Embed Size (px)
Citation preview

ENVIRONMENTAL MICROBIOLOGY
Isolation of UVC-Tolerant Bacteria from the HyperaridAtacama Desert, Chile
Ivan Gláucio Paulino-Lima & Armando Azua-Bustos &
Rafael Vicuña & Carlos González-Silva & Loreto Salas &
Lia Teixeira & Alexandre Rosado &
Alvaro Augusto da Costa Leitao & Claudia Lage
Received: 2 August 2012 /Accepted: 29 August 2012 /Published online: 23 September 2012# Springer Science+Business Media, LLC 2012
Abstract Martian surface microbial inhabitants would bechallenged by a constant and unimpeded flux of UV radiation,and the study of analog model terrestrial environments may beof help to understand how such life forms could survive underthis stressful condition. One of these environments is theAtacama Desert (Chile), a well-known Mars analog due toits extreme dryness and intense solar UV radiation. Here, wereport the microbial diversity at five locations across this desertand the isolation of UVC-tolerant microbial strains found inthese sites. Denaturing gradient gel electrophoresis (DGGE) of16S rDNA sequences obtained from these sites showed band-ing patterns that suggest distinct and complex microbial com-munities. Analysis of 16S rDNA sequences obtained fromUV-tolerant strains isolated from these sites revealed species relat-ed to theBacillus and Pseudomonas genera. Vegetative cells ofone of these isolates, Bacillus S3.300-2, showed the highestUV tolerance profile (LD100318 Jm2), tenfold higherthan a wild-type strain of Escherichia coli. Thus, our
results show that the Atacama Desert harbors a noteworthymicrobial community that may be considered for futureastrobiological-related research in terms of UV tolerance.
Introduction
Life is continuously threatened by abiotic stresses in desertenvironments, among them, wide temperature fluctuations,desiccation, lack of nutrients, and intense solar radiation[11]. The Atacama Desert, located between 17° S and 27°S latitude in northern Chile, is the driest and oldest desert onEarth, having experienced extreme aridity for the past 150million years and hyperaridity for the last 15 million years[12, 13]. To cope with these conditions, its inhabitantsadapted to various harsh conditions, such as very low airhumidity, a nearly complete absence of rains, and high-fluxsolar radiation [12, 13, 19]. In addition, soils of the Atacama
I. G. Paulino-Lima :A. A. da Costa Leitao : C. Lage (*)Carlos Chagas Filho Biophysics Institute,Rio de Janeiro Federal University,Centro de Ciências da Saúde, Bld G. 373 Carlos Chagas FilhoAve., Cidade Universitária,21941-902, Rio de Janeiro, RJ, Brazile-mail: [email protected]
I. G. Paulino-Limae-mail: [email protected]
A. Azua-Bustos : R. Vicuña : L. SalasFaculty of Biological Sciences, Department of Molecular Geneticsand Microbiology, Pontificia Universidad Católica de Chile,Santiago, Chile
C. González-SilvaCentro de Investigación del Medio Ambiente (CENIMA),Universidad Arturo Prat,Iquique, Chile
L. Teixeira :A. RosadoProf. Paulo de Goes Microbiology Institute,Rio de Janeiro Federal University,Centro de Ciências da Saúde, Bld I. 373 Carlos Chagas Filho Ave.,Cidade Universitária,21941-902, Rio de Janeiro, RJ, Brazil
A. Azua-Bustos : R. VicuñaMillennium Institute for Fundamental and Applied Biology(MIFAB),Santiago, Chile
Present Address:I. G. Paulino-LimaNASA Ames Research Center,Mail Stop 239-20, Bldg. N239, Rm 377, P.O. Box 1, Moffett Field,CA 94035-0001, USA
Microb Ecol (2013) 65:325–335DOI 10.1007/s00248-012-0121-z

contain chemically aggressive sulfates, chlorides, and per-chlorates in concentrations well above the limits tolerated bymesophilic microorganisms [2]. Annual average sunlight inthe Atacama core is 335 Wm−2 with a consistent dailymaximum over 1,000 Wm−2 [16]. Maximum values forphotosynthetic active radiation (PAR) recorded in 2006summer at Yungay and Salar Grande were as high as 2.37and 2.21 mmolm−2s−1, respectively [6].
These harsh physical and chemical environmental con-straints apparently caused those extensive regions of theAtacama Desert to be almost devoid even of microbial life,with abundances of one or two orders of magnitude belowthose found in any other arid region on Earth [19].
Several studies have been conducted on the Atacama Des-ert in order to obtain a better profiling of its extremophilicmicrobial communities [2, 17, 19, 24, 26]. Navarro-Gonzalezet al. [19] were among the first to report the unique propertiesof soils in the extreme arid core region of the Atacama Desert.Samples from this region exhibited only trace levels oforganics and variable scores of culturable and noncul-turable bacteria [10].
However, little has been published on indigenous UVradiation-tolerant microbial populations of the AtacamaDesert. An impressive silica-encased filamentous cyanobac-teria was recently found in El Tatio geothermal fields at theAndes Mountains in the Atacama. The silica matrices weresuggested to provide cells with an efficient UV shield,without PAR attenuation [22]. In a different study conductedin the dry core (Yungay) of the Atacama Desert, nonnativemicroorganisms quickly died upon exposure to solar radia-tion at the soil surface [4, 9]. However, when protected bygypsum and mineral grain coverings, survival was extendedup to 8 days, suggesting that UV-tolerant microorganismsmay be able to survive below the soil surface of theAtacama. One well-known UV-tolerant microorganismwhich members have been found across the Atacama Desertis Chroococcidiopsis. These cyanobacterial species havebeen found in endolithic and hypolithic habitats [1, 26].Besides this Cyanobacterium, there are no reports on thedescription and further enumeration of UV-tolerant micro-organisms sampled from this Desert. Here, we report onnovel bacterial isolates (mainly from the Bacillus genus)collected from different sites of the Atacama. The UVtolerance of one of these isolates is about tenfold that of awild-type Escherichia coli strain.
Material and Methods
Sampling Sites
Five different sites in the Atacama Desert (Chile) weresampled (Fig. 1) using sterile 50-ml Falcon tubes.
Except for site 3 in which an epilithic biofilm wassampled, at the remaining sites, subsurface soils werecollected by first removing and discarding the topmost5 cm of the soil surface. These samples were thenimmediately shipped to the Molecular RadiobiologyLab at Carlos Chagas Filho Institute of Biophysics,Rio de Janeiro Federal University, Brazil. This samplingwas performed in July of 2008, which is the wet seasonin this area of the Atacama Desert.
