Embed Size (px)
Citation preview

Interleukin-17 PromotesDevelopmentofCastration-Resistant ProstateCancer Potentially
ThroughCreating an Immunotolerant and Pro-AngiogenicTumorMicroenvironment
Qiuyang Zhang,1 Sen Liu,1 Qingsong Zhang,1,2 Zhenggang Xiong,6 Alun R. Wang,6
Leann Myers,7 Jonathan Melamed,8 Wendell W. Tang,9 and Zongbing You1,2,3,4,5*1Departmentof Structural&Cellular Biology,TulaneUniversity,NewOrleans, Louisiana
2DepartmentofOrthopaedic Surgery,TulaneUniversity,NewOrleans, Louisiana3TulaneCancerCenterand LouisianaCancer ResearchConsortium,TulaneUniversity,NewOrleans, Louisiana4TulaneCenter for StemCell ResearchandRegenerativeMedicine,TulaneUniversity,NewOrleans, Louisiana
5TulaneCenter for Aging,TulaneUniversity,NewOrleans, Louisiana6Departmentof Pathologyand LaboratoryMedicine, SchoolofMedicine,TulaneUniversity,NewOrleans,
Louisiana7Departmentof BiostatisticsandBioinformatics, Schoolof PublicHealthandTropicalMedicine,Tulane
University,NewOrleans, Louisiana8Departmentof Pathology,NewYorkUniversity SchoolofMedicine,NewYork,NewYork
9Departmentof Pathology,OchsnerClinic Foundation,NewOrleans, Louisiana
BACKGROUND. Interleukin-17 (IL-17) has been demonstrated to promote formation andgrowth of hormone-na€ıve prostate adenocarcinoma in mice. IL-17’s role in development ofcastration-resistant prostate cancer is unknown. In the present study, we investigated IL-17’srole in castration-resistant prostate cancer in a mouse model.METHODS. IL-17 receptor C (IL-17RC) deficient mice were interbred with Pten conditionalmutant mice to produce RCþ mice that maintained IL-17RC expression and RC� mice thatwere IL-17RC deficient. Male RCþ and RC� mice were Pten-null and were castrated at
Abbreviations: AR, androgen receptor; Bcl-2; B-cell, lymphoma 2; COX-2, cyclooxygenase-2; Cre, Cre recombinase; CRPC, castration-resistantprostate cancer; CXCL, C-X-C motif ligand; GU, genitourinary; Gr-1, granulocyte-differentiation antigen-1; H&E, hematoxylin and eosin; HER-2,human epidermal growth factor receptor 2; HIF, hypoxia inducible factor; IL, interleukin; IL-17RC, IL-17 receptor C; iNOS, inducible nitric oxidesynthase; KO, knockout; MDSCs, myeloid-derived suppressor cells; MMP, matrix metalloproteinase; MTA1, metastasis associated 1; PIN,prostatic intraepiethelial neoplasia; Pten, phosphatase and tensin homolog; Ras, rat sarcoma gene; SMA, smooth muscle actin; TH17, T helphercells expressing IL-17; TMPRSS2-ERG, transmembrane protease, serine 2 - E-twenty six related gene; TUNEL, terminal deoxynucleotidyltransferase-mediated dUTP nick end labeling; UBE2C, ubiquitin-conjugating enzyme E2C; VEGF, vascular endothelial growth factor; YBX1, Y-box binding protein 1.
Grant sponsor: National Institute of General Medical Sciences; Grant number: P20GM103518; Grant sponsor: National Cancer Institute;Grant number: R01CA174714; Grant sponsor: Department of Defense; Grant numbers: PC121647; PC131448; Grant sponsor: The DevelopmentalFund of Tulane Cancer Center (TCC); Grant sponsor: Louisiana Cancer Research Consortium (LCRC) Fund.
The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutesof Health.�Correspondence to: Zongbing You, Department of Structural & Cellular Biology, Tulane University School of Medicine, 1430 Tulane Ave SL 49,New Orleans, LA 70112. E-mail: [email protected] 27 January 2014; Accepted 5 March 2014DOI 10.1002/pros.22805Published online in Wiley Online Library(wileyonlinelibrary.com).
The Prostate
� 2014 Wiley Periodicals, Inc.

16 weeks of age when invasive prostate cancer had already formed. At 30 weeks of age, allmale mice were analyzed for the prostate phenotypes.RESULTS. RC� mice displayed prostates that were smaller than RCþ mice. Approximately23% of prostatic glands in RC� mice, in contrast to 65% of prostatic glands in RCþ mice,developed invasive adenocarcinomas. Compared to castrate RCþ mice, castrate RC� mouseprostate had lower rates of cellular proliferation and higher rates of apoptosis as well as lowerlevels of MMP7, YBX1, MTA1, and UBE2C proteins. In addition, castrate RC� mouse prostatehad less angiogenesis, which was associated with decreased levels of COX-2 and VEGF.Moreover, castrate RC� mouse prostate had fewer inflammatory cells including lymphocytes,myeloid-derived suppressor cells, and macrophages.CONCLUSIONS. Taken together, our findings suggest that IL-17 promotes development ofinvasive prostate adenocarcinomas under castrate conditions, potentially through creating animmunotolerant and pro-angiogenic tumor microenvironment. Prostate# 2014 Wiley Periodicals, Inc.
KEY WORDS: prostate cancer; castration-resistant; interleukin-17; tumor immunology;tumor microenvironment
INTRODUCTION
Locally confined prostate cancer is treated bysurgery or radiation. At the advanced stage whenmetastases occur, prostate cancer is treated withandrogen deprivation therapy. However, Castration-induced regression of tumor is typically followed byre-growth with castrate levels of androgens, a statusknown as castration-resistant prostate cancer(CRPC) [1]. The mechanisms of how hormone-sensi-tive prostate cancer develops into CRPC remain to bedefined. Alterations of androgen receptor (AR) signal-ing pathways, such as AR gene amplification, increasein AR expression, and AR gene mutations [2], maycause hypersensitivity of AR to low levels of bothendocrine and intracrine androgens [3]. AR splicingvariants may constitutively activate AR signaling inligand-independent manners [4]. AR signaling mayalso be activated by growth factors in the absence ofandrogens [5]. Activation of HER-2/neu and Ras/mitogen-activated protein kinase pathways causesandrogen-independent AR activities [6]. Transcription-al coactivators may lead to ligand-independent ARactivation [7]. Focal neuroendocrine differentiationseems to be a common feature of prostate cancer. Bysecretion of a number of growth factor-like molecules(such as bombesin, calcitonin, and parathyroid hor-mone-related peptide), neuroendocrine cells can sup-port the growth and progression of surroundingprostate cancer cells toward the castration-resistantstate [8]. Androgen ablation up-regulates expression ofthe anti-apoptotic Bcl-2 gene [9] and clusteringene [10], whereas the pro-apoptotic p53 gene is oftenmutated [11]. Decreased PTEN or increased Akt activi-ties are linked to castration-resistant progression ofprostate cancer [12,13]. Expression of TMPRSS2-ERG(transmembrane protease, serine 2-E26 related gene)
fusion protein [14,15] and some microRNAs [16,17]has also been associated with CRPC.
