Embed Size (px)
Citation preview

Mutation Research 553 (2004) 111–124
Immunomodulatory effects of mineral fibres inoccupationally exposed workers
Jana Tulinskaa,∗, Eva Jahnovaa, Maria Dusinskaa, Miroslava Kuricovaa,Aurelia Liskovaa, Silvia Ilavskaa, Mira Horvathovaa, Ladislava Wsolovaa,
Soterios A. Kyrtopoulosb, Andrew Collinsc, Vikki Harringtond, Laurence Fuortesea Research Base of the Slovak Medical University, Institute of Preventive and Clinical Medicine,
Limbova 12, 833 03 Bratislava, Slovak Republicb National Hellenic Research Foundation, Institute of Biological Research and Biotechnology,
48 Vassileos Constantinou Avenue, Athens 11635, Greecec Department of Nutrition, University of Oslo, P.O. Box 1046 Blindern, 0316 Oslo, Norway
d The Robert Gordon University, Schoolhill, Aberdeen, AB10 1FR Scotland, UKe College of Public Health, University of Iowa, 2941 Steindler Bldg., Iowa City, IO, USA
Received 16 April 2004; received in revised form 4 May 2004; accepted 7 May 2004
Abstract
In the context of a large-scale molecular epidemiology study, the possible immunomodulatory effects of mineral fibres,in workers occupationally exposed to asbestos, rockwool and glass fibres, were examined. In each plant, 61, 98 and 80exposed workers and 21, 43 or 36 control clerical subjects, respectively, were recruited. In the case of the asbestos-exposedsubjects, an additional town-control group of 49 people was included. Evidence of pulmonary fibrosis was found in 42%of the asbestos-exposed workers, while evidence of pleural fibrosis was found in 24%. The asbestos-exposed cohort hadsignificantly decreased forced vital capacity of lungs as well as forced expiratory volume per first second.
Our findings indicate that exposure to all three types of fibres examined modulates to different degrees the immune response.Suppression of T-cell immunity and to a lesser extent, B-cell immunity was found in the case of workers from a former asbestoscement plant, while stimulation of T-cell response was observed in rockwool workers, and stimulation of T- and B-cell responsewas seen in glass fibre workers. Depression of the percentage of lymphocyte subpopulation of CD 16+56 (natural killer cells)in peripheral blood was found in glass fibre workers. Statistical analysis showed increased levels of proinflammatory cytokines(IL-6 asbestos; IL-8 all three fibres), expression of adhesion molecule L-selectin on granulocytes and monocytes (asbestos),levels of soluble adhesion molecules (SAMs) in sera (ICAM-1 all three fibres; E-selectin glass fibres), increased levels ofimmunoglobulin E (asbestos and rockwool) and elevated expression of activation markers on eosinophils (CD66b asbestos,glass fibres; CD69 asbestos). Significant correlations were observed between lymphocyte proliferation and markers of DNA
Abbreviations:MMMF, man-made mineral fibres; Con A, concanavalin A; ICAM, intercellular adhesion molecule; PHA, phytohemmag-glutinin; PWM, pokeweed mitogen; SAM, soluble adhesion molecule; TET, tetanus toxoid-antigen
∗ Corresponding author. Tel.:+421 2 59369 227.E-mail address:[email protected] (J. Tulinska).
0027-5107/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.mrfmmm.2004.06.030

112 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
damage and repair. Increased levels of proinflammatory cytokines, SAMs, immunoglobulin E and elevated expression ofactivation markers on eosinophils was found in people with symptoms of hypersensitivity and an elevated inflammatorystatus.© 2004 Elsevier B.V. All rights reserved.
Keywords:Immune system; Mineral fibres; Asbestos; Rockwool; Glass fibres; Occupational exposure; DNA damage; DNA repair
1. Introduction
Asbestos has long been recognized as a cause ofboth benign and malignant lung disease (interstitialand pleural fibrosis, lung cancer and mesothelioma).The evidence for adverse health effects following ex-posure to asbestos has prompted a drastic reduction inthe use of asbestos, resulting in the increased use ofsubstitutes composed of both naturally occurring andsynthetic materials which are thought to have lowertoxicity. However, some reports have implicated alsosome of these potential asbestos substitutes, as well asother natural and man-made mineral fibres (MMMF)as causes of lung disease[1,2]. For these reasons, im-proved understanding, assessment and monitoring ofthe toxic effects of MMMFs is important. Glass fibresand rockwool belong to the family of MMMFs used asinsulation materials and, owning to their widespreaduse, their possible toxicity is of significant interest forpublic health[3,4].
Immunotoxicity may play an important role in theinduction of fibre-related lung disease, including can-cer. Correlations between genotoxic parameters andparameters of immune response were found in workersoccupationally exposed to PAHs in rubber factory[5]and a population exposed to styrene[6]. Informationon the immunotoxicity and immunomodulatory abil-ity of MMMFs is still limited, and the investigation ofsuch possible effects may provide new knowledge onthe toxicity of mineral fibres.
In study reported here, biomarkers of immunotox-icity were examined in workers exposed to asbestosas well as MMMFs. The markers examined includedlymphocyte subset analysis by flow cytometry, lym-phocyte function, phagocytic activity of leukocytes,natural killer cell activity, expression of adhesionmolecules on peripheral blood leukocytes, activa-tion markers on eosinophils, serum levels of solubleadhesion molecules and serum concentrations of im-munoglobulins. In the present paper, a short overviewof the most important findings is presented.
2. Materials and methods
2.1. Subjects
Markers of the immune response were assessed inworkers with at least 5 years exposure to asbestos,rockwool or glass fibres, at three industrial plants in theSlovak Republic, using a cross-sectional study design.Asbestos-exposed workers had worked in an asbestoscement factory, which ceased operation 9 months priorto the conduct of the study. On the other hand, pro-duction of rockwool and glass fibres was ongoing inthe corresponding plants at the time of the study. Theselection criteria for control participants were similarage, sex and duration of employment to exposed sub-jects but having worked in office areas, rather thanthe production plants, of the same factories. Each per-son was interviewed about possible confounding fac-tors such as smoking habits, alcohol consumption andmedication. Blood was collected by venepuncture intotubes.
2.2. Exposure
Public Health Institute kindly provided data onhistorical exposure to asbestos in workplace. Currentexposure to the asbestos, rockwool and glass mineralfibres was assessed using stationary area samplingas well as personal monitoring. Quantification of fi-bre exposure and identification of fibres with focuson fibre morphology was done by electron scanningmicroscopy. Air-borne particles were collected in theworking area of the asbestos cement, rockwool andglass fibre plants, sampling took place in all fourseasons. Samples were used for asbestos fibre andMMMF identification, fibre morphology and quan-tification, using a microscope with phase contrast(Nikon, Japan) according to the Reference Methodfor the Determination of Airborne Asbestos FibreConcentration at Workplaces by Light Microscopy

J. Tulinska et al. / Mutation Research 553 (2004) 111–124 113
(Membrane Filter Method), AIA 1979, London,UK.
2.3. Immune assays
2.3.1. Lymphoproliferative response to mitogens andantigens
The proliferation of peripheral blood lymphocytesin response to mitogens [25�g/ml concanavalin A(Con A, Sigma), 25�g/ml phytohemagglutinin (PHA,Sigma), 2,5�g/ml pokeweed mitogen (PWM, Sigma)]and antigens [tetanus toxoid (TET, Sevac, Praha) andmonoclonal anti-human CD3 antibody (Sigma)] wasevaluated using a microculture assay[7] as describedpreviously[8]. Venous heparinized blood (drawn asep-tically and kept at room temperature) was diluted 1:15(v/v) with complete RPMI 1640 medium. Results wereexpressed as disintegrations per minute (dpm)/numberof lymphocytes. The stimulation index was calculatedby dividing the mean dpm/number of lymphocytes inresponse to stimulation by the mean dpm/number oflymphocytes of the cells cultured alone.
