Embed Size (px)
Citation preview

FTY720 induces apoptosis of human hepatoma cell lines through PI3-K-mediatedAkt dephosphorylation
Terence K.Lee1, Kwan Man1,5, Joanna W.Ho1, ChrisK.Sun1, Kevin T.Ng1, Xiang Hong Wang2, Yong ChuanWong2, Irene O.Ng3, Ray Xu4 and Sheung Tat Fan1
Centre for the Study of Liver Disease and 1Department of Surgery,2Department of Anatomy, 3Department of Pathology and 4Institute ofMolecular Biology, University of Hong Kong, Pokfulam, Hong Kong, China
5To whom correspondence should be addressed at: Department of Surgery,University of Hong Kong Medical Centre, L9-55, Faculty of MedicineBuilding, 21 Sassoon Road, Hong Kong. Tel: þ852 2819 9646;Fax: þ852 2819 9634;Email: [email protected]
Our aim was to study the anticancer effect of the novelimmunomodulator FTY720 in vitro and in vivo by investiga-tion of cell cycle entry, cell cycle regulation, cell survivaland apoptosis pathways. Three hepatoma cell lines withdifferent p53 statuses (HepG2, Huh-7 and Hep3B) and onenon-tumorigenic immortalized liver cell line (MIHA) wereused for an in vitro study. The in vivo effects of FTY720wereevaluated in a nude mouse tumor model. Cell cycle distri-bution and cell cycle regulator proteins p27Kip1 and cyclinD1, together with the PI3-K/Akt pathway, mitogen-activated protein kinases and cleaved caspase-3 andcaspase-9, were evaluated. FTY720 selectively induced cellapoptosis in hepatoma cell lines with overexpression ofcleaved caspase-3 and caspase-9, but the same phenomenawere not found inMIHA cells. FTY720 inducedAkt dephos-phorylation at Ser473 mediated by phosphoinositide3-kinase (PI3-K) inhibition. Dephosphorylation led to down-regulation of p42/p44 and dephosphorylation of Forkheadtranscription factor and GSK-3b and, subsequently, up-regulation of p27Kip1 and down-regulation of cyclin D1.In our in vivo model FTY720 induced apoptosis of tumorcells by down-regulation of the Akt pathway. FTY720 sup-pressed tumor growth without notable side-effects in nor-mal liver. In conclusion, FTY720 is a novel anticancer agentthat induces apoptosis of hepatoma cell lines both in vitroand in vivo through PI3-K-mediated Akt dephosphoryla-tion in a p53-independent manner.
Introduction
Hepatocellular carcinoma (HCC), which is particularly preva-lent in Asia and Africa, is a major malignancy world wide andis one of the most common causes of cancer mortality (1).Although surgical resection is the first treatment of choice forHCC, most cases present at an advanced stage when surgerycan no longer cure the disease. Chemotherapy is commonly
applied for inoperable HCC, but its results are often discourag-ing because of chemoresistance conferred by p53 mutation andtoxicity to normal cells. Therefore, a novel pharmaceuticaltherapy needs to be developed. Recently, FTY720, a novelimmunomodulator, has been reported to have a strong anti-tumor effect for breast cancer (2), bladder cancer (3) andleukemia (4). So far, the feasibility of using this drug in HCCtreatment has not been studied. The precise cellular mechan-isms of FTY720 on cancer cells are not completely under-stood. Therefore, in this study we aimed to investigate thein vitro and in vivo anticancer potential of FTY720 and ascer-tain its exact mechanism of inducing apoptosis in HCC cells.Phosphoinositide 3-kinase (PI3-K), an upstream effector of
Ser/Thr kinase Akt (also known as protein kinase B), is themajor mediator of survival signals that protect cells fromapoptosis (5). PI3-K activity is opposed by the dual specificityphosphatase PTEN (also known as MMAC1 and TEP1), whichacts on position 3 of the inositol ring (6). Mutations ofPTEN have been demonstrated in many cancers and, in parti-cular, in HCC (7), suggesting that PI3-K and Akt are criticalfactors in the genesis of cancers (8). Therefore, PI3-K acting incellular signal transduction pathways plays a central role incancer cell survival.The immunomodulator FTY720, a synthetic structural ana-
log of myriocin, a metabolite in ascomycetes, was originallydemonstrated to induce apoptosis of infiltrated lymphocytes,especially T cells, and to prolong allograft survival after solidorgan transplantation (9,10). Recently, FTY720 was found tobe effective at inducing cancer cell apoptosis via caspase-3activation in the human prostate cancer cell line DU145 (11).FTY720 was also reported to induce apoptosis through mito-chondria by inhibition of bcl-2, independent of the Fas path-way (12,13). Furthermore, FTY720 induced G0/G1 arrest inHL60RG and Jurkat cells via dephosphorylation of retino-blastoma protein (14).The PI3-K and Akt pathway is very important in governing
cell survival and prevention of apoptosis in tumors. In this studywe have investigated the effect of FTY720 on the Akt cellsurvival pathway and found that Akt dephosphorylation plays acentral role in cell growth arrest and apoptosis upon administra-tion of FTY720 to hepatoma cell lines. Dephosphorylation ofAktSer473 but not AktThr308 might lead to dephosphorylation ofForkhead transcription factor (FKHR), GSK-3b and p42/p44,resulting in p53-independentG1 arrest, possibly by up-regulationof p27Kip1 and down-regulation of cyclin D1. It might alsoactivate the apoptotic pathway by up-regulation of caspase-3and caspase-9, both in vitro and in vivo.
Materials and methods
Cell lines and reagents
The human HCC cell line Huh-7 (15) (a gift from Dr H.Nakabayashi,Hokkaido University School of Medicine, Japan), human HCC cell lineHep3B (American Type Culture Collection, HB-8064), and the human
Abbreviations: DAPI, 4,6-diamindino-2-phenylindole; DMEM, Dulbecco’smodified Eagle’s medium; FKHR, Forkhead transcription factor; HCC,hepatocellular carcinoma; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; PBS, phosphate-buffered saline; PI(3,4,5)P3,phosphatidylinositol 3,4,5-trisphosphate; PI3-K, phosphoinositide 3-kinase.
