Embed Size (px)
Citation preview

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolo
Macroalgal-mediated transfers of water column nitrogen to
intertidal sediments and salt marsh plants
Katharyn E. BoyerT, Peggy Fong
Department of Organismic Biology, Ecology and Evolution, 621 Charles E. Young Drive, South, University of California, Los Angeles,
Los Angeles, California 90095-1606, USA
Received 28 May 2004; received in revised form 1 December 2004; accepted 5 January 2005
Abstract
In many temperate estuaries, mats of opportunistic macroalgae accumulate on intertidal flats and in lower elevations of salt
marshes, perhaps playing a role in linking water column nitrogen (N) supply to these benthic habitats. Using a flow-through
seawater system and tidal simulator, we varied densities (equivalent to 0, 1, 2, or 3 kg m�2 wet mass) of 15N-labelled
macroalgae (Enteromorpha intestinalis) on estuarine sediments in microcosms with/without pickleweed (Salicornia virginica)
to assess N transfers from algae. In the 6-week experiment, macroalgal biomass increased from initial levels in the lower density
treatments but all algae lost N mass, probably through both leakage and decomposition. With all densities of algae added,
sediments and pickleweed became enriched in 15N. With increasing mat density, losses of algal N mass increased, resulting in
stepwise increases in 15N labeling of the deeper sediments and pickleweed. While we did not detect a growth response in
pickleweed with macroalgal addition during the experiment, N losses from algal mats that persist over many months and/or
recur each year could be important to the mineral nutrition of N-limited marsh plants. We conclude that N dynamics of intertidal
sediments and lower salt marsh vegetation are linked to the N pools of co-occurring macroalgae and that further study is needed
to assess the magnitude and importance of N transfers.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Enteromorpha; N isotopes; N limitation; Salicornia; Salt marsh
0022-0981/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.jembe.2005.01.005
T Corresponding author. Current address: Romberg Tiburon
Center for Environmental Studies and Department of Biology, San
Francisco State University, 3152 Paradise Drive, Tiburon, California
94920, USA. Tel.: +1 415 338 3751; fax: +1 510 558 0167.
E-mail address: [email protected] (K.E. Boyer).
1. Introduction
Many studies have documented algal-mediated
linkages between nutrient dynamics of the water
column and sediments in the subtidal zone of
estuaries. For example, phytoplankton and macro-
algae suspended in the water column influence
benthic nutrient processes through interception of
gy and Ecology 321 (2005) 59–69

K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–6960
light and water column nutrients (Fong et al., 1993;
McGlathery et al., 1997; Hauxwell et al., 2001).
Macroalgae can also intercept nutrients fluxing from
the sediments (Bierzychudek et al., 1993) and
promote such fluxes by depleting water column
nutrients (Thybo-Christesen et al., 1993; Fong and
Zedler, 2000). In addition, macroalgae have been
found to release nutrients and organic matter to
underlying sediments when decomposing (Hansen
and Kristensen, 1997; Tyler et al., 2001), fueling
remineralization, and nutrient fluxes back to the water
column (Lavery and McComb, 1991; Hansen and
Blackburn, 1992; Enoksson, 1993).
While many of the same processes are at work
in the intertidal zone of estuaries, alternating
immersion and emersion of the sediment surface
may uniquely influence the relationship between
algae and benthic/pelagic nutrient exchanges. For
example, benthic microalgae at the surface of
intertidal flats access both the water column and
sediment nutrient supply when submerged, but
direct deposition of N from the air during exposure
can be several orders of magnitude higher (Paerl,
1997). Further, macroalgae in the intertidal become
desiccated when exposed at low tide, resulting in
reduction of photosynthesis, light attenuation within
the compressed algal mat, and inorganic carbon
limitation (Pregnall, 1983; Pregnall and Rudy,
1985). These stresses are likely to influence
nutrient dynamics both within and outside of the
algal mat.
Growth and accumulation of macroalgae on
intertidal flats are common in many estuaries through-
out the world, with percent cover up to 100%,
especially where anthropogenic N supply is pro-
nounced (e.g., Pregnall and Rudy, 1985; Sfriso et
al., 1992; Hernandez et al., 1997; Kamer et al., 2001;
Kennison et al., 2003). Opportunistic macroalgae,
typically species in the green (Division Chlorophyta)
genera Ulva, Enteromorpha, Cladophora, and Chae-
tomorpha, have rapid nutrient uptake, high growth
rates, and the capacity to store bexcessQ nutrients for
future growth (Rosenberg and Ramus, 1984; Fujita,
1985; Fong et al., 1994; Peckol et al., 1994). Such
bloom-forming macroalgae represent a large pool of
nutrients potentially available for release, and perhaps
more prone to be released in the stressful intertidal
zone.
Another distinguishing feature of the intertidal
zone of estuaries is the presence of emergent vascular
plants. In many locations, macroalgae can be found
growing or deposited among the bases of the stems of
these marsh plants at their lower elevations. One or
more species of macroalgae can be observed among
culms of smooth cordgrass (Spartina alterniflora
Loisel) in many areas of the US, including the coastal
bays of Maryland and Delaware (J. Gallagher, pers.
comm.), Jamaica Bay, New York (D. Franz, pers.
comm.), Bogue Sound, North Carolina (K. Boyer,
pers. obs.), and Waquoit Bay, Massachusetts (P.
Peckol, pers. comm.). In southern California, the
major bloom-forming genera Enteromorpha and Ulva
are found among Salicornia virginica L. (pickle-
weed), Salicornia bigelovii Torr. (annual pickleweed),
Spartina foliosa Trin. (Pacific cordgrass), Batis
maritima L. (saltwort), and Distichlis spicata L.
(saltgrass) (K. Boyer, pers. obs., H. Page, pers.
comm., Zedler, 1980).
While such associations may be fairly common in
the lower marsh, we know of only one study that has
explicitly examined the effects of macroalgae occur-
ring on sediments among salt marsh plants. Gerard
(1999) found lower biomass of S. alterniflora in plots
where the brown alga Ascophyllum nodosum Scorpi-
odes was removed experimentally; algal nutrient
releases to sediments upon decomposition were
suggested, but not assessed, as the mechanism for
enhancement of cordgrass growth.
