of sea urchineggs. Dev Biol 206:232–247.
Churchill GC, Okada Y, Thomas JM,Genazzani AA, Patel S, Galione A. 2002.NAADP mobilizes Ca2� from reservegranules, a lysosome-related organelle,in sea urchin eggs. Cell 111:703–708.
Churchill GC, O’Neill JS, Masgrau R, PatelS, Thomas JM, Genazzani AA, GalioneA. 2003. Sperm deliver a new secondmessenger. NAADP Curr Biol 13:125–128.
Ciapa B, Chiri S. 2000. Egg activation: up-stream of the fertilization calcium sig-nal. Biol Cell 92:215–233.
Ciapa B, Epel D. 1991. A rapid change inphosphorylation on tyrosine accompa-nies fertilization of sea urchin eggs.FEBS Lett 295:167–170.
Ciapa B, Whitaker M. 1986. Two phases ofinositol polyphosphate and diacylglyc-erol production at fertilisation. Febs Lett195:347–351.
Clapper DL, Walseth TF, Dargie PJ, LeeHC. 1987. Pyridine nucleotide metabo-lites stimulate calcium release from seaurchin egg microsomes desensitized to
inositol trisphosphate. J Biol Chem 262:9561–9568.
Coward K, Campos-Mendoza A, LarmanM, Hibbitt O, McAndrew B, Bromage N,Parrington J. 2003. Teleost fish sperma-tozoa contain a cytosolic protein factorthat induces calcium release in sea ur-chin egg homogenates and triggers cal-cium oscillations when injected intomouse oocytes. Biochem Biophys ResCommun 305:299–304.
Coward K, Ponting CP, Chang HY, HibbittO, Savolainen P, Jones KT, Parrington J.2005. Phospholipase Czeta, the trigger ofegg activation in mammals, is present ina non-mammalian species. Reproduction130:157–163.
Cox RL, Mariano T, Heck DE, Laskin JD,Stegeman JJ. 2001. Nitric oxide syn-thase sequences in the marine fishStenotomus chrysops and the sea urchinArbacia punctulata, and phylogeneticanalysis of nitric oxide synthase calmod-ulin-binding domains. Comp BiochemPhysiol B Biochem Mol Biol 130:479–491.
Cran DG, Moor RM, Irvine RF. 1988. Ini-tiation of the cortical reaction in hamsterand sheep oocytes in response to inositoltrisphosphate. J Cell Sci 91:139–144.
Creton R, Jaffe LF. 2001 Chemilumines-cence microscopy as a tool in biomedicalresearch. Biotechniques 31:1098–1100.
Crossley I, Whalley T, Whitaker M. 1991.Guanosine 5-thiotriphosphate maystimulate phosphoinositide messengerproduction in sea urchin eggs by a differ-ent route than the fertilizing sperm. CellRegul 2:121–133.
Dale B. 1988. Primary and secondary mes-sengers in the activation of ascidianeggs. Exp Cell Res 177:205–211.
Dale B, DeFelice LJ, Ehrenstein G. 1985.Injection of a soluble sperm fraction intosea-urchin eggs triggers the cortical re-action. Experientia 41:1068–1070.
Derbes AA. 1847. Observations sur lemechanisme et les phenomenes qui ac-compagnent la formation de l’embryonchez l’oursin comestible. Ann Sci NatZool 8:80–98.
Dinerman JL, Lowenstein CJ, Snyder SH.1993. Molecular mechanisms of nitric ox-ide regulation. Potential relevance tocardiovascular disease. Circ Res 73:217–222.
Dong JB, Tang TS, Sun FZ. 2000. Xenopusand chicken sperm contain a cytosolicsoluble protein factor which can triggercalcium oscillations in mouse eggs. Bio-chem Biophys Res Commun. 268:947–951.
Dupont G, McGuinness OM, Johnson MH,Berridge MJ, Borgese F. 1996. Phospho-lipase C in mouse oocytes: characteriza-tion of beta and gamma isoforms andtheir possible involvement in sperm-in-duced Ca2� spiking. Biochem J 316:583–591.
Evans JP, Kopf GS. 1998. Molecular mech-anisms of sperm-egg interactions andegg activation. Andrologia 30:297–307.
Fissore RA, Robl JM. 1994. Mechanism ofcalcium oscillations in fertilized rabbiteggs. Dev Biol 166:634–642.
