Embed Size (px)
Citation preview

Increased dopamine after mating impairs olfaction andprevents odor interference with pregnancy
Che Serguera, Viviana Triaca, Jakki Kelly-Barrett, Mumna Al Banchaabouchi & Liliana Minichiello
In rodents, social odor sensing influences female reproductive status by affecting neuroendocrine cascades. The odor of male
mouse urine can induce ovulation or block pregnancy within 3 d post coitus. Females avoid the action of such olfactory stimuli
after embryonic implantation. The mechanisms underlying these changes are unknown. Here we report that shortly after mating,
a surge in dopamine in the mouse main olfactory bulb impairs the perception of social odors contained in male urine. Treatment
of females at 6.5 d post coitus with a dopamine D2 receptor antagonist restores social odor sensing and favors disruption of
pregnancy by inhibition of prolactin release, when administered in the presence of alien male urine odors. These results show
that an active sensory barrier blocks social olfactory cues detrimental to pregnancy, consistent with the main olfactory bulb
being a major relay through which social odor modulates reproductive status.
The length of the female rodent ovarian cycle is altered by environ-
mental cues, such as urine odors. These cuesmodulate the release of the
pituitary hormones, such as luteinizing hormone and prolactin (PRL),
that are required for ovulation and pregnancy1. In rodents, the
vaginocervical stimulations produced during copulation block cycling
and trigger a biphasic day and night surge of PRL, the primary
luteotrophic factor2. The luteotrophic action of PRL until midpreg-
nancy is characterized by increased progesterone secretion, which is
essential for implantation of the fertilized ovum, maintenance of
pregnancy3 and inhibition of ovulation2,4. The scent of the urine of
an alien male, but not that of the stud male, blocks embryo implanta-
tion between days 0 and 3 of pregnancy (the Bruce effect)5 by inhibiting
PRL secretion6, which results in a wane in progesterone and a return to
ovulation. This effect is mediated by odorants conveyed through the
vomeronasal organ (VNO)7, a sensory structure located in the nasal
cavity specialized in the detection of pheromones1. In contrast, on days
4 to 6 of pregnancy (the implantation stage), females become refractory
to the Bruce effect8. At this stage, the implanted embryos are still
dependent on progesterone production by the corpus luteum3,9,10.
Thus, it remains elusive why the odor of alien male urine can induce
luteolysis before implantation capability but becomes unable to com-
promise pregnancy soon after implantation.
In the nasal cavity of rodents, the VNO and the main olfactory
epithelium (MOE) contain two sets of neuronal populations with
exclusive chemosensory functions, such as detection of pheromones by
the VNO and detection of odorants by the MOE. Recent evidence
suggests, however, that both structures are essential for pheromone-
mediated responses through specific and nonredundant mechanisms11,
which suggest a higher level of complexity in the integration of
chemical information in rodents. The MOE and the VNO project
parallel fibers that innervate the main olfactory bulb (MOB) and the
accessory olfactory bulb (AOB), respectively. Although the MOB and
the AOB share substantial cellular organization, they are characterized
by divergent projecting areas11 and other specific differences. One such
difference is the absence from the AOB of a prominent dopaminergic
neuronal population, the juxtaglomerular dopaminergic interneurons
(JGD), that is characteristic of the MOB12,13.
MOB JGD cells have been shown to be involved in processing
odorant signaling14, and dopamine is known to regulate odor dis-
crimination and some form of olfactory learning15–17. Tyrosine hydro-
xylase, the rate-limiting enzyme in the synthesis of dopamine18, is
modulated in the mouse MOB by estrogen19, and sex hormones and
vaginocervical stimulations have been shown to modulate tyrosine
hydroxylase synthesis and dopamine release in several nuclei of the
hypothalamus, the limbic system and the midbrain20–22. Given these
previous findings, we asked whether JGD cells influence social
odor perception after mating and whether this, in turn, protects
pregnancy. Here we show that shortly after coitus, tyrosine hydroxylase
increases in femalemouseMOB JGD cells and reaches a plateau at 3.5 d
post coitus (dpc), which coincides with the end of the Bruce effect.
This rise in tyrosine hydroxylase expression is associated with an
impairment in social odor discrimination. Treatment of females at
6.5 dpc with a dopamine D2 receptor (D2R) antagonist restores
olfactory perception abilities and allows alien male urine odors to
disrupt the pregnancy by inhibiting PRL secretion. Our results uncover
an important function of JGD cells in the modulation of social odors
perception detrimental to pregnancy. In addition, these results
strengthen recent evidence and emphasize the role of the MOB in
gating stimuli that affect female reproductive status11 together with the
vomeronasal system1,11.
