Embed Size (px)
Citation preview

http://france.elsevier.com/direct/GEOBIO/
Geobios 39 (2006) 372–384
* Auteur correspondant.Adresse e-mail : gomm
0016-6995/$ - see front madoi:10.1016/j.geobios.2005.
La phalange distale du pouce d’Orrorin tugenensis
(Miocène supérieur du Kenya)The terminal thumb phalanx of Orrorin tugenensis
(Upper Miocene of Kenya)Dominique Gommery a,b,*, Brigitte Senut c
aUPR 2147 du CNRS, 44, rue de l’amiral-Mouchez, 75014 Paris, FrancebHOPE (Human Origins and Past Environments), Transvaal Museum, Prétoria, Afrique du Sud
cDépartement « Histoire de la Terre », Muséum national d’Histoire naturelle, USM 0203-UMR 5143 du CNRS,
Case Postale 38, 57, rue Cuvier, 75231 Paris cedex 05, FranceReçu le 14 juin 2004 ; accepté le 8 mars 2005Disponible sur internet le 09 mars 2006
Résumé
La quasi-totalité des articles traitant de l’anatomie du pouce chez les hominidés plio-pléistocènes est focalisée sur la capacité à manipuler et àfabriquer des outils tout en négligeant l’aspect fonctionnel locomoteur. Or, chez ces hominidés, le membre antérieur joue encore un rôle dans lalocomotion. Certains de leurs caractères anatomiques classiquement associés à la manipulation et/ou la fabrication d’outils sont déjà présents auMiocène supérieur chez Orrorin tugenensis comme le prouve la phalange de pouce (BAR 1901’01). Cette dernière apporte donc des informationsessentielles : elle permet de conclure que la morphologie du pouce n’est probablement pas liée uniquement à la manufacture d’outils, maispourrait plutôt refléter une adaptation à une saisie fine nécessaire pour équilibrer le corps dans les mouvements de grimper chez des animauxfréquemment bipèdes et différents de ceux des grands singes.© 2006 Elsevier SAS. Tous droits réservés.
Abstract
The lion’s share of articles dealing with the thumb anatomy of Plio-Pleistocene hominids has focussed on the capacity to manipulate andmanufacture tools, and has largely neglected the locomotor aspects. However, in these hominids, the forelimb was still employed in locomotion.Certain of the anatomical characters classically associated with manipulation and/or fabrication of tools are already present in the Late Miocenespecies Orrorin tugenensis as shown by the terminal thumb phalanx BAR 1901’01. This specimen reveals crucial information suggesting thatthumb morphology is not exclusively related to such tool using and manufacturing activities but reflects in a frequently bipedal creature a deeperadaptation concerning the precision grip essential for climbing and balancing, different from that of apes.© 2006 Elsevier SAS. Tous droits réservés.
Mots clés : Orrorin tugenensis ; Hominidé ; Phalange distale du pouce ; Miocène supérieur ; Kapsomin ; Kenya
Keywords: Orrorin tugenensis; Hominid; Thumb distal phalanx; Upper Miocene; Kapsomin; Kenya
[email protected] (D. Gommery).
tter © 2006 Elsevier SAS. Tous droits réservés.03.002

Fig. 1. Vue dorsale de BAR 1901’01.Fig. 1. Dorsal view of BAR 1901’01.
D. Gommery, B. Senut / Geobios 39 (2006) 372–384 373
1. Introduction
Le site de Kapsomin se trouve dans la Formation de Lukei-no (Pickford et Senut, 2001) dont l’âge, estimé par radiométrieet paléomagnétisme, est compris entre 5,9 et 5,8 millions d’an-nées (Sawada et al., 2002), confirmant l’âge biochronologique(Pickford, 1974, 1975 ; Pickford et Senut, 2001). Depuis 2000,près d’une vingtaine de restes d’hominidés, attribués à Orrorintugenensis y ont été découverts (Senut et al., 2001). Les quel-ques restes postcrâniens connus suggèrent que cet hominidéétait bipède, tout en conservant des adaptations au grimpercomme l’indiquent son humérus et sa phalange proximale demain (Pickford et al., 2002 ; Senut et al., 2001, 2002).
Durant la saison de fouille de l’automne 2001 à Kapsomin,plusieurs fossiles d’hominidés ont été découverts dont une pha-lange distale de pouce (BAR 1901’01) parfaitement conservée.Le même niveau a livré des restes dentaires d’un hominoïdenon humain proche en taille et en morphologie d’un gorille(Pickford et Senut, 2005). La phalange par sa gracilité et samorphologie presque humaine est rapportée à Orrorin tugenen-sis (Gommery, 2003 ; Gommery et Senut, 2002a, 2002b).
Les phalanges distales du pollex soulèvent de nombreusesquestions sur les liens entre la morphologie osseuse et les fonc-tions de manipulation et éventuellement de fabrication d’outilspar les différentes espèces d’Hominidés plio-pléistocènes(Bimson et al., 1997 ; Napier, 1962, 1993 ; Marzke, 1997 ;Marzke et Shackley, 1986 ; Panger et al., 2002 ; Stern et Sus-man, 1983 ; Shrewsbury et al., 2003 ; Shrewsbury et Johnson,1983 ; Smith, 1995, 2000 ; Susman, 1979, 1988a, 1988b,1998). Les fossiles sont généralement comparés aux hommesmodernes, aux grands singes africains et aux babouins, plusrarement à l’orang-outang (Shrewsbury et al., 2003) et jamaisaux hominoïdes miocènes. L’aspect locomoteur n’est pas envi-sagé par les auteurs précédents alors que les premiers homini-dés pratiquaient une bipédie associée au grimper (Senut, 1981 ;Stern et Susman, 1983 ; Stern, 2000). La main et donc le pouceétaient sollicités lors des déplacements dans les arbres. Lapièce de Kapsomin permet aujourd’hui d’éclairer le débat.
2. Méthode et matériel
Les phalanges distales de pouce restent rares dans les col-lections d’hominoïdes fossiles. Pour le Miocène, on n’enconnaît que deux attribuées à Oreopithecus bambolii d’Italie(Moyà-Solà et al., 1999) et plusieurs de Rusinga au Kenya ap-partenant au genre Proconsul (Begun et al., 1994). Napier etDavis en 1959 ont décrit la main de Proconsul de Rusinga« KNM RU 2036 », mais il n’existe pas de phalange terminalepolliciale ; même chose pour Walker et Pickford, 1983. Pour lePlio-Pléistocène, deux phalanges sud-africaines sont rapportéesà des australopithèques : Stw 294 et SKX 5016 dont les origi-naux ont pu être étudiés. Stw 294, rapporté à Australopithecusafricanus provient du membre 4 du site de Sterkfontein enAfrique du Sud, daté de 2,5 à 2,6 millions d’années (Collectionde la Medical School de l’Université de Witwatersrand à Jo-hannesburg en Afrique du Sud) (Ricklan, 1987, 1988). SKX
5016, rapporté à Paranthropus robustus découvert dans lemembre 1 du Site de Swartkrans est daté d’environ 1,8 milliond’années (Collection du Transvaal Museum à Pretoria enAfrique du Sud) (Susman, 1988a, 1988b). Le squelette proba-blement complet Stw 573 du membre 2 de Sterkfontein récem-ment annoncé (Clarke, 2002) possède une main complète quin’est pas encore accessible à l’étude, mais nous faisons réfé-rence aux caractères décrits. La phalange distale d’OH 7 (Na-pier, 1962 ; Susman et Creel, 1979) n’est pas incluse ici, carson attribution anatomique reste problématique. Elle appartient,en effet, à un sujet sub-adulte ; en outre, mal conservée, elle nepermet pas des mesures fiables. Susman et Creel (1979) n’ontpas pu trancher sur son attribution à une phalange de pied oude main, et nous n’avons pas pu lever l’incertitude.
Des comparaisons anatomiques ont été effectuées avec desgrands singes africains provenant des collections du laboratoired’Anatomie Comparée du Muséum National d’Histoire Natu-relle.
3. Description de BAR 1901’01
3.1. Identification
L’identification des phalanges terminales isolées de main oude pied n’est pas toujours aisée chez les grands singes. Toute-fois, la morphologie particulière de BAR 1901’01 suggèrequ’elle appartient à un pouce : phalange aplatie, allongée, detaille relativement importante et non trapue (comme c’est le casde la phalange distale du gros orteil), robustesse et aspect ar-rondi de la surface dorsale de la diaphyse (qui l’isole des pha-langes distales d’orteils ou des autres doigts), une surface arti-culaire basse et large.
3.2. Description
Il s’agit d’une phalange complète, non déformée d’un poucedroit de grandes dimensions (L : 18,8 mm et l : 11 mm)(Figs. 1– 4).
Pour faciliter la lecture, nous adoptons la terminologie sui-vante pour les bords latéraux de la phalange : bord radial (bordgauche de la phalange en vue dorsale) et bord ulnaire (borddroit de la phalange en vue dorsale).

Fig. 3. Vue proximale de BAR 1901’01.Fig. 3. Proximal view of BAR 1901’01.
Fig. 2. Vue palmaire de BAR 1901’01.Fig. 2. Palmar view of BAR 1901’01.
