Embed Size (px)
Citation preview

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
Long-term data for endemic frog genera revealpotential conservation crisis in the Bale Mountains,Ethiopia
DAV I D J . G OW E R , R O M A N K . A B E R R A , S I L V I A S C HWA L L E R , M A L C O L M J . L A R G E N
B E N C O L L E N , S T E P H E N S P AW L S , M I C H E L E M E N E G O N , B R E D A M . Z I M K U S
R A F A E L D E S Á , A B E B E A . M E N G I S T U , F I K I R T E G E B R E S E N B E T , R O B I N D . MO O R E
S A M Y A . S A B E R and S I M O N P . L O A D E R
Abstract Populations of many frogs have declined alar-mingly in recent years, placing nearly one third of the. 6,000 species under threat of extinction. Declines havebeen attributed largely to habitat loss, environmentaldegradation and/or infectious diseases such as chytridio-mycosis. Many frogs undergo dramatic natural populationfluctuations such that long-term data are required todetermine population trends without undue influenceof stochastic factors. We present long-term quantitativedata (individuals encountered per person hour of search-ing) for four monotypic frog genera endemic to anAfromontane region of exceptional importance but growingconservation concern: one endemic to the Ethiopianhighlands (Spinophrynoides osgoodi) and three endemic tothe Bale Mountains (Altiphrynoides malcolmi, Balebrevicepshillmani, Ericabatrachus baleensis), collected during 15 fieldtrips to the Bale Mountains between 1971 and 2009. Only asingle confirmed sighting of S. osgoodi has been made since
1995. The other three species have also declined, at leastlocally. E. baleensis appears to have been extirpated at itstype locality and at the same site B. hillmani has declined.These declines are in association with substantial habitatdegradation caused by a growing human population.Chytrid fungus has been found on several frog speciesin Bale, although no dead or moribund frogs have beenencountered. These results expose an urgent need for moreamphibian surveys in the Bale Mountains. Additionally, weargue that detrimental human exploitation must be haltedimmediately in at least some parts of the Harenna Forest if aconservation crisis is to be averted.
Keywords Africa, Afromontane, amphibians, BaleMountains National Park, chytrid, decline, Ethiopia,management
This paper contains supplementary material that can befound online at http://journals.cambridge.org
Introduction
Populations of many frogs have declined alarmingly inrecent years, placing nearly one third of the . 6,000
species under threat of extinction (IUCN, 2010). Declineshave been attributed largely to habitat loss, environmentaldegradation and/or infectious diseases such as chytridio-mycosis (Stuart et al., 2008; Kilpatrick et al., 2010). Manyfrogs undergo dramatic natural population fluctuations (e.g.Pechmann et al., 1991; Semlitsch et al., 1996) such that long-term data are required to determine population trendswithout undue influence of stochastic factors (e.g. Meyeret al., 1998; Marsh, 2001; Green, 2003). Long-term quan-titative data are generally lacking for the vast majority offrog species, particularly in the tropics (Adeba et al., 2010;Largen & Spawls, 2010). For Africa this is exacerbated by ageneral relative scarcity of attention to conservation biologyresearch despite many threats to its globally important biota(Pimm, 2007). Here we present long-term quantitative datafor four frog genera endemic to an Afromontane region ofexceptional importance but growing conservation concern.
DAVID J. GOWER (Corresponding author) Department of Zoology, The NaturalHistory Museum, London SW7 5BD, UK. E-mail [email protected]
ROMAN K. ABERRA Ethiopian Wildlife Conservation Authority, Addis Ababa,Ethiopia
SILVIA SCHWALLER, ABEBE A. MENGISTU* and SIMON P. LOADER University ofBasel, Institute of Biogeography, Department of Environmental Sciences, Basel,Switzerland
MALCOLM J. LARGEN Formby, UK
BEN COLLEN Institute of Zoology, Zoological Society of London, London, UK
STEPHEN SPAWLS Spixworth, Norwich, UK
MICHELE MENEGON Sezione di Zoologia dei Vertebrati, Museo Tridentino diScienze Naturali, Trento, Italy
BREDA M. ZIMKUS Museum of Comparative Zoology, Harvard University,Cambridge, Massachusetts, USA
RAFAEL DE SÁ Department of Biology, University of Richmond, Richmond,Virginia, USA
FIKIRTE GEBRESENBET and SAMY A. SABER† Department of Biology, Addis AbabaUniversity, Addis Ababa, Ethiopia
ROBIN D. MOORE Conservation International, Arlington, Virginia, USA
*Also at: Department of Biology, Addis Ababa University, Addis Ababa,Ethiopia†Also at: Zoology Department, Faculty of Science, Al-Azhar University, Assiut,Egypt
Received 1 April 2011. Revision requested 16 May 2011.Accepted 26 July 2011.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69 doi:10.1017/S0030605311001426

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
The Bale Mountains lie in southern Ethiopia, east of theRift Valley (Fig. 1). The Mountains are a high (up to nearly4,400 m) and extensive massif with rich biodiversity,including many unique, rare, spectacular and threatenedtaxa. Hillman (1986: 255) considered theMountains ‘a centreof faunal endemicity, probably with the highest rate ofanimal endemicity for a terrestrial habitat anywhere inthe world’. The region incorporates the largest single area(. 4,000 km2) in Africa over 3,000 m altitude, and is acrucially important reservoir of globally threatenedAfromontane and Afroalpine habitats (Largen & Spawls,2010).
The highest part of the region, the Sanetti Plateau andassociated peaks, comprises Afroalpine grasslands with ahighly endemic biota. To the south, the Sanetti Plateauterminates at the Harenna escarpment that drops from3,200 to 2,000 m in 8 km. This escarpment exhibits adramatic vegetation zonation from the plateau grasslandsthrough low heather scrub, giant heather (Erica spp.) forest,Hagenia, bamboo and Podocarpus forest, high altitude
woodland and into medium altitude rainforest (Friis, 1986;Miehe &Miehe, 1993, 1994). This is one of the largest naturalareas of forest remaining in Ethiopia. Towards the west,north and east the Sanetti Plateau falls away more gentlyand change in vegetation is less abrupt, with Afroalpinegrasslands giving way to heather moorlands and, belowc. 3,000 m, Hagenia forest.
A substantial area (c. 2,200 km2) of the region is coveredby the Bale Mountains National Park, established in 1971 butnot yet formally gazetted by the Ethiopian government. ThePark is the most important conservation area in Ethiopia(Frankfurt Zoological Society, 2007), one of themost import-ant in Africa (Largen & Spawls, 2010), and one of 34
Conservation International global biodiversity hotspots.The Park is a BirdLife International Important Bird Area,and on the Tentative List to qualify as aWorld Heritage Site.Many people live and exploit natural resources withinthe Park, and their activities are the primary threat to thenative biodiversity (Frankfurt Zoological Society, 2007).Conservation efforts are currently focused on habitats, andon flagship threatened bird and mammal species, which areconcomitantly the taxa that are best known scientifically.For example, the Park is the focus of the Ethiopian Wolf(Canis simensis) Conservation Programme (e.g. Sillero-Zubiri et al., 1997; Atickem et al., 2009), and the mainremaining stronghold of the mountain nyala Tragelaphusbuxtoni (Malcolm & Evangelista, 2005) and Bale monkeyChlorocebus djamdjamensis (Mekonnen, 2008; Mekonnenet al., 2010). The Park is important as a water source for c. 12million people and of outstanding social, scenic and culturalvalue (Hillman, 1988; Frankfurt Zoological Society, 2007).A management plan for the Park was published recently(Frankfurt Zoological Society, 2007).
About 15 amphibian species (all anurans) have beenreported from the Bale Mountains, including 10 endemics(Largen, 2001; Largen & Spawls, 2010). The Bale MountainsNational Park is an Alliance for Zero Extinction site,partly because of its endemic amphibians (Alliance for ZeroExtinction, 2010). Given the Park’s importance for theirconservation we focus here on four monotypic frog genera:all endemic to Ethiopia and three endemic to the BaleMountains.
Previous work up to 1986
Although there is a long history of human habitation in thearea, the Bale Mountains remained largely unexploredscientifically until the middle of the 20th century. TheChicago Field Museum’s W.H. Osgood collected some am-phibians and reptiles in 1926–1927. Partly in association withother British colleagues, M.J. Largen carried out extensivecollections between 1971 and 1975. Early expeditions werecentred around Dinsho/Dinshu and the northern slopesbelow the Sanetti Plateau. In 1983 it become feasible to reach
Dinsho
Goba
Robe
Legend
Road
4,300 m
1,500 m
To Dola Mena
0 3.75 7.5 15
km
Rira
H1
H2
G1
G2
D1D2
C1
C2F1
E1E2
E3
A1
A2
A3A4
A5
B2
B1
A2–A5
Altitude
N
FIG. 1 Map of the Bale Mountains National Park, Ethiopiashowing Park boundary, local villages and towns (squares), andsampling localities A–H (dots) referred to in the text andTable 1. The dots labelled A2 to A5 are based on coordinatesgiven in error by Grandison (1978) and Largen (2001); theselocalities are actually close to the Goba–Rira road, approximatelywithin the area marked by the ellipse labelled A2–A5. Rira, in thecentre of the Park, is at 6.771°N, 39.725°E.
60 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
the Harenna Forest on the southern side of the massif byroad. Conscious of the conservation threat posed by the newroad the Ethiopian Wildlife Conservation Organizationarranged an expedition with Addis Ababa University toconduct a preliminary survey of the Harenna Forest inAugust 1986.