Interleukin-6 (IL-6) and IL-8 have been found to playa role in development of CRPC [18,19]. Both IL-6 and IL-8 are downstream targets of IL-17, a cytokine that isproduced by TH17 cells, gd T cells, and other immunecells [20]. IL-17 acts through a heterodimer of receptorsIL-17RA and IL-17RC [21–23], thus, either Il17ra knock-out (KO) or Il17rc KO completely abolishes IL-17 signal-ing [24,25]. We have previously reported that IL-17RCprotein expression as detected by the anti-IL-17RCintracellular domain antibodies is significantly increasedin CRPC, compared to hormone-sensitive prostate can-cer [26,27]. Recently, we cross-bred Il17rc KO (Il17rc�/�)mice with Pten conditional KO mice (PtenL/L;Creþ) andfound that, in Pten-deficient context, Il17rc KO micedeveloped significantly smaller prostate tumors com-pared to Il17rc wild-type mice [28]. Our findings suggestthat IL-17 promotes formation and growth of hormone-na€ıve prostate adenocarcinoma. However, it is unknownwhether IL-17 plays any role in the development ofCRPC. In the present study, we castrated the mice at16 weeks of age and examined them at 30 weeks of age.We found that Il17rc KO mice developed significantlysmaller prostates compared to Il17rc wild-type miceunder castrate conditions.
MATERIALSANDMETHODS
Mice
Animal protocol was approved by the Animal Careand Use Committee of Tulane University. The breedingstrategy and genotyping protocols have been de-scribed previously, using Ptenloxp/loxp (PtenL/L) mice, PB-Cre4 mice, and Il17rc�/� mice [28]. Male RCþ (n¼ 9)and RC� (n¼ 9) mice at 16 weeks of age were
2 Zhang et al.
The Prostate

castrated. This age was chosen because a majority ofRCþ and RC� mice had already developed invasiveadenocarcinomas by this age [28,29]. The castrationprocedures were as the following: mice were anesthe-tized with 4% isoflurane; the skin over the scrotumwas disinfected by 70% ethanol and Betadine solution;a 0.5-cm incision was made over the scrotum; thetestes were exposed by pulling the adipose tissue; ahemostat was applied to curtail blood flow followedby silk ligation of blood vessels; the ductus deferenteswere ligated and cut; the testes were excised; and theskin incision was closed with #5–0 nylon suture thatwas removed 7 days later. All instruments used weresterile. To alleviate pain, Carprofen (2mg/kg) wasinjected subcutaneously at the end of surgical proce-dure and then every 12 hr up to 48 hr.
Histopathology
Mice were euthanized and weighed at 30 weeks ofage. The genitourinary (GU) blocs were photographed,weighed with an empty bladder, and fixed en bloc asdescribed previously [28,30]. Twenty-eight consecutive4-mm sections of each prostate were cut and foursections (from every seventh section) were stainedwith hematoxylin and eosin (H&E) for histopathologicassessment in a blinded fashion according to the BarHarbor Classification [30]. The prostatic glands wereassessed under low- and high-power magnifications,and approximately 27–94 prostatic glands in eachprostate were counted, with a total of over 500prostatic glands in nine mouse prostates per genotype.The number of inflammatory cells in the connectivetissue space between the prostatic glands was countedin ten high-power fields (�400 magnification) of eachdorsal prostatic lobe; the average number of inflamma-tory cells per high-power field in nine mouse prostatesper genotype was compared.
Immunohistochemicaland TerminalDeoxynucleotidyl Transferase-MediateddUTP
Nick End Labeling Staining
Immunohistochemical staining and double immuno-fluorescent staining were performed as described previ-ously [28]. The antibodies used were: rabbit anti-p-Akt(1:100), mouse anti-PTEN (26H9, 1:50), rabbit anti-YB1(D299, 1:50), and rabbit anti-MTA1 (D40D1, 1:25) fromCell Signaling Technology, Inc., Danvers, MA; rabbitanti-Ki-67 (1:100, EMD Millipore, Billerica, MA); rabbitanti-VEGF (A-20, sc-152, 1:200), goat anti-HIF-1a (Y-15,sc-12542, 1: 50), rabbit anti-NOS2 (iNOS, N-20, sc-651,1:100), rabbit anti-Integrin aM (CD11b, H-61, sc-28664,1:100), goat anti-Ly6C (P-12, sc-23080, 1:100), goat anti-Ly6G (Y-11, sc-103603, 1:100), goat anti-arginase I (V-20,
sc-18345, 1:100), and goat anti-COX-2 (C-20, sc-1745,1:200) from Santa Cruz Biotechnology, Inc., Santa Cruz,CA; rabbit anti-CD31 (Ab28364, 1:50, Abcam, PLC.,Cambridge, MA); rabbit anti-laminin (1:100, Sigma–Aldrich, St. Louis, MO); rabbit anti-a-smooth muscleactin (1:200, Pierce Biotechnology, Rockford, IL); goatanti-MMP7 (1:200, R&D systems, Minneapolis, MN);rabbit anti-UbcH10/UBE2C (1:200, Boston Biochem,Cambridge, MA); and Cy3-conjugated anti-goat IgG andDyLight 488-conjugated anti-rabbit IgG (Jackson Immu-noResearch Laboratories, Inc., West Grove, PA). Terminaldeoxynucleotidyl transferase-mediated dUTP nick endlabeling (TUNEL) staining was performed using TACS.XL1 Blue Label In Situ Apoptosis Detection Kits(Trevigen, Inc., Gaithersburg, MD) according to themanufacturer’s instructions [28]. To quantify Ki-67-posi-tive and TUNEL-positive cells, three animals from eachgenotype group were randomly selected; three represen-tative prostate sections from each animal were stained;approximately 200 cells per field of ten high-power fields(�400 magnification) of each prostate lobe were counted;and the percentages of positive cells were calculated asthe number of positive cells divided by the total numberof cells. The density of microvessels was evaluated bycounting the CD31-positive microvessels in ten high-power fields per lobe; the average number of CD31-positive microvessels per high-power field in threerandom mouse prostates per genotype was compared.The myeloid-derived suppressor cells (MDSCs) weredefined as CD11b/Gr-1 (CD11b/Ly6C and CD11b/Ly6G) double positive cells. The M1 and M2 macro-phages were defined as iNOS-positive and arginase I-positive cells, respectively. The numbers of MDSCs, M1macrophages, and M2 macrophages in the connectivetissue space between the prostatic glands were countedin ten high-power fields (�400 magnification) in eachdorsal prostatic lobe; the average numbers of MDSCs,M1 and M2 macrophages per high-power field in threerandommouse prostates per genotype were compared.
Statistical Analysis
Comparisons of the GU-bloc weights were analyzedusing Student’s t-test. Kruskal–Wallis test was used tocompare the incidences of normal, PIN and invasiveadenocarcinoma. Student’s t-test was used to analyzethe remaining data.