The sample from site 1 was collected at the shores ofthe Salar de Llamara, a hypersaline environment in theCentral Valley, northeast of the city of Iquique, which isknown for its microbial mats formed by several speciesof bacteria algae and archaea found at the bottom of itshypersaline lagoons and beneath the surrounding saltcrusts (Fig. 1a) [7]. This sample was composed of amixture of clay and salt crusts. The sample from site 2was collected in a quartz-covered field at the CoastalRange of the Atacama, south of the city of Antofagastaand close to the Pacific Ocean (Fig. 1b). This samplewas mostly soil and small pieces of rocks. Sample 3was collected from the region of La Portada, north ofAntofagasta, from biofilm-covered rocks directly con-fronting the Pacific Ocean (Fig. 1c). This sample wascomposed of brittle chips of petrified sea shells coveredby black biofilms. Sample 4 was collected from a fieldcovered with dispersed gypsum patches and basalticrocks, adjacent to site 2 (Fig. 1d). Sample from site 4included both gypsum chips and the surrounding soils.Sample 5, a different quartz-covered field was collectedclose to site 2. This sample was mostly soil, similar tosample taken from site 2.
Microbial Enumerations
A one-gram sample from each site was placed separate-ly in five conic flasks containing 10 ml saline solution(0.9 % NaCl) and glass beads. After shaking (100 rpm)for 3 h at room temperature (~22 °C), each mixture wassubmitted to serial dilution and plating on three differ-ent culture media: LB (1.0 % tryptone, 0.5 % NaCl,0.5 % yeast extract, 1.5 % agar), TGY (0.5 % tryptone,0.3 % yeast extract, 0.1 % glucose, 1.5 % agar), andMarine Agar (MA) 2216 (Difco) for enumeration ofcultivable microorganisms.
Microbial Screening of UV Tolerance
As the focus of this work was the isolation of micro-organisms tolerant to UV radiation, the same soil sus-pensions used for microbial enumeration were exposedseparately to UVC radiation using a germicidal lampemitting a flux of 2.0 Jm−2s−1, as measured with a
326 I. G. Paulino-Lima et al.

Vilber Lourmat radiometer using a photocell CX-254. Avolume of 5 ml from each soil suspension was placedin 9 mm diameter glass Petri dishes and exposed to atotal fluency of 300 Jm−2 UVC (λ0254 nm). Thistreatment was performed under continuous shaking tohomogenize the cell population being irradiated and tominimize the protection effects caused by the particulatematerial of the soil suspension. After the UVC
irradiation, samples were subjected to serial dilutionand plating on culture media (LB, TGY, and MA) formicrobial enumeration of survivors (A in Fig. 2). Afterup to 7 days of incubation at 28 °C, the number ofcolony-forming units (CFU) corresponding to the irradi-ated aliquots (N) was divided by the number of CFUcorresponding to the nonirradiated control (N0), resultingin survival fractions (N/N0) for each sample.
Fig. 1 a Geographical location of the sampled sites (modified from aNASA MODIS image). Site labeling: b Salar de Llamara (21°16′07.54″S, 69°37′03.90″W. Altitude: 751 m); c quartz field 1 (23°48′59.15″S, 70°29′25.59″W. Altitude: 538 m); d cliff biofilms, Capa
Negra La Portada (23°29′58.61″S, 70°25′42.10″W. Altitude: 27 m); esoil gypsum field (23°49′01.14″S, 70°29′22.98″W. Altitude: 531 m); fquartz field 2 (23°49′10.76″S, 70°28′36.77″W. Altitude: 736 m)
UVC-Tolerant Bacteria from the Hyperarid Atacama Desert 327

In order to check the efficiency of the irradiationtreatment, the sample from site 4 was subjected to anadditional irradiation experiment with fractionated fluen-cies (Joule per square meters) of 0, 25, 50, 100, 200,300, and 400 (B in Fig. 2). This sample was chosen forthis procedure because it showed a higher number ofpigmented morphotypes after the single exposure irradi-ation experiment. Survival fractions (N/N0) were deter-mined as described above.
A third approach for screening UV-resistant microorgan-isms was applied by sprinkling about 100 mg samples fromeach site onto three agar plates (LB, TGY, and MA). Afterup to 7 days of incubation at 28 °C, pigmented morphotypeswere isolated for subsequent analysis (C in Fig. 2).
Determination of UVC Tolerance
Since pigmentation is a known cellular protection mech-anism against UV radiation [14], different colored mor-photypes were chosen among the survivors forsubsequent analysis (D in Fig. 2). These isolates weregrown in liquid culture medium (LB, TGY, and MarineBroth (MB) 2216, Difco) under shaking at 150 rpm,
28 °C. The isolates able to grow under these conditionswere subjected to a new step of UVC irradiation, thistime in saline solution without the interference of theparticulate material from the soil samples. For this,cultures of each isolate were grown under shaking at150 rpm, 28 °C for up to 48 h, until reaching OD600
~0.8. A volume of 5 ml from each culture was washedtwice in saline solution through centrifugation at 8,000×g for 5 min at 4 °C, followed by replacement ofsupernatant by a new saline solution and vortexing for10 s. The resulting cell suspension was placed in 9 mmdiameter glass Petri dishes, exposed to UVC irradiation(300 Jm−2) and recovered for survival fraction analysis,as described previously. Results were plotted on a graphshowing different survival fractions.
Survival Curve
The most UV-tolerant isolate (S3.300-2) was grown in MBunder shaking at 150 rpm, 28 °C for up to 48 h, untilreaching OD600 ~0.8. A volume of 5 ml from each culturewas washed twice in saline solution as described above. Theresulting cell suspension was placed in 9 mm diameter glass
Fig. 2 Scheme of the isolationand irradiation procedures usedin this study. A Irradiation of theliquid phase of each samplewith 300 Jm−2 of UVCradiation. B Irradiation ofsample from site 4 with severaldoses of UVC radiation. CInoculation of soil in natura onagar plates. D Visual inspectionand selection of pigmentedmorphotypes. E Irradiation ofisolate S3.300-2 with severaldoses of UVC radiation
328 I. G. Paulino-Lima et al.

Petri dishes, exposed to UVC radiation using fractionatedfluences (Joule per square·meters) of 0, 50, 100, 200, 300,400, 500, and 600 (E in Fig. 2). Aliquots of eachtreatment were recovered for survival analysis, as de-scribed previously. In order to compare its survivalprofile with other biological models, aliquots of UVC-tolerant Deinococcus radiodurans and UV-sensitive E.coli K12 wild-type strain (StrR) [8] were also submittedto the same protocol. Results were plotted on a graphshowing survival curves.
