Embed Size (px)
Citation preview

Research Article
Functional analysis of the Neurospora crassa PZL-1protein phosphatase by expression in budding and®ssion yeast
Emese Vissi1, Josep Clotet2, Eulalia de Nadal2, Anna BarceloÂ2, EÂva BakoÂ1, PaÂl Gergely1, Viktor DombraÂdi1
and JoaquõÂn ArinÄo2*1 Department of Medical Chemistry, Faculty of Medicine, Medical and Health Science Centre, University of Debrecen, H-4026 Debrecen,Hungary
2 Departament de BioquõÂmica i Biologia Molecular, Universitat AutoÁnoma de Barcelona, Bellaterra 08193 Barcelona, Spain
*Correspondence to:J. ArinÄo, Dept. BioquõÂmica iBiologia Molecular, Facultat deVeterinaÁria, Ed. V, UniversitatAutoÁnoma de Barcelona,Bellaterra 08193, Barcelona,Spain.E-mail: [email protected]
Received: 9 June 2000
Accepted: 2 August 2000
Abstract
The gene pzl-1 from the ®lamentous fungus Neurospora crassa encodes a putative Ser/Thr
protein phosphatase that is reminiscent of the Ppz1/Ppz2 and Pzh1 phosphatases from
Saccharomyces cerevisiae and Schizosaccharomyces pombe, respectively. The entire PZL-
1 protein, as well as its carboxyl-terminal domain, have been expressed in Escherichia colias active protein phosphatases. To characterize its cellular role, PZL-1 was also expressed
in Sz. pombe and in S. cerevisiae. Expression of PZL-1 in S. cerevisiae from the PPZ1promoter was able to rescue the altered sensitivity to caffeine and lithium ions of a ppz1strain. Furthermore, high copy number expression of PZL-1 alleviated the lytic phenotype
of a S. cerevisiae slt2/mpk1 mitogen-activated protein (MAP) kinase mutant, similarly to
that described for PPZ1, and mimicked the effects of high levels of Ppz1 on cell growth.
Expression of PZL-1 in ®ssion yeast from a weak version of the nmt1 promoter fully
rescued the growth defect of a pzh1D strain in high potassium, but only partially
complemented the sodium-hypertolerant phenotype. Strong overexpression of the N. crassaphosphatase in Sz. pombe affected cell growth and morphology. Therefore, PZL-1 appears
to ful®l every known function carried out by its S. cerevisiae counterpart, despite the
marked divergence in sequence within their NH2-terminal moieties. Copyright # 2000
John Wiley & Sons, Ltd.
Keywords: Ppz phosphatases; functional expression; yeast; salt tolerance
Introduction
The PPZ enzymes de®ne a novel type of Ser/Thrprotein phosphatases that have been identi®ed inbudding yeast (Saccharomyces cerevisiae; Posaset al., 1992) and ®ssion yeast (Schizosaccharomycespombe; Balcells et al., 1997). These phosphatasesdisplay a COOH-terminal catalytic domain ofabout 300 residues, which is about 60% identicalto the catalytic subunit of protein phosphatase-1,and a NH2-terminal moiety that is unrelated toother known phosphatases. Budding yeast containstwo genes for PPZ, encoding the Ppz1 (692residues) and Ppz2 (710 residues) proteins (Posaset al., 1992; Lee et al., 1993, Hughes et al., 1993).
They are about 93% identical within their COOH-terminal portion, whereas their NH2-terminalregions are less related (43%). Strains lackingPPZ1 are prone to cell lysis under some stresssituations, such as exposure to low concentrationsof caffeine (Posas et al., 1993), and the deletion ofPPZ1 is synthetically lethal in combination with theabsence of the Slt2/Mpk1 MAP kinase (Lee et al.,1993). In addition, lack of PPZ1 results in increasedtolerance to sodium and lithium ions and this effecthas been attributed to an increased expression ofthe Na+-ATPase encoded by the gene ENA1 (Posaset al., 1995a). Deletion of PPZ2 in a ppz1 back-ground intensi®es these effects, although it does notresult in a detectable phenotype in an otherwise
YeastYeast 2001; 18: 115±124.
Copyright # 2000 John Wiley & Sons, Ltd.

wild-type cell. Recent evidences suggest that Ppz1
might play a role in the regulation of the cell cycle,
as we have observed that the deletion of PPZ1
improves the defective growth of mutants in the
SIT4 gene (Clotet et al., 1999). SIT4 encodes a Ser/
Thr protein phosphatase involved in the G1±S
transition of the cell cycle (Arndt et al., 1989;
Sutton et al., 1991; FernaÂndez-Sarabia et al., 1992).
It has been proposed that Ppz1 might integrate
diverse stimuli relevant for cell cycle progression
(Clotet et al., 1999).The Sz. pombe gene pzh1+ encodes a homologue
of the PPZ phosphatases (Balcells et al., 1997). Pzh1
is a 515-residue protein, whose COOH-terminal
segment is 78% identical to Ppz1 and Ppz2, whereas
its NH2-terminal domain is shorter and, with the
exception of the ®rst 40 amino acids, shows a low
level of similarity to other proteins (Figure 1). Pzh1
is also involved in the regulation of salt tolerance,
although its role in Sz. pombe might differ substan-
tially from that of the PPZ phosphatases in S.
cerevisiae (Balcells et al., 1997, 1998, 1999). In fact,
in our hands the Pzh1 protein has been unable to
restore the wild-type phenotype of S. cerevisiae ppz1
strain, or to substitute for Ppz1 under different
conditions (Balcells et al., 1997; Clotet J, ArinÄo J,
unpublished results).Recently, a phosphatase gene and cDNA encod-
ing a PPZ-like protein (pzl-1) have been identi®ed
and cloned in Neurospora crassa (SzoÈoÄr et al.,
1998). The predicted open reading frame is similar
in size to ®ssion yeast Pzh1 (531 residues) and can
be also divided in two segments. The COOH-terminal portion, spanning about 300 residues, ishighly similar to the budding and ®ssion yeast PPZphosphatases (Figure 1). The NH2-terminal moietyof PZL-1 is rich in Ser and Pro and, althoughoverall it is quite unrelated to Ppz1/Ppz2 or Pzh1, itretains a signi®cant level of identity over the ®rst 50residues. This includes a NH2-terminal myristoyla-tion consensus sequence, which appears as acommon trait for all Ppz phosphatases, and isimportant for the proper function of Ppz1 (Clotetet al., 1996).