RESULTS
Castrate RC�MiceDeveloped Smaller ProstateGlandsThanCastrate RCþMice
Previously, we found that there were no signifi-cant differences in the expression of Il17rc mRNA
IL-17 Promotes CRPC 3
The Prostate
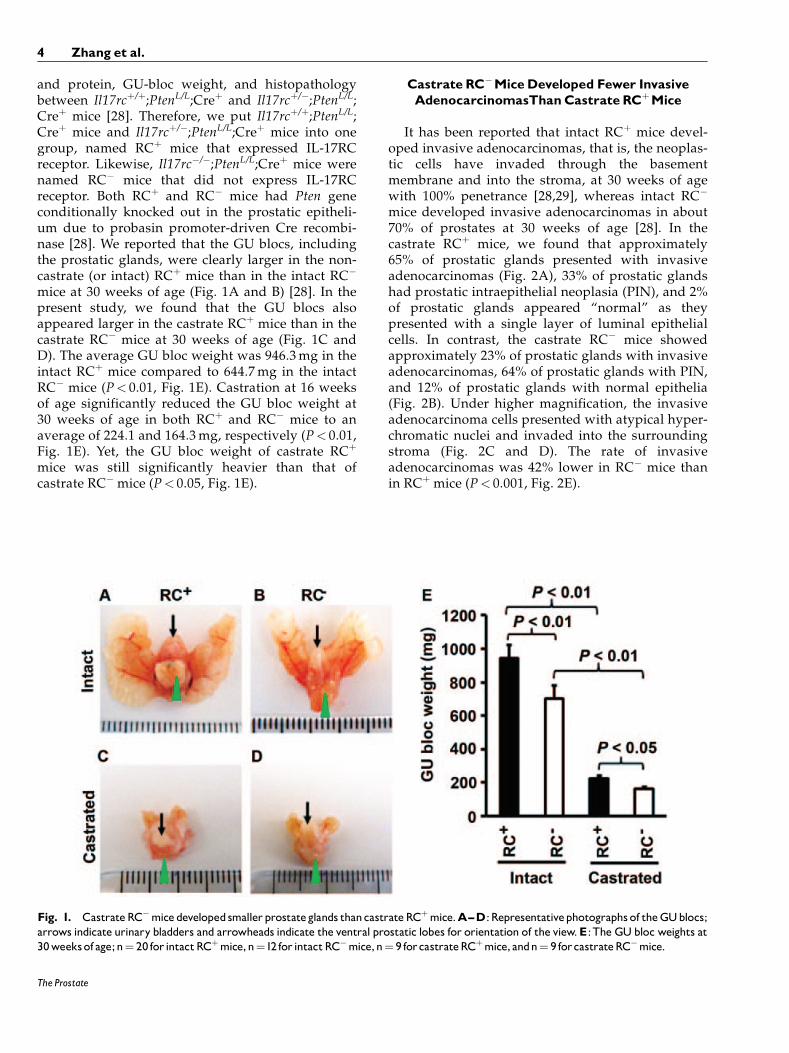
and protein, GU-bloc weight, and histopathologybetween Il17rcþ/þ;PtenL/L;Creþ and Il17rcþ/�;PtenL/L;Creþ mice [28]. Therefore, we put Il17rcþ/þ;PtenL/L;Creþ mice and Il17rcþ/�;PtenL/L;Creþ mice into onegroup, named RCþ mice that expressed IL-17RCreceptor. Likewise, Il17rc�/�;PtenL/L;Creþ mice werenamed RC� mice that did not express IL-17RCreceptor. Both RCþ and RC� mice had Pten geneconditionally knocked out in the prostatic epitheli-um due to probasin promoter-driven Cre recombi-nase [28]. We reported that the GU blocs, includingthe prostatic glands, were clearly larger in the non-castrate (or intact) RCþ mice than in the intact RC�
mice at 30 weeks of age (Fig. 1A and B) [28]. In thepresent study, we found that the GU blocs alsoappeared larger in the castrate RCþ mice than in thecastrate RC� mice at 30 weeks of age (Fig. 1C andD). The average GU bloc weight was 946.3mg in theintact RCþ mice compared to 644.7mg in the intactRC� mice (P< 0.01, Fig. 1E). Castration at 16 weeksof age significantly reduced the GU bloc weight at30 weeks of age in both RCþ and RC� mice to anaverage of 224.1 and 164.3mg, respectively (P< 0.01,Fig. 1E). Yet, the GU bloc weight of castrate RCþ
mice was still significantly heavier than that ofcastrate RC� mice (P< 0.05, Fig. 1E).
Castrate RC�MiceDeveloped Fewer InvasiveAdenocarcinomasThanCastrate RCþMice
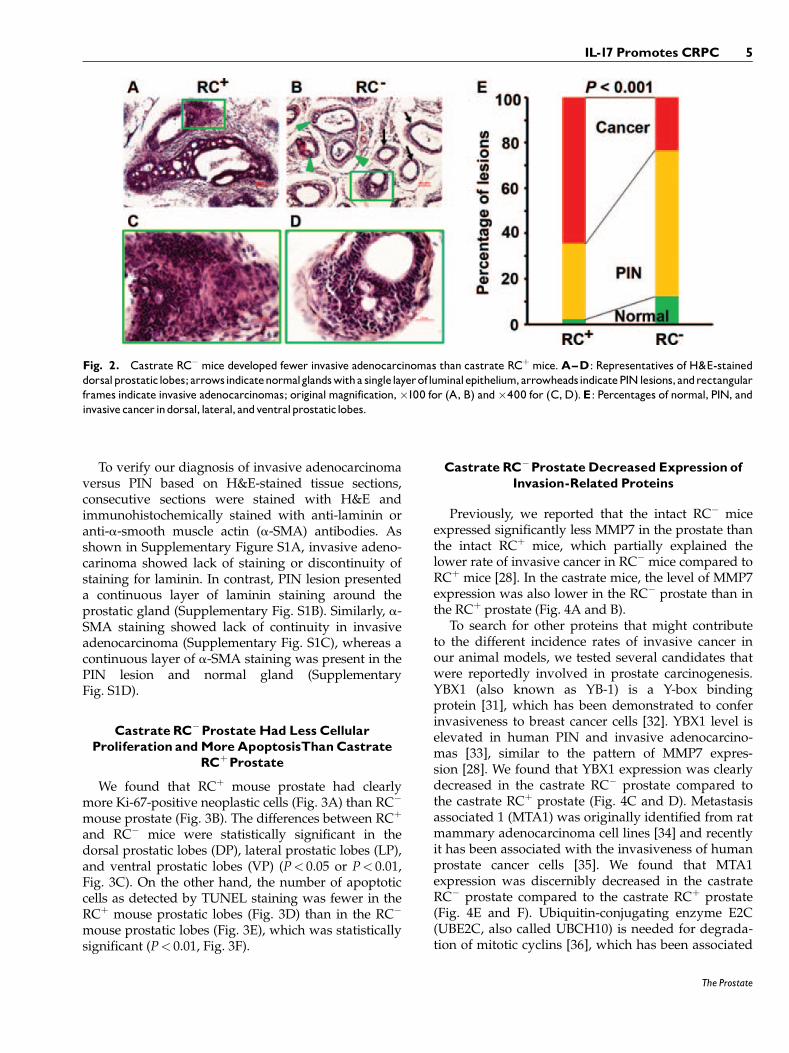
It has been reported that intact RCþ mice devel-oped invasive adenocarcinomas, that is, the neoplas-tic cells have invaded through the basementmembrane and into the stroma, at 30 weeks of agewith 100% penetrance [28,29], whereas intact RC�
mice developed invasive adenocarcinomas in about70% of prostates at 30 weeks of age [28]. In thecastrate RCþ mice, we found that approximately65% of prostatic glands presented with invasiveadenocarcinomas (Fig. 2A), 33% of prostatic glandshad prostatic intraepithelial neoplasia (PIN), and 2%of prostatic glands appeared “normal” as theypresented with a single layer of luminal epithelialcells. In contrast, the castrate RC� mice showedapproximately 23% of prostatic glands with invasiveadenocarcinomas, 64% of prostatic glands with PIN,and 12% of prostatic glands with normal epithelia(Fig. 2B). Under higher magnification, the invasiveadenocarcinoma cells presented with atypical hyper-chromatic nuclei and invaded into the surroundingstroma (Fig. 2C and D). The rate of invasiveadenocarcinomas was 42% lower in RC� mice thanin RCþ mice (P< 0.001, Fig. 2E).
Fig. 1. CastrateRC�micedevelopedsmallerprostateglands thancastrateRCþmice.A^D:Representativephotographs of theGUblocs;arrows indicate urinary bladders and arrowheads indicate the ventral prostatic lobes for orientation of the view.E:The GUbloc weights at30weeksof age;n¼ 20 for intactRCþmice,n¼12 for intactRC�mice,n¼ 9 forcastrateRCþmice, andn¼ 9 forcastrateRC�mice.
