Embed Size (px)
Citation preview

Journal of Archaeological Science: Reports 2 (2015) 106–113
Contents lists available at ScienceDirect
Journal of Archaeological Science: Reports
j ourna l homepage: ht tp : / /ees.e lsev ie r .com/ jas rep
Fungal spores located in 18th century human dental calculi in the church“La Concepción” (Tenerife, Canary Islands)
José Afonso-Vargas a, Irene La Serna-Ramos b, Matilde Arnay-de-la-Rosa a,⁎a Dpto. de Geografía e Historia, Universidad de La Laguna, 38071 La Laguna, Tenerife, Canary Islands, Spainb Dpto. de Biología Vegetal (Botánica), Universidad de La Laguna, 38071 La Laguna, Tenerife, Canary Islands, Spain
⁎ Corresponding author.E-mail address: [email protected] (M. Arnay-de-la-Ros
http://dx.doi.org/10.1016/j.jasrep.2015.01.0032352-409X/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 31 July 2014Received in revised form 4 January 2015Accepted 4 January 2015Available online xxxx
Keywords:SporesUstilago maydisCornDental calculusPhytolithsPalynomorphsCanary Islands
Wepresent the results of our study of fungal spores found in two samples ofmineralized dental calculi or “tartar”identified during the analysis of plant microfossils (phytoliths and starch granules), taken from individuals fromthe late 18th century, buried in graves in the church “La Concepción” (Santa Cruz de Tenerife). The identificationof palynomorphs motivated the application of a specific methodology to investigate the nature of their presencein the dental tartar, seeking to discover whether this was the result of archaeological sediment contamination orof particles trapped within it. Comparative analysis of the palynomorphs found in the calculi, using referencematerial from the Palynotheque in the Department of Plant Biology, University of La Laguna, and analysis ofthe archaeological sediment, allowedus to confirm that the fungal sporeswere exclusively located inside thema-trix of the calculi and did not originate from a contaminant source. Morphometric study of the spores, referencematerial and bibliographic descriptions allow us to propose that these are spores ofUstilagomaydis (D.C) Corda, aparasitic corn (Zea mays L.) fungus. These results confirm, on the one hand, the historical consumption of corn asopposed to cereals produced locally until that time, such as barley and wheat, and, on the other hand, consump-tion of some shipments of maize contaminated by the so-called “corn smut” (U. maydis).
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
The study of microfossils contained in partially mineralized humandental tartar or dental calculi is one of the sources of direct informationabout palaeodietary aspects (Lalueza Fox and Pérez-Pérez, 1994). Froman archaeological perspective, the study of plant microfossils in dentalcalculus yields information on the diet of prehistoric ungulates(Armitage, 1975), historic ungulates (Middleton, 1990) and primateslinked to the evolutionary chain (Ciochon et al., 1990), finally beingapplied to human populations in different contexts and periods oftime (Scott-Cummings and Magenis, 1997; Juan-Tresserras, 1997).The first microfossils detected in these studies were silicophytolithsand starch granules, which also cause tooth enamel striation (LaluezaFox and Pérez-Pérez, 1994; Lalueza et al., 1996) and both indicate theconsumption of plants or plant products. As for palynomorphs as partof the microfossil record, distinguishing them from those found innon-human calculi (Middleton and Rovner, 1994), the first referenceseems to be that of Torok et al. (1999), in this case in corpses from the18th and 19th centuries. Fungal basidiospores were detected amongmicrofossils such as oxalates and phytoliths, but without referring tospecific taxa. More recently, Blatt et al. (2010) have also identifiedcotton fibre microfossils and other plant microfossils in historic
a).
human dental calculi from Ohio (USA). The history of oral hygieneshows the attention given in the past to the removal of dental tartar,with the use of awide variety of specific instruments that varied inmor-phology and sophistication throughout history (González et al., 2003).
Dental calculus is mineralized bacterial plaque adhered to thesurface of the tooth. Diet affects the formation of tartar, but it is noteasy to establish the processes because of the numerous ways tartarcan be formed. Some authors relate its formation almost exclusively toa protein-rich diet, for example, a diet based on meat products (Lillie,1996; Malgosa and Subirá, 1996), while others link it to starch-richdiets, such as those based on cereals (Hanikara et al., 1994; Eshedet al., 2006; Afonso, 2007).
In terms of its composition, organic and inorganic components can bedistinguished. Among the former there are remains of cells, food, bacteriaand protein components from saliva, while the latter comprise calciumsalts deposited on this matrix (Jin and Ying, 2002; Pérez et al., 2004).The components trapped in the tartar are an important source of informa-tion about the food consumed, because the presence of saliva is necessaryfor this process to take place. Therefore, many researchers suggest thatthis rules out the possibility that many of the components of the calculiwere acquired after death and they defend its validity as a source of directinformation about the products consumed (Afonso, 2007).
The extraction and study of this type of material has required nu-merous methodological reviews (Boyadjian et al., 2007; Afonso, 2007)focussing on the above-mentioned aspects. Unlike other archaeological
107J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
materials, the samples involved must be destroyed to be analysed, inorder to recover as much information as possible. In this case, theinformation takes the form of microfossils such as starch granules andsilica phytoliths, which are not affected by the chemical treatmentsused, as opposed to calcium oxalates (Juan-Tresserras, 1997) whichare completely dissolved. In this regard, we have evidenced the partialalteration of reference spores after chemical treatment, which suppos-edly also occurred in spores found within the dental calculi of thepresent study, but to our knowledge the literature contains no informa-tion on this.
From a historical research perspective, the information provided byplantmicrofossils contained in dental calculi allows a direct approxima-tion of the diet at different periods of time and historical processes(González et al., 2003; Flandrin and Montanari, 2004; Juan-Tresserras,1997).
In the case of the calculi from the church “La Concepción”, the studyprovides insight into dietary patterns of the inhabitants of the CanaryIslands and of Tenerife in particular, during the 18th century, a boomtime for the city of Santa Cruz de Tenerife as a port and commercial cen-tre, connected to the rest of the Canary Islands' capital cities, the Spanishmainland and the Spanish colonies in America. In terms of food, it alsomeant the expansion of habits that gradually changed the diet of the Ca-nary population, probably starting with the upper classes, as suggestedby the differential content of strontium/barium (Arnay et al., 2009)among individuals buried near the altar or far from the altar. As previ-ously reported, tombs near the Altar were destined to individualsbelonging to the high social class (Cioranescu, 1998). Among dietarychange, it is important to highlight the increased consumption of cornor maize (Zea mays) compared to other common grains such as wheator barley. The gradual production of the former from the late 16th cen-tury in islands such asGranCanaria, resulted in exportation to the rest ofthe archipelago (Alzola, 1984). The 18th century was a period of greatsocial crises and famines that led not only to the importation of cerealssuch as corn (or “millo” in the Canary Islands) but also to the prolifera-tion of domestic cultivation and consumption.
The numerous human remains found in the subsoil of the church “LaConcepción” have provided a considerable number of teeth that havebeen subjected to various palaeopathological and anthropologicalanalyses, including the study of tartar as a variable of great interest in re-vealing the diet and oral health conditions of the population of the 18thcentury (Afonso, 2007; Gámez Mendoza, 2004; Arnay et al., 2009).
