Embed Size (px)
Citation preview

Int. J. Devl Neuroscience 21 (2003) 309–320
High affinity neurotrophin receptors in the human pre-termnewborn, infant, and adult cerebellum
Marina Quartua, Maria Pina Serraa, Annalisa Mancaa, Paolo Follesab,Rossano Ambua, Marina Del Fiaccoa,∗
a Department of Cytomorphology, University of Cagliari, Cittadella Universitaria di Monserrato, 09042 Monserrato, Italyb Department of Experimental Biology ‘Bernardo Loddo’, University of Cagliari, Cittadella Universitaria di Monserrato, 09042 Monserrato, Italy
Received 17 February 2003; received in revised form 2 June 2003; accepted 4 June 2003
Abstract
The immunohistochemical occurrence of the high affinity neurotrophin (NT) receptors trkA, trkB, and trkC is shown in the pre-termnewborn, infant, and adult human post-mortem cerebellum. Immunoreactive neuronal perikarya and processes were observed in allspecimens examined, where they appeared unevenly distributed in the cerebellar cortical layers and deep nuclei, and showed regionaldifferences among cerebellar lobules and folia. The trk receptor-antibodies, tested by Western blot on human cerebellum homogenates,revealed multiple immunoreactive bands for trkA and single bands for trkB and trkC. The results obtained show the tissue localization ofthe trk receptor-like immunoreactivity in the human cerebellum from prenatal to adult age. The analysis for codistribution of the receptorswith the relevant ligand and among the receptors in discrete cortical and deep nuclei tissue fields shows a wide variety of conditions,from a good similarity in terms of type and density of labeled structures, to a lack of correspondence, and suggests the possibility ofcolocalization of trk receptors with the relevant neurotrophin and among them in the cerebellar cortex. These results sustain the conceptthat the neurotrophin trophic system participates in the development, differentiation, and maintenance of the human cerebellar connectivityand support the possibility of a multifactorial trophic support for the neurotrophins through target-derived and local mechanisms.© 2003 ISDN. Published by Elsevier Ltd. All rights reserved.
Keywords: trkA, trkB, and trkC receptors; Neurotrophins; Newborn, infant, and adult cerebellum; Man; Immunohistochemistry; Western blot
1. Introduction
The trk family of protein-tyrosine kinases consists of threetransmembrane receptors, known as trkA, trkB, and trkC,each of which interacts specifically, but not exclusively, withdifferent members of the neurotrophin (NT) family and me-diates their effects on distinct and partially overlapping setsof central and peripheral neurons (Bothwell, 1991; Chao andHempstead, 1995). trkA is the high affinity receptor for nervegrowth factor (NGF), trkB is the one for both brain-derivedneurotrophic factor (BDNF) and neurotrophin-4 (NT-4),and trkC is the preferred receptor for neurotrophin-3 (NT-3)(Bothwell, 1991, 1995; Soppet et al., 1991; Chao andHempstead, 1995; Neet and Campenot, 2001). Binding ofneurotrophins to their relevant trk receptors results in bio-logical activity spanning from regulation of survival, prolif-eration and differentiation in development to maintenance,modulation of synaptic efficacy, and repair in mature neu-
∗ Corresponding author. Tel.:+39-070-6754080;fax: +39-070-6754003.
E-mail address: [email protected] (M. Del Fiacco).
rons (Bothwell, 1995; Thoenen, 1995; Lewin and Barde,1996). The altered expression of the NTs or their receptorsin several neuropathological conditions, such as a variety ofneurodegenerative diseases, peripheral neuropathies, painstates and neoplastic syndromes, provides grounds for theiruse as therapeutic tools (Apfel, 1999; Siegel and Chauhan,2000; seeSaragovi and Gehring, 2000). A rich literatureshows that NTs (Hofer et al., 1990; Maisonpierre et al.,1990; Ernfors et al., 1992; Rocamora et al., 1993; Timmusket al., 1993; Das et al., 2001) and their receptors (Pioroand Cuello, 1988; Klein et al., 1989; Wanaka and Johnson,1990; Lamballe et al., 1991; Mufson et al., 1991; Segalet al., 1992; Merlio et al., 1992; Rocamora et al., 1993;Ohira et al., 1999) are widely expressed in the developingand adult cerebellum of experimental animals. Most of theresults relating to cellular actions of the NTs in the cere-bellum are derived from studies on primary cultures of ratgranule cells, in which NTs have been found to sustainsurvival, prevent apoptosis, promote neurite extension andprotect partially against glutamate-induced neurotoxicity(Segal et al., 1992, 1997; Lindholm et al., 1993; Gao et al.,1995; Kubo et al., 1995; Tanaka et al., 2000). Effects of
0736-5748/$30.00 © 2003 ISDN. Published by Elsevier Ltd. All rights reserved.doi:10.1016/S0736-5748(03)00086-8

310 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
exogenously applied NTs on survival and/or differentiationof other cerebellar neuronal types have also been reported(Lärkfors et al., 1996; Mertz et al., 2000). Gene-targetingstudies have shown that NT-3 (Bates et al., 1999) andBDNF (Schwartz et al., 1997; Borghesani et al., 2002;Carter et al., 2002) are involved in normal cerebellar devel-opment and foliation, whereas a massive death of cerebellargranule neurons occurs in double trkB and trkC mutantmice (Minichiello and Klein, 1996), and trkB is requiredfor the development of GABA-ergic cerebellar neurons(Rico et al., 2002). Studies on organotypic slice cultures ofrat cerebellum (Segal et al., 1992) and in situ hybridizationand immunohistochemical analyses (Rocamora et al., 1993;Das et al., 2001) indicate that BDNF and NT-3 may affectdeveloping granule cells at distinct stages of differentiation,suggesting a sequential influence, critical for a coordinatedmaturation of the cerebellar circuitry. In addition, severallines of evidence indicate that some NTs have a role in pro-tection (Tong and Perez-Polo, 1998) and repair of cerebellarneurons and connectivity (Skaper et al., 1998), and exert atrophic influence on cerebellar afferent neurons (Shibayamaet al., 1998; Rabacchi et al., 1999; Sherrard and Bower,2001). Altered levels of NTs and/or trk receptors have beenreported in experimental pathological states. Thus, reducedlevels of trkB and BDNF have been correlated to impairedcerebellar development after early postnatal ethanol expo-sure (Heaton et al., 1999; Light et al., 2001, 2002) and ininherited neurological disorders (Qiao et al., 1996, 1998;Bao et al., 1998; Wullner et al., 1998). Interestingly, anincreased trkB immunoreactivity has been found in thegranular layer of patients with the Macado–Joseph disease(Kumada et al., 2000), and reduced levels of NT-4 havebeen reported in the cerebellum of Alzheimer’s disease pa-tients (Hock et al., 2000), whereas the expression of trkCreceptor is associated with a more favorable life expectancyin medulloblastoma (Goumnerova, 1996; Pomeroy et al.,1997; Eberhart et al., 2001).
