Embed Size (px)
Citation preview

Macrophytic, epipelic and epilithic primary productionin a semiarid Mediterranean stream
J . VELASCO*, A. MILLAN*, M. R. VIDAL-ABARCA*, M. L. SUAREZ*, C. GUERRERO*
AND M. ORTEGA †
*Department of Ecology and Hydrology, University of Murcia, Murcia, Spain
†Department of Biology and Ecology, University of Almeria, Almeria, Spain
SUMMARY
1. Primary production by Chara vulgaris and by epipelic and epilithic algal assemblages
was measured in a semiarid, Mediterranean stream (Chicamo stream, Murcia, Spain)
during one annual cycle.
2. The rates of gross primary production (GPP) and community respiration (CR) were
determined for each algal assemblage using oxygen change in chambers. The net daily
metabolism (NDM) and the GPPd)1 : CR24 ratio were estimated by patch-weighting the
assemblage-level metabolism values.
3. Gross primary production and CR showed significant differences between assemblages
and dates. The highest rates were measured in summer and spring, while December was
the only month when there were no significant differences in either parameters between
assemblages. GPP was strongly correlated with respiration, but not with algal biomass.
4. Chara vulgaris showed the highest mean annual metabolic rates (GPP ¼ 2.80 ± 0.83 gC
m)2 h)1, CR ¼ 0.76 ± 0.29 gC m)2 h)1), followed by the epilithic assemblage
(GPP ¼ 1.97 ± 0.73 gC m)2 h)1, CR ¼ 0.41 ± 0.12 gC m)2 h)1) and epipelic algae
(GPP ¼ 1.36 ± 0.22 gC m)2 h)1, CR ¼ 0.39 ± 0.06 gC m)2 h)1).
5. The epipelic assemblage dominated in terms of biomass (82%) and areal cover (88%),
compared with the other primary producers. Epipelic algae contributed 84% of gross
primary production and 86% of community respiration in the stream.
6. Mean monthly air temperature was the best single predictor of macrophyte respiration
and of epipelic GPP and CR. However, ammonium concentration was the best single
predictor of C. vulgaris GPP, and suspended solid concentration of epilithon GPP and CR.
7. Around 70% of the variation in both mean GPP and mean CR was explained by the
mean monthly air temperature alone. A multiple regression model that included
conductivity, PAR and nitrates in addition to mean monthly air temperature, explained
99.99% of the variation in mean CR.
8. Throughout the year, NDM was positive (mean value 7.03 gC m)2 day)1), while the
GPP : CR24 ratio was higher than 1, confirming the net autotrophy of the system.
Keywords: chamber method, Chara vulgaris, gross primary production, Mediterranean stream, netdaily metabolism
Introduction
Important ecological questions concerning the energy
flow in stream ecosystems require estimates of algal
primary production (Morin, Lamoureux & Busnarda,
1999). Such estimates of primary production have
been made in lotic systems across biomes and
Correspondence: J. Velasco, Department of Ecology and
Hydrology, University of Murcia, Campus of Espinardo,
30100 Murcia, Spain. E-mail: [email protected]
Freshwater Biology (2003) 48, 1408–1420
1408 � 2003 Blackwell Publishing Ltd

latitudes in North America (Bott et al., 1985; Mulhol-
land et al., 2001) and in most regions of the world
(Lamberti & Steinman, 1997). However, primary
production studies in Mediterranean-climate regions
(areas surrounding the Mediterranean Sea, parts of
western North America, parts of west and south
Australia, southwestern South Africa and parts of
central Chile) are largely lacking. Rainfall seasonality
and variability are the principal attributes of the
Mediterranean-type climate, most precipitation result-
ing from a few major storm events that may produce
flooding (Gasith & Resh, 1999). Annual precipitation
is highly variable among Mediterranean areas, usually
ranging between 275 and 900 mm, while certain
Mediterranean-climate regions fall into the category
of semiarid regions (between 200 and 500 mm;
UNESCO, 1979).
The importance of primary production in Mediter-
ranean streams is generally expected to increase with
increasing gradients of aridity (Gasith & Resh, 1999),
following the pattern found in an analysis of 30
streams over a large geographic range (Lamberti &
Steinman, 19971 ), where GPP increased significantly
with mean temperature and declined with total
precipitation. To our knowledge, there are few data
on primary production in Mediterranean streams.
Four streams in the Spanish Mediterranean area have
been studied: La Solana and Riera Major, in the NE of
the Iberian Peninsula with 800–1000 mm mean annual
precipitation (Guasch & Sabater, 1994, 1998); Monte-
sina stream, in the south (Molla, Malchik & Casado,
1994) with 527 mm of mean annual precipitation; and
Chicamo stream (Suarez & Vidal-Abarca, 20002 ),
located in southeastern Spain, with a semiarid climate
(250 mm mean annual precipitation). Early estimates
of primary production and respiration in Chicamo
stream (Suarez & Vidal-Abarca, 2000) were higher
than those reported in the other Mediterranean
streams of wetter areas, but were similar to those
obtained in the warm desert stream, Sycamore Creek
(Busch & Fisher, 1981; Grimm & Fisher (1984).
Although autotrophy may be important in arid and
semiarid streams, the patchiness of aquatic vegeta-
tion, its seasonal growth pattern and the contribution
of periphyton and macrophytes to autochthonous
sources during the year may differ. The objectives of
this paper were to determine the contribution of
different algal assemblages to annual primary pro-
duction in a semiarid Mediterranean stream (Chicamo
stream), to confirm its autotrophic metabolism, to
identify climatic and environmental variables that
might govern primary production in this stream, and
to compare metabolic rates with those of other
Mediterranean and desert streams.
Study area
Chicamo stream is a saline and intermittent tributary
of the Segura River located in southeast Spain (38�Nlatitude), in the most arid area of the province of
Murcia. The climate is characterised by a mean annual
precipitation below 300 mm and a mean annual
temperature of 18�C. A long warm and dry summer
season is interrupted by spring and autumn rains, the
latter followed by a short and temperate winter.
Annual and interannual variations in rainfall can lead
to a high discharge variability, with extreme condi-
tions of flooding and drying. The highest flows
normally occur in early autumn, followed by a second
peak in spring.
Chicamo stream is a 4th-order stream, draining a
sedimentary watershed of 502 km2. Surface water
flow is intermittent in 10 km of its 59.4 km total
length. The natural cover on the watershed is open
Mediterranean scrub, although much is dedicated to
citrus and horticultural crops.
Studies were conducted in a permanent, braided,
shallow and unshaded reach, 100 m length, typical of
the middle section. The substrate consisted of con-
solidated and impermeable marls, with deposits of
gravels and sand in the erosional zones of runs, and
silts in pools and depositional zones. Chicamo
stream water is hyposaline and hard (7.6 g L)1 mean
salinity, 219.5 mg L)1 mean alkalinity), well oxygen-
ated and rich in nutrients, especially nitrates and
ammonium (Vidal-Abarca et al., 2000). Riparian
vegetation is sparse because of frequent floods.
There are no trees, but isolated shrubs such as
Phragmites australis, Tamarix canariensis and Juncus
maritimus are present.
Aquatic primary producers in the study site include
the macrophyte Chara vulgaris (with epiphytic algae)
in pools, an extensive diatom assemblage on fine
sediments, including Nitzschia, Amphora, Navicula,
Gyrosigma and Pleurosigma among the most abundant
genera (D. Ros, per. com.3 ), and epilithic periphyton
dominated by the cyanobacteria Calothrix and the
chlorophyte Kentrosphaera facciolae, although in spring
Primary production in a semiarid stream 1409
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

the filamentous green alga Cladophora glomerata pre-
dominates.
Fine benthic particulate organic matter (FPOM) is
the principal benthic fraction (about 70% of the total)
while coarse particulate organic matter (CPOM) rep-
resents only 6% of the total because of the scarcity of
riparian vegetation and the consequent low input of
allochthonous materials (Martınez et al., 19984 ). Dis-
solved organic carbon (DOC) is the principal organic
carbon source flowing in the Chicamo stream, partic-
ularly when flash-floods occur (Vidal-Abarca et al.,
2001).
