Embed Size (px)
Citation preview

www.elsevier.com/locate/yclim
Clinical Immunology
Interleukin-7 improves reconstitution of antiviral CD4 T cells$
Hailing Lu, Zhao Zhao, Tomas Kalina, Thurman Gillespy III, Denny Liggitt, Robert G. Andrews,
David G. Maloney, Hans-Peter Kiem, Jan Storek1
Fred Hutchinson Cancer Research Center and University of Washington, Seattle, WA, United States
Received 11 June 2004; accepted with revision 10 August 2004
Abstract
We evaluated whether long-term (2 months) administration of interleukin-7 (IL7) hastens immune recovery in baboons rendered severely
lymphopenic by total body irradiation and antithymocyte globulin (ATG). Four baboons were treated with recombinant baboon IL7 and three
baboons with placebo. Median CD4 T cell count at the end of IL7/placebo treatment was higher in the IL7-treated animals (2262 vs. 618/Al,P = 0.03). This appeared to be a result of peripheral expansion rather than de novo generation. Median cytomegalovirus (CMV)-specific
IFNg-producing CD4 T cell count at the end of IL7/placebo treatment was higher in the IL7-treated animals (122 vs. 1/Al, P = 0.03). All
animals were pretransplant cytomegalovirus-seropositive. One animal died at the end of IL7 treatment; necropsy showed extensive T cell
infiltration of kidneys and lungs. In conclusion, IL7 stimulates the expansion of CD4 T cells, including functional antiviral cells. Clinical
risk–benefit ratio needs to be evaluated.
D 2004 Elsevier Inc. All rights reserved.
Keywords: T cell reconstitution; Interleukin-7; Hematopoietic cell transplantation; Antiviral immunity; Cytomegalovirus; T lymphocytes; Immunodeficiency
diseases; Viral diseases; Cytokines
Introduction
CD4 T-lymphopenia in hematopoietic cell transplant
recipients and HIV-infected patients is associated with
infections [1–3], including cytomegalovirus (CMV) infec-
tions [4,5]. IL7 has been shown to increase CD4 T cell
counts in both murine and nonhuman primate models [6–
11]. Both de novo generation of T cells (from hematopoietic
cells, typically in the thymus) and peripheral expansion of
existing T cells were reported to be stimulated by IL7 in
mice [9]. However, only peripheral expansion was stimu-
lated in the two primate studies reported, that is, Fry et al.’s
[10] study using normal and SIV-infected macaques and our
study using irradiated baboons [11]. It is possible that de
1521-6616/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.clim.2004.08.008
$ Supported by National Institutes of Health Grants No. HL68710 and
CA18221.1 Present address: University of Calgary, Health Science Centre, 3330
Hospital Drive NW, Calgary, Alberta, Canada T2N 4N1. Fax: +1 403 283
1267.
E-mail address: [email protected] (J. Storek).
novo generation was not observed in the two primate studies
because the treatment was too short (10 days in Fry et al.’s
study and 28 days in our study), because xenogeneic IL7
was used or because only moderately lymphopenic monkeys
were treated.
The present study was thus performed for the following
reasons: (1) to study the effect of IL7 in severely
lymphopenic monkeys, as stimulation of de novo generation
by IL7 was not observed in T cell-replete mice [12]; (2) to
administer baboon instead of human IL7, as baboon–
antihuman IL7 antibodies [11] or, less likely, relative
incompatibility between human IL7 and baboon IL7
receptor could diminish the IL7 effect; (3) to treat the
animals for a longer period (8 weeks) than in the previous
studies; as thymocyte precursor to T cell differentiation
takes at least 12 days [13], only prolonged treatment might
result in a detectable rise of recent thymic emigrants,
represented by T cell receptor excision circle-containing
(TREC+) T cells; (4) to add data on potential side effects of
IL7 in primates, as clinical trials of IL7 in patients with
iatrogenic or HIV-induced lymphopenia are being planned;
114 (2005) 30–41

H. Lu et al. / Clinical Immunology 114 (2005) 30–41 31
(5) most important, to determine whether the IL7-stimulated
increase in the total CD4 T cell counts might be functionally
relevant. To evaluate this, we treated CMV-seropositive
baboons and asked whether higher anti-CMV CD4 T cell
counts were achieved in the IL7 than placebo-treated
animals. Reconstitution of anti-CMV CD4 T cells is
essential for the reconstitution of anti-CMV CD8 T cells
(reviewed by Gratama and Cornelissen [14]), and increased
numbers of CMV-specific CD4 T cells have been associated
with decreased incidence of symptomatic CMV disease
[15].
Materials and methods
Study design and animals
Seven male baboons (Papio cynocephalus anubis), age 2
years (equivalent to 5–7 human years [16–18]), underwent
total body irradiation (TBI, 10.2 Gy) followed by autolo-
gous transplantation of CD34 cells marked with yellow
fluorescent protein (YFP) gene. Before starting IL7/placebo
treatment, T cells were further depleted by antithymocyte
globulin (ATG, thymoglobulin, 5 mg/kg intravenously on
days 32, 34, 36, and 39 posttransplant, kindly donated by
Sangstat, Fremont, CA). Importantly, in vivo ATG causes
apoptosis of circulating splenic and lymphonodal T cells but
does not affect thymocytes, probably because of limited
access of ATG to the thymus [19]. Three baboons received
placebo, and four baboons received recombinant baboon
IL7 (kindly donated by Cytheris, Vanves, France) 80 Ag/kgonce a day subcutaneously from days 42 to 98 after
transplantation. In the Results, day 42 is referred to as
bimmediately pre-IL7/placebo treatmentQ and day 98 as
bweek 8 of treatment.Q Lyophilized recombinant baboon IL7
was reconstituted with a diluent containing 4 mg/ml sucrose
and 125 mM sodium citrate, pH 6.2. The same diluent was
used as placebo. Pretransplant, the animals were sero-
positive for CMV, seronegative for Babesia and STLV-I, and
tuberculin test-negative. They were housed and cared for at
the University of Washington Regional Primate Research
Center under conditions approved by the American Asso-
ciation for the Accreditation of Laboratory Animal Care.
The study was approved by the UW Animal Care and Use
Committee and the FHCRC Institutional Review Board.
Transplantation and posttransplant treatment
Filgrastim (G-CSF), 100 Ag/kg daily, and stem cell factor
(SCF), 50 Ag/kg daily, were given subcutaneously from days
8 to 4 to increase the number of CD34 cells in the marrow.
On day 3, bone marrow was harvested into preservative-free
heparin from both humeri and femora. CD34 cells were
immunomagnetically enriched to approximately 80% purity,
using antibody 12.8 and goat antimouse IgM-coated
microbeads (Miltenyi, Auburn, CA). Transduction of
CD34 cells with an oncoretrovirus carrying the enhanced
YFP gene was done as described [20]. Approximately 50%
transduced and 50% untransduced CD34 cells were infused.
(The utility of transplanting the YFP gene-transduced CD34
cells was limited, as sustained engraftment of the transduced
cells was achieved in only four of the seven animals.) After
total body irradiation (10.2 Gy in two fractions at 0.07 Gy/
min), the cells were reinfused on day 0. Supportive care
included irradiated whole blood transfusions for thrombo-
cytopenia and prophylactic antibiotics (acyclovir and fluco-
nazole from days 0 to 112, ceftazidime + gentamycin +
vancomycin from days 0 to 41 and levofloxacin from days
41 to 112 posttransplant).
