Embed Size (px)
Citation preview

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
Contents lists available at ScienceDirect
HOMO - Journal of ComparativeHuman Biology
j o urnal homepage: www.elsev ier .com/ locate / j chb
Heritability estimates for somatic traitsdetermined perinatally with the twin method
Małgorzata Waszaka,∗, Krystyna Cieslika,Marzena Skrzypczak-Zielinskab, Marlena Szalatab,Karolina Wielgusc, Joanna Kempiakd, Grzegorz Breborowiczd,Ryszard Słomskib
a Department of Functional Anatomy, University School of Physical Education in Poznan, Królowej Jadwigi27/39, 61-871 Poznan, Polandb Institute of Human Genetics, Polish Academy of Sciences, Strzeszynska 32, 60-479 Poznan, Polandc Department of Biotechnology and Molecular Biology, Institute of Natural Fibres and Medicinal Plants,Wojska Polskiego 71b, 60-630 Poznan, Polandd Department of Perinatology and Gynecology in Poznan, University of Medical Sciences, Polna 33, 60-535Poznan, Poland
a r t i c l e i n f o
Article history:Received 20 August 2013Accepted 18 November 2014Available online xxx
a b s t r a c t
The aim of this study was to establish estimates for heritabilityof somatic traits determined perinatally with the twin method.The studied material, including 1263 twin pairs live-born at var-ious stages of prenatal life, i.e. between the 22nd and 41st weekof gestation, was collected at the Department of Perinatology andGynecology, Poznan University of Medical Sciences, between 2002and 2009. The zygosity of 821 pairs of same-sex twins was deter-mined at the Laboratory of Molecular Genetics in Poznan usinganalysis of single and multiple nucleotide polymorphisms.
The proportion of phenotypic variance explained by genetic vari-ability was determined using heritability indices based on intrapairvariances and correlations. Analysis of these indices revealed thatgenetic variability predominated over environmental variability indetermining the phenotypic variance of all studied traits. Heritabil-ity was the highest for body weight and shoulder width.
∗ Corresponding author. Tel.: +48 61 835 52 26; fax: +48 61 835 52 20.E-mail address: [email protected] (M. Waszak).
http://dx.doi.org/10.1016/j.jchb.2015.04.0010018-442X/© 2015 Elsevier GmbH. All rights reserved.

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
2 M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
The high values of heritability coefficients documented in our studywere to a certain extent associated with an inconsistency of theassumption on the additive effect of genetic and environmentaleffects. While analyzing the phenotypic variance of a trait, the roleof genotype–environment interactions and covariance cannot beignored, as these are their effects which increase the genetic vari-ance and related coefficients of heritability. Therefore, it can beconcluded that the genotypic component of the phenotypic vari-ance of the trait is enhanced by the effects of various environmentalfactors.
© 2015 Elsevier GmbH. All rights reserved.
s t r e s z c z e n i e
Celem pracy było oszacowanie odziedziczalnosci cech somaty-cznych w okresie perinatalnym przy wykorzystaniu metodyblizniat. Materiał badawczy zgromadzony w Klinice Perinatologiii Ginekologii Uniwersytetu Medycznego w Poznaniu w latach2002–2009 stanowiły zywo urodzone, w róznych etapach zyciapłodowego, tj. od 22 do 41 tygodnia ciazy 1263 pary blizniat. Ocenazygotycznosci 821 par blizniat jednopłciowych została wykonanaw Laboratorium Genetyki Molekularnej w Poznaniu na podstawieanalizy polimorfizmu DNA metoda jedno i wielopunktowa.Proporcje wariancji fenotypowej wyjasnianej zmiennoscia gene-tyczna okreslono posługujac sie wskaznikami odziedziczalnosciopartymi na wariancjach i korelacjach wewnatrzparowych. Anal-iza obliczonych współczynników odziedziczalnosci pokazuje, zew zmiennosci fenotypowej wszystkich omawianych w pracycech somatycznych przewaza udział zmiennosci genetycznejnad udziałem zmiennosci srodowiskowej. Najwiekszym stopniemodziedziczalnosci charakteryzuje masa ciała i szerokosc barków.Wysokie wartosci obliczonych przez nas współczynnikówodziedziczalnosci prawdopodobnie w pewnym stopniu wynikajaz niespełnienia załozenia o addytywnosci wpływów genetycznychi srodowiskowych. Analizujac wariancje fenotypowa cechy niemozna pominac w jej składzie wariancji interakcji i kowari-ancji genotyp-srodowisko. Bo to własnie ich efekty zwiekszajawariancje genetyczna, a wiec przyczyniaja sie do zwiekszeniawartosci współczynników odziedziczalnosci. Mozna zatempowiedziec, ze składnik genotypowy zmiennosci fenotypowejcechy jest potegowany poprzez wpływy róznorodnych czynnikówsrodowiskowych.
© 2015 Elsevier GmbH. All rights reserved.
Introduction
The concept of heritability originates from an attempt to describe a degree to which the phenotypicdifferences between individuals result from differences in their genotype rather than from differentenvironmental influences (Kang et al., 1978). Heritability can be relatively easily determined in thecase of animals, whose way of mating and environment can be controlled under experimental con-ditions. However, similar experiments would be unethical in the case of humans. This obstacle canbe eliminated by finding human samples which are naturally exposed to conditions required by agiven experimental method. Twin studies constitute one method of explaining the role of genes and

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx 3
environment in determining a given trait. However, they suffer from some methodological limitationsand are based on certain assumptions.