Early studies of microfossils in these samples confirmed the pres-ence of silicophytoliths and starch granules, as well as palynomorphsin two of them, which were provisionally classified as spores (Afonso,2007). A detailed study revealed that they could be identified taxonom-ically using morphometric and statistical analyses, including that ofreference materials and sediments surrounding the human remainsfrom which the dental calculi came.
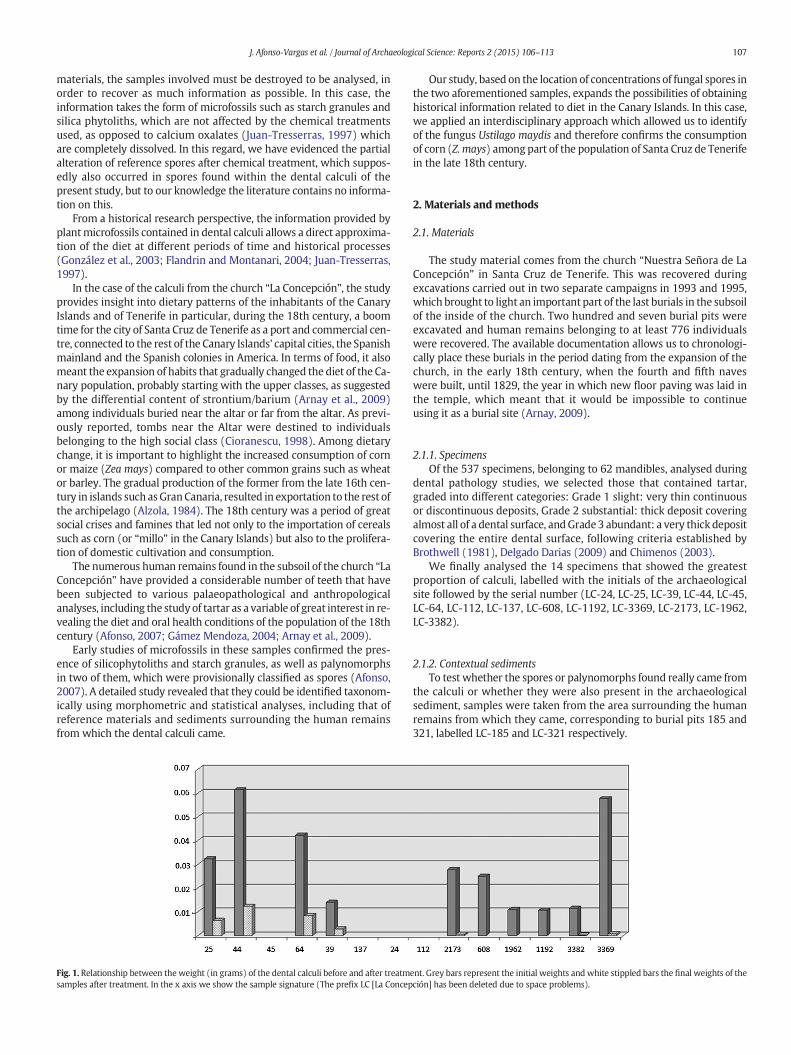
Fig. 1. Relationship between the weight (in grams) of the dental calculi before and after treatmsamples after treatment. In the x axis we show the sample signature (The prefix LC [La Concep
Our study, based on the location of concentrations of fungal spores inthe two aforementioned samples, expands the possibilities of obtaininghistorical information related to diet in the Canary Islands. In this case,we applied an interdisciplinary approach which allowed us to identifyof the fungus Ustilago maydis and therefore confirms the consumptionof corn (Z. mays) among part of the population of Santa Cruz de Tenerifein the late 18th century.
2. Materials and methods
2.1. Materials
The study material comes from the church “Nuestra Señora de LaConcepción” in Santa Cruz de Tenerife. This was recovered duringexcavations carried out in two separate campaigns in 1993 and 1995,which brought to light an important part of the last burials in the subsoilof the inside of the church. Two hundred and seven burial pits wereexcavated and human remains belonging to at least 776 individualswere recovered. The available documentation allows us to chronologi-cally place these burials in the period dating from the expansion of thechurch, in the early 18th century, when the fourth and fifth naveswere built, until 1829, the year in which new floor paving was laid inthe temple, which meant that it would be impossible to continueusing it as a burial site (Arnay, 2009).
2.1.1. SpecimensOf the 537 specimens, belonging to 62 mandibles, analysed during
dental pathology studies, we selected those that contained tartar,graded into different categories: Grade 1 slight: very thin continuousor discontinuous deposits, Grade 2 substantial: thick deposit coveringalmost all of a dental surface, andGrade 3 abundant: a very thick depositcovering the entire dental surface, following criteria established byBrothwell (1981), Delgado Darias (2009) and Chimenos (2003).
We finally analysed the 14 specimens that showed the greatestproportion of calculi, labelled with the initials of the archaeologicalsite followed by the serial number (LC-24, LC-25, LC-39, LC-44, LC-45,LC-64, LC-112, LC-137, LC-608, LC-1192, LC-3369, LC-2173, LC-1962,LC-3382).
2.1.2. Contextual sedimentsTo test whether the spores or palynomorphs found really came from
the calculi or whether they were also present in the archaeologicalsediment, samples were taken from the area surrounding the humanremains from which they came, corresponding to burial pits 185 and321, labelled LC-185 and LC-321 respectively.
ent. Grey bars represent the initial weights and white stippled bars the final weights of theción] has been deleted due to space problems).
