Embed Size (px)
Citation preview

www.elsevier.com/locate/jneuroim
Journal of Neuroimmunolo
Mimotopes of conformational epitopes in fibrillar h-amyloid
Goar Gevorkiana,1, Irina Petrushinab,1, Karen Manoutchariana, Anahit Ghochikyanc,
Gonzalo Aceroa, Vitaly Vasilevkob, David H. Cribbsb,2, Michael G. Agadjanyanc,*,2
aInstituto de investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, MexicobThe Institute for Brain Aging and Dementia, University of California Irvine, Irvine, CA 92697, United States
cDepartment of Immunology, The Institute for Molecular Medicine, 16371 Gothard Street, Huntington Beach, CA 92649, United States
Received 21 April 2004; received in revised form 7 June 2004; accepted 7 June 2004
Abstract
In Alzheimer’s disease (AD) h-amyloid peptide accumulates in the brain in different forms including fibrils. Amyloid fibrils could be
recognized as foreign by the mature immune system since they are not present during its development. Thus, using mouse antisera raised
against the fibrillar form of Ah42, we have screened two phage peptide libraries for the presence of foreign conformational mimotopes of Ah.Antisera from wild type animals recognized predominately peptides with the EFRH motif from Ah42 sequence, whereas amyloid precursor
protein (APP) transgenic mice recognized mainly phage clones that mimic epitopes (mimotopes) within the fibrillar Ah42 but lack sequence
homology with this peptide.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Antibodies; Transgenic mice; Antigens; Epitopes; Mimotopes; Immunotherapy
1. Introduction
Alzheimer’s disease (AD) is the most common form
of0 dementia in the elderly and is characterized by two
distinct neuropathological lesions in the brain: extracel-
lular senile plaques and intracellular neurofibrillary tangles
(NFT). The plaques are composed primarily of the h-amyloid peptide (Ah), whereas the NFT are composed of
hyperphosphorylated tau protein (Selkoe, 2001). The Ahpeptide is the product of the proteolytic processing of the
amyloid precursor protein (APP), and there is now a
general consensus that this highly amyloidogenic peptide
is a critical element in the onset and progression of AD
(Hardy and Selkoe, 2002). Thus, reducing the production
or enhancing the degradation and clearance of this peptide
has been a major focus of many laboratories. However,
0165-5728/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jneuroim.2004.06.004
* Corresponding author. Tel.: +1 714 596 7821; fax: +1 714 596 3791
E-mail address: [email protected] (M.G. Agadjanyan).1 Both authors contributed equally to this work.2 These senior authors contributed equally to this work.
.
key tenets of the bamyloid hypothesisQ of AD have
evolved over the last 12 years and important tests of the
hypothesis are now underway in animal models and
humans (Hardy, 2002; Hardy and Selkoe, 2002). In
particular, Ah-immunotherapy has received considerable
attention as one possible strategy for reducing Ah levels
in the brain (Monsonego and Weiner, 2003; Schenk,
2002; Weiner and Selkoe, 2002). The seminal report by
Schenk et al., 1999, showing that active immunization of
APP transgenic (APP/Tg) mice with fibrillar Ah peptide
in combination with a powerful adjuvant induced anti-
bodies specific for Ah that promoted the clearance of Ahfrom the mouse brain, initiated a new field of study in
AD.
Subsequent studies using passive transfer of anti-Ahantibodies also reduced Ah plaque deposition, neuritic
dystrophy, astrogliosis in the brains of APP/Tg mice, and
diminished learning deficits in mice confirmed the role of
the anti-Ah antibodies in the clearance of Ah (Bard et al.,
2000; DeMattos et al., 2001; Janus et al., 2000; Morgan
et al., 2000; Schenk, 2002). A number of groups have
demonstrated that active immunization induces antibodies
gy 156 (2004) 10–20

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–20 11
that preferentially recognize a linear epitope in the N-
terminal region of Ah42 (Cribbs et al., 2003; Dickey et
al., 2001; Lemere et al., 2000, 2001; Town et al., 2001).
These results have been confirmed using proteolytic
cleavage of intact, immobilized immune complexes
consisting of polyclonal antibodies from APP/Tg mice
bound to Ah42, followed by high-resolution mass-spec-
trometry. In this report, the linear Ah42 epitope spanning
residues 4–10 was identified as the sequence binding to
the antibodies (McLaurin et al., 2002). Previously,
monoclonal antibodies to residues 3EFRH6 within the
Ah42 sequence were shown to be effective in disaggre-
gation of fibrillar Ah42 in vitro (Solomon et al., 1996,
1997). Thus, the major linear B cell antigenic epitope in
Ah42 is within the sequence containing residues 1–10.
However, there is considerable evidence that fibrillar
Ah42 possesses conformational epitopes that can induce a
humoral response in humans as wells as mice. Gaskin et
al. reported that four human monoclonal IgM antibodies
derived from one AD patient recognized conformational
antigenic determinants in fibrillar Ah42. This antigenic
epitope/s was conserved in amyloid plaques in situ, which
indicates that humans may recognize this unique con-
formational antigenic determinant/s in fibrillar Ah42 as
non-self (Gaskin et al., 1993b). Recently, two conforma-
tion-specific mouse monoclonal IgM antibodies against
different disease-related amyloid fibrils including Ah40
fibrils were characterized (O’Nuallain and Wetzel, 2002),
although binding of these antibodies to amyloid plaques
in brain tissue from AD patients has not yet been
demonstrated. More recently, polyclonal rabbit antibodies
to the fibrillar Ah42 have been generated, and these
antibodies bound to the fibrillar full-length Ah. Although
the same antisera were capable of binding to linear Ah1–8
sequence, the affinity of binding to fibrillar Ah42 was
almost 1000 times higher. These authors suggested that
the anti-fibrillar antibodies might preferentially bind to the
conformation of Ah1–8 that is stabilized in fibrillar form
of Ah42 (Miller et al., 2003). These results collectively
suggest that the full-length fibrillar Ah42 contains
conformational antigenic determinants that are not formed
by the shorter linear peptides.
In the current study we assayed for conformational B
cell epitopes within the fibrillar form of Ah42 using anti-
Ah42 antisera raised in APP/Tg or wild type mice with
the help of two phage display peptide libraries. We report
that in APP/Tg mice immunization with fibrillar Ah42
generates antibodies that recognize not only the linear
EFRH epitope of this antigen but also a number of
mimotopes expressed by phage clones. One such linear
mimotope expressed by several phage clones was
synthesized, and binding to anti-Ah42 antibodies was
confirmed. Interestingly, anti-Ah42 antisera generated in
immunized wild type mice recognized predominately
linear peptides with the EFRH motif from the Ah42
sequence.