Molecular Identification of UV-Tolerant Isolates
Total genomic DNA was extracted from colonies using theMicrobial DNA Isolation Kit (Mo Bio Laboratories, SolanoBeach, CA, USA) according to the manufacturer's instruc-tions. 16S rDNA sequences present in the total extractedDNA were amplified using universal oligonucleotide pri-mers 515Fw (5′-GTG CCA GCA GCC GCG GTA A-3′)and 1492Rv (5′-GGT TAC CTT GTT ACG ACT T-3′). Forthe amplification of template DNA, the GoTaq colorlessMaster Mix (Promega Corporation, Madison, WI, USA)was used according to the manufacturer's instructions.PCR conditions were as follows: an initial denaturing stepat 94 °C for 5 min followed by four consecutive cycles of94 °C for 1 min, n degrees Celsius for 1 min, and 72 °C for1 min, in which n048–49–50–51 °C. This was then fol-lowed by 31 cycles of 94 °C for 1 min, 52 °C for1 min, 72 °C for 1 min, and a final extension step at72 °C for 15 min, thus totaling 35 cycles of amplifica-tion. PCR products were then separately digested withrestriction enzymes AluI and MspI. PCR products withunique restriction patterns were stored and ligated to thepGEM-T Easy Vector System (Promega Corporation,Madison, WI, USA), and cloned in E. coli XL1-Bluecells. The resulting plasmid vectors were isolated andpurified using the Invisorb Spin Plasmid Mini Two(Invitek GmbH, Berlin, Germany) according to the man-ufacturer's instructions. The automated sequencing ofthe clones was conducted by Macrogen DNA Sequenc-ing Inc. (Seoul, South Korea) using the M13 forwardprimer site of the pGEM-T Easy Vector. Strains withunique 16S rDNA restriction patterns were identifiedand maintained on LB or Marine Broth agar plates. Toidentify the closest relatives of the queried 16S rRNAgene sequences, the nucleotide sequence of the isolated16S rRNA genes were analyzed using the Mega BLASToption for highly similar sequences of the BLASTn algorithmagainst the National Centre for Biotechnology Informationnonredundant database (www.ncbi.nlm.nih.gov).Phylogeny was assigned by comparing the maximumidentity values of known species in the list of resultswith the corresponding new clone.
DGGE Analysis of Soil Communities
Total genomic DNA was extracted from 0.5 g of soil/biofilm from each site using the FastDNA Spin Kit forsoil (Qbiogene, Carlsbad, CA) following the manufac-turer's instructions. A 16S rRNA gene fragmentcorresponding to nucleotide positions 968–1401 (E. colinumbering) was amplified using the universal bacterialprimers: 968f (5′-AAC GCG AAG AAC CTT AC-3′),which contains a 40-bp GC clamp (5′-CGC CCG CCGCGC GCG GCG GGC GGG GCG GGG GCA CGGGGG G-3′) attached to its 5′ end, making it suitable fordenaturing gradient gel electrophoresis (DGGE) and1401r (5′-CGG TGT GTA CAA GAC CC-3′). ThePCR mixture consisted of 1 μl DNA (10–30 ng), 25pmol of each primer, 5 μl 10× PCR buffer (Fermentas),2 .5 mM MgCl2 , 2 .5 U Taq DNA polymerase(Fermentas), 0.2 mM of each deoxynucleoside triphos-phate (Promega), 1 % formamide, 5 μg bovine serumalbumin (BSA) and sterile, filtered MilliQ water to afinal volume of 50 μl. Negative controls consisted ofsterile MilliQ water instead of sample. PCR amplifica-tion was performed in a DNA thermocycler (Master-cycler Personal, Eppendorf, Hamburg, Germany). Thetemperature profile included an initial denaturation stepat 94 °C for 2 min, 35 cycles of a denaturation step at94 °C for 1 min, a primer annealing step at 55 °C for1 min, and an extension step at 72 °C for 2 min,followed by a final step of 72 °C for 10 min. BeforeDGGE analysis, the presence of PCR products wasconfirmed by electrophoresis in a 1.2 % agarose gelrun at 80 V in Tris–borate–EDTA buffer. The gel wasstained for 15 min with 0.5 μg.ml−1 ethidium bromideand viewed under short-wavelength UV. A 100-bp DNAladder (Fermentas) served as the molecular sizestandard.
DGGE of the PCR products generated with the 968f-GC/1401r primer set was performed using the DCodeuniversal mutation detection system (Bio-Rad DCode,Richmond, VA, USA) at 75 V and 60 °C for 16 h in0.5× TAE buffer. PCR products (30 μl) were loaded on6 % (w/v) polyacrylamide gels containing a lineargradient ranging from 45 to 65 % denaturant (100 %denaturant corresponded to 7 M urea and 40 % (v/v)formamide), increasing in the direction of electrophore-sis. A 10-ml stacking gel without denaturant was addedon top. After electrophoresis, the gels were stainedwith SYBR green I nucleic acid gel stain (MolecularProbes, Leiden, The Netherlands) for 40 min and werethen scanned using a Storm PhosphorImager (Amer-sham Biosciences, Uppsala, Sweden). Analysis of theDGGE profiles was performed using the BioNumericsversion 5.10 software package (Applied Maths). A
UVC-Tolerant Bacteria from the Hyperarid Atacama Desert 329
dendrogram was constructed using Pearson correlationcoefficients (r) and cluster analysis performed by theunweighted pair group method with average linkages(UPGMA).