The relevance of NTs activity in the correct developmentand function of the cerebellum contrasts with the paucity ofinformation regarding NT responsive neuronal populations
Table 1List of specimens
Case Sex Cause of death Post-mortemdelay (h)
Regions examined Method
1 Pre-term newborn 26 d (24 w.g.) F Cardio-respiratory failure 48 Whole cerebellum IHC2 Pre-term newborn 3 d (30 w.g.) F Cardio-respiratory failure 28 Temporal cortex WB3 Pre-term newborn 1 d (32 w.g.) M Cardio-respiratory failure 43 Hemisphere WB4 Pre-term newborn 22 d (33 w.g.) F Cardio-respiratory failure 28 Vermis and adjacent hemispheres, deep nuclei IHC5 Full-term newborn 8 m M Pneumonitis 45 Vermis IHC6 Adult 43 y F Myocardial infarction 34 Temporal cortex WB7 Adult 47 y F Myocardial infarction 45 Posterior lobe of the hemisphere WB8 Adult 51 y M Cardio-respiratory failure 42 Posterior lobe of the hemisphere, deep nuclei IHC9 Adult 52 y M Myocardial infarction 45 Posterior lobe of the hemisphere, deep nuclei IHC
10 Adult 56 y F Pneumonitis 34 Posterior lobe of the hemisphere, deep nuclei IHC11 Adult 79 y F Cardio-respiratory failure 42 Posterior lobe of the hemisphere, deep nuclei IHC
d: days; F: female; m: months; M: male; w.g.: weeks of gestation; y: years.
in the human cerebellum, and the possible imbalances ofNTs and their relevant receptors in pathology. Occurrenceof trk receptors in human cerebellum has been substantiatedby northern blot and RT-PCR studies (Segal et al., 1995;Pomeroy et al., 1997; Hock et al., 1998; Dubus et al., 2000).However, information on their tissue distribution is eithernon selective for each molecule (Muragaki et al., 1995;Chen et al., 1996) or limited to some of them (Allen et al.,1994; Benisty et al., 1998; Savaskan et al., 2000; Kumadaet al., 2000) and controversial (Savaskan et al., 2000;Kumada et al., 2000). We have recently reported on thetissue localization of NGF, BDNF, NT-4, and NT-3 in thehuman post-mortem cerebellum at pre-, perinatal and adultage (Quartu et al., 2003). This investigation probes thesame specimens examined in that study for the immuno-histochemical occurrence of the high affinity NT receptorstrkA, trkB and trkC, and reports observations on the codis-tribution of trk receptors with the relevant NT ligand andamong trk receptors.
2. Materials and methods
Autopsy specimens of human cerebellum were obtainedfrom subjects of different ages, with no signs of neu-ropathology (Table 1). The sampling and handling of humanspecimens has been conducted under the appropriate ethi-cal safeguards and protocols of the Section of PathologicalAnatomy, Department of Cytomorphology, University ofCagliari. Cerebellar samples comprised the whole cerebel-lum in case 1, the vermis in cases 4 and 5, and part of thehemispheral posterior lobe, including also deep nuclei, incases 8–11. The flocculo-nodular lobe was absent in allspecimens examined.
2.1. Western blot
Human autoptic tissue blocks of cerebellum and cere-bral temporal cortex from pre-term newborns (cases 3 and2) and adult subjects (cases 7 and 6) were collected and

M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320 311
stored at−80◦C until required. Tissue homogenates wereprepared by homogenization in 50 mM Tris–HCl, pH 7.5,centrifuged at 10,000× g for 20 min, and resuspended in3 ml of Tris–HCl, pH 7.5. Protein concentrations were de-termined using the Lowry method of protein assay (Lowryet al., 1951) with bovine serum albumin as standard. Pro-teins for each tissue homogenate (40�g) diluted 1:1 inloading buffer were heated to 95◦C for 3 min and separatedby SDS-polyacrilamide gel electrophoresis (SDS-PAGE)using a 10% (w/v) polyacrylamide resolving gel. Internalmolecular weight standards (Kaleidoscope Prestained Stan-dards, Bio-Rad, Hercules, CA, USA) were run in parallel.Two gels at a time were run for Coomassie staining andimmunoblotting, respectively. Proteins for immunoblottingwere electrophoretically transferred on a polyvinylidenefluoride membrane (Biorad) using the Mini Trans Blot Cell(Biorad). Blots were blocked by immersion in 20 mM Trisbase and 137 mM sodium chloride (TBS) containing 5%milk powder and 0.05% Tween 20 (TBS-T), for 60 min atroom temperature, and incubated overnight at 4◦C with thesame rabbit polyclonal antisera against trk receptors usedfor the immunohistochemical analysis, all diluted 1:100 inTBS containing 5% milk powder and 5% MOP (BoehringerMannheim). After TBS-T rinse, blots were incubated for60 min, at room temperature, with a peroxidase-conjugatedanti-rabbit serum (Amersham Corp.), diluted 1:10,000. Af-ter TBS-T rinse, protein bands were visualized on a film(Kodak X-Omat LS, Kodak, Rochester, NY) using the ECLmethod (Amersham Corp.). In order to control for non spe-cific staining, blots were stripped and incubated with therelevant secondary antiserum.
2.2. Immunohistochemistry
Fixation in 4% freshly prepared phosphate-bufferedformaldehyde, pH 7.3, for 4–6 h at 4◦C was followedby overnight rinsing in 0.1 M phosphate buffer (PB),pH 7.3, containing 5–30% sucrose. Cryostat sections of10�m, cut in frontal plane from specimen of case 1 andin sagittal planes from the remainders, were collected onchrome alum-gelatin coated slides, and processed by theavidin–biotin–peroxidase complex (ABC) immunohisto-chemical technique. To detect the different trk receptors,rabbit polyclonal antisera directed against the residues763–777, 794–808, 798–812 of trkA of human origin, trkBof mouse origin (precursor form), and trkC of mouse origin,respectively (SantaCruz Biotechnology, USA), were usedas primary antiserum at dilution 1:400. For the analysis ofcodistribution, adjacent sections were incubated with rab-bit polyclonal antisera directed against the residues 1–20,128–147, 80–99, 139–158 of NGF (precursor form), BDNF,NT-4 (precursor form), and NT-3 (precursor form) of hu-man origin, respectively (SantaCruz Biotechnology, USA),all diluted 1:400. Biotin-conjugated goat anti-rabbit serum(Vector), diluted 1:200, was used as secondary antiserum.The reaction product was revealed with ABC (BioSpa Div.),
diluted 1:250, followed by incubation with a solution of0.1 M PB, pH 7.3, containing 0.05% 3-3′-diaminobenzidine(Sigma), 0.04% nickel ammonium sulphate, and 0.01% hy-drogen peroxide. Incubations with primary antiserum werecarried out overnight at 4◦C. Incubations with secondaryantiserum and ABC lasted 60 and 30 min, respectively, andwere performed at room temperature. All antisera and theABC were diluted in phosphate-buffered saline containing0.2% Triton X-100 (PBS-T). Control preparations were runin parallel by incubating tissue sections with either PBS-Talone or with the relevant primary antiserum preabsorbedwith the corresponding peptide antigen (SantaCruz Biotech-nology, USA). Slides were dehydrated, cover-slipped, andobserved with a Leitz Dialux 20 microscope. Some ofthe immunostained sections were stained with modifiedMayer’s hematoxylin for assistance in defining the anatomicboundaries of the cortical layers.
3. Results
3.1. Western blot
As shown inFig. 1, the trkA antibody detected multiplebands in both the cerebellar (lanes a and b) and temporal(lanes c and d) cortical tissue homogenates; one of them, atapproximately 165 kDa, occurs in pre-term cerebellum andpre-term and adult temporal cortex; two or three additionalbands of molecular weight (Mw) falling within a range be-tween 80 and 44 kDa occur in all lanes. In the cerebellum,these are placed at two levels at about 75–80 and 44 kDa,respectively. The anti-trkB serum labeled a single band atapproximately 80 kDa in both cerebellar and cerebral cor-tical tissue homogenates, at all examined ages. The trkCantiserum detected a single immunoreactive band at about80 kDa in the cerebellar and cerebral cortical homogenatesof the pre-term newborns, whereas it failed to recognize anyband in the adult tissue homogenates.