The identi®cation of a PPZ-like enzyme in the®lamentous fungus N. crassa raised the question ofits functional role. We have addressed this questionby expressing this protein in bacteria, to character-ize its catalytic activity, and in budding and ®ssionyeast, to evaluate whether or not the PZL-1 proteincan ful®l the biological functions of Ppz1 and/orPzh1.
Materials and methods
Growth of Escherichia coli and yeast strains
E. coli cells (strain DH5a) were grown at 37uC inLB medium (containing 50 mg/ml ampicillin forplasmid selection), unless otherwise stated. Sz.pombe cells were grown at 28uC in YES (0.5%yeast extract, 3% glucose, plus 225 mg/ml of ade-nine, uracil and leucine) or essential minimalmedium (EMM) supplemented with the necessaryrequirements (Moreno et al., 1991). The pH of themedium was buffered at 5.2 with 20 mM MES. S.cerevisiae cells were grown at 28uC in YPD mediumor, when indicated, in CM synthetic medium (Sher-man et al., 1986). The relevant genotypes of thedifferent S. cerevisiae strains used in this work areshown in Table 1. The Sz. pombe strain tested forexpression of PZL-1 was strain LB2 (pzh1::ura4+),described earlier (Balcells et al., 1997).
Recombinant DNA techniques
E. coli cells were transformed using the standardcalcium chloride method (Sambrook et al., 1989). S.cerevisiae cells were transformed by a modi®cationof the lithium acetate method (Schiestl andGietz, 1989), that includes treatment with dimethylsulphoxide. Sz. pombe cells were transformed bya modi®cation of the lithium acetate method
Figure 1. Schematic alignment of the S. cerevisiae Ppz1/Ppz2,Sz. pombe Pzh1 and N. crassa PZL-1 proteins. The dottedareas represent the conserved NH2-terminal element. Theasterisk denotes the conserved myristoylation consensussequence surrounding Gly-2. Numbers indicate the percen-tage of amino acid identities within the highly conservedphosphatase carboxyl-terminal domains, represented byblack boxes
116 E. Vissi et al.
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.

(Norbury and Moreno, 1997). Restriction diges-tions, DNA ligations and other standard recombi-nant DNA techniques were performed essentially asdescribed (Sambrook et al., 1989).
Expression of PZL-1 in bacterial cells
The entire open reading frame of N. crassa pzl-1was ampli®ed by PCR using a forward oligo-nucleotide 5k-CGGAATTCATGGGCAACTCATCCTC-3k and the reverse oligonucleotide 5k-GATGCGGCCGCTCAGACACTCTGGGGG-3k (Eco-RI and NotI sites, respectively, which are under-lined, were added to facilitate cloning). Theampli®ed 1.6 kbp fragment was cloned into theplasmid pGEX 4T1 (Pharmacia), previouslydigested with EcoRI and NotI, to yield plasmidpGEX±Pzl1. The carboxy-terminal moiety of PZL-1was ampli®ed with the forward primer 5k-CGGAATTCGGGTGTTGCCATGAG-3k (addedEcoRI site underlined) and the reverse primerdescribed above. The ampli®ed fragment (about1.0 kbp) was cloned into EcoRI and NotI digestedpGEX 4T3 vector (Pharmacia), to generatepGEX±Pzl1/CT.
For bacterial expression, DH5a E. coli cells weretransformed with plasmids and grown up to OD600
of 1.0 in 2rYT-G media (16 g/l tryptone, 10 g/lyeast extract, 5 g/l NaCl and 20 g/l glucose) contain-ing 1 mM MnCl2. Cells were induced by adding1 mM IPTG (0.2 mM for cells bearing pGEX-Pzl1/CT) and growth was resumed for 2 h at 28uC. Cellswere collected and disrupted with an Ultra Thuraxhomogenizer (4r1 min at 20 000 rpm) in thepresence of a buffer containing 50 mM Tris±HCl,pH 8.0, 1 mM MnCl2, 1% Triton X-100, 10mM
DTT, 0.2 mM phenylmethylsulphonyl ¯uoride,5 mM benzamidine and 1 mM o-phenantroline at0uC. Extracts were centrifuged at 3000rg for15 min at 4uC and the glutathione-S-transferase(GST) fusion proteins were puri®ed from the
supernatants by incubation with glutathione-Sepharose beads (Pharmacia) for 1 h at 4uC. Thebeads were washed three times with 50 mM Tris,pH 8.0, 0.1% b-mercaptoethanol, and the fusionproteins were eluted with the same buffer contain-ing 20 mM reduced glutathione.
