Embed Size (px)
Citation preview

ARTICLE IN PRESS
Quaternary International 122 (2004) 57–68
*Correspondin
1040-6182/$ - see
doi:10.1016/j.qua
Holocene vegetation history and climate change on the northern KolaPeninsula, Russia: a case study from a small tundra lake
Konstantin V. Kremenetskia,*, Glen M. MacDonalda, Bruce R. Gervaisa,Olga K. Borisovab, Jeffrey A. Snyderc
aDepartments of Geography and Organismic Biology, Ecology and Evolution University of California, Los Angeles, CA 90095-1524, USAb Institute of Geography, Russian Academy of Sciences, 29 Staromonetny Lane, Moscow 109017, Russia
cDepartment of Geology, Bowling Green State University, 190 Overman Hall, Bowling Green, OH 43403, USA
Abstract
A sediment core from Lake KP3 (unofficial name) located near the coastline on the northern Kola Peninsula, 100 km east-south-
east fromMurmansk, Russia (69�0401900N, 36�0004000E) provides palynological evidence for the vegetation and climate history of the
last 14,000 years. During the Younger Dryas time the climate was cold and vegetation cover around the lake consisted of a shrub
birch-willow tundra. With warming during the early Holocene birch forest expanded across the Kola Peninsula and by
9500 cal yr BP a dense birch tree forests likely extended as far north as the modern Barents Sea coastline. By about 7800 cal yr BP
Alnus incana was present as an admixture to birch in wet places as far north as the Barents Sea shoreline. Scots pine expanded to
north of its modern limits on the Kola Peninsula between 7900 and 3700 cal yr BP with its maximum northward progress and
abundance at 7300–6200 cal yr BP. Although scattered pines may have grown near the coastline, pine forests did not reach the
Barents Sea coastal area and the region remained covered with birth forest-tundra as much as during earlier Holocene. The
expansion of birch forest and pine forest in the early to mid Holocene likely reflects the impact of higher amounts of summer
radiation due to Milankovitch forcing and warmer temperatures in the North Atlantic Ocean and Barents Sea. The mid-Holocene
summer warming in the Barents Sea coastal area likely ranged from 1�C to 1.5�C above modern conditions. Following
5900 cal yr BP shrub tundra communities began to replace birch forest and pines on the northernmost Kola Peninsula. The modern
tundra which occupies the coastal region near Lake KP3 was established at about 3100 cal yr BP. The vegetation and climatic
changes evident on the northern Kola Peninsula are similar to those evident in northernmost Fennoscandia.
r 2004 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Arctic lacustrine deposits can be particularly goodrepositories of paleoenvironmental records of climateand vegetation change (MacDonald et al., 1993, 2000a).Many lakes ideal for such purposes can be found on theKola Peninsula and adjacent Fennoscandia. The recon-struction of climate and vegetation change in this regionis of special interest as it is sensitive to climatic changescaused by Milankovitch cycles and variations in seasurface temperatures in the North Atlantic. The north-ern Kola-Fennoscandian region is the gateway by whichclimatic changes in the North Atlantic sector areteleconnected into the Eurasian Arctic (MacDonaldet al., 2000b, c). The vegetation of the northern Kola isparticularly sensitive to climatic changes because it
g author.
front matter r 2004 Elsevier Ltd and INQUA. All rights
int.2004.01.031
possesses a tundra-forest ecotone that includes climati-cally induced tree lines for tree-birch and pine.The postglacial history of vegetation and tree line
change is well studied in northern Fennoscandia, mainlythrough pollen and macrofossil analyses (Hyv.arinen,1976, 1993; Eronen, 1979; Eronen et al., 1999a; Prentice,1982; Sepp.a, 1996; Sepp.a and Hammarlund, 2000). Adata set of radiocarbon dated pine macrofossils has beenrecovered from the Kola Peninsula (MacDonald et al.,2000b). However, palynological studies with good chron-ological control remain sparse from the Kola Peninsula.Those that are available were obtained mainly from peatsections (Yelina et al., 1995, 2000; Kremenetski et al.,1997, 1999; Kremenetski and Patyk-Kara, 1997). Veryrecently high-resolution palynological analyses of lakecores from the Kola have began to become available(Snyder et al., 2000; Gervais et al., 2002) and demonstratethe great potential of lacustrine deposits to providevaluable paleoenvironmental records from the region.
reserved.

ARTICLE IN PRESSK.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6858
This paper presents the pollen record obtained from asediment core taken from a small lake in the moderntundra belt of the north-central Kola Peninsula. The siteis adjacent to the Barents Sea coastline. The pollenstratigraphy from the site extends back to approxi-mately 14,000 cal yr BP and provides evidence of vegeta-tion, tree line and climate history of the north KolaPeninsula. These data are compared with other recentlypublished evidence from the Kola Peninsula and similarrecords from northernmost Fennoscandia.
2. Study area
Lake KP3 (69�0401900N, 36�0004000E, 75m a.s.l.) islocated in the tundra of the central Murman coast atabout 100 km east-south-east from Murmansk not farfrom a previously studied site at Yarnyshnoe Lake(Snyder et al., 2000) (Fig. 1). Numerous glacier-scouredlakes of different sizes and shape characterise theregional landscape. The topography of the area is flatto rolling with surrounding hills rising to approximately250m a.s.l. Bedrock surrounding the lake consists ofPrecambrian crystalline igneous rocks of the BalticShield (Atlas Murmanskoi Oblasti, 1971). The area wasglaciated during the last glacial maximum but is locatednorth of the Younger Dryas end moraine (Rainio et al.,1995). The climate is cold-temperate and modified bythe proximity to the Barents Sea. The January and Julymean temperature is �11�C and +9�C, respectively.Annual precipitation amounts to 600–700mm at theMurmansk coast. Most of the precipitation occurs inJuly–October (Atlas Murmanskoi Oblasti, 1971).The size of Lake KP3 is about 5 ha with maximum
depth 5.5m. The surrounding vegetation is shrub tundrawith scattered mountain birches (Betula pubescens Ehrh.ssp. czerepanovii (Orlova) H.amit-Ahti (Betula tortuosa
Ledeb.)) at low-elevations in protected areas. Such
Fig. 1. Map of the Kola Peninsula and sites mentioned in the paper (1) KP
Poteryanny Zub Lake (Gervais et al., 2002); (4) Rybachiy mire (Kremenetsk
scattered birches are a common feature for the tundraof the Murman coast (Kremenetski et al., 1997). Shrubsspecies include Ericaceae, Rubus chamaemorus, andSalix. The ground cover consists of sedges, grasses,diverse tundra herbs and mosses. Large areas of barebedrock are also present.
3. Methods
3.1. Coring and sampling
Lake coring was conducted from the lake ice in thecentral part of the lake in April 1998, using a modifiedLivingstone piston corer. The cores were extruded andwrapped in the field and subsampled in the UCLAlaboratory.
3.2. Radiocarbon dating and holocene chronology
Radiocarbon ages was established by accelerator massspectrometry (AMS) dating of plant macrofossil andbulk organic sediments. Radiocarbon dating was con-ducted by the INSTAAR Laboratory of the Universityof Colorado at Boulder. Calibration of radiocarbondates into calendar years was done using CALIB.4program (Stuiver et al., 1998) (Table 1). The calibratedchronology is used throughout the paper.