2.3.2. Phagocytic activity of monocytes in peripheralblood
The phagocytic activity of monocytes was assayedby the method of Fornusek[9] as previously described[8]. Blood smears were made and stained by panop-tic staining of Pappenheim. Two hundred cells wereexamined for each sample. Cells with three or moreparticles were considered as phagocytic leukocytes. Adifferential leukocyte count was carried out on bloodsmears stained with Giemsa-Romanowski; again 200leukocytes were examined. Leukocytes were differ-entiated into polymorphonuclear leukocytes, lympho-cytes, large granular lymphocytes, monocytes andeosinophils.
2.3.3. Natural killer cell assayThe activity of natural killer cells (NK-cells) was
assayed by the method of Bromelow et al.[10]. Bloodwas first dispensed in triplicate in a range of fivedoubling dilutions, starting at 100�l, to which corre-sponding volumes of complete medium were addedup to 200�l, and then 50�l of 1 × 104 cells K 562labelled with51Cr were added to the wells. Sponta-neous (S) and total (T)51Cr release wells were alsoset up in a total volume of 250�l/well. The plates
were incubated at 37◦C in 5% CO2 for 18 h. Afterco-culture, 100�l aliquots of the culture supernatantswere transferred to tubes and measured on a gammacounter. Reading and calculations for percent of lysiswere performed.
2.3.4. Flow cytometryCells from whole peripheral blood were stained
and analysed by flow cytometry using an EPICS XLCoulter (Coulter, USA) running under System II soft-ware. For direct staining, the following conjugatedantibodies were employed: CD3-Pe, CD4-Pe, HLADR-Fitc, CD19-Fitc, CD16+CD56-Fitc, CD11b-Pe,CD62L-Fitc, CD18-Fitc, CD49d-Pe, CD25-Fitc,CD69-Fitc (all from Becton Dickinson, USA), andICAM-1-Fitc, CD66b-Fitc, CD81-Pe (all from Im-munotech, A Coulter company, France). The ap-propriate antibody negative controls (anti-IgG1 oranti-IgG2a) were used for every tested sample.
The antibodies and whole blood samples wereadded to tubes and incubated for 15 min at roomtemperature. For the lysis of red blood cells OptilyseC lysing solution (Immunotech, France) was usedand after 10 min incubation at room temperature thelysing process was stopped with PBS. Finally, thesamples were evaluated by flow cytometry.
2.3.5. ELISAFor the quantitative determination of the concen-
trations of human interleukins (IL-1, IL-6 and IL-8)and human soluble adhesion molecules (ICAM-1,E-selectin) in serum, solid phase ELISA test (Quan-tikine and Parameter; R&D Systems Inc., USA,respectively) were used. Levels of total immunoglob-ulin E were measured by ELISA (Total IgE; BeckmanCoulter, made by Immunotech, France).
2.3.6. Measurement of DNA damage and repair of8-oxoguanine
DNA damage and repair were measured using themodified comet assay (single cell alkaline gel elec-trophoresis) as described in detail elsewhere in thisvolume (Dusinska et al., in press).
2.4. Statistical analysis
Statistical analysis was done using SPSS software(SPSS Co., USA). Normality was verified using the
114 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
Kolmogorov–Smirnov test. Data were compared us-ing one-way and multifactorial analysis of variance(Manova) for significant differences between groups.Non-parametric tests such as Mann–WhitneyU-testand Wilcox test were used for data that were not dis-tributed normally. Experimental data from control andexposed workers were considered statistically differ-ent atP < 0.05.
3. Results
3.1. Subjects
Table 1summarizes the characteristics of the stud-ied populations from asbestos, rockwool and glass fi-bre factories and controls. In each plant, 61, 98 and80 exposed workers and 21, 43 or 36 control clericalsubjects, respectively, were recruited. In the case ofasbestos-exposed subjects, an additional town-controlgroup of 49 people was included.
Table 1Participants characteristics and duration of asbestos, rockwool and glass fibres exposure
Exposure Group Number ofdonors
Sex(male/female)
Smoking status Age (years),median(range)
Duration ofexposure (years),median (range)
Asbestos Town control 42 13/29 Smoker 13 53 (39–88) 0Non-smoker 29All 42
Factory control 18 11/7 Smoker 6 58 (28–72) 0Non-smoker 12All 18
Exposed 60 44/16 Smoker 24 59.5 (39–80) 25 (5–40)Non-smoker 36All 60
Rockwool Factory control 36 15/21 Smoker 13 43 (29–58) 0Non-smoker 23All 36
Exposed 87 65/22 Smoker 31 43 (24–57) 16 (5–44)Non-smoker 56All 87
Glass fibres Factory control 30 14/16 Smoker 10 50 (31–59) 0Non-smoker 20All 30
Exposed 75 36/39 Smoker 32 46 (25–58) 16 (5–34)Non-smoker 43All 75
Clinical investigation and spirometry was per-formed in all exposed and control cohorts, whilean X-ray examination was given only to personsexposed to asbestos, rockwool and glass fibres. Evi-dence of pulmonary fibrosis was found in 42% of theasbestos-exposed workers, while evidence of pleuralfibrosis was found in 24%. Nine asbestos-exposedworkers (15%) had both interstitial and pleural fi-brosis. The asbestos-exposed cohort had significantlydecreased forced vital capacity of lungs as well asforced expiratory volume per first second. No changeswere found by X-ray and spirometry examinationamong workers in the rockwool or glass fibre plants.
3.2. Exposure to mineral fibres
Data from the historical records of asbestosworkplace exposure indicate that during the years1956–1977 only total dust concentrations were mea-sured. Between the years 1956–1960, they were inthe range 40–60 mg/m3, while during 1960–1977
J. Tulinska et al. / Mutation Research 553 (2004) 111–124 115
they were in the range 10–34 mg/m3. In the years1978–1998, the concentration of asbestos fibres wasmeasured (1978–1990: 2–11 fibres/cm3; 1991–1994:1–4 fibres/cm3; 1994–1998: 0.2–1 fibres/cm3). In1996, a gradual replacement of asbestos by other fi-brous materials began, which was completed by 2000.At the time of the presently reported study, the as-bestos fibre concentration measured in the productionhall of the former asbestos cement plant exceededby three to five times the Slovak occupational limit(0.001 fibre/cm3 for indoor areas where asbestos wasused as building material) (data not shown). The con-centrations in the administrative area were below thislimit.
Only the presence of basalt glass (rockwool) fibreswas detected in the rockwool producing factory and,similarly, only the presence of glass fibres was de-tected in the glass fibre production plant. All mea-sured levels of rockwool and glass fibres were verylow (10–1000 times below the Slovak occupationallimits) (data not shown).