Methods
Metabolism experiments were conducted six times
between July 1998 and May 1999 to record the annual
variability. Macrophyte, epipelic and epilithic primary
production and community respiration were meas-
ured using oxygen change in clear plastic chambers
(25.5 · 13.8 · 8 cm) placed in the stream. Because of
the low discharge and current in the stream, we did
not use a pump for water recirculation. Chamber
temperature never exceeded stream temperature by
more than 3 �C. Four replicate estimates of production
and respiration were obtained from each assemblage.
The macrophyte, C. vulgaris, and epipelic samples
were collected 15 days before the experiment, tran-
ferred to plastic trays (21.5 · 12 · 2 cm) and placed
on the streambed contiguous with the sediment
surface for colonisation to continue. At the start of
the experiment, the trays were removed from the
stream and transferred to the chambers with minimal
disturbance to the community during removal. For
the epilithon, the stones used for each incubation were
collected from the stream immediately before the
experiment started.
All the chambers were filled with stream water,
closed and placed in situ. Each measurement of net
photosynthetic rate was made over a 3-h period (2 h
dark and 1 h light) during the morning. Chambers for
dark incubations were covered with aluminium foil.
Changes in O2 concentration were detected using
standard Winkler titration scaled to 10 mL sample
sizes taken with a needle and syringe. Winkler deter-
minations of dissolved oxygen (DO) were made before
and after the dark incubations, and after the light
incubations. Water temperature in the stream and in
each chamber was recorded at each sampling time.
Gross primary production (GPP) was calculated by
adding the change in DO measured during the dark
incubation to the change measured during the light
incubation. Because respiration rates include the
metabolism of heterotrophs such as microbes and
insects as well as autotrophs, respiration is termed
community respiration (CR). Oxygen generation was
converted to carbon fixation assuming a photosyn-
thetic quotient of 1.2 (Bott, 1997a).
After the incubation experiments, C. vulgaris was
removed from the chambers and stored on ice until it
was washed with tap water in the laboratory and
weighed. Three samples of 3 g of fresh macrophyte
with its epiphitic algae were taken from each chamber
to determine chlorophyll a (Chla) concentration by
spectrophotometry, following extraction in boiling
90% ethanol as described by Biggs (1995). Concentra-
tions were converted to algal carbon biomass using a
factor of 30, which is suitable for communities in
nutrient-rich, unshaded environments (Vollenweider,
1974; Bott et al., 1997). The rest of the macrophyte was
dried at 60 �C to constant dry weight (DW) and then
ashed at 450 �C for 4 h to estimate the content of ash
free dry weight (AFDW) in each chamber. Triplicate
sediment cores of 2.7 cm diameter were also collected
from each epipelic chamber for Chla and AFDW
analyses. The rocks were removed and returned to the
laboratory for determination of surface area and Chla
and AFDW content. Rock surface area was deter-
mined by tracing the outline of the top of the rock on
aluminium paper, cutting out the shape, and compar-
ing its weight with the weight of a known area of
paper. Periphyton on rocks was scraped, brushed and
washed from the rock surface. The extracts were
filtered onto Whatman A filters for Chla concentration
and AFDW determinations. Autochthonous detritus
standing crop was determined as the difference
between AFDW measurements (assuming a 50%
carbon content) and the periphyton biomass obtained
from Chla concentrations.
On each date, the area covered by each primary
producer assemblage in the stream was measured in
the study reach (100 m length) to extrapolate the
metabolism rates estimated for each assemblage to the
reach. Measurements of discharge, photosynthetically
active radiation (PAR), air and water temperatures,
conductivity, salinity and dissolved oxygen were
made in the stream during the experiments. Two
water samples were taken to determine suspended
1410 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

solids, ammonium, nitrate and soluble reactive phos-
phorus (SRP) concentration on each date. The samples
were kept cool until their arrival at the laboratory,
where they were filtered onto preashed and pre-
weighed GF/F glass-fiber filters5 (Whatman) and
oven-dried at 60 �C to constant dry weight. Nutrients
were determined according to standard methods
(American Public Health Association (APHA), 1992):
nitrate by the cadmium reduction method, nitrites by
sulfanylic acid colorimetry, ammonium by phenol
nitroprussiate colorimetry and phosphate by ascorbic
acid colorimetry.
The days on which the experiments were performed
were sunny with mean PAR values during the experi-
ments of more than 1000 lE m)2 s)1, except in August
(914.78 lE m)2 s)1) and December (595.94 lE m)2 s)1).
At the stream reach scale, we estimated the net
daily metabolism (NDM) and the GPPd)1/CR24 ratio
by patch-weighting the assemblage-level metabolism
values. Chamber measurements of community res-
piration were extrapolated to a 24-h period, assu-
ming the respiratory rate during the night to be half
daylight respiration because of the decrease in
temperature:
GPP d�1 ¼ ðNPP h�1 � light hoursÞþ ðCR h�1 � light hoursÞ;
and
CR24 ¼ ðCR h�1 � light hoursÞþ ðCR h�1=2Þ � dark hours� �
Differences in gross primary production and res-
piration between primary producer assemblages
were analysed using analysis of variance (one way
ANOVAANOVA) and Tukey’s honest significant difference
test (HSD). All data were log(x + 1) transformed to
normalise distributions and equalise variance. Rela-
tionships of climatic and environmental variables to
metabolic rates were examined using bivariate cor-
relation and multiple linear regression approaches.
Pearson correlation analysis was used to indentify
relationships between single factors and metabolic
rates. Finally, a stepwise multiple linear regression
used all significant (P £ 0.05) correlated factors and the
response variables (GPP and CR). Statistical analyses
were performed using Systat (Wilkinson, 1996).
Results
Climatic and environmental conditions
Table 1 shows the mean values of physical and
chemical parameters measured for the six experiment
dates in Chicamo stream, together with monthly air
temperature and precipitation recorded in a nearby
metereological station. Total precipitation during the
study period (from July 1998 to May 1999) was
143.5 mm, mainly concentrated in autumn and winter
peaks. Discharge variation during the study period
was closely related to daily precipitation. Discharge
was very low (£ 3.5 L s)1) except following rainfall in
December and March. The mean annual temperature
Table 1 Mean (±1SE) values of physical and chemical parameters measured on the experiment dates in Chicamo stream, together
with monthly air temperature and precipitation recorded at a nearby meterological station
17/07/1998 19/08/1998 15/10/1998 28/12/1998 20/03/1999 28/05/1999
Monthly mean air temperature (�C) 26.4 26.5 18.9 9.7 14 20.7
Monthly precipitation (mm) 0 0 0 51.5 22.5 0
PAR during the experiment (lE m)2 s)1) 1788 915 1437 596 1512 1760
Air temperature during the experiment (�C) 33 27 25.5 7.5 23.5 30.5
Water temperature during the experiment (�C) 32.5 30 20 10 19.5 27.5
Discharge (L s)1) 3.5 0.17 0 73 48.6 3.3
Conductivity (mS cm)1) 14 13.5 13.5 11 12.7 13
Salinity 10.9 10 11 9 9.3 11
Total suspended solids (mgDW L)1) 69.6 ± 6.1 10.85 ± 5.7 33.82 ± 3.3 6.20 ± 1.45 18.13 ± 8.13 302 ± 98
Alkalinity (meq L)1) 4.88 ± 0 5.24 ± 0.04 4.28 ± 0.52 5.32 ± 0.28 5.4 ± 0.04 6.44 ± 0.12
Dissolved oxygen (mg L)1) 6.47 ± 0.03 5.35 ± 0 10.38 ± 0.45 9.78 ± 0.03 7.28 ± 0.06 6.08 ± 0.18
NH4-N (lg L)1) 173 ± 12 70 ± 1.2 55.5 ± 6.7 19.12 ± 1.64 0 46 ± 3.9
NO3-N (lg L)1) 8555 ± 529 549 ± 14 59.15 ± 4.9 2611 ± 129 3481 ± 145 1239 ± 124
SRP (lg L)1) 8.24 ± 0.4 255 ± 11 6.38 ± 0.74 2.17 ± 0.12 4.78 ± 0.5 7.71 ± 2.72
Primary production in a semiarid stream 1411
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420
was 18.2 �C with August being the hottest month,
when the maximum temperature reached over 32 �C,
while December was the coldest month, with the
minimum recorded temperature being 4 �C. The
mean water temperature was 20 �C (max ¼ 30 �C in
August; min ¼ 7.5 �C in December).