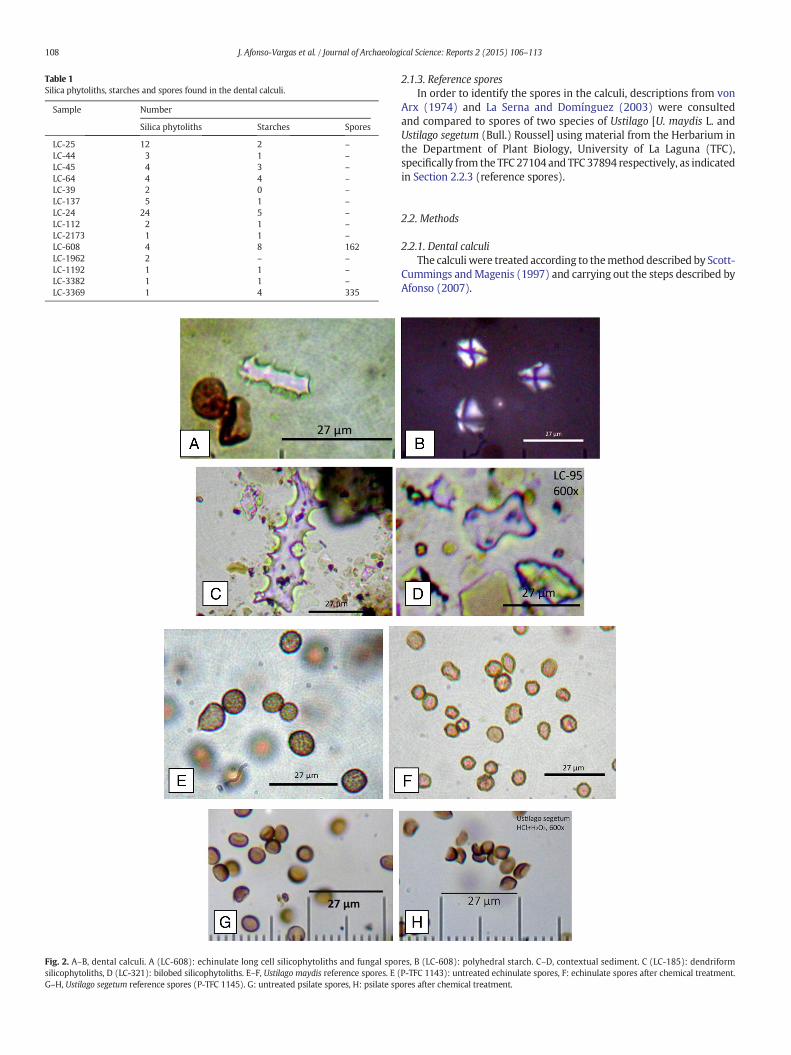
Fig. 2. A–B, dental calculi. A (LC-608): echinulate long cell silicophytoliths and fungal sposilicophytoliths, D (LC-321): bilobed silicophytoliths. E–F, Ustilago maydis reference spores. E (G–H, Ustilago segetum reference spores (P-TFC 1145). G: untreated psilate spores, H: psilate sp
Table 1Silica phytoliths, starches and spores found in the dental calculi.
Sample Number
Silica phytoliths Starches Spores
LC-25 12 2 –
LC-44 3 1 –
LC-45 4 3 –
LC-64 4 4 –
LC-39 2 0 –
LC-137 5 1 –
LC-24 24 5 –
LC-112 2 1 –
LC-2173 1 1 –
LC-608 4 8 162LC-1962 2 – –
LC-1192 1 1 –
LC-3382 1 1 –
LC-3369 1 4 335
108 J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
2.1.3. Reference sporesIn order to identify the spores in the calculi, descriptions from von
Arx (1974) and La Serna and Domínguez (2003) were consultedand compared to spores of two species of Ustilago [U. maydis L. andUstilago segetum (Bull.) Roussel] using material from the Herbarium inthe Department of Plant Biology, University of La Laguna (TFC),specifically from the TFC 27104 and TFC 37894 respectively, as indicatedin Section 2.2.3 (reference spores).
2.2. Methods
2.2.1. Dental calculiThe calculi were treated according to themethod described by Scott-
Cummings andMagenis (1997) and carrying out the steps described byAfonso (2007).
res, B (LC-608): polyhedral starch. C–D, contextual sediment. C (LC-185): dendriformP-TFC 1143): untreated echinulate spores, F: echinulate spores after chemical treatment.ores after chemical treatment.
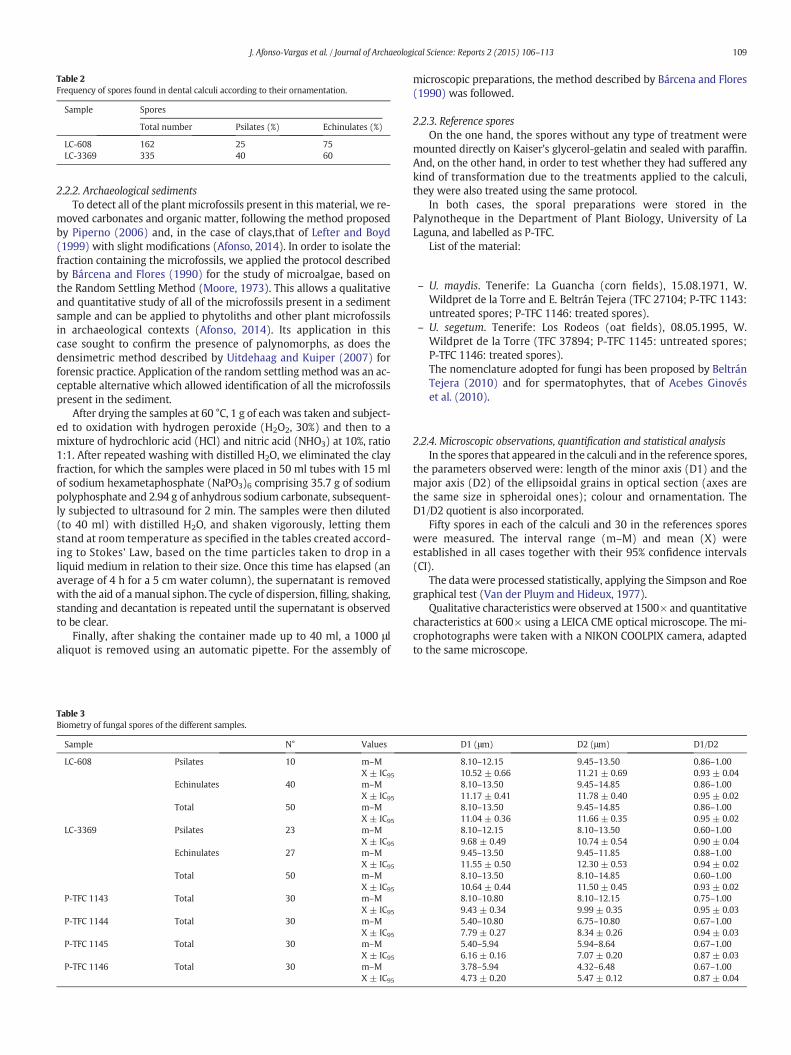
Table 2Frequency of spores found in dental calculi according to their ornamentation.
Sample Spores
Total number Psilates (%) Echinulates (%)
LC-608 162 25 75LC-3369 335 40 60
109J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
2.2.2. Archaeological sedimentsTo detect all of the plant microfossils present in this material, we re-
moved carbonates and organic matter, following the method proposedby Piperno (2006) and, in the case of clays,that of Lefter and Boyd(1999) with slight modifications (Afonso, 2014). In order to isolate thefraction containing the microfossils, we applied the protocol describedby Bárcena and Flores (1990) for the study of microalgae, based onthe Random Settling Method (Moore, 1973). This allows a qualitativeand quantitative study of all of the microfossils present in a sedimentsample and can be applied to phytoliths and other plant microfossilsin archaeological contexts (Afonso, 2014). Its application in thiscase sought to confirm the presence of palynomorphs, as does thedensimetric method described by Uitdehaag and Kuiper (2007) forforensic practice. Application of the random settling method was an ac-ceptable alternative which allowed identification of all the microfossilspresent in the sediment.