4 Zhang et al.
The Prostate
To verify our diagnosis of invasive adenocarcinomaversus PIN based on H&E-stained tissue sections,consecutive sections were stained with H&E andimmunohistochemically stained with anti-laminin oranti-a-smooth muscle actin (a-SMA) antibodies. Asshown in Supplementary Figure S1A, invasive adeno-carinoma showed lack of staining or discontinuity ofstaining for laminin. In contrast, PIN lesion presenteda continuous layer of laminin staining around theprostatic gland (Supplementary Fig. S1B). Similarly, a-SMA staining showed lack of continuity in invasiveadenocarcinoma (Supplementary Fig. S1C), whereas acontinuous layer of a-SMA staining was present in thePIN lesion and normal gland (SupplementaryFig. S1D).
Castrate RC�ProstateHad Less CellularProliferation andMoreApoptosisThanCastrate
RCþProstate
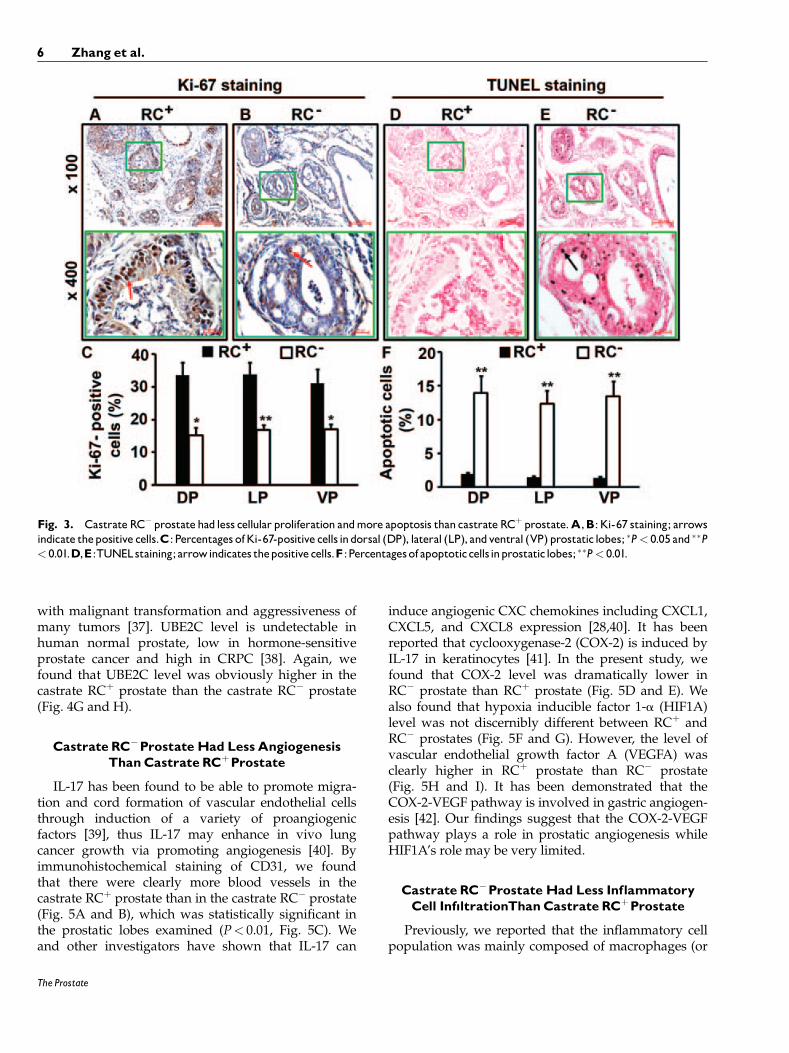
We found that RCþ mouse prostate had clearlymore Ki-67-positive neoplastic cells (Fig. 3A) than RC�
mouse prostate (Fig. 3B). The differences between RCþ
and RC� mice were statistically significant in thedorsal prostatic lobes (DP), lateral prostatic lobes (LP),and ventral prostatic lobes (VP) (P< 0.05 or P< 0.01,Fig. 3C). On the other hand, the number of apoptoticcells as detected by TUNEL staining was fewer in theRCþ mouse prostatic lobes (Fig. 3D) than in the RC�
mouse prostatic lobes (Fig. 3E), which was statisticallysignificant (P< 0.01, Fig. 3F).
Castrate RC�ProstateDecreased Expression ofInvasion-Related Proteins
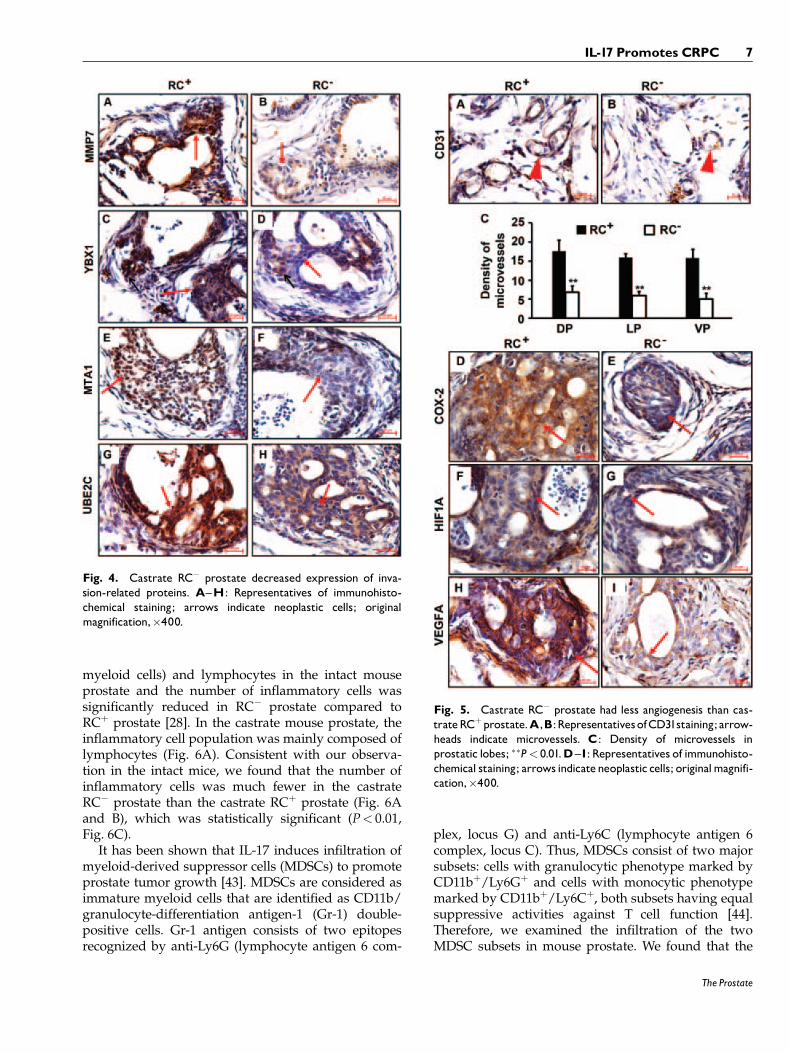
Previously, we reported that the intact RC� miceexpressed significantly less MMP7 in the prostate thanthe intact RCþ mice, which partially explained thelower rate of invasive cancer in RC� mice compared toRCþ mice [28]. In the castrate mice, the level of MMP7expression was also lower in the RC� prostate than inthe RCþ prostate (Fig. 4A and B).