Areal coverage and biomass of algal assemblages
Epipelic algae were the most widespread assemblage
throughout the study period, representing more than
70% of the total wetted surface (Table 2). The cover-
age by C. vulgaris was peaked in October, though
subsequent high winter flows buried it with sedi-
ments and reduced it to small patches. In March, the
macrophyte was absent, but it started to grow again in
May. Maximum epilithon cover was attained in
December and minimum in October. C. vulgaris cover
was negatively correlated with discharge and
monthly precipitation (r ¼ )0.92, P £ 0.005 and
r ¼ )0.84, P £ 0.05) while it was positively correlated
with ammonium (r ¼ 0.85, P £ 0.02). Epilithic algal
cover showed a positive correlation with discharge
and precipitation (r ¼ 0.95, P £ 0.001; r ¼ 0.89,
P £ 0.005, respectively) and a negative corrrelation
with conductivity (r ¼ )0.76, P £ 0.05). The epipelic
algal cover showed a negative correlation with the
dissolved oxygen (r ¼ )0.78, P £ 0.05).
Mean annual biomass values for macrophyte,
epipelic and epilithic assemblages were 25.26, 5.05
and 4.62 gC m)2, respectively (Table 2). C. vulgaris
registered the highest biomass values on all the dates
except in March when it was absent. Its biomass
peak occurred in December, and the minimum in
summer. It was positively correlated with the stand-
ing crop of detritus and ammonium concentration
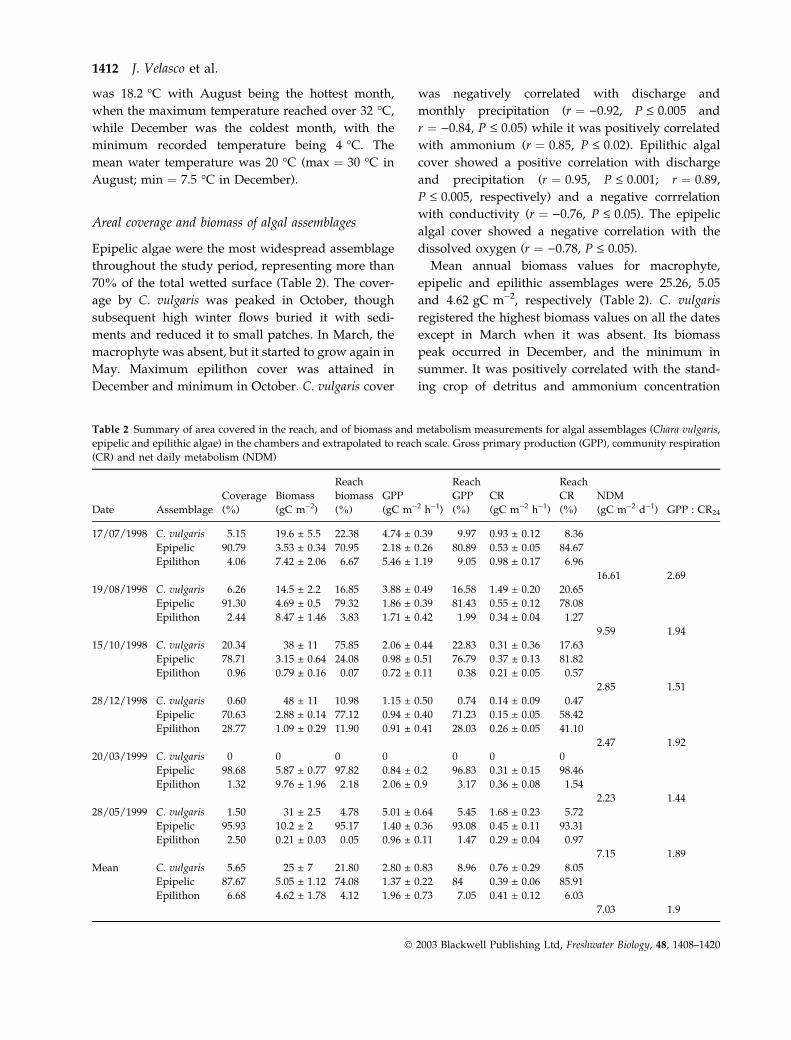
Table 2 Summary of area covered in the reach, and of biomass and metabolism measurements for algal assemblages (Chara vulgaris,
epipelic and epilithic algae) in the chambers and extrapolated to reach scale. Gross primary production (GPP), community respiration
(CR) and net daily metabolism (NDM)
Date Assemblage
Coverage
(%)
Biomass
(gC m)2)
Reach
biomass
(%)
GPP
(gC m)2 h)1)
Reach
GPP
(%)
CR
(gC m)2 h)1)
Reach
CR
(%)
NDM
(gC m)2 d)1) GPP : CR24
17/07/1998 C. vulgaris 5.15 19.6 ± 5.5 22.38 4.74 ± 0.39 9.97 0.93 ± 0.12 8.36
Epipelic 90.79 3.53 ± 0.34 70.95 2.18 ± 0.26 80.89 0.53 ± 0.05 84.67
Epilithon 4.06 7.42 ± 2.06 6.67 5.46 ± 1.19 9.05 0.98 ± 0.17 6.96
16.61 2.69
19/08/1998 C. vulgaris 6.26 14.5 ± 2.2 16.85 3.88 ± 0.49 16.58 1.49 ± 0.20 20.65
Epipelic 91.30 4.69 ± 0.5 79.32 1.86 ± 0.39 81.43 0.55 ± 0.12 78.08
Epilithon 2.44 8.47 ± 1.46 3.83 1.71 ± 0.42 1.99 0.34 ± 0.04 1.27
9.59 1.94
15/10/1998 C. vulgaris 20.34 38 ± 11 75.85 2.06 ± 0.44 22.83 0.31 ± 0.36 17.63
Epipelic 78.71 3.15 ± 0.64 24.08 0.98 ± 0.51 76.79 0.37 ± 0.13 81.82
Epilithon 0.96 0.79 ± 0.16 0.07 0.72 ± 0.11 0.38 0.21 ± 0.05 0.57
2.85 1.51
28/12/1998 C. vulgaris 0.60 48 ± 11 10.98 1.15 ± 0.50 0.74 0.14 ± 0.09 0.47
Epipelic 70.63 2.88 ± 0.14 77.12 0.94 ± 0.40 71.23 0.15 ± 0.05 58.42
Epilithon 28.77 1.09 ± 0.29 11.90 0.91 ± 0.41 28.03 0.26 ± 0.05 41.10
2.47 1.92
20/03/1999 C. vulgaris 0 0 0 0 0 0 0
Epipelic 98.68 5.87 ± 0.77 97.82 0.84 ± 0.2 96.83 0.31 ± 0.15 98.46
Epilithon 1.32 9.76 ± 1.96 2.18 2.06 ± 0.9 3.17 0.36 ± 0.08 1.54
2.23 1.44
28/05/1999 C. vulgaris 1.50 31 ± 2.5 4.78 5.01 ± 0.64 5.45 1.68 ± 0.23 5.72
Epipelic 95.93 10.2 ± 2 95.17 1.40 ± 0.36 93.08 0.45 ± 0.11 93.31
Epilithon 2.50 0.21 ± 0.03 0.05 0.96 ± 0.11 1.47 0.29 ± 0.04 0.97
7.15 1.89
Mean C. vulgaris 5.65 25 ± 7 21.80 2.80 ± 0.83 8.96 0.76 ± 0.29 8.05
Epipelic 87.67 5.05 ± 1.12 74.08 1.37 ± 0.22 84 0.39 ± 0.06 85.91
Epilithon 6.68 4.62 ± 1.78 4.12 1.96 ± 0.73 7.05 0.41 ± 0.12 6.03
7.03 1.9
1412 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420
(r ¼ 0.96, P £ 0.001 and r ¼ 0.80, P £ 0.05, respect-
ively).