Carcinogenesis vol.25 no.12 # Oxford University Press 2004; all rights reserved. 2397
Carcinogenesis vol.25 no.12 pp.2397--2405, 2004doi:10.1093/carcin/bgh250
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

hepatoblastoma cell line HepG2 (American Type Culture Collection, HB-8065) were maintained in Dulbecco’s modified Eagle’s medium (DMEM)with high glucose (Gibco BRL, Grand Island, NY) supplemented with 10%heat-inactivated fetal bovine serum (Gibco BRL), 50 U/ml penicillin G and50 mg/ml streptomycin (Gibco BRL) at 37�C in a humidified atmospherecontaining 5% CO2. The human non-tumorigenic, immortalized liver cellline MIHA (16) (kindly provided by Dr J.R.Chowdhury, Albert EinsteinCollege of Medicine, New York) was maintained in Chee’s medium(Gibco BRL) supplemented with 5% fetal bovine serum, 100 U/ml penicillin,100 mg/ml streptomycin, 2 mmol/l L-glutamine, 50 mmol/l dexamethasoneand 20 mU/ml recombinant insulin (Boehringer Mannheim, Indianapolis, IN).FTY720 was kindly provided as a powder by Novartis Pharmaceuticals Ltd(Basel, Switzerland). It was added to the culture medium at different concen-trations after dissolution in normal saline. The constitutively active PI3-Kplasmid (CD2-p110) was a gift from Dr D.Cantrell (University of Dundee,Dundee, UK).
3-(4,5-Cimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay
The hepatoma cell lines and non-tumorigenic, immortalized MIHA cells wereseeded on 96-well plates and appropriate concentrations of FTY720 ranging from0.078 to 200 mM were then added. After 4--24 h 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) dye, at a concentration of 5 mg/ml(Sigma-Aldrich, St Louis,MO), was added and the plates were incubated for 12 hin a moist chamber at 37�C. Optical density was determined by eluting the dyewith dimethyl sulfoxide (Sigma-Aldrich) and the absorbance was measured at570 nm. At least three independent experiments were performed.
Cell cycle analysis
After FTY720 treatment, the DNA content and cell cycle distribution ofhepatoma cells grown in 6-well plates were determined by flow cytometry.The cells were plated at a low density (5 � 104 cells/well) and were harvestedat 0, 4, 8, 16 and 24 h. Another set of controls was employed without FTY720treatment. They were trypsinized and washed once in phosphate-bufferedsaline (PBS). They were then fixed in cold 70% ethanol and stored at 4�C.Before testing, the ethanol was removed and the cells were resuspended inPBS. The fixed cells were then washed with PBS and treated with RNase(1 mg/ml) and stained with propidium iodide (50 mg/ml) for 30 min at 37�C.Cell cycle analysis was performed in an EPICS profile analyzer, using ModFitLT2.0 software (Coulter Electronics, Hialeah, FL).
Detection of apoptotic cells by 4,6-diamidino-2-phenylindole (DAPI) stainingand TUNEL assay
Hep3B cells were plated onto 12 mm coverslips in DMEM medium at ~70%confluence for 24 h. The cells were then treated with FTY720 at the IC10 andIC50 doses for 0, 4, 8, 16 and 24 h. They were fixed in ice-cold acetone andmethanol (1:1), washed with PBS and then stained with DAPI for 5 min. Thestained cells were examined under a fluorescence microscope and cells wereconsidered to undergo apoptosis based on the appearance of nuclear fragmenta-tion. A total of 500 cells were counted in 5 fields/sample. The percentage ofapoptotic cells was calculated as (number of apoptotic cells � total number ofcells counted) � 100. For the TUNEL assay the procedure was the same as forDAPI staining except that the cells were stained in situ with a POD cell deathdetection kit (Boehringer Manheim, Manheim, Germany) and counterstainedwith propidium iodide.
Assay for PI3-K activity
Cultured Hep3B cells were treated with FTY720 at the IC10 and IC50 doses for4 h and then stimulated with 100 nM insulin for 10 min at 37�C. The cells werethen lysed in 1 ml of NP-40 containing lysis buffer with protease inhibitors.Equal amounts of protein lysate (Protein Assay Kit, Bio-Rad, Hercules, CA)were incubated with 5 ml of anti-PI3-kinase antibody (Upstate Biotechnology,Lake Placid, NY) and bound with protein A--agarose beads for 1 h at 4�C.Immunoprecipitates were washed and evaluated for PI3-K activity by compe-titive ELISA (Echelon Biosciences, Salt Lake City, UT) as described (17).Briefly, PI3-K bound to the beads was incubated for 2 h with 10 mMdiacylglycerol phosphatidylinositol 4,5-diphosphate (diC8PI(4,5)P2) substrateat room temperature in 50 ml of buffer containing 5 mMHEPES (pH 7), 25 mMATP and 2.5 mM MgCl2. The beads removed by centrifugation and thesupernatant, or known concentrations of phosphatidylinositol 3,4,5-trispho-sphate [PI(3,4,5)P3], were incubated for 1 h with 50 ml of PI(3,4,5)P3-bindingreagent and then transferred to a detection plate coated with PI(3,4,5)P3. Plate-bound binding reagent was quantitated using a secondary detection reagent,peroxidase, and peroxidase substrate, with the reaction product measured byabsorbance at 450 nm.
In vivo effect of FTY720 in the nude mouse tumor model
We studied the in vivo effect of FTY720 on the cell survival and apoptosispathway using a nude mouse tumor model. The mice were housed in
microisolator cages under positive air pressure and maintained at a constanttemperature (22�C) and humidity. Surgical operations and drug administrationwere performed in a laminar flow cabinet. Four-week-old male BALB/c nu/numice were used to establish the tumor model by s.c. implantation of 1--2 mm3
tumor tissue originating from the Hep3B strain. FTY720 was given at10 mg/kg by i.p. injection when the tumor size reached 32 mm3 (L � W2/2)at 10 mg/kg for 10 or 20 days. Tumor tissue and liver were sampled fordetection of protein expression and morphological examination. Tumor growthwas also compared between the rats with and without FTY720 treatment.
Western blotting
The cells were lysed and protein extraction was performed. The samples wereseparated in a 10% SDS--acrylamide gel and electrophoretically transferred toPVDF membrane (Amersham, Little Chalfont, UK). The membrane wasblotted with 10% non-fat milk, washed and then probed with PI3-K (1:1000)(Upstate Biotechnology), p53 (1:1000) (Dako, Carpinteria, CA), p21WAF1(1:500) (Dako), p27Kip1 (1:2000) (Transduction Laboratories, Lexington, KY),phospho-p42/p44 and p42/p44 (Cell Signaling Technology, Beverly, MA),phospho-Akt and Apoptosis Pathway Sampler Kits (Cell Signaling Technol-ogy) and actin (Santa Cruz Biotechnology, Santa Cruz, CA). After washing,the membrane was then incubated with horseradish peroxidase-conjugatedrabbit anti-mouse antibody (Amersham) and then visualized by enhancedchemiluminescense plus according to the manufacturer’s protocol.
Morphological examination
TUNEL staining was performed in situ using a POD cell death detection kit(Boehringer Manheim) following the manufacturer’s protocol. Apoptotic cellswere determined with careful observation of TUNEL-positive nuclei. Forhematoxylin and eosin (H&E) staining the samples were stained with H&E.The images were captured by an image analysis system (Eclipse E600; Nikon,Japan).