Received 1 April; accepted 3 June; published online 20 July 2008; doi:10.1038/nn.2154
European Molecular Biology Laboratory, Mouse Biology Unit, Via Ramarini 32, 00015 Monterotondo, Italy. Correspondence should be addressed to L.M.
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 8 [ AUGUST 2008 949
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
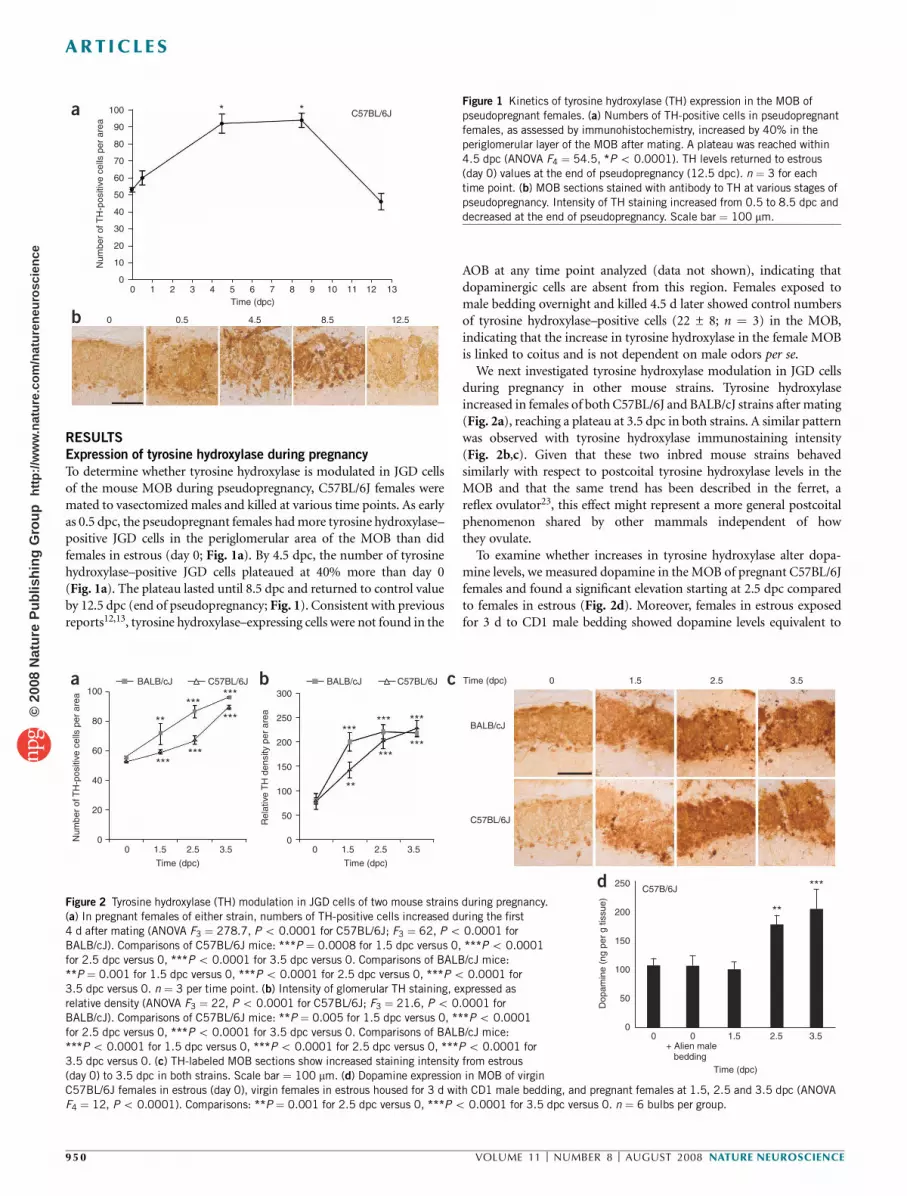
RESULTS
Expression of tyrosine hydroxylase during pregnancy
To determine whether tyrosine hydroxylase is modulated in JGD cells
of the mouse MOB during pseudopregnancy, C57BL/6J females were
mated to vasectomized males and killed at various time points. As early
as 0.5 dpc, the pseudopregnant females hadmore tyrosine hydroxylase–
positive JGD cells in the periglomerular area of the MOB than did
females in estrous (day 0; Fig. 1a). By 4.5 dpc, the number of tyrosine
hydroxylase–positive JGD cells plateaued at 40% more than day 0
(Fig. 1a). The plateau lasted until 8.5 dpc and returned to control value
by 12.5 dpc (end of pseudopregnancy; Fig. 1). Consistent with previous
reports12,13, tyrosine hydroxylase–expressing cells were not found in the
AOB at any time point analyzed (data not shown), indicating that
dopaminergic cells are absent from this region. Females exposed to
male bedding overnight and killed 4.5 d later showed control numbers
of tyrosine hydroxylase–positive cells (22 ± 8; n ¼ 3) in the MOB,
indicating that the increase in tyrosine hydroxylase in the female MOB
is linked to coitus and is not dependent on male odors per se.
We next investigated tyrosine hydroxylase modulation in JGD cells
during pregnancy in other mouse strains. Tyrosine hydroxylase
increased in females of both C57BL/6J and BALB/cJ strains after mating
(Fig. 2a), reaching a plateau at 3.5 dpc in both strains. A similar pattern
was observed with tyrosine hydroxylase immunostaining intensity
(Fig. 2b,c). Given that these two inbred mouse strains behaved
similarly with respect to postcoital tyrosine hydroxylase levels in the
MOB and that the same trend has been described in the ferret, a
reflex ovulator23, this effect might represent a more general postcoital
phenomenon shared by other mammals independent of how
they ovulate.
To examine whether increases in tyrosine hydroxylase alter dopa-
mine levels, we measured dopamine in the MOB of pregnant C57BL/6J
females and found a significant elevation starting at 2.5 dpc compared
to females in estrous (Fig. 2d). Moreover, females in estrous exposed
for 3 d to CD1 male bedding showed dopamine levels equivalent to
C57BL/6J* *
90
100
80
70
60
Num
ber
of
TH
-positiv
e c
ells
per
are
a
50
40
30
20
10
0 1 2 3
0
4 5 6
Time (dpc)
0.5 4.5 8.5 12.5
7 8 9 10 11 12 130
a
b
Figure 1 Kinetics of tyrosine hydroxylase (TH) expression in the MOB of
pseudopregnant females. (a) Numbers of TH-positive cells in pseudopregnant
females, as assessed by immunohistochemistry, increased by 40% in the
periglomerular layer of the MOB after mating. A plateau was reached within
4.5 dpc (ANOVA F4 ¼ 54.5, *Po 0.0001). TH levels returned to estrous
(day 0) values at the end of pseudopregnancy (12.5 dpc). n ¼ 3 for each
time point. (b) MOB sections stained with antibody to TH at various stages of
pseudopregnancy. Intensity of TH staining increased from 0.5 to 8.5 dpc and
decreased at the end of pseudopregnancy. Scale bar ¼ 100 mm.
100 300
250
200
150
100
50
0Num
ber
of T
H-p
ositiv
e c
ells
per
are
a
Rela
tive
TH
density p
er
are
a
Do
pam
ine (
ng p
er
g tis
sue)
BALB/cJ C57BL/6J
80
60
40
20
0 1.5 2.5 3.5 0 1.5 2.5 3.5
Time (dpc) Time (dpc)
Time (dpc)
0
BALB/cJ
BALB/cJ
0 1.5 2.5 3.5C57BL/6J
C57BL/6J
C57B/6J
0 0+ Alien male bedding
Time (dpc)
1.5 2.5
**
***
3.5
250
200
150
100
50
0
******
*** ***
***
***
***
***
**
******
**
a b c
dFigure 2 Tyrosine hydroxylase (TH) modulation in JGD cells of two mouse strains during pregnancy.
(a) In pregnant females of either strain, numbers of TH-positive cells increased during the first
4 d after mating (ANOVA F3 ¼ 278.7, Po 0.0001 for C57BL/6J; F3 ¼ 62, Po 0.0001 for
BALB/cJ). Comparisons of C57BL/6J mice: ***P ¼ 0.0008 for 1.5 dpc versus 0, ***Po 0.0001
for 2.5 dpc versus 0, ***Po 0.0001 for 3.5 dpc versus 0. Comparisons of BALB/cJ mice:
**P ¼ 0.001 for 1.5 dpc versus 0, ***Po 0.0001 for 2.5 dpc versus 0, ***Po 0.0001 for
3.5 dpc versus 0. n ¼ 3 per time point. (b) Intensity of glomerular TH staining, expressed as
relative density (ANOVA F3 ¼ 22, Po 0.0001 for C57BL/6J; F3 ¼ 21.6, Po 0.0001 for
BALB/cJ). Comparisons of C57BL/6J mice: **P ¼ 0.005 for 1.5 dpc versus 0, ***Po 0.0001
for 2.5 dpc versus 0, ***Po 0.0001 for 3.5 dpc versus 0. Comparisons of BALB/cJ mice:
***Po 0.0001 for 1.5 dpc versus 0, ***Po 0.0001 for 2.5 dpc versus 0, ***Po 0.0001 for
3.5 dpc versus 0. (c) TH-labeled MOB sections show increased staining intensity from estrous
(day 0) to 3.5 dpc in both strains. Scale bar ¼ 100 mm. (d) Dopamine expression in MOB of virgin
C57BL/6J females in estrous (day 0), virgin females in estrous housed for 3 d with CD1 male bedding, and pregnant females at 1.5, 2.5 and 3.5 dpc (ANOVA
F4 ¼ 12, Po 0.0001). Comparisons: **P ¼ 0.001 for 2.5 dpc versus 0, ***Po 0.0001 for 3.5 dpc versus 0. n ¼ 6 bulbs per group.