We follow the taxonomy of the latest comprehensivetreatment of Ethiopian amphibians (Largen, 2001; Largen &Spawls, 2010). The localities reported for Spinophrynoides,Altiphrynoides and some other Bale Mountain frogs assouth-east of Goba (Grandison, 1978; Largen, 2001) are inerror. These localities are south of Goba along the Goba–Rira road; the reported altitudes are approximately correctbut the coordinates are not.
Focal species
Fuller accounts of the systematics, natural history and con-servation biology of the four species are given in Appendix 1.
Spinophrynoides osgoodi (Plate 1a) was described in 1932
from specimens collected between 1926 and 1927, froman imprecise type locality (‘Gedeb Mountains’, Loveridge,1932). It has been recorded in Ethiopia from beyond the Bale
Mountains but the vast majority of records are from northand south of the Sanetti Plateau, typically in montane forestbut extending also into open moorland at 1,950–3,520 m(Largen, 2001). It is a small to medium (snout–vent length to62 mm) species that lays strings of eggs in small pools thatdevelop into tadpoles before metamorphosing into froglets(Grandison, 1978; Largen, 2001). The presence of thisEthiopian endemic genus in the Bale Mountains has beenoverlooked by some non-herpetologists and conservationmanagers (e.g. WCMC, 1993; Miehe &Miehe, 1994) becauseS. osgoodi was not classified in the genus Spinophrynoidesuntil 1987. It has been categorized as Vulnerable on theIUCNRed List since the first Global Amphibian Assessmentin 2004 (Stuart et al., 2008; IUCN, 2010).
Altiphrynoides malcolmi (Plate 1b) was described in 1978
from . 160 specimens collected between 1971 and 1975
from the northern side of the Sanetti Plateau. The specieswas collected subsequently from the Harenna escarpment.It is endemic to the Bale Mountains, reported from altitudesof 3,200–4,000 m, spanning the upper limits of theSchefflera–Hagenia–Hypericum forest to Afroalpine moor-land (Largen, 2001). This small species (snout–vent length, 32 mm, Largen, 2001) has internal fertilization and lays
(a) (b)
(c) (d)
(a) (b)
(c) (d)
PLATE 1 The four endemic Ethiopian monotypic frog genera studied: (a) Spinophrynoides osgoodi, (b) Altiphrynoides malcolmi,(c) Ericabatrachus baleensis, and (d) Balebreviceps hillmani. S. osgoodi is endemic to the highlands of Ethiopia; the others are endemicto the Bale Mountains. (a) by S. Spawls, (b–d) by M. Menegon.
Endemic frogs in the Bale Mountains 61
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
small clutches of large eggs communally in moist terrestrialsites that hatch into non-feeding tadpoles that remain onland (Grandison, 1978). Because A. malcolmi has beenclassified in Altiphrynoides only since 1987 the presence ofthis endemic genus has been overlooked in some summariesof the fauna (e.g. WCMC, 1993; Miehe &Miehe, 1994). It hasbeen categorized as Endangered on the IUCN Red List sincethe first Global Amphibian Assessment in 2004 (Stuartet al., 2008; IUCN, 2010).
Ericabatrachus baleensis (Plate 1c) is endemic to theHarenna escarpment and was described in 1991 fromspecimens collected in 1986. Twenty-two of the types werecollected from the margins of a stream in giant Erica forestat the type locality (Tulla Negesso) for Balebrevicepshillmani (see below) at 3,200 m, with one specimen comingfrom a stream at Katcha, at 2,400 m. We know of nopublished records beyond the type series. The reproductionand life history of E. baleensis are unknown but it hasbeen postulated to lay terrestrial eggs that possiblydevelop directly without a tadpole stage (Largen, 1991).The presence of E. baleensis and its endemicity have beenoverlooked by at least some important works on the BaleMountains (e.g. Miehe & Miehe, 1994). It has beencategorized as Endangered on the IUCN Red List sincethe first Global Amphibian Assessment in 2004 (Stuartet al., 2008; IUCN, 2010).
B. hillmani (Plate 1d) was described in 1989 fromspecimens collected in 1986 from a single locality (TullaNegesso). To the best of our knowledge no subsequentcollections had been made, nor any reports published forany other locality (e.g. Largen & Spawls, 2010). The typeswere collected from beneath logs and boulders within giantErica forest 50 m below the timber line on the Harennaescarpment (Largen & Drewes, 1989). The presence ofB. hillmani and its endemicity were overlooked by at leastsome important works on the Bale Mountains (e.g. Miehe &Miehe, 1994). The breeding biology and life history ofB. hillmani are unknown but it is predicted to have terrestrialeggs with direct development (Largen & Drewes, 1989;Müller et al., 2007). It has been categorized as Endangeredon the IUCN Red List since the first Global AmphibianAssessment in 2004 (Stuart et al., 2008; IUCN, 2010).
Methods
We evaluated data on the change in abundance of the fourendemic genera, combining information from five previousfield visits (1971–1986) with that from seven teams thatconducted amphibian and reptile survey fieldwork on 10
subsequent occasions (1994–2009). All six expeditions thattook place this century included the four focal species asspecific targets, and the teams were equipped with detailedhistorical locality information for these taxa. Fieldworktook place by day and night, employing visual and audio
encounter surveys. Searching included netting of streamsand ponds, inspecting trees and shrubs, digging leaf litterand surface soil, rolling logs and rocks, raking leaf litter, andsearching through ground vegetation by hand, includingsome intensive searches in 25 m2 quadrats. Particular effortwas applied along the edge of water bodies. Details of thefield effort undertaken and frogs encountered are providedin Appendix 2. All teams recorded numbers of individualsof the four endemic frog genera. Teams 11–14 recordedsearch effort (as number of person hours searching) while inthe field. Search effort data for the other teams, includingexpeditions between 1971 and 1999, were compiled retro-spectively from the field notebooks of expedition members.Frogs other than the four focal species were noted butprecise numbers of individuals encountered were generallynot recorded.
Field data were converted into numbers of frogsencountered per person hour of searching. These datawere grouped into eight localities (Fig. 1): (A) North ofSanetti (2,800–3,900 m), mostly around Dinsho and southof Goba up to the northern edge of the Sanetti Plateau,(B) Sanetti Plateau (c. 3,800–4,150 m), (C) Tulla Negessoand areas close to the road, down to the lower edge of themain centre of Rira village (2,900–3,250 m), (D) Fute andKotinsa, superficially less disturbed (than Tulla Negesso)giant Erica forest along the Harenna escarpment east ofthe road (3,200–3,255 m), (E) Degebato and Galema,superficially less disturbed giant Erica forest along theHarenna escarpment west of the road (3,200–3,300 m),(F) In the vicinity of the WWF hut below Rira, down tothe lower rim of the Rira basin (2,700–2,850 m) but closeto the road, (G) in and immediately around theKatcha clearing (c. 2,350–2,450 m), and (H) around theShawe River bridge and areas close to the road south ofhere, including the marshy clearing at Magano (c. 1,700–1,950 m).
Likelihood of site-level extinction of frogs was assessedwith optimal linear estimation (Cooke, 1980; Solow, 2005), anon-parametric method that allows extinction date to beestimated based on the distribution of most recent sightings.We applied the technique reported by Solow (2005) togenerate an estimate of extinction date, with 95% confidencelimits (see Appendix 3 for full details). Change in numbersof individuals over time was assessed by fitting models in ageneralized additive modelling (GAM) framework. Weused GAM to model sightings as a function of effort trendas a smooth, non-linear function of time, and to providea framework for testing the statistical significance ofchanges. GAM is advantageous in long-term trend analysesbecause it allows change in mean abundance to followany smooth curve, not only linear forms (Fewster et al.,2000). Calculations were carried out using the MGCVpackage (Wood, 2006) in R v. 2.11 (R Development CoreTeam, 2010).
62 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69
http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
TABLE 1 Numbers (and individuals per hour in parentheses) of the four species of frogs surveyed in the Bale Mountains across eightlocalities (A–H; see Methods for further details and Fig. 1) during 15 numbered field trips between 1971 and 2009.