Results
FTY720 selectively induces significant hepatoma cell deathin vitro
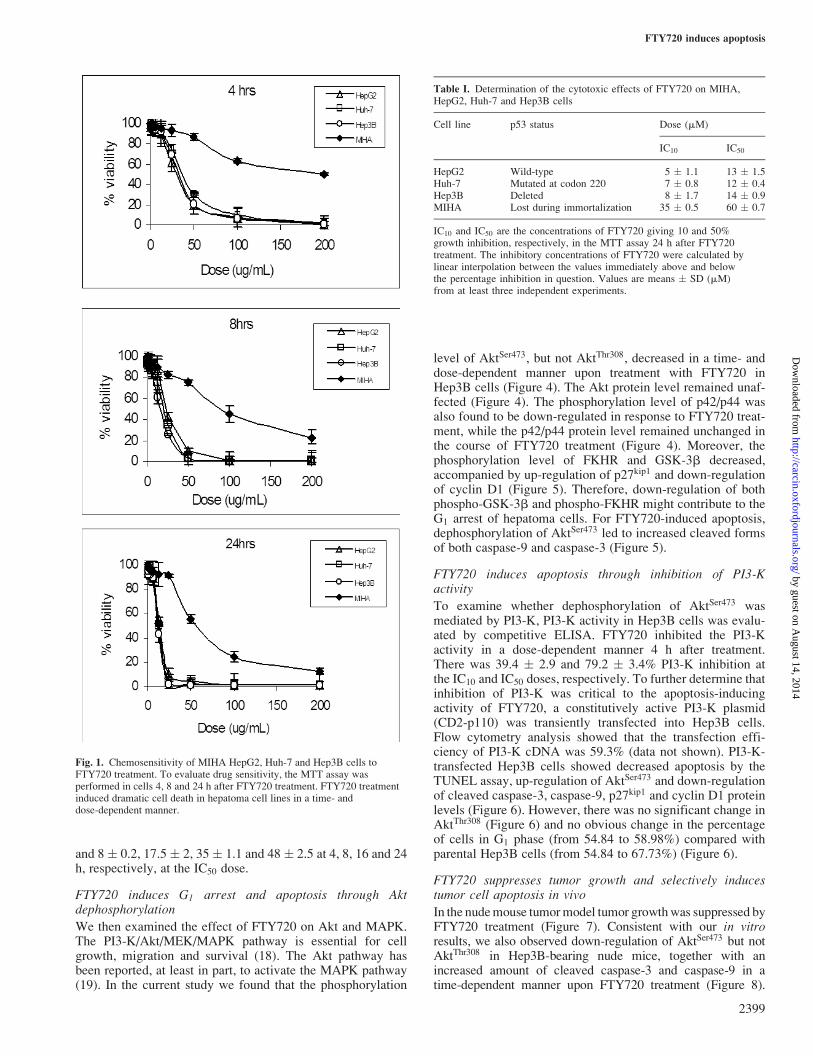
To determine the cytotoxic effect of FTY720 on hepatoma celllines, the MTT assay was performed to determine the IC10 andIC50 doses. The MTT assay demonstrated that FTY720 treat-ment induced dramatic cell death in hepatoma cell lines in atime- and dose-dependent manner (Figure 1). As shown inTable I, there was no significant difference in drug sensitivity(IC50 range 12--14 mM) among the three hepatoma cell lines(HepG2, Huh-7 and Hep3B) with different p53 statuses. Toexamine its toxicity, the effect of FTY720 on MIHA cells wasalso investigated. The results showed that FTY720 conferredgreater chemoresistance to MIHA, with an IC50 up to 60 mM(Table I).
FTY720 induces p53-independent G1 arrest in hepatoma celllines
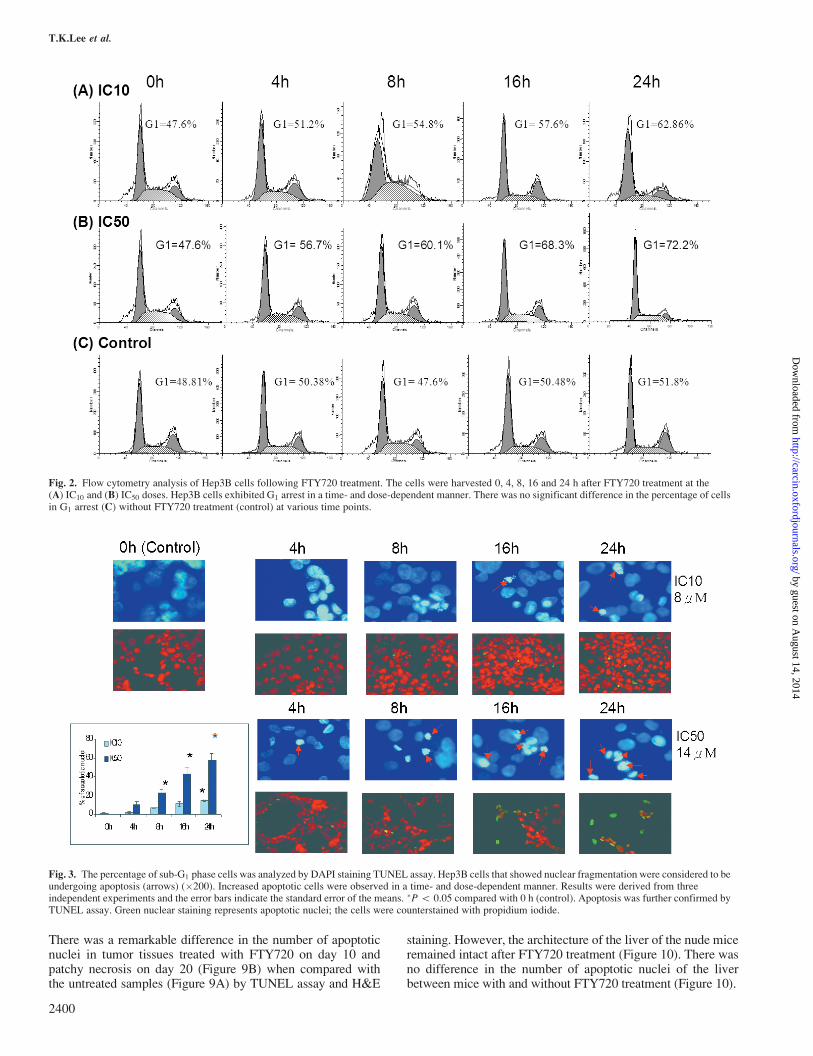
FTY720 was previously reported to induce G1 arrest in pro-state and breast cancers (2,11). To evaluate its possible effectson the cell cycle of cells with different p53 statuses, cell cyclechanges were analyzed by flow cytometry in HepG2, Huh-7and Hep3B cells. The effect of FTY720 on G1 arrest wasexamined by excluding cells of small size in the sub-G1
phase. When FTY720 was administered at the IC10 and IC50
doses, all three hepatoma cell lines showed increased G1 arrestin a dose- and time-dependent manner. The cell cycle distribu-tion of Hep3B cells upon FTY720 treatment is shown inFigure 2. The percentage of sub-G1 phase cells was analyzedby DAPI staining and TUNEL assay. The DAPI result showedthat the number of apoptotic nuclei increased significantlyafter FTY720 administration compared with the control start-ing at 8 h at the IC50 dose and at 24 h at the IC10 dose(Figure 3). The percentages of apoptotic nuclei as determinedby the TUNEL assay were 1.2 � 1.0, 4.3 � 0.3, 7.9 � 0.9 and10.5 � 1.5 at 4, 8, 16 and 24 h, respectively, at the IC10 dose
T.K.Lee et al.