The epilithic assemblage registered its highest bio-
mass value in March, when C. glomerata was the
dominant alga. A second peak of epilithon biomass
occurred in summer, when the dominant alga was the
cyanobacterium Calothrix. Epilithon biomass also
showed a positive correlation with the standing crop
of detritus (r ¼ 0.88, P £ 0.01). The peak of epipelon
biomass was reached in spring. Epipelon biomass was
positively correlated with alkalinity (r ¼ 0.83, P £ 0.05).
At the reach level, epipelic biomass accounted for
the highest portion of biomass on all dates, except in
October when C. vulgaris represented 76% of the total
biomass of the reach. Differences in algal biomass
between assemblages were significant on all dates
(Table 3).
Gross primary production and respiration rates
The mean gross primary production rate was
2.05 ± 0.42 gC m)2 h)1 and the mean respiration rate
0.52 ± 0.12 gC m)2 h)1, resulting in a mean net pri-
mary production rate of 1.53 ± 0.31 gC m)2 h)1. Res-
piration represented 25.36% of mean gross primary
production.
At the assemblage level, C. vulgaris showed the
highest mean metabolic rates (Table 2), followed by
the epilithic and the epipelic assemblages. Summer
and spring were the most productive seasons
(Fig. 1a). The maximum values measured correspon-
ded to epilithon in July (5.46 gC m)2 h)1) and C.
vulgaris in May (5.01 gC m)2 h)1).
Gross primary production rates showed highly
significant differences between assemblages, except
in December (Table 3). Macrophyte (Chara) production
exceeded epilithic and epipelic production for most of
the year (Fig. 1a), significantly so in August, October
and May (Table 3). Epipelic and epilithic production
rates were similar except in July, when epilithic
production reached its highest value (Fig. 1a).
Respiration and gross primary production were
highly correlated for all the assemblages (r ¼ 0.9,
P £ 0.005 for C. vulgaris; r ¼ 0.83, P £ 0.02 for epipelic
algae, and r ¼ 0.97, P £ 0.001, for epilithic algae).
However, no significant correlations were found for
any algal assemblage between metabolic rates and
biomass.
Community respiration rates followed a similar
pattern to the gross primary production rates
(Fig. 1b). The highest respiration rates were also
found in spring and summer, and the lowest in
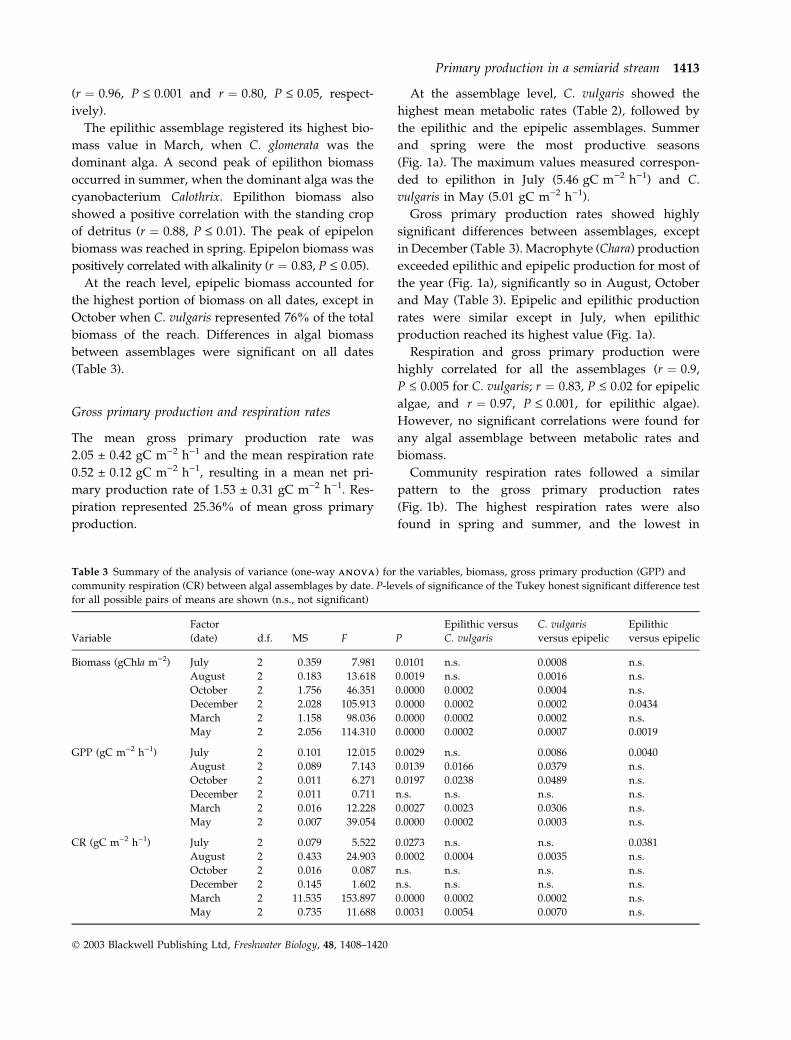
Table 3 Summary of the analysis of variance (one-way A N O V AA N O V A) for the variables, biomass, gross primary production (GPP) and
community respiration (CR) between algal assemblages by date. P-levels of significance of the Tukey honest significant difference test
for all possible pairs of means are shown (n.s., not significant)
Variable
Factor
(date) d.f. MS F P
Epilithic versus
C. vulgaris
C. vulgaris
versus epipelic
Epilithic
versus epipelic
Biomass (gChla m)2) July 2 0.359 7.981 0.0101 n.s. 0.0008 n.s.
August 2 0.183 13.618 0.0019 n.s. 0.0016 n.s.
October 2 1.756 46.351 0.0000 0.0002 0.0004 n.s.
December 2 2.028 105.913 0.0000 0.0002 0.0002 0.0434
March 2 1.158 98.036 0.0000 0.0002 0.0002 n.s.
May 2 2.056 114.310 0.0000 0.0002 0.0007 0.0019
GPP (gC m)2 h)1) July 2 0.101 12.015 0.0029 n.s. 0.0086 0.0040
August 2 0.089 7.143 0.0139 0.0166 0.0379 n.s.
October 2 0.011 6.271 0.0197 0.0238 0.0489 n.s.
December 2 0.011 0.711 n.s. n.s. n.s. n.s.
March 2 0.016 12.228 0.0027 0.0023 0.0306 n.s.
May 2 0.007 39.054 0.0000 0.0002 0.0003 n.s.
CR (gC m)2 h)1) July 2 0.079 5.522 0.0273 n.s. n.s. 0.0381
August 2 0.433 24.903 0.0002 0.0004 0.0035 n.s.
October 2 0.016 0.087 n.s. n.s. n.s. n.s.
December 2 0.145 1.602 n.s. n.s. n.s. n.s.
March 2 11.535 153.897 0.0000 0.0002 0.0002 n.s.
May 2 0.735 11.688 0.0031 0.0054 0.0070 n.s.