Month Year TripSpinophrynoidesosgoodi
Altiphrynoidesmalcolmi
Ericabatrachusbaleensis
Balebrevicepshillmani
Totalhours
(A) North of SanettiDec. 1971 1 2 (0.13) 2 (0.13) 16Apr. 1972 2 3 (1) 3Apr. 1974 3 49 (9.8) 115 (23) 5Apr. 1974 4 32 (1.6) 75 (3.75) 20Dec./Jan. 1994/1995 6 6Sep. 1996 7 3 (0.38) 8Dec. 1996 8 6Oct. 1997 9 8July 2006 10 3Sep. 2006 11 1 (0.05) 21.5July/Aug. 2008 12 4(B) Sanetti PlateauDec. 1971 1 3 (0.5) 6Dec./Jan. 1994/1995 6 6Dec. 1996 8 6Oct. 1997 9 8July 2006 10 3Sep. 2006 11 1.5July/Aug. 2008 12 9June 2009 14 15(C) Tulla Negesso & environsAug. 1986 5 10 (0.33) 30 (1) 23 (0.77) 18 (0.6) 30Dec./Jan. 1994/1995 6 ? 1 (0.5) 2Sep. 1996 7 20 + (1 +) 1 (0.05) 20Dec. 1996 8 18 (3) 1 (0.17) 6Oct. 1997 9 8July 2006 10 15Sep. 2006 11 3 (0.1) 4 (0.13) 30.5July/Aug. 2008 12 1 (0.04) 4 (0.17) 23Feb./Mar. 2009 13 3 (0.26) 2 (0.17) 11.5June 2009 14 1 (0.1) 1 (0.1) 10.5Sep. 2009 15 1 (0.07) 14(D) Fute & KotinsaJuly/Aug. 2008 12 4 (0.1) 1 (0.03) 3 (0.08) 39Feb./Mar. 2009 13 5 (0.48) 1 (0.1) 10.5June 2009 14 5 (0.2) 2 (0.08) 14 (0.56) 25Sep. 2009 15 1 (0.14) 7(E) Degebato & GalemaSep. 2006 11 5July/Aug. 2008 12 11Feb./Mar. 2009 13 2 (0.22) 9(F) Vicinity of WWF hut below RiraDec./Jan. 1994/1995 6 40 (10) 4July 2006 10 4July/Aug. 2008 12 15Feb./Mar. 2009 13 8.5June 2009 14 5(G) Katcha clearingAug. 1986 5 25 (1.32) 1 (0.05) 19July 2006 10 9Sep. 2006 11 9July/Aug. 2008 12 8Feb./Mar. 2009 13 5June 2009 14 8Sep. 2009 15 6
Endemic frogs in the Bale Mountains 63
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69
http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
Results
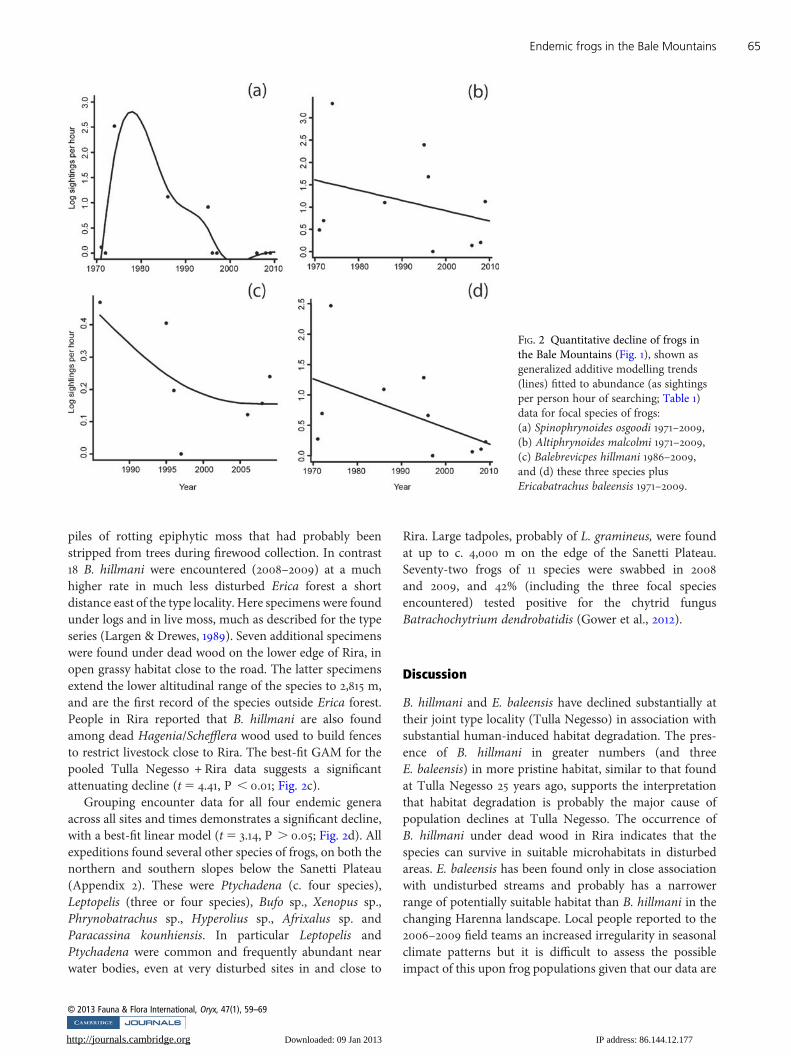
Raw encounter data are reported in Appendix 2 andsummarized in Table 1. A single S. osgoodi (identified from aphotograph provided by M. Hoffman) was found at Katchaon 14 October 2003 but we were aware of this visit and findonly after our study had been completed and so it is notincluded in our quantitative analyses. S. Spawls possiblyfound three S. osgoodi in a clearing 3 km south of Rira(January 1995) but their identity has not been confirmed(possibly Altiphrynoides instead). No specimens were foundby later expeditions during substantial fieldwork, so that tothe best of our knowledge S. osgoodi has not been collectedsince 1986, perhaps observed only twice since then, and notobserved since 2003 despite concerted attempts to locatespecimens. From the quantitative data, Cooke’s estimatorgives an estimated date of extirpation of S. osgoodi fromthe sampled region of 2016 but the confidence interval, CI, iswide (95%CI 1999–2781; 80%CI 2003–2100). The encounterrate data are too non-linear to use simple regression to testfor significant decline but a GAM using sightings pooledacross study sites showed a good fit, illustrating significantdecline since the mid 1970s (t5 2.95, P . 0.05; Fig. 2a).
More than 100 A. malcolmi were recorded in four visitsduring 1994–1999, most on the Harenna escarpment butalso on the north-east edge of the Sanetti Plateau. A further25 were found in six subsequent visits (2006–2009), all butone on the Harenna escarpment. Together these specimensextend the altitudinal range of the species to c. 2,500–4,000 m (from 3,200–4,000 m, Largen, 2001). The Harennaspecimens were found during the day, almost all under logs,and some (including all from the north-east edge of Sanetti)
under rocks. Analysis of encounter data with GAM showeda significant decline in abundance using sightings pooledacross study sites (t5 3.25, P, 0.05; Fig. 2b).
Only three E. baleensis were found by the 10 expeditionspost-1986 (1994–2009), and all of these in relatively undis-turbed habitat east of the type locality. Despite dedicatedsearches no specimens have been found at the type localityor at Katcha since their original discovery. Statisticalanalysis is not appropriate for the patchy quantitative databut E. baleensis is probably extirpated or rare at the typelocality. Comparison of the stream at the type locality at thetime of discovery (1986) and the recent expeditions (2006–2009) reveals substantial habitat alteration (Plate 2). Thestream is no longer a swift mountain torrent (Largen, 1991)but is now a broad, boggy, muddy and relatively slow-flowing waterway heavily trampled by domesticated cattle.The stream is also now used by local people to wash clothes,with one section diverted to form a leat that serves thenearby dwelling. This habitat change happened between1999 and 2006.
Six specimens of B. hillmani have been found at the typelocality since the original collection in 1986. Only three ofthese were found in the six visits since 2006, despiteconsiderable effort. Comparison of the type locality giantErica forest in 1986 and 2006–2009 demonstrates substantialchange (Plate 2). There is now a greatly reduced leaf litter,fewer trees, an almost compete absence of logs (removed forfirewood and fencing), a human settlement on the edge ofthe forest, and an introduction of cows and other livestockthat has resulted in the forest floor being heavily grazed,trampled and crossed by muddy tracks. At least two of therecent B. hillmani from the type locality were found under
Table 1 (Continued)
Month Year TripSpinophrynoidesosgoodi
Altiphrynoidesmalcolmi
Ericabatrachusbaleensis
Balebrevicepshillmani
Totalhours
(H) Shawe River bridge & MaganoAug. 1986 5 1 (0.06) 16July 2006 10 4Sep. 2006 11 3June 2009 14 19Sep. 2009 15 3Tulla Negesso only*Aug. 1986 5 10 (0.33) 30 (1) 23 (0.77) 18 (0.6) 30Dec./Jan. 1994/1995 6 1 (0.5) 2Sep. 1996 7 1 (0.25) 4Dec. 1996 8 1 (0.5) 2July 2006 10 4Sep. 2006 11 1 (0.06) 16.5July/Aug. 2008 12 1 (0.08) 13Feb./Mar. 2009 13 3 (0.67) 1 (0.22) 4.5June 2009 14 1 (0.18) 5.5Sep. 2009 15 1 (0.17) 6
*Also part of locality (C) and is the type locality of E. baleensis and B. hillmani.
64 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69
http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
piles of rotting epiphytic moss that had probably beenstripped from trees during firewood collection. In contrast18 B. hillmani were encountered (2008–2009) at a muchhigher rate in much less disturbed Erica forest a shortdistance east of the type locality. Here specimens were foundunder logs and in live moss, much as described for the typeseries (Largen & Drewes, 1989). Seven additional specimenswere found under dead wood on the lower edge of Rira, inopen grassy habitat close to the road. The latter specimensextend the lower altitudinal range of the species to 2,815 m,and are the first record of the species outside Erica forest.People in Rira reported that B. hillmani are also foundamong dead Hagenia/Schefflera wood used to build fencesto restrict livestock close to Rira. The best-fit GAM for thepooled Tulla Negesso + Rira data suggests a significantattenuating decline (t5 4.41, P , 0.01; Fig. 2c).