2398
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
and 8� 0.2, 17.5� 2, 35� 1.1 and 48 � 2.5 at 4, 8, 16 and 24h, respectively, at the IC50 dose.
FTY720 induces G1 arrest and apoptosis through Aktdephosphorylation
We then examined the effect of FTY720 on Akt and MAPK.The PI3-K/Akt/MEK/MAPK pathway is essential for cellgrowth, migration and survival (18). The Akt pathway hasbeen reported, at least in part, to activate the MAPK pathway(19). In the current study we found that the phosphorylation
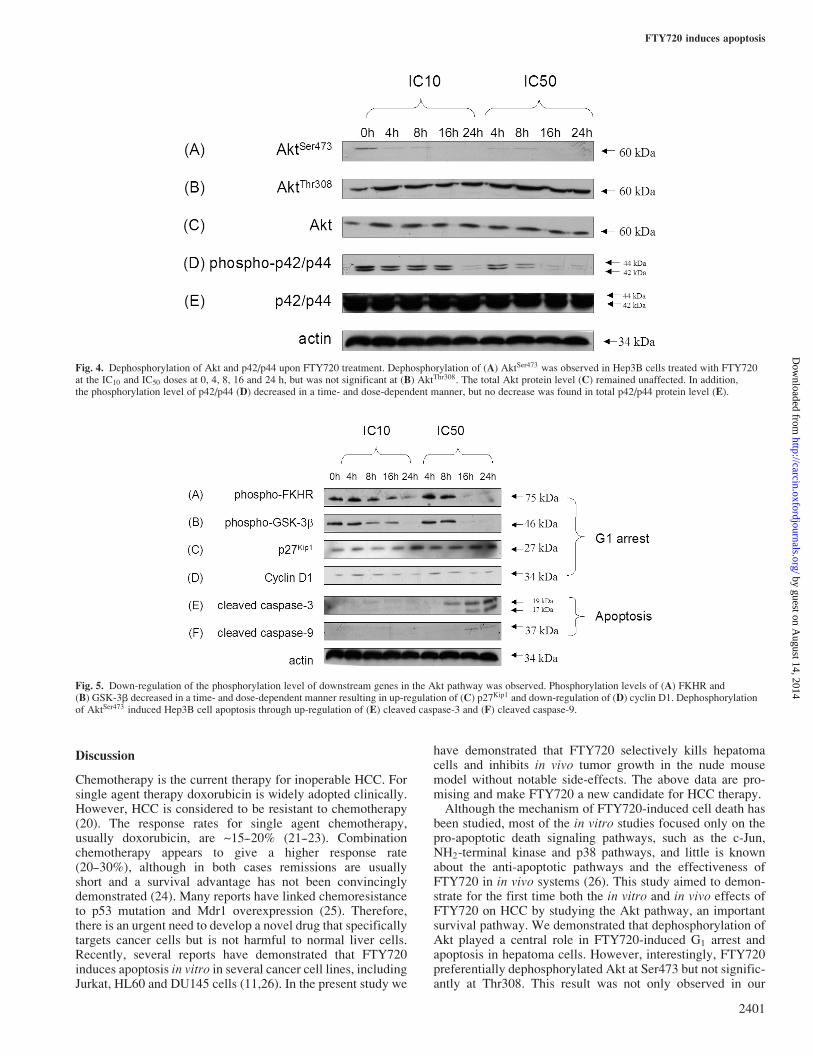
level of AktSer473, but not AktThr308, decreased in a time- anddose-dependent manner upon treatment with FTY720 inHep3B cells (Figure 4). The Akt protein level remained unaf-fected (Figure 4). The phosphorylation level of p42/p44 wasalso found to be down-regulated in response to FTY720 treat-ment, while the p42/p44 protein level remained unchanged inthe course of FTY720 treatment (Figure 4). Moreover, thephosphorylation level of FKHR and GSK-3b decreased,accompanied by up-regulation of p27kip1 and down-regulationof cyclin D1 (Figure 5). Therefore, down-regulation of bothphospho-GSK-3b and phospho-FKHR might contribute to theG1 arrest of hepatoma cells. For FTY720-induced apoptosis,dephosphorylation of AktSer473 led to increased cleaved formsof both caspase-9 and caspase-3 (Figure 5).