Enumeration of blood mononuclear cell subsets
Mononuclear cells (MNCs) were separated from EDTA-
anticoagulated blood using density gradient (Ficoll, density
1.073 kg/l) centrifugation. The cells, suspended in flow
buffer (PBS with 1% bovine serum albumin and 0.1%
sodium azide), were stained with fluorochrome-conjugated
monoclonal antibodies and analyzed by five-color flow
cytometry. The following antibodies were used: CD3 from
Biosource (clone FN18), CD4 from BD Biosciences (clone
SK3), CD8 from Beckman-Coulter (clone B9.11), CD11a
from Beckman-Coulter (clone 25.3.1), CD14 from Beck-
man-Coulter (clone RMO52), CD16 from Beckman-Coulter
(clone 3G8), CD20 from BD Biosciences (clone L27),
CD45RA from Beckman-Coulter (clone ALB11), and CD56
from Beckman-Coulter (clone N901). Residual red blood
cells were lysed in hemolytic buffer (8.3% ammonium
chloride + 1% potassium bicarbonate + 0.4% EDTA). No
fixation was done. Cells were resuspended in flow buffer
and kept on ice until flow cytometry, which occurred within
12 h from blood drawing. FACSCalibur (BD Biosciences)
flow cytometer was used for data acquisition. List-mode
data were analyzed using WinList software (Verity Software
House, Topsham, ME). The FSc � SSc gate was set to
encompass MNCs. B cells were defined as CD20+ MNCs.
CD4 T cells were defined as CD3+CD4+ MNCs, and CD8 T
cells were defined as CD3+CD8+ MNCs. NK cells were
defined as MNCs expressing CD16 or CD56 and not
expressing CD3 or CD14. Monocytes were defined as
CD14+ MNCs. Each absolute MNC subset count was
calculated as the absolute MNC count multiplied by the
percent of the MNC subset divided by 100. The absolute
MNC count represented the sum of absolute lymphocyte
count and absolute monocyte count determined by our
clinical hematology laboratory, using Sysmex XE2100 cell
counter and Giemsa-stained blood smears.
Lymph nodes (LN) were obtained immediately before
starting IL7 (right axilla) at week 8 of treatment (left axilla)
and approximately 3 months later (right axilla). Axillary
LNs were chosen as they were distant from the IL7/placebo
injection sites (thighs). The LNs were teased in RPMI with
10% fetal calf serum. The resulting cell suspension was

H. Lu et al. / Clinical Immunology 114 (2005) 30–4132
filtered through a 70-Am nylon cell strainer. Percentages of
cell subsets were determined by flow cytometry as in the
blood.
Thymic biopsy was done in one placebo-treated and two
IL7-treated animals at week 8 of treatment, using upper
sternal split approach under general anesthesia. Approx-
imately 3000 mm3 of thymic tissue was removed and
processed for flow cytometry as for lymph nodes and for
histology as described below.
CMV serology
The presence of antibodies against baboon CMV was
tested by Simian Diagnostic Laboratory (San Antonio, TX)
using rapid dot-immunobinding assay [21]. In brief, virus or
control antigens were spotted on the nitrocellulose sheet and
allowed to absorb for 30 min at room temperature. Then the
nitrocellulose sheet was submerged in 5% nonfat milk in
PBS to block nonspecific binding for 1 h followed by
washing with PBS-Tween (0.1%). After test sera were
applied over the test antigen dots, the nitrocellulose sheet
was incubated for 1 h at 378C and then washed three times
in PBS-Tween. Then staphylococcal protein A-conjugated
horseradish peroxidase was used to detect primate IgG.
Following incubation and washing, the substrate, 4-chloro-
1-naphthol-hydrogen peroxide, was used, and a blue spot
was developed as a positive result. Controls consisted of
uninfected cell culture antigens and known positive and
negative sera.
Intracellular cytokine staining for detection of CMV-specific
CD4 T cells
We used CMV lysate to stimulate baboon MNCs. CMV
lysate was prepared using MRC5 human lung fibroblast cell
line (ATCC CCL-171) infected with baboon CMV, strain
PGH-01 (ATCC VR-1530) or OCOM4-37 (kindly provided
by Dr. Earl Blewett, Oklahoma State University). After
infection had progressed to N90% cytopathic effect, the cells
were harvested and subjected to multiple freeze–thaw
cycles. Then the cells were washed twice in glycine-
buffered saline, and the pellets were sonicated on ice. The
resulting suspension of cell organelles/particles was centri-
fuged at 2000 rpm for 10 min, and the supernatant was
harvested. The supernatants from both CMV strains were
mixed and heat-inactivated at 568C for 30 min. Protein
concentration was determined with a BCA protein assay kit
(Pierce, Rockford, IL). Aliquots of the CMV lysate were
stored in �808C. Control lysate with the same protein
concentration was prepared from uninfected MRC5 cells.
One million PBMC in 1 ml of RPMI 1640 supplemented
with 2 mM glutamine, 25 mM HEPES, 10% fetal bovine
serum (Hyclone, Logan, UT) were incubated with CMV
lysate (5 Ag protein/ml), uninfected MRC5 lysate (5 Agprotein/ml, negative control), or staphylococcal enterotoxin
B (1 Ag/ml, Sigma, positive control) in the presence of anti-
CD28 and anti-CD49d antibodies (0.5 Ag/ml each, BD
Biosciences, San Jose, CA) overnight at 378C in a
humidified atmosphere. Brefeldin A (10 Ag/ml; Sigma-
Aldrich, St. Louis, MO) was added 2 h after the initiation of
the culture. After the overnight incubation, cells were
treated with 1 mM EDTA for 10 min at RT. Then the cells
were sequentially fixed and permeabilized using FACS
lysing solution and FACS permeabilizing solution (BD
Biosciences). The cells were then stained with anti-IFNg-
phycoerythrin (clone 45.15, Beckman Coulter [Immuno-
tech], Miami, FL) or anti-TNFa-phycoerythrin (clone
Mab11, BD Biosciences), anti-CD3-PerCPCy5.5 (clone
SK34-2, BD Biosciences), anti-CD8 APCCy7 (clone SK1,
BD Biosciences), and anti-CD4-APC (clone SK3, BD
Biosciences) for 30 min at room temperature in the dark.
After final wash with flow buffer, data acquisition was done
on LSR II cytometer (BD Biosciences). At least 200,000
events were acquired. Data were analyzed using FlowJo
software (Treestar, San Carlos, CA).
Detection of CMV by PCR
DNA was extracted from 200 Al serum using QIAamp
DNA Blood Mini Kit (Qiagen, Valencia, CA). The
following primers were used: 5V-TACGTCATTGG-
TACCCTCC-3V(forward) and 5V-GGAGTACTGCCAATG-TACTA-3V(reverse) [22]. Each reaction contained 2.5 U
platinum-Taq polymerase (Invitrogen, Carlsbad, CA), 2.5
mM MgCl2, 500 uM dNTPs, 500 nM each primer, 20 Alextracted DNA (of total 100 Al yielded by the QIAamp
kit) in a total volume of 50 Al. The conditions of each
reaction were 1 cycle of 958C for 9 min, followed by 40
cycles of 948C/60 s; 588C/60 s; 728C/60 s, and finally 1
cycle of 728C for 9 min. PCR products were separated
on 2% agarose gel and visualized by ethidium bromide
staining. As a positive control, we used DNA extracted
from the CMV lysate (derived from baboon CMV-
infected MRC5 cells). As negative control, we used
DNA extracted from baboon lymphocryptovirus (Epstein-
Barr virus-like virus)-transformed baboon B lymphoblas-
toid cells.
