Embed Size (px)
Citation preview

Genomic Priming of the Antisecretory Response toEstrogen in Rat Distal Colon throughout theEstrous Cycle
Fiona O’Mahony, Rodrigo Alzamora, Ho-Lam Chung, Warren Thomas,and Brian J. Harvey
Department of Molecular Medicine, Royal College of Surgeons in Ireland, Beaumont Hospital,Dublin 9, Ireland
The secretion of Cl� across distal colonic crypt cells provides the driving force for the movement offluid into the luminal space. 17�-Estradiol (E2) produces a rapid and sustained reduction in secretionin females, which is dependent on the novel protein kinase C� (PKC�) isozyme and PKA isoform Itargeting of KCNQ1 channels. This sexual dimorphism in the E2 response is associated with a higherexpression level of PKC� in female compared with the male tissue. The present study revealed theantisecretory response is regulated throughout the female reproductive (estrous) cycle and is primedby genomic regulation of the kinases. E2 (1–10 nM) decreased cAMP-dependent secretion in colonicepithelia during the estrus, metestrus, and diestrus stages. A weak inhibition of secretion was dem-onstrated in the proestrus stage. The expression levels of PKC� and PKA fluctuated throughout theestrous cycle and correlated with the potency of the antisecretory effect of E2. The expression of PKC�
and PKA were up-regulated by estrogen at a transcriptional level via a PKC�-MAPK-cAMP responseelement-binding protein-regulated pathway indicating a genomic priming of the antisecretory re-sponse. PKC� was activated by the membrane-impermeant E2-BSA, and this response was inhibited bythe estrogen receptor antagonist ICI 182,780. The 66-kDa estrogen receptor-� isoform was present atthe plasma membrane of female colonic crypt cells with a lower abundance found in male coloniccrypts. The study demonstrates estrogen regulation of intestinal secretion both at a rapid and tran-scriptional level, demonstrating an interdependent relationship between both nongenomic andgenomic hormone responses. (Molecular Endocrinology 23: 1885–1899, 2009)
The absorption and secretion of salt and water is amajor physiological role of the large intestine. Estro-
gen is known to be a salt-retaining steroid hormone inmany tissues. Clinically, salt and water retention is ob-served during high estrogenic states (1). The syntheticderivative of estradiol, ethinylestradiol, which is found inoral contraceptives, hormone replacement therapy, andthe morning after pill, has been demonstrated to causeretention of salt and water in the female body (2). Formany years, it has been recognized that cyclical changes inthe levels of ovarian hormone secretions effect whole-body water and electrolyte homeostasis (3, 4). The kid-neys, lungs, and large intestine are the major regulators of
whole-body water and salt balance. Previously, these or-gans were not recognized as being targeted by circulatingsex steroids. It is now known that estrogen can influenceNa� retention in all three tissues (5–7). Certain phases ofthe reproductive cycle have been implicated in influencingintestinal physiology in three ways: function, cell prolif-eration/profile, and electrolyte and fluid movement. Inaddition, differences in the colonic transit time of the fecesbetween phases of the menstrual cycle have been docu-mented and in particular a slowing in the luteal phase dueto less fluid content (8, 9). The luteal phase of the men-strual cycle occurs after ovulation lasting from 10–16 dand is referred to as the window of implantation. The
ISSN Print 0888-8809 ISSN Online 1944-9917Printed in U.S.A.Copyright © 2009 by The Endocrine Societydoi: 10.1210/me.2008-0248 Received July 22, 2008. Accepted August 18, 2009.First Published Online October 21, 2009
Abbreviations: CRE, cAMP response element; CREB, CRE-binding protein; DAPI, 4�,6-diamidino-2-phenylindole; E2, 17�-estradiol; ER�, estrogen receptor-�; MEK, MAPK ki-nase; PKACI, protein kinase A catalytic subunit isoform I; PKC�, protein kinase C�; qRT,quantitative real-time; WGA, wheat germ agglutinin.
O R I G I N A L R E S E A R C H
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1885
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.

estrogen peak occurs previous to the luteal phase duringovulation. The uterus continues to expand throughoutthis phase, which involves retention of bodily fluid (10)and fluid expansion in the endometrium (11). Given theimpact that high-estrogen states has on whole-body fluidvolume and uterine swelling, it is surprising so little isknown of the molecular mechanisms for estrogen effectson fluid and electrolyte transport in the body and, inparticular, the intestine. It is possible that the regulationof Cl� secretion by estrogen in the distal colon and thecomponents of the molecular mechanism involved mayvary depending on the hormonal background of thefemale.
Cl� secretion in the distal colon can be up-regulated bytwo distinct mechanisms: an increase in intracellularCa2� (12) or cAMP activity (13). Cl� enters the cell acrossthe basolateral membrane by cotransport with K� andNa�. Cl� is then secreted via CFTR channels at the apicalmembrane and other Cl� channels. A basolateral K� con-ductance maintains the favorable negative membrane po-tential to drive Cl� electrodiffusion into the lumen. Themajor K� channels providing the route for K� recyclingare cAMP-dependent KCNQ1 channels and Ca2�-acti-vated KCNN4 channels. In human colonic tissue, it hasbeen demonstrated that blocking basolateral K� activitydecreases forskolin-induced Cl� secretion (14). Similarly,pretreatment of colonic tissue with 17�-estradiol (E2)(0.1–100 nM) significantly reduced forskolin-stimulatedCl� secretion (7). We have recently demonstrated that theE2 inhibition of secretion in rat distal colonic epithelia isfemale sex specific (15). The mechanism was shown to in-volve protein kinase C� (PKC�) and PKA isoform I-depen-dent modulation of KCNQ1 channel activity.
The genomic action of estrogen involves binding of thehormone to the cytosolic estrogen receptor-� (ER�) fol-lowed by translocation of this complex to the nucleuswhereupon it binds the estrogen response element induc-ing transcription. It is known that rapid actions of E2 mayprime gene transcription via the activation of second mes-sengers, which in turn target transcription factors. ThecAMP response element (CRE)-binding protein (CREB)was found in many tissues to be phosphorylated in re-sponse to PKA and MAPK activation by estrogen (16).Activated CREB binds to the CRE found in the promoterregion of many genes. The outcome of CREB activationand binding is an increase/decrease in the transcription ofcertain genes. Upon binding to the CRE, the CREB-bind-ing protein is recruited and coactivates CREB. Binding ofthe CREB-binding protein to the CREB-DNA complex isenhanced by further binding of the steroid coactivatorreceptor 1 (SRC1) in the case of steroid receptor-depen-dent transcription (17). Steroid receptor-independent ac-
tivation of this transcription factor occurs in response toa wide variety of extracellular signals such as growth hor-mones, steroid hormones, and neurotransmitters. RapidCREB phosphorylation by estrogen independent of theclassical receptor has previously been reported (18). Prim-ing of the genomic response during the rapid nongenomicphase of action of steroid hormones and transcriptionalregulation of the nongenomic responses are hot topics inendocrinology. Cross talk between the rapid nongenomicand genomic actions of estrogen occurring via membraneER�, protein kinases, and modulation of the CREB tran-scription factor was investigated in this paper.
Results
The potency of E2 inhibition of cAMP-dependentCl� secretion in female rat colonic epithelia variesthroughout the estrous cycle
We have previously demonstrated that E2 inhibitscAMP-dependent Cl� secretion via a PKC�/PKA signal-ing pathway (15). Basolateral addition of heat-stable en-terotoxin (STa) (E.Coli, potent activator of cAMP-depen-dent secretion) (1 �g/ml) induced an instantaneous andsustained increase (100%) in short circuit current (ISC).E2 (10 nM) reduced the STa effect on ISC with maximalinhibition of secretion observed at estrus, metestrus, anddiestrus (percent secretion: estrus, 57 � 4%; metestrus,60 � 3%; diestrus, 57 � 4%; n � 5; P � 0.001; Fig. 1A).The E2 effect on secretion at proestrus was insignificantand comparable to that of the male tissue (percent secre-tion: proestrus, 87 � 5%; male, 101 � 7%; Fig. 1A).Inhibition of secretion in proestrus became significant atsupraphysiological levels of E2 (percent secretion: 100 nM
E2, 70 � 3%; n � 5; P � 0.001). No effects upon secre-tion were noted with the addition of the estrogen vehiclecarrier MeOH (data not shown). These results demon-strate the antisecretory effect of E2 is estrous cycledependent.
Estrous cycle differences in PKA activation inresponse to E2 in female rat distal colonic crypts
Isolated female rat distal colonic crypts from each stage ofthe cycle were exposed to E2 (10 nM) or equivalent vehicle(0.01%) for the duration of 2, 5, and 15 min. Total lysateswere prepared and analyzed using the PepTag assay forthe nonradioactive detection of cAMP-dependent pro-tein. Differences were recorded as fold values of treatedover control.
PKA was activated in female distal colonic crypts at theestrus stage in response to E2 at 5 min (2.8 � 0.4-foldhigher, n � 5; P � 0.001; Fig. 1B). In contrast, PKA wasnot activated at the proestrus stage in response to E2 (n �
1886 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
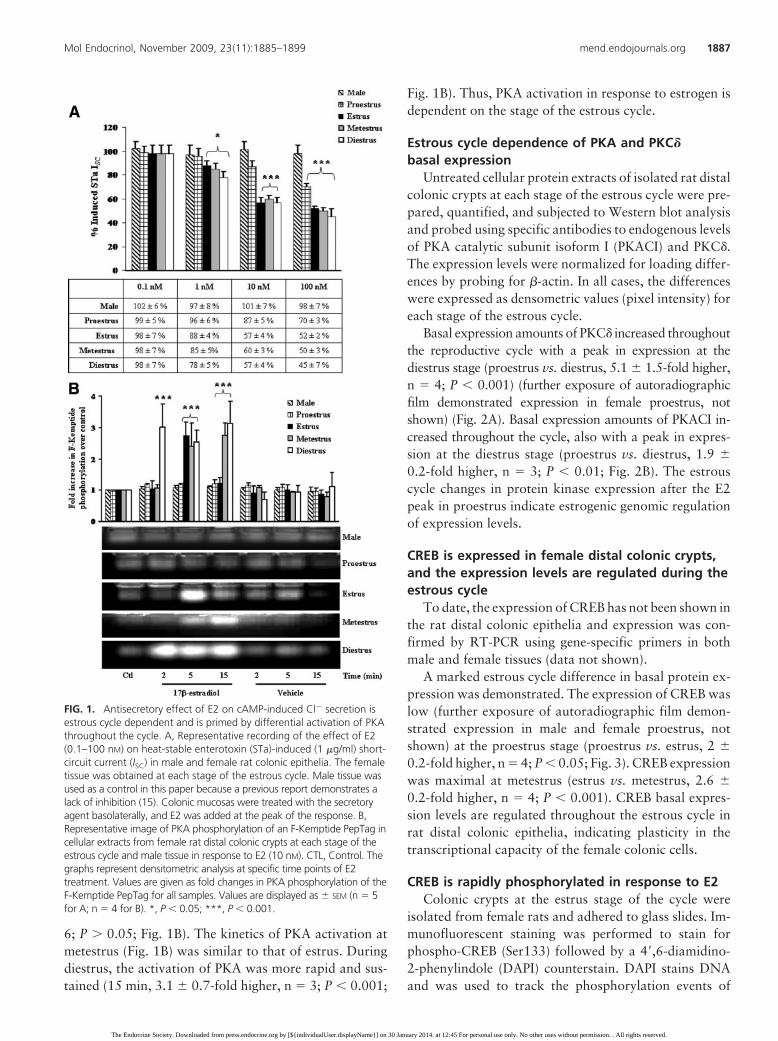
6; P � 0.05; Fig. 1B). The kinetics of PKA activation atmetestrus (Fig. 1B) was similar to that of estrus. Duringdiestrus, the activation of PKA was more rapid and sus-tained (15 min, 3.1 � 0.7-fold higher, n � 3; P � 0.001;
Fig. 1B). Thus, PKA activation in response to estrogen isdependent on the stage of the estrous cycle.