If macroalgae release substantial quantities of
nutrients in the intertidal zone, then enhancement of
the nutrient supply to intertidal sediments and the
plants rooted therein is plausible. In view of the fact
that lower marsh plants are typically N limited (e.g.,
Broome et al., 1975; Boyer et al., 2001), access to
water column N formerly sequestered in algal tissues
could be important to marsh plant mineral nutrition
where macroalgae co-occur. However, flooding tides
may wash algal-derived nutrients out of the intertidal
zone, limiting their accumulation in sediments and
their uptake by associated vascular plants. Further,
algal mats in the intertidal may initiate nutrient fluxes
from sediments as can occur subtidally (e.g., Lavery
and McComb, 1991), reducing sediment nutrient
pools. Even if N is being supplied to the sediments
by algal mats, if algal decomposition depletes oxygen
and leads to accumulation of hydrogen sulfide in

K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–69 61
sediments, marsh plant productivity may be reduced
(Koch et al., 1990).
We are particularly interested in the fate of macro-
algal N in tidal flats and marshes dominated by coarse
sediments. Where such habitats are created for
restoration purposes, sediments are often sandy due
to terrestrial origin or as a product of dredging (Langis
et al., 1991; Craft et al., 1999). Coarse-grained
sediments can also be found in marshes and tidal
flats in geologically young systems such as those that
form along barrier islands (Tyler and Zieman, 1998),
in areas of estuaries where storms episodically deposit
sandy terrestrial or dune/beach-derived sediments
(Osgood and Zieman, 1993, 1995), or in areas where
there are higher velocity flows (e.g., Bolam et al.,
2000). Sandy sediments lack the nutrient adsorption
capacity of clay particles, and store and supply N
poorly (Brady and Weil, 1999). Therefore, processes
that supply N from external sources, e.g., advective
flux of enriched tidal water through sediments
(Osgood and Zieman, 1998), can be especially
important to nutrient dynamics of coarse sediment
systems. Macroalgal mats on sandy tidal flats could
similarly act as an external N source.
The objective of our study was to investigate the
potential role of macroalgal mats in influencing N
supply to intertidal sand flats and salt marsh habitats.
We hypothesized that algal mat density influences
algal N transfers, with thicker mats declining more in
underlying layers and releasing a greater pool of
nutrients than thinner mats. We used Enteromorpha
intestinalis Link isotopically-enriched in N to assess
N transfers to sediments and S. virginica tissues.
2. Methods
We performed a 4�2 factorial experiment varying
E. intestinalis (hereafter called macroalgae) mat
density (0, 1, 2, or 3 kg macroalgae m�2, see below)
and presence/absence of S. virginica (hereafter called
pickleweed) in pots subjected to simulated tides (Fig.
1). In April 2000, we collected sediment and
pickleweed from a sandy restoration marsh at Mugu
Lagoon in Ventura County, California. Sediments
were collected from an unvegetated slope at a point
just below the highest tidal elevation. Lower on the
slope, a trowel was used to collect pickleweed
recruits and soil surrounding their roots (~4 cm
diameter, 4 cm deep).
Sediments were added to nursery pots and half were
planted with pickleweed. Five extra pots were planted
for later analysis of initial conditions. To acclimate
plants, pots were set into pans of low nutrient (b3.57
AM NO3, b1.61 AM PO4) seawater in a greenhouse at
the University of CA Los Angeles for three weeks; de-
ionized water was added to counter evaporation.
Macroalgae were also collected at Mugu Lagoon,
cleaned of visible fauna and debris, and cultured in
low nutrient seawater to equalize internal nutrient
stores. After 1 week, 15N-labelled NaNO3 was added
to produce a 200 AM culture solution, to simulate
moderately enriched conditions common to southern
CA estuarine waters (concentrations can exceed 800
AM in upper Newport Bay (Boyle et al., 2004) and
3000 AM at Carpinteria marsh, near Santa Barbara
(Kennison et al., 2003)). Two more equivalent nitrate
additions were made over the next 2 weeks, along
with deionized water to counter evaporation.
The 6-week experiment was conducted outdoors
at the Redondo Beach Generating Station in
Redondo Beach, CA, from early May to mid-June
2000. Algal tissue was spun for 1 min in a lettuce
spinner, then divided into 20, 40, or 60 g portions
(equivalent to 1, 2, or 3 kg algae m�2) to reflect the
range in densities typical of estuaries in southern
CA and elsewhere where algal blooms occur (e.g.,
Pregnall and Rudy, 1985; Hernandez et al., 1997;
Kamer et al., 2001; Tyler et al., 2001). Algal tissues
were spread evenly on the sediment surface and the
pots were placed in a random array on a table. With
5-fold replication, there were a total of 40 pots, each
independently plumbed with seawater using drip
irrigation tubing and emitters (Fig. 1B). A swim-
ming pool timer controlled a pump to simulate
twice-daily tides of equal length. Pots were
scrubbed weekly to reduce algal/bacterial films,
then re-randomized.
At the start of the experiment, water samples were
collected from the seawater system, filtered through
Whatman GF/C filters, and frozen for later analysis of
total Kjeldahl N (TKN), NH4–N, NO3–N, and total P
at the University of CA, Davis Division of Agriculture
and Natural Resources (DANR) laboratory using
standard methods. As the TKN method does not
digest N from oxidized forms (e.g., nitrate and nitrite),

Fig. 1. (A) Schematic of experimental design with four levels of algal addition (0–3 kg m�2) and presence/absence of pickleweed in pots of
sediments. (B) Cross-sectional view; water entered and exited the outer pots through irrigation tubing and emitters, and the inner pot through
holes (covered with mesh to retain sediments) in the bottom. Wooden blocks elevated the shorter inner pots such that water did not reach their
top edges, and so that the water level dropped below the inner pots at simulated low tide. Pump timing permitted approximately equal periods of
submergence and exposure of the sediment surface.
K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–6962
dissolved organic N was calculated by subtraction of
NH4–N from TKN. Water nutrients (Table 1) were
extremely low compared to levels in southern CA
estuaries (Fong, 1986; Fong and Zedler, 2000; Boyle
et al., 2004; Kennison et al., 2003).
Initial sediment subsamples were dried (55 8C),ground, and sieved (1-mm sieve), and grain size
distribution, organic matter, and total N and P
concentration were determined at DANR (Table 1).