Analysis and interpretation of the results of twin studies from the heritability point of view requiresatisfying the assumptions of the so-called additive model. The assumptions of this model are simul-taneously the prerequisites of heritability estimation. According to one principal assumption of themodel, genetic and environmental factors act additively, independently from each other, and unidi-rectionally (i.e. constantly in the same manner). Therefore, this model is not adjusted for potentialcooperation between genetic and environmental factors, i.e. it does not include a component describ-ing potential interaction or correlation between genotype and environment, the so-called non-additiveeffect. This is a weak point of the additive model, since the effects of genotype–environment interac-tion and correlation are relatively frequently observed among twins (Bell and Saffery, 2012; Bergman,1987; Bergman and Sawicki, 1988; Bergman et al., 1983, 1987; Dubois et al., 2012; McCaffery et al.,2009; Pinker, 2004; Ridley, 2003; Rutter, 2006; Silventoinen et al., 2010). In such cases, total pheno-typic variance is not only a simple sum of genetic and environmental variances, but should be adjustedfor non-additive effects. However, the latter is difficult to achieve. The genotype–environment interac-tion occurs whenever individuals with different genotypes – dizygotic (DZ) twins respond differentlyto the same environment. In contrast, the genotype–environment correlation takes place if individualswith different genotypes – DZ twins stay or are placed in different environments.
Using the heritability index, one can determine the proportion of phenotypic variance which isexplained by genetic variability (Bell and Saffery, 2012; Bell and Spector, 2011; Falconer and Mackay,1996; Schousboe et al., 2003). It should be noted, however, that the coefficients of heritability deter-mined by comparisons of monozygotic (MZ) and DZ twins should be interpreted as heritability inthe wide sense, as heritability estimates obtained from twin studies reflect jointly additive effects,dominance, and epistasis.
The aim of this study was to determine the heritability of somatic traits in twins during theirfetal development, and to analyze the heritability measures in the context of their methodologicallimitations.
Materials and methods
The studied material, including 1263 twin pairs live-born at various stages of prenatal life, i.e.between the 22nd and 41st week of gestation, was collected at the Department of Perinatology andGynecology, Poznan University of Medical Sciences, between 2002 and 2009. Individuals with congen-ital anomalies or mechanical injuries were not included. The gestational age at birth was determinedby the time elapsed since the first day of the last menstrual period. The number of male and femaleneonates born at various gestational weeks, and the number of twin pairs per each lunar month arepresented in Table 1.
The degree of morphological development of studied twins was characterized by the followingsix somatic traits: (1) body weight, (2) total body length, (3) crown-rump length (si), (4) shoulderwidth, (5) head circumference, and (6) chest circumference. The definitions of these traits and themethod of measurement were consistent with the traditional measurement technique proposed byMartin (1988). The measurements were taken immediately after birth by appropriately trained med-ical personnel. Birth weight (in g) was determined with a medical scale, to the nearest 10 g. Bodylength was measured with a metric tape; total body length (v-pl) was defined as a chord of a circle
Table 1Number of male and female newborns from twin pregnancies born at various gestational weeks (22–41) and the number oftwin pairs born in various lunar months.
Lunar month 6th 7th 8th 9th 10th
Weeks ofpregnancy
22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41
♂ 3 7 9 13 23 20 16 22 22 46 69 61 109 131 206 294 144 63 19 5♀ 3 3 3 5 23 16 14 32 36 42 47 65 85 159 216 276 128 67 15 9
Pairs of twins 14 65 158 516 510

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
4 M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
having endpoints on the vertex (apex of the head) and plantare (the point at the foot surface, locatedat an extension of the anatomical crural axis). Maximum circumferences of the head and chest weredetermined with the metric tape. The lengths and circumferences were measured to the nearest 5 mm.
The zygosity of 821 pairs of same-sex twins was determined at the Laboratory of Molecular Geneticsin Poznan on the basis of the analysis of single and multiple nucleotide polymorphisms. One milliliterof the umbilical cord blood collected in EDTA (10 ml 10% EDTA per 1 ml of blood) immediately afterbirth was subjected to the DNA analysis. Following DNA isolation from umbilical blood lymphocytesall twins had DNA polymorphism analyses carried out – both through hybridization with a molecularprobe and with PCR. The hybridization method involved the analysis of one locus, i.e. D7S21 (7p22) andD12S11 (12q24.3), as well as multiple loci analysis with (CAC)5 probe. PCR technique was used in theanalysis of DNA polymorphism within short tandem repeats (STR) for the following genes: pancreaticlipase (HUMPLA2A1, 12q23), P450 cytochrome P450 (HUMCYARO, 15q21.1), von Willebrandt’s fac-tor (HUMvWF, 12p13), thyroid peroxidase (HUMTPOX, 2p23) and tyrosine hydroxylase (HUMTH01,11p15.5). In total, the analysis involved loci located on 5 different chromosomes.
All the procedures were approved by the Local Ethics Committee of the Medical University inPoznan.
Three types of coefficients were selected from an array of heritability measures reported in theliterature:
1. The h2 heritability coefficient calculated from the formula proposed by Clark (Bergman, 1987):
h2 = VG
VG + VE= VDZ − VMZ
VDZ,
where VG – genetic variance, VE – environmental variance, and VMZ and VDZ – intrapair variancesof a given trait in monozygotic (MZ) and dizygotic twins (DZ).