3.3. Immune parameters
Using immune assays, markers of lymphocytefunction were found to differ significantly betweenfibre-exposed cohorts and corresponding controls.The most pronounced changes in lymphocyte prolif-
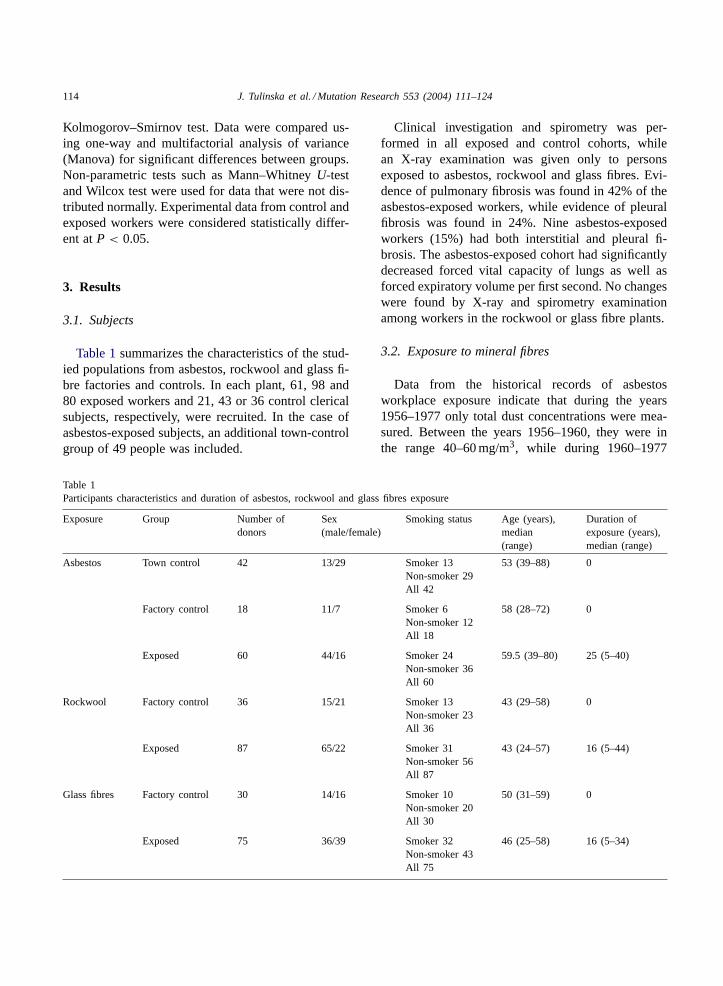
Fig. 1. Proliferative response of T- and B-lymphocytes in workers occupationally exposed (EX) to asbestos, rockwool or glass fibres incomparison with factory control (FC) or town control (TC). Proliferation of T-lymphocytes was induced by mitogen phytohaemmagglutinin(PHA) and proliferation of B-lymphocytes by pokeweed (PWM) mitogen. Samples were measured as incorporation of [3H]-thymidine intoproliferating cells. Statistically significance: *P < 0.05; **P < 0.01.
erative activity were found in cultures stimulated withmitogens phytohemagglutinin (PHA) and pokeweedmitogen (PWM). Workers from the former asbestoscement plant had significantly decreased proliferativecapacity of lymphocytes stimulated by T-cell mito-gen PHA. In contrast, the proliferative activity ofT-lymphocytes in people from the rockwool and glassfibre factories was stimulated (Fig. 1).
Concerning effects on B-lymphocytes, a significantstimulatory effect was observed in cultures of periph-eral blood derived from the glass fibre workers in vitrostimulated with PWM mitogen, while no such effectwas found in workers from the asbestos and rockwoolplants in comparison with the corresponding controls(Fig. 1).
The phagocytic activity of polymorphonuclearleukocytes and monocytes individually, as well asof leukocytes as a whole, did not differ significantlybetween exposed and control groups. Similarly, theresults of the natural killer cell assay indicated no sig-nificant differences in NK-activity between exposedand controls in the cohort exposed to asbestos androckwool (assay in glass fibre workers not done).
Phenotypic analysis of peripheral blood leuko-cytes was performed to assess the proportions of themain lymphocytes subsets. Flow cytometry analysisshowed significantly decreased expression of markersCD16+56 (natural killer cells) in exposed workers
116 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
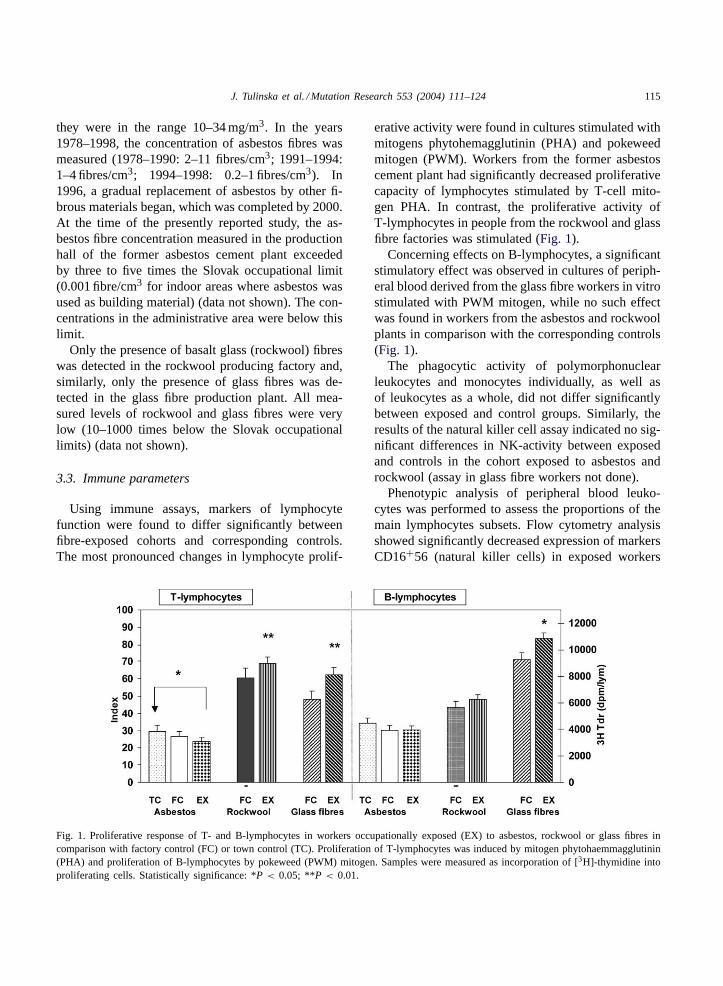
Fig. 2. Expression of marker CD16+56 (natural killer cells) inworkers occupationally exposed (EX) to asbestos, rockwool orglass fibres in comparison with factory control (FC) or town control(TC). Statistically significance: *P < 0.05.
from the glass fibre plant in comparison with thecorresponding controls (Fig. 2).