The high proportion of sand particles was comparable
to that of other marshes formed naturally on sandy
deposits or newly constructed from dredged material
(Osgood and Zieman, 1993, Boyer and Fong,
unpublished data). There had been little accumulation
of organic matter in these sediments (Table 1;
compare to 0.8–3% in sandy marshes N10 years old,
Osgood and Zieman, 1993; Craft et al., 1999). As
expected for coarse sediments, N concentration was at
the low end of the range for low elevation marshes
(often greater than 0.2% dry mass and as high at
0.8%; reviewed in Boyer et al., 2001), but similar to
other constructed and natural coarse grained sites
(Boyer and Zedler, 1998; Craft et al., 1999). Total P
was in the middle of the range found in lower
elevation marshes (0.01–0.14% dry mass: Mendels-
sohn and Marcellus, 1976; Ellison et al., 1986;
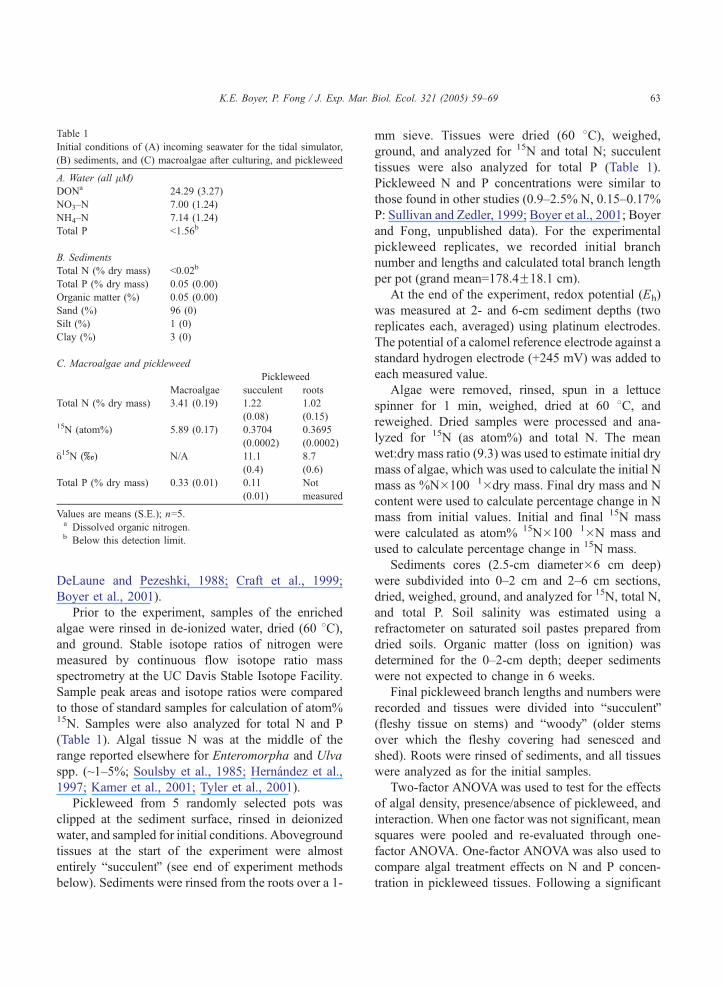
Table 1
Initial conditions of (A) incoming seawater for the tidal simulator,
(B) sediments, and (C) macroalgae after culturing, and pickleweed
A. Water (all lM)
DONa 24.29 (3.27)
NO3–N 7.00 (1.24)
NH4–N 7.14 (1.24)
Total P b1.56b
B. Sediments
Total N (% dry mass) b0.02b
Total P (% dry mass) 0.05 (0.00)
Organic matter (%) 0.05 (0.00)
Sand (%) 96 (0)
Silt (%) 1 (0)
Clay (%) 3 (0)
C. Macroalgae and pickleweed
Pickleweed
Macroalgae succulent roots
Total N (% dry mass) 3.41 (0.19) 1.22
(0.08)
1.02
(0.15)15N (atom%) 5.89 (0.17) 0.3704
(0.0002)
0.3695
(0.0002)
y15N (x) N/A 11.1
(0.4)
8.7
(0.6)
Total P (% dry mass) 0.33 (0.01) 0.11
(0.01)
Not
measured
Values are means (S.E.); n=5.a Dissolved organic nitrogen.b Below this detection limit.
K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–69 63
DeLaune and Pezeshki, 1988; Craft et al., 1999;
Boyer et al., 2001).
Prior to the experiment, samples of the enriched
algae were rinsed in de-ionized water, dried (60 8C),and ground. Stable isotope ratios of nitrogen were
measured by continuous flow isotope ratio mass
spectrometry at the UC Davis Stable Isotope Facility.
Sample peak areas and isotope ratios were compared
to those of standard samples for calculation of atom%15N. Samples were also analyzed for total N and P
(Table 1). Algal tissue N was at the middle of the
range reported elsewhere for Enteromorpha and Ulva
spp. (~1–5%; Soulsby et al., 1985; Hernandez et al.,
1997; Kamer et al., 2001; Tyler et al., 2001).
Pickleweed from 5 randomly selected pots was
clipped at the sediment surface, rinsed in deionized
water, and sampled for initial conditions. Aboveground
tissues at the start of the experiment were almost
entirely bsucculentQ (see end of experiment methods
below). Sediments were rinsed from the roots over a 1-
mm sieve. Tissues were dried (60 8C), weighed,
ground, and analyzed for 15N and total N; succulent
tissues were also analyzed for total P (Table 1).
Pickleweed N and P concentrations were similar to
those found in other studies (0.9–2.5% N, 0.15–0.17%
P: Sullivan and Zedler, 1999; Boyer et al., 2001; Boyer
and Fong, unpublished data). For the experimental
pickleweed replicates, we recorded initial branch
number and lengths and calculated total branch length
per pot (grand mean=178.4F18.1 cm).
At the end of the experiment, redox potential (Eh)
was measured at 2- and 6-cm sediment depths (two
replicates each, averaged) using platinum electrodes.
The potential of a calomel reference electrode against a
standard hydrogen electrode (+245 mV) was added to
each measured value.
Algae were removed, rinsed, spun in a lettuce
spinner for 1 min, weighed, dried at 60 8C, and
reweighed. Dried samples were processed and ana-
lyzed for 15N (as atom%) and total N. The mean
wet:dry mass ratio (9.3) was used to estimate initial dry
mass of algae, which was used to calculate the initial N
mass as %N�100�1�dry mass. Final dry mass and N
content were used to calculate percentage change in N
mass from initial values. Initial and final 15N mass
were calculated as atom% 15N�100�1�N mass and
used to calculate percentage change in 15N mass.
Sediments cores (2.5-cm diameter�6 cm deep)
were subdivided into 0–2 cm and 2–6 cm sections,
dried, weighed, ground, and analyzed for 15N, total N,
and total P. Soil salinity was estimated using a
refractometer on saturated soil pastes prepared from
dried soils. Organic matter (loss on ignition) was
determined for the 0–2-cm depth; deeper sediments
were not expected to change in 6 weeks.