950 VOLUME 11 [ NUMBER 8 [ AUGUST 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

those of unexposed females in estrous. This result indicates that the
postcoital tyrosine hydroxylase increase in female mouse MOB is
translated into a local elevation in dopamine.
Increased tyrosine hydroxylase impairs odor perception
JGD cells in the MOB modulate sensory input from olfactory sensory
neurons (OSNs) to mitral cells (the output neurons of the MOB)
through D2R-mediated presynaptic inhibition14. Functionally, JGD
cells modulate the threshold of odor detection and the discrimination
between odors. D2R antagonists increase odor discrimination and
reduce odor detection threshold, whereas D2R agonists have an
opposite effect24. We examined whether the increase in tyrosine
hydroxylase expression in the MOB during pregnancy is associated
with an impairment of social odorant perception.
The interest of pregnant females in male or female urine odors was
observed at 0.5 and 6.5 dpc, stages at which tyrosine hydroxylase levels
in the MOB are respectively low and high (Fig. 1). Urine can be
separated into two fractions: the low-molecular-weight (LMW) frac-
tion, which causes OSN- and MOB-dependent exploratory behavior
for which the VNO is dispensable25,26; and the high-molecular-weight
(HMW) fraction, the exploration of which requires an intact VNO25.
Female mice prefer to explore male over female urine odors of either
fraction25. Accordingly, at 0.5 dpc, females spent significantly more
time sniffing male urine of either fraction (Fig. 3a,b). At 6.5 dpc,
females explored the HMW fraction of male
urine more than that of females (Fig. 3c) but
were no longer interested in the LMW frac-
tion of male urine (Fig. 3d), indicating that
the processing of inputs through the MOB,
but not the AOB, is altered during pregnancy.
Because tyrosine hydroxylase levels in the
JGD cells at 6.5 dpc were high (Fig. 1a), which
is linked to increased levels of dopamine
(Fig. 2d), we hypothesized that the lack of
social odor discrimination at this stage of
pregnancy is caused by increased dopamine
levels in the MOB. To verify this hypothesis,
we treated pregnant females with the D2R
antagonist spiperone. In rats, spiperone has
been used to increase olfactory discrimination
at 0.6 mg per kg of body weight17. This dose
in mice caused pronounced cataleptic
motor side effects that impaired exploratory
behavior. A lower dosage (0.03 mg per kg
intraperitoneally) did not cause motor side effects, and instead allowed
exploratory behavior and restored in pregnant mice the interest for the
LMWmale urine fraction when administered at 6.5 dpc (Fig. 3e).
To determine whether increased dopamine in the MOB can modify
olfactory sensitivity, we measured the threshold of odor detection
during early gestation. C57BL/6J females, either in estrous or at
6.5 dpc, were given the choice to explore cotton buds soaked with
water or increasing concentrations of the LMW fraction of CD1 male
urine. Females in estrous spent the same amount of time smelling water
and urine, up to a 10–3 dilution of urine (Fig. 4a). In contrast,
females at 6.5 dpc needed at least a tenfold greater concentration
(10–2 dilution) of urine to start exploring (Fig. 4b). Spiperone (0.03 mg
per kg) restored and even improved odor detection in pregnant females
to a lower threshold (10–4 dilution of urine) compared to females
in estrous (Fig. 4c). Moreover, spiperone did not affect tyrosine
hydroxylase levels in the MOB of pregnant females at 4.5 dpc (Supple-
mentary Fig. 1 online), although it did antagonize the action of
dopamine. Because the MOB receives no centrifugal dopaminergic
innervations, the effect of spiperone on olfactory perception is likely to
have been induced by attenuating dopaminergic inhibition of
OSNs. These results indicate that olfaction is altered in pregnant
mice because of a local increase in dopamine in the MOB. The
findings also strongly suggest that the lack of preference for LMW
male urine at 6.5 dpc is a consequence of lower discrimination
30
25
20
15
10
Seconds s
pent
sniffing
HMW urine fraction, 0.5 dpc (n = 9)
***
5
0
HMW urine fraction, 6.5 dpc (n = 13)
**
30
25
20
15
10
Seconds s
pent
sniffing
Seconds s
pent
sniffing
5
0
LMW urine fraction, 6.5 dpc (n = 13)
30
25
20
15
10
Seconds s
pent
sniffing
5
0
LMW urine fraction, 6.5 dpc + spiperone (n = 13)
30
25
20
15
10
Seconds s
pent
sniffing
5
0
***
LMW urine fraction, 0.5 dpc (n = 9)
30
25
20
15
10
5
0
a b c d e
*
Figure 3 Progression of pregnancy is associated with loss of social olfactory discrimination. (a�d) Time spent by females at 0.5 dpc (a,b) and 6.5 dpc (c,d)
sniffing HMW (a,c) or LMW (b,d) odor cues of urine of fertile males or females in estrous. (a), ***P ¼ 0.0006; (b), ***P ¼ 0.0002; (c), **P ¼ 0.002;
(d), P ¼ 0.2 for male versus female urine. (e) Females at 6.5 dpc treated with spiperone 15 min before the social odor discrimination test gained interest
for the LMW male urine fraction (*P ¼ 0.01). Male and female urine fractions are indicated by gender symbols. n ¼ number of females analyzed per group.
C57BL/6 females in estrous C57BL/6 females at 6.5 dpc C57BL/6 females at 6.5 dpc + spiperone
Urine dilutions 10–7
10–5
10–4
10–3
10–2
**
10–5
10–4
10–5
10–4
10–3
10–2
Ratio o
f tim
e s
pent sniffing
uri
ne v
ers
us w
ate
r
3.0
2.0 2.0
1.0
0
3.0
2.0
1.0
0
3.0
1.0
0
***
a b c
Figure 4 Threshold of detection of LMW male urine fraction by C57BL/6J estrous or pregnant females.
Shown are ratios of seconds spent sniffing increasing concentrations of LMW male urine fraction to
seconds spent sniffing water. (a) Females in estrous (n ¼ 5) started detecting the LMW male urine
fraction at 10–3 dilution (ANOVA F4 ¼ 3.4, P ¼ 0.02; *P ¼ 0.04 for 10–3 versus 10–5 or 10–7).
(b) Females at 6.5 dpc (n ¼ 6) needed at least tenfold (10–2) more concentrated solution to start
exploring the LMW male urine fraction (ANOVA F3 ¼ 1.8, P ¼ 0.2; P ¼ 0.07 for 10–2 versus 10–3).
(c) Females at 6.5 dpc treated with spiperone (0.03 mg per kg; n ¼ 5) detected the LMW male urine
fraction at 10–4 dilution (ANOVA F1 ¼ 31, P ¼ 0.0005; ***P ¼ 0.0005 for 10–4 versus 10–5).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 8 [ AUGUST 2008 951
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

and sensitivity to odors rather than of pregnancy-related motivational
changes or habituation to male urine odors.