Estrous cycle dependence of PKA and PKC�
basal expressionUntreated cellular protein extracts of isolated rat distal
colonic crypts at each stage of the estrous cycle were pre-pared, quantified, and subjected to Western blot analysisand probed using specific antibodies to endogenous levelsof PKA catalytic subunit isoform I (PKACI) and PKC�.The expression levels were normalized for loading differ-ences by probing for �-actin. In all cases, the differenceswere expressed as densometric values (pixel intensity) foreach stage of the estrous cycle.
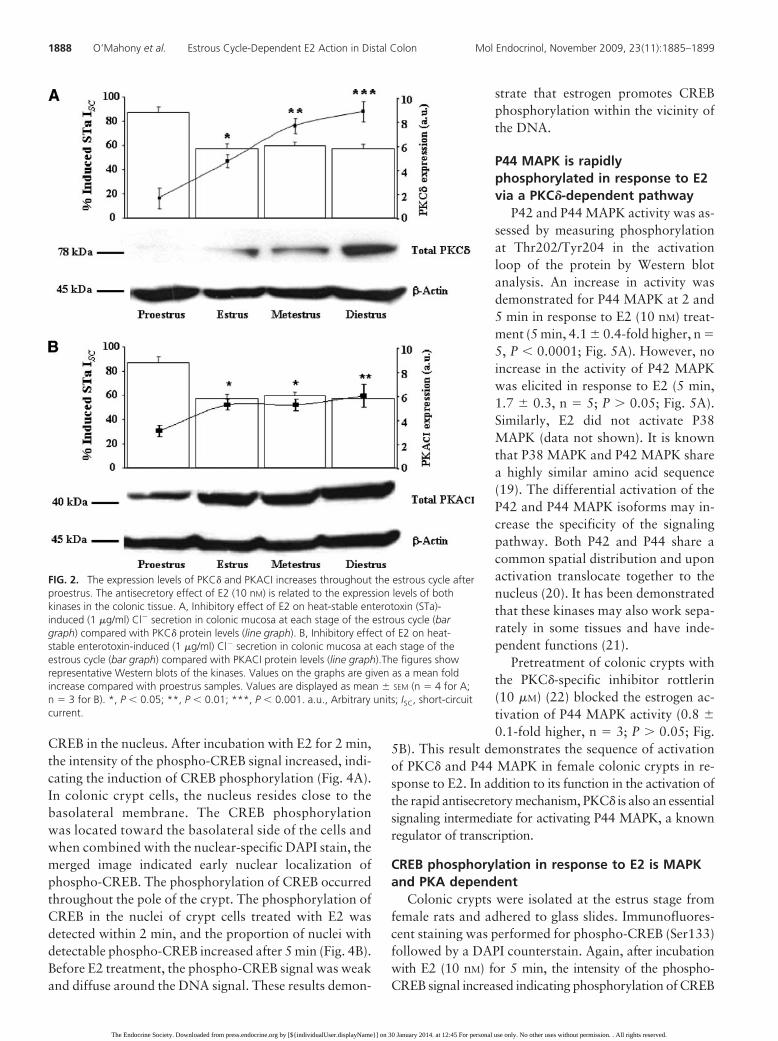
Basal expression amounts of PKC� increased throughoutthe reproductive cycle with a peak in expression at thediestrus stage (proestrus vs. diestrus, 5.1 � 1.5-fold higher,n � 4; P � 0.001) (further exposure of autoradiographicfilm demonstrated expression in female proestrus, notshown) (Fig. 2A). Basal expression amounts of PKACI in-creased throughout the cycle, also with a peak in expres-sion at the diestrus stage (proestrus vs. diestrus, 1.9 �0.2-fold higher, n � 3; P � 0.01; Fig. 2B). The estrouscycle changes in protein kinase expression after the E2peak in proestrus indicate estrogenic genomic regulationof expression levels.
CREB is expressed in female distal colonic crypts,and the expression levels are regulated during theestrous cycle
To date, the expression of CREB has not been shown inthe rat distal colonic epithelia and expression was con-firmed by RT-PCR using gene-specific primers in bothmale and female tissues (data not shown).
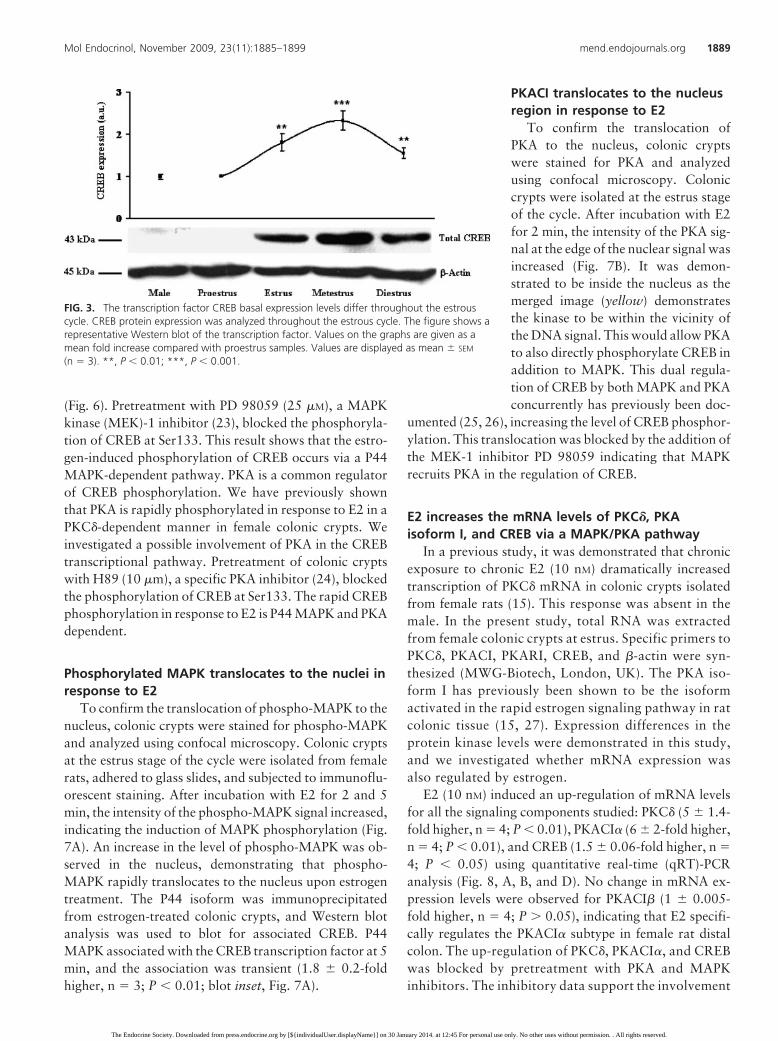
A marked estrous cycle difference in basal protein ex-pression was demonstrated. The expression of CREB waslow (further exposure of autoradiographic film demon-strated expression in male and female proestrus, notshown) at the proestrus stage (proestrus vs. estrus, 2 �0.2-fold higher, n � 4; P � 0.05; Fig. 3). CREB expressionwas maximal at metestrus (estrus vs. metestrus, 2.6 �0.2-fold higher, n � 4; P � 0.001). CREB basal expres-sion levels are regulated throughout the estrous cycle inrat distal colonic epithelia, indicating plasticity in thetranscriptional capacity of the female colonic cells.
CREB is rapidly phosphorylated in response to E2Colonic crypts at the estrus stage of the cycle were
isolated from female rats and adhered to glass slides. Im-munofluorescent staining was performed to stain forphospho-CREB (Ser133) followed by a 4�,6-diamidino-2-phenylindole (DAPI) counterstain. DAPI stains DNAand was used to track the phosphorylation events of
FIG. 1. Antisecretory effect of E2 on cAMP-induced Cl� secretion isestrous cycle dependent and is primed by differential activation of PKAthroughout the cycle. A, Representative recording of the effect of E2(0.1–100 nM) on heat-stable enterotoxin (STa)-induced (1 �g/ml) short-circuit current (ISC) in male and female rat colonic epithelia. The femaletissue was obtained at each stage of the estrous cycle. Male tissue wasused as a control in this paper because a previous report demonstrates alack of inhibition (15). Colonic mucosas were treated with the secretoryagent basolaterally, and E2 was added at the peak of the response. B,Representative image of PKA phosphorylation of an F-Kemptide PepTag incellular extracts from female rat distal colonic crypts at each stage of theestrous cycle and male tissue in response to E2 (10 nM). CTL, Control. Thegraphs represent densitometric analysis at specific time points of E2treatment. Values are given as fold changes in PKA phosphorylation of theF-Kemptide PepTag for all samples. Values are displayed as � SEM (n � 5for A; n � 4 for B). *, P � 0.05; ***, P � 0.001.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1887
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
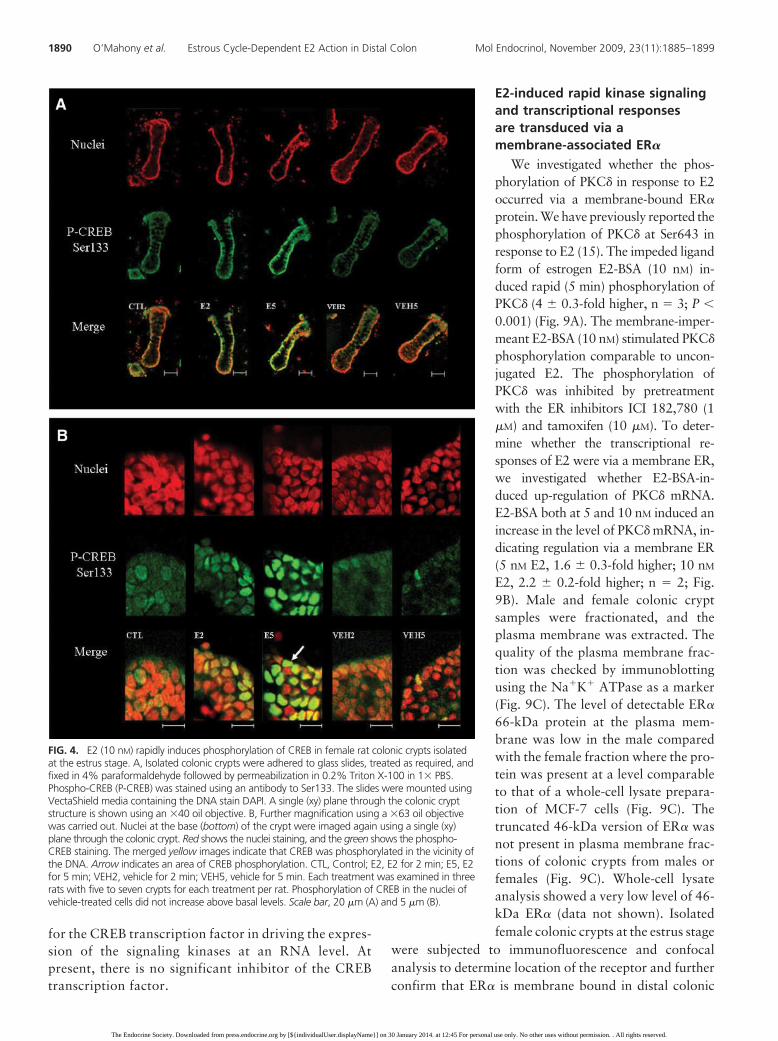
CREB in the nucleus. After incubation with E2 for 2 min,the intensity of the phospho-CREB signal increased, indi-cating the induction of CREB phosphorylation (Fig. 4A).In colonic crypt cells, the nucleus resides close to thebasolateral membrane. The CREB phosphorylationwas located toward the basolateral side of the cells andwhen combined with the nuclear-specific DAPI stain, themerged image indicated early nuclear localization ofphospho-CREB. The phosphorylation of CREB occurredthroughout the pole of the crypt. The phosphorylation ofCREB in the nuclei of crypt cells treated with E2 wasdetected within 2 min, and the proportion of nuclei withdetectable phospho-CREB increased after 5 min (Fig. 4B).Before E2 treatment, the phospho-CREB signal was weakand diffuse around the DNA signal. These results demon-
strate that estrogen promotes CREBphosphorylation within the vicinity ofthe DNA.
