Embed Size (px)
Citation preview

IrrepressibleMONOPTEROS/ARF5 promotes de novo shootformation
Wenzislava Ckurshumova, Tatiana Smirnova, Danielle Marcos, Yara Zayed and Thomas Berleth
Department of Cell and Systems Biology, University of Toronto, 25 Willcocks Street, Toronto, ON M5S 3B2, Canada
Author for correspondence:Thomas BerlethTel: +1 416 946 3734
Email: [email protected]
Received: 4 July 2014
Accepted: 9 July 2014
New Phytologist (2014) 204: 556–566doi: 10.1111/nph.13014
Key words: Arabidopsis, Auxin ResponseFactors, CYTOKININ RESPONSE FACTOR2
(CRF2), in vitro shoot formation, monopteros(MP), plant regeneration.
Summary
� In vitro regeneration of complete organisms from diverse cell types is a spectacular property
of plant cells. Despite the great importance of plant regeneration for plant breeding and bio-
technology, its molecular basis is still largely unclear and many important crop plants have
remained recalcitrant to regeneration.� Hormone-exposure protocols to trigger the de novo formation of either roots or shoots
from callus tissue demonstrate the importance of auxin and cytokinin signaling pathways, and
genetic differences in these pathways may contribute to the highly divergent responsiveness
of plant species to regeneration protocols.� In this study, we show that signaling through MONOPTEROS (MP)/AUXIN RESPONSE
FACTOR 5 is necessary for the formation of shoots from Arabidopsis calli. Most strikingly, an
irrepressible variant of MP, MPD, is sufficient for promoting de novo shoot formation through
pathways involving the genetically downstream functions of SHOOT MERISTEMLESS (STM)
and CYTOKININ RESPONSE FACTOR2 (CRF2).� We conclude that the MPD genotype can promote de novo shoot formation and can be
used to probe corresponding signaling pathways.
Introduction
Many plant improvement strategies, including in vitro propaga-tion, selection and transformation are based on the remarkableability of plant cells to regenerate whole organisms from smallsamples (explants) of adult tissue. Although in many plant speciesshoots and roots are readily regenerated from callus tissue, shootformation in particular is still a major bottleneck, especially incells from woody plants (Steeves & Sussex, 1989; Busov et al.,2005; Poupin & Arce-Johnson, 2005).
Plant growth occurs via the activity of two localized growthcenters of self-maintaining stem cells: the root apical and shootapical meristems (SAMs). Continuous activity of those meristemsgives rise to multiple cell types including cells needed for organinitiation. Apart from species-specific variations, the establishedway of inducing shoot formation in tissue culture involves a two-step process (Valvekens et al., 1988). In step one, explants areincubated on Callus Inducing Media (CIM), typically compris-ing a nontransported auxin, such as 2,4-dichlorophenoxy aceticacid (2,4-D) (Delbarre et al., 1996). Incubation on CIM willinduce the formation of cell masses called calli, which initiallydisplay levels of organization resembling lateral root primordia(Sugimoto et al., 2010; Fan et al., 2012). Inductive signals in thesecond (cytokinin rich) Shoot Inducing Medium (SIM) generatesshoot meristem-like structures, wherein cells depending on theirposition enter a specific developmental pathway – cells of the
center remain meristematic and cells on the flanks become organprimordia (Gordon et al., 2007; Sugimoto et al., 2010). Althoughcallus cultures in Arabidopsis are readily established from nearlyall organs and accessions of Arabidopsis (Acedo, 1986; Feldmann& David Marks, 1986), the likelihood of shoot induction is morevariable and influenced by the origin of the explant and the prop-erties of the accession. For example post-embryonic cotyledons ofthe Arabidopsis accession Col-0 are recalcitrant to regeneration,whereas explants from other accessions show higher organogeniccapacity (Akama et al., 1992; Zhao et al., 2002; Chatfield &Raizada, 2008).
In normal development, new shoot apical meristems (SAMs)arise very reproducibly in specific locations relative to pre-existingstructures, such as in the axils of newly developing leaf primordia(Barton, 2010). This high degree of reproducibility is probablydue to the synergistic influences of multiple local factors, not allof which can be coordinately supplied in tissue culture. Theemergence of new SAMs in tissue culture is thus not preciselypredictable and can be rather infrequent, but the genetic dissec-tion of SAM function in normal development has shed light onthe roles of critical components in the auxin and cytokinin path-ways in positioning the expression domains as key regulators ofshoot stem cell fate (Duclercq et al., 2011). As the role of planthormone signaling networks in normal development has becomeclearer (Vanstraelen & Benkov�a, 2012), a number of studies havelinked these findings to observations during in vitro
556 New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research

organogenesis (Birnbaum & Alvarado, 2008; Sugimoto et al.,2011) with emphasis on cross-talks between the auxin and cyto-kinin pathways (Kakani et al., 2009; Pernisov�a et al., 2009;Buechel et al., 2010; Cheng et al., 2010, 2012; Pernisova et al.,2011; Su et al., 2011). Although some reports assign functions inin vitro regeneration to several components of hormone signalingpathways (Chatfield & Raizada, 2008; Duclercq et al., 2011; Fanet al., 2012; Qiao et al., 2012; Nameth et al., 2013), these rolesare still unknown for most components.
Among the Arabidopsis transcription factors implicated inauxin signaling, MONOPTEROS/AUXIN RESPONSEFACTOR5/MP/ARF5, is well-characterized with regard todownstream target genes, which have implicated it not only asa mediator of auxin responses, but also as a key factor inhighly specialized local developmental decisions and as a mod-ulator of cytokinin signaling and production (Cole et al., 2009;Donner et al., 2009; Schlereth et al., 2010; Zhao et al., 2010;Cheng et al., 2012; Yamaguchi et al., 2013). The activity ofARFs is at least partially controlled through binding of nega-tive regulators of the Aux/IAA class to their interactiondomains III/IV (Guilfoyle & Hagen, 2012). In a variant ofMP, MPD, we have expressed a novel transgene deprived ofthese regulatory domains. Plants carrying MPD showedincreased transcription of target genes, prolonged expression ofauxin induced gene activities and the expression of phenotypicfeatures normally dependent on the supply of external auxin,suggesting that MPD represents an irrepressible variant of nor-mal MP gene activity (Krogan et al., 2012). In line with previ-ous reports about self-regulation of MP (Lau et al., 2011), theMP gene itself is enhanced and prolonged in its expression inMPD plants; in addition, patterning processes, and processesthat are thought to involve auxin-based feedback controls, suchas the formation of procambial strands in leaves, are vastlyintensified in MPD plants (Krogan & Berleth, 2012; Kroganet al., 2012).
Because of the pivotal role of auxin in the regeneration pro-cesses, and because of the genetically defined contribution of MPin the specification of the SAM stem cell niche (Zhao et al.,2010), we evaluated the role of MP during in vitro shoot forma-tion. Analysis of the organogenic responses define a role of MP inde novo shoot initiation and characterize MPD as a genetic tool toincrease the shoot formation.
Materials and Methods
Plant material and culture conditions
Regeneration frequency was defined as the percentage of explantsthat regenerated at least one shoot. The regeneration frequency/explant, was defined as, the number of shoots per explant.