T cell receptor excision circle (TREC) assay
FACS-sorted CD4 T cell pellets of 200,000 cells were
lysed in 40 Al of lysis buffer containing 100 Ag/ml
proteinase K and 10 mM Tris–HCl (pH 8.0). The 5V-nuclease (Taqman) assay for the baboon ay signal joint
TREC [11] was performed on 5 Al of cell lysate using
forward primer, 5VCACATCCCTTTCAACCATGCT3V,reverse primer, 5VGCCAGCTGCAGGGTTTAGG3V, and
probe FAM-ACGCATTTGGTTTTTGTAAAGGTGCT-
CACT-QSY7 (MegaBases, Chicago, IL). PCR reactions
contained 0.5 U Taq polymerase, 3.5 mM MgCl2, 0.2 mM
dNTPs, 400 nM each primer, 200 nM probe, and Blue-636
reference (MegaBases). The reactions were run at 958C for

H. Lu et al. / Clinical Immunology 114 (2005) 30–41 33
5 min, then 958C/30 s, and 608C/1 min for 40 cycles, using
ABI7700 system (PE Biosystems, Norwalk, CT). Samples
were analyzed in quadruplicates. Plasmids containing the
baboon signal joint region were used as standards. A
standard curve was plotted, and the TREC level (the number
of TREC copies in 5 Al of cell lysate, which corresponds to
25,000 cells) was calculated using the ABI7700 software.
The absolute count of TREC+ CD4 (CD8) T cells (per
microliter) was calculated as the TREC level (per 25,000
cells) multiplied by the absolute CD4 T cell count (per
microliter) and divided by 25,000.
Computed tomography (CT)
Volumes of the thymus, the spleen, and the lymph
nodes and the density of the cancellous bone of L3, L4,
and L5 vertebrae were measured as described [11]. The
calculated density of the torso phantom ranged from 94.6
to 96.7 mg/cm3, suggesting adequate reproducibility. The
volume of the kidneys was determined analogously to the
thymus. Briefly, the area of kidneys (mm2) was determined
on each slice, using IV contrast images and GE Pathspeed
software (GE Medical Systems). The area was multiplied
by the slice thickness (1.25 mm) to get the volume at that
slice level (mm3). Kidney volume (of both kidneys
together) was calculated as the sum of the volumes at
each slice level. Increase of volume of an organ was
calculated as the volume at the end (week 8) of IL7/
placebo treatment minus the volume immediately pre-IL7/
placebo treatment.
Histology
Using standard techniques, 10% neutral buffered for-
malin-fixed, paraffin-embedded, and in the case of bone
marrow, decalcified tissue sections were stained with
hematoxylin and eosin (H and E) or immunostained with
CD3 (rabbit–antihuman CD3 polyclonal antibody, Dako,
Carpinteria, CA) or CD20 (clone L26, Dako). OTC-
embedded snap-frozen tissues were immunostained with
CD4 (clone 1F6, Novocastra, Newcastle upon Tyne, UK) or
CD8 (clone G10.1, kind gift of Dr. Bing Hu, University of
Washington). Slides were analyzed by one pathologist
(D.L.) blinded to IL7 versus placebo treatment. Bone
marrow cellularity was estimated from the H and E-stained
sections and expressed as the percentage of nonfat cells
among fat plus nonfat cells. Percentage of T or B cells
among bone marrow hematopoietic cells was estimated
from the immunostained sections.
IL7 levels and IL7 neutralizing antibodies
Serum concentration of IL7 was determined by ELISA
using the kit for human IL7 (R and D Systems, Minneapolis,
MN). In case of values exceeding the concentration of the
highest standard, diluted serum was used.
IL7 neutralizing antibodies were determined by the
inhibition of the growth of IL7-dependent cells (PB1) as
described [11].
Statistics
To compare the counts of CD4 T cells or their subsets at
the end of IL7/placebo treatment for the animals that
survived until the week 8 evaluation, we used the average
count from weeks 7 and 8 of the IL7/placebo treatment. For
the animal that died 1 day before the week 8 evaluation
(Z00158), we used the count from week 7. Significance of
differences was tested by the Mann-Whitney–Wilcoxon
rank sum test. As the hypothesis was that the cell counts
were higher in the IL7-treated animals, one-tailed P value
was used.
Results
IL7 treatment improved the reconstitution of CD4 T cells but
not CD8 T cells
After total body irradiation and further T cell depletion
by ATG, the baboons were severely lymphopenic.
Immediately before starting IL7/placebo treatment, median
CD4 T cell count in the blood was 3/Al, and lymph nodes
were severely lymphocyte-depleted (composed of mostly
histiocytes). At the end of IL7/placebo treatment (weeks
7–8), median CD4 T cell count in the blood was higher in
the IL7-treated animals (2262 vs. 618/Al, P = 0.03) (Fig.
1). The median number of weeks it took to reach
individual pretransplant CD4 T cell count was 2.5 weeks
for IL7-treated versus 8 weeks for placebo-treated animals
(P = 0.03). The improved CD4 T cell reconstitution
appeared to be primarily, if not entirely, due to the
stimulation of peripheral expansion, as the count of
TREC+ CD4 T cells (per microliter of blood which
reflects thymic output irrespective of peripheral expansion
[not the percent of TREC+ cells among CD4 T cells which
is influenced by peripheral expansion]) was not signifi-
cantly higher in IL7 than placebo-treated animals (Fig. 1).
Both CD45RAhigh and CD45RAlow/� CD4 T cell counts
were significantly higher at the end of treatment (weeks
7–8) in IL7 than placebo-treated animals (P = 0.03) (Fig.
1). This, together with the fact that TREC+ CD4 T cell
counts were not significantly higher in the IL7-treated
animals, suggests that IL7 induced expansion of both
naive as well as memory/effector cells. The naive cells
expanded by IL7 likely included cells generated de novo
since the time of transplant; as in the four animals with
sustained YFP marking (two IL7- and two placebo-
treated), there was a trend toward higher YFP+ CD4 T
cell counts at the end of treatment (weeks 7–8) in the IL7-
than placebo-treated animals (median 294 vs. 68/Al, datanot shown).

Fig. 1. Counts per microliter blood (top and center) or percentages (bottom) of CD4 T cells or their subsets in IL7-treated (black thin lines) and placebo-treated
(gray thick lines) animals. On each x-axis, bPre-TxQ denotes pretransplant, bPre-IL7Q denotes immediately pre-IL7/placebo treatment, and subsequent numbers
denote weeks after starting IL7/placebo treatment. The period of IL7 treatment is indicated in each graph by the black horizontal bar. The TREC+CD4Tcell counts
could not be reliably enumerated immediately before and in the first 3 weeks of IL7/placebo treatment due to low total CD4 counts; b0.01Q is arbitrarily displayedfor the immediate pretreatment time point. BMdenotes bonemarrow, and LN denotes lymph nodes. In both organs, the percentages are among total nucleated cells
and were determined by flow cytometry. For a clearer interpretation of this figure, the reader is referred to the figure in the Web version of this article.
H. Lu et al. / Clinical Immunology 114 (2005) 30–4134
The increased blood CD4 T cell counts in IL7-treated
animals were not due to redistribution of CD4 T cells from
tissues to blood; as in the bone marrow of IL7-versus
placebo-treated animals, there was a trend toward a higher
percentage of CD4 T cells (Fig. 1) and a higher overall
marrow cellularity (median 62.5% vs. 55% nonfat cells
among fat plus nonfat cells at week 4 and 72.5% vs. 40% at
week 8). In the lymph nodes at week 8, the percentage of
CD4 T cells was similar (Fig. 1), while there was a trend
toward larger lymph node volume in IL7- than placebo-
treated animals (Fig. 2). These results suggest that IL7
treatment increased the number of total body CD4 T cells.
