Embed Size (px)
Citation preview

Gene Therapy (2002) 9, 327–336 2002 Nature Publishing Group All rights reserved 0969-7128/02 $25.00
www.nature.com/gt
RESEARCH ARTICLE
LCR-mediated, long-term tissue-specific geneexpression within replicating episomal plasmid andcosmid vectors
C-M Chow1,4, A Athanassiadou1,2, S Raguz1,4, L Psiouri1,2, L Harland1,5, M Malik1, MA Aitken1,F Grosveld3 and M Antoniou1
1Nuclear Biology Group, Division of Medical and Molecular Genetics, GKT School of Medicine, Guy’s Hospital, London, UK;2Department of General Biology, Faculty of Medicine, University of Patras, Patras, Greece; and 3Erasmus University MedicalGenetics Centre, Department of Cell Biology, Rotterdam, The Netherlands
Locus control regions (LCRs) are transcriptional regulatoryelements, which possess a dominant chromatin remodellingand transcriptional activating capability conferring fullphysiological levels of expression on a gene linked in cis,when integrated into the host cell genome. Using the human�-globin LCR (�LCR) as a model, we show that this class ofcontrol element can drive high levels of tissue-specific geneexpression in stably transfected cultured cells from within anEpstein–Barr virus-based plasmid REV. Furthermore, a 38-kb �LCR minilocus-REV cosmid vector was efficientlyretained and maintained therapeutic levels of �-globin trans-gene expression in the absence of drug selective pressure
Keywords: episomal vectors; EBV; locus control region
IntroductionThe use of replicating episomal vectors (REVs)1 offers anattractive alternative to integrating retroviral and adeno-associated viral (AAV) vectors for producing long-termgene expression within a gene therapy context. First,REVs do not pose the same size limitations on the thera-peutic transcription unit as do viral vectors, with insertsin excess of 300 kb being a possibility.2 Second, being epi-somal, REVs do not suffer from potential hazards associa-ted with insertional mutagenesis that is an inherent prob-lem with integrating viral vectors. Lastly, REVs areintroduced into the target cells using nonviral deliverysystems that can be produced more cheaply at scale thanwith viral vectors.3–5
REVs based on viral origins of replication such as thosefrom EBV,6 human papovavirus BK,7,8 BPV-19 and SV4010
hold the greatest promise. Combinations of viral and
Correspondence: M Antoniou, Nuclear Biology Group, Division of Medi-cal and Molecular Genetics, GKT School of Medicine, 8th Floor Guy’sTower, Guy’s Hospital, London Bridge, London SE1 9RT, UKCurrent addresses: 4Gene Regulation and Chromatin Group and Mem-brane Transport Biology Group, MRC Clinical Sciences Centre, Ham-mersmith Hospital, Du Cane Road, London W12 0NN, UK; and 5Bioinfor-matics Group, Central Research Division, Pfizer Limited, Sandwich, KentCT13 9NJ, UKReceived 21 September 2001; accepted 18 December 2001
over a 2-month period of continuous culture equivalent to atleast 60 generations. This demonstrates for the first time thefeasibility of using REVs for gene therapy of the haemo-globinopathies. Importantly, our results demonstrate that asin the case of integrated transgenes, expression from withinREVs is prone to silencing but that the inclusion of the �LCRprevented this repression of gene function. Therefore,appropriate control elements to provide and maintain tissue-specific gene expression, as well as the episomal status ofREVs is a crucial feature in vector design. Our data suggestthat LCRs can contribute to this vital function.Gene Therapy (2002) 9, 327–336. DOI: 10.1038/sj/gt/3301654
mammalian chromosomal origins of replication havealso been found to improve nuclear retention of theepisomes.11,12
It has been demonstrated that both non-replicating,transiently transfected plasmids,13,14 as well asREVs,13,15,16 assemble nucleosomes. Assembly on REVs ismore organised and resembles native chromatin, whereasnucleosomes on transient plasmids are less well orderedallowing greater access of transcription factors to targetsequences.14 These observations imply that transcrip-tional control elements with a potent chromatinremodelling capacity may be required for efficient long-term tissue-specific gene expression from within REVs.One such class of elements are locus control regions(LCRs).17 LCRs are tissue-specific regulatory elementswhich are able to overcome chromatin position effectsand confer upon a gene linked in cis, site-of-integration,physiological levels of expression directly proportional totransgene copy number in mice.18 LCRs are able toobstruct the spread of heterochromatin and prevent pos-ition effect variegation (PEV).19 This pattern of expressionconferred by LCRs suggests that these elements possessa powerful chromatin remodelling capability and are ableto establish and maintain a transcriptionally competent,open chromatin domain. Therefore, LCRs are seen as themost efficient means of achieving reproducible, physio-logical levels of gene expression from stably integratedtransgenes.

Episomal vectors and locus control regionsC-M Chow et al
328
Gene Therapy
The requirements for tight tissue-specific geneexpression from within REVs, at a sustained level to beof long-term therapeutic benefit, has to date been poorlyaddressed. We have therefore assessed the ability ofLCRs to drive efficient, tissue-specific gene expressionfrom within REVs using the human �-globin LCR (�LCR)as a model system. The �LCR consists of five elementsspread over 16 kb of DNA located 5� of the entire �-glo-bin gene family and which are characterised by high sen-sitivity to digestion with DNaseI.17 Components of the�LCR have been incorporated into both retroviral andAAV vectors.20 The �LCR is seen as an essential part oftranscription units to reproducibly generate sufficientlyhigh levels of �-globin to be of value in the gene therapyfor the haemoglobinopathies.20,21
In this study, we have assessed the expression of thehuman �-globin gene either alone or in combination withelements of the �LCR within plasmid REVs and showthat long-term, high levels of erythroid-specificexpression can be conferred by this element. In addition,a 38-kb �LCR minilocus cosmid REV is efficiently propa-gated in an episomal state and retains high levels of �-globin gene expression. This demonstrates the potentialutility of this system for gene therapy applications.
Results
Plasmid episomal replication and copy number ofLCR/�-globin gene constructs in K562 and HeLa cellsThe �-globin gene either alone or in combination withone or more of the three transcriptionally most potentDNaseI hypersensitive (HS) sites (HS2, 3 and 4) that con-stitute the �LCR17 were inserted into the EBV-based plas-mid vector p220.2 (Figure 1).6 These constructs were thenused to generate stable transfected pools of the humanmyelogenous leukaemia cell line K562. These cells consti-tutively express the embryonic �- and foetal �-globin, butnot the adult �-globin genes.22 However, it has beenshown that a stably integrated human �-globin transgeneunder control of the �LCR linked in cis, is unlike theendogenous �-globin genes, efficiently expressed in K562cells.23 Current models suggest that this is due to theabsence of a �-globin gene on the transgene construct thatwould otherwise compete for the �LCR and be preferen-tially transcribed.17 This allows the �-globin reporter onthe REVs to be quantified without interference from thehomologous endogenous genes. In addition, four keyconstructs (�, 2�, 3� and 432�) were introduced intohuman HeLa cells, which acted as a non-erythroidcontrol.
The episomal status of the �LCR/�-globin gene plas-mid constructs was determined as follows. Total DNAfrom the transfected pools of K562 cells was digestedwith BamHI and HindIII and Southern blotted. Theserestriction enzymes liberate the p220.2 vector backboneas an 8.9 kb fragment (Figure 1a) only if the constructsremain as independent episomes. These blots wereprobed using the plasmid pBluescript that possessesextensive homology to the pBR322 region of p220.2. Theresults (Figure 1b), show a single band corresponding tothe size of linearised p220.2 indicating that within K562cells, the �LCR/�-globin gene episomal constructs werepresent only as non-integrated episomal DNA. Similarresults were obtained for constructs transfected intoHeLa cells (data not shown).