3.3. Loss on ignition and pollen analysis
The loss on ignition analysis followed the standardprocedure (Dean, 1974). Standard methods (Faegri andIversen, 1989) have been used for the preparation ofsamples for pollen analysis and for pollen counting.Pretreated Lycopodium tablets were added to eachsubsample to calculate pollen influx values (Stockmarr,1971). A minimum count of 500 grains per sample was
3 Lake (this paper); (2) Yarnyshnoe-3 Lake (Snyder et al., 2000); (3)
i et al., 1997).
ARTICLE IN PRESS
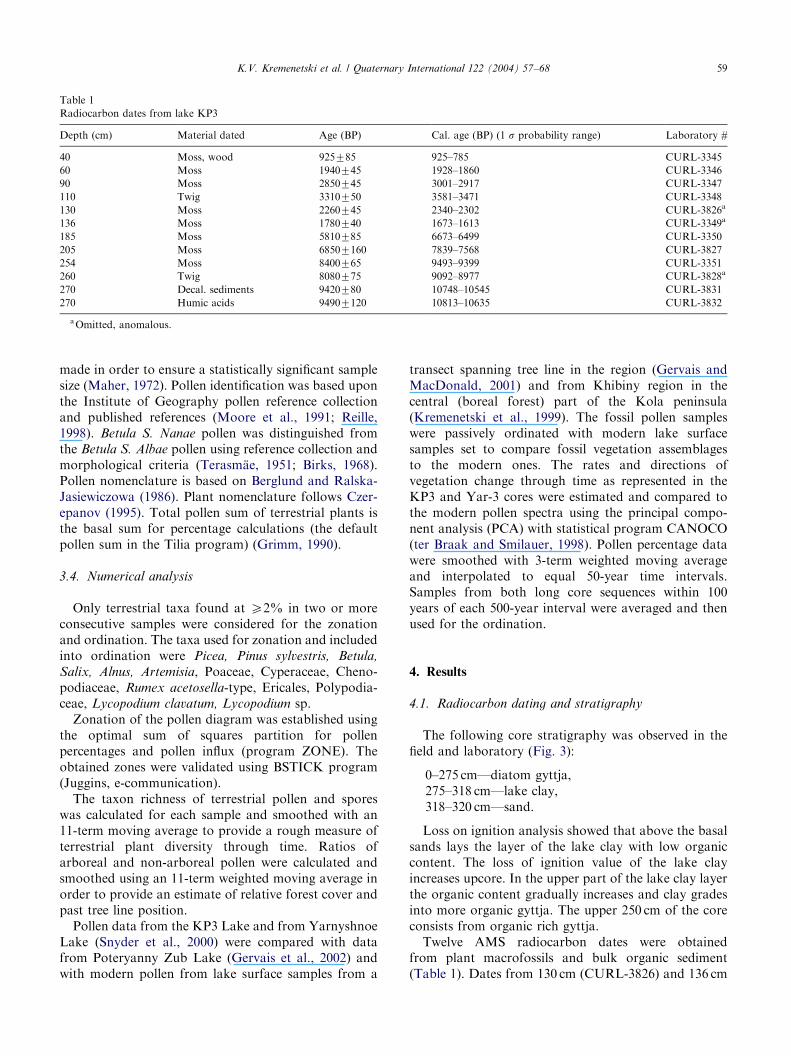
Table 1
Radiocarbon dates from lake KP3
Depth (cm) Material dated Age (BP) Cal. age (BP) (1 s probability range) Laboratory #
40 Moss, wood 925785 925–785 CURL-3345
60 Moss 1940745 1928–1860 CURL-3346
90 Moss 2850745 3001–2917 CURL-3347
110 Twig 3310750 3581–3471 CURL-3348
130 Moss 2260745 2340–2302 CURL-3826a
136 Moss 1780740 1673–1613 CURL-3349a
185 Moss 5810785 6673–6499 CURL-3350
205 Moss 68507160 7839–7568 CURL-3827
254 Moss 8400765 9493–9399 CURL-3351
260 Twig 8080775 9092–8977 CURL-3828a
270 Decal. sediments 9420780 10748–10545 CURL-3831
270 Humic acids 94907120 10813–10635 CURL-3832
aOmitted, anomalous.
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–68 59
made in order to ensure a statistically significant samplesize (Maher, 1972). Pollen identification was based uponthe Institute of Geography pollen reference collectionand published references (Moore et al., 1991; Reille,1998). Betula S. Nanae pollen was distinguished fromthe Betula S. Albae pollen using reference collection andmorphological criteria (Terasm.ae, 1951; Birks, 1968).Pollen nomenclature is based on Berglund and Ralska-Jasiewiczowa (1986). Plant nomenclature follows Czer-epanov (1995). Total pollen sum of terrestrial plants isthe basal sum for percentage calculations (the defaultpollen sum in the Tilia program) (Grimm, 1990).
3.4. Numerical analysis
Only terrestrial taxa found at X2% in two or moreconsecutive samples were considered for the zonationand ordination. The taxa used for zonation and includedinto ordination were Picea, Pinus sylvestris, Betula,
Salix, Alnus, Artemisia, Poaceae, Cyperaceae, Cheno-podiaceae, Rumex acetosella-type, Ericales, Polypodia-ceae, Lycopodium clavatum, Lycopodium sp.Zonation of the pollen diagram was established using
the optimal sum of squares partition for pollenpercentages and pollen influx (program ZONE). Theobtained zones were validated using BSTICK program(Juggins, e-communication).The taxon richness of terrestrial pollen and spores
was calculated for each sample and smoothed with an11-term moving average to provide a rough measure ofterrestrial plant diversity through time. Ratios ofarboreal and non-arboreal pollen were calculated andsmoothed using an 11-term weighted moving average inorder to provide an estimate of relative forest cover andpast tree line position.Pollen data from the KP3 Lake and from Yarnyshnoe
Lake (Snyder et al., 2000) were compared with datafrom Poteryanny Zub Lake (Gervais et al., 2002) andwith modern pollen from lake surface samples from a
transect spanning tree line in the region (Gervais andMacDonald, 2001) and from Khibiny region in thecentral (boreal forest) part of the Kola peninsula(Kremenetski et al., 1999). The fossil pollen sampleswere passively ordinated with modern lake surfacesamples set to compare fossil vegetation assemblagesto the modern ones. The rates and directions ofvegetation change through time as represented in theKP3 and Yar-3 cores were estimated and compared tothe modern pollen spectra using the principal compo-nent analysis (PCA) with statistical program CANOCO(ter Braak and Smilauer, 1998). Pollen percentage datawere smoothed with 3-term weighted moving averageand interpolated to equal 50-year time intervals.Samples from both long core sequences within 100years of each 500-year interval were averaged and thenused for the ordination.
4. Results
4.1. Radiocarbon dating and stratigraphy
The following core stratigraphy was observed in thefield and laboratory (Fig. 3):
0–275 cm—diatom gyttja,275–318 cm—lake clay,318–320 cm—sand.