To search for other proteins that might contributeto the different incidence rates of invasive cancer inour animal models, we tested several candidates thatwere reportedly involved in prostate carcinogenesis.YBX1 (also known as YB-1) is a Y-box bindingprotein [31], which has been demonstrated to conferinvasiveness to breast cancer cells [32]. YBX1 level iselevated in human PIN and invasive adenocarcino-mas [33], similar to the pattern of MMP7 expres-sion [28]. We found that YBX1 expression was clearlydecreased in the castrate RC� prostate compared tothe castrate RCþ prostate (Fig. 4C and D). Metastasisassociated 1 (MTA1) was originally identified from ratmammary adenocarcinoma cell lines [34] and recentlyit has been associated with the invasiveness of humanprostate cancer cells [35]. We found that MTA1expression was discernibly decreased in the castrateRC� prostate compared to the castrate RCþ prostate(Fig. 4E and F). Ubiquitin-conjugating enzyme E2C(UBE2C, also called UBCH10) is needed for degrada-tion of mitotic cyclins [36], which has been associated
Fig. 2. Castrate RC� mice developed fewer invasive adenocarcinomas than castrate RCþ mice.A^D: Representatives of H&E-staineddorsalprostatic lobes; arrowsindicatenormalglandswitha singlelayerofluminalepithelium,arrowheadsindicatePINlesions, andrectangularframes indicate invasive adenocarcinomas; originalmagnification,�100 for (A, B) and�400 for (C,D).E: Percentages of normal, PIN, andinvasivecancer indorsal, lateral, andventralprostatic lobes.
IL-17 Promotes CRPC 5
The Prostate
with malignant transformation and aggressiveness ofmany tumors [37]. UBE2C level is undetectable inhuman normal prostate, low in hormone-sensitiveprostate cancer and high in CRPC [38]. Again, wefound that UBE2C level was obviously higher in thecastrate RCþ prostate than the castrate RC� prostate(Fig. 4G and H).
Castrate RC�ProstateHad LessAngiogenesisThanCastrate RCþProstate
IL-17 has been found to be able to promote migra-tion and cord formation of vascular endothelial cellsthrough induction of a variety of proangiogenicfactors [39], thus IL-17 may enhance in vivo lungcancer growth via promoting angiogenesis [40]. Byimmunohistochemical staining of CD31, we foundthat there were clearly more blood vessels in thecastrate RCþ prostate than in the castrate RC� prostate(Fig. 5A and B), which was statistically significant inthe prostatic lobes examined (P< 0.01, Fig. 5C). Weand other investigators have shown that IL-17 can
induce angiogenic CXC chemokines including CXCL1,CXCL5, and CXCL8 expression [28,40]. It has beenreported that cyclooxygenase-2 (COX-2) is induced byIL-17 in keratinocytes [41]. In the present study, wefound that COX-2 level was dramatically lower inRC� prostate than RCþ prostate (Fig. 5D and E). Wealso found that hypoxia inducible factor 1-a (HIF1A)level was not discernibly different between RCþ andRC� prostates (Fig. 5F and G). However, the level ofvascular endothelial growth factor A (VEGFA) wasclearly higher in RCþ prostate than RC� prostate(Fig. 5H and I). It has been demonstrated that theCOX-2-VEGF pathway is involved in gastric angiogen-esis [42]. Our findings suggest that the COX-2-VEGFpathway plays a role in prostatic angiogenesis whileHIF1A’s role may be very limited.
Castrate RC�ProstateHad Less InflammatoryCell Inf|ltrationThanCastrate RCþProstate
Previously, we reported that the inflammatory cellpopulation was mainly composed of macrophages (or
Fig. 3. Castrate RC� prostatehad less cellular proliferation andmore apoptosis than castrate RCþ prostate.A,B: Ki-67 staining; arrowsindicate thepositive cells.C: Percentages ofKi-67-positive cells indorsal (DP), lateral (LP), andventral (VP) prostatic lobes; �P< 0.05 and ��P< 0.01.D,E:TUNELstaining;arrowindicates thepositive cells.F :Percentagesof apoptoticcellsinprostatic lobes; ��P< 0.01.
6 Zhang et al.
The Prostate
myeloid cells) and lymphocytes in the intact mouseprostate and the number of inflammatory cells wassignificantly reduced in RC� prostate compared toRCþ prostate [28]. In the castrate mouse prostate, theinflammatory cell population was mainly composed oflymphocytes (Fig. 6A). Consistent with our observa-tion in the intact mice, we found that the number ofinflammatory cells was much fewer in the castrateRC� prostate than the castrate RCþ prostate (Fig. 6Aand B), which was statistically significant (P< 0.01,Fig. 6C).
It has been shown that IL-17 induces infiltration ofmyeloid-derived suppressor cells (MDSCs) to promoteprostate tumor growth [43]. MDSCs are considered asimmature myeloid cells that are identified as CD11b/granulocyte-differentiation antigen-1 (Gr-1) double-positive cells. Gr-1 antigen consists of two epitopesrecognized by anti-Ly6G (lymphocyte antigen 6 com-
plex, locus G) and anti-Ly6C (lymphocyte antigen 6complex, locus C). Thus, MDSCs consist of two majorsubsets: cells with granulocytic phenotype marked byCD11bþ/Ly6Gþ and cells with monocytic phenotypemarked by CD11bþ/Ly6Cþ, both subsets having equalsuppressive activities against T cell function [44].Therefore, we examined the infiltration of the twoMDSC subsets in mouse prostate. We found that the
Fig. 4. Castrate RC� prostate decreased expression of inva-sion-related proteins. A^H: Representatives of immunohisto-chemical staining; arrows indicate neoplastic cells; originalmagnification,�400.
Fig. 5. Castrate RC� prostate had less angiogenesis than cas-trateRCþprostate.A,B:RepresentativesofCD31staining;arrow-heads indicate microvessels. C: Density of microvessels inprostatic lobes; ��P< 0.01.D^I: Representatives of immunohisto-chemical staining; arrows indicateneoplastic cells; originalmagnifi-cation,�400.
IL-17 Promotes CRPC 7
The Prostate
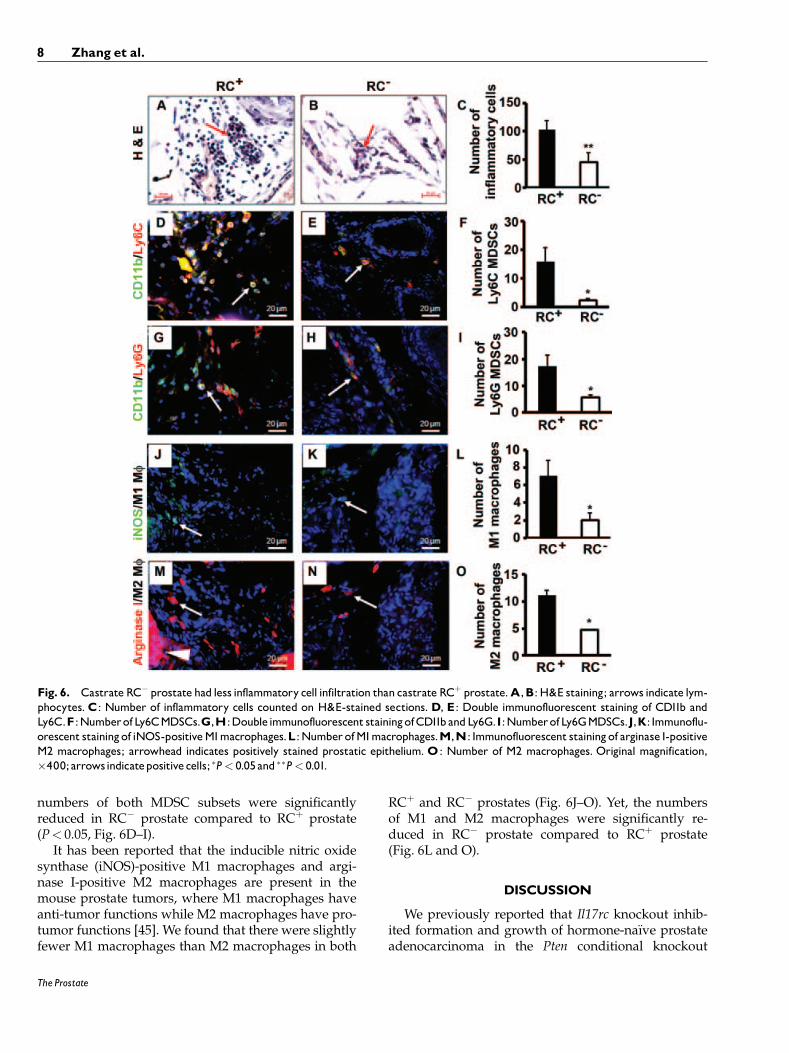
numbers of both MDSC subsets were significantlyreduced in RC� prostate compared to RCþ prostate(P< 0.05, Fig. 6D–I).