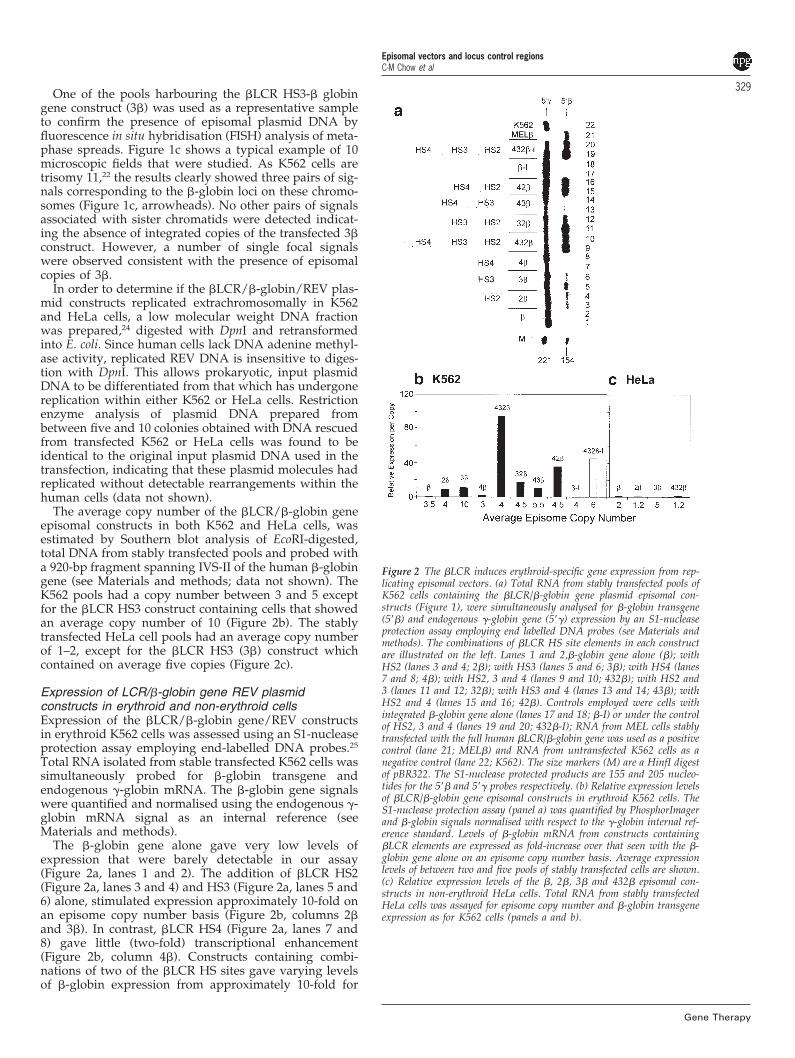
Figure 1 Structure of the EBV-based plasmid, �LCR/�-globin gene con-structs and confirming episomal status in stably transfected tissue culturecells. (a) Illustration of �LCR/�-globin gene episomal constructs. Thehuman �-globin gene as a 4.1-kb EcoRV fragment was inserted into ablunted SalI site in the polylinker of the EBV-based plasmid vector p220.2.DNaseI hypersensitive site numbers 2, 3 and 4 of the �-globin LCR eitheralone or in combinations (see Figure 2) were then cloned into the newlyintroduced SalI site upstream of the �-globin gene. (b) Southern blotanalysis of K562 cells stably transfected with the �LCR/�-globin geneepisomal constructs. Total DNA from stable transfected pools of K562cells was digested with BamHI and HindIII to separate �-globin fromEBV vector sequences, resolved on a 0.7% agarose gel, Southern blottedand hybridised with 32P-labelled pBluescript plasmid as a probe to assessthe integrity of the p220.2 vector backbone. Only a single band of 8.9 kbcorresponding to the size of linearised p220.2 is observed in all pools indi-cating that only episomal copies of the constructs are present. Lanes 1–8correspond to samples from cells transfected respectively with constructs�, 2�, 3�, 4b, 432�, 32�, 43� and 42� (Figure 2). Lane C, untransfectedK562 cells as a negative control. (c) Confirmation of episomal status byfluorescence in situ hybridisation (FISH). The �LCR HS3-� globin geneconstruct (3�) was used as a representative sample to confirm the presenceof episomal plasmid DNA by FISH analysis of metaphase spreads. A typi-cal example of 10 microscopic fields that were studied is shown. The sig-nals corresponding to the �-globin loci on the three copies of chromosome11 present in K562 cells are seen (yellow arrowheads). No other pairs ofsignals associated with sister chromatids were detected indicating theabsence of integrated copies of the transfected 3� construct. A number ofsingle focal signals are observed, also visible in the interphase nuclei tothe right, consistent with the presence of episomal copies of 3�.
Episomal vectors and locus control regionsC-M Chow et al
329One of the pools harbouring the �LCR HS3-� globingene construct (3�) was used as a representative sampleto confirm the presence of episomal plasmid DNA byfluorescence in situ hybridisation (FISH) analysis of meta-phase spreads. Figure 1c shows a typical example of 10microscopic fields that were studied. As K562 cells aretrisomy 11,22 the results clearly showed three pairs of sig-nals corresponding to the �-globin loci on these chromo-somes (Figure 1c, arrowheads). No other pairs of signalsassociated with sister chromatids were detected indicat-ing the absence of integrated copies of the transfected 3�construct. However, a number of single focal signalswere observed consistent with the presence of episomalcopies of 3�.
In order to determine if the �LCR/�-globin/REV plas-mid constructs replicated extrachromosomally in K562and HeLa cells, a low molecular weight DNA fractionwas prepared,24 digested with DpnI and retransformedinto E. coli. Since human cells lack DNA adenine methyl-ase activity, replicated REV DNA is insensitive to diges-tion with DpnI. This allows prokaryotic, input plasmidDNA to be differentiated from that which has undergonereplication within either K562 or HeLa cells. Restrictionenzyme analysis of plasmid DNA prepared frombetween five and 10 colonies obtained with DNA rescuedfrom transfected K562 or HeLa cells was found to beidentical to the original input plasmid DNA used in thetransfection, indicating that these plasmid molecules hadreplicated without detectable rearrangements within thehuman cells (data not shown).
The average copy number of the �LCR/�-globin geneepisomal constructs in both K562 and HeLa cells, wasestimated by Southern blot analysis of EcoRI-digested,total DNA from stably transfected pools and probed witha 920-bp fragment spanning IVS-II of the human �-globingene (see Materials and methods; data not shown). TheK562 pools had a copy number between 3 and 5 exceptfor the �LCR HS3 construct containing cells that showedan average copy number of 10 (Figure 2b). The stablytransfected HeLa cell pools had an average copy numberof 1–2, except for the �LCR HS3 (3�) construct whichcontained on average five copies (Figure 2c).
Expression of LCR/�-globin gene REV plasmidconstructs in erythroid and non-erythroid cellsExpression of the �LCR/�-globin gene/REV constructsin erythroid K562 cells was assessed using an S1-nucleaseprotection assay employing end-labelled DNA probes.25
Total RNA isolated from stable transfected K562 cells wassimultaneously probed for �-globin transgene andendogenous �-globin mRNA. The �-globin gene signalswere quantified and normalised using the endogenous �-globin mRNA signal as an internal reference (seeMaterials and methods).