Results
Microbial Enumerations
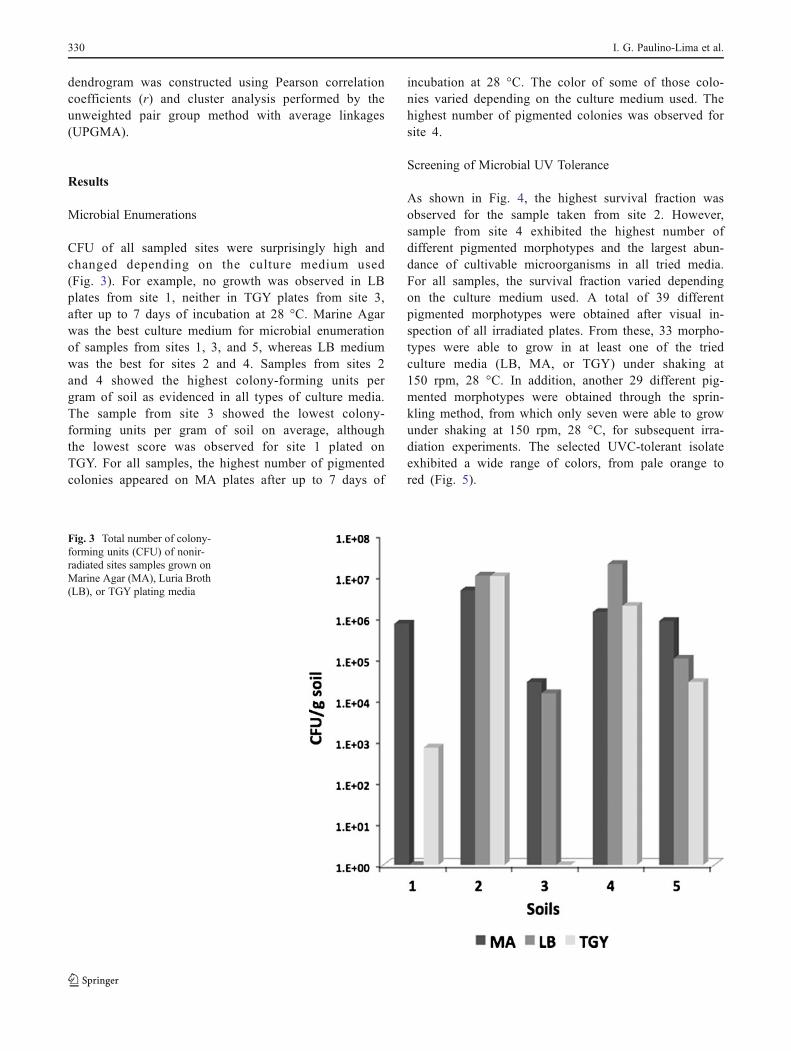
CFU of all sampled sites were surprisingly high andchanged depending on the culture medium used(Fig. 3). For example, no growth was observed in LBplates from site 1, neither in TGY plates from site 3,after up to 7 days of incubation at 28 °C. Marine Agarwas the best culture medium for microbial enumerationof samples from sites 1, 3, and 5, whereas LB mediumwas the best for sites 2 and 4. Samples from sites 2and 4 showed the highest colony-forming units pergram of soil as evidenced in all types of culture media.The sample from site 3 showed the lowest colony-forming units per gram of soil on average, althoughthe lowest score was observed for site 1 plated onTGY. For all samples, the highest number of pigmentedcolonies appeared on MA plates after up to 7 days of
incubation at 28 °C. The color of some of those colo-nies varied depending on the culture medium used. Thehighest number of pigmented colonies was observed forsite 4.
Screening of Microbial UV Tolerance
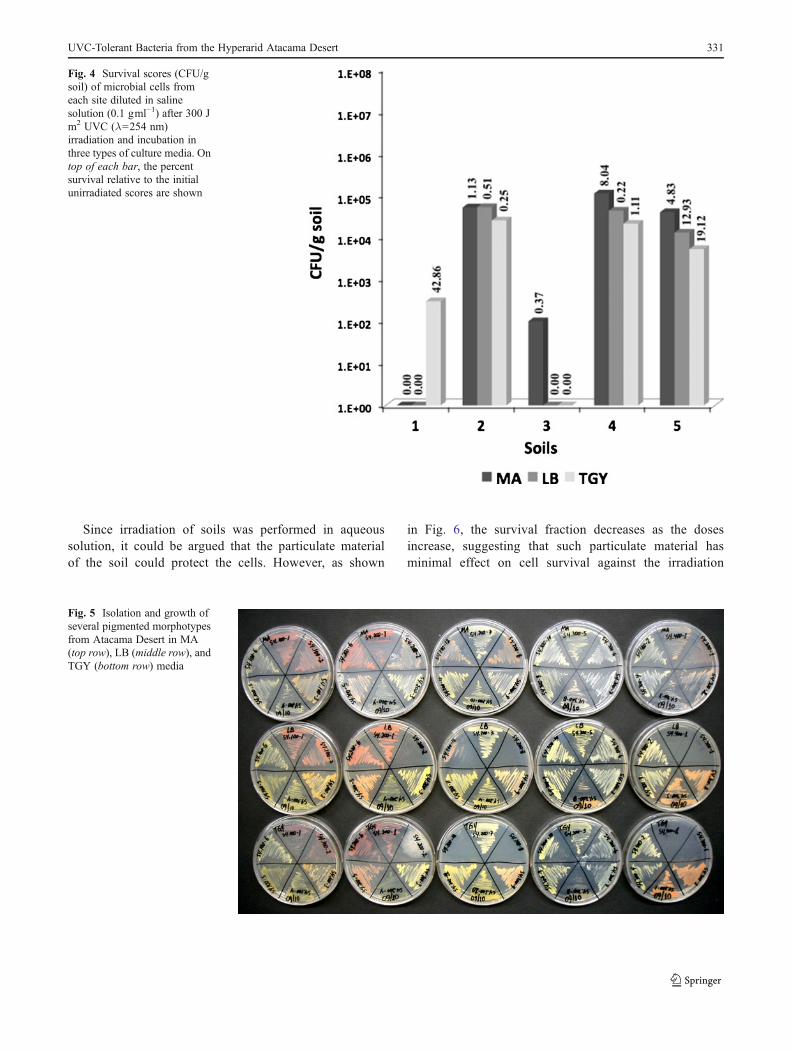
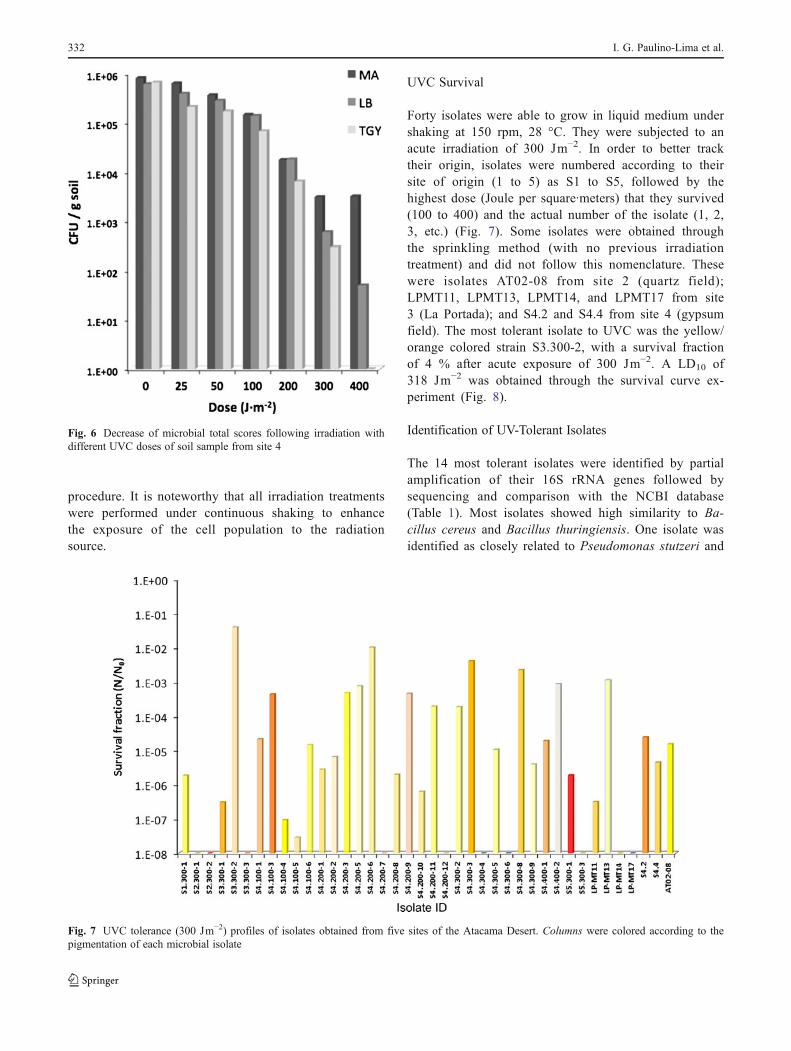
As shown in Fig. 4, the highest survival fraction wasobserved for the sample taken from site 2. However,sample from site 4 exhibited the highest number ofdifferent pigmented morphotypes and the largest abun-dance of cultivable microorganisms in all tried media.For all samples, the survival fraction varied dependingon the culture medium used. A total of 39 differentpigmented morphotypes were obtained after visual in-spection of all irradiated plates. From these, 33 morpho-types were