It has been reported that the inducible nitric oxidesynthase (iNOS)-positive M1 macrophages and argi-nase I-positive M2 macrophages are present in themouse prostate tumors, where M1 macrophages haveanti-tumor functions while M2 macrophages have pro-tumor functions [45]. We found that there were slightlyfewer M1 macrophages than M2 macrophages in both
RCþ and RC� prostates (Fig. 6J–O). Yet, the numbersof M1 and M2 macrophages were significantly re-duced in RC� prostate compared to RCþ prostate(Fig. 6L and O).
DISCUSSION
We previously reported that Il17rc knockout inhib-ited formation and growth of hormone-na€ıve prostateadenocarcinoma in the Pten conditional knockout
Fig. 6. Castrate RC� prostate had less inflammatory cell infiltration than castrate RCþ prostate.A,B: H&E staining; arrows indicate lym-phocytes.C: Number of inflammatory cells counted on H&E-stained sections.D, E: Double immunofluorescent staining of CD11b andLy6C.F :NumberofLy6CMDSCs.G,H:Doubleimmunofluorescent stainingofCD11bandLy6G.I:NumberofLy6GMDSCs.J,K: Immunoflu-orescent stainingof iNOS-positiveM1macrophages.L:NumberofM1macrophages.M,N: Immunofluorescent stainingof arginase I-positiveM2 macrophages; arrowhead indicates positively stained prostatic epithelium.O: Number of M2 macrophages. Original magnification,�400;arrowsindicatepositivecells; �P< 0.05and ��P< 0.01.
8 Zhang et al.
The Prostate

mouse model [28]. In the present study, we used thesame mouse model and castrated the animals at16 weeks of age. Fourteen weeks after castration, wefound that invasive adenocarcinomas were present inboth RCþ and RC� mouse prostates, albeit at differentincidence rates. The GU bloc size, judged by the GUbloc weight, was significantly smaller in the castratemice than the intact mice (Fig. 1). The remaininginvasive adenocarcinomas in the castrate mice pre-sumably are CRPC based on the direct evidence thatcellular proliferation was still present 14 weeks afterthe absence of testicular androgens. Another line ofindirect evidence is that UBE2C level was very high inthe castrate RCþ mouse prostate as UBE2C was onlyexpressed at high levels in CRPC [38].
The most significant phenotypic difference betweenthe castrate RCþ and RC� mice is the incidence rate ofinvasive adenocarcinomas. RCþ mice presented inva-sive adenocarcinomas in 65% of prostatic glands, insharp contrast to 23% of prostatic glands in RC� mice(Fig. 2E). These incidence rates are much lower thanthe rates in the non-castrate RCþ and RC� mice (i.e.,100% and 70%, respectively) [28,29]. One possiblereason is that the AR-positive prostatic epitheliumundergoes increased apoptosis in response to castra-tion. A 10-times increase of apoptotic cells in the Pten-null prostate cancer was found compared to the intactmouse prostate cancer, and the apoptotic rate waseven higher in the prostate cancer than the normalepithelium 3 days post-castration [29]. Apoptosis maypossibly lead to regression of some invasive adenocar-cinomas and PIN lesions, resulting in a heterogeneousappearance of normal epithelium, PIN, and invasiveadenocarcinomas.
There are several possible reasons to explain thedifferent incidence rates of invasive adenocarcinomasbetween the castrate RCþ and RC� mice. First, thecellular proliferation rate is higher in RCþ prostate thanRC� prostate while the apoptotic rate is lower in RCþ
prostate than RC� prostate. These differences confer anadvantage to the tumor growth in RCþ mice over RC�
mice. Second, several proteins, namely, MMP7, YBX1,MTA1, and UBE2C, are expressed at higher levels inRCþ prostate than RC� prostate. These proteins havebeen associated with the invasiveness and aggres-siveness of human prostate cancer cells [28,32,33,35,38].While we have demonstrated that MMP7 is a directdownstream target of IL-17 signaling pathway [28], itremains to be determined if YBX1, MTA1, and UBE2Care also IL-17 downstream targets. Third, angiogenesis isreduced in RC� prostate compared to RCþ prostate.Angiogenesis is an integral hallmark of cancer and it hasrecently been associated with early neoplastic progres-sion besides its well-known role in macroscopictumors [46]. One mechanism by which IL-17 promotes
lung cancer growth is through induction of angiogene-sis [40]. The reduced levels of COX-2 and VEGFA inRC� prostate may be responsible for the decreasedangiogenesis, as the COX-2-VEGF pathway has beenassociated with gastric angiogenesis [42]. COX-2 is an IL-17 downstream target in keratinocytes [41]. And last,inflammatory cell infiltration is reduced in RC� prostatecompared to RCþ prostate. The number of inflammatorycells appears to be more in the prostate of the castratemice than the intact mice. It has been reported thatandrogen ablation increased infiltration of CD4þ T cellsand macrophages in human prostate tumors [47]. Theinflammatory cell population shifts from myeloid cells/lymphocytes in the intact mice to mainly lymphocytes inthe castrate mice. This finding is in line with a recentreport that castration elicits infiltration of TH1 cellsfollowed by predominantly TH17 cells in rat prostate[48]. The subtypes of lymphocytes in our animal modelsare the subjects of our ongoing studies. Nevertheless, wehave shown that the numbers of two major myeloid celltypes, MDSCs and macrophages, are significantly re-duced in RC� mice compared to RCþ mice. We haverecently demonstrated that IL-17 is a chemoattractant formonocytes/macrophages [49]. The reduced infiltrationof MDSCs and macrophages may be caused by lack ofIL-17RC receptor on these cells and/or indirectly by thedecreased chemokine levels in the tumor microenviron-ment of RC� mice. Since MDSCs and M2 macrophagesare pro-tumor inflammatory cells, a decrease in theirnumbers may partially contribute to the phenotype ofreduced incidence rate of invasive adenocarcinomas inRC� mice.
In summary, the present study demonstrates thatIL-17 promotes development of CRPC in the Ptenconditional knockout mouse model. IL-17 may affectseveral hallmark capabilities of cancer, including sus-taining proliferation, resisting cell death, activatinginvasion, inducing angiogenesis, and recruiting pro-tumor inflammatory cells [46]. These findings suggestthat blocking IL-17 signaling through pharmacologicalinterventions may have potentials in the preventionand treatment of CRPC.