Flaherty SP, Payne D, Matthews CD. 1998.Fertilization failures and abnormal fer-tilization after intracytoplasmic sperminjection. Hum Reprod Suppl 1:155–164.
Fujimoto S, Yoshida N, Fukui T, AmanaiM, Isobe T, Itagaki C, Izumi T, Perry AC.2004. Mammalian phospholipase Czetainduces oocyte activation from the spermperinuclear matrix. Dev Biol 274:370–383.
Fulton BP, Whittingham DG. 1978. Activa-tion of mammalian oocytes by intracellu-lar injection of calcium. Nature 273:149–151.
Galione A, Lee HC, Busa WB. 1991. Ca2�-induced Ca2� release in sea urchin egghomogenates: modulation by cyclic ADP-ribose. Science 253:1143–1146.
Galione A, McDougall A, Busa WB, Will-mott N, Gillot I, Whitaker M. 1993. Re-dundant mechanisms of calcium-inducedcalcium release underlying calciumwaves during fertilization of sea urchineggs. Science 261:348–352.
Genazzani AA, Mezna M, Dickey DM,Michelangeli F, Walseth TF, Galione A.1997. Pharmacological properties of theCa2�-release mechanism sensitive toNAADP in the sea urchin egg. Br J Phar-macol 121:1489–1495.
Gerachshenko T, Blackmer T, Yoon EJ,Bartleson C, Hamm HE, Alford S. 2005.Gbetagamma acts at the C terminus ofSNAP-25 to mediate presynaptic inhibi-tion. Nat Neurosci 8:597–605.
Giusti AF, Carroll DJ, Abassi YA, TerasakiM, Foltz KR, Jaffe LA. 1999. Require-ment of a Src family kinase for initiatingcalcium release at fertilization in star-fish eggs. J Biol Chem 274:29318–29322.
Giusti AF, O’Neill FJ, Yamasu K, FoltzKR, Jaffe LA. 2003. Function of a seaurchin egg Src family kinase in initiatingCa2� release at fertilization. Dev Biol256:367–378.
Hall RA, Premont RT, Lefkowitz RJ. 1999.Heptahelical receptor signaling: beyondthe G protein paradigm. J Cell Biol 145:927–932.
Harlan JE, Hajduk PJ, Yoon HS, FesikSW. 1994. Pleckstrin homology domainsbind to phosphatidylinositol-4,5-bisphos-phate. Nature 371:168–170.
Heilbrunn LV. 1952. An outline of generalphysiology, 3rd ed. Philadelphia: Saun-ders.
Heindryckx B, Van der Elst J, De Sutter P,Dhont M. 2005. Treatment option forsperm- or oocyte-related fertilizationfailure: assisted oocyte activation follow-ing diagnostic heterologous ICSI. HumReprod 20:2237–2241.
Hyslop LA, Carroll M, Nixon VL, McDou-gall A, Jones KT. 2001. Simultaneousmeasurement of intracellular nitric ox-ide and free calcium levels in chordateeggs demonstrates that nitric oxide hasno role at fertilization. Dev Biol 234:216–230.
Ireland DR. Abraham WC. 2002. Group ImGluRs increase excitability of hip-
EGG ACTIVATION AT FERTILIZATION 2035

pocampal CA1 pyramidal neurons by aPLC-independent mechanism. J Neuro-physiol 88:107–116.
Jaffe LA. 1976. Fast block to polyspermy insea urchin eggs is electrically mediated.Nature 261:68–71.
Jaffe LF. 1983. Sources of calcium in eggactivation: a review and hypothesis. DevBiol 99:265–276.
Jaffe LA. 1990. First messengers at fertil-ization. J Reprod Fertil Suppl 42:107–116.
Jaffe LF. 1991. The path of calcium in cy-tosolic calcium oscillations: a unifyinghypothesis. Proc Natl Acad Sci USA 88:9883–9887.
Jaffe LA, Turner PR, Kline D, Kado RT,Shilling F. 1988. G-proteins and egg ac-tivation. Cell Differ Dev 25(Suppl):15–18.
Jones KT, Soeller C, Cannell MB. 1998.The passage of Ca2� and fluorescentmarkers between the sperm and egg af-ter fusion in the mouse. Development.125:4627–4635.
Jones KT, Matsuda M, Parrington J, Ka-tan M, Swann K. 2000. Different Ca2�-releasing abilities of sperm extractscompared with tissue extracts and phos-pholipase C isoforms in sea urchin egghomogenate and mouse eggs. Biochem J346:743–749.