Tissue culture procedures
Seeds were surface sterilized by the vapor-phasemethod (Clough&Bent, 1998) for 4 h in a desicator jar. Parfilm (Bioshop, Burlington,ON, Canada) sealed plates were stratified at 4°C for 3 d and seeds
were germinated and grown at 23°C under continuous light(150 lmol m�2 s�1). Growth medium was composed of half-strength Murashige and Skoog salts (½ MS) (Sigma), 15 g l�1
sucrose (Bioshop), 0.5 mg l�1 morpholino ethane sulfonic acid(MES, Sigma), pH 5.7. Root segments (2 cm), cotyledons, leavesor petioles were cut from 7–10-d-old seedlings and explanted ontosimplified callus induction medium (Gordon et al., 2007) (CIM;½ MS supplemented with 15 g l�1 sucrose (Bioshop), 0.5 g l�1
MES, 0.5 mg l�1 2,4-dichlorophenoxyacetic acid (2,4-D, Sigma)and 1 g l�1 Phytagel (Sigma). Cotyledons and leaves were placedadaxial side up on the media. Adventitious roots were harvestedfrom 12–14-d-old seedlings. mp mutant roots were obtained byincubating mp seedlings on growth medium supplemented with0.3 mg l�1 IBA (Sigma). For shoot induction, after 4 d of CIMincubation, unless otherwise specified, explants were transferred tosimplified shoot induction medium (SIM; ½ MS supplementedwith 1 mg l�1 6-a,a-dimethylallylamino-purine (2-IP, Sigma)(experiments reported in Figs 2, 4, 5) and 5 mg l�1 6-a,a-dim-ethylallylamino-purine (2-IP, Sigma) (experiments reported in Fig.3). For Fig. 1 shoot conversion was done as in Chatfield et al.(2013). All hormones were supplied to the medium after autoclav-ing. Culture plates were incubated at 22°Cunder constant light.
Quantitative real-time RT–PCR
Total RNA was extracted from calli after 6 d of SIM incubationwith the RNeasyTM kit (Qiagen) according to the manufac-turer’s instructions, and reverse transcription was performedwith 3 mg of total RNA using Superscript II reverse transcrip-tase (Invitrogen). Amplification was monitored in real timewith the CFX96 Real Time PCR System (Bio-Rad). Dataanalysis was done with the Bio-Rad CFX Manager software.Fold induction was calculated as a ratio of the signal fromselected genotypes to wt Col control. ACT2 was used to nor-malize each sample.
Microscopy
Explants were imaged directly on the media using a Leica MZFLIII stereomicroscope for either bright or dark field or epifluo-rescence (GFP2 filter) (Leica Microsystems, Concord, ON,Canada) or Canon EOS D60 for larger samples or plates. Forconfocal laser scanning microscopy, explants were mounted inwater and observed with a Zeiss Axiovert 100M microscopeequipped with a Zeiss LSM510 laser module confocal unit (Ze-iss, Canada). Confocal settings and band pass filters were as pre-viously described (Scarpella et al., 2006). pMP-GFP was imagedon a ZeisAX10 (Zeiss) equipped with an Olympus Q5 camera(Olympus, Richmond Hill, ON, Canada). MP:MP-GFPexpression in lateral roots was done on 3–5-d-old roots andstages were determined based on (Malamy & Benfey, 1997).
Histochemical detection of GUS activity was performed asdescribed (Scarpella et al., 2004). Samples were viewed with aLeica MZFLIII stereomicroscope (Leica Microsystems) equippedwith a Canon EOS D60 digital camera (Canon, Mississauga,ON, Canada)
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 556–566
www.newphytologist.com
NewPhytologist Research 557

Plant materials and genotyping
Allele mp1058 was genotyped using primers Lba1 TGGTTCACGTAGTGGGCCATCG and Afl-ins-r ATCGGAACAACACATCAAATGC. Homozygous crf2 (SAIL_371_D04) withprimers CRFOER:AACAGCTAAAAGAGGATCCGACCCGAATAG and CRFOE F:ATGGAAGCGGAGAAGAAAATGGTTCTACCG. 35S:CRF2 was generated using primers CRFOER:AACAGCTAAAAGAGGATCCGACCCGAATAG and CRFOE F:ATGGAAGCGGAGAAGAAAATGGTTCTACCG inpEGAD (Cutler et al., 2000).
MP:MP-GFP, pMP-GFP were previously described (Schlerethet al., 2010; Rademacher et al., 2011). 35S:STM:GR and the stm2allele were previously described (Endrizzi et al., 1996; Galloiset al., 2002).MPD (Krogan et al., 2012) was previously described.MPD was selected on plates containing 10 mg l�1 Basta (Gold-bio, St Louis, MO, USA) and 35S:STM:GR on Kanamycin(25 mg l�1) (Sigma). DEX induction was done by supplementingthe media with 1 lM dexamethasone (Sigma) for 35S:STM:GR.
Results
MP expression in root explants during callus and shootformation in vitro
During culture on CIM, explants from various organs displayreproducible gene expression patterns while forming lateral rootmeristem-like structures that later turn into calli (Cary et al.,2002; Che et al., 2006, 2007). As MP is expressed during lateralroot meristem formation (De Smet et al., 2010a,b), we comparedits expression during lateral root and callus formation and corre-lated its expression to the organogenic capacities of the resultingcalli.
As shown in Fig. 1(a–h), MP is expressed in a distinct cellularpattern in the formation of lateral root meristems. A functionalMP-GFP fusion gene (Schlereth et al., 2010) was switched onspecifically in short daughter cells of lateral root founder cells inthe pericycle (Fig. 1a,e), which are generated by asymmetricalanticlinal divisions in these locations (Malamy & Benfey, 1997).
(a) (b) (c) (d)
(h)(g)(f)(e)
(i) (j) (k) (l)
(p)(o)(n)(m)
Fig. 1 Expression ofMONOPTEROS (MP)during lateral root formation, callus inductionand shoot formation in Arabidopsis. (a–h)Successive stages of lateral root emergence,stage (Malamy & Benfey, 1997) indicated onthe bottom. (i–m) Callus formation; redarrowheads point at decliningMP
expression. Incubation times and media typeindicated on the bottom. (n–p) Shootformation on calli; white arrowhead points atnascentMP expression domain. (a, k, m)MP:MP-GFP; (l, n, p) pMP:GFP; (a–k, m)confocal laser scanning image, (a–d, i–k, m)expression shown with DIC overlay. (l, n, p)Epifluorescence. (o) Look-up-table (LUT)diagram for red box in (n). 5–10 explants percondition were imaged to ensurereproducibility of expression patterns. Bars:(a–m) 20 lm;(n, p) 200 lm.
New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist558

During the following stages, MP expression was strongest in themost central cell files of the stage IV (Malamy & Benfey, 1997)lateral root primordium, and much more weakly in peripheralcell layers (arrowheads in Fig. 1c,d,g,h). As a structured lateralroot meristem emerged, MP expression became graduallyrestricted and remained strong in cells of the columella and cen-tral cylinder (Fig. 1d,h), at this stage matching the reported MPexpression in the primary root (Rademacher et al., 2011).