The �-globin gene alone gave very low levels ofexpression that were barely detectable in our assay(Figure 2a, lanes 1 and 2). The addition of �LCR HS2(Figure 2a, lanes 3 and 4) and HS3 (Figure 2a, lanes 5 and6) alone, stimulated expression approximately 10-fold onan episome copy number basis (Figure 2b, columns 2�and 3�). In contrast, �LCR HS4 (Figure 2a, lanes 7 and8) gave little (two-fold) transcriptional enhancement(Figure 2b, column 4�). Constructs containing combi-nations of two of the �LCR HS sites gave varying levelsof �-globin expression from approximately 10-fold for
Gene Therapy
Figure 2 The �LCR induces erythroid-specific gene expression from rep-licating episomal vectors. (a) Total RNA from stably transfected pools ofK562 cells containing the �LCR/�-globin gene plasmid episomal con-structs (Figure 1), were simultaneously analysed for �-globin transgene(5��) and endogenous �-globin gene (5��) expression by an S1-nucleaseprotection assay employing end labelled DNA probes (see Materials andmethods). The combinations of �LCR HS site elements in each constructare illustrated on the left. Lanes 1 and 2,�-globin gene alone (�); withHS2 (lanes 3 and 4; 2�); with HS3 (lanes 5 and 6; 3�); with HS4 (lanes7 and 8; 4�); with HS2, 3 and 4 (lanes 9 and 10; 432�); with HS2 and3 (lanes 11 and 12; 32�); with HS3 and 4 (lanes 13 and 14; 43�); withHS2 and 4 (lanes 15 and 16; 42�). Controls employed were cells withintegrated �-globin gene alone (lanes 17 and 18; �-I) or under the controlof HS2, 3 and 4 (lanes 19 and 20; 432�-I); RNA from MEL cells stablytransfected with the full human �LCR/�-globin gene was used as a positivecontrol (lane 21; MEL�) and RNA from untransfected K562 cells as anegative control (lane 22; K562). The size markers (M) are a HinfI digestof pBR322. The S1-nuclease protected products are 155 and 205 nucleo-tides for the 5�� and 5�� probes respectively. (b) Relative expression levelsof �LCR/�-globin gene episomal constructs in erythroid K562 cells. TheS1-nuclease protection assay (panel a) was quantified by PhosphorImagerand �-globin signals normalised with respect to the �-globin internal ref-erence standard. Levels of �-globin mRNA from constructs containing�LCR elements are expressed as fold-increase over that seen with the �-globin gene alone on an episome copy number basis. Average expressionlevels of between two and five pools of stably transfected cells are shown.(c) Relative expression levels of the �, 2�, 3� and 432� episomal con-structs in non-erythroid HeLa cells. Total RNA from stably transfectedHeLa cells was assayed for episome copy number and �-globin transgeneexpression as for K562 cells (panels a and b).

Episomal vectors and locus control regionsC-M Chow et al
330
Gene Therapy
43� (Figure 2a, lanes 13 and 14; Figure 2b, columns 43�)to 35-fold for 42� (Figure 2a, lanes 15 and 16; Figure 2b,columns 42�). The highest levels of expression, anapproximately 95-fold increase over the �-globin genealone, were observed under the control of all three of theHS sites tested (432�; Figure 2a, lanes 9 and 10; Figure2b) giving a maximum of 13.6% of endogenous �-globinmRNA levels per episome copy.
In order to determine whether the �LCR HS sites main-tain their ability to drive strict erythroid-specific geneexpression from within an REV context, a number of keyconstructs were analysed in non-erythroid HeLa cells.Expression of the human �-globin gene was determinedby the same S1-nuclease protection assay that was usedto analyse the transfected K562 cells. The ubiquitouslyexpressed hnRNPA2 mRNA was used as internal refer-ence to normalise the �-globin signal (see Materials andmethods).
Although a signal of human �-globin mRNA could bedetected from cells transfected with the �-globin genealone, the addition of HS2, HS3 or the HS234 combi-nation, failed to enhance expression (Figure 2c). Thesedata demonstrate that the �LCR confers tissue-specificexpression from within REVs.
Maintenance of episomal status and gene expressionIn order to test the maintenance and stability of geneexpression of the �LCR/�-globin/REVs within K562 cellsas a function of time, stable transfected pools containingthe �-globin gene with �LCR HS2, HS3 and the HS234combination were cultured continuously for 30 days inthe absence, as well as presence of hygromycin drug-selective pressure. Southern blot analysis was used asbefore (Figure 1b) to assess episome copy number overthe 30-day period of culture. The results showed that inthe presence of hygromycin there was no significant dropof episomal copy number, whereas in the absence ofselection there was a gradual decrease (data not shown).However, this rate of episomal loss varied between poolsand appeared to largely depend upon the copy numberat the initial stages of the transfection before removal ofhygromycin from the culture medium. In some casesthere was almost complete loss of episomes (�1 copy percell) by day 30 (data not shown).
Since the rate of episome loss was different for eachpool, �-globin gene expression was determined at thepoint where 50% of the episomes were lost from the cul-tures. In the absence of hygromycin, the 432� constructmaintained �-globin gene expression per copy at a simi-lar (80–100%) level to that observed before commence-ment of the 30-day period of culture in the absence of thedrug (Figure 3a, �Hygromycin; 432� columns). How-ever, the 2� or 3� episomes both showed a significantdecline in �-globin gene expression after hygromycinwas removed (Figure 3a, �Hygromycin). The pools withthe 2� construct gave 10–20% of the original value of �-globin gene expression per episome copy, whilst the 3�construct showed only 4–6%. Surprisingly, similar resultswere obtained with the parallel cultures growing in thepresence of hygromycin (Figure 3a, +Hygromycin).Despite no obvious drop in episomal copy number, �-globin gene expression markedly decreased over 30 daysof culture in the pools harbouring either 2� (41–59%) or3� (6.5%) when compared with the level seen on day 1(Figure 3a, +Hygromycin; columns 2� and 3�). In con-
Figure 3 Combinations of �LCR elements are required to maintain geneexpression. (a) Pools of stable transfected K652 cells harbouring constructs2�, 3� and 432� (Figures 1 and 2) were grown continuously in the pres-ence (+Hygromycin) and absence (�Hygromycin) of drug-selective press-ure for 30 days. Total RNA/DNA isolated at periodic intervals duringthis time was analysed for episome copy number and �-globin transgeneexpression (Figure 2a). The +Hygromycin results shown correspond tothe 30-day time-point. The results from cultures grown in the absence ofhygromycin are taken after a 50% decrease in episome copy number wasobserved and correspond to 30 days for the 2� and 3� pools and 10 daysfor the 432� pools. Levels of �-globin mRNA are expressed as a percentageof the starting value present after the initial selection of transfected clonesand before the commencement of the 30-day culture period with and with-out drug-selective pressure. Each bar in the histogram represents an inde-pendently generated, transfected pool of cells. (b) K562 cells stably trans-fected with 432� and grown continuously for 30 days in hygromycincontaining medium (panel a, +Hygromycin), were analysed by DNAFISH (Figure 1c; see Materials and methods) to assess for the presence ofintegrated copies of this construct. A typical metaphase spread of 10–12microscopic fields analysed is shown. The �-globin loci on the three copiesof chromosome 11 are clearly visible (yellow arrowheads). No other pairsof signals associated with sister chromatids were seen indicating theabsence of transgene integration events.
trast, 432� maintained �-globin gene expression at thesame level throughout the 30-day culture period (Figure3a, +Hygromycin; columns 432�). FISH analysis con-firmed that this resulted from episomal, rather than inte-grated copies of the 432� construct (Figure 3b).