D. Gommery, B. Senut / Geobios 39 (2006) 372–384374
3.2.1. Vue dorsale (Figs. 1,4)L’extrémité distale ou apex (Tuberositas phalangis distalis)
est développée, robuste, large et en forme de « fer à cheval ».La diaphyse (Corpus phalangis distalis) est convexe transver-salement. Proximalement, on note un fort renflement, à la baseduquel s’insère le muscle long extenseur du pouce (m. extensorpollicis longus). L’extrémité proximale de la phalange est pluslarge que la distale. Elle est encadrée par deux renflements (unradial et un ulnaire) correspondant aux tubercules basaux. Lebord proximal présente deux concavités liées aux deux partiesde la facette articulaire situées sur la face proximale. Ces der-nières sont séparées par un renflement convexe formé parl’éminence de la base de la phalange (Eminentia basis phalan-gis). La partie ulnaire est la plus longue et la plus inclinée parrapport à l’axe longitudinal de la phalange ; ce qui permet d’at-tribuer la pièce au côté droit (Shrewsbury et al., 2003). Dansune dépression bien marquée, allongée transversalement lon-geant le bord dorsal de l’articulation, s’insèrent les ligamentslatéraux (Ligg. collateralia) qui participent à l’articulation entrela phalange distale et la phalange proximale. Des deux côtés decette dépression, on distingue deux tubercules basaux d’impor-tance inégale : l’ulnaire, érodé, est plus développé transversa-lement que le radial plus allongé proximodistalement. Ces tu-bercules reçoivent les ligaments palmaires (Ligg. palmaria) etinterosseux latéraux (ces derniers s’attachent sur des excrois-sances osseuses développées à la partie distale des tubercules).
3.2.2. Vue palmaire (Figs. 2,4)La tubérosité apicale (Tuberositas phalangis distalis), bien
développée en vue palmaire est asymétrique : plus renflée ducôté radial et plus allongée du côté ulnaire. Son bord proximalsurplombe la diaphyse. Ses bords radial et ulnaire portent des
épines unguéales érodées ; l’ulnaire s’étendant un peu plus bassur l’os que la radiale.
Le corps de la phalange est divisé en deux par un fort ren-flement transversal : distalement, on trouve une fossette allon-gée, la fossette unguéale (Shrewsbury et al., 2003), située sousla tubérosité apicale et proximalement une grande dépressionventrale qui reçoit le m. flexor pollicis longus (muscle longfléchisseur propre du pouce) dont la zone principale d’insertionest située sur le renflement lui-même (Susman, 1998). Dans ladépression, on remarque un surcreusement arrondi dans la par-tie radiale qui pourrait être dû à la fossilisation ou bien à unacte de prédation. Sur le bord proximal, du côté radial commeulnaire, on trouve les tubercules basaux où s’insèrent les liga-ments palmaires (Ligg. palmaria).
3.2.3. Vues radiale et ulnaire (Fig. 4)La diaphyse conserve une épaisseur constante sur toute sa
longueur ; la partie médiane étant renforcée par le renflementlié à l’insertion du long fléchisseur du pouce. La tubérositéapicale, asymétrique est légèrement inclinée dorsalement.
Des deux tubercules basaux, le radial est le mieux conservé.Légèrement plus développé dorsoventralement que l’ulnaire, ilfait légèrement saillie dorsalement. Sur le bord radial, on ob-serve une excroissance osseuse allongée bien développée quicorrespond à l’insertion du ligament interosseux latéral. Envue radiale, une fine crête étirée le long du bord palmaire reliel’épine unguéale au tubercule basal. Elle reçoit le ligament in-terosseux latéral et se confond proximalement avec le bord ra-dial de la fossette du m. flexor pollicis longus.
En vue ulnaire, une petite crête mousse, où s’insère le liga-ment interosseux latéral, est visible sur le bord palmaire.
3.2.4. Vue proximale (Figs. 3,4)La surface d’articulation proximale (Facies articularis ossis
basis phalangis) est allongée transversalement ; son bord ven-tral est presque rectiligne alors que le dorsal est bombé. Elle estdivisée en deux par une crête assez nette, l’éminence de la basede la phalange (Eminentia basis phalangis) et forme deux sur-faces faiblement inclinées vers la face palmaire, la radialel’étant toutefois plus nettement que l’ulnaire ; l’ulnaire carréeest la plus haute et la radiale la plus concave en raison de lasaillie de son bord externe.
4. Les données métriques
Les mesures linéaires confirment les observations sur lesdimensions générales (Tableaux 1,2). En effet, la stature esti-mée d’O. tugenensis est d’environ 1,50 m ce qui correspond àla taille d’un chimpanzé femelle (Senut et al., 2001).
L’aspect effilé de BAR 1901’01 et celui plus massif deSKX 5016 sont confirmés par l’indice de forme exprimé parle rapport « Longueur totale/Largeur base » qui est respective-ment de 1,71 et de 1,48 (Tableau 3). L’homme présente unevaleur moyenne de 1,55, le chimpanzé de 2,15 et l’orang-ou-tang se rapproche de celle de l’homme (1,50). Shrewsbury et
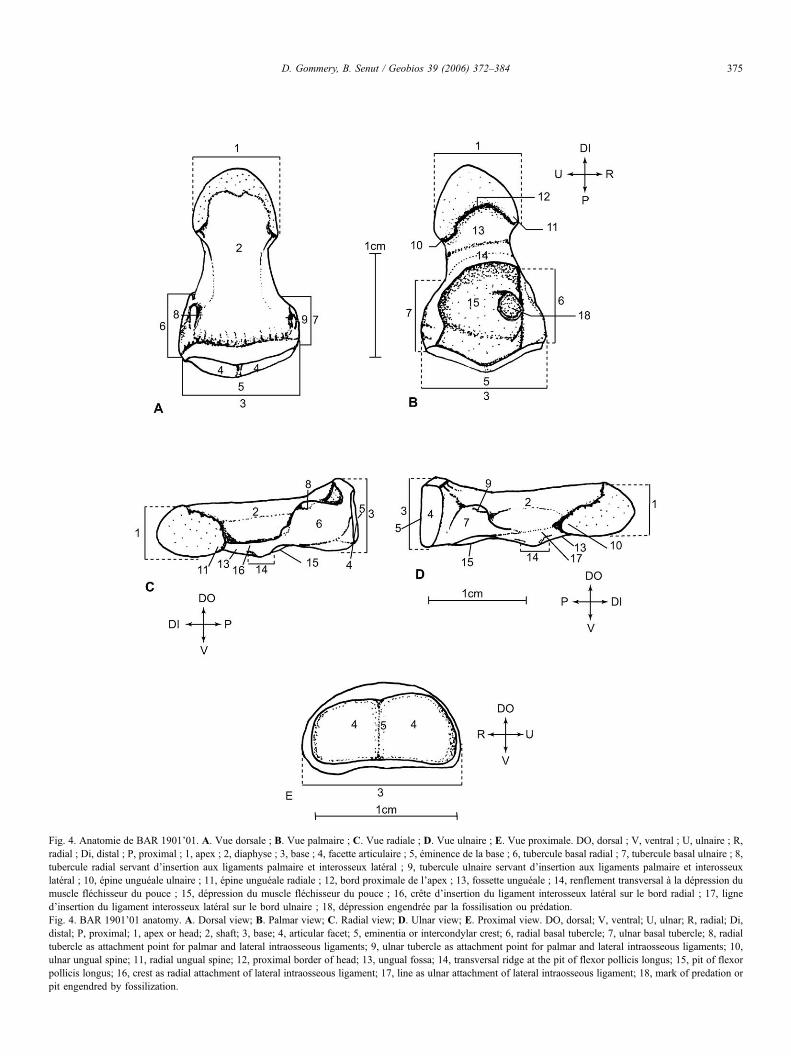
Fig. 4. Anatomie de BAR 1901’01. A. Vue dorsale ; B. Vue palmaire ; C. Vue radiale ; D. Vue ulnaire ; E. Vue proximale. DO, dorsal ; V, ventral ; U, ulnaire ; R,radial ; Di, distal ; P, proximal ; 1, apex ; 2, diaphyse ; 3, base ; 4, facette articulaire ; 5, éminence de la base ; 6, tubercule basal radial ; 7, tubercule basal ulnaire ; 8,tubercule radial servant d’insertion aux ligaments palmaire et interosseux latéral ; 9, tubercule ulnaire servant d’insertion aux ligaments palmaire et interosseuxlatéral ; 10, épine unguéale ulnaire ; 11, épine unguéale radiale ; 12, bord proximale de l’apex ; 13, fossette unguéale ; 14, renflement transversal à la dépression dumuscle fléchisseur du pouce ; 15, dépression du muscle fléchisseur du pouce ; 16, crête d’insertion du ligament interosseux latéral sur le bord radial ; 17, ligned’insertion du ligament interosseux latéral sur le bord ulnaire ; 18, dépression engendrée par la fossilisation ou prédation.Fig. 4. BAR 1901’01 anatomy. A. Dorsal view; B. Palmar view; C. Radial view; D. Ulnar view; E. Proximal view. DO, dorsal; V, ventral; U, ulnar; R, radial; Di,distal; P, proximal; 1, apex or head; 2, shaft; 3, base; 4, articular facet; 5, eminentia or intercondylar crest; 6, radial basal tubercle; 7, ulnar basal tubercle; 8, radialtubercle as attachment point for palmar and lateral intraosseous ligaments; 9, ulnar tubercle as attachment point for palmar and lateral intraosseous ligaments; 10,ulnar ungual spine; 11, radial ungual spine; 12, proximal border of head; 13, ungual fossa; 14, transversal ridge at the pit of flexor pollicis longus; 15, pit of flexorpollicis longus; 16, crest as radial attachment of lateral intraosseous ligament; 17, line as ulnar attachment of lateral intraosseous ligament; 18, mark of predation orpit engendred by fossilization.