Expression of N. crassa PZL-1 in S. cerevisiaeand Sz. pombe
For expression of PZL-1 in S. cerevisiae, thepromoter region (±525/±10) of S. cerevisiae PPZ1was ampli®ed by PCR using oligonucleotides A1 5k-GCG AGC TCG TCC TCC AAT TCA AC-3k andA2 5k-CGT CTA GAA GGA AAG ATA AGCAGA G-3k (added SacI and XbaI sites are under-lined) and Expand High Fidelity DNA polymerase(Roche). The 0.6 kbp fragment was cloned into theSacI and XbaI sites of plasmids YCplac111 andYEplac181 (which contain a LEU2 selectionmarker). The complete open reading frame of N.crassa pzl-1 was ampli®ed from the cloned cDNA(SzoÈoÄr et al., 1998) by PCR using oligonucleotidesA3 5k-CGT CTAGA A ATC ATG GGC AACTCA TC-3k and A4 5k-CAG AAG CTT TGG CTAGCG TGC AG-3k (added XbaI and HindIII sitesunderlined). This fragment was cloned into theabove-mentioned constructs to yield YCplac111±Pzl1 and YEplac181±Pzl1. The hybrid fragment(PPZ1 S. cerevisiae promoter plus N. crassa pzl-1open reading frame) was released by digestion withSacI±HindIII and used for cloning in the alternativeyeast vector YEplac195, bearing a URA3 selectionmarker, to generate YEplac195±Pzl1.
For expression of PZL-1 in ®ssion yeast, theentire pzl-1 open reading frame was ampli®ed witholigonucleotides A5 5k-CGC TGA TCA TGG GCAACT CAT CCT C-3k (added BclI site underlined)and A4 (see above). The fragment was digestedwith BclI, to create a BamHI-compatible end,and ligated into vectors pREP3x, pREP41x and
Table 1. S. cerevisiae strains used in this work
Strains Relevant genotype Reference
JA-100* MATa PPZ1 PPZ2 SIT4 MPK1 De Nadal et al., 1998
JA-101 MATa ppz1::URA3 PPZ2 SIT4 MPK1 De Nadal et al., 1998JC-10 MATa PPZ1 PPZ2 mpk1::LEU2 SIT4 This work
JA-110 MATa PPZ1 PPZ2 MPK1 sit4::TRP1 Clotet et al., 1999
JA-112 MATa ppz1::URA3 PPZ2 MPK1 sit4::TRP1 Clotet et al., 1999
*JA-100 is a haploid derivative from strain DL790 (Lee et al., 1993) and is ura3-52 leu2-3,112 trp1-1 his4 can-1r.
Expression of the N. crassa PZL-1 phosphatase 117
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.

pREP81x (Forsburg, 1993), previously digestedwith BamHI and SmaI, to yield pREP3x±Pzl1,pREP41x±Pzl1 and pREP81x±Pzl1. The insertswere sequenced using an automated DNA sequen-cer to verify the absence of unwanted mutations.These constructs were used to transform wild-typeand LB2 (pzh1) Sz. pombe cells. Positive clones weremaintained in medium containing 4 mm thiamine,to avoid undesired expression from the nmt1promoter, unless otherwise stated.
Other techniques
Sensitivity of S. cerevisiae strains to caffeine, NaCland LiCl was tested as previously reported (Clotetet al., 1996). Recovery from a-factor arrest wasmonitored as in Clotet et al. (1999). Sensitivity ofSz. pombe cells to ions was tested as describedearlier (Balcells et al., 1997). Growth rate determi-nations were performed essentially as in Clotet et al.(1996). Measurements of protein phosphatase activ-ity were carried out essentially as described pre-viously (Posas et al., 1995b), except that theincubation assay contained no MnCl2. One unit ofphosphatase is de®ned as the amount of enzymewhich releases one mmol phosphate/min from 32P-labelled myelin basic protein.
Results and discussion
Characterization of bacterially expressedPZL-1
The entire open reading frame encoding the PZL-1protein, as well as the region from residue 196 tothe stop codon (comprising the presumed phospha-tase catalytic domain), were successfully expressedas a soluble GST fusion protein in E. coli. The yieldwas relatively low (about 1.5±2 mg/l of culture). Thesoluble protein was puri®ed through glutathione-Sepharose columns, yielding essentially homoge-neous proteins (not shown). The estimated molecu-lar mass was 84 kDa for the full-length GST±Pzl1construct, and 61 kDa for the GST±catalyticdomain fusion protein, in good agreement with thecalculated values. Activity measurements demon-strated that the recombinant proteins were able todephosphorylate 32P-labelled myelin basic protein,as previously shown for Ppz1 and Pzh1 (Posas et al.,1995b; Balcells et al., 1997). The speci®c activity ofthe catalytic moiety was higher than that of the
entire protein (3.55 vs. 0.22 mU/mg protein), sug-gesting an inhibitory function for the NH2-terminalsegment. Extracts of bacteria harbouring no phos-phatase-containing plasmids showed a negligiblephosphatase activity in this assay (0.002 mU/mg).As indicated in Table 2, the phosphatase activity ofPZL-1 was inhibited by ATP and ¯uoride, similarlyto its homologues, but was less sensitive to micro-cystin LR and okadaic acid than bacteriallyexpressed Ppz1 or Pzh1 (Posas et al., 1995b; Balcellset al., 1997).
Functional analysis of PZL-1 in budding yeast
To test the possibility that N. crassa PZL-1phosphatase might functionally replace Ppz1, weexpressed the pzl-1 open reading frame from thePPZ1 promoter in ppz1 S. cerevisiae mutants.Budding yeast strains lacking PPZ1 display aphenotype of increased tolerance to sodium andlithium (Posas et al., 1995a). We ®rst tested whetherPZL-1 was able to rescue the lithium-tolerantphenotype of ppz1 cells. As can be observed inFigure 2, expression of pzl-1 from a low-copy,centromeric plasmid resulted in a reduction in thelithium tolerance of a ppz1 strain, similarly to theexpression of budding yeast PPZ1. The effect ofhigh-copy number expression of pzl-1 on salttolerance was dif®cult to assess because it resultedin a reduced cell growth rate, even in the absence ofsalt stress (see Figure 5).
