Embed Size (px)
Citation preview

American Association of Stratigraphic Palynologists
Late Cretaceous Megaspores from Southern Sweden: Morphology and PaleoenvironmentalSignificanceAuthor(s): Eva B. Koppelhus and David J. BattenReviewed work(s):Source: Palynology, Vol. 13 (1989), pp. 91-120Published by: American Association of Stratigraphic PalynologistsStable URL: http://www.jstor.org/stable/3687535 .
Accessed: 26/02/2013 12:49
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Association of Stratigraphic Palynologists is collaborating with JSTOR to digitize, preserve andextend access to Palynology.
http://www.jstor.org
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

LATE CRETACEOUS MEGASPORES FROM SOUTHERN SWEDEN: MORPHOLOGY AND PALEOENVIRONMENTAL SIGNIFICANCE EVA B. KOPPELHUS Geological Survey of Denmark Thoravej 8 DK-2400 Copenhagen NV Denmark
DAVID J. BATTEN* Department of Geology and Mineralogy Marischal College Aberdeen University Aberdeen AB9 IAS Scotland, United Kingdom
Abstract
The poorly consolidated clastic sediments of Late Cretaceous age in the vicinity of Asen, southern Sweden (Scania) are noted for the variety of fossil flowers and Normapolles pollen grains they have yielded. Megaspores have also been recovered from a num- ber of horizons. Comments on, and illustrations of all but a few indeterminate remains of these are provided in this paper, and four new species, Bacutriletes ferulus, B. majorinus, Erlan- sonisporites scanicus and E. septus, are erected. Comparison of the total recovery with the relatively small number of Late Cretaceous assemblages that have been described from elsewhere reveals only a few taxa in common. As a result, it is not possible to improve on the late Santonian-early Campanian age determi- nation that has already been established for the Asen deposits. It is reasonable to infer from the wall structure of the majority of the spores that they were shed by plants of either selaginellalean or isoetalean affinity. Their distribution within the Asen sequence supports the previously held view that the general environment of deposition changed from floodplain-lacustrine to predominantly fluviatile during the time represented.
INTRODUCTION
The fluviatile-lacustrine deposits of Asen in Scania, southern Sweden, are well known to those
who work on Cretaceous plants and palynomorphs for the diversity of their floral content. During the past several decades both macro- and microfossil remains have been described in a series of papers (Ross, 1949; Nykvist, 1957; Skarby, 1964, 1968, 1974, 1978, 1986) culminating in headline-making news about charcoalified flowers with pollen grains in situ, some of the latter being of the Normapolles group (Friis and Skarby, 1981, 1982; Skarby and Nilsson, 1982; Friis, 1983, 1984, 1985; Friis et al., 1986, 1988). The megaspore content of these depos- its has not, however, been described hitherto.
Our paper is based on specimens recovered from two suites of samples. One of these was collected comparatively recently (1983 onwards), mainly by E. M. Friis and K. Raunsgaard Pedersen but with contributions from E. Koppelhus. The other is from a borehole drilled many years ago; samples from the cores recovered were forwarded by N. E. Ross to S. J. Dijkstra during the 1950s. There was an agree- ment that the former would work on the miospores and that Dijkstra would describe the megaspores but then Ross died and no publication ensued.
Present Address: Institute of Earth Sciences UCW University of Wales Aberystwyth SY23 3DB United Kingdom
Palynology, 13. 91-120
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

92 PALYNOLOGY, VOLUME 13 - 1989
580N 2
SWEDEN
Asen. 560
S0'O 50 100 km DENMARK
120 140 160E
Text-Figure 1. Map of southern Sweden and adjacent countries showing the location of the Hoganas AB quarry at Asen.
The Asen locality is the Higanis AB kaolin quarry which is situated in the Kristianstad Basin (lat. 5609'N, long. 14?30'E; Text-Figure 1). The plant-bearing sequence consists of about 20 m of unconsolidated sands and clays that are divisible into a lower and an upper series, the boundary between the two being a distinctive weathered hori- zon. The lower unit overlies the kaolinite which is unconformable on Lower Palaeozoic basement. Similar deposits are known from two other, now abandoned, quarries nearby in the vicinity at Ax- eltorp and Ivo Klack.
The geology of the area has been described on several occasions (e.g., by Gronwall, 1914; Lundegren, 1931, 1934; Christensen, 1975, 1984), and Ross's (1949) geological cross-section, which is redrawn here (Text-Figure 2), is still pertinent. The plant-bearing beds have been dated on the basis of palynomorphs (Skarby, 1968; Friis and Skarby, 1981) and paleomagnetic evidence (MOrner, 1983) as late Santonian or early Campanian, a determina-
tion that we cannot better on the basis of our study of the megaspores. We comment on this and on the paleobotanical and evolutionary significance of the megaspores recovered from Scania in the sections following the systematic part of our paper.
MATERIAL AND METHODS
Twenty-two samples (GI32140-32162) were col- lected during 1983 from a 15 m sequence in the western part of the quarry (Plate 13, fig. 17). Their distribution and the number of megaspores recov- ered from 800 g of each are given alongside a lithological column on Text-Figure 3. Additional assemblages extracted by E.M. Friis from samples collected elsewhere in the quarry (GI32106, 32107, 32117, 32123, 32188, 32189) have also been ex- amined (Table 1). Other fossils recovered from some of these have recently been described by Friis (1985) and Friis et al. (1988), namely Platananthus scanicus (GI32107, 32189), Platanocarpus sp. (GI32107, 32189) and Actinocalyx bohrii (GI32107). The megaspores were isolated from the encompassing sediments by washing onto a sieve of 120 pm mesh. The removal of adhering inorganic detritus was achieved by placing them in hydroflu- oric acid to digest silicate minerals followed by immersion in hydrochloric acid. All of the spores are now stored in the Department of Palaeobotany, Swedish Museum of Natural History, Stockholm. Each of the figured specimens is numbered (prefixed by S; see plate explanations).
Dijkstra's collection originally consisted of single cell slides containing dry, unmounted specimens and small amounts of dry organic detritus in card- board pill-boxes. The few megaspores and fragments thereof in the latter were picked out from the resi- dues and, together with the specimens that had been isolated previously, were stored in a new set of
PLATE 1
Trileites persimilis (Harris) Dijkstra. Figs. 1-3 and 5, scanning electron micrographs. 1, 2 One specimen, S100500, sample GI32123. 1, proximal
view, x 150; 2, surface of exoexine, x 5000. 3, 5 One specimen, S100501, GI32123. 3, transverse section
of spore showing compact peripheral "skin," open outer network and densely packed inner layers of exoexine, x 2000; 5, detail of transition from outer to inner exoex- ine, x5000.
4, 7 Transmitted light photographs of transverse section of spore showing thick exoexine and thin, membraneous and largely detached intexine, S100502; GI32146; 4, x125, 7, x500.
6, 8 Transmission electron micrographs of ultra-thin sections of megaspore wall; S100503, GI32146. 6, part of outer, but mainly inner exoexine, x 3000; 8, detail of inner exoexine showing regularly arranged spherical elements of sporopollenin, x 10,000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 1
,-C ....- ;. j ?
.,*g: 1E! F:a~
tI~k
+,~~.. :.cr
ie
.... ....i.:i..i:;.:ncFIrstj~ Q~
.. ?il .:.... ar
?i., ,..i. .: :lld ~
?i'i~"~~ ;,~Y +liFn ./all~~l~~3l ~ ? -ryb!E~
? ,o
?
... .: . i- :,
.- . . : ?
.
..~2 '. . "
'i. "." -i:.
:-.: :i.. .-:.:
? "iiii:.j i :.. E; .:i ~..:.. .
- I L-PI L .. ~ ~ ~ i
?~~: .: .
.I,
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

94 PALYNOLOGY, VOLUME 13 - 1989
W10 S EO10 N Blaksudden 8sen Axeltorp
Archean lKaolin nd d clae + + + +taceous ++++ __uU
-on tainin lant fossil---s--- l lY++ + +li +m+est S+ $++ + + Y~ II + + + t- + 4-t S+ + + + +- 4" 'r + + + + + + -i-
Archean Kaolincontaining plant fossils r- limestone
Text-Figure 2. Schematic cross-section of the geology of Asen and surrounding area (after Ross, 1949, fig. 2).
slides. There are no rock samples. The collection is on loan to D.J. Batten from the Geologisch Bureau of the Rijks Geologische Dienst, Heerlen, The Netherlands.
Scanning electron microscopy was undertaken us- ing both Cambridge Stereoscan-2 and Hitachi S800 instruments. Prior to examination, all specimens were coated with either gold or gold-palladium. Megaspores selected for sectioning were embedded in epoxy resin prior to being thin-sectioned on an ultramicrotome. Sections between 700 and 800 A thick were mounted on grids and examined under a JEM-100U transmission electron microscope. Thicker (2-3 rtm) sections were mounted on glass slides in glycerine jelly for study in transmitted light.
SYSTEMATIC PALYNOLOGY
Almost all of the forms of megaspores encoun- tered are described or commented upon in this section. The few omitted are too fragmentary or degraded to merit consideration. Descriptions of previously published species have been omitted if the specimens recovered are virtually indistinguish- able from the type material or are represented by only one or two specimens. Observations on these are instead largely confined to the characters that serve to distinguish them from other taxa. Several of the rare spores may belong to new species but it has
not been possible to formally describe these because of the dearth of specimens.
Although most of the megaspores can be classified without difficulty using Potoni6's (1956) system, a few present problems. It is, for example, not possible to place Cabochonicus satisfactorily in any of the existing sub- or infraturmas, and Ricinospora can be included in either of the infraturmas Murornati or Perinotriletes depending on rather arbitrary criteria. As more data on the wall structure of Cretaceous megaspores become available, it is likely that most species will ultimately be classifiable in a semi- natural system. Since Potoni6's method of grouping taxa is based purely on morphological criteria, some spores that are likely to have been produced by closely related plant taxa may be separated on the basis of unimportant characters and others which are probably unrelated are brought together under a single infraturma. There seems little point, there- fore, in continuing to use a system of categorization that hinders rather than enhances our understand- ing of the evolution of heterosporous plants as re- corded by their reproductive organs.
We have attempted to determine the structure of the walls of the megaspores wherever possible. This depended on the number of whole and fragmented specimens available. In general we were reluctant to fracture undamaged specimens in order to obtain cross-sections of their walls that were suitable for
PLATE 2
All scanning electron micrographs apart from fig. 9. 1, 2, 4 Trileites murrayi (Harris) Marcinkiewicz. 1, 2 One specimen, sample/slide 1587, Dijkstra's (SJD) col-
lection. 1, proximal view, x 100; 2, detail of surface, x 5000.
4 Specimen with undulating proximal face, S100504, sam- ple GI32117, x 150.
3 Trileites sp., proximal view, S100505, GI32123, x 150. 5, 6, 10 Maexisporites soldanellus (Dijkstra) Potoni6. One speci-
men, 1594 (SJD). 5, proximal view, x200; 6, part of surface at higher magnification, x 1000; 10, detail, x 10,000.
7-9 Maexisporites hammaphorus Schultz & Noll. 7, 8 One specimen, S100506, GI32123. 7, proximal view,
x 150; 8, detail of surface, x 1500. 9 Transmitted light photograph of thin section of mega-
spore wall showing intexine (i) and exoexine (e), S100507, GI32145, x400.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 2
~?F~ ~,;i~::u~~~~r~;liJJ~?~~?:. . L P''~ "
:::~::' :b~z; '
: GJ)? "X"' '""
C" c? ' ' ' ?
2 ~1~. ?~i~f~I~ac .8:?: ~El~..u?~sr~%lLY?~msIlrrsrp~sl~"~s~8~ia I_
2:_?~P*d, ?
,. :,ilDaB~Pi?l' C; ,. ,sL~ :~Ji~' r r ~dJz~2b~ss;:' ~i~C~ 1~?-r? g
, ? t r '?~i'* .i
i ??
'"'' ~~tiggE~i i ~E~~f~CS ~rt~~t~lE~r:i T~s~ t: * t
; ..'9"' .rS r'r. ~ ~ r: '' t lr?? i'.' P
C ~P; .i
?; ii~yi- ~ ----~l~Ell~-?e 'Iri~Ill 131 ~ -? ~tFbssp rsI df~il;~tl~+ ?.r 1* -i 3
r
i~-*IPl~i '~"esl~pr~ic""4iqlewr I~ ii
'~tii~ 4~
'C- ~E* rl~' "YEYi.~?~?C"I~:?~i~~r~jrpl~i~ ~r~3j~., i. :~.~:?ll~t~E~3bsau~-sP~- Ir r~lr I ?* * ~~ ??:?. * 5~. r\ :4 ? " B ,S r
1 Li ' !, ? , 'f Il ;tl? .''*?~...*~ .) r~lllElii;S?+*+???i ? ~fi~ ...., .. 1.,
: ??"?"f~~.??? ? ,L'~~ I
i :r~~?~ .I~ ~I~?111~I*? :u
~*~'3~' I* .1 r*f ?-~ ~*L*- t n~4` r. ?~ ., r ?I * B~i*L L ?;i~ Ibi :I !I i *' 'Ja 9i :: ti. ? .!?
:~ ::: .. i
,, ~`,??~: r ..? .~*'?"; d'? ,.1 ti ;? br 1, r; *I
8.7. ?~? i; a "' ~ .?( *? ~??S~i~~~?'~~ ,? 31~E~ESgi~WR;~k'
~ * r?- ~14~:9. :C?*J?.~E~ r `'s r *?' a :?*
4 "" .~- 5 ~ r
rij211~(: i v L T~E:I&I :I' C i. .**
I . ` ~~.?.~?~.~LQlse~iS~=?r -?E?~? I .? r .*;-i?ICr :i: U ',
P' i :'j t ,.~~?L 3 * ??~ ,a?? i4 ?r " :?? ?s, * c ,~. ?, 4*
a dek-'1 s 9i?
..*E r '? '~54? ,? ,? ?j~?
::iii ,,, ~E*rF *IP?~??l IYihF.: *;i~ ~? ?? +CIic;'
'~j~i i1 U *?? ~ :I P' ? ?~J ?rre~? P,~r~ a~'i?i ??) .le~r~?gS1*71 fl~
** j f??? jC*~' *r r ??il *'1EI' ~I
'' " '~L* ??? *,~IR ?? I~4?~rYp ?rlP *' r .?
e .?iJs~ ~ i i:
i
~i d g *: ?"' .r
?r 1 r! - - ??:iik?l?~:
7 ,?-- ?? ?? 8: r 9 ~ " _ ~g~ol
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