2. The formula for estimating the heritability of quantitative traits based on the intrapair coefficientsof correlation in MZ and DZ twins, proposed by Holzinger (Bergman, 1987) and referred to as the Hcoefficient:
H = rMZ − rDZ
1 − rDZ
where rMZ and rDZ – intrapair coefficients of correlation in MZ and DZ twins.3. The Falconer’s (1974) coefficient of heritability (Hdr), being a doubled difference between the intra-
pair coefficients of correlation of analyzed variable in groups of MZ and DZ twins:
Hdr = 2(rMZ − rDZ)
The following criteria must be satisfied in order to provide the reliability of the estimates mentionedabove:
a. equality of environmental correlation in MZ and DZ twinsb. equality of genotype–environment correlation in MZ and DZ twinsc. lack of dominanced. random mating.
If these criteria are not satisfied, or in the case of any inconsistency with the additive method, thecomparison of MZ and DZ twins may not produce an objective, close to reality estimates of heritability,and the h2, H, and Hdr coefficients can differ from one another. Some authors tighten the criterionb, stating that the heritability can be estimated solely when the effects of genotype–environmentcorrelation and interaction are absent.

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx 5
Table 2Heritability coefficients (h2, H, and Hdr) of studied somatic traits in three analyzed groups of twins.
Research group Generally twins Male Female
Feature h2 H Hdr h2 H Hdr h2 H Hdr
Body mass .82 .80 .36 .85 .83 .38 .78 .80 .39Total body length .73 .73 .33 .74 .72 .34 .71 .77 .37Crown rump length .78 .76 .35 .76 .70 .31 .76 .83 .47Shoulder width .81 .76 .41 .73 .72 .33 .76 .78 .44Head circumference .72 .71 .30 .75 .73 .29 .66 .70 .34Chest circumference .72 .69 .27 .78 .75 .32 .69 .73 .35
Significance of differences between intrapair variances of MZ and DZ twins was tested with an F-test, while significance of differences between correlation coefficients for MZ and DZ twins was testedusing Statistica package. All differences were significant at 0.01 level.
Results
Due to the low number of twin pairs from some gestational weeks, heritability was estimated forlunar months, which corresponded to the following weeks of pregnancy: 6th month – weeks 22–24,7th month – weeks 25–28, 8th month – weeks 29–32, 9th month – weeks 33–36, and 10th month –weeks 37–41.
Heritability coefficients were calculated for all analyzed somatic traits of DZ twins and mono-and dichorionic MZ twins, except for those affected with twin-to-twin transfusion syndrome (TTTS).The heritability coefficients, h2, H, and Hdr determined in three analyzed groups (all twins, males,and females) are presented in Table 2. While the values of the first two coefficients were similar,the values of the Falconer’s coefficient were approximately twice lower. Hdr is calculated from thedifference between the coefficients of intrapair correlation in MZ and DZ twins, and not from thevalues of these coefficients.
Analysis of the heritability coefficients determined in our study revealed that all analyzed somatictraits were characterized by relatively high heritability; namely, the involvement of genetic variabil-ity in the phenotypic variance of these traits predominated over the involvement of environmentalvariability. This was confirmed by high values of h2 coefficients, ranging from 0.72 to 0.82, and Hcoefficients, which amounted to 0.69–0.80. The h2 value of traits analyzed in male twins ranged from0.73 to 0.85, and was higher than in female twins (0.66–0.78, Table 2). In contrast, only the H valuesof body weight and circumferences of the head and chest were higher in males, while those char-acterizing the remaining traits were higher in female twins. Body weight and shoulder width werecharacterized by the highest heritability among all analyzed traits, as suggested by the highest valuesof h2 and H for these traits documented in the whole group of twins.
Some intertrait differences in h2 and H were sex-specific. In the group of male twins, the values ofthese coefficients were the highest in the case of body weight and chest circumference, while in femaletwins in the case of body weight, crown-rump length, and shoulder width. Comparison between thetwo types of heritability coefficients (h2 and H) revealed that in the case of all analyzed traits of maletwins the values of h2 were higher than H, whereas the inverse relationship was documented in femaletwins in whom the values of H coefficients were higher than the respective values of h2 coefficients.
The heritability coefficient H, based on intrapair correlations, increases with decreasing coefficientsof intrapair correlation in DZ twins, which is particularly evident in the case of female twins, suggestinggreater involvement of factors modifying the traits analyzed in this group. The coefficients of intrapaircorrelation in DZ twins showed a decreasing tendency with fetal age, especially in the case of bodyweight and head circumference (being the lowest at the 10th month; Table 3). During the terminalphase of fetal development, the rDZ coefficients oscillated around 0.5 or even 0.4 for the majority ofanalyzed traits. Assuming the lack of dominance with regard to the analyzed anthropometric traits, itshould be assumed that the decrease in rDZ coefficients resulted from genotype–environment inter-action or correlation. DZ twins, i.e. individuals characterized by different heritability, can respond
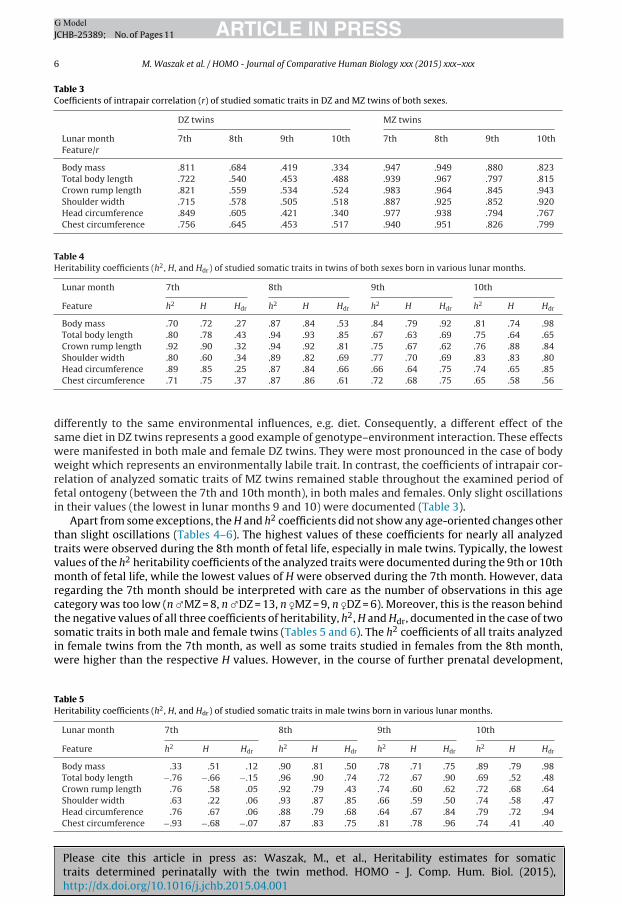
Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
6 M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
Table 3Coefficients of intrapair correlation (r) of studied somatic traits in DZ and MZ twins of both sexes.