In addition to the salt tolerance phenotype, inbudding yeast the absence of Ppz1 results inhypersensitivity to caffeine, as it has been reportedthat exposure to low concentrations of this drugcauses cell lysis (Posas et al., 1993). We show inFigure 3 that expression of N. crassa PZL-1 from
Table 2. Enzymological characteristics of bacteriallyexpressed PZL-1
Inhibitor*
Expressed protein
Pzl1 Pzl1/CT
ATP (mM) 23t2 40t4
NaF (mM) 25t2 20t2
Microcystin (nM) 66t3 66t5
*Data represent the IC50 values for each compound and are
meantSD from three to seven determinations. 2 mm okadaic acidhas no signi®cant effect on the phosphatase activity. Pzl1/CT refers to
the C-terminal catalytic fragment of Pzl1.
118 E. Vissi et al.
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.

Figure 2. Expression of N. crassa PZL-1 rescues the lithium hypertolerant phenotype of a ppz1 S. cerevisiae strain. StrainsJA-100 (PPZ1) or JA-101 (ppz1) were transformed with plasmids YEplac181 (YEp), YCplac111±PPZ1 (YCp±PPZ1),YEplac181±PPZ1 (YEp±PPZ1), YCplac111±Pzl1 (YCp±Pzl1), or YEplac181±Pzl1 (YEp±Pzl1) and plated (initial A660 of 0.05) onCM plates (lacking leucine) containing LiCl at the indicated concentrations. Plates were incubated at 30uC and growth wasscored after 2 days (except for cells grown in the absence of LiCl, which were scored for growth after 32 h to show theeffect of high-level expression of pzl-1 in the absence of salt)
Figure 3. Expression of N. crassa PZL-1 rescues the caffeine-sensitive phenotype of a budding yeast ppz1 strain. StrainsJA-100 (PPZ1) or JA-101 (ppz1) were transformed with plasmids YEplac181 (YEp), YCplac111±PPZ1 (YCp±PPZ1),YCplac111±Pzl1 (YCp±Pzl1), or YEplac181±Pzl1 (YEp±Pzl1) and plated on CM plates (lacking leucine) containing caffeine atthe indicated concentrations. Plates were incubated at 30uC and growth was scored after 2 days
Expression of the N. crassa PZL-1 phosphatase 119
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.

the S. cerevisiae PPZ1 promoter results in a
complete rescue of the caffeine-sensitive phenotype
of a ppz1 strain. It is remarkable that the presence
of PZL-1 virtually mimics the expression of bona
®de Ppz1, and that rescue is achieved even when
PZL-1 is expressed from a low-copy, centromeric
plasmid. By contrast, rescue of the caffeine-sensitive
phenotype in budding yeast is achieved by ®ssion
yeast Pzh1 only when expressed from a high copy
number plasmid (Balcells et al., 1997). It must be
emphasized that the Pzh1 phosphatase was also
expressed from the PPZ1 promoter in budding
yeast.The caffeine-sensitive phenotype of ppz1 mutants
has been related to the role of this phosphatase in
the maintenance of cell integrity (Posas et al., 1993).
A functional relationship between Ppz1 and the
PKC1-regulated Slt2±Mpk1 MAP kinase pathway
has been established (Lee et al., 1993). mpk1
mutants, like ppz1 mutants, are caffeine-sensitive.
In addition, mpk1 cells are temperature-sensitive
and cannot grow at 37uC unless osmotically
stabilized. High copy number expression of PPZ1
allows mpk1 to grow in the absence of osmotic
stabilizers (Lee et al., 1993). Figure 4 shows that
high copy number expression of PZL-1 is also able
to allow growth of a mpk1 mutant under non-
permissive conditions in the absence of 1 M sorbitol.
This observation suggests that the N. crassa PZL-1
phosphatase is able to perform certain function(s)
related to the maintenance of cell integrity in
budding yeast, similarly to endogenous Ppz1. It is
remarkable that, by contrast, even high copy
number expression of ®ssion yeast pzh1+ is unable
to rescue the temperature-sensitive phenotype of
mpk1 cells (J. Clotet & J. ArinÄo, unpublished).We have recently reported that, in budding yeast,
Ppz1 is negatively regulated by the Hal3 protein
(De Nadal et al., 1998). The gene HAL3/SIS2 was
isolated as a determinant of salt tolerance (Fer-
rando et al., 1995). On the basis that high copy
number expression of HAL3/SIS2 was able to
rescue the growth defect of sit4 mutants, it was
also identi®ed as a component of the machinery
that regulates the progression of the cell cycle
through the G1±S transition (Di Como et al.,
1995). This evidence led us to postulate that the
effect of Hal3 on the cell cycle might be mediated
by Ppz1 and that, in consequence, Ppz1 might be a
regulatory component of the cell cycle. This
hypothesis is in agreement with the fact that high
levels of Ppz1 are detrimental for growth (Clotet
Figure 4. High-copy number expression of N. crassa PZL-1 rescues the temperature-sensitive lytic phenotype of a MAPkinase-de®cient mpk1 S. cerevisiae strain. Wild-type strain JA-100 (wt) was transformed with plasmid YEplac195 (YEp),whereas the isogenic strain JC10 (mpk1::LEU2) was transformed with the same plasmid or with plasmid YEplac195±Pzl1(YEp±Pzl1). Cells were plated on CM plates lacking uracil, with or without 1 M sorbitol, and were grown at 37uC for 2 days
120 E. Vissi et al.
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.