P44 MAPK is rapidlyphosphorylated in response to E2via a PKC�-dependent pathway
P42 and P44 MAPK activity was as-sessed by measuring phosphorylationat Thr202/Tyr204 in the activationloop of the protein by Western blotanalysis. An increase in activity wasdemonstrated for P44 MAPK at 2 and5 min in response to E2 (10 nM) treat-ment (5 min, 4.1 � 0.4-fold higher, n �5, P � 0.0001; Fig. 5A). However, noincrease in the activity of P42 MAPKwas elicited in response to E2 (5 min,1.7 � 0.3, n � 5; P � 0.05; Fig. 5A).Similarly, E2 did not activate P38MAPK (data not shown). It is knownthat P38 MAPK and P42 MAPK sharea highly similar amino acid sequence(19). The differential activation of theP42 and P44 MAPK isoforms may in-crease the specificity of the signalingpathway. Both P42 and P44 share acommon spatial distribution and uponactivation translocate together to thenucleus (20). It has been demonstratedthat these kinases may also work sepa-rately in some tissues and have inde-pendent functions (21).
Pretreatment of colonic crypts withthe PKC�-specific inhibitor rottlerin(10 �M) (22) blocked the estrogen ac-tivation of P44 MAPK activity (0.8 �0.1-fold higher, n � 3; P � 0.05; Fig.
5B). This result demonstrates the sequence of activationof PKC� and P44 MAPK in female colonic crypts in re-sponse to E2. In addition to its function in the activation ofthe rapid antisecretory mechanism, PKC� is also an essentialsignaling intermediate for activating P44 MAPK, a knownregulator of transcription.
CREB phosphorylation in response to E2 is MAPKand PKA dependent
Colonic crypts were isolated at the estrus stage fromfemale rats and adhered to glass slides. Immunofluores-cent staining was performed for phospho-CREB (Ser133)followed by a DAPI counterstain. Again, after incubationwith E2 (10 nM) for 5 min, the intensity of the phospho-CREB signal increased indicating phosphorylation of CREB
FIG. 2. The expression levels of PKC� and PKACI increases throughout the estrous cycle afterproestrus. The antisecretory effect of E2 (10 nM) is related to the expression levels of bothkinases in the colonic tissue. A, Inhibitory effect of E2 on heat-stable enterotoxin (STa)-induced (1 �g/ml) Cl� secretion in colonic mucosa at each stage of the estrous cycle (bargraph) compared with PKC� protein levels (line graph). B, Inhibitory effect of E2 on heat-stable enterotoxin-induced (1 �g/ml) Cl� secretion in colonic mucosa at each stage of theestrous cycle (bar graph) compared with PKACI protein levels (line graph).The figures showrepresentative Western blots of the kinases. Values on the graphs are given as a mean foldincrease compared with proestrus samples. Values are displayed as mean � SEM (n � 4 for A;n � 3 for B). *, P � 0.05; **, P � 0.01; ***, P � 0.001. a.u., Arbitrary units; ISC, short-circuitcurrent.
1888 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
(Fig. 6). Pretreatment with PD 98059 (25 �M), a MAPKkinase (MEK)-1 inhibitor (23), blocked the phosphoryla-tion of CREB at Ser133. This result shows that the estro-gen-induced phosphorylation of CREB occurs via a P44MAPK-dependent pathway. PKA is a common regulatorof CREB phosphorylation. We have previously shownthat PKA is rapidly phosphorylated in response to E2 in aPKC�-dependent manner in female colonic crypts. Weinvestigated a possible involvement of PKA in the CREBtranscriptional pathway. Pretreatment of colonic cryptswith H89 (10 �m), a specific PKA inhibitor (24), blockedthe phosphorylation of CREB at Ser133. The rapid CREBphosphorylation in response to E2 is P44 MAPK and PKAdependent.
Phosphorylated MAPK translocates to the nuclei inresponse to E2
To confirm the translocation of phospho-MAPK to thenucleus, colonic crypts were stained for phospho-MAPKand analyzed using confocal microscopy. Colonic cryptsat the estrus stage of the cycle were isolated from femalerats, adhered to glass slides, and subjected to immunoflu-orescent staining. After incubation with E2 for 2 and 5min, the intensity of the phospho-MAPK signal increased,indicating the induction of MAPK phosphorylation (Fig.7A). An increase in the level of phospho-MAPK was ob-served in the nucleus, demonstrating that phospho-MAPK rapidly translocates to the nucleus upon estrogentreatment. The P44 isoform was immunoprecipitatedfrom estrogen-treated colonic crypts, and Western blotanalysis was used to blot for associated CREB. P44MAPK associated with the CREB transcription factor at 5min, and the association was transient (1.8 � 0.2-foldhigher, n � 3; P � 0.01; blot inset, Fig. 7A).
PKACI translocates to the nucleusregion in response to E2
To confirm the translocation ofPKA to the nucleus, colonic cryptswere stained for PKA and analyzedusing confocal microscopy. Coloniccrypts were isolated at the estrus stageof the cycle. After incubation with E2for 2 min, the intensity of the PKA sig-nal at the edge of the nuclear signal wasincreased (Fig. 7B). It was demon-strated to be inside the nucleus as themerged image (yellow) demonstratesthe kinase to be within the vicinity ofthe DNA signal. This would allow PKAto also directly phosphorylate CREB inaddition to MAPK. This dual regula-tion of CREB by both MAPK and PKAconcurrently has previously been doc-
umented (25, 26), increasing the level of CREB phosphor-ylation. This translocation was blocked by the addition ofthe MEK-1 inhibitor PD 98059 indicating that MAPKrecruits PKA in the regulation of CREB.
E2 increases the mRNA levels of PKC�, PKAisoform I, and CREB via a MAPK/PKA pathway
In a previous study, it was demonstrated that chronicexposure to chronic E2 (10 nM) dramatically increasedtranscription of PKC� mRNA in colonic crypts isolatedfrom female rats (15). This response was absent in themale. In the present study, total RNA was extractedfrom female colonic crypts at estrus. Specific primers toPKC�, PKACI, PKARI, CREB, and �-actin were syn-thesized (MWG-Biotech, London, UK). The PKA iso-form I has previously been shown to be the isoformactivated in the rapid estrogen signaling pathway in ratcolonic tissue (15, 27). Expression differences in theprotein kinase levels were demonstrated in this study,and we investigated whether mRNA expression wasalso regulated by estrogen.
E2 (10 nM) induced an up-regulation of mRNA levelsfor all the signaling components studied: PKC� (5 � 1.4-fold higher, n � 4; P � 0.01), PKACI� (6 � 2-fold higher,n � 4; P � 0.01), and CREB (1.5 � 0.06-fold higher, n �4; P � 0.05) using quantitative real-time (qRT)-PCRanalysis (Fig. 8, A, B, and D). No change in mRNA ex-pression levels were observed for PKACI� (1 � 0.005-fold higher, n � 4; P � 0.05), indicating that E2 specifi-cally regulates the PKACI� subtype in female rat distalcolon. The up-regulation of PKC�, PKACI�, and CREBwas blocked by pretreatment with PKA and MAPKinhibitors. The inhibitory data support the involvement
FIG. 3. The transcription factor CREB basal expression levels differ throughout the estrouscycle. CREB protein expression was analyzed throughout the estrous cycle. The figure shows arepresentative Western blot of the transcription factor. Values on the graphs are given as amean fold increase compared with proestrus samples. Values are displayed as mean � SEM
(n � 3). **, P � 0.01; ***, P � 0.001.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1889
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
for the CREB transcription factor in driving the expres-sion of the signaling kinases at an RNA level. Atpresent, there is no significant inhibitor of the CREBtranscription factor.
E2-induced rapid kinase signalingand transcriptional responsesare transduced via amembrane-associated ER�
We investigated whether the phos-phorylation of PKC� in response to E2occurred via a membrane-bound ER�
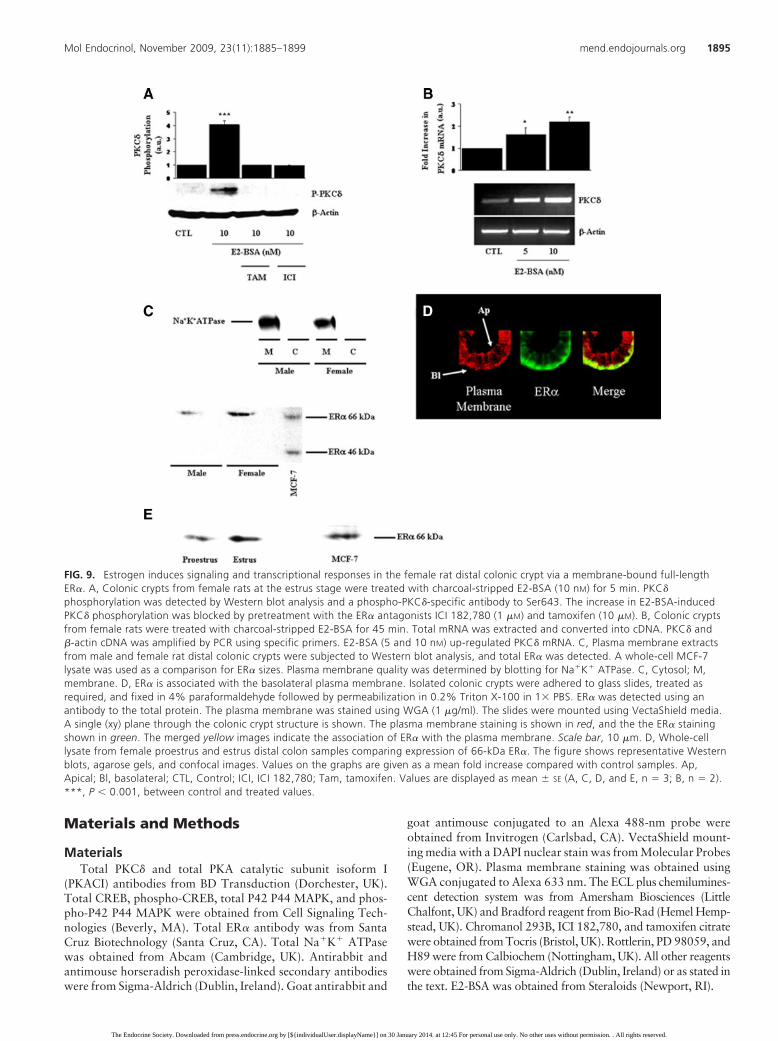
protein. We have previously reported thephosphorylation of PKC� at Ser643 inresponse to E2 (15). The impeded ligandform of estrogen E2-BSA (10 nM) in-duced rapid (5 min) phosphorylation ofPKC� (4 � 0.3-fold higher, n � 3; P �0.001) (Fig. 9A). The membrane-imper-meant E2-BSA (10 nM) stimulated PKC�
phosphorylation comparable to uncon-jugated E2. The phosphorylation ofPKC� was inhibited by pretreatmentwith the ER inhibitors ICI 182,780 (1�M) and tamoxifen (10 �M). To deter-mine whether the transcriptional re-sponses of E2 were via a membrane ER,we investigated whether E2-BSA-in-duced up-regulation of PKC� mRNA.E2-BSA both at 5 and 10 nM induced anincrease in the level of PKC� mRNA, in-dicating regulation via a membrane ER(5 nM E2, 1.6 � 0.3-fold higher; 10 nM
E2, 2.2 � 0.2-fold higher; n � 2; Fig.9B). Male and female colonic cryptsamples were fractionated, and theplasma membrane was extracted. Thequality of the plasma membrane frac-tion was checked by immunoblottingusing the Na�K� ATPase as a marker(Fig. 9C). The level of detectable ER�
66-kDa protein at the plasma mem-brane was low in the male comparedwith the female fraction where the pro-tein was present at a level comparableto that of a whole-cell lysate prepara-tion of MCF-7 cells (Fig. 9C). Thetruncated 46-kDa version of ER� wasnot present in plasma membrane frac-tions of colonic crypts from males orfemales (Fig. 9C). Whole-cell lysateanalysis showed a very low level of 46-kDa ER� (data not shown). Isolatedfemale colonic crypts at the estrus stage
were subjected to immunofluorescence and confocalanalysis to determine location of the receptor and furtherconfirm that ER� is membrane bound in distal colonic
FIG. 4. E2 (10 nM) rapidly induces phosphorylation of CREB in female rat colonic crypts isolatedat the estrus stage. A, Isolated colonic crypts were adhered to glass slides, treated as required, andfixed in 4% paraformaldehyde followed by permeabilization in 0.2% Triton X-100 in 1� PBS.Phospho-CREB (P-CREB) was stained using an antibody to Ser133. The slides were mounted usingVectaShield media containing the DNA stain DAPI. A single (xy) plane through the colonic cryptstructure is shown using an �40 oil objective. B, Further magnification using a �63 oil objectivewas carried out. Nuclei at the base (bottom) of the crypt were imaged again using a single (xy)plane through the colonic crypt. Red shows the nuclei staining, and the green shows the phospho-CREB staining. The merged yellow images indicate that CREB was phosphorylated in the vicinity ofthe DNA. Arrow indicates an area of CREB phosphorylation. CTL, Control; E2, E2 for 2 min; E5, E2for 5 min; VEH2, vehicle for 2 min; VEH5, vehicle for 5 min. Each treatment was examined in threerats with five to seven crypts for each treatment per rat. Phosphorylation of CREB in the nuclei ofvehicle-treated cells did not increase above basal levels. Scale bar, 20 �m (A) and 5 �m (B).