96 PALYNOLOGY, VOLUME 13 - 1989
Sample no.
GI 32162* ..212~.....~...~.... .
61 _ * 1\Clay
Silty clay
60-
59 _ _ _ _ _ _
__.Silt/fine sand
58
57* "j- Medium sand
56 -. . Coarse sand
55- ................'
Z"Roots 54 - . '. ' '. .' .-
..................
53 .[.1 Coalified wood fragments 52*
51 - .......-'..
Weathered horizon 50 .................
48 - - ........
- -
-4 47. _
-3
411
600
0 10 20 30 40 50 60 70 80 90 100 110
Number of megaspores recovered from 800 g of sediment
5 ............... 5
55..........
... - 4ICaiidwo rget
. . . . . . . -
51 ~ ~~~~~~ _%???? etee oio .. .. .. .. . ..????
4? _
47 ----- . .
45 -:
44 - .--~;
43 1---"------ ---- -C
42 -' . ................. 1
41 - .- - GI32140 .% -._
0 10 20 30 40 50 60 70 80 90 100 110
Number of megaspores recovered from 800g of sediment
Text-Figure 3. Lithological sequence in the western part of the Hoganais AB quarry (see Plate 13, fig. 17), sampling horizons (compiled by E.M. Friis) and numbers of specimens recovered. * = sample yielded only indeterminate fragments.
scanning electron microscopy, or to destroy them completely by thin-sectioning.
Genus Trileites Erdtman 1947 ex Potoni6 1956
Trileites persimilis (Harris 1935) Potoni6 1956 Plate 1, figs. 1-8; Plate 13, fig. I1
Triletes persimilis Harris, 1935, p. 165, text-fig. 52R, pl. 25, fig. 4.
Trileites persimilis (Harris) Potoni6, 1956, p. 24.
Description. Trilete megaspore, outline in polar view triangular with slightly convex sides (Plate 1, fig. 1). Maximum diameter 440(590)640 jtm (10 specimens). Proximal face slightly flattened, distal face rounded (Plate 1, fig. 4). Trilete ridge promi-
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden 97
C./)
32162 61 F o 0 0
60 o 0 o o e o o o
59 o o F o F o oo
57
56 o o * o o o F
55 o F o * *
54 o o o o 53 0 o C
52
51 F F o 50 0 o C
49
48 o 0
47 o oo o F
46 o o o o o o o 0 45 o o o o ? * o
44 o o o o o o 43 o o o o 0
42 o o o F o o
41 o F o F F o o o o
40 o o F F o o o o
060 o 0o o 0 o0 *o 0o 0 o 0o 07 o 0 0 0 C
17 C) C 0 o 0 0o
23 0o o o * * o00
88 F 0 0 0
32189 0 00 0
SJD o 0 o o 0 0
53 0 00v v
52 S J v, v SiF F, 0, v
500, 0
49vv 48 0 47 0 0 0 460 v 0 0 450 0 , 0P 0 @ 0 44 00 0 y0
41 0 F 0 0
400o 0 F F 00 0 0
060 00 00 0 @ 000 0
070 0 0 00 0
170 * o 0 @0
88 00 0
3218 0 0 0
SJD 0 0 0i 0 0 0 0
o present, 1-10 specimens * common, >10 specimens F 1-10 fragments -* indeterminate fragments only
TABLE 1. Distribution of megaspore taxa in the measured section at Asen (32140-32162), recovered from isolated samples from other parts of the quarry (32106-32189), and present in Dijkstra's collection (SJD).
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

98 PALYNOLOGY, VOLUME 13- 1989
nent, extending to >2/3 radius. Exine ca. 35 pm thick (may be more than this adjacent to triradiate mark; see Plate 1, fig. 4), surface essentially smooth but commonly displays pits and irregular grooves which are presumed to be the result of degradation (Plate 1, fig. 1). Surface layer of exoexine is made up of a mat of threads separated by both small (0.2-2.0 im) and comparatively large spaces which are rounded or elongate (Plate 1, fig. 2) in outline. Immediately below, outer exoexine is composed of small spheri- cal elements ca. 0.5 jm in diameter connected together to form a three dimensional reticulate meshwork which becomes more compact towards interior giving a very characteristic pattern of alter- nating rows and layers connected by short rods (Plate 1, figs. 3, 5, 6, 8); inner exoexine is more compact (Plate 1, figs. 3, 4). Intexine thin (ca. 3 jm), membraneous (Plate 1, figs. 4, 7).
Discussion. Our specimens are similar to those referred to "Triletes cf. persimilis" by Dijkstra (1949; see Batten, 1988, for new illustrations). There is also some resemblance to Trileites subrotundus (Miner 1932) Potonie 1956.
Trileites murrayi (Harris 1961) Marcinkiewicz 1971
Plate 2, figs. 1, 2, 4; Plate 13, fig. 2
Triletes murrayi Harris, 1961, p. 64, 66, figs. 20A-F, 21B.
Trileites murrayi (Harris) Marcinkiewicz, 1971, p. 31, pl. 1, figs. 6-8.
Remarks. The two specimens illustrated are typi- cal of the very few forms encountered at Asen and present in Dijkstra's collection whose characters fall more or less within the circumscription of T. mur- rayi. The rays of the triradiate ridge are, however, rather longer than those on the type (Middle Juras- sic) material, being approximately 1/2, as opposed to
13, of the radius of the spore. The surface of the exoexine appears to be smooth to slightly roughened at low magnifications (Plate 2, figs. 1, 4) but, in common with many other kinds of megaspores, it is seen to be spongiose to foveo-reticulate when viewed at high power under the SEM (Plate 2, fig. 2).
Trileites sp. Plate 2, fig. 3; Plate 13, fig. 3
Remarks. The gross morphology of the spore identified as Trileites carbunculus (Dijkstra 1949) in Knobloch (1984b, pl. 7, fig. 7) appears to be similar to the specimen illustrated on Plate 2, fig. 3 but it is nearly twice as large.
Genus Maexisporites Potoni& 1956 Maexisporites hammaphorus Schultz & Noll 1987
Plate 2, figs. 7-9; Plate 3, fig. 1
Maexisporites hammaphorus Schultz & Noll, 1987, p. 91, pl. 3, figs. 6-7.
Remarks. Schultz and Noll (1987) did not provide a size range for Maexisporites hammaphorus. The specimen they illustrate is ca. 280 jm in diameter. The few spores from the Asen section that have been referred to this species are somewhat larger (320-400 jm), and are sculptured with a mixture of granulate, papillate and irregularly murornate elements (Plate 2, fig. 7; Plate 3, fig. 1). At high magnifications (e.g., Plate 2, fig. 8) these are seen to be constructed of a three-dimensional mesh of sporopollenin threads that is finer than that of the unsculptured, outer part of the exoexine. They also tend to be capped by an irregular but largely imperforate membrane. In common with all of the other megaspores examined in thin section, the intexine is much thinner than the exoexine (Plate 2, fig. 9).
PLATE 3
All scanning electron micrographs. 1 Maexisporites hammaphorus Schultz & Noll, oblique
view, S100508, sample GI32146, x280. 2-7 Maexisporites spp. 2, 5, 6 One specimen, S100509, GI32123, 2, oblique view,
x 100; 5, detail of surface excrescence, x 2000; 6, part of equatorial region, x 300.
3, 4 One specimen, S100510, GI32106. 3, proximal view, x 100; 4, surface detail showing three-dimensional sporopollenin network, x 2000.
7 Oblique proximal view, sample/slide 1594, Dijkstra's collection, x 200.
8-11 Cabochonicus carbunculus (Dijkstra) Batten & Ferguson. 8, 10, One specimen, S100511, GI32123. 8, proximal view, 11 x 150; 10, detail of surface of gemmate excrescences,
x 2000; 11, gemmae adjacent to roughened contact area of proximal face, x 1000.
9 Transverse section through exoexine, specimen same as that illustrated on Plate 4, figs. 1-3, S100512, GI32106, x 5000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 3
?r,
, g,,, o, ,~ ?"~ LU~
?ii.j.;%
. *
. :
+ .+ . . ;.,+,+ +~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~~:i ....~ ,+?; ++ .. +-++++....+! , .
+: .r "c~
?*??' ,5 .+: :+:I .. - ,-, c?
+"~~ +,++- , ++ ++ .,++ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ,
+.::" ?I ....:...w...+++- : .
. ...++. ...+ ,+J , ? + ...
:: pr *': +:++
i;*
: . .+ " ++ ++ + +
+ + ,. +
++ ... ++<+\ .+ '' ++ ++ + ++ ,, ;;R ++'~i~ ?,)l+ ++++ +': + +++ + ..... . ..; ? + . ,,++
+,
+ +,++
* + :i i: + . .. +;+ '
rc,
,
sl
+?s",+
?? .:;;s*C ,.+. ?I .<...:.++
~,' _
. r +, .<.
. + =,,,~ Im+~C; ??I' + +++
o.+:~" ?sJI
::+ +<i++ i?~ '
1* +TL? ?? ,+.r+ I L,~?%: ~ ? .,+*
P
... + 3+ +
?r"U. +: Ik
+,,,,) + .. + I _,,,?
? ~*r
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