Episomal vectors and locus control regionsC-M Chow et al
331General DNaseI sensitivity of 432� and 3�The silencing of transgene expression in K562 cells con-taining human �-globin under �LCR HS3 control (3b;Figure 3a), suggests changes in the chromatin structureof these REV molecules13,15,16 to a transcriptionally non-permissive state. We therefore determined the generalDNaseI sensitivity of the 3� REV episomes in K562 cellsafter 30 days of continuous culture in the presence ofdrug-selective pressure, during which time �-globin geneexpression is silenced, and compared this with cells con-taining the �LCR HS234-driven construct (432�), wheregene expression is retained over the same period (Figure3a). Nuclei were prepared from these transfected poolsof cells and digested with limiting amounts of DNaseI.Total DNA from these nuclei was then digested with XbaIand after gel electrophoresis and Southern blotting, washybridised with a probe consisting of a genomic fragmentspanning the second intron of the human �-globin gene.This probe detects an 11-kb fragment from the endogen-ous �-globin locus, a 4.5-kb band from 432� and 6-kbband from the 3� REV episomes (Figure 4a and b). Theintensities of these signals were then quantified by Phos-phorImager. Since endogenous �-globin does not expressin K562 cells, the chromatin structure around this genewithin the native locus should remain similarly accessibleto DNaseI digestion over the 30-day period of culture.Therefore, the ratio of the intensities of the endogenous�-globin band with respect to that of the episome-derivedband was used as a measure of the latter’s rate of disap-pearance and hence overall sensitivity to DNaseI diges-tion (see Materials and methods). If DNaseI digests theendogenous and episome-derived �-globin fragments ata similar rate before and after the 30-day period of con-tinuous culture, their ratio over the DNaseI digestion ser-ies will remain similar at these two time-points. How-ever, if the episomal chromatin structure becomes moreresistant to DNaseI digestion, the ratio between the twobands will be altered. As shown in Figure 4c, in the caseof 432�, the trend in the ratios over the DNaseI digestionseries before and after 30 days of culture, are similar sug-gesting that there is no obvious change in the generalDNaseI sensitivity of the plasmid REV chromatin. How-ever, in the case of 3� (Figure 4d), the �-globin genebecomes more resistant to DNaseI digestion after 30 daysas demonstrated by an increasing endogenous to episo-mal �-globin gene ratio indicating the formation of a lessDNaseI-sensitive structure. Similar results were obtainedfrom two independently transfected pools.
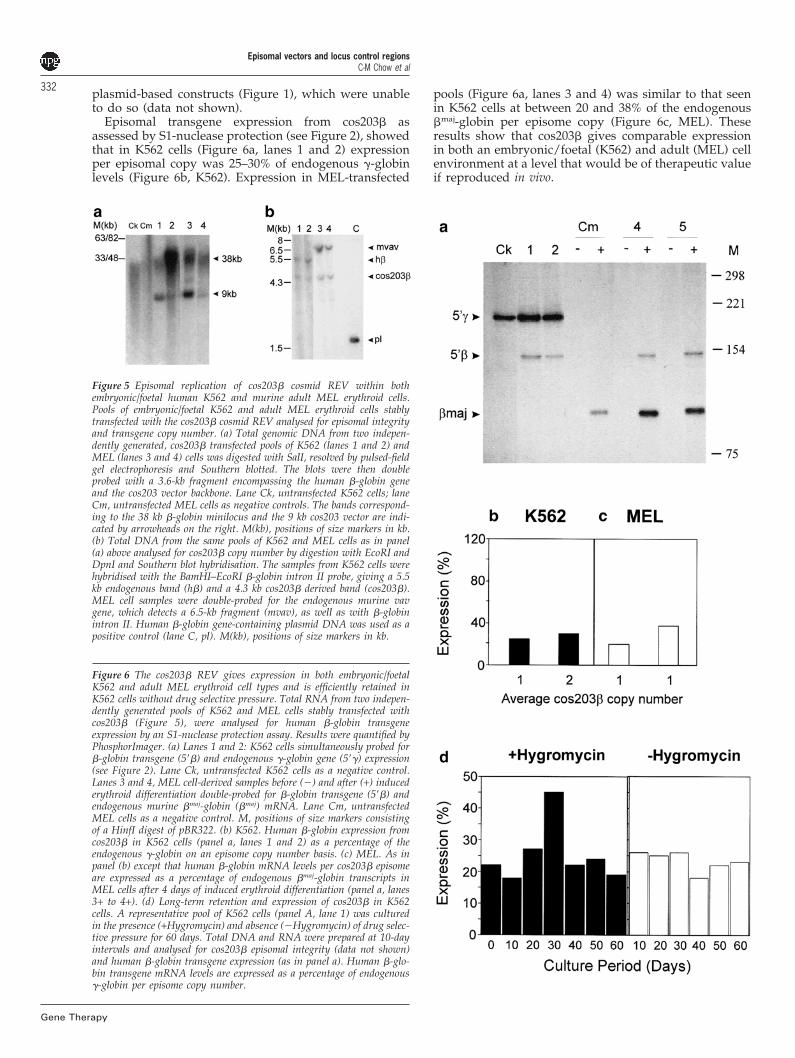
High-level gene expression from �LCR minilocus REVsThe results obtained with the p220.2 plasmid-based REVs(Figure 2) show increasing efficiency of gene expressionas the �LCR employed becomes more complete. Wetherefore decided to test the efficacy of the minilocus�LCR26 within a REV context. The principal features ofthis 38-kb construct are a 21-kb genomic region spanningthe complete �LCR region linked to a 4.98-kb �-globingene fragment with approximately 1.5-kb 5� and 1.8-kb3� flanking sequences. This �-globin minilocus was incor-porated into the cos203 REV cosmid vector (cos203�; seeMaterials and methods) and analysed in both thefoetal/embryonic and adult stage erythroid environ-ments of K562 and murine erythroleukaemia (MEL) cells,respectively. Analysis of total genomic DNA from twoindependently generated pools of stable transfected cells
Gene Therapy
Figure 4 The 3�, but not the 432�, plasmid REV becomes less sensitiveto DNaseI digestion in K562 cells over a prolonged (30-day) period ofculture. Nuclei were harvested from pools of K562 cells stably transfectedwith either the 432� or 3� plasmid REVs (Figures 1 and 2), before (day 0)and after 30-days of continuous culture (day 30). Aliquots of the nuclearsuspension were subjected to digestion with increasing, but limitingamounts of DNaseI (indicated by the sloping triangle from left to rightabove the lanes). Total DNA from these nuclei was digested with XbaI,Southern blotted and probed with a 920 bp BamHI–EcoRI fragment span-ning the second intron of the human �-globin gene. This gives a fragmentof 11 kb from the endogenous (Endo) �-globin locus and a variable sizedproduct corresponding to the episomal (Epi) �-globin transgene. Southernblot hybridisation signals were quantified by PhosphorImager. (a) DNaseIsensitivity of 432�. The episomal (Epi) �-globin transgene fragment is4.5 kb in length. Lanes 0, no DNaseI control. (b) DNaseI sensitivity of3�. The episomal (Epi) �-globin transgene is a 6-kb fragment. Lanes 0,no DNaseI control. (c) Relative DNaseI sensitivity of 432b REVs. TheEndo/Epi signal ratios from panel (a) were calculated to give the relativeDNaseI sensitivity after comparison with the no-DNaseI control (lanes 0;see text and Materials and methods). This reflects the trend of DNaseIsensitivity over the digestion series at day 0 (�) and day 30 (�). (d)Relative DNaseI sensitivity of 3� REVs. The Endo/Epi signal ratios frompanel (b) were calculated as in (c) above. Day 0, �; day 30, �. Noteincreased resistance to DNaseI digestion of the day 30 sample.
of both types by digestion with SalI followed by pulsed-field gel electrophoresis and Southern blot hybridisation,showed the presence of the 38-kb �-globin minilocus con-struct, as well as the cos203 cosmid vector backbone inboth K562 (Figure 5a, lanes 1 and 2) and MEL (Figure 5a,lanes 3 and 4) cells confirming the presence of episomes3–4 weeks after transfection and drug selection. FurtherSouthern blot analysis of genomic DNA digested withEcoRI and DpnI, showed that replication of episomes hadoccurred and were present at an average copy numberof 1–2 per cell in K562 (Figure 5b, lanes 1 and 2) and onecopy per cell in MEL (Figure 5b, lanes 3 and 4) pools. Theability of the cos203� cosmid vector to establish episomalstatus in MEL cells is in marked contrast to the p220.2
Episomal vectors and locus control regionsC-M Chow et al
332
Gene Therapy
plasmid-based constructs (Figure 1), which were unableto do so (data not shown).