FTY720 induces apoptosis through inhibition of PI3-Kactivity
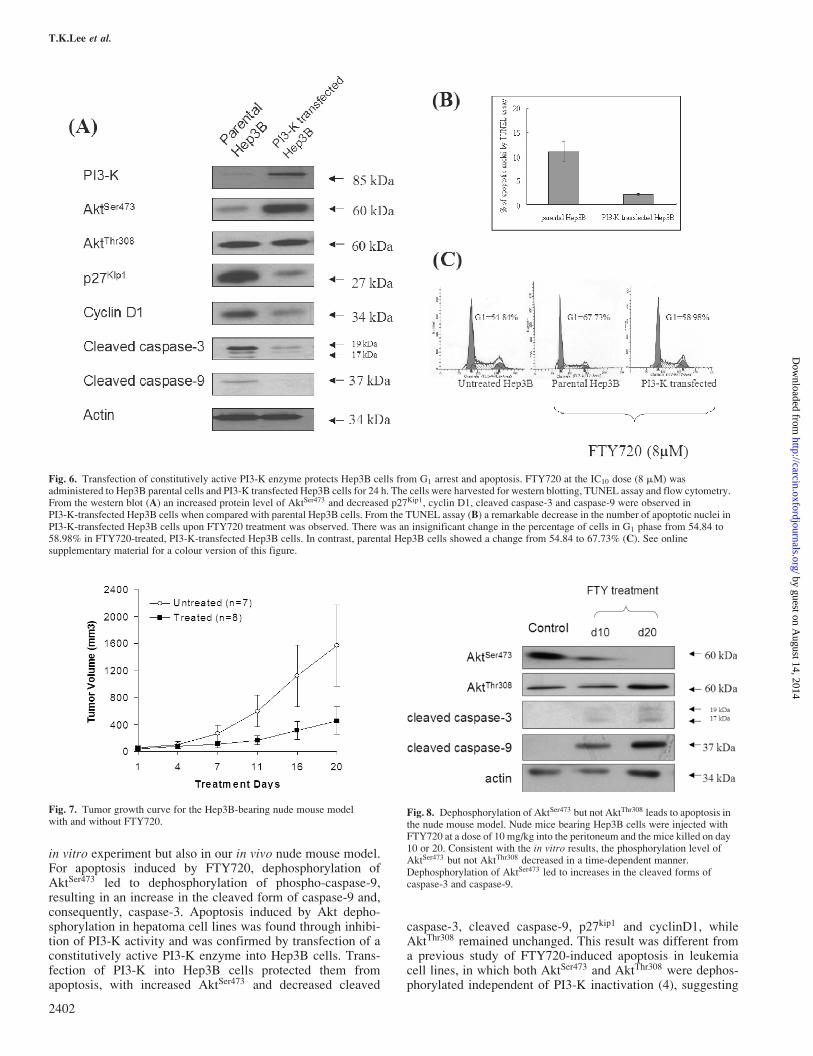
To examine whether dephosphorylation of AktSer473 wasmediated by PI3-K, PI3-K activity in Hep3B cells was evalu-ated by competitive ELISA. FTY720 inhibited the PI3-Kactivity in a dose-dependent manner 4 h after treatment.There was 39.4 � 2.9 and 79.2 � 3.4% PI3-K inhibition atthe IC10 and IC50 doses, respectively. To further determine thatinhibition of PI3-K was critical to the apoptosis-inducingactivity of FTY720, a constitutively active PI3-K plasmid(CD2-p110) was transiently transfected into Hep3B cells.Flow cytometry analysis showed that the transfection effi-ciency of PI3-K cDNA was 59.3% (data not shown). PI3-K-transfected Hep3B cells showed decreased apoptosis by theTUNEL assay, up-regulation of AktSer473 and down-regulationof cleaved caspase-3, caspase-9, p27kip1 and cyclin D1 proteinlevels (Figure 6). However, there was no significant change inAktThr308 (Figure 6) and no obvious change in the percentageof cells in G1 phase (from 54.84 to 58.98%) compared withparental Hep3B cells (from 54.84 to 67.73%) (Figure 6).
FTY720 suppresses tumor growth and selectively inducestumor cell apoptosis in vivo
In the nudemouse tumormodel tumor growthwas suppressed byFTY720 treatment (Figure 7). Consistent with our in vitroresults, we also observed down-regulation of AktSer473 but notAktThr308 in Hep3B-bearing nude mice, together with anincreased amount of cleaved caspase-3 and caspase-9 in atime-dependent manner upon FTY720 treatment (Figure 8).
Fig. 1. Chemosensitivity of MIHA HepG2, Huh-7 and Hep3B cells toFTY720 treatment. To evaluate drug sensitivity, the MTT assay wasperformed in cells 4, 8 and 24 h after FTY720 treatment. FTY720 treatmentinduced dramatic cell death in hepatoma cell lines in a time- anddose-dependent manner.
Table I. Determination of the cytotoxic effects of FTY720 on MIHA,HepG2, Huh-7 and Hep3B cells
Cell line p53 status Dose (mM)
IC10 IC50
HepG2 Wild-type 5 � 1.1 13 � 1.5Huh-7 Mutated at codon 220 7 � 0.8 12 � 0.4Hep3B Deleted 8 � 1.7 14 � 0.9MIHA Lost during immortalization 35 � 0.5 60 � 0.7
IC10 and IC50 are the concentrations of FTY720 giving 10 and 50%growth inhibition, respectively, in the MTT assay 24 h after FTY720treatment. The inhibitory concentrations of FTY720 were calculated bylinear interpolation between the values immediately above and belowthe percentage inhibition in question. Values are means � SD (mM)from at least three independent experiments.
FTY720 induces apoptosis
2399
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
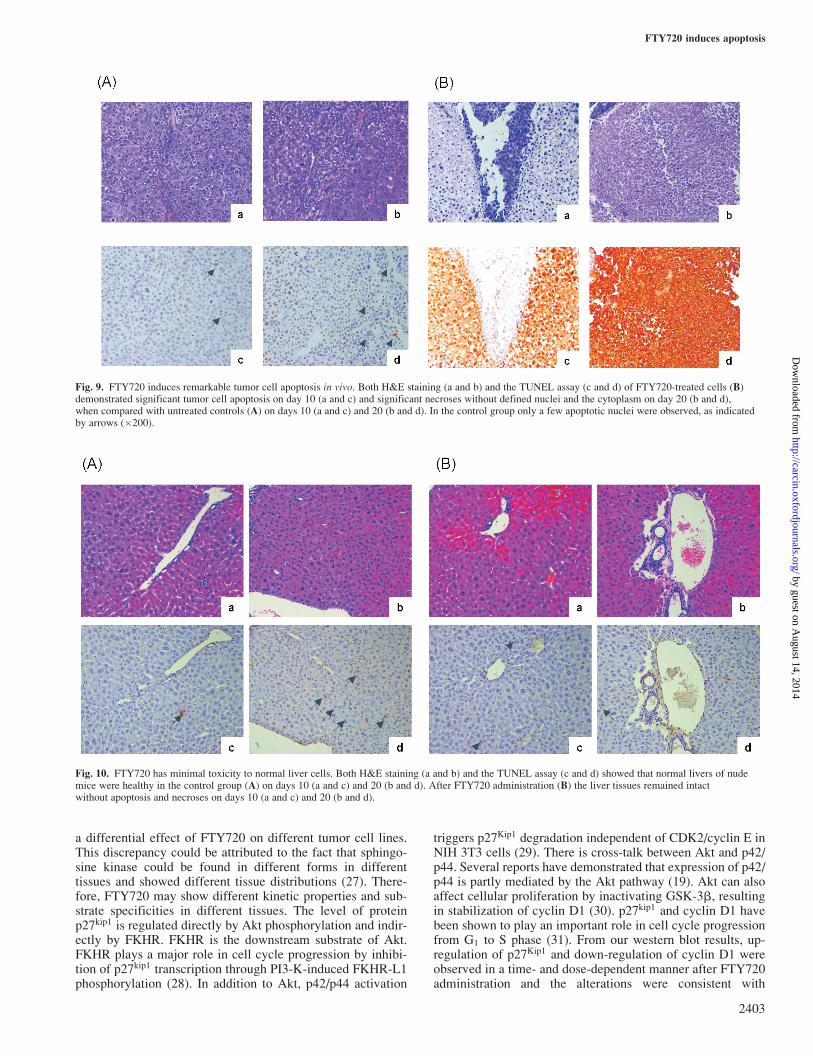
There was a remarkable difference in the number of apoptoticnuclei in tumor tissues treated with FTY720 on day 10 andpatchy necrosis on day 20 (Figure 9B) when compared withthe untreated samples (Figure 9A) by TUNEL assay and H&E
staining. However, the architecture of the liver of the nude miceremained intact after FTY720 treatment (Figure 10). There wasno difference in the number of apoptotic nuclei of the liverbetween mice with and without FTY720 treatment (Figure 10).