Final pickleweed branch lengths and numbers were
recorded and tissues were divided into bsucculentQ(fleshy tissue on stems) and bwoodyQ (older stems
over which the fleshy covering had senesced and
shed). Roots were rinsed of sediments, and all tissues
were analyzed as for the initial samples.
Two-factor ANOVAwas used to test for the effects
of algal density, presence/absence of pickleweed, and
interaction. When one factor was not significant, mean
squares were pooled and re-evaluated through one-
factor ANOVA. One-factor ANOVA was also used to
compare algal treatment effects on N and P concen-
tration in pickleweed tissues. Following a significant
0.5
1
1.5
2
2.5
3A
conce
ntr
atio
n (
atom
%)
p = 0.0001
a
b
bb
K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–6964
one-factor ANOVA, multiple comparisons were made
using Fisher’s PLSD method. Data not meeting
assumptions of ANOVA after transformation were
analyzed using the non-parametric Kruskal–Wallis (K–
W) test. When one factor was not significant, mean
squares were pooled for analysis by K–W or Mann–
Whitney U tests. Non-parametric multiple compari-
sons by simultaneous test procedure followed signifi-
cant K–W tests. All errors presented are F1 S.E.
00
0.5
1
1.5
sedim
ent
15N
0 1 2 3
kg algae m-2
B
p = 0.0001
ba
c
c
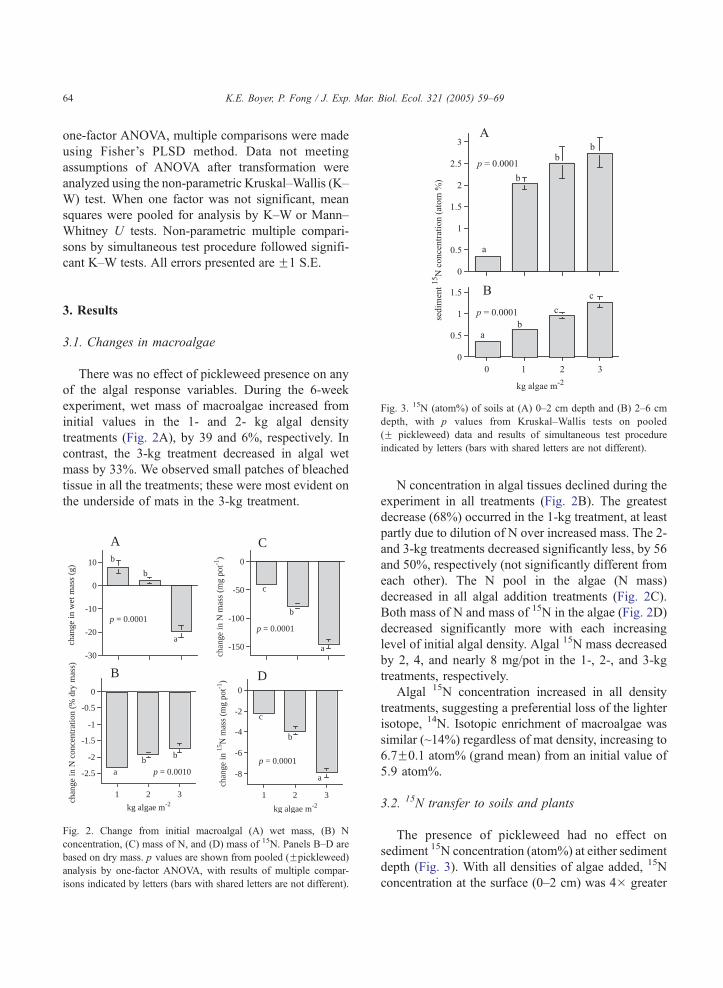
Fig. 3. 15N (atom%) of soils at (A) 0–2 cm depth and (B) 2–6 cm
depth, with p values from Kruskal–Wallis tests on pooled
(F pickleweed) data and results of simultaneous test procedure
indicated by letters (bars with shared letters are not different).
3. Results
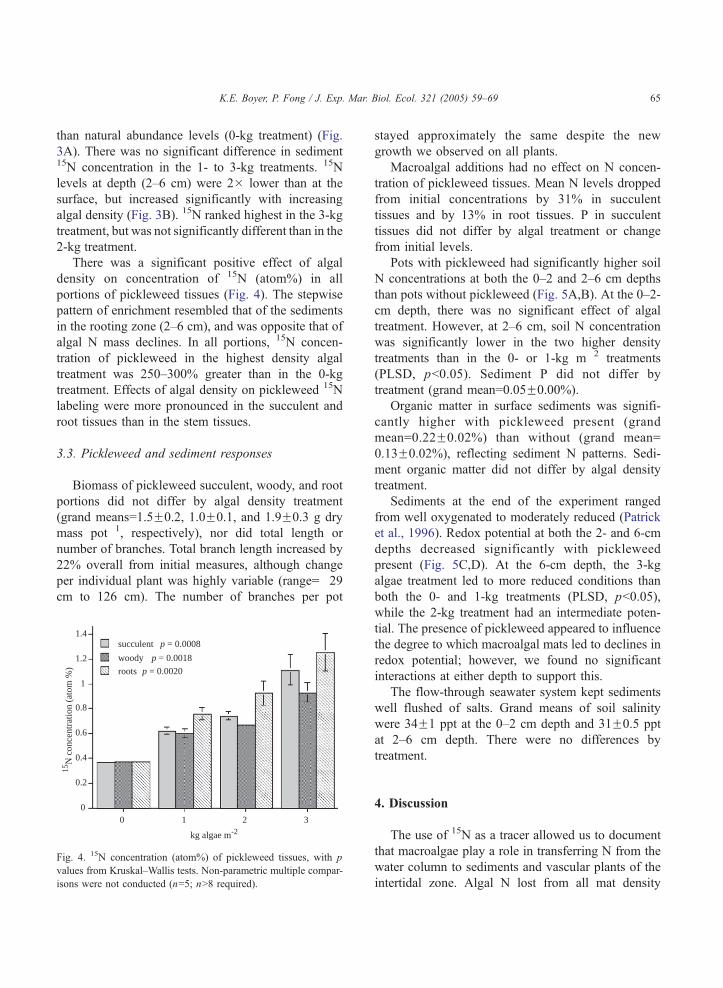
3.1. Changes in macroalgae
There was no effect of pickleweed presence on any
of the algal response variables. During the 6-week
experiment, wet mass of macroalgae increased from
initial values in the 1- and 2- kg algal density
treatments (Fig. 2A), by 39 and 6%, respectively. In
contrast, the 3-kg treatment decreased in algal wet
mass by 33%. We observed small patches of bleached
tissue in all the treatments; these were most evident on
the underside of mats in the 3-kg treatment.