Pregnancy reduces neuronal activation in the MOB
We next examined whether increased presynaptic inhibition of OSNs
by dopamine hampers neuronal activation in theMOB. To that end, we
evaluated neuronal activation through c-Fos immunoreactivity in the
MOB of C57BL/6J females, either in estrous or pregnant (4.5 dpc),
stimulated or not with CD1 male bedding and treated or not treated
with spiperone. As expected, c-Fos immunoreactivity was significantly
higher in the glomerular, mitral and granular MOB layers of females in
estrous stimulated withmale bedding (Fig. 5a–d). Notably, the number
of c-Fos–immunoreactive neurons in the MOB layers of 4.5-dpc
females stimulated with male bedding was significantly lower than
that of stimulated estrous females. However, in females at 4.5 dpc
treated with spiperone and stimulated with male bedding, c-Fos
immunoreactivity was restored to levels similar to those of females in
estrous stimulated with male bedding. In contrast, spiperone did not
modify c-Fos immunoreactivity in theMOB of unstimulated females in
estrous. These results indicate that neuronal activation is significantly
reduced in all MOB layers of pregnant mice as a consequence of local
increases in dopamine acting through D2R activation. It seems that a
sensory barrier occurs at early stages of pregnancy in the mouse MOB,
presumably linked to the process of pregnancy itself.
Mitral cells of the MOB project to several cortical structures and
limbic nuclei, known as the olfactory cortex and the olfactory amyg-
dala, that in turn extend projections to many other areas of the brain,
including hypothalamic structures that regulate sexual behavior and
reproductive neuroendocrine cascades27–30. We examined whether
decreased neuronal activation in the MOB during pregnancy would
equally affect c-Fos immunoreactivity in the main brain targets of the
MOB in the presence of olfactory cues. c-Fos immunoreactivity was
significantly higher in the anterior olfactory nucleus of C57BL/6J
females in estrous stimulated with CD1 male bedding compared to
control mice, but remained at low levels in pregnant females (4.5 dpc)
stimulated with CD1 male bedding (Fig. 5e). However, in females at
4.5-dpc treated with spiperone and stimulated with CD1male bedding,
c-Fos immunoreactivity was restored to levels similar to that of females
in estrous stimulated with CD1 male bedding (Fig. 5e). This is
consistent with the observation that the MOB hardly responds to
urine odorants at this stage of pregnancy, thus allowing limited output.
Moreover, spiperone in the absence of male urine odorants did not
affect c-Fos immunoreactivity in the anterior olfactory cortex of
females in estrous (Fig. 5e), indicating that c-Fos immunoreactivity
in those mice was induced by odors. In the arcuate nucleus, a structure
pivotal in regulating neuroendocrine cascades associated with cycling
and pregnancy1,4,27, CD1 male odors did not induce significant c-Fos
immunoreactivity in females in estrous or pregnant mice (Fig. 5f), as
observed previously31. c-Fos immunoreactivity in the arcuate nucleus
increased significantly in females at 4.5-dpc treated with spiperone and
stimulated with CD1 male bedding, whereas the drug alone had no
effect on c-Fos immunoreactivity in females in estrous.
The mitral cells of the AOB, in contrast to those of the MOB, avoid
cortical structures and instead send projections to nuclei of the limbic
Figure 5 c-Fos immunoreactivity (c-Fos–ir) is a
marker of neuronal activation. (a�c) Bars from
left to right indicate average number of c-Fos–
positive cells in C57BL/6J females in estrous not
exposed to male bedding (1), in estrous
stimulated with CD1 male bedding (2), in estrous
stimulated with CD1 male bedding and treated
with spiperone (3), pregnant (4.5 dpc) and
stimulated with CD1 male bedding (4) and
4.5 dpc stimulated with CD1 male bedding and
treated with spiperone (5). (a), Glomerular layer of
the MOB (ANOVA F4 ¼ 42.3, Po 0.0001).
(b), Mitral layer (ANOVA F4 ¼ 28, Po 0.0001).
(c), Granular layer (ANOVA F4 ¼ 37, Po
0.0001). Comparisons of stimulated versus
unstimulated: (a), ***Po 0.0001; (b), ***P ¼
0.0002; (c), ***Po 0.0001. Comparisons of
estrous stimulated versus 4.5 dpc stimulated:
(a), **P ¼ 0.002; (b), **P ¼ 0.005;
(c), ***Po 0.0001. Comparisons of 4.5 dpc
treated with spiperone and stimulated versus
estrous stimulated: (a), P ¼ 0.3; (b), *P ¼ 0.05;
(c), *P ¼ 0.04. NS, not significant. (d) Repre-
sentative c-Fos–labeled MOB sections showing
part of the granular (gr) and mitral (mi) layers of
groups 1–5. Scale bar ¼ 100 mm. (e�g) Average
numbers of c-Fos–positive cells in cortical and
hypothalamic nuclei of groups 1–5. (e), Anterior
olfactory nucleus (ANOVA F4 ¼ 13, P ¼ 0.0006).
**P ¼ 0.002 for estrous stimulated versus
estrous unstimulated; **P ¼ 0.003 for estrous
stimulated versus 4.5 dpc stimulated; P ¼ 0.5 for
4.5 dpc treated with spiperone and stimulated versus estrous stimulated; P ¼ 0.7 for estrous unstimulated versus estrous unstimulated and treated with
spiperone. (f), Arcuate nucleus (ANOVA F4 ¼ 44, Po 0.0001). ***Po 0.0001 for 4.5 dpc treated with spiperone and stimulated versus estrous or 4.5 dpc
stimulated; P ¼ 0.2 for estrous unstimulated versus estrous unstimulated and treated with spiperone. (g), Ventromedial hypothalamus (ANOVA F4 ¼ 77,
Po 0.0001). ***Po 0.0001 for estrous stimulated versus estrous unstimulated; ***P ¼ 0.0002 for estrous stimulated versus 4.5 dpc stimulated;
*P ¼ 0.02 for 4.5 dpc treated with spiperone and stimulated versus estrous stimulated; P ¼ 0.2 for estrous unstimulated versus estrous unstimulated and
treated with spiperone. n ¼ 3 per group.
Glo
meru
lar
Num
ber
of
c-F
os–ir c
ells
Ante
rior
olfa
cto
ry n
ucle
us
Num
ber
of
c-F
os–ir c
ells
160
140
120
100
80
60
40
20
0
Mitra
lN
um
ber
of
c-F
os–ir c
ells
0
10
20
30
40
50
60
70
80
90
Gra
nula
rN
um
ber
of c-F
os–ir c
ells
0 Ventr
om
edia
l hypoth
ala
mus
Num
ber
of c-F
os–ir c
ells
05
101520253035404550
Arc
uate
nucle
us
Num
ber
of
c-F
os–ir c
ells
0
5
10
15
20
25
30
35
40
45
100
200
300
400
500
600
700
800
020406080
100120140160180200
MOB layers MOBns
*****
***
*** ***
*** ****
*
*****
*
Different brain regionsns
****
1 2 3 4 5 1 2 3 4 5
+ + + – –
– – – + +
– – + + +
–
Estrous
4.5 dpc
Alien male bedding
Spiperone + – – +
+ + + – –
– – – + +
– – + + +
–
Estrous
4.5 dpc
Alien male bedding
Spiperone + – – +
a
b
c
ed
f
g
952 VOLUME 11 [ NUMBER 8 [ AUGUST 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

system, including the medial amygdaloid nucleus. Neurons from these
nuclei project to hypothalamic nuclei such as the ventromedial
hypothalamus11,32,33. CD1 male bedding significantly increased c-Fos
immunoreactivity in the AOB of females in estrous compared to
unstimulated females in estrous (Supplementary Fig. 2 online).