1890 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
crypts. Wheat germ agglutinin (WGA) was used to stainfor plasma membrane, and ER� association at the baso-lateral membrane was investigated. ER� was detectedand localized at the basolateral plasma membrane. Asso-ciation of the receptor with the plasma membrane isapparent from the merged image of plasma membraneand ER� (Fig. 9D). This is the first report of a mem-brane-associated full-length ER� in the distal colon.Figure 9E demonstrates estrous cycle differences in 66-kDa ER� in whole-cell lysates from female coloniccrypts.
Discussion
E2 induces an antisecretory responsein the female rat distal colon (15).The antisecretory action of E2 is es-trous cycle dependent, indicating theresponse is differentially primed de-pending on the level of sex steroids inthe blood plasma. Reproductive cycleregulation of cross-organ interactionshas been reported between the repro-ductive, gastrointestinal, and urinarytracts. A previous study has shown alevel of functional communication be-tween the uterus, colon, and bladder(28). In another study, inflammation ofthe colon has been demonstrated to af-fect uterine contractility and in turn in-flammation is also affected by the es-trous cycle (29).
The endometrial implantation win-dow is associated with expansion ofthe extracellular fluid volume. Here weshow that the fluctuating hormone lev-els throughout the estrous cycle arecorrelated with changes in the poten-tial for estrogen to inhibit secretion inthe colon.
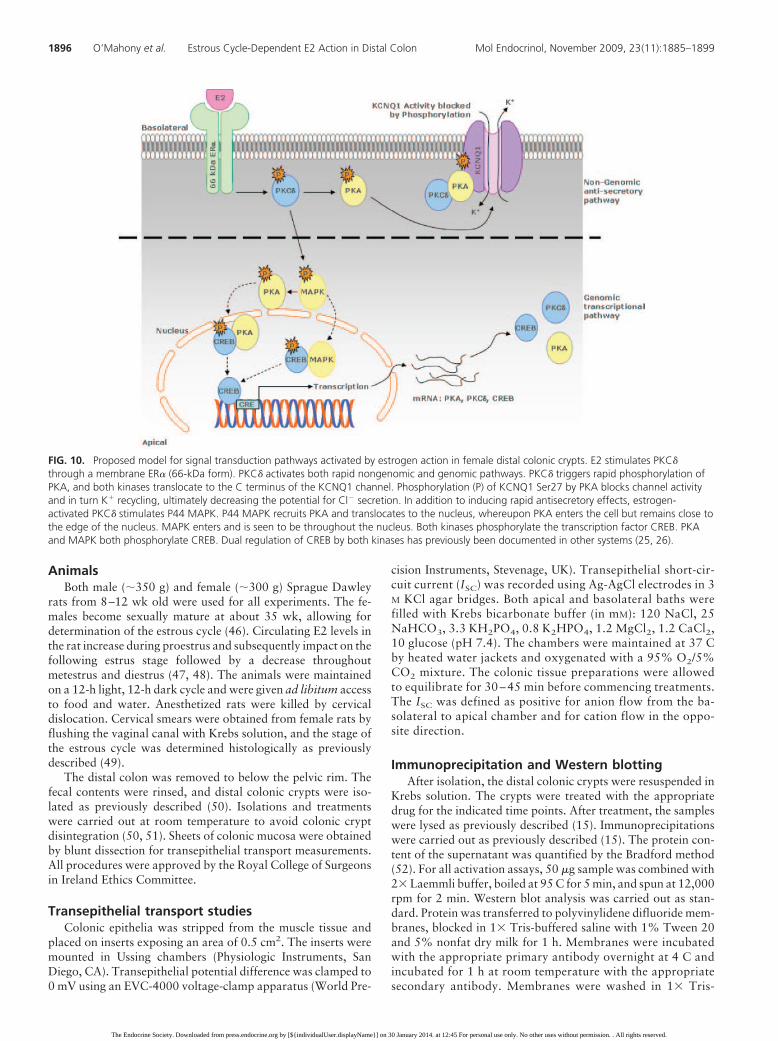
We have previously described themolecular mechanism of estrogen in-hibition of Cl� secretion in the femaledistal colon (15). The antisecretorymechanism involves the rapid andtransient up-regulation of PKC�/PKAactivity. It was demonstrated thatPKC� phosphorylates PKA, which inturn associates with the KCNQ1 chan-nel inducing serine phosphorylation ofthe channel. Phosphorylation of theKCNQ1 channel results in a decreasein basolateral K� recycling, which re-duces the electrical driving force for
Cl� secretion. The key signaling factor underlying thesexual dimorphism in E2 action is the differential regula-tion of PKC� at a transcriptional level. PKC� expressionwas higher in the female compared with the male. Thepresent study identified key estrous cycle-dependentchanges in the expression of PKC� and PKA both at tran-script and protein levels.
There are few reports in the literature on estradiol reg-ulation of kinase expression throughout the reproductivecycle. Estrogen has previously been shown to regulate
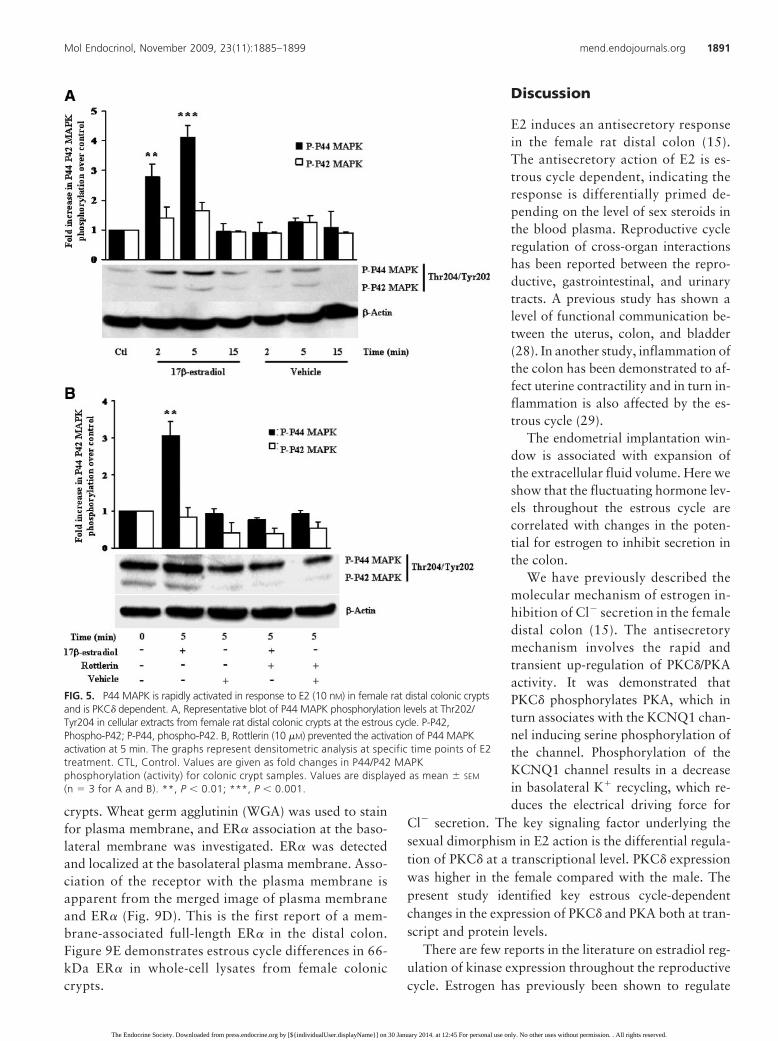
FIG. 5. P44 MAPK is rapidly activated in response to E2 (10 nM) in female rat distal colonic cryptsand is PKC� dependent. A, Representative blot of P44 MAPK phosphorylation levels at Thr202/Tyr204 in cellular extracts from female rat distal colonic crypts at the estrous cycle. P-P42,Phospho-P42; P-P44, phospho-P42. B, Rottlerin (10 �M) prevented the activation of P44 MAPKactivation at 5 min. The graphs represent densitometric analysis at specific time points of E2treatment. CTL, Control. Values are given as fold changes in P44/P42 MAPKphosphorylation (activity) for colonic crypt samples. Values are displayed as mean � SEM
(n � 3 for A and B). **, P � 0.01; ***, P � 0.001.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1891
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
PKC� in the rat ovary where PKC� increased during preg-nancy and also fluctuated at different stages of luteal dif-ferentiation (30). A previous study in uterine tissue dem-onstrated that PKB was regulated by the sex steroidhormones, and expression fluctuated throughout the re-productive cycle (31). Here we show both PKC� andPKACI expression levels varied throughout the estrouscycle. PKC� and PKACI are the major signaling compo-nents recruited by estrogen in the antisecretory response.The increased expression of PKC� and PKACI fromproestrus toward the diestrus stage was associated withincreased potency of the estrogen inhibition of Cl� secre-tion at diestrus. PKC� and PKACI expression was signif-icantly reduced in proestrus and correlated with minimalinhibition of Cl� secretion by estrogen. This is the first re-port of estrous cycle-dependent regulation of protein kinaseexpression in colonic tissue. It appears that the expressionlevel of the kinases is the rate-limiting factor in determiningthe potency of the antisecretory response to estrogen.