The conclusion that IL7 stimulated peripheral expansion
rather than de novo generation was further supported by the
determination of the volumes of thymi, spleens, and lymph
nodes (Fig. 2). There was a trend toward higher median
increase of splenic volume (30,063 vs. 13,914 mm3) and
lymph node volume (1150 vs. 714 mm3) but not thymic
volume (191 vs. 2682 mm3) from the start to the end of IL7
versus placebo treatment. This was consistent with the
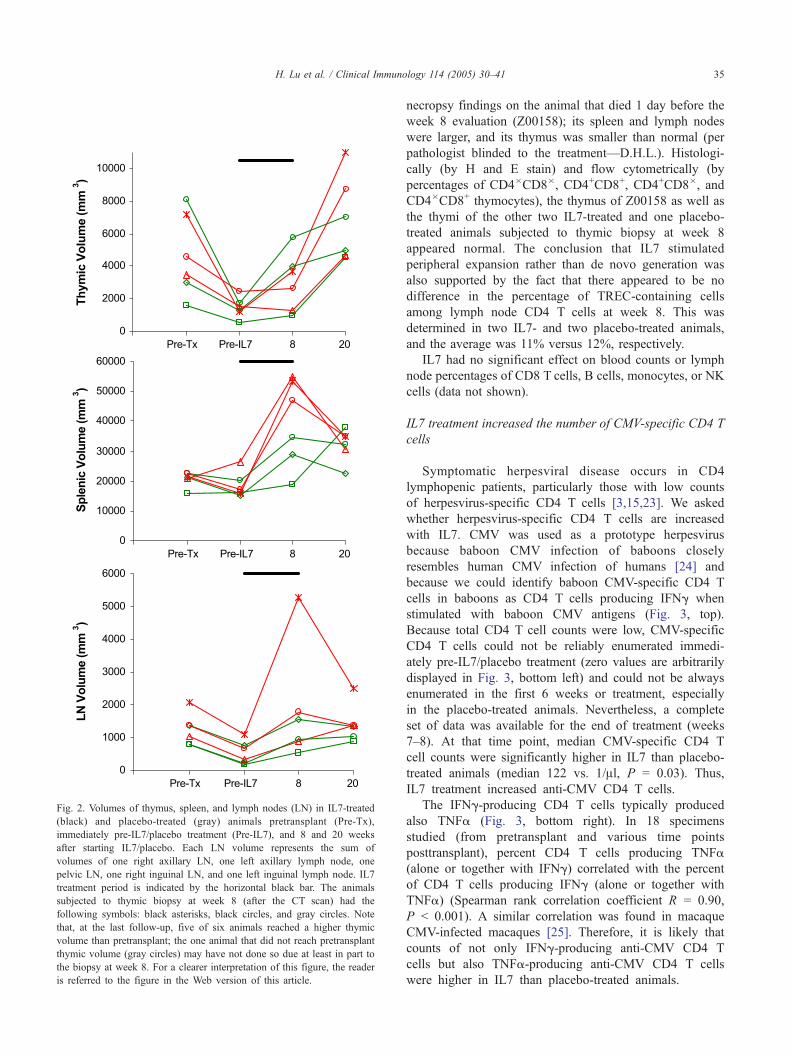
Fig. 2. Volumes of thymus, spleen, and lymph nodes (LN) in IL7-treated
(black) and placebo-treated (gray) animals pretransplant (Pre-Tx),
immediately pre-IL7/placebo treatment (Pre-IL7), and 8 and 20 weeks
after starting IL7/placebo. Each LN volume represents the sum of
volumes of one right axillary LN, one left axillary lymph node, one
pelvic LN, one right inguinal LN, and one left inguinal lymph node. IL7
treatment period is indicated by the horizontal black bar. The animals
subjected to thymic biopsy at week 8 (after the CT scan) had the
following symbols: black asterisks, black circles, and gray circles. Note
that, at the last follow-up, five of six animals reached a higher thymic
volume than pretransplant; the one animal that did not reach pretransplant
thymic volume (gray circles) may have not done so due at least in part to
the biopsy at week 8. For a clearer interpretation of this figure, the reader
is referred to the figure in the Web version of this article.
H. Lu et al. / Clinical Immunology 114 (2005) 30–41 35
necropsy findings on the animal that died 1 day before the
week 8 evaluation (Z00158); its spleen and lymph nodes
were larger, and its thymus was smaller than normal (per
pathologist blinded to the treatment—D.H.L.). Histologi-
cally (by H and E stain) and flow cytometrically (by
percentages of CD4�CD8�, CD4+CD8+, CD4+CD8�, and
CD4�CD8+ thymocytes), the thymus of Z00158 as well as
the thymi of the other two IL7-treated and one placebo-
treated animals subjected to thymic biopsy at week 8
appeared normal. The conclusion that IL7 stimulated
peripheral expansion rather than de novo generation was
also supported by the fact that there appeared to be no
difference in the percentage of TREC-containing cells
among lymph node CD4 T cells at week 8. This was
determined in two IL7- and two placebo-treated animals,
and the average was 11% versus 12%, respectively.
IL7 had no significant effect on blood counts or lymph
node percentages of CD8 T cells, B cells, monocytes, or NK
cells (data not shown).
IL7 treatment increased the number of CMV-specific CD4 T
cells
Symptomatic herpesviral disease occurs in CD4
lymphopenic patients, particularly those with low counts
of herpesvirus-specific CD4 T cells [3,15,23]. We asked
whether herpesvirus-specific CD4 T cells are increased
with IL7. CMV was used as a prototype herpesvirus
because baboon CMV infection of baboons closely
resembles human CMV infection of humans [24] and
because we could identify baboon CMV-specific CD4 T
cells in baboons as CD4 T cells producing IFNg when
stimulated with baboon CMV antigens (Fig. 3, top).
Because total CD4 T cell counts were low, CMV-specific
CD4 T cells could not be reliably enumerated immedi-
ately pre-IL7/placebo treatment (zero values are arbitrarily
displayed in Fig. 3, bottom left) and could not be always
enumerated in the first 6 weeks or treatment, especially
in the placebo-treated animals. Nevertheless, a complete
set of data was available for the end of treatment (weeks
7–8). At that time point, median CMV-specific CD4 T
cell counts were significantly higher in IL7 than placebo-
treated animals (median 122 vs. 1/Al, P = 0.03). Thus,
IL7 treatment increased anti-CMV CD4 T cells.
The IFNg-producing CD4 T cells typically produced
also TNFa (Fig. 3, bottom right). In 18 specimens
studied (from pretransplant and various time points
posttransplant), percent CD4 T cells producing TNFa
(alone or together with IFNg) correlated with the percent
of CD4 T cells producing IFNg (alone or together with
TNFa) (Spearman rank correlation coefficient R = 0.90,
P b 0.001). A similar correlation was found in macaque
CMV-infected macaques [25]. Therefore, it is likely that
counts of not only IFNg-producing anti-CMV CD4 T
cells but also TNFa-producing anti-CMV CD4 T cells
were higher in IL7 than placebo-treated animals.