DZ twins MZ twins
Lunar month 7th 8th 9th 10th 7th 8th 9th 10thFeature/r
Body mass .811 .684 .419 .334 .947 .949 .880 .823Total body length .722 .540 .453 .488 .939 .967 .797 .815Crown rump length .821 .559 .534 .524 .983 .964 .845 .943Shoulder width .715 .578 .505 .518 .887 .925 .852 .920Head circumference .849 .605 .421 .340 .977 .938 .794 .767Chest circumference .756 .645 .453 .517 .940 .951 .826 .799
Table 4Heritability coefficients (h2, H, and Hdr) of studied somatic traits in twins of both sexes born in various lunar months.
Lunar month 7th 8th 9th 10th
Feature h2 H Hdr h2 H Hdr h2 H Hdr h2 H Hdr
Body mass .70 .72 .27 .87 .84 .53 .84 .79 .92 .81 .74 .98Total body length .80 .78 .43 .94 .93 .85 .67 .63 .69 .75 .64 .65Crown rump length .92 .90 .32 .94 .92 .81 .75 .67 .62 .76 .88 .84Shoulder width .80 .60 .34 .89 .82 .69 .77 .70 .69 .83 .83 .80Head circumference .89 .85 .25 .87 .84 .66 .66 .64 .75 .74 .65 .85Chest circumference .71 .75 .37 .87 .86 .61 .72 .68 .75 .65 .58 .56
differently to the same environmental influences, e.g. diet. Consequently, a different effect of thesame diet in DZ twins represents a good example of genotype–environment interaction. These effectswere manifested in both male and female DZ twins. They were most pronounced in the case of bodyweight which represents an environmentally labile trait. In contrast, the coefficients of intrapair cor-relation of analyzed somatic traits of MZ twins remained stable throughout the examined period offetal ontogeny (between the 7th and 10th month), in both males and females. Only slight oscillationsin their values (the lowest in lunar months 9 and 10) were documented (Table 3).
Apart from some exceptions, the H and h2 coefficients did not show any age-oriented changes otherthan slight oscillations (Tables 4–6). The highest values of these coefficients for nearly all analyzedtraits were observed during the 8th month of fetal life, especially in male twins. Typically, the lowestvalues of the h2 heritability coefficients of the analyzed traits were documented during the 9th or 10thmonth of fetal life, while the lowest values of H were observed during the 7th month. However, dataregarding the 7th month should be interpreted with care as the number of observations in this agecategory was too low (n ♂MZ = 8, n ♂DZ = 13, n ♀MZ = 9, n ♀DZ = 6). Moreover, this is the reason behindthe negative values of all three coefficients of heritability, h2, H and Hdr, documented in the case of twosomatic traits in both male and female twins (Tables 5 and 6). The h2 coefficients of all traits analyzedin female twins from the 7th month, as well as some traits studied in females from the 8th month,were higher than the respective H values. However, in the course of further prenatal development,
Table 5Heritability coefficients (h2, H, and Hdr) of studied somatic traits in male twins born in various lunar months.
Lunar month 7th 8th 9th 10th
Feature h2 H Hdr h2 H Hdr h2 H Hdr h2 H Hdr
Body mass .33 .51 .12 .90 .81 .50 .78 .71 .75 .89 .79 .98Total body length −.76 −.66 −.15 .96 .90 .74 .72 .67 .90 .69 .52 .48Crown rump length .76 .58 .05 .92 .79 .43 .74 .60 .62 .72 .68 .64Shoulder width .63 .22 .06 .93 .87 .85 .66 .59 .50 .74 .58 .47Head circumference .76 .67 .06 .88 .79 .68 .64 .67 .84 .79 .72 .94Chest circumference −.93 −.68 −.07 .87 .83 .75 .81 .78 .96 .74 .41 .40

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx 7
Table 6Heritability coefficients (h2, H, and Hdr) of studied somatic traits in female twins born in various lunar months.
Lunar month 7th 8th 9th 10th
Feature h2 H Hdr h2 H Hdr h2 H Hdr h2 H Hdr
Body mass .94 .61 .16 .83 .87 .61 .89 .86 .98 .64 .66 .78Total body length .95 .89 .22 .91 .91 .60 .66 .66 .73 .71 .72 .76Crown rump length .88 .70 .11 .96 .95 .98 .79 .79 .94 .70 .77 .77Shoulder width .58 .50 .08 .71 .74 .54 .79 .77 .85 .71 .82 .93Head circumference .88 .36 .06 .84 .83 .35 .67 .62 .68 .60 .49 .56Chest circumference .81 .50 .09 .89 .93 .70 .70 .69 .84 .62 .59 .48
the values of these coefficients were similar, and H even exceeds h2 in the case of body weight andshoulder width.
