Embed Size (px)
Citation preview

of September 25, 2014.This information is current as
Transcription Factors Sp1 and Sp3IL-10 Gene Expression Is Controlled by the
Thompson and Herman WaldmannMasahide Tone, Mark J. Powell, Yukiko Tone, Sara A. J.
http://www.jimmunol.org/content/165/1/286doi: 10.4049/jimmunol.165.1.286
2000; 165:286-291; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/1/286.full#ref-list-1
, 27 of which you can access for free at: cites 41 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

IL-10 Gene Expression Is Controlled by the TranscriptionFactors Sp1 and Sp31
Masahide Tone,2 Mark J. Powell, Yukiko Tone, Sara A. J. Thompson, and Herman Waldmann
IL-10 is an 18-kDa cytokine with a key role in homeostatic control of inflammatory and immune responses. We have investigatedhow transcription of the IL-10 gene is regulated, so as to be able to understand the circumstances of IL-10 expression in bothhealth and disease. In the mouse, IL-10 gene expression is regulated by a TATA-type promoter with a criticalcis-acting elementcontaining GGA repeats located at289 to 277. Its complementary sequence is similar to thecis-acting elements (TCC repeats)in the promoters of genes encoding epidermal growth factor receptor and CD58. All these elements comprise a common CCTCCTsequence with less conserved C1 T-rich sequences. Eliminating this CCTCCT sequence results in a marked reduction in pro-moter activity, suggesting a necessary role in IL-10 gene expression. Despite its dissimilarity to the G1 C-rich Sp1 consensussequence (GC box), Sp1 and Sp3 transcription factors could be shown to bind to this motif. The requirement for Sp1 and Sp3 intranscription of IL-10 was confirmed using DrosophilaSL2 cells, which lack endogenous Sp factors. These results suggest that thetranscription of IL-10 is positively regulated by both Sp1 and Sp3. The Journal of Immunology,2000, 165: 286–291.
I nterleukin-10 is an 18-kDa glycoprotein (1, 2), initially iden-tified as a product of Th2 cells that has subsequently beenshown to be produced by a wide range of cell types, includ-
ing Th1 cells, Tr1 cells, B cells, monocytes, macrophages, kera-tinocytes, and many tumor cells (2–8). It behaves like a potentantiinflammatory and immunosuppressive molecule, inhibiting theproduction and release of IL-2 and IFN-g and a range of proin-flammatory cytokines such as TNF-a, IL-1, IL-6, and IL-12 (2, 9).Despite its crucial role in immunoregulation, the factors regulatingexpression of IL-10 gene are poorly understood at both the tran-scriptional and posttranscriptional levels. Control at the posttran-scriptional level has been implied from the identification of poten-tial mRNA-destabilizing sequences (AUUUA with A1 U-richsequences) in 39-untranslated regions of mouse and human IL-10mRNA (10–12). In the accompanying paper (13), we show directevidence for such posttranscriptional control. In this paper, how-ever, we examine how IL-10 is controlled at the transcriptionallevel.
The mouse IL-10 gene was cloned, and its DNA sequence in-cluding the 59-flanking region was determined (14). This genecomprises five exons distributed over 5.1 kb of DNA. A typicalTATA box sequence is located 97 bp upstream of the first ATGwithin a corresponding region previously shown to have promoteractivity for human IL-10 (15).
We have made the novel and surprising observation that, unlikemost other cytokines, IL-10 gene expression is regulated by theconstitutively and ubiquitously expressed Sp1 and Sp3 as key tran-scription factors. This may explain how the IL-10 gene can con-
tinue to be transcribed when other cytokine genes are inactive.This combination of constitutive expression and fine control at theposttranscriptional level may permit a rapid homeostatic responseto inflammation.
Materials and MethodsReverse transcriptase-PCR
IL-10 mRNA levels were determined by RT-PCR. First strand cDNA wasprepared from 1mg total RNA using an oligo(dT) primer. This reactionmixture (20ml) was diluted with 80 or 180ml water, and then 0, 5, 10, or20 ml of the cDNA solution were utilized for PCR amplification usingsense and antisense primers (IL-10; 25 cycles, hypoxanthine phosphoribo-syltransferase (HPRT)3; 17 cycles). The primer sequences used were:IL-10 sense primer, CCAGTTTTACCTGGTAGAAGTGATG; IL-10 an-tisense primer, TGTCTAGGTCCTGGAGTCCAGCAGACTC; HPRTsense primer, ACAGCCCCAAAATGGTTAAGG; and HPRT antisenseprimer, TCTGGGGACGCAGCAACTGAC. The amplified cDNAs weredetected by Southern blot hybridization using cDNA probes.
Mapping of transcription start sites
To determine transcription start sites, we performed the rapid amplificationof cDNA ends procedure (RACE) as described previously (16). cDNA for59-RACE was prepared using an antisense primer (GGAGTCGGTTAGCAGTATGTTGTCC) and 10mg total RNA. After adding a G-tail at 59-end, IL-10 cDNA was amplified using a poly(C) primer and an antisenseprimer (CACCTGGCTGAAGGCAGTCCGCAGCTC). Amplified frag-ments were cloned, and the DNA sequences were determined.
Assessment of promoter activity using the luciferase assay
Luciferase reporter plasmids were constructed using deletion mutants ofthe 59-flanking region of the IL-10 gene and pGL3-Basic Vector (Promega,Madison, WI). This 59-flanking fragment was subcloned from al IL-10genomic clone, a kind gift from Dr. K. Moore (DNAX Research Institute,Palo Alto, CA). These deletion mutants (D1 to D11) were generated by theexonuclease III/mung bean nuclease procedure or by PCR. The structuresof these deletion mutants are indicated in Fig. 2A. The resulting deletionmutants,KpnI/SacI fragments, were cloned upstream of the luciferase genein pGL3-Basic Vector. TheSacI site (39-end of the deletion mutants) islocated at163 in the IL-10 gene.
To construct plasmid D1 CCT KO and D6 CCT KO, the CCTCCTsequences (285 to280) in plasmid D1 and D6 were replaced byGAATTC (EcoRI site) by PCR.