Fig. 2. Flow cytometry analysis of Hep3B cells following FTY720 treatment. The cells were harvested 0, 4, 8, 16 and 24 h after FTY720 treatment at the(A) IC10 and (B) IC50 doses. Hep3B cells exhibited G1 arrest in a time- and dose-dependent manner. There was no significant difference in the percentage of cellsin G1 arrest (C) without FTY720 treatment (control) at various time points.
Fig. 3. The percentage of sub-G1 phase cells was analyzed by DAPI staining TUNEL assay. Hep3B cells that showed nuclear fragmentation were considered to beundergoing apoptosis (arrows) (�200). Increased apoptotic cells were observed in a time- and dose-dependent manner. Results were derived from threeindependent experiments and the error bars indicate the standard error of the means. �P5 0.05 compared with 0 h (control). Apoptosis was further confirmed byTUNEL assay. Green nuclear staining represents apoptotic nuclei; the cells were counterstained with propidium iodide.
T.K.Lee et al.
2400
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
Discussion
Chemotherapy is the current therapy for inoperable HCC. Forsingle agent therapy doxorubicin is widely adopted clinically.However, HCC is considered to be resistant to chemotherapy(20). The response rates for single agent chemotherapy,usually doxorubicin, are ~15--20% (21--23). Combinationchemotherapy appears to give a higher response rate(20--30%), although in both cases remissions are usuallyshort and a survival advantage has not been convincinglydemonstrated (24). Many reports have linked chemoresistanceto p53 mutation and Mdr1 overexpression (25). Therefore,there is an urgent need to develop a novel drug that specificallytargets cancer cells but is not harmful to normal liver cells.Recently, several reports have demonstrated that FTY720induces apoptosis in vitro in several cancer cell lines, includingJurkat, HL60 and DU145 cells (11,26). In the present study we
have demonstrated that FTY720 selectively kills hepatomacells and inhibits in vivo tumor growth in the nude mousemodel without notable side-effects. The above data are pro-mising and make FTY720 a new candidate for HCC therapy.Although the mechanism of FTY720-induced cell death has
been studied, most of the in vitro studies focused only on thepro-apoptotic death signaling pathways, such as the c-Jun,NH2-terminal kinase and p38 pathways, and little is knownabout the anti-apoptotic pathways and the effectiveness ofFTY720 in in vivo systems (26). This study aimed to demon-strate for the first time both the in vitro and in vivo effects ofFTY720 on HCC by studying the Akt pathway, an importantsurvival pathway. We demonstrated that dephosphorylation ofAkt played a central role in FTY720-induced G1 arrest andapoptosis in hepatoma cells. However, interestingly, FTY720preferentially dephosphorylated Akt at Ser473 but not signific-antly at Thr308. This result was not only observed in our
Fig. 4. Dephosphorylation of Akt and p42/p44 upon FTY720 treatment. Dephosphorylation of (A) AktSer473 was observed in Hep3B cells treated with FTY720at the IC10 and IC50 doses at 0, 4, 8, 16 and 24 h, but was not significant at (B) AktThr308. The total Akt protein level (C) remained unaffected. In addition,the phosphorylation level of p42/p44 (D) decreased in a time- and dose-dependent manner, but no decrease was found in total p42/p44 protein level (E).
Fig. 5. Down-regulation of the phosphorylation level of downstream genes in the Akt pathway was observed. Phosphorylation levels of (A) FKHR and(B) GSK-3b decreased in a time- and dose-dependent manner resulting in up-regulation of (C) p27Kip1 and down-regulation of (D) cyclin D1. Dephosphorylationof AktSer473 induced Hep3B cell apoptosis through up-regulation of (E) cleaved caspase-3 and (F) cleaved caspase-9.
FTY720 induces apoptosis
2401
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
in vitro experiment but also in our in vivo nude mouse model.For apoptosis induced by FTY720, dephosphorylation ofAktSer473 led to dephosphorylation of phospho-caspase-9,resulting in an increase in the cleaved form of caspase-9 and,consequently, caspase-3. Apoptosis induced by Akt depho-sphorylation in hepatoma cell lines was found through inhibi-tion of PI3-K activity and was confirmed by transfection of aconstitutively active PI3-K enzyme into Hep3B cells. Trans-fection of PI3-K into Hep3B cells protected them fromapoptosis, with increased AktSer473 and decreased cleaved
caspase-3, cleaved caspase-9, p27kip1 and cyclinD1, whileAktThr308 remained unchanged. This result was different froma previous study of FTY720-induced apoptosis in leukemiacell lines, in which both AktSer473 and AktThr308 were dephos-phorylated independent of PI3-K inactivation (4), suggesting
Fig. 6. Transfection of constitutively active PI3-K enzyme protects Hep3B cells from G1 arrest and apoptosis. FTY720 at the IC10 dose (8 mM) wasadministered to Hep3B parental cells and PI3-K transfected Hep3B cells for 24 h. The cells were harvested for western blotting, TUNEL assay and flow cytometry.From the western blot (A) an increased protein level of AktSer473 and decreased p27Kip1, cyclin D1, cleaved caspase-3 and caspase-9 were observed inPI3-K-transfected Hep3B cells when compared with parental Hep3B cells. From the TUNEL assay (B) a remarkable decrease in the number of apoptotic nuclei inPI3-K-transfected Hep3B cells upon FTY720 treatment was observed. There was an insignificant change in the percentage of cells in G1 phase from 54.84 to58.98% in FTY720-treated, PI3-K-transfected Hep3B cells. In contrast, parental Hep3B cells showed a change from 54.84 to 67.73% (C). See onlinesupplementary material for a colour version of this figure.
Fig. 7. Tumor growth curve for the Hep3B-bearing nude mouse modelwith and without FTY720.
Fig. 8. Dephosphorylation of AktSer473 but not AktThr308 leads to apoptosis inthe nude mouse model. Nude mice bearing Hep3B cells were injected withFTY720 at a dose of 10 mg/kg into the peritoneum and the mice killed on day10 or 20. Consistent with the in vitro results, the phosphorylation level ofAktSer473 but not AktThr308 decreased in a time-dependent manner.Dephosphorylation of AktSer473 led to increases in the cleaved forms ofcaspase-3 and caspase-9.