Loss on ignition analysis showed that above the basalsands lays the layer of the lake clay with low organiccontent. The loss of ignition value of the lake clayincreases upcore. In the upper part of the lake clay layerthe organic content gradually increases and clay gradesinto more organic gyttja. The upper 250 cm of the coreconsists from organic rich gyttja.Twelve AMS radiocarbon dates were obtained
from plant macrofossils and bulk organic sediment(Table 1). Dates from 130 cm (CURL-3826) and 136 cm
ARTICLE IN PRESS
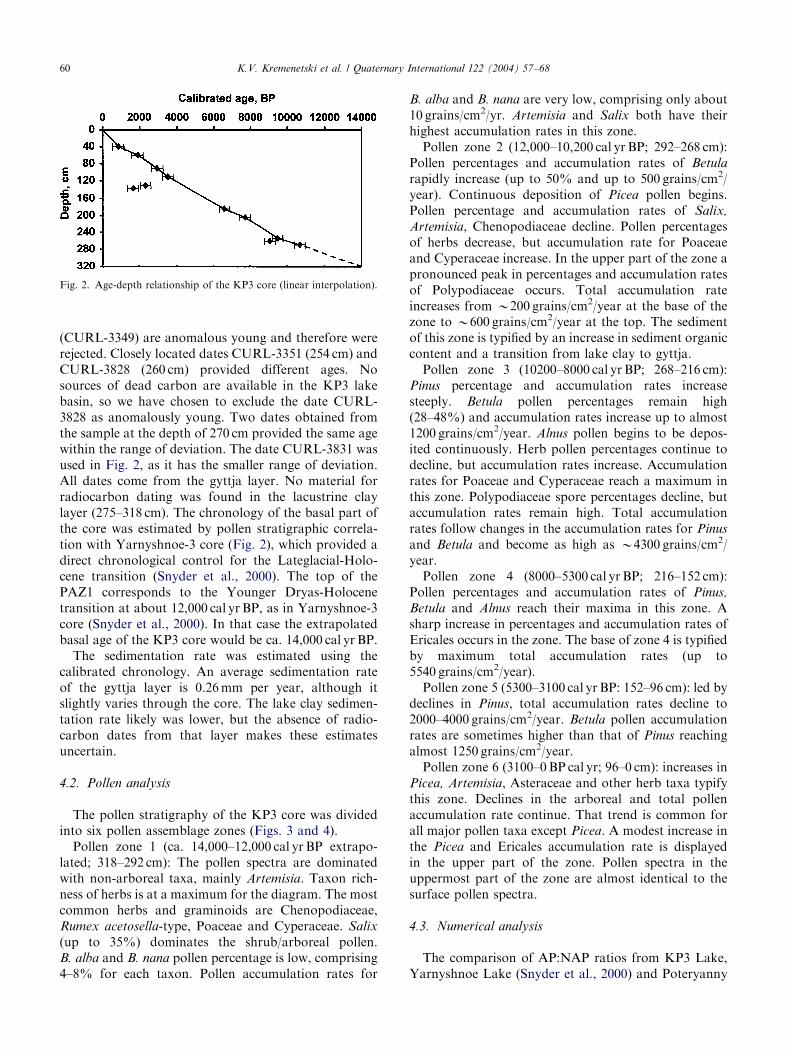
Fig. 2. Age-depth relationship of the KP3 core (linear interpolation).
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6860
(CURL-3349) are anomalous young and therefore wererejected. Closely located dates CURL-3351 (254 cm) andCURL-3828 (260 cm) provided different ages. Nosources of dead carbon are available in the KP3 lakebasin, so we have chosen to exclude the date CURL-3828 as anomalously young. Two dates obtained fromthe sample at the depth of 270 cm provided the same agewithin the range of deviation. The date CURL-3831 wasused in Fig. 2, as it has the smaller range of deviation.All dates come from the gyttja layer. No material forradiocarbon dating was found in the lacustrine claylayer (275–318 cm). The chronology of the basal part ofthe core was estimated by pollen stratigraphic correla-tion with Yarnyshnoe-3 core (Fig. 2), which provided adirect chronological control for the Lateglacial-Holo-cene transition (Snyder et al., 2000). The top of thePAZ1 corresponds to the Younger Dryas-Holocenetransition at about 12,000 cal yr BP, as in Yarnyshnoe-3core (Snyder et al., 2000). In that case the extrapolatedbasal age of the KP3 core would be ca. 14,000 cal yr BP.The sedimentation rate was estimated using the
calibrated chronology. An average sedimentation rateof the gyttja layer is 0.26mm per year, although itslightly varies through the core. The lake clay sedimen-tation rate likely was lower, but the absence of radio-carbon dates from that layer makes these estimatesuncertain.
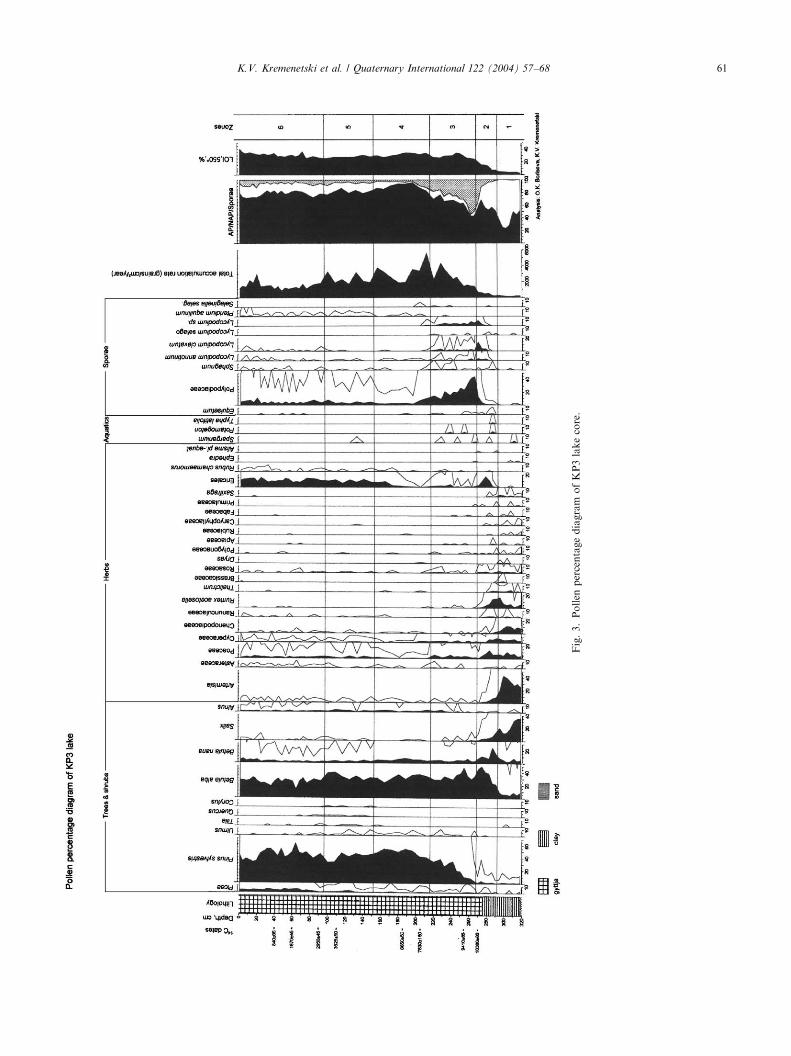
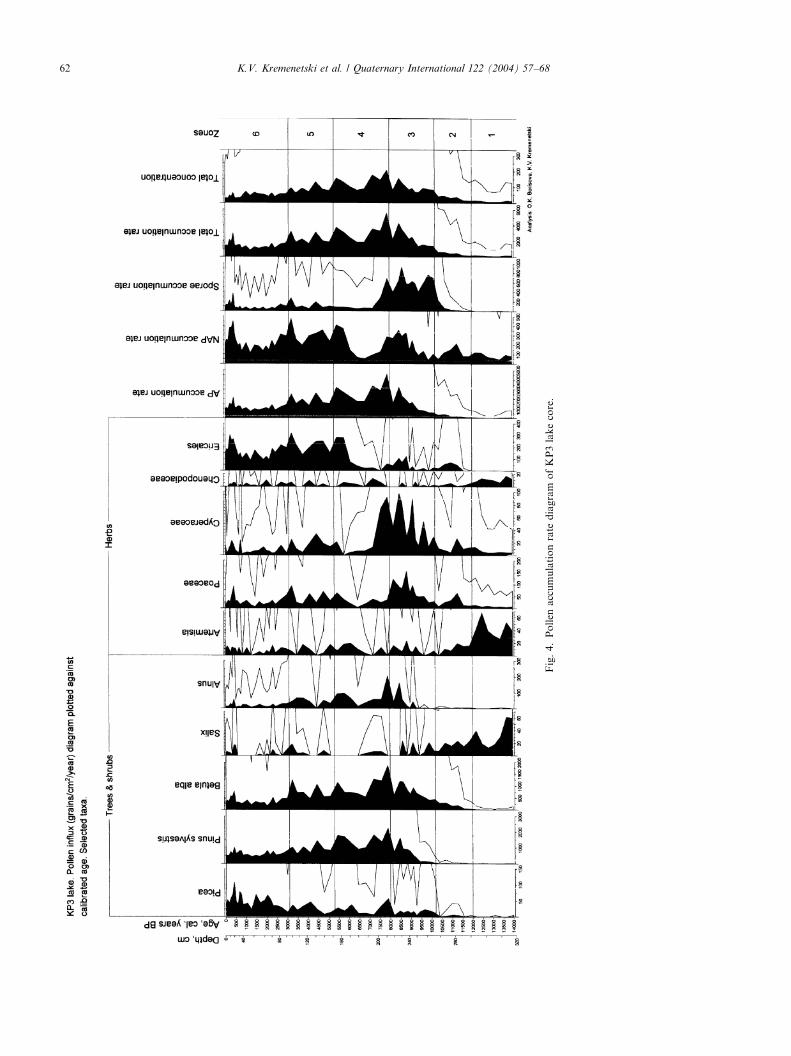
4.2. Pollen analysis
The pollen stratigraphy of the KP3 core was dividedinto six pollen assemblage zones (Figs. 3 and 4).Pollen zone 1 (ca. 14,000–12,000 cal yr BP extrapo-
lated; 318–292 cm): The pollen spectra are dominatedwith non-arboreal taxa, mainly Artemisia. Taxon rich-ness of herbs is at a maximum for the diagram. The mostcommon herbs and graminoids are Chenopodiaceae,Rumex acetosella-type, Poaceae and Cyperaceae. Salix
(up to 35%) dominates the shrub/arboreal pollen.B. alba and B. nana pollen percentage is low, comprising4–8% for each taxon. Pollen accumulation rates for
B. alba and B. nana are very low, comprising only about10 grains/cm2/yr. Artemisia and Salix both have theirhighest accumulation rates in this zone.Pollen zone 2 (12,000–10,200 cal yr BP; 292–268 cm):