Sir William Dunn School of Pathology, University of Oxford, Oxford, UnitedKingdom
Received for publication December 21, 1999. Accepted for publication April13, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Medical Research Council and The WellcomeTrust.2 Address correspondence and reprint requests to Dr. Masahide Tone, Sir WilliamDunn School of Pathology, University of Oxford, South Parks Road, Oxford, OX13RE, U.K. E-mail address: [email protected]
3 Abbreviations used in this paper: HPRT, hypoxanthine phosphoribosyltransferase;RACE, the rapid amplification of cDNA ends procedure; bmDC, bone marrow-de-rived dendritic cell; EGFR, epidermal growth factor receptor.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
EL-4 and RAW 264 cells (23 107) were transfected by electroporationusing 10mg of luciferase reporter plasmids with 0.1mg pRL-SV40 orpRL-TK (Promega) as an internal control plasmid. If required, cells werestimulated with PMA (50 ng/ml)/ionomycin (1mM) or LPS (20mg/ml) 7 hpostelectroporation. Cells were harvested 48 h postelectroporation, andpromoter activities were analyzed by the Dual-luciferase Reporter AssaySystem (Promega). These assays were repeated more than three times, andthe activities were normalized toRenilla luciferase activities.
DrosophilaSL2 cells (23 106) were transfected using 0.5mg of theluciferase reporter plasmid containing the IL-10 promoter (D6;2305 to163) with pPac, pPacSp1, and pPacUSp3. Scheider SL2 cells were ob-tained from American Type Culture Collection (Manassas, VA). The ex-pression plasmids were kind gifts from Dr. G. Suske (Philipps-Universitat,Marburg, Germany). Transfection was performed using Pfx-20 transfectionreagent (Promega). Luciferase activities were analyzed 48 h posttransfec-tion. These assays were repeated more than three times.
Gel mobility band shift assays
A nuclear extract from EL-4 cells was prepared using methods describedby Dignam et al. (17). Gel mobility band shift assay was performed asdescribed previously (18). To perform super gel mobility band shift assay,the reaction mixture was incubated with anti-Sp1 (Santa Cruz, PEP2)and/or anti-Sp3 (Santa Cruz D-20) Abs.
DNase I footprinting
A DNase I footprinting assay was performed according to the methoddescribed by Dynan andTjian (19). A DNA fragment of the promoter regionwas amplified by PCR using a32P-labeled primer (CTGAAGGCTCAGTGGGGCCTTCC) and an unlabeled primer (CCAGTTCTTTAGCGCTTACAATGC). The PCR product was isolated and incubated with 0–50mg of anuclear extract in 20 mM HEPES (pH 7.9), 2 mM MgCl2, 50 mM NaCl, 1 mMDTT, 20% glycerol, and 4mg poly[d(I-C)]-poly[d(I-C)]. The mixtures weretreated with DNase I at 25°C for 1 min. Purified DNAs from these mixtureswere analyzed by sequencing gel with sequence ladders generated using thesame labeled primer.
Resultscis-Acting elements for IL-10 gene expression
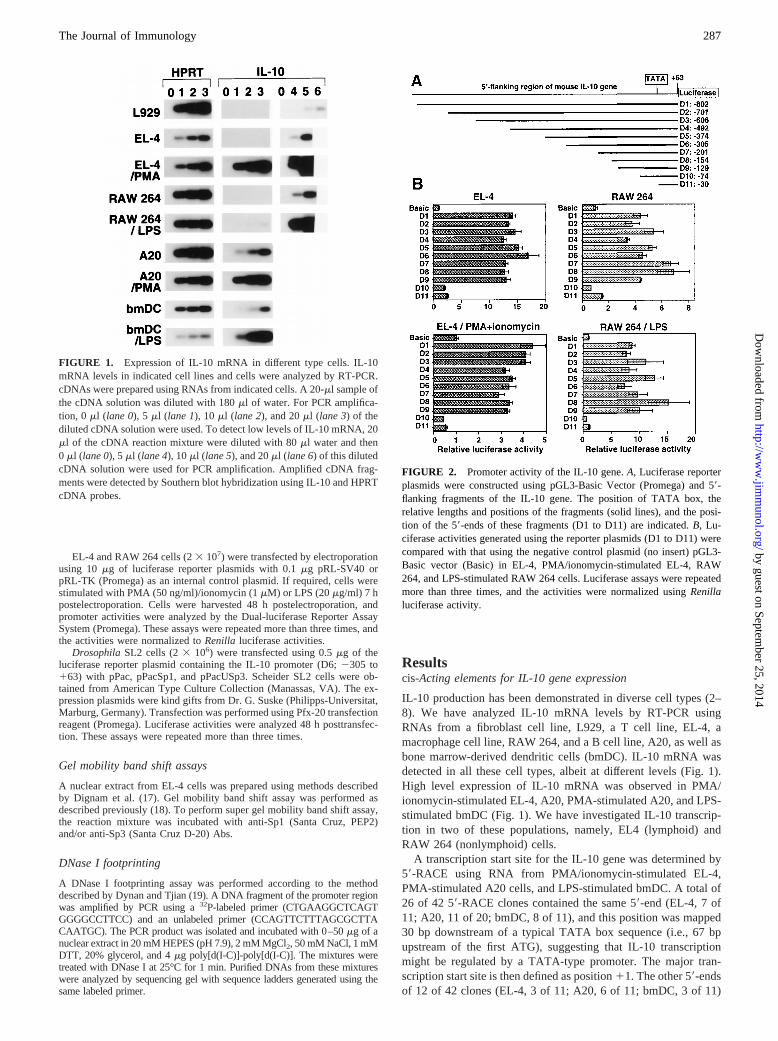
IL-10 production has been demonstrated in diverse cell types (2–8). We have analyzed IL-10 mRNA levels by RT-PCR usingRNAs from a fibroblast cell line, L929, a T cell line, EL-4, amacrophage cell line, RAW 264, and a B cell line, A20, as well asbone marrow-derived dendritic cells (bmDC). IL-10 mRNA wasdetected in all these cell types, albeit at different levels (Fig. 1).High level expression of IL-10 mRNA was observed in PMA/ionomycin-stimulated EL-4, A20, PMA-stimulated A20, and LPS-stimulated bmDC (Fig. 1). We have investigated IL-10 transcrip-tion in two of these populations, namely, EL4 (lymphoid) andRAW 264 (nonlymphoid) cells.