Grouping encounter data for all four endemic generaacross all sites and times demonstrates a significant decline,with a best-fit linear model (t5 3.14, P . 0.05; Fig. 2d). Allexpeditions found several other species of frogs, on both thenorthern and southern slopes below the Sanetti Plateau(Appendix 2). These were Ptychadena (c. four species),Leptopelis (three or four species), Bufo sp., Xenopus sp.,Phrynobatrachus sp., Hyperolius sp., Afrixalus sp. andParacassina kounhiensis. In particular Leptopelis andPtychadena were common and frequently abundant nearwater bodies, even at very disturbed sites in and close to
Rira. Large tadpoles, probably of L. gramineus, were foundat up to c. 4,000 m on the edge of the Sanetti Plateau.Seventy-two frogs of 11 species were swabbed in 2008
and 2009, and 42% (including the three focal speciesencountered) tested positive for the chytrid fungusBatrachochytrium dendrobatidis (Gower et al., 2012).
Discussion
B. hillmani and E. baleensis have declined substantially attheir joint type locality (Tulla Negesso) in association withsubstantial human-induced habitat degradation. The pres-ence of B. hillmani in greater numbers (and threeE. baleensis) in more pristine habitat, similar to that foundat Tulla Negesso 25 years ago, supports the interpretationthat habitat degradation is probably the major cause ofpopulation declines at Tulla Negesso. The occurrence ofB. hillmani under dead wood in Rira indicates that thespecies can survive in suitable microhabitats in disturbedareas. E. baleensis has been found only in close associationwith undisturbed streams and probably has a narrowerrange of potentially suitable habitat than B. hillmani in thechanging Harenna landscape. Local people reported to the2006–2009 field teams an increased irregularity in seasonalclimate patterns but it is difficult to assess the possibleimpact of this upon frog populations given that our data are
FIG. 2 Quantitative decline of frogs inthe Bale Mountains (Fig. 1), shown asgeneralized additive modelling trends(lines) fitted to abundance (as sightingsper person hour of searching; Table 1)data for focal species of frogs:(a) Spinophrynoides osgoodi 1971–2009,(b) Altiphrynoides malcolmi 1971–2009,(c) Balebrevicpes hillmani 1986–2009,and (d) these three species plusEricabatrachus baleensis 1971–2009.
Endemic frogs in the Bale Mountains 65
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
from sites that have undergone rapid habitat change. Morework is needed if the impact of future expected climatechange is to be understood.
S. osgoodi has declined greatly in the Bale Mountains tothe extent that although previously considered to be‘particularly common in the Bale Mountains NationalPark’ (Largen & Spawls, 2010; see also IUCN, 2010), itscontinued existence is not yet established. The abundance ofA. malcolmi has also decreased significantly. Given ourevidence, the IUCN Red List categories of these four speciesshould be reassessed. The current IUCN assessmentsinclude statements about abundance and threat that arenow known to be incorrect. Although B. dendrobatidiswas recorded no dead or moribund frogs were found, andit remains to be seen to what extent this pathogen isimplicated in the declines. Although B. dendrobatidisis known to kill frogs elsewhere it has been associatedwith the decline of only a single species of African frog,Nectophrynoides asperginus in Tanzania, and even thenhabitat deterioration was also implicated (Weldon & duPreez, 2004; Channing et al., 2006).
The Bale Mountains National Park is facing majorchallenges (Frankfurt Zoological Society, 2007), causedchiefly by the rapidly growing human population. Evenwithin the short period of 2006–2009 the negative impact ofhumans on the ecosystems of the Park was apparent, withdeforestation at Harenna and increasing effects of livestock.Given that most conservation in the Bale Mountains hashitherto focused on large mammals (as elsewhere inEthiopia, Loader et al., 2009), the threats to Bale ecosystemsmore generally have been under appreciated. The continuedpresence of endemic mammals and patches of, for example,giant Erica, could lead to the superficial conclusion that Baleforests are relatively stable but our examination of a smallerfaunal element, the frogs, presents a different perspective.
We suggest several priority research objectives for theBale amphibian fauna. More intensive and temporally andspatially broader searches for S. osgoodi are required, withhunts for breeding aggregations such as those described byGrandison (1978) perhaps the best target. More surveys arerequired to complete an inventory of the amphibians, withsurveys away from roads and in more pristine habitat as well
(a) (b)
(c) (d)
(a) (b)
(c) (d)
PLATE 2 Habitat degradation in the Harenna Forest, Bale Mountains (Fig. 1): (a) stream trampled by cattle in Erica forest at the typelocality (Tulla Negesso) of Balebreviceps and Ericabatrachus, September 2006, (b) livestock trails, cut wood and lack of ground cover inErica forest between Tulla Negesso and Fute, December 2010, (c) collection site of Balebreviceps under rotten wood forming part of oldlivestock fence (new bamboo fence in background) in heavily grazed clearance just below Rira, December 2010, and (d) house andbamboo fencing in man-made clearance in Erica forest, Fute, June 2009.
66 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
as in degraded ecosystems; these surveys should includeareas surrounding the National Park. More basic ecologicaldata on frogs need to be gathered, especially on thereproductive biology of B. hillmani and E. baleensis. Moresurveys are required across a range of localities and habitatsto determine where monitoring efforts could best befocused. Research on other Bale organisms is required,with the works of Miehe & Miehe (1993, 1994), studies ofendemic mammals (see Introduction) and the BaleMountains National Park General Management Plan(Frankfurt Zoological Society, 2007) providing a frameworkwith which this can be integrated.
In terms of conservation management the growingnumbers and destructive activities of people living withinthe National Park need to be addressed. When the Park isformally gazetted all settlement will be illegal but until thentheir status remains a grey area. Protection of key habitats isan immediate priority until human disturbance is haltedacross the Park. Human population growth in Ethiopia ishigh (Largen & Spawls, 2010; UN, Department of Economicand Social Affairs, Population Division, 2011), and this isreflected in the increase in people living within the Parkfrom 2,500 in 1986 (Hillman, 1986) to . 40,000 in 2003
(Frankfurt Zoological Society, 2007). The Erica forests of theBale Mountains have been limited historically by fire andgrazing (Miehe & Miehe, 1993, 1994) but more activedeforestation is now taking place, extended grazing andwood collecting are altering the forest floor, and regener-ation of these forests remains poorly understood. Theimportance of working properly with local people inconservation management in Africa has been recognized(Abrams et al., 2009) but we believe that within theBale Mountains National Park the right balance has not yetbeen reached. The General Management Plan (FrankfurtZoological Society, 2007) already recognizes much of whatwe conclude here but our data for endemic frogs emphasizesthe urgency of conducting further biological research anddeveloping appropriate conservation management in theregion.
Acknowledgements
For advice, help with fieldwork, and permits for researchand export, we thank (in no particular order) Hailu Tadesse,Kifle Argaw, Yeneneh Teka, Daniel Pawlos and BirutesfaYimer (Ethiopian Wildlife Conservation Authority),Afework Bekele and Abebe Getahun (Addis AbabaUniversity), Isa (WWF), Daniel Tilaye (Bale MountainsNational Park), Ryan Kerney (Harvard University), AlastairNelson, Anouska Kinahan and Thadaigh Baggallay(Frankfurt Zoological Society), Michael Geiser (Universityof Basel), Zelealem Tefera, Addis Berhanu, SisayShewamene, Red Jackal Tour Operators, the people of
Rira, Colin Tilbury, Bill Branch, Chris Smith, Jon Gower,Alexia Kerney, Jonathan and Tim Spawls, and our guidesin 2008–2009 (Issa, Ahmed, Hussein, Hussein andMohammed). The Harvard University expedition receivedfunding from the Museum of Comparative Zoology’sPutnam Expedition Grants. Fieldwork in 2008 and2009 was supported in part by a National GeographicExpedition Grant (CRE Grant #8532–08). The followingbodies also funded surveys and laboratory work: EthiopianWildlife Conservation Authority, University of Basel,European Union Synthesys project (AAM), The NaturalHistory Museum London (DJG, SPL), University ofRichmond Faculty Research Committee (RdS), andStipendienkommission für Nachwuchskräfte ausEntwicklungsländern, Basel (AAM).
References
ABRAMS, R.W., ANWANA, E.D., ORMSBY, A., DOVIE, D.B.K.,AJAGBE, A. & ABRAMS, A. (2009) Integrating top-down withbottom-up conservation policy in Africa. Conservation Biology, 23,799–804.
ADEBA, P.J., KOUASSI , P. & RÖDEL, M.-O. (2010) Anuran amphibiansin a rapidly changing environment—revisiting Lamto, Cote d’Ivoire,40 years after the first herpetofaunal investigations. African Journalof Herpetology, 59, 1–16.
ALLIANCE FOR ZERO EXTINCTION (2010) 2010 AZE Update. Http://www.zeroextinction.org [accessed 1 July 2012].
ATICKEM, A., BEKELE, A. & WILLIAMS, S.D. (2009) Competitionbetween domestic dogs and Ethiopian wolf (Canis simensis) in theBale Mountains National Park, Ethiopia. African Journal of Ecology,48, 401–407.
CHANNING, A., FINLOW-BATES, K.S., HAARKLAU, S.E. &HAWKES, P.G. (2006) The biology and recent history of theCritically Endangered Kihansi spray toad Nectophrynoidesasperginis in Tanzania. Journal of East African Natural History, 95,117–138.
COLLEN, B., PURVIS, A. & MACE, G.M. (2010) When is a species reallyextinct? Inferring extinction from a sightings record to informconservation assessment. Diversity and Distributions, 16, 755–764.
COOKE, P. (1980) Optimal linear estimation of bounds of randomvariables. Biometrika, 67, 257–258.