Fig. 3. IL7 treatment increased CMV-specific CD4 T cells. At the top, examples of CD4 versus IFNg dotplots from the end of treatment (week 7 or 8) are
displayed, showing the trend toward higher percentages of CMV-specific IFNg-producing CD4 T cells (among total CD4 T cells) in IL7- than placebo-
treated animals. The bNormalQ blood was obtained pretransplant. Blood MNCs were stimulated with CMV lysate prepared from infected MRC5 cells and
processed as described in the Materials and methods. Stimulation with uninfected MRC5 lysate resulted in less than 0.05% IFNg-producing CD4 cells (not
shown); this was subtracted when calculating the absolute number of CMV-specific CD4 T cells displayed below. In each dotplot, the gate is set on CD4
lymphocytes, and 10,000 events are displayed. At the bottom left, the absolute numbers of CMV-specific CD4 T cells in IL7 (black)- and placebo (gray)-treated
baboons are shown. IL7 treatment period is indicated by the horizontal black bar. CMV-specific cells could not be reliably enumerated immediately pre-IL7/
placebo treatment; b0Q is arbitrarily displayed for each animal. At the bottom right, a typical TNFa versus IFNg dotplot gated on CD4 lymphocytes is
displayed, showing that most IFNg CD4 T cells produced also TNFa. For a clearer interpretation of this figure, the reader is referred to the figure in the Web
version of this article.
H. Lu et al. / Clinical Immunology 114 (2005) 30–4136
After transplantation in humans, recovery of CMV-
specific T cells is boosted by CMV reactivation (detection
of CMV in posttransplant blood using a technique that
usually does not detect CMV in blood from normal
seropositive individuals, for example, pp65 immunostain-
ing) [26–28]. In our experiment, CMV reactivation could
have occurred in the IL7-treated and not in the placebo-
treated baboons. If true, the higher counts of CMV-specific
CD4 T cells in the IL7-treated baboons could be due to
CMV reactivation and not necessarily due to IL7. Contrary
to that, reactivation of CMV during IL7/placebo treatment
was documented in all (both IL7- and placebo-treated)
baboons (Fig. 4). Therefore, the increased number of CMV-
specific CD4 T cells in the IL7-treated animals was likely
due to IL7.
Potential toxicity of IL7
One animal (Z00158) died unexpectedly at the end of
IL7 administration (1 day before the scheduled week 8

Fig. 5. Extensive lung (top) and kidney (center) infiltration with CD3+ cells
(brown). This was found on the necropsy of animal Z00158 that died 1 day
before week 8 evaluation. In the lungs, the T cells were localized primarily
around blood vessels (asterisks), but also around bronchioles and in the
parenchyma. In the kidneys, the T cells were localized also primarily
around blood vessels (asterisks), but also around tubules and glomeruli.
Among the surviving animals, kidney enlargement (bilateral) at week 8 of
treatment was found in the IL7-treated but not the placebo-treated animals
(bottom). The kidney volume is a composite volume of right and left
kidneys. Image (top and center) information: Nikon E-400 microscope,
Nikon 10� lens with 0.25 numerical aperture, digital imaging medium,
Nikon Coolpix 5000 camera with Nikon software, Microsoft Photo Editor
(for adjustment of general brightness and contrast only—no other
operations performed).
Fig. 4. Reactivation of CMVoccurred in all seven baboons (four IL7-treated
baboons, upper gels; and three placebo-treated baboons, lower gels). Viral
DNA was detected in serum by PCR immediately before IL7/placebo
treatment (bPre IL7Q) and at most time points during IL7/placebo treatment
(weeks 1–7) but not pretransplant (bPre TxQ). CMV lysate (derived from
baboon CMV-infected MRC5 cells) was used as the positive control (+) and
baboon Epstein-Barr virus-transformed baboon B lymphoblastoid cells
were used as the negative control (�). N/A indicates serum not available.
H. Lu et al. / Clinical Immunology 114 (2005) 30–41 37
evaluation). The only clinical symptom or sign noted was
anorexia in the last week of life, attributed to mild renal
insufficiency (serum creatinine rose from baseline 0.5–1.0
mg/dl to maximum 1.6 mg/dl). On necropsy, macroscopic
findings included large lymph nodes and spleen, small
thymus, marginally enlarged kidneys with pale, radial
streaks in the cortex, and firm and nonfloating lungs.
Microscopically, most organs were infiltrated with CD3+
cells. The infiltration was most prominent in the lungs and
the kidneys (Fig. 5, top). Approximately one half of the cells
infiltrating the kidneys and the lungs were CD4+ and one
half CD8+. No inclusions or syncytia were noted, suggesting
autoimmune rather than viral etiology. Of the total of nine
baboons we have treated with IL7, death associated with T
cell infiltration of multiple organs occurred in two animals
(A00066 reported in Ref. [11] and Z00158 reported here).
This complication may have resulted from overtreatment, as
we continued to give IL7 to animals per protocol even after
they have reached a high CD4 T cell count. The death
occurred in two of five animals that reached a CD4 T cell
count of greater than 2,500/Al and none of four animals that
did not reach CD4 T cell count of 2500/Al. CD8 T cell
counts and the deaths appeared to be unrelated.
Because of the renal dysfunction of Z00158 premortem
with the postmortem finding of T cell infiltration of the
kidneys reported here and because of a mild kidney
enlargement from immediately pre-IL7 to the end of 4-
week treatment noted on CT in all five animals treated
with IL7 in our previous experiment [11] (unpublished),
we retrospectively measured kidney volumes of the three
surviving IL7-treated and the three placebo-treated animals
in the present experiment (Fig. 5, bottom). Median
increase in the kidney volume was 31,155 mm3 in the
IL7-treated group compared to �4301 mm3 in the

Fig. 6. Hematocrit and platelet counts in IL7 (black)- and placebo (gray)-
treated baboons. The high hematocrit immediately pre-IL7/placebo treat-
ment is a result of overtransfusion (using whole blood for thrombocyto-
penia) in the first 6 weeks after transplant. The sudden increases in
hematocrit in the animal denoted by black triangles between 2 and 3 weeks
and in the animal denoted by gray squares between 3 and 4 weeks into IL7/
placebo treatment resulted from blood transfusions. The sudden drop in
hematocrit in the animal denoted by gray squares between weeks 14 and 15
resulted from babesiosis, and the subsequent rise in hematocrit coincided
with doxycycline treatment. As in Fig. 1, bPre-TxQ denotes pretransplant,
bPre-IL7Q denotes immediately pre-IL7/placebo treatment, and subsequent
numbers denote weeks after starting IL7/placebo treatment. The period of
IL7 treatment is indicated by the black horizontal bar. For a clearer
interpretation of this figure, the reader is referred to the figure in the Web
version of this article.
Fig. 7. Density of cancellous bone in IL7 (black)- and placebo (gray)-
treated baboons. Average density of the cancellous bone of vertebral bodies
L3, L4, and L5 showed no decrease during IL7 treatment. IL7 treatment
period is indicated by the horizontal black bar. For a clearer interpretation of
this figure, the reader is referred to the figure in the Web version of this
article.
H. Lu et al. / Clinical Immunology 114 (2005) 30–4138
placebo-treated group. Concurrent rise in serum creatinine
was not observed. The kidney volume returned close to
baseline within 3 months after discontinuation of IL7. We
speculate that the kidney enlargement was due to T cell
infiltration, although histology was only available for
Z00158.
Thrombocytopenia and possibly neutropenia and anemia
were associated with IL7 treatment in the previous experi-
ment. In this experiment, we observed a trend toward lower
hematocrit and platelet counts (Fig. 6) but not neutrophil
counts (data not shown) in IL7- than placebo-treated
animals. Perhaps, the hematologic toxicity in this experi-
ment was not as obvious as in the previous experiment
because one of the placebo-treated animals had an unusually
poor engraftment (gray squares in Fig. 6).