3.2. Immunohistochemistry
Immunoreactive material for trkA, trkB, and trkC recep-tors was detectable in all specimens examined (Figs. 2–4).Control preparations resulted in negative staining. Morpho-logically, the immunostained structures appeared as labeledcell bodies, dendritic processes, and thread-like and punctateelements, interpreted as nerve fibers and terminals, respec-tively. The immunoreactivity for each trk receptor showedan uneven distribution in the cerebellar cortex and deep nu-clei, with regional differences, in terms of density of labeledstructures, among cerebellar lobules and folia. Because ofthe small number of cases we could examine, most of whichconsisting in parts of the organ, we were often unable to as-cribe precisely the immunoreactive elements to distinct lob-ules and folia of the cerebellum. As a whole, for each trkreceptor the labeling appeared to persist in cortical layers

312 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
Fig. 1. trkA, trkB and trkC Western blot analysis of cerebellar (lanes aand b) and cerebral temporal cortex (lanes c and d) tissue homogenate(protein concentration: 60�g/20�l) from human pre-term newborn (case3, lane a and case 2, lane c) and adult samples (case 7, lane b and case6, lane d).
and deep nuclei throughout life.Table 2summarizes the oc-currence and localization of trkA-, trkB-, and trkC-like im-munoreactive (LI) neuronal perikarya and fibers.
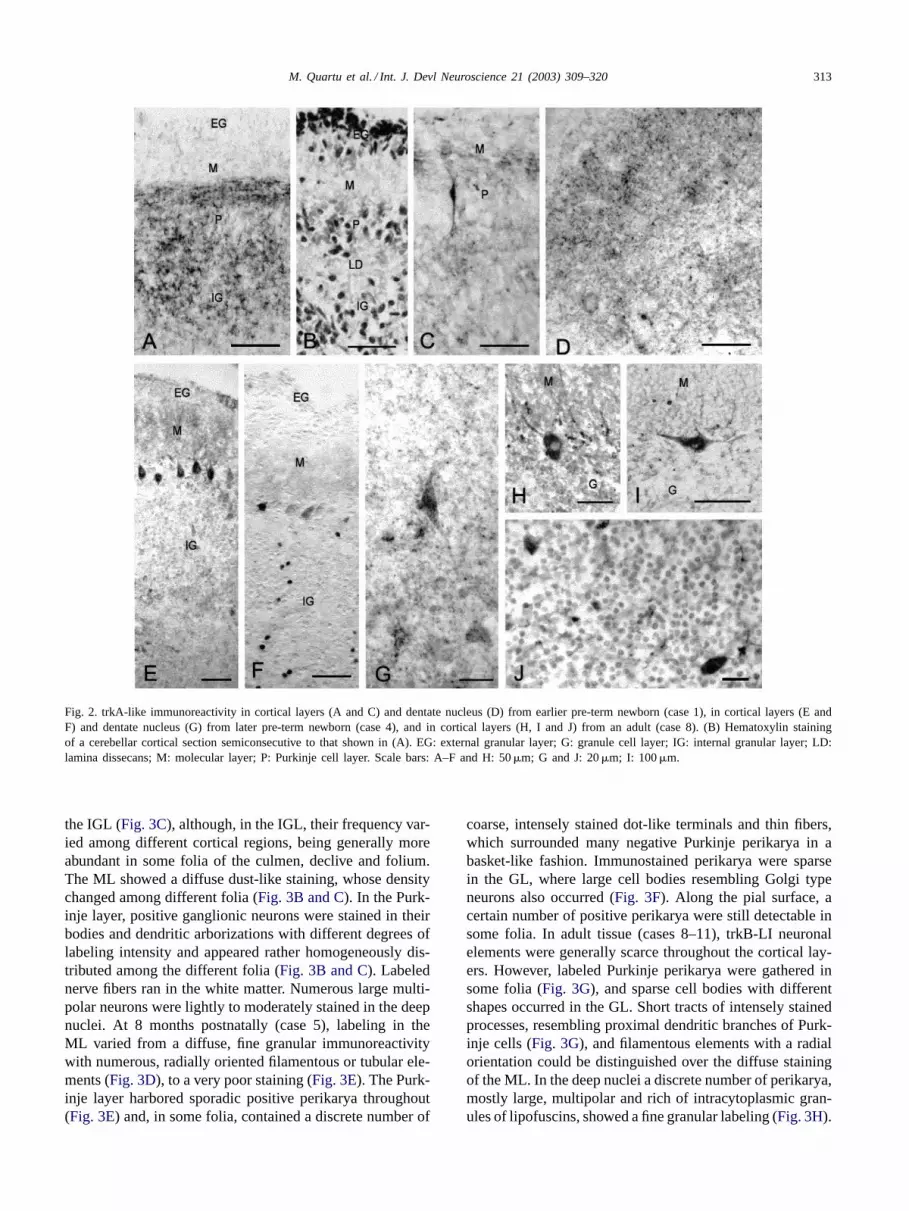
3.2.1. trkA (Fig. 2)In the earlier pre-term newborn (case 1) trkA-like im-
munoreactivity labeled a certain number of neuronal ele-ments unevenly distributed throughout the cortical layers,whose density was higher in the cortex of the vermis(Fig. 2A) as compared to that of the lateral hemisphere(Fig. 2C). As a general rule, a poor staining occurred in theexternal granular layer (EGL), wherein some positive cellswere distinguishable. In the vermal cortex, punctate andfiber-like structures were frequent; they ran parallel to thepial surface in the inner part of the molecular layer (ML)and formed loose networks in the lamina dissecans (LD) andin the internal granular layer (IGL) (Fig. 2A). By contrast,in the hemispheral cortex only sparse dot-like terminals andthin varicose fibers were observed (Fig. 2C). Rare positivefusiform neurons, resembling immature Purkinje cells, weredetected in the Purkinje layer (Fig. 2C). trkA-LI fibers of
different caliber and direction were present in the white mat-ter, being particularly appreciable medially to the dentatenucleus. In the latter, a few lightly labeled cell bodies, pref-erentially located along its peripheral borders, and punctateimmunoreactivity of moderate intensity could be observed(Fig. 2D). At later stages of gestation (case 4), positive neu-ronal cell bodies in the cortical layers increased in number.A few labeled neurons were unevenly distributed withinthe EGL, and a fine, dust-like trkA-LI material occurred inthe ML. Labeled Purkinje neuronal perikarya occurred withdifferent frequency among different folia (Fig. 2E and F).Positive granule cells were detectable in the IGL (Fig. 2Eand F). Sparse neuronal cell bodies and punctate elementswere detected in the deep cerebellar nuclei (Fig. 2G). In theinfant (case 5), labeled cell bodies could be seen over a fair,dot-like immunoreactivity within the ML, where positivenerve fibers with the distinctive morphology of parallel fiberscould be observed occasionally. In the Purkinje layer, sparseperikarya were labeled only in some lobules such as theuvula, culmen and central lobule. Coarse punctate elementsand positive fibers organized in a basket-like fashion aroundthe soma of negative Purkinje cells could be observed often.Labeled granule neurons, perikarya with different shapes,sometimes reminishent of Golgi type neurons, and frequentcoarse punctate elements were detectable in the granule celllayer (GL). In the adult tissue (cases 8–11), trkA-LI neuronalelements had a distribution pattern generally similar to thatobserved in the infant. In some folia, occasional neuronalcell bodies and numerous labeled filaments with a radial ori-entation could be detected in the ML. In the Purkinje layer, anumber of ganglionic perikarya (Fig. 2H) and rare horizon-tally extended cell bodies, resembling Lugaro cells (Fig. 2I),were detectable. Filamentous and coarse terminal-like ele-ments occurred within the Purkinje cell layer (Fig. 2H),sometimes outlining a basket-like mesh around the somaof negative Purkinje cells. Small positive cell bodies withthe aspect of granule cells and larger perikarya of variousshapes were scattered throughout the GL (Fig. 2J). Coarsepunctate elements also occurred with varied density in dif-ferent fields (cfrFigs. 2J and 6A). In the deep nuclei, anumber of lightly stained multipolar neurons together withfine positive dots and short tracts of positive fibers weredetectable.