Pollen percentages and accumulation rates of Betula
rapidly increase (up to 50% and up to 500 grains/cm2/year). Continuous deposition of Picea pollen begins.Pollen percentage and accumulation rates of Salix,
Artemisia, Chenopodiaceae decline. Pollen percentagesof herbs decrease, but accumulation rate for Poaceaeand Cyperaceae increase. In the upper part of the zone apronounced peak in percentages and accumulation ratesof Polypodiaceae occurs. Total accumulation rateincreases from B200 grains/cm2/year at the base of thezone to B600 grains/cm2/year at the top. The sedimentof this zone is typified by an increase in sediment organiccontent and a transition from lake clay to gyttja.Pollen zone 3 (10200–8000 cal yr BP; 268–216 cm):
Pinus percentage and accumulation rates increasesteeply. Betula pollen percentages remain high(28–48%) and accumulation rates increase up to almost1200 grains/cm2/year. Alnus pollen begins to be depos-ited continuously. Herb pollen percentages continue todecline, but accumulation rates increase. Accumulationrates for Poaceae and Cyperaceae reach a maximum inthis zone. Polypodiaceae spore percentages decline, butaccumulation rates remain high. Total accumulationrates follow changes in the accumulation rates for Pinus
and Betula and become as high as B4300 grains/cm2/year.Pollen zone 4 (8000–5300 cal yr BP; 216–152 cm):
Pollen percentages and accumulation rates of Pinus,
Betula and Alnus reach their maxima in this zone. Asharp increase in percentages and accumulation rates ofEricales occurs in the zone. The base of zone 4 is typifiedby maximum total accumulation rates (up to5540 grains/cm2/year).Pollen zone 5 (5300–3100 cal yr BP: 152–96 cm): led by
declines in Pinus, total accumulation rates decline to2000–4000 grains/cm2/year. Betula pollen accumulationrates are sometimes higher than that of Pinus reachingalmost 1250 grains/cm2/year.Pollen zone 6 (3100–0BP cal yr; 96–0 cm): increases in
Picea, Artemisia, Asteraceae and other herb taxa typifythis zone. Declines in the arboreal and total pollenaccumulation rate continue. That trend is common forall major pollen taxa except Picea. A modest increase inthe Picea and Ericales accumulation rate is displayedin the upper part of the zone. Pollen spectra in theuppermost part of the zone are almost identical to thesurface pollen spectra.
4.3. Numerical analysis
The comparison of AP:NAP ratios from KP3 Lake,Yarnyshnoe Lake (Snyder et al., 2000) and Poteryanny
ARTICLE IN PRESS
Fig.3.PollenpercentagediagramofKP3lakecore.
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–68 61
ARTICLE IN PRESS
Fig.4.PollenaccumulationratediagramofKP3lakecore.
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6862
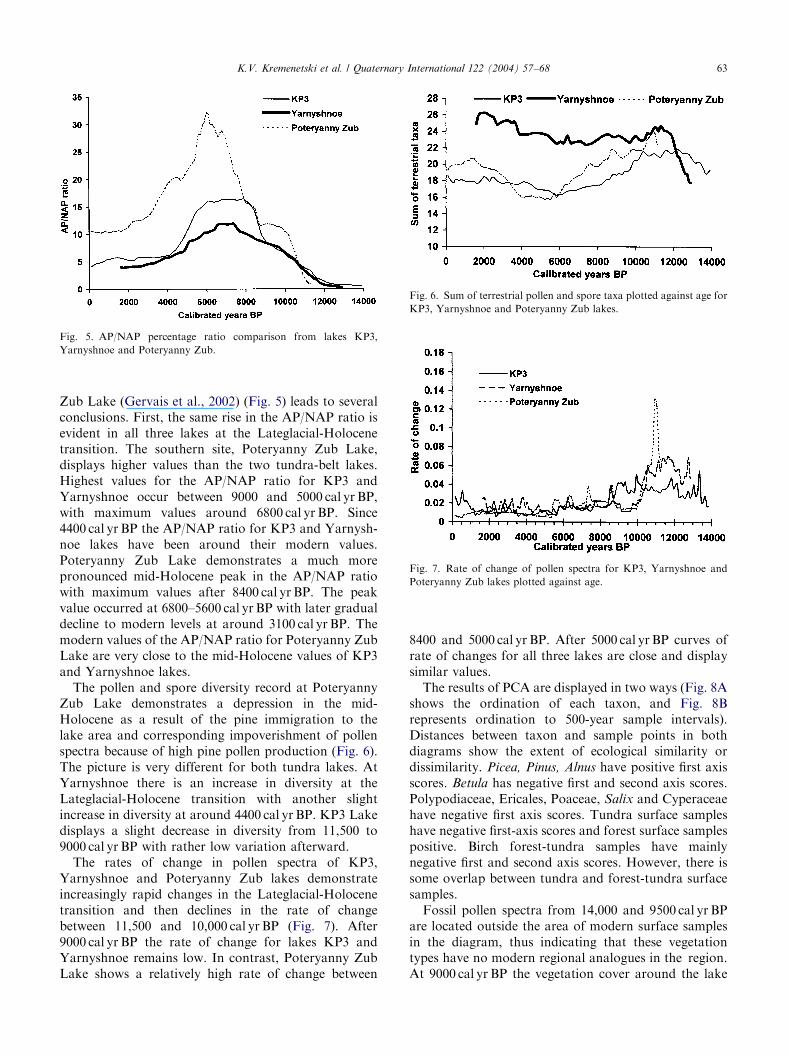
ARTICLE IN PRESS
Fig. 5. AP/NAP percentage ratio comparison from lakes KP3,
Yarnyshnoe and Poteryanny Zub.