CONCLUSIONS
IL-17 promotes development of invasive prostateadenocarcinomas in Pten conditional knockout miceunder castrate conditions, potentially through creatingan immunotolerant and pro-angiogenic tumor micro-environment.
ACKNOWLEDGMENTS
We thank Drs. Prescott L. Deininger, Asim B.Abdel-Mageed, Steven M. Hill, David E. Blask, Brian
IL-17 Promotes CRPC 9
The Prostate

G. Rowan, and Oliver Sartor (Tulane University) fortheir advices and comments on the manuscript. TulaneCancer Center Core Facilities were used in this study.We thank Dr. Wenjun Ouyang and Genentech forproviding the Il17rc�/� mice and NCI MMHCC forproviding the PB-Cre4 mice.
REFERENCES
1. Harris WP, Mostaghel EA, Nelson PS, Montgomery B. Androgendeprivation therapy: Progress in understanding mechanisms ofresistance and optimizing androgen depletion. Nat Clin PractUrol 2009;6(2):76–85.
2. Newmark JR, Hardy DO, Tonb DC, Carter BS, Epstein JI, IsaacsWB, Brown TR, Barrack ER. Androgen receptor gene mutationsin human prostate cancer. Proc Natl Acad Sci USA 1992;89(14):6319–6323.
3. Mohler JL, Gregory CW, Ford OH III, Kim D, Weaver CM,Petrusz P, Wilson EM, French FS. The androgen axis in recurrentprostate cancer. Clin Cancer Res 2004;10(2):440–448.
4. Hu R, Dunn TA, Wei S, Isharwal S, Veltri RW, Humphreys E,Han M, Partin AW, Vessella RL, Isaacs WB, Bova GS, Luo J.Ligand-independent androgen receptor variants derived fromsplicing of cryptic exons signify hormone-refractory prostatecancer. Cancer Res 2009;69(1):16–22.
5. Culig Z, Hobisch A, Cronauer MV, Radmayr C, Trapman J,Hittmair A, Bartsch G, Klocker H. Androgen receptor activationin prostatic tumor cell lines by insulin-like growth factor-I,keratinocyte growth factor, and epidermal growth factor. CancerRes 1994;54(20):5474–5478.
6. Craft N, Shostak Y, Carey M, Sawyers CL. A mechanism forhormone-independent prostate cancer through modulation ofandrogen receptor signaling by the HER-2/neu tyrosine kinase.Nat Med 1999;5(3):280–285.
7. Debes JD, Schmidt LJ, Huang H, Tindall DJ. p300 mediatesandrogen-independent transactivation of the androgen receptorby interleukin 6. Cancer Res 2002;62(20):5632–5636.
8. Jin RJ, Wang Y, Masumori N, Ishii K, Tsukamoto T, Shappell SB,Hayward SW, Kasper S, Matusik RJ. NE-10 neuroendocrinecancer promotes the LNCaP xenograft growth in castrated mice.Cancer Res 2004;64(15):5489–5495.
9. McDonnell TJ, Troncoso P, Brisbay SM, Logothetis C, Chung LW,Hsieh JT, Tu SM, Campbell ML. Expression of the protooncogenebcl-2 in the prostate and its association with emergence ofandrogen-independent prostate cancer. Cancer Res 1992;52(24):6940–6944.
10. July LV, Akbari M, Zellweger T, Jones EC, Goldenberg SL,Gleave ME. Clusterin expression is significantly enhanced inprostate cancer cells following androgen withdrawal therapy.Prostate 2002;50(3):179–188.
11. Heidenberg HB, Bauer JJ, McLeod DG, Moul JW, Srivastava S.The role of the p53 tumor suppressor gene in prostate cancer: Apossible biomarker? Urology 1996;48(6):971–979.
12. Abate-Shen C, Banach-Petrosky WA, Sun X, Economides KD,Desai N, Gregg JP, Borowsky AD, Cardiff RD, Shen MM.Nkx3.1; Pten mutant mice develop invasive prostate adenocarci-noma and lymph node metastases. Cancer Res 2003;63(14):3886–3890.
13. Graff JR, Konicek BW, McNulty AM, Wang Z, Houck K, Allen S,Paul JD, Hbaiu A, Goode RG, Sandusky GE, Vessella RL,
Neubauer BL. Increased AKT activity contributes to prostatecancer progression by dramatically accelerating prostate tumorgrowth and diminishing p27Kip1 expression. J Biol Chem2000;275(32):24500–24505.
14. Cai C, Wang H, Xu Y, Chen S, Balk SP. Reactivation of androgenreceptor-regulated TMPRS S2: ERG gene expression in castra-tion-resistant prostate cancer. Cancer Res 2009;69(15):6027–6032.
15. Tomlins SA, Rhodes DR, Perner S, Dhanasekaran SM, Mehra R,Sun XW, Varambally S, Cao X, Tchinda J, Kuefer R, Lee C,Montie JE, Shah RB, Pienta KJ, Rubin MA, Chinnaiyan AM.Recurrent fusion of TMPRSS2 and ETS transcription factor genesin prostate cancer. Science 2005;310(5748):644–648.
16. Shi XB, Tepper CG, White RW. MicroRNAs and prostate cancer.J Cell Mol Med 2008;12(5A):1456–1465.
17. Sun T, Wang Q, Balk S, Brown M, Lee GS, Kantoff P. The role ofmicroRNA-221 and microRNA-222 in androgen-independentprostate cancer cell lines. Cancer Res 2009;69(8):3356–3363.
18. Corcoran NM, Costello AJ. Interleukin-6: Minor player orstarring role in the development of hormone-refractory prostatecancer? BJU Int 2003;91(6):545–553.
19. Lee LF, Louie MC, Desai SJ, Yang J, Chen HW, Evans CP, KungHJ. Interleukin-8 confers androgen-independent growth andmigration of LNCaP: Differential effects of tyrosine kinases Srcand FAK. Oncogene 2004;23(12):2197–2205.
20. Onishi RM, Gaffen SL. Interleukin-17 and its target genes:Mechanisms of interleukin-17 function in disease. Immunology2010;129(3):311–321.
21. Toy D, Kugler D, Wolfson M, Vanden Bos T, Gurgel J, Derry J,Tocker J, Peschon J. Cutting edge: Interleukin 17 signals througha heteromeric receptor complex. J Immunol 2006;177(1):36–39.
22. Haudenschild D, Moseley T, Rose L, Reddi AH. Soluble andtransmembrane isoforms of novel interleukin-17 receptor-likeprotein by RNA splicing and expression in prostate cancer. J BiolChem 2002;277(6):4309–4316.
23. Ely LK, Fischer S, Garcia KC. Structural basis of receptor sharingby interleukin 17 cytokines. Nat Immunol 2009;10(12):1245–1251.
24. Ye P, Rodriguez FH, Kanaly S, Stocking KL, Schurr J, Schwarzen-berger P, Oliver P, Huang W, Zhang P, Zhang J, Shellito JE,Bagby GJ, Nelson S, Charrier K, Peschon JJ, Kolls JK. Require-ment of interleukin 17 receptor signaling for lung CXC chemo-kine and granulocyte colony-stimulating factor expression,neutrophil recruitment, and host defense. J Exp Med 2001;194(4):519–527.
25. Hu Y, Ota N, Peng I, Refino CJ, Danilenko DM, Caplazi P,Ouyang W. IL-17RC is required for IL-17A- and IL-17F-depen-dent signaling and the pathogenesis of experimental autoim-mune encephalomyelitis. J Immunol 2010;184(8):4307–4316.