Osteoporosis due to stimulation of osteoclasts developed
after only 20 days of treatment with IL7 in mice. [29]. In our
previous baboon experiment, treatment with IL7 for 28 days
was not associated with decreased bone density. In the
present experiment, treatment for 56 days was also not
associated with decreased bone density (Fig. 7).
Lymphoma did not develop in the three IL7-treated
animals that did not die unexpectedly. This was determined
by both CTand necropsy at 3 months after discontinuation of
IL7.
IL7 levels and IL7 neutralizing antibodies
Median serum IL7 level (of all seven animals) rose from
6 pg/ml pretransplant to 27 pg/ml immediately pre-IL7/
placebo treatment, reflecting endogenous response to the
severe CD4 lymphopenia (Fig. 8). In the IL7-treated
animals, IL7 levels approximately 3 logs higher than at
baseline (pretransplant) were reached within 1 week and
were maintained throughout the remaining 7 weeks of
treatment. In the placebo-treated animals, IL7 levels
returned to baseline within approximately 1 month.
IL7 neutralizing antibodies may have theoretically been
generated in the IL7-treated animals, although the IL7 was
species-specific, as human cytokine neutralizing antibodies
have developed in humans treated with erythropoietin,
GM-CSF and thrombopoietin [30–32], and IL7 might
stimulate antibody production by stimulating helper T
cells. Therefore, sera from all four IL7-treated baboons

Fig. 8. Serum levels of IL7 in IL7 (black)- and placebo (gray)-treated
baboons. As in Fig. 1, bPre-TxQ denotes pretransplant, bPre-IL7Q denotesimmediately pre-IL7/placebo treatment, and subsequent numbers on the x-
axis denote weeks after starting IL7/placebo treatment. The period of IL7
treatment is indicated by the black horizontal bar. For a clearer
interpretation of this figure, the reader is referred to the figure in the Web
version of this article.
H. Lu et al. / Clinical Immunology 114 (2005) 30–41 39
collected at the end of IL7 treatment were assayed for the
presence of IL7 neutralizing antibodies. None were
detected.
Discussion
This study shows that, even in the setting of severe
lymphopenia and with 8-week treatment (longer than
previously reported), IL7 does not substantially stimulate
thymopoiesis. The positive finding of this study is that
IL7 administered to primates expands anti-CMV CD4 T
cells that are functional (produce IFNg and TNFa when
stimulated). This gives hope that IL7 administered to
patients will expand antiviral or antitumor CD4 T cells
capable of producing IFNg and TNFa. IFNg- or TNFa-
producing CD4 T cells can directly kill virus-infected or
tumor cells [15,33–44]. Virus/tumor-specific CD4 T cells
may also facilitate protection against viral disease or
cancer indirectly by sustaining virus/tumor-specific cyto-
toxic CD8 T cells (reviewed in Ref. [14]). Major
application of IL7 will likely be in immunotherapy. For
example, after high-dose chemo/radiotherapy for cancer
with autologous transplantation of CD34 cells (purified to
minimize the likelihood of reinfusing tumor cells), T cells
specific for the cancer and for viruses that may cause
disease after autologous CD34 cell transplantation like
CMV [45] or Epstein-Barr virus [46] could be infused
and expanded in vivo by the administration of IL7. In
herpes simplex virus-infected mice treated with antiviral
T cells, IL7 treatment resulted in 20-fold reduction in
viral load compared to controls treated with antiviral T
cells alone [47].
Regarding potential toxicities of IL7, it is reassuring that
even after 8-week treatment, lymphoma (detectable as a
mass on CT or necropsy) or osteoporosis did not develop,
and anemia and thrombocytopenia appeared to be mild.
However, the two deaths associated with T cell infiltration
of multiple organs, occurring in nine IL7-treated baboons,
are of concern. In both cases, the organs were infiltrated
with both CD4 and CD8 T cells, suggesting against
monoclonal/malignant lymphoproliferation. Viral etiology
is also unlikely as no inclusions or syncytia were noted on H
and E-stained sections of the most involved organs
(intestines in the first case and lungs and kidneys in the
second case) or other organs. Direct autoimmune damage of
organs by the infiltrating T cells is a plausible mechanism; in
the first case, the T cells were concentrated in the areas of
damaged intestinal crypts (Fig. 10 in Ref. [11]). Alter-
natively, the IL7-expanded T cells may have secreted other
cytokines, leading to a bcytokine storm.Q This would be
reminiscent of interleukin-2 toxicity which appears to be
mediated through the release of secondary cytokines like
TNF, IFNg, interleukin-1, and interleukin-6 [48]. The latter
mechanism is less likely as in normal monkeys treated with
IL7 (up to 1200 Ag/kg/day for less than 1 month) for the
determination of IL7 toxicity, no increase in serum levels of
TNFa, IFNg, or interleukin-6 was observed (M. Morre,
personal communication, March 2004). Despite the fact that
the pathogenesis of the death associated with T lympho-
proliferation has not been elucidated, the important obser-
vation is that it may be related to the degree of the IL7-
induced CD4 T cell expansion. The deaths occurred in two
of five animals whose CD4 T cell count during IL7
treatment exceeded 2500/Al and in none of four animals
with lower maximum CD4 T cell count during IL7
treatment. Among 21 normal 2-year-old baboons in which
we determined CD4 T cell counts, the two highest counts
were 2005 and 2130/Al. Our suggestion for the design of
clinical trials is to monitor CD4 T cell counts during IL7
treatment and either discontinue IL7 or proceed with
extreme caution after the CD4 T cell count has exceeded
the upper normal limit.
In conclusion, given the potential for increasing the
number of functional antiviral or antitumor T cells,
clinical testing of IL7 is indicated. Analogous to the
fact that life-threatening side effects of erythropoietin
(e.g., occlusion of a cerebral vessel) occur in patients
reaching high erythrocyte counts and those of filgrastim
(e.g., dyspnea associated with pulmonary X-ray infil-
trates) occur in patients reaching high granulocyte counts,
it is possible that life-threatening side effects of IL7 may
occur in patients reaching high CD4 T cell counts.

H. Lu et al. / Clinical Immunology 114 (2005) 30–4140
Acknowledgments
We thank Monja D. Metcalf, Roxanne Velez, Julia
Morris, and Renee Kendall for excellent technical assis-
tance. This work could not be done without the dedication
of the staff of the University of Washington National
Primate Research Center, in particular, Michael Gough,
Leslie Falch, Ed Novak, Mac Durning, Peggy Smith, Judy
Johnson, Dr. David Anderson, Dr. Steven Kelley, and Dr.
Maggie Gillen. The dedication and help of the computer
tomogram staff were also essential, namely, Olivia Hicks,
Mario Ramos, and Pat Manion. We also greatly acknowl-
edge Drs. Michel Morre and Renaud Buffet of Cytheris for
providing us with the recombinant baboon IL7.
References
[1] T.N. Small, E.B. Papadopoulos, F. Boulad, P. Black, H. Castro-
Malaspina, B.H. Childs, N. Collins, A. Gillio, D. George, A.
Jakubowski, G. Heller, M. Fazzari, N. Kernan, S. MacKinnon, P.
Szabolcs, J.W. Young, R.J. O’Reilly, Comparison of immune
reconstitution after unrelated and related T-cell-depleted bone marrow
transplantation: effect of patient age and donor leukocyte infusions,
Blood 93 (1999) 467.