Episomal transgene expression from cos203� asassessed by S1-nuclease protection (see Figure 2), showedthat in K562 cells (Figure 6a, lanes 1 and 2) expressionper episomal copy was 25–30% of endogenous �-globinlevels (Figure 6b, K562). Expression in MEL-transfected
Figure 5 Episomal replication of cos203� cosmid REV within bothembryonic/foetal human K562 and murine adult MEL erythroid cells.Pools of embryonic/foetal K562 and adult MEL erythroid cells stablytransfected with the cos203� cosmid REV analysed for episomal integrityand transgene copy number. (a) Total genomic DNA from two indepen-dently generated, cos203� transfected pools of K562 (lanes 1 and 2) andMEL (lanes 3 and 4) cells was digested with SalI, resolved by pulsed-fieldgel electrophoresis and Southern blotted. The blots were then doubleprobed with a 3.6-kb fragment encompassing the human �-globin geneand the cos203 vector backbone. Lane Ck, untransfected K562 cells; laneCm, untransfected MEL cells as negative controls. The bands correspond-ing to the 38 kb �-globin minilocus and the 9 kb cos203 vector are indi-cated by arrowheads on the right. M(kb), positions of size markers in kb.(b) Total DNA from the same pools of K562 and MEL cells as in panel(a) above analysed for cos203� copy number by digestion with EcoRI andDpnI and Southern blot hybridisation. The samples from K562 cells werehybridised with the BamHI–EcoRI �-globin intron II probe, giving a 5.5kb endogenous band (h�) and a 4.3 kb cos203� derived band (cos203�).MEL cell samples were double-probed for the endogenous murine vavgene, which detects a 6.5-kb fragment (mvav), as well as with �-globinintron II. Human �-globin gene-containing plasmid DNA was used as apositive control (lane C, pl). M(kb), positions of size markers in kb.
Figure 6 The cos203� REV gives expression in both embryonic/foetalK562 and adult MEL erythroid cell types and is efficiently retained inK562 cells without drug selective pressure. Total RNA from two indepen-dently generated pools of K562 and MEL cells stably transfected withcos203� (Figure 5), were analysed for human �-globin transgeneexpression by an S1-nuclease protection assay. Results were quantified byPhosphorImager. (a) Lanes 1 and 2: K562 cells simultaneously probed for�-globin transgene (5��) and endogenous �-globin gene (5��) expression(see Figure 2). Lane Ck, untransfected K562 cells as a negative control.Lanes 3 and 4, MEL cell-derived samples before (�) and after (+) inducederythroid differentiation double-probed for �-globin transgene (5��) andendogenous murine �maj-globin (�maj) mRNA. Lane Cm, untransfectedMEL cells as a negative control. M, positions of size markers consistingof a HinfI digest of pBR322. (b) K562. Human �-globin expression fromcos203� in K562 cells (panel a, lanes 1 and 2) as a percentage of theendogenous �-globin on an episome copy number basis. (c) MEL. As inpanel (b) except that human �-globin mRNA levels per cos203� episomeare expressed as a percentage of endogenous �maj-globin transcripts inMEL cells after 4 days of induced erythroid differentiation (panel a, lanes3+ to 4+). (d) Long-term retention and expression of cos203� in K562cells. A representative pool of K562 cells (panel A, lane 1) was culturedin the presence (+Hygromycin) and absence (�Hygromycin) of drug selec-tive pressure for 60 days. Total DNA and RNA were prepared at 10-dayintervals and analysed for cos203� episomal integrity (data not shown)and human �-globin transgene expression (as in panel a). Human �-glo-bin transgene mRNA levels are expressed as a percentage of endogenous�-globin per episome copy number.
pools (Figure 6a, lanes 3 and 4) was similar to that seenin K562 cells at between 20 and 38% of the endogenous�maj-globin per episome copy (Figure 6c, MEL). Theseresults show that cos203� gives comparable expressionin both an embryonic/foetal (K562) and adult (MEL) cellenvironment at a level that would be of therapeutic valueif reproduced in vivo.

Episomal vectors and locus control regionsC-M Chow et al
333We also assessed the retention and stability ofexpression of the cos203� construct by culturing a rep-resentative pool of stably transfected K562 (Figure 5, lane1) and MEL (Figure 5, lane 3) cells in the presence andabsence of drug selective pressure for a period of 60 dayscorresponding to at least 60 generations. Similar pulsed-field gel and Southern blot analysis to that shown in Fig-ure 5 showed that cos203� episomes were retained atbetween two and three copies in K562, but not in MELcells without any detectable transgene integration events(data not shown). RNA analysis from this pool of K562cells showed that even in the absence of drug-selectivepressure, expression of cos203� as a percentage of theendogenous �-globin on a episome copy basis, was main-tained at a similar level (18–26%) over the 60-day periodof culture to the day 0 starting value (22%; Figure 6d).
DiscussionREVs based on viral origins of replication are showingincreasing promise for use in gene therapy.1,3–5,27 Inaddition to improving the efficiency of REV delivery,advances in two areas need to be made in order for theutility of this system to be realised: episome maintenanceand regulated gene expression. The majority of studiesaddressing gene expression from within REVs, havefocused on the use of ubiquitously acting classical pro-moter–enhancer combinations, such as those from RSV28
and CMV.8 The question of how to obtain efficient tissue-specific expression has not been thoroughly addressed.Inducible systems including the heavy metal responsivehuman metallothionein IIA promoter,28 the ionophore-stimulated rat GRP78 promoters28 and the incorporationof the dioxin response element7 have also been testedwith varying degrees of success. In this report we addressthe problem of obtaining strict tissue-specific, long-termgene expression from within REVs. We demonstrate thatthe DNaseI HS sites that constitute the �-globin locuscontrol region (�LCR) are able to confer cell-type specificgene expression from within REVs. HS sites 2, 3 and 4act synergistically to give an approximately 95-foldincrease in expression compared with the human �-glo-bin gene alone and the level of which corresponded togreater than 13% of endogenous �-globin mRNA per epi-some copy (Figure 2b). In contrast these elements areinactive in promoting enhanced transcription in non-erythroid HeLa cells (Figure 2c). Generally, the perform-ance of individual and combinations of �LCR HS sites inREVs reflects that seen in stably transfected tissue culturecells,29–32 rather than transient transfection assays.33–35
This difference in the activity of �LCR elements whenassayed by transient transfection or within REVs, may bedue to the fact that REVs adopt a well-ordered, nucleoso-mal configuration akin to that of native chromatin.14,15
Since most (HS1, 3 and 4) �LCR elements enhance tran-scription only when integrated within the host cell gen-ome, the chromatin configuration adopted by REVswould appear to provide the necessary environment fortheir function. In support of this idea is the report thatHS2 can remodel chromatin on REVs36 in K562 cells,although this study addressed neither tissue-specificity,the cumulative effect of combining �LCR HS sites nor themaintenance of the open remodelled chromatin stateupon prolonged periods of culture. Importantly in thisregard, our own findings show that a �LCR HS 234 site
Gene Therapy
combination, but not HS 3 alone, retain a DNaseI-sensi-tive structure (Figure 4) and gene expression (Figure 3)over prolonged periods of time, suggesting that multipleLCR elements are required to maintain a transcriptionallycompetent open chromatin configuration.
Our data also show that the constructs containingsmall variants of �LCR elements do not contribute to themaintenance of episomes as they are lost from cells cul-tured in the absence of drug-selective pressure (Figure3a; data not shown). EBV-based vectors disappear fromdividing cells over a period of approximately 2 monthswithout selection.37,38 Our results showing the episomalloss in K562 cells within 1 to 2 months are, therefore, con-sistent with previous studies. However, in marked con-trast to the small plasmid-based constructs (Figure 1a),the cos203� 38 kb minilocus-REV cosmid vector, wasefficiently retained in K562 cells over 60 days of culturein the absence of drug-selective pressure (Figure 6d; datanot shown). Other vector systems with improved persist-ence inside cells have been reported. An SV40-based epi-somal vector carrying a scaffold/matrix attachmentregion (S/MAR) from the human interferon �-gene wasshown to be able to replicate and maintain in CHO cellsover more than 100 generations without selective press-ure.39 The mechanism by which the cos203� cosmid con-struct is retained is currently unknown. However, ourfindings are consistent with previous observations thatfound plasmid EBV-based vectors containing randomlyselected large (�16 kb) human genomic DNA fragments,conferred efficient episomal replication in mammaliancells.11,40 This may involve the presence of eitheradditional genomic origins of replication,11,12,40 orS/MAR elements39 or both.