After drying the samples at 60 °C, 1 g of each was taken and subject-ed to oxidation with hydrogen peroxide (H2O2, 30%) and then to amixture of hydrochloric acid (HCl) and nitric acid (NHO3) at 10%, ratio1:1. After repeated washing with distilled H2O, we eliminated the clayfraction, for which the samples were placed in 50 ml tubes with 15 mlof sodium hexametaphosphate (NaPO3)6 comprising 35.7 g of sodiumpolyphosphate and 2.94 g of anhydrous sodium carbonate, subsequent-ly subjected to ultrasound for 2 min. The samples were then diluted(to 40 ml) with distilled H2O, and shaken vigorously, letting themstand at room temperature as specified in the tables created accord-ing to Stokes' Law, based on the time particles taken to drop in aliquid medium in relation to their size. Once this time has elapsed (anaverage of 4 h for a 5 cm water column), the supernatant is removedwith the aid of a manual siphon. The cycle of dispersion, filling, shaking,standing and decantation is repeated until the supernatant is observedto be clear.
Finally, after shaking the container made up to 40 ml, a 1000 μlaliquot is removed using an automatic pipette. For the assembly of
Table 3Biometry of fungal spores of the different samples.
Sample N° Values
LC-608 Psilates 10 m–MX ± IC95
Echinulates 40 m–MX ± IC95
Total 50 m–MX ± IC95
LC-3369 Psilates 23 m–MX ± IC95
Echinulates 27 m–MX ± IC95
Total 50 m–MX ± IC95
P-TFC 1143 Total 30 m–MX ± IC95
P-TFC 1144 Total 30 m–MX ± IC95
P-TFC 1145 Total 30 m–MX ± IC95
P-TFC 1146 Total 30 m–MX ± IC95
microscopic preparations, the method described by Bárcena and Flores(1990) was followed.
2.2.3. Reference sporesOn the one hand, the spores without any type of treatment were
mounted directly on Kaiser's glycerol-gelatin and sealed with paraffin.And, on the other hand, in order to test whether they had suffered anykind of transformation due to the treatments applied to the calculi,they were also treated using the same protocol.
In both cases, the sporal preparations were stored in thePalynotheque in the Department of Plant Biology, University of LaLaguna, and labelled as P-TFC.
List of the material:
– U. maydis. Tenerife: La Guancha (corn fields), 15.08.1971, W.Wildpret de la Torre and E. Beltrán Tejera (TFC 27104; P-TFC 1143:untreated spores; P-TFC 1146: treated spores).
– U. segetum. Tenerife: Los Rodeos (oat fields), 08.05.1995, W.Wildpret de la Torre (TFC 37894; P-TFC 1145: untreated spores;P-TFC 1146: treated spores).The nomenclature adopted for fungi has been proposed by BeltránTejera (2010) and for spermatophytes, that of Acebes Ginovéset al. (2010).
2.2.4. Microscopic observations, quantification and statistical analysisIn the spores that appeared in the calculi and in the reference spores,
the parameters observed were: length of the minor axis (D1) and themajor axis (D2) of the ellipsoidal grains in optical section (axes arethe same size in spheroidal ones); colour and ornamentation. TheD1/D2 quotient is also incorporated.
Fifty spores in each of the calculi and 30 in the references sporeswere measured. The interval range (m–M) and mean (X) wereestablished in all cases together with their 95% confidence intervals(CI).
The data were processed statistically, applying the Simpson and Roegraphical test (Van der Pluym and Hideux, 1977).
Qualitative characteristics were observed at 1500× and quantitativecharacteristics at 600× using a LEICA CME optical microscope. The mi-crophotographs were taken with a NIKON COOLPIX camera, adaptedto the same microscope.
D1 (μm) D2 (μm) D1/D2
8.10–12.1510.52 ± 0.66
9.45–13.5011.21 ± 0.69
0.86–1.000.93 ± 0.04
8.10–13.5011.17 ± 0.41
9.45–14.8511.78 ± 0.40
0.86–1.000.95 ± 0.02
8.10–13.5011.04 ± 0.36
9.45–14.8511.66 ± 0.35
0.86–1.000.95 ± 0.02
8.10–12.159.68 ± 0.49
8.10–13.5010.74 ± 0.54
0.60–1.000.90 ± 0.04
9.45–13.5011.55 ± 0.50
9.45–11.8512.30 ± 0.53
0.88–1.000.94 ± 0.02
8.10–13.5010.64 ± 0.44
8.10–14.8511.50 ± 0.45
0.60–1.000.93 ± 0.02
8.10–10.809.43 ± 0.34
8.10–12.159.99 ± 0.35
0.75–1.000.95 ± 0.03
5.40–10.807.79 ± 0.27
6.75–10.808.34 ± 0.26
0.67–1.000.94 ± 0.03
5.40–5.946.16 ± 0.16
5.94–8.647.07 ± 0.20
0.67–1.000.87 ± 0.03
3.78–5.944.73 ± 0.20
4.32–6.485.47 ± 0.12
0.67–1.000.87 ± 0.04
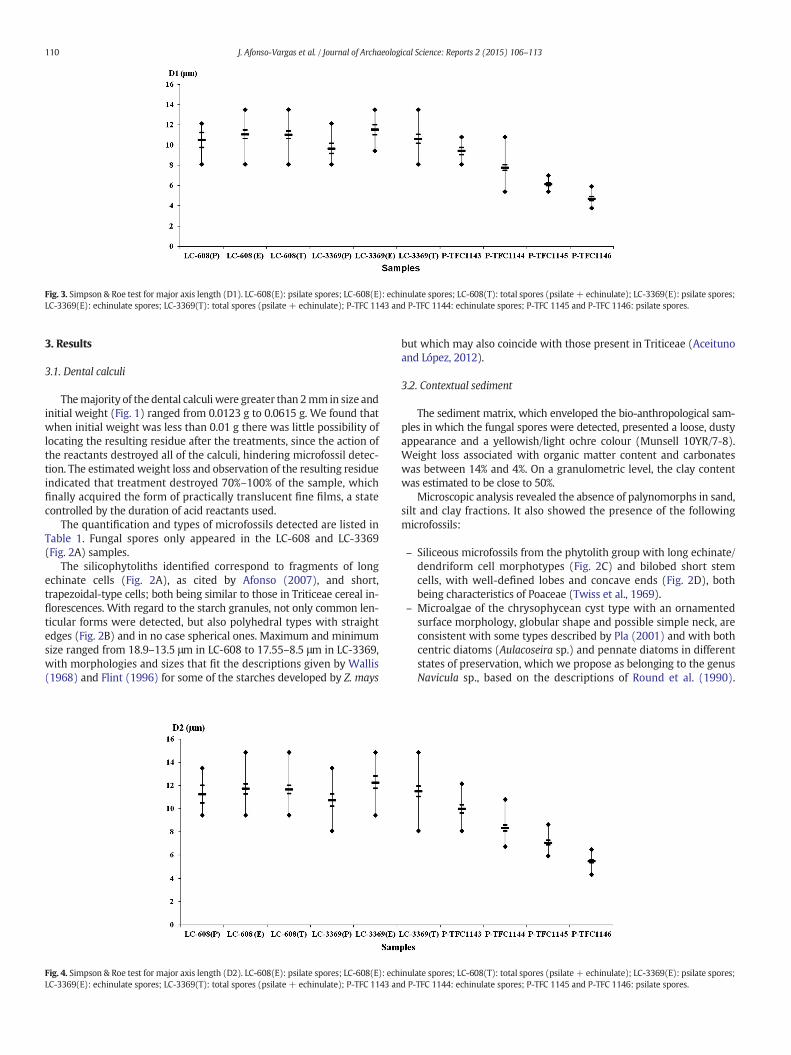
Fig. 3. Simpson & Roe test for major axis length (D1). LC-608(E): psilate spores; LC-608(E): echinulate spores; LC-608(T): total spores (psilate + echinulate); LC-3369(E): psilate spores;LC-3369(E): echinulate spores; LC-3369(T): total spores (psilate + echinulate); P-TFC 1143 and P-TFC 1144: echinulate spores; P-TFC 1145 and P-TFC 1146: psilate spores.