[2] J. Storek, T. Gooley, R.P. Witherspoon, K.M. Sullivan, R. Storb,
Infectious morbidity in long-term survivors of allogeneic marrow
transplantation is associated with low CD4 T cell counts, Am. J.
Hematol. 54 (1997) 131.
[3] T.C.J. Merigan, J.G. Bartlett, D. Bolognesi, Textbook of AIDS
Medicine, Baltimore, Baltimore, 1999.
[4] M. Hakki, S.R. Riddell, J. Storek, R.A. Carter, T. Stevens-Ayers, P.
Sudour, K. White, L. Corey, M. Boeckh, Immune reconstitution to
cytomegalovirus after allogeneic hematopoietic stem cell transplanta-
tion: impact of host factors, drug therapy, and subclinical reactivation,
Blood 102 (2003) 3060.
[5] B. Ledergerber, M. Egger, V. Erard, R. Weber, B. Hirschel, H. Furrer,
M. Battegay, P. Vernazza, E. Bernasconi, M. Opravil, D. Kaufmann, P.
Sudre, P. Francioli, A. Telenti, AIDS-related opportunistic illnesses
occurring after initiation of potent antiretroviral therapy: the Swiss
HIV Cohort Study, JAMA 282 (1999) 2220.
[6] O.C. Boerman, T.A. Gregorio, K.J. Grzegorzewski, C.R. Faltynek, J.J.
Kenny, R.H. Wiltrout, K.L. Komschlies, Recombinant human IL-7
administration in mice affects colony-forming units-spleen and
lymphoid precursor cell localization and accelerates engraftment of
bone marrow transplants, J. Leukocyte Biol. 58 (1995) 151.
[7] E. Bolotin, M. Smogorzewska, S. Smith, M. Widmer, K. Weinberg,
Enhancement of thymopoiesis after BMT by in vivo interleukin-7,
Blood 88 (1996) 1887.
[8] A. Abdul-Hai, R. Or, S. Slavin, G. Friedman, L. Weiss, D. Matsa, A.
Ben-Yehuda, Stimulation of immune reconstitution by IL-7 after
syngeneic BMT in mice, Exp. Hematol. 24 (1996) 1416.
[9] C.L. Mackall, T.J. Fry, C. Bare, P. Morgan, A. Galbraith, R.E. Gress,
IL-7 increases both thymic-dependent and thymic-independent T-cell
regeneration after bone marrow transplantation, Blood 97 (2001)
1491.
[10] T.J. Fry, M. Moniuszko, S. Creekmore, S.J. Donohue, D.C. Douek, S.
Giardina, T.T. Hecht, B.J. Hill, K. Komschlies, J. Tomaszewski, G.
Franchini, C.L. Mackall, IL-7 therapy dramatically alters peripheral T-
cell homeostasis in normal and SIV-infected nonhuman primates,
Blood 101 (2003) 2294.
[11] J. Storek, T. Gillespy, H. Lu, A. Joseph, M.A. Dawson, M. Gough,
J.C. Morris, R.C. Hackman, P.A. Horn, G.E. Sale, R.G. Andrews,
D.G. Maloney, H.-P. Kiem, Interleukin-7 improves CD4 T cell
reconstitution after autologous CD34 cell transplantation in monkeys,
Blood 101 (2003) 4209.
[12] A.E. Broers, S.J. Posthumus-van Sluijs, H. Spits, B. van der Holt, B.
Lowenberg, E. Braakman, J.J. Cornelissen, Interleukin-7 improves T-
cell recovery after experimental T-cell-depleted bone marrow trans-
plantation in T-cell-deficient mice by strong expansion of recent
thymic emigrants, Blood 102 (2003) 1534.
[13] R. Scollay, D.I. Godfrey, Thymic emigration: conveyor belts or lucky
dips? Immunol. Today 16 (1995) 268.
[14] J.W. Gratama, J.J. Cornelissen, Diagnostic potential of tetramer-
based monitoring of cytomegalovirus-specific CD8+ T lymphocytes
in allogeneic stem cell transplantation, Clin. Immunol. 106 (2003)
29.
[15] L.E. Gamadia, E.B. Remmerswaal, J.F. Weel, F. Bemelman, R.A. van
Lier, I.J. Ten Berge, Primary immune responses to human CMV: a
critical role for IFN-gamma-producing CD4+ T cells in protection
against CMV disease, Blood 101 (2003) 2686.
[16] L.D. Chen, R.S. Kushwaha, H.C. McGill Jr., K.S. Rice, K.D. Carey,
Effect of naturally reduced ovarian function on plasma lipoprotein and
27-hydroxycholesterol levels in baboons (Papio sp.), Atherosclerosis
136 (1998) 89.
[17] B.A. Crawford, D.J. Handelsman, Androgens regulate circulating
levels of insulin-like growth factor (IGF)-I and IGF binding protein-3
during puberty in male baboons, J. Clin. Endocrinol. Metab. 81 (1996)
65.
[18] B.A. Crawford, J. Spaliviero, J. Simpson, D.J. Handelsman, Androgen
effects on bioactive and immunoreactive gonadotrophin levels during
puberty in male baboons, J. Pediatr. Endocrinol. Metab. 10 (1997)
401.
[19] X. Preville, M. Flacher, B. LeMauff, S. Beauchard, P. Davelu, J.
Tiollier, J.P. Revillard,Mechanisms involved in antithymocyte globulin
immunosuppressive activity in a nonhuman primate model, Trans-
plantation 71 (2001) 460.
[20] J.C. Morris, M. Conerly, B. Thomasson, J. Storek, S.R. Riddell, H.P.
Kiem, Induction of cytotoxic T-lymphocyte responses to enhanced
green and yellow fluorescent proteins after myeloablative condition-
ing, Blood 103 (2004) 492.
[21] R.L. Heberling, S.S. Kalter, Rapid dot-immunobinding assay on
nitrocellulose for viral antibodies, J. Clin. Microbiol. 23 (1986) 109.
[22] M.G. Michaels, F.J. Jenkins, K. St. George, M.A. Nalesnik, T.E.
Starzl, C.R. Rinaldo Jr., Detection of infectious baboon cytomegalo-
virus after baboon-to-human liver xenotransplantation, J. Virol. 75
(2001) 2825.
[23] K.G. Blume, S.J. Forman, F.R. Appelbaum, Thomas’s Hematopoietic
Cell Transplantation, Malden, Malden, 2003.
[24] M.G. Michaels, Nonhuman primate herpesviruses: importance for
xenotransplantation, Curr. Top. Microbiol. Immunol. 278 (2003) 73.
[25] A. Kaur, C.L. Hale, B. Noren, N. Kassis, M.A. Simon, R.P. Johnson,
Decreased frequency of cytomegalovirus (CMV)-specific CD4+ T
lymphocytes in simian immunodeficiency virus-infected rhesus
macaques: inverse relationship with CMV viremia, J. Virol. 76
(2002) 3646.
[26] J.W. Gratama, J.W. van Esser, C.H. Lamers, C. Tournay, B.
Lowenberg, R.L. Bolhuis, J.J. Cornelissen, Tetramer-based quantifi-
cation of cytomegalovirus (CMV)-specific CD8+ T lymphocytes in T-
cell-depleted stem cell grafts and after transplantation may identify
patients at risk for progressive CMV infection, Blood 98 (2001) 1358.
[27] K. Cwynarski, J. Ainsworth, M. Cobbold, S. Wagner, P. Mahendra, J.
Apperley, J. Goldman, C. Craddock, P.A. Moss, Direct visualization
of cytomegalovirus-specific T-cell reconstitution after allogeneic stem
cell transplantation, Blood 97 (2001) 1232.