Primary production in a semiarid stream 1413
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420
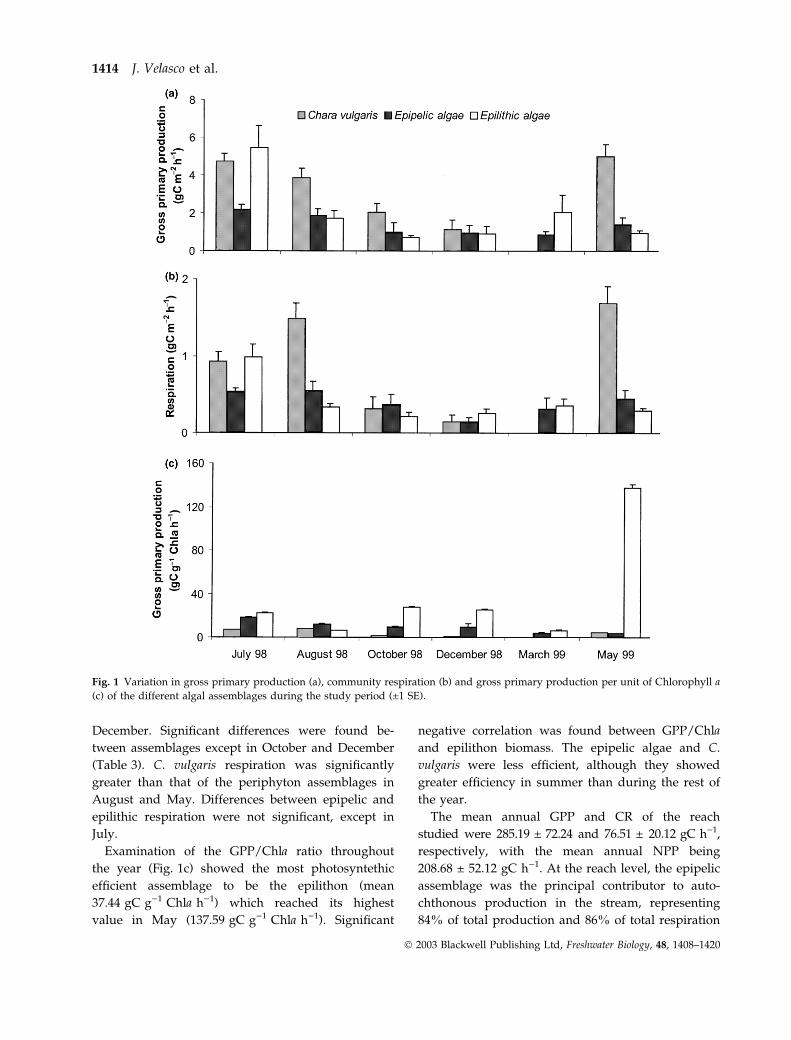
December. Significant differences were found be-
tween assemblages except in October and December
(Table 3). C. vulgaris respiration was significantly
greater than that of the periphyton assemblages in
August and May. Differences between epipelic and
epilithic respiration were not significant, except in
July.
Examination of the GPP/Chla ratio throughout
the year (Fig. 1c) showed the most photosyntethic
efficient assemblage to be the epilithon (mean
37.44 gC g)1 Chla h)1) which reached its highest
value in May (137.59 gC g)1 Chla h)1). Significant
negative correlation was found between GPP/Chla
and epilithon biomass. The epipelic algae and C.
vulgaris were less efficient, although they showed
greater efficiency in summer than during the rest of
the year.
The mean annual GPP and CR of the reach
studied were 285.19 ± 72.24 and 76.51 ± 20.12 gC h)1,
respectively, with the mean annual NPP being
208.68 ± 52.12 gC h)1. At the reach level, the epipelic
assemblage was the principal contributor to auto-
chthonous production in the stream, representing
84% of total production and 86% of total respiration
Fig. 1 Variation in gross primary production (a), community respiration (b) and gross primary production per unit of Chlorophyll a
(c) of the different algal assemblages during the study period (±1 SE).
1414 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

(Table 2). Its contribution to GPP varied during the
year from 71.23% in December to 96.83% in March.
C. vulgaris and epilithon production represented
only 8.96 and 7.05% of the total, although the
contribution of the macrophyte and its epiphitic
algae was more important in summer and autumn,
while the epilithon showed a maximum in Decem-
ber. Patch respiration followed a similar pattern to
patch GGP.
Relationships between metabolic rates and climatic
and environmental variables
The primary production of C. vulgaris showed a
positive correlation with ammonium (r ¼ 0.92,
P £ 0.005), standing crop of detritus (0.82, P £ 0.01),
and mean monthly air temperature (r ¼ 0.76,
P £ 0.05), and was negatively correlated with monthly
precipitation (r ¼ )0.82, P £ 0.05). Ammonium con-
centration was the best single predictor of GPP in the
simple regression analysis, explaining 81% of its
variation, although the best multiple regression model
included detritus standing crop and monthly air
temperature, which explained 92% of the variation
(Table 4).
The respiration rate of C. vulgaris was positively
correlated with mean monthly air temperature
(r ¼ 0.78, P £ 0.05), and negatively with dissolved
oxygen concentration (r ¼ )0.76, P £ 0.05) and
monthly precipitation (r ¼ )0.77, P £ 0.05). Mean
monthly air temperature was the only significant
predictor of macrophyte community respiration,
explaining 51% of its variation (Table 4).
Epipelic production and respiration were also
positively correlated with mean monthly air tempera-
ture (r ¼ 0.84, P £ 0.05, and r ¼ 0.99, P £ 0.001,
respectively) and water temperature (r ¼ 0.78,
P £ 0.05, and r ¼ 0.98, P £ 0.001, respectively). Res-
piration rate was also positively correlated with
conductivity (r ¼ 0.92, P £ 0.005) and negatively with
monthly precipitation (r ¼ )0.89, P £ 0.005), dis-
charge (r ¼ )0.77, P £ 0.05) and dissolved oxygen
(r ¼ )0.766, P £ 0.05). Mean monthly air temperature
was the best single predictor of the epipelic produc-
tion and respiration, explaining 63 and 98%, respect-
ively, of its variation. Multiple regression analyses
provided a predictive model of GPP that included,
besides monthly air temperature, nitrate, ammonium
concentration and PAR as predictor variables,
accounting for 99.99% of the variation (Table 4). The
best multiple regression model for epipelic respiration
(r2adj ¼ 99.99%) included monthly air temperature,
dissolved oxygen and conductivity as independent
variables (Table 4).
Table 4 Results of stepwise multiple
regression analysis for rates of gross pri-
mary production (GPP) and community
respiration (CR) for each algal assemblage
and for mean assemblage rates (n ¼ 6 for
each regression)
Community
Dependent
variable Independent variable r2adj P
C. vulgaris GPP Detritus standing crop 0.582 0.0146
Mean monthly air temperature 0.342 0.0223
Full model 0.924 0.0097
CR Mean monthly air temperature 0.507 0.0683
Epipelic GPP Mean monthly air temperature 0.629 0.0095
Nitrate 0.0016
Ammonium 0.0034
PAR 0.0041
Full model 0.999 0.0018
CR Mean monthly air temperature 0.980 0.0299
Dissolved oxygen 0.0279
Conductivity 0.0429
Full model 0.999 0.0066
Epilithon GPP Suspended solids 0.690 0.0253
CR Suspended solids 0.753 0.0157
Mean assemblages GPP Mean monthly air temperature 0.638 0.0352
CR Mean monthly air temperature 0.712 0.0018
Conductivity 0.0032
PAR 0.0053
Nitrates 0.0219
Full model 0.999 0.0029
Primary production in a semiarid stream 1415
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

However, epilithon GPP and CR only showed a
positive correlation with the suspended solid concen-
tration (r ¼ 0.87, P £ 0.05, and r ¼ 0.78, P £ 0.05,
respectively). This independient variable explained
69% of epilithon production and 75% of variation in
its respiration rate, and it was the only predictor
variable in the multiple regression analysis (Table 4).
In general, the mean rates of gross primary pro-
duction and respiration of algal assemblages were
positively correlated (P £ 0.05) with mean monthly air
temperature (r ¼ 0.88 and r ¼ 0. 87, respectively), and
respiration was also negatively correlated with
monthly precipitation (r ¼ )0.79) and dissolved oxy-
gen (r ¼ )0.83). The results obtained by simple
regression analysis using the mean values for the
assemblages as a whole, indicated that 64% of the
variation in mean GPP and 71 % of the variation of
mean CR could be explained by the mean monthly air
temperature alone. For mean CR, a significant mul-
tiple regression model including, besides mean
monthly air temperature, conductivity, PAR and
nitrate concentration, explained 99.99% of its vari-
ation (Table 4).
Net ecosystem metabolism and GPP : CR24 ratio
Daily net metabolism in the reach was positive on all
the dates studied, ranging from 2.23 gC m)2 day)1 in
March to 16.61 gC m)2 day)1 in July, with a mean
value of 7.03 gC m)2 day)1 (Table 2).
The maximum rate of daily GPP was
26.41 gC m)2 day)1 in July, and the minimum
5.15 gC m)2 day)1 in December. The rates of daily
benthic respiration were much lower than those of
GPP, with values ranging from 2.67 to
10.22 gC m)2 day)1. Mean daily GPP and CR24 were
13.70 and 6.88 gC m)2 day)1, respectively. The
GPP : CR24 ratios on all dates were higher than 1,
with a mean value of 1.9, a maximum value of 2.7
being reached in July.