2. Materials and methods
2.1. Preparation of peptides
All peptides were synthesized at the Peptide Core
Facility at the Institute for Brain Aging and Dementia at
UCI. We routinely obtain very high quality Ah synthetic
peptides (N99% purity). Peptides were synthesized by the
solid-phase Fmoc method, purified by reverse-phase
HPLC, as previously described (Cribbs et al., 1997,
2003). h-Amyloid peptides 1–42 (Ah42), 1–40 (Ah1–40),
1–28 (Ah1–28), and 25–35 (Ah25–35), as well as 15- and 17-
mer overlapping small peptides spanning aa 1–15 (Ah1–15),
6–20 (Ah6–20), 11–25 (Ah11–25), 16–30 (Ah16–30), 21–35
(Ah21–35), and 26–42 (Ah26–42) of Ah were prepared. In
addition, we synthesized (Multiple Peptide Systems, San
Diego, CA) high quality (N99% purity) 11- and 16-mer
synthetic mimotopic antigens (MAG) GDLKRTTGGGC
(MAG11) and LDLKRTTTTHTSGGGC (MAG16). Both
MAG peptides were based on data generated from screen-
ing of phage display libraries against anti-Ah42 sera
produced in APP/Tg mice (see Results).
In addition, we conjugated MAG11 to Imject Malei-
mide-Activated Ovalbumin (OVA) to obtain MAG11–
OVA. Two milligrams of peptide MAG11 was conjugated
with powdered Imject Maleimide-Activated Ovalbumin,
and the conjugate was purified by gel filtration according
to the manufacturer’s protocol (Pierce, Rockford, IL).
Briefly, 10 fractions of eluate were collected and the
amount of protein in each fraction was tested by Bio-Rad
protein assay (Bio-Rad, Hercules, CA). Two fractions of
the eluate showing the highest presence of protein were
combined and stored at 4 8C.
2.2. Preparation of antigen and immunization of mice
Six- to eight-week-old APP/Tg 2576 {(H2bxs) UCI
Transgenic Mouse Facility)}, BALB/c (H2d), C57Bl/6
(H2b), C3H (H-2k), and F1B6SJL (H2bxs) mice (Jackson
Laboratories, Bar Harbor, ME) were housed in the animal
facility at UCI in a temperature and light-cycle controlled
facility, and their care was under the guidelines of the NIH
and UCI. Each mouse was injected subcutaneously (s.c.)
with 100 Ag of fibrillar Ah42 formulated in Th2-type
adjuvant Alum (Cribbs et al., 2003). The protocol for
preparation of fibrillar Ah42 was adapted from Kayed et al.
(2003). Briefly, we dissolved lyophilized Ah42 peptide in
1,1,1,3,3,3-hexafluoro-2-propanol (HFIP, Sigma) and incu-
bated it at room temperature, 15 min. The seedless Ahsolution was added to ddH2O (V/V=1:10) and after addi-
tional 15 min incubation at room temperature the samples
were centrifuged (14,000�g, 15 min). The supernatant
fraction was transferred to a siliconized tube and subjected
to a gentle stream of nitrogen to evaporate the HFIP (10
min). The samples were then stirred at 500 rpm using a
Teflon coated micro stir bar for 9 days. Fibril formation was

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–2012
monitored by thioflavin T fluorescence and UV light
scattering. The solutions were centrifuged at 14,000�g for
20 min, the fibril pellet was washed 3� with the ddH20, and
then resuspended in the desired buffer. The morphology was
verified by AFM or negative stain EM. After the first
immunization, two boosts were performed at 2-week
intervals with the same peptide. Mice from each exper-
imental group (six mice of APP/Tg, six C57BL/6, six C3H,
eight F1B6SJL and twelve BALB/c) were bled 8 days after
last boost, and individual sera were used to detect the
presence of specific anti-Ah42 antibodies by ELISA and/or
for screening of the phage display peptide libraries.
2.3. Detection of the titer of anti-Ab42 antibodies and
peptide competition assay
The same fibrillar Ah42 used for immunization was used
for ELISA. Binding of polyclonal sera to fibrillar Ah42 was
detected as previously described (Kayed et al., 2003; Cribbs
et al., 2003). Briefly, fibrillar Ah42 peptide was diluted in
coating buffer (0.1 M sodium bicarbonate, pH 9.6) to 2.5
AM concentration, and 96 wells plates were coated at 37 8Cwithin only 2 h (the fibril formation was checked by
thioflavin T fluorescence). Plates were then washed,
blocked, and primary sera from experimental and control
mice were added in duplicate at an initial dilution of 1:1000,
or as indicated, and diluted serially in Tween-20 Tris Buffer
Solution (TTBS) to 1:64,000. After incubation and washing,
anti-mouse IgG (whole molecule) conjugated with horse-
radish peroxidase (HRP) was added as recommended by
manufacturer (Jackson Laboratories). Plates were incubated,
washed, and OPD substrate solution (o-phenylenediamine
in 0.05 M phosphate–citrate buffer, pH 5.0, Sigma) was
added to develop the reaction. All plates were analyzed
spectrophotometrically at 405 nm.
Using anti-Ah42 antisera generated in APP/Tg animals
and mice of different haplotypes, we also analyzed binding
of antibodies to fibrillar Ah42 in the presence of small
overlapping peptides Ah1–15, Ah6–20, Ah11–25, Ah16–30,
Ah21–35, and Ah26–42 of Ah42. In this competition ELISA
we also used peptides spanning Ah1–28, Ah25–35 and
fibrillar Ah42. Briefly, plates coated with fibrillar Ah42
were washed and blocked, as described above. Antisera
from immunized and control mice, at approximately the
equal concentrations, were mixed with small overlapping
peptides, Ah1–28, Ah25–35 or with control fibrillar Ah42
peptide (all peptides at final concentration from 0.019 to 5
AM as indicated in Figs. 2 and 3 legends). After incubation
(1 h, 37 8C) the peptide-treated antisera were added in
duplicate to Ah42-coated plates. Using secondary anti-
mouse IgG-HRP antibodies and OPD substrate the reaction
was developed, the plates were analyzed for binding as
described above, and the percent of inhibition by small
peptides, as well as by control peptides were calculated
using binding of sera without competing peptides to fibrillar
Ah42 as 100% (Cribbs et al., 2003).
2.4. Binding of anti-Ab42 antibodies to mimotopes MAG11
and MAG16
In order to correctly compare the levels of binding of
anti-Ah42 antisera from APP/Tg and wild type mice to
MAG11 and MAG16 peptides, we first detected the concen-
tration of anti-Ah42 antibodies in the experimental sera.