3.2.2. trkB (Fig. 3)In the earlier pre-term newborn (case 1), a discrete number
of labeled granule cells, many of which fusiform in shape,were detectable in the EGL and IGL (Fig. 3A). In the MLand in the LD, short fiber tracts and punctate immunore-activity could be observed over a diffuse dust-like staining(Fig. 3A). No positive perikarya were observed in the Purk-inje layer. Rare positive fibers of different caliber occurredin the white matter. In the pre-term newborn of 33 weeks ofgestation (w.g.) (case 4), labeled structures were generallymore abundant than at earlier stages of life (Fig. 3B and C).Positive neurons were detectable throughout the EGL and
M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320 313
Fig. 2. trkA-like immunoreactivity in cortical layers (A and C) and dentate nucleus (D) from earlier pre-term newborn (case 1), in cortical layers (E andF) and dentate nucleus (G) from later pre-term newborn (case 4), and in cortical layers (H, I and J) from an adult (case 8). (B) Hematoxylin stainingof a cerebellar cortical section semiconsecutive to that shown in (A). EG: external granular layer; G: granule cell layer; IG: internal granular layer; LD:lamina dissecans; M: molecular layer; P: Purkinje cell layer. Scale bars: A–F and H: 50�m; G and J: 20�m; I: 100�m.
the IGL (Fig. 3C), although, in the IGL, their frequency var-ied among different cortical regions, being generally moreabundant in some folia of the culmen, declive and folium.The ML showed a diffuse dust-like staining, whose densitychanged among different folia (Fig. 3B and C). In the Purk-inje layer, positive ganglionic neurons were stained in theirbodies and dendritic arborizations with different degrees oflabeling intensity and appeared rather homogeneously dis-tributed among the different folia (Fig. 3B and C). Labelednerve fibers ran in the white matter. Numerous large multi-polar neurons were lightly to moderately stained in the deepnuclei. At 8 months postnatally (case 5), labeling in theML varied from a diffuse, fine granular immunoreactivitywith numerous, radially oriented filamentous or tubular ele-ments (Fig. 3D), to a very poor staining (Fig. 3E). The Purk-inje layer harbored sporadic positive perikarya throughout(Fig. 3E) and, in some folia, contained a discrete number of
coarse, intensely stained dot-like terminals and thin fibers,which surrounded many negative Purkinje perikarya in abasket-like fashion. Immunostained perikarya were sparsein the GL, where large cell bodies resembling Golgi typeneurons also occurred (Fig. 3F). Along the pial surface, acertain number of positive perikarya were still detectable insome folia. In adult tissue (cases 8–11), trkB-LI neuronalelements were generally scarce throughout the cortical lay-ers. However, labeled Purkinje perikarya were gathered insome folia (Fig. 3G), and sparse cell bodies with differentshapes occurred in the GL. Short tracts of intensely stainedprocesses, resembling proximal dendritic branches of Purk-inje cells (Fig. 3G), and filamentous elements with a radialorientation could be distinguished over the diffuse stainingof the ML. In the deep nuclei a discrete number of perikarya,mostly large, multipolar and rich of intracytoplasmic gran-ules of lipofuscins, showed a fine granular labeling (Fig. 3H).

314 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
Fig. 3. trkB-like immunoreactivity in cortical layers (A) from earlier pre-term newborn (case 1), in cortical layers (B and C) from later pre-term newborn(case 4), in cortical layers (D and E) and dentate nucleus (F) from infant (case 5), and in cortical layers (G) and dentate nucleus (H) from an adult (case11). EG: external granular layer; G: granule cell layer; IG: internal granular layer; LD: lamina dissecans; M: molecular layer; P: Purkinje cell layer.Scale bars: A, B, C and E: 50�m; D and G: 100�m; F and H: 25�m.
3.2.3. trkC (Fig. 4)In the earlier pre-term newborn (case 1), few labeled gran-
ule cells were detectable in the EGL and IGL (Fig. 4A).Sporadic trails of varicose fibers and isolated punctate ele-ments were also observed in the IGL. A faint diffuse label-ing occurred in the ML (Fig. 4A) and LD. In the Purkinjecell layer, trkC-LI perikarya occurred frequently in somecerebellar regions (Fig. 4B). Numerous labeled perikaryaand isolated fibers were detectable in the dentate nucleus.In the later pre-term (case 4), a small number of positiveneurons occurred in the EGL and, only in some folia, in theIGL (Fig. 4C). In the latter both granule cells and neuronswith different morphology were present. In the ML a dif-fuse dust-like staining of moderate intensity was detectable(Fig. 4C). Rare Purkinje neurons were stained (Fig. 4C).A few large trkC-LI perikarya occurred in the deep nuclei(Fig. 4D). In the infant (case 5) and in adult tissue, trkC-LIstructures showed a similar distribution pattern. In the ML,
a certain number of positive cell bodies, Purkinje cell prox-imal dendrites, and radially oriented, intensely labeled thickfilaments could be detected (Fig. 4E and F). Single and smallsets of Purkinje cells were sporadically labeled throughout(Fig. 4E and F). Their frequency varied among different foliaand in the infant they were more abundant in the culmen andcentral lobule. Thin fibers and dot-like terminals were occa-sionally organized in a basket-like fashion around the somaof negative Purkinje cells. Immunostained granule cells weresparse in the GL; however, neuronal perikarya with differ-ent morphology were rather abundant. In the deep nuclei ofadult cerebellum, a discrete number of lightly labeled mul-tipolar neurons and sporadic tracts of varicose fibers weredetected (Fig. 4G).
3.2.4. Analysis of codistributionAs no successful double labeling immunostaining was
obtained with the available antisera, the analysis was

M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320 315
Fig. 4. trkC-like immunoreactivity in cortical layers (A and B) from earlier pre-term newborn (case 1), in cortical layers (C) and dentate nucleus (D)from later pre-term newborn (case 4), in cortical layers (E) from infant (case 5), and in cortical layers (F) and dentate nucleus (G) from an adult (case11). EG: external granular layer; G: granule cell layer; IG: internal granular layer; M: molecular layer; P: Purkinje cell layer. Scale bars: A: 20�m; B,C, D and G: 50�m; E and F: 100�m.
performed on adjacent sections. This hampered the possi-bility of detecting colocalization on punctate elements andsmall neurons, such as granule cells. In fact, only Purkinjeneuronal somata could be recognized in two adjacent sec-tions.Figs. 5 and 6show a number of Purkinje cells whichmay possibly contain immunoreactive material for bothtrkA and NGF (Fig. 5A and B), trkB and NT-4 (Fig. 5Cand D), and trkA and trkB (Fig. 6). However, even in theseelements it was difficult to ascertain a double labeling.Instead, this analysis provided information as to the codis-tribution of trk receptors and relevant ligands, and amongtrk receptors. In this respect, when examining discrete fieldsof the cortex and deep nuclei, a variety of conditions couldbe seen, from a good similarity in terms of type and densityof labeled elements, as noticeable inFig. 5A and Bfor trkAand NGF in cortical layers, to a less tight equivalence anda plain lack of correspondence.Fig. 6, for instance, showsa different distribution of trkA and trkB in the ML and GLin a cortical field where the two receptors are codistributed(and possibly colocalized) in the Purkinje cell layer.