Fig. 6. Sum of terrestrial pollen and spore taxa plotted against age for
KP3, Yarnyshnoe and Poteryanny Zub lakes.
Fig. 7. Rate of change of pollen spectra for KP3, Yarnyshnoe and
Poteryanny Zub lakes plotted against age.
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–68 63
Zub Lake (Gervais et al., 2002) (Fig. 5) leads to severalconclusions. First, the same rise in the AP/NAP ratio isevident in all three lakes at the Lateglacial-Holocenetransition. The southern site, Poteryanny Zub Lake,displays higher values than the two tundra-belt lakes.Highest values for the AP/NAP ratio for KP3 andYarnyshnoe occur between 9000 and 5000 cal yr BP,with maximum values around 6800 cal yr BP. Since4400 cal yr BP the AP/NAP ratio for KP3 and Yarnysh-noe lakes have been around their modern values.Poteryanny Zub Lake demonstrates a much morepronounced mid-Holocene peak in the AP/NAP ratiowith maximum values after 8400 cal yr BP. The peakvalue occurred at 6800–5600 cal yr BP with later gradualdecline to modern levels at around 3100 cal yr BP. Themodern values of the AP/NAP ratio for Poteryanny ZubLake are very close to the mid-Holocene values of KP3and Yarnyshnoe lakes.The pollen and spore diversity record at Poteryanny
Zub Lake demonstrates a depression in the mid-Holocene as a result of the pine immigration to thelake area and corresponding impoverishment of pollenspectra because of high pine pollen production (Fig. 6).The picture is very different for both tundra lakes. AtYarnyshnoe there is an increase in diversity at theLateglacial-Holocene transition with another slightincrease in diversity at around 4400 cal yr BP. KP3 Lakedisplays a slight decrease in diversity from 11,500 to9000 cal yr BP with rather low variation afterward.The rates of change in pollen spectra of KP3,
Yarnyshnoe and Poteryanny Zub lakes demonstrateincreasingly rapid changes in the Lateglacial-Holocenetransition and then declines in the rate of changebetween 11,500 and 10,000 cal yr BP (Fig. 7). After9000 cal yr BP the rate of change for lakes KP3 andYarnyshnoe remains low. In contrast, Poteryanny ZubLake shows a relatively high rate of change between
8400 and 5000 cal yr BP. After 5000 cal yr BP curves ofrate of changes for all three lakes are close and displaysimilar values.The results of PCA are displayed in two ways (Fig. 8A
shows the ordination of each taxon, and Fig. 8Brepresents ordination to 500-year sample intervals).Distances between taxon and sample points in bothdiagrams show the extent of ecological similarity ordissimilarity. Picea, Pinus, Alnus have positive first axisscores. Betula has negative first and second axis scores.Polypodiaceae, Ericales, Poaceae, Salix and Cyperaceaehave negative first axis scores. Tundra surface sampleshave negative first-axis scores and forest surface samplespositive. Birch forest-tundra samples have mainlynegative first and second axis scores. However, there issome overlap between tundra and forest-tundra surfacesamples.Fossil pollen spectra from 14,000 and 9500 cal yr BP
are located outside the area of modern surface samplesin the diagram, thus indicating that these vegetationtypes have no modern regional analogues in the region.At 9000 cal yr BP the vegetation cover around the lake
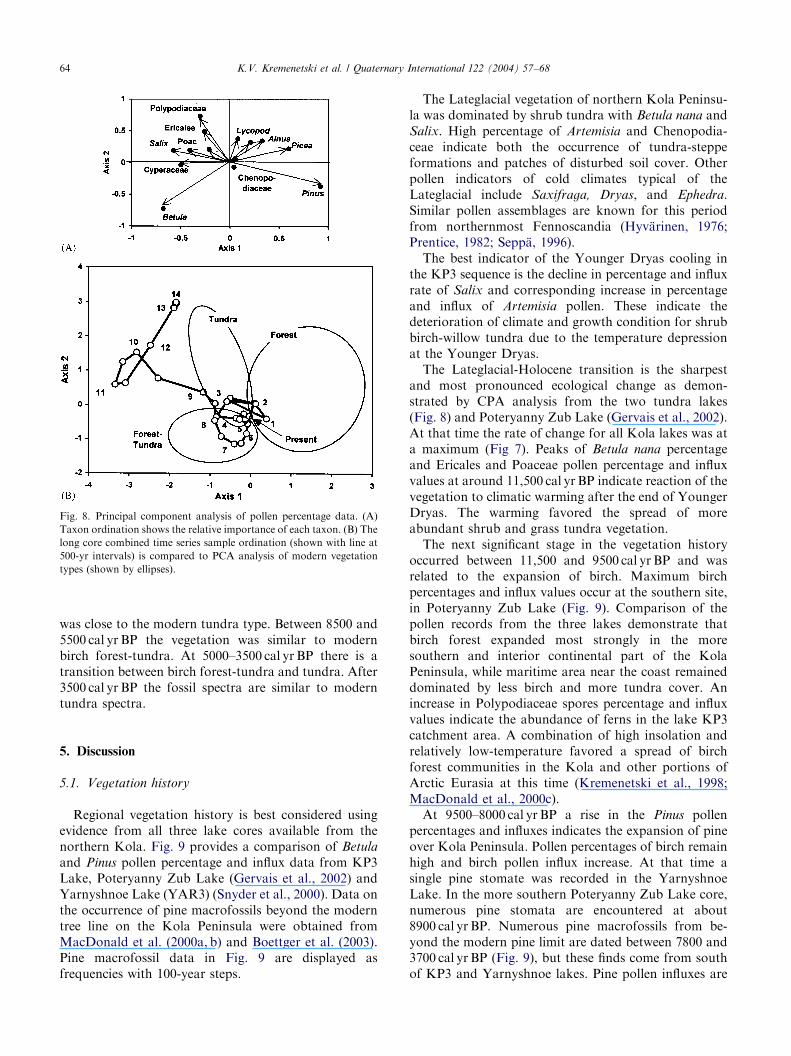
ARTICLE IN PRESS
Fig. 8. Principal component analysis of pollen percentage data. (A)
Taxon ordination shows the relative importance of each taxon. (B) The
long core combined time series sample ordination (shown with line at
500-yr intervals) is compared to PCA analysis of modern vegetation
types (shown by ellipses).