After 12 h incubation of root explants on CIM, MP wasturned on in pericycle cells along the entire length of the explant(Fig. 1i). After 24 h on CIM, wherever the pericycle had under-gone a periclinal cell division, MP was also expressed in the newpericycle-derived cells (Fig. 1j). After 5 d on CIM, MP-GFP wasobserved in all rapidly proliferating cells and from day 6 onwardbecame progressively restricted to the callus front (Fig. 1l).Expression of MP subsided from the callus mass between 10 and16 d on CIM (Fig. 1m).
During culture on SIM, MP expression was very faint orabsent in most of the callus, but was instead strongly upregulatedin discrete spots (Fig. 1n–p), all of which later produced shoots(followed up individually 23/23). Expression was observed grad-ually increasing before overt shoot formation, commencingalready during the second day on SIM, and typically comprised aring-shaped zone of stronger expression (outer diameter 200–250 lm), enclosing a weaker expressing center (c. 50 lm)(Fig. 1n–o). After c. 6 d on SIM, leaf primordia became visible inthese positions (Fig. 1p). In summary, MP expression in emerg-ing calli initially resembled the expression in lateral root primor-dia, but soon became increasingly associated with proliferatingcells predominantly near the callus surface. In calli on SIM, MPexpression presaged the appearance of shoots, first highlightingthe incipient peripheral zone and finally the emerging leaf pri-mordia.
Shoot formation capacity andMP activity
We next measured the ability of calli to give rise to new SAMsafter culture on CIM over a period of 1–16 d. As shown inFig. 2(a), the propensity of calli to produce shoots increased withpre-incubation on CIM from 1 to 6 d, but sharply declined after> 10 d on CIM. Because of the correlation between the sharpdecline in both organogenic capacity and MP expression in calliafter > 10 d on CIM, and because of the high expression of MPin spots presaging the formation of new SAMs, we next askedwhether MP activity may be required for the de novo formationof SAMs in calli. To this end, we compared the probability ofSAM formation on calli of mpG12 mutant and wt explants. Asshown in Fig. 2(a), calli derived from mp mutant adventitiousroots never displayed a profile of shoot formation competencecomparable to that of wild-type calli. Regardless of the pre-incubation time on CIM, the proportion of shoot-forming calliremained far lower than even during the least competent stages inwild-type callus development. The consistently low shoot-form-ing capacity of mp mutant calli argues in favor of a generallyimpaired response of the tissue as opposed to a merely delayedresponse to hormone signals from the media.
Because the requirement for MP in regeneration could dependon the tissue of origin, we determined those values independentlyin explants from cotyledons, leaves and petioles. As shown inFig. 2(b), mp mutant explants from all organs showed a dramati-cally diminished ability to form shoots, except for explants fromcotyledons, which do not form shoots even in the Col-0 wild-type background (Chatfield & Raizada, 2008). The fact that sim-ilar genotype-specific differences in shoot formation capacitywere observed using explants from diverse organs argues in favorof a direct connection between MP activity and shooting capac-ity. It seems unlikely that genotype-specific differences in the tis-sue composition of the explants had a major role, because nosuch differences have been observed between wild-type andmutant petioles and roots (De Smet et al., 2010a,b).
Finally, we asked whether the MP genotype may just affect theproportion of a hypothetical subfraction of calli that were com-pletely recalcitrant to shoot formation, for example because ofsome unrecognized difference in their anatomy. If this was thecase, one would expect that those calli that produce shoots ineither genotype, should produce similar numbers of shoots percallus. However, as shown in Fig. 2(c), even those mp mutantcalli that generated shoots, hardly ever produced more than oneshoot per callus, whereas the average number of shoot-producingwild-type calli was close to three shoots per callus. In summary,the consistently strongly diminished shoot-forming capacity ofmp mutant calli, regardless of growth stage and tissue of originsuggests that MP activity is necessary for the proper signal trans-duction in calli in the de novo generation of SAMs.
(a)
(b) (c)
Fig. 2 MONOPTEROS (MP) activity indenovo shoot formation inArabidopsis. (a) Percentageof shoot-formingcalli in explants fromwt (wild-type)andmpadventitious rootsafter indicateddaysof incubationonCallusInducingMedia (CIM)before transfer toSIM.Valueswere scoredafter3 wkonSoot InducingMedia (SIM). (b)Percentageof shoot-formingcalli inexplants from indicatedorgans (4 dCIM,3 wkSIM). (c)Numberof shoots/organogenic callus (procedureas inb).Columns (wt,black;mp,white)represent themeanof four independent replicas (n = 25explantseach)� SD.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 556–566
www.newphytologist.com
NewPhytologist Research 559
MPD confers organogenic properties to explantsrecalcitrant to regeneration
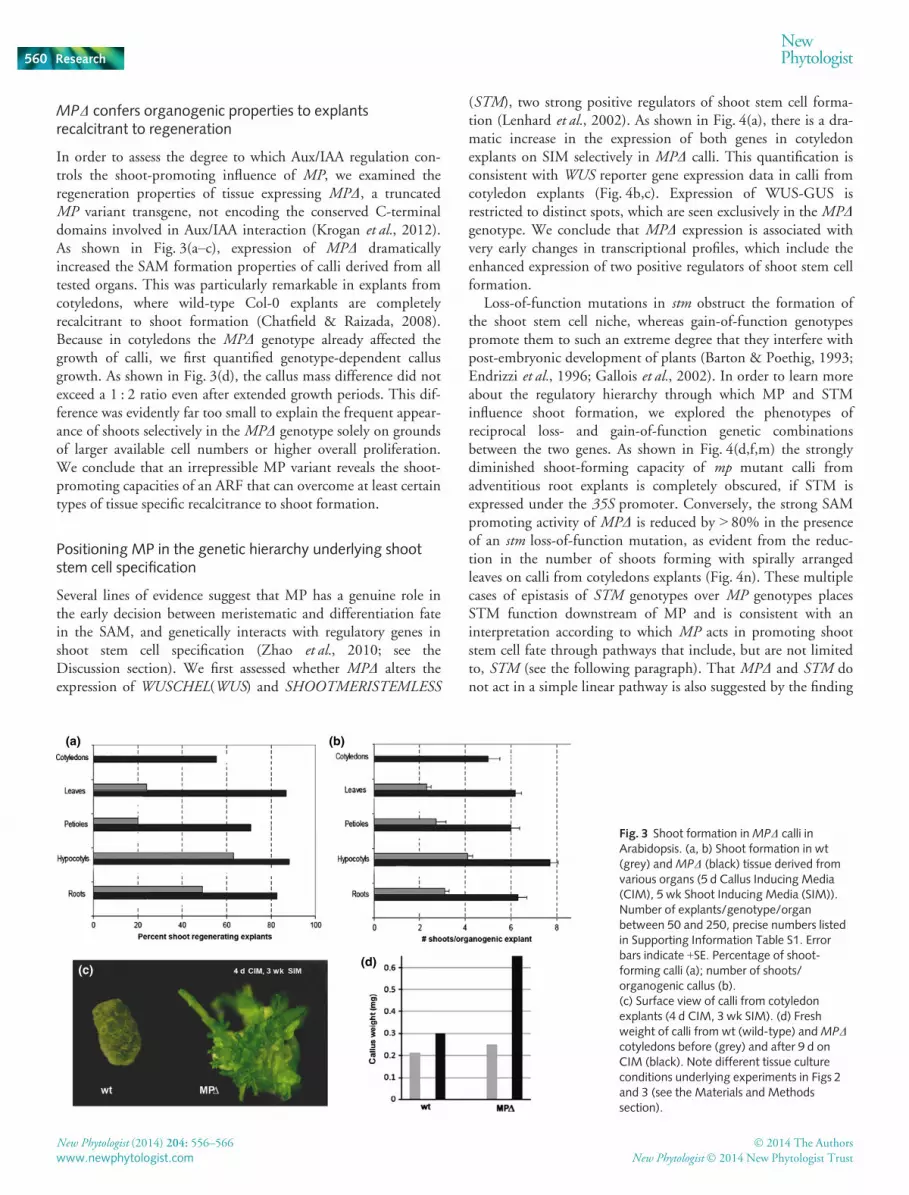
In order to assess the degree to which Aux/IAA regulation con-trols the shoot-promoting influence of MP, we examined theregeneration properties of tissue expressing MPD, a truncatedMP variant transgene, not encoding the conserved C-terminaldomains involved in Aux/IAA interaction (Krogan et al., 2012).As shown in Fig. 3(a–c), expression of MPD dramaticallyincreased the SAM formation properties of calli derived from alltested organs. This was particularly remarkable in explants fromcotyledons, where wild-type Col-0 explants are completelyrecalcitrant to shoot formation (Chatfield & Raizada, 2008).Because in cotyledons the MPD genotype already affected thegrowth of calli, we first quantified genotype-dependent callusgrowth. As shown in Fig. 3(d), the callus mass difference did notexceed a 1 : 2 ratio even after extended growth periods. This dif-ference was evidently far too small to explain the frequent appear-ance of shoots selectively in the MPD genotype solely on groundsof larger available cell numbers or higher overall proliferation.We conclude that an irrepressible MP variant reveals the shoot-promoting capacities of an ARF that can overcome at least certaintypes of tissue specific recalcitrance to shoot formation.