D. Gommery, B. Senut / Geobios 39 (2006) 372–384 375
D. Gommery, B. Senut / Geobios 39 (2006) 372–384376
al. (2003) ont obtenu des valeurs plus faibles pour des primatesquadrupèdes terrestres comme Papio anubis (1,31) et Papiohamadryas (1,21). Smith (1995) rappelle que si le rapport entrela longueur et la largeur change pour un os, les forces des brasde levier changent, et donc les fonctions. La largeur de l’apexde BAR 1901’01 (7,5 mm), est faible par rapport à celle desautres hominidés (homme compris) et des grands singes. Tou-tefois, le rapport « Largeur de l’apex/Largeur de la base » estde 68,18 chez BAR 1901’01 (Tableau 3), une valeur proche de
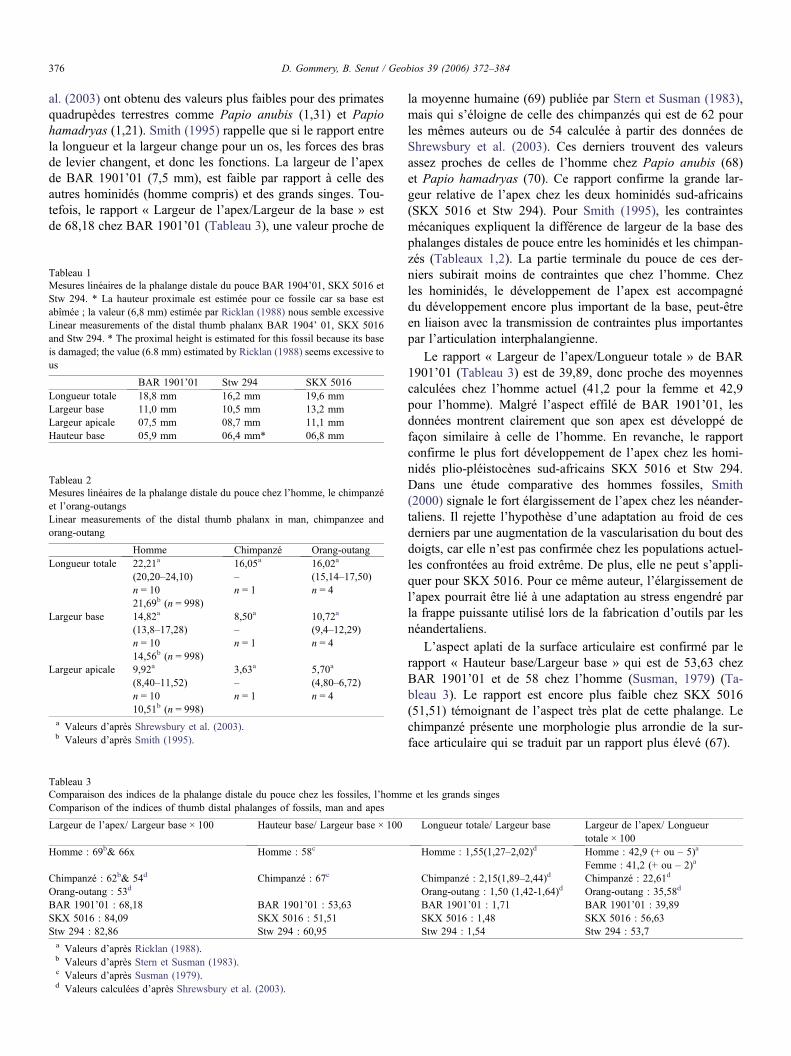
Tableau 1Mesures linéaires de la phalange distale du pouce BAR 1904’01, SKX 5016 etStw 294. * La hauteur proximale est estimée pour ce fossile car sa base estabîmée ; la valeur (6,8 mm) estimée par Ricklan (1988) nous semble excessiveLinear measurements of the distal thumb phalanx BAR 1904’ 01, SKX 5016and Stw 294. * The proximal height is estimated for this fossil because its baseis damaged; the value (6.8 mm) estimated by Ricklan (1988) seems excessive tous
BAR 1901’01 Stw 294 SKX 5016Longueur totale 18,8 mm 16,2 mm 19,6 mmLargeur base 11,0 mm 10,5 mm 13,2 mmLargeur apicale 07,5 mm 08,7 mm 11,1 mmHauteur base 05,9 mm 06,4 mm* 06,8 mm
Tableau 2Mesures linéaires de la phalange distale du pouce chez l’homme, le chimpanzéet l’orang-outangsLinear measurements of the distal thumb phalanx in man, chimpanzee andorang-outang
Homme Chimpanzé Orang-outangLongueur totale 22,21a 16,05a 16,02a
(20,20–24,10) – (15,14–17,50)n = 10 n = 1 n = 421,69b (n = 998)
Largeur base 14,82a 8,50a 10,72a
(13,8–17,28) – (9,4–12,29)n = 10 n = 1 n = 414,56b (n = 998)
Largeur apicale 9,92a 3,63a 5,70a
(8,40–11,52) – (4,80–6,72)n = 10 n = 1 n = 410,51b (n = 998)
a Valeurs d’après Shrewsbury et al. (2003).b Valeurs d’après Smith (1995).
Tableau 3Comparaison des indices de la phalange distale du pouce chez les fossiles, l’hommComparison of the indices of thumb distal phalanges of fossils, man and apes
Largeur de l’apex/ Largeur base × 100 Hauteur base/ Largeur base × 100
Homme : 69b& 66x Homme : 58c
Chimpanzé : 62b& 54d Chimpanzé : 67c
Orang-outang : 53d
BAR 1901’01 : 68,18 BAR 1901’01 : 53,63SKX 5016 : 84,09 SKX 5016 : 51,51Stw 294 : 82,86 Stw 294 : 60,95a Valeurs d’après Ricklan (1988).b Valeurs d’après Stern et Susman (1983).c Valeurs d’après Susman (1979).d Valeurs calculées d’après Shrewsbury et al. (2003).
la moyenne humaine (69) publiée par Stern et Susman (1983),mais qui s’éloigne de celle des chimpanzés qui est de 62 pourles mêmes auteurs ou de 54 calculée à partir des données deShrewsbury et al. (2003). Ces derniers trouvent des valeursassez proches de celles de l’homme chez Papio anubis (68)et Papio hamadryas (70). Ce rapport confirme la grande lar-geur relative de l’apex chez les deux hominidés sud-africains(SKX 5016 et Stw 294). Pour Smith (1995), les contraintesmécaniques expliquent la différence de largeur de la base desphalanges distales de pouce entre les hominidés et les chimpan-zés (Tableaux 1,2). La partie terminale du pouce de ces der-niers subirait moins de contraintes que chez l’homme. Chezles hominidés, le développement de l’apex est accompagnédu développement encore plus important de la base, peut-êtreen liaison avec la transmission de contraintes plus importantespar l’articulation interphalangienne.
Le rapport « Largeur de l’apex/Longueur totale » de BAR1901’01 (Tableau 3) est de 39,89, donc proche des moyennescalculées chez l’homme actuel (41,2 pour la femme et 42,9pour l’homme). Malgré l’aspect effilé de BAR 1901’01, lesdonnées montrent clairement que son apex est développé defaçon similaire à celle de l’homme. En revanche, le rapportconfirme le plus fort développement de l’apex chez les homi-nidés plio-pléistocènes sud-africains SKX 5016 et Stw 294.Dans une étude comparative des hommes fossiles, Smith(2000) signale le fort élargissement de l’apex chez les néander-taliens. Il rejette l’hypothèse d’une adaptation au froid de cesderniers par une augmentation de la vascularisation du bout desdoigts, car elle n’est pas confirmée chez les populations actuel-les confrontées au froid extrême. De plus, elle ne peut s’appli-quer pour SKX 5016. Pour ce même auteur, l’élargissement del’apex pourrait être lié à une adaptation au stress engendré parla frappe puissante utilisé lors de la fabrication d’outils par lesnéandertaliens.
L’aspect aplati de la surface articulaire est confirmé par lerapport « Hauteur base/Largeur base » qui est de 53,63 chezBAR 1901’01 et de 58 chez l’homme (Susman, 1979) (Ta-bleau 3). Le rapport est encore plus faible chez SKX 5016(51,51) témoignant de l’aspect très plat de cette phalange. Lechimpanzé présente une morphologie plus arrondie de la sur-face articulaire qui se traduit par un rapport plus élevé (67).