c-Fos immunoreactivity was also significantly increased by male
bedding in females at 4.5 dpc, although to a lesser extent than in
females in estrous; this effect was not significantly modulated by
spiperone (Supplementary Fig. 2). c-Fos immunoreactivity was
similarly increased in the medial amygdaloid nucleus of females in
estrous or at 4.5 dpc stimulated with CD1 male bedding, and as in the
AOB, this effect was not modulated by spiperone (Supple-
mentary Fig. 2). These results indicate that spiperone does not alter
social odor signaling through the VNO-AOB system in pregnant
females at 4.5 dpc
Notably, after stimulation with CD1 male bedding, c-Fos immuno-
reactivity was significantly lower in the ventromedial hypothalamus of
females at 4.5 dpc compared to females in estrous (Fig. 5g). Moreover,
spiperone restored c-Fos immunoreactivity in pregnant mice stimu-
lated with CD1 male bedding to levels observed in stimulated females
in estrous (Fig. 5g). In contrast, spiperone had no effect on unstimu-
lated females in estrous, indicating a direct response of the ventrome-
dial hypothalamus to MOB stimuli. These results indicate that during
pregnancy, the activation of many brain structures by social odors
undergoes changes similar to those measured in the MOB, implying
that the postcoital dopaminergic block of olfaction modifies the central
influence of social odors.
Male urine odor blocks PRL secretion after implantation
Because male odors can induce ovulation and preclude pregnancy6,34,
we hypothesized that the sensory block of olfaction experienced by a
female mouse soon after coitus is a physiological feature that protects
gestation from male odor stimuli. In rodents, both surges of PRL—the
diurnal surge that peaks around 5 p.m. and the nocturnal surge that
peaks around 3 a.m.—are necessary to maintain pregnancy, and
blocking one of the surges compromises gestation2,4,6. We thus sought
to determine whether detection of male urine odors at 7 dpc by a
pregnant female treated with spiperone would hamper the day and
night surges of PRL necessary at that stage for progesterone production
and embryonic development2–4,9,10.
We first assessed whether spiperone restores olfaction throughout
the time of both surges of PRL. C57BL/6J females at 6.5 dpc were
treated with spiperone (0.3 mg per kg) or control vehicle at 7 a.m., and
their threshold detection for the LMW male urine fraction was
measured 12 h later. Pregnant females treated with spiperone could
still detect the LMWmale urine fraction, starting at a 10–4 dilution, in
contrast to females treated with vehicle (Supplementary Fig. 3 online).
Thus, spiperone has long-term effects on olfaction.
We then tested whether spiperone affects PRL secretion at 7 dpc
when administered in the presence of alien male urine odors. The
LMWconstituents of alienmale urine are sufficient to block pregnancy,
but the HMW constituents improve their effectiveness by concentrat-
ing and fixing them30. We therefore used CD1 male soiled bedding
containing both urine fractions. C57BL/6J females were treated at 7 dpc
with spiperone (0.3 mg per kg) at 3 p.m., 2 h before the peak of the day
surge but 12 h before the peak of the night surge. Serum levels of PRL
were measured at both the day surge peak (5 p.m.) and the night surge
peak (3 a.m.). PRL levels in C57BL/6J females treated with spiperone
and housed with alien (CD1) male bedding were not affected at the
peak of the day surge compared to control females housed with CD1
male bedding, control females treated with spiperone or control
females left undisturbed (Fig. 6a). In contrast, the combination of
spiperone and alien male bedding induced a significant drop in PRL
serum concentration during the night surge compared to control
females left undisturbed, housed with CD1 male bedding or treated
with spiperone only (Fig. 6b). We confirmed pregnancy in each animal
by uterine dissection. These results indicate that spiperone alone is not
able to modify the amplitude of the PRL surges but, presumably by
improving olfactory perception, allows the alien male odors to disrupt
the night surge of PRL. Together, these findings suggest that a maternal
MOB dopaminergic block prevents environmental modulation of PRL
during pregnancy.
Olfactory dopaminergic barrier protects pregnancy
Given the above results, we determined whether restoring olfaction
at 7 and 8 dpc would compromise pregnancy in the presence of
alien male bedding containing both the LMW and HMW urine
fractions. C57BL/6J and BALB/cJ females were treated with spiperone
(0.3 mg per kg) for 2 successive days (6.5 and 7.5 dpc) at 3 p.m. to
restore their ability to discriminate urine odors during this period.
Females of either strain only exposed to the male bedding for 48 h
(from 6.5 to 8.5 dpc) carried their pregnancies to term at a rate of
90–100%, similar to pregnant females left undisturbed (Fig. 7). In
contrast, only 50% of females treated with spiperone at 6.5 and 7.5 dpc
and exposed to alien male bedding for 48 h (from 6.5 to 8.5 dpc)
carried their pregnancies to term. However, females either treated
with spiperone at 6.5 and 7.5 dpc and exposed to original stud male
bedding for 48 h (from 6.5 to 8.5 dpc) or treated with spiperone at
6.5 and 7.5 dpc with no exposure to soiled bedding carried their
pregnancies to term at a rate of 90–100%. This experiment indicates
that odors contained in the bedding of an alien male can still
induce pregnancy failure after embryos are implanted if the dopa-
minergic barrier in the mother’s MOB is concomitantly disrupted.
Moreover, given the results of the previous experiment (Fig. 6b), it is
conceivable that this effect occurs through a drop of the night surge of
PRL at 7 and 8 dpc.
400 Day surge PRL Night surge PRL
350
300
250
200
150
100
Seru
m P
RL (
ng m
l–1)
Seru
m P
RL (
ng m
l–1)
50
Spiperone Spiperone
CD1 soiledbedding
CD1 soiledbedding
0– –
– – – –
––
400
350
300
250
200
150
100
50
0
+ + +
+ +
+
**
++
a bFigure 6 After embryonic implantation, the night surge of PRL is hampered
by alien male odor in females treated with the D2R antagonist spiperone.
Shown are serum PRL levels during day and night surge peaks in C57BL/6J
females 1 week after fecundation (7 dpc). (a) PRL levels during the day surge
peak (5 p.m.) were similarly high in females that were left undisturbed,
treated with spiperone, housed with alien CD1 male bedding or treated with
spiperone and housed with CD1 male bedding (n ¼ 3 for all groups). (b) In
contrast, PRL levels during the night surge peak (3 a.m.) were significantly
lower in females treated with spiperone and housed with CD1 male bedding
compared to control females left undisturbed (ANOVA F3 ¼ 5.5, P ¼ 0.01;
**P ¼ 0.006; n ¼ 6), treated with spiperone (P ¼ 0.01; n ¼ 3) or housed
with CD1 male bedding (P ¼ 0.01; n ¼ 3).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 8 [ AUGUST 2008 953
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

DISCUSSION
In mammals, exchange of body odors favors the establishment of social
and sexual interactions and contributes to the regulation of reproduc-
tion. In rodents, these odorants are mainly contained in urine, affecting
neuroendocrine status, ovulation, opposite-sex tethering and
mating1,27,35,36. Urine constituents activate the VNO-AOB and the
MOE-MOB olfactory systems30, which, through their respective out-
puts11, modulate innate responses such as sexual receptivity (lordo-
sis)25 and ovulation1 and adaptive responses such as sex discrimination
and mate choice35,37. An effective mating therefore requires both
systems to be functional25,37,38. Because odors favor fecundity, they
might harm pregnancy, a condition that is not compatible with cycling.