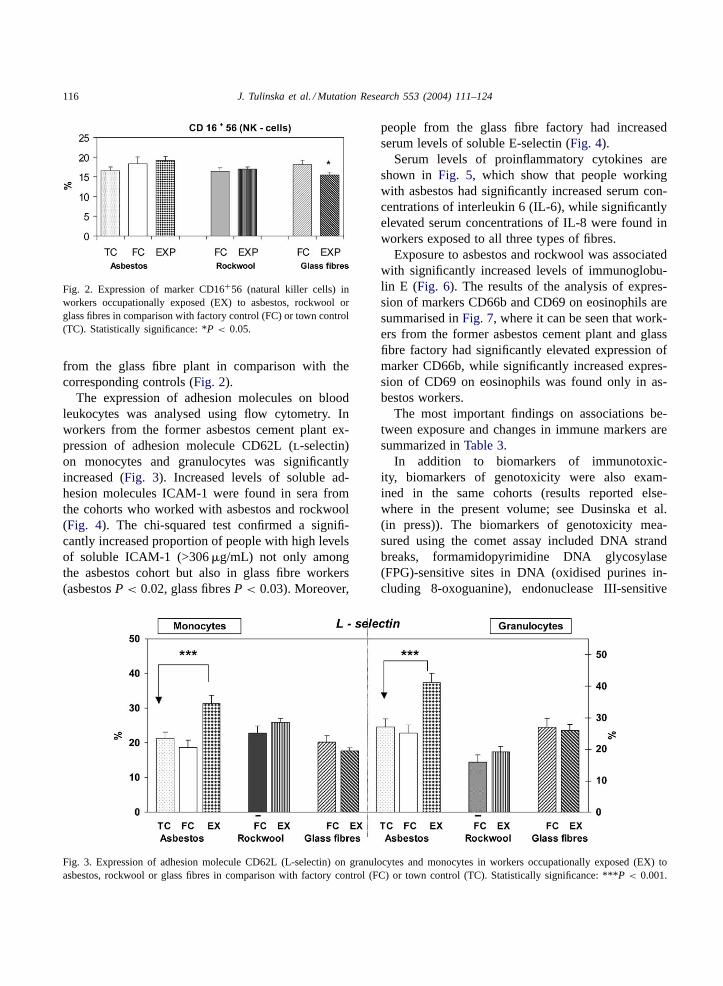
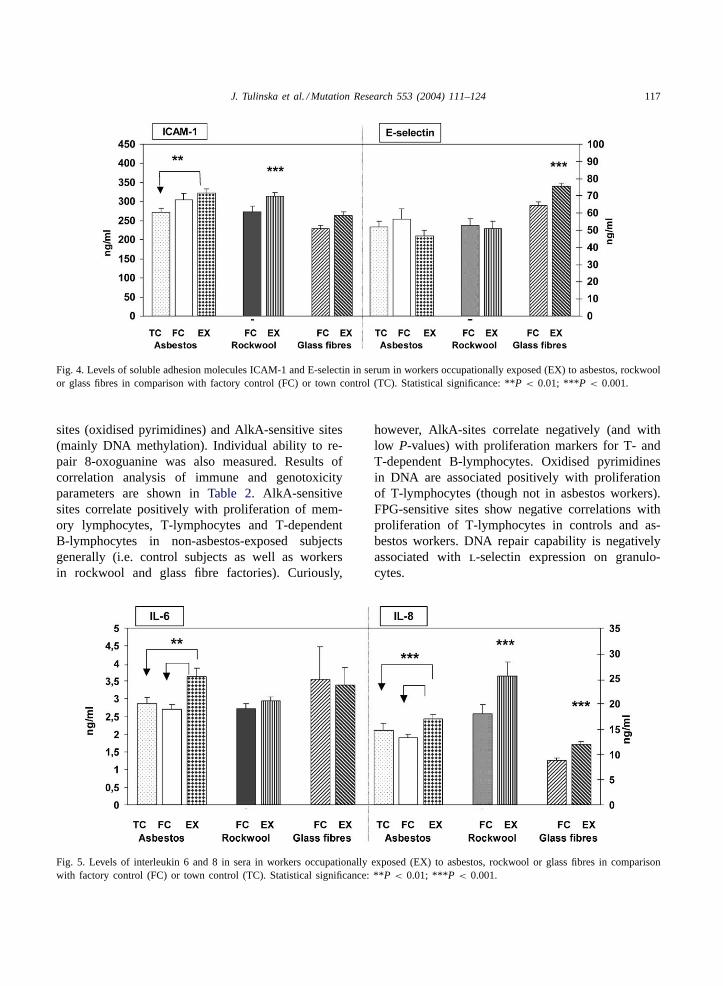
The expression of adhesion molecules on bloodleukocytes was analysed using flow cytometry. Inworkers from the former asbestos cement plant ex-pression of adhesion molecule CD62L (l-selectin)on monocytes and granulocytes was significantlyincreased (Fig. 3). Increased levels of soluble ad-hesion molecules ICAM-1 were found in sera fromthe cohorts who worked with asbestos and rockwool(Fig. 4). The chi-squared test confirmed a signifi-cantly increased proportion of people with high levelsof soluble ICAM-1 (>306�g/mL) not only amongthe asbestos cohort but also in glass fibre workers(asbestosP < 0.02, glass fibresP < 0.03). Moreover,
Fig. 3. Expression of adhesion molecule CD62L (L-selectin) on granulocytes and monocytes in workers occupationally exposed (EX) toasbestos, rockwool or glass fibres in comparison with factory control (FC) or town control (TC). Statistically significance: ***P < 0.001.
people from the glass fibre factory had increasedserum levels of soluble E-selectin (Fig. 4).
Serum levels of proinflammatory cytokines areshown in Fig. 5, which show that people workingwith asbestos had significantly increased serum con-centrations of interleukin 6 (IL-6), while significantlyelevated serum concentrations of IL-8 were found inworkers exposed to all three types of fibres.
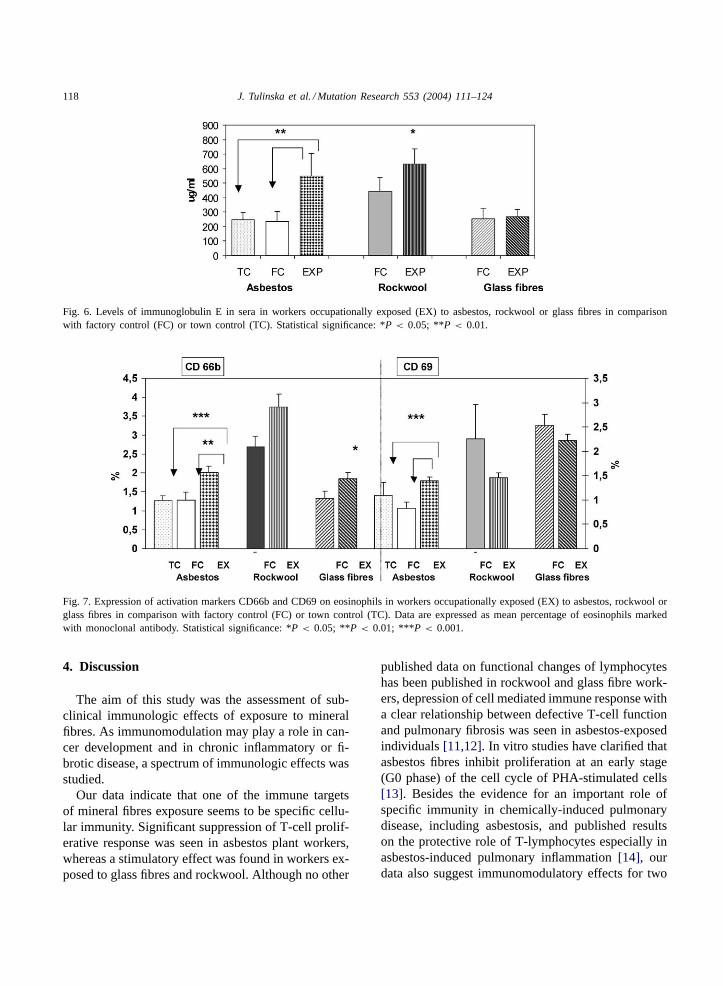
Exposure to asbestos and rockwool was associatedwith significantly increased levels of immunoglobu-lin E (Fig. 6). The results of the analysis of expres-sion of markers CD66b and CD69 on eosinophils aresummarised inFig. 7, where it can be seen that work-ers from the former asbestos cement plant and glassfibre factory had significantly elevated expression ofmarker CD66b, while significantly increased expres-sion of CD69 on eosinophils was found only in as-bestos workers.