T.K.Lee et al.
2402
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
a differential effect of FTY720 on different tumor cell lines.This discrepancy could be attributed to the fact that sphingo-sine kinase could be found in different forms in differenttissues and showed different tissue distributions (27). There-fore, FTY720 may show different kinetic properties and sub-strate specificities in different tissues. The level of proteinp27kip1 is regulated directly by Akt phosphorylation and indir-ectly by FKHR. FKHR is the downstream substrate of Akt.FKHR plays a major role in cell cycle progression by inhibi-tion of p27kip1 transcription through PI3-K-induced FKHR-L1phosphorylation (28). In addition to Akt, p42/p44 activation
triggers p27Kip1 degradation independent of CDK2/cyclin E inNIH 3T3 cells (29). There is cross-talk between Akt and p42/p44. Several reports have demonstrated that expression of p42/p44 is partly mediated by the Akt pathway (19). Akt can alsoaffect cellular proliferation by inactivating GSK-3b, resultingin stabilization of cyclin D1 (30). p27kip1 and cyclin D1 havebeen shown to play an important role in cell cycle progressionfrom G1 to S phase (31). From our western blot results, up-regulation of p27Kip1 and down-regulation of cyclin D1 wereobserved in a time- and dose-dependent manner after FTY720administration and the alterations were consistent with
Fig. 9. FTY720 induces remarkable tumor cell apoptosis in vivo. Both H&E staining (a and b) and the TUNEL assay (c and d) of FTY720-treated cells (B)demonstrated significant tumor cell apoptosis on day 10 (a and c) and significant necroses without defined nuclei and the cytoplasm on day 20 (b and d),when compared with untreated controls (A) on days 10 (a and c) and 20 (b and d). In the control group only a few apoptotic nuclei were observed, as indicatedby arrows (�200).
Fig. 10. FTY720 has minimal toxicity to normal liver cells. Both H&E staining (a and b) and the TUNEL assay (c and d) showed that normal livers of nudemice were healthy in the control group (A) on days 10 (a and c) and 20 (b and d). After FTY720 administration (B) the liver tissues remained intactwithout apoptosis and necroses on days 10 (a and c) and 20 (b and d).
FTY720 induces apoptosis
2403
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

induction of G1 arrest. Therefore, up-regulation of p27Kip1 and
down-regulation of cyclin D1 might be correlated withFTY720-induced G1 arrest. This conclusion was further con-firmed by transfection of constitutively active PI3-K enzymeinto Hep3B cells. There was no apparent change in p27kip1 andcyclin D1 protein levels in PI3-K-transfected Hep3B cells,with only an insignificant change in the percentage in G1
phase from 54.84 to 58.98%, in contrast to parental Hep3Bcells, which changed from 54.84 to 67.73%, thus suggestingthe role of p27kip1 and cyclin D1 in FTY720-induced G1 arrest.Therefore, we propose that FTY720-induced apoptosis ismediated by PI3-K Akt dephosphorylation (Figure 11).The main hurdle to chemotherapy for HCC is chemo-
resistance and toxicity to normal liver cells (32). In this studywe have found that FTY720 induces hepatoma cell death in ap53-independent manner. This result is an advantage in pre-venting chemoresistance induced by p53 mutation in HCC(33). FTY720 selectively induces hepatoma cell death butnot that of immortalized, non-tumorigenic liver cell lines.Consistent with the in vitro study, we found that FTY720selectively induces apoptosis in tumors in a time- and dose-dependent manner, but not in normal liver in the nude mousetumor model.In conclusion, FTY720 is a novel anticancer agent that
induces apoptosis of hepatoma cell lines in a p53-independentmanner through PI3-K-mediated Akt dephosphorylation.FTY720 may be a promising agent for the treatment of HCCpatients.
Supplementary material
Supplementary material is available online at http://www.carcin.oupjournals.org/.
Acknowledgements
This work was supported by a Distinguished Research Achievement Awardand the Sun C.Y. Research Foundation for Hepatobiliary and PancreaticSurgery of the University of Hong Kong.
References
1.Ching,Y.P., Wong,C.M., Chan,S.F., Leung,T.H., Ng,D.C., Jin,D.Y. andNg,I.O. (2003) Deleted in liver cancer (DLC) 2 encodes a RhoGAP proteinwith growth suppressor function and is underexpressed in hepatocellularcarcinoma. J. Biol. Chem., 278, 10824--10830.
2.Azuma,H., Takahara,S., Ichimaru,N. et al. (2002) Marked preventionof tumor growth and metastasis by a novel immunosuppressiveagent, FTY720, in mouse breast cancer models. Cancer Res., 62,1410--1419.
3.Azuma,H., Takahara,S., Horie,S., Muto,S., Otsuki,Y. and Katsuoka,Y.(2003) Induction of apoptosis in human bladder cancer cells in vitro andin vivo caused by FTY720 treatment. J. Urol., 169, 2372--2377.
4.Matsuoka,Y., Nagahara,Y., Ikekita,M. and Shinomiya,T. (2003) A novelimmunosuppressive agent FTY720 induced Akt dephosphorylation inleukemia cells. Br. J. Pharmacol., 138, 1303--1312.
5.Toker,A. (2000) Protein kinases as mediators of phosphoinositide 3-kinasesignaling. Mol. Pharmacol., 57, 652--658.
6.Wen,J., Ramadevi,N., Nguyen,D., Perkins,C., Worthington,E. andBhalla,K. (2000) Antileukemic drugs increase death receptor 5 levelsand enhance Apo-2L-induced apoptosis of human acute leukemia cells.Blood, 96, 3900--3906.
7.Yao,Y.J., Ping,X.L., Zhang,H. et al. (1999) PTEN/MMAC1 mutations inhepatocellular carcinomas. Oncogene, 18, 3181--3185.
8.Simpson,L. and Parsons,R. (2001) PTEN: life as a tumor suppressor. Exp.Cell Res., 264, 29--41.
9.Azuma,H., Horie,S., Muto,S. et al. (2003) Selective cancer cell apoptosisinduced by FTY720; evidence for a Bcl-dependent pathway andimpairment in ERK activity. Anticancer Res., 23, 3183--3193.
10.Kimura,T., Hasegawa,T., Nakai,H., Azuma,T., Usui,N., Sasaki,T. andOkada,A. (2003) FTY720 reduces T-cell recruitment into murine intestinalallograft and prevents activation of graft-infiltrating cells.Transplantation, 75, 1469--1474.