Here we report that female mice experience a postcoital dopaminergic
surge in the MOB that hampers olfaction. We provide evidence that
male urine odors, channeled through the MOB, decrease PRL release at
7 dpc. This further implicates mouse MOB projections in the regula-
tion of reproductive neuroendocrine status and suggests that in mice,
as well as in hamsters27 and rats39, both sensory systems send inputs to
shared structures such as the amygdala and the hypothalamus and so
interact in eliciting behavioral and neuroendocrine responses. This
suggestion is strengthened by our finding that c-Fos is proportionally
activated in the MOB and the ventromedial hypothalamus by male
odorants during estrous and/or pregnancy. The ventromedial hypotha-
lamus is typically a target of the medial amygdala nucleus32
and contributes, together with other hypothalamic nuclei of the
vomeronasal system, to the integration of olfactory33,35 and vagino-
cervical inputs36 controlling female sexual behavior and postcoital PRL
surges. Thus, odors gated through the MOB might reach the ventro-
medial hypothalamus either directly through projections from the
olfactory cortex or indirectly through MOB efferences that modulate
inputs from the medial amygdala to the ventromedial hypothalamus
and tune mating behaviors35,37 and cycling40.
How odorant cues modulate PRL release is not clear. Because
dopamine release by tuberoinfundibular neurons, located in the
arcuate nucleus, is a major PRL inhibitor through D2R activation in
lactotrophs2, it has been proposed that the
tuberoinfundibular neurons are the final
target of luteolytic male odorants6. We showed
that administration of the D2R antagonist
spiperone concomitant with exposure to
alien male bedding affects the night surge,
but not the day surge, of PRL in females
at 7 dpc, presumably a result of the
PRL-releasing effect of spiperone administered
shortly before the day surge41. In contrast, 12 h
later, because induction of PRL is acute and
brief after spiperone injection41,42, no such
interference occurred; spiperone, which has a
long-term facilitating effect on olfaction,
allowed disruption of the night surge of PRL.
The day and the night surges of PRL in rats are
differently affected by a D2R antagonist42,
indicating that there is variation in the
effects of dopamine and other distinct
factors modulating PRL release at each
surge2. Male odors may have a different impact
on each surge, for example by modu-
lating hypothalamic dopamine during the
day surge and modulating other neuro-
transmitters with effector properties during
the night surge2.
Our observation that MOB inputs can affect PRL release is contrary
to the current view of the MOB being dispensable for blocking
pregnancy in the context of the Bruce effect, with VNO-AOB projec-
tions transmitting the luteolytic stimulus (blocking PRL)6,7,43,44. As
this is true until 3�4 dpc but not later8, the circuits leading to blockade
of PRL surges and return to estrous in response to male odor may be
progressively conditioned by progestational hormonal changes after
mating36. One main effect linked to such changes could be hormonal19
induction of the postcoital dopaminergic surge in the MOB that
effectively obstructs olfaction, which would consequently block
VNO-AOB outputs. In fact, deafferentation of the MOB prevents
c-Fos expression in mouse AOB in response to opposite-sex urine
odorants45, suggesting that themain olfactory pathway indeed provides
modulatory inputs to VNO efferent circuits. This is consistent with our
observation of decreased c-Fos immunoreactivity in the AOB of
females at 4.5 dpc, compared to estrous females, after exposure to
male bedding. It is also notable that the plateau of the dopaminergic
surge in the MOB coincides with the end of the Bruce effect (3�4 dpc)
and the embryonic implantation stage, when maternal investment
prevails over mate choice and estrous-inducing male odors. Thus,
the dopaminergic surge in the MOB may have evolved to prevent
interference of male odors with progestational status from implanta-
tion on. To this extent, the Bruce effect might be an emblematic
manifestation of mate choice that benefits both sexes at periovulation
but becomes deleterious to the species at peri-implantation time.
Our findings emphasize the importance of the MOB in regulating
reproductive processes in mice. VNO ablation, for example, does not
affect estrous or mating46, whereas bulbectomy induces ovarian
atrophy47 and infertility48. In addition, recent studies have challenged
the common view of exclusive functions for the MOB and the AOB,
showing that classical odorants and pheromones can activate either
system49,50. It has also been shown that the mouse MOB not only
projects fibers to neurons expressing luteinizing hormone–releasing
hormone, a key regulator of the ovarian cycle28,29, but also transduces
sensory inputs that regulate sexual and aggressive behaviors38. Finally,
100
a bP
erc
enta
ge o
f pre
gnancie
s t
o t
erm
in C
57B
L/6
J
90
80
***
70
60
50
40
30
20
10
0
100
Perc
enta
ge o
f pre
gnancie
s t
o t
erm
in B
ALB
/cJ
90
80
70
60
50
40
30
20
10
01
+
– –
––
– – – –
–
Plugged Plugged
Spiperone Spiperone
Original studbedding (C57BL/6J)
Original studbedding (BALB/cJ)
Alien(CD1)
Alien(C57BL/6J)
+
+ +
++ +
+
+ + +
2 3 4 5 1
+
– –
––
– – – –
–
+
+ +
++ +
+
+ + +
2
*
3 4 5
bedding bedding
Figure 7 Alien male odors compromise pregnancy. Shown are percentages of pregnancies carried to term
in C57BL/6J (a) and BALB/cJ (b) females. Group 1, females plugged and left undisturbed; group 2,
females plugged and exposed to alien male bedding for 48 h from 6.5 to 8.5 dpc; group 3, females
plugged and injected with spiperone at 6.5 and 7.5 dpc; group 4, females plugged, injected with
spiperone at 6.5 and 7.5 dpc (at 3 p.m.) and exposed to alien male bedding for 48 h from 6.5 to
8.5 dpc (w2 test with 4 degrees of freedom; ***P ¼ 0.0007 for C57BL/6J; *P ¼ 0.01 for BALB/cJ);
group 5, females plugged, injected with spiperone and exposed to original stud male bedding for 48 h
from 6.5 to 8.5 dpc. n ¼ 10 for each group, except n ¼ 12 for group 5 in b.
954 VOLUME 11 [ NUMBER 8 [ AUGUST 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

our results support the idea that the vomeronasal and the olfactory
systems convey convergent sensory inputs27–29 rather than being relays
of unrelated cognitive and instinctive information.
The postcoital rise of dopamine in the MOB and consequent
decrease of olfactory perception abilities are an unexpected mechanism
by which female mice prevent male odor interference with neuroendo-
crine luteotrophic processes. Our results indicate that stimuli gated
through the MOB affect female reproductive status, strengthening the
view that the main olfactory system, together with the vomeronasal
system, contributes to modulation of reproductive behavior and
physiology in rodents.
METHODSExperimental animals. We used 6- to 10-week-old C57BL/6J and BALB/cJ
males and females and CD1 males (Harlan). The EMBL Monterotondo ethical
committee approved all animal procedures.
Dopamine tissue levels. Levels of endogenous dopamine were measured as
described in Supplementary Methods online.