Positioning MP in the genetic hierarchy underlying shootstem cell specification
Several lines of evidence suggest that MP has a genuine role inthe early decision between meristematic and differentiation fatein the SAM, and genetically interacts with regulatory genes inshoot stem cell specification (Zhao et al., 2010; see theDiscussion section). We first assessed whether MPD alters theexpression of WUSCHEL(WUS) and SHOOTMERISTEMLESS
(STM), two strong positive regulators of shoot stem cell forma-tion (Lenhard et al., 2002). As shown in Fig. 4(a), there is a dra-matic increase in the expression of both genes in cotyledonexplants on SIM selectively in MPD calli. This quantification isconsistent with WUS reporter gene expression data in calli fromcotyledon explants (Fig. 4b,c). Expression of WUS-GUS isrestricted to distinct spots, which are seen exclusively in the MPDgenotype. We conclude that MPD expression is associated withvery early changes in transcriptional profiles, which include theenhanced expression of two positive regulators of shoot stem cellformation.
Loss-of-function mutations in stm obstruct the formation ofthe shoot stem cell niche, whereas gain-of-function genotypespromote them to such an extreme degree that they interfere withpost-embryonic development of plants (Barton & Poethig, 1993;Endrizzi et al., 1996; Gallois et al., 2002). In order to learn moreabout the regulatory hierarchy through which MP and STMinfluence shoot formation, we explored the phenotypes ofreciprocal loss- and gain-of-function genetic combinationsbetween the two genes. As shown in Fig. 4(d,f,m) the stronglydiminished shoot-forming capacity of mp mutant calli fromadventitious root explants is completely obscured, if STM isexpressed under the 35S promoter. Conversely, the strong SAMpromoting activity of MPD is reduced by > 80% in the presenceof an stm loss-of-function mutation, as evident from the reduc-tion in the number of shoots forming with spirally arrangedleaves on calli from cotyledons explants (Fig. 4n). These multiplecases of epistasis of STM genotypes over MP genotypes placesSTM function downstream of MP and is consistent with aninterpretation according to which MP acts in promoting shootstem cell fate through pathways that include, but are not limitedto, STM (see the following paragraph). That MPD and STM donot act in a simple linear pathway is also suggested by the finding
(a) (b)
(c)(d)
Fig. 3 Shoot formation inMPD calli inArabidopsis. (a, b) Shoot formation in wt(grey) andMPD (black) tissue derived fromvarious organs (5 d Callus Inducing Media(CIM), 5 wk Shoot Inducing Media (SIM)).Number of explants/genotype/organbetween 50 and 250, precise numbers listedin Supporting Information Table S1. Errorbars indicate +SE. Percentage of shoot-forming calli (a); number of shoots/organogenic callus (b).(c) Surface view of calli from cotyledonexplants (4 d CIM, 3 wk SIM). (d) Freshweight of calli from wt (wild-type) andMPDcotyledons before (grey) and after 9 d onCIM (black). Note different tissue cultureconditions underlying experiments in Figs 2and 3 (see the Materials and Methodssection).
New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist560

that MPD and 35S:STM-GR show additive effects. As shown inFig. 4(g–i) MPD 35S:STM-GR plants were able to convert lateralroot meristems on roots to shoot meristems in the absence ofexternal hormone application, a phenomenon not observed ineither genotype alone Second, neither MPD nor 35S:STM-GRformed multiple meristems around the shoot apical meristem,
but in MPD 35S:STM-GR plants, multiple meristems often aroseon the adaxial side of cotyledons (compare Fig. 4h with 4i).
Consistent with a role as a transcriptional target of MP(Schlereth et al., 2010), we found that the expression level ofCytokinin Response Factor2/CRF2 was drastically reduced in mpmutant background (Supporting Information Fig. S1). We then
(a) (b) (c)
(d)
(h)(g)
(f)(e)
(i)
(j) (k)
Fig. 4 MONOPTEROS (MP)-dependent pathways of shoot regeneration in Arabidopsis. (a) Gene expression profiles ofWUSCHEL (WUS) andSHOOTMERISTEMLESS (STM) in indicated genotypes determined by qRT-PCR (cotyledon derived calli, 8 d Callus Inducing Media (CIM), 6 d ShootInducing Media (SIM), note that in cotyledon-derived calli, anatomically recognizable shoot apical meristems (SAMs) arise after 6–9 d SIM; ACT7,transcript abundance control), bars are mean of three technical replicas � SD. MPD, black bars; crf2MPD, grey bars. (b, c)WUS:GUS expression after 6 don SIM (culture conditions as in a). (d–f) Shoot formation in calli from adventitious roots in genetic combinations involving 35S:STM-GR andmpG12 (4 dCIM, 2wk SIM). (g–i) Shoot formation in calli from adventitious roots in genetic combinations involving 35S:STM-GR andMPD (4 wk on ½MS, nophytohormones) Inset in (g) shows direct lateral root primordium conversion to SAM (red arrowheads), (h) shows ectopic SAMs on cotyledons; no suchfeatures are observed in either single genotypes (i and Krogan et al., 2012). (j, k) Quantification of shoot formation in indicated genotypes. Columnsrepresent means � SD. Culture conditions and explants in (j) as for (g–i), in (k) as for Fig. 2.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 556–566
www.newphytologist.com
NewPhytologist Research 561

tested whether CRF2 acts genetically downstream of MP inreciprocal genetic combinations. As shown in Fig. 5(c,d), the crf2mutation acted as a recessive extragenic suppressor of MPD,nearly abolishing the formation of SAMs on calli of MPD cotyle-don explants. Further, mutations in crf2 also strongly diminishedthe WUS and STM expression in MPD calli from cotyledon ex-plants (Fig. 4a). Inversely, uncoupling of CRF2 expression fromMP activity could partially overcome the absolute recalcitrantproperties of mp mutant cotyledon explants. In two out of 1535S:CRF2 mp lines, shoots were produced on cotyledon explantsand 35S:CRF2 expression could confer even higher shoot-form-ing properties in Col-0 background (Fig. 5d). We conclude thatCRF2 acts genetically downstream of MPD in shoot formationand, hence, that the shoot-promoting influence of MP is likely tobe partially conferred by CRF2.