The increase in the H coefficients for body weight and shoulder width documented during the fetalontogeny of female twins resulted from a simultaneous decrease in the coefficients of intrapair correla-tion of these traits in DZ twins. This phenomenon is associated with increased genotype–environmentcorrelation observed during this period.
The involvement of genotype–environment covariance in determining the genetic variance ofquantitative traits is probably highly variable, since it is affected by considerable changes of environ-mental conditions during various periods of life. The terminal stage of fetal development is additionallyconfounded by the effects of perinatal factors; probably, the role of genotype–environment interac-tions and correlations in determining the genetic variance of analyzed somatic traits increases inconcert with the variability of environmental factors affecting a pair of twins.
Discussion
To function correctly, heritability coefficients must meet certain conditions. Hdr is calculated fromthe difference between the coefficients of intrapair correlation in MZ and DZ twins, and not from thevalues of these coefficients. According to some authors, this formula based on a doubled differencebetween the coefficients of correlation of a given variable in a group of MZ and DZ twins can be consid-ered as a heritability estimate only providing the additive effect of genes (Allen, 1979). Oniszczenko(2002), in turn, blames the Falconer’s formula for its excessive genetic determinism and imprecisedistinguishing between genetic and environmental factors.
The h2 coefficient should not be interpreted separately from the variances on the basis of which itwas calculated. The assumption that a trait characterized by lower h2 is more susceptible to the influ-ence of environmental factors than that with higher h2 value is not always true (Skibinska and Skład,1979). A low value of h2 coefficient can result from an extremely low value of VDZ or an extremely highvalue of VMZ, and vice versa: a high value of h2 can be associated with an extremely high VDZ or anextremely low VMZ. This makes the h2 coefficient ambiguous, and causes difficulties in its interpreta-tion. Therefore, the Fisher’s intrapair coefficient of correlation is a better and more univocal measureof ecosensitivity within the pairs of MZ twins.
From a practical point of view, the heritability coefficient calculated on the basis of intrapair corre-lation is more useful than the h2 index. Any deviation (increase or decrease) of rDZ coefficient from thetheoretical expected value 0.5 signals inconsistence with the assumptions of additive model (e.g. thepresence of genotype–environment interaction or correlation) or with the prerequisites of heritabilitycoefficient estimation (e.g. random mating of twin’s parents, lack of dominance, etc.) (Bergman, 1987).
Studies on the role of genetic and environmental factors in determining the phenotypic variabilityof anthropometric traits have intensified during recent years. Most of these studies dealt with bodyweight and body height (Boziłow et al., 1992; Chumlea and San Guo, 2002; Hanisch et al., 2004;Ijzerman et al., 2005; Johnston et al., 1976; Levine et al., 1987; McCaffery et al., 2009; Mueller, 1998;Pfäffle, 2006; Pietiläinen et al., 2002; Sammalisto, 2008; Schell and Knutsen, 2002; Silventoinen, 2003;Silventoinen et al., 2003, 2007; Van Dommelen et al., 2004; Vogler et al., 1995).

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
8 M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
The study of heritability amongst adults, conducted by Lettre (2009), revealed that most differencesin body height are controlled genetically. Although body height is a classic polygenic trait studiedfor more than 100 years, the genetic factors responsible for its variability remained unknown until2007. Identification of genes which control body height markedly improved our understanding of thedeterminants of this trait. Studies of genome-wide associations become potent tool enabling identi-fication of genes that determine complex quantitative traits. Recent meta-analyses of genome-wideassociations identified 47 loci responsible for the variability of body height (Lettre, 2009; Zhao et al.,2010).
The proportional involvement of genetic and environmental factors in determining the variance of agiven trait changes with age of examined individuals (Oniszczenko, 2005). According to the literature,the heritability coefficient of body weight increases throughout consecutive stages of development.Mook-Kanamori et al. (2012) studied the heritability of body weight during intrauterine life and earlychildhood in a group of 33 thousand twins. They observed that between the second and third trimester,the heritability of body weight in a fetus from twin pregnancy increased from 17% to 27%. It equaled29% at birth, increasing to 71% after 36 months of life. However, in another study the heritability ofbirth weight determined in a group of neonates was low (between 6.4% and 8.7% in boys, and between4.8% and 7.9% in girls). Nevertheless, the values of this trait increased with age; at the age of 5 months,irrespective of sex, the heritability explained nearly half of the variance in body weight (Dubois et al.,2012).
The analysis of more than two thousand twins from Great Britain conducted by Johnson andcolleagues (2011) showed that the heritability of body weight ranges from only 38% at birth andat 3 months of age, to 62% at the age of 6 months. The results of this study suggest that the environ-mentally determined variability of birth weight results mostly from the influence of factors uniquefor each neonate. A similar pattern was documented in the study of Norwegian families, in which theinfluence of shared and unique environmental factors on birth weight was estimated at 15% and 32%,respectively (Lunde et al., 2007). However, the effect of unique environmental factors on birth weightcan be stronger due to the influence of placental characteristics determining the transfer of nutrients(including the type of placenta, the occurrence of placental fusion, and central or eccentric insertion ofthe umbilical cord), despite the same genome and placenta in MZ twins (Van Baal and Boomsma, 1998;Loos et al., 2005). Adjustment of the outcome of twin analysis for individual variability of placentalfactors was reflected by higher degree of birth weight heritability (Gielen et al., 2008).