4. Discussion
4.1. Western blot analysis
All trk antibodies used in this study are directed againstan intracytoplasmic C-terminal epitope of the full-lengthcatalytic isoform of the receptors. The trkA-LI bands cor-responding to aMw of about 165 kDa are likely to rep-resent the NGF-trkA receptor complex (seeMeakin andShooter, 1992), whereas those at about 75–80 kDa might rep-resent the unglycosilated polypeptide (Martin-Zanca et al.,1989). As for the trkA-LI bands of about 44 kDa, it ispossible that they represent a truncated version of trkAcontaining all or part of the C-terminal intracellular do-main, as suggested for the trkA-LI 38 kDa protein, tyrosinephosphorylated upon NGF stimulation, observed in PC12cells (Zhou et al., 1995). trkA-LI bands at about 40 and35 kDa have been detected in the human post-mortem hip-pocampus with the same antibody we used (Connor et al.,1996).

316 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
Table 2Distribution of trkA-, trkB-, and trkC-LI neurons (�) and nervous fibres (+) in the human cerebellum
External granular layer Molecular layer Purkinje layer Granular layer Deep nuclei
trkAPre-term newborn F 26 d (24 w.g.) � + � + � +Pre-term newborn F 22 d (33 w.g.) � + � � � +Full-term newborn M 8 m – � + � + � + n.d. n.d.Adult M 51 y – + � + � + + +Adult M 52 y – + � � � +Adult F 56 y – + + + � +Adult F 79 y – � + � + � � +
trkBPre-term newborn F 26 d (24 w.g.) � + �Pre-term newborn F 22 d (33 w.g.) � + � � � +Full-term newborn M 8 m – + � + � n.d. n.d.Adult M 51 y – + �Adult M 52 y – + � � +Adult F 56 y – � +Adult F 79 y – + � + � � +
trkCPre-term newborn F 26 d (24 w.g.) � + � � + � +Pre-term newborn F 22 d (33 w.g.) � + � � �Full-term newborn M 8 m – � + � n.d. n.d.Adult M 51 y – +Adult M 52 y – � + � +Adult F 56 y – � � � � +Adult F 79 y – + � + � � +
d: days; F: female; m: months; M: male; n.d.: not determined; w.g.: weeks of gestation; y: years. No quantification is given.
Fig. 5. The distribution of trkA-like immunoreactivity (A) and NGF-like immunoreactivity (B), and trkB-like immunoreactivity (C) and NT-4-likeimmunoreactivity (D) is seen in two pairs (A, B and C, D) of adjacent sections of cerebellar cortex from infant (case 4). Arrows point to possible sitesof colocalization in Purkinje cell bodies. Arrowheads point to Purkinje cell bodies labelled for only one substance. G: granule cell layer; M: molecularlayer. Scale bar: A–D: 100�m.

M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320 317
Fig. 6. The distribution of trkA-like immunoreactivity (A) and trkB-like immunoreactivity (B) is seen in two adjacent sections of cerebellar cortexfroman adult (case 11). Arrows point to a possible site of colocalization in a Purkinje cell body. G: granule cell layer; M: molecular layer. The asterisksindicate a blood vessel. Scale bar: A and B: 100�m.
The outcome of our trkB and trkC analysis shows a sin-gle band in each homogenate, whose molecular identity isuncertain. By means of trkB antibodies against an intracel-lular portion of the rat receptor molecule different from thatused as immunogen for the antibody applied in this study,several protein bands of mw around the value we obtainedhave been reported in post-synaptic densities from rat cere-bral cortex, (Aoki et al., 2000). The possibility exists thatwe are detecting truncated forms of trkB and trkC. In fact,it has been shown that in the rat postnatal cochlea the sametrkB and trkC antibodies used in this study cross-react withthe truncated trkB isoform trkB-T2 and with truncated trkCisoforms, respectively (Gestwa et al., 1999), and expressionof trkB-T2 has been reported in the rat (Light et al., 2001)and monkey cerebellum (Ohira et al., 1999). Several stud-ies have reported the expression of different forms of trkreceptor proteins (Tsoulfas et al., 1993; Barker et al., 1993;Ehlers et al., 1995; Connor et al., 1996; Aoki et al., 2000)and mRNAs (Shelton et al., 1995; Dubus et al., 2000), whichmight reflect authentic differences in the mw of these re-ceptors among different subcellular compartments, variousbrain regions, different species, and in tissue specimens ver-sus cultured cells. In the present study, the lack of trkC bandsin the adult tissue homogenates conflicts with the trkC-LIstaining obtained in the tissue sections. While it is possiblethat levels of trkC in mature cerebellar tissue were too lowto be detected by Western blotting, it should be consideredthat the conditions and milieu where antigen and antibodymay interact in the two methods are very different. This mayperhaps explain why the visualization of a given antigenby Western blot does not always reflect a satisfactory im-munohistochemical detection of it (Connor et al., 1997; DelFiacco et al., 2002) and vice versa (Nishio et al., 1994;Quartu et al., 2003).
4.2. trkA, trkB, and trkC tissue localization
This study shows the occurrence of trkA-, trkB-, andtrkC-like immunoreactivity in the pre-term newborn, infant,and adult human cerebellum, and provides information on
the codistribution among trk receptors and their ligands,thus adding to the knowledge of the organization of the NTtrophic system in the organ and supporting the concept ofits involvement in development, differentiation, and mainte-nance of the cerebellar circuitry in man. Positive structuresfor each trk receptor were observed in all specimens ex-amined. The paucity of samples tested did not allow theirprecise mapping, nor the determination of the existenceof age-related changes in their distribution and frequency.However, our observations indicate that (1) immature cellsimmunoreactive for each trk receptor occur in the EGL inthe premature newborn; (2) immunoreactivity for each trkreceptor characterizes various neuronal types of the corticallayers and deep nuclei both at developmental age and in themature tissue; (3) trk receptors and NTs show a complexpattern of codistribution in the cerebellar cortex and deepnuclei, which varies in different tissue fields. Finally, thepossibility is suggested that different trk receptors and trkreceptors and the relevant trophin colocalize in Purkinjecells.
Occurrence of the three trk receptors in a variety ofcortical and deep nuclei neuronal elements supports thefunctional relevance of the NT trophic system in the humancerebellum from prenatal life to adulthood, already pro-posed on the basis of the presence of the trophins (Quartuet al., 2003). In this context, it is relevant that altered levelsof trk receptors occur in inherited neurological disordersinvolving the cerebellum in man (Kumada et al., 2000) andthat reduced levels of NT-4 are found in the cerebellum ofpatients with Alzheimer’s disease (Hock et al., 2000).