able to grow in at least one of the triedculture media (LB, MA, or TGY) under shaking at150 rpm, 28 °C. In addition, another 29 different pig-mented morphotypes were obtained through the sprin-kling method, from which only seven were able to growunder shaking at 150 rpm, 28 °C, for subsequent irra-diation experiments. The selected UVC-tolerant isolateexhibited a wide range of colors, from pale orange tored (Fig. 5).
Fig. 3 Total number of colony-forming units (CFU) of nonir-radiated sites samples grown onMarine Agar (MA), Luria Broth(LB), or TGY plating media
330 I. G. Paulino-Lima et al.
Since irradiation of soils was performed in aqueoussolution, it could be argued that the particulate materialof the soil could protect the cells. However, as shown
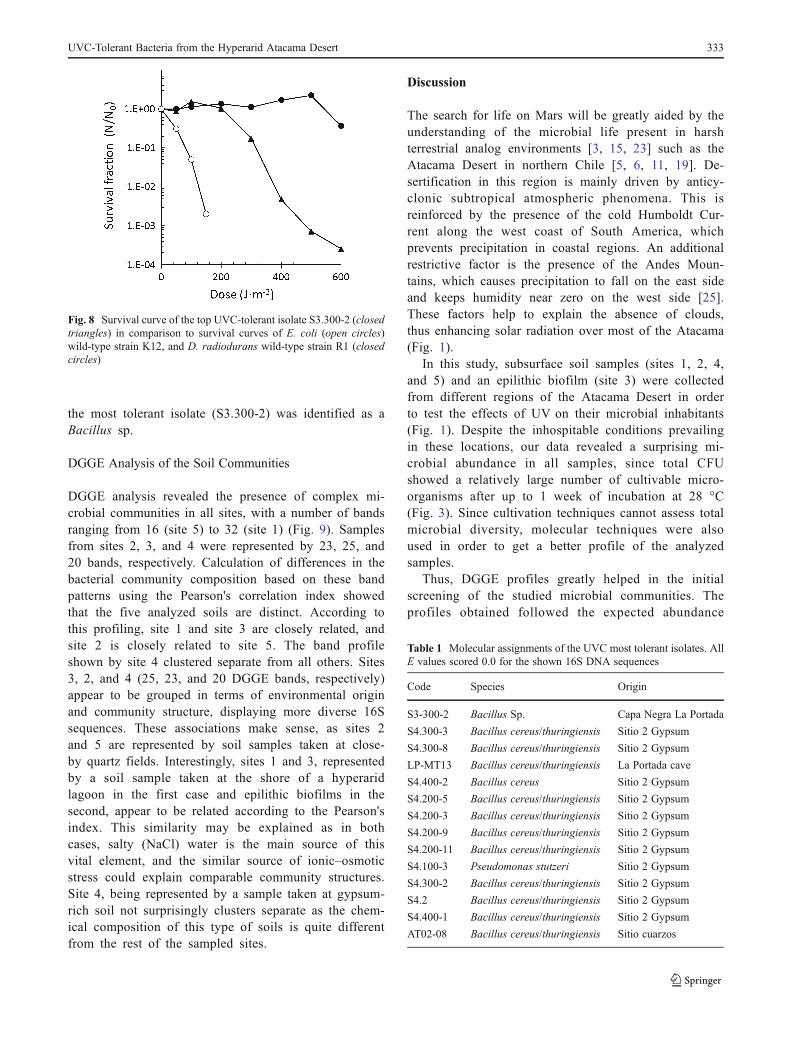
in Fig. 6, the survival fraction decreases as the dosesincrease, suggesting that such particulate material hasminimal effect on cell survival against the irradiation
Fig. 4 Survival scores (CFU/gsoil) of microbial cells fromeach site diluted in salinesolution (0.1 gml−1) after 300 Jm2 UVC (λ0254 nm)irradiation and incubation inthree types of culture media. Ontop of each bar, the percentsurvival relative to the initialunirradiated scores are shown
Fig. 5 Isolation and growth ofseveral pigmented morphotypesfrom Atacama Desert in MA(top row), LB (middle row), andTGY (bottom row) media
UVC-Tolerant Bacteria from the Hyperarid Atacama Desert 331
procedure. It is noteworthy that all irradiation treatmentswere performed under continuous shaking to enhancethe exposure of the cell population to the radiationsource.