A
-8
-6
-4
-2
0
chan
ge in
15N
mas
s (m
g po
t-1)
1 2 3
kg algae m-2
D
a
b
c
p = 0.0001
-150
-100
-50
0
chan
ge in
N m
ass
(mg
pot-1
)
C
p = 0.0001
a
b
c
-30
-20
-10
0
10
chan
ge in
wet
mas
s (g
)
b
b
a
p = 0.0001
B
-2.5
-2
-1.5
-1
-0.5
0
chan
ge in
N c
once
ntra
tion
(% d
ry m
ass)
1 2 3
kg algae m-2
p = 0.0010ab
b
Fig. 2. Change from initial macroalgal (A) wet mass, (B) N
concentration, (C) mass of N, and (D) mass of 15N. Panels B–D are
based on dry mass. p values are shown from pooled (Fpickleweed)
analysis by one-factor ANOVA, with results of multiple compar-
isons indicated by letters (bars with shared letters are not different).
N concentration in algal tissues declined during the
experiment in all treatments (Fig. 2B). The greatest
decrease (68%) occurred in the 1-kg treatment, at least
partly due to dilution of N over increased mass. The 2-
and 3-kg treatments decreased significantly less, by 56
and 50%, respectively (not significantly different from
each other). The N pool in the algae (N mass)
decreased in all algal addition treatments (Fig. 2C).
Both mass of N and mass of 15N in the algae (Fig. 2D)
decreased significantly more with each increasing
level of initial algal density. Algal 15N mass decreased
by 2, 4, and nearly 8 mg/pot in the 1-, 2-, and 3-kg
treatments, respectively.
Algal 15N concentration increased in all density
treatments, suggesting a preferential loss of the lighter
isotope, 14N. Isotopic enrichment of macroalgae was
similar (~14%) regardless of mat density, increasing to
6.7F0.1 atom% (grand mean) from an initial value of
5.9 atom%.
3.2. 15N transfer to soils and plants
The presence of pickleweed had no effect on
sediment 15N concentration (atom%) at either sediment
depth (Fig. 3). With all densities of algae added, 15N
concentration at the surface (0–2 cm) was 4� greater
K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–69 65
than natural abundance levels (0-kg treatment) (Fig.
3A). There was no significant difference in sediment15N concentration in the 1- to 3-kg treatments. 15N
levels at depth (2–6 cm) were 2� lower than at the
surface, but increased significantly with increasing
algal density (Fig. 3B). 15N ranked highest in the 3-kg
treatment, but was not significantly different than in the
2-kg treatment.
There was a significant positive effect of algal
density on concentration of 15N (atom%) in all
portions of pickleweed tissues (Fig. 4). The stepwise
pattern of enrichment resembled that of the sediments
in the rooting zone (2–6 cm), and was opposite that of
algal N mass declines. In all portions, 15N concen-
tration of pickleweed in the highest density algal
treatment was 250–300% greater than in the 0-kg
treatment. Effects of algal density on pickleweed 15N
labeling were more pronounced in the succulent and
root tissues than in the stem tissues.
3.3. Pickleweed and sediment responses
Biomass of pickleweed succulent, woody, and root
portions did not differ by algal density treatment
(grand means=1.5F0.2, 1.0F0.1, and 1.9F0.3 g dry
mass pot�1, respectively), nor did total length or
number of branches. Total branch length increased by
22% overall from initial measures, although change
per individual plant was highly variable (range=�29
cm to 126 cm). The number of branches per pot
0
0.2
0.4
0.6
0.8
1
1.2
1.4
15N
con
cent
ratio
n (a
tom
%)
210 3
kg algae m-2
roots
woody
succulent p = 0.0008
p = 0.0018
p = 0.0020
Fig. 4. 15N concentration (atom%) of pickleweed tissues, with p
values from Kruskal–Wallis tests. Non-parametric multiple compar-
isons were not conducted (n=5; nN8 required).
stayed approximately the same despite the new
growth we observed on all plants.
Macroalgal additions had no effect on N concen-
tration of pickleweed tissues. Mean N levels dropped
from initial concentrations by 31% in succulent
tissues and by 13% in root tissues. P in succulent
tissues did not differ by algal treatment or change
from initial levels.
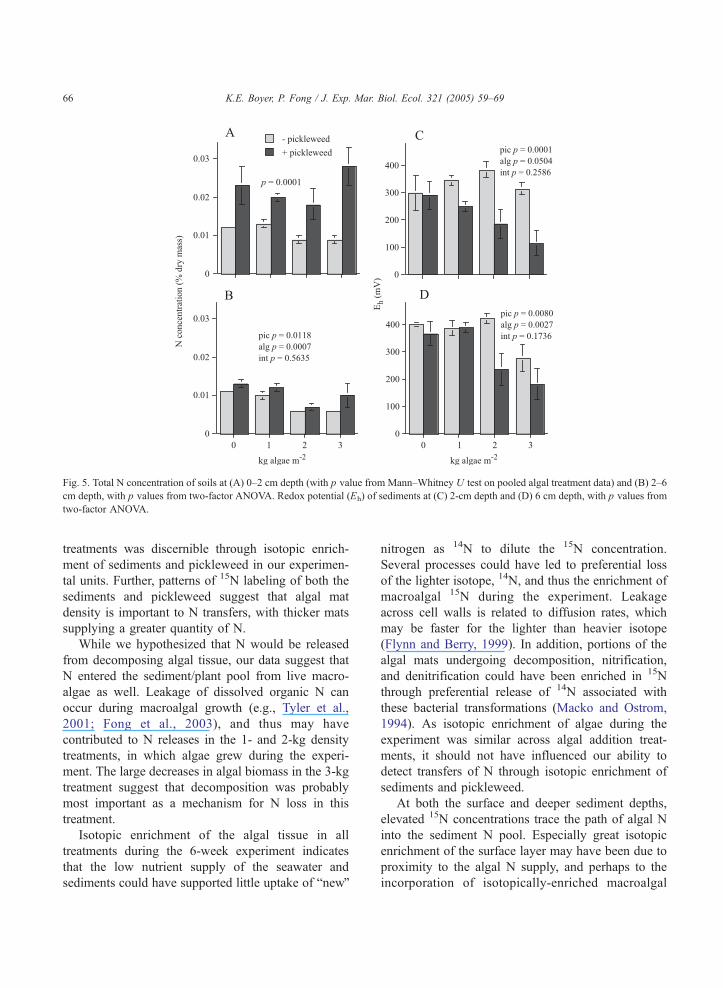
Pots with pickleweed had significantly higher soil
N concentrations at both the 0–2 and 2–6 cm depths
than pots without pickleweed (Fig. 5A,B). At the 0–2-
cm depth, there was no significant effect of algal
treatment. However, at 2–6 cm, soil N concentration
was significantly lower in the two higher density
treatments than in the 0- or 1-kg m�2 treatments
(PLSD, pb0.05). Sediment P did not differ by
treatment (grand mean=0.05F0.00%).