Kerwin JF Jr, Lancaster JR Jr, FeldmanPL. 1995. Nitric oxide: a new paradigmfor second messengers. J Med Chem 38:4343–4362.
Kline D. 1988. Calcium-dependent eventsat fertilization of the frog egg: injectionof a calcium buffer blocks ion channelopening, exocytosis, and formation ofpronuclei. Dev Biol 126:346–361.
Kline D, Kline JT. 1992. Repetitive calciumtransients and the role of calcium in exo-cytosis and cell cycle activation in themouse egg. Dev Biol 149:80–89.
Knott JG, Kurokawa M, Fissore RA,Schultz RM, Williams CJ. 2005. Trans-genic RNA interference reveals role formouse sperm phospholipase Czeta intriggering Ca2� oscillations during fer-tilization. Biol Reprod 72:992–996.
Kouchi Z, Shikano T, Nakamura Y,Shirakawa H, Fukami K, Miyazaki S.2005. The role of EF-hand domains andC2 domain in regulation of enzymaticactivity of phospholipase Czeta. J BiolChem 280:21015–21021.
Kuo RC, Baxter GT, Thompson SH,Stricker SA, Patton C, Bonaventura J,Epel D. 2000. NO is necessary and suffi-cient for egg activation at fertilization.Nature 406:633–636.
Kurokawa M, Sato K, Wu H, He C, MalcuitC, Black SJ, Fukami K, Fissore RA.2005. Functional, biochemical, and chro-matographic characterization of the com-plete [Ca2�]i oscillation-inducing activ-ity of porcine sperm. Dev Biol 285:376–392.
Kyozuka K, Deguchi R, Mohri T, MiyazakiS. 1998. Injection of sperm extract mim-ics spatiotemporal dynamics of Ca2� re-sponses and progression of meiosis at
fertilization of ascidian oocytes. Develop-ment 125:4099–4105.
Larman MG, Saunders CM, Carroll J, LaiFA, Swann K. 2004. Cell cycle-depen-dent Ca2� oscillations in mouse em-bryos are regulated by nuclear targetingof PLCzeta. J Cell Sci 117:2513–2521.
Lawrence Y, Whitaker M, Swann K. 1997.Sperm-egg fusion is the prelude to theinitial Ca2� increase at fertilization inthe mouse. Development 124:233–241.
Leckie C, Empson R, Becchetti A, ThomasJ, Galione A, Whitaker M. 2003. The NOpathway acts late during the fertiliza-tion response in sea urchin eggs. J. Biol.Chem 278:12247–12254.
Lee CW, Lee KH, Lee SB, Park D, RheeSG. 1994. Regulation of phospholipaseC-beta 4 by ribonucleotides and the al-pha subunit of Gq. J Biol Chem 269:25335–25338.
Lee HC. 1993. Potentiation of calcium- andcaffeine-induced calcium release by cy-clic ADP-ribose. J Biol Chem 268:293–299.
Lee HC, Aarhus R. 1995. A derivative ofNAADP mobilizes calcium stores insen-sitive to inositol trisphosphate and cyclicADP-ribose. J Biol Chem 270:2152–2157.
Lee HC, Aarhus R. 2000. Functional visu-alization of the separate but interactingcalcium stores sensitive to NAADP andcyclic ADP-ribose. J Cell Sci 113:4413–4420.
Lee HC, Aarhus R, Graeff RM. 1993. Cal-cium mobilization by dual receptors dur-ing fertilization of sea urchin eggs. Sci-ence 261:352–355.
Lee HC, Aarhus R, Graeff RM. 1995. Sen-sitization of calcium-induced calcium re-lease by cyclic ADP-ribose and calmodu-lin. J Biol Chem 270:9060–9066.
Lee SJ, Madden PJ, Shen SS. 1998.U73122 blocked the cGMP-induced cal-cium release in sea urchin eggs. Exp CellRes 242:328–340
Leguia M, Wessel GM. 2006. The Hista-mine H1 receptor activates the nitric ox-ide pathway at fertilization. Mol ReprodDev 73:1550–1563.
Li H. Poulos TL. 2005. Structure-functionstudies on nitric oxide synthases. J InorgBiochem 99:293–305.