11.Permpongkosol,S., Wang,J.D., Takahara,S., Matsumiya,K., Nonomura,N.,Nishimura,K., Tsujimura,A., Kongkanand,A. and Okuyama,A. (2002)Anticarcinogenic effect of FTY720 in human prostate carcinoma DU145cells: modulation of mitogenic signaling, FAK, cell-cycle entry andapoptosis. Int. J. Cancer, 98, 167--172.
12.Suzuki,S., Enosawa,S., Kakefuda,T., Shinomiya,T., Amari,M., Naoe,S.,Hoshino,Y. and Chiba,K. (1996) A novel immunosuppressant,FTY720, with a unique mechanism of action, induces long-term graftacceptance in rat and dog allotransplantation. Transplantation, 61,200--205.
13.Suzuki,S., Li,X.K., Enosawa,S. and Shinomiya,T. (1996) A newimmunosuppressant, FTY720, induces bcl-2-associated apoptotic celldeath in human lymphocytes. Immunology, 89, 518--523.
14.Nagahara,Y., Matsuoka,Y., Saito,K., Ikekita,M. and Higuchi,S. (2001)Coordinate involvement of cell cycle arrest and apoptosis strengthen theeffect of FTY720. Jpn. J. Cancer Res., 92, 680--687.
15.Nakabayashi,H., Taketa,K., Miyano,K., Yamane,T. and Sato,J. (1982)Growth of human hepatoma cell lines with differentiated functions inchemically defined medium. Cancer Res., 42, 3858--3863.
16.Brown,J.J., Parashar,B., Moshage,H., Tanaka,K.E., Engelhardt,D.,Rabbani,E., Roy-Chowdhury,N. and Roy-Chowdhury,J. (2000) A long-term hepatitis B viremia model generated by transplanting nontumorigenicimmortalized human hepatocytes in Rag-2-deficient mice. Hepatology, 31,173--181.
17.Oganesian,A., Poot,M., Daum,G., Coats,S.A., Wright,M.B., Seifert,R.A.and Bowen-Pope,D.F. (2003) Protein tyrosine phosphatase RQ is aphosphatidylinositol phosphatase that can regulate cell survival andproliferation. Proc. Natl Acad. Sci. USA, 13, 7563--7568.
18.Wu,H., Goel,V. and Haluska,F.G. (2003) PTEN signaling pathways inmelanoma. Oncogene, 22, 3113--3122.
19.Chien,M.W., Chien,C.S., Hsiao,L.D., Lin,C.H. and Yang,C.M. (2003)OxLDL induces mitogen-activated protein kinase activation mediated viaPI3-kinase/Akt in vascular smooth muscle cells. J. Lipid Res., 44,1667--1675.
20.Bruix,J., Sherman,M., Llovet,J.M., Beaugrand,M., Lencioni,R.,Burroughs,A.K., Christensen,E., Pagliaro,L., Colombo,M., Rodes,J.;EASL Panel of Experts on HCC (2001) Clinical management ofhepatocellular carcinoma. Conclusions of the Barcelona-2000 EASLconference. European Association for the Study of the Liver. J. Hepatol.,35, 421--430.
21. Johnson,P.J., Williams,R., Thomas,H., Sherlock,S. and Murray-Lyon,J.M.(1978) Induction of remission in hepatocellular carcinoma withdoxorubicin. Lancet, 1, 1006--1009.
Fig. 11. The possible mechanism of FTY720 induced human hepatomacell line apoptosis is through PI3-K-mediated Akt dephosphorylation.
T.K.Lee et al.
2404
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

22.Lai,C.L., Wu,P.C., Chan,G.C., Lok,A.S. and Lin,H.J. (1988) Doxorubicinversus no antitumor therapy in inoperable hepatocellular carcinoma. Aprospective randomized trial. Cancer, 62, 479--483.
23.Nerenstone,S.R., Ihde,D.C. and Friedman,M.A. (1988) Clinical trials inprimary hepatocellular carcinoma: current status and future directions.Cancer Treat. Rev., 15, 1--31.
24.Leung,T.W. and Johnson,P.J. (2001) Systemic therapy for hepatocellularcarcinoma. Semin. Oncol., 28, 514--520.
25.Bush,J.A. and Li,G. (2002) Cancer chemoresistance: the relationshipbetween p53 and multidrug transporters. Int. J. Cancer, 98, 323--330.
26.Matsuda,S., Minowa,A., Suzuki,S. and Koyasu,S. (1999) Differentialactivation of c-Jun NH2-terminal kinase and p38 pathways duringFTY720-induced apoptosis of T lymphocytes that is suppressed by theextracellular signal-regulated kinase pathway. J. Immunol., 162,3321--3326.
27.Meacci,E., Cencetti,F., Donati,C., Nuti,F., Becciolini,L. and Bruni,P.(2004) Sphingosine kinase activity is required for sphingosine-mediatedphospholipase D activation in C2C12 myoblasts. Biochem. J., 381,655--663.
28.Dijkers,P.F., Medema,R.H., Pals,C. et al. (2000) Forkhead transcriptionfactor FKHR-L1 modulates cytokine-dependent transcriptional regulationof p27 (KIP1). Mol. Cell. Biol., 20, 9138--9148.
29.Delmas,C., Manenti,S., Boudjelal,A., Peyssonnaux,C., Eychene,A. andDarbon,J.M. (2001) The p42/p44 mitogen-activated protein kinaseactivation triggers p27Kip1 degradation independently of CDK2/cyclinE in NIH 3T3 cells. J. Biol. Chem., 276, 34958--34965.
30.Sulis,M.L. and Parsons,R. (2003) PTEN: from pathology to biology.Trends Cell Biol., 13, 478--483.
31.Toyoshima,H. and Hunter,T. (1994) p27, a novel inhibitor of G1 cyclin-Cdk protein kinase activity, is related to p21. Cell, 78, 67--74.
32. Johnson,P.J. (2002) Hepatocellular carcinoma: is current therapy reallyaltering outcome? Gut, 51, 459--462.
33.Ng,I.O., Chung,L.P., Tsang,S.W., Lam,C.L., Lai,E.C., Fan,S.T. and Ng,M.(1994) p53 gene mutation spectrum in hepatocellular carcinomas in HongKong Chinese. Oncogene, 9, 985--990.
Received March 31, 2004; revised June 30, 2004; accepted July 25, 2004
FTY720 induces apoptosis
2405
by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

by guest on August 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from







![Novel pyrazolo[1,5-a]pyridines as p110α-selective PI3 kinase inhibitors: Exploring the benzenesulfonohydrazide SAR](https://img.pdfslide.net/doc/110x75/634c7245d2302b71f40cd024/novel-pyrazolo15-apyridines-as-p110-selective-pi3-kinase-inhibitors-exploring.jpg)





