The rapid estrogen activation of PKA by PKC� andsubsequent association of PKA with the KCNQ1 channel
has been shown to be PKC� depen-dent in colonic crypts (15). PKA acti-vation appears to be the final step inthe inhibition of KCNQ1 channel ac-tivity. Indeed, PKA has previouslybeen shown to directly interact withand phosphorylate the channel atSer27 (32), further indicating it is thelast step in the signaling cascade thatimpacts on the channel. We thereforeinvestigated PKA activation as the end-point of the rapid antisecretory path-way and whether this activation wasalso dependent on the stage of the es-trous cycle. The results indicated thatthe timing and intensity of PKA activa-tion by estrogen is dependent on theestrous cycle stage. The failure of estro-gen to produce PKA activation atproestrus correlated with minimal in-hibition of secretion. Conversely, themaximal PKA activation at diestruscorrelated with maximal inhibition ofsecretion. We hypothesize that the lackof kinase activation by estrogen atproestrus may be due to reduced basalexpression levels of PKC� and PKA. Ithas previously been determined thatthe lack of estrogen effects on Cl� se-cretion in the male colonic mucosa isdue to the low expression of PKC�
compared with the high level in the fe-male tissue (15). PKA activity has been shown to be reg-ulated by the reproductive cycle in the hypothalamus(33). The largest cAMP-induced PKA activation in thehypothalamus was observed during diestrus, similar tothe sustained diestrus activation demonstrated in thisstudy. A recent study showed that estrogen-induced ac-celeration of the oviductal transport of the oocytes incycling rats was dependent on a successive activation ofnongenomic PKA signaling pathways (34). The latterstudy and the estrous cycle data presented in this paperdemonstrate a physiological role for rapid nongenomicPKA activation. The potential for estrogen to inhibit in-testinal Cl� secretion is dependent on a combined effectof enhanced PKC� and PKA expression and rapid activa-tion of the kinases.
The differences in kinase expression during the estrouscycle indicate the levels are regulated at a genomic level bythe sex steroid hormones. It was hypothesized that in addi-tion to causing the rapid antisecretory response by inhibitingKCNQ1 channels, estrogen may also induce a rapid tran-
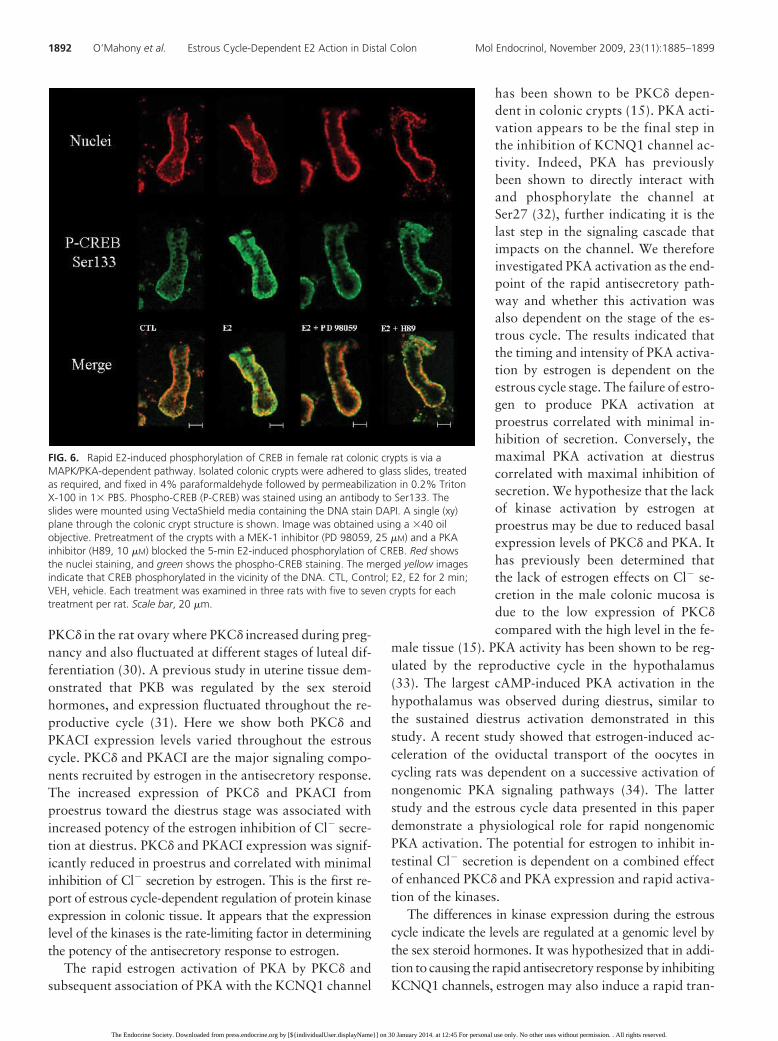
FIG. 6. Rapid E2-induced phosphorylation of CREB in female rat colonic crypts is via aMAPK/PKA-dependent pathway. Isolated colonic crypts were adhered to glass slides, treatedas required, and fixed in 4% paraformaldehyde followed by permeabilization in 0.2% TritonX-100 in 1� PBS. Phospho-CREB (P-CREB) was stained using an antibody to Ser133. Theslides were mounted using VectaShield media containing the DNA stain DAPI. A single (xy)plane through the colonic crypt structure is shown. Image was obtained using a �40 oilobjective. Pretreatment of the crypts with a MEK-1 inhibitor (PD 98059, 25 �M) and a PKAinhibitor (H89, 10 �M) blocked the 5-min E2-induced phosphorylation of CREB. Red showsthe nuclei staining, and green shows the phospho-CREB staining. The merged yellow imagesindicate that CREB phosphorylated in the vicinity of the DNA. CTL, Control; E2, E2 for 2 min;VEH, vehicle. Each treatment was examined in three rats with five to seven crypts for eachtreatment per rat. Scale bar, 20 �m.
1892 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
scriptional pathway to induce expres-sion of the kinases required to drive theantisecretory response throughout thereproductive cycle. Cross talk betweenrapid nongenomic and genomic ac-tions of steroid hormones in epithelia isstill a mystery. Estrogen may rapidlyinduce alterations in gene transcriptionas a result of the activation of secondmessengers, rather than binding to theclassical receptor, and in turn modu-late certain transcription factors. Weinvestigated whether the expression ofthe CREB transcription factor was reg-ulated throughout the estrous cycle.Previous work has demonstrated a re-productive cycle dependence of CREBexpression in human endometrium(35) and in rat hypothalamic tissue(36). We hypothesized that regulationof protein kinase expression by estra-diol throughout the estrous cycleprimes the rapid nongenomic antise-cretory action of estrogen. Conversely,the rapid protein kinase activation re-sponses to estrogen also prime thegenomic transcriptional pathway. Inthis way, estradiol works in a bidirec-tional manner throughout the repro-ductive cycle. The expression of CREBwas shown to be sex specific with ap-proximately eight times more of theprotein expressed in the female tissuein comparison with the male. Expres-sion of CREB was also shown to beregulated throughout the estrous cy-cle. Expression of CREB was low toabsent at proestrus followed by a dra-matic increase during estrus and amaximal expression at the metestrusstage. The estrous cycle modulationof the regulation of CREB indicatesthat the rapid phase of estrogen actionin the distal colon also regulates tran-scription. Estrogen induced the phos-phorylation of CREB and induction ofthe expression of CREB itself, PKC�,and PKA via a rapid MAPK-PKA sig-naling pathway. P42 P44 MAPK is animportant regulator of transcription inepithelial cells. Rapid sex-specific P42P44 MAPK activation in response to
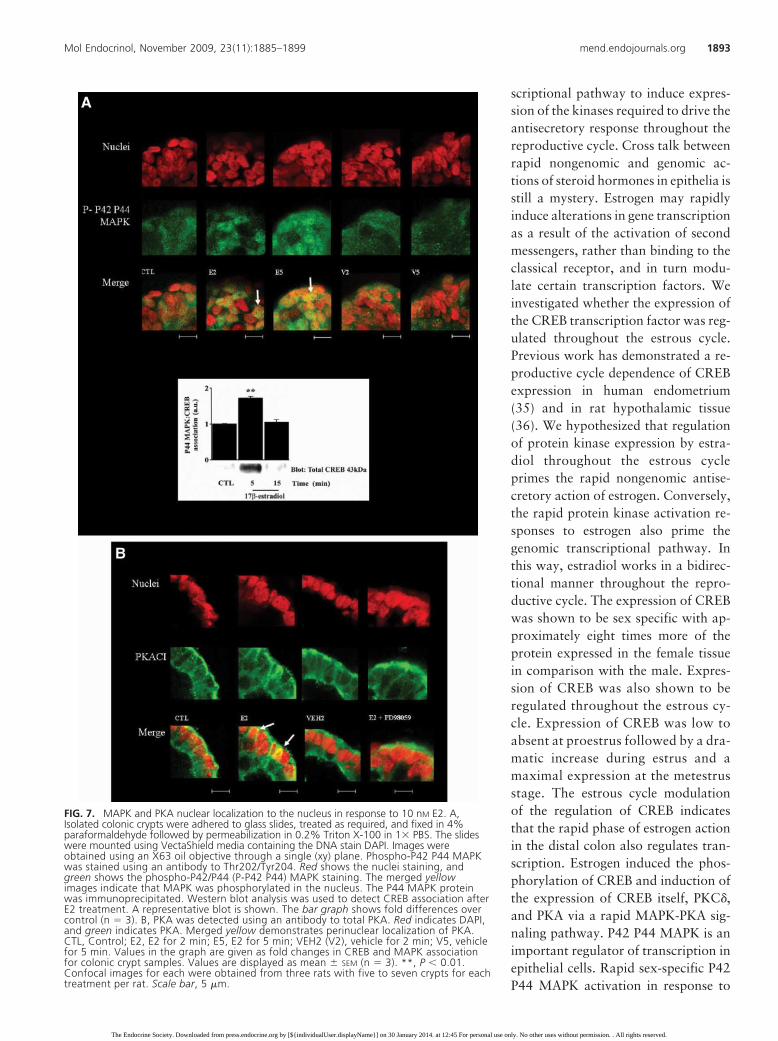
FIG. 7. MAPK and PKA nuclear localization to the nucleus in response to 10 nM E2. A,Isolated colonic crypts were adhered to glass slides, treated as required, and fixed in 4%paraformaldehyde followed by permeabilization in 0.2% Triton X-100 in 1� PBS. The slideswere mounted using VectaShield media containing the DNA stain DAPI. Images wereobtained using an X63 oil objective through a single (xy) plane. Phospho-P42 P44 MAPKwas stained using an antibody to Thr202/Tyr204. Red shows the nuclei staining, andgreen shows the phospho-P42/P44 (P-P42 P44) MAPK staining. The merged yellowimages indicate that MAPK was phosphorylated in the nucleus. The P44 MAPK proteinwas immunoprecipitated. Western blot analysis was used to detect CREB association afterE2 treatment. A representative blot is shown. The bar graph shows fold differences overcontrol (n � 3). B, PKA was detected using an antibody to total PKA. Red indicates DAPI,and green indicates PKA. Merged yellow demonstrates perinuclear localization of PKA.CTL, Control; E2, E2 for 2 min; E5, E2 for 5 min; VEH2 (V2), vehicle for 2 min; V5, vehiclefor 5 min. Values in the graph are given as fold changes in CREB and MAPK associationfor colonic crypt samples. Values are displayed as mean � SEM (n � 3). **, P � 0.01.Confocal images for each were obtained from three rats with five to seven crypts for eachtreatment per rat. Scale bar, 5 �m.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1893
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
E2 has previously been demonstrated in female growthplate chondrocytes (37). PKC is a known regulator ofcellular transcription, and PKC� has been demonstratedto induce transcription in bronchial epithelial cells via aP42 P44 MAPK pathway (38). A more recent study inuterine tissue demonstrated rapid E2 activation of P42P44 MAPK via a PKC�-dependent pathway (39). Thus, apositive feed-forward mechanism for CREB, PKC�, andPKA expression is driven by rapid E2 activation of PKC�.This is further evidence for dependence of a genomic re-sponse on the rapid induction of kinase activity. P44MAPK appears to be the signaling protein linking therapid nongenomic pathway to the genomic pathway. Inthis study, MAPK is clearly seen to enter and pool withinthe nucleus. PKA is also responsible for phosphorylatingCREB. PKA localizes just inside the nuclear envelope (asseen with merged image of PKA and DAPI signal), and itmay phosphorylate CREB within this region. RapidCREB phosphorylation at Ser133 in response to estrogenhas previously been demonstrated to occur via both PKAand MAPK concurrently. A previous study in testiculartissue demonstrated rapid CREB phosphorylation via themembrane ER�, leading to both P42 P44 MAPK and PKAactivation, which in turn both modulate the phosphory-lation of CREB (blocked by PD 98059 and H89) (26).Another study demonstrated cooperative effects betweenPKA and MAPK in the phosphorylation of CREB atSer133 in the nucleus similar to this study (25).