Li X, Lerea KM, Li J, Olson SC. 2004. Srckinase mediates angiotensin II-depen-dent increase in pulmonary endothelialnitric oxide synthase. Am J Respir CellMol Biol 31:365–372.
Lillie FR. 1913. The mechanism of fertili-zation. Science 38:524–528.
Lim D, Kyozuka K, Gragnaniello G, Car-afoli E, Santella L. 2001. NAADP� ini-tiates the Ca2� response during fertiliza-tion of starfish oocytes. Faseb. J 15: 2257–2267.
Liu WS, Heckman CA. 1998. The sevenfoldway of PKC regulation. Cell Signal 10:529–542.
Loeb J. 1899. On the nature of the processof fertilization and the artificial produc-tion of normal larvae (Plutei) from theunfertilized eggs of the sea urchin. Am JPhysiol 3:135–138.
Luttrell DK, Luttrell LM. 2004. Not sostrange bedfellows: G-protein-coupledreceptors and Src family kinases. Onco-gene 23:7969–7978.
Luttrell DK, Hausdorff WP, Moyers JE,Gilmer TM, Parsons SJ, Caron MG,Lefkowitz RJ. 1992. Overexpression ofpp60c-src is associated with altered reg-ulation of adenylyl cyclase. Cell Signal4:531–541.
Luttrell LM, Lefkowitz RJ. 2002. The roleof beta-arrestins in the termination andtransduction of G-protein-coupled recep-tor signals. J Cell Sci 115:455–465.
Ma YC, Huang J, Ali S, Lowry W, HuangXY. 2000. Src tyrosine kinase is a noveldirect effector of G proteins. Cell 102:635–646.
Mahutte NG, Arici A. 2003. Failed fertili-zation: is it predictable? Curr Opin Ob-stet Gynecol 15:211–218.
Markoulaki S, Matson S, Abbott AL, Duci-bella T. 2003. Oscillatory CaMKII activ-ity in mouse egg activation. Dev Biol 258:464–474.
Markoulaki S, Matson S, Ducibella T.2004. Fertilization stimulates long-last-ing oscillations of CaMKII activity inmouse eggs. Dev Biol 272:15–25.
Matsubara M, Hayashi N, Jing T, Titani K.2003. Regulation of endothelial nitric ox-ide synthase by protein kinase C. J Bio-chem (Tokyo) 133:773–781.
Mazia D. 1937. The release of calcium inArbacia eggs on fertilization. J CellComp Physiol 10:291–304.
McCulloh DH, Chambers EL. 1992. Fusionof membranes during fertilization. In-creases of the sea urchin egg’s mem-brane capacitance and membrane con-ductance at the site of contact with thesperm. J Gen Physiol 99:137–175.
Mehlmann LM, Carpenter G, Rhee SG,Jaffe LA. 1998. SH2 domain-mediatedactivation of phospholipase Cgamma isnot required to initiate Ca2� release atfertilization of mouse eggs. Dev Biol 203:221–232.
Mehlmann LM, Chattopadhyay A, Carpen-ter G, Jaffe LA. 2001. Evidence thatphospholipase C from the sperm is notresponsible for initiating Ca(2�) releaseat fertilization in mouse eggs. Dev Biol236:492–501.
Michel T. Feron O. 1997. Nitric oxide syn-thases: which, where, how, and why?J Clin Invest 100:2146–2152.
Miyazaki S. 1988. Inositol 1,4,5-trisphos-phate-induced calcium release and gua-nine nucleotide-binding protein-medi-ated periodic calcium rises in goldenhamster eggs. J Cell Biol 106:345–353.
Miyazaki S, Yuzaki M, Nakada K,Shirakawa H, Nakanishi S, Nakade S,Mikoshiba K 1992. Block of Ca2� waveand Ca2� oscillation by antibody to theinositol 1,4,5-trisphosphate receptor infertilized hamster eggs. Science 257:251–255.
Moccia F, Nusco GA, Lim D, Ercolano E,Gragnaniello G, Brown ER, Santella L.2003. Ca2� signaling and membrane cur-rent activated by cADPr in starfish oo-cytes. Pflugers Arch 446:541–552.
2036 PARRINGTON ET AL.

Moncada S, Palmer RM, Higgs EA. 1991.Nitric oxide: physiology, pathophysiol-ogy, and pharmacology. Pharmacol Rev43:109–142.