Organic matter in surface sediments was signifi-
cantly higher with pickleweed present (grand
mean=0.22F0.02%) than without (grand mean=
0.13F0.02%), reflecting sediment N patterns. Sedi-
ment organic matter did not differ by algal density
treatment.
Sediments at the end of the experiment ranged
from well oxygenated to moderately reduced (Patrick
et al., 1996). Redox potential at both the 2- and 6-cm
depths decreased significantly with pickleweed
present (Fig. 5C,D). At the 6-cm depth, the 3-kg
algae treatment led to more reduced conditions than
both the 0- and 1-kg treatments (PLSD, pb0.05),
while the 2-kg treatment had an intermediate poten-
tial. The presence of pickleweed appeared to influence
the degree to which macroalgal mats led to declines in
redox potential; however, we found no significant
interactions at either depth to support this.
The flow-through seawater system kept sediments
well flushed of salts. Grand means of soil salinity
were 34F1 ppt at the 0–2 cm depth and 31F0.5 ppt
at 2–6 cm depth. There were no differences by
treatment.
4. Discussion
The use of 15N as a tracer allowed us to document
that macroalgae play a role in transferring N from the
water column to sediments and vascular plants of the
intertidal zone. Algal N lost from all mat density
0
0.01
0.02
0.03
N c
once
ntr
atio
n (
% d
ry m
ass)
A
0
0.01
0.02
0.03
0 1 2 3
kg algae m-2 kg algae m-2
B
p = 0.0001
pic p = 0.0118
alg p = 0.0007
int p = 0.5635
0
100
200
300
400
Eh (
mV
)
C
+ pickleweed
- pickleweed
0
100
200
300
400
0 1 2 3
D
pic p = 0.0001
alg p = 0.0504
int p = 0.2586
pic p = 0.0080
alg p = 0.0027
int p = 0.1736
Fig. 5. Total N concentration of soils at (A) 0–2 cm depth (with p value from Mann–Whitney U test on pooled algal treatment data) and (B) 2–6
cm depth, with p values from two-factor ANOVA. Redox potential (Eh) of sediments at (C) 2-cm depth and (D) 6 cm depth, with p values from
two-factor ANOVA.
K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–6966
treatments was discernible through isotopic enrich-
ment of sediments and pickleweed in our experimen-
tal units. Further, patterns of 15N labeling of both the
sediments and pickleweed suggest that algal mat
density is important to N transfers, with thicker mats
supplying a greater quantity of N.
While we hypothesized that N would be released
from decomposing algal tissue, our data suggest that
N entered the sediment/plant pool from live macro-
algae as well. Leakage of dissolved organic N can
occur during macroalgal growth (e.g., Tyler et al.,
2001; Fong et al., 2003), and thus may have
contributed to N releases in the 1- and 2-kg density
treatments, in which algae grew during the experi-
ment. The large decreases in algal biomass in the 3-kg
treatment suggest that decomposition was probably
most important as a mechanism for N loss in this
treatment.
Isotopic enrichment of the algal tissue in all
treatments during the 6-week experiment indicates
that the low nutrient supply of the seawater and
sediments could have supported little uptake of bnewQ
nitrogen as 14N to dilute the 15N concentration.
Several processes could have led to preferential loss
of the lighter isotope, 14N, and thus the enrichment of
macroalgal 15N during the experiment. Leakage
across cell walls is related to diffusion rates, which
may be faster for the lighter than heavier isotope
(Flynn and Berry, 1999). In addition, portions of the
algal mats undergoing decomposition, nitrification,
and denitrification could have been enriched in 15N
through preferential release of 14N associated with
these bacterial transformations (Macko and Ostrom,
1994). As isotopic enrichment of algae during the
experiment was similar across algal addition treat-
ments, it should not have influenced our ability to
detect transfers of N through isotopic enrichment of
sediments and pickleweed.
At both the surface and deeper sediment depths,
elevated 15N concentrations trace the path of algal N
into the sediment N pool. Especially great isotopic
enrichment of the surface layer may have been due to
proximity to the algal N supply, and perhaps to the
incorporation of isotopically-enriched macroalgal

K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–69 67
detritus into the sediment surface layer. Ammonia
volatilization, which results in preferential release of14N (Wada et al., 1981), may have further enhanced15N concentration in the surface sediments. Condi-
tions suitable for ammonia volatilization were perhaps
provided by the heightened pH that can result in water
during algal photosynthesis and the position of the
NH4–N source (the algal mat) on the sediment surface
as it dried during simulated low tide (Brady and Weil,
1999). If so, this process was not differentially
affected by algal mat density, but rather led to
statistically similar isotopic signatures across algal
addition treatments.
With increasing macroalgal mat density, a striking
stepwise 15N enrichment of the deeper sediment zone
suggests that sediments intercepted a greater supply of
N from denser macroalgal mats. Our data are not
supportive of an alternative explanation for this
pattern, that denitrification beneath thicker mats led
to isotopic enrichment of sediments (Handley and
Raven, 1992). Redox potential fell to levels associated
with denitrification (b250 mV; Patrick and Jugsujinda,
1992) in some of the pickleweed pots with higher
density algal additions, but sediment Eh explained
little of the variation in 15N (regression on sediments
from pickleweed pots with 1–3 kg/m2 of algae; in
surface sediments, n=15, p=0.8740, R2=�0.002; at
depth, n=15, p=0.1290, R2=�0.168).
Pickleweed enrichment in 15N with Enteromorpha
present at all three densities suggests that N transferred
from this alga may contribute to the plant’s mineral
nutrition whenever algal mats occur on sediments in
close proximity. Increased isotopic enrichment with
increasing mat density provides evidence that a greater
quantity of N was available to pickleweed in the
presence of the thickest algal mats.