Using 6E10 monoclonal antibody that recognizes 1–17aa of
Ah42 (Signet Labs, Dedham, MA), we generated a standard
curve and determined the anti-Ah42 antibody concentration
in the sera of immunized mice. Briefly, individual mouse
serum and the 6E10 antibody were incubated in the wells
coated with fibrillar Ah42. The plates were washed, and
secondary anti-mouse IgG-HRP antibodies were added
followed by OPD substrate solution. All plates were
analyzed spectrophotometrically, average dataFS.D. were
calculated for each group of mice, and results were recorded
in Ag per ml. Equal concentrations of antisera from wild
type and APP/Tg mice were used to detect binding of anti-
Ah42 antibodies to mimotopes. The same ELISA procedure
outlined above was used in these experiments, except
MAG11, MAG16, and MAG11–OVA were used for coating
the 96 wells plates (2.5AM peptide in bicarbonate coating
buffer, pH 9.7).
2.5. Screening of phage display peptide library with
polyclonal anti-Ab42 antibodies
Selection of phage by biopanning was performed
essentially as previously described (Gevorkian et al.,
2000; Gomez-Roman et al., 2002). Briefly, Maxi Sorp
microtiter plates (Nunc, Roskilde, Denmark) were coated
with goat anti-mouse IgG (Zymed, CA, USA) at a
concentration of 5 Ag/ml and blocked with PBS supple-
mented with 2% BSA (bovine serum albumin). After
washing, polyclonal anti-Ah42 antisera (1:200 dilution)
from wildtype or APP/Tg mice were added, and plates
were incubated for 1 h at 37 8C. Two phage display
peptide libraries (New England Biolabs, Beverly, MA,
USA) of linear heptapeptides and dodecapeptides were
screened against polyclonal anti-Ah42 sera raised in APP/
Tg mice, whereas sera generated in C57BL/6 mice were
used only with the heptapeptides library. In both libraries
random peptides were expressed at the N-terminal region
of the minor coat protein pIII of the M13 phage. After
washing, 1011 plaque-forming units (PFU) from each
library were added to the plates and incubated for 3 h
at 4 8C, followed by 1 h at room temperature. Non-
specific phages were washed off, and affinity-selected
phages were eluted by glycine–HCl (pH 2.2) and
neutralized by Tris–base. A total of three rounds of
biopanning were performed, and the eluate from each
round was amplified and used for each subsequent round
(second and third) or for characterization of clones (third
round). Individual phage clones were picked at random
from the plated eluates, and used in ELISA to detect the
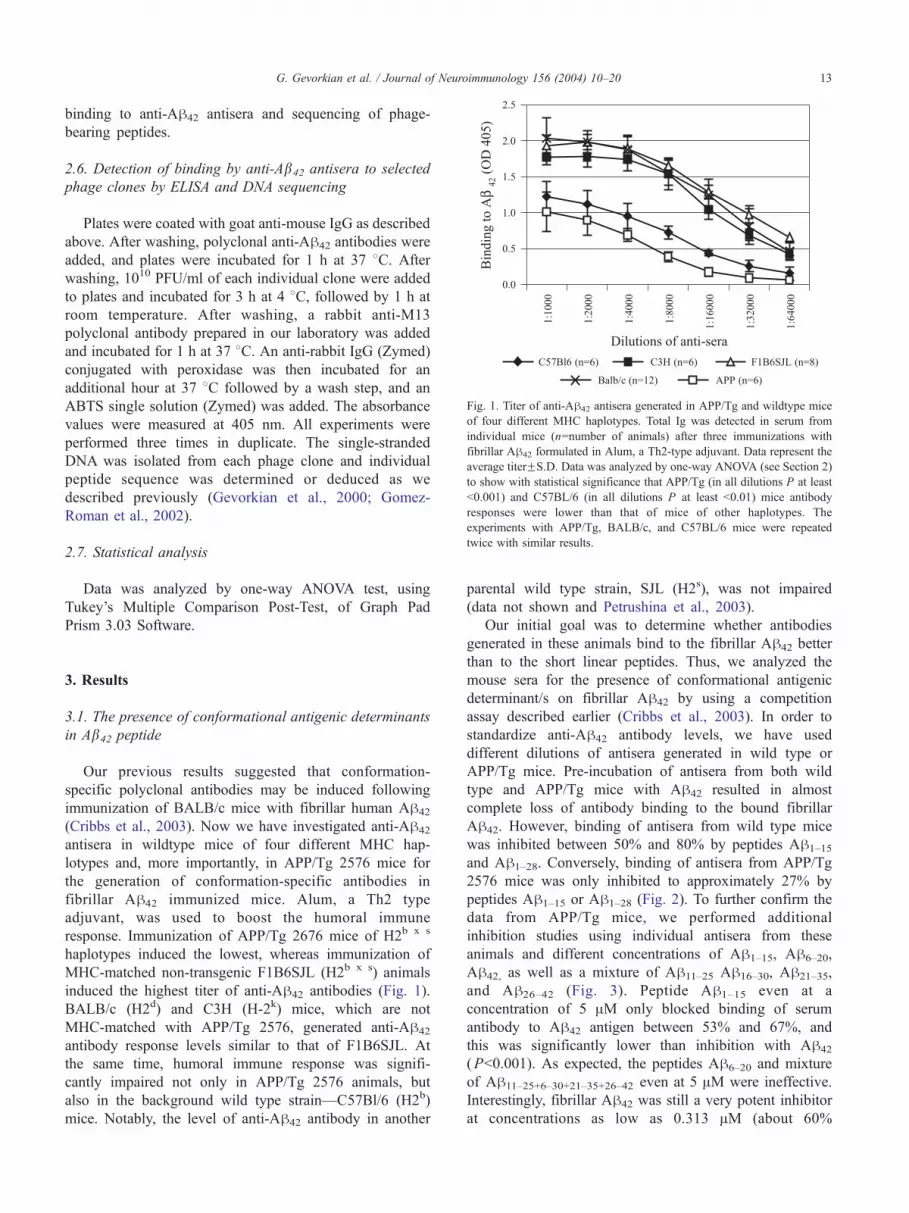
Fig. 1. Titer of anti-Ah42 antisera generated in APP/Tg and wildtype mice
of four different MHC haplotypes. Total Ig was detected in serum from
individual mice (n=number of animals) after three immunizations with
fibrillar Ah42 formulated in Alum, a Th2-type adjuvant. Data represent the
average titerFS.D. Data was analyzed by one-way ANOVA (see Section 2)
to show with statistical significance that APP/Tg (in all dilutions P at least
b0.001) and C57BL/6 (in all dilutions P at least b0.01) mice antibody
responses were lower than that of mice of other haplotypes. The
experiments with APP/Tg, BALB/c, and C57BL/6 mice were repeated
twice with similar results.
G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–20 13
binding to anti-Ah42 antisera and sequencing of phage-
bearing peptides.