As already suggested (Quartu et al., 2003), this trophicsystem appears to meet the conditions for a concurrentactivity of multiple trophins. In fact, the observed codistri-bution of trk receptors indicate that multiple trophic factorsmay act simultaneously in the same region and possibly inthe same neuron. The simultaneous presence of trkA, trkB,and trkC receptors in the EGL of pre-term newborns maydenote a cooperating activity among NTs in the promotionof survival and migration of immature granule cells towardstheir final position in the IGL, as suggested in the mouse

318 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
(Minichiello and Klein, 1996). In a similar way, Purkinjecells differentiation may also benefit of a multiple trophicsupport (Minichiello and Klein, 1996). On the other hand,the finding that trk receptors (present study) and their ligandsNGF, NT-4 and NT-3 (Quartu et al., 2003) are still de-tectable in Purkinje and granule cells of the human cerebel-lum in infant and adult life extends to the whole ontogenesisthe possibility that a multifactorial trophic support is activein the maintenance of cerebellar neuronal connectivity.
Our results agree with and deepen previous data showingpan-trk immunoreactivity in EGL granule neurons and inmature granule and Purkinje cells (Muragaki et al., 1995),and are consistent with the detectability of trkA, trkB, andtrkC mRNAs (Hock et al., 2000) in human adult cerebel-lum. Yet, previous studies, where the immunohistochemicallocalization of either trk B and trkC receptors (Kumadaet al., 2000) or all trk receptors (Savaskan et al., 2000)is examined in the human cerebellum with the same an-tibodies we used, report different data between them andcompared to our results. The most likely explanation forthese discrepancies is that they result from the differentconditions in the tissue sampling and processing, and inthe immunostaining protocol. Also, in situ hybridizationstudies on developing and adult human cerebellum showlack of expression of trkB mRNA in Purkinje cells (Allenet al., 1994; Benisty et al., 1998). However, the issue ofdifferences between mRNA expression and detectabilityof the translated protein has been looked at carefully, andtemporal and spatial differences between the dynamics ofprotein and mRNA levels have been reported (seeNawaet al., 1995; Narisawa-Saito and Nawa, 1996; Das et al.,2001), which may account for this apparent inconsistency.
Comparison between the distribution of trk receptors andthat of NTs provides hints regarding the mechanisms oftrophic support. The trk receptors render the cells that ex-press them sensitive to the relevant NT, their localization al-lowing therefore the identification of the neurons that utilizethe appropriate NT. Colocalization of a certain trk receptorwith its proper NT may be interpreted as the result of boththe classical target-derived and the paracrine mode of sup-port, the NT being there as an internalized molecule. Alter-natively, it may be the result of an autocrine trophic mech-anism, the NT being then produced by the neuron togetherwith its receptor. Our results substantiate the likelihood ofboth the classical target-derived mechanism and local trophicinfluences of paracrine and, possibly, autocrine type. Theexistence of local modes of trophic support can be proposedfor the cells of the EGL, where immunoreactivity to both thetrk receptors and the NTs (Quartu et al., 2003) occur. In therat, the expression of BDNF mRNA in neurons of the deepcerebellar nuclei (Rocamora et al., 1993) has led to suggesta role for BDNF in the maturation of the Purkinje cells-deepcerebellar nuclei pathway, probably involving the classicaltarget-derived mechanism. Accordingly, our results allow theview that Purkinje neurons endowed with trk receptors, maycapture and internalize trophins at the level of the deep nu-
clei, where NT-LI neurons are present (Quartu et al., 2003).On the other hand, expression of trk receptors by perikaryaof the deep nuclei suggests further that these cells may alsodepend on NTs and this trophic activity may occur vialocal influences. Additional trophic mechanisms maybe also envisaged, such as the anterograde transport oftrophins, as suggested for BDNF in the development ofthe olivo-cerebellar pathway (Rocamora et al., 1993). Thecombined tissue localization of the trophin and receptorproteins, together with the in situ hybridization analysis forthe identification of the trophin expression sites should helpin defining the organization of the NT trophic system in thehuman cerebellum.
Acknowledgements
This work was funded by the CNR (MDF) and FondazioneBanco di Sardegna (MQ).
References
Allen, S.J., Dawbarn, D., Eckford, S.D., Wilcock, G.K., Ashcroft, M.,Colebrook, S.M., Feeney, R., MacGowan, S.H., 1994. Cloning of anon-catalytic form of human trkB in human brain. Neuroscience 60,825–834.
Aoki, C., Wu, K., Elste, A., Len, G., Lin, S., McAuliffe, G., Black,I.B., 2000. Localization of brain-derived neurotrophic factor and trkBreceptors to postsynaptic densities of adult rat cerebral cortex. J.Neurosci. Res. 59, 454–463.
Apfel, S., 1999. Neurotrophic factors in peripheral neuropathies:therapeutic implications. Brain Pathol. 9, 393–413.
Bao, S., Chen, L., Qiao, X., Knusel, B., Thompson, R.F., 1998. Impairedeye-blink conditioning in waggler, a mutant mouse with cerebellarBDNF deficiency. Learn Mem. 5, 355–364.
Barker, P.A., Lomen-Hoerth, C., Gensch, E.M., Meakin, S.O., Glass, D.J.,Shooter, E.M., 1993. Tissue-specific alternative splicing generates twoisoforms of the trkA receptor. J. Biol. Chem. 268, 15150–15157.
Bates, B., Rios, M., Trumpp, A., Chen, C., Fan, G., Bishop, J.M., Jaenisch,R., 1999. Neurotrophin-3 is required for proper cerebellar development.Nat. Neurosci. 2, 115–117.
Benisty, S., Boissiere, F., Faucheux, B., Agid, Y., Hirsch, E.C., 1998.trkB messenger RNA expression in normal human brain and in thesubstantia nigra of parkinsonian patients: an in situ hybridization study.Neuroscience 86, 813–826.
Borghesani, P.R., Peyrin, J.M., Klein, R., Rubin, J., Carter, A.R., Schwartz,P.M., Luster, A., Corfas, G., Segal, R.A., 2002. BDNF stimulatesmigration of cerebellar granule cells. Development 129, 1435–1442.
Bothwell, M., 1991. Keeping track of neurotrophin receptors. Cell 65,915–918.
Bothwell, M., 1995. Functional interactions of neurotrophins andneurotrophin receptors. Annu. Rev. Neurosci. 18, 223–253.
Carter, A.R., Chen, C., Schwartz, P.M., Segal, R.A., 2002. Brain-derivedneurotrophic factor modulates cerebellar plasticity and synapticultrastructure. J. Neurosci. 22, 1316–1327.
Chao, M.V., Hempstead, B.L., 1995. p75 and Trk: a two-receptor system.TINS 18, 321–326.
Chen, E., Mufson, E.J., Kordower, J.H., 1996. trk and p75 neurotrophinreceptor systems in the developing human brain. J. Comp. Neurol.369, 591–618.
Connor, B., Young, D., Lawlor, P., Gai, W., Waldvogel, H., Faull, R.L.M.,Dragunow, M., 1996. Trk receptor alterations in Alzheimer’s disease.Mol. Brain Res. 42, 1–17.

M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320 319
Connor, B., Young, D., Yan, Q., Faull, R.L.M., Sunek, B., Dragunow,M., 1997. Brain-derived neurotrophic factor is reduced in Alzheimer’sdisease. Mol. Brain Res. 49, 71–81.
Das, K.P., Chao, S.L., White, L.D., Haines, W.T., Harry, G.J., Tilson, H.A.,Barone Jr., S., 2001. Differential pattern of nerve growth factor, brain-derived neurotrophic factor and neurotrophin-3 mRNA and proteinlevels in developing regions of rat brain. Neuroscience 103, 739–761.
Del Fiacco, M., Quartu, M., Serra, M.P., Follesa, P., Lai, M.L., Bachis, A.,2002. Topographical localization of glial cell line-derived neurotrophicfactor in the normal human brain stem: immunohistochemical studyat prenatal, neonatal and adult age. J. Chem. Neuroanat. 23, 29–41.