et al., 1996), and it has recently received furthersupport from a number of observations, includingthe evidence that deletion of Ppz1 results in therescue of the sit4 growth defect similarly to the
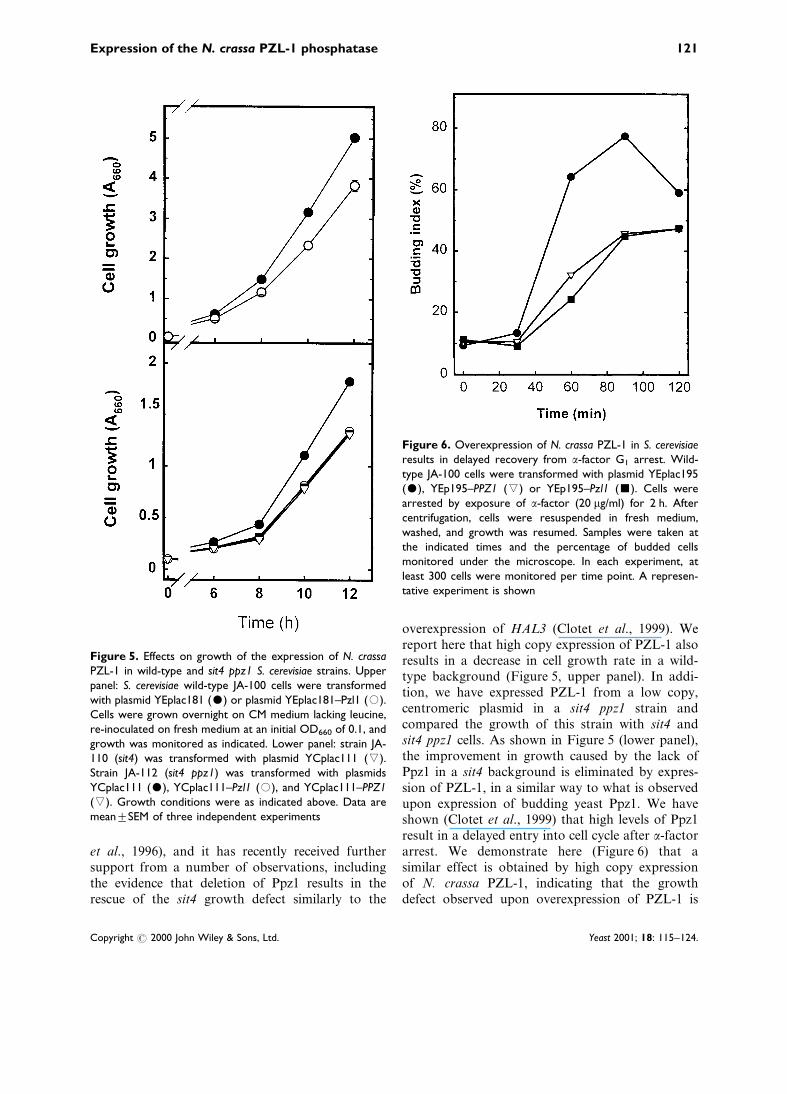
overexpression of HAL3 (Clotet et al., 1999). Wereport here that high copy expression of PZL-1 alsoresults in a decrease in cell growth rate in a wild-type background (Figure 5, upper panel). In addi-tion, we have expressed PZL-1 from a low copy,centromeric plasmid in a sit4 ppz1 strain andcompared the growth of this strain with sit4 andsit4 ppz1 cells. As shown in Figure 5 (lower panel),the improvement in growth caused by the lack ofPpz1 in a sit4 background is eliminated by expres-sion of PZL-1, in a similar way to what is observedupon expression of budding yeast Ppz1. We haveshown (Clotet et al., 1999) that high levels of Ppz1result in a delayed entry into cell cycle after a-factorarrest. We demonstrate here (Figure 6) that asimilar effect is obtained by high copy expressionof N. crassa PZL-1, indicating that the growthdefect observed upon overexpression of PZL-1 is
Figure 5. Effects on growth of the expression of N. crassaPZL-1 in wild-type and sit4 ppz1 S. cerevisiae strains. Upperpanel: S. cerevisiae wild-type JA-100 cells were transformedwith plasmid YEplac181 ($) or plasmid YEplac181±Pzl1 (#).Cells were grown overnight on CM medium lacking leucine,re-inoculated on fresh medium at an initial OD660 of 0.1, andgrowth was monitored as indicated. Lower panel: strain JA-110 (sit4) was transformed with plasmid YCplac111 (().Strain JA-112 (sit4 ppz1) was transformed with plasmidsYCplac111 ($), YCplac111±Pzl1 (#), and YCplac111±PPZ1((). Growth conditions were as indicated above. Data aremeantSEM of three independent experiments
Figure 6. Overexpression of N. crassa PZL-1 in S. cerevisiaeresults in delayed recovery from a-factor G1 arrest. Wild-type JA-100 cells were transformed with plasmid YEplac195($), YEp195±PPZ1 (() or YEp195±Pzl1 (&). Cells werearrested by exposure of a-factor (20 mg/ml) for 2 h. Aftercentrifugation, cells were resuspended in fresh medium,washed, and growth was resumed. Samples were taken atthe indicated times and the percentage of budded cellsmonitored under the microscope. In each experiment, atleast 300 cells were monitored per time point. A represen-tative experiment is shown
Expression of the N. crassa PZL-1 phosphatase 121
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.
the result of a delayed G1±S transition. Therefore,all of the experimental evidences strongly suggestthat the heterologous phosphatase is functionallyreplacing Ppz1.