A transcription start site for the IL-10 gene was determined by59-RACE using RNA from PMA/ionomycin-stimulated EL-4,PMA-stimulated A20 cells, and LPS-stimulated bmDC. A total of26 of 42 59-RACE clones contained the same 59-end (EL-4, 7 of11; A20, 11 of 20; bmDC, 8 of 11), and this position was mapped30 bp downstream of a typical TATA box sequence (i.e., 67 bpupstream of the first ATG), suggesting that IL-10 transcriptionmight be regulated by a TATA-type promoter. The major tran-scription start site is then defined as position11. The other 59-endsof 12 of 42 clones (EL-4, 3 of 11; A20, 6 of 11; bmDC, 3 of 11)
FIGURE 1. Expression of IL-10 mRNA in different type cells. IL-10mRNA levels in indicated cell lines and cells were analyzed by RT-PCR.cDNAs were prepared using RNAs from indicated cells. A 20-ml sample ofthe cDNA solution was diluted with 180ml of water. For PCR amplifica-tion, 0 ml (lane 0), 5ml (lane 1), 10ml (lane 2), and 20ml (lane 3) of thediluted cDNA solution were used. To detect low levels of IL-10 mRNA, 20ml of the cDNA reaction mixture were diluted with 80ml water and then0 ml (lane 0), 5ml (lane 4), 10ml (lane 5), and 20ml (lane 6) of this dilutedcDNA solution were used for PCR amplification. Amplified cDNA frag-ments were detected by Southern blot hybridization using IL-10 and HPRTcDNA probes.
FIGURE 2. Promoter activity of the IL-10 gene.A, Luciferase reporterplasmids were constructed using pGL3-Basic Vector (Promega) and 59-flanking fragments of the IL-10 gene. The position of TATA box, therelative lengths and positions of the fragments (solid lines), and the posi-tion of the 59-ends of these fragments (D1 to D11) are indicated.B, Lu-ciferase activities generated using the reporter plasmids (D1 to D11) werecompared with that using the negative control plasmid (no insert) pGL3-Basic vector (Basic) in EL-4, PMA/ionomycin-stimulated EL-4, RAW264, and LPS-stimulated RAW 264 cells. Luciferase assays were repeatedmore than three times, and the activities were normalized usingRenillaluciferase activity.
287The Journal of Immunology
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
and 4 of 42 clones (EL-4, 1 of 11; A20, 3 of 20) were mapped at12 and13, respectively.
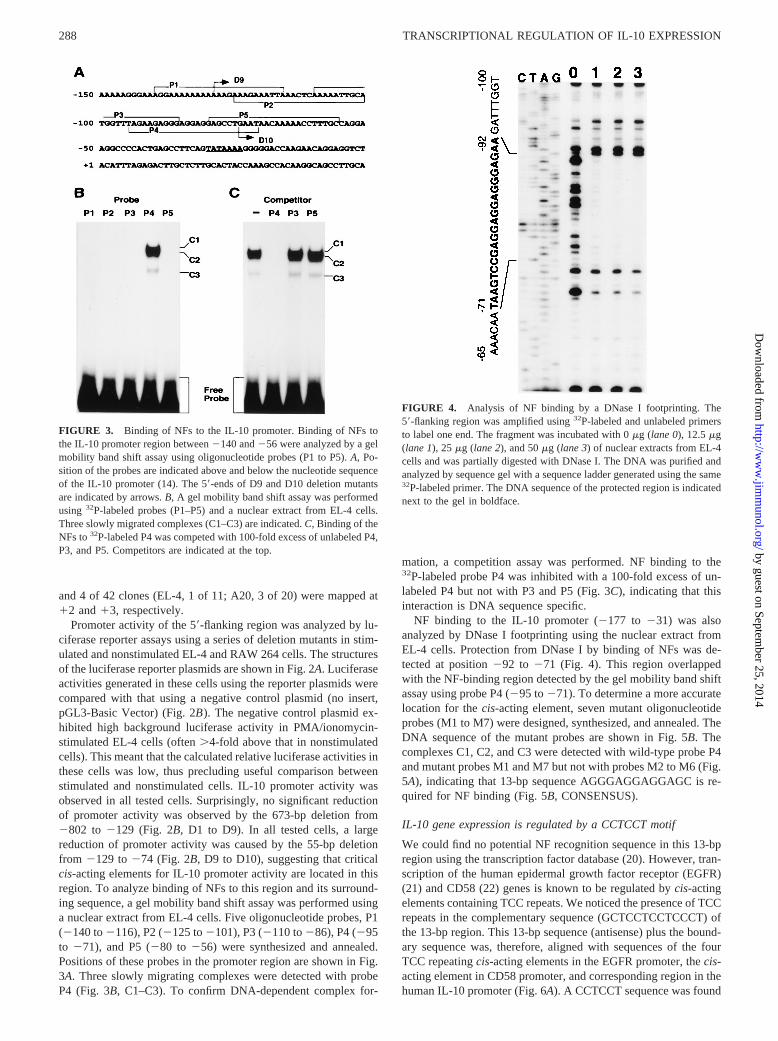
Promoter activity of the 59-flanking region was analyzed by lu-ciferase reporter assays using a series of deletion mutants in stim-ulated and nonstimulated EL-4 and RAW 264 cells. The structuresof the luciferase reporter plasmids are shown in Fig. 2A. Luciferaseactivities generated in these cells using the reporter plasmids werecompared with that using a negative control plasmid (no insert,pGL3-Basic Vector) (Fig. 2B). The negative control plasmid ex-hibited high background luciferase activity in PMA/ionomycin-stimulated EL-4 cells (often.4-fold above that in nonstimulatedcells). This meant that the calculated relative luciferase activities inthese cells was low, thus precluding useful comparison betweenstimulated and nonstimulated cells. IL-10 promoter activity wasobserved in all tested cells. Surprisingly, no significant reductionof promoter activity was observed by the 673-bp deletion from2802 to 2129 (Fig. 2B, D1 to D9). In all tested cells, a largereduction of promoter activity was caused by the 55-bp deletionfrom 2129 to274 (Fig. 2B, D9 to D10), suggesting that criticalcis-acting elements for IL-10 promoter activity are located in thisregion. To analyze binding of NFs to this region and its surround-ing sequence, a gel mobility band shift assay was performed usinga nuclear extract from EL-4 cells. Five oligonucleotide probes, P1(2140 to2116), P2 (2125 to2101), P3 (2110 to286), P4 (295to 271), and P5 (280 to256) were synthesized and annealed.Positions of these probes in the promoter region are shown in Fig.3A. Three slowly migrating complexes were detected with probeP4 (Fig. 3B, C1–C3). To confirm DNA-dependent complex for-
mation, a competition assay was performed. NF binding to the32P-labeled probe P4 was inhibited with a 100-fold excess of un-labeled P4 but not with P3 and P5 (Fig. 3C), indicating that thisinteraction is DNA sequence specific.