While the retention of episomes within cells is clearlyimportant, our work indicates that the maintenance oftissue-specific transgene expression from within REVs isalso a crucial consideration. Expression of the �-globingene from REVs with either �LCR HS2 or HS3 alone sig-nificantly declines over time in the presence or absenceof drug selective pressure (Figure 3a). The silencing of �-globin gene expression in the presence of drug selectionis intriguing as it implies that regions of the same epi-some can be repressed while others (the hygromycinresistance gene in this case), can remain active.Importantly, our findings imply that as in the case of inte-grated transgenes,41 expression from within REVs isprone to silencing but that the inclusion of the �LCR pre-vented this repression of gene function. Therefore,inclusion of elements to counteract the cell’s selectivegene silencing mechanism will be necessary for the useof episomal constructs in gene therapy. Our data showthat combinations of LCR elements can efficiently fulfilthis requirement. In this study, we have shown that thecos203� minilocus cosmid vector is not only efficientlyretained in stably transfected tissue culture cells, but thata significant level of �-globin expression (18–26% of �-globin per episome copy), was maintained throughoutthe culture period (Figure 6d). Although MEL and K562cells have been used extensively as model systems tostudy globin gene expression and gene therapy,20 futuredevelopments in technology that will allow the transferof the large cos203� cosmid construct into haematopo-ietic stem cells are required to extend our observationsin a more clinically relevant context. Nevertheless, ourresults indicate for the first time the feasibility of using

Episomal vectors and locus control regionsC-M Chow et al
334
Gene Therapy
REVs for gene therapy of �-thalassaemia and sickle celldisease since levels of �-globin gene expression thatwould be therapeutic in such cases can be obtained withas few as 1–2 copies of cos203�.
Materials and methods
Cells and cell cultureK562,22 MEL42 and HeLa cells were maintained in DMEM(Life Technologies, Paisley, UK) containing 10% foetalcalf serum, 2 mM L-glutamine, 50 �g/ml penicillin and50 units/ml streptomycin. Cells were cultured at 37°C in5% CO2. Transfection of plasmid p220.2-based constructsin K562 and HeLa cells was performed by electroporationusing a BioRad Gene Pulser (BioRad, Hertfordshire, UK)at a setting of 0.25 kV and 960 �F, with 20–50 �g of super-coiled DNA. Transfection of the cos203� cosmid �-globinminilocus construct into K562 and MEL cells was by lipo-fection using Lipofectin (Life Technologies) or FuGene6(Roche, Lewes, UK) according to the manufacturers’instructions. Stably transfected cells were selected in thepresence of 250 �g/ml hygromycin B. K562 cells har-bouring stably integrated transgenes were generated bylinearising gene constructs at the unique PvuI site (withinthe ampicillin resistance gene on the vector) before elec-troporation. In general, stable pools were analysed 3–4weeks after transfection, except for long-term cultureexperiments when approximately 1 × 107 cells were div-ided into media with or without hygromycin and grownfor up to a further 60 days.
Replicating episomal constructsA human �-globin gene extending from a 5� HpaI site at�815 bp to an EcoRV site 1685 bp downstream of thepoly(A)-addition site in the plasmid GSE175830 wasremoved as a 4.1-kb EcoRV fragment and inserted into ablunted SalI site in the polylinker of p220.2 (Figure 1a), anEpstein–Barr virus-based vector containing oriP, EBNA-1and hygromycin resistance genes.6 This cloning stepbrings a number of extra restriction sites (including aunique SalI site) 5� to the �-globin gene (Figure 1a). Theelements of the �LCR were isolated as a 1.5-kb KpnI–BglIIHS2 fragment, a 1.9-kb HindIII HS3 fragment and a 2.1-kb BamHI–XbaI HS4 fragment.29 These elements eitheralone or in combination were inserted by blunt-end lig-ation into the unique SalI site 5� of the �-globin gene(Figure 1a). Constructs which contain more than one HSsite were joined, such that their order reflected that foundin the native �-globin locus on chromosome 11 (Figure2a).
The human �LCR minilocus construct26 was cloned asa 38-kb SalI fragment into the SalI site of the cos203 EBV-based vector which is as p220.2 (Figure 1a), but includesa cos site for cosmid cloning. This construct was desig-nated as cos203�.
Isolation of low molecular weight DNA fractionsLow molecular weight DNA was prepared by themethod of Hirt.24 Briefly, cells were washed with PBS andsubsequently incubated with lysis buffer (0.6% SDS, 10mM EDTA) for 15 min at room temperature. NaCl wasthen added to a concentration of 1.4 M. After precipi-tation overnight at 4°C, genomic DNA and cell debriswere pelleted by centrifugation at 10 000 r.p.m. for 10 min
in a Beckman J21 high-speed centrifuge. The supernatantwas extracted with an equal volume of phenol:chloro-form (1:1 v/v) and the DNA precipitated by adding 2.5volumes of ethanol and storage at �20°C. The precipi-tated DNA was collected by centrifugation and resus-pended in TE (10 mM Tris-HCl, 1 mM EDTA, pH 7.5)buffer.
Analysis of extrachromosomally replicating episomesThe presence of the REV plasmid constructs (Figures 1and 2) in stably transfected K562 and HeLa cells wasassessed as follows. Total DNA (10 �g) was prepared asdescribed,42 digested with BamHI and HindIII andresolved by electrophoresis on a 0.7% agarose gel fol-lowed by Southern transfer on to Hybond N+(Amersham-Pharmacia, Buckingamshire, UK). Blots werethen hybridised with 32P-labeled pBluescript (Stratagene,Amsterdam, The Netherlands), which detects the ampic-illin resistance gene on the p220.2 vector backbone. Gelelectrophoresis, blotting and hybridisation were carriedout using standard procedures.43 In order to assess theintegrity of cos203� cosmid episomes in K562 and MELcells, total genomic DNA (15 �g) was digested with SalI,resolved by pulsed field gel electrophoresis employing aBioRad CHEF DRII system (BioRad) and Southern blot-ted. These blots were then double probed with a 3.6 kbsub-clone of the human �-globin gene extending from�265 to +3400 from the transcriptional start site and theempty cos203 vector backbone. All hybridisation probeswere labelled by random priming using a commerciallyavailable kit (Amersham-Pharmacia).
Southern blot analysis for episome copy numberdeterminationPlasmid, p220.2-based episomal copy number wasdetermined in total genomic DNA (10 �g) isolated fromK562 and HeLa cells digested with EcoRI and DpnI,resolved by electrophoresis on a 0.7% agarose gels,Southern blotted and probed with a 920-bp BamHI–EcoRIfragment spanning the second intron of the human �-globin gene. This probe detects both the endogenous �-globin genes as a 5.5-kb fragment and a band of varyingsize from the different transfected plasmid constructs.The fragment sizes for each construct (see Figure 2a) are:6.45 kb (�); 7.95 kb (2�); 8.35 kb (3�); 8.55 kb (4�); 11.9kb (432�); 9.85 kb (32�); 10.45 kb (43�) and 10.05 kb (42�).
Total DNA (10 �g) from K562 and MEL cells trans-fected with the cos203� cosmid construct, was digestedwith EcoRI and DpnI and subjected to Southern blotanalysis as described above. The samples from K562 cellswere hybridised with the BamHI–EcoRI �-globin intron IIprobe alone giving a 4.3-kb cos203� derived band. Thosefrom MEL cells were double probed for the endogenousmurine vav, which acted as an internal 2 copy genereference standard.44
The intensities of bands corresponding to episomal andthe endogenous genes were quantified by Phosphor-Imager and their ratio used to determine episome copynumber taking into consideration that K562 cells are tri-somy for chromosome 1122 and therefore possess threecopies of the �-globin locus.