Discussion
The exceptionally high rates of primary production
found in Chicamo stream are only possible as a result
of high temperatures, high light availability and the
intensive internal recycling of nutrients. Mean
monthly air temperature was the best single predictor
of mean GPP and mean CR in Chicamo stream,
explaining more than 63 and 71% of their variation,
respectively. Morin et al. (1999), using multiple regres-
sion models fitted to predict the primary production
of stream periphyton, found that the production of
algae increased with both Chla standing crop and
water temperature, although Chla was most strongly
correlated with primary production, accounting for
65% of the variation, while temperature accounted for
a much smaller portion (4%) of the variability. Other
comparative studies of streams from different biomes
found that temperature explained 33 (Bott et al., 1985)
and 38% (Sinsabaugh, 1997) of the variation in R,
although Mulholland et al. (2001) found no evidence
for any effect of water temperature on R or GPP. On a
local-scale, Uehlinger, Konig & Reichert (2000)6 during
one annual cycle of a Swiss river, found that R was
significantly related to water temperature, although
temperature explained only 22% of its variation.
Temperature interacts with other environmental fac-
tors, such as nutrients, gases, metabolites, develop-
ment stage, trophic interactions and, especially, light,
although the individual effect of each factor is not
clear (DeNicola, 1996). In Chicamo stream, high
temperatures were associated with high irradiance,
low or null precipitation, low discharge, high con-
ductivity, low dissolved oxygen levels and high
ammonium concentrations.
Although photoinhibition usually ocurrs at irradi-
ances greater than 600 lmol m)2 s)1 (Hill, 1996), in
our study, exposure of the experimental chambers to
higher natural light levels did not appear to cause any
photoinhibition. Regarding photosynthesis-irradiance
responses, benthic algae inhabiting ‘‘high-light’’ envi-
ronments (such as open sites in clear streams) are
probably under considerable selective pressure to
develop mechanisms that reduce the potentially
damaging effects of high irradiances, whether in the
form of accessory pigments (carotenids) or sheath
pigments such as scytonemin (Garcia-Pichel &
Castenholz, 1991). On the other hand, high irradiance
at the surface of benthic algal mats may inhibit
photosynthesis by surface cells, while shaded subsur-
face cells receive only saturating or subsaturating
levels of irradiance. Photosynthesis by underlayers at
high surface irradiance may compensate for inhibition
in surface layers, so there is no evidence of photoin-
hibition at community level (Hill, 1996).
In our study, PAR was a secondary predictor
variable of both epipelic GPP and mean CR in the
1416 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420
Ta
ble
5S
um
mar
yo
fd
aily
gro
ssp
rim
ary
pro
du
ctio
n(G
PP
ing
Cm
)2
day
)1)
and
dai
lyco
mm
un
ity
resp
irat
ion
(CR
ing
Cm
)2
day
)1)
fro
mst
ream
so
fd
iffe
ren
td
eser
tan
d
Med
iter
ran
ean
area
s
Stu
dy
area
GP
PC
RC
om
mu
nit
yS
tud
yp
erio
dM
eth
od
Ref
eren
ce
Sy
cam
ore
Cre
ek(S
C),
Ho
td
eser
tst
ream
,A
Z,
U.S
.A.
4.68
2.50
Wh
ole
com
mu
nit
y27
May
1997
Tw
o-s
tati
on
diu
rnal
ox
yg
ench
ang
e
Mu
lho
llan
det
al.,
2001
Dee
pC
reek
(DC
),
Co
ol
des
ert
stre
am,
ID,
U.S
.A.
1.18
1.15
Mac
rop
hy
tes
and
per
iph
yto
nA
nn
ual
aver
age
(mo
nth
lym
easu
res)
Tw
o-s
tati
on
diu
rnal
ox
yg
ench
ang
e
Min
shal
l,19
78
Rat
tles
nak
eC
reek
(RC
),
Co
ol
des
ert
stre
am,
WA
,U
.S.A
.
9.3
8.1
Wh
ole
com
mu
nit
y
(per
iph
yto
nan
dw
ater
cres
s)
An
nu
alav
erag
e
(mo
nth
lym
easu
res)
Tw
o-s
tati
on
die
l
curv
e-p
H-C
O2
Cu
shin
g&
Wo
lf,
1984
Tec
op
aB
ore
(TB
),
Th
erm
ald
eser
tsp
rin
g,
CA
,U
.S.A
.
3.25
Alg
alm
ats
An
nu
alav
erag
e
(mo
nth
lym
easu
res)
14C
up
tak
eN
aim
an,
1976
Rie
raM
ajo
r(R
M),
Med
iter
ran
ean
stre
am,
Sp
ain
0.11
0.08
Ep
hil
itic
alg
aeA
nn
ual
aver
age
(mo
nth
lym
easu
res)
Ph
oto
syn
thes
is-i
rrad
ian
cecu
rve
Gu
asch
&S
abat
er,
1998
La
So
lan
a(L
S),
Med
iter
ran
ean
stre
am,
Sp
ain
0.27
0.23
Ep
hil
itic
alg
aeA
nn
ual
aver
age
(mo
nth
lym
easu
res)
Ph
oto
syn
thes
is-i
rrad
ian
cecu
rve
Gu
asch
&S
abat
er,
1998
Mo
nte
sin
a(M
),
Med
iter
ran
ean
stre
am,
Sp
ain
0.71
0.69
Mac
rop
hy
tes
and
per
iph
yto
nF
ou
rd
ates
aver
age
(J,
M,
M,
J)
Ox
yg
ench
ang
ein
cham
ber
sM
oll
aet
al.,
1994
16
Ch
icam
o(C
H),
Sem
iari
d
Med
iter
ran
ean
stre
am,
Sp
ain
1.27
1.18
Wh
ole
com
mu
nit
yF
ou
rd
ates
aver
age
(J,
N,
F,
J)
On
e-st
atio
nd
iurn
alo
xy
gen
chan
ge
Su
arez
&V
idal
-Ab
arca
,20
0017
13.7
6.88
Cha
ravu
lgar
isS
ixd
ates
aver
age
(J,
A,
O,
D,
M,
M)
Ox
yg
ench
ang
ein
cham
ber
sT
his
stu
dy
Ep
hil
itic
alg
ae
Ep
ipel
ical
gae
Primary production in a semiarid stream 1417
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420
multiple regression models. Of the nutrients exam-
ined, only the ammonium concentration was signifi-
cantly related to C. vulgaris GPP and the nitrate and
ammonium concentration to epipelic GPP.
Measurements of GPP and CR showed within-year
differences between assemblages that reflected the
phenological characteristics of the primary producers
studied in relation to seasonal variations of tempera-
ture and precipitation. In addition, proximate factors
such as resource availability, ecophysiology, life his-
tory characteristics, stress, allelophathy, competition
and predation, are causes of differential species
performance (Stevenson, 1996). In unshaded streams,
the flood disturbance regime is perhaps the funda-
mental factor determining habitat suitability and the
spatial and temporal patterns of benthic algae, and is
a major biomass loss mechanism (Biggs, 1996). Floods
in the Chicamo stream increase the habitat suitability
for epilithon and nutrient availability in the water
column, but have a negative effect on the production
of C. vulgaris which is buried by transported sedi-
ment. Although epipelic algae biomass was largely
removed by flooding, recolonisation was rapid and
biomass and production values returned to predis-
turbance levels in less than one month. The availab-
ility of algal propagules and rapid growth rates make
epipelic algae resilient in the face of flooding.
In Chicamo stream, the greater extent of epipelic
assemblages, the high abundance of gathering collec-
tors and the low diversity of algal grazers (Martınez
et al. 19987 ) suggest that neither the macrophyte
C. vulgaris nor epilithic algae are an important food
source, although epipelic algae with detritus derived
from sloughing periphyton are. High rates of primary
production in Chicamo stream support correspond-
ingly high rates of secondary production of aquatic
macroinvertebrates. The annual production estimates
of the gathering-collector Caenis luctuosa presented the
highest value ever reported for Caenis species
(6.35 gDW m)2 year)1; Peran, Velasco & Millan,
1999).