NF binding to the IL-10 promoter (2177 to 231) was alsoanalyzed by DNase I footprinting using the nuclear extract fromEL-4 cells. Protection from DNase I by binding of NFs was de-tected at position292 to 271 (Fig. 4). This region overlappedwith the NF-binding region detected by the gel mobility band shiftassay using probe P4 (295 to271). To determine a more accuratelocation for thecis-acting element, seven mutant oligonucleotideprobes (M1 to M7) were designed, synthesized, and annealed. TheDNA sequence of the mutant probes are shown in Fig. 5B. Thecomplexes C1, C2, and C3 were detected with wild-type probe P4and mutant probes M1 and M7 but not with probes M2 to M6 (Fig.5A), indicating that 13-bp sequence AGGGAGGAGGAGC is re-quired for NF binding (Fig. 5B, CONSENSUS).
IL-10 gene expression is regulated by a CCTCCT motif
We could find no potential NF recognition sequence in this 13-bpregion using the transcription factor database (20). However, tran-scription of the human epidermal growth factor receptor (EGFR)(21) and CD58 (22) genes is known to be regulated bycis-actingelements containing TCC repeats. We noticed the presence of TCCrepeats in the complementary sequence (GCTCCTCCTCCCT) ofthe 13-bp region. This 13-bp sequence (antisense) plus the bound-ary sequence was, therefore, aligned with sequences of the fourTCC repeatingcis-acting elements in the EGFR promoter, thecis-acting element in CD58 promoter, and corresponding region in thehuman IL-10 promoter (Fig. 6A). A CCTCCT sequence was found
FIGURE 3. Binding of NFs to the IL-10 promoter. Binding of NFs tothe IL-10 promoter region between2140 and256 were analyzed by a gelmobility band shift assay using oligonucleotide probes (P1 to P5).A, Po-sition of the probes are indicated above and below the nucleotide sequenceof the IL-10 promoter (14). The 59-ends of D9 and D10 deletion mutantsare indicated by arrows.B, A gel mobility band shift assay was performedusing 32P-labeled probes (P1–P5) and a nuclear extract from EL-4 cells.Three slowly migrated complexes (C1–C3) are indicated.C, Binding of theNFs to32P-labeled P4 was competed with 100-fold excess of unlabeled P4,P3, and P5. Competitors are indicated at the top.
FIGURE 4. Analysis of NF binding by a DNase I footprinting. The59-flanking region was amplified using32P-labeled and unlabeled primersto label one end. The fragment was incubated with 0mg (lane 0), 12.5mg(lane 1), 25mg (lane 2), and 50mg (lane 3) of nuclear extracts from EL-4cells and was partially digested with DNase I. The DNA was purified andanalyzed by sequence gel with a sequence ladder generated using the same32P-labeled primer. The DNA sequence of the protected region is indicatednext to the gel in boldface.
288 TRANSCRIPTIONAL REGULATION OF IL-10 EXPRESSION
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
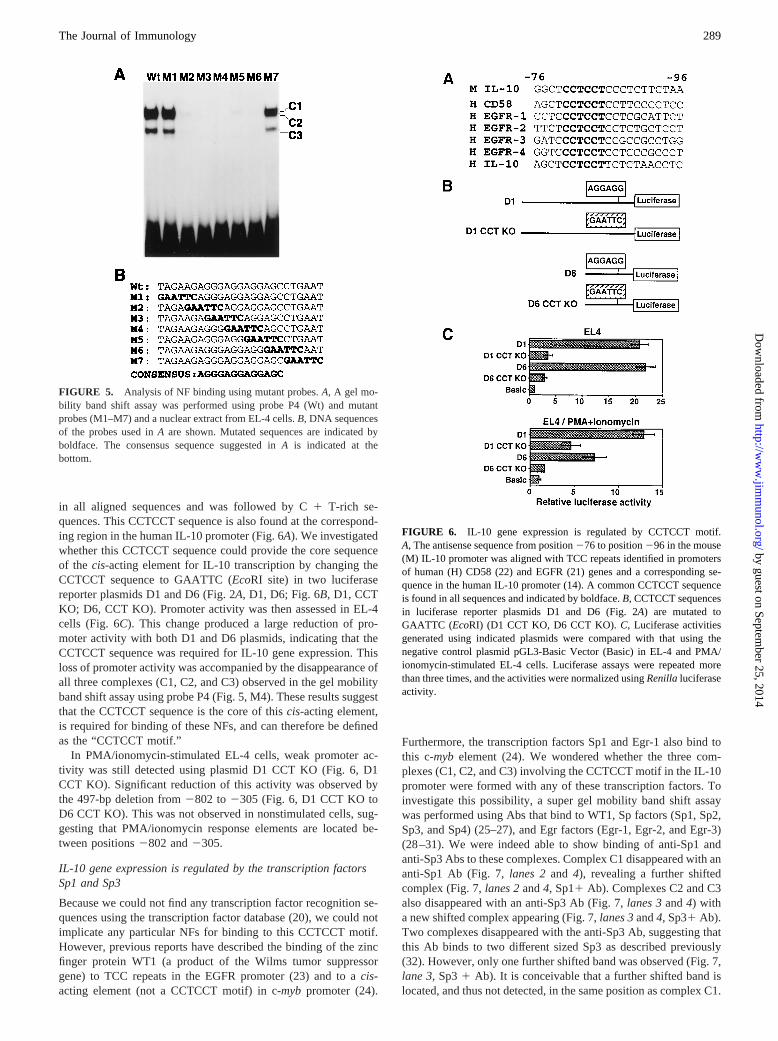
in all aligned sequences and was followed by C1 T-rich se-quences. This CCTCCT sequence is also found at the correspond-ing region in the human IL-10 promoter (Fig. 6A). We investigatedwhether this CCTCCT sequence could provide the core sequenceof the cis-acting element for IL-10 transcription by changing theCCTCCT sequence to GAATTC (EcoRI site) in two luciferasereporter plasmids D1 and D6 (Fig. 2A, D1, D6; Fig. 6B, D1, CCTKO; D6, CCT KO). Promoter activity was then assessed in EL-4cells (Fig. 6C). This change produced a large reduction of pro-moter activity with both D1 and D6 plasmids, indicating that theCCTCCT sequence was required for IL-10 gene expression. Thisloss of promoter activity was accompanied by the disappearance ofall three complexes (C1, C2, and C3) observed in the gel mobilityband shift assay using probe P4 (Fig. 5, M4). These results suggestthat the CCTCCT sequence is the core of thiscis-acting element,is required for binding of these NFs, and can therefore be definedas the “CCTCCT motif.”