Moore GD, Ayabe T, Visconti PE, SchultzRM, Kopf GS. 1994. Roles of heterotri-meric and monomeric G proteins insperm-induced activation of mouse eggs.Development 120:3313–3323.
Moser F. 1939. Studies on a cortical layerresponse to stimulating agents in the Ar-bacia egg. I. Response to insemination. JExp Zool 80:423–445.
Nakahara M, Shimozawa M, Nakamura Y,Irino Y, Morita M, Kudo Y, Fukami K.2005. A novel phospholipase C, PL-C(eta)2, is a neuron-specific isozyme.J Biol Chem 280:29128–29134.
Noh DY, Shin SH, Rhee SG. 1995. Phos-phoinositide-specific phospholipase Cand mitogenic signaling. Biochim Bio-phys Acta 1242:99–113.
Ozil JP, Markoulaki S, Toth S, Matson S,Banrezes B, Knott JG, Schultz RM, Hu-neau D, Ducibella T. 2005. Egg activa-tion events are regulated by the durationof a sustained [Ca2�]cyt signal in themouse. Dev Biol 282:39–54.
Parrington J. 2001. Does a soluble spermfactor trigger calcium release in the eggat fertilization? J Androl 22:1–11.
Parrington J, Jones KT, Lai A, Swann K.1999. The soluble sperm factor thatcauses Ca2� release from sea-urchin(Lytechinus pictus) egg homogenatesalso triggers Ca2� oscillations after in-jection into mouse eggs. Biochem J 341:1–4.
Parrington J, Jones KT, Lai A, Swann K.2002. Phospholipase C isoforms in mam-malian spermatozoa: potential compo-nents of the sperm factor that causesCa2� release in eggs. Reproduction 123:31–39.
Patel S. 2004. NAADP-induced Ca2� re-lease: a new signalling pathway. BiolCell 96:19–28.
Pauly PJ. 1987. Controlling life: JacquesLoeb and the engineering ideal in biol-ogy. New York: Oxford University Press.
Perez-Terzic CM, Chini EN, Shen SS,Dousa TP, Clapham DE. 1995. Ca-2�release triggered by nicotinate adeninedinucleotide phosphate in intact sea ur-chin eggs. Biochem J 312:955–959.
Poteser M, Romanin C, Schreibmayer W,Mayer B, Groschner K. 2001. S-nitrosa-tion controls gating and conductance ofthe alpha 1 subunit of class C L-typeCa(2�) channels.JBiolChem276:14797–14803.
Rakow TL, Shen SS. 1990. Multiple storesof calcium are released in the sea urchinegg during fertilization. Proc Natl AcadSci USA 87:9285–9289.
Rameh L E,Cantley LC. 1999. The role ofphosphoinositide 3-kinase lipid productsin cell function. J Biol Chem 274:8347–8350.
Rhee SG. 2001. Regulation of phosphoino-sitide-specific phospholipase C. AnnuRev Biochem 70:281–312.
Ridgway EB, Gilkey JC, Jaffe LF. 1977.Free calcium increases explosively in ac-
tivating medaka eggs. Proc Natl AcadSci USA 74:623–627.
Rongish BJ, Wu W, Kinsey WH 1999. Fer-tilization-induced activation of phospho-lipase C in the sea urchin egg. Dev Biol215:147–154.
Roux MM, Townley IK, Raisch M, Reade A,Bradham C, Humphreys G, GunaratneHJ, Killian CE, Moy G, Su YH, Etten-sohn CA, Wilt F, Vacquier VD, BurkeRD, Wessel G, Foltz KR. 2006. A func-tional genomic and proteomic perspec-tive of sea urchin calcium signaling andegg activation. Dev Biol 300:416–433.
Runft LL, Jaffe LA. 2000. Sperm extractinjection into ascidian eggs signalsCa(2�) release by the same pathway asfertilization. Development 127:3227–3236.
Runft LL, Watras J, Jaffe LA. 1999. Cal-cium release at fertilization of Xenopuseggs requires type I IP(3) receptors, butnot SH2 domain-mediated activation ofPLCgamma or G(q)-mediated activationof PLCbeta. Dev Biol 214:399–411.
Runft LL, Jaffe LA, Mehlmann LM. 2002.Egg activation at fertilization: where itall begins. Dev Biol 245:237–254.
Runft LL, Carroll DJ, Gillett J, Giusti AF,O’Neill FJ, Foltz KR. 2004. Identificationof a starfish egg PLC-gamma that regu-lates Ca2� release at fertilization. DevBiol 269:220–236.