The distal colon was previouslythought not to be a target of estrogenuntil the confirmation of ER expres-sion (40). Increasing evidence suggeststhat E2 acts via a membrane-bound ERin many cell types (41–44). We haveprovided evidence for membrane-bound ER� involvement in transduc-ing rapid E2 phosphorylation of PKC�.The expression of 66-kDa ER� waslow in male colonic tissue in compari-son with female. We hypothesize that amembrane-bound 66-kDa ER� is thenongenomic receptor transducing therapid antisecretory actions of E2 inthe female distal colon. This is the firstreport of a gender difference in the ex-pression of the membrane 66-kDa ER�
protein. Plasma membrane preparationsdemonstrated that ER� is present at theplasma membrane to the basolateral sideof the cell (with an absence of the trun-cated 46-kDa ER form) and showed ahigher abundance in the female in com-
parison with the male. Based on this observation, it appearsthat a membrane-bound 66-kDa ER� is the nongenomicreceptor transducing the rapid actions of E2 in the femaledistal colon.
The fact that the reproductive cycle regulates interre-lated nongenomic and genomic actions in the intestinefurther strengthens the case for a physiological impact ofrapid responses to sex steroid hormones. The results pre-sented in this study clearly demonstrated the impact of thereproductive cycle on the intestinal function in the femaleboth at a molecular level through the regulation of proteinexpression and at a physiological level through the inhi-bition of transepithelial Cl� secretion. This is the firstreport of the modulation of rapid responses to steroidhormones throughout the estrous cycle in the gastrointes-tinal tract. A previous report demonstrated that estrouscycle differences in dopamine release from rat striatumwere primed by differential regulation of the level of crosstalk between acute and long-term effects of estrogenthrough each phase of the cycle (45). In our study, therapid antisecretory action of E2 was primed throughout theestrous cycle. In addition to the impact of sexual dimor-phism on the rapid responses to steroid hormones, the workreported in this paper has demonstrated the reproductivecycle as a new physiological regulatory element in the rapidresponses to estrogen. The study also provides novel insightsinto the molecular and cellular basis for fluid retention in thefemale in high-estrogen states (Fig. 10).
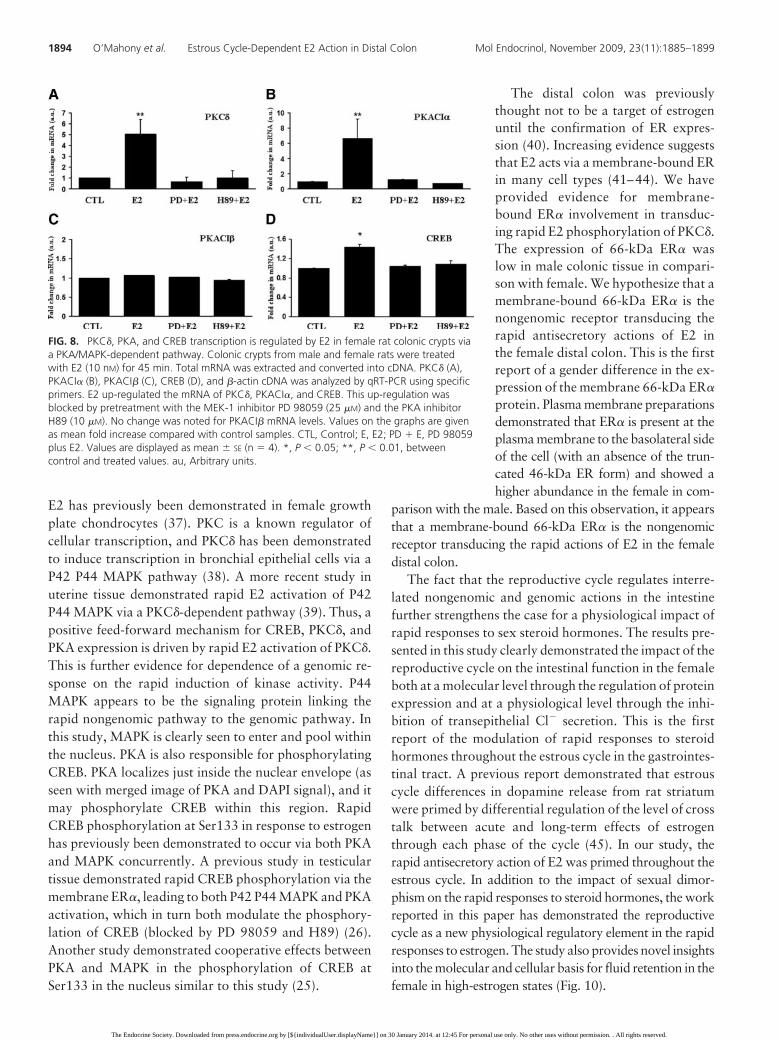
FIG. 8. PKC�, PKA, and CREB transcription is regulated by E2 in female rat colonic crypts viaa PKA/MAPK-dependent pathway. Colonic crypts from male and female rats were treatedwith E2 (10 nM) for 45 min. Total mRNA was extracted and converted into cDNA. PKC� (A),PKACI� (B), PKACI� (C), CREB (D), and �-actin cDNA was analyzed by qRT-PCR using specificprimers. E2 up-regulated the mRNA of PKC�, PKACI�, and CREB. This up-regulation wasblocked by pretreatment with the MEK-1 inhibitor PD 98059 (25 �M) and the PKA inhibitorH89 (10 �M). No change was noted for PKACI� mRNA levels. Values on the graphs are givenas mean fold increase compared with control samples. CTL, Control; E, E2; PD � E, PD 98059plus E2. Values are displayed as mean � SE (n � 4). *, P � 0.05; **, P � 0.01, betweencontrol and treated values. au, Arbitrary units.
1894 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
Materials and Methods
MaterialsTotal PKC� and total PKA catalytic subunit isoform I
(PKACI) antibodies from BD Transduction (Dorchester, UK).Total CREB, phospho-CREB, total P42 P44 MAPK, and phos-pho-P42 P44 MAPK were obtained from Cell Signaling Tech-nologies (Beverly, MA). Total ER� antibody was from SantaCruz Biotechnology (Santa Cruz, CA). Total Na�K� ATPasewas obtained from Abcam (Cambridge, UK). Antirabbit andantimouse horseradish peroxidase-linked secondary antibodieswere from Sigma-Aldrich (Dublin, Ireland). Goat antirabbit and
goat antimouse conjugated to an Alexa 488-nm probe wereobtained from Invitrogen (Carlsbad, CA). VectaShield mount-ing media with a DAPI nuclear stain was from Molecular Probes(Eugene, OR). Plasma membrane staining was obtained usingWGA conjugated to Alexa 633 nm. The ECL plus chemilumines-cent detection system was from Amersham Biosciences (LittleChalfont, UK) and Bradford reagent from Bio-Rad (Hemel Hemp-stead, UK). Chromanol 293B, ICI 182,780, and tamoxifen citratewere obtained from Tocris (Bristol, UK). Rottlerin, PD 98059, andH89 were from Calbiochem (Nottingham, UK). All other reagentswere obtained from Sigma-Aldrich (Dublin, Ireland) or as stated inthe text. E2-BSA was obtained from Steraloids (Newport, RI).
FIG. 9. Estrogen induces signaling and transcriptional responses in the female rat distal colonic crypt via a membrane-bound full-lengthER�. A, Colonic crypts from female rats at the estrus stage were treated with charcoal-stripped E2-BSA (10 nM) for 5 min. PKC�
phosphorylation was detected by Western blot analysis and a phospho-PKC�-specific antibody to Ser643. The increase in E2-BSA-inducedPKC� phosphorylation was blocked by pretreatment with the ER� antagonists ICI 182,780 (1 �M) and tamoxifen (10 �M). B, Colonic cryptsfrom female rats were treated with charcoal-stripped E2-BSA for 45 min. Total mRNA was extracted and converted into cDNA. PKC� and�-actin cDNA was amplified by PCR using specific primers. E2-BSA (5 and 10 nM) up-regulated PKC� mRNA. C, Plasma membrane extractsfrom male and female rat distal colonic crypts were subjected to Western blot analysis, and total ER� was detected. A whole-cell MCF-7lysate was used as a comparison for ER� sizes. Plasma membrane quality was determined by blotting for Na�K� ATPase. C, Cytosol; M,membrane. D, ER� is associated with the basolateral plasma membrane. Isolated colonic crypts were adhered to glass slides, treated asrequired, and fixed in 4% paraformaldehyde followed by permeabilization in 0.2% Triton X-100 in 1� PBS. ER� was detected using anantibody to the total protein. The plasma membrane was stained using WGA (1 �g/ml). The slides were mounted using VectaShield media.A single (xy) plane through the colonic crypt structure is shown. The plasma membrane staining is shown in red, and the the ER� stainingshown in green. The merged yellow images indicate the association of ER� with the plasma membrane. Scale bar, 10 �m. D, Whole-celllysate from female proestrus and estrus distal colon samples comparing expression of 66-kDa ER�. The figure shows representative Westernblots, agarose gels, and confocal images. Values on the graphs are given as a mean fold increase compared with control samples. Ap,Apical; Bl, basolateral; CTL, Control; ICI, ICI 182,780; Tam, tamoxifen. Values are displayed as mean � SE (A, C, D, and E, n � 3; B, n � 2).***, P � 0.001, between control and treated values.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1895
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.
AnimalsBoth male (350 g) and female (300 g) Sprague Dawley
rats from 8–12 wk old were used for all experiments. The fe-males become sexually mature at about 35 wk, allowing fordetermination of the estrous cycle (46). Circulating E2 levels inthe rat increase during proestrus and subsequently impact on thefollowing estrus stage followed by a decrease throughoutmetestrus and diestrus (47, 48). The animals were maintainedon a 12-h light, 12-h dark cycle and were given ad libitum accessto food and water. Anesthetized rats were killed by cervicaldislocation. Cervical smears were obtained from female rats byflushing the vaginal canal with Krebs solution, and the stage ofthe estrous cycle was determined histologically as previouslydescribed (49).