110 J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
3. Results
3.1. Dental calculi
Themajority of the dental calculi were greater than 2mm in size andinitial weight (Fig. 1) ranged from 0.0123 g to 0.0615 g. We found thatwhen initial weight was less than 0.01 g there was little possibility oflocating the resulting residue after the treatments, since the action ofthe reactants destroyed all of the calculi, hindering microfossil detec-tion. The estimated weight loss and observation of the resulting residueindicated that treatment destroyed 70%–100% of the sample, whichfinally acquired the form of practically translucent fine films, a statecontrolled by the duration of acid reactants used.
The quantification and types of microfossils detected are listed inTable 1. Fungal spores only appeared in the LC-608 and LC-3369(Fig. 2A) samples.
The silicophytoliths identified correspond to fragments of longechinate cells (Fig. 2A), as cited by Afonso (2007), and short,trapezoidal-type cells; both being similar to those in Triticeae cereal in-florescences. With regard to the starch granules, not only common len-ticular forms were detected, but also polyhedral types with straightedges (Fig. 2B) and in no case spherical ones. Maximum and minimumsize ranged from 18.9–13.5 μm in LC-608 to 17.55–8.5 μm in LC-3369,with morphologies and sizes that fit the descriptions given by Wallis(1968) and Flint (1996) for some of the starches developed by Z. mays
Fig. 4. Simpson & Roe test for major axis length (D2). LC-608(E): psilate spores; LC-608(E): echLC-3369(E): echinulate spores; LC-3369(T): total spores (psilate + echinulate); P-TFC 1143 an
but which may also coincide with those present in Triticeae (Aceitunoand López, 2012).
3.2. Contextual sediment
The sediment matrix, which enveloped the bio-anthropological sam-ples in which the fungal spores were detected, presented a loose, dustyappearance and a yellowish/light ochre colour (Munsell 10YR/7-8).Weight loss associated with organic matter content and carbonateswas between 14% and 4%. On a granulometric level, the clay contentwas estimated to be close to 50%.
Microscopic analysis revealed the absence of palynomorphs in sand,silt and clay fractions. It also showed the presence of the followingmicrofossils:
– Siliceous microfossils from the phytolith group with long echinate/dendriform cell morphotypes (Fig. 2C) and bilobed short stemcells, with well-defined lobes and concave ends (Fig. 2D), bothbeing characteristics of Poaceae (Twiss et al., 1969).
– Microalgae of the chrysophycean cyst type with an ornamentedsurface morphology, globular shape and possible simple neck, areconsistent with some types described by Pla (2001) and with bothcentric diatoms (Aulacoseira sp.) and pennate diatoms in differentstates of preservation, which we propose as belonging to the genusNavicula sp., based on the descriptions of Round et al. (1990).
inulate spores; LC-608(T): total spores (psilate + echinulate); LC-3369(E): psilate spores;d P-TFC 1144: echinulate spores; P-TFC 1145 and P-TFC 1146: psilate spores.
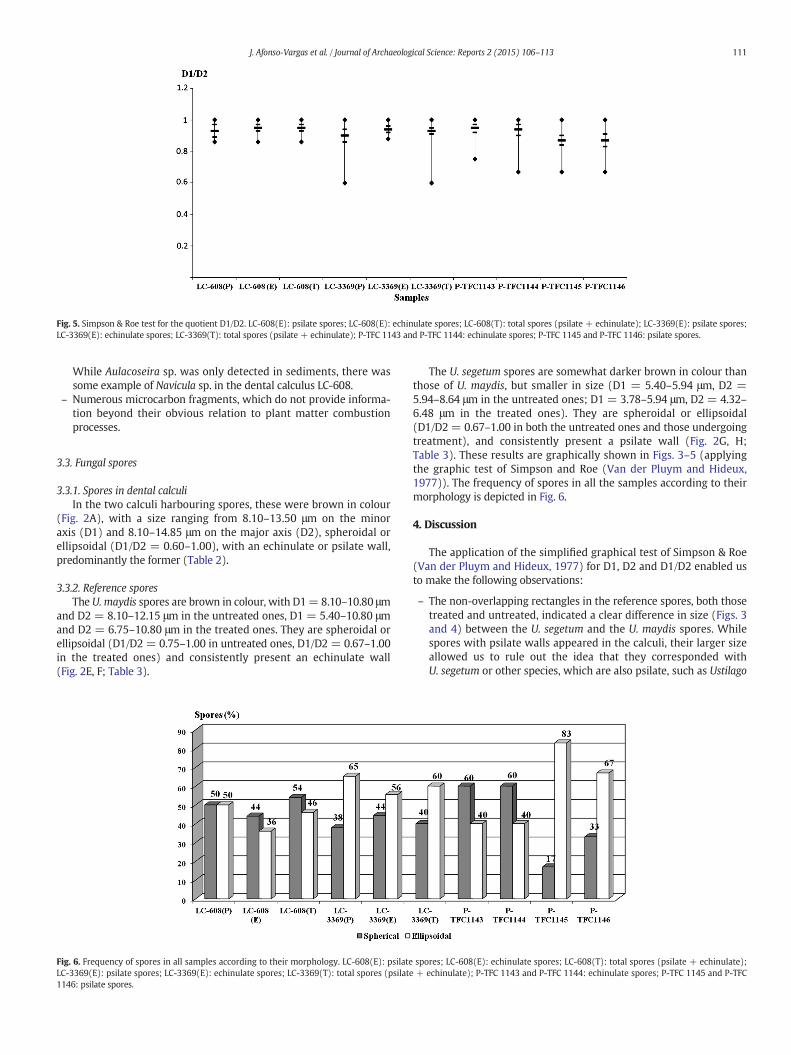
Fig. 5. Simpson & Roe test for the quotient D1/D2. LC-608(E): psilate spores; LC-608(E): echinulate spores; LC-608(T): total spores (psilate + echinulate); LC-3369(E): psilate spores;LC-3369(E): echinulate spores; LC-3369(T): total spores (psilate + echinulate); P-TFC 1143 and P-TFC 1144: echinulate spores; P-TFC 1145 and P-TFC 1146: psilate spores.
111J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
While Aulacoseira sp. was only detected in sediments, there wassome example of Navicula sp. in the dental calculus LC-608.