UVC Survival
Forty isolates were able to grow in liquid medium undershaking at 150 rpm, 28 °C. They were subjected to anacute irradiation of 300 Jm−2. In order to better tracktheir origin, isolates were numbered according to theirsite of origin (1 to 5) as S1 to S5, followed by thehighest dose (Joule per square·meters) that they survived(100 to 400) and the actual number of the isolate (1, 2,3, etc.) (Fig. 7). Some isolates were obtained throughthe sprinkling method (with no previous irradiationtreatment) and did not follow this nomenclature. Thesewere isolates AT02-08 from site 2 (quartz field);LPMT11, LPMT13, LPMT14, and LPMT17 from site3 (La Portada); and S4.2 and S4.4 from site 4 (gypsumfield). The most tolerant isolate to UVC was the yellow/orange colored strain S3.300-2, with a survival fractionof 4 % after acute exposure of 300 Jm−2. A LD10 of318 Jm−2 was obtained through the survival curve ex-periment (Fig. 8).
Identification of UV-Tolerant Isolates
The 14 most tolerant isolates were identified by partialamplification of their 16S rRNA genes followed bysequencing and comparison with the NCBI database(Table 1). Most isolates showed high similarity to Ba-cillus cereus and Bacillus thuringiensis. One isolate wasidentified as closely related to Pseudomonas stutzeri and
Fig. 6 Decrease of microbial total scores following irradiation withdifferent UVC doses of soil sample from site 4
Fig. 7 UVC tolerance (300 Jm−2) profiles of isolates obtained from five sites of the Atacama Desert. Columns were colored according to thepigmentation of each microbial isolate
332 I. G. Paulino-Lima et al.
the most tolerant isolate (S3.300-2) was identified as aBacillus sp.
DGGE Analysis of the Soil Communities
DGGE analysis revealed the presence of complex mi-crobial communities in all sites, with a number of bandsranging from 16 (site 5) to 32 (site 1) (Fig. 9). Samplesfrom sites 2, 3, and 4 were represented by 23, 25, and20 bands, respectively. Calculation of differences in thebacterial community composition based on these bandpatterns using the Pearson's correlation index showedthat the five analyzed soils are distinct. According tothis profiling, site 1 and site 3 are closely related, andsite 2 is closely related to site 5. The band profileshown by site 4 clustered separate from all others. Sites3, 2, and 4 (25, 23, and 20 DGGE bands, respectively)appear to be grouped in terms of environmental originand community structure, displaying more diverse 16Ssequences. These associations make sense, as sites 2and 5 are represented by soil samples taken at close-by quartz fields. Interestingly, sites 1 and 3, representedby a soil sample taken at the shore of a hyperaridlagoon in the first case and epilithic biofilms in thesecond, appear to be related according to the Pearson'sindex. This similarity may be explained as in bothcases, salty (NaCl) water is the main source of thisvital element, and the similar source of ionic–osmoticstress could explain comparable community structures.Site 4, being represented by a sample taken at gypsum-rich soil not surprisingly clusters separate as the chem-ical composition of this type of soils is quite differentfrom the rest of the sampled sites.
Discussion
The search for life on Mars will be greatly aided by theunderstanding of the microbial life present in harshterrestrial analog environments [3, 15, 23] such as theAtacama Desert in northern Chile [5, 6, 11, 19]. De-sertification in this region is mainly driven by anticy-clonic subtropical atmospheric phenomena. This isreinforced by the presence of the cold Humboldt Cur-rent along the west coast of South America, whichprevents precipitation in coastal regions. An additionalrestrictive factor is the presence of the Andes Moun-tains, which causes precipitation to fall on the east sideand keeps humidity near zero on the west side [25].These factors help to explain the absence of clouds,thus enhancing solar radiation over most of the Atacama(Fig. 1).
In this study, subsurface soil samples (sites 1, 2, 4,and 5) and an epilithic biofilm (site 3) were collectedfrom different regions of the Atacama Desert in orderto test the effects of UV on their microbial inhabitants(Fig. 1). Despite the inhospitable conditions prevailingin these locations, our data revealed a surprising mi-crobial abundance in all samples, since total CFUshowed a relatively large number of cultivable micro-organisms after up to 1 week of incubation at 28 °C(Fig. 3). Since cultivation techniques cannot assess totalmicrobial diversity, molecular techniques were alsoused in order to get a better profile of the analyzedsamples.