Direct regulation of A-type ARRs, which act as negative regu-lators of cytokinin signaling, by MP has been established (Zhaoet al., 2010). If this is interpreted as a cytokinin-sensitizing func-tion of MP in the stem cell zone, it is interesting to note thatexpression of MPD allows for the generation of increased num-bers of SAMs not only on standard SIM, but also on media withconsiderably reduced cytokinin concentrations (Fig. S2). Weconclude that the promotion of de novo shoot formation by MPoccurs through several parallel inputs that have been associatedwith cytokinin signaling.
Discussion
Shoot meristems and the sizes of their stem cell niches are main-tained by the parallel action of several feedback mechanisms(reviewed in Barton, 2010). How these niches are set up duringplant development, or without reference to pre-existing structuresin tissue culture, is far less understood, although de novo shoot
formation constitutes a time-consuming bottleneck for in vitrotechnologies in many agronomically important species (Duclercqet al., 2011). In this study, we report that the introduction of anirrepressible variant of ARF5, MPD, significantly increases thefrequencies of de novo shoot formation in tissue from Arabidopsisand can be used to overcome organogenic recalcitrance of callifrom certain sources. Because in normal development, MP is animportant component in pattern-generating feedback processesinvolving auxin, its implication in regeneration processes mayprovide new entries towards a molecular understanding of theSAM self-organization in tissue culture. Besides ARF3, MP is thesecond ARF to be implicated in de novo shoot formation and, likeARF3 (Cheng et al., 2012), it seems to do so through affectingnot only auxin but also cytokinin pathways in the shoot meri-stem.
Control of shoot stem cell niches
In normal development, new SAMs appear in the axils of leavespromoted by signaling from the adaxial side of shoot organs(Barton, 2010). A first highly reproducibly specified SAM arisesin the center of a heart stage embryo in close proximity to aWUS expression domain between the cotyledon primordia (Lauxet al., 1996; Mayer et al., 1998; Schoof et al., 2000). In this natu-ral context, the expression of WUS and STM act as strongpositive regulators of shoot stem cell fate. Their localized expres-sion is critical for the scheduled formation of a stem cell niche inthe heart-stage embryo, whereby WUS expression commenceslong before overt SAM formation (Lau et al., 2012)
Among the classical plant hormones, cytokinins are longknown for promoting shoot formation (Skoog & Miller, 1957).Genetic manipulation of cytokinin levels or responsiveness influ-ence the size of shoot stem cell zones and the cytokinin signaling
(a) (b)
(c) (d)
Fig. 5 Shoot formation in calli fromArabidopsis cotyledons in geneticcombinations involvingMPD and crf2
(4 d CIM, 2 wk SIM). (a–c) Explants after3 wk on SIM (genotypes, bottom left). Notethe presence of meristems and leaf primordiain (a) as opposed to (b) and (c).(d) Quantification of shoot formation inindicated genotypes. Origin of explants andculture conditions as described in Fig. 2.
New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist562

pathway is an integral part of the SAM patterning mechanisms,which includes the regulation of STM and WUS (Rupp et al.,1999; Gordon et al., 2009) as well as of other canonical regula-tors of stem cell fate. Conversely, STM can promote cytokininbiosynthesis directly (Jasinski et al., 2005; Yanai et al., 2005) andWUS influences cytokinin signaling locally, as it diminishes thetranscript levels of negative regulators of cytokinin signaling ofthe A-type ARR class (Leibfried et al., 2005). Consistent withthese genetic findings, cytokinins have long been known to be themost critical component of media promoting shoot formation intissue culture (Skoog & Miller, 1957). Cytokinin-insensitivemutants are impaired in shooting, but by far not all recalcitranceto shoot formation in tissue culture can be explained in this way,as they show multiple developmental defects (Werner et al.,2003; Duclercq et al., 2011).
Analogies to other stem cell niches suggest that spatial arrange-ments of multiple signals might be necessary for the localizedspecification of stem cell fate and the influence of lateral shootorgans and local signaling through sRNAs and peptides has beendocumented (reviewed in Sablowski, 2011). Among other geneticinfluences that could promote SAM formation, ARFs have beenreported to act synergistically in the generation of the shoot stemcell niche, whereas they act antagonistically in the peripheral zoneof organ initiation (Furutani et al., 2004; Nordstr€om et al., 2004;Heisler et al., 2005; Zhao et al., 2010). Evidence for a role ofauxin is derived from direct experimental evidence in plant tissueculture (Heisler et al., 2005), global gene expression studies(Zhao et al., 2010) and genetic findings (Furutani et al., 2004;Leibfried et al., 2005), and are also supported by conceptual con-siderations, because experimental data as well as mathematicalmodeling have established auxin-based feedback controls in thede novo self-organization of many cellular patterns in plants(reviewed in Krupinski & Jonsson, 2010).
The role of MPD in de novo SAM formation
Cell biological and expression data have provided direct evidencefor a role of MP in the formation of shoot stem cell niches. Bybeing expressed near the stem cell zone, both WUS and MP havebeen shown to stabilize stem cell fate by suppressing the tran-scription of A-type ARRs (Zhao et al., 2010) as one prerequisitefor the formation of stable stem cell niches. In normal plantdevelopment, MP is expressed in the center of the heart-stageembryo, in leaf axils, flower buds and any other instance of sched-uled meristem formation in the shoot, processes that becomeirregular or lacking upon reduction of MP activity in mp mutants(Przemeck et al., 1996; Hardtke & Berleth, 1998; Wenzel et al.,2007). Thus, genetic evidence supports a strong positive influ-ence of MP on shoot stem cell formation. In fully structuredestablished SAMs, MP expression is strongest in the peripheralzone, especially in emerging lateral organs, whereas its expressionin the center is far weaker, and targeted central expression is actu-ally detrimental (Zhao et al., 2010). The fact that MP is compar-atively weakly expressed in the actual stem cell niche does notargue against its stem cell-promoting role. Just like WUS, itcould act best in this specific profile or act non-cell-
autonomously over the distance of a few cells (Mayer et al., 1998;Groß-Hardt et al., 2002). A stem cell-promoting influence fromthe adaxial side of lateral organs is well established by experimen-tal and genetic data (reviewed in Barton, 2010), and adaxiallyexpressed MP (Vidaurre et al., 2007; Wenzel et al., 2007; Kroganet al., 2012) could be crucial, as suggested by vastly delayed meri-stem development that is frequently observed in the axils of mpmutant leaves (Vidaurre et al., 2007). In the heart-stage embryo,for example, central WUS expression in combination with theadaxial cotyledon expression of MP would be ideally positionedfor defining the position of the shoot stem cell zone. If MP isinvolved in exerting a promoting influence from lateral shootorgans, its irrepressible variant may simply exert stronger activity,that might be beneficial in creating a stem cell niche. A dimin-ished influence from Aux/IAA protein-mediated regulation maynot be detrimental, as no vegetative SAM defects have beenobserved inMPD plants (Krogan et al., 2012).