Dubus, P., Faucheaux, B., Boissière, F., Groppi, A., Vital, C., Vital, A.,Agid, Y., Hirsch, E.C., Merlio, J.P., 2000. Expression of trk isoforms inbrain regions and in the striatum of patients with Alzheimer’s disease.Exp. Neurol. 165, 285–294.
Eberhart, C.G., Kaufman, W.E., Tihan, T., Burger, P.C., 2001. Apoptosis,neuronal maturation, and neurotrophin expression within medullo-blastoma nodules. J. Neuropathol. Exp. Neurol. 60, 462–469.
Ehlers, M.D., Kaplan, D.R., Price, D.L., Koliatsos, V.E., 1995.NGF-stimulated retrograde transport of trkA in the mammalian nervoussystem. J. Cell Biol. 130, 149–156.
Ernfors, P., Merlio, J.P., Persson, H., 1992. Cells expressing mRNAfor neurotrophins and their receptors during embryonic and postnataldevelopment. Eur. J. Neurosci. 4, 1140–1158.
Gao, W.-Q., Zheng, J.L., Karihaloo, M., 1995. Neurotrophin 4/5 (NT-4/5)and brain-derived neurotrophic factor (BDNF) act at later stages ofcerebellar granule cell differentiation. J. Neurosci. 15, 2656–2667.
Gestwa, G., Wiechers, B., Zimmermann, U., Praetorius, M., Rohboch, K.,Köpschall, I., Zenner, H.-P., Knipper, M., 1999. Differential expressionof trkB.T1 and trkB.T2, truncated trkC, and p75NGFR in the cochleaprior to hearing function. J. Comp. Neurol. 414, 33–49.
Goumnerova, L.C., 1996. Growth factor receptors and medulloblastoma.J. Neurooncol. 29, 85–89.
Heaton, M.B., Mitchell, J.J., Paiva, M., 1999. Ethanol-induced alterationsin neurotrophin expression in developing cerebellum: relationships toperiods of temporal susceptibility. Alchohol. Clin. Exp. Res. 23, 1637–1642.
Hock, C., Heese, K., Muller-Spahn, F., Hulette, C., Rosenberg, C., Otten,U., 1998. Decreased trkA neurotrophin receptor expression in theparietal cortex of patients with Alzheimer’s disease. Neurosci. Lett.241, 151–154.
Hock, C., Heese, K., Hulette, C., Rosenberg, C., Otten, U., 2000.Region-specific neurotrophin imbalances in Alzheimer disease:decreased levels of brain-derived neurotrophic factor and increasedlevels of nerve growth factor in hippocampus and cortical areas. Arch.Neurol. 57, 846–851.
Hofer, M., Pagliusi, S.R., Hohn, A., Leibrock, J., Barde, Y.-A., 1990.Regional distribution of brain-derived neurotrophic factor mRNA inthe adult mouse brain. EMBO J. 9, 2459–2464.
Klein, R., Parada, L.F., Coulier, F., Barbacid, M., 1989. trkB, a noveltyrosine protein kinase receptor gene expressed during mouse neuraldevelopment. EMBO J. 8, 3701–3709.
Kubo, T., Nonomura, T., Enokido, Y., Hatanaka, H., 1995. Brain-derivedneurotrophic factor (BDNF) can prevent apoptosis of rat cerebellargranule neurons in culture. Dev. Brain Res. 85, 249–258.
Kumada, S., Hayashi, M., Mizuguchi, M., Nakano, I., Morimatsu, Y.,Oda, M., 2000. Cerebellar degeneration in hereditary dentatorubral-pallidoluysian atrophy and Machado-Joseph disease. Acta Neuropathol.99, 48–54.
Lamballe, F., Klein, R., Barbacid, M., 1991. trkC, a new member of thetrk family of tyrosine protein kinases, is a receptor for neurotrophin-3.Cell 66, 967–979.
Lärkfors, L., Lindsay, R.M., Alderson, R.F., 1996. Characterization of theresponses of Purkinje cells to neurotrophin treatment. J. Neurochem.66, 1362–1373.
Lewin, G.R., Barde, Y.A., 1996. Physiology of the neurotrophins. Annu.Rev. Neurosci. 19, 289–317.
Light, K.E., Ge, Y., Belcher, S.M., 2001. Early postnatal ethanol exposureselectively decreases BDNF and truncated trkB-T2 receptor mRNAexpression in the rat cerebellum. Mol. Brain Res. 93, 46–55.
Light, K.E., Brown, D.P., Newton, B.W., Belcher, S.M., Kane, C.J.M.,2002. Ethanol-induced alterations of neurotrophin receptor expressionon Purkinje cells in the neonatal rat cerebellum. Brain Res. 924, 71–81.
Lindholm, D., Dechant, G., Heisenberg, C.P., Thoenen, H., 1993.Brain-derived neurotrophic factor is a survival factor for cultured ratcerebellar granule neurons and protects them against glutamate-inducedneurotoxicity. Eur. J. Neurosci. 5, 1455–1464.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Proteinmeasurements with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Maisonpierre, P.E., Belluscio, L., Friedman, B., Alderson, R.F., Wiegand,S.J., Furth, M.E., Lindsay, R.M., Yancopoulos, G.D., 1990. NT-3,BDNF, and NGF in the developing rat nervous system: parallel as wellas reciprocal patterns of expression. Neuron 5, 501–509.
Martin-Zanca, D., Oskam, R., Mitra, G., Copeland, T., Barbacid, M.,1989. Molecular and biochemical characterization of the human trkproto-oncogene. Mol. Cell Biol. 9, 24–33.
Meakin, S.O., Shooter, E.M., 1992. The nerve growth factor family ofreceptors. TINS 15, 323–331.
Merlio, J.P., Ernfors, P., Jaber, M., Persson, H., 1992. Molecular cloningof rat trkC and distribution of cells expressing messenger RNAsfor members of the trk family in the rat central nervous system.Neuroscience 51, 513–532.
Mertz, K., Koscheck, T., Schilling, K., 2000. Brain-derived neurotrophicfactor modulates dendritic morphology of cerebellar basket and stellatecells: an in vitro study. Neuroscience 97, 303–310.
Minichiello, L., Klein, R., 1996. TrkB and TrkC neurotrophin receptorscooperate in promoting survival of hippocampal and cerebellar granuleneurons. Genes Dev. 10, 2849–2858.
Mufson, E.J., Higgins, G.A., Kordower, J.H., 1991. Nerve growth factorreceptor immunoreactivity in the new world monkey (Cebus Apella)and human cerebellum. J. Comp. Neurol. 308, 555–575.
Muragaki, Y., Timothy, N., Leight, S., Hempstead, B.L., Chao, M.V.,Trojanowski, J.Q., Lee, V.M.-Y., 1995. Expression of trk receptors indeveloping and adult human central and peripheral nervous system. J.Comp. Neurol. 356, 387–397.
Narisawa-Saito, M., Nawa, H., 1996. Differential regulation ofhippocampal neurotrophins during aging in rats. J. Neurochem. 67,1124–1131.
Nawa, H., Carnahan, J., Gall, C., 1995. BDNF protein measured by anovel enzyme immunoassay in normal brain and after seizure: partialdisagreement with mRNA levels. Eur. J. Neurosci. 7, 1527–1535.
Neet, K.E., Campenot, R.B., 2001. Receptor binding, internalization, andretrograde transport of neurotrophic factors. Cell. Mol. Life Sci. 58,1021–1035.