Santella L, Kyozuka K, Genazzani AA, DeRiso L, Carafoli E. 2000. Nicotinic acidadenine dinucleotide phosphate-inducedCa2� release. Interactions among dis-tinct Ca2� mobilizing mechanisms instarfish oocytes. J Biol Chem 275:8301–8306.
Sato K, Tokmakov AA, Iwasaki T, FukamiY. 2000. Tyrosine kinase-dependent acti-vation of phospholipase Cgamma is re-quired for calcium transient in Xenopusegg fertilization. Dev Biol 224:453–469.
Saunders CM, Larman MG, Parrington J,Cox LJ, Royse J, Blayney LM, Swann K,Lai FA. 2002. PLC zeta: a sperm-specifictrigger of Ca(2�) oscillations in eggs andembryo development. Development 129:3533–3544.
Schneider JC, El Kebir D, Chereau C,Lanone S, Huang XL, De Buys Roess-ingh AS, Mercier JC, Dall’Ava-SantucciJ, Dinh-Xuan AT. 2003. Involvement ofCa2�/calmodulin-dependent protein ki-nase II in endothelial NO productionand endothelium-dependent relaxation.Am J Physiol Heart Circ Physiol 284:H2311–2319.
Schneider SW. 2001. Kiss and run mecha-nism in exocytosis. J Membr Biol 181:67–76.
Schultz RM, Kopf GS. 1995. Molecular ba-sis of mammalian egg activation. CurrTop Dev Biol 30:21–62.
Sethi JK, Empson RM, Galione A. 1996.Nicotinamide inhibits cyclic ADP-ribose-mediated calcium signalling in sea ur-chin eggs. Biochem J 319:613–617.
Shen SS, Buck WR. 1993. Sources of cal-cium in sea urchin eggs during the fer-tilization response. Dev Biol 157:157–169.
Smrcka AV, Sternweis PC. 1993. Regula-tion of purified subtypes of phosphatidy-linositol-specific phospholipase C beta byG protein alpha and beta gamma sub-units. J Biol Chem 268:9667–9674.
Sodergren E. 2006. The Sea Urchin Ge-nome Sequencing Consortium. The ge-nome of the sea urchin Strongylocentro-tus purpuratus. Science 314:941–952.
Sprang SR. 1997. G protein mechanisms:insights from structural analysis. AnnuRev Biochem 66:639–678.
Steinhardt RA, Epel D 1974. Activation ofsea-urchin eggs by a calcium ionophore.Proc Natl Acad Sci USA.71:1915–1919.
Steinhardt RA, Epel D, Carroll EJ Jr,Yanagimachi R. 1974. Is calcium iono-phore a universal activator for unfertil-ised eggs? Nature 252:41–43.
Stith BJ, Espinoza R, Roberts D, Smart T.1994. Sperm increase inositol 1,4,5-trisphosphate mass in Xenopus laeviseggs preinjected with calcium buffers orheparin. Dev Biol 165:206–215.
Stricker SA. 1999. Comparative biology ofcalcium signaling during fertilizationand egg activation in animals. Dev Biol211:157–176.
Swann KA. 1990. Cytosolic sperm factorstimulates repetitive calcium increasesand mimics fertilization in hamster eggs.Development 110:1295–1302.
Swann K, Ozil JP. 1994. Dynamics of thecalcium signal that triggers mammalianegg activation. Int Rev Cytol152:183–222.
Swann K, Saunders CM, Rogers NT, LaiFA. 2006. PLCzeta(zeta): a sperm pro-tein that triggers Ca2� oscillations andegg activation in mammals. Semin CellDev Biol 17:264–273.
Tahara M, Tasaka K, Masumoto N, Mam-moto A, Ikebuchi Y, Miyake A. 1996. Dy-namics of cortical granule exocytosis atfertilization in living mouse eggs. Am JPhysiol 270:C1354–1361.
Tao L, English AM. 2003. Mechanism ofS-nitrosation of recombinant humanbrain calbindin D28K. Biochemistry 42:3326–3334.
Tatone C, Delle Monache S, Iorio R, Case-rta D, Di Cola M, Colonna R. 2002. Pos-sible role for Ca(2�) calmodulin-depen-dent protein kinase II as an effector ofthe fertilization Ca(2�) signal in mouseoocyte activation. Mol Hum Reprod 8:750–757.