2.6. Detection of binding by anti-Ab42 antisera to selected
phage clones by ELISA and DNA sequencing
Plates were coated with goat anti-mouse IgG as described
above. After washing, polyclonal anti-Ah42 antibodies were
added, and plates were incubated for 1 h at 37 8C. Afterwashing, 1010 PFU/ml of each individual clone were added
to plates and incubated for 3 h at 4 8C, followed by 1 h at
room temperature. After washing, a rabbit anti-M13
polyclonal antibody prepared in our laboratory was added
and incubated for 1 h at 37 8C. An anti-rabbit IgG (Zymed)
conjugated with peroxidase was then incubated for an
additional hour at 37 8C followed by a wash step, and an
ABTS single solution (Zymed) was added. The absorbance
values were measured at 405 nm. All experiments were
performed three times in duplicate. The single-stranded
DNA was isolated from each phage clone and individual
peptide sequence was determined or deduced as we
described previously (Gevorkian et al., 2000; Gomez-
Roman et al., 2002).
2.7. Statistical analysis
Data was analyzed by one-way ANOVA test, using
Tukey’s Multiple Comparison Post-Test, of Graph Pad
Prism 3.03 Software.
3. Results
3.1. The presence of conformational antigenic determinants
in Ab42 peptide
Our previous results suggested that conformation-
specific polyclonal antibodies may be induced following
immunization of BALB/c mice with fibrillar human Ah42
(Cribbs et al., 2003). Now we have investigated anti-Ah42
antisera in wildtype mice of four different MHC hap-
lotypes and, more importantly, in APP/Tg 2576 mice for
the generation of conformation-specific antibodies in
fibrillar Ah42 immunized mice. Alum, a Th2 type
adjuvant, was used to boost the humoral immune
response. Immunization of APP/Tg 2676 mice of H2b x s
haplotypes induced the lowest, whereas immunization of
MHC-matched non-transgenic F1B6SJL (H2b x s) animals
induced the highest titer of anti-Ah42 antibodies (Fig. 1).
BALB/c (H2d) and C3H (H-2k) mice, which are not
MHC-matched with APP/Tg 2576, generated anti-Ah42
antibody response levels similar to that of F1B6SJL. At
the same time, humoral immune response was signifi-
cantly impaired not only in APP/Tg 2576 animals, but
also in the background wild type strain—C57Bl/6 (H2b)
mice. Notably, the level of anti-Ah42 antibody in another
parental wild type strain, SJL (H2s), was not impaired
(data not shown and Petrushina et al., 2003).
Our initial goal was to determine whether antibodies
generated in these animals bind to the fibrillar Ah42 better
than to the short linear peptides. Thus, we analyzed the
mouse sera for the presence of conformational antigenic
determinant/s on fibrillar Ah42 by using a competition
assay described earlier (Cribbs et al., 2003). In order to
standardize anti-Ah42 antibody levels, we have used
different dilutions of antisera generated in wild type or
APP/Tg mice. Pre-incubation of antisera from both wild
type and APP/Tg mice with Ah42 resulted in almost
complete loss of antibody binding to the bound fibrillar
Ah42. However, binding of antisera from wild type mice
was inhibited between 50% and 80% by peptides Ah1–15
and Ah1–28. Conversely, binding of antisera from APP/Tg
2576 mice was only inhibited to approximately 27% by
peptides Ah1–15 or Ah1–28 (Fig. 2). To further confirm the
data from APP/Tg mice, we performed additional
inhibition studies using individual antisera from these
animals and different concentrations of Ah1–15, Ah6–20,
Ah42, as well as a mixture of Ah11–25 Ah16–30, Ah21–35,
and Ah26–42 (Fig. 3). Peptide Ah1–15 even at a
concentration of 5 AM only blocked binding of serum
antibody to Ah42 antigen between 53% and 67%, and
this was significantly lower than inhibition with Ah42
(Pb0.001). As expected, the peptides Ah6–20 and mixture
of Ah11–25+6–30+21–35+26–42 even at 5 AM were ineffective.
Interestingly, fibrillar Ah42 was still a very potent inhibitor
at concentrations as low as 0.313 AM (about 60%

Fig. 3. Conformation-specific polyclonal anti-Ah42 antibodies in the
immune sera from individual APP/Tg mice. Inhibition studies were
performed with indicated concentrations of Ah1–15, Ah6–20,
Ah11–25+16–30+21–35+26–42, and Ah42 peptides and anti-Ah42 sera.
Averaged data with antisera at dilution of 1:4000FS.D. are presented
(n=6). The ANOVA analysis (see Section 2) showed with statistical
significance that linear Ah1–15 peptide inhibited significantly weaker than
fibrillar Ah42 peptide ( Pb0.001 at concentrations 5–0.313 AM; Pb0.05 at
concentrations 0.078 AM; and Pb0.01 at concentrations 0.019 AM).
Fig. 2. The presence of conformation-specific polyclonal anti-Ah42 antibodies in the sera of Ah42 immunized APP/Tg 2576 mice and wild type animals of
different haplotypes. An ELISAwas performed to measure the ability of six small overlapping peptides, as well as Ah1–28, Ah25–35, and Ah42 peptides (all at
final concentration 2.5 AM) to inhibit the binding of polyclonal anti-Ah42 antibodies to Ah42 (concentrations of pooled sera are indicated in parenthesis). The
inhibitory potency of both Ah1–28 and Ah1–15 were similar, whereas the inhibition with Ah42 was significantly higher, especially with sera from APP/Tg mice.
These experiments were repeated twice with similar results.
G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–2014
inhibition), while Ah1–15 was virtually inactive at this
concentration (Pb0.001). The results from competition
assays argue for the presence of conformation-specific
polyclonal anti-Ah42 antibodies generated in wild type
mice of different haplotypes and at much higher levels in
APP/Tg 2576 animals.
3.2. Screening of phage display libraries against polyclonal
anti-Ab42 antibodies raised in wild type mice
In order to identify the linear peptides that mimic
conformational epitope/s possessed in fibrillar Ah42 peptide,
we used the well-characterized phage display peptide library
technique. Phage display peptide libraries provide an
excellent source of a large number of epitopes and have
been widely used in studies on antigen–antibody binding
(Manoutcharian et al., 2001; Mertens et al., 2001; Scott and
Smith, 1990). Accordingly, a phage display library of linear
heptapeptides was screened against polyclonal anti-Ah42
sera raised in C57BL/6 mice, the weakest responder among
wild type lines. Three rounds of biopanning were per-
formed, and enrichment of affinity-selected clones was
selected in each round. After the third round of biopanning,
22 plaques were randomly picked and amplified. DNA
sequences of the heptapeptide inserts, and the deduced

Fig. 4. (a, b, c) Binding of anti-Ah42 antibodies generated in C57BL/6 (a) and APP/Tg (b, c) mice to phage clones, isolated after screening of 7-mer (a, b) and
12-mer (c) phage display libraries (see details in Section 2). The experiments with isolated phage clones were repeated three times with the similar results.