e et les grands singes
Longueur totale/ Largeur base Largeur de l’apex/ Longueurtotale × 100
Homme : 1,55(1,27–2,02)d Homme : 42,9 (+ ou – 5)a
Femme : 41,2 (+ ou – 2)a
Chimpanzé : 2,15(1,89–2,44)d Chimpanzé : 22,61d
Orang-outang : 1,50 (1,42-1,64)d Orang-outang : 35,58d
BAR 1901’01 : 1,71 BAR 1901’01 : 39,89SKX 5016 : 1,48 SKX 5016 : 56,63Stw 294 : 1,54 Stw 294 : 53,7
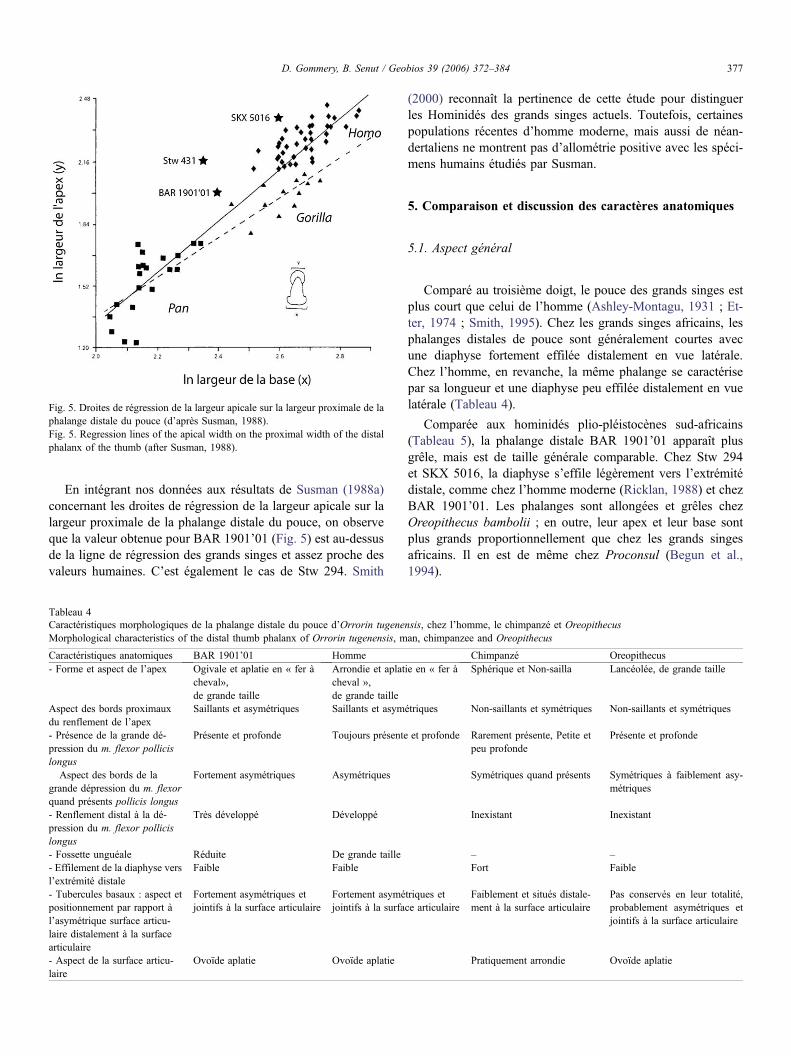
Fig. 5. Droites de régression de la largeur apicale sur la largeur proximale de laphalange distale du pouce (d’après Susman, 1988).Fig. 5. Regression lines of the apical width on the proximal width of the distalphalanx of the thumb (after Susman, 1988).
D. Gommery, B. Senut / Geobios 39 (2006) 372–384 377
En intégrant nos données aux résultats de Susman (1988a)concernant les droites de régression de la largeur apicale sur lalargeur proximale de la phalange distale du pouce, on observeque la valeur obtenue pour BAR 1901’01 (Fig. 5) est au-dessusde la ligne de régression des grands singes et assez proche desvaleurs humaines. C’est également le cas de Stw 294. Smith
Tableau 4Caractéristiques morphologiques de la phalange distale du pouce d’Orrorin tugeneMorphological characteristics of the distal thumb phalanx of Orrorin tugenensis, m
Caractéristiques anatomiques BAR 1901’01 Homme- Forme et aspect de l’apex Ogivale et aplatie en « fer à
cheval»,de grande taille
Arrondie et aplatcheval »,de grande taille
Aspect des bords proximauxdu renflement de l’apex
Saillants et asymétriques Saillants et asym
- Présence de la grande dé-pression du m. flexor pollicislongus
Présente et profonde Toujours présente
Aspect des bords de lagrande dépression du m. flexorquand présents pollicis longus
Fortement asymétriques Asymétriques
- Renflement distal à la dé-pression du m. flexor pollicislongus
Très développé Développé
- Fossette unguéale Réduite De grande taille- Effilement de la diaphyse versl’extrémité distale
Faible Faible
- Tubercules basaux : aspect etpositionnement par rapport àl’asymétrique surface articu-laire distalement à la surfacearticulaire
Fortement asymétriques etjointifs à la surface articulaire
Fortement asyméjointifs à la surfa
- Aspect de la surface articu-laire
Ovoïde aplatie Ovoïde aplatie
(2000) reconnaît la pertinence de cette étude pour distinguerles Hominidés des grands singes actuels. Toutefois, certainespopulations récentes d’homme moderne, mais aussi de néan-dertaliens ne montrent pas d’allométrie positive avec les spéci-mens humains étudiés par Susman.
5. Comparaison et discussion des caractères anatomiques
5.1. Aspect général
Comparé au troisième doigt, le pouce des grands singes estplus court que celui de l’homme (Ashley-Montagu, 1931 ; Et-ter, 1974 ; Smith, 1995). Chez les grands singes africains, lesphalanges distales de pouce sont généralement courtes avecune diaphyse fortement effilée distalement en vue latérale.Chez l’homme, en revanche, la même phalange se caractérisepar sa longueur et une diaphyse peu effilée distalement en vuelatérale (Tableau 4).
Comparée aux hominidés plio-pléistocènes sud-africains(Tableau 5), la phalange distale BAR 1901’01 apparaît plusgrêle, mais est de taille générale comparable. Chez Stw 294et SKX 5016, la diaphyse s’effile légèrement vers l’extrémitédistale, comme chez l’homme moderne (Ricklan, 1988) et chezBAR 1901’01. Les phalanges sont allongées et grêles chezOreopithecus bambolii ; en outre, leur apex et leur base sontplus grands proportionnellement que chez les grands singesafricains. Il en est de même chez Proconsul (Begun et al.,1994).
nsis, chez l’homme, le chimpanzé et Oreopithecusan, chimpanzee and Oreopithecus
Chimpanzé Oreopithecusie en « fer à Sphérique et Non-sailla Lancéolée, de grande taille
étriques Non-saillants et symétriques Non-saillants et symétriques
et profonde Rarement présente, Petite etpeu profonde
Présente et profonde
Symétriques quand présents Symétriques à faiblement asy-métriques
Inexistant Inexistant
– –Fort Faible
triques etce articulaire
Faiblement et situés distale-ment à la surface articulaire
Pas conservés en leur totalité,probablement asymétriques etjointifs à la surface articulaire
Pratiquement arrondie Ovoïde aplatie

Tableau 5Caractéristiques morphologiques de la phalange distale du pouce BAR 1901’01, comparaison avec les hominidés fossiles sud-africains (Stw 294 et SKX 5016)Morphological characteristics of the distal thumb phalanx BAR 1901’ 01, comparison with South African hominids (Stw 294 and SKX 5016)
Ressemblances Différences- Forme de l’apex aplatie en « fer à cheval » comme chez l’homme et nonsphérique comme chez les grands singes. Le bord proximal saillant surplombeprésente un décrochement la diaphyse. L’apex est symétrique et est inclinédorsalement en vue latérale.
- L’extrémité de la partie distale est plus robuste et à contour ogival chez BAR1901’01.- Largeur plus réduite de l’apex chez BAR 1901’01.
- Grande dépression du m. flexor pollicis longus et ses bords sont aymétriques. - L’asymétrie est plus importante sur BAR 1901’01. Le bord radial forme une crête.- Existence distalement d’un renflement à la dépression du m. flexor pollicislongus et d’une fossette unguéale réduite.
- Distalement, le renflement transversal de la dépression du m. flexor pollicislongus est plus robuste chez BAR 1901’01.
- La diaphyse s’effile légèrement vers l’extrémité distale.- Les tubercules basaux sont asymétriques et la surface articulaire est aplatie. - L’insertion des ligaments interosseux sur les tubercules basaux est très forte.
D. Gommery, B. Senut / Geobios 39 (2006) 372–384378
5.2. Tubérosité apicale
La morphologie de la tubérosité apicale est utilisée classi-quement en systématique pour déterminer l’appartenance d’unephalange terminale de pouce à un taxon précis. Chez l’homme,la forme aplatie en « fer à cheval » est distincte de la formesphérique de celle des chimpanzés (Tableau 4). Le bord proxi-mal saillant de l’apex surplombe la diaphyse chez l’homme,mais non chez le chimpanzé (Ricklan, 1988). Chez l’homme,la tubérosité apicale est inclinée dorsalement en vue latéralealors qu’elle est dans le prolongement de la diaphyse chez lechimpanzé. La forme en « fer à cheval » est une conséquencede la saillie du rebord proximal de l’apex qui surplombe la
Fig. 6. Vue dorsale de Stw 294 avec le moulage de BAR 1901’01.Fig. 6. Dorsal view of Stw 294 with the cast of BAR 1901’01.
Fig. 7. Vue palmaire de Stw 294 avec le moulage de BAR 1901’01.Fig. 7. Palmar view of Stw 294 with the cast of BAR 1901’01.
diaphyse en vue palmaire, l’asymétrie doit y jouer probable-ment aussi un rôle.
Dans la collection de Hadar (Bush et al., 1982 ; Stern etSusman, 1983), il n’existe pas de phalange distale depouce mais des autres doigts et elles présentent les mêmescaractères généraux. La tubérosité apicale est arrondie en formede sphère aplatie et ne montre pas de saillie du bord proximalau-dessus de la diaphyse (Ricklan, 1988). En vue radiale ouulnaire, la tubérosité apparaît située dans le même plan quecelui de la diaphyse voire légèrement inclinée. Il est doncprobable que la phalange terminale chez les Hominidés deHadar présentait la même morphologie que celle des autresdoigts.
Fig. 8. Vue dorsale de SKX 5016 avec le moulage de BAR 1901’01.Fig. 8. Dorsal view of SKX 5016 with the cast of BAR 1901’01.
Fig. 9. Vue palmaire de SKX 5016 avec le moulage de BAR 1901’01.Fig. 9. Palmar view of SKX 5016 with the cast of BAR 1901’01.

D. Gommery, B. Senut / Geobios 39 (2006) 372–384 379
Chez Stw 294 (Australopithecus africanus) (Ricklan, 1988 ;Susman, 1998) (Figs. 6,7) et SKX 5016 (Paranthropus robus-tus) (Susman, 1988a, 1998) (Figs. 8,9), l’apex est large, aplativoire spatulé et est incliné dorsalement (Tableau 5). Sur la pha-lange distale du pouce de Stw 573, Clarke (2002) note la formelarge et spatulée de la phalange distale. Chez BAR 1901’01, latubérosité apicale présente une forme de « fer à cheval » àcontour ogival. Elle est robuste et moyennement inclinée dor-salement en vue latérale ; toutefois, l’apex pourrait être un peuplus incliné, car masqué par la robustesse de l’os. Son bordproximal surplombe la diaphyse comme c’est aussi le cas chezSKX 5016 et Stw 294 (Tableau 5) mais ne le semble pas surStw 573.