100 PALYNOLOGY, VOLUME 13 - 1989
Most species of Maexisporites are morphologi- cally "conservative" in appearance in the sense that similar spores occur in both much older and younger rocks. M. hammaphorus is no exception. Hence, although originally described from north German palaeokarst infill deposits of Aptian age, its occurrence at Asen in beds that were laid down some 25 million years or more later is thought to be within an acceptable age-range for the species.
Maexisporites soldanellus (Dijkstra 1951) Potoni6 1956
Plate 2, figs. 5, 6, 10; Plate 13, fig. 4
Triletes soldanellus Dijkstra, 1951, p. 10, pl. 2, fig. 17.
Maexisporites soldanellus (Dijkstra) Potoni6, 1956, p. 25, pl. 2, figs. 17-19.
Remarks. Our specimens are somewhat larger than those on which Dijkstra (1951) based his description of M. soldanellus (400-480 jim as op- posed to 300-360 jim), and the sculptural elements are perhaps a little more variable in form. They are, however, indistinguishable in other respects.
Maexisporites spp. Plate 3, figs. 2-7; Plate 13, fig. 5
Remarks. Apart from the species of Maexisporites named above, two others have also been recovered from Asen and a third is present in Dijkstra's collec- tion. Unfortunately, all three are represented by only single specimens, thus hindering identification at specific level. Under the SEM the globular sculp- tural elements on one of these (Plate 3, figs. 2, 5, 6) are similar in form and distribution to those that occur on some specimens of Cabochonicus but they do not have the glossy, resinous appearance that is characteristic of this genus, and the shape of the
spore, with its prominent triradiate ridge, is more typical of Maexisporites. Our reference to this genus is, however, only tentative pending the recovery of more specimens.
Genus Cabochonicus Batten & Ferguson 1987
Cabochonicus carbunculus (Dijkstra 1949) Batten & Ferguson 1987
Plate 3, figs. 8-11; Plate 4, figs. 1-3; Plate 13, fig. 9
Triletes carbunculus Dijkstra, 1949, p. 22, pl. 1, fig. 12.
Verrutriletes carbunculus (Dijkstra) Potoni6, 1956, p. 28, pl. 3, fig. 26.
Trileites carbunculus (Dijkstra) Knobloch, 1984b, p. 156, pl. 1, figs. 1-5, 7, 8, pl. 7, fig. 7.
Cabochonicus carbunculus (Dijkstra) Batten & Fer- guson, 1987, p. 72, pl. 1, figs. 1-4.
Description. Trilete megaspore, triangular in equatorial outline, light to dark brown, sometimes orange-brown in color with shiny surface. Maxi- mum diameter 400 (504) 560 jim (3 specimens). Trilete ridge ca. 35 jtm wide and 30 jim high, extends to 3/4 of radius of spore. Numerous gemmae 5(15)30 pm in diameter occur in equatorial region and may also be scattered over distal face (Plate 3, fig. 8; Plate 4, fig. 3). They can be yellow, orange or occasionally black in color and nearly always have a glossy appearance. Granulate to vermiculate ele- ments are commonly scattered over their surface (Plate 3, fig. 10; Plate 4, figs. 1, 2); they may also be irregularly depressed or punctured (Plate 4, fig. 1) and densely packed with fused bases. "Unsculp- tured" surface of spore can be either more or less smooth or also have a roughened/scabrate, or granu- late to vermiculate appearance (Plate 3, figs. 8, 9; Plate 4, fig. 1).
PLATE 4
All scanning electron micrographs. 1-3 Cabochonicus carbunculus (Dijkstra) Batten & Ferguson. 1-3 One damaged specimen, S100512, sample GI32106. 1,
gemmae with partly granular and pitted surfaces sur- rounded by "collars"; surface of exoexine vermiculate, x 2000; 2, detail, x 5000; 3, general view showing scat- tered gemmae of varying size, x 200.
4-6, Bacutriletes ferulus sp. nov. 8, 9 4, 6, 9 Holotype, S100513, GI32145. 4, proximal view, x 150;
6, close-up of bacule, x 750; 9, detail of arrangement of sculptural elements, x 300.
5, 8 Parts of broken specimen, S100514, GI32145. 5, show- ing membraneous intexine partly separated from much thicker exoexine, x 100; 8, detail, x 500.
7 Verrutriletes sp. cf. V. compositipunctatus (Dijkstra) Potoni6, detail of sculpture of specimen illustrated on Plate 5, fig. 8, sample/slide 1594, Dijkstra's collection, x 3500.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 4
:liiiii!ii .ii,~ii~~~s~*s~---~l"'~~~ss~~iiiili:::i::i: ::::? ::
???:7ii I~li~ri;(i.il(liiiiiii :it ~:ilg~ppg~ 1 assr~- ~911j3s~a ~
?h:;i ::r: ::;-: ::""": :::i '::?'"
,i~~:?3i:~ii~a' ..~ia~kllarr~3lr~l~as~jii~CIIIICqgBa~i~ C :i :: .!il :'" :ti
:i' ?:lii~L' 2;
:~3il
C, "?ir~ .. ~ Jq~3~?~ Ir ?. r*?:''?? ??:' a:. ??~?
'?? ;iiitllr"~. *IY .C ;r? -'" :I;
?:fPllfP~~11.,. P"T~~i~' ~i~iiiL~iiJ~iP*E ~ ''"
C. ; ~i?-- :c~ 1 ??r? ?ii~L~i~E~Tt~? * i!lii ?i i: yil~6~ :? * ?? ~oirif ..J
,, ~1 l~~t ?:3 ii :.: :::;.I -e wi-' i~~IQ~?i~ 4? :?
fC .:.. Il~a~?? 2 ?:? ?:: -;'? ~;~r- #LW,`E rC~Ci~b: ~fii~ 1: ~1!'it ..::: .,::??r':? ??? :a:i:
?!, ::::?? * ,.ii :ii
:'i:~ 3R~astlll~sllBB~,~i~'~FB~:~?i, ';'?: *rx
d ;; *I;r" i;. ~?~sl~j~urau~B.sz;~E~' iIij!: .dw d: ....r 4 ?-? i:::
e
d;i"l -ic i? \s
~A JFi :Z ?1.1 4. :ib:? 9*;. .,?? a :5:1
"
rird* tl C~t~ljiii
?u ?: ?~:??? ;*r"'* ii:l~~ c ' I:'J
I::~:s:? ?:ii:: ~ ? 4'?f
.::?: ?, :A?: :i.?.i :ii: t?~ ?,,~ ?i ~+ ;*?I :?:;:?
;.?i?:;I'D' .';i P)Jk.: CIJ *;' ? :Lii~ iI? F-- .ii:
-;r r? 3 *ir~ f? ?~. a "5 ?k,.. ;a Eza~rI-
?:."i "lii ::::. .r I ,? ii
.r,.h
.i~s~L~ .Ir; p ?ii 5 ii* ..i"P r: i: Q.
?If?? ~r~ :":iE:::: .s!~ "
::i:
If
4: 5 .iiiF j. ~iZ ?:' iii :?1:?
.iii Fu~: ~AlJ
r? I'F~ ""':I~PS~:'~JSp~; 8,
rfrt *r ** ir~cf-~ L * *..~k?? ~ ~~ ' .'* I puc ~urs - ... :: *? "'Y ~ 511 ;i", Q
r* ~r? ??j~ r r. a ~ V'? .? i?? * .~i,?* "? ... ', ...*. i? ~? a 1:? 3 ,~ '?~ '
?L. ' d*~i. " -r- ?-;rr; ? - "'C... z ?*L?;*P i "jsr i*l ?::?:i: a ?"~~F~i! ii~ i. ,*? ??i it
.IL-~r i?- R ~ C t I'V** .$" +? JI~F~L'3 ~ t' ;'* :lr ??Y
c3f r? - r= ?;?*x?..?' .li "r,*f * ,, ~ ???*C i? ?r. ?: ? ?i, ~ a L? ;7~~ s .;I ? ? Lt ~i iii: "?? : -I ??r Y O r.F ? - r r ?*: .,? r 11 . ~1"; ,?
?tl- ,?.* , Lt~;~, ,risa~ ii ?. ; ~F~m~,-rs lan ?.* *)~IJt~ -*- r n ?? * ri~a~- s ?r~ ~x.g,* :~a~~ ,, '~* d
~r~li~~ ?irZ* ::? ?:;?' *' ?P ??I ;'" ? Li!
) Q* ~* .. ?* :*~ ??;i::s?:: ?a: ?1**;bi r . : E a :~: ~-C *'* :.?
.b ns:I~ a~r.: ~-a~~ ri ?:a ,e .. ... . I rirqtie ~~8~sl~Rt~?? ?1I1: *v *r I ha
t j(Ir/+ .E t~': 1. ?1 *? .I:
-1 J~ ,,.sa ??? ,?ir a??? "ir 8 ?,
,C:? d ???:: g ?? II: ~I
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

102 PALYNOLOGY, VOLUME 13 - 1989
Remarks. The few specimens recovered from the Asen locality are similar in appearance to those in Dijkstra's collection (Dijkstra, 1949; Batten, 1988). They differ in being generally smaller and in show- ing greater morphological variation. There are, how- ever, no reliable criteria at present on which to provide a basis for distinguishing another north European Late Cretaceous species of the carbuncu- lus-type.
Genus Verrutriletes van der Hammen 1955 ex Potoni& 1956
Verrutriletes sp. cf. V. compositipunctatus (Dijkstra 1949) Potoni6 1956 Plate 4, fig. 7; Plate 5, fig. 8
Remarks. This form is morphologically similar to, but smaller (by ca. 100 jim) and more strongly sculptured on the proximal face than the presumed holotype of Verrutriletes compositipunctatus which is, in turn, slightly different from paratype speci- mens of this species in Dijkstra's (1949) collection (see Batten, 1988). It is also smaller, and sculptured with more bulbous verrucae than the single speci- men (holotype) of Verrutriletes dasyplocoides fig- ured by Schultz and Noll (1987, pl. 4, figs. 1,2).
Genus Bacutriletes van der Hammen 1955 ex Potoni6 1956
Bacutriletes ferulus sp. nov. Plate 4, figs. 4-6, 8, 9; Plate 13, figs. 14, 15
Diagnosis. Trilete megaspore, triangular in equa- torial outline with straight to convex sides and rounded angles. Maximum diameter 402-440 jLm (10 specimens). Triradiate ridge usually prominent, extending to more than 3/4 radius. Exoexine very thick (ca. 60 jlm), sculptured with bacula up to 17
jim in width and 30 jim high. Structure of sculptural elements and outer surface of exoexine is an open reticulate meshwork of sporopollenin threads but wall is much more compact beneath, consisting of laminae of globular elements connected in long rows. Intexine ca. 2 jim thick with reticulate outer, and smooth inner surfaces.
Holotype. Plate 4, figs. 4, 6, 9; sample GI32145. Derivation of Name. Latin ferulus, rod, staff, in
reference to the shape of the sculptural elements. Remarks. This species is distinguished from all of
the others that have been placed in Bacutriletes on the basis of its sculpture of densely distributed, elongate bacula.
Bacutriletes majorinus sp. nov. Plate 5, figs. 1-7; Plate 13, fig. 12
Diagnosis. Trilete megaspore, subcircular in equa- torial outline, subspherical in uncompressed state; proximal face slightly flattened. Maximum diameter 720(830)920 jim (15 specimens). Triradiate ridge relatively narrow (20 jim) but prominent (up to 60 jim high) and ornamented with bacula and papillae that are mostly ca. 15-30 jim in basal diameter and 10-50 jim in height. Sculpture similar elsewhere but elements may be somewhat shorter (<30 jim) or longer (up to 60 jim) in places and more widely spaced on distal face. Exoexine thick (ca. 60 jim), structure of outer part including surface features is an open reticulate meshwork of sporopollenin threads but towards interior it is more compact and granular in aspect. Adjacent to intexine, elements of reticulate meshwork are mainly orientated subparal- lel to inner surface to which a thin (ca. 2 jlm) intexine is attached.
Holotype. Plate 5, fig. 1; sample/slide 1597a, Dijkstra's collection.
Derivation of Name. Latin majorinus, of a larger kind of form, in reference to its overall dimensions.
PLATE 5
All electron scanning micrographs. 1-7 Bacutriletes majorinus sp. nov. 1 Holotype, x 100, sample/slide 1597a, Dijkstra's (SJD)
collection, x 100. 2 Sculpture adjacent to, and on, triradiate ridge, 1597a
(SJD), x 1000. 3, 6 Part of fragment showing exoexine in cross-section,
1597a (SJD). 3, sporopollenin meshwork compact in middle part of wall, becomes more open in construction towards outer surface, x 1500; 6, detail, x 5000.
4, 7 Fragment, 1597a (SJD). 4, illustrating membraneous intexine partly separated from thick exoexine, x 100; 7, part of papilla showing detail of construction, x 5000.
5 Intexine partly detached and folded back from exoexine of fragmented specimen, 1597a (SJD), x 2500.
8 Verrutriletes sp. cf. V. compositipunctatus (Dijkstra) Potoni6, proximal view, 1594 (SJD), x 250.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 5
" "ab~ " *C ~rtr.?*. * ?t.~~ ? sil rl~*rr~e~t rr?~~Ei~etlcaa~*~~~irr- i I ~l~sB C~'';ii~"lb~~lt~BII~~~d~S~E~jat~?I;~a~i* ?---' *r .1 a I R4r
i? .i r? .?1~1. * ,, !-~Es~ .!* r " **** :t: *E .;%
~ils: 'F 'Y r~?F ** r I d ~ ~.~.~?
~-? ?:i ii r
?i *a + * ~f3~ u a
Bi?I. I ~i"t! h iTo .. r
r. ...!- " ...-.*~I~V;t) ~.I): 1~?4 '* '", ???::::?r:: 1?: :?~* ?:e ~f ''i *" `*
~ax f~QP~' uZi?*; -~? Irilbr7. s' ':" ~Y~ ~e-~;?' C ??.q ?: "~n~il
i**. ?:::: i? *? 'P
*fi~t '~?*E:. ';~ '~s*? Y9a~i~~ ,,? "*r~
?, *?z ;? ~c~~Qw, :' c.av* I E :+*C; :" ?'.i 'L ~~*i? ;?
"arrt:rr*a *I;,*:c~ a If~k~TJ*I;X~: ?: :? ;* ~J*Cf~ '* r~ll? I*?*L+;C* *J ~+
I?. I ,.?? ~:*ltt,~.. ~ .i? ?ir 1C~+r~ *+ rr? ; 21:, ?f I ~ prr lc.~:, I* * * ?b * i 'I;+* Ii .J~:EZj~s -~r?
:?* u ,??1 .*+*;ISf~r I .* ?:? *i a.+0:..' '. ir~ rli, J:?r;JZ I?C * .' r r
.~ **r*~, ? *+* t J;
,* Irt I rrs~c. ' ri
~?:
'::: ?"t: :II i+ * .~.d.C::: -?? :-+:~i~qY~-~i~ I~c~~jq* i: ?-? i
i ~?t~Rk~"~;i~~ ~I r~~r?l~?~a?~j~;;d~F~*L *c?~?- *?i~ I? t?lpi- ~??:. +~~Ckls rE".. :;h :*C i*~Ys *IZ*~_. i *ll~-J '~Y T ~??; I-]~IFY.I .. I* ; ~--ru. ?. Y
-*,'L' I'u r ~F~;~,~"~~?~~~ ~I~Ell~*;ii~p,~aclt~a~"~?~rpl ~L~ia$BIB~g~_~tj' II~%s~B~I~~ i. J 2~ L?.?J' " _: ~I~ 4illil i
r* *'' ,rw;:5 ,? F*?.*.'I*L
it r~e~rr' ?..z .~ *r Z~c~~i*irr. ~s~c-,g~ia~;baQ?r~b.
q .,~ .. :F ~Ea(~i
P ??; ?E it: t .L '* r;~yrz:*t~ ?1 ?: a ?;? ~ .P
:???'~;. i:: t- '~5~ . I,~.~""i i: *Pi "' -~c~lm' s; ,t *Ct" ~"F~::r: ~s~!~,ci *? '~E~- .L*; --* ,ii'Z: C.* *~C). -?rlJ~~ ~~;iur~: V" ~ p~~g. ~
~i? '?*
,?C' ~i~~Sl~llli~Ol~~j~jr~ I~
L*? ~ r ~cPlb?~' ci~~- U1' & -, ?ii :i
~t; a a
~CZ~1 ~il~L~:!t~rlr~?rrrfa~~~r "1P?11 +~ ~II:: :??: 1 a .:::. II- .i::::!:l::: ~Ei~~a~ ??
??i~~cLI s?-~ , i:,,, , ? :" ~p?L,'-~? f J .* ~t~~i?~ ?: ::: "b i ??~: ll:iii 'f~?
i ?:? ;i:: I;ri ?::;:::: ?1.
?:I~. ~Ss~L~d~!: :i~:l:":?": ?.." :?' :~r -:? r .:"'"
~i ),ili~ii ~P :";? I" '" ';? * :::: I:::
:? ?:?~ 4f~a~"+d"i~a"~l~n~~~s~ri~'f sii? ~:: .:.. ??i:?i.:i:? I rllil ;'i .i. 'i~Pii .1?i~ ?- i.: * :::: :rS." ::-::. ~i~ij i*YB*~~ * S i iiii :.:i: n:
I ?r
?: ::::?P::' k a ~J~~ :r * ~ ?" :??
it~i?~" ?oi f'l n .. :i
J i~F~jq~q; ??:
.* ILL r C .." +? ??r r?P 1! ?:: ~ile~kr?l~e~-F?~s~~'~S~"lli~e;slt~,:l,~g ~~~_~fl~F~,YIIL-,F~'~*~~ 9E i?8 ::: 8:?: ??
_ ~ P :i ii? I i.. i ' * i: :gj:?;? r~? *
1: 4 5 o
'5'
?r
:1 P r~t~! T
i~p~S1~~f~. ??s ?;:? r ?~Ir ~ib?s t:k 'ii:
~i~:"u sv ~I
.i,. s
,.t* t * ...,;
a-tl.''?..L~L~s?? ilk ,3 ,i :: :~b~.? ~~:liii r ,ru~ ..... I I:
:i ?, * ;;~3? .~ 3 ;;; ~p~iL"
t, j ::::i??:
~i~ " 4iIYr~i~i~.z ;?s ,? ?,? ? ;..., ::
't;tQIIPk~"i~ll~)~Ei ..,.i~PY~ c~: j S .t ~9, i;E ~;-:? ..?f?? !:..~.1E.
Zi: t 7f ?r?;
?I;? 1~ d I" -~r
;':f!v1~. `;~;;; '~-~? ~jl$L' ' .??;. * %~4. rr. J~i~fi~i~ L?!':~Sl~.~s*i~lE~*c~PTI; i:l
::r:??? ?:.:? ;II?"i)
": :::.
ziig, :" ~? -' L"UL~Eors~a~r~rY- r8a ?i:i;lisiiil ?i?? ;; ~i
'Zh'l 'Yr '; ;;; *i:? ~s :Z ,.g. P ~Cii'i t
..., ~j~lbrc ~a~Fj~L~: i s
:J I .U'
9 ;1
P ;b r _;qC a; r6~1~ 8 -: * a
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