[28] G. Aubert, A.F. Hassan-Walker, J.A. Madrigal, V.C. Emery, C. Morte,
S. Grace, M.B. Koh, M. Potter, H.G. Prentice, I.A. Dodi, P.J. Travers,
Cytomegalovirus-specific cellular immune responses and viremia in
recipients of allogeneic stem cell transplants, J. Infect. Dis. 184 (2001)
955.

H. Lu et al. / Clinical Immunology 114 (2005) 30–41 41
[29] C. Miyaura, Y. Onoe, M. Inada, K. Maki, K. Ikuta, M. Ito, T. Suda,
Increased B-lymphopoiesis by interleukin 7 induces bone loss in mice
with intact ovarian function: similarity to estrogen deficiency, Proc.
Natl. Acad. Sci. U. S. A. 94 (1997) 9360.
[30] N. Casadevall, J. Nataf, B. Viron, A. Kolta, J.J. Kiladjian, P. Martin-
Dupont, P. Michaud, T. Papo, V. Ugo, I. Teyssandier, B. Varet, P.
Mayeux, Pure red-cell aplasia and antierythropoietin antibodies in
patients treated with recombinant erythropoietin, N. Engl. J. Med. 346
(2002) 469.
[31] P. Ragnhammar, H.J. Friesen, J.E. Frodin, A.K. Lefvert, M. Hassan, A.
Osterborg, H. Mellstedt, Induction of anti-recombinant human gran-
ulocyte–macrophage colony-stimulating factor (Escherichia coli-
derived) antibodies and clinical effects in nonimmunocompromised
patients, Blood 84 (1994) 4078.
[32] R.L. Basser, E. O’Flaherty, M. Green, M. Edmonds, J. Nichol, D.M.
Menchaca, B. Cohen, C.G. Begley, Development of pancytopenia
with neutralizing antibodies to thrombopoietin after multicycle
chemotherapy supported by megakaryocyte growth and development
factor, Blood 99 (2002) 2599.
[33] P.D. Murray, D.B. McGavern, L.R. Pease, M. Rodriguez, Cellular
sources and targets of IFN-gamma-mediated protection against viral
demyelination and neurological deficits, Eur. J. Immunol. 32 (2002)
606.
[34] J.D. Ahlers, I.M. Belyakov, S. Matsui, J.A. Berzofsky, Mechanisms of
cytokine synergy essential for vaccine protection against viral
challenge, Int. Immunol. 13 (2001) 897.
[35] R.J. Hogan, W. Zhong, E.J. Usherwood, T. Cookenham, A.D.
Roberts, D.L. Woodland, Protection from respiratory virus infections
can be mediated by antigen-specific CD4(+) T cells that persist in the
lungs, J. Exp. Med. 193 (2001) 981.
[36] A.M. Harandi, B. Svennerholm, J. Holmgren, K. Eriksson, Differ-
ential roles of B cells and IFN-gamma-secreting CD4(+) T cells in
innate and adaptive immune control of genital herpes simplex virus
type 2 infection in mice, J. Gen. Virol. 82 (2001) 845.
[37] J.I. Sin, J. Kim, K. Dang, D. Lee, C. Pachuk, C. Satishchandran, D.B.
Weiner, LFA-3 plasmid DNA enhances Ag-specific humoral- and
cellular-mediated protective immunity against herpes simplex virus-2
in vivo: involvement of CD4+ T cells in protection, Cell. Immunol.
203 (2000) 19.
[38] C. Munz, K.L. Bickham, M. Subklewe, M.L. Tsang, A. Chahroudi,
M.G. Kurilla, D. Zhang, M. O’Donnell, R.M. Steinman, Human
CD4(+) T lymphocytes consistently respond to the latent Epstein-Barr
virus nuclear antigen EBNA1, J. Exp. Med. 191 (2000) 1649.
[39] S.S. Wimalasundera, D.R. Katz, B.M. Chain, Characterization of the
T cell response to human rhinovirus in children: implications for
understanding the immunopathology of the common cold, J. Infect.
Dis. 176 (1997) 755.
[40] F. Ren, K. Hino, Y. Yamaguchi, K. Funatsuki, A. Hayashi, H. Ishiko,
M. Furutani, T. Yamasaki, K. Korenaga, S. Yamashita, T. Konishi, K.
Okita, Cytokine-dependent anti-viral role of CD4-positive T cells in
therapeutic vaccination against chronic hepatitis B viral infection,
J. Med. Virol. 71 (2003) 376.
[41] L. Zitvogel, J.I. Mayordomo, T. Tjandrawan, A.B. DeLeo, M.R.
Clarke, M.T. Lotze, W.J. Storkus, Therapy of murine tumors with
tumor peptide-pulsed dendritic cells: dependence on T cells, B7
costimulation, and T helper cell 1-associated cytokines, J. Exp. Med.
183 (1996) 87.
[42] A.E. Patterson, R. Korngold, Cross-protective murine graft-versus-
leukemia responses to phenotypically distinct myeloid leukemia lines,
Biol. Blood Marrow Transplant. 6 (2000) 537.
[43] T. Morisaki, D.L. Morton, A. Uchiyama, D. Yuzuki, A. Barth, D.S.
Hoon, Characterization and augmentation of CD4+ cytotoxic T cell
lines against melanoma, Cancer Immunol. Immunother. 39 (1994)
172.
[44] E. Landais, X. Saulquin, E. Scotet, L. Trautmann, M.A. Peyrat,
J.L. Yates, W.W. Kwok, M. Bonneville, E. Houssaint, Direct
killing of Epstein-Barr virus (EBV)-infected B cells by CD4 T cells
directed against the EBV lytic protein BHRF1, Blood 103 (2004)
1408.
[45] L.A. Holmberg, M. Boeckh, H. Hooper, W. Leisenring, S. Rowley, S.
Heimfeld, O. Press, D.G. Maloney, P. McSweeney, L. Corey, R.T.
Maziarz, F.R. Appelbaum, W. Bensinger, Increased incidence of
cytomegalovirus disease after autologous CD34-selected peripheral
blood stem cell transplantation, Blood 94 (1999) 4029.
[46] R.A. Nash, R. Dansey, J. Storek, G.E. Georges, J.D. Bowen, L.A.
Holmberg, G.H. Kraft, M.D. Mayes, K.T. McDonagh, C.S. Chen, J.
Dipersio, C.F. Lemaistre, S. Pavletic, K.M. Sullivan, J. Sunderhaus,
D.E. Furst, P.A. McSweeney, Epstein-Barr virus-associated posttrans-
plantation lymphoproliferative disorder after high-dose immunosup-
pressive therapy and autologous CD34-selected hematopoietic stem
cell transplantation for severe autoimmune diseases, Biol. Blood
Marrow Transplant. 9 (2003) 583.
[47] P. Wiryana, T. Bui, C.R. Faltynek, R.J. Ho, Augmentation of cell-
mediated immunotherapy against herpes simplex virus by interleu-
kins: comparison of in vivo effects of IL-2 and IL-7 on adoptively
transferred T cells, Vaccine 15 (1997) 561.
[48] E.S. Vitetta, E. Coleman, M.-A. Ghetie, V. Ghetie, J. Michalek, L.M.
Pop, J.E. Smallshaw, C. Spiridon, Immunotherapy, in: W.E. Paul
(Ed.), Fundamental Immunology, Lippinkott Williams and Wilkins,
Philadelphia, 2003, pp. 1621.





