CRAWLEY, M.J. (2007) The R Book. Wiley, Chichester, UK.DUBOIS , A. (1987) Miscellanea taxinomica batrachologica (I). Alytes, 5,
7–95.DUBOIS , A. (2005) Amphibia Mundi. 1.1. An ergotaxonomy of recent
amphibians. Alytes, 23, 1–24.FEWSTER, R.M., BUCKLAND, S.T., SIRIWARDENA, G.M., BAILLIE, S.R.
& WILSON, J.D. (2000) Analysis of population trends for farmlandbirds using generalized additive models. Ecology, 81, 1970–1984.
FRANKFURT ZOOLOGICAL SOCIETY (2007) Bale Mountains NationalPark. General Management Plan 2007–2017. Frankfurt ZoologicalSociety Report, Frankfurt, Germany. Http://www.zgf.de/download/166/BMNP_GMP_2007.pdf [accessed 1 July 2012].
FRI IS , I. (1986) Zonation of forest vegetation on the south slopes of BaleMountains, south Ethiopia. SINET: Ethiopian Journal of Science, 9,29–44.
FROST, D.R. (2011) Amphibian Species of the World: An OnlineReference. Version 5.5. Http://research.amnh.org/vz/herpetology/amphibia/ [accessed 1 July 2012].
Endemic frogs in the Bale Mountains 67
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
FROST, D.R., GRANT, T., FAIVOVICH, J., BAIN, R.H., HAAS, A.,HADDAD, C.F.B. et al. (2006) The amphibian tree of life. Bulletin ofthe American Museum of Natural History, 297, 1–370.
GOWER, D.J., DOHERTY-BONE, T.M., ABERRA, R.K., MENGISTU, A.A.,SCHWALLER, S., MENEGON, M. et al. (2012) High prevalence of theamphibian chytrid fungus (Batrachochytrium dendrobatidis) acrossmultiple taxa and localities in the highlands of Ethiopia.Herpetological Journal, 22, 225–233.
GRANDISON, A.G.C. (1978) The occurrence of Nectophrynoides(Anura Bufonidae) in Ethiopia. A new concept of the genus witha description of a new species. Monitore Zoologico Italiano(Nuova Serie) Supplemento, 11, 119–172.
GREEN, D.M. (2003) The ecology of extinction: population fluctuationand decline in amphibians. Biological Conservation, 111, 331–343.
HILLMAN, J.C. (1986) Bale Mountains National Park. ManagementPlan. Ethiopian Wildlife Conservation Organization, Addis Ababa,Ethiopia.
HILLMAN, J.C. (1988) The Bale Mountains National Park area,southeast Ethiopia, and its management. Mountain Research andDevelopment, 8, 253–258.
IUCN (2004) IUCN Red List of Threatened Species 2004. Http://www.iucnredlist.org [accessed 15 October 2005].
IUCN (2010) IUCN Red List of Threatened Species v. 2010.3. Http://www.iucnredlist.org [accessed 22 September 2010].
KILPATRICK, A.M., BRIGGS, C.J. & DASZAK, P. (2010) The ecology andimpact of chytridiomycosis: an emerging disease of amphibians.Trends in Ecology & Evolution, 25, 109–118.
LARGEN, M.J. (1991) A new genus and species of petropedatine frog(Amphibia Anura Ranidae) from high altitude in the mountains ofEthiopia. Tropical Zoology, 4, 139–152.
LARGEN, M.J. (2001) Catalogue of the amphibians of Ethiopia,including a key for their identification. Tropical Zoology, 14,307–402.
LARGEN, M.J. & DREWES, R.C. (1989) A new genus and speciesof brevicpitine frog (Amphibia Anura Microhylidae) from highaltitude in the mountains of Ethiopia. Tropical Zoology, 2, 13–30.
LARGEN, M.J. & SPAWLS, S. (2010) The Amphibians and Reptiles ofEthiopia and Eritrea. Chimaira, Frankfurt Am Main, Germany.
LOADER, S.P., GOWER, D.J., HOWELL, K.H., DOGGART, N.,RÖDEL, M.-O., CLARKE, B.T. et al. (2004) Phylogenetic relationshipsof African microhylid frogs inferred from DNA sequences ofmitochondrial 12S and 16S rRNA genes. Organisms, Diversity &Evolution, 4, 227–235.
LOADER, S.P., MENGISTU, A., SCHWALLER, S., GOWER, D.,NAGEL, P.N., GETAHUN, A. et al. (2009) Are Ethiopian highlandschanging? Amphibians as ecosystem indicators. Mountain ForumBulletin, 9, 5–6.
LOVERIDGE, A. (1932) Eight new toads of the genus Buho fromEast andCentral Africa. Occasional Papers of the Boston Society of NaturalHistory, 8, 43–54.
MALCOLM, J. & EVANGELISTA, P.H. (2005) The Range and Status of theMountain Nyala. EthiopianWolf Conservation Programme Report.Http://www.ethiopianwolf.org/publications/Malcolm-Evangelista-2005-Mountain-Nyala-Report.pdf [accessed 1 July 2012].
MARSH, D.M. (2001) Fluctuations in amphibian populations: a meta-analysis. Biological Conservation, 101, 327–335.
MEKONNEN, A. (2008) Distribution of the Bale monkey (Chlorocebusdjamdjamensis) in the Bale Mountains and its ecology in theOdobullu Forest, Ethiopia—A study of habitat preference, populationsize, feeding behaviour, activity and ranging patterns. MSc thesis.Addis Ababa University, Addis Ababa, Ethiopia. Http://www.
ethiopianwolf.org/publications/Addisu%20Bale%20monkey%20Thesis%20July%202008.pdf [accessed 28 June 2012].
MEKONNEN, A., BEKELE, A., HEMSON, G., TESHOME, E. &ATICKEM, A. (2010) Population size and habitat preference of theVulnerable Bale monkey Chlorocebus djamdjamensis in OdobulluForest and its distribution across the Bale Mountains, Ethiopia.Oryx, 44, 558–563.
MEYER, A.H., SCHMIDT, B.R. & GROSSENBACHER, K. (1998) Analysisof three amphibian populations with quarter-century long time-series. Proceedings of the Royal Society, B, 265, 523–528.
MIEHE, G. & MIEHE, S. (1993) On the physiognomic and floristicdifferentiation of ericaceous vegetation in the Bale Mountains, SEEthiopia. Opera Botanica, 121, 85–117.
MIEHE, S. & MIEHE, G. (1994) Ericaceous Forests and Heathlands inthe Bale Mountains of South Ethiopia. Stiftung walderhaltung inAfrika & Bundesforschungsansalt für Forst- und Holzwirtschaft,Hamburg, Germany.
MÜLLER, H., LOADER, S.P., NGALASON, W., HOWELL, K.M. &GOWER, D.J. (2007) Reproduction in brevicipitid frogs (Amphibia:Anura: Brevicipitidae)—evidence from Probreviceps m.macrodactylus. Copeia, 2007, 726–733.
PECHMANN, J.H.K., SCOTT, D.E., SEMLITSCH, R.D., CALDWELL, J.P.,VITT, L.J. & GIBBONS, J.W. (1991) Declining amphibianpopulations: the problems of separating human impacts fromnatural fluctuations. Science, 253, 892–895.
PIMM, S.L. (2007) Africa: still the dark continent. Conservation Biology,21, 567–569.
R DEVELOPMENT CORE TEAM (2010) R: A language and Environmentfor Statistical Computing. R Foundation for Statistical Computing,Vienna, Austria.
SEMLITSCH, R.D., SCOTT, D.E., PECHMANN, J.H.K. & GIBBONS, J.W.(1996) Structure and dynamics of an amphibian community.Evidence from a 16-year study of a natural pond. In Long-TermStudies of Vertebrate Communities (eds M.L. Cody & J.A.Smallwood), pp. 217–248. Academic Press, San Diego, USA.
SILLERO-ZUBIRI , C., MACDONALD, D. & IUCN/SPECIES SURVIVAL
COMMISSION CANID SPECIALIST GROUP (1997) The EthiopianWolf—Status Survey and Conservation Action Plan. IUCN, Gland,Switzerland.
SOLOW, A.R. (2005) Inferring extinction from a sighting record.Mathematical Biosciences, 195, 47–55.
STUART, S.N., HOFFMAN, M., CHANSON, J.S., COX, N.A.,BERRIDGE, R.J., RAMANI, P. & YOUNG, B.E. (2008) ThreatenedAmphibians of the World. Lynx Edicions, Barcelona, Spain,IUCN, Gland, Switzerland &Conservation International, Arlington,USA.
TANDY, M. & KEITH, R. (1972) Bufo of Africa. In Evolution in theGenus Bufo (ed. W.F. Blair), pp. 119–170. University of Texas Press,Austin, USA.
UN, DEPARTMENT OF ECONOMIC AND SOCIAL AFFAIRS,POPULATION DIVIS ION (2011)World Population Prospects: The 2010Revision. Http://esa.un.org/unpd/wpp/Documentation/publications.htm [accessed 1 July 2012].
WELDON, C. & DU PREEZ, L.H. (2004) Decline of Kihansi spray toad,Nectophrynoides asperginis, from the Udzungwa mountains,Tanzania. Froglog, 62, 2–3.
WOOD, S.N. (2006) Generalized Additive Models: An Introduction.Chapman and Hall/CRC Press, Boca Raton, Florida, USA.
WCMC (WORLD CONSERVATION MONITORING CENTRE) (1993)World Checklist of Threatened Amphibians. Joint NatureConservation Committee, Peterborough, UK.