Beyond this role, MPD could provide a positional anchor forother gene activities during early steps of SAM initiation. Auxinsignaling has been shown to be instrumental in many instances ofplant patterning, involving feedback mechanisms through whichproteins in auxin signal transduction and transport become local-ized to specific cells (reviewed in Leyser, 2010). In multiple pro-cesses, MP seems to have an inherent tendency to focus its ownexpression into lines or spots of cells (Wenzel et al., 2007; Lauet al., 2011) and we observed small discs of MP-expressing cellsemerging in calli shortly after transfer to SIM. These discs thenturned into ring-shaped expression domains, apparently repre-senting the peripheral zones of incipient primordia (Fig. 1n,o).As such, they would be expected to exert a positive influence onstem cell fate in the center, subsequently maintained by the devel-oping lateral organs. How the discs, and then rings of early MPexpression are specified is unknown, but if they involve auxin-based feedback loops, comparable, for example, to the formationof new veins in leaves and new lateral organs on SAMs, their den-sity reflecting the amount of MP activity should be increasedupon additition of an irrepressible MP variant to the system(Krogan et al., 2012). In the same way as the amount of MPactivity is critical for the frequency at which procambial cell fateis selected within the responsive environment of leaf groundmeri-stem cells, our MP loss- and gain-of-function genotypes suggestthat the incidence of forming spots and later rings of MP expres-sion might be controlled by the amount MP activity and elevatedby MPD.
Influence of MPD on STM and CRF2
Our observations in reciprocal combinations of gain- and loss-of-function genotypes position STM downstream of MP in shootstem cell specification and assign another surprisingly clear rolefor CRF2, an already molecularly established downstream targetgene of MP (Schlereth et al., 2010). These findings are not incon-sistent with the formation of SAMs in mp mutants, because thoseare often delayed and further delayed or absent when the activityof related ARFs is also reduced (Hardtke et al., 2004; Vidaurreet al., 2007). Both findings underscore the hypothesis that MP,
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 556–566
www.newphytologist.com
NewPhytologist Research 563

similar to WUS, has a role in locally modulating cytokinin sig-nalling (Zhao et al., 2010), and possibly through this, promotingSTM expression. Overlapping roles of WUS and MP are alsosuggested by the identical effects that expressions of 35S:WUSand of MPD have in 35S:STM background (Gallois et al., 2002and Fig. 4g,h).
The family of Cytokinin Response Factors (CRFs) comprises sixcytokinin inducible AP2-type transcription factors, which collec-tively have decisive roles in cytokinin-dependent gene regulation(Rashotte et al., 2006). The fully penetrant epistasis of crf2 loss-of-function mutations overMPD in the formation of shoots fromcotyledon explants is consistent with the previous characteriza-tion of CRF2 as a direct target of MP regulation (Schlereth et al.,2010) and provides first evidence for a stringent requirement of aspecific CRF gene in cell fate specification. The fact that theMPD-CRF2 pathway is strictly required and operates throughmolecularly tractable transcriptional regulation provides newexperimental opportunities to study recalcitrance of tissue sam-ples to shoot regeneration at the molecular level.
Acknowledgements
We would like to acknowledge project support from a NaturalSciences and Engineering Research Council of Canada (NSERC)discovery grant to T.B., and support from the Center for Analysisof Genome Evolution and Function (CAGEF). We would like tothank Drs T. Laux, P. Gallois and Shin-ya Miyagishima formaterials.
References
Acedo GN. 1986. Regeneration of Arabidopsis callus in vitro. Plant cell tissue andorgan culture 2: 109–114.
Akama K, Shiraishi H, Ohta S, Nakamura K, Okada K, Shimura Y. 1992.
Efficient transformation of Arabidopsis thaliana: comparison of the efficiencies
with various organs, plant ecotypes and Agrobacterium strains. Plant Cell Reports12: 7–11.
Barton M. 2010. Twenty years on: the inner workings of the shoot apical
meristem, a developmental dynamo. Developmental Biology 341: 95–113.Barton MK, Poethig RS. 1993. Formation of the shoot apical meristem in
Arabidopsis thaliana: an analysis of development in the wild type and in the
shoot meristemless mutant. Development 119: 823–831.Birnbaum KD, Alvarado AS. 2008. Slicing across Kingdoms: regeneration in
plants and animals. Cell 132: 697–710.Buechel S, Leibfried A, To JPC, Zhao Z, Andersen SU, Kieber JJ, Lohmann JU.
2010. Role of A-type ARABIDOPSIS RESPONSE REGULATORS in
meristem maintenance and regeneration. European Journal of Cell Biology 89:279–284.
Busov VB, Brunner AM, Meilan R, Filichkin S, Ganio L, Gandhi S, Strauss SH.
2005. Genetic transformation: a powerful tool for dissection of adaptive traits
in trees. New Phytologist 167: 9–18.Cary AJ, Che P, Howell SH. 2002. Developmental events and shoot apical
meristem gene expression patterns during shoot development in Arabidopsisthaliana. Plant Journal 32: 867–877.
Chatfield S, Raizada M. 2008. Ethylene and shoot regeneration: hookless1modulates de novo shoot organogenesis in Arabidopsis thaliana. Plant CellReports 27: 655–666.
Chatfield SP, Capron R, Severino A, Penttila PA, Alfred S, Nahal H, Provart
NJ. 2013. Incipient stem cell niche conversion in tissue culture: using a systems
approach to probe early events in WUSCHEL-dependent conversion of lateral
root primordia into shoot meristems.conversion of lateral root primordia into
shoot meristems. Plant Journal 73: 798–813.Che P, Lall S, Howell SH. 2007. Developmental steps in acquiring competence
for shoot development in Arabidopsis tissue culture. Planta 226: 1183–1194.Che P, Lall S, Nettleton D, Howell SH. 2006. Gene expression programs during
shoot, root, and callus development in Arabidopsis tissue culture. AnnualReview of Plant Physiology 141: 620–637.
Cheng ZJ, Wang L, Sun W, Zhang Y, Zhou C, Su YH, Li W, Sun TT, Zhao
XY, Li XG et al. 2012. Pattern of auxin and cytokinin responses for shoot
meristem induction results from regulation of cytokinin biosynthesis by Auxin
Response Factor 3. Plant Physiology 161: 240–251.Cheng ZJ, Zhu SS, Gao XQ, Zhang XS. 2010. Cytokinin and auxin regulates
WUS induction and inflorescence regeneration in vitro in Arabidopsis. PlantCell Reports 29: 927–933.