104 PALYNOLOGY, VOLUME 13 - 1989
Remarks. This species is distinguished from B. ferulus on the basis of its larger size and more globular shape, and in being sculptured with more widely dispersed, mainly conical rather than paral- lel-sided baculate elements.
Bacutriletes sp. Plate 6, figs. 1-3
Remarks. The tetrad illustrated is the only record of this form. The specimens of which it is composed are similar in size to Bacutriletes nanus (Dijkstra 1949) Potoni6 1956. They differ from those on which Dijkstra (1949) based his species (see scan- ning electron micrographs in Batten, 1988) mainly in being sculptured with smaller, more sharply pointed spinose elements and in having a very pro- nounced fine surface granulation; in addition, none of the spines appears to be basally constricted.
Genus Echitriletes van der Hammen 1955 ex Potoni6 1956
Echitriletes sp. Plate 6, figs. 4, 5
Remarks. This form is sculptured both proximally and distally with short (up to 25 iim), closely spaced echinae. These are most weakly developed and widely scattered on the prominent, narrow triradiate ridge, the rays of which are about half the radius of the spore in length.
Genus Erlansonisporites Potoni6 1956
Erlansonisporites scanicus sp. nov. Plate 6, figs. 6, 7; Plate 7, figs. 1-9,
Plate 8, figs. 1, 2; Plate 13, figs. 10, 11
Diagnosis. Trilete megaspore, circular to subcir- cular in equatorial outline. Maximum diameter 520(560)640 jtm (10 specimens). Triradiate ridge
simple, rays ca. '/2 radius, may be difficult to distin- guish from muri of robust reticulum which extend over both proximal and distal surfaces and are of similar width (26-48 jm at base) and height (30-60
jtm). Contact areas commonly delineated by arcuate
muri. Lumina and lower elevations of muri finely ornamented with granula, rugulae and/or a reticu- lum. At high magnification, structure of exine ap- pears reticulate at surface and muri are seen to be composed of a fine reticulate meshwork of sporopol- lenin threads and some granular elements, particu- larly where they connect. Outer part of exoexine is of similar construction but most of this thick wall (ca. 20-65 jtm)
is composed of a compact frame- work of regularly distributed spherical elements ar- ranged in variably orientated layers. Inner exoexine is difficult to differentiate from intexine, which ap- pears to vary from ca. 1-5 jlm in thickness; structure of both is a fine three-dimensional reticulum of sporopollenin threads.
Holotype. Plate 6, fig. 6; sample GI32107. Derivation of Name. After the province of Scania,
southern Sweden. Remarks. About half of the specimens of Erlan-
sonisporites scanicus recovered from the Asen pit have contact areas that are delineated by arcuate muri and lack any murornate incursions within them (e.g., Plate 7, fig. 4). The regular arrangement of layers of granular sporopollenin in the exoexine (Plate 7, figs. 5, 7-9) renders the majority iridescent in reflected light.
Associated with the specimens in Dijkstra's collec- tion are smaller spores, sometimes united (remains of tetrads; Plate 8, fig. 1), that are enveloped in closely-spaced muri with little or no intervening lumina; some are little more than crumpled murornate bodies apparently devoid of a triradiate flange. These are thought to be immature and shrunken, malformed specimens respectively.
No published species of Cretaceous megaspores bears much resemblance to E. scanicus. Erlan- sonisporites dubius Knobloch 1984b may be similar
PLATE 6
All scanning electron micrographs. 1-3 Bacutriletes sp., one specimen, S100515, sample
GI32117. 1, general view of tetrad, x 220; 2, close-up showing that entire outer surface consists of granular elements, x 1100; 3, detail, x 11,000.
4, 5 Echitriletes sp., one specimen, S100516, GI32107. 4,
general view, x 150; 5, detail of surface sculpture, x 1500.
6, 7 Erlansonisporites scanicus sp. nov. 6 Holotype, proximal view, S100517, GI32107, x 150. 7 Close-up of part of specimen illustrated on Plate 7, fig. 4,
showing sculpture of muri and lumina, S100518, GI32106, x500.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 6
??.di??:??i;il;l:?i:?i. i~?~~ *? """II''?!??:i;*???-?: 1 x.. ,r I : i'?116iii':i c-1* ; ? L. ;rira!i:?aa
:;;I~"" Lii~l ~1.. ~Jlr* ~,:. I~~*~iE~'I~LPaa~?r"?~sp;~is :ir
b' .I,
r
?*1C ? r
~ :i;,??,
1~ g t
z , r? ?rl r. 3;-~-~,',f ?t~~Cr*
,I*s~t~-~ ,;? '?'?r ?~ ,
*i ...a.~,L .. dii5! ~F~c :V ?": ??
:il ~ dFi
~,~F~??a~ ~FP g: 13 ~L~c*19 s -?l~a;?L~~'?.~e~Y~~~i Cbil ??? rt ~s~910, ~p ~e~-~-~ ?ae~b?-E-I?i~" gL
;p;~l~.~y~gE;, L '"*'::; *~?
,p... ??c: ~ 1..,. ,~
2 L? I ?-e 'L~ ~df~
r~~i~ ~e~3"~"l~""sl~"P"~c~~ ~Et~t~i"l?
~il .P
a ~ Sr?~jl)l? f** ?I' *~.C *.~* *
11J C* :r. B "t: *"* ii8'
, ~. r3d. ~t~lu
?'%i~*tj ~,."t: f *5 C '*' J ,,. "i'
?'? -~t~R u 5*t +. ,, ~pr 3 *nl: '''
,?* ?* ~Plrca~*l;'~lcl~urr---~IC;~~rlcrllPF~LI1
rJ". i -ae~Qlllap--4, I~E!t '"::a~,._-.,,.?;ll~?Z if: .I 'g r ?r:?1 rj 5 :y*r:~ rsl~s~ra~~ --r rr .:dfl~BEl~l.. ? + i?~;?da~b~*
II *~ '*" b ;~
;JL~: .~L
,trc; -?? ,~v ~r ~-_ila~rs~L -cr~p~ ,..P s* ..r ?5~s*- ~f?
1* r j?-_ -FT ~-~s~E~~IP~L~hL~Jlb '- 's-~bjlL~"lrr~~ '~yjC~;1E?~~ZC~j~~T~i:~
?'" *t 4 " 5 1 IP'Jt ?I?C X i C; iabh" a' r r ~ tr? ?,:
-i:ia a. - pu: 15 *1:1.%: 5~
*.~ ~Z~,~L~IF~Y~_ . ~? '5 ~. P: ?B -'~?:~PEarl~wrt~t~81~'$j: T:L'*i~p~7~li~&f*~~~i~.
i .r ?I. ~r , S;iCb ~E~
.aI:. .?, ?.?,??
-3LP~lrp~'i
.~~': ~i~:1?"~?~r~-s~?asee~ ~n~Ak~~:~ ~,,1YPr~.~;i~-?'~t~~: ~8~"~ii~i~rs~ ~'1? ?,::?
a .::;: ..:??:?:
I'i~P:;Y~J~k~ i~:j~!~%~l~~J'F~FYC~~ ~ '~.ysitL - -- -~~-?~~L1 -r~s~c~~: ~ ' - ~ eL C~ 'IpS-~~)i~'_~llS~T*"l:-~'W ~P~r~f~ b~.
:?r?:?
'P' r~? r- ;? I .il ;*.:??:
~E~l ~J~L~~J$ ~~ 1~J~p~ ??:??
~Eil:
I I III~C~EPI 4 I~U~3?PI~%PP???-Y"slrss: II ; I- d JT ~-?1I~JI?;~glF~':j:9i'*::: ?
lg'I ~F?I --""P84swas%;a;sllasi-` ir-~r~ ~F-s~ 7 ;:? i: i:;;i??`""":?? Is~ar~-~~ ar*~ra~?? ~;; 1 Ir I?:
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