68 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

http://journals.cambridge.org Downloaded: 09 Jan 2013 IP address: 86.144.12.177
Appendices 1–3
The appendices for this article are available online at http://journals.cambridge.org
Biographical sketches
DAVID GOWER is a herpetologist specializing in systematics andevolution. ROMAN ABERRA works in Ethiopian conservation, with aspecial interest in frogs. SILVIA SCHWALLER conducted her Masters thesison environmental change in the Harenna Forest. MALCOLM LARGEN
is a leading authority on herpetology in Ethiopia. BEN COLLEN isa conservation biologist with expertise in quantitative analysis.STEVEN SPAWLS is an expert in East African herpetology.MICHELE MENEGON researches the diversity and evolution of EastAfrican amphibians and reptiles. BREDA ZIMKUS is a herpetologist withexpertise in African frogs. RAFAEL DE SÁ is an amphibian biologistwith expertise in tadpole biology. ABEBE MENGISTU has expertise withEthiopian fish and frogs. FIKIRTE GEBRESENBET is a conservationbiologist working in her native Ethiopia. ROBIN MOORE is an amphibianconservation biologist. SAMY SABER is an expert on the ecology ofAfrican amphibians and reptiles. SIMON LOADER studies African(especially Afromontane) amphibian systematics and evolution.
Endemic frogs in the Bale Mountains 69
© 2013 Fauna & Flora International, Oryx, 47(1), 59–69

Long-term data for endemic frog genera revealpotential conservation crisis in the Bale Mountains,Ethiopia
DAV I D J . G OW E R , R O M A N K . A B E R R A , S I L V I A S C HWA L L E R , M A L C O L M J . L A R G E N
B E N C O L L E N , S T E P H E N S P AW L S , M I C H E L E M E N E G O N , B R E D A M . Z I M K U S
R A F A E L D E S Á , A B E B E A . M E N G I S T U , F I K I R T E G E B R E S E N B E T , R O B I N D . MO O R E
S A M Y A . S A B E R and S I M O N P . L O A D E R
Appendix I Species accounts
Spinophrynoides osgoodi (Loveridge, 1932)Bufo osgoodi Loveridge, 1932: Loveridge (1932: 47)Nectophrynoides osgoodi (Loveridge, 1932): Grandison(1978)Spinophrynoides osgoodi (Loveridge, 1932): Dubois(1987: 26)Altiphrynoides osgoodi (Loveridge, 1932): Frost et al.(2006: 363)
Spinophrynoides osgoodiwas described originally by Loveridge(1932) as a new species of the widespread genus Bufo, basedon material collected by W.H. Osgood in 1926–27. TandyandKeith (1972: 156) consideredBufo osgoodi to be amemberof the Bufo taitanus (5 Mertensophryne taitana of Frostet al., 2006) complex. Grandison (1978) included osgoodi inan expanded concept of the East African toad genusNectophrynoides, currently comprising 13 nominal species,all endemic to Tanzania (e.g. Frost, 2011). Dubois (1987)erected the monotypic genus Spinophrynoides forosgoodi based on it having a different breeding biology toNectophrynoides. Most recently, Frost et al. (2006) trans-ferred osgoodi toAltiphrynoides (see also below), but this wasnot based on any new data, and here we follow Largen’s(2001) taxonomy. The presence of this Ethiopian endemicgenus in the Bale Mountains has been overlooked by non-herpetologists and conservationmanagers because S. osgoodiwas not classified in Spinophrynoides until 1987 (e.g.WCMC,1993; Miehe & Miehe, 1994).
The original description of S. osgoodi was based on twospecimens collected in 1926–27 from an imprecise locality(‘Gedeb Mountains’, Loveridge, 1932), but it has sub-sequently been recorded from beyond the Bale Mountains,even including a specimen from the Gughe Mountains onthe western side of the Rift Valley (Largen, 2001: map 7). Inthe Bale Mountains, where the vast majority of specimenshave been recorded, S. osgoodi has been reported from bothnorth and south of the Sanetti Plateau, typically in montaneforest but extending also into open moorland at 1,950–3,520m (Largen, 2001). It is a small to medium (snout–ventlength, SVL, to 62mm) species whose large clutches of eggsin strings have been found laid in small pools in an openglade within Hypericum woodland at 3,200 m, these eggs
developing into free-swimming aquatic tadpoles thatmetamorphose into froglets (Grandison, 1978; Largen,2001).
In the 2004 and 2006 Global Amphibian Assessments(Stuart et al., 2008) and current Red List (IUCN, 2010)S. osgoodi was assessed as Vulnerable because its ‘Extentof Occurrence is less than 20,000 km2 and its Area ofOccupancy is less than 200 km2, its distribution is severelyfragmented, and there is continuing decline in the extentand quality of its forest habitat in the Ethiopian Mountains’.The population trend was stated to be decreasing althoughto the best of our knowledge there have been no quantitativeassessments of population size or density. Major threatsto the species were considered to be ‘environmental de-gradation resulting from human settlement and expansion’.With respect to necessary conservation measures, theGlobal Amphibian Assessments stated ‘the main priorityis prevention of further loss of primemontane forest habitat.The species is relatively common in the Bale MountainsNational Park, but there is a need to identify further sitessuitable for conservation’.
S. osgoodi is listed in CITES Appendix I becausethe species was formerly included within the genusNectophrynoides, all species of which are listed underAppendix I of CITES. However, there is ‘no knownutilization’ of S. osgoodi (IUCN, 2004).
Altiphrynoides malcolmi (Grandison, 1978)Nectophrynoides malcolmi Grandison, 1978Altiphrynoides malcolmi (Grandison, 1978): Dubois(1987: 27)
Altiphynoides malcolmi was originally described as anew species of the East African genus Nectophrynoidesby Grandison (1978). Dubois (1987) erected the genusAltiphrynoides for Grandison’s species on the basis that itsreproduction differs to that of Nectophrynoides (andSpinophrynoides). We follow Largen (2001) in consideringAltiphynoides to be a monotypic genus, despite Frostet al.’s (2006) inclusion of Spinophrynoides osgoodi inAltiphrynoides (see above). Because A. malcolmi has beenclassified in Altiphrynoides only since 1987 the presence ofthis endemic genus has been overlooked in some summariesof the fauna (e.g. WCMC, 1993; Miehe & Miehe, 1994).
© 2013 Fauna & Flora International, Oryx, 47(1), 1–5 doi:10.1017/S0030605311001426

The type series of . 160 A. malcolmi was collected byM.J. Largen and colleagues in December 1971, April 1972,April 1974 and April 1975 at 3,200–4,000 m from localitieson the northern and north-eastern (Dinsho/Dinshu andGoba) side of the Sanetti Plateau. An additional 20
specimens of A. malcolmi were collected in August 1986from the Harenna escarpment during the original HarennaForest Expedition (Largen, 2001). Overall, this species isthus reported from 3,200–4,000 m, spanning the trans-ition from the upper limits of the Schefflera–Hagenia–Hypericum forest to open Afroalpine moorland (Largen,2001).
This small species (SVL , 32 mm, Largen, 2001) wasobserved by Largen and colleagues forming large breedingaggregations in a forest glade in April 1975 (Grandison,1978). Following internal fertilization, small clutches (, 32)of large eggs are laid communally in moist places at the baseof herbaceous vegetation or under leaves/logs close tostreams but not in water. Tadpoles hatch from the eggs,but these are non-feeding and remain on land until meta-morphosis.
The Global Amphibian Assessments (IUCN, 2004, 2010;Stuart et al., 2008) categorized A. malcolmi as Endangered‘because its extent of occurrence is less than 5,000 km2 andits area of occupancy is less than 500 km2, all individualsare in fewer than five locations, and there is continuingdecline in the extent and quality of its habitat in the BaleMountains’. As with S. osgoodi, A. malcolmi is a CITESAppendix I species because it was formerly classified inNectophrynoides, even though there is ‘no known utilizationof this species’ (IUCN et al., 2006).
Ericabatrachus baleensis (Largen, 1991)
Ericabatrachus is a monotypic, possibly petropedetine/id(Largen, 2001) or phrynobatrachine/id (Dubois, 2005; Frostet al., 2006) frog endemic to the Bale Mountains. Dubois’(2005) and Frost et al.’s (2006) classifications are apparentlybased on Largen’s (1991: 141) description of the holotype ofEricabatrachus baleensis as ‘Habitus Phrynobatrachus-like’,although Largen clearly did not view E. baleensis as aphrynobatrachine/id. Scott (2005) provided morphologicalevidence that Ericabatrachus is more closely related to thepyxicephalid Cacosternum but its relationships are ob-viously not yet well understood. E. baleensis was describedfrom 23 specimens collected by M.J. Largen and D.W.Yalden in August during the 1986 Harenna ForestExpedition. Twenty-two of the types were collected fromthe margins of a stream in giant Erica forest in a single day(15 August) at the type locality for Balebreviceps hillmani(see below) at 3,200m, with one immature paratype comingfrom a stream at Katcha, at 2,400 m in the Harenna Forest.The original type locality collection data are very precise:daytime, in damp grass at edge of swiftest parts of mountaintorrent, into which they hopped when disturbed, surfacing
to cling to the bank at the water level (Largen, 1991). Thislocality has the name Tulla Negesso.