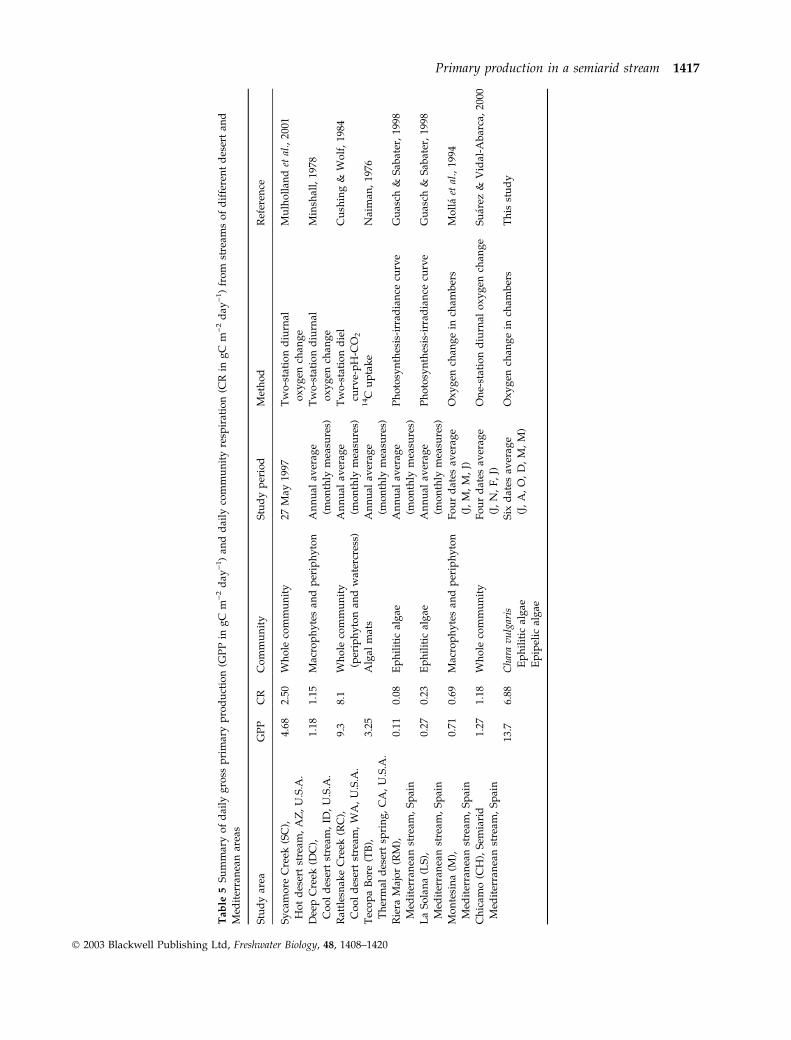
A comparison of the primary production values
obtained in this study with those reported for other
desert and Mediterranean streams (Table 5) shows
that our rates of GPP were higher than those reported
earlier from other streams, including desert streams
which had higher GPP than a variety of streams from
different biomes and locations throughout the world
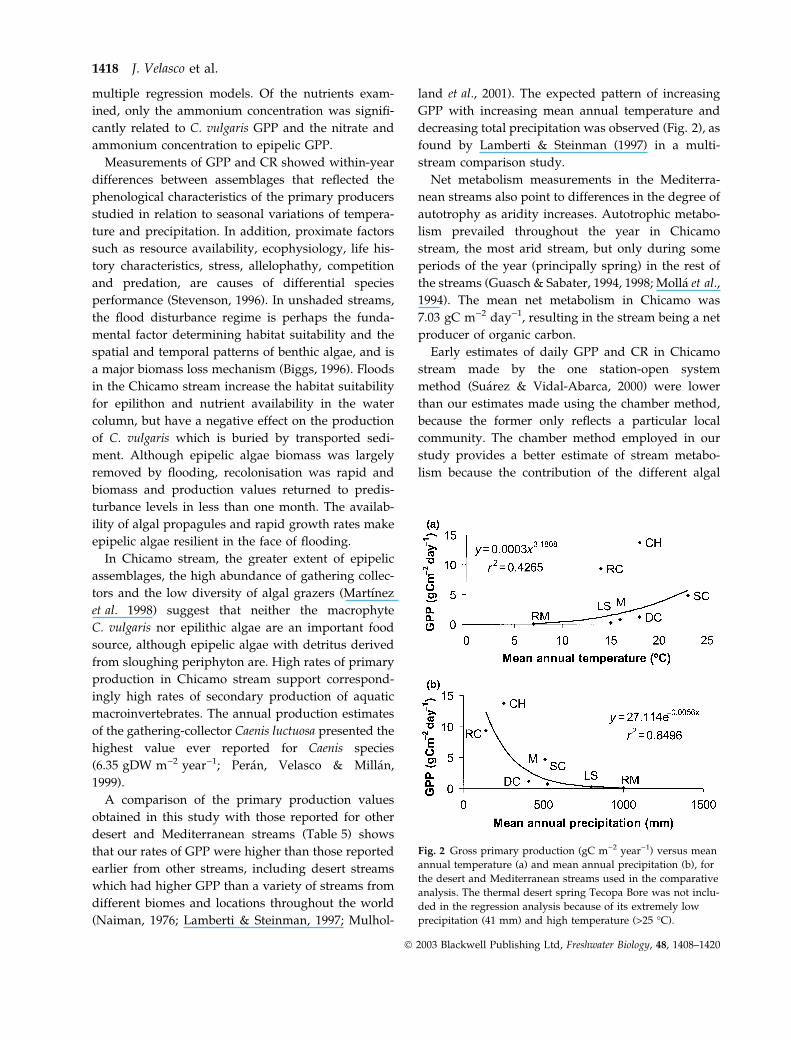
(Naiman, 19768 ; Lamberti & Steinman, 1997; Mulhol-
land et al., 2001). The expected pattern of increasing
GPP with increasing mean annual temperature and
decreasing total precipitation was observed (Fig. 2), as
found by Lamberti & Steinman (1997) in a multi-
stream comparison study.
Net metabolism measurements in the Mediterra-
nean streams also point to differences in the degree of
autotrophy as aridity increases. Autotrophic metabo-
lism prevailed throughout the year in Chicamo
stream, the most arid stream, but only during some
periods of the year (principally spring) in the rest of
the streams (Guasch & Sabater, 1994, 1998; Molla et al.,
1994). The mean net metabolism in Chicamo was
7.03 gC m)2 day)1, resulting in the stream being a net
producer of organic carbon.
Early estimates of daily GPP and CR in Chicamo
stream made by the one station-open system
method (Suarez & Vidal-Abarca, 2000) were lower
than our estimates made using the chamber method,
because the former only reflects a particular local
community. The chamber method employed in our
study provides a better estimate of stream metabo-
lism because the contribution of the different algal
Fig. 2 Gross primary production (gC m)2 year)1) versus mean
annual temperature (a) and mean annual precipitation (b), for
the desert and Mediterranean streams used in the comparative
analysis. The thermal desert spring Tecopa Bore was not inclu-
ded in the regression analysis because of its extremely low
precipitation (41 mm) and high temperature (>25 �C).
1418 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

assemblages present is recorded and can be extra-
polated to the total reach. Bott et al. (1978), in a
comparison of methods for measuring primary
productivity and community respiration in streams,
found that open methods provided lower NPP
estimates than the chamber methods. Therefore,
inter and intra stream differences should be inter-
preted with caution because comparative analyses of
primary production are difficult because of the large
differences in analytical methods, conversion factors,
communities and sampling frequencies employed
(Wetzel & Ward, 1992).
In conclusion, Chicamo stream is a net producer
of organic carbon, with autotrophic metabolism
throughout the year, epipelic algae being the most
important primary producer in the stream. Tem-
perature dominates the control of mean GPP and
mean CR in the stream, perhaps because it integ-
rates many features relevant to primary production.
In addition, the frequency and intensity of flood
disturbance has a considerable influence on the
biomass and production of different algal assem-
blages. Nutrient concentrations, especially of ammo-
nium and nitrates, appeared to be a secondary
determinant of GPP.