Functional analysis of PZL-1 in ®ssion yeast
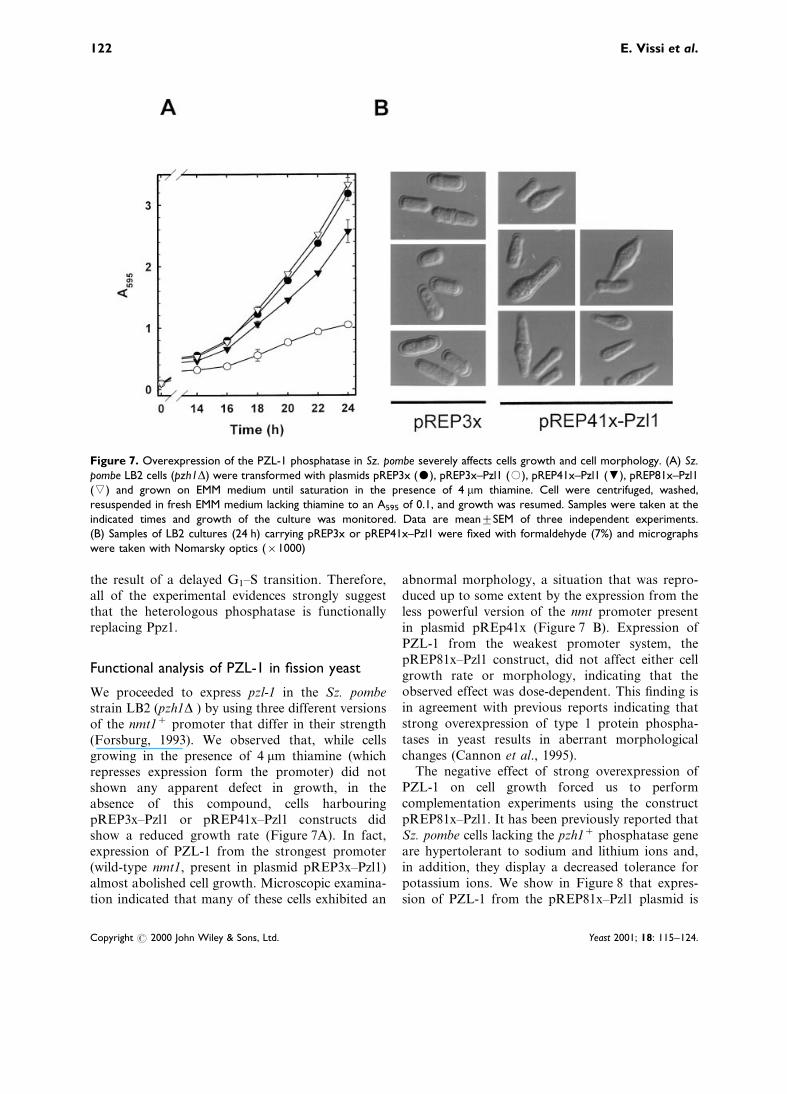
We proceeded to express pzl-1 in the Sz. pombestrain LB2 (pzh1D ) by using three different versionsof the nmt1+ promoter that differ in their strength(Forsburg, 1993). We observed that, while cellsgrowing in the presence of 4 mm thiamine (whichrepresses expression form the promoter) did notshown any apparent defect in growth, in theabsence of this compound, cells harbouringpREP3x±Pzl1 or pREP41x±Pzl1 constructs didshow a reduced growth rate (Figure 7A). In fact,expression of PZL-1 from the strongest promoter(wild-type nmt1, present in plasmid pREP3x±Pzl1)almost abolished cell growth. Microscopic examina-tion indicated that many of these cells exhibited an
abnormal morphology, a situation that was repro-duced up to some extent by the expression from theless powerful version of the nmt promoter presentin plasmid pREp41x (Figure 7 B). Expression ofPZL-1 from the weakest promoter system, thepREP81x±Pzl1 construct, did not affect either cellgrowth rate or morphology, indicating that theobserved effect was dose-dependent. This ®nding isin agreement with previous reports indicating thatstrong overexpression of type 1 protein phospha-tases in yeast results in aberrant morphologicalchanges (Cannon et al., 1995).
The negative effect of strong overexpression ofPZL-1 on cell growth forced us to performcomplementation experiments using the constructpREP81x±Pzl1. It has been previously reported thatSz. pombe cells lacking the pzh1+ phosphatase geneare hypertolerant to sodium and lithium ions and,in addition, they display a decreased tolerance forpotassium ions. We show in Figure 8 that expres-sion of PZL-1 from the pREP81x±Pzl1 plasmid is
Figure 7. Overexpression of the PZL-1 phosphatase in Sz. pombe severely affects cells growth and cell morphology. (A) Sz.pombe LB2 cells (pzh1D) were transformed with plasmids pREP3x ($), pREP3x±Pzl1 (#), pREP41x±Pzl1 (,), pREP81x±Pzl1(() and grown on EMM medium until saturation in the presence of 4 mm thiamine. Cell were centrifuged, washed,resuspended in fresh EMM medium lacking thiamine to an A595 of 0.1, and growth was resumed. Samples were taken at theindicated times and growth of the culture was monitored. Data are meantSEM of three independent experiments.(B) Samples of LB2 cultures (24 h) carrying pREP3x or pREP41x±Pzl1 were ®xed with formaldehyde (7%) and micrographswere taken with Nomarsky optics (r1000)
122 E. Vissi et al.
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.