As macroalgal presence/density did not lead to
differences in pickleweed total branch length or tissue
N concentration, the contribution of macroalgal N to
pickleweed productivity over short time periods
remains uncertain. In this study, pickleweed declined
in N concentration due, in part, to growth, which
would have bdilutedQ N across biomass. However,
growth was highly variable among individuals, with
some even decreasing in total length. In a study at
Mugu Lagoon, Onuf (unpublished manuscript) found
that only ~30% of new branch tips survived for N3
months and that biomass loss by shedding of branches
could be up to 4� the amount of new biomass
produced. Greater sediment N and organic matter
where pickleweed was present further suggests that
the plant contributed detritus to the sediments. With
high variability in growth and uncertainty about tissue
losses, we are unable to adequately assess pickleweed
productivity or, therefore, to estimate the absolute
contribution of algal N to it.
In conclusion, this experiment provides evidence
that mats of an opportunistic green macroalga link
water column and benthic N pools through uptake of
water column N and transfers to underlying tidal flat
or vegetated saltmarsh sediments. Transfers of N
occurred from algal mats that were both growing and
declining in biomass, suggesting that macroalgae may
contribute N to sediments and marsh plants whenever
mats are present. In addition, the persistence of algal
blooms in the intertidal zone of estuaries over periods
of months may have a cumulative effect on N pools of
both sediments and emergent vascular plants, perhaps
over many years of recurring algal blooms. As has
been suggested previously for a US Atlantic coast
marsh (Gerard, 1999), the N dynamics of intertidal
sediments and marsh plants appear to be linked to the
N pools of co-occurring macroalgal mats, and further
study is needed to assess the significance of N
transfers.
Acknowledgements
We thank Larry Elkins for providing space at the
Redondo Beach Generating Station, Wayne Shipman
for help in developing the tidal simulator, and Karleen
Boyle for sampling assistance. Phil Rundel, Christy
Tyler, Rick Vance, Rich Ambrose, and anonymous
reviewers provided helpful comments. Funding was
provided by the United States Environmental Protec-
tion Agency (fellowship #U915399 to K. Boyer and
grant #R827637 to P. Fong), the Society of Wetland
Scientists, the California Native Plant Society, and the
American Association of University Women. [SS]
References
Bierzychudek, A., D’Avanzo, C., Valiela, I., 1993. Effects of
macroalgae, night and day, on ammonium profiles in Waquoit
Bay. Biol. Bull. 185, 330–331.

K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–6968
Bolam, S.F., Fernandes, T.F., Read, P., Raffaelli, D., 2000. Effects
of macroalgal mats on intertidal sandflats: an experimental
study. J. Exp. Mar. Biol. Ecol. 249, 123–137.
Boyer, K.E., Zedler, J.B., 1998. Effects of nitrogen additions on the
vertical structure of a constructed cordgrass marsh. Ecol. Appl.
8, 692–705.
Boyer, K.E., Fong, P., Vance, R.R., Ambrose, R.F., 2001.
Salicornia virginica in a southern California saltmarsh: seasonal
patterns and a nutrient enrichment experiment. Wetlands 21,
315–326.
Boyle, K.A., Fong, P., Kamer, K., 2004. Spatial and temporal
patterns in sediment and water column nutrients in an eutrophic
southern California estuary. Estuaries 27, 378–388.
Brady, N.C., Weil, R.R., 1999. The nature and properties of soils,
12th ed. Prentice Hall, New Jersey.
Broome, S.W., Woodhouse, W.W., Seneca, E.D., 1975. The
relationship of mineral nutrients to growth of Spartina alterni-
flora in North Carolina: II. The effects of N, P, and Fe fertilizers.
Soil Sci. Soc. Am. Proc. 39, 301–307.
Craft, C., Reader, J., Sacco, J.N., Broome, S.W., 1999. Twenty-five
years of ecosystem development of constructed Spartina
alterniflora (Loisel) marshes. Ecol. Appl. 9, 1405–1419.
DeLaune, R.D., Pezeshki, S.R., 1988. Relationship of mineral
nutrients to growth of Spartina alterniflora in Louisiana salt
marshes. Northeast Gulf Sci. 10, 55–60.
Ellison, A.M., Bertness, M.D., Miller, T., 1986. Seasonal patterns in
the belowground biomass of Spartina alterniflora (Graminae)
across a tidal gradient. Am. J. Bot. 73, 1548–1554.
Enoksson, V., 1993. Nutrient recycling by coastal sediments: effects
of added algal material. Mar. Ecol. Prog. Ser. 92, 245–254.
Flynn, K.J., Berry, L.S., 1999. The loss of organic nitrogen during
marine primary production may be significantly overestimated
when using N-15 substrates. Proc. R. Soc. Lond., B Biol. Sci.
266, 641–647.
Fong, P. 1986. Monitoring and manipulation of phytoplankton
bloom dynamics in a southern California estuary. MS Thesis,
San Diego State University. 106 pp.
Fong, P., Zedler, J.B., 2000. Sources, sinks, and fluxes of nutrients
(N + P) in a small, highly-modified estuary in southern
California. Urban Ecosyst. 4, 125–144.
Fong, P., Zedler, J.B., Donohoe, R.M., 1993. Nitrogen vs.
phosphorus limitation of algal biomass in shallow coastal
lagoons. Limnol. Oceanogr. 38 (5), 906–923.
Fong, P., Donohoe, R.M., Zedler, J.B., 1994. Nutrient concentration
in tissue of the macroalga Enteromorpha as a function of
nutrient history: an experimental evaluation using field micro-
cosms. Mar. Ecol. Prog. Ser. 106, 273–281.
Fong, P., Fong, J., Fong, C., 2003. Growth, nutrient storage, and
release of DON by Enteromorpha intestinalis in response to
pulses of nitrogen and phosphorus. Aquat. Bot. 78, 83–95.
Fujita, R.M., 1985. The role of nitrogen status in regulating transient
ammonium uptake and nitrogen storage by macroalgae. J. Exp.
Mar. Biol. Ecol. 92, 283–301.
Gerard, V.A., 1999. Positive interactions between cordgrass,
Spartina alterniflora, and the brown alga, Ascophyllum nodo-
sum ecad scorpioides, in a mid-Atlantic coast salt marsh. J. Exp.
Mar. Biol. Ecol. 239, 157–164.
Handley, L.L., Raven, J.A., 1992. The use of natural abundance of
nitrogen isotopes in plant physiology and ecology. Plant Cell
Environ. 15, 965–985.
Hansen, L.S., Blackburn, T.H., 1992. Effect of algal bloom
deposition on sediment respiration and fluxes. Mar. Biol. 112,
147–152.