Quantification of human �-globin mRNATotal RNA was extracted from K562, MEL and HeLa cellsby selective precipitation in the presence of 3 M LiCl and

Episomal vectors and locus control regionsC-M Chow et al
3356 M urea.42,45 Analysis of human �-globin transgeneexpression in these samples was by an S1-nuclease pro-tection assay using 10 �g total RNA and end-labelled,double-stranded DNA probes.25 The 5� human �-globinprobe was a 525-bp AccI cDNA fragment that gives anS1-nuclease protected product of 155 nucleotides.46 The5� human �-globin probe was an 1819-p HindIII–BamHIgenomic fragment giving rise to a 205 nucleotide S1-pro-tected product from exon II. The 5� murine �maj-globinprobe was a 700 bp HindIII–NcoI genomic fragment,which gives a 96 nucleotide S1-protected product.46 Theprobe for human hnRNPA2 was prepared as follows: aPCR fragment starting from 330 bp upstream of exon 9to the 3� end of this exon47 was cloned into the EcoRVsite of pBluescript. The probe was prepared by PCRamplification using primers corresponding to a position545 within pBluescript (5�-TGC TGC AAG GCG ATTAA-3�) and the 3� end of hnRNPA2 exon 9 (5�-CTC CTCCAT AGT TGT CAT AAC C-3�). This generated a 627-bp probe that results in a 122 nucleotide S1-nuclease pro-tected product. Products were resolved after S1-nucleasedigestion on a 6% polyacrylamide gel in the presence of8 M urea, dried, exposed to Kodak XAR 5 X-ray film andquantified by PhosphorImager (Molecular Dynamics/Amersham-Pharmacia) analysis.
DNA fluorescence in situ hybridisation (FISH)DNA FISH was carried out as described48 with modifi-cations. K562 cells were washed in PBS, resuspended inhypotonic buffer (10 mM HEPES pH7.5, 30 mM glycerol,1 mM CaCl2, 0.8 mM MgCl2) for 10 min. Cells were thenwashed in methanol/acetic acid (3:1) between three andfour times over 30 min. Slides with spread cells were air-dried, dehydrated through successive washes in 70%,90% and 100% ethanol, before treatment with 100 �g/mlof RNase A at 37°C for 15 min. The slides were thenrinsed in water, dehydrated by successive ethanol washesas before, air-dried and denatured in 70% formamide, 2 ×SSC for 2 min at 70°C. The slides were then immediatelyimmersed in ice-cold 70%, 90% and 100% ethanol (3 mineach step) and air-dried. The probe used for hybridis-ation was cos203� (see above), which simultaneouslydetects the endogenous �-globin loci and the episomalconstructs. DNA (2 �g) was labelled with digoxygeninby nick-translation in a 25-�l reaction consisting of 0.5 MTris-HCl pH 7.5, 0.1 M MgSO4, 1 mM dithiothreitol, 500�g/ml bovine serum albumin, 0.5 mM dNTPs (dATP,dCTP, dGTP), 0.7 nM digoxygenin-11-dUTP, 5 unitsDNA polymerase (Roche) and 0.004 units DNaseI (Sigma,Poole, UK) for 40 min at 15°C. Digoxygenin-labelledprobes were dissolved in hybridisation mix (50% forma-mide, 2 × SSC, 200 ng/�l of salmon sperm DNA, 5 ×Denhardt’s solution, 1 mM EDTA, 50 mM sodium phos-phate pH 7.0). Probe (20–50 ng) was denatured at 75°Cfor 5 min in 40 �l of hybridisation mix and allowed toanneal at 37°C with 20-fold excess of human Cot-1 DNAfor 30 min before applying to the dried and denaturedslides. Hybridisation was performed overnight. Slidewashes and probe detection were carried out asdescribed.48
General DNaseI sensitivity analysisNuclei were prepared from a minimum of 1 × 108 K562cells as described previously49 using 20 strokes of a B-type Dounce homogenizer. The nuclei were resuspended
Gene Therapy
in 1 ml of 1 × reticulocyte standard buffer (10 mM Tris-HCl pH 7.5, 1 mM NaCl, 3 mM MgCl2) and CaCl2 wasadded to a final concentration of 1 mM. Aliquots of 100�l were digested with increasing amounts (0–0.26 �g)DNaseI (Sigma) for 2 min at 37°C. The DNaseI digestionwas stopped by addition of a stop solution (0.3 M NaCl,10 mM Tris-HCl, pH 8, 5 mM EDTA, 0.5% SDS finalconcentrations), made 0.5 �g/�l proteinase K and incu-bated overnight at 37°C. Genomic DNA was thenextracted using a standard phenol:chloroform and etha-nol precipitation procedure. The DNA was resuspendedin 100 �l of water, 20 �l aliquots digested with XbaIrestriction enzyme and subjected to Southern blot analy-sis using a 920-bp BamHI–EcoRI fragment spanning thesecond intron of the human �-globin gene as a hybridis-ation probe. This gives internal fragments extending overthe human �-globin gene and part of the �LCR elementsof 4.5 kb and 6 kb from the 432� and 3� episomal plas-mid constructs, respectively. In addition, an 11 kb bandcorresponding to the endogenous �-globin gene is alsoobtained, which acts as a reference. The intensity of thehybridised bands was quantified by PhosphorImager.DNaseI digests were performed with nuclei from at leasttwo, independently generated transfected pools of K562cells for each construct analysed. Since there may alsohave occurred a slight change in REV copy numberwithin a given pool of cells before and after 30 days ofculture, the ratio of the endogenous to episomal �-globinbands at each concentration of DNaseI was further com-pared with that of the no-DNaseI control. This new ratioobtained by dividing the value of the undigested controlby each subsequent point within the same DNaseI diges-tion series, reflects the trend of DNaseI sensitivity forthat sample of nuclei allowing comparisons betweendifferent cultures.
AcknowledgementsWe thank Bill Sugden for the p220.2 and cos203 EBV-based vectors. This work was initiated with support fromCobra Therapeutics (Keele, UK) and continued withfunding under the European Union Framework IVBiomed-2 (contract No. BMH4-CT96-1279) and Biotech-nology (contract No. BIO4-CT96-0414) programmes. AAwas funded for this work by a European Union MarieCurie Fellowship (contract No. HPMF-CT-1994-00091).
References1 Sclimenti CR, Calos MP. Epstein–Barr virus vectors for gene
expression and transfer. Curr Opin Biotechnol 1998; 9: 476–479.2 Sun TQ, Fernstermacher DA, Vos JM. Human artificial episomal
chromosomes for cloning large DNA fragments in human cells.Nat Genet 1994; 8: 33–41.
3 Satoh E et al. Efficient gene transduction by Epstein–Barr virus-based vectors coupled with cationic liposome and HVJ-liposom.Biochem Biophys Res Commun 1997; 238: 795–799.
4 Saeki Y, Wataya-Kaneda M, Tanaka K, Kaneda Y. Sustainedtransgene expression in vitro and in vivo using an Epstein–Barrvirus replicon vector system combined with HVJ liposomes.Gene Therapy 1998; 5: 1031–1037.
5 Tsukamoto H et al. Enhanced expression of recombinant dystro-phin following intramuscular injection of Epstein–Barr virus(EBV)-based mini-chromosome vectors in mdx mice. Gene Ther-apy 1999; 6: 1331–1335.
6 Yates JL, Warren N, Sugden B. Stable replication of plasmids

Episomal vectors and locus control regionsC-M Chow et al
336
Gene Therapy
derived from Epstein–Barr virus in various mammalian cells.Nature 1985; 313: 812–815.
7 De Benedetti A, Rhoads RE. A novel BK virus-based episomalvector for expression of foreign genes in mammalian cells.Nucleic Acids Res 1991; 19: 1925–1931.
8 Sabbioni S et al. A BK virus episomal vector for constitutive highexpression of exogenous cDNAs in human cells. Arch Virol 1995;140: 335–339.