DOD=experimental OD (binding to a phage clone displaying a peptide) minus background (binding to a control phage clone with irrelevant peptide). Number
of positive phage clones presented in parentheses.
G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–20 15

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–2016
amino acid sequences were determined. All selected phage
clones were amplified and used in ELISA to confirm
binding of selected phage clones with appropriate anti-Ah42
and control sera. Peptide inserts of 17 positive clones (from
22 selected) were identified. Sixteen clones have peptide
inserts comprised of the linear 3EFRH6 immunodominant
region of Ah42: seven clones had EFRH, five clones XFRH,
two clones EXRH, one clone XXRH, one clone EFRX (Fig.
4a). One of 17 positive clones had peptide insert YELPRLT,
non-homologous to immunodominant linear 3EFRH6 epit-
ope. However, this phage clone binds to Ah42 antibodies
very weakly (Fig. 4a). None of the negative phage clones
had homology with this immunodominant region (data not
shown). Similar results were generated with anti-Ah42 sera
isolated from high responder mice of H2k haplotype (data
not shown). Therefore, wild type mice of two haplotypes
predominately recognized the linear immunodominant
epitope EFRH spanning N-terminal amino acids 3–6.
3.3. Screening of phage display libraries against polyclonal
anti-Ab42 antibodies raised in APP/Tg 2576 mice
Next, we screened anti-Ah42 antisera from APP/Tg2576
mice against the same phage display peptide library
(heptapeptides) that had been used against antisera from
Fig. 5. (a, b, c, d) Binding of anti-Ah42 antisera (equivalent to 0.4 Ag/ml antibod
haplotypes (n=4) to MAG11 and MAG16 mimotopic peptides (mimotopes). Dilu
generated with monoclonal antibody 6E10.
wild type mice. Sixteen positive clones were selected from
biopanning and DNA (data not shown), and amino acid
sequences of their inserts were determined (Fig. 4b). Only 4
out of 16 ELISA-positive clones had the sequence EFRH,
and one clone had EFRH motif (XFXH). The inserts of the
other positive clones had sequences that did not show
homology with Ah42, suggesting that they may represent
peptides that mimic Ah42 epitopes. From these phage
clones, five had similar inserts with the GDXKRTT motif:
four clones with GDLKRTT sequence and one clone with
GDFKRTT sequence. To confirm these results, we screened
anti-Ah42 antisera from APP/Tg mice against a phage
display peptide library of linear dodecapeptides. Thirteen
ELISA-positive clones were generated, and only two of
them had the EFRH sequence motif, whereas all the other
clones had inserts that lacked homology with the Ah42
peptide. Interestingly, two clones had the sequence LDLK-
RTTTTHTS (two clones). Thus, DXKRTT motif was
encountered in seven phage clones isolated collectively
from two different libraries: GDFKRTT (one clone),
GDLKRTT (four clones), and LDLKRTTTTHTS (Fig.
4b,c). Thus, anti-Ah42 sera from APP/Tg mice recognized
predominantly phage clones carrying non-self peptides
(mimotopes) and very few clones containing the linear
epitope EFRH of Ah42.
ies) generated in individual APP/Tg (n=6) and wild type mice of different
tions of antisera in ELISA were adjusted according to the standard curve

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–20 17
3.4. Binding of mimotopic peptide MAG11 or MAG16 to anti-
Ab42 sera isolated from mice of different haplotypes and
APP/Tg animals
Two of the mimotopes with DLKRTT motif identified in
screening of phage display libraries against anti-Ah42 sera
from APP/Tg 2576 mice were synthesized as following: the
7-mer (GDLKRTT) and 12-mer (LDLKRTTTTHTS) pep-
tides cores plus three amino acids from the N-terminal
region of the minor coat protein of the M13 phage (GGG)
and Cys residue (MAG11 and MAG16). Along with these
peptides, we generated MAG11 and MAG16 peptides
conjugated to OVA and all four peptides were used for
detection of binding to anti-Ah42 antibodies from wild type
and APP/Tg 2576 immunized mice. Because titers of anti-
human Ah42 antibodies in the sera of APP/Tg and wild type
mice were very different (see Fig. 1), we first determined the
concentrations of these antibodies in the antisera of
individual BALB/c, C57BL6, F1B6SJL, and APP/Tg mice
using a standard curve generated with monoclonal antibody
6E10 (data not shown). Immunization of C57BL6 and APP/
Tg 2576 mice with fibrillar Ah42 induced approximately six
times less anti-Ah42 antibodies than in BALB/c and
F1B6SJL animals (41.6F9.01 vs. 250.2F31.77 Ag/ml),
and these data correlated with results previously reported
with immunization of C57BL6 (H2b) and B6D2F1 (H2bxd)
mice by Ah immunogen (Spooner et al., 2002). Accord-
ingly, to compare binding of anti-Ah42 antibodies to MAG11
and MAG16 peptides, we used approximately 0.4 Ag/ml
anti-Ah42 antibodies in each ELISA. We demonstrated that
anti-Ah42 antibodies from two APP/Tg mice bound to
MAG11 and MAG16 peptides strongly. Antisera from two
other mice bound these peptides weakly, whereas the sera
from the remaining two mice did not show significant
binding (Fig. 5a). Anti-Ah42 sera from F1B6SJL mice also
Fig. 6. Conjugation of MAG11 and MAG16 to the carrier protein (OVA) had
masked some antigenic determinants of mimotopes. Binding of anti-Ah42
antisera (equivalent to 0.4 Ag/ml of antibodies) generated in APP/Tg (n=4,
responders only) mice to MAG11, and to OVA–MAG11 or OVA–MAG16,
was measured by ELISA three times with similar results. The data analysis
(see Section 2) showed with statistical significance ( Pb0.05) that
conjugation with OVA inhibited binding of the antisera to MAG peptides.
reacted to MAG11 and MAG16 peptides. One animal
responded significantly, whereas the other three mice had
a very weak response (Fig. 5b). All four BALB/c and the
four C57BL6 animals immunized with fibrillar Ah42
peptide did not recognize MAG11 and MAG16 mimotope
peptides (Fig. 5c,d). Next, we compared binding of four
positive antisera from APP/Tg mice (two high and two
intermediate responders) to MAG11 directly or to MAG11
and MAG16 conjugated with OVA. It appears that anti-Ah42
antibodies bound to MAG11 peptide significantly (pb0.05)
better than to MAG11–OVA or MAG16–OVA coated plates
(Fig. 6).