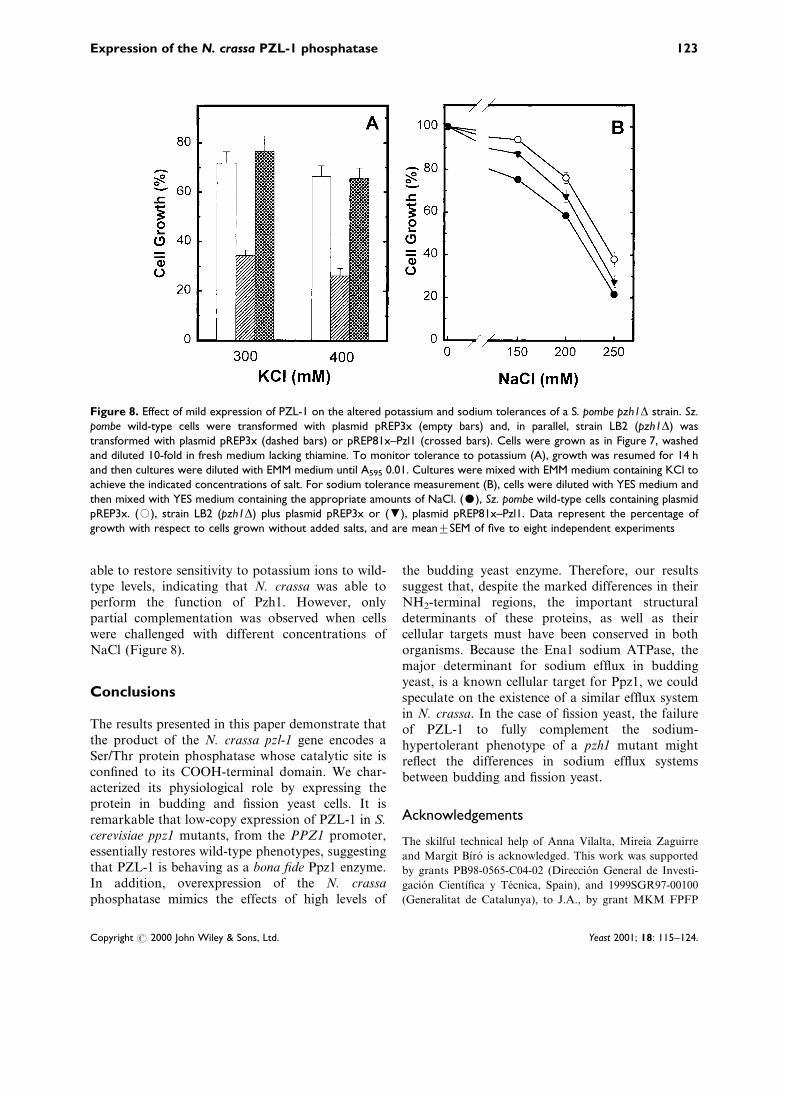
able to restore sensitivity to potassium ions to wild-type levels, indicating that N. crassa was able toperform the function of Pzh1. However, onlypartial complementation was observed when cellswere challenged with different concentrations ofNaCl (Figure 8).
Conclusions
The results presented in this paper demonstrate thatthe product of the N. crassa pzl-1 gene encodes aSer/Thr protein phosphatase whose catalytic site iscon®ned to its COOH-terminal domain. We char-acterized its physiological role by expressing theprotein in budding and ®ssion yeast cells. It isremarkable that low-copy expression of PZL-1 in S.cerevisiae ppz1 mutants, from the PPZ1 promoter,essentially restores wild-type phenotypes, suggestingthat PZL-1 is behaving as a bona ®de Ppz1 enzyme.In addition, overexpression of the N. crassaphosphatase mimics the effects of high levels of
the budding yeast enzyme. Therefore, our resultssuggest that, despite the marked differences in theirNH2-terminal regions, the important structuraldeterminants of these proteins, as well as theircellular targets must have been conserved in bothorganisms. Because the Ena1 sodium ATPase, themajor determinant for sodium ef¯ux in buddingyeast, is a known cellular target for Ppz1, we couldspeculate on the existence of a similar ef¯ux systemin N. crassa. In the case of ®ssion yeast, the failureof PZL-1 to fully complement the sodium-hypertolerant phenotype of a pzh1 mutant mightre¯ect the differences in sodium ef¯ux systemsbetween budding and ®ssion yeast.
Acknowledgements
The skilful technical help of Anna Vilalta, Mireia Zaguirre
and Margit BõÂro is acknowledged. This work was supported
by grants PB98-0565-C04-02 (DireccioÂn General de Investi-
gacioÂn Cientõ®ca y TeÂcnica, Spain), and 1999SGR97-00100
(Generalitat de Catalunya), to J.A., by grant MKM FPFP
Figure 8. Effect of mild expression of PZL-1 on the altered potassium and sodium tolerances of a S. pombe pzh1D strain. Sz.pombe wild-type cells were transformed with plasmid pREP3x (empty bars) and, in parallel, strain LB2 (pzh1D) wastransformed with plasmid pREP3x (dashed bars) or pREP81x±Pzl1 (crossed bars). Cells were grown as in Figure 7, washedand diluted 10-fold in fresh medium lacking thiamine. To monitor tolerance to potassium (A), growth was resumed for 14 hand then cultures were diluted with EMM medium until A595 0.01. Cultures were mixed with EMM medium containing KCl toachieve the indicated concentrations of salt. For sodium tolerance measurement (B), cells were diluted with YES medium andthen mixed with YES medium containing the appropriate amounts of NaCl. ($), Sz. pombe wild-type cells containing plasmidpREP3x. (#), strain LB2 (pzh1D) plus plasmid pREP3x or (,), plasmid pREP81x±Pzl1. Data represent the percentage ofgrowth with respect to cells grown without added salts, and are meantSEM of ®ve to eight independent experiments
Expression of the N. crassa PZL-1 phosphatase 123
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.

0784/1997 (Ministry of Education) to G.P, and by grant
OTKA 22675 (Hungarian Research Foundation) to V.D. E.
de N. was recipient of a Fellowship from the `Ministerio de
EducacioÂn y Cultura', Spain. E.V. was recipient of a
fellowship within the TEMPUS-PHARE and ERASMUS
frameworks.