In PMA/ionomycin-stimulated EL-4 cells, weak promoter ac-tivity was still detected using plasmid D1 CCT KO (Fig. 6, D1CCT KO). Significant reduction of this activity was observed bythe 497-bp deletion from2802 to2305 (Fig. 6, D1 CCT KO toD6 CCT KO). This was not observed in nonstimulated cells, sug-gesting that PMA/ionomycin response elements are located be-tween positions2802 and2305.
IL-10 gene expression is regulated by the transcription factorsSp1 and Sp3
Because we could not find any transcription factor recognition se-quences using the transcription factor database (20), we could notimplicate any particular NFs for binding to this CCTCCT motif.However, previous reports have described the binding of the zincfinger protein WT1 (a product of the Wilms tumor suppressorgene) to TCC repeats in the EGFR promoter (23) and to acis-acting element (not a CCTCCT motif) in c-mybpromoter (24).
Furthermore, the transcription factors Sp1 and Egr-1 also bind tothis c-mybelement (24). We wondered whether the three com-plexes (C1, C2, and C3) involving the CCTCCT motif in the IL-10promoter were formed with any of these transcription factors. Toinvestigate this possibility, a super gel mobility band shift assaywas performed using Abs that bind to WT1, Sp factors (Sp1, Sp2,Sp3, and Sp4) (25–27), and Egr factors (Egr-1, Egr-2, and Egr-3)(28–31). We were indeed able to show binding of anti-Sp1 andanti-Sp3 Abs to these complexes. Complex C1 disappeared with ananti-Sp1 Ab (Fig. 7,lanes 2and 4), revealing a further shiftedcomplex (Fig. 7,lanes 2and4, Sp11Ab). Complexes C2 and C3also disappeared with an anti-Sp3 Ab (Fig. 7,lanes 3and4) witha new shifted complex appearing (Fig. 7,lanes 3and4, Sp31Ab).Two complexes disappeared with the anti-Sp3 Ab, suggesting thatthis Ab binds to two different sized Sp3 as described previously(32). However, only one further shifted band was observed (Fig. 7,lane 3, Sp31 Ab). It is conceivable that a further shifted band islocated, and thus not detected, in the same position as complex C1.
FIGURE 5. Analysis of NF binding using mutant probes.A, A gel mo-bility band shift assay was performed using probe P4 (Wt) and mutantprobes (M1–M7) and a nuclear extract from EL-4 cells.B, DNA sequencesof the probes used inA are shown. Mutated sequences are indicated byboldface. The consensus sequence suggested inA is indicated at thebottom.
FIGURE 6. IL-10 gene expression is regulated by CCTCCT motif.A, The antisense sequence from position276 to position296 in the mouse(M) IL-10 promoter was aligned with TCC repeats identified in promotersof human (H) CD58 (22) and EGFR (21) genes and a corresponding se-quence in the human IL-10 promoter (14). A common CCTCCT sequenceis found in all sequences and indicated by boldface.B, CCTCCT sequencesin luciferase reporter plasmids D1 and D6 (Fig. 2A) are mutated toGAATTC (EcoRI) (D1 CCT KO, D6 CCT KO).C, Luciferase activitiesgenerated using indicated plasmids were compared with that using thenegative control plasmid pGL3-Basic Vector (Basic) in EL-4 and PMA/ionomycin-stimulated EL-4 cells. Luciferase assays were repeated morethan three times, and the activities were normalized usingRenillaluciferaseactivity.
289The Journal of Immunology
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
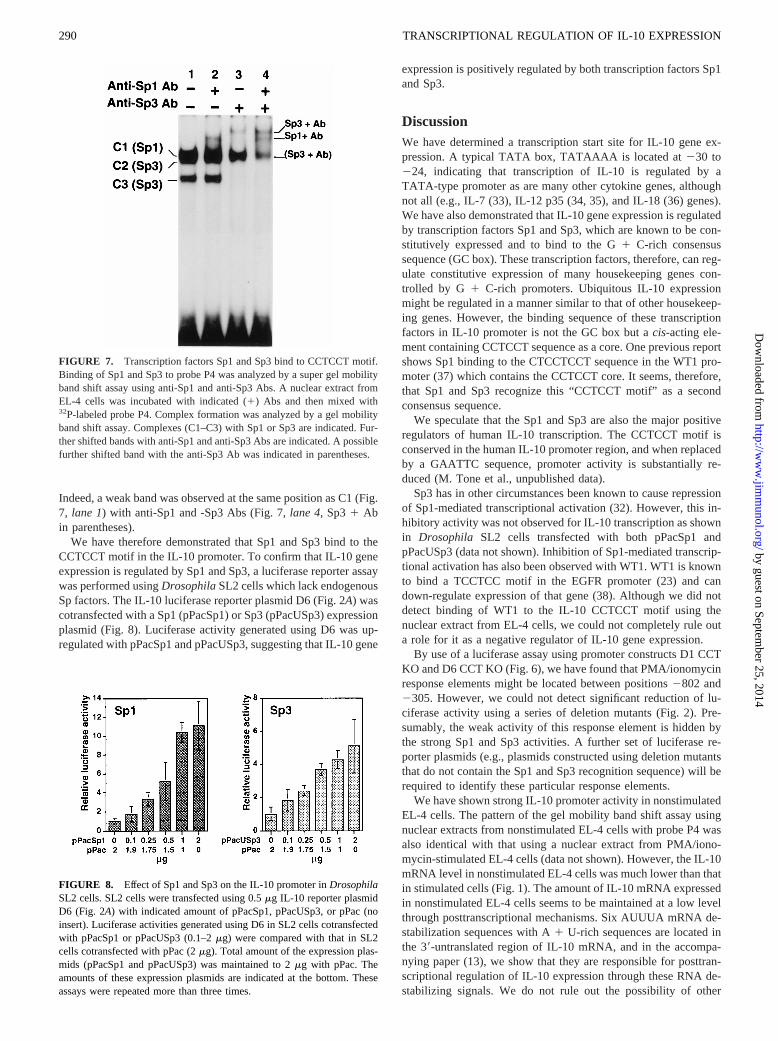
Indeed, a weak band was observed at the same position as C1 (Fig.7, lane 1) with anti-Sp1 and -Sp3 Abs (Fig. 7,lane 4, Sp31 Abin parentheses).