Thaler CD, Kuo RC, Patton C, Preston CM,Yagisawa H, Epel D. 2004. Phosphoino-sitide metabolism at fertilization of seaurchin eggs measured with a GFP-probe.Dev Growth Differ 46:413–423.
Thorn P, Fogarty KE, Parker I. 2004. Zy-mogen granule exocytosis is character-ized by long fusion pore openings andpreservation of vesicle lipid identity.Proc Natl Acad Sci USA 101:6774–6779.
Townley IK, Roux MM, Foltz KR. 2006.Signal transduction at fertilization: theCa2� release pathway in echinodermsand other invertebrate deuterostomes.Semin Cell Dev Biol. 17:293–302.
Turner PR, Jaffe LA, Fein A. 1986. Regu-lation of cortical vesicle exocytosis in sea
EGG ACTIVATION AT FERTILIZATION 2037

urchin eggs by inositol 1,4,5-trisphos-phate and GTP-binding protein. J CellBiol 102:70–76.
Voronina E, Wessel GM. 2004. betagammasubunits of heterotrimeric G-proteinscontribute to Ca2� release at fertiliza-tion in the sea urchin. J Cell Sci 117:5995–6005.
Wang S, Gebre-Medhin S, Betsholtz C,Stalberg P, Zhou Y, Larsson C, Weber G,Feinstein R, Oberg K, Gobl A, SkogseidB. 1998. Targeted disruption of themouse phospholipase C beta3 gene re-sults in early embryonic lethality. FEBSLett 441:261–265.
Weiss A. 1993. T cell antigen receptor sig-nal transduction: a tale of tails and cyto-plasmic protein-tyrosine kinases. Cell 73:209–212.
Whalley T, McDougall A, Crossley I,Swann K, Whitaker M. 1992. Internalcalcium release and activation of sea ur-chin eggs by cGMP are independent ofthe phosphoinositide signaling path-wayy. Mol Biol Cell 3:373–383.
Whitaker M. 2006. Calcium microdomainsand cell cycle control. Cell Calcium 40:585–592.
Whitaker MJ, Steinhardt RA. 1982. Ionicregulation of egg activation. Q Rev Bio-phys 15:593–666.
Whitaker MJ, Swann K. 1993. Lighting thefuse at fertilization. Development 117:1–12.
Williams CJ, Schultz RM, Kopf GS. 1992.Role of G proteins in mouse egg activa-tion: stimulatory effects of acetylcholineon the ZP2 to ZP2f conversion and pro-nuclear formation in eggs expressing afunctional m1 muscarinic receptor. DevBiol 151:288–296.
Willmott N, Sethi JK, Walseth TF, Lee HC,White AM, Galione A. 1996. Nitric oxideinduced mobilization of intracellular cal-cium via the cyclic ADP-ribose signallingpathway. J Biol Chem 271:3699–3705.
Whittingham DG. 1980. Parthenogenesisin mammals. Oxford Rev Reprod Biol 2:205–231.
Wong J, Wessel GM. 2006. Defending thezygote: Search for the ancestral animal
block to polyspermy. In: Schatten G, ed-itor. Current topics in developmental bi-ology, vol. 72. New York: AcademicPress. p 1–151.
Wu H, Smyth J, Luzzi V, Fukami K, Tak-enawa T, Black SL, Allbritton NL, Fis-sore RA. 2001. Sperm factor induces in-tracellular free calcium oscillations bystimulating the phosphoinositide path-way. Biol Reprod 64:1338–1349.
Xu L, Eu JP, Meissner G, Stamler JS.1998.Activation of the cardiac calcium releasechannel (ryanodine receptor) by poly-S-nitrosylation. Science 279:234–237.
Yanagida K. 2004. Complete fertilizationfailure in ICSI. Hum Cell 17:187-193.
Yanagimachi R. 1994. Fertility of mamma-lian spermatozoa: its development andrelativity. Zygote 2:371–372.
Yoda A, Oda S, Shikano T, Kouchi Z, AwajiT, Shirakawa H, Kinoshita K, Miyazaki S.2004. Ca2� oscillation-inducing phospho-lipase C zeta expressed in mouse eggs isaccumulated to the pronucleus during eggactivation. Dev Biol 268:245–257.
2038 PARRINGTON ET AL.





