106 PALYNOLOGY, VOLUME 13 - 1989
but Knobloch's illustrations do not show the form of the triradiate flange or reveal sculptural details, and there is no mention of the latter in his descrip- tion apart from a note on the presence of coni on the "Y-mark". His specimens appear to be rather poorly preserved.
Erlansonisporites septus sp. nov. Plate 8, figs. 3-7; Plate 13, fig. 13
Diagnosis. Megaspore circular to subcircular in equatorial outline. Maximum diameter 800(900)1000 jm (10 specimens). Triradiate flange distinct, ca. I/2 radius in length, 80-130 jm high, composed of a three-dimensional network of sporo- pollenin threads which expands outwards slightly along the upper extremities of the rays. Outer exoex- ine sculptured with muri of varying length that meander over surface and which, together with in- tervening lumina, form an irregular reticulum. Muri of similar construction to that of triradiate flange, but not so high (up to 1 jm).
Outer surface of exoexine and reticulum of muri composed of a fine meshwork of sporopollenin threads, but most of the thick (40-60 jtm) exoexine is made up of tightly packed laminae of granular elements.
Derivation of Name. Latin septus, fence in, en- close, in reference to the murornate sculpture.
Holotype. Plate 8, figs. 3, 4, 6; sample GI32123. Remarks. Some degraded specimens of this spe-
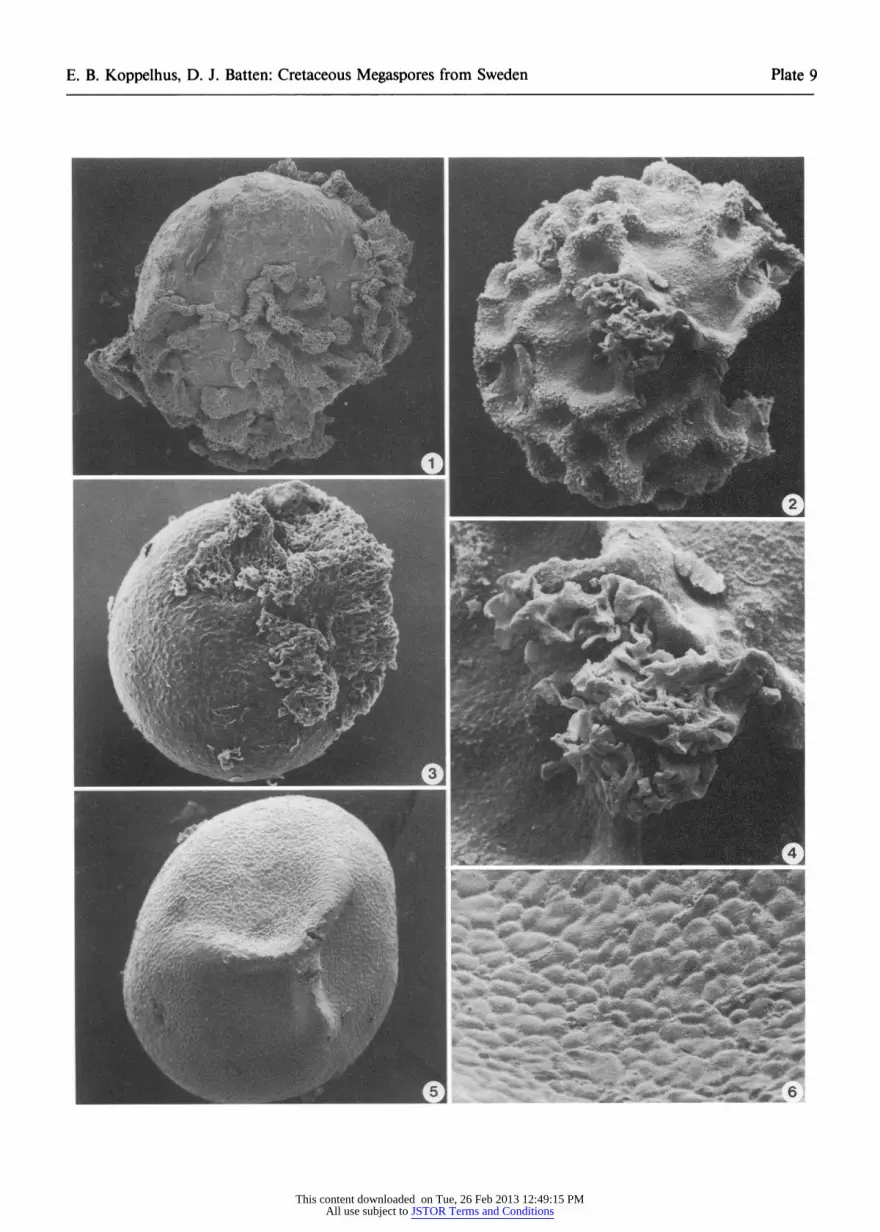
cies have very reduced muri but these remain firmly attached to the body of the spore (Plate 8, fig. 5). By contrast, the sculpture of other spores referred here to Rugotriletes costatus (see below) forms part of an only vaguely reticulate perisporal layer which tends to lift off the body of the spore (Plate 9, figs. 1, 3). The triradiate flange of these specimens is com- monly not readily discernible.
Genus Rugotriletes van der Hammen 1955 ex Potoni6 1956
Rugotriletes costatus (Dijkstra 1952) Potoni6 1956 Plate 9, figs. 1, 3
Triletes costatus Dijkstra, 1952, p. 167, pl. 7, fig. 4. Rugotriletes costatus (Dijkstra) Potoni6, 1956, p. 51.
Remarks. This species differs from Erlan- sonisporites septus in being much smaller and in having a poorly delineated triradiate flange and reticulate sculpture. Associated with the specimens that have lost some of their sculpture are bodies that have scabrate to weakly reticulate surfaces (Plate 9, figs. 5, 6). These are probably the completely de- nuded remains of either R. costatus or Ricinospora (Plate 10, figs. 1-6). Rugotriletes costatus should probably be transferred to another genus but we are reluctant to do this until further specimens have been found; Dijkstra based the species on only three spores (Batten, 1988).
Rugotriletes sp. Plate 9, figs. 2, 4; Plate 13, fig. 6
Remarks. In common with Rugotriletes diktyotus (Dijkstra 1949) Potoni6 1956 (see for example pl. 13, fig. 1 in Batten, 1988) the apical region of the trilete ridge of this form is obscured by an irregular, coarsely reticulate mass of sporopollenin. It differs, however, in that the outer surface of the exoexine is ornamented with finely granulate, rugulate and rather irregular elements; these are weakly devel- oped and widely dispersed on the lumina, but be- come larger and more closely spaced towards the top of the muri.
Genus Ricinospora Bergad 1978
PLATE 7
Erlansonisporites scanicus sp. nov. Figs. 1-5 are scanning elec- tron micrographs. 1-3 One specimen, S100519, sample GI32123. 1, oblique
orientation showing character of proximal and equatori- al regions, x 150; 2, close-up of surface of exine, x 500; 3, part of murus and adjacent lumen, x 2000.
4 Proximal view, S100518, GI32106, x 100. 5 Transverse section of spore; spherical elements of bulk of
exoexine arranged regularly in long rows; inner exoexine and intexine composed of threads orientated mainly
subparallel to inner surface forming a reticulate mesh- work; S100520, GI32123, x 5000.
6 Transmitted light photograph of transverse section of spore showing muri, exoexine and intexine, S100521, GI32146, x 500.
7-9 Transmission electron micrographs of ultra-thin sections of megaspore wall, S100522, GI32146. 7, section through entire wall, intexine on left-hand side, x 3000; 8, detail of inner to middle exoexine, x 6000; 9, part of outer exoexine, x 10,000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 7
.ii f~Si' ..
i?: c ?; s ~~E~ C. 'g r. '~3~6~E~L~ t.) ?IEP."
?i~E~~?t
*;?~i"~~ ?a: i* *i: ~Zc r?_.... .,,
'-~BBl[sF~F~ ~E? ~i3~ r ;5~F .J
i s2i~ ?*. d i. *
:,, ,. d:, .*f" I* * ,? i-??r ~ ]9~
;O i Fw~E,; r ?t r, .f:i~
?*' ,,Jr: .;?? ,?~.,.. ,,
??-I-?? ?ji?- ?:~. ?' *'~L~.: ~ r~t~?- ? i' ~~,~i13iB~s~Y~;~&o, '1~?3?IP~Lli-;81~?~~i?r~ I?,--?8%: ---- ILa srnf' C?l~~;~ ..i:..4 Y * * t,
?~?Tirrr~a. --~~8~8ba~rrs-~a~ ~~s-l ~rj jlt*
:"k~ll~IJIF ~~~YBF~::? .aa I ~- ~~1R~t~ll~ &B~C*jLlES.C
t '?: r "pr.
~'?i~?Li~
?rt jlF~' -' ?~ ?ifS~ZIQ~Fi~W~J~tiC$~~t~t~.f??_l
1 26 i~j~ll 31 U~a ,? :'.~jruY'C"'--?ll?rII i" .~.:1:~:~~:~i:iiiiiij(:": ""4r ;:::?? :::?.??:::::.
::::::::: ::?~ iiiiiiiiCijiii(:;ili ;diiP::::.::?:? :;::;
~;? v 4~Pl"r~3T! -~ I~P~~L~.`Sn ~ ilss~Asl~llYI~- - -u~"~- I. r... a i .I? ?~'4 * :? ::::: :illiiiiiiiiil :ai-? ""'' :::::. '* .:I P i
?rr? *' ~iswrr~J11~: ?L.li **:i" .. ,i
- ?Il~*t? : '~E?~' ~, ?.,.
J: ~; ct~l"li;i~Y~s~ ? ~" :1: : i::
r *f: ?rii r? L ??;:i~nL?" ..?. .a~n~_, ? ? ? . I
:?::, i'(
'" ~.lm~ ~C~;;: ~C i; r ' iiCJ :?-ii::::::.::.: ?:: : ?
??~hi? * ::??: _II ~?- ;? i' i: : **. *~ ~t~
1.1. evil ?::?.; . ?? ":'.
4]~I~C~I~GP111~5 --,- re i:.iii
r"~;~~l~r~~"4 I~i~Y 1~~~E~~~"r ~:?~j311LP;~i~r .~*IFL ~yrc;lly, I~l~l~~;?-~a~?rr;;f -""l?~k;j*~rrs~~ "YEL~?~PZPI i~fB~I~~~B~1?Clr
:. ly~ r' ~c~SIAL..1? ~ ~~----~?rr~c~.r rr~rr~wp~-? ~ ~-ILE111~ , 4.-?1 ~i I IIIF~IF .ill~ ~llsl'~lF?.~E*--I C 991t~ ~jgii:. y
j ~~Z ~NI~L?' i r
.I o 'F?
?,~-- LS.Y*E C~ iek,
rJ% I' ne.*x ~C~3Z: ,?r 7
''~Fg ~t~UIP~II*J~31~ ~L~fIICP~ ?Illlii ;*II~LUIF~IUIWI,'~I~~ 1IIIFBIFi~UsY~L ~y~Si~' ~I)Ei& ~il*C*l~srJBl~rPlsl~U~l~~~bl~lyll~Y ~IR~ _Z
r?
a ~:
:'Jd~i
~.1~~`7ha*r ''' iP*S I` ..~' ~L~P~FjlA'~P~ *PEE`IIIJPW;l~sa ~i~e~r;z~`~i" ~i~s ?ik
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