4. Discussion
Conformation-specific monoclonal anti-Ah42 antibodies
generated in APP/Tg mice and AD patient have been
previously described (Gaskin et al., 1993a; O’Nuallain and
Wetzel, 2002). Data recently generated by our group also
suggests that active immunization of wild type mice with
fibrillar human Ah42 induces antibodies that may recognize
conformational antigenic determinants on this peptide
(Cribbs et al., 2003). More recently, it was demonstrated
that rabbits injected with fibrillar self-Ah42 peptide induced
antibodies that bind to fibrillar form of Ah42 about 1000-
fold stronger than to linear peptide Ah1–8 (Miller et al.,
2003). These results collectively suggest that the full-length
fibrillar Ah42 contains some conformational antigenic
determinant/s that is not present in the shorter linear
peptides. In this study we directly investigated the presence
of such conformational epitope/s (mimotope/s) on fibrillar
Ah42 peptide using mouse polyclonal anti-Ah42 sera and
phage display library technique.
First, we raised polyclonal anti-fibrillar Ah42 antibodies
in wild type mice of four different haplotypes and APP/Tg
2576 animals, and detected the titer of these antibodies
(Fig. 1). Next, we analyzed the binding of antibodies to
different short linear peptides or fibrillar Ah42. Because
adsorption of a peptide to an ELISA plate may mask some
of the peptide’s epitopes, we decided to detect the presence
of conformational antigenic determinants in fibrillar Ah42
by using a competition assay where the small peptides are
in solution and the fibrillar Ah42 is bound to the ELISA
plate. As we expected from our previous data (Cribbs et al.,
2003), fibrillar Ah42 peptide almost completely inhibited
binding of antisera from all wild type and APP/Tg mice to
the bound Ah42. Only two short peptides, Ah1–15 and Ah1–
28, also had substantial inhibitory activity, although it was
less than that for fibrillar Ah42, specifically in case of APP/
Tg mice (Fig. 2). These data were confirmed with antisera
from APP/Tg mice when we used different concentrations
of Ah1–15, Ah6–20, Ah11–25+ 16–30+21–35+26–42, and fibrillar
Ah42 peptides (Fig. 3). Thus, the results from competition
assays argue for the presence of conformation-specific
polyclonal anti-Ah42 antibodies generated in wild type

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–2018
mice of different haplotypes, and especially in APP/Tg
2576 mice.
To identify linear peptides that mimic conformational
epitope/s within fibrillar Ah42 we utilized the phage display
technique which has proven useful in identifying mimotopes
of conformational epitopes. Antisera from C57BL6 wild
type mice frequently recognized the linear immunodominant
epitope EFRH spanning the N-terminal amino acids 3–6 of
Ah42. In total, 16 positive phage clones were identified, and
from all of these clones seven had the EFRH motif (Fig. 4a).
Previously, phage clones carrying the 3EFRH6 epitope of
Ah42 were described by binding to two different mono-
clonal antibodies (Frenkel et al., 1998). In our report, we
demonstrated that polyclonal anti-fibrillar Ah42 serum from
wild type mice of two different haplotypes (low and high
responders) also recognized this linear antigenic determi-
nant of Ah42 (Fig. 4a and data not shown, respectively). In
this mouse model, human fibrillar Ah42 is not a self-antigen,
so it was not surprising that the majority of antibodies
recognize the N-terminal region of Ah42 that differs from
the rodent sequence by three aa at positions 5, 10, and 13
(Hilbich et al., 1991). Thus, at least some of the immune
responses were likely due to recognition of the non-self
human residues in the human peptide used for immuniza-
tion. Next, we decided to screen two different phage
libraries against anti-fibrillar Ah42 serum from APP/Tg
2576 mice. In two separate experiments we detected only 6
out of 29 ELISA positive clones that had a sequence EFRH,
and one clone that had a EFRH motif (XFXH). These results
were similar to those generated with M13 phage display
system and polyclonal antibodies from rabbits immunized
with fibrillar Ah42 (Miller et al., 2003) (rabbit and human
Ah42 are identical). These authors identified five phage
clones that expressed peptides with a 2AEFRH6 motif.
However, we also detected 23 positive clones that did not
have any homology with the Ah42 immunogen. Whether or
not these data represent differences in animal models
remains to be determined, however in our experiments the
DXKRTT motif was encountered in seven phage clones
isolated from two different libraries. Thus, antisera from
wild type animals recognized primarily the EFRH epitope
(Fig. 4a), whereas anti-Ah42 sera from APP/Tg mice
recognized very few clones carrying the linear epitope
EFRH (Fig. 4b). On the contrary, the sera from immunized
APP/Tg mice identified predominantly phage clones carry-
ing mimotopes of fibrillar Ah42 (Fig. 4b,c), which suggests
that different antibody repertoires were induced following
Ah-immunization of transgenic and wild type mice.
Next we demonstrated binding of anti-fibrillar Ah42 sera
from APP/Tg mice to the mimotopes GDLKRTT and
LDLKRTTTTHTS, frequently identified in screening of
two phage display libraries with anti-Ah42 sera from APP/
Tg mice. In an attempt to conserve the native structure of
these peptides, we added three amino acids from N-terminal
region of the minor coat protein of the M13 phage (GGG) to
the C-terminus. In addition, we added a cysteine residue to
facilitate conjugation to the carrier protein. Therefore, we
generated MAG11 and MAG16 peptides along with MAG11–
OVA and MAG16–OVA conjugates, which were used for
detection of binding by anti-Ah42 antisera. To compare
binding of anti-Ah42 antibodies from each mouse to MAG11
and MAG16 peptides in ELISA, we used anti-Ah42 sera
equivalent to approximately 0.4 Ag/ml of antibody. Sera
from at least two APP/Tg mice recognized MAG11 and
MAG16 strongly, whereas sera from two other animals
bound these peptides weakly, and the remaining two sera
did not show appreciable binding (Fig. 5a). Surprisingly,
one out of four B6SJLF1 mice, that shared the same MHC
haplotype with the APP/Tg mice, also had antibodies
binding to MAG11 and MAG16 peptides. In contrast, anti-
Ah42 sera from two other mice strains (BALB/c or C57BL6)
with MHC haplotypes different from APP/Tg animals did
not recognize either MAG11 or MAG16 (Fig. 5c,d).
These results identified for the first time the linear
foreign peptides that mimic fibrillar Ah42 without having
any homology. Molecular mimicry is defined as similar
structures within very different macromolecules. The con-
cept of molecular mimicry is taken primarily from the
classical biochemistry of receptor–ligand or antibody–
antigen interactions. Previously, we reported that immuni-
zation with a peptide that mimics a carbohydrate molecule
induced antibodies against a viral glycoprotein (Agadjanyan
et al., 1997). In the current report we demonstrate that
conformation-specific polyclonal anti-Ah42 antibodies can
bind to non-homologous linear peptides, which mimic
conformational epitopes of fibrillar Ah42. Thus, we have
identified antigenic mimotopes of fibrillar Ah42. We realize
that antigenic and immunogenic properties of epitope/
mimotopes may not correlate directly (Deroo and Muller,
2001; Van Regenmortel, 2001), and in order to demonstrate
the immunogenicity of mimotopic peptides we need to
determine whether anti-Ah42 antibodies are induced after
immunization of mice with these peptides. However, first
we analyzed whether conjugation with the carrier protein
influences binding of anti-Ah42 antisera from APP/Tg mice
to the mimotopic peptides MAG11 or MAG16. Anti-Ah42
antibodies bound to non-conjugated MAG peptide signifi-
cantly better than to conjugated Mag11 and MAG16 (Fig. 6).