– Numerous microcarbon fragments, which do not provide informa-tion beyond their obvious relation to plant matter combustionprocesses.
3.3. Fungal spores
3.3.1. Spores in dental calculiIn the two calculi harbouring spores, these were brown in colour
(Fig. 2A), with a size ranging from 8.10–13.50 μm on the minoraxis (D1) and 8.10–14.85 μm on the major axis (D2), spheroidal orellipsoidal (D1/D2 = 0.60–1.00), with an echinulate or psilate wall,predominantly the former (Table 2).
3.3.2. Reference sporesThe U. maydis spores are brown in colour, with D1= 8.10–10.80 μm
and D2 = 8.10–12.15 μm in the untreated ones, D1 = 5.40–10.80 μmand D2 = 6.75–10.80 μm in the treated ones. They are spheroidal orellipsoidal (D1/D2 = 0.75–1.00 in untreated ones, D1/D2 = 0.67–1.00in the treated ones) and consistently present an echinulate wall(Fig. 2E, F; Table 3).
Fig. 6. Frequency of spores in all samples according to their morphology. LC-608(E): psilateLC-3369(E): psilate spores; LC-3369(E): echinulate spores; LC-3369(T): total spores (psilate1146: psilate spores.
The U. segetum spores are somewhat darker brown in colour thanthose of U. maydis, but smaller in size (D1 = 5.40–5.94 μm, D2 =5.94–8.64 μm in the untreated ones; D1 = 3.78–5.94 μm, D2 = 4.32–6.48 μm in the treated ones). They are spheroidal or ellipsoidal(D1/D2 = 0.67–1.00 in both the untreated ones and those undergoingtreatment), and consistently present a psilate wall (Fig. 2G, H;Table 3). These results are graphically shown in Figs. 3–5 (applyingthe graphic test of Simpson and Roe (Van der Pluym and Hideux,1977)). The frequency of spores in all the samples according to theirmorphology is depicted in Fig. 6.
4. Discussion
The application of the simplified graphical test of Simpson & Roe(Van der Pluym and Hideux, 1977) for D1, D2 and D1/D2 enabled usto make the following observations:
– The non-overlapping rectangles in the reference spores, both thosetreated and untreated, indicated a clear difference in size (Figs. 3and 4) between the U. segetum and the U. maydis spores. Whilespores with psilate walls appeared in the calculi, their larger sizeallowed us to rule out the idea that they corresponded withU. segetum or other species, which are also psilate, such as Ustilago
spores; LC-608(E): echinulate spores; LC-608(T): total spores (psilate + echinulate);+ echinulate); P-TFC 1143 and P-TFC 1144: echinulate spores; P-TFC 1145 and P-TFC

112 J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
hordei (a parasite found in barley and other grasses) and accordingto von Arx (1987) measure 5–8 μm.
– Comparing the spores from the calculi with those of U. maydis, wecan say that, although the rectangles are not completely overlapping,the variations in size (Figs. 3 and 4), are minor and therefore boththe echinulate (ornamentation typical of U. maydis) and the psilatespores coincide in size with those of “corn smut”.
– While there were no significant differences in the D1/D2 quotient(Fig. 5) between the calculus spores and the U. maydis andU. segetum ones, in this case, shape was not a determinant in identi-fying these spores. This was also clearly evident in Fig. 6, where, inone of the calculi (LC-608), the spherical spores predominated,whether echinulate or psiladas, as opposed to those of the calculusLC-3369, where they appeared in smaller proportions.
The absence of fungal spores in the contextual sediment and thepresence of microfossils that were not detected in the calculi, rule outthe possibility of contamination.
Furthermore, thesemicrofossils allowed us to confirm that the burialpitswere filledwith soil obtained fromoutside the church. The presenceof diatoms and chrysophyte cysts, indicators of wet conditions likethose of the nearby gully bed “Barranco de Santos” (Afonso, 2004) con-firms that the burial pits were filled with soil from outside the church.These data could also indicate the origin of some of the phytolithsfound, as in the case of short bilobed cells. Until we have more accuratemorphometric studies, we will consider two interpretative approaches,one linked to grasses of the Arundinoideae subfamily, typical in wet en-vironments, such as the edges of the gully bed mentioned above; theother could refer to a Panicoideae grass adapted to drier environments,such asHyparrhenia hirta or “cerrillo”, a very common species in coastaland mid-mountain regions of the Canary Islands (Afonso, 2014).
The presence of echinate long cell phytoliths, common in the inflores-cences of cultivated grasses (Poaceae of the Triticeae tribe), indicated theexistence of an agricultural sedimentary context, either near the churchorlocated at higher altitudes and arriving there by seasonal rainfall runoff.
The presence of microcarbons in the sediment may be related toactivities carried out in the vicinity of the church, such as an “asepticburning” of waste materials, or it could be evidence of previous fires,both nearby and inside the church premises.
Morphometric analysis of the echinulate spores and comparisonwith the reference preparations (P-TFC 1143 and P-TFC 1144) allowedus to determine that they were associated with corn smut (U. maydis).Regarding the psilate spores, aswe do not currently have the herbariummaterial on other species of Ustilago (e.g.: Ustilago crameri, Ustilagocynodontis, U. hordei, Ustilago longissima) and their much larger sizedoes not coincidewith those described in the literature for these speciesor the reference U. segetum studied, we cannot guarantee whether theywere of another Ustilago or U. maydis that had lost their ornamentationthrough taphonomic processes.
On an archaeo-botanical level, several interpretations can be made:
– The phytoliths and starches found in the calculi confirm the con-sumption of cereals, both of the Triticeae family, to which wheatand barley belong, and of the Panicoideae subfamily, to which cornbelongs. Although these phytoliths, characteristic of Triticeae inflo-rescences, are absent in Z. mays, the polyhedral starches are less nu-merous in cereals of thewheat group,which reinforces the existenceof corn smut spores.
– The fungal spores also provide direct information about theconsumption of cereals, particularly of corn (“millo”) by the twoindividuals with spores. These individuals were buried in tombs lo-cated in different parts of the church, at different distances fromthe altar. Therefore, from the location of the burial pits we cannotinfer that they belonged to the privileged social class. The cornthey consumed came from cobs infected with the U. maydis fungus,which has no harmful effects for humans.
While it is believed that by the mid to late 16th century corn wasbeing cultivated in the Canary Islands, it was in the 18th centurywhen its cultivation and consumption as “"gofio” (toasted flour of oneor more grains) became generalized, surpassing that of wheat andbarley, which were already consumed in aboriginal times, Gran Canariabeing the main supplier (Alzola, 1984).
As already indicated, the type of diet is one of the main factorsinvolved in the formation of tartar. There are other factors besides dietthat can intervene in the presence and severity of tartar buildup, suchas oral hygiene, because if the bacterial plaque is removed, mineraliza-tion is avoided (Gámez Mendoza, 2010).