The reproduction and life history of E. baleensisare unknown but Largen (1991) interpreted the presenceof large ovarian eggs in some female paratypes ofE. baleensis as indicative of terrestrial clutches, potentiallywith direct development. No larvae have been reportedfor E. baleensis. The presence of E. baleensis and itsendemicity have been overlooked by at least some impor-tant works on the Bale Mountains (e.g., Miehe & Miehe,1994).
In the Global Amphibian Assessments (IUCN et al.,2004, 2006; Stuart et al., 2008), E. baleensis was categorizedas Endangered on the basis of an extent of occurrence of, 5,000 km2 and ‘continuing decline in the extent andquality of its habitat’. Accompanying notes state that aCritically Endangered assessment was narrowly avoidedbecause the species ‘almost certainly occurs more widely’.Ericabatrachus is not a CITES listed taxon. We know of nopublished records beyond the type series.
Balebreviceps hillmani (Largen & Drewes, 1989)
The monotypic Balebreviceps is a member of theBrevicipitinae/dae, a group of frogs endemic to sub-Saharan Africa. Its precise relationships to other brevicipi-tines/ds, most species of which occur in the Eastern ArcMountains of Kenya and Tanzania, remain unclear (e.g.Largen & Drewes, 1989; Loader et al., 2004). Balebrevicepshillmani was described originally from 16 individuals, allcollected by M.J. Largen and colleagues from a singlelocality during one day (15 August) of the Harenna ForestExpedition of 1986. To the best of our knowledge nosubsequent collections had been made, nor any reportspublished of this species being encountered at the type orany other locality (e.g. Largen & Spawls, 2010). All of thetype specimens were collected by day from beneath logs andboulders within giant Erica forest 50 m below the timberline on the Harenna escarpment (Largen & Drewes, 1989).The precise locality is immediately east of the road (betweenthe Sanetti Plateau and Rira), at Tulla Negesso, particularlyon the floor of the giant Erica forest between the road andthe part of the stream that is the type locality of E. baleensis.The presence of B. hillmani and its endemicity wereoverlooked by at least some important works on the BaleMountains (e.g., Miehe & Miehe, 1994).
The breeding biology and life history of B. hillmani areunknown but, based on close relatives, is predicted to haveterrestrial eggs with direct-development into froglets(Largen & Drewes, 1989; Müller et al., 2007). B. hillmani iscategorized as Endangered, narrowly missing CriticallyEndangered, for the same reasons noted above forEricabatrachus (IUCN et al., 2004, 2006; Stuart et al.,2008). Balebreviceps is not a CITES listed taxon.We know ofno published records beyond the type series.
2 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 1–5

Appendix 2 Details of the fieldwork undertaken tolocate amphibians in the Bale Mountains
For each record, the data are given in the following format:Date (as day/month/year): Locality: Length and timing ofsearch: (frogs found). Where one non-herpetologist foundfrogs by day while looking for other organisms, this isarbitrarily recorded here as c. two person hours. Expeditions10–14 also involved local assistants from Harenna (Issa,Hussein, Hussein and Mohammed).
Abbreviations: AAU, Addis Ababa University; BMNP, BaleMountains National Park; CI, Conservation International;EWCA, Ethiopian Wildlife Conservation Authority;FZS, Frankfurt Zoological Society; MCZ, Museum ofComparative Zoology, Harvard University; NHM, TheNatural History Museum, London; PH, person hours; TM,Trento Museum; UB, University of Basel; UR, University ofRichmond, Virginia; WWF, near the WWF hut, below Rira;Aa, Amieta angolensis; Am, Altiphrynoides malcomi; Ax,Afrixalus sp.; Bs, Bufo sp.; Bh, Balebreviceps hillmani;Eb, Ericabatrachus baleensis; Hs, Hyperolius sp.; Lg,Leptopelis gramineus; Lr, L. ragazzii and/or L. cf. ragazzii;Ls, Leptopelis sp.; Lss, Leptopelis spp.; Ofs-X, other (not thefour species of particular interest) unspecified frog species(where X is number of species); Pe, Ptychadena erlangeri;Phs, Phrynobatrachus sp.; Pk, Paracassina kounhiensis; Pn,Pychadena neumanni and P. cf. neumanni; Ps, Ptychadenasp.; Pss, Ptychadena spp.; So, Spinophrynoides osgoodi; UE,unidentified eggs; Xp, Xenopus sp.
M.J. Largen, D.W. Yalden, P.A. Morris and others
1. (December 1971)17/12/1971: Dinsho area: 6 PH day: (1 Lg, 1 Lr, 2 Am, 6Ofs-3).18/12/1971: Dinsho andMt Gaysay: 9 PH day: (1 So, 18Ofs-3).19/12/1971: Dinsho: 1 PH day: (1 So, 1 Lg, 2 Ofs-2). 21/12/1971:Urgana Valley and Little Batu (Sanetti Plateau): 2 PH day:(1 Am, 2 Ofs-1). 23/12/1971: Garba Guracha (Sanetti Plateau)and E of Dinsho: 2 PH day: (2 Am, 2Ofs-1). 24/12/1971: LittleBatu (Sanetti Plateau): 2 PH (1 Lg).
2. (April 1972)29/4/1972: Near Dinsho: 3 PH day (17 Lg, 1 Lr, 3Am, 2Ofs-1).
3. (April 1974)28/4/1974: S of Goba: 5 PH day (115 Am; 49 So, 1 Lr)
4. (April 1975)16/4/1975: Dinsho and S of Goba: 6 PH day (8 Lg, 4 Ofs-2).17/4/1975: S of Goba:8 PH day (75 Am, 1 Lg, 2 Ofs-1). 19/4/1975: S of Goba: 6 PHday and night (32 So).
5. Harenna expedition (August 1986)3/8/86: Katcha: 6 PH day and night (3 So, 2 Lg, 4 Lr, 9Ofs-2).4/8/1986: Katcha: 4 PH night (19 So, 3 Lr, 1 Lg, 1 Eb, 20 Ax,
4 Ofs-2). 7/8/1986: Katcha: 3 PH night (15 Lr, 9 Ofs-4).3/8/1986: Magano: 3 PH day (24 Lg, 5 Ofs-1). 8/8/1986:S of Katcha: 6 PH (3 So, 6 Lr, 1 Ofs-1). 9/8/1986: Shawebridge and Magano: 4 PH day (1 So, 11 Ofs-4). 15/8/1986:Tulla Negesso: 30 PH day (30 Am, 10 So, 18 Bh, 23 Eb,1 Lr, 3 Ofs-2). 19/8/1986: Magano: 9 PH day (16 Xp,17 Ofs-3).
Steve Spawls (SS) and others
6. SS, Jonathan and Tim Spawls (Dec 1994 to Jan 1995)30/12/1994: Saneti Plateau: 6 PH day. 31/12/1994: Dinsho:6 PH day. 1/1/1995: Harenna escarpment, close to road: 6 PHday (40 Am, 1 Bh, 6 or 7 So?, some Pk, Pn, Lg).
7. SS, Colin Tilbury (Sept 1996)21/9/1996: NE edge of Sanetti Plateau: 2 PH early morning(3 Am). 21/9/1996: Harenna escarpment: 20 PH daytime toearly evening (20 + Am, 1 Bh; some Lg). 22/9/1996: Dinsho:6 PH (Bs).
8. SS, Jonathan and Tim Spawls (Dec 1996)29/12/1996: Sanetti Plateau: 6 PH day. 30/12/1996: Dinsho:6 PH day. 31/12/1996: Harenna escarpment: 6 PH day(18 + Am, 1 Bh).
9. SS, Jonathan and Tim Spawls, Chris Smith (Oct 1999)26/10/1999: Sanetti Plateau: 8 PH day. 27/10/1999: Dinsho:8 PH day. 28/10/1999: near Rira: 8 PH day (Ps).
10. MCZ (July 2006). Breda Zimkus and Ryan Kerney(MCZ); Daniel Pawlos (EWCA)
1/7/2006: 3,055 m, Gaysay, 2 km west of Dinsho, and nearBMNP lodge: 3 PH day pm (Pc, UE, T). 2/7/2006: 2801 m,South of Rira: 2 PH day pm (Pn): 2 PH night. 3/7/2006:2,816 m, Kolala Lake, nr. Rira: 2 PH day am (Lr, Ls, Ps, T):2,378 m, Katcha Camp:4 PH day pm (Pn, T). 4/7/2006: 3,400–4,000 m HarennaEscarpment and S side of Sanetti Plateau: 4 PH day am:Tulla Negesso: 4 PH am/pm: 2,810m, Telila Stream, nr. Rira:2 PH pm (Lr, T). 5/7/2006: 3,800–4,100 + m, Sanetti Plateau:3 PH day am (Ls, T). 13/7/2006: 1,750 m, 18 km south ofKatcha Camp: 4 PH day am (Pe). 14/7/2006: 2,300–2,400m,north of Katcha:5 PH am/pm (Pe, Pn, Lg, Lr, T). 15/7/2006: 2,700–2,800 m,Gussa River, nr. Rira: 3 PH day am (Ls).