Chez les Hominidés, l’asymétrie de la tubérosité est nette ;la partie ulnaire étant plus étendue proximalement. Elle in-fluence aussi le développement des épines unguéales quandelles existent. Leur asymétrie reflète, en effet, celle de la tubé-rosité apicale. L’asymétrie se retrouve chez BAR 1901’01, oùle bord ulnaire est plus allongé proximalement, mais devait êtreabsente chez les spécimens d’Hadar (d’après la morphologiesphérique) et chez Stw 573. Les spécimens d’O. bambolii,BA#130 et IGF 11778 gauches figurés dans Moyà-Solà et al.(1999) présentent un apex lancéolé (Tableau 4). La saillieproximale de la tubérosité semble absente. L’asymétrie nesemble pas exister chez au moins un des spécimens d’O. bam-bolii, il est plus difficile de conclure pour l’autre spécimen(IGF 11778 gauche) car une partie de l’apex manque. Ce der-nier est arrondi chez deux spécimens sub-adultes de Proconsul,mais pas aussi allongé que celui des autres phalanges termina-les (Begun et al., 1994). Ceci suggérerait que l’apex de la pha-lange de pouce de ces hominoïdes était plus allongé.
5.3. La dépression du m. flexor pollicis longus et ses partiesannexes
5.3.1. Dépression du m. flexor pollicis longusLa présence d’une dépression du m. flexor pollicis longus
est classiquement considérée comme un caractère humain. Ce-pendant, elle est variable chez les primates actuels : on latrouve occasionnellement chez les grands singes et chez d’au-tres primates non hominoïdes (Shrewsbury et al., 2003 ; et ob-servations personnelles). Mais elle est toutefois plus fréquentechez Papio (Shrewsbury et al., 2003). Quand elle existe, elleest en général peu profonde et limitée. En outre, Susman(1998) a montré chez l’homme l’association entre la fosse etle m. flexor pollicis longus (généralement distinct du m. flexordigitorum profundus), et l’absence ou la présence rudimentairede celui-ci chez les grands singes. Toutefois, chez Papio où lafosse est fréquente, le m. flexor pollicis longus n’est pas diffé-rencié du m. flexor digitorum profundus (Shrewsbury et al.,2003).
Chez Stw 294 (Fig. 7), SKX 5016 (Fig. 9), Stw 531 et BAR1901’01 (Fig. 2), elle est grande, profonde et occupe une bonnepartie de la face palmaire de la diaphyse. Sur les deux spéci-mens d’Oreopithecus bambolii (BA#130 ; IGF 11778 gauche),elle est grande et semble profonde (Moyà-Solà et al., 1999).
Begun et al. (1994) ne la signalent que sur une phalange distale(PH 96) de Proconsul chez qui ils décrivent une fosse profondepour le tendon du long fléchisseur (Begun et al., 1994; page115) : « Ventrally, there is a sharp median keel and a deep pitto the left for the long flexor tendon ».
L’asymétrie du bord radial de la dépression du m. flexorpollicis longus est marquée chez SKX 5016 et Stw 294, maiselle l’est encore plus chez Stw 573 (Clarke, 2002 ; Fig. 8) etbien plus encore chez BAR 1901’01 où le relief du bord radial,plus développé que sur le bord ulnaire, forme une crête. L’asy-métrie est présente mais moins prononcée chez l’homme.Shrewsbury et al. (2003) ont noté l’asymétrie de l’insertiondu tendon m. flexor pollicis longus chez l’homme.
5.3.2. Renflement distal à la dépression du m. flexor pollicislongus
Susman (1998) a montré que la différence essentielle entrel’homme et le chimpanzé réside non pas seulement dans laprésence d’une dépression du m. flexor pollicis longus, maisaussi dans l’existence d’un renflement transversal qui bordedistalement la dépression chez l’homme (Tableau 4). Il s’agiten fait de la zone d’insertion principale du tendon du m. flexorpollicis longus (Susman, 1998 ; Shrewsbury et al., 2003). Lechimpanzé et l’homme présentent la même zone d’insertionprincipale pour ce muscle mais elle s’exprime différemmentpar sa nature et par l’impact sur la morphologie osseuse. Lerenflement est marqué chez Stw 294 (Fig. 7), SKX 5016(Fig. 9) et particulièrement chez BAR 1901’01 (Fig. 2), où ilest très robuste (Tableau 5). Il semble faible chez Stw 573 quiest érodé (Clarke, 2002 ; Fig. 8 et la photo de couverture). Ilparaît inexistant chez Oreopithecus bambolii et n’est pas décritchez Proconsul. Sa présence est une caractéristique d’homi-nidé ; il ne se retrouve chez aucun primate non humain.
5.3.3. Fossette unguéale
Cette fossette est définie par la présence du pont osseuxtransversal et elle n’existe que chez les hominidés. Elle estgrande chez l’homme et beaucoup plus réduite chez les fossilessud-africains et BAR 1901’01 (Tableaux 4,5).
5.4. Les tubercules basaux
Chez l’homme, les tubercules basaux, situés proximalement,semblent jointifs à la surface articulaire ; ils sont asymétriques,l’ulnaire étant le plus développé transversalement (Tableau 5).Chez les chimpanzés, ces tubercules presque symétriques, sontplacés plus distalement par rapport à la surface articulaire quechez l’homme (Moyà-Solà et al., 1999) ; ce qui confère unaspect de bobine à la phalange distale des grands singes. ChezBAR 1901’01, mais aussi chez les hominidés sud-africains etchez Oreopithecus bambolii, ils sont situés proximalement etjointifs à la surface articulaire. Le tubercule basal ulnaire estplus développé transversalement que le radial chez les homini-dés fossiles, comme chez l’homme actuel. Chez Stw 573, Clar-ke (2002) note la puissance des insertions musculaires (et liga-

D. Gommery, B. Senut / Geobios 39 (2006) 372–384380
mentaires) en liaison avec le développement des tubercules ba-saux. BAR 1901’01 présente des excroissances osseuses surses tubercules basaux où s’insèrent les ligaments interosseuxlatéraux. Les phalanges terminales, dont celle du pouce, deProconsul sont décrites comme larges dans leur partie proxi-male (Begun et al., 1994). Ceci est dû à la présence de tuber-cules basaux plus développés que chez les hominidés et proba-blement jointifs à la surface articulaire.
5.5. Morphologie de la surface articulaire
La partie proximale de la phalange distale de pouce étantaplatie chez l’homme, la surface articulaire est ovoïde très apla-tie ou allongée (Tableau 4). Elle présente une inclinaison plusdistale de la partie ulnaire, une partie radiale plus concave etplus inclinée vers la face palmaire (Shrewsbury et al., 2003).Chez les chimpanzés, la surface articulaire proximale est pres-que arrondie avec des reliefs peu prononcés.
La phalange BAR 1901’01 (Fig. 3) montre toutes les carac-téristiques observées chez l’homme, mais la partie radiale estmoins inclinée que chez ce dernier. Toutefois, elle diffère de laphalange humaine par la projection proximale plus prononcéedes bords de la surface articulaire au niveau de l’éminence,visible en vues dorsale (Fig. 1) et palmaire (Fig. 2). Pour cecaractère, elle se rapproche plutôt des bases de phalanges mé-dianes et distales des autres doigts humains. Stw 294 et SKX5016 présentent également cette morphologie.
Chez IGF 11778 gauche d’Oreopithecus bambolii, la sur-face articulaire est allongée et basse, comme chez Proconsul(Begun et al., 1994), qui montre une surface peu profonde etdivisée en deux fosses moins prononcées que sur les phalangesde pied. Chez Proconsul, la partie proximale présente des« becs » au-dessus et au-dessous de l’éminence de la surfacearticulaire, caractère qui est plus estompé chez les Hominidés.
6. Discussion générale et aspect fonctionnel
La main est une extrémité de membre complexe dont lepouce est une composante essentielle. Cette complexité est liéeen partie à son rôle dans la préhension.
On identifie classiquement six types principaux de préhen-sion dont quatre font intervenir le pouce et sa phalange (Ka-pandji, 1971 ; Napier, 1993) :
● la préhension par opposition terminale ou préhension pulpo-unguéale : le pouce et l’index s’opposent par l’extrémité dela pulpe, voire la tranche de l’ongle. Le muscle longfléchisseur propre du pouce stabilise la phalange distale enflexion ;
● la préhension par opposition subterminale : c’est le mode depréhension le plus courant chez l’homme. Le pouce et l’in-dex s’opposent par la face palmaire de la pulpe. L’articula-tion interphalangienne distale peut-être en extension ou blo-quée en demi-flexion ;
● la préhension par opposition subterminolatérale ou préhen-sion pulpolatérale : la face palmaire du pouce s’appuie sur laface externe de la première phalange de l’index ;
● la préhension palmaire à « pleine main » ou à « pleinepaume » : c’est la préhension de force. Le pouce constituela butée unique à la force des quatre autres doigts, et sonefficacité est d’autant plus grande qu’il est plus fléchi. Lemuscle long fléchisseur propre du pouce verrouille la prisegrâce à la flexion de la phalange distale.