K.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6864
was close to the modern tundra type. Between 8500 and5500 cal yr BP the vegetation was similar to modernbirch forest-tundra. At 5000–3500 cal yr BP there is atransition between birch forest-tundra and tundra. After3500 cal yr BP the fossil spectra are similar to moderntundra spectra.
5. Discussion
5.1. Vegetation history
Regional vegetation history is best considered usingevidence from all three lake cores available from thenorthern Kola. Fig. 9 provides a comparison of Betula
and Pinus pollen percentage and influx data from KP3Lake, Poteryanny Zub Lake (Gervais et al., 2002) andYarnyshnoe Lake (YAR3) (Snyder et al., 2000). Data onthe occurrence of pine macrofossils beyond the moderntree line on the Kola Peninsula were obtained fromMacDonald et al. (2000a, b) and Boettger et al. (2003).Pine macrofossil data in Fig. 9 are displayed asfrequencies with 100-year steps.
The Lateglacial vegetation of northern Kola Peninsu-la was dominated by shrub tundra with Betula nana andSalix. High percentage of Artemisia and Chenopodia-ceae indicate both the occurrence of tundra-steppeformations and patches of disturbed soil cover. Otherpollen indicators of cold climates typical of theLateglacial include Saxifraga, Dryas, and Ephedra.Similar pollen assemblages are known for this periodfrom northernmost Fennoscandia (Hyv.arinen, 1976;Prentice, 1982; Sepp.a, 1996).The best indicator of the Younger Dryas cooling in
the KP3 sequence is the decline in percentage and influxrate of Salix and corresponding increase in percentageand influx of Artemisia pollen. These indicate thedeterioration of climate and growth condition for shrubbirch-willow tundra due to the temperature depressionat the Younger Dryas.The Lateglacial-Holocene transition is the sharpest
and most pronounced ecological change as demon-strated by CPA analysis from the two tundra lakes(Fig. 8) and Poteryanny Zub Lake (Gervais et al., 2002).At that time the rate of change for all Kola lakes was ata maximum (Fig 7). Peaks of Betula nana percentageand Ericales and Poaceae pollen percentage and influxvalues at around 11,500 cal yr BP indicate reaction of thevegetation to climatic warming after the end of YoungerDryas. The warming favored the spread of moreabundant shrub and grass tundra vegetation.The next significant stage in the vegetation history
occurred between 11,500 and 9500 cal yr BP and wasrelated to the expansion of birch. Maximum birchpercentages and influx values occur at the southern site,in Poteryanny Zub Lake (Fig. 9). Comparison of thepollen records from the three lakes demonstrate thatbirch forest expanded most strongly in the moresouthern and interior continental part of the KolaPeninsula, while maritime area near the coast remaineddominated by less birch and more tundra cover. Anincrease in Polypodiaceae spores percentage and influxvalues indicate the abundance of ferns in the lake KP3catchment area. A combination of high insolation andrelatively low-temperature favored a spread of birchforest communities in the Kola and other portions ofArctic Eurasia at this time (Kremenetski et al., 1998;MacDonald et al., 2000c).At 9500–8000 cal yr BP a rise in the Pinus pollen
percentages and influxes indicates the expansion of pineover Kola Peninsula. Pollen percentages of birch remainhigh and birch pollen influx increase. At that time asingle pine stomate was recorded in the YarnyshnoeLake. In the more southern Poteryanny Zub Lake core,numerous pine stomata are encountered at about8900 cal yr BP. Numerous pine macrofossils from be-yond the modern pine limit are dated between 7800 and3700 cal yr BP (Fig. 9), but these finds come from southof KP3 and Yarnyshnoe lakes. Pine pollen influxes are
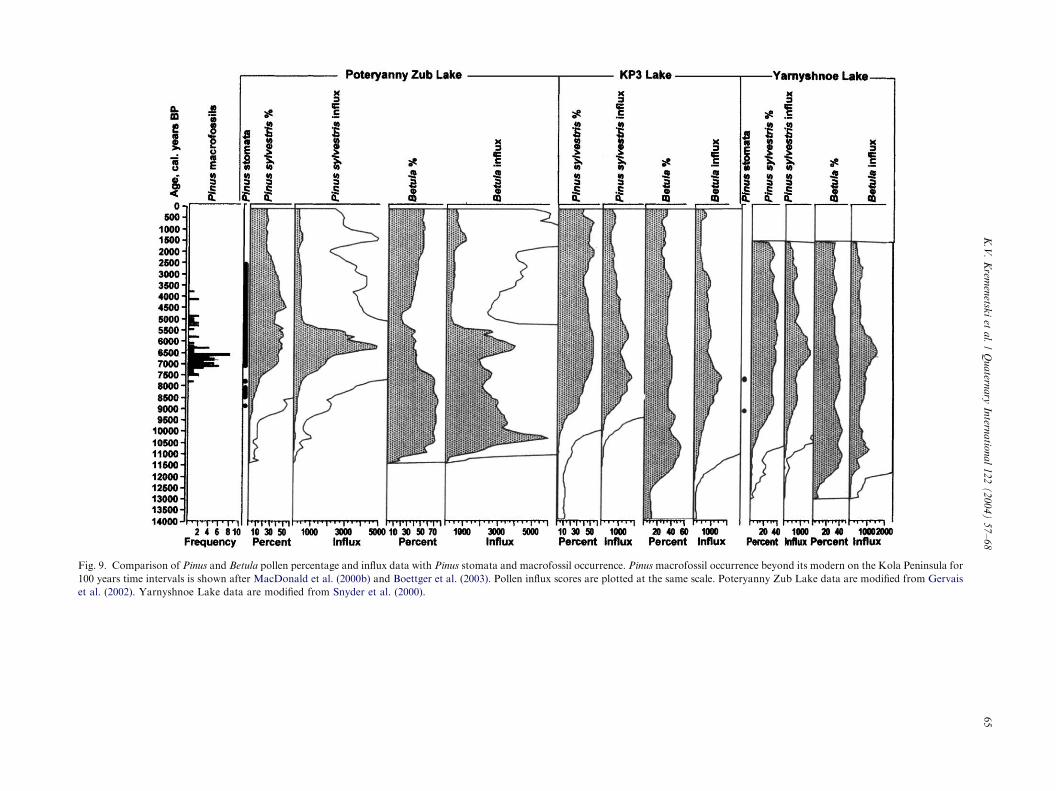
ARTIC
LEIN
PRES
S
Fig. 9. Comparison of Pinus and Betula pollen percentage and influx data with Pinus stomata and macrofossil occurrence. Pinusmacrofossil occurrence beyond its modern on the Kola Peninsula for
100 years time intervals is shown after MacDonald et al. (2000b) and Boettger et al. (2003). Pollen influx scores are plotted at the same scale. Poteryanny Zub Lake data are modified from Gervais
et al. (2002). Yarnyshnoe Lake data are modified from Snyder et al. (2000).