Our study revealed that heritability of body weight is high. Probably, this resulted from properselection of analyzed material. After excluding the cases of TTTS, all the studied MZ twins were exposedto the same intrauterine conditions. Therefore, our material met the assumptions allowing for properassessment of the role of genetic factors and environmental influences in the variability of phenotypictraits during the perinatal period. Our previous studies revealed that the analysis of same-sex pairsof DZ twins and MZ twins from mono- and dichorionic pregnancies, free from TTTS, undoubtedlyprovides the comparability of intrauterine environment conditions and enables accurate analysis ofintrapair differences (Waszak, 2013). Moreover, high level of heritability can be a consequence ofgenotype–environment interactions and correlations, which increase a widely understood geneticvariance.
The results of quantitative genetic studies based on a single measurement of body weight betweenbirth and reaching the age of 3 months suggest that the variability of body weight is mostly determinedby environmental factors; genetic factors explain only 20–44% of variability in this trait (Beardsallet al., 2009; Dubois et al., 2007; Gielen et al., 2008; Levine et al., 1987; Lunde et al., 2007; Pietiläinenet al., 2002; Vlietinck et al., 1989; Whitfield et al., 2001). In contrast, genes play a greater role indetermining the variability of body weight in neonates aged 5 months and older (66–90% heritability)(Demerath et al., 2007; Dubois et al., 2007; Levine et al., 1987). A sudden change of developmentalcharacteristics occurs between birth and completing the third month of age. As a result, body weightdetermined at a single time point of this period cannot reflect the size of a newborn as accurately asat later developmental stages, when the rate of growth stabilizes (Howe et al., 2009). Lower reliabilityis reflected by the underestimated influence of heritability and shared environmental factors and bythe overestimated influence of unique environmental effects; this may lead to apparent decrease inthe heritability of body weight during early life.

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx 9
The influence of genetic and environmental factors on the variability of body weight in twins isdetermined not only by their age but also by sex (Silventoinen et al., 2001). Recently, Dubois et al.(2012:1) reported that the effect of genetics is low at birth and increases no earlier than after the fifthmonth of life: “Genetics appear to play an increasingly important role in explaining the variation inweight, height, and BMI from early childhood to late adolescence, particularly in boys”. The valuesof heritability coefficients (h2) for birth weight determined in a sample of twins suggest that geneticfactors play an important role as a cause of individual variability of this trait, and are higher in malenewborns.
It should be remembered that heritability is a statistical parameter, describing the pattern of geneticdifferences observed between individuals from a given population, studied at a given point of time.Therefore, both genetic effects and estimated heritability can be different if studied in another sampleor at a different time.
Summary
Analysis of the coefficients of genetic determination revealed that all somatic traits analyzed in ourstudy were characterized by relatively high heritability. Probably, subtracting the VMZ variance fromthe VDZ variance produced an overestimate of genetic variance, from which the genetic componentwas not entirely excluded. During terminal stages of fetal ontogeny, the rDZ coefficients are usuallylower than the theoretically expected values, thus being lower than 0.5. Consequently, the heritabilitycoefficients which are based on rMZ − rDZ difference can be falsely overestimated. This seems to resultfrom the occurrence of genotype–environment interactions and correlations, which increased a widelyunderstood genetic variance and the related heritability measures.
Interpreting coefficients of heritability, one should be aware that the inconsistency with theassumptions of twin method can “deform” their values. Probably, the high values of heritabilitycoefficients documented in our study were, to a certain extent, associated with the inconsistencywith the assumptions of twin method.
The results of our study put into question the assumption of the additive effect of genetic andenvironmental influences as too much of a simplification. Analyzing the phenotypic variance of atrait, one cannot ignore the role of genotype–environment interactions and covariance, as these arethe effects which increase the genetic variance and related coefficients of heritability.
Acknowledgement
Grant sponsor: Polish Ministry of Science and Higher Education, grant NN303457238.
References
Allen, G., 1979. Holzinger’s Hc revised. Acta Genet. Med. Gemellol. 28, 161–164.Beardsall, K., Ong, K.K., Murphy, N., Ahmed, M.L., Zhao, J.H., Peeters, M.W., Dunger, D.B., 2009. Heritability of childhood weight
gain from birth and risk markers for adult metabolic disease in prepubertal twins. J. Clin. Endocrinol. Metab. 94, 3708–3713.Bell, J.T., Saffery, R., 2012. The value of twins in epigenetic epidemiology. Int. J. Epidemiol. 41, 140–150.Bell, J.T., Spector, T.D., 2011. A twin approach to unraveling epigenetics. Trends Genet. 27, 116–125.Bergman, P., 1987. The problem of genetic determination of growth at adolescence. Mater. Pr. Antropol. 108, 165–216.Bergman, P., Sawicki, K., 1988. Zarys metody blizniat. Mater. Pr. Antropol. 108, 11–50.Bergman, P., Goracy, M., Sawicki, K., 1983. Unterschiede der Körperhohe und Körpergewichts bei Zwilllingen aus günstigen und
ungünstigen Lebensbedingungen. HOMO – J. Comp. Hum. Biol. 34, 99–104.Bergman, P., Gronkiewicz, L., Lebioda, H., 1987. Genetic and environmental conditioning of growth of some head diameters.
Mater. Pr. Antropol. 108, 217–237.Boziłow, W., Sawicki, K., Poradnik, E., Kurlej, W., Gworys, B., 1992. Zmiennosc masy ciała podczas rozwoju prenatalnego i
okołoporodowego. Prz. Antropol. 55, 45–55.Chumlea, W.C., San Guo, S., 2002. The assessment of human growth. In: Cameron, N. (Ed.), Human Growth and Development.