Dans la littérature, la majorité des discussions porte surl’importance du m. flexor pollicis longus et son activité (Ham-rick et al., 1998 ; Shrewsbury et al., 2003 ; Susman, 1998)largement impliqué dans plusieurs types de préhension. Chezl’homme, ce muscle présente la particularité d’être distinct dum. flexor digitorum profundus (Susman, 1998) alors qu’il estrudimentaire ou absent chez les grands singes. Il semble plussollicité chez l’homme. Hamrick et al. (1998) soulignent l’im-portance de l’activité électromyographique du m. flexor pollicislongus lors des préhensions fortes comme dans le cas de lafabrication et/ou de l’utilisation d’outils. Ces dernières activitésnécessitent de maintenir la phalange distale du pouce fortementfléchie.
La profonde dépression du m. flexor pollicis longus n’estpas à mettre en relation directe avec l’importance fonctionnellede ce muscle (Susman, 1998). Ce dernier ne s’y insère pas.Chez l’homme, des fibres du tendon de ce dernier vont du ren-flement transversal situé distalement à la dépression du m. fle-xor pollicis longus, correspondant à la zone d’insertion ma-jeure, au bord proximal saillant de la tubérosité distale maisaussi aux épines unguéales et aux tubercules basaux. Ces ca-ractéristiques sont révélatrices de l’importance du muscle.Chez les primates non humains, des fibres partent de l’insertionmajeure du m. flexor pollicis longus pour s’attacher à la tubé-rosité distale et de façon plus variable aux tubercules basaux ouaux épines unguéales quand elles existent. Shrewsbury et al.(2003) ont noté une grande fréquence de l’asymétrie de l’inser-tion de ce muscle chez l’homme et très rarement chez les autresprimates, puisqu’ils ne l’ont retrouvé que chez un orang-ou-tang, qui est un grimpeur lourd et lent. Shrewsbury et al.(2003) ont observé un développement et une insertion plus im-portants de la partie radiale du tendon du m. flexor pollicislongus qui marque radialement l’os de façon prononcée. Cecidiffère de l’insertion symétrique du m. flexor digitorum profun-dus observée sur les phalanges distales de l’index. La fractionde fibre plus importante du côté radial peut améliorer la prona-tion et contrebalancer les charges ainsi que la déformation del’aspect ulnaire de la pulpe durant la préhension comme l’ad-mettent Shrewsbury et al. (2003). Ces derniers suggèrent éga-lement que ce seul caractère semble être un témoin du compor-tement de manipulation chez les espèces fossiles chez qui il yaurait une opposition maximum de la pulpe de la phalangedistale du pouce contre celle des autres doigts. En fait, il s’agitdu même mouvement pour saisir une branche pendant le grim-per d’où la similitude avec l’orang-outang qui pratique réguliè-rement ce mode de locomotion. L’asymétrie des bords radial et

D. Gommery, B. Senut / Geobios 39 (2006) 372–384 381
ulnaire de la dépression du m. flexor pollicis longus est pronon-cée chez Orrorin.
La présence du renflement transversal et l’asymétrie desbords radial et ulnaire de la dépression du m. flexor pollicislongus montrent que l’action de ce dernier permet d’obtenirune opposition maximale de la pulpe du pouce avec les autresdoigts et elles témoignent du stress important enduré par cemuscle.
Shrewsbury et al. (2003) ont observé une compartimenta-tion fonctionnelle de la pulpe du pouce. Il existe une pulpeunguéale proximale grande et mobile et une pulpe unguéaledistale plus réduite, mais plus stable. La limite entre les deuxse situe au niveau de l’insertion principale du m. flexor pollicislongus, donc du renflement transversal. Cette compartimenta-tion se traduit par une différence de consistance de la pulpe quiest plus graisseuse proximalement et plus fibreuse distalement.Elle est située principalement au niveau de la saillie ou sur-plomb du bord proximal de la tubérosité apicale et corresponden grande partie à la fossette unguéale. La compartimentationde la pulpe est associée aux caractéristiques de l’apex, de laface palmaire de la diaphyse et des tubercules basaux. Cheztous les primates non humains actuels, il n’y a pas de compar-timentation de la pulpe unguéale ; en effet, on trouve un tissufibrograisseux indifférencié (Shrewsbury et al., 2003).
Les caractéristiques observées chez l’homme se retrouventchez SKX 5016, Stw 294 et BAR 1901’01. Toutefois, la partiereprésentée par la fossette unguéale est plus restreinte et, parconséquent, la pulpe unguéale distale devait être plus limitée.
Un autre aspect débattu dans la littérature concerne l’aspectet la taille de l’apex de la phalange distale qui est en partieassociée au m. flexor pollicis longus. La largeur de la pulpedépend en partie de la largeur de l’apex de la phalange distale.Pour Shrewsbury et Johnson (1983), une extrémité apicalelarge permet une saisie puissante avec des pressions minimaleset la possibilité de saisir avec précision des objets de grandetaille entre le pouce et les autres doigts. La présence et le rôledes épines unguéales dans la manipulation et dans la fabrica-tion d’outils ont été largement débattus (Marzke, 1997 ; Marz-ke et Shackley, 1986 ; Shrewsbury et al., 2003 ; Shrewsbury etJohnson, 1983 ; Susman, 1998). Les épines sont aussi présen-tes chez le babouin (Susman, 1998), mais chez ces derniers,leur présence ne peut être associée à la fabrication d’outils maisplutôt à la locomotion et à la manipulation d’objets et de nour-riture.
Les ligaments interosseux s’insèrent sur les épines unguéa-les et sur la face dorsale des tubercules basaux. L’épine un-guéale ulnaire de BAR 1901’01 est la plus allongée proxima-lement. Shrewsbury et al. (2003) ont noté que le ligamentinterosseux ulnaire était le plus large chez l’homme et doncqu’il y avait aussi une asymétrie pour ce ligament. Le dévelop-pement asymétrique des épines unguéales et des ligaments in-terosseux est corrélé aux forces importantes exercées sur lapartie proximale de la pulpe du pouce humain du côté ulnaireau cours de la préhension. La pulpe et l’ongle s’attachent aumême endroit sur les ligaments interosseux. Cette asymétriereflète la pression sur la partie ulnaire de la pulpe de la pha-
lange distale durant les manipulations, alors que l’asymétrie dum. flexor pollicis longus permet de contrebalancer la pressionexercée conjointement par de fortes pronations et flexions de laphalange distale, mettant ainsi en contact la plus grande partiede surface palmaire avec l’objet mais aussi en opposant lepouce aux autres doigts afin de manipuler. La tubérosité api-cale de BAR 1901’01 est différente de celle des hominidésplio-pléistocènes et de l’homme actuel, car plus robuste. Tou-tefois, elle est large et asymétrique comme chez eux. L’archi-tecture osseuse de la partie distale de BAR 1901’01 présenteles mêmes caractéristiques que celles des hominidés plio-pléis-tocènes sud-africains et de l’homme actuel. Il devait en être demême pour les tissus mous. Les excroissances osseuses sur lestubercules basaux montrent que les ligaments interosseux laté-raux ont du être très fortement sollicités.
Orrorin tugenensis devait utiliser sa main pour se mouvoirdans les arbres en grimpant. La main est alors employéecomme une pince dans laquelle le pouce est utilisé commepour la préhension palmaire à « pleine main » pour hisserson corps et maintenir l’équilibre, d’une manière différente decelle des grands singes. L’humérus d’Orrorin présente des ca-ractères anatomiques liés au grimper (Senut et al., 2001). Lamorphologie des tubercules basaux de la phalange pollicale,sur lesquels s’insèrent les ligaments interphalangiens et interos-seux latéraux, suggèrent que ces ligaments étaient très sollicitésen particulier lors de la flexion de la phalange distale. Ces tu-bercules et le grand développement du pont osseux de la dé-pression permettent de forts mouvements de flexion–extensionde la première phalange sur la deuxième. Chez les grands sin-ges, le pouce est petit en taille absolue, mais aussi relative encomparaison avec les autres doigts. Les grands singes africainsse déplacent souvent en pratiquant le knuckle-walking. Lepouce est alors moins sollicité et est placé contre le doigt II(Susman, 1979 ; Tuttle, 1967, 1969). Le grimper, mais aussila suspension, sont caractérisés chez les grands singes par unemain qui est utilisée comme un crochet où le pouce n’est passollicité (Napier, 1993 ; Aiello et Dean, 1999).
Les données métriques ont aussi leur importance ; nousavons pu montrer que le rapport de la largeur de l’apex sur lalongueur de BAR 1901’01 est proche de celui de l’homme ac-tuel. La largeur de l’apex augmente proportionnellement à salongueur (allométrie). Les hominidés sud-africains présente-raient une convergence avec les néandertaliens pour la largeurde l’apex (Smith, 1995, 2000). La grande différence concernela largeur relative de la base de BAR 1901’01 et donc du rap-port de la largeur de la base sur la longueur. Chez les homi-nidés, l’augmentation de la largeur de l’apex s’accompagned’une augmentation de la largeur de la base afin de transmettrele stress plus important induit par les contraintes mécaniquespassant par l’articulation interphalangienne. BAR 1901’01pourrait démontrer que cette adaptation ne s’est pas encore pro-duite et/ou que la morphologie des os sous-jacents ne pouvaitle permettre. Les conditions mécaniques s’en trouveraient dif-férentes et expliqueraient assez aisément que les structures os-seuses en relations avec les ligaments interphalangiens et inte-

D. Gommery, B. Senut / Geobios 39 (2006) 372–384382
rosseux latéraux mais aussi en rapport avec le m. flexor pollicislongus soient accentuées.