K.V
.K
remen
etski
eta
l./
Qu
atern
ary
Intern
atio
na
l1
22
(2
00
4)
57
–6
865

ARTICLE IN PRESSK.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6866
never as high at the two northern lakes as they are atPoteryanny Zub Lake in the mid-Holocene. It can beconcluded that although a few pines may have grown inthe coastal tundra area in the mid-Holocene, pine forestnever occupied this zone in the Holocene.High birch pollen influx suggests that birch tree
communities covered the area around KP3 and Yar-nyshnoe Lake in the early and mid-Holocene. Birchforest-tundra thus reached the coastal Barents Sea area.Birch pollen influx scores are close to the modern valuesfrom the birch forest-tundra belt (Gervais and MacDo-nald, 2001 and Fig. 9). This interpretation is supportedby the results of the PCA analysis (Fig. 8). Alnus polleninflux peaks at about 7800 cal yr BP. That suggests thatAlnus incana was present as an admixture to birch in wetplaces.A dramatic decline in Polypodiaceae percentages and
influx values after 8000 cal yr BP could indicate drierconditions. That evidence correlates well with datasuggesting lower lake levels in northernmost Fennos-candia in the mid-Holocene (Eronen et al., 1999b;Korhola et al., 2000). Lower lake levels are alsoreconstructed for the northwest Kola Peninsula at thetime of Scots pine expansion beyond its modern limit(Boettger et al., 2003).Between 5300 and 3100 cal yr BP no major changes in
the vegetation cover of the area around KP3 andYarnyshnoe lakes occurred. A decline in the Pinus
pollen influx (Fig. 4) reflects rather dramatic changes invegetation cover in the more southern parts of the KolaPeninsula. At 5300–5000 cal yr BP the dramatic decreasein Pinus and Betula pollen influx occurred in thePoteryanny Zub Lake sequence (Fig. 9). This likelyreflects a degradation of pine forests. After 3700 ca-l yr BP no pine macrofossils are found beyond moderntree line on the Kola Peninsula (MacDonald et al.,2000b; Boettger et al., 2003) and stomata are absentfrom the Poteryanny Zub core.By 3100 cal yr BP modern tundra cover was estab-
lished around KP3. This conclusion is supported byPCA analysis (Fig. 8). At the same time, birch forest-tundra became dominant in the region around Poter-yanny Zub Lake (Gervais et al., 2002). PCA suggests aslight reversion of vegetation toward warmer types atB1000 cal yr BP (Fig. 8). There is similar evidence fromPoteryanny Zub Lake (Gervais et al., 2002). That mightreflect the Medieval warming recorded in interior part ofthe Kola Peninsula at around 1000 AD, when the upperpine limit in Khibiny Mountains was 100–120m higherthan its present position (Hiller et al., 2001).Recently obtained data demonstrate a synchronous
change in the vegetation pattern in forest, forest-tundraand tundra belts of the Kola Peninsula. Data from theKola Peninsula show a large similarity to the establishedtrend in the vegetation history in northernmost Fennos-candia. The development of forest cover on the Kola
Peninsula and northward expansion of northern pinelimit and of birch forest-tundra were favoured by earlyHolocene warming and increase of North Atlantic seasurface temperature (Ko@ et al., 1993; Hald et al., 1999).Oxygen and carbon isotope analysis of foraminiferalassemblages from northern Barents Sea indicate anincrease in water temperature from 10,000 cal yr BPculminating in a temperature optimum between 7800and 6800 cal yr BP (Duplessy et al., 2001). That corre-sponds well to the optimal climate conditions inferredfor the northern Kola Peninsula. The mid-Holocenewarming for northern Sweden and the interior partof the Kola Peninsula is estimated as 2�C (Barnekow,1999; Gervais et al., 2002). The warming in the coastalarea was less pronounced and was within the range of1–1.5�C, which is partly explained by a cooling effectof Barents Sea. The modern tundra belt on theKola Peninsula was formed as a result of late Holoceneclimate deterioration and southward retreat of pinetree line and birch forests around 3100 cal yr BP.This event was synchronous all over the northernEurasia (Kremenetski et al., 1998; MacDonald et al.,2000c) and is related to the end of mid-Holocenewarming due to decreasing summer insolation and seasurface temperatures in the North Atlantic (Gano-polski et al., 1998; MacDonald et al. 2000a; Brovkinet al., 2003).
Acknowledgements
We would like to thank Academician GennadyMatishov and his colleagues at the Murmansk MarineBiological Institute of Russian Academy of Sciences fortheir logistical help. We thank David Porinchu at theUCLA Geography Department for very helpful com-ments on this manuscript. We are most grateful to KeithBennett, Matti Saarnisto and Achim Brauer for valuablereviews. Financial support for the research was providedby the NSF Paleoclimatology of Arctic Lakes andEstuaries (PALE) grant (ATM-9632926). This paper isPARCS contribution # .
References
Atlas Murmanskoi Oblasti, 1971. Moscow, State Administration of
Geodesy and Cartography (GUGK), 33pp.
Barnekow, L., 1999. Holocene tree-line dynamics and inferred climatic
changes in the Abisko area, northern Sweden, based on macrofossil
and pollen records. The Holocene 9, 253–265.
Berglund, B.E., Ralska-Jasiewiczowa, M., 1986. Pollen analysis and
pollen diagrams. In: Berglund, B.E. (Ed.), 1986. Handbook of
Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester,
pp. 455–484.

ARTICLE IN PRESSK.V. Kremenetski et al. / Quaternary International 122 (2004) 57–68 67
Birks, H.J.B., 1968. The identification of Betula nana pollen. New
Phytologist 67, 309–314.
Boettger, T., Hiller, A., Kremenetski, K., 2003. Mid-Holocene
warming in the northwest Kola Peninsula, Russia: northern pine
limit movement and stable isotope evidence. The Holocene 13,
403–410.
Brovkin, V., Levis, S., Loutre, M.-F., Crucifix, M., Claussen, M.,
Ganopolski, A., Kubatzki, C., Petoukhov, P., 2003. Stability
analysis of the climate-vegetation system in the northern high
latitudes. Climatic Research 57, 119–138.
Czerepanov, S.K., 1995. Vascular Plants of Russia and Adjacent States
(The former USSR). Cambridge University Press, Cambridge
(516pp).
Dean, W.E., 1974. Determination of carbonate and organic matter in
calcareous sediments and sedimentary rocks by loss on ignition:
comparison with other methods. Journal of Sediment Petrology 44,
242–248.
Duplessy, J.-C., Ivanova, E., Murdmaa, I., Paterne, M., Labeyrie, L.,
2001. Holocene paleoceanography of the northern Barents Sea and
variations of the northward heat transport by Atlantic Ocean.
Boreas 30, 2–16.
Eronen, M., 1979. The retreat of pine forest in Finnish Lapland
since the Holocene climatic optimum: a general discussion
with radiocarbon evidence from subfossil pines. Fennia 157,
93–114.
Eronen, M., Lindholm, M., Saastamoinen, S., Zetterberg, P., 1999a.
Variable Holocene treeline dynamics and changes in natural
environments in northern Finnish Lapland. Chemosphere: Global
Change Science 1, 377–387.
Eronen, M., Hyv.arinen, H., Zetterberg, P., 1999b. Holocene humidity
changes in northern Finnish Lapland inferred from lake sediments
and submerged Scots pines dated by tree-rings. The Holocene 9,
569–580.
Faegri, K., Iversen, J., 1989. Textbook of Pollen Analysis 4th Edition.
Wiley, Chichester (328pp).
Ganopolski, A., Kubatzki, C., Claussen, M., Brovkin, V., Petoukhov,
V., 1998. The influence of vegetation–atmosphere–ocean in-
teraction on climate during the mid-Holocene. Science 280,
1916–1919.
Gervais, B.R., MacDonald, G.M., 2001. Modern pollen and stomate
deposition in lake across treeline on the Kola Peninsula, Russia.