26. You Z, Dong Y, Kong X, Zhang Y, Vessella RL, Melamed J.Differential expression of IL-17RC isoforms in androgen-depen-dent and androgen-independent prostate cancers. Neoplasia2007;9(6):464–470.
27. You Z, Shi XB, DuRaine G, Haudenschild D, Tepper CG, Lo SH,Gandour-Edwards R, de Vere White RW, Reddi AH. Interleukin-17 receptor-like gene is a novel antiapoptotic gene highlyexpressed in androgen-independent prostate cancer. Cancer Res2006;66(1):175–183.
28. Zhang Q, Liu S, Ge D, Xue Y, Xiong Z, Abdel-Mageed AB, MyersL, Hill SM, Rowan BG, Sartor O, Melamed J, Chen Z, You Z.Interleukin-17 promotes formation and growth of prostateadenocarcinoma in mouse models. Cancer Res 2012;72(10):2589–2599.
10 Zhang et al.
The Prostate

29. Wang S, Gao J, Lei Q, Rozengurt N, Pritchard C, Jiao J, ThomasGV, Li G, Roy-Burman P, Nelson PS, Liu X. Wu H. Prostate-specific deletion of the murine Pten tumor suppressor gene leadsto metastatic prostate cancer. Cancer Cell 2003;4(3):209–221.
30. Shappell SB, Thomas GV, Roberts RL, Herbert R, Ittmann MM,Rubin MA, Humphrey PA, Sundberg JP, Rozengurt N, Barrios R,Ward JM, Cardiff RD. Prostate pathology of genetically engi-neered mice: Definitions and classification. The consensus reportfrom the Bar Harbor meeting of the Mouse Models of HumanCancer Consortium Prostate Pathology Committee. Cancer Res2004;64(6):2270–2305.
31. Didier DK, Schiffenbauer J, Woulfe SL, Zacheis M, Schwartz BD.Characterization of the cDNA encoding a protein binding to themajor histocompatibility complex class II Y box. Proc Natl AcadSci USA 1988;85(19):7322–7326.
32. Lovett DH, Cheng S, Cape L, Pollock AS, Mertens PR. YB-1alters MT1-MMP trafficking and stimulates MCF-7 breast tumorinvasion and metastasis. Biochem Biophys Res Commun 2010;398(3):482–488.
33. Gimenez-Bonafe P, Fedoruk MN, Whitmore TG, Akbari M,Ralph JL, Ettinger S, Gleave ME, Nelson CC. YB-1 is upregulatedduring prostate cancer tumor progression and increases P-glycoprotein activity. Prostate 2004;59(3):337–349.
34. Toh Y, Pencil SD, Nicolson GL. A novel candidate metastasis-associated gene, mta1, differentially expressed in highly metastaticmammary adenocarcinoma cell lines. cDNA cloning, expression,and protein analyses. J Biol Chem 1994;269(37):22958–22963.
35. Kai L, Wang J, Ivanovic M, Chung YT, Laskin WB, Schulze-Hoepfner F, Mirochnik Y, Satcher RL Jr, Levenson AS. Targetingprostate cancer angiogenesis through metastasis-associated pro-tein 1 (MTA1). Prostate 2011;71(3):268–280.
36. Townsley FM, Aristarkhov A, Beck S, Hershko A, Ruderman JV.Dominant-negative cyclin-selective ubiquitin carrier protein E2-C/UbcH10 blocks cells in metaphase. Proc Natl Acad Sci USA1997;94(6):2362–2367.
37. Hao Z, Zhang H, Cowell J. Ubiquitin-conjugating enzymeUBE2C: Molecular biology, role in tumorigenesis, and potentialas a biomarker. Tumour Biol 2012;33(3):723–730.
38. Wang Q, Li W, Zhang Y, Yuan X, Xu K, Yu J, Chen Z, BeroukhimR, Wang H, Lupien M, Wu T, Regan MM, Meyer CA, Carroll JS,Manrai AK, Janne OA, Balk SP, Mehra R, Han B, ChinnaiyanAM, Rubin MA, True L, Fiorentino M, Fiore C, Loda M, KantoffPW, Liu XS, Brown M. Androgen receptor regulates a distincttranscription program in androgen-independent prostate cancer.Cell 2009;138(2):245–256.
39. Numasaki M, Fukushi J, Ono M, Narula SK, Zavodny PJ, KudoT, Robbins PD, Tahara H, Lotze MT. Interleukin-17 promotesangiogenesis and tumor growth. Blood 2003;101(7):2620–2627.
40. Numasaki M, Watanabe M, Suzuki T, Takahashi H, NakamuraA, McAllister F, Hishinuma T, Goto J, Lotze MT, Kolls JK, SasakiH. IL-17 enhances the net angiogenic activity and in vivo growthof human non-small cell lung cancer in SCID mice throughpromoting CXCR-2-dependent angiogenesis. J Immunol 2005;175(9):6177–6189.
41. Kanda N, Koike S, Watanabe S. IL-17 suppresses TNF-alpha-induced CCL27 production through induction of COX-2 inhuman keratinocytes. J Allergy Clin Immunol 2005;116(5):1144–1150.
42. Miura S, Tatsuguchi A, Wada K, Takeyama H, Shinji Y,Hiratsuka T, Futagami S, Miyake K, Gudis K, Mizokami Y,Matsuoka T, Sakamoto C. Cyclooxygenase-2-regulated vascularendothelial growth factor release in gastric fibroblasts. Am JPhysiol Gastrointest Liver Physiol 2004;287(2):G444–G451.
43. He D, Li H, Yusuf N, Elmets CA, Li J, Mountz JD. Xu H. IL-17promotes tumor development through the induction of tumorpromoting microenvironments at tumor sites and myeloid-derived suppressor cells. J Immunol 2010;184(5):2281–2288.
44. Youn JI, Nagaraj S, Collazo M, Gabrilovich DI. Subsets ofmyeloid-derived suppressor cells in tumor-bearing mice. JImmunol 2008;181(8):5791–5802.
45. Redente EF, Dwyer-Nield LD, Merrick DT, Raina K, Agarwal R,Pao W, Rice PL, Shroyer KR, Malkinson AM. Tumor progressionstage and anatomical site regulate tumor-associated macrophageand bone marrow-derived monocyte polarization. Am J Pathol2010;176(6):2972–2985.
46. Hanahan D, Weinberg RA. Hallmarks of cancer: The nextgeneration. Cell 2011;144(5):646–674.
47. Mercader M, Bodner BK, Moser MT, Kwon PS, Park ES,Manecke RG, Ellis TM, Wojcik EM, Yang D, Flanigan RC, WatersWB, Kast WM, Kwon ED. T cell infiltration of the prostateinduced by androgen withdrawal in patients with prostatecancer. Proc Natl Acad Sci USA 2001;98(25):14565–14570.
48. Morse MD, McNeel DG. T cells localized to the androgen-deprived prostate are T(H) 1 and T(H) 17 biased. Prostate2012;72(11):1239–1247.
49. Liu L, Ge D, Ma L, Mei J, Liu S, Zhang Q, Ren F, Liao H, Pu Q,Wang T, You Z. Interleukin-17 and prostaglandin E2 areinvolved in formation of an M2 macrophage-dominant microen-vironment in lung cancer. J Thorac Oncol 2012;7(7):1091–1100.
Supporting Information
Additional supporting information may be found inthe online version of this article at the publisher ’s web-site.
IL-17 Promotes CRPC 11
The Prostate





