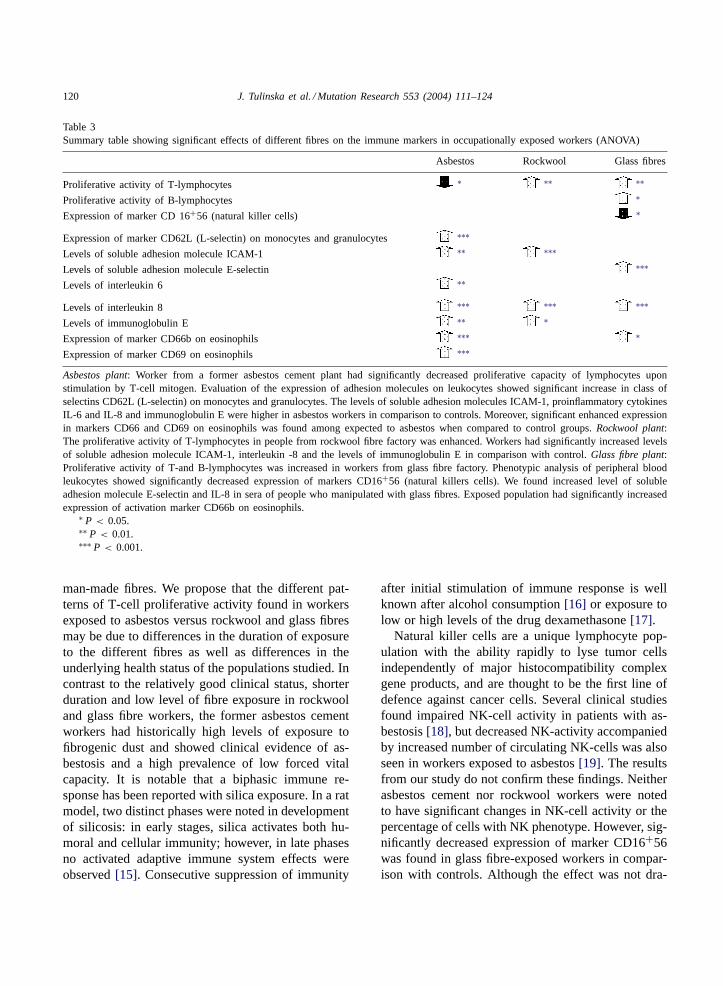
The most important findings on associations be-tween exposure and changes in immune markers aresummarized inTable 3.
In addition to biomarkers of immunotoxic-ity, biomarkers of genotoxicity were also exam-ined in the same cohorts (results reported else-where in the present volume; see Dusinska et al.(in press)). The biomarkers of genotoxicity mea-sured using the comet assay included DNA strandbreaks, formamidopyrimidine DNA glycosylase(FPG)-sensitive sites in DNA (oxidised purines in-cluding 8-oxoguanine), endonuclease III-sensitive
J. Tulinska et al. / Mutation Research 553 (2004) 111–124 117
Fig. 4. Levels of soluble adhesion molecules ICAM-1 and E-selectin in serum in workers occupationally exposed (EX) to asbestos, rockwoolor glass fibres in comparison with factory control (FC) or town control (TC). Statistical significance: **P < 0.01; ***P < 0.001.
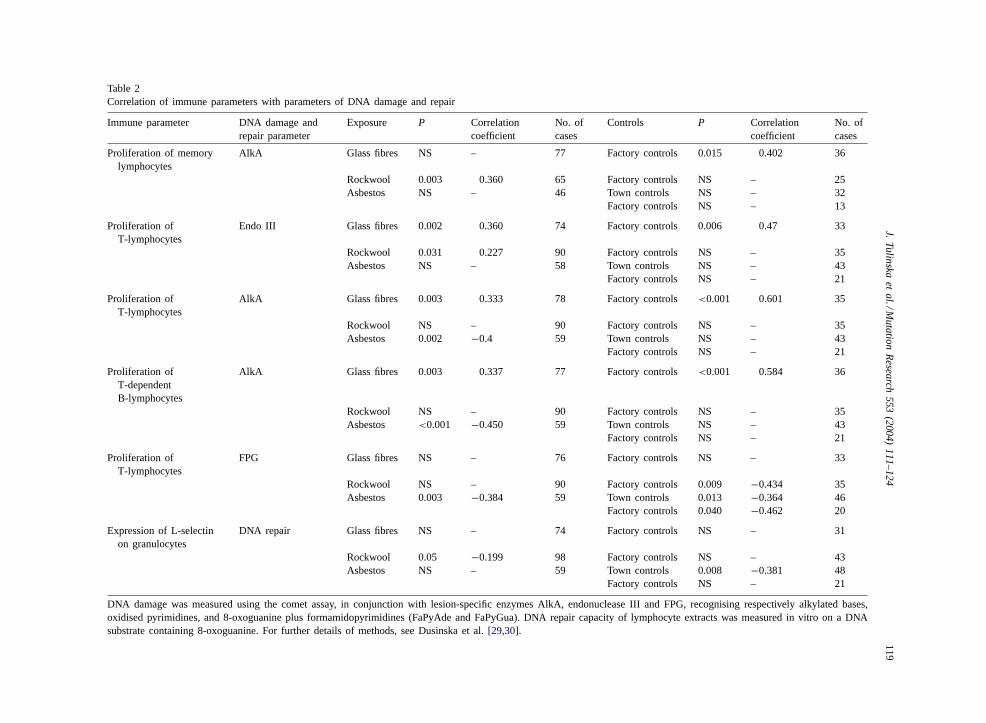
sites (oxidised pyrimidines) and AlkA-sensitive sites(mainly DNA methylation). Individual ability to re-pair 8-oxoguanine was also measured. Results ofcorrelation analysis of immune and genotoxicityparameters are shown inTable 2. AlkA-sensitivesites correlate positively with proliferation of mem-ory lymphocytes, T-lymphocytes and T-dependentB-lymphocytes in non-asbestos-exposed subjectsgenerally (i.e. control subjects as well as workersin rockwool and glass fibre factories). Curiously,
Fig. 5. Levels of interleukin 6 and 8 in sera in workers occupationally exposed (EX) to asbestos, rockwool or glass fibres in comparisonwith factory control (FC) or town control (TC). Statistical significance: **P < 0.01; ***P < 0.001.
however, AlkA-sites correlate negatively (and withlow P-values) with proliferation markers for T- andT-dependent B-lymphocytes. Oxidised pyrimidinesin DNA are associated positively with proliferationof T-lymphocytes (though not in asbestos workers).FPG-sensitive sites show negative correlations withproliferation of T-lymphocytes in controls and as-bestos workers. DNA repair capability is negativelyassociated withl-selectin expression on granulo-cytes.
118 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
Fig. 6. Levels of immunoglobulin E in sera in workers occupationally exposed (EX) to asbestos, rockwool or glass fibres in comparisonwith factory control (FC) or town control (TC). Statistical significance: *P < 0.05; **P < 0.01.
Fig. 7. Expression of activation markers CD66b and CD69 on eosinophils in workers occupationally exposed (EX) to asbestos, rockwool orglass fibres in comparison with factory control (FC) or town control (TC). Data are expressed as mean percentage of eosinophils markedwith monoclonal antibody. Statistical significance: *P < 0.05; **P < 0.01; ***P < 0.001.
4. Discussion
The aim of this study was the assessment of sub-clinical immunologic effects of exposure to mineralfibres. As immunomodulation may play a role in can-cer development and in chronic inflammatory or fi-brotic disease, a spectrum of immunologic effects wasstudied.