108 PALYNOLOGY, VOLUME 13 - 1989
Ricinospora sp. Plate 10, figs. 1-6
Remarks. The surface of this subspherical to ellip- tical megaspore is covered with a reticulate perispore of complex construction (Plate 10, figs. 2-5), the outermost parts of which are ornamented with pa- pillae (Plate 10, figs. 4, 5). The elevated triradiate ridge is enveloped by a spongiose mass that is also finely papillate on its outer extremities (Plate 10, figs. 1, 6). Where the perisporal layer has been eroded away, the surface of the exoexine is seen to be smooth to faintly reticulate (Plate 10, fig. 3).
This form bears some resemblance to Ricinospora cryptoreticulata Bergad 1978, but it differs in the manner of construction and ornamentation of the perispore.
Genus Thylakosporites Potoni6 1956
Thylakosporites retiarius (Hughes 1955) Potoni6 1956
Plate 10, figs. 7, 8; Plate 11, figs. 1-7; Plate 13, fig. 16
Triletes retiarius Hughes, 1955, p. 213, pl. 11, figs. 3, 4
Thylakosporites retiarius (Hughes) Potoni6, 1956, p. 49, pl. 6, figs. 55-57.
Description. Spores probably originally subspheri- cal but all are compressed. Maximum diameter 1120(1260)1400 pm (5 specimens). Trilete mark obscured by a spongiose layer of perispore. Surface of exoexine smooth, enveloped by a reticulate per- isporal net which may be partly or entirely removed from poorly preserved specimens. Muri of reticulum are composed of a robust network of sporopollenin threads (Plate 10, fig. 8) whereas surface of exoexine is made up of more slender elements (Plate 11, fig. 2). Width of muri ca. 30 pm; diameter (up to 90 pm) and shape of lumina very variable. Exoexine
light brown in color, often iridescent, consists of an outer layer of closely packed globular elements con- nected by short rods of sporopollenin (Plate 10, fig. 7; Plate 11, figs. 3, 5, 7). Structure becomes more open towards interior except adjacent to intexine where threads and globules are again tightly ad- pressed (Plate 11, fig. 5). Intexine is much thinner (ca. 5 pm) and constructed of interwoven threads (Plate 11, figs. 3-6).
Remarks. The specimens of Thylakosporites reti- arius recovered from Asen differ from those illus- trated by Hughes (1955) in that the triradiate ridge is obscured by a perisporal mesh. Their wall layering is, however, entirely comparable with that recorded previously for both this species (Hueber, 1982) and some forms of Erlansonisporites (in Bergad, 1978; Collinson et al., 1985).
Genus Tenellisporites Potoni6 1956
Tenellisporites sp. cf. T. aachenensis (Dijkstral949) Potoni6 1956
Plate 11, fig. 8; Plate 12, figs. 1, 2, 4, 5; Plate 13, figs. 7, 8
Description. Trilete megaspore, circular to subtri- angular in equatorial outline. Maximum diameter including zona and capilli (appendages) 378(509)810 pm (12 specimens). Trilete mark reaches equator and is bordered by capilli that are closely spaced and fused at their bases, 54(64)81 pm in length and 10(11)16 jim wide. Capilli 64(75)108 pm long and 10(19)32 jlm wide also extend from equatorial flange. Contact area of proximal face is irregularly reticulate (Plate 11, fig. 8; Plate 12, figs. 5, 8). Distal surface bears ca. 20 to 25 appendages with bluntly rounded tips (bacula) 48(65)81 jm in length and 16(21)27 jim in diameter. Exine is yel- lowish brown in color, consists of an exoexine ca. 12-14 jim thick, which is made up of interwoven threads orientated mainly subparallel to outer sur-
PLATE 8
All scanning electron micrographs. 1, 2 Erlansonisporites scanicus sp. nov. 1 Presumed immature specimens with poorly developed
lumina, sample/slide 1597b, Dijkstra's (SJD) collection, x 150.
2 Detail of lumina on another specimen showing structure of outer exoexine, 1591a (SJD), x 10,000.
3-7 Erlansonisporites septus sp. nov.
3, 4, 6 Holotype, S100523, sample GI32123. 3, oblique view, x 80; 4, structure of outer surface of exoexine, x 5500; 6, enlargement of murus to show structure, x 550.
5 Lateral view of degraded specimen, much of sculpture and triradiate flange removed, S100524, GI32106, x 100.
7 Detail of structure of middle part of exoexine of frag- mented specimen, S100525, GI32123, x 10,000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 8
r*? *) '?r'-e L~* 8: L ?~-~"~ :~fi~ ?o i.. t~? r~jr~?? ? : ?~
~ ..II 1~I r ?i? 'i QLILF~ i,, ?:; i
:"?? Ii?i! ?;r ..9 ?* 61i~i~)?
::~rllii~ ':" ~t ? ;? ~iirh. :::.
.:?:;"::: ?::: ?:* ?;"' 'iii~i~ii?ii? :: ? ii:.
I Yir! ':7 :?ii :s ?:. : t
.Ylftlij~jii~ii;Bii ji~i(ia ~c~i~l~ij~i~lk~ a;:::: r.*. - jrsg~iwr~s~i~-i ?-, iii~i~'~ ?1 Pb rll u:r~
:c. ?,,
, ?r? 2~??t,
~ki~
t ?.3L . * Iri -4J ~~?
t 1C; ?( .??~? -..s*
11...:i :,? : i t ?? "(I .i
sl-?: ~ ri Y -- k '1: 1
.n? ?:? ? f :?diP: U 'ti6iiiii Clfi; ~~ia~j~i .I C~? 'il ~" :: t liriE1Cr:i ??: ?r: I?* 1~1Si~i~~llp;~Lirr: i~i. :i? re ?? ?i :;;;; 9; ?..:? i? :: 4!' ?:~
~i~J~i~(?r ?; I' 71i
i:i
*f: ~II~YI ,i:
r~z~- f) -i~8~19~:? ,I-
e, P ::
iiiiil;. .i. 1F: iii. r* :i:"i'f'i6 ????:II
.r. ?~jpg~~a7L .?:l~::'~lii~ii*IILII8HKuruunr::~;it~ -::: ?:ii::i;~;~i~Fl1C~EEffFi~P: I?iiiliii;iai .i ::::?; ?? ?w' :i:::? -:
'1 I sC~i13t~ 2;
?::i,~:iiiiiitjii'iir'lii-~ilr. i:; j ) '1 I::::. .a?.lii??? ...::?:? i? ? . ~ :% j ".....""'' :!8ij,,i..?:?':i:..iilli:?:?::::::' ..::'"iikCiIlrYB!!I:? ?I:? irl~i~?l~?~:?il?ilPEIl:O"ti ifiiiiitiiriiiiiiiiii;;riiil.siiiii:?lill:::i:: ''X* :???????
"
:ii~iii~Yi, P a?;I' ..s:. ,~I
*:?e~J"e~l ~Jr llrcs :4 ~1~-
:?~p~' Its - -te , *L
*lr. Lf; .rCr7~lt~F;~ F~ 1* L? ** ~* ?!?C
" ~ ::i st L
~A :pr E:. I ? :?, :`*tl ?, :~
~i~ak ::. * * *L
;i3 i?: i?*r? 1? ~ i: *
QF: *?. ?, ~1151 :I *s? 2~~ v ?liSvi+?i, ,? r ri?: .??,.
; *. i-? * ~i 1~' ~? 1* ;? Lli
"? ??. k' *? jf *? C'
.... "I "'?lti~~ ."-. .1
I` ,::. cr? t -?! -.??* ~I? ~Ci Y?.~;?~3~2E::f:9r?.LJ'
I * :'rt I? ~"?~F.I, 9~: Ip;l i a(I) ?t?:;. :IlitiiEi~L~~.~?L
~y~ -p?~. .. *. n
... ??:..
, ": ; ~-~- t .B ill~~ *; . S ?~~. ."*sqlj~.c
:ai ii: ii:. ::? i ?~1~... c, ii:::::l. ?, Y5kd;? .:; ii
ii: * ?-- ????l?,lii. L~ac, i
~n ~c~~,~ , ?" -a~ '` ~*?~fJ~ '* ,. ~"' * t~ t;; =r %? ?;; 1 r?''P -4: "irr. "* ~B~p?:, ;~
;.i I*'r; I?':LEL $ ?a ??* ru?? * 1?
" --~~ L"lsar~l~c:l - '* ~I(k ?~i~ta~?~p?"?si;~~f .* ?:? *1 (? ~? i- -rJ ?: C.
r 3" .. ~~ 2- .* ~lll~k~ ;1 ~i~? :?~**crk~ ??-
I*r 1ICZ~Y$IIP a~t~. ?Ir 1 ,.
:1 t~:,* r r
,r P' ""' 1) "'?r ilr; r~ I f F 5
Y ~Qli: I? h'
r ~I i. rir..;i
C t*.5 't *?r
r: ,k ~-~ .:. .:: ,~ a? ,~t? " ..
*IE: SLll~li~ * iis ~p?~ .~ ::: ~ :~~?t~ ?C, ** ~L~-~I~~E rJ ~C~rs~- if ~'fll :?~P~IB s--,-rern~e~g*~ r
;? r *
r- -Iri~il r ~i~ r F' :~lt s ...:
?:? h~?i~?'~ X r 1 r ??e ~;s~J;il&~~ 7 ii'.... 13 ILC"IL:~' ;ji :Jrr* 1
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

110 PALYNOLOGY, VOLUME 13 - 1989
face, and a thin (ca. I rim) intexine that lacks any visible structure (Plate 12, fig. 2).
Remarks. Vangerow's (1954) specimens are simi- lar to ours but those on which Dijkstra based the species (Dijkstra, 1949; Batten, 1988) differ in bear- ing coni 10-15 jm in basal diameter and 10-20 jm high. The undivided inner zone of the equatorial flange is also less pronounced. At present, however, we do not think that these differences are suffi- ciently important to justify the separation of a new species for our material.
Tenellisporites sp. cf. T. tenellus (Dijkstra 1949) Potoni6 1956
Plate 12, figs. 3, 4, 6, 7
Remarks. The morphology of the two specimens found is similar to that of T. tenellus but the sculp- ture, especially that on the distal face, is more strongly developed. One specimen is also larger (480 jim) than any of those in the type assemblage (maxi- mum 450 jLm; see Batten, 1988, p. 8).
BOTANICAL RELATIONSHIPS
The structure of the wall of modern megaspores has been shown in a number of publications (e.g., Martens, 1960 a, b; Stainier, 1965, 1967; Pettitt, 1966; Kempf, 1970; Sievers and Buchen, 1970; Robert et al., 1973; Morbelli, 1977; Tryon and Lugardon, 1978; Tryon and Tryon, 1982; Minaki, 1985; Kovach, 1989) to be characteristic for particu- lar species and groups of heterosporous plants. This has encouraged some palynologists to examine frag- mented fossil spores under the SEM and/or ultra- thin sections in the TEM in the hope of finding a basis for suggesting the botanical affinities of the plants that produced them (e.g., Kempf, 1971a, b, 1973; Hall 1974, 1975; Bergad, 1978; Hueber, 1982; Kovach and Dilcher, 1985, 1988; Li and Batten, 1986; Li et al., 1987; Batten and Ferguson, 1987;
Batten, 1988; Taylor and Taylor, 1987, 1988). We adopted a similar approach in our study of the Asen assemblage.
We were unable to observe the wall structure of some of the species because of a dearth of suitable specimens. Of those for which useful information was obtained, none suggests affinities with either the Salviniaceae or the Marsileaceae. This is not surpris- ing in view of the external morphology of the species concerned, which does not conform to that of the spores referred so far to these groups (e.g., in Hall, 1975; Dorofeev, 1981). The lack of representation of "water ferns" at Asen is noteworthy, however, be- cause their spores do occur in beds of probable Santonian age within the Aachen Formation of the Limburg region of the Netherlands and neighboring Belgium and West Germany (Dijkstra, 1949, 1952; Batten, 1988). Since the two deposits are probably closely similar in age, perhaps even contemporane- ous, the fact that they have also yielded relatively few other taxa in common may be attributable to environmental rather than stratigraphic differences.
We have shown that the exoexine of Tenel- lisporites sp. cf. T aachenensis consists of a three- dimensional network of sporopollenin threads, the dominant orientation of which is parallel to the outer surface of the spore. This construction is identical to that described for other species of the genus (Batten, 1988) and is also typical of several other genera which are characterized by having membraneous equatorial and triradiate flanges (e.g., Minerisporites, Herbosisporites and Paxillitriletes in Li et al., 1987, and Batten, 1988). It is, in addition, comparable to the form of the exine beneath the siliceous perispore of modern isoetalean megaspores (see Pettitt, 1966). Despite the lack of a mineralized outer layer, it is quite possible that all of these Cretaceous spores were produced by plants related to Isoetes or its ancestors.
Further evidence in support of such an interpreta- tion for Tenellisporites is provided by the fact that small monolete microspores have been found to occur in abundance with these megaspores in the
PLATE 9
All scanning electron micrographs. 1, 3 Rugotriletes costatus (Dijkstra) Potonim. 1 Sculptured perisporal layer partly removed, sample/slide
1587b, Dijkstra's collection, x 100. 3 Almost all of sculptured perispore missing from distal
and equatorial regions, S100526, GI32189, x 100.
2, 4 Rugotriletes sp., one specimen, S100527, GI32106. 2, proximal view, x200; 4, detail of reticulate mass of sporopollenin at apex of triradiate ridge, x 500.
5, 6 Denuded exoexinal remains of either Rugotriletes cos- tatus or Ricinospora, S100528, GI32156. 5, general view, x 150; 6, close-up of reticulate surface, x 700.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions
E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 9
? ~ .
. ,?
.. .. ...:.?.
?
... . , ..... '.. ...
:ii ' ' i:.i ......... .. . .. ......... .... .
?: .... ? , ?...
?i ... ? ':
..i
'. *. ? . .? i'
.'i?? .J
._YC
"rri .-..- .=":i .... ?
:? " - ...i , : ? ...
.: .. : j -~ ~~ , 'i ',i: s
?r :a~ ........;.' ::: ... .
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