References
Arndt KT, Styles CA, Fink GR. 1989. A suppressor of a HIS4
transcriptional defect encodes a protein with homology to the
catalytic subunit of protein phosphatases. Cell 56: 527±537.
Balcells Ll, GoÂmez N, Casamayor A, Clotet J, ArinÄo J. 1997.
Regulation of salt tolerance in ®ssion yeast by a PPZ-like Ser/
Thr protein phosphatase. Eur J Biochem 250: 476±483.
Balcells L, MartõÂn R, Ruiz MC, GoÂmez N, Ramos J, ArinÄo J.
1998. The Pzh1 protein phosphatase and the Spm1 protein
kinase are involved in the regulation of the plasma membrane
H+-ATPase in ®ssion yeast. FEBS Lett 435: 241±244.
Balcells L, Calero F, GoÂmez N, Ramos J, ArinÄo J. 1999. The
Schizosaccharomyces pombe Pzh1 protein phosphatase regu-
lates sodium in¯ux in a Trk1-independent fashion. Eur
J Biochem 260: 31±37.
Cannon JF, Clemens KE, Morcos PA, Nair BM, Pearson JL,
Khalil M. 1995. Type 1 protein phosphatase systems of yeast.
Adv Prot Phosphatases 9: 211±232.
Clotet J, Posas F, De Nadal E, ArinÄo J. 1996. The NH2-terminal
extension of protein phosphatase PPZ1 has an essential
functional role. J Biol Chem 271: 26349±26355.
Clotet J, GarõÂ E, Aldea M, ArinÄo J 1999. The yeast Ser/Thr
phosphatases Sit4 and Ppz1 play opposite roles in the
regulation of the cell cycle. Mol Cell Biol 19: 2408±2415.
De Nadal E, Clotet J, Posas F, Serrano R, GoÂmez N, ArinÄo J.
1998. The yeast halotolerance determinant Hal3p is an
inhibitory subunit of the Ppz1p Ser/Thr protein phosphatase.
Proc Natl Acad Sci USA 95: 7357±7362.
Di Como CJ, Bose R, Arndt KT. 1995. Overexpression of SIS2,
which contains an extremely acidic region, increases the
expression of SWI4, CLN1 and CLN2 in sit4 mutants.
Genetics 139: 95±107.
FernaÂndez-Sarabia MJ, Sutton A, Zhong T, Arndt KT. 1992.
SIT4 protein phosphatase is required for the normal accumu-
lation of SWI4, CLN1, CLN2, and HCS26 RNAs during late
G1. Genes Dev 6: 2417±2428.
Ferrando A, Kron SJ, Rios G, Fink GR, Serrano R. 1995.
Regulation of cation transport in Saccharomyces cerevisiae by
the salt tolerance gene HAL3. Mol Cell Biol 15: 5470±5481.
Forsburg SL. 1993. Comparison of Schizosaccharomyces pombe
expression systems. Nucleic Acids Res 21: 2955±2956.
Hughes V, MuÈ ller A, Stark MJ, Cohen PTW. 1993. Both
isoforms of protein phosphatase Z are essential for the
maintenance of cell size and integrity in Saccharomyces
cerevisiae in response to osmotic stress. Eur J Biochem 216:
269±279.
Lee KS, Hines LK, Levin DE. 1993. A pair of functionally
redundant yeast genes (PPZ1 and PPZ2) encoding type 1-
related protein phosphatases function within the PKC1-
mediated pathway. Mol Cell Biol 13: 5843±5853.
Moreno S, Amar K, Nurse P. 1991. Molecular genetic analysis of
®ssion yeast Schizosaccharomyces pombe. In Guide to Yeast
Genetics and Molecular Biology, Guthrie C, Fink GR (Eds).
795±823. Academic Press Inc., San Diego, CA.
Norbury C, Moreno S. 1997. Cloning cell cycle regulatory genes
by trans-complementation in yeast. Methods Enzymol 283:
44±59.
Posas F, Casamayor A, Morral N, ArinÄo J. 1992. Molecular
cloning and analysis of a yeast protein phosphatase with an
unusual amino-terminal region. J Biol Chem 267: 11734±11740.
Posas F, Casamayor A, ArinÄo J. 1993. The PPZ protein
phosphatases are involved in the maintenance of osmotic
stability of yeast cells. FEBS Lett 318: 282±286.
Posas F, Camps M, ArinÄo J. 1995a. The PPZ protein
phosphatases are important determinants of salt tolerance in
yeast cells. J Biol Chem 270: 13036±13041.
Posas F, Bollen M, Stalmans W, ArinÄo J 1995b. Biochemical
characterization of recombinant yeast PPZ1, a protein phos-
phatase involved in salt tolerance. FEBS Lett 368: 39±44.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor Laboratory Press:
New York.
Schiestl RH, Gietz RD. 1989. High ef®ciency transformation of
intact yeast cells using single stranded nucleic acids as a
carrier. Curr Genet 16: 339±346.
Sherman F, Fink GR, Hicks JB. 1986. Laboratory Course
Manual for Methods in Yeast Genetics. Cold Spring Harbor
Laboratory Press: New York.
Sutton A, Immanuel, D, Arndt KT. 1991. The SIT4 protein
phosphatase functions in late G1 for progression into S phase.
Mol Cell Biol 11: 2133±2148.
SzoÈoÄr B, FeheÂr Z, Zeke T, Gergely P, Yatzkan E, Yarden O,
DombraÂdi V. 1998. pzl-1 encodes a novel protein phosphatase-
Z-like Ser/Thr protein phosphatase in Neurospora crassa.
Biochim Biophys Acta 1388: 260±266.
124 E. Vissi et al.
Copyright # 2000 John Wiley & Sons, Ltd. Yeast 2001; 18: 115±124.





