Our data indicate that one of the immune targetsof mineral fibres exposure seems to be specific cellu-lar immunity. Significant suppression of T-cell prolif-erative response was seen in asbestos plant workers,whereas a stimulatory effect was found in workers ex-posed to glass fibres and rockwool. Although no other
published data on functional changes of lymphocyteshas been published in rockwool and glass fibre work-ers, depression of cell mediated immune response witha clear relationship between defective T-cell functionand pulmonary fibrosis was seen in asbestos-exposedindividuals[11,12]. In vitro studies have clarified thatasbestos fibres inhibit proliferation at an early stage(G0 phase) of the cell cycle of PHA-stimulated cells[13]. Besides the evidence for an important role ofspecific immunity in chemically-induced pulmonarydisease, including asbestosis, and published resultson the protective role of T-lymphocytes especially inasbestos-induced pulmonary inflammation[14], ourdata also suggest immunomodulatory effects for two
J.T
ulin
skae
ta
l./Mu
tatio
nR
ese
arch
55
3(2
00
4)
11
1–
12
4119
Table 2Correlation of immune parameters with parameters of DNA damage and repair
Immune parameter DNA damage andrepair parameter
Exposure P Correlationcoefficient
No. ofcases
Controls P Correlationcoefficient
No. ofcases
Proliferation of memorylymphocytes
AlkA Glass fibres NS – 77 Factory controls 0.015 0.402 36
Rockwool 0.003 0.360 65 Factory controls NS – 25Asbestos NS – 46 Town controls NS – 32
Factory controls NS – 13
Proliferation ofT-lymphocytes
Endo III Glass fibres 0.002 0.360 74 Factory controls 0.006 0.47 33
Rockwool 0.031 0.227 90 Factory controls NS – 35Asbestos NS – 58 Town controls NS – 43
Factory controls NS – 21
Proliferation ofT-lymphocytes
AlkA Glass fibres 0.003 0.333 78 Factory controls <0.001 0.601 35
Rockwool NS – 90 Factory controls NS – 35Asbestos 0.002 −0.4 59 Town controls NS – 43
Factory controls NS – 21
Proliferation ofT-dependentB-lymphocytes
AlkA Glass fibres 0.003 0.337 77 Factory controls <0.001 0.584 36
Rockwool NS – 90 Factory controls NS – 35Asbestos <0.001 −0.450 59 Town controls NS – 43
Factory controls NS – 21
Proliferation ofT-lymphocytes
FPG Glass fibres NS – 76 Factory controls NS – 33
Rockwool NS – 90 Factory controls 0.009 −0.434 35Asbestos 0.003 −0.384 59 Town controls 0.013 −0.364 46
Factory controls 0.040 −0.462 20
Expression of L-selectinon granulocytes
DNA repair Glass fibres NS – 74 Factory controls NS – 31
Rockwool 0.05 −0.199 98 Factory controls NS – 43Asbestos NS – 59 Town controls 0.008 −0.381 48
Factory controls NS – 21
DNA damage was measured using the comet assay, in conjunction with lesion-specific enzymes AlkA, endonuclease III and FPG, recognising respectivelyalkylated bases,oxidised pyrimidines, and 8-oxoguanine plus formamidopyrimidines (FaPyAde and FaPyGua). DNA repair capacity of lymphocyte extracts was measuredin vitro on a DNAsubstrate containing 8-oxoguanine. For further details of methods, see Dusinska et al.[29,30].
120 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
Table 3Summary table showing significant effects of different fibres on the immune markers in occupationally exposed workers (ANOVA)
Asbestos Rockwool Glass fibres
Proliferative activity of T-lymphocytes ∗ ∗∗ ∗∗
Proliferative activity of B-lymphocytes ∗
Expression of marker CD 16+56 (natural killer cells) ∗
Expression of marker CD62L (L-selectin) on monocytes and granulocytes ∗∗∗
Levels of soluble adhesion molecule ICAM-1 ∗∗ ∗∗∗
Levels of soluble adhesion molecule E-selectin ∗∗∗
Levels of interleukin 6 ∗∗
Levels of interleukin 8 ∗∗∗ ∗∗∗ ∗∗∗
Levels of immunoglobulin E ∗∗ ∗
Expression of marker CD66b on eosinophils ∗∗∗ ∗
Expression of marker CD69 on eosinophils ∗∗∗
Asbestos plant: Worker from a former asbestos cement plant had significantly decreased proliferative capacity of lymphocytes uponstimulation by T-cell mitogen. Evaluation of the expression of adhesion molecules on leukocytes showed significant increase in class ofselectins CD62L (L-selectin) on monocytes and granulocytes. The levels of soluble adhesion molecules ICAM-1, proinflammatory cytokinesIL-6 and IL-8 and immunoglobulin E were higher in asbestos workers in comparison to controls. Moreover, significant enhanced expressionin markers CD66 and CD69 on eosinophils was found among expected to asbestos when compared to control groups.Rockwool plant:The proliferative activity of T-lymphocytes in people from rockwool fibre factory was enhanced. Workers had significantly increased levelsof soluble adhesion molecule ICAM-1, interleukin -8 and the levels of immunoglobulin E in comparison with control.Glass fibre plant:Proliferative activity of T-and B-lymphocytes was increased in workers from glass fibre factory. Phenotypic analysis of peripheral bloodleukocytes showed significantly decreased expression of markers CD16+56 (natural killers cells). We found increased level of solubleadhesion molecule E-selectin and IL-8 in sera of people who manipulated with glass fibres. Exposed population had significantly increasedexpression of activation marker CD66b on eosinophils.
∗ P < 0.05.∗∗ P < 0.01.∗∗∗ P < 0.001.
man-made fibres. We propose that the different pat-terns of T-cell proliferative activity found in workersexposed to asbestos versus rockwool and glass fibresmay be due to differences in the duration of exposureto the different fibres as well as differences in theunderlying health status of the populations studied. Incontrast to the relatively good clinical status, shorterduration and low level of fibre exposure in rockwooland glass fibre workers, the former asbestos cementworkers had historically high levels of exposure tofibrogenic dust and showed clinical evidence of as-bestosis and a high prevalence of low forced vitalcapacity. It is notable that a biphasic immune re-sponse has been reported with silica exposure. In a ratmodel, two distinct phases were noted in developmentof silicosis: in early stages, silica activates both hu-moral and cellular immunity; however, in late phasesno activated adaptive immune system effects wereobserved[15]. Consecutive suppression of immunity
after initial stimulation of immune response is wellknown after alcohol consumption[16] or exposure tolow or high levels of the drug dexamethasone[17].
Natural killer cells are a unique lymphocyte pop-ulation with the ability rapidly to lyse tumor cellsindependently of major histocompatibility complexgene products, and are thought to be the first line ofdefence against cancer cells. Several clinical studiesfound impaired NK-cell activity in patients with as-bestosis[18], but decreased NK-activity accompaniedby increased number of circulating NK-cells was alsoseen in workers exposed to asbestos[19]. The resultsfrom our study do not confirm these findings. Neitherasbestos cement nor rockwool workers were notedto have significant changes in NK-cell activity or thepercentage of cells with NK phenotype. However, sig-nificantly decreased expression of marker CD16+56was found in glass fibre-exposed workers in compar-ison with controls. Although the effect was not dra-

J. Tulinska et al. / Mutation Research 553 (2004) 111–124 121
matic, this observation suggests that exposed workersneed to be screened preventively for this marker.
Pathologically relevant increases in the expres-sion and function of adhesion molecules have beenobserved in humans with such pulmonary dis-ease/conditions as bronchial hyperreactivity, allergicrhinitis, idiopathic pulmonary fibrosis or neoplasia[20,21]. In workers from the former asbestos cementplant, the expression of adhesion molecule L-selectinon monocytes and granulocytes was significantly in-creased, while increased levels of soluble adhesionmolecule ICAM-1 were found in people who workedwith all three types of MMMFs. Exposure to glassfibres enhanced the level of soluble E-selectin inworkers’ sera. Thus, adhesion molecules seem to bea sensitive indicator of activation of the immune sys-tem and inflammatory response in humans exposedto mineral fibres.