The distal colon was removed to below the pelvic rim. Thefecal contents were rinsed, and distal colonic crypts were iso-lated as previously described (50). Isolations and treatmentswere carried out at room temperature to avoid colonic cryptdisintegration (50, 51). Sheets of colonic mucosa were obtainedby blunt dissection for transepithelial transport measurements.All procedures were approved by the Royal College of Surgeonsin Ireland Ethics Committee.
Transepithelial transport studiesColonic epithelia was stripped from the muscle tissue and
placed on inserts exposing an area of 0.5 cm2. The inserts weremounted in Ussing chambers (Physiologic Instruments, SanDiego, CA). Transepithelial potential difference was clamped to0 mV using an EVC-4000 voltage-clamp apparatus (World Pre-
cision Instruments, Stevenage, UK). Transepithelial short-cir-cuit current (ISC) was recorded using Ag-AgCl electrodes in 3M KCl agar bridges. Both apical and basolateral baths werefilled with Krebs bicarbonate buffer (in mM): 120 NaCl, 25NaHCO3, 3.3 KH2PO4, 0.8 K2HPO4, 1.2 MgCl2, 1.2 CaCl2,10 glucose (pH 7.4). The chambers were maintained at 37 Cby heated water jackets and oxygenated with a 95% O2/5%CO2 mixture. The colonic tissue preparations were allowedto equilibrate for 30 – 45 min before commencing treatments.The ISC was defined as positive for anion flow from the ba-solateral to apical chamber and for cation flow in the oppo-site direction.
Immunoprecipitation and Western blottingAfter isolation, the distal colonic crypts were resuspended in
Krebs solution. The crypts were treated with the appropriatedrug for the indicated time points. After treatment, the sampleswere lysed as previously described (15). Immunoprecipitationswere carried out as previously described (15). The protein con-tent of the supernatant was quantified by the Bradford method(52). For all activation assays, 50 �g sample was combined with2� Laemmli buffer, boiled at 95 C for 5 min, and spun at 12,000rpm for 2 min. Western blot analysis was carried out as stan-dard. Protein was transferred to polyvinylidene difluoride mem-branes, blocked in 1� Tris-buffered saline with 1% Tween 20and 5% nonfat dry milk for 1 h. Membranes were incubatedwith the appropriate primary antibody overnight at 4 C andincubated for 1 h at room temperature with the appropriatesecondary antibody. Membranes were washed in 1� Tris-
FIG. 10. Proposed model for signal transduction pathways activated by estrogen action in female distal colonic crypts. E2 stimulates PKC�through a membrane ER� (66-kDa form). PKC� activates both rapid nongenomic and genomic pathways. PKC� triggers rapid phosphorylation ofPKA, and both kinases translocate to the C terminus of the KCNQ1 channel. Phosphorylation (P) of KCNQ1 Ser27 by PKA blocks channel activityand in turn K� recycling, ultimately decreasing the potential for Cl� secretion. In addition to inducing rapid antisecretory effects, estrogen-activated PKC� stimulates P44 MAPK. P44 MAPK recruits PKA and translocates to the nucleus, whereupon PKA enters the cell but remains close tothe edge of the nucleus. MAPK enters and is seen to be throughout the nucleus. Both kinases phosphorylate the transcription factor CREB. PKAand MAPK both phosphorylate CREB. Dual regulation of CREB by both kinases has previously been documented in other systems (25, 26).
1896 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.

buffered saline with 1% Tween 20 three times for 15 min.Bands were detected using autoradiographic film and chemi-luminescence. The membranes were stripped using a high-saltstripping buffer (Promega, Southampton, UK) to obtain load-ing levels of �-actin.
PKA activation assayPKA activation was measured using a PepTag Assay for the
nonradioactive detection of cAMP-dependent protein kinase(Promega) according to the manufacturer’s instructions withminor modifications; 2.5 �l F-Kemptide PepTag and 2.5 �lcAMP activator solution were added instead of 5 �l.
Immunofluorescence and confocal microscopyIsolated female crypts were adhered to eight-well slides
(Nunc, Rochester, NY) using Cell-Tak (BD Biosciences, UK)using the adsorption technique (according to the manufacturer’sinstructions). Adhered crypts were treated as required andwashed with ice-cold 1� PBS to halt the treatment. The cellswere fixed in 4% paraformaldehyde solution (dissolved at 70 Cin 1� PBS) at room temperature. The crypts were then perme-abilized in 0.2% Triton X-100 in 1� PBS. The crypts wererinsed in 1� PBS and incubated at room temperature for 15 minin a 3% BSA blocking buffer to avoid nonspecific binding ofantibodies and fluorescent conjugates. After this, the antibodysolution (phospho-CREB, 1:100 dilution; phospho-MAPK P42P44, 1:100 dilution; PKACI, 1:100 dilution; ER�, 1:100 dilu-tion) was added in 2% BSA blocking buffer for 1 h at roomtemperature. The crypts were washed twice in 0.2% BSA buffer,and the rabbit secondary antibody (Alexa Fluor 488 conju-gated) was added in a 2% BSA buffer. The cells were washedthree times with a 0.2% BSA solution and twice with 1� PBS.The slides were then mounted in VectaShield (Vector Laborato-ries, Burlingame, CA) with DAPI DNA stain. Single crypts wereimaged using a �40 oil objective on a Zeiss LSM 510 confocalmicroscope. Detailed images of colonic crypt nuclei were ob-tained using a �63 oil objective. The excitation wavelengths forWGA, Alexa Fluor 488, and DAPI were 633, 488, and 345 nm,respectively.
Total RNA preparation, RT-PCR analysis, andreal-time PCR analysis
RNA extracts were prepared from female rat colonic cryptsat the estrus stage of the estrous cycle using the QIAGENRNeasy kit (QIAGEN, Crawley, UK). Because the RNA wasfrom primary tissue, we carried out a genomic DNA treatmentas a precaution. Any contaminating genomic DNA was digestedusing the DNA-free kit as per manufacturer’s instructions(Ambion, Huntingdon, UK). Single-strand cDNA was synthe-sized using the Improm II reverse transcriptase system (Pro-mega). The synthesized cDNA was quantified at 260 nm andcorrected for loading differences in RT-PCR reaction mixes.PKA isoform I has three catalytic subunits (�, �, and �) and tworegulatory subunits (� and �). PKACI� and PKARI� were notexamined because these subunit isotypes are primarily ex-pressed in nervous and adipose tissues. PKC� (NM_133307;forward, 5�-caccatcttccagaaagaacg-3�; reverse, 5�-cttgccatag-gtcccgttgttg-3�; product size, 352 bp), PKACI� (X57986; for-ward, 5�-tcctttgggcgagtgatgc-3�; reverse, 5�-gcgaagaagggcggg-taac-3�; product size, 563 bp), PKACI� (D10770; forward, 5�-cagatcgtgctaacatttgag-3�; reverse, 5�-gtcatcgaagttgctggtatc-3�;product size, 543 bp), PKARI� (NM_013181; forward, 5�-
gcaagacagattcagagcc-3�; reverse, 5�-ggttgcccattcattattg-3�;product size,393 bp), and CREB (NM_134443; forward, 5�-agcccctgccatcaccact-3�; reverse, 5�-tgctgcttccctgttcttcattag-3�;product size, 409 bp) primers were designed using primer 3analysis (http://frodo.wi.mit.edu/primer3/). GoTaq polymerasemix from Promega was used in the amplification. TouchdownPCR was used to amplify the cDNA for 25 cycles over an an-nealing temperature range of 59–49 C in the case of PKACI�,PKACI�, PKARI�, and CREB. PKC� was amplified over arange of 65–55 C for 25 cycles. �-Actin (NM_031144; forward,5�-cagtaatctccttctgcatcc-3�; reverse, 5�-actacctcatgaagatcctga-3�; product size, 350 bp) was amplified for 25 cycles at anannealing temperature of 52 C. Amplicons were analyzed on a2% 1� Tris-acetate-EDTA agarose gel and imaged using a UVlight source. A 100-bp marker was employed to determinewhether the amplicons were the correct size. PCR analysis wascarried out using the RNA preparations as the template to en-sure an absence of interfering genomic DNA. Real-time PCRanalysis was carried out using the Bio-Rad iCycler and SYBRGreenER qPCR SuperMix (Invitrogen) according to manufac-turer’s instructions. The PCR program was set up as described inthe SuperMix protocol (Invitrogen). A melting-curve analysiswas performed for all primer sets after PCR analysis to ensurethe absence of nonspecific amplicons and also primer-dimers. Achange in transcript levels was determined by the Ct method.�-Actin was used as an internal control.
Plasma membrane extractionPlasma membrane was extracted using the Mem-PER eu-
karyotic membrane protein extraction kit from Promega. Theplasma membrane was extracted according to the kit’s instruc-tions. The quality of extract was compared between cytosolicfraction and membrane fraction by immunoblotting for theplasma membrane marker Na�K� ATPase.
Statistical and densitometric analysisFor the study, the data are presented as mean � SEM for a
series of the indicated number of experiments. Statistical anal-ysis of the data was obtained by analysis using one-wayANOVA and Tukeys post hoc test for multiple analysis of morethan two groups. Densitometric analysis of Western blots, PKAassays, and RT-PCR images were performed using GeneToolssoftware (Syngene, Cambridge, UK).
Acknowledgments
Address all correspondence and requests for reprints to: FionaO’Mahony, Ph.D., Department of Molecular Medicine, RoyalCollege of Surgeons in Ireland, Education and Research CentreSmurfit Building, Beaumont Hospital, P.O. Box 9063, Dublin 9,Ireland. E-mail: [email protected].
This work was supported by The Wellcome Trust ProgramGrant 06089/Z/00/Z and Higher Education Authority of Ire-land (HEA) PRTLI Grant Cycle 3. R.A. was recipient of a Well-come Trust Studentship Prize 06379/Z/01/Z. F.O.M. is sup-ported by a postdoctoral fellowship grant from the NationalBiophotonics and Imaging Platform of Ireland.
Disclosure Summary: The authors have nothing to disclose.
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1897
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.