We have therefore demonstrated that Sp1 and Sp3 bind to theCCTCCT motif in the IL-10 promoter. To confirm that IL-10 geneexpression is regulated by Sp1 and Sp3, a luciferase reporter assaywas performed usingDrosophilaSL2 cells which lack endogenousSp factors. The IL-10 luciferase reporter plasmid D6 (Fig. 2A) wascotransfected with a Sp1 (pPacSp1) or Sp3 (pPacUSp3) expressionplasmid (Fig. 8). Luciferase activity generated using D6 was up-regulated with pPacSp1 and pPacUSp3, suggesting that IL-10 gene
expression is positively regulated by both transcription factors Sp1and Sp3.
DiscussionWe have determined a transcription start site for IL-10 gene ex-pression. A typical TATA box, TATAAAA is located at230 to224, indicating that transcription of IL-10 is regulated by aTATA-type promoter as are many other cytokine genes, althoughnot all (e.g., IL-7 (33), IL-12 p35 (34, 35), and IL-18 (36) genes).We have also demonstrated that IL-10 gene expression is regulatedby transcription factors Sp1 and Sp3, which are known to be con-stitutively expressed and to bind to the G1 C-rich consensussequence (GC box). These transcription factors, therefore, can reg-ulate constitutive expression of many housekeeping genes con-trolled by G 1 C-rich promoters. Ubiquitous IL-10 expressionmight be regulated in a manner similar to that of other housekeep-ing genes. However, the binding sequence of these transcriptionfactors in IL-10 promoter is not the GC box but acis-acting ele-ment containing CCTCCT sequence as a core. One previous reportshows Sp1 binding to the CTCCTCCT sequence in the WT1 pro-moter (37) which contains the CCTCCT core. It seems, therefore,that Sp1 and Sp3 recognize this “CCTCCT motif” as a secondconsensus sequence.
We speculate that the Sp1 and Sp3 are also the major positiveregulators of human IL-10 transcription. The CCTCCT motif isconserved in the human IL-10 promoter region, and when replacedby a GAATTC sequence, promoter activity is substantially re-duced (M. Tone et al., unpublished data).
Sp3 has in other circumstances been known to cause repressionof Sp1-mediated transcriptional activation (32). However, this in-hibitory activity was not observed for IL-10 transcription as shownin Drosophila SL2 cells transfected with both pPacSp1 andpPacUSp3 (data not shown). Inhibition of Sp1-mediated transcrip-tional activation has also been observed with WT1. WT1 is knownto bind a TCCTCC motif in the EGFR promoter (23) and candown-regulate expression of that gene (38). Although we did notdetect binding of WT1 to the IL-10 CCTCCT motif using thenuclear extract from EL-4 cells, we could not completely rule outa role for it as a negative regulator of IL-10 gene expression.
By use of a luciferase assay using promoter constructs D1 CCTKO and D6 CCT KO (Fig. 6), we have found that PMA/ionomycinresponse elements might be located between positions2802 and2305. However, we could not detect significant reduction of lu-ciferase activity using a series of deletion mutants (Fig. 2). Pre-sumably, the weak activity of this response element is hidden bythe strong Sp1 and Sp3 activities. A further set of luciferase re-porter plasmids (e.g., plasmids constructed using deletion mutantsthat do not contain the Sp1 and Sp3 recognition sequence) will berequired to identify these particular response elements.
We have shown strong IL-10 promoter activity in nonstimulatedEL-4 cells. The pattern of the gel mobility band shift assay usingnuclear extracts from nonstimulated EL-4 cells with probe P4 wasalso identical with that using a nuclear extract from PMA/iono-mycin-stimulated EL-4 cells (data not shown). However, the IL-10mRNA level in nonstimulated EL-4 cells was much lower than thatin stimulated cells (Fig. 1). The amount of IL-10 mRNA expressedin nonstimulated EL-4 cells seems to be maintained at a low levelthrough posttranscriptional mechanisms. Six AUUUA mRNA de-stabilization sequences with A1 U-rich sequences are located inthe 39-untranslated region of IL-10 mRNA, and in the accompa-nying paper (13), we show that they are responsible for posttran-scriptional regulation of IL-10 expression through these RNA de-stabilizing signals. We do not rule out the possibility of other
FIGURE 7. Transcription factors Sp1 and Sp3 bind to CCTCCT motif.Binding of Sp1 and Sp3 to probe P4 was analyzed by a super gel mobilityband shift assay using anti-Sp1 and anti-Sp3 Abs. A nuclear extract fromEL-4 cells was incubated with indicated (1) Abs and then mixed with32P-labeled probe P4. Complex formation was analyzed by a gel mobilityband shift assay. Complexes (C1–C3) with Sp1 or Sp3 are indicated. Fur-ther shifted bands with anti-Sp1 and anti-Sp3 Abs are indicated. A possiblefurther shifted band with the anti-Sp3 Ab was indicated in parentheses.