Plaque mineralization is intensified in an alkaline medium and,therefore, in high protein diets, because these diets increase alkalinity(Lieverse, 1999). However, in calculi formation, the oral microorgan-isms that inhabit dental plaque are also involved, which are favouredby diets rich in carbohydrates. Among the carbohydrates, starch hasan important role on the mineralization of plaque. One can thereforeconsider that populations with a diet rich in grains, and therefore instarch, would therefore present a greater buildup of tartar (Littletonand Frohlich, 1993; Lukacs, 1989; Delgado Darias, 2009).
Previous studies carried out on human remains from “LaConcepción” demonstrated the importance of the plant-based diet inthe population studied, despite the proximity to the sea and the possi-bility of consuming seafood. Thus, the average of the values obtainedby analysing tooth decay approaches that of societieswith primarily ag-ricultural economies. Chemical studies of the bone, to determinebariumand strontium content, have also shown the importance of plantproducts in the diet. This diet, fundamentally rich in carbohydrates,was found to be the same among children (Arnay et al., 2009; GámezMendoza, 2010).
All of this is consistent with the historical documentation studied.From the second half of the 18th century onwards, there was a signifi-cant increase in the consumption of plant products, especially cerealssuch as corn and tubers such as potatoes (Gámez Mendoza, 2010).
5. Conclusions
The discovery of these phytopathogenic spores reveals a new line ofpalynological research, at an archaeo-botanical level, in human dentalcalculi, which seeks to extend the analysis of the teeth with low tartarbuildup from this very site and that of others from different sourcesand time periods.
The absence of fungal spores in the contextual sediment and thepresence of microfossils that were not detected in the calculi rule outpossible contamination and allowed us to extract data related to thediet from the 18th century population.
In the present study we detected U. maydis, a parasitic corn fungus,using an interdisciplinary approach that supported the proposedarchaeo-botanical interpretation. The results of the present study areconsistent with the increased consumption of corn on the island ofTenerife from the 18th century onwards, introducing dietary changeswhich subsequently spread to all social classes.
Acknowledgements
This research was conducted as part of the project HAR2011-27413supported by the Ministerio de Economía y Competitividad (Spain).
References
Acebes Ginovés, J.R., León Arencibia, M.C., Rodríguez Navarro, M.L., del Arco Aguilar, M.,García Gallo, A., Pérez de Paz, P.L., Rodríguez Delgado, O., Martín Osorio, V.E.,Wildpret de la Torre, W., 2010. Spermatophyta. In: Arechavaleta, M., Rodríguez, S.,Zurita, N., García, A. (Eds.), Lista de especies silvestres de Canarias. Hongos, plantasy animales terrestres. Gobierno de Canarias, Santa Cruz de Tenerife, pp. 122–172(2009, http://escalera.bio.ucm.es/usuarios/bba/cont/docs/456.pdf).
Aceituno, F.J., López, J.A., 2012. Caracterización morfológica de los almidones de losgéneros Triticum y Hordeum de la Península Ibérica. Trab. Prehist. 69, 332–348.

113J. Afonso-Vargas et al. / Journal of Archaeological Science: Reports 2 (2015) 106–113
Afonso, J.A., 2004. Aportaciones del análisis de fitolitos, almidones y otros referentesmicroscópicos al estudio de la Prehistoria y Arqueología de las Islas Canarias:resultados preliminares. Tabona 9, 69–96 (http://publica.webs.ull.es/publicaciones/detalle/revista-tabona/tabona-prehistoria-y-antropologia/).
Afonso, J.A., 2007. Silicofitolitos y gránulos de almidón en cálculos dentales de antiguaspoblaciones de Tenerife: propuesta para la ampliación del estudio de la dieta yalimentación históricas. Tabona 15, 143–162 (http://publica.webs.ull.es/l).
Afonso, J.A., 2014. Aplicación del análisis de fitolitos y otros microfósiles al estudio deyacimientos, materiales arqueológicos y edáficos de las Islas Canarias. Los ejemplosde Las Cañadas del Teide (Tenerife), La Cerera (Arucas, Gran Canaria) y otras zonasde aplicación experimental. (Tesis Doctoral), Servicio de Publicaciones de laUniversidad de La Laguna, La Laguna (http://riull.ull.es/xmlui/discover?filtertype=author&filter_relational_operator=equals&filter=Afonso+Vargas%2C+Jos%C3%A9+%C3%81nge).
Alzola, J.M., 1984. El millo en Gran Canaria. Colección Viera y Clavijo 10. El Museo Canario,Las Palmas de Gran Canaria (http://www.elmuseocanario.com/index.php/es/publicaciones/coleccion-viera-y-clavijo).
Armitage, P.L., 1975. The extraction and identification of opal phytoliths from the teeth ofungulates. J. Archeol. Sci. 2, 187–197.
Arnay, M., 2009. La Arqueología Histórica en Canarias. El yacimiento sepulcral de la Iglesiade Nuestra Señora de La Concepción de Santa Cruz de Tenerife. Arqueol. Iberoam. 3,21–36.
Arnay, M., González-Reimers, E., Gámez, A., Galindo, L., 2009. The Ba/Sr ratio, carious le-sions, and dental calculus among the population buried in the church La Concepción(Tenerife, Canary Islands). J. Archaeol. Sci. 36, 351–358.
Bárcena,M.A., Flores, J.A., 1990. Ensayo de una técnica para la preparación y cuantificaciónde diatomeas fósiles. In: Civis, J., Flores, J.A. (Eds.), Actas de IV Jornadas dePaleontología. Ed. Universidad de Salamanca, Salamanca, pp. 75–83.
Beltrán Tejera, E., 2010. Fungi (sensu lato). In: Arechavaleta, M., Rodríguez, S., Zurita, N.,García, A. (Eds.), Lista de especies silvestres de Canarias. Hongos, plantas y animalesterrestres. Gobierno de Canarias, Santa Cruz de Tenerife, pp. 25–70 (http://escalera.bio.ucm.es/usuarios/bba/cont/docs/456.pdf).
Blatt, S.H., Redmon, B.G., Cassman, V., Sciulli, P.W., 2010. Dirty Teeth and Ancient Trade:Evidence of Cotton Fibres in Human Dental Calculus From Late Woodland, Ohio.Int. J. Osteoarchaeol. 21, 669–678.
Boyadjian, C.H.C., Eggers, S., Reinhard, K., 2007. Dental wash: a problematic method forextracting microfossils from teeth. J. Archaeol. Sci. 34, 1622–1628.
Brothwell, D., 1981. Desenterrando huesos. La excavación, tratamiento y estudio delesqueleto humano. Fondo Cultura Económica, Madrid.
Chimenos, E., 2003. Perspectiva odontoestomatológica en paleopatología. In: Isidro, A.,Malgosa, A. (Eds.), Paleopatología. La enfermedad no escrita. Masson, Barcelona,pp. 151–162.
Ciochon, R.L., Piperno, D., Thompson, R., 1990. Opal phytoliths found on the teeth of theextinct ape Gigantopithecus blacki: implications for paleodietary studies. Proc. Natl.Acad. Sci. 87, 8120–8124.
Cioranescu, A., 1998. Historia de Santa Cruz. Servicio de Publicaciones de la Caja deAhorros de Canarias, Santa Cruz de Tenerife.