La partie proximale des phalanges distales de pouceconnues d’hominidés et d’hominoïdes non hominidés fossilesprésentent des caractères communs comme la largeur plus im-portante que la hauteur conférant un aspect aplati à la surfacearticulaire, le développement des tubercules basaux jointifs à lasurface articulaire, la double concavité des bords ventraux etdorsaux de l’extrémité proximale. Le plus fort développementde cette concavité chez Proconsul signalé par Begun et al.(1994) limiterait la mobilité lors des mouvements de flexion–extension dans l’articulation interphalangienne distale. Laconcavité semble plus limitée chez Oreopithecus bambolii ; lamobilité devait être donc plus importante dans l’articulationinterphalangienne et peut-être en rapport avec une vie partiel-lement arboricole.
Récemment, Alba et al. (2003) ont émis l’hypothèque sui-vante : à partir du moment où la locomotion arboricole domi-née par le membre supérieur est devenue de moins en moinsimportante chez des formes adaptées à la bipédie, la morpho-logie humaine de la main serait le produit de la résultante despressions de sélection de manipulation complexe déjà présentechez les primates non humains fossiles. En d’autres termeschez les formes bipèdes la diminution des contraintes biomé-caniques imposées par le grimper arboricole et la suspensionont permis d’améliorer l’efficacité des mouvements de manipu-lation que peuvent effectuer tous les primates non humains,mais moins efficacement car leur main est adaptée à la loco-motion. Mais, Alba et al. (2003) ont basé leur étude sur unecomparaison des proportions de la main de l’hominidé de lalocalité 333 de Hadar qui ne possède pas de phalange terminalede pouce avec celles de mains de grands singes actuels. Lesproportions de la main de certains Hominoïdes du Miocène,comme Proconsul (Napier, 1993), sont différentes de cellesdes grands singes modernes et se rapprochent plus des Cerco-pithecoidea terrestres actuels. En outre, ces derniers montrentdes proportions comparables à celles de l’homme.
Nous ne sommes pas totalement en désaccord avec l’hypo-thèse d’Alba et al. mais Orrorin tugenensis n’est pas un Aus-tralopithèque. L’étude de celui-ci montre qu’il était engagédans une voie différente de celle de ces derniers : Orrorin estun être microdonte (petites dents – grand squelette), et non pasmégadonte (grandes dents – petit squelette) comme le sont lesAustralopithèques (Senut et al., 2001). Ses proportions sontdonc différentes, comme son répertoire locomoteur. L’humé-rus, comme la phalange proximale (longue et courbe) suggè-rent qu’il n’était pas complètement libéré du milieu arboré (Se-nut et al., 2001) ne serait-ce que pour se nourrir ou se protégerdes prédateurs. Les caractères d’hominidés de la phalange dis-tale du pouce d’Orrorin sont pour certains très accentués parrapport à l’homme. On pourrait prétexter qu’ils le sont pourpalier d’autres différences comme la morphologie de la tubéro-sité apicale. Mais cette explication n’est pas valable pour SKX5016. Tout ceci impliquerait indirectement que ces êtres ont lesmêmes capacités manipulatrices que l’homme actuel. D’autrepart, la manipulation est aussi une contrainte fonctionnelle. Il
est bien évident que tous les primates manipulent par exemplepour la recherche d’alimentation ou dans les activités sociales.Dans ce cas, à quoi pouvait servir ce puissant pouce d’Orro-rin ?
Par ailleurs, Orrorin n’a pas nécessairement pratiqué ungrimper identique à celui des grands singes modernes. Les ho-minoïdes miocènes ont été beaucoup plus diversifiés que lesactuels et sans nul doute leurs répertoires locomoteurs. Il y aprobablement des différences dans le grimper arboricole liéesau poids, mais aussi aux animaux qui le pratiquaient. On nepeut donc pas tenir exclusivement compte de la présence d’unedépression associée au m. flexor pollicis longus, que l’on re-trouve chez tous les hominidés, pour suggérer que Orrorin nepratiquait pas une forme de locomotion arboricole.
Le pouce d’Orrorin était différent de celui des grands singescomme en témoigne les dimensions et les proportions de saphalange terminale. Ses membres postérieurs sont bien diffé-rents de ceux des grands singes grimpeurs. Certains caractèresd’hominidé, liés aux actions musculaires et ligamentaires, sontaccentués sur BAR 1901’01 (asymétrie et développement),suggérant de fortes contraintes mécaniques du côté ulnaire. Ilest difficile d’imaginer que ces dernières sont seulement res-ponsables d’un rapprochement du pouce contre le deuxièmedoigt ou contre la paume de la main. On peut aussi supposerqu’elles sont liées à de puissants mouvements de saisie, néces-saire pour le grimper. On ne peut pas les imputer aux possibi-lités de manipuler et/ou fabriquer des objets et/ou des outils,sinon comment expliquer que ces caractères soient moins ac-centués chez l’homme ?
7. Conclusion
Susman (1998) et Shrewsbury et al. (2003) rappellent ladifficulté des choix tant pour la détermination et la spécificitédes taxons que pour les interprétations fonctionnelles. Iln’existe pas un caractère « magique » unique.
BAR 1901’01 présente certains caractères communs à Oreo-pithecus bambolii du Miocène supérieur : présence d’une pro-fonde dépression du m. flexor pollicis longus, facette articulaireaplatie, tubercules basaux asymétriques et situés proximale-ment tout en étant jointifs à la facette articulaire, tubérositéapicale robuste. Certains de ces derniers se rencontrent aussichez Proconsul mais leurs phalanges distales ne sont pas com-plètement décrites pour en apprécier l’ensemble. La présenced’une profonde dépression du m. flexor pollicis longus sembleêtre exceptionnelle pour ce genre à l’opposé de ce qui existechez Oreopithecus. Il est difficile de conclure pour ce caractèrecar les phalanges distales de pouce sont très rares chez les ho-minoïdes fossiles, même chez les squelettes les plus completscomme Dryopithecus ou Nacholapithecus. La présence d’asy-métrie du bord radial de la dépression, d’un renflement trans-versal situé distalement à cette dépression, d’une tubérosité api-cale en forme de « fer à cheval » et asymétrique avec undéveloppement de la partie ulnaire (épine unguéale), et enfinde la présence d’une fosse unguéale forment une associationde caractères qui confirment l’attribution de BAR 1901’01 à

Aie
Alb
As
Be
Bim
Bu
Cla
Ett
Go
Go
Go
Ha
Ka
D. Gommery, B. Senut / Geobios 39 (2006) 372–384 383
un hominidé (Gommery, 2003 ; Gommery et Senut, 2002a,2002b) et non à un grand singe ou à un ancêtre des grandssinges actuels.
Les caractères observés chez BAR 1901’01 mais aussi chezSKX 5016 et Stw 294 montrent des ressemblances avecl’homme qui, chez ce dernier, sont associées à une pulpe del’extrémité du pouce différenciée en deux parties distinctes :une partie proximale plus graisseuse et une partie distale plusfibreuse. Cette distinction n’existe pas chez les primates nonhumains. Il semble que chez les trois hominidés fossiles consi-dérés, la différenciation en deux parties de la pulpe existaitmais avec une partie distale plus réduite.
D’autres caractéristiques liées à l’action de muscles commele m. flexor pollicis longus ou à des ligaments comme les liga-ments interosseux latéraux se retrouvent dans la morphologieosseuse de BAR 1901’01 et SKX 5016. C’est le cas du renfle-ment transversal situé distalement à la dépression du m. flexorpollicis longus, de l’asymétrie des bords de cette dernière, maisaussi de la morphologie du bord ulnaire de la phalange et desfortes insertions ligamentaires. Elles prouvent que les phalan-ges devaient subir de fortes contraintes en flexion.
À partir des résultats obtenus, on peut considérer que lescaractères d’Orrorin pourraient être liés à la locomotion et enparticulier au grimper. Ce type de locomotion, associé à de labipédie, a été suggéré par la morphologie humérale d’O. tuge-nensis et de la phalange proximale (Senut et al., 2001).
Dans la littérature, le débat sur le pouce des hominidés re-pose presque uniquement sur la possibilité ou non de manipu-lation et/ou de fabrication d’outils. La manipulation d’outilsn’est que la conséquence de la possibilité de manipulationd’objets et/ou d’aliments ; tous les primates en sont capablesmais avec des moyens différents. Il faut donc distinguer le typede préhension comme la préhension de « type humain » ou lapréhension de « type grand singe » (Susman, 2004). Nous pou-vons affirmer sans contestation qu’Orrorin pouvait manipulerdes aliments et des objets mais nous ne nous risquerons pas àparler d’outils sans preuve archéologique. Pour conclure plusdéfinitivement sur l’aspect fonctionnel, il faudrait avoir la tota-lité des os du pouce qui constituent un ensemble anatomiquefonctionnel. Quoiqu’il en soit, la phalange terminale BAR1901’01 apporte des renseignements primordiaux sur l’évolu-tion et l’écologie des premiers Hominidés. L’étude du matérielpostcrânien d’Orrorin tugenensis (Pickford et al., 2002) montrequ’il est indispensable d’intégrer des fossiles d’hominoïdesmiocènes aux comparaisons classiques effectuées avec lesgrands singes et les hommes modernes. L’étude des fémursd’Orrorin tugenensis a démontré que certains caractères dit dé-rivés humains sont, en fait, hérités des hominoïdes du Mio-cène ; par ailleurs, elle a montré que les fémurs de chimpanzéspossédaient un grand nombre de caractères dérivés qui permet-tent d’affirmer que la morphologie fémorale des chimpanzés nepeut être prise comme modèle ancestral de l’homme, comme lesuggèrent malheureusement, indirectement de nombreux tra-vaux sur l’évolution des hominidés. Il en est de même pourla phalange distale de pouce.