Social olfactory discrimination test and threshold detection of odors. Urine
was collected from fertile BALB/cJ males and females. The LMWurine fraction
was separated from the HMW fraction using a Microcon YM10 centrifugal
filter for 30 min at 14,000g to retain proteins heavier than 10 kDa. The LMW
fraction was collected in the flow-through and the HMW fraction retained in
the filter and resuspended in a volume of water equivalent to the original
volume of urine. The fractions were stored at �80 1C until use.
To measure discrimination for social odors, mice were kept in their own
cages. Cotton buds were impregnated with either 2 ml of male urine or 2 ml of
female urine (either the LMWor HMW fraction) and placed at opposite ends
of the cage. Mice were subjected to two successive trials of 5 min during which
the time spent sniffing the cotton buds impregnated with the LMW, and
90 min later the HMW, urine was quantified using a chronometer. To restore
social odor discrimination, 6.5-dpc females were treated with spiperone
(0.03 mg per kg intraperitoneally; Sigma) 15 min before the social
odor discrimination test.
To measure the lowest concentration of LMWmale urine fraction that could
be detected by C57BL/6J females, the mice were exposed to increasing
concentrations of urine on one side of their cage and water on the other side.
More details are given in Supplementary Methods.
c-Fos immunoreactivity. The induction of c-Fos in the olfactory bulbs and
other brain areas of 2-month-old C57BL/6J virgin estrous females exposed to
alien male (CD1) urine odor was measured and compared to unstimulated
control estrous females or pregnant (4.5 dpc) females, either treated or not with
spiperone (0.03 mg per kg) and exposed or not to alien male urine odor. The
odor exposure always started at 3:30 p.m., and c-Fos expression was measured
after 90 min (time of the second daily prolactin surge) in the five experimental
groups. More details about the experimental groups are given in Supplemen-
tary Methods.
Histology. Histology, cell counts, and tyrosine hydroxylase staining intensity
were analyzed as described in Supplementary Methods.
Pregnancy block experiment. Virgin females (C57BL/6J and BALB/cJ) in
proestrous or early estrous, as assessed by vaginal smears, were housed in a cage
with syngeneic stud males (one male per female) overnight. Females with
vaginal plugs were considered 0.5-dpc. Plugged females were removed from the
stud male and housed individually. For the pregnancy block experiment, five
experimental groups were used. Group 1 females were left undisturbed until the
end of pregnancy. Group 2 females were exposed at 6.5 dpc, in the morning, to
alien male bedding for 48 h (CD1 males for C57BL/6J females and C57BL/6J
males for BALB/cJ females). Group 3 females were injected with spiperone
(0.3 mg per kg) at 6.5 and 7.5 dpc at 3 p.m. and left undisturbed. Group 4
females were injected with spiperone as for group 3 and exposed at 6.5 dpc to
alien male bedding for 48 h. Group 5 females were injected with spiperone as
for group 3 and returned at 6.5 dpc to the original stud male bedding for 48 h.
Pregnancy block was inferred by rating the number of completed gestations
3 weeks after plug. Females not giving birth at the expected time were deemed
not pregnant.
PRL dosing was done as described in Supplementary Methods.
Statistical analysis. A two-way ANOVA test (Statview 5.0) was used to evaluate
statistical significance, followed by a post hoc Fisher’s probable least-squares
difference test. The unpaired Student t test was used when an experiment had
only two groups to compare. The alpha level for all statistical tests wasr0.05.
Error bars of graphs represent standard error of the mean.
Note: Supplementary information is available on the Nature Neuroscience website.
ACKNOWLEDGMENTS
We are grateful to M. Hamon and F. Saurini (INSERM U288) for the tissue
dopamine dosage and C. Sciarretta for comments. C.S. was supported by an
EMBO fellowship.
AUTHOR CONTRIBUTIONS
C.S. planned and performed most of the experiments and, together with L.M.,
drafted the manuscript. V.T. conducted the c-Fos experiment with the help of
J.K.-B. and provided conceptual input. J.K.-B. also contributed to the PRL dosage
experiments. M.A.B. provided the expertise for and helped conduct the behavioral
experiments. L.M. is the PI; she contributed to the experimental plans, supervised
the project, provided theoretical input and wrote the manuscript.
Published online at http://www.nature.com/natureneuroscience/
Reprints and permissions information is available online at http://npg.nature.com/
reprintsandpermissions/
1. Halpern,M. &Martinez-Marcos, A. Structure and function of the vomeronasal system: an
update. Prog. Neurobiol. 70, 245–318 (2003).
2. Freeman, M.E., Kanyicska, B., Lerant, A. & Nagy, G. Prolactin: structure, function and
regulation of secretion. Physiol. Rev. 80, 1523–1631 (2000).
3. Galosy, S.S. & Talamantes, F. Luteotropic actions of placental lactogens atmidpregnancy
in the mouse. Endocrinology 136, 3993–4003 (1995).
4. Erskine, M.S. Prolactin release after mating and genitosensory stimulation in females.
Endocr. Rev. 16, 508–528 (1995).
5. Bruce, H.M. An exteroceptive block to pregnancy in themouse.Nature 184, 105 (1959).
6. Rosser, A.E., Remfry, C.J. & Keverne, E.B. Restricted exposure of mice to primer
pheromones coincident with prolactin surges blocks pregnancy by changing
hypothalamic dopamine release. J. Reprod. Fertil. 87, 553–559 (1989).
7. Lloyd-Thomas, A. & Keverne, E.B. Role of the brain and accessory olfactory system in the
block to pregnancy in mice. Neuroscience 7, 907–913 (1982).
8. Chung, H.J., Reyes, A.B., Watanabe, K., Tomogane, H. & Wakasugi, N. Embryonic
abnormality caused bymale pheromonal effect in pregnancy block inmice.Biol. Reprod.
57, 312–319 (1997).
9. Arkaravichien, W. & Kendle, K.E. Critical progesterone requirement for maintenance of
pregnancy in ovariectomized rats. J. Reprod. Fertil. 90, 63–70 (1990).
10.Milligan, S.R. & Finn, C.A. Minimal progesterone support required for the maintenance
of pregnancy in mice. Hum. Reprod. 12, 602–607 (1997).
11.Dulac, C. & Wagner, S. Genetic analysis of brain circuits underlying pheromone
signaling. Annu. Rev. Genet. 40, 449–467 (2006).
12.Meisami, E. & Bhatnagar, K.P. Structure and diversity inmammalian accessory olfactory
bulb. Microsc. Res. Tech. 43, 476–499 (1998).
13.Baker, H. Species differences in the distribution of substance P and tyrosine hydroxylase
immunoreactivity in the olfactory bulb. J. Comp. Neurol. 252, 206–226 (1986).
14.Ennis, M. et al. Dopamine D2 receptor–mediated presynaptic inhibition of olfactory
nerve terminals. J. Neurophysiol. 86, 2986–2997 (2001).
15.Coopersmith, R., Weihmuller, F.B., Kirstein, C.L., Marshall, J.F. & Leon, M. Extracellular
dopamine increases in the neonatal olfactory bulb during odor preference training.Brain
Res. 564, 149–153 (1991).
16.Harley, C.W. Norepinephrine and dopamine as learning signals. Neural Plast. 11,
191–204 (2004).