Therefore, conjugation of these peptides with OVA appears
to mask some antigenic determinants of the mimotopes.
While other carrier proteins and/or methods of conjuga-
tion may be required for preparation of immunogenic
versions of mimotopes and testing for their antigenic
properties, we believe that it will be extremely important
to detect both antigenic and immunogenic mimotopes in
fibrillar Ah42 in humans. It is likely that different organisms
may see the mimotopic structures of the same antigen quite
differently. The characterization of antibodies generated
after vaccination of AD patients and immunization of APP/
Tg mice supports this hypothesis. Although the Ah42
epitope recognized by human antibodies was not determined
directly, it was shown that, while sera from APP/Tg mice

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–20 19
recognized Ah42 monomers, tetramers, hexamers and large
oligomers, the human antibodies appear to bind only to the
aggregated form of Ah42 (Hock et al., 2002; McLaurin et
al., 2002; Spooner et al., 2002; Dodart et al., 2003).
Accordingly, we are planning to screen the same phage
display libraries against human anti-Ah42 sera in order to
detect human mimotopes of conformational epitopes in h-amyloid fibrils. Identification of linear peptides that mimic
human Ah42 structures may provide significant advantages
over immunization with fibrillar Ah42. First, the mimotope
will be non-toxic. Second, the immunogenicity may be
significantly enhanced in humans, because it will be a non-
self antigen. Third, the mimotopic short peptide (non-self B
cell epitope) can be attached to a foreign T cell epitope to
amplify the humoral immune response to the antigenic
mimotopes. Thus, the identification of human mimotopes of
fibrillar or possibly oligomeric forms of Ah42 may provide
novel molecular targets for generating a safe and effective
epitope vaccine against AD.
Acknowledgements
This work was supported by R01 grants from NIH
AG20241 for D.H. Cribbs and M.G. Agadjanyan; AI
44809 for M.G. Agadjanyan, and IIRG grant by Alzheimer’s
Association (IIRG-03-6279) for M.G. Agadjanyan and D.H.
Cribbs.
References
Agadjanyan, M., Luo, P., Westerink, M.A.J., Carey, L.A., Hutchins, W.,
Steplewski, Z., Weiner, D.B., Kieber-Emmons, T., 1997. Peptide
mimicry of carbohydrate epitopes on human immunodeficiency virus.
Nat. Biotechnol. 15, 547–551.
Bard, F., Cannon, C., Barbour, R., Burke, R.L., Games, D., Grajeda, H.,
Guido, T., Hu, K., Huang, J., Johnson-Wood, K., Khan, K., Kholodenko,
D., Lee,M., Lieberburg, I., Motter, R., Nguyen,M., Soriano, F., Vasquez,
N., Weiss, K., Welch, B., Seubert, P., Schenk, D., Yednock, T., 2000.
Peripherally administered antibodies against amyloid beta-peptide enter
the central nervous system and reduce pathology in a mouse model of
Alzheimer disease. Nat. Med. 6, 916–919.
Cribbs, D.H., Pike, C.J., Weinstein, S.L., Velazquez, P., Cotman, C.W.,
1997. h-Amyloid stereoisomers exhibit similar structural and biological
properties: implications for mechanisms of toxicity. J. Biol. Chem. 272,
7431–7436.
Cribbs, D.H., Ghochikyan, A., Tran, M., Vasilevko, V., Petrushina, I.,
Sadzikava, N., Kesslak, P., Kieber-Emmons, T., Cotman, C.W.,
Agadjanyan, M.G., 2003. Adjuvant-dependent modulation of Th1 and
Th2 responses to immunization with beta-amyloid. Int. Immunol. 15,
505–514.
DeMattos, R.B., Bales, K.R., Cummins, D.J., Dodart, J.C., Paul, S.M.,
Holtzman, D.M., 2001. Peripheral anti-A beta antibody alters CNS and
plasma A beta clearance and decreases brain A beta burden in a mouse
model of Alzheimer’s disease. Proc. Natl. Acad. Sci. U. S. A. 98,
8850–8855.
Deroo, S., Muller, C.P., 2001. Antigenic and immunogenic phage displayed
mimotopes as substitute antigens: applications and limitations. Comb.
Chem. High Throughput Screen. 4, 75–110.
Dickey, C.A., Morgan, D.G., Kudchodkar, S., Weiner, D.B., Bai, Y., Cao,
C., Gordon, M.N., Ugen, K.E., 2001. Duration and specificity of
humoral immune responses in mice vaccinated with the Alzheimer’s
disease-associated beta-amyloid 1– 42 peptide. DNA Cell, 723–729.
Dodart, J.C., Bales, K.R., Paul, S.M., 2003. Immunotherapy for Alzheim-
er’s disease: will vaccination work? Trends Mol. Med. 9, 85–87.
Frenkel, D., Balass, M., Solomon, B., 1998. N-terminal EFRH sequence of
Alzheimer’s beta-amyloid peptide represents the epitope of its anti-
aggregating antibodies. J. Neuroimmunol. 88, 85–90.
Gaskin, F., Finley, J., Fang, Q., Xu, S., Fu, S.M., 1993a. Human antibodies
reactive with beta-amyloid protein in Alzheimer’s disease. J. Exp. Med.
177, 1181–1186.
Gaskin, F., Xu, S., Finley, J., 1993b. Human antibodies reactive with beta-
amyloid in Alzheimer’s disease. J. Immunol. 150, 238A.
Gevorkian, G., Manoutcharian, K., Govezensky, T., Cano, J.A., Domi-
nguez, V., Santamaria, H., Larralde, C., 2000. Identification of
mimotopes of platelet autoantigens associated with autoimmune
thrombocytopenic purpura. J. Autoimmun. 15, 33–40.
Gomez-Roman, V.R., Cao, C., Bai, Y., Santamaria, H., Acero, G.,
Manoutcharian, K., Weiner, D.B., Ugen, K.E., Gevorkian, G., 2002.
Phage-displayed mimotopes recognizing a biologically active anti-HIV-
1 gp120 murine monoclonal antibody. J. Acquir. Immune Defic. Syndr.
31, 147–153.
Hardy, J., 2002. Testing times for the bamyloid cascade hypothesisQ.Neurobiol. Aging 23, 1073–1074. (author reply 1101–1075).