11. NHM (September 2006). David Gower and SimonLoader (NHM), Roman Aberra (EWCA), Daniel Tilaye(BMNP)
15/9/2006: Dinsho: 3 PH day pm: 3 PH night (Lg, T).16–19/9/2006: Dinsho: locals and park staff casualdaytime searching (Pss, Lg). 16/9/2006: 3,900 m on Nside of Sanetti Plateau: 6 PH day am (1 Am): 4,000 + m,Sanetti Plateau: 1.5 PH day am: Tulla Negesso: 6 PH
Endemic frogs in the Bale Mountains 3
© 2013 Fauna & Flora International, Oryx, 47(1), 1–5

day pm: (Lr): Katcha: 3 PH night (Lr, Ls). 17/9/2006: Katcha:1 PH day am (Ps): 2,400–2,500 m, above Katcha: 5 PH dayam: 2900–3200 m, W of Rira: 10 PH day pm: 2,750 m, Westof Rira: 5 PH day pm: 2,815 m, nr. Rira: locals day (3 Bh,3 Am). 18/9/2006: 1,900 m, Harenna Forest: 1 PH : TullaNegesso: 6 PH day pm: 1,900m, Shawe River: locals day pm(Ls): 2,800m, nr. Rira: locals day pm (Ps. Ls): Tulla Negesso:overnight four terrestrial funnel traps on ground: TullaNegesso: 3 PH night (1 Bh). 19/9/2006: Tulla Negesso: 1.5 PHday am: 3,060 m, Gaysay: 3.5 PH (Ps).
12. UB, EWCA and AAU (July–August 2008). SimonLoader, Abebe Mengistu, Silvia Schwaller, Michael Geiser(UB), Roman Aberra (AAU, EWCA), Samy Saber, FikirteGebresenbet (AAU)
14/7/2008: Dinsho: 2 PH daytime (1 Lg). 15/7/2008: TullaNegesso: 4 PH daytime (1 Am): WWF: 4PH night (1 Lg,1 Lr, T). 16/7/2008: Tulla Negesso: 6PH daytime:WWF: 9PHdaytime (6 Lr, 5 Lg, T). 18/7/2008: 3,200 m, Fute: 3 PHdaytime: 3,145 m, nr. Fute: 5 PH daytime (1 Bh). 19/7/2008:3,255 m, Kotinsa: 1.5 PH daytime: 3,200 m nr. Kotinsa:0.5 PH daytime. 20/7/2008: 3,200 m Kotinsa: 3 PH dayime(UE). 21/7/2008: 3,200 m, nr. Fute: 3 PH daytime (3 Am).22/7/2008: 3,950m, Sanetti Plateau: 1 PH daytime. 23/7/2008:3,950 m, Sanetti Plateau: 2 PH daytime. 26/7/2008: 3,200 m,Kotinsa: 4 PH night. 27/7/2008: 3,200 m, Degebato: 3 PHday + 1 PH night. 28/7/2008: 3,200 m, Degebato: 3PH day.29/7/2008: Rira: locals, day (1 Lg). 30/7/2008: 2,370 m,Katcha: 8PH day (3 Lg, 1 Lr, T): 2,800 m, nr. WWF: night(2 Lr, T). 31/7/2008: 3,300m, Galema: 1 PH night: Rira: localsday (Ps). 1/8/2008: 3,301m,Galema: 3 PH day (1 Lg, 1 Ps). 2/8/2008: 3,200 m, Tulla Negesso: 3 PH day: 3,200 m, nr. Fute:2 PH day (1 Bh, 1 Eb): Rira: locals day (1 Lg, 1 Ps). 4/8/2008:3,200 m, Kotinsa: 3PH day: Rira: locals day (5 Ps, 1 Lg).5/8/2008: 3,200mKotinsa: 6.5 PH day: 3,200m, Fute: 3.5 PHday (1Am): Fute: 4 PH day (1 Bh). 29/7–5/8/2008: Rira: localsday (4 Bh, 8 Lr, 1 Lg). 7/8/2008: 3,160 m, Dinsho: park staffday (2 Phs). 27–28/8/2008: Weyib Valley, Sanetti Plateau:6PH day.
13. AAU and EWCA (February–March 2009). RomanAberra (AAU, EWCA)
22/2/2009: 2,780m, WWF: 1.5 PH day (10 Ps, 1 Lg): 2,850m,nr. WWF: 1.5 PH day (2 Ps). 23/2/2009: Tulla Negesso: 3 PHday (1 Am, 1 Bh): 2,790 m, nr. WWF: 1.5 PH pm (1 Ps, 1 Lg).24/2/2009: 3,200 m, Fute: 1.5 PH day: 2,800 m, nr. WWF:locals (Ps): nr. Rira: 0.5 PH day (5 Ps, T): 2,900 m, nr. Rira:1.5 PH night (4 Ps, 1 Ls). 25/2/2009: 3,150 m, Fute: 3 PH, day(5 Am). 26/2/2009: 3,250 m, Kotinsa: 6 PH day (1 Bh). 27/2/2009: 3,300 m, Galema: 3 PH day (2 Bh, 5 Lg, T). 28/2/2009:Tulla Negesso: 1.5 PH day (2 Am): 2,900 m, nr. Rira: 2 PHday (10 s Lg, 4 Ps). 1/3/2009: 2.800 m, nr. WWF: localsday (7 Lg, 4 Ps). 2/3/2009: 3,300 m, Degebato: 6 PH day.
3/3/2009: 2,350 m, Katcha: 5 PH day (10 Lr, 9 Ps, 2 Lg).4/3/2009: 2,900 m, nr. Rira: 1.5 PH day: 2,800 m, nr. Rira:0.5 PH day (50 T): 2,800 m, nr. Rira: 1 PH day (1 Bh).
14. UB, AAU, TM, UR and NHM (June 2009). SimonLoader, Abebe Mengistu (UB), Michele Menegon (TM),Rafael de Sá (UR), David Gower (NHM), Bruhtesfa Yimer(EWCA)
18/6/2009: 2,750m, nr. WWF: 3.75 PH night (10 s Lr, 10 s T).19/6/2009: c. 4,000 m, Sanetti Plateau: 3 PH day: TullaNegesso: 5.5 PH day: (1 Bh): 2,380 m, Katcha: 8 PH dusk/night (Ps, Lr, Ax, 100s T). 20/6/2009: 3,100m, nr. Fute: 10 PHday (10 Bh, 2 Eb): 3,000–2,750 m, Fute to Rira: 5 PH day(1 Am): nr. WWF: 1.25 PH (T): 1,890 m, Shawe river: 4 PHnight (Lr, Ps, Xp, 10s T). 21/6/2009: 3,100 m, Fute: 9 PH day(4 Bh, 5 Am): 3,100 m, Fute: 5 PH night: c. 1,600 m, DolaMena to Rira: 3 PH day: 1,900 m, Magano: 1.5 PH day(Ps, Xp, Pk): 1,900m,Magano: 3 PH night (Xp, Pk, Ps). 22/6/2009: 3,100 m, Fute: 1 PH day: 1,890 m, Shawe river: 1 PHday: 1,900 m, Magano: 4 PH day (Xp, Ps, Phs): 1,900 m,Magano: 2.5 PH night (Xp, Ps, Pk, Hs). 25–26/6/2009:c. 4,000m, nr. Garba Guracha: 10 PH: 26/6/2009: c. 4,000m,Sanetti Plateau: 2 PH (Lg T).
15. CI and FZS (September 2009) Robin Moore (CI),Thadaigh Bagallay (FZS)
6/9/2009: Tella Negesso: 6PH day (1 Am): 2,380 Katcha:6PH dusk (Phs, Ls). 7/9/2009: 3,100, nr. Rira: 8PH night:Fute: 7 PH dusk (1 Am). 8/9/2009: nr. Dola Mena: 3PH day(Phs, Aa).
Appendix 3 Statistical analysis
The approach used to infer time of extinction is based onthe Weibull distribution, a two-parameter model. Its originis in weakest link analysis (Crawley, 2007); its use hastraditionally been in engineering industrial risk analysis (forexample, the failure of a component). The attraction for ananalysis aimed at inferring extinction comes from the resultthat, provided the sample of sightings data are large enough,it is reasonable to assume that the k most recent sightingscome from the same Weibull extreme value distribution,regardless of the parent distribution (Solow, 2005; see alsoCollen et al., 2010). We implemented the following fromSolow (2005). The optimal linear estimate of time ofextinction, T̂E based on the k most recent sightings has theform
T̂E =∑ki=1
witn−i+1 (1)
The weight vector w 5 (w1, w2, . . . , wk)′ is given by
w = (e′Λ−1e)−1Λ−1e (2)
4 D. J. Gower et al.
© 2013 Fauna & Flora International, Oryx, 47(1), 1–5

where e is a vector of k 1s and Λ is the symmetric k-by-kmatrix with typical element
Λij = Γ(2v̂ + i)Γ(v̂ + j)Γ(v̂ + i)Γ( j) j ≤ i (3)
where Γ is the gamma function and
v̂ = 1k− 1
∑k−2
i=1
logtn − tn−k+1
tn − ti+1(4)
is an estimate of the shape parameter of theWeibull extremevalue distribution, where tn is the n times a species is sightedduring an observation period t.
Last, under the assumption that the species is extinct,the upper bound of an approximate 1 − α confidence interval
for T̂E is
TuE = tn − c(α)tn−k+1
1− c(α) (5)
where
c(α) = k−log α
( )−v̂
(6)
We looked at two levels of certainty: 95 and 80%. Variationsin the sighting rate reflect variations in both ‘sightability’,which may be connected to abundance, and sighting effort.The main assumption of the technique is that althoughsighting effort may vary, it never falls to zero over an annualtime step, particularly around the time of extinction (Solow,2005).
Endemic frogs in the Bale Mountains 5
© 2013 Fauna & Flora International, Oryx, 47(1), 1–5





