17. Yue, E.L., Cleland, T.A., Pavlis, M. & Linster, C. Opposing effects of D1 and D2 receptor
activation on odor discrimination learning. Behav. Neurosci. 118, 184–190 (2004).
18.Halasz, N., Johansson, O., Hokfelt, T., Ljungdahl, A. & Goldstein, M. Immunohisto-
chemical identification of two types of dopamine neuron in the rat olfactory bulb as seen
by serial sectioning. J. Neurocytol. 10, 251–259 (1981).
19.Dluzen, D.E., Park, J.H. & Kim, K. Modulation of olfactory bulb tyrosine hydroxylase and
catecholamine transporter mRNA by estrogen. Brain Res. Mol. Brain Res. 108,
121–128 (2002).
20.Arbogast, L.A. & Voogt, J.L. Progesterone reverses the estradiol-induced decrease in
tyrosine hydroxylase mRNA levels in the arcuate nucleus. Neuroendocrinology 58,
501–510 (1993).
21. Tashiro, Y., Kaneko, T., Nagatsu, I., Kikuchi, H. & Mizuno, N. Increase of tyrosine
hydroxylase–like immunoreactive neurons in the nucleus accumbens and the olfactory
bulb in the rat with the lesion in the ventral tegmental area of the midbrain. Brain Res.
531, 159–166 (1990).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 8 [ AUGUST 2008 955
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

22. Thanky, N.R., Son, J.H. & Herbison, A.E. Sex differences in the regulation of tyrosine
hydroxylase gene transcription by estrogen in the locus coeruleus of TH9-LacZ trans-
genic mice. Brain Res. Mol. Brain Res. 104, 220–226 (2002).
23.Wersinger, S.R. & Baum, M.J. Sexually dimorphic activation of midbrain tyrosine
hydroxylase neurons after mating or exposure to chemosensory cues in the ferret.
Biol. Reprod. 56, 1407–1414 (1997).
24.Wei, C.J., Linster, C. & Cleland, T.A. Dopamine D(2) receptor activation modulates
perceived odor intensity. Behav. Neurosci. 120, 393–400 (2006).
25.Keller, M., Pierman, S., Douhard, Q., Baum, M.J. & Bakker, J. The vomeronasal organ is
required for the expression of lordosis behaviour, but not sex discrimination in female
mice. Eur. J. Neurosci. 23, 521–530 (2006).
26. Spehr, M. et al. Essential role of the main olfactory system in social recognition of
major histocompatibility complex peptide ligands. J. Neurosci. 26, 1961–1970
(2006).
27.Meredith, M. Vomeronasal, olfactory, hormonal convergence in the brain. Cooperation or
coincidence? Ann. NY Acad. Sci. 855, 349–361 (1998).
28. Yoon, H., Enquist, L.W. & Dulac, C. Olfactory inputs to hypothalamic neurons controlling
reproduction and fertility. Cell 123, 669–682 (2005).
29.Boehm, U., Zou, Z. & Buck, L.B. Feedback loops link odor and pheromone signaling with
reproduction. Cell 123, 683–695 (2005).
30.Brennan, P.A. & Zufall, F. Pheromonal communication in vertebrates. Nature 444,
308–315 (2006).
31.Halem, H.A., Cherry, J.A. & Baum, M.J. Central forebrain Fos responses to familiar male
odors are attenuated in recently mated female mice. Eur. J. Neurosci. 13, 389–399
(2001).
32. Choi, G.B. et al. Lhx6 delineates a pathway mediating innate repro-
ductive behaviors from the amygdala to the hypothalamus. Neuron 46, 647–660
(2005).
33. Pierman, S., Douhard, Q. & Bakker, J. Evidence for a role of early oestrogens in the
central processing of sexually relevant olfactory cues in female mice. Eur. J. Neurosci.
27, 423–431 (2008).
34.Marchlewska-Koj, A. Pheromones andmammalian reproduction. Oxf. Rev. Reprod. Biol.
6, 266–302 (1984).
35.Robarts, D.W. & Baum, M.J. Ventromedial hypothalamic nucleus lesions disrupt
olfactory mate recognition and receptivity in female ferrets. Horm. Behav. 51,
104–113 (2007).
36. Lehmann, M.L. & Erskine, M.S. Glutamatergic stimulation of the medial amygdala
induces steroid-dependent c-fos expression within forebrain nuclei responsive to mating
stimulation. Neuroscience 136, 55–64 (2005).
37.Keller, M., Douhard, Q., Baum, M.J. & Bakker, J. Destruction of the main olfactory
epithelium reduces female sexual behavior and olfactory investigation in female mice.
Chem. Senses 31, 315–323 (2006).
38.Mandiyan, V.S., Coats, J.K. & Shah, N.M. Deficits in sexual and aggressive behaviors in
Cnga2 mutant mice. Nat. Neurosci. 8, 1660–1662 (2005).
39.Pro-Sistiaga, P. et al. Convergence of olfactory and vomeronasal projections in the rat
basal telencephalon. J. Comp. Neurol. 504, 346–362 (2007).
40. La Vaque, T.J. & Rodgers, C.H. Recovery of mating behavior in the female rat following
VMH lesions. Physiol. Behav. 14, 59–63 (1975).
41.Arey, B.J., Averill, R.L. & Freeman, M.E. A sex-specific endogenous stimulatory
rhythm regulating prolactin secretion. Endocrinology 124, 119–123 (1989).
42.Mathiasen, J.R. & Voogt, J.L. Differential regulation of the nocturnal and diurnal
prolactin surges in pregnant rats revealed by dopamine receptor antagonism. Neuroen-
docrinology 56, 704–711 (1992).
43.Keverne, E.B. & de la Riva, C. Pheromones in mice: reciprocal interaction between the
nose and brain. Nature 296, 148–150 (1982).
44.Ma, D. et al. Selective ablation of olfactory receptor neurons without functional
impairment of vomeronasal receptor neurons in OMP-ntr transgenic mice. Eur. J.
Neurosci. 16, 2317–2323 (2002).
45.Martel, K.L. & Baum, M.J. Sexually dimorphic activation of the accessory, but not the
main, olfactory bulb in mice by urinary volatiles. Eur. J. Neurosci. 26, 463–475
(2007).
46.Rajendren, G. & Dominic, C.J. Evaluation of involvement of accessory olfactory
(vomeronasal) system in estrous cyclicity and mating in female mice. Indian J. Exp.
Biol. 24, 573–577 (1986).
47.Whitten, W.K. The effect of removal of the olfactory bulbs on the gonads of mice.
J. Endocrinol. 14, 160–163 (1956).
48. Lamond, D.R. Infertility associated with extirpation of the olfactory bulbs in female
albino mice. Aust. J. Exp. Biol. Med. Sci. 36, 103–108 (1958).
49. Lin, D.Y., Zhang, S.Z., Block, E. & Katz, L.C. Encoding social signals in the mouse main
olfactory bulb. Nature 434, 470–477 (2005).
50. Xu, F. et al. Simultaneous activation of mouse main and accessory olfactory bulbs by
odors or pheromones. J. Comp. Neurol. 489, 491–500 (2005).
956 VOLUME 11 [ NUMBER 8 [ AUGUST 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e





