9 Ohe Y, Zhao D, Saijo N, Podack ER. Construction of a novelbovine papillomavirus vector without detectable transformingactivity suitable for gene transfer. Hum Gene Ther 1995; 6:325–333.
10 Cooper MJ et al. Safety-modified episomal vectors for humangene therapy. Proc Natl Acad Sci USA 1997; 94: 6450–6455.
11 Krysan PJ, Haase SB, Calos MP. Isolation of human sequencesthat replicate autonomously in human cells. Mol Cell Biol 1989;9: 1026–1033.
12 Wohlgemuth JG et al. Long-term expression from autonomouslyreplicating vectors in mammalian cells. Gene Therapy 1996; 3:503–512.
13 Reeves R, Gorman CM, Howard B. Minichromosome assemblyof non-integrated plasmid DNA transfected into mammaliancells. Nucleic Acids Res 1985; 13: 3599–3615.
14 Archer TK, Lefebvre P, Wolford RG, Hager GL. Transcriptionfactor loading on the MMTV promoter: a bimodal mechanismfor promoter activation. Science 1992; 255: 1573–1576.
15 Cereghini S, Yaniv M. Assembly of transfected DNA intochromatin: structural changes in the origin-promoter-enhancerregion upon replication. EMBO J 1984; 3: 1243–1253.
16 Smith CL, Archer TK, Hamlin-Green G, Hager GL. Newlyexpressed progesterone receptor cannot activate stable, repli-cated mouse mammary tumor virus templates but acquirestransactivation potential upon continuous expression. Proc NatlAcad Sci USA 1993; 90: 11202–11206.
17 Fraser P, Grosveld F. Locus control regions, chromatin acti-vation and transcription. Curr Opin Cell Biol 1998; 10: 361–365.
18 Li Q, Harju S, Peterson KR. Locus control regions: coming ofage at a decade plus. Trends Genet 1999; 15: 403–408.
19 Kioussis D, Festenstein R. Locus control regions: overcomingheterochromatin-induced gene inactivation in mammals. CurrOpin Genet Dev 1997; 7: 614–619.
20 Antoniou M, Grosveld F. Genetic approaches to therapy for thehaemoglobinopathies. In: Fairbairn LJ, Testa NG (eds). Blood CellBiochemistry, Volume 8: Hematopoiesis and Gene Therapy. KluwerAcademic/Plenum: New York, 1999, pp 219–242.
21 May C et al. Therapeutic haemoglobin synthesis in �-thalassa-emic mice expressing lentivirus-encoded human �-globin. Nat-ure 2000; 406: 82–86.
22 Gahmberg CG, Andersson LC. K562-a human leukemia cell linewith erythroid features. Semin Hematol 1981; 18: 72–77.
23 BlomvanAssendelft G, Hanscombe O, Grosveld F, Greaves DR.The �-globin dominant control region activates homologous andheterologous promoters in a tissue-specific manner. Cell 1989;56: 969–977.
24 Hirt B. Selective extraction of polyma DNA from infected mousecell culture. J Mol Biol 1967; 26: 365–369.
25 Antoniou M, deBoer E, Grosveld F. Fine mapping of genes: thecharacterization of the transcriptional unit. In: Davies KE (ed.).Human Genetic Disease Analysis. A Practical Approach, 2nd edn.IRL Press: Oxford, 1993, pp 83–108.
26 Grosveld F, BlomvanAssendelft GB, Greaves DR, Kollias G. Pos-ition-independent high level expression of the human �-globingene in transgenic mice. Cell 1987; 51: 975–985.
27 WadeMartins R et al. Stable correction of a genetic deficiency inhuman cells by an episome carrying a 115 kb genomic trans-gene. Nature Biotechnol 2000; 18: 1311–1314.
28 Hauer CA, Getty RR, Tykocinski ML. Epstein–Barr virus epi-
some-based promoter function in human myeloid cells. NucleicAcids Res 1989; 17: 1989–2003.
29 Collis P, Antoniou M, Grosveld F. Definition of the minimalrequirements within the human �-globin gene and the dominantcontrol region for high level expression. EMBO J 1990; 9: 233–240.
30 Talbot D, Philipsen S, Fraser P, Grosveld F. Detailed analysis ofthe site 3 region of the human �-globin dominant control region.EMBO J 1990; 9: 2169–2178.
31 Philipsen S, Talbot D, Fraser P, Grosveld F. The �-globin domi-nant control region: hypersensitive site 2. EMBO J 1990; 9:2159–2167.
32 Pruzina S et al. Hypersensitive site 4 of the human �-globinlocus control region. Nucleic Acids Res 1991; 19: 1413–1419.
33 Tuan DY, Solomon WB, London IM, Lee DP. An erythroid-spe-cific, developmental stage-independent enhancer far upstreamof the human ‘�-like globin’ genes. Proc Natl Acad Sci USA 1989;86: 2554–2558.
34 Ney PA, Sorrentino BP, McDonagh KT, Nienhuis AW. TandemAP-1-binding sites within the human �-globin dominant controlregion function as an inducible enhancer in erythroid cells.Genes Dev 1990; 4: 993–1006.
35 Ney PA, Sorrentino BP, Lowrey CH, Nienhuis AW. Inducibilityof the HS II enhancer depends on binding of an erythroid spe-cific nuclear protein. Nucleic Acids Res 1990; 18: 6011–6017.
36 Gong Q, McDowell JC, Dean A. Essential role of NF-E2 inremodelling of chromatin structure and transcriptional acti-vation of the �-globin gene in vivo by 5�hypersensitive site 2 ofthe �-globin locus control region. Mol Cell Biol 1996; 16: 6055–6064.
37 Lei DC et al. Episomal expression of wild-type CFTR correctscAMP-dependent chloride transport in respiratory epithelialcells. Gene Therapy 1996; 3: 427–436.
38 Banerjee S, Livanos E, Vos JMH. Therapeutic gene delivery inhuman B-lymphoblastoid cells by engineered non-transforminginfectious Epstein–Barr virus. Nature Med 1995; 1: 1303–1308.
39 Piechaczek C et al. A vector based on the SV40 origin of repli-cation and chromosomal S/MARs replicates episomally in CHOcells. Nucleic Acids Res 1999; 27: 426–428.
40 Heinzel SS, Krysan PJ, Tran CT, Calos MP. Autonomous DNAreplication in human cells is affected by the size and the sourceof the DNA. Mol Cell Biol 1991; 11: 2263–2272.
41 Verma IM, Somia N. Gene therapy – promises, problems andprospects. Nature 1997; 389: 239–242.
42 Antoniou M. Induction of erythroid-specific expression inmurine erythroleukemia (MEL) cell lines. In: Murray EJ (ed.).Methods in Molecular Biology: vol 7 Gene Transfer and ExpressionProtocols. Humana Press: New York, 1991, pp 421–434.
43 Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Labora-tory Manual 2nd edn. Cold Spring Harbor Laboratory Press:Cold Spring Harbor, NY, 1989.
44 Raguz S et al. Muscle-specific locus control region activity asso-ciated with the human desmin gene. Dev Biol 1998; 201: 26–42.
45 Auffray C, Rougeon F. Purification of mouse immunoglobulinheavy-chain RNAs from total myeloma tumor RNA. Eur JBiochem 1980; 107: 303–324.
46 Antoniou M, DeBoer E, Habets G, Grosveld F. The human �-globin gene contains multiple regulatory regions: identificationof one promoter and two downstream enhancers. EMBO J 1988;7: 377–384.
47 Kozu T, Henrich B, Schafer KP. Structure and expression of thegene (HNRPA2B1) encoding the human hnRNP protein A2/B1.Genomics 1995; 25: 365–371.
48 Ashe HL et al. Intergenic transcription and transinduction of thehuman �-globin locus. Genes Dev 1997; 11: 2494–2509.
49 Reitman M, Lee E, Westphal H, Felsenfeld G. Anenhancer/locus control region is not sufficient to open chroma-tin. Mol Cell Biol 1993; 13: 3990–3998.





