Hansen, K., Kristensen, E., 1997. Impact of macrofaunal recoloni-
zation on benthic metabolism and nutrient fluxes in a shallow
marine sediment previously overgrown with macroalgal mats.
Estuar. Coast. Shelf Sci. 45, 613–628.
Hauxwell, J., Cebrian, J., Valiela, I., 2001. Macroalgal canopies
contribute to eelgrass (Zostera marina) decline in temperate
estuarine ecosystems. Ecology 82, 1007–1022.
Hernandez, I., Peralta, G., Perez-Llorens, J.L., Vergara, J.J., 1997.
Biomass and dynamics of growth of Ulva species in Palmones
River Estuary. J. Phycol. 33, 764–772.
Kamer, K., Boyle, K.A., Fong, P., 2001. Macroalgal bloom
dynamics in a highly eutrophic southern California estuary.
Estuaries 24, 623–635.
Kennison, R., Kamer, K., Fong, P., 2003. Nutrient dynamics and
macroalgal blooms: a comparison of five southern California
estuaries. Southern California Coastal Water Research Project
Technical Report #416, Westminster, California.
Koch, M.S., Mendelssohn, I.A., McKee, K.L., 1990. Mechanisms
for the hydrogen sulfide-induced growth limitation of Spartina
alterniflora and Panicum hemitomon. Limnol. Oceanogr. 35,
399–408.
Langis, R., Zalejko, M., Zedler, J.B., 1991. Nitrogen assessments in
a constructed and a natural salt marsh of San Diego Bay,
California. Ecol. Appl. 1, 40–51.
Lavery, P.S., McComb, A.J., 1991. Macroalgal-sediment nutrient
interactions and their importance to macroalgal nutrition in a
eutrophic estuary. Estuar. Coast. Shelf Sci. 32, 281–295.
Macko, S.A., Ostrom, N.E., 1994. Pollution studies using stable
isotopes. In: Lajtha, K., Michener, R.H. (Eds.), Stable Isotopes
in Ecology and Environmental Science. Methods in Ecology
Series. Blackwell Scientific, Oxford, pp. 45–62.
McGlathery, K.J., Krause-Jensen, D., Rysgaard, S., Christensen,
P.B., 1997. Patterns of ammonium uptake within dense mats of
the filamentous macroalga Chaetomorpha linum. Aquat. Bot.
59, 99–115.
Mendelssohn, I.A., Marcellus, K.L., 1976. Angiosperm production
of three Virginia marshes in various salinity and soil nutrient
regimes. Chesap. Sci. 17, 15–23.
Onuf, C.P., Unpublished manuscript. Demography and production
of Salicornia virginica L. in a southern California salt marsh.
Osgood, D.T., Zieman, J.C., 1993. Factors controlling aboveground
Spartina alterniflora (smooth cordgrass) tissue element compo-
sition and production in different-age barrier island marshes.
Estuaries 16, 815–826.
Osgood, D.T., Zieman, J.C., 1995. Sediment physico-chemistry
associated with natural marsh development on a storm-deposited
sand flat. Mar. Ecol. Prog. Ser. 20, 271–283.
Osgood, D.T., Zieman, J.C., 1998. The influence of subsurface
hydrology on nutrient supply and smooth cordgrass (Spartina
alterniflora) production in a developing barrier island marsh.
Estuaries 21, 767–783.

K.E. Boyer, P. Fong / J. Exp. Mar. Biol. Ecol. 321 (2005) 59–69 69
Paerl, H., 1997. Coastal eutrophication and harmful algal blooms:
importance of atmospheric deposition and groundwater as bnewQnitrogen and other nutrient sources. Limnol. Oceanogr. 42,
1154–1165.
Patrick Jr., W.H., Jugsujinda, A., 1992. Sequential reduction and
oxidation of nitrogen, manganese, and iron in flooded soil. Soil
Sci. Soc. Am. J. 56, 1071–1073.
Patrick, W.H., Gambrell, R.P., Faulkner, S.P., 1996. Redox
measurements of soils. Methods of Soil Analysis: Part 3.
Chemical methods. American Society of Agronomy, Madison,
pp. 255–1273.
Peckol, P., DeMeo-Anderson, B., Rivers, J., 1994. Growth, nutrient
uptake capacities, and tissue constituents of the macroalgae
Cladophora vagabunda and Gracilaria tikvahiae, related to
site-specific nitrogen loading rates. Mar. Biol. 121, 175–185.
Pregnall, A.M., 1983. Release of dissolved organic carbon from the
estuarine intertidal macroalga Enteromorpha prolifera. Mar.
Biol. 73, 37–42.
Pregnall, A.M., Rudy, P.P., 1985. Contribution of green macroalgal
mats (Enteromorpha spp.) to seasonal production in an estuary.
Mar. Ecol. Prog. Ser. 24, 167–176.
Rosenberg, G., Ramus, J., 1984. Uptake of inorganic nitrogen and
seaweed surface area: volume ratios. Aquat. Bot. 19, 65–72.
Sfriso, A., Pavoni, B., Marcomini, A., Orio, A.A., 1992. Macro-
algae, nutrient cycles, and pollutants in the lagoon of Venice.
Estuaries 15, 517–528.
Soulsby, P.G., Lowthion, D., Houston, M., Montgomery, H.A.C.,
1985. The role of sewage effluents in the accumulation of
macroalgal mats on intertidal mudflats in two basins in southern
England. Neth. J. Sea Res. 19, 257–263.
Sullivan, G., Zedler, J.B., 1999. Functional redundancy among tidal
marsh halophytes: a test. Oikos 84 (2), 246–260.
Thybo-Christesen, M., Rasmussen, M.N., Blackburn, T.H., 1993.
Nutrient fluxes and growth of Cladophora sericea in a shallow
Danish bay. Mar. Ecol. Prog. Ser. 100, 273–281.
Tyler, A.C., Zieman, J.C., 1998. Patterns of development in the
creekbank region of a barrier island Spartina alterniflora marsh.
Mar. Ecol. Prog. Ser. 180, 161–177.
Tyler, A.C., McGlathery, K.J., Anderson, I.C., 2001. Macroalgae
mediation of dissolved organic nitrogen fluxes in a temperate
coastal lagoon. Estuar. Coast. Shelf Sci. 53, 155–168.
Wada, E., Shibata, R., Torri, T., 1981. 15N abundance in Antarctica:
origin of soil nitrogen and ecological implications. Nature 392,
327–329.
Zedler, J.B., 1980. Algal mat productivity: comparisons in a salt
marsh. Estuaries 3, 122–131.





