Hardy, J., Selkoe, D.J., 2002. The amyloid hypothesis of Alzheimer’s
disease: progress and problems on the road to therapeutics. Science 297,
353–356.
Hilbich, C., Kisters-Woike, B., Reed, J., Masters, C.L., Beyreuther, K.,
1991. Human and rodent sequence analogs of Alzheimer’s amyloid beta
A4 share similar properties and can be solubilized in buffers of pH 7.4.
Eur. J. Biochem. 201, 61–69.
Hock, C., Konietzko, U., Papassotiropoulos, A., Wollmer, A., Streffer, J.,
Von Rotz, R.C., Davey, G., Moritz, E., Nitsch, R.M., 2002. Generation
of antibodies specific for beta-amyloid by vaccination of patients with
Alzheimer disease. Nat. Med. 8, 1270–1275.
Janus, C., Chishti, M.A., Westaway, D., 2000. Transgenic mouse models of
Alzheimer’s disease. Biochim. Biophys. Acta 1502, 63–75.
Kayed, R., Head, E., Thompson, J.L., McIntire, T.M., Milton, S.C.,
Cotman, C.W., Glabe, C.G., 2003. Common structure of soluble
amyloid oligomers implies common mechanism of pathogenesis.
Science 300, 486–489.
Lemere, C.A., Maron, R., Spooner, E.T., Grenfell, T.J., Mori, C., Desai, R.,
Hancock, W.W., Weiner, H.L., Selkoe, D.J., 2000. Nasal Ah treatment
induces anti-Ah antibody production and decreases cerebral amyloid
burden in PD-APP mice. Ann. N. Y. Acad. Sci. 920, 328–331.
Lemere, C.A., Maron, R., Selkoe, D.J., Weiner, H.L., 2001. Nasal
vaccination with beta-amyloid peptide for the treatment of Alzheimer’s
disease. DNA Cell Biol. 20, 705–711.
Manoutcharian, K., Gevorkian, G., Cano, A., Almagro, J.C., 2001. Phage
displayed biomolecules as preventive and therapeutic agents. Curr.
Pharm. Biotechnol. 2, 217–223.
McLaurin, J., Cecal, R., Kierstead, M.E., Tian, X., Phinney, A.L., Manea,
M., French, J.E., Lambermon, M.H., Darabie, A.A., Brown, M.E.,
Janus, C., Chishti, M.A., Horne, P., Westaway, D., Fraser, P.E., Mount,
H.T., Przybylski, M., St George-Hyslop, P., 2002. Therapeutically
effective antibodies against amyloid-beta peptide target amyloid-beta
residues 4 –10 and inhibit cytotoxicity and fibrillogenesis. Nat. Med. 8,
1263–1269.
Mertens, P., Walgraffe, D., Laurent, T., Deschrevel, N., Letesson, J.J., De
Bolle, X., 2001. Selection of phage-displayed peptides recognised by
monoclonal antibodies directed against the lipopolysaccharide of
Brucella. Int. Rev. Immunol. 20, 181–199.
Miller, D.L., Currie, J.R., Mehta, P.D., Potemska, A., Hwang, Y.W., Wegiel,
J., 2003. Humoral immune response to fibrillar beta-amyloid peptide.
Biochemistry 42 (40), 11682–11692.

G. Gevorkian et al. / Journal of Neuroimmunology 156 (2004) 10–2020
Monsonego, A., Weiner, H.L., 2003. Immunotherapeutic approaches to
Alzheimer’s disease. Science 302, 834–838.
Morgan, D., Diamond, D.M., Gottschall, P.E., Ugen, K.E., Dickey, C.,
Hardy, J., Duff, K., Jantzen, P., DiCarlo, G., Wilcock, D., Connor, K.,
Hatcher, J., Hope, C., Gordon, M., Arendash, G.W., 2000. A beta
peptide vaccination prevents memory loss in an animal model of
Alzheimer’s disease. Nature 408, 982–985.
O’Nuallain, B., Wetzel, R., 2002. Conformational Abs recognized a generic
amyloid fibril epitope. PNAS 99, 1485–1490.
Petrushina, I., Tran, M., Sadzikava, N., Ghochikyan, A., Vasilevko, V.,
Agadjanyan, M.G., Cribbs, D.H., 2003. Importance of IgG2c isotype in
the immune response to b-amyloid in APP/Tg mice. Neurosci. Lett.
338, 5–8.
Schenk, D., 2002. Opinion: amyloid-beta immunotherapy for Alzheimer’s
disease: the end of the beginning. Nat. Rev. Neurosci. 3, 824–828.
Schenk, D., Barbour, R., Dunn, W., Gordon, G., Grajeda, H., Guido, T., Hu,
K., Huang, J., Johnson-Wood, K., Khan, K., Kholodenko, D., Lee, M.,
Liao, Z., Lieberburg, I., Motter, R., Mutter, L., Soriano, F., Shopp, G.,
Vasquez, N., Vandevert, C., Walker, S., Wogulis, M., Yednock, T.,
Games, D., Seubert, P., 1999. Immunization with amyloid-beta
attenuates Alzheimer-disease-like pathology in the PDAPP mouse.
Nature 400, 173–177. [see comments].
Scott, J.K., Smith, G.P., 1990. Searching for peptide ligands with an epitope
library. Science 249, 386–390.
Selkoe, D.J., 2001. Alzheimer’s disease: genes, proteins, and therapy.
Physiol. Rev. 81, 741–766.
Solomon, B., Koppel, R., Hanan, E., Katzav, T., 1996. Monoclonal
antibodies inhibit in vitro fibrillar aggregation of the Alzheimer beta-
amyloid peptide. Proc. Natl. Acad. Sci. U. S. A. 93, 452–455.
Solomon, B., Koppel, R., Frankel, D., Hanan-Aharon, E., 1997. Disag-
gregation of Alzheimer beta-amyloid by site-directed mAb. Proc. Natl.
Acad. Sci. U. S. A. 94, 4109–4112.
Spooner, E.T., Desai, R.V., Mori, C., Leverone, J.F., Lemere, C.A., 2002.
The generation and characterization of potentially therapeutic Abeta
antibodies in mice: differences according to strain and immunization
protocol. Vaccine 21, 290–297.
Town, T., Tan, J., Sansone, N., Obregon, D., Klein, T., Mullan, M., 2001.
Characterization of murine immunoglobulin G antibodies against
human amyloid-b 1– 42. Neurosci. Lett. 307, 101–104.
Van Regenmortel, M.H., 2001. Antigenicity and immunogenicity of
synthetic peptides. Biologicals 29, 209–213.
Weiner, H.L., Selkoe, D.J., 2002. Inflammation and therapeutic vaccination
in CNS diseases. Nature 420, 879–884.





