Clough SJ, Bent AF. 1998. Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant Journal16: 735–743.
Cole M, Chandler J, Weijers D, Jacobs B, Comelli P, Werr W. 2009.
DORNR€OSCHEN is a direct target of the auxin response factor
MONOPTEROS in the Arabidopsis embryo. Development 136: 1643–1651.Cutler SR, Ehrhardt DW, Griffitts JS, Somerville CR. 2000. Random
GFP∷cDNA fusions enable visualization of subcellular structures in cells of
Arabidopsis at a high frequency. Proceedings of the National Academy of Sciences,USA 97: 3718–3723.
De Smet I, Lau S, Mayer U, J€urgens G. 2010b. Embryogenesis – the humble
beginnings of plant life. Plant Journal 61: 959–970.De Smet I, Lau S, Voss U, Vanneste S, Benjamins R, Rademacher EH,
Schlereth A, De Rybel B, Vassileva V, Grunewald W et al. 2010a. Bimodular
auxin response controls organogenesis in Arabidopsis. Proceedings of theNational Academy of Sciences, USA 107: 2705–2710.
Delbarre A, Muller P, Imhoff V, Guern J. 1996. Comparison of mechanisms
controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid,
naphthalene-1-acetic acid, and indole-3-acetic acid in suspension-cultured
tobacco cells. Planta 198: 532–541.Donner TJ, Sherr I, Scarpella E. 2009. Regulation of preprocambial cell state
acquisition by auxin signaling in Arabidopsis leaves. Development 136: 3235–3246.
Duclercq J, Sangwan-Norreel B, Catterou M, Sangwan RS. 2011. De novo shootorganogenesis: from art to science. Trends in Plant Science 16: 597–606.
Endrizzi K, Moussian B, Haecker A, Levin JZ, Laux T. 1996. The SHOOTMERISTEMLESS gene is required for maintenance of undifferentiated cells in
Arabidopsis shoot and floral meristems and acts at a different regulatory level
than the meristem genesWUSCHEL and ZWILLE. Plant Journal 10: 967–979.Fan M, Xu C, Xu K, Hu Y. 2012. LATERAL ORGAN BOUNDARIES
DOMAIN transcription factors direct callus formation in Arabidopsis
regeneration. Cell Research 22: 1169–1180.Feldmann KA, David Marks M. 1986. Rapid and efficient regeneration of plants
from explants of Arabidopsis thaliana. Plant Science 47: 63–69.Furutani M, Vernoux T, Traas J, Kato T, Tasaka M, Aida M. 2004.
PIN-FORMED1 and PINOID regulate boundary formation and cotyledon
development in Arabidopsis embryogenesis. Development 131: 5021–5030.Gallois JL, Woodward C, Reddy GV, Sablowski R. 2002. Combined SHOOT
MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in
Arabidopsis. Development 129: 3207–3217.Gordon SP, Chikarmane VS, Ohno C, Meyerowitz EM. 2009.Multiple
feedback loops through cytokinin signalling control stem cell number within
the Arabidopsis shoot meristem. Proceedings of the National Academy of Sciences,USA 106: 16529–16534.
Gordon SP, Heisler MG, Reddy GV, Ohno C, Das P, Meyerowitz EM. 2007.
Pattern formation during de novo assembly of the Arabidopsis shoot meristem.
Development 134: 3539–3548.Groß-Hardt R, Lenhard M, Laux T. 2002.WUSCHEL signaling functions in
interregional communication during Arabidopsis ovule development. Genes &Development 16: 1129–1138.
Guilfoyle TJ, Hagen G. 2012. Getting a grasp on domain III/IV responsible for
Auxin Response Factor–IAA protein interactions. Plant Science 190: 82–88.
New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist564

Hardtke CS, Berleth T. 1998. The Arabidopsis geneMONOPTEROS encodes atranscription factor mediating embryo axis formation and vascular
development. EMBO Journal 17: 1405–1411.Hardtke CS, Ckurshumova W, Vidaurre DP, Singh SA, Stamatiou G, Tiwari
SB, Hagen G, Guilfoyle TJ, Berleth T. 2004.Overlapping and non-redundant
functions of the Arabidopsis auxin response factors MONOPTEROS and
NONPHOTOTROPIC HYPOCOTYL 4. Development 131: 1089–1100.Heisler MG, Ohno C, Das P, Sieber P, Reddy GV, Long JA, Meyerowitz EM.
2005. Patterns of auxin transport and gene expression during primordium
development revealed by live imaging of the Arabidopsis inflorescence
meristem. Current Biology 15: 1899–1911.Jasinski S, Piazza P, Craft J, Hay A, Woolley L. 2005. KNOX action in
Arabidopsis is mediated by coordinate regulation of cytokinin and gibberellin
activities. Current Biology 15: 1560.Kakani A, Li G, Peng Z. 2009. Role of AUX1 in the control of organ identity
during in vitro organogenesis and in mediating tissue specific auxin and
cytokinin interaction in Arabidopsis. Planta 229: 645–657.Krogan N, Berleth T. 2012. A dominant mutation reveals asymmetry in MP/
ARF5 function along the adaxial–abaxial axis of shoot lateral organs. PlantSignaling & Behavior 7: 940–943.
Krogan NT, Ckurshumova W, Marcos D, Caragea AE, Berleth T. 2012.
Deletion of MP/ARF5 domains III and IV reveals a requirement for Aux/IAA
regulation in Arabidopsis leaf vascular patterning. New Phytologist 194: 391–401.
Krupinski P, Jonsson H. 2010.Modeling auxin-regulated development. ColdSpring Harbor Perspectives in Biology 2: a001560.
Lau S, Slane D, Heru O, Kong J, Jurgens G. 2012. Early embryogenesis in
flowering plants: setting up the basic body pattern. Annual Review of PlantBiology 63: 483–506.
Lau S, Smet ID, Kolb M, Meinhardt H, J€urgens G. 2011. Auxin triggers a
genetic switch. Nature Cell Biology 13: 611–615.Laux T, Mayer KF, Berger J, J€urgens G. 1996. TheWUSCHEL gene is required
for shoot and floral meristem integrity in Arabidopsis. Development 122: 87–96.
Leibfried A, To JPC, Busch W, Stehling S, Kehle A, Demar M, Kieber JJ,
Lohmann JU. 2005.WUSCHEL controls meristem function by direct
regulation of cytokinin-inducible response regulators. Nature 438: 1172–1175.Lenhard M, J€urgens G, Laux T. 2002. TheWUSCHEL and
SHOOTMERISTEMLESS genes fulfil complementary roles in Arabidopsis
shoot meristem regulation. Development 129: 3195–3206.Leyser O. 2010. The power of auxin. Plant Physiology 154: 501–505.Malamy JE, Benfey PN. 1997.Organization and cell differentiation in lateral
roots of Arabidopsis thaliana. Development 124: 33–44.Mayer K, Schoof H, Haecker A, Lenhard M, J€urgens G, Laux T. 1998. Role of
WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell95: 805–815.