Thus, DGGE profiles greatly helped in the initialscreening of the studied microbial communities. Theprofiles obtained followed the expected abundance
Fig. 8 Survival curve of the top UVC-tolerant isolate S3.300-2 (closedtriangles) in comparison to survival curves of E. coli (open circles)wild-type strain K12, and D. radiodurans wild-type strain R1 (closedcircles)
Table 1 Molecular assignments of the UVC most tolerant isolates. AllE values scored 0.0 for the shown 16S DNA sequences
Code Species Origin
S3-300-2 Bacillus Sp. Capa Negra La Portada
S4.300-3 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.300-8 Bacillus cereus/thuringiensis Sitio 2 Gypsum
LP-MT13 Bacillus cereus/thuringiensis La Portada cave
S4.400-2 Bacillus cereus Sitio 2 Gypsum
S4.200-5 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.200-3 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.200-9 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.200-11 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.100-3 Pseudomonas stutzeri Sitio 2 Gypsum
S4.300-2 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.2 Bacillus cereus/thuringiensis Sitio 2 Gypsum
S4.400-1 Bacillus cereus/thuringiensis Sitio 2 Gypsum
AT02-08 Bacillus cereus/thuringiensis Sitio cuarzos
UVC-Tolerant Bacteria from the Hyperarid Atacama Desert 333
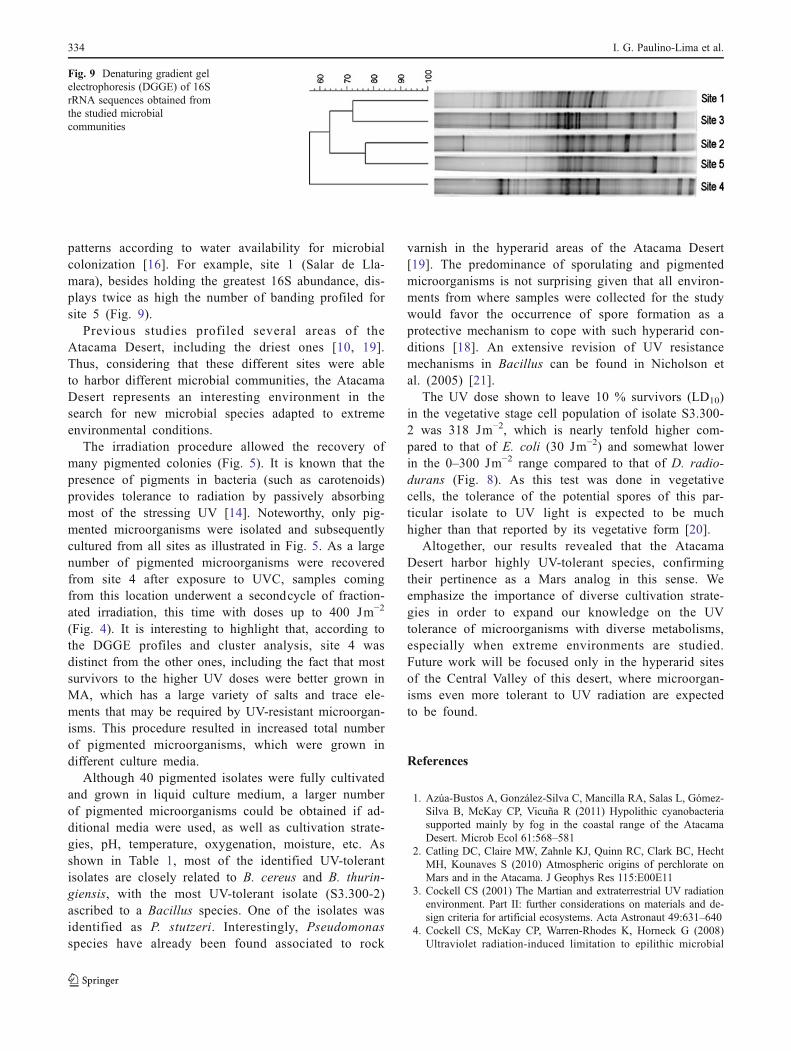
patterns according to water availability for microbialcolonization [16]. For example, site 1 (Salar de Lla-mara), besides holding the greatest 16S abundance, dis-plays twice as high the number of banding profiled forsite 5 (Fig. 9).
Previous studies profiled several areas of theAtacama Desert, including the driest ones [10, 19].Thus, considering that these different sites were ableto harbor different microbial communities, the AtacamaDesert represents an interesting environment in thesearch for new microbial species adapted to extremeenvironmental conditions.
The irradiation procedure allowed the recovery ofmany pigmented colonies (Fig. 5). It is known that thepresence of pigments in bacteria (such as carotenoids)provides tolerance to radiation by passively absorbingmost of the stressing UV [14]. Noteworthy, only pig-mented microorganisms were isolated and subsequentlycultured from all sites as illustrated in Fig. 5. As a largenumber of pigmented microorganisms were recoveredfrom site 4 after exposure to UVC, samples comingfrom this location underwent a secondcycle of fraction-ated irradiation, this time with doses up to 400 Jm−2
(Fig. 4). It is interesting to highlight that, according tothe DGGE profiles and cluster analysis, site 4 wasdistinct from the other ones, including the fact that mostsurvivors to the higher UV doses were better grown inMA, which has a large variety of salts and trace ele-ments that may be required by UV-resistant microorgan-isms. This procedure resulted in increased total numberof pigmented microorganisms, which were grown indifferent culture media.
Although 40 pigmented isolates were fully cultivatedand grown in liquid culture medium, a larger numberof pigmented microorganisms could be obtained if ad-ditional media were used, as well as cultivation strate-gies, pH, temperature, oxygenation, moisture, etc. Asshown in Table 1, most of the identified UV-tolerantisolates are closely related to B. cereus and B. thurin-giensis, with the most UV-tolerant isolate (S3.300-2)ascribed to a Bacillus species. One of the isolates wasidentified as P. stutzeri. Interestingly, Pseudomonasspecies have already been found associated to rock
varnish in the hyperarid areas of the Atacama Desert[19]. The predominance of sporulating and pigmentedmicroorganisms is not surprising given that all environ-ments from where samples were collected for the studywould favor the occurrence of spore formation as aprotective mechanism to cope with such hyperarid con-ditions [18]. An extensive revision of UV resistancemechanisms in Bacillus can be found in Nicholson etal. (2005) [21].
The UV dose shown to leave 10 % survivors (LD10)in the vegetative stage cell population of isolate S3.300-2 was 318 Jm−2, which is nearly tenfold higher com-pared to that of E. coli (30 Jm−2) and somewhat lowerin the 0–300 Jm−2 range compared to that of D. radio-durans (Fig. 8). As this test was done in vegetativecells, the tolerance of the potential spores of this par-ticular isolate to UV light is expected to be muchhigher than that reported by its vegetative form [20].
Altogether, our results revealed that the AtacamaDesert harbor highly UV-tolerant species, confirmingtheir pertinence as a Mars analog in this sense. Weemphasize the importance of diverse cultivation strate-gies in order to expand our knowledge on the UVtolerance of microorganisms with diverse metabolisms,especially when extreme environments are studied.Future work will be focused only in the hyperarid sitesof the Central Valley of this desert, where microorgan-isms even more tolerant to UV radiation are expectedto be found.