Inflammatory cytokines are rapidly induced andexpressed early in a disease or injury process. Theymediate and modulate the healing processes but,if overexpressed, may exacerbate the severity of adisease condition as well as give rise to oxidativestress. Up-regulation of IL-8 secretion was foundin patients with fibrosing lung disease and, becauseIL-8 is the main chemotactic and activation factorfor neutrophils, secretion of IL-8 was associated withneutrophil accumulation in the lower respiratory tract[22]. Since the presence of neutrophils is frequentlyreported in humans with asbestosis[23,18], changesin levels of inflammatory cytokines were examined inthe context of the present study. In vitro studies haveshown that asbestos-caused activation of NF�B inA549 cell line and normal human bronchial epithelialcells induces changes in gene expression and releaseof cytokines IL-6 and IL-8[18]. Similarly, Luster[24]found a concentration-dependent increase in secre-tion of immunoreactive IL-8 and IL-6 in pulmonaryepithelial cells type II (line A546) cultivated withcrocidolite asbestos. The increase in concentrationof IL-8 was higher than that of IL-6. The enhancedproduction of IL-6 in the human pulmonary epithelialcell line (BEAS-2B) after cultivation with crocidoliteconfirmed also the findings of Lang[25]. No data onthe effect of mineral fibres on production of cytokinesIL-1, IL-6 and IL-8 have been published.
Our findings show significantly elevated levels ofinterleukin 8 in workers exposed to all three types of
fibres. Levels of interleukin 6 were significantly in-creased only in people from the former asbestos ce-ment plant, while no changes in IL-1� were recordedin exposed populations. Our data indicate that the mostsensitive immune marker of inflammation in mineralfibre exposed-subjects was IL-8. Despite the fact thathighly significant differences were observed betweenpeople exposed to mineral fibres and controls, mostindividual values, as well as group means, were withinthe normal reference range. Proinflammatory cytokineIL-6 was a sensitive marker only in people exposedto asbestos. Although published results from in vitroas well as animal studies indicate possible sensitivityof IL-1� in sera as an indicator of inflammatory re-sponse, our data did not confirm the sensitivity of thismarker for human studies.
Immunoglobulin E is a well known as being in-volved in the mechanisms of development of allergicdiseases. In our study, levels of total immunoglob-ulin E were significantly increased in two popula-tions of workers exposed to fibres-i.e. asbestos-androckwool-exposed workers, while the levels of IgE inthe glass fibre-exposed workers did not differ fromthe corresponding controls. The observation of sig-nificantly increased levels of total immunoglobulin Ein asbestos workers is in agreement with publishedresults of Rosenthal et al.[18] who concluded thatasbestos appears to produce a hyperresponsive state,with chronically exposed individuals manifesting anelevation in circulating immunoglobulins (IgG, IgM,IgE). No data are available on populations occupation-ally exposed to rockwool for comparison.
Another set of indicators of activation of allergic re-sponse is activation markers on eosinophils. In peoplefrom the asbestos cement plant, we observed signifi-cantly increased expression of two markers, CD69 andCD66b, on eosinophils. Workers exposed to glass fi-bres had also elevated expression of activation markerCD66b on eosinophils. Circulating eosinophils are ele-vated in patients with asthma and allergic rhinitis[26].
In contrast to high asbestos exposures of past times,all measured workplace levels of rockwool and glassfibres were lower than Slovak occupational limit. Un-der such conditions, the finding of significant changesin several immune parameters in exposed workers issurprising. With the aim to exclude other possiblefactors of immunostimulation, subjects were matchedwith equal proportions of smokers, since smoking is

122 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
a known stimulator of immune response[27]. Con-sidering the large biological variability in the humanpopulation, we selected large study populations andcompared our results with the normal physiologicalrange. Significant changes were seen in many param-eters, such as levels of soluble adhesion moleculesE-selectin, ICAM-1 or interleukin 6 in sera of exposedworkers, which were above the reference range. In thecase of percentage of CD16+56 cells (NK-cells) inperipheral blood, the mean value for exposed popula-tion was under limit. On the other hand, the levels ofinterlekin 8 recorded in subjects exposed to all threefibres, while highly significantly increased, remainedwithin normal values.
Exposure to asbestos (and possibly other fibres) cangive rise to oxidative stress, which contributes to theinduction of oxidative DNA damage as well as im-munotoxicity. Thus, while markers of oxidative DNAdamage and immunomodulation may not be mecha-nistically linked, examination of their correlations mayprovide supporting evidence of effects of fibres andtheir molecular basis.
Significant correlations were observed betweenlymphocyte proliferation and markers of DNA dam-age. While oxidised pyrimidines and AlkA-sensitivesites generally showed positive associations with pro-liferation in non-asbestos-exposed subjects, in thecase of asbestos-exposed workers, correlations be-tween AlkA-sites and proliferation were negative.FPG-sensitive sites also correlated negatively withT-lymphocyte proliferation-in asbestos-exposed butalso in control subjects. DNA repair showed a neg-ative association withl-selectin expression on gran-ulocytes. Thus, complex but consistent patterns ofassociation are seen. A rational mechanistic explana-tion must await deeper understanding of the involve-ment of immunology in the body’s response to DNAdamage.
5. Conclusions
Immunological lung disorders are accompanied byan array of molecular abnormalities, some of whichcontribute to disease pathogenesis. Understanding theimmunological abnormalities of lung disorders maylead to more effective therapies[28].
Based on our findings, we conclude that exposureto all three types of fibres examined may modulateto different degrees the immune responses. While im-mune changes in workers with high past exposure toasbestos are to be expected, the observed alterations ofimmune responses in people with current exposure tovery low doses of rockwool and glass fibres need to beinterpret with certain degree of reservation in case theyreflect effects of other, confounding factors or changefindings. With these reservations in mind, we can sum-marize our findings as follows: suppression of T-cellimmunity and, to a lesser extent, B-cell immunity werefound in the case of workers from a former asbestoscement plant, while stimulation of T-cell response wasobserved in rockwool workers, and stimulation of T-and B-cell response was seen in glass fibre workers.Depression of the percentage of lymphocyte subpop-ulation of CD 16+56 (natural killer cells) in periph-eral blood was found in glass fibre workers. Statisticalanalysis showed increased levels of proinflammatorycytokines (IL-6 asbestos; IL-8 all three fibres), expres-sion of adhesion molecule L-selectin on granulocytesand monocytes (asbestos), levels of soluble adhesionmolecules in sera (ICAM-1 all three fibres; E-selectinglass fibres), increased levels of immunoglobulin E(asbestos and rockwool) and elevated expression ofactivation markers on eosinophils (CD66b asbestos,glass fibres; CD69 asbestos). Significant correlationswere observed between lymphocyte proliferation andmarkers of DNA damage and repair. Finally, increasedlevels of proinflammatory cytokines, levels of solu-ble adhesion molecules in sera, increased levels ofimmunoglobulin E and elevated expression of activa-tion markers on eosinophils was found in people withsymptoms of hypersensitivity and an elevated inflam-matory status.
Acknowledgements
We would like to express our gratitude to VieraVachálková, Helena Turazová, Anna Morávková,Anna Gažiová, Renáta Mátéová, Kristına Gaval’ová,Zuzana Roštášová, Lubica Miklošková, Jarmila Jan-tošková and Mikuláš Krnác for their excellent tech-nical help. We thank Dr. Katarına Volkovová, Dr.Marta Staruchová, Dr. Alexandra Horská, Dr. ZuzanaDžupinková, Dr. Kornélia Burghardtová and Dr. He-

J. Tulinska et al. / Mutation Research 553 (2004) 111–124 123
lena Petrovská for their help with sampling and withthe comet assay. We thank also Dr. Petrık and Dr.Kocan for measurement of exposure, Dr. Machata,Dr. Šedivcová, Dr. Kopernická, Dr. Holecká for theirhelp with sampling and analysis of clinical data.
This work was supported by the European Union(project no. QLK4-1999-01629), NIEHS Grant # 51020 5240 00000 1 15010 9220, US NIH # 2 D43TW00621-006 and Slovak Grant Agency for Science# 04.92.11.08.
References
[1] M.J. Gardner, C. Magnani, B. Pannett, A.C. Fletcher, P.D.Winter, Lung cancer among glass fibre production workers:a case-control study, Br. J. Ind. Med. 45 (9) (1988) 613–618.