Nameth B, Dinka SJ, Chatfield SP, Morris A, English J, Lewis D, Oro R, Raizada
MN. 2013. The shoot regeneration capacity of excised Arabidopsis cotyledons is
established during the initial hours after injury and is modulated by a complex
genetic network of light signalling. Plant, Cell & Environment 36: 68–86.Nordstr€om A, Tarkowski P, Tarkowska D, Norbaek R, Astot C, Dolezal K,
Sandberg G. 2004. Auxin regulation of cytokinin biosynthesis in Arabidopsisthaliana: a factor of potential importance for auxin-cytokinin-regulated
development. Proceedings of the National Academy of Sciences, USA 101: 8039–8044.
Pernisov�a M, Kl�ıma P, Hor�ak J, V�alkov�a M, Malbeck J, Soucek P, Reichman P,
Hoyerov�a K, Dubov�a J, Friml J et al. 2009. Cytokinins modulate
auxin-induced organogenesis in plants via regulation of the auxin efflux.
Proceedings of the National Academy of Sciences, USA 106: 3609–3614.Pernisova M, Kuderova A, Hejatko J. 2011. Cytokinin and auxin Iiteractions in
plant development: metabolism, signalling, transport and gene expression.
Current Protein and Peptide Science 12: 137–147.Poupin MJ, Arce-Johnson P. 2005. Transgenic trees for a new era. In VitroCellular & Developmental Biology-Plant 41: 91–101.
Przemeck GK, Mattsson J, Hardtke CS, Sung ZR, Berleth T. 1996. Studies on
the role of the Arabidopsis geneMONOPTEROS in vascular development and
plant cell axialization. Planta 200: 229–237.
Qiao M, Zhao Z, Song Y, Liu Z, Cao L, Yu Y, Li S, Xiang F. 2012. Proper
regeneration from in vitro cultured Arabidopsis thaliana requires themicroRNA-directed action of an auxin response factor. Plant Journal 71: 14–22.
Rademacher EH, M€oller B, Lokerse AS, Llavata-Peris CI, van den Berg W,
Weijers D. 2011. A cellular expression map of the Arabidopsis AUXINRESPONSE FACTOR gene family. Plant Journal 68: 597–606.
Rashotte AM, Mason MG, Hutchison CE, Ferreira FJ, Schaller GE, Kieber JJ.
2006. A subset of Arabidopsis AP2 transcription factors mediates cytokinin
responses in concert with a two-component pathway. Proceedings of theNational Academy of Sciences, USA 103: 11 081–11 085.
Rupp H-M, Frank M, Werner T, Strnad M, Schm€ulling T. 1999. Increased
steady state mRNA levels of the STM and KNAT1 homeobox genes in
cytokinin overproducing Arabidopsis thaliana indicate a role for cytokinins inthe shoot apical meristem. Plant Journal 18: 557–563.
Sablowski R. 2011. Plant stem cell niches: from signalling to execution. CurrentOpinion in Plant Biology 14: 4–9.
Scarpella E, Francis P, Berleth T. 2004. Stage-specific markers define early steps
of procambium development in Arabidopsis leaves and correlate termination of
vein formation with mesophyll differentiation. Development 131: 3445–3455.Scarpella E, Marcos D, Friml J, Berleth T. 2006. Control of leaf vascular
patterning by polar auxin transport. Genes & Development 20: 1015–1027.Schlereth A, M€oller B, Liu W, Kientz M, Flipse J, Rademacher EH, Schmid
M, J€urgens G, Weijers D. 2010. MONOPTEROS controls embryonic
root initiation by regulating a mobile transcription factor. Nature 464:913–916.
Schoof H, Lenhard M, Haecker A, Mayer K, J€urgens G, Laux T. 2000. The
stem cell population of Arabidopsis shoot meristems is maintained by a
regulatory loop between the CLAVATA andWUSCHEL genes. Cell 100: 635–644.
Skoog F, Miller CO. 1957. Chemical regulation of growth and organ formation
in plant tissues cultured in vitro. Symposia of the Society for Experimental Biology11: 118–130.
Steeves TA, Sussex IM. 1989. Patterns in plant development. Cambridge, UK:
Cambridge University Press.
Su YH, Liu YB, Zhang XS. 2011. Auxin-cytokinin interaction regulates meristem
development.Molecular Plant 4: 616–625.Sugimoto K, Gordon SP, Meyerowitz EM. 2011. Regeneration in plants and
animals: dedifferentiation, transdifferentiation, or just differentiation? Trends inCell Biology 21: 212–218.
Sugimoto K, Jiao Y, Meyerowitz EM. 2010. Arabidopsis regeneration from
multiple tissues occurs via a root development pathway. Developmental Cell 18:463–471.
Valvekens D, Montagu MV, Lijsebettens MV. 1988. Agrobacterium
tumefaciens-mediated transformation of Arabidopsis thaliana root explants byusing kanamycin selection. Proceedings of the National Academy of Sciences, USA85: 5536–5540.
Vanstraelen M, Benkov�a E. 2012.Hormonal interactions in the regulation of
plant development. Annual Review of Cell and Developmental Biology 28: 463–487.
Vidaurre DP, Ploense S, Krogan NT, Berleth T. 2007. AMP1 and MP
antagonistically regulate embryo and meristem development in Arabidopsis.
Development 134: 2561–2567.Wenzel CL, Schuetz M, Yu Q, Mattsson J. 2007. Dynamics of
MONOPTEROS and PIN-FORMED1 expression during leaf vein pattern
formation in Arabidopsis thaliana. Plant Journal 49: 387–398.Werner T, Motyka V, Laucou V, Smets R, Van Onckelen H, Schm€ulling T.
2003. Cytokinin-deficient transgenic Arabidopsis plants show multiple
developmental alterations indicating opposite functions of cytokinins in the
regulation of shoot and root meristem activity. Plant Cell 15: 2532–2550.Yamaguchi N, Wu MF, Winter CM, Berns MC, Nole-Wilson S, Yamaguchi A,
Coupland G, Krizek BA, Wagner D. 2013. A molecular framework for
auxin-mediated initiation of flower primordia. Developmental Cell 24: 271–282.
Yanai O, Shani E, Dolezal K, Tarkowski P, Sablowski R, Sandberg G, Samach
A, Ori N. 2005. Arabidopsis KNOXI proteins activate cytokinin biosynthesis.
Current Biology 15: 1566–1571.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 204: 556–566
www.newphytologist.com
NewPhytologist Research 565

Zhao Q, Fisher R, Auer C. 2002. Developmental phases and STM
expression during Arabidopsis shoot organogenesis. Plant Growth Regulation37: 223–231.
Zhao Z, Andersen SU, Ljung K, Dolezal K, Miotk A, Schultheiss SJ, Lohmann
JU. 2010.Hormonal control of the shoot stem-cell niche. Nature 465: 1089–1092.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Expression profile of CRF2 in wt and mp determined byqRT-PCR.
Fig. S2 Shoot formation in MPD cotyledon explants under vary-ing cytokinin conditions.
Table S1 Explant numbers for Fig. 3
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
New Phytologist (2014) 204: 556–566 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist566





