References
1. Westerholm B 1980 Clinical toxicology of estrogens. PharmacolTher 10:337–349
2. Weenink GH, Ten Cate JW, Kahle LH, Lamping RJ, Treffers PE1981 “Morning after pill” and antithrombin III. Lancet 1:1105
3. Stachenfeld NS, DiPietro L, Palter SF, Nadel ER 1998 Estrogeninfluences osmotic secretion of AVP and body water balance inpostmenopausal women. Am J Physiol 274:R187–R195
4. Harvey BJ, Condliffe S, Doolan CM 2001 Sex and salt hormones:rapid effects in epithelia. News Physiol Sci 16:174–177
5. Johnson JA, Davis JO 1976 The effect of estrogens on renal sodiumexcretion in the dog. Perspect Nephrol Hypertens 5:239–248
6. Sweezey N, Tchepichev S, Gagnon S, Fertuck K, O’Brodovich H1998 Female gender hormones regulate mRNA levels and functionof the rat lung epithelial Na channel. Am J Physiol 274:C379–C386
7. Condliffe SB, Doolan CM, Harvey BJ 2001 17�-Oestradiol acutelyregulates Cl� secretion in rat distal colonic epithelium. J Physiol530:47–54
8. Jung HK, Kim DY, Moon IH 2003 Effects of gender and menstrualcycle on colonic transit time in healthy subjects. Korean J InternMed 18:181–186
9. Cordova-Fraga T, Huerta-Franco R, Gutierrez-Juarez G, Sosa-Aquino M, Vargas-Luna M 2004 The colon transit time in differentphases of the menstrual cycle: assessed with biomagnetic technique.Neurol Clin Neurophysiol 2004:31
10. Woods NF, Lentz MJ, Mitchell ES, Shaver J, Heitkemper M 1998Luteal phase ovarian steroids, stress arousal, premenses perceivedstress, and premenstrual symptoms. Res Nurs Health 21:129–142
11. Christofferson RH, Nilsson BO 1988 Morphology of the endome-trial microvasculature during early placentation in the rat. CellTissue Res 253:209–220
12. Halm ST, Liao T, Halm DR 2006 Distinct K� conductive pathwaysare required for Cl� and K� secretion across distal colonic epithe-lium. Am J Physiol Cell Physiol 291:C636–C648
13. Kunzelmann K, Hubner M, Schreiber R, Levy-Holzman R, GartyH, Bleich M, Warth R, Slavik M, von Hahn T, Greger R 2001Cloning and function of the rat colonic epithelial K� channel KV-LQT1. J Membr Biol 179:155–164
14. McNamara B, Winter DC, Cuffe JE, O’Sullivan GC, Harvey BJ1999 Basolateral K� channel involvement in forskolin-activatedchloride secretion in human colon. J Physiol 519(Pt 1):251–260
15. O’Mahony F, Alzamora R, Betts V, LaPaix F, Carter D, Irnaten M,Harvey BJ 2007 Female gender-specific inhibition of KCNQ1 chan-nels and chloride secretion by 17�-estradiol in rat distal coloniccrypts. J Biol Chem 282:24563–24573
16. Johannessen M, Moens U 2007 Multisite phosphorylation of thecAMP response element-binding protein (CREB) by a diversity ofprotein kinases. Front Biosci 12:1814–1832
17. Smith CL, Onate SA, Tsai MJ, O’Malley BW 1996 CREB bindingprotein acts synergistically with steroid receptor coactivator-1 toenhance steroid receptor-dependent transcription. Proc Natl AcadSci USA 93:8884–8888
18. Beyer C, Karolczak M 2000 Estrogenic stimulation of neuritegrowth in midbrain dopaminergic neurons depends on cAMP/pro-tein kinase A signalling. J Neurosci Res 59:107–116
19. Roux PP, Blenis J 2004 ERK and p38 MAPK-activated proteinkinases: a family of protein kinases with diverse biological func-tions. Microbiol Mol Biol Rev 68:320–344
20. Lenormand P, Pages G, Sardet C, L’Allemain G, Meloche S, Pouysse-gur J 1993 MAP kinases: activation, subcellular localization and rolein the control of cell proliferation. Adv Second Messenger Phospho-protein Res 28:237–244
21. Lenormand P, Sardet C, Pages G, L’Allemain G, Brunet A, PouyssegurJ 1993 Growth factors induce nuclear translocation of MAP kinases(p42mapk and p44mapk) but not of their activator MAP kinase kinase(p45mapkk) in fibroblasts. J Cell Biol 122:1079–1088
22. Gschwendt M, Muller HJ, Kielbassa K, Zang R, Kittstein W,
Rincke G, Marks F 1994 Rottlerin, a novel protein kinase inhibitor.Biochem Biophys Res Commun 199:93–98
23. Alessi DR, Cuenda A, Cohen P, Dudley DT, Saltiel AR 1995 PD098059 is a specific inhibitor of the activation of mitogen-activatedprotein kinase kinase in vitro and in vivo. J Biol Chem 270:27489–27494
24. Lochner A, Moolman JA 2006 The many faces of H89: a review.Cardiovasc Drug Rev 24:261–274
25. Costes S, Longuet C, Broca C, Faruque O, Hani EH, Bataille D,Dalle S 2004 Cooperative effects between protein kinase A andp44/p42 mitogen-activated protein kinase to promote cAMP-re-sponsive element binding protein activation after �-cell stimulationby glucose and its alteration due to glucotoxicity. Ann NY Acad Sci1030:230–242
26. Bouskine A, Nebout M, Mograbi B, Brucker-Davis F, Roger C,Fenichel P 2008 Estrogens promote human testicular germ cell can-cer through a membrane-mediated activation of extracellular reg-ulated kinase and protein kinase A. Endocrinology 149:565–573
27. Doolan CM, Condliffe SB, Harvey BJ 2000 Rapid non-genomicactivation of cytosolic cyclic AMP-dependent protein kinase activ-ity and [Ca2�]i by 17�-oestradiol in female rat distal colon. Br JPharmacol 129:1375–1386
28. Winnard KP, Dmitrieva N, Berkley KJ 2006 Cross-organ interac-tions between reproductive, gastrointestinal, and urinary tracts:modulation by estrous stage and involvement of the hypogastricnerve. Am J Physiol Regul Integr Comp Physiol 291:R1592–R1601
29. Houdeau E, Larauche M, Monnerie R, Bueno L, Fioramonti J 2005Uterine motor alterations and estrous cycle disturbances associatedwith colonic inflammation in the rat. Am J Physiol Regul IntegrComp Physiol 288:R630–R637
30. Cutler Jr RE, Maizels ET, Hunzicker-Dunn M 1994 Delta proteinkinase-C in the rat ovary: estrogen regulation and localization. En-docrinology 135:1669–1678
31. Dery MC, Leblanc V, Shooner C, Asselin E 2003 Regulation of Aktexpression and phosphorylation by 17�-estradiol in the rat uterusduring estrous cycle. Reprod Biol Endocrinol 1:47
32. Kurokawa J, Motoike HK, Rao J, Kass RS 2004 Regulatory actionsof the A-kinase anchoring protein Yotiao on a heart potassiumchannel downstream of PKA phosphorylation. Proc Natl Acad SciUSA 101:16374–16378
33. Ostrowska A 1994 The activation of protein kinase A by cyclicAMP is influenced by oestradiol and progesterone in supernatantsfrom the anterior pituitary but not from hypothalamus of the fe-male rat. Biol Cell 81:223–226
34. Orihuela PA, Parada-Bustamante A, Zuniga LM, Croxatto HB2006 Inositol triphosphate participates in an oestradiol non-genomic signalling pathway involved in accelerated oviductal trans-port in cycling rats. J Endocrinol 188:579–588
35. Vienonen A, Miettinen S, Blauer M, Martikainen PM, Tomas E,Heinonen PK, Ylikomi T 2004 Expression of nuclear receptors andcofactors in human endometrium and myometrium. J Soc GynecolInvestig 11:104–112
36. Mogi K, Funabashi T, Mitsushima D, Hagiwara H, Kimura F 2005Sex difference in the response of melanin-concentrating hormoneneurons in the lateral hypothalamic area to glucose, as revealed bythe expression of phosphorylated cyclic adenosine 3�,5�-mono-phosphate response element-binding protein. Endocrinology 146:3325–3333
37. McMillan J, Fatehi-Sedeh S, Sylvia VL, Bingham V, Zhong M,Boyan BD, Schwartz Z 2006 Sex-specific regulation of growth platechondrocytes by estrogen is via multiple MAP kinase signalingpathways. Biochim Biophys Acta 1763:381–392
38. Vuong H, Patterson T, Shapiro P, Kalvakolanu DV, Wu R, Ma WY,Dong Z, Kleeberger SR, Reddy SP 2000 Phorbol ester-induced ex-pression of airway squamous cell differentiation marker, SPRR1B,is regulated by protein kinase C�/Ras/MEKK1/MKK1-dependent/AP-1 signal transduction pathway. J Biol Chem 275:32250–32259
39. Shum JK, Melendez JA, Jeffrey JJ 2002 Serotonin-induced
1898 O’Mahony et al. Estrous Cycle-Dependent E2 Action in Distal Colon Mol Endocrinol, November 2009, 23(11):1885–1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.

MMP-13 production is mediated via phospholipase C, protein ki-nase C, and ERK1/2 in rat uterine smooth muscle cells. J Biol Chem277:42830–42840
40. Thomas ML, Xu X, Norfleet AM, Watson CS 1993 The presence offunctional estrogen receptors in intestinal epithelial cells. Endocri-nology 132:426–430
41. Pappas TC, Gametchu B, Watson CS 1995 Membrane estrogenreceptors identified by multiple antibody labeling and impeded-ligand binding. FASEB J 9:404–410
42. Pietras RJ, Nemere I, Szego CM 2001 Steroid hormone receptors intarget cell membranes. Endocrine 14:417–427
43. Razandi M, Pedram A, Merchenthaler I, Greene GL, Levin ER 2004Plasma membrane estrogen receptors exist and functions as dimers.Mol Endocrinol 18:2854–2865
44. Yang JZ, O’Flatharta C, Harvey BJ, Thomas W 2008 MembraneER�-dependent activation of PKC� in endometrial cancer cells byestradiol. Steroids 73:1110–1122
45. Becker JB, Rudick CN 1999 Rapid effects of estrogen or progester-one on the amphetamine-induced increase in striatal dopamine areenhanced by estrogen priming: a microdialysis study. PharmacolBiochem Behav 64:53–57
46. Nequin LG, Alvarez J, Schwartz NB 1979 Measurement of serum
steroid and gonadotropin levels and uterine and ovarian variablesthroughout 4 day and 5 day estrous cycles in the rat. Biol Reprod20:659–670
47. Haim S, Shakhar G, Rossene E, Taylor AN, Ben-Eliyahu S 2003Serum levels of sex hormones and corticosterone throughout 4- and5-day estrous cycles in Fischer 344 rats and their simulation inovariectomized females. J Endocrinol Invest 26:1013–1022
48. Mannino CA, South SM, Inturrisi CE, Quinones-Jenab V 2005Pharmacokinetics and effects of 17�-estradiol and progesteroneimplants in ovariectomized rats. J Pain 6:809–816
49. Hubscher CH, Brooks DL, Johnson JR 2005 A quantitative methodfor assessing stages of the rat estrous cycle. Biotech Histochem80:79–87
50. Doolan CM, Harvey BJ 1996 Modulation of cytosolic protein ki-nase C and calcium ion activity by steroid hormones in rat distalcolon. J Biol Chem 271:8763–8767
51. Schultheiss G, Lan Kocks S, Diener M 2002 Methods for the studyof ionic currents and Ca2�-signals in isolated colonic crypts. BiolProced Online 3:70–78
52. Bradford MM 1976 A rapid and sensitive method for the quanti-tation of microgram quantities of protein using the principle ofprotein-dye binding. Anal Biochem 72:248–254
NIDDK Workshop on Circadian Rhythms and Metabolic Disease
April 12–13, 2010Marriott North Bethesda Hotel and Conference Center, Bethesda, MD
This workshop seeks to gain a better understanding of the link between circadian rhythms and human health anddisease. Emphasis will be placed on the influence of central and cellular clocks on the physiology of behavior andmetabolism, specifically on overall energy balance and obesity. The goal will be to identify areas of future research tobetter elucidate the contribution of circadian rhythms to metabolism and disease.
Further information may be obtained at http://www3.niddk.nih.gov/fund/other/circadian2010. For additional infor-mation contact Ronald Margolis, Ph.D., 301-594-8819/[email protected].
Mol Endocrinol, November 2009, 23(11):1885–1899 mend.endojournals.org 1899
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 30 January 2014. at 12:45 For personal use only. No other uses without permission. . All rights reserved.





