Thus, Chicamo stream resembles desert and prairie
streams as regards the dominant role of autochtho-
nous organic matter as the base of the food web.
Acknowledgments
We thank J.L. Moreno, B. Martınez, A. Peran and A.
Mellado for sampling assistance and D. Ros for algal
identification. Comments made on a early version of
the manuscript by Professor R.I. Jones and two
anonymous reviewers have enabled us to improved
the paper. This research was supported by Projectt
PB96-1113 (National Program CICYT).
References
American Public Health Association (APHA). (1992)
Standard Methods for the Examination of Water and
Wastewater. American Public Health Association,
Washington, DC.
Biggs B.J.F. (1995) The contribution of flood disturbance,
catchment geology and land use to the habitat tem-
plate of periphyton in stream ecosystems. Freshwater
Biology, 33, 419–438.
Biggs B.J.F. (1996) Patterns in Benthic Algae of
Streams. In: Algal Ecology (Eds R.J. Stevenson, M.L.
Bothwell & R.L. Lowe), pp. 31–56. Academic Press,
San Diego, CA.9
Bott T.L. (1997a10 ) Primary Productivity and Community
Respiration. In: Methods in Stream Ecology (Eds F.R.
Hauer & G.A. Lamberti), pp. 533–556. Academic Press,
San Diego, CA.
Bott T.L., Brock J.T., Cushing C.E., Gregory S.V., King D.
& Petersen R.C. (1978) A comparison of methods for
measuring primary productivity and community
respiration in streams. Hydrobiologia, 60, 3–12.
Bott T.L., Brock J.T., Dunn C.S., Naiman R.J., Ovink R.W.
& Petersen R.C. (1985) Benthic community metabolism
in four temperate stream systems: an inter-biome
comparison and evaluation of the river continuum
concept. Hydrobiologia, 123, 3–45.
Bott T.L., Brock J.T., Baattrup-Pedersen A., Chambers
P.A., Dodds W.K., Himbeault K.T., Lawrence J.R.,
Planas D., Snyder E. & Wolfaardt G.M. (1997b11 ) An
evaluation of techniques for measuring periphyton
metabolism in chambers. Canadian Journal of Fisheries
and Aquatic Sciences, 57, 715–725.
Busch D.E. & Fisher S.G. (1981) Metabolism of a desert
stream. Freshwater Biology, 11, 301–307.
Cushing C.E. & Wolf E.G. (1984) Primary production in
Rattlesnake Springs, a cold desert spring-stream.
Hydrobiologia, 114, 229–236.
DeNicola D.M. (1996) Periphyton responses to tempera-
ture at different eccological levels. In: Algal Ecology (Eds
R.J. Stevenson, M.L. Bothwell & R.L. Lowe), pp. 150–
182. Academic Press, San Diego, CA12 .
Garcia-Pichel F. & Castenholz R.W. (1991) Characteriza-
tion and biological implications of scytonemin, a
cyanobacterial sheath pigment. Journal of Phycology,
27, 395–409.
Gasith A. & Resh V.H. (1999) Streams in Mediterranean
climate regions: abiotic influences and biotic responses
to predictable seasonal events. Annual Review of Ecology
and Systematic, 30, 51–81.
Grimm N.B. & Fisher S.G. (1984) Exchange between
interstitial and surface water: implications for stream
metabolism and nutrient cycling. Hydrobiologia, 111,
219–228.
Guasch H. & Sabater S. (1994) Primary production of
epilithic communities in undisturbed Mediterranean
streams. Verhandlungen Internationale Vereinigung fur
Theorische und Angewandte Limnologie, 25, 1761–1764.
Guasch H. & Sabater S. (1998) Estimation of the annual
primary production of stream epilithic biofilms based
on photosynthesis-irradiance relations. Archiv fur Hy-
drobiologie, 141, 469–481.
Primary production in a semiarid stream 1419
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420

Hill W. (1996) Effects of Light. In: Algal Ecology (Eds R.J.
Stevenson, M.L. Bothwell & R.L. Lowe), pp. 121–148.
Academic Press, San Diego, CA13 .
Lamberti G.A. & Steinman A.D. (1997) A comparison of
primary production in stream ecosystems. In: Stream
organic matter budgets (Eds J.R. Webster & J.L. Meyer),
pp. 95–104. Journal of the North American Benthological
Society, 16, 3–161.14
Martınez B., Velasco J., Suarez M.L. & Vidal-Abarca M.R.
(1998) Benthic organic matter dynamics in an inter-
mittent stream in South-East Spain. Archiv fur Hydro-
biologie, 141, 303–320.
Minshall G.W. (1978) Autotrophy in stream ecosystems.
BioScience, 28, 767–771.
Molla S., Malchik L. & Casado C. (1994) Primeros
datos sobre el metabolismo de un arroyo temporal
mediterraneo de Sierra Morena (Cordoba). Limnetica,
10, 59–67.
Morin A., Lamoureux W. & Busnarda J. (1999) Empirical
models predicting primary productivity from chlor-
ophyll a and water temperature for stream periphyton
and lake and ocean phytoplankton. Journal of the North
American Benthological Society, 18, 299–307.
Mulholland P.J., Fellows, C.S., Tank J.L. et al. (2001) Inter-
biome comparison of factors controlling stream meta-
bolism. Freshwater Biology, 46, 1503–1517.
Naiman R.J. (1976) Primary production, standing crop,
and export of organic matter in a Mohave Desert
thermal stream. Limnology and Oceanography, 21, 60–
73.
Peran A., Velasco J. & Millan A. (1999) Life cycle and
secondary production of Caenis luctuosa (Ephemerop-
tera) in a semiarid stream (Southeast Spain). Hydro-
biologia, 400, 187–194.
Sinsabaugh R.L. (1997) Large-scale trends for stream
benthic respiration. In: Stream organic matter budgets
(Eds J.R. Webster & J.L. Meyer), pp. 119–122. Journal of
the North American Benthological Society, 16, 3–161.
Stevenson R.J. (1996) An introduction to algal ecology in
freshwater benthic habitats. In: Algal Ecology (Eds R.J.
Stevenson, M.L. Bothwell & R.L. Lowe), pp. 3–30.
Academic Press, San Diego, CA.15
Suarez M.L. & Vidal-Abarca M.R. (2000) Metabolism of a
semi-arid stream of south-east Spain. Verhandlungen
Internationale Vereinigung fur Theorische und Angewandte
Limnologie, 27, 756–761.
UNESCO (1979) Map of the World Distribution of Arid
Regions. MAB Technical Note 7, Paris.
Uehlinger U., Konig C. & Reichert P. (2000). Variability of
photosynthesis-irradiance curves and ecosystem
respiration in a small river. Freshwater Biology, 44,
493–507.
Vidal-Abarca M.R., Suarez M.L., Moreno J.L., Gomez, R.
& Sanchez, I. (2000) Hidroquımica de un rıo de
caracterısticas semiaridas (Rıo Chıcamo, Murcia).
Analisis espacio-temporal. Limnetica, 18, 57–73.
Vidal-Abarca M.R., Suarez M.L., Guerrero C., Velasco J.,
Moreno J.L., Millan M. & Peran A. (2001) Dynamics of
dissolved and particulate organic carbon in a saline
and semiarid stream of southeast Spain (Chicamo
stream). Hydrobiologia, 455, 71–78
Vollenweider R.A. (1974) A Manual of Methods for Meas-
uring Primary Production in Aquatic Environments. IBP
Handbook 12 (2nd edn.), Blackwell, Oxford, UK.
Wetzel R.G. & Ward A.K. (1992) Primary production. In:
The Rivers Handbook (Eds P. Calow & G.E. Petts), pp.
354–369, Blackwell Science, Oxford.
Wilkinson L. (1996) Systat 6 for Windows. SPPP Inc.,
Chicago, USA.
(Manuscript accepted 8 May 2003)
1420 J. Velasco et al.
� 2003 Blackwell Publishing Ltd, Freshwater Biology, 48, 1408–1420




