112 PALYNOLOGY, VOLUME 13 - 1989
Hergenrath Clay Member of the Aachen Formation (Batten et al., 1988). Similar associations have also been recorded previously for both Minerisporites and Paxillitriletes (Batten, 1969, 1975; Kovach and Dilcher, 1985). All suggest that the affinities of the parent plants lie with the Isoetales. By contrast, microspores of selaginellalean affinity are typically triradiate (see, for example, Tryon and Tryon, 1982).
The exoexine of the megaspores produced by extant Selaginella galeotii and S. myosurus has received special attention since the 1960s (e.g., Stainier, 1965; Kempf, 1970; Tryon and Lugardon, 1978; Minaki, 1985) because, along with a few other species, it is composed in part of tightly packed, interconnected granular layers of sporopollenin which give the wall an iridescence in reflected light. When viewed in cross-section under a scanning electron microscope or in thin section under a TEM, these laminae have a very distinctive appear- ance that varies according to their orientation.
Walls of similar construction have been encoun- tered previously in some fossil species of the mega- spore genera Erlansonisporites, Horstisporites, Rici- nospora and Rugotriletes (Bergad, 1978; Hueber, 1982; Batten, 1988; Taylor and Taylor, 1988). The exoexine of both of our new species of Erlan- sonisporites (E. scanicus and E. septus) is of this type, as is that of the smooth walled form identified as Trileites persimilis. Hence, another name can now be added to the generic list above.
It is not possible to compare the construction of the exoexine of the other Asen spores so closely with that of modern species. The robust but generally less granular, and sometimes very open meshwork of sporopollenin in part, is nevertheless consistent with a selaginellalean interpretation (cf. Minaki, 1985). This is supported in some instances by the gross morphology of the spores. There seems little doubt in our opinion, for example, that the affinity of Bacutriletes majorinus lies with the Selaginellales.
ENVIRONMENT OF DEPOSITION
In common with interpretations of the occurrence of probable isoetalean spores in other sequences, the presence of Tenellisporites in the majority of the assemblages recovered from the lower part of the Asen section and their dominance in two of these (GI32188 and 89; Text-Figure 3 and Table 1) sug- gest that a low energy environment of deposition prevailed at this time. A floodplain marsh to lacus- trine habitat may be inferred from both present-day distributions of species of Isoetes and previous sedi- mentological interpretations of the sequence (Grionwall, 1914; Lundegren, 1931, 1934; Ross, 1949; Nykvist, 1957).
By contrast, all but one of the beds sampled higher up the section lack Tenellisporites. Those species that have been recorded in largest numbers, namely Trileites persimilis, Erlansonisporites scanicus, E. septus and Thylakosporites retiarius, are all presumed to have selaginellalean origins, as indicated in the preceding section. Coupled with a greater variation in sedimentary facies, including the presence of several root-bearing horizons, this sug- gests that conditions of deposition were generally more fluviatile than they had been earlier. It follows that the plants which shed these spores may well have been mostly riparian.
CONCLUSIONS
There are very few Late Cretaceous deposits in Europe and Scandinavia that are suitable for the recovery of megaspores. This is because much of what had been land and areas of non-marine deposi- tion during the Early Cretaceous was submerged by the "Chalk Sea." Although the composition of the assemblage described does not at present refine pre- vious age determinations of late Santonian or early Campanian for the Asen succession, it may do so in
PLATE 10
All scanning electron micrographs. 1-6 Ricinospora sp. One specimen, S100529, sample
GI32123. 1, proximal view, x 150; 2, part of sculptured perispore, x 1000; 3, higher magnification showing retic- ulate surface of exoexine, x 3000; 4, showing construc- tion of complex columellate-reticulate and papillate perispore, x 1000; 5, papillae, x 1000; 6, lateral view, x 200.
7, 8 Thylakosporites retiarius (Hughes) Potoni6. 7 Transmission electron micrograph of ultra-thin section
of exoexine showing tightly packed laminae of globular sporopollenin elements, S100530, GI32188, x 4000.
8 Surface of murus, sporopollenin elements more robust than those of the lumina (see Plate 11, fig. 2), S100531, GI32123, x 10,000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 10
rrr ??,: t+l; C'*C *"*: ?litm" ;?
* :F .. P ??I~?: ..L
~ ~e~t~ jg c? i ~
11 31i. ~t4i c ;r iiY*pi~ :~C~1~J~8~3"F~
~7nr.11~ rt ~ ~i ~L~..s
14i: 'b'~i;k2~ ?r-ak~J
.;r tl ". ~'? f ?". ,~jic* 6* c?
;~iir :crt i~r :ii
.3 i~?~ ,'* ~P aBt~,~$~E~a~: ~ ,I* f :r
?. , 6?~ I r. :.~CF~ r ?d;; i~iakc .??riiii )11 rEr~rr~r~rsl~~lvt~"?lilr*
r?" ~ n''1
I ?~ ji~k? ~tr ?: ? llS ~k~: r " :i:
3 ~21 u t Ji~ ?I ?i? ~~r;-:~:i a~i~8 ~e2~8i~aC~ . -!!t ?.
*.. ? ?? 3 .f~;? L ~i ???;: ..,. d: rE~;drr '" ?ii? :i ~ ..) c??
I '? " + b ?? : ?? ~ :? U? ~r~f~c.
r i! ?I ~ rj: c ??d ::*~t i ~.? ,e ,"~sI ,.. , 4 ,: ?Iti, i. a' ~t;C~ I, c ~ ~ i. r? -.
';** * ~Z": ~j"-~~
rj +~1E " 5 i" ? 6~ i .O ai~ f;* a? :~sc3jr" i:4 ?:: .J
a ?CX * o? :pa if --P ~:?
1 a?yo;? ? i?- i 3 J, Br ?..:b?
?:t'i ~~* '*
i~ ilrr ~. - i
.r, ,. ? ??s ? : :t? ic~ ~I?*~ ~p
?r
i ~C 9~,.rii 3E. utiY*
.i4 :j~- 'rCYc. rctk a ?E!
; it" 2* r '~? ~5~?*""lsS ~~E I~?~am-- ~i~?~i~B~84~f~?~~~~II
:?' ~r :r
t~i"?il~8Ee~&~i~i8$iii~lPf~$"~?~i~, i ri
r f
~ ? ,:*f ~~.i~t~t ~ r? ~ :ii ? ~ixn -~~~g~~Elsr~a~ II- - I i: ~II C'
I 'Z r i'" .~E;: T"s ~ L::: 4 O c ?i I :i,
-* i; ~&a?~ li:~r*~??
,.i i: u ;;; -% ????1 rr? ,, ;~o~ :* C r j:i n
~* ~:~ ?~b~ f. ?lilr
()10 '" ?::i'L! * r h .?i;.. b:? *? .*?'9~ ~iir~ I r ?i~x ? . CI,
1, - i i
;fl II I les~ ~ilr~i91 rlwrrr~i ~' : *
P~~YII~1~:ir 6
cg~6~,~ Ft r"Ci~Y~ Is~
~.:~ -;~; r ?r
i:?: ? 1FE?3;1~"~::ar ---?l?-i?~lrus"sm~.~Bss~ReU~si~" .?-~' ...:?
;:.r:i,?t~?t~~~i~???? ~'~:;(i ?P' fn .:: .~i~;???~
:e, i? srl? ? ..:k .:.'? !I
pbi ~~.
7 JII~Pk~S~S~:8 U
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

114 PALYNOLOGY, VOLUME 13 - 1989
the future once the stratigraphic ranges of Senonian megaspores become better known.
The discovery of Selaginella galeotii-type exoex- ine structure in a species of Trileites means that it is now known to occur in five fossil genera. It appears to be more widespread in Cretaceous spores than perhaps expected when first encountered. Previous sedimentological interpretations of a change in depositional regime from lacustrine to predomi- nantly fluvial conditions are broadly supported by differences in the composition of the megaspore assemblages recovered from the lower and upper parts of the succession.
ACKNOWLEDGMENTS
This paper is based in part on a thesis by E. B. Koppelhus prepared at the University of Copenha- gen under the supervision of Dr. Kaj Raunsgaard Pedersen and Prof. Else Marie Friis, whose advice and encouragement is gratefully acknowledged. The collaborative study was made possible by financial support from the Geological Survey of Denmark which covered both time for research and travel costs. D. J. Batten is grateful to Drs. J.G.M. van de Laar and H.W.J. van Amerom of the Geologisch Bureau, Rijks Geologische Dienst, for permission to work on S.J. Dijkstra's collection of Cretaceous and Palaeocene megaspores.
References Cited
BATTEN, D.J. 1969 Some British Wealden megaspores and their facies dis-
tribution. Palaeontology, 12: 333-350, 6 pl. 1975 Wealden palaeoecology from the distribution of plant
fossils. Proceedings of the Geologists' Association, 85 (for 1974): 433-458, 1 pl.
1988 Revision of S.J. Dijkstra's Late Cretaceous megaspores and other plant microfossils from Limburg, The Netherlands. Mededelingen Rijks Geologische Dienst, 41-3: 1-55.
BATTEN, D.J., and FERGUSON, D.J.P. 1987 Cabochonicus, a new genus for species of gemminate
megaspores previously referred to Verrutriletes. Journal of Micropalaeontology, 6(1): 65-75.
BATTEN, D.J., DUPAGNE-KIEVITS, J., and LISTER, J.K. 1988 Palynology of the Upper Cretaceous Aachen Formation
of northeast Belgium. In: Streel, M. and Bless, M.J.M. (eds.), The Chalk district of the Euregio Meuse-Rhine. Natuurhistorische Museum Maastricht and Laboratoire de Palontologie de l'Universit6 d'Etat & Liege, p. 95- 103.
BERGAD, R.D. 1978 Ultrastructural studies of selected North American Cre-
taceous megaspores of Minerisporites, Erlan- sonisporites, Horstisporites and Ricinospora, n. gen. Palynology, 2: 39-51.
CHRISTENSEN, W.K. 1975 Upper Cretaceous belemnites from the Kristianstad
area in Scania. Fossils and Strata, 7: 1-69.
1984 The Albian to Maastrichtian of southern Sweden and Bornholm, Denmark: a review. Cretaceous Research, 5: 313-327.
COLLINSON, M.E., BATTEN, D.J., SCOTT, A.C., and AYONGHE, S.N.
1985 Palaeozoic, Mesozoic and contemporaneous mega- spores from the Tertiary of southern England: indica- tors of sedimentary provenance and ancient vegetation. Journal of the Geological Society, London, 142: 375- 395.
PLATE 11
1-3 and 8 are scanning electron micrographs. 1-7 Thylakosporites retiarius (Hughes) Potoni6. 1-3 One specimen, S100531, sample GI32123. 1, equatorial
view; triradiate ridge partly obscured by spongiose peri- spore on right-hand side, x 75; 2, surface of exoexine between muri of reticulum, showing three-dimensional network of sporopollenin threads, x 10,000; 3, trans- verse section of damaged part of spore coat; tightly and regularly packed granular exoexine and underlying fi- brous intexine, x 5500.
4 Transmitted light photograph of transverse section
through spore showing thick exoexine and membrane- ous intexine, S100532, GI32188, x 125.
5-7 Transmission electron micrographs of ultra-thin sec- tions; S100533, GI32188. 5, part of tightly packed mid- dle layer of exoexine; inner exoexine of more open structure with thin intexine (i) closely adhering to this, x 3000; 6, part of inner zone of exoexine and intexine; x 10,000; 7, middle zone of exoexine x 10,000.
8 Tenellisporites sp. cf. T aachenensis (Dijkstra) Potoni&, proximal view, S100534, GI32189, x 150.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 11
i
~ ~ .......
Ii~L
~~ ir" C
:il
.= +.i~?..
~tx~`"u~~ "~~rn
"%W ~J~.c~ r
i i!.13 .li a -i~i Il~ *...
.:=Si~i~i@.(!i x~c a :~ ~ ~ ~~~E
-i:i:i;iu is
....
.. . .: .,,...
.. .: r~ ?;
%' 13
-i
*I:i-.
B f
Tj[
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