[2] R. Wilson, A.M. Langer, R.P. Nolan, A risk assessment forexposure to glass wool, Regul. Toxicol. Pharmacol. 33 (2001)102–103.
[3] G. Oberdorster, Determinants of the pathogenicity of man-made vitreous fibres (MMVF), Int. Arch. Occup. Environ.Health 73 (Suppl) (2000) S60–S68.
[4] Y. Ishihara, In vitro studies on biological effects of fibrousminerals, Ind. Health 39 (2001) 94–105.
[5] M. Somorovska, E. Szabova, P. Vodicka, J. Tulinska,M. Barancokova, R. Fabry, J. Kubova, Z. Riegerova, H.Petrovska, A. Liskova, K. Rausova, M. Dusinska, A. Collins,Biomonitoring of genotoxic risk in rubber factory comparisonof comet assay with cytogenetic methods and immunology,Mutat. Res. 445 (1999) 181–192.
[6] M. Somorovska, E. Jahnova, J. Tulinska, M. Zamecnikova,J. Sarmanova, A. Terenova, L. Vodickova, A. Liskova, B.Vallova, P. Soucek, K. Hemminki, H. Norpa, A.D. Tates, M.Dusinska, P. Vodicka, Biomonitoring of occupational exposureto styrene in a plastics lamination plant, Mutat. Res. 428(1999) 255–269.
[7] R.J. Smialowicz, In vitro lymphocyte proliferation assays:the mitogen-stimulated response and the mixed-lymphocytereaction in immunotoxicity testing, in: B.R. Burleson, J.H.Dean, A.E. Munson (Eds.), Methods in Immunotoxicology,Wiley-Liss, New York, 1995, pp. 71–108.
[8] J. Tulinska, M. Dusinska, E. Jahnova, A. Liskova, M.Kuricova, P. Vodicka, L. Vodickova, M. Sulcova, L. Fuortes,Changes in cellular immunity among workers occupationallyexposed to styrene in a plastics lamination plant, Am. J. Ind.Med. 5 (2000) 576–583.
[9] L. Fornusek, V. Vetvicka, J. Kopecek, Differences inphagocytic activity of methacrylate copolymer particles innormal and stimulated macrophages, Experientia 37 (1981)418–420.
[10] K.V. Bromelow, J. Galea-Lauri, M.E. O’Brien, B.E.Souberbielle, A highly sensitive whole blood natural killercell assay, J. Immunol. Methods 217 (1998) 177–184.
[11] G.J. Rosenthal, E. Corsini, P. Simeonova, Selected newdevelopments in asbestos immunotoxicity, Environ. HealthPerspect. 106 (Suppl) (1998) 159–169.
[12] M.J. Campbel, M.M. Wagner, M.P. Scott, D.G. Brown,Sequential immunological studies in an asbestos-exposedpopulation. II. Factors affecting lymphocyte function, Clin.Exp. Immunol. 39 (1980) 176–182.
[13] F. Hyodoh, K. Kinugawa, A. Ueki, Effects of asbestos onthe cell cycle of PHA-stimulated human peripheral bloodlymphocytes, Nippon. Eiseigaku Zasshi. 45 (1991) 1074–1081.
[14] E. Corsini, M.I. Luster, J. Mahler, W.A. Craig, M.E.Blazka, G.J. Rosenthal, A protection role for T-lymphocytesin asbestos induced pulmonary inflammation and collagendeposition, Am. J. Respir. Cell Mol. Biol. 5 (1994) 531–539.
[15] R.J. Langley, R. Kalra, N.C. Mishra, F.F. Hahn, S. Razani-Boroujerdi, S.P. Singh, J.M. Benson, J.C. Pena-Philippides,E.B. Barr, M.L. Sopori, A biphasic response to silica. I.Immunostimulation is restricted to the early stage of silicosis,Am. J. Respir. Cell. Mol. Biol. 30 (2004) 823–829.
[16] G. Szabo, Monocytes, alcohol use, and altered immunity,Alcohol Clin. Exp. Res. Aug. 22 (1998) 216S–219S.
[17] M.P. Nambiar, E.J. Enyedy, C.U. Fisher, V.G. Warke,Y.T. Juang, G.C. Tsokos, Dexamethasone modulates TCRzeta chain expression and antigen receptor-mediated earlysignaling events in human T-lymphocytes, Cell Immunol.208 (1) (2001) 62–71.
[18] G.J. Rosenthal, P. Simeonova, E. Corsini, Asbestos toxicity:an immunologic perspective, Rev. Environ. Health 14 (1999)11–20.
[19] P. Froom, N. Lahat, E. Kristal-Boneh, C. Cohen, Y. Lerman,J. Ribak, Circulating natural killer cells in retired asbestoscement workers, J. Occup. Environ. Med. 42 (1) (2000) 19–24.
[20] I. Baumer, G. Zissel, M. Schlaak, J. Muller-Quernheim,Shed soluble ICAM-1 molecules in bronchoalveolar lavagecell supernatants and serum of patients with pulmonarysarcoidosis, Lung 175 (1997) 105–116.
[21] J. Hamacher, T. Schaberg, Adhesion molecules in lungdiseases, Lung 172 (1994) 189–213.
[22] P. Pantelidis, A.M. Southcott, C.M. Black, R.M. du Bois, Up-regulation of IL-8 secretion by alveolar macrophages frompatients with fibrosing alveolitis: a subpopulation analysis,Clin. Exp. Immunol. 108 (1997) 95–104.
[23] V. Galani, S. Constantopoulos, C. Manda-Stachouli,M. Frangou-Lazaridis, A. Mavridis, M. Vassiliou, Y.Dalavanga, Additional proteins in BAL fluid of Metsovitesenvironmentally exposed to asbestos: more evidence of“protection” against neoplasia? Chesta 121 (2002) 273–278.
[24] M.I. Luster, P.P. Simeonova, Asbestos induces inflammatorycytokines in the lung through redox sensitive transcriptionfactors, Toxicol. Lett. 102/103 (1998) 271–275.
[25] D.S. Lang, H. Schocker, S. Hockertz, Effects of crocidoliteasbestos on human bronchoepithelial-dependent fibroblaststimulation in coculture: the role of IL-6 and GM-CSF,Toxicology 159 (1–2) (2001) 81–98.
[26] D. Venarske, R.D. deShazo, Molecular mechanisms of allergicdisease, South Med. J. 96 (11) (2003) 1049–1054.

124 J. Tulinska et al. / Mutation Research 553 (2004) 111–124
[27] A. Zeidel, B. Beilin, I. Yardeni, E. Mayburd, G. Smirnov,H. Bessler, Immune response in asymptomatic smokers, ActaAnaesthesiol. Scand. 46 (8) (2002) 959–964.
[28] P.A. Greenberger, Immunologic aspects of lung diseases andcystic fibrosis, JAMA 278 (22) (1997) 1924–1930.
[29] M. Dušinská, A. Collins, A. Kažimırová, M. Barancková,V. Harrington, K. Volkovová, M. Starchová, A. Horská, L.Wsólová, A. Kocan, J. Petrık , M. Machata, B. Ratcluiffe, S.
Kyrtopoulos, Genotoxic effects of asbestos in humans, Mutat.Res. 553 (1–2) (2004) 91–102.
[30] M. Dušinská, M. Barancková, A. Kažimırová, V. Harrington,K. Volkovová, M. Staruchová, A. Horská, L. Wsólová, A.Collins, Does occupational exposure to mineral fibres causeDNA or chromosome damage? Mutat. Res. 553 (1–2) (2004)103–110.





