References
1. Azúa-Bustos A, González-Silva C, Mancilla RA, Salas L, Gómez-Silva B, McKay CP, Vicuña R (2011) Hypolithic cyanobacteriasupported mainly by fog in the coastal range of the AtacamaDesert. Microb Ecol 61:568–581
2. Catling DC, Claire MW, Zahnle KJ, Quinn RC, Clark BC, HechtMH, Kounaves S (2010) Atmospheric origins of perchlorate onMars and in the Atacama. J Geophys Res 115:E00E11
3. Cockell CS (2001) The Martian and extraterrestrial UV radiationenvironment. Part II: further considerations on materials and de-sign criteria for artificial ecosystems. Acta Astronaut 49:631–640
4. Cockell CS, McKay CP, Warren-Rhodes K, Horneck G (2008)Ultraviolet radiation-induced limitation to epilithic microbial
Fig. 9 Denaturing gradient gelelectrophoresis (DGGE) of 16SrRNA sequences obtained fromthe studied microbialcommunities
334 I. G. Paulino-Lima et al.

growth in arid deserts—dosimetric experiments in the hyperaridcore of the Atacama Desert. J Photochem Photobiol B Biol 90:79–87
5. Connon SA, Lester ED, Shafaat HS, Obenhuber DC, Ponce A(2007) Bacterial diversity in hyperarid Atacama Desert soils. JGeophys Res Biogeosci 112:G04S17. doi:10.1029/2006JG000311
6. Dávila AF, Gómez-Silva B, de los Rios A, Ascaso C, Olivares H,McKay CP CP, Wierzchos J (2008) Facilitation of endolithicmicrobial survival in the hyperarid core of the Atacama Desertby mineral deliquescence. J Geophys Res 113:G01028
7. Demergasso C, Chong G, Galleguillos P, Escudero L, Martínez-Alonso M, Esteve I (2003) Tapetes microbianos del Salar DeLlamará, norte de Chile. Rev Chil Hist Nat 76:485–499, Spanish
8. DeWitt SK, Adelbeg EA (1962) The occurrence of a genetictransposition in a strain of Escherichia coli. Genetics 47:577–585
9. Dose K, Bieger-Dose A, Ernst B, Feister U, Gómez-Silva B, KleinA, Risi S, Stridde C (2001) Survival of microorganisms under theextreme conditions of the Atacama Desert. Orig Life Evol Biosph31:287–303
10. Drees KP, Neilson JW, Betancourt JL, Quade J, Henderson DA,Pryor BM, Maier RM (2006) Bacterial community structure in thehyperarid core of the Atacama Desert, Chile. Appl Environ Micro-biol 72:7902–7908
11. Gorbushina AA (2007) Life on the rocks. Environ Microbiol9:1613–1631
12. Hartley A, Chong G, Houston J, Mather A (2005) 150 millionyears of climatic stability: evidence from the Atacama Desert,northern Chile. J Geol Soc Lond 162:421–424
13. Houston J, Hartley AJ (2003) The central Andean west-sloperainshadow and its potential contribution to the origin of hyper-aridity in the Atacama Desert. Int J Climatol 23:1453–1464.doi:10.1002/joc.938
14. Jacobs JL, Carroll TL, Sundin GW (2005) The role of pigmenta-tion, ultraviolet radiation tolerance, and leaf colonization strategiesin the epiphytic survival of phyllosphere bacteria. Microb Ecol49:104–113
15. Kuhlman KR, Allenbach LB, Ball CL, Fusco WG, La Duc MT,Kuhlman GM, Anderson RC, Stuecker T, Erickson IK, BenardiniJ, Crawford RL (2005) Enumeration, isolation, and characteriza-tion of ultraviolet (UV-C) resistant bacteria from rock varnish inthe Whipple Mountains, California. Icarus 174:585–595
16. McKay CP, Friedmann EI, Gómez-Silva B, Cáceres L, AndersenDT, Landheim R (2003) Temperature and moisture conditions forlife in the extreme arid region of the Atacama Desert: four years of
observations including the El Niño of 1997–1998. Astrobiology3:393–406
17. Meunier D, Sternberg R, Mettetal F, Buch A, Coscia D, Szopa C,Rodier C, Coll P, Cabanec M, Raulin F (2007) A laboratory pilotfor in situ analysis of refractory organic matter in Martian soil bygas chromatography mass spectrometry. Adv Space Res 39:337–344
18. Myasnik M, Manasherob R, Ben-Dov E, Zaritsky A, Margalith Y,Barak Z (2001) Comparative sensitivity to UV-B radiation of twoBacillus thuringiensis subspecies and other Bacillus sp. CurrMicrobiol 43:140–143
19. Navarro-González R, Rainey FA, Molina P, Bagaley DR, HollenBJ, de la Rosa J, Small AM, Quinn RC, Grunthaner FJ, Cáceres L,Gomez-Silva B, McKay CP (2003) Mars-like soils in the AtacamaDesert, Chile, and the dry limit of microbial life. Science302:1018–1021
20. Nicholson WL, Munakata N, Horneck G, Melosh HJ, Setlow P(2000) Resistance of Bacillus endospores to extreme terrestrial andextraterrestrial environments. Microbiol Mol Biol Rev 64:548–572
21. Nicholson WL, Schuerger AC, Setlow P (2005) The solar UVenvironment and bacterial spore UV resistance: considerationsfor Earth-to-Mars transport by natural processes and human space-flight. Mutat Res 571:249–264
22. Phoenix VR, Bennett PC, Engel AS, Tyler SW, Ferris FG (2006)Chilean high-altitude hot-spring sinters: a model system for UVscreening mechanisms by early Precambrian Cyanobacteria. Geo-biology 4:15–28
23. Schuerger AC, Mancinelli RL, Kern RG, Rothschild LJ, McKayCP (2003) Survival of endospores of Bacillus subtilis on spacecraftsurfaces under simulated Martian environments: implications forthe forward contamination of Mars. Icarus 165:253–276
24. Skelley AM, Scherer JR, Aubrey AD, Grover WH, Ivester RH,Ehrenfreund P, Grunthaner FJ, Bada JL, Mathies RA (2005) De-velopment and evaluation of a microdevice for amino acid bio-marker detection and analysis on Mars. Proc Natl Acad Sci U S A102(4):1041–1046
25. Thompson RD (1975) The climatology of the arid world. Univer-sity of Reading, Department of Geography, Geographical Papers,No. 35, p 39
26. Warren-Rhodes KA, Rhodes KL, Pointing SB, Ewing SA, LacapDC, Gómez-Silva B, Amundson R, Friedmann EI, McKay CP(2006) Hypolithic cyanobacteria, dry limit of photosynthesis, andmicrobial ecology in the hyperarid Atacama Desert. Microb Ecol52:389–398
UVC-Tolerant Bacteria from the Hyperarid Atacama Desert 335





