Elsevier Science, USA, pp. 349–361.Demerath, E.W., Choh, A.C., Czerwinski, S.A., Lee, M., Sun, S.S., Chumlea, W.C., Duren, D., Sherwood, R.J., Blangero, J., Towne,
B., Siervogel, R.M., 2007. Genetic and environmental influences on infant weight and weight change: the Fels longitudinalstudy. Am. J. Hum. Biol. 19, 692–702.
Dubois, L., Girard, M., Girard, A., Tremblay, R., Boivin, M., Pérusse, D., 2007. Genetic and environmental influences on body sizein early childhood: a twin birth-cohort study. Twin Res. Hum. Genet. 10, 479–485.

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
10 M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx
Dubois, L., Ohm Kyvik, K., Girard, M., Tatone-Tokuda, F., Pérusse, D., Hjelmborg, J., Skytthe, A., Rasmussen, F., Wright, M.J.,Lichtenstein, P., Martin, N.G., 2012. Genetic and environmental contributions to weight, height, and BMI from birth to 19years of age: an international study of over 12,000 twin pairs. PLoS ONE 7, e30153.
Falconer, D.S., 1974. Dziedziczenie cech ilosciowych. PWN, Warszawa.Falconer, D.S., Mackay, T.F.C., 1996. Introduction to Quantitative Genetics, 4th ed. Longmans Green, Harlow, Essex, UK.Gielen, M., Lindsey, P.J., Derom, C., Smeets, H.J., Souren, N.Y., Paulussen, A., Derom, R., Nijhuis, J.G., 2008. Modeling genetic and
environmental factors to increase heritability and ease the identification of candidate genes for birth weight: a twin study.Behav. Genet. 38, 44–54.
Hanisch, D., Dittmar, M., Höhler, T., Alt, K.W., 2004. Contribution of genetic and environmental factors to variation in bodycompartments – a twin study in adults. Anthropol. Anz. 62, 51–60.
Howe, L.D., Tilling, K., Lawlor, D.A., 2009. Accuracy of height and weight data from child health records. Arch. Dis. Child. 94,950–954.
Ijzerman, R.G., Boomsma, D.I., Stehouwer, C.D.A., 2005. Intrauterine environmental and genetic influences on the associationbetween birthweight and cardiovascular risk factors: studies in twins as a means of testing the fetal origins hypothesis.Paediatr. Perinat. Epidemiol. 19, 10–14.
Johnson, L., Llewellyn, C.H., van Jaarsveld, C.H.M., Cole, T.J., Wardle, J., 2011. Genetic and environmental influences on infantgrowth: prospective analysis of the Gemini twin birth cohort. PLoS ONE 6, e19918.
Johnston, F.E., Wainer, H., Thissen, D., Macvean, R.B., 1976. Hereditary and environmental determinants of growth in height ina longitudinal sample of children and youth of Guatemalan and European ancestry. Am. J. Phys. Anthropol. 44, 469–475.
Kang, K.W., Christian, J.C., Norton, J.A., 1978. Haritability estimates from twin studies. I. Formuale of heritability estimates. ActaGenet. Med. Gemellol. 27, 39–44.
Lettre, G., 2009. Genetic regulation of adult stature. Curr. Opin. Pediatr. 21, 515–522.Levine, R.S., Hennekens, C.H., Jesse, M.J., 1987. Genetic variance of weight and length in infant twins. Am. J. Epidemiol. 126,
929–935.Loos, R.J., Derom, C., Derom, R., Vlietinck, R., 2005. Determinants of birthweight and intrauterine growth in liveborn twins.
Paediatr. Perinat. Epidemiol. 19, 115–122.Lunde, A., Melve, K.K., Gjessing, H.K., Skjaerven, R., Irgens, L.M., 2007. Genetic and environmental influences on birth weight,
birth length, head circumference, and gestational age by use of population-based parent-offspring data. Am. J. Epidemiol.165, 734–741.
Martin, R., 1988. Anthropologie. Handbuch der Vergleichenden Biologie des Menchen. Gustav Fischer Verlag, Stuttgart/NewYork.
McCaffery, J.M., Papandonatos, G.D., Bond, D.S., Lyons, M.J., Wing, R.R., 2009. Gene X environment interaction of vigorous exerciseand body mass index among male Vietnam-era twins. Am. J. Clin. Nutr. 89, 1011–1018.
Mook-Kanamori, D.O., van Beijsterveldt, C.E., Steegers, E.A., Aulchenko, Y.S., Raat, H., Hofman, Eilers, P.H., Boomsma, D.I., Jaddoe,V.W., 2012. Heritability estimates of body size in fetal life and early childhood. PLoS ONE 7, e39901.
Mueller, W.H., 1998. Genetic and environmental influences on fetal growth. In: Ulijaszek, S.J., Johnston, F.E., Preece, M.A. (Eds.),The Cambridge Encyclopedia of Human Growth and Development. Cambridge University Press, Cambridge, pp. 133–136.