Review of Palaeobotany and Palynology 114, 223–237.
Gervais, B.R., MacDonald, G.M., Snyder, J.A., Kremenetski, K.V.,
2002. Pinus sylvestris treeline development and movement on the
Kola Peninsula of Russia: pollen and stomate evidence. Journal of
Ecology 90, 627–638.
Grimm, E.C., 1990. TILIA and TILIA�GRAPH.PC spreadsheet andgraphics software for pollen data. INQUA, Working Group on
Data-Handling Methods, Newsletter 4, pp. 5–7.
Hald, M., Kolstad, V., Olyak, L., Forman, S.L., Herlihy, F.A.,
Ivanov, G., Nescheretov, A., 1999. Lateglacial and Holocene
paleoceanography and sedimentary environmennts in the St. Anna
Trough, eurasian Arctic Margin. Paleogeography, Paleoclimatol-
ogy, Paleoecology 146, 229–249.
Hiller, A., Boettger, T., Kremenetski, K., 2001. Medieval climate
warming recorded by radiocarbon-dated alpine tree-line shift on
the Kola Peninsula, Russia. The Holocene 11, 491–497.
Hyv.arinen, H., 1976. Flandrian pollen deposition rates and tree-line
history in northernmost Fennoscandia. Boreas 5, 163–175.
Hyv.arinen, H., 1993. Holocene pine and birch limits near Kilpisj.arvi,
Western Finnish Lapland: pollen stratigraphical evidence. Pal-
.aoklimaforschung 9, 19–27.
Ko@, N., Jansen, E., Haflidason, H., 1993. Paleoceonographicreconstructions of surface ocean conditions in the Greenland,
Iceland and Norwegian seas through the last 14 ka based on
diatoms. Quaternary Science Reviews 12, 115–140.
Korhola, A., Olander, H., Blom, T., 2000. Cladoceran and chironomid
assemblages as quantitative indicators of water depth in sub-Arctic
Fennoscandian lakes. Journal of Palaeolimnology 24, 43–54.
Kremenetski, C., Patyk-Kara, N., 1997. Holocene vegetation dyna-
mics of the southeast Kola Peninsula, Russia. The Holocene 7,
473–479.
Kremenetski, C., Vaschalova, T., Goriachkin, S., Cherkinsky, A.,
Sulerzhitsky, L., 1997. Holocene pollen stratigraphy and bog
development in the western part of the Kola Peninsula, Russia.
Boreas 26, 91–102.
Kremenetski, C.V., Sulerzhitsky, L.D., Hantemirov, R., 1998.
Holocene history of the northern range limits of some trees and
shrubs in Russia. Arctic and Alpine Research 30, 317–333.
Kremenetski, C.V., Vaschalova, T., Sulerzhitsky, L., 1999. The
Holocene vegetation history of the Khibiny mountains: implica-
tions for the post-glacial expansion of spruce and alder on the Kola
Peninsula, northwestern Russia. Journal of Quaternary Science 14,
29–43.
MacDonald, G.M., Edwards, T.W.D., Moser, K.A., Pienitz, R., Smol,
J.P., 1993. Rapid response of treeline vegetation to past climate
warming. Nature 361, 243–246.
MacDonald, G.M., Felzer, B., Finney, B.P., Forman, S.L., 2000a.
Holocene lake records of arctic hydrology. Journal of Paleolimnol-
ogy 24, 1–14.
MacDonald, G.M., Gervais, B.R., Snyder, J.A., Tarasov, G.A.,
Borisova, O.K., 2000b. Radiocarbon dated Pinus sylvestris L.
wood from beyond tree-line on the Kola Peninsula, Russia. The
Holocene 10, 143–147.
MacDonald, G.M., Velichko, A.A., Kremenetski, K.V., Borisova,
O.K., Goleva, A.A., Andreev, A.A., Cwynar, L.S., Riding, R.T.,
Forman, S.L., Edwads, T.W.D., Aravena, R., Hammarlund, D.,
Szeicz, J.M., Gattaulin, V., 2000c. Holocene treeline history and
climate change across Northern Eurasia. Quaternary Research 53,
302–311.
Maher Jr., L.J., 1972. Nomograms for computing 95% limits of pollen
data. Review of Palaeobotany and Palynology 13, 85–93.
Moore, P.D., Webb, J.A., Collinson, M.E., 1991. Pollen analysis, 2nd
Edition. Oxford Blackwell Scientific Publications, London, Edin-
bourgh, Boston, Melbourne, Paris, Berlin, Vienna, 216pp.
Prentice, H.C., 1982. Late Weichselian and early Flandrian vegeta-
tional history of Varanger peninsula, northeast Norway. Boreas 11,
187–208.
Rainio, H., Saarnisto, M., Ekman, I., 1995. Younger Dryas end
moraines in Finland and NW Russia. Quaternary International 28,
179–192.
Reille, M., 1998. Pollen et spores d’Europe et de l’Afrique du Nord.
Laboratoire de Botanique Historique et Palynologie, Marseille,
520pp.
Sepp.a, H., 1996. Post-glacial dynamics of vegetation and tree-line in
the far north of Fennoscandia. Fennia 174 (1), 1–96.
Sepp.a, H., Hammarlund, D., 2000. Pollen-stratigraphical evidence of
Holocene hydrological change in northern Fennoscandia sup-
ported by independent isotopic data. Journal of Paleolimnology 24,
69–79.
Snyder, J.A., MacDonald, G.M., Forman, S.L., Tarasov, G.A., Mode,
W.N., 2000. Postglacial climate and vegetation history, north-
central Kola Peninsula, Russia: pollen and diatom records from
lake yarnyshnoe-3. Boreas 29, 261–271.
Stockmarr, J., 1971. Tablets with spores used in absolute pollen
analysis. Pollen and Spores 13, 615–621.
Stuiver, M., Reimer, P., Bard, E., Warren-Beck, J., Burr, G.S.,
Hughen, K.A., Kromer, B., McCormac, G., van der Plicht, J.,
Spurk, M., 1998. INTCAL 98 Radiocarbon age calibration,
24,000–0 cal BP. Radiocarbon 40, 1041–1083.
Terasm.ae, J., 1951. On the pollen morphology of Betula nana. Svensk
Botanisk Tidskrift 45, 358–361.

ARTICLE IN PRESSK.V. Kremenetski et al. / Quaternary International 122 (2004) 57–6868
ter Braak, C.J.F., Smilauer, P., 1998. CANOCO Reference Manual
and User’s Guide to Canoco for Windows. Software for Canonical
Community Ordination (version 4). Centre for Biometry Wagenin-
gen (Wageningen, NL) and Microcomputer Power (Ithaca NY,
USA), 352pp.
Yelina, G.F., Arslanov, Kh.A., Klimanov, V.A., Usova, L.I., 1995.
Paleorastitelnost i klimatostratigrafiya golotsena Lovozetskoi
ravniny Kolskogo poluostrova (po sporovo-pyltsevym diagram-
mam bugristo-topyanogo bolota). Botanicheskiy Zhurnal 80 (3),
1–16.
Yelina, G.F., Lukashov, A.D., Yurkovskaya, T.K., 2000. Pozdneled-
nikovye i golotsen Vostochnoi Fennoscandii (paleorastitelnost i
paleogeografiya). Petrozavodsk, Karelskiy Nauchnyi tsentr RAN,
240pp.





