Delgado Darias, T., 2009. La Historia en los dientes. Una aproximación a la prehistoria deGran Canaria desde la Antropología DentalCuadernos de Patrimonio Histórico 8.Gobierno de Canarias, Las Palmas de Gran Canaria (http://mdc.ulpgc.es/cgi-bin/showfile.exe?CISOROOT=/MDC&CISOPTR=136872&filename=174316.pdf).
Eshed, V., Gopher, A., Hershkovitz, I., 2006. Tooth wear and dental pathology at the adventof agriculture: new evidence from the Levant. Am. J. Phys. Anthropol. 130, 145–159.
Flandrin, J.L., Montanari, M., 2004. Historia de la Alimentación. Ediciones Trea, Gijón.Flint, O., 1996. Microscopía de los Alimentos. Acribia, Zaragoza.Gámez Mendoza, A., 2004. Las investigaciones bioarqueológicas para ámbitos históricos
en Canarias. La Iglesia de Nuestra Señora de La Concepción de Santa Cruz de Tenerifecomo ejemplo. Tabona 13, 279–299.
Gámez Mendoza, A., 2010. Estudios bioantropológicos de una población arqueológicahistórica de las Islas Canarias. La Iglesia de La Concepción de Santa Cruz de Tenerife.(Tesis Doctoral), Facultad de Geografía e Historia. Universidad de La Laguna, LaLaguna (http://publica.webs.ull.es/).
González, J., Ucha, M., González, J.J., 2003. Historia General de la Higiene Bucodentaria.Yeltes Soluciones Gráficas, Madrid.
Hanikara, T., Ishida, H., Ohshima, N., Kondo, O., Masuda, T., 1994. Dental calculus andother dental disease in a human skeleton of the Okhotsk culture unearthed alHamanaka-2 site, Rebun-Island, Hokkaido, Japan. Int. J. Osteoarchaeol. 4, 343–351.
Jin, Y., Ying, H., 2002. Supragingival calculus: formation and control. Crit. Rev. Oral Biol.Med. 13, 426–441.
Juan-Tresserras, J., 1997. Procesado y preparación de Alimentos vegetales para consumohumano. Aportaciones del estudio de fitolitos, almidones y lípidos en yacimientosarqueológicos prehistóricos y protohistóricos del cuadrante NE de la PenínsulaIbérica. (Tesis Doctoral). Universidad de Barcelona, Barcelona.
La Serna, I., Domínguez, M.D., 2003. Pólenes y esporas aerovagantes en Canarias:incidencia en alergias, Serie Botánica/1, Materiales Didácticos Universitarios, Serviciode Publicaciones. Universidad de La Laguna, La Laguna (http://www.ull.es/view/departamentos/biovegetal/2010/es).
Lalueza Fox, C., Pérez-Pérez, A., 1994. Dietary information through the examination ofplant phytoliths on the enamel surface of human dentition. J. Archaeol. Sci. 21, 29–34.
Lalueza, C., Juan-Tresserras, J., Albert, R.M., 1996. Phytolith analysis on dental calculus,enamel surface and burial soil: information about diet and paleoenvironment. Am.J. Phys. Anthropol. 101, 101–113.
Lefter, C.J., Boyd, W.E., 1999. An assessment of techniques for the deflocculation and re-moval of clays from sediments used in phytoliths analysis. J. Archaeol. Sci. 26, 31–44.
Lieverse, A., 1999. Diet and aetiology of dental calculus. Int. J. Osteoarchaeol. 9, 219–232.Lillie, M.C., 1996.Mesolithic and Neolithic populations of Ukraine: indications of diet from
dental pathology. Curr. Anthropol. 37, 135–142.Littleton, J., Frohlich, B., 1993. Fish eaters and farmers: dental pathology in the Arabian
Gulf. Am. J. Phys. Anthropol. 92, 427–447.Lukacs, J.R., 1989. Dental paleopathology: methods for reconstructing dietary patterns. In:
Iscan, M.Y., Kennedy, K.A. (Eds.), Reconstruction of Life From the Skeleton. Alan R.Liss, Inc., New York, pp. 261–286.
Malgosa, A., Subirá, M.E., 1996. Antropología i dieta: metodologies per a la reconstrucciónde alimentación de poblacions antigues. Cota Zero 12, 15–27.
Middleton, W., 1990. An improved method for extraction of opal phytoliths from tartarresidues on herbivore teeth. Phytolitharien 6, 2–6.
Middleton, W., Rovner, I., 1994. Extraction of opal phytoliths from herbivore dental calcu-lus. J. Archaeol. Sci. 21, 469–473.
Moore, T.C., 1973. Method of randomly distributing grain for microscopic examination.J. Sediment. Petrol. 43, 904–906.
Pérez, C., Sánchez, H., Barrea, R., Grenón, M., Abraham, J., 2004. Microscopic X-ray fluores-cence analysis of human dental calculus using syncroton radiation. J. Anal. At.Spectrom. 19, 392–397.
Piperno, D., 2006. Phytoliths, A Comprehensive Guide for Archaeologists and ecologists.Altamira Press, Lanham, MD (USA).
Pla, S., 2001. Chrysophycean cysts From the Pyrennes. Biblioteca Phycologica, Band 109J.Cramer, Sttutgart.
Round, F.E., Crawford, R.M., Mann, D.G., 1990. The Diatoms, Biology & Morphology of theGenera. Cambridge University Press, Cambridge, (UK).
Scott-Cummings, L., Magenis, A., 1997. A phytolith and starch record of food and grit inMayan human tooth tartar. Estado Actual de los estudios de fitolitos en suelos y plantas.Monografías 4. Centro de Ciencias Medioambientales, CSIC, Madrid, pp. 211–218.
Torok, K., Pap, I., Jozsa, L., 1999. Századi múmiák fogköveinek mikroszkópos vizsgálata.Fogorv. Sz. 92, 317–327.
Twiss, P., Suess, E., Smith, R., 1969. Morphological classification of grass phytoliths. SoilSci. Soc. Am. Proc. 33, 321–322.
Uitdehaag, S.C.A., Kuiper, I., 2007. Amethod for the combined extraction of pollen, diatomsand phytoliths from forensic soil samples. 2nd Soil Forensics Conference, Edinburgh(http://www.soilforensicsinternational.org/sites/www.soilforensicsinternational.org/files/documents/outputs/Session%205%20Biological%20and%20Chemical%20Analytical%20Diagnostics/SUitdehaag%20poster.pdf).
Van der Pluym, A., Hideux, M., 1977. Application d'une méthodologie quantitative à lapalynologie d'Eryngium maritimum (Umbelliferae). Plant Syst. Evol. 127, 55–85.
von Arx, J.A., 1974. The genera of fungi. Sporulating in Pure Culture. J. Cramer (Ed),Valduz.
von Arx, J.A., 1987. Plan Pathogenic Fungi. Gebruder Borntraeger Verlagsbuchhandlung,Suttgart, Germany.
Wallis, T.E., 1968. Microscopía Analítica. Acribia, Zaragoza.





