116 PALYNOLOGY, VOLUME 13 - 1989
DIJKSTRA, S.J. 1949 Megaspores and some other fossils from the Aachenian
(Senonian) in south Limburg, Netherlands. Mededel- ingen van de Geologische Stichting, Nieuwe Serie, 3:19- 32, 2 pl.
1951 Wealden megaspores and their stratigraphical value. Mededelingen van de Geologische Stichting, Nieuwe Serie, 5, 7-22, 2 pl.
1952 The stratigraphical value of megaspores. Compte Rendu Troisidme Congrbs pour I'Avancement des Atudes de Stratigraphie et de Giologie du Carbonifere, Heerlen, 1951, 1: 163-168, 3 pl.
DOROFEEV, P.I. 1981 On the systematics of Tertiary Marsileaceae.
Botanicheskiy Zhurnal, 66: 792-801, 4 pl. (in Russian). ERDTMAN, G.
1947 Suggestions for the classification of fossil and recent pollen grains and spores. Svensk Botanisk Tidskrift, 41: 104-114.
FRIIS, E.M. 1983 Upper Cretaceous (Senonian) floral structures of
Juglandalean affinity containing Normapolles pollen. Review ofPalaeobotany and Palynology, 39: 161-188.
1984 Preliminary report of Upper Cretaceous angiosperm reproductive organs from Sweden and their level of organisation. Annals of the Missouri Botanical Garden, 71: 403-418.
1985 Actinocalyx gen. nov., sympetalous angiosperm flowers from the Upper Cretaceous of southern Sweden. Review of Palaeobotany and Palynology, 45: 171-183.
FRIIS, E.M., and SKARBY, A. 1981 Structurally preserved angiosperm flowers from the Up-
per Cretaceous of southern Sweden. Nature, 291: 485- 486.
1982 Scandianthus gen. nov., angiosperm flowers of saxifra- galean affinity from the Upper Cretaceous of southern Sweden. Annals of Botany, 50: 569-583.
FRIIS, E.M., CRANE, P.R., and PEDERSEN, K.R. 1986 Floral evidence for Cretaceous chloranthoid angio-
sperms. Nature, 320: 163-164. 1988 Reproductive structures of Cretaceous Platanaceae. Bi-
ologiske Skrifter, Det Kongelige Danske Videnskabernes Selskab, 31: 1-55.
GRONWALL, K.A. 1914 Nord6stra Skines kaolin och kridtbildningar. Sveriges
Geologiska Undersc'kning Arsbok, 8: 77-100.
HALL, J.W. 1974 Cretaceous Salviniaceae. Annals of the Missouri Botani-
cal Garden, 61: 354-367. 1975 Ariadnaesporites and Glomerisporites in the Late Creta-
ceous: ancestral Salviniaceae. American Journal ofBot- any, 62: 359-369.
HARRIS, T.M. 1935 The fossil flora of Scoresby Sound, East Greenland. Part
4: Ginkgoales, Coniferales, Lycopodiales and isolated fructifications. Meddelelser om Gr 4land, 112, 1: 176 p., 29 pl.
1961 The Yorkshire Jurassic Flora I. Thal- lophyta-Pteridophyta. British Museum (Natural His- tory), London, ix + 212 p.
HUEBER, F.M. 1982 Megaspores and a palynomorph from the Lower Poto-
mac Group in Virginia. Smithsonian Contributions to Paleobiology, 49: iii + 69 p.
HUGHES, N.F. 1955 Wealden plant microfossils. Geological Magazine, 92:
201-217, 3 pl. KEMPF, E.K.
1970 Elektronenmikroskopie der Sporodermis von Mega- sporen der Gattung Selaginella (Pteridophyta). Review ofPalaeobotany and Palynology, 10: 99-116.
1971a Electron microscopy of the megaspore Horstisporites semireticulatus from Liassic strata of Germany. Grana, 11: 18-22.
197 1b Electron microscopy of Mesozoic megaspores from Denmark. Grana, 11: 151-163.
1973 Transmission electron microscopy of fossil spores. Palaeontology, 16: 787-797.
KNOBLOCH, E. 1984a Megasporen aus der Kreide von Mitteleuropa. Sbornik
Geologickf)ch VId, Paleontologie, 26: 157-195, 12 pl. 1984b Megasporen aus der Kreide (Oberconiak-Santon) der
Bohrung Volfartice (Nordbohmen). Casopis pro Miner- alogii a Geologii, 29: 155-165, 8 pl.
KOVACH, W.L. 1989 Quantitative methods for the study of lycopod mega-
spore ultrastructure. Review of Palaeobotany and Paly- nology, 57: 233-246.
KOVACH, W.L., and DILCHER, D.L. 1985 Morphology, ultrastructure, and paleoecology of Paxil-
litriletes vittatus sp. nov. from the mid-Cretaceous (Ce- nomanian) of Kansas. Palynology, 9: 85-94.
PLATE 12
All scanning electron micrographs. 1, 2 Tenellisporites sp. cf. T aachenensis (Dijkstra) Potoni6. 5, 8 1 Distal view, S100535, sample GI32189, x 150. 2 Transverse section showing spongiose exoexine un-
derlain by a thin, membraneous intexine, S100536, GI32189, x 5500.
5 Polar view, S100537, GI32189, x 150.
8 Lateral view, S100538, GI32189, x 150. 3, 4 Tenellisporites sp. cf. T. tenellus (Dijkstra) Potoni6. 6, 7 3 Proximal surface between muri showing three-dimen-
sional sporopollenin meshwork, S100539, GI32106, x 10,000.
4, 6, 7 One specimen, S100540, GI32106. 4, lateral view, x 150; 6, close-up of proximal surface, x 2000; 7, detail of bases of equatorial capilli, x 5000.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 12
,.i ....' . :+. &? .-
?L ? " ,i':: .j
?
+++ t+,+,i --I~i .
.,,
P +: .,,:, +++. ,.+.. . ..
?:'" . , ,'
,t:i~ . .m, :
". ? ?
'6 Fk F ?-p
'~? "+:' . .. .
10 +? +L?~ i.,+ " :
??ii~' ,-: ?. +? ? + ,,+,..
? :
... <. .P ,+!.+ +++ , .+. ..,. ,,
?L ' ,
, i "::IE~~ I~F;
*~~~ ....... '+.-
-.. .;i;a ,
?~~ . <+ .
? . t
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

118 PALYNOLOGY, VOLUME 13 - 1989
1988 Megaspores and other dispersed plant remains from the Dakota Formation (Cenomanian) of Kansas, U.S.A. Palynology, 12: 89-119.
LI WENBEN, and BATTEN, D.J. 1986 The Early Cretaceous megaspore Arcellites and closely
associated Crybelosporites microspores from northeast Inner Mongolia, P.R. China. Review of Palaeobotany and Palynology, 46: 189-208.
LI WENBEN, BATTEN, D.J., ZHANG DAHUA, and ZHANG LIANGDE.
1987 Early Cretaceous megaspores from the Jalainor Group of northeast Inner Mongolia, P.R. China. Palaeonto- graphica, Abteilung B, 206: 117-135, 9 pl.
LUNDEGREN, A. 1931 De Kretaceiska ler- och sandfbrekomsterna N om Ivos-
jon. Geologiska Fireningens i Stockholm Fdrhandlin- gar, 53: 298-320.
1934 Kristianstadsomridets Kridtbildningar. Geologiska Foreningens i Stockholm Forhandlingar, 56: 126-307.
MARCINKIEWICZ, T. 1971 The stratigraphy of the Rhaetian and Lias in Poland
based on megaspore investigations. Instytut Geo- logiczny, Prace, 65: 1-58, 22 pl. (in Polish, English summary).
MARTENS, P. 1960a Sur une structure microscopique orientre, dans la paroi
mrgasporale d'une S6laginelle. Compte Rendu Hebdomadaire des Siances de l'Acadimie des Sciences, Paris, 250: 1599-1602, 2 pl.
1960b Nouvelles observations sur la structure des parois mrgasporales de Selaginella myosurus (Sow.) Alston. Compte Rendu Hebdomadaire des Scances de l'Acadimie des Sciences, Paris, 250: 1774-1775.
MINAKI, M. 1985 Macrospore morphology and taxonomy of Selaginella
(Selaginellaceae). Pollen et Spores, 26 (for 1984): 421- 479.
MINER, E.L. 1932 Megaspores ascribed to Selaginellites, from the Upper
Cretaceous coals of western Greenland. Journal of the Washington Academy of Sciences, 22: 497-506
MORBELLI, M.A. 1977 Esporas de las especies Argentinas de Selaginella (Se-
laginellaceae-Pteridophyta). Obra del Centenario del Museo de La Plata, 3:121-150.
MORNER, N.-A. 1983 The Santonian/Campanian boundary; paleomagnet-
ism, sea level changes, biostratigraphy and sedimentolo- gy in SE Sweden. Abstracts, Symposium on Cretaceous Stage Boundaries, University of Copenhagen: 128-131.
NYKVIST, N. 1957 Kretaceiska vedrester vid Asen i Skine. Svenska Skog-
svcrdsfOireningens Tidskrift, 55: 477-481. PETTITT, J.M.
1966 Exine structure in some fossil and recent spores and pollen as revealed by light and electron microscopy. Bulletin of the British Museum (Natural History), Geol- ogy, 13: 221-257, 21 pl.
POTONIE, R. 1956 Synopsis der Gattungen der Sporae dispersae. I.
Sporites. Beihefte zum Geologischen Jahrbuch, 23: 103 p, 11 pl.
ROBERT D., ROLAND-HEYDACKER, F., DENIZOT, J., LAROCHE, J., FOUGEROUX, P., and DAVIGNON, L.
1973 La paroi m~gasporale de l'Isoetes setacea Bose ex De- lile. Adansonia, serie 2, 13: 313-332.
ROSS, N.-E. 1949 Investigations of the Senonian of the Kristianstad Dis-
trict, S. Sweden I. On a Cretaceous pollen and spore bearing clay deposit of Scania. Bulletin of the Geological Institution of the University of Uppsala, 34: 25-43.
SCHULTZ, G., and NOLL, H. 1987 Die Megasporen-assoziation in den unterkretazischen
Sedimenten einer Paliokarsthohle bei Nehden im Sauerland (Rheinisches Schiefergebirge). Palaeontographica, Abteilung B, 203: 83-107, 7 pl.
SIEVERS, A., and BUCHEN, B. 1970 Uber den Feinbau der wachsenden Megaspore von
Selaginella. Protoplasma, 71: 267-279. SKARBY, A.
1964 Revision of Gleicheniidites senonicus Ross. Stockholm Contributions in Geology, 11: 59-77, 3 pl.
1968 Extratriporopollenites (Pflug) emend. from the Upper Cretaceous of Scania, Sweden. Stockholm Contributions in Geology, 16: 1-60, 30 pl.
1974 The status of the spore genus Cibotiidites Ross. Stock- holm Contributions in Geology, 28: 1-7, 3 pl.
1978 Optical properties of fossil Schizaea spores from the Upper Cretaceous of Scania. Grana, 17: 111-123.
PLATE 13
1-16 Transmitted light photographs; all magnifications ca. x 60 except where stated.
1 Trileites persimilis. 2 Trileites murrayi. 3 Trileites sp. 4 Maexisporites soldanellus. 5 Maexisporites sp. 6 Rugotriletes sp. 7, 8 Tenellisporites sp. cf. T aachenensis, proximal (7) and
distal (8) surfaces. 9 Cabochonicus carbunculus. 10 Erlansonisporites scanicus.
11 Erlansonisporites scanicus, tetrad, x 30. 12 Bacutriletes majorinus, x 30. 13 Erlansonisporites septus, x 30. 14, 15 Bacutriletes ferulus, one specimen; proximal and distal
faces respectively. 16 Thylakosporites retiarius, x 30. 17 The western part of the Hoganis AB quarry from which
most of the samples were collected (see Text-Figure 3). The complete section (arrow) consists of ca. 20 m of lacustrine and fluviatile sediments separated into lower and upper units respectively by a clearly delineated weathered horizon (star).
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

E. B. Koppelhus, D. J. Batten: Cretaceous Megaspores from Sweden Plate 13
.....
.. "~~'1 ICI
...?rr~~1 411~pr 1 " l
?~ ?., .
.. .
!~ i ??
"'
??
'' ""
C
" : . . , .
1 2 ? i" 13 ' , 1 " '... ? . ....
1
? '- '~~? ',. . " L /~~" .. . 1
; , . .. ? ... . . ,. : .r: ?
:.I..,. o.. .,
?,. r:~~" *7I ""~F~
o.. '*?U "
*~?a
:'... . ' 2 , .. .
:? ? t i]
.........e! : . .
.. . .. . . ... . ... . . . .. . . . ... ..! i ii ; ! : ii ,. .. .. ;+ , .
. ..,..v.. . =... ,....... ...
"? ..j . .? . .zg . ='. .. ".
F ''i ;..?..=::', :'::,-, y .
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions

120 PALYNOLOGY, VOLUME 13 - 1989
1986 Normapolles anthers from the Upper Cretaceous of southern Sweden. Review ofPalaeobotany and Palynol- ogy, 46: 235-256.
SKARBY, A., and NILSSON, L. 1982 Krittidsblommor frin Skine - 78 miljoner Ar gamla.
Fauna och Flora, 77: 281-290. STAINIER, F.
1965 Structure et infrastructure des parois sporales chez deux S6laginelles (Selaginella myosurus et S. kraussiana). La Cellule, 65: 221-244, 5 pl.
1967 Morphological study of the walls of the mega- and micro-spores of Selaginella myosurus (Sw.) Alston. Re- view ofPalaeobotany and Palynology, 3: 47-50.
TAYLOR, W.A., and TAYLOR, T.N. 1987 Subunit construction of the spore wall in fossil and
living lycopods. Pollen et Spores, 29: 241-247.
1988 Ultrastructural analysis of selected Cretaceous mega- spores from Argentina. Journal of Micropalaeontology, 7(1): 73-87.
TRYON, A.F., and LUGARDON, B. 1978 Wall structure and mineral content in Selaginella
spores. Pollen et Spores, 20: 315-340. TRYON, R.M., and TRYON, A.F.
1982 Ferns and allied plants, with special reference to tropical America. Springer Verlag, New York, xiv + 857 p.
VAN DER HAMMEN, T. 1955 Principios para la nomenclatura palinologica sistemati-
ca. Boletin Geologico, 2(2) (for 1954): 3-21. VANGEROW, E.F.
1954 Megasporen und andere pflanzliche Mikrofossilien aus der Aachener Kreide. Palaeontographica, Abteilung B, 96: 24-38, 1 pl.
This content downloaded on Tue, 26 Feb 2013 12:49:15 PMAll use subject to JSTOR Terms and Conditions





