Remerciements
Cette étude a été financée par les projets « Paléontologie del’Afrique Sub-Saharienne » au Kenya et « Mission KromdraaiB, Greensleeves » en Afrique du Sud de la Division des Scien-ces sociales et de l’Archéologie du Ministère des AffairesEtrangères, l’UPR 2147, l’UMR 5143 et le PICS 1048 duCNRS, USM 203 Muséum et le Collège de France. Nous re-mercions le Community Museums of Kenya à Nairobi, le Dé-partement de Paléontologie du Transvaal Museum à Pretoria enAfrique du Sud, le département d’Anatomie de la MedicalSchool de l’Université de Witwatersrand à Johannesburg enAfrique du Sud et le Laboratoire d’Anatomie Comparée duMuséum National d’Histoire Naturelle à Paris, de nous avoirpermis d’étudier leur matériel. Nous sommes reconnaissants àE. Gitonga, M. Pickford, B. Kramer, P.V. Tobias, R. Clarke,Qian Wang, F. Thackeray, S. Potze, L. Rook, F. Renoult, D.Robineau, D. Fouchier pour leur aide et leurs conseils.
Références
llo, L., Dean, C., 1999. An introduction to Human Evolutionary anatomy.Academic Press, London.
a, D.M., Moyà-Solà, S., Köhler, M., 2003. Morphological affinities of theAustralopithecus afarensis hand on the basis of manual proportions andrelative thumb length. Journal of Human Evolution 44, 225–254.
hley-Montagu, M.F., 1931. On the primate thumb. American Journal ofPhysical Anthropology 15, 291–314.
gun, D., Teaford, M., Walker, A., 1994. Comparative and functional ana-tomy of Proconsul phalanges from the Kaswanga primate site, RusingaIsland, Kenya. Journal of Human Evolution 26, 89–165.
son, B., Ottevanger, J., Roberts, N., Macho, G., Percy, D., Whitehouse,G., 1997. Hominid thumb strength predicted by high resolution magneticresonance imaging and force measurements in living subjects. MagneticResonance Imaging 15, 899–908.
sh, M., Lovejoy, C., Johanson, D., Coppens, Y., 1982. Hominid carpal,metacarpal, and phalangeal bones recovered from the Hadar Formation:1974–1977 collections. American Journal of Physical Anthropology 57,651–677.
rke, R., 2002. Newly revealed information on the Sterkfontein Member 2Australopithecus skeleton. South African Journal of Science 98, 523–526.
er, H.-U., 1974. Morphologisch- und metrisch-vergleichende Untersuchungam Handskelet rezenter Primaten. Teil III. Gegenbaurs morphologischeJahrbuch 3, 299–322.
mmery, D., 2003. Rachis, bipedalism and freedom of the hands. Book ofabstracts, International Round Table, From Tools to Symbols. “FromEarly Hominids to Modern Humans – Des outils aux symboles. Des pre-miers Hominidés aux hommes modern. University of Witwatersrand,South Africa 16-18, March 2003, p. 8.
mmery, D., Senut, B., 2002a. Orrorin tugenensis distal thumb phalanx.Book of abstracts, International Worshop at Bogorio, From Samburupithe-cus to Orrorin: origins of hominids. Geological and palaeontological back-ground. Kenya 28-30, September 2002, p. 2.
mmery, D., Senut, B., 2002b. L’extrémité du pouce de l’ancêtre du millé-naire (Orrorin tugenensis-Kenya). XIVe colloque de la SFDP, 23-25 octo-bre 2002, Doué-la-Fontaine, p. 15.
mrick, M., Churchill, S., Schmitt, D., Hylander, W., 1998. EMG of thehuman flexor pollicis longus muscle: implications for the evolution of thehominid tool use. Journal of Human Evolution 34, 123–136.
pandji, I., 1971. Membre supérieur. Physiologie articulaire, schémas com-mentés de mécanique humaine, fascicule 1. Librairie Maloine S.A., Paris.

Ma
Ma
Mo
NaNa
Na
Pan
Pic
Pic
Pic
Pic
Pic
Ric
Ric
Saw
Sen
Sen
Sen
Sh
Sh
Sm
Sm
Ste
Ste
Su
Su
Su
Su
Su
Su
Tu
Tu
Wa
D. Gommery, B. Senut / Geobios 39 (2006) 372–384384
rzke, M., 1997. Precision grips, hand morphology, and tools. AmericanJournal of Physical Anthropology 102, 91–110.rzke, M., Shackley, M., 1986. Hominid hand use in the Pliocene and Pleis-tocene: Evidence from experimental archaeology and comparative morpho-logy. Journal of Human Evolution 15, 439–460.yà-Solà, S., Köhler, M., Rook, L., 1999. Evidence of hominid-like preci-sion grip capability in the hand of Miocene ape Oreopithecus. Proceedingsof the National Academy of Sciences USA 96, 313–317.pier, J., 1962. Fossil Hand bones from Olduvai Gorge. Nature 3, 409–411.pier, J., 1993. Hands. Revised by R. Tuttle. Princeton Science Library,Princeton.pier, J., Davis, P., 1959. The forelimb skeleton and associated remains ofProconsul africanus. Fossil Mammals of Africa 16, 1–70.ger, M., Brooks, A., Richmond, B., Wood, B., 2002. Older than the Oldo-wan? Rethinking the emergence of Hominin tool use. EvolutionaryAnthropology 11, 235–245.kford, M, 1974. Stratigraphy and Palaeoecology of Five Late CainozoicFormations in the Kenya Rift Valley. PhD thesis, University of London.kford, M., 1975. Late Miocene sediments and fossils from the northern RiftValley. Nature 256, 279–284.kford, M., Senut, B., 2001. The geological and faunal context of Late Mio-cene hominid remains from Lukeino, Kenya. Comptes Rendus de l’Acadé-mie des Sciences de Paris (2a) 332, 145–152.kford, M., Senut, B., 2005. Hominoid teeth with chimpanzee and gorilla-like features from the Miocene of Kenya: Implications for the chronologyof the ape-human divergence and biogeography of Miocene hominoids.Anthropological Sciences, juillet 2004, 113, 95–102.kford, M., Senut, B., Gommery, D., Treil, J., 2002. Bipedalism in Orrorintugenensis revealed by its femora. Comptes Rendus Palevol 1, 191–203.klan, D., 1987. Functional anatomy of the hand of Australopithecus africa-nus. Journal of Human Evolution 16, 643–664.klan, D., 1988. A functional and morphological study of the hand bones ofearly and recent South African hominids. Ph D Thesis, University of theWitwatersrand, Johannesburg.ada, Y., Pickford, M., Senut, B., Itaya, T., Hyodo, M., Miura, T., Kashine,C., Chujo, T., Fujii, H., 2002. The age of Orrorin tugenensis, an earlyhominid from the Tugen Hills, Kenya. Comptes Rendus Palevol 1, 293–303.ut, B., 1981. L’humérus et ses articulations chez les hominidés pliopléisto-cènes. Cahiers de Paléoanthropologie, Éditions du CNRS, Paris, 1–141.ut, B., Gommery, D., Pickford, M. 2002. The locomotion of Orrorin tuge-nensis: implications for the origins of bipedalism. Book of abstracts, Inter-national Worshop at Bogorio, From Samburupithecus to Orrorin: origins
of hominids. Geological and palaeontological background. Kenya 28-30,September 2002, p. 18.ut, B., Pickford, M., Gommery, D., Mein, P., Cheboi, K., Coppen, Y.,2001. First hominid from the Miocene (Lukeino Formation, Kenya).Comptes Rendus de l’Académie des Sciences de Paris (2a) 332, 137–144.rewsbury, M., Johnson, R., 1983. Form, function, and evolution of the dis-tal phalanx. Journal of Hand Surgery 8, 475–479.rewsbury, M., Marzke, M., Linscheid, R., Reece, S., 2003. Comparativemorphology of the pollical distal phalanx. American Journal of PhysicalAnthropology 121, 30–47.ith, S., 1995. Pattern profile analysis of Hominid and chimpanzee handbones. American Journal of Physical Anthropology 96, 283–300.ith, S., 2000. Shape variation of the human pollical distal phalanx andmetacarpal. American Journal of Physical Anthropology 113, 329–348.rn, J., 2000. Climbing to the top: a personal memoir of Australopithecusafarensis. Evolutionary Anthropology 9, 113–133.rn, J., Susman, R., 1983. The locomotor anatomy of Australopithecus afa-rensis. American Journal of Physical Anthropology 60, 279–317.sman, R., 1979. Comparative and functional morphology of hominoid fin-gers. American Journal of Physical Anthropology 50, 215–236.sman, R., 1988a. Hand of Paranthropus robustus from Member 1, Swart-krans: fossil evidence for tool behavior. Science 240, 781–784.sman, R., 1988b. New postcranial remains from Swartkrans and their bea-ring on the functional morphology and behavior of Paranthropus robustus.In: Grine, F. (Ed.), Evolutionary History of the Robust Australopithecines.Aldine de Gruyer, New York, pp. 149–172.sman, E., 1998. Hand function and tool behavior in early hominids. Journalof Human Evolution 35, 23–46.sman, R., 2004. Oreopithecus bambolii: an unlikely case of hominid likegrip capability in a Miocene ape. Journal of Human Evolution 46, 105–117.sman, R., Creel, N., 1979. Functional and morphological affinities of thesubadult hand (O.H. 7) from Olduvai Gorge. American Journal of PhysicalAnthropology 51, 311–332.ttle, R., 1967. Knuckle-walking and the evolution of hominoids hands.American Journal of Physical Anthropology 26, 171–206.ttle, R., 1969. Knuckle-walking and the problem of human origins. Science166, 953–961.lker, A.C., Pickford, M., 1983. New postcranial fossils of Proconsul afri-canus and Proconsul nyanzae. In: Ciochon, R.L., Corruccini, R.S. (Eds.),New interpretations of ape and human ancestry. Plenum Press, New York,pp. 325–351.





