FIGURE 8. Effect of Sp1 and Sp3 on the IL-10 promoter inDrosophilaSL2 cells. SL2 cells were transfected using 0.5mg IL-10 reporter plasmidD6 (Fig. 2A) with indicated amount of pPacSp1, pPacUSp3, or pPac (noinsert). Luciferase activities generated using D6 in SL2 cells cotransfectedwith pPacSp1 or pPacUSp3 (0.1–2mg) were compared with that in SL2cells cotransfected with pPac (2mg). Total amount of the expression plas-mids (pPacSp1 and pPacUSp3) was maintained to 2mg with pPac. Theamounts of these expression plasmids are indicated at the bottom. Theseassays were repeated more than three times.
290 TRANSCRIPTIONAL REGULATION OF IL-10 EXPRESSION
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

transcriptional regulatory elements not included in the promoterfragment (e.g., enhancers).
IL-10 seems to be a major homeostatic regulator of inflamma-tion, immune responses, and involved in prevention of autoimmu-nity (39). Why then should one find that the transcriptional controlof the IL-10 gene is so dependent on these ubiquitously and con-stitutively expressed transcription factors, Sp1 and Sp3? It may bethat the immune system requires rapid availability of homeostaticregulators such as IL-10 and achieves this by ensuring that thegene is actively transcribing in a variety of cell types, but deter-mines protein availability through posttranscriptional mechanisms.
AcknowledgmentsWe thank Dr. Paul Fairchild for providing bmDC and Mark Frewin fortechnical assistance.
AddendumSince submission of our manuscript, Brightbill et al. (40) have publishedon transcriptional control of IL-10 gene expression. They too have estab-lished a role for Sp1 in the transcriptional control of the IL-10 gene, butonly in stimulated RAW 264 cells. Our conclusions differ insofar as wedetect promoter activity in both stimulated and nonstimulated RAW 264cells, perhaps as a result of the use of luciferase vectors in our study (41).In the accompanying paper (13), we explain the effects of stimulation onIL-10 expression, as a consequence of posttranscriptional control.
References1. Fiorentino, D. F., M. W. Bond, and T. R. Mosmann. 1989. Two types of mouse
T helper cell. IV. Th2 clones secrete a factor that inhibits cytokine production byTh1 clones.J. Exp. Med. 170:2081.
2. Moore, K. W., A. O’Garra, R. de Waal-Malefyt, P. Vieira, and T. R. Mosmann.1993. Interleukin-10.Annu. Rev. Immunol. 11:165.
3. Del Prete, G., M. De Carli, F. Almerigogna, M. G. Giudizi, R. Biagiotti, andS. Romagnani. 1993. Human IL-10 is produced by both type 1 helper (Th1)and type 2 (Th2) T cell clones and inhibits their antigen-specific proliferationand cytokine production.J. Immunol. 150:353.
4. O’Garra, A., R. Chang, N. Go, R. Hastings, G. Haughton, and M. Howard. 1992.Ly-1 B (B-1) cells are the main source of B cell-derived interleukin 10.Eur.J. Immunol. 22:711.
5. de Waal-Malefyt, R., J. Abrams, B. Bennett, C. G. Figdor, and J. E. de Vries.1991. Interleukin 10 (IL-10) inhibits cytokine synthesis by human monocytes: anautoregulatory role of IL-10 produced by monocytes.J. Exp. Med. 174:1209.
6. Enk, A. H., and S. I. Katz. 1992. Identification and induction of keratinocyte-derived IL-10.J. Immunol. 149:92.
7. Gastl, G. A., J. S. Abrams, D. M. Nanus, R. Oosterkamp, J. Silver, F. Liu,M. Chen, A. P. Albino, and N. H. Bander. 1993. Interleukin-10 production byhuman carcinoma cell lines and its relationship to interleukin-6 expression.Int. J.Cancer 55:96.
8. Groux, H., A. O’Garra, M. Bigler, M. Rouleau, S. Antonenko, J. E. de Vries, andM. G. Roncarolo. 1997. A CD41T-cell subset inhibits antigen-specific T-cellresponses and prevents colitis.Nature 389:737.
9. D’Andrea, A., M. Aste-Amezaga, N. M. Valiante, X. Ma, M. Kubin, andG. Trinchieri. 1993. Interleukin 10 (IL-10) inhibits human lymphocyte interferongamma-production by suppressing natural killer cell stimulatory factor/IL-12synthesis in accessory cells.J. Exp. Med. 178:1041.
10. Moore, K. W., P. Vieira, D. F. Fiorentino, M. L. Trounstine, T. A. Khan, andT. R. Mosmann. 1990. Homology of cytokine synthesis inhibitory factor (IL-10)to the Epstein-Barr virus geneBCRFI. Science 248:1230.
11. Vieira, P., R. de Waal-Malefyt, M. N. Dang, K. E. Johnson, R. Kastelein,D. F. Fiorentino, J. E. de Vries, M. G. Roncarolo, T. R. Mosmann, andK. W. Moore, 1991. Isolation and expression of human cytokine synthesis in-hibitory factor cDNA clones: homology to Epstein-Barr virus open reading frameBCRFI. Proc. Natl. Acad. Sci. USA 88:1172.
12. Chen, C.-Y. A., and A.-B. Shyu. 1995. AU-rich elements: characterization andimportance in mRNA degradation.Trends Biochem. Sci. 20:465.
13. Powell, M. J., S. A. J. Thompson, Y. Tone, H. Waldmann, and M. Tone. 2000.Posttranscriptional regulation of IL-10 gene expression through sequences in the39-untranslated region.J. Immunol. 165:292–296.
14. Kim, J. M., C. I. Brannan, N. G. Copeland, N. A. Jenkins, T. A. Khan, andK. W. Moore. 1992. Structure of the mouse IL-10 gene and chromosomal local-ization of the mouse and human genes.J. Immunol. 148:3618.
15. Kube, D., C. Platzer, A. von Knethen, H. Straub, H. Bohlen, M. Hafner, andH. Tesch. 1995. Isolation of the human interleukin 10 promoter: characterizationof the promoter activity in Burkitt’s lymphoma cell lines.Cytokine 7:1.
16. Frohman, M. A., M. K. Dush, and G. R. Martin. 1988. Rapid production offull-length cDNAs from rare transcripts: amplification using a single gene-spe-cific oligonucleotide primer.Proc. Natl. Acad. Sci. USA 85:8998.
17. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder.1983. Accurate transcriptioninitiation by RNA polymerase II in a soluble extract from isolated mammaliannuclei.Nucleic Acids Res. 11:1475.
18. Tone, M., L. E. Diamond, L. A. Walsh, Y. Tone, S. A. J. Thompson,E. M. Shanahan, J. S. Logan, and H. Waldmann. 1999. High level transcriptionof the complement regulatory protein CD59 requires an enhancer located in in-tron 1.J. Biol. Chem. 274:710.
19. Dynan, W. S., and R. Tjian. 1983. The promoter-specific transcription factor Sp1binds to upstream sequences in the SV40 early promoter.Cell 35:79.
20. Ghosh, D. 1993. Status of the transcription factors database (TFD).Nucleic AcidsRes. 21:3117.
21. Johnson, A. C., Y. Jinno, and G. T. Merlino. 1988. Modulation of epidermalgrowth factor receptor proto-oncogene transcription by a promoter site sensitiveto S1 nuclease.Mol. Cell. Biol. 8:4174.
22. Ewulonu, U. K., L. Ravi, and M. E. Medof. 1991. Characterization of the decay-accelerating factor gene promoter region.Proc. Natl. Acad. Sci. USA 88:4675.
23. Wang, Z.-Y., Q.-Q. Qiu, K. T. Enger, and T. F. Deuel. 1993. A second tran-scriptionally active DNA-binding site for the Wilms tumor gene product, WT1.Proc. Natl. Acad. Sci. USA 90:8896.
24. McCann, S., J. Sullivan, J. Guerra, M. Arcinas, and L. M. Boxer. 1995. Repres-sion of the c-mybgene by WT1 protein in T and B cell lines.J. Biol. Chem.270:23785.
25. Kadonaga, J. T., K. R. Carner, F. R. Masiarz, and R. Tjian. 1987. Isolation ofcDNA encoding transcription factor Sp1 and functional analysis of the DNAbinding domain.Cell 51:1079.
26. Kingsley, C., and A. Winoto. 1992. Cloning of GT box-binding proteins: a novelSp1 multigene family regulating T-cell receptor gene expression.Mol. Cell. Biol.12:4251.
27. Hagen, G., S. Muller, M. Beato, and G. Suske. 1992. Cloning by recognition sitescreening of two novel GT box binding proteins: a family of Sp1 related genes.Nucleic Acids Res. 20:5519.
28. Sukhatme, V. P., X. Cao, L. C. Chang, C.-H. Tsai-Morris, D. Stamenkovich,P. C. P. Ferreira, D. R. Cohen, S. A. Edwards, T. B. Shows, T. Curran, et al.1988.A zinc finger-encoding gene coregulated with c-fosduring growth and differen-tiation, and after cellular depolarization.Cell 53:37.
29. Milbrandt, J. 1987. A nerve growth factor-induced gene encodes a possible tran-scriptional regulatory factor.Science 238:797.
30. Joseph, L. J., M. M. Le Beau, G. A. Jamieson, S. Acharya, T. B. Shows,J. D. Rowley, and V. P. Sukhatme. 1988. Molecular cloning, sequencing, andmapping of EGR2, a human early growth response gene encoding a protein with“zinc-binding finger” structure.Proc. Natl. Acad. Sci. USA 85:7164.
31. Patwardhan, S., A. Gashler, M. G. Siegel, L. C. Chang, L. J. Joseph, T. B. Shows,M. M. Le Beau, and V. P. Sukhatme. 1991. EGR3, a novel member of the Egrfamily of genes encoding immediate-early transcription factors.Oncogene 6:917.
32. Hagen, G., S. Muller, M. Beato, and G. Suske. 1994. Sp1-mediated transcrip-tional activation is repressed by Sp3.EMBO J. 13:3843.
33. Lupton, S. D., S. Gimpel, R. Jerzy, L. L. Brunton, K. A. Hjerrild, D. Cosman, andR. G. Goodwin. 1990. Characterization of the human and murine IL-7 genes.J. Immunol. 144:3592.
34. Tone, Y., S. A. J. Thompson, J. M. Babik, K. F. Nolan, M. Tone, C. Raven, andH. Waldmann.1996. Structure and chromosomal location of the mouse interleu-kin-12 p35 and p40 subunit genes.Eur. J. Immunol. 26:1222.
35. Babik, J.M., E. Adams, Y. Tone, P.J. Fairchild, M. Tone, and H. Waldmann.1999. Expression of murine IL-12 is regulated by translational control of the p35subunit.J. Immunol. 162:4069.
36. Tone, M., S. A. J. Thompson, Y. Tone, P. J. Fairchild, and H. Waldmann. 1997.Regulation of IL-18 (IFN-g-inducing factor) gene expression.J. Immunol. 159:6156.
37. Cohen, H. T., S. A. Bossone, G. Zhu, G. A. McDonald, and V. P. Sukhatme.1997. Sp1 is a critical regulator of the Wilms’ tumor-1 gene.J. Biol. Chem.272:2901.
38. Englert, C., X. Hou, S. Maheswaran, P. Bennett, C. Ngwu, G. G. Re, A. J. Garvin,M. R. Rosner, and D. A. Haber. 1995. WT1 suppresses synthesis of the epidermalgrowth factor receptor and induces apoptosis.EMBO J. 14:4662.
39. Segal, B. M., B. K. Dwyer, and E. M. Shevach. 1998. An interleukin (IL)-10/IL-12 immunoregulatory circuit controls susceptibility to autoimmune disease.J. Exp. Med. 187:537.
40. Brightbill, H. D., S. E. Plevy, R. L. Modlin, and S. T. Smale. 2000. A prominentrole for Sp1 during lipopolysaccharide-mediated induction of the IL-10 promoterin macrophages.J. Immunol. 164:1949.
41. Plevy, S. E., J. H. Gemberling, S. Hsu, A. J. Domer, and S. T. Smale. 1997.Multiple control elements mediate activation of the murine and human interleukin12 p40 promoters: evidence of functional synergy between C/EBP and Rel pro-teins.Mol. Cell. Biol. 17:4572.
291The Journal of Immunology
by guest on September 25, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from





