Nishio, T., Furukawa, S., Akiguchi, I., Oka, N., Ohnishi, K., Tomimoto,H., Nakamura, S., Kimura, J., 1994. Cellular localization of nervegrowth factor-like immunoreactivity in adult rat brain: quantitative andimmunohistochemical study. Neuroscience 60, 67–84.
Ohira, K., Shimizu, K., Hayashi, M., 1999. Change of expression offull-length and truncated trkBs in the developing monkey centralnervous system. Dev. Brain Res. 112, 21–29.
Pioro, E.P., Cuello, A.C., 1988. Purkinje cells of adult cerebellum expressnerve growth factor receptor immunoreactivity: light microscopicobservations. Brain Res. 455, 182–186.
Pomeroy, S.L., Sutton, M.E., Goumnerova, L.C., Segal, R.A.,1997. Neurotrophins in cerebellar granule cell development andmedulloblastoma. J. Neurooncol. 35, 347–352.
Qiao, X., Hefti, F., Knusel, B., Noebels, J.L., 1996. Selective failure ofbrain-derived neurotrophic factor mRNA expression in the cerebellumof stargazer, a mutant mouse with ataxia. J. Neurosci. 16, 640–648.
Qiao, X., Chen, L., Gao, H., Bao, S., Hefti, F., Thompson, R.F., Knusel,B., 1998. Cerebellar brain-derived neurotrophic factor-TrkB defectassociated with impairment of eyeblink conditioning in Stargazermutant mice. J. Neurosci. 18, 6990–6999.

320 M. Quartu et al. / Int. J. Devl Neuroscience 21 (2003) 309–320
Quartu, M., Serra, M.P., Manca, A., Follesa, P., Lai, M.L., Del Fiacco,M., 2003. Neurotrophin-like immunoreactivity in the human pre-termnewborn, infant, and adult cerebellum. Int. J. Dev. Neurosci. 21, 23–33.
Rabacchi, S.A., Kruk, B., Hamilton, J., Carney, C., Hoffman, J.R., Meyer,S.L., Springer, J.E., Baird, D.H., 1999. BDNF and NT4/5 promotesurvival and neurite outgrowth of pontocerebellar mossy fiber neurons.J. Neurobiol. 40, 254–269.
Rico, B., Xu, B., Reichardt, L.F., 2002. TrkB receptor signaling is requiredfor establishment of GABAergic synapses in the cerebellum. Nat.Neurosci. 5, 225–233.
Rocamora, N., Garcia-Ladona, F.J., Palacios, J.M., Mengod, G.,1993. Differential expression of brain-derived neurotrophic factor,neurotrophin-3, and low-affinity nerve growth factor receptor duringthe postnatal development of the rat cerebellar system. Mol. Brain Res.17, 1–8.
Saragovi, H.U., Gehring, K., 2000. Development of pharmacologicalagents for targeting neurotrophins and their receptors. TIPS 21, 93–98.
Savaskan, E., Muller-Spahn, F., Olivieri, G., Bruttel, S., Otten, U.,Rosenberg, C., Hulette, C., Hock, C., 2000. Alterations in trkA, trkBand trkC receptor immunoreactivities in parietal cortex and cerebellumin Alzheimer’s disease. Eur. Neurol. 44, 172–180.
Schwartz, P.M., Borghesani, P.R., Levy, R.L., Pomeroy, S.L., Segal, R.A.,1997. Abnormal cerebellar development and foliation in BDNF-/-micereveals a role for neurotrophins in CNS patterning. Neuron 19, 269–281.
Segal, R.A., Takahashi, H., McKay, R.D., 1992. Changes in neurotrophinresponsiveness during the development of cerebellar granule neurons.Neuron 9, 1041–1052.
Segal, R.A., Pomeroy, S.L., Stiles, C.D., 1995. Axonal growth andfasciculation linked to differential expression of BDNF and NT-3receptors in developing cerebellar granule cells. J. Neurosci. 15, 4970–4981.
Segal, R.A., Rua, L., Schwartz, P., 1997. Neurotrophins and programmedcell death during cerebellar development. Adv. Neurol. 72, 79–86.
Shelton, D.L., Sutherland, J., Gripp, J., Camerato, T., Armanini, M.P.,Phillips, H.S., Carroll, K., Spencer, S.D., Levinson, A.D., 1995.Human trks: molecular cloning, tissue distribution, and expression ofextracellular domain immunoadhesins. J. Neurosci. 156, 477–491.
Sherrard, R.M., Bower, A.J., 2001. BDNF and NT-3 extend the criticalperiod for developmental climbing fiber plasticity. NeuroReport 12,2871–2874.
Shibayama, M., Hattori, S., Himes, B.T., Murray, M., Tessler, A., 1998.Neurotrophin-3 prevents death of axotomized Clarke’s nucleus neuronsin adult rat. J. Comp. Neurol. 390, 102–111.
Siegel, G.J., Chauhan, N.B., 2000. Neurotrophic factors in Alzheimer’sand Parkinson’s disease brain. Brain Res. Rev. 33, 199–227.
Skaper, S.D., Floreani, M., Negro, A., Facci, L., Giusti, P., 1998.Neurotrophins rescue cerebellar granule neurons from oxidativestress-mediated apoptotic death: selective involvement of phospha-tidylinositol 3-kinase and the mitogen-activated protein kinase pathway.J. Neurochem. 70, 1859–1868.
Soppet, D., Escandon, E., Maragos, J., Middlemas, D.S., Reid, S.W., Balir,J., Burton, L.E., Stanton, B.R., Kaplan, D.R., Hunter, T., Nikolics, K.,Parada, L.F., 1991. The neurotrophic factors brain-derived neurotrophicfactor and neurotrophin-3 are ligands for the trkB tyrosine kinasereceptor. Cell 65, 895–903.
Tanaka, S., Sekino, Y., Shirao, T., 2000. The effects of neurotrophin-3 andbrain-derived neurotrophic factor on cerebellar granule cell movementand neurite extension in vitro. Neuroscience 97, 727–734.
Thoenen, H., 1995. NTs and neuronal plasticity. Science 270, 593–598.
Timmusk, T., Belluardo, N., Metsis, M., Persson, H., 1993. Widespreadand developmentally regulated expression of neurotrophin-4 mRNA inrat brain and peripheral tissues. Eur. J. Neurosci. 5, 605–613.
Tong, L., Perez-Polo, R., 1998. Brain-derived neurotrophic factor(BDNF) protects cultured rat cerebellar granule neurons againstglucose deprivation-induced apoptosis. J. Neural Transm. 105, 905–914.
Tsoulfas, P., Soppet, D., Escandon, E., Tessarollo, L., Mendoza-Ramirez,J.-L., Rosenthal, A., Nikoliks, K., Parada, L.F., 1993. The rat trkClocus encodes multiple neurogenic receptors that exhibit differentialresponse to neurotrophin-3 in PC12 cells. Neuron 10, 975–990.
Wanaka, A., Johnson Jr., E.M., 1990. Developmental study of nervegrowth factor receptor mRNA expression in the postnatal rat cere-bellum. Dev. Brain Res. 55, 288–292.
Wullner, U., Isenmann, S., Gleichmann, M., Klockgether, T., Bahr, M.,1998. Expression of neurotrophins and neurotrophin receptors in thecerebellum of mutant weaver and lurcher mice. Dev. Brain Res. 110,1–6.
Zhou, J., Valletta, J.S., Grimes, M., Mobley, W.C., 1995. Multiple levelsfor regulation of trkA in PC12 cells by nerve growth factor. J.Neurochem. 65, 1146–1156.





