Oniszczenko, W., 2002. Geny, srodowisko a zachowanie. PWN, Warszawa.Oniszczenko, W., 2005. Genetyczne podstawy ludzkich zachowan. GWP, Gdansk.Pfäffle, R., 2006. Genetic of growth in the normal child. Eur. J. Epidemiol. 155, 27–33.Pietiläinen, K.H., Kaprio, J., Räsänen, M., Rissanen, Rose, R.J., 2002. Genetic and environmental influences on the tracking of body
size from birth to early adulthood. Obesity 10, 875–884.Pinker, S., 2004. Why nature and nurture won’t go away. Daedalus 133, 5–17, http://dx.doi.org/10.1162/0011526042365591.Ridley, M., 2003. Nature Via Nurture: Genes, Experience, and What Makes Us Human. Harper Collins, New York.Rutter, M., 2006. Genes and Behavior: Nature–Nurture Interplay Explained. Blackwell, Oxford, pp. 101–113.Sammalisto, S., (Academic Dissertation) 2008. Search for genetic variants influencing human height. Medical Faculty, University
of Helsinki.Schell, L.M., Knutsen, K.L., 2002. Environmental effects on growth. In: Cameron, N. (Ed.), Human Growth and Development.
Elsevier Science, USA, pp. 165–196.Schousboe, K., Willemsen, G., Kyvik, K.O., Mortensen, J., Boomsma, D.I., Cornes, B.K., Davis, C.J., Fagnani, C., Hjelmborg, J., Kaprio,
J., De Lange, M., Luciano, M., Martin, N.G., Pedersen, N., Pietiläinen, K.H., Rissanen, A., Saarni, S., Sørensen, T.I., Van Baal, G.C.,Harris, J.R., 2003. Sex differences in heritability of BMI: a comparative study of results from twin studies in eight countries.Twin Res. 6, 409–421.
Silventoinen, K., 2003. Determinants of variation in adult body height. J. Biosoc. Sci. 35, 263–285.Silventoinen, K., Kaprio, E., Lahelma, R.J., Viken, R.J., Rose, R.J., 2001. Sex differences in genetic and environmental factors
contribution to body height. Twin Res. 4, 25–29.Silventoinen, K., Sammalisto, S., Perola, M., Boomsma, D.I., Cornes, B.K., Davis, C., Dunkel, L., De Lange, M., Harris, J.R., Hjelmborg,
J.V.B., Luciano, M., Martin, N.G., Mortensen, J., Nistico, L., Pedersen, N.L., Skytthe, A., Spector, T.D., Stazi, M.A., Willemsen,G., Kaprio, J., 2003. Heritability of adult body height: a comparative study of twin cohorts in eight countries. Twin Res. 6,399–408.
Silventoinen, K., Bartels, M., Posthuma, D., Estourgie-Van Burk, G.F., Willemsen, G., Van Beijsterveldt, T.C.E.M., Boomsma, D.I.,2007. Genetic regulation of growth in height and weight from 3 to 12 years of age: a longitudinal study of Dutch twinchildren. Twin Res. Hum. Genet. 10, 354–363.
Silventoinen, K., Rokholm, B., Kaprio, J., Sørensen, T.I., 2010. The genetic and environmental influences on childhood obesity: asystematic review of twin and adoption studies. Int. J. Obes. (Lond.) 34, 29–40.
Skibinska, A., Skład, M., 1979. Genetyczne uwarunkowanie somatotypu Sheldona. Wych. Fizyczne Sport 23, 3–12.Van Baal, C.G., Boomsma, D.I., 1998. Etiology of individual differences in birth weight of twins as a function of maternal smoking
during pregnancy. Twin Res. 1, 123–130.Van Dommelen, P., Boomsma, D.I., De Gunst, M.C.M., van der Vaart, A.W., 2004. Genetic study of the height and weight process
during infancy. Twin Res. 7, 607–616.

Please cite this article in press as: Waszak, M., et al., Heritability estimates for somatictraits determined perinatally with the twin method. HOMO - J. Comp. Hum. Biol. (2015),http://dx.doi.org/10.1016/j.jchb.2015.04.001
ARTICLE IN PRESSG ModelJCHB-25389; No. of Pages 11
M. Waszak et al. / HOMO - Journal of Comparative Human Biology xxx (2015) xxx–xxx 11
Vlietinck, R., Derom, R., Neale, M.C., Maes, H., van Loon, H., Derom, C., Thiery, M., 1989. Genetic and environmental variation inthe birth weight of twins. Behav. Genet. 19, 151–161.
Vogler, G.P., Sørensen, T.I., Stunkard, A.J., Srinivasan, M.R., Rao, D.C., 1995. Influences of genes and shared family environmenton adult body mass index assessed in an adoption study by a comprehensive path model. Int. J. Obes. Relat. Metab. Disord.19, 40–45.
Waszak, M., 2013. Wykorzystanie metody blizniat do okreslenia udziału czynników genetycznych i srodowiskowych w zmi-ennosci cech ilosciowych w okresie okołourodzeniowym. Monografie, Seria, vol. 422. AWF, Poznan, pp. 98–107.
Whitfield, J.B., Treloar, S.A., Zhu, G., Martin, N.G., 2001. Genetic and non-genetic factors affecting birth-weight and adult BodyMass Index. Twin Res. 4, 365–370.
Zhao, J., Li, M., Bradfield, J.P., Zhang, H., Mentch, F.D., Wang, K., Sleiman, P.M., Kim, C.E., Glessner, J.T., Hou, C., Keating, B.J.,Thomas, K.A., Garris, M.L., Deliard, S., Frackelton, E.C., Otieno, F.G., Chiavacci, R.M., Berkowitz, R.I., Hakonarson, H., Grant,S.F., 2010. The role of height-associated loci identified in genome wide association studies in the determination of pediatricstature. BMC Med. Genet. 11, 96, http://dx.doi.org/10.1186/1471-2350-11-96.





